Submitted:
18 December 2025
Posted:
19 December 2025
You are already at the latest version
Abstract
Kynurenic acid (KYNA) is a byproduct of the kynurenine pathway, which breaks down the amino acid tryptophan. KYNA acts as an antagonist of glutamate ionotropic excitatory amino acid (EAA) receptors and α7 nicotinic acetylcholine receptors (nAChRs). There is evidence that KYNA plays a significant role in various pathological conditions and the ageing process. It has also been suggested that KYNA contributes to memory impairment. This study investigated the impact of L-kynurenine, D-cycloserine, and Cerebrolysin on KYNA synthesis in the liver homogenate of Helix pomatia snails in vitro and in their ganglia in vivo. Furthermore, a memory model was established using these snails, wherein tentacle shortening served as an indicator of memory activity. In vitro experiments on Helix pomatia demonstrated the significant impact of L-kynurenine and anti-dementia drugs on KYNA synthesis. KYNA levels increased significantly in the presence of L-kynurenine, a bio-precursor, in liver homogenate. However, KYNA formation decreased when anti-dementia drugs, including Cerebrolysin or D-cycloserine, were administered to the snails’ liver homogenate. L-kynurenine has been shown to impair memory in vivo in snails, but an anti-dementia drug has been demonstrated to reverse this effect. Significant inhibition of tentacle lowering was observed in response to L-kynurenine treatment, which corresponded with elevated KYNA levels in the central nervous system. Administering D-cycloserine or Cerebrolysin alongside L-kynurenine reversed its effects. The Helix pomatia memory model is a valuable tool for studying learning and memory in various conditions and in the presence of different pharmacological agents. A drug or natural extract that blocks KYNA synthesis could be considered an anti-dementia agent. It may also protect against ageing and delay damage to the central nervous system related to memory.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Compounds
2.2. Animals
2.3. Methods
2.3.1. Age Determination of Snails ARS—Middle-Aged Group of Snails
2.3.2. Preparation of Region and Homogenates
3. Biochemical Methods
3.1. KAT Assay
3.2. High-Performance Liquid Chromatography (HPLC) Method for KYNA Detection
3.3. Pharmacological Study—In Vitro
3.3.1. Synthesis of KYNA from L-Kynurenine in Snail Liver Homogenate
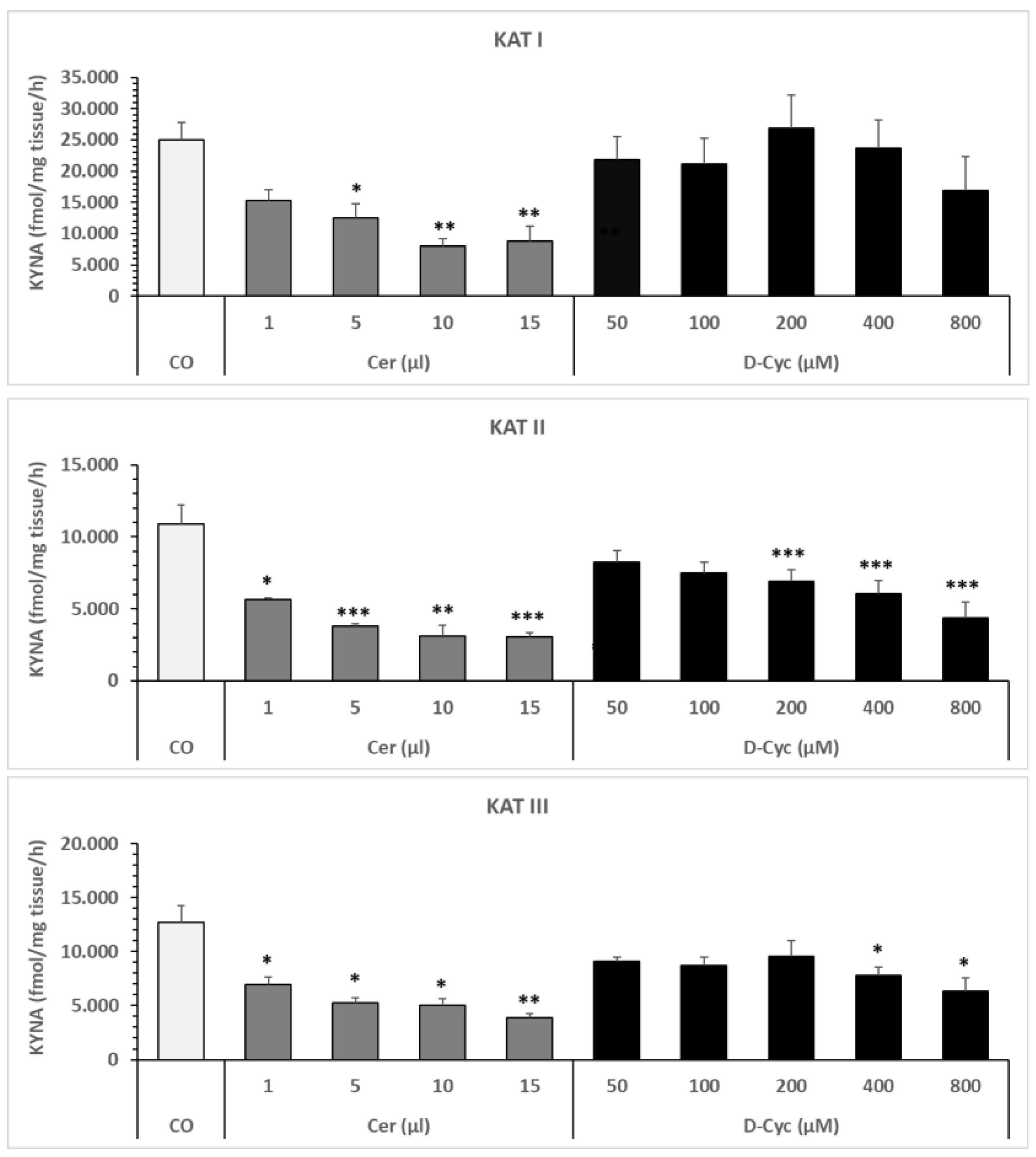
3.3.2. Effect of Cerebrolysin and D-Cycloserine on Snail Liver KAT I, KAT II, and KAT III Activities
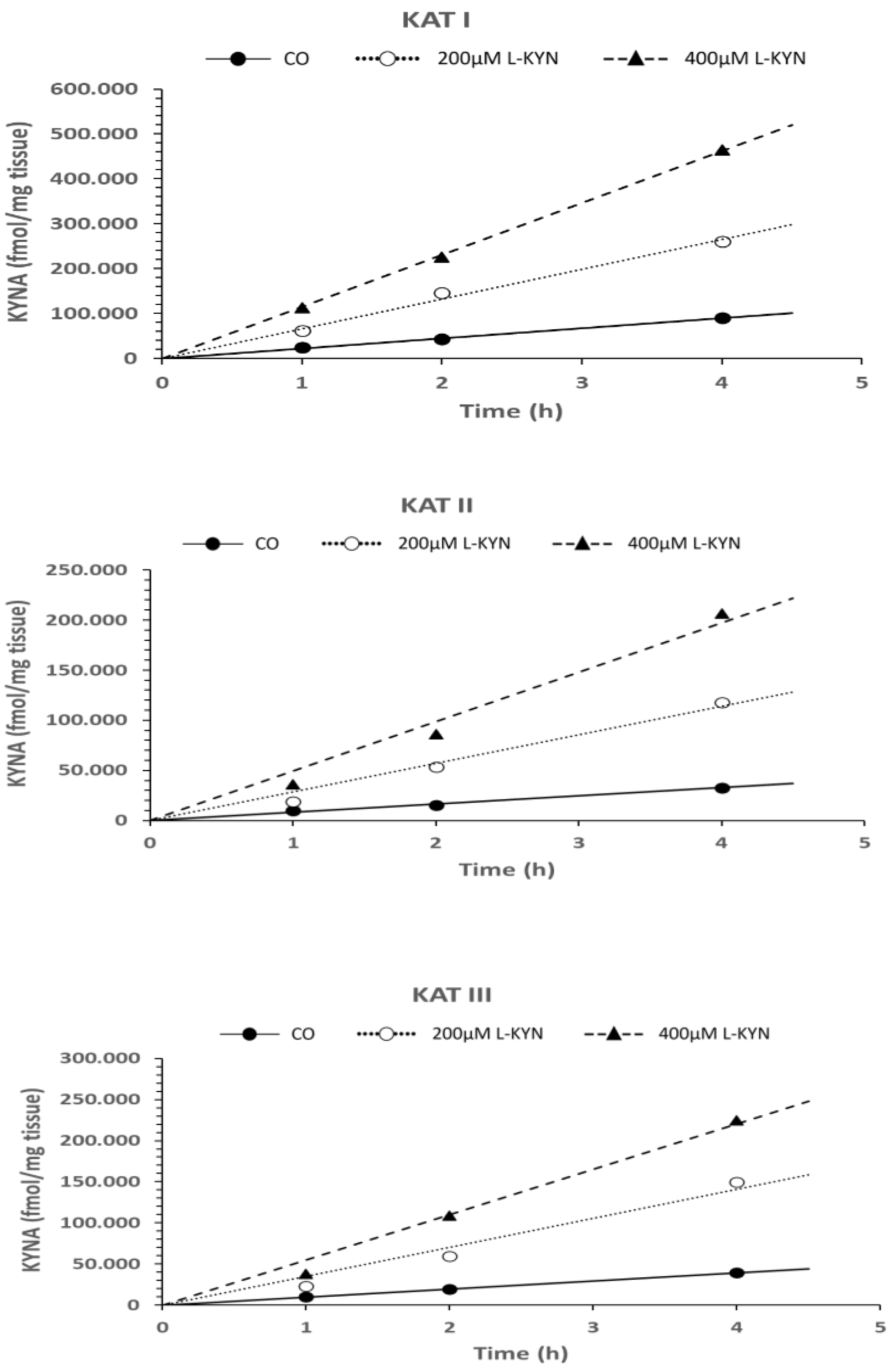
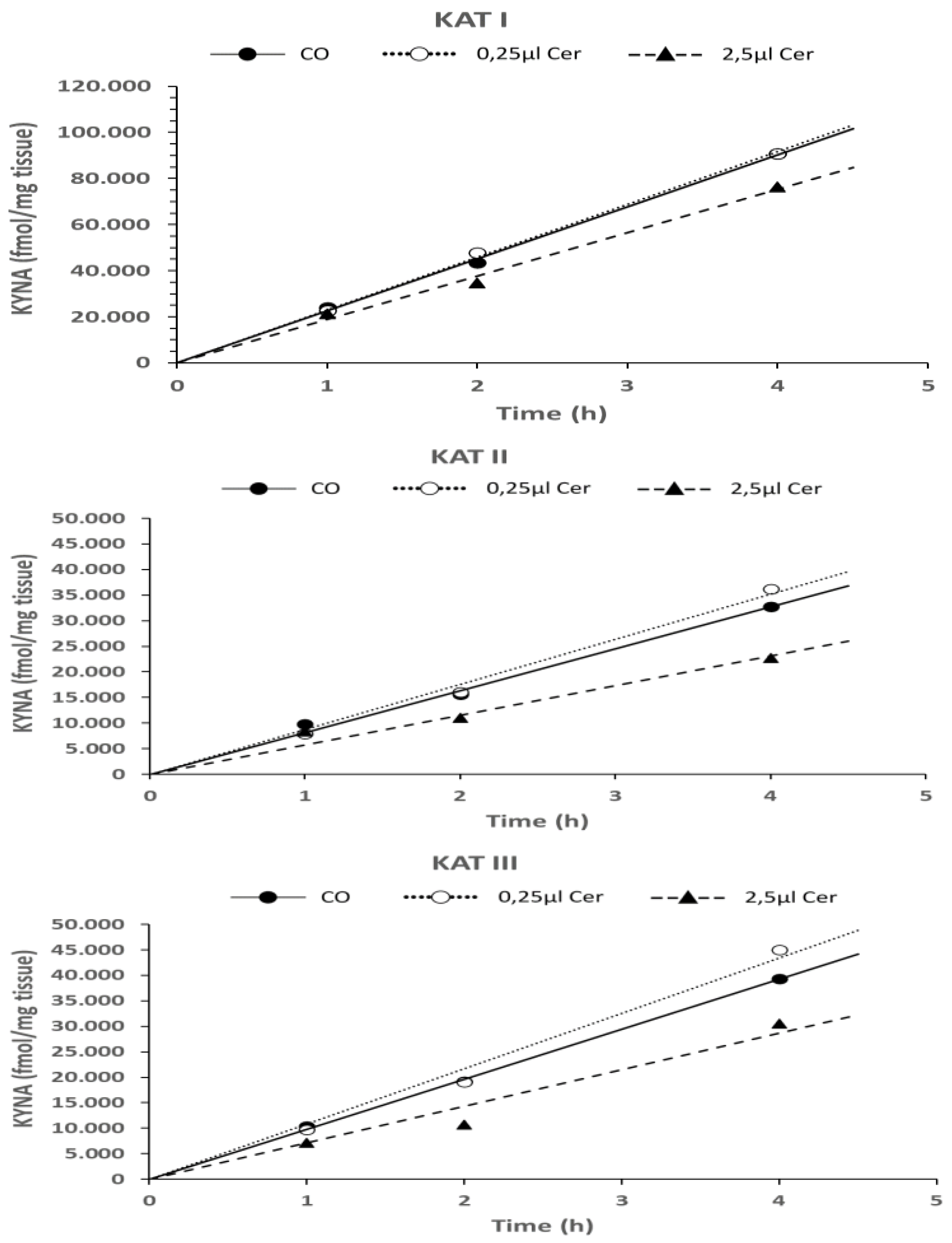
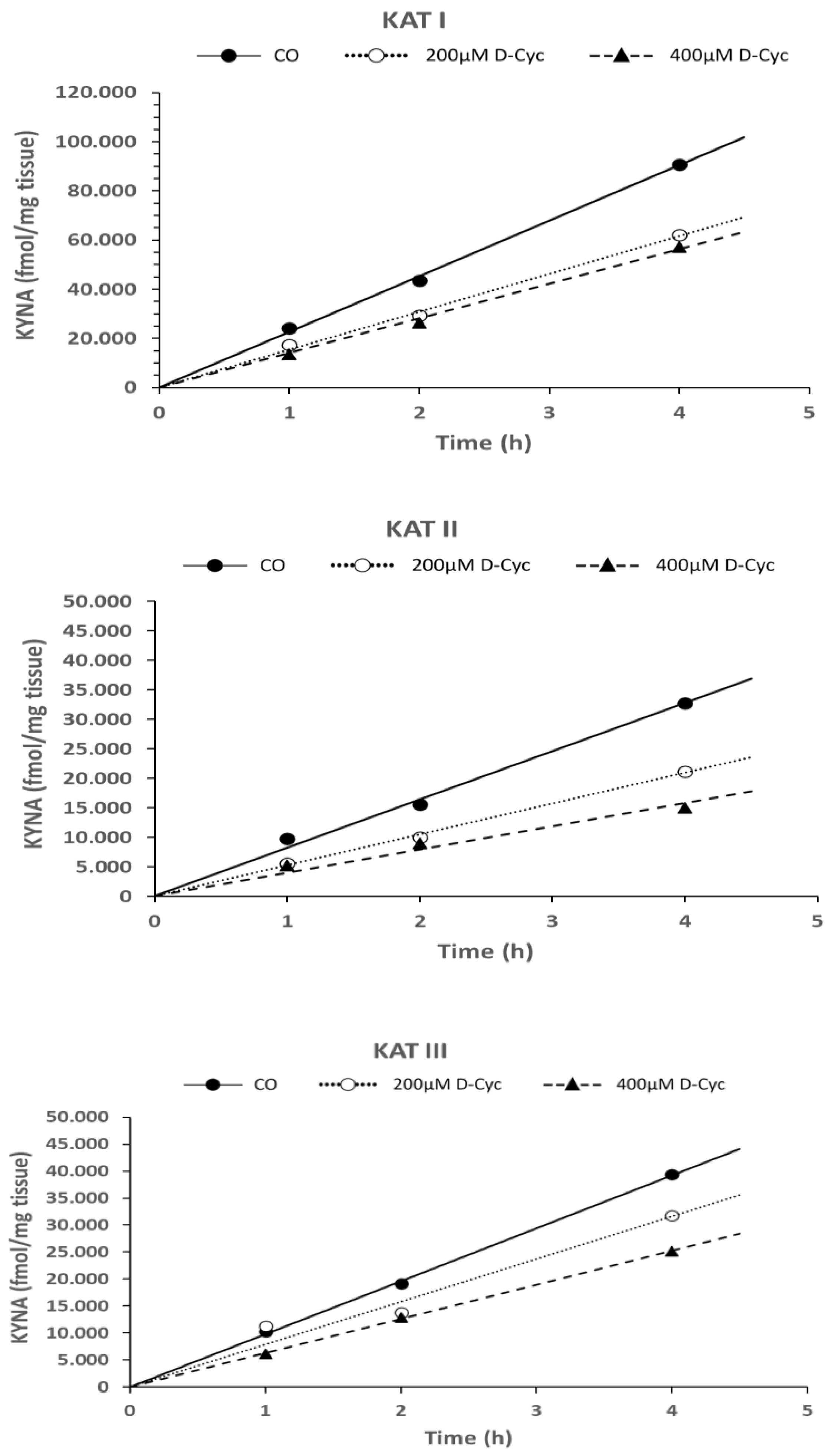
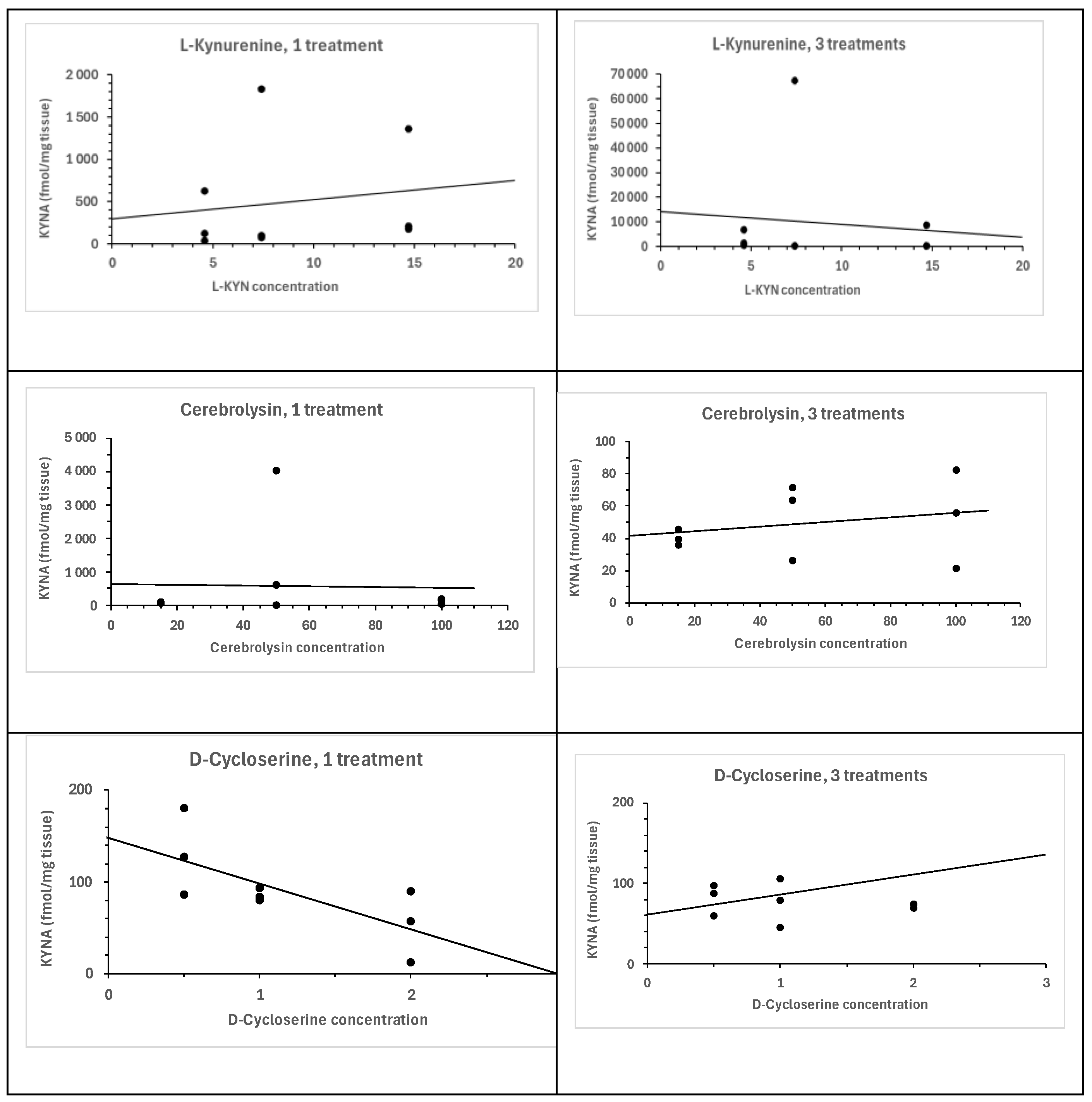
3.3.3. Time-Dependent Formation of KYNA in the Presence of L-Kynurenine, Cerebrolysin, and D-Cycloserine In Vitro
4. Pharmacological Study—In Vivo
4.1. Effect of Treatment with L-Kynurenine, Cerebrolysin, and D-Cycloserine on KYNA
4.2. Experimental Design and Pharmacological Treatment In Vivo
4.2.1. Behaviour
4.2.2. Experimental Treatment
4.2.3. Time Course and Conditioning
4.2.4. Testing
4.2.5. Video Analysis
5. Data Analysis
6. In Vitro Study Results
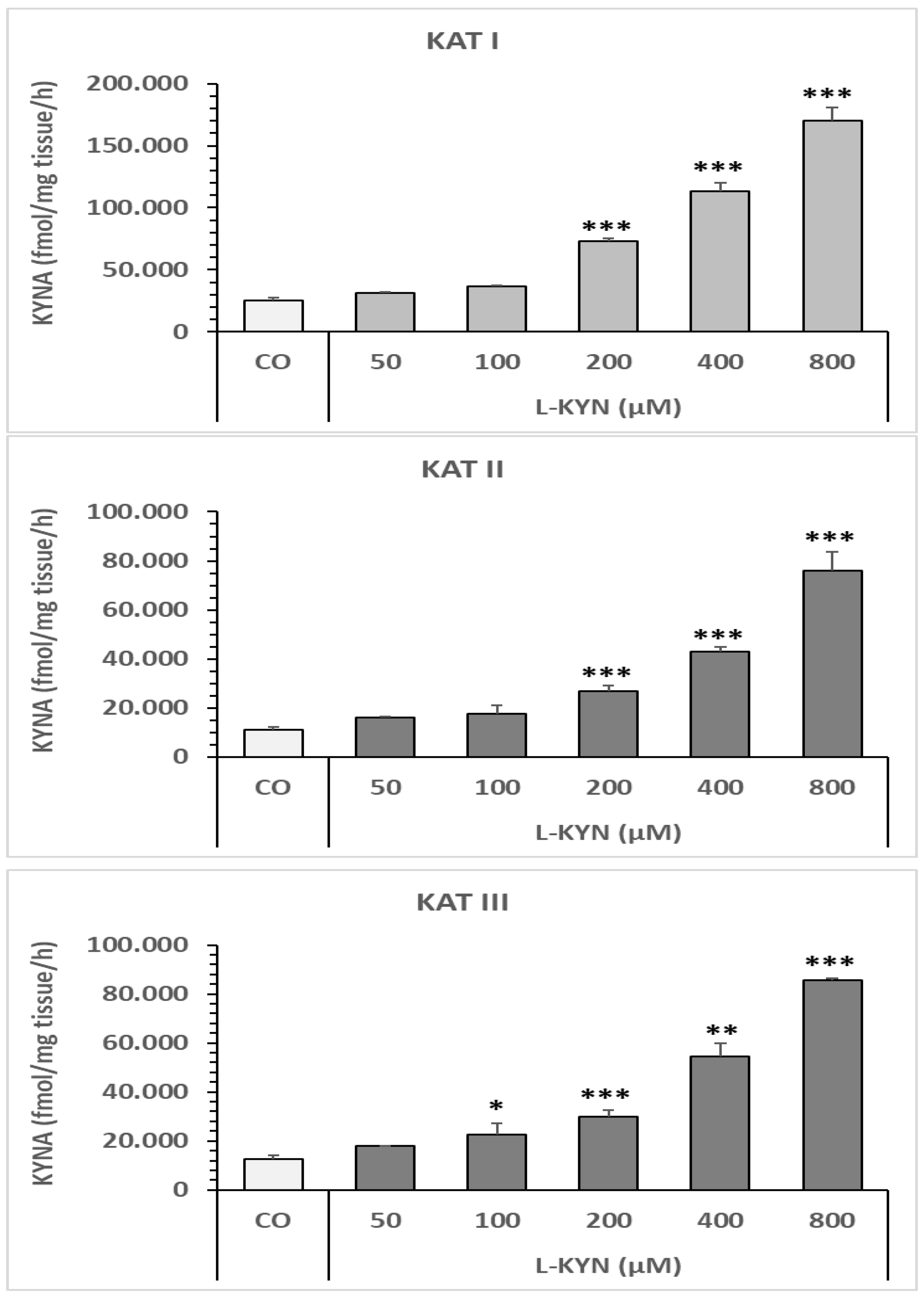
6.1. Dose–Response of L-Kynurenine and Anti-Dementia Drugs in an In Vitro Assay
6.1.1. Effect of L-Kynurenine on Snail Liver KAT I, KAT II, and KAT III In Vitro
6.1.2. Effect of Cerebrolysin or D-Cycloserine on Snail Liver KAT I, KAT II, and KAT III In Vitro
6.1.3. Time Dependence of KYNA Formation in the Presence of L-Kynurenine, Cerebrolysin, and D-Cycloserine in In Vitro Conditions

| Treatment | 1 h–2 h | 1 h–4 h | 2 h–4 h |
| CO | ns. | 0.001 | 0.009 |
| Cer 0.25 µL | 0.008 | 0.021 | ns |
| Cer 2.5 µL | ns | 0.012 | 0.029 |
| D-Cyc 200 µM | ns | 0.03 | ns |
| D-Cyc 400 µM | ns | 0.018 | 0.048 |
| L-Kyn 200 µM | ns | ns | ns |
| L-Kyn 400 µM | ns | 0.001 | 0.003 |
| Treatment | 1 h–2 h | 1 h–4 h | 2 h–4 h |
| CO | 0.007 | 0.004 | 0.007 |
| Cer 0.25 µL | ns | ns | 0.049 |
| Cer 2.5 µL | ns | ns | ns |
| D-Cyc 200 µM | ns | 0.004 | 0.013 |
| D-Cyc 400 µM | 0.016 | 0.041 | ns |
| L-Kyn 200 µM | 0.030 | 0.001 | 0.002 |
| L-Kyn 400 µM | 0.002 | <0.001 | <0.001 |
| Treatment | 1 h–2 h | 1 h–4 h | 2 h–4 h |
| CO | 0.034 | <0.001 | <0.001 |
| Cer 0.25 µL | ns | ns | ns |
| Cer 2.5 µL | ns | ns | ns |
| D-Cyc 200 µM | ns | 0.044 | ns |
| D-Cyc 400 µM | ns | ns | 0.023 |
| L-Kyn 200 µM | 0.038 | <0.001 | 0.001 |
| L-Kyn 400 µM | 0.027 | 0.001 | 0.004 |
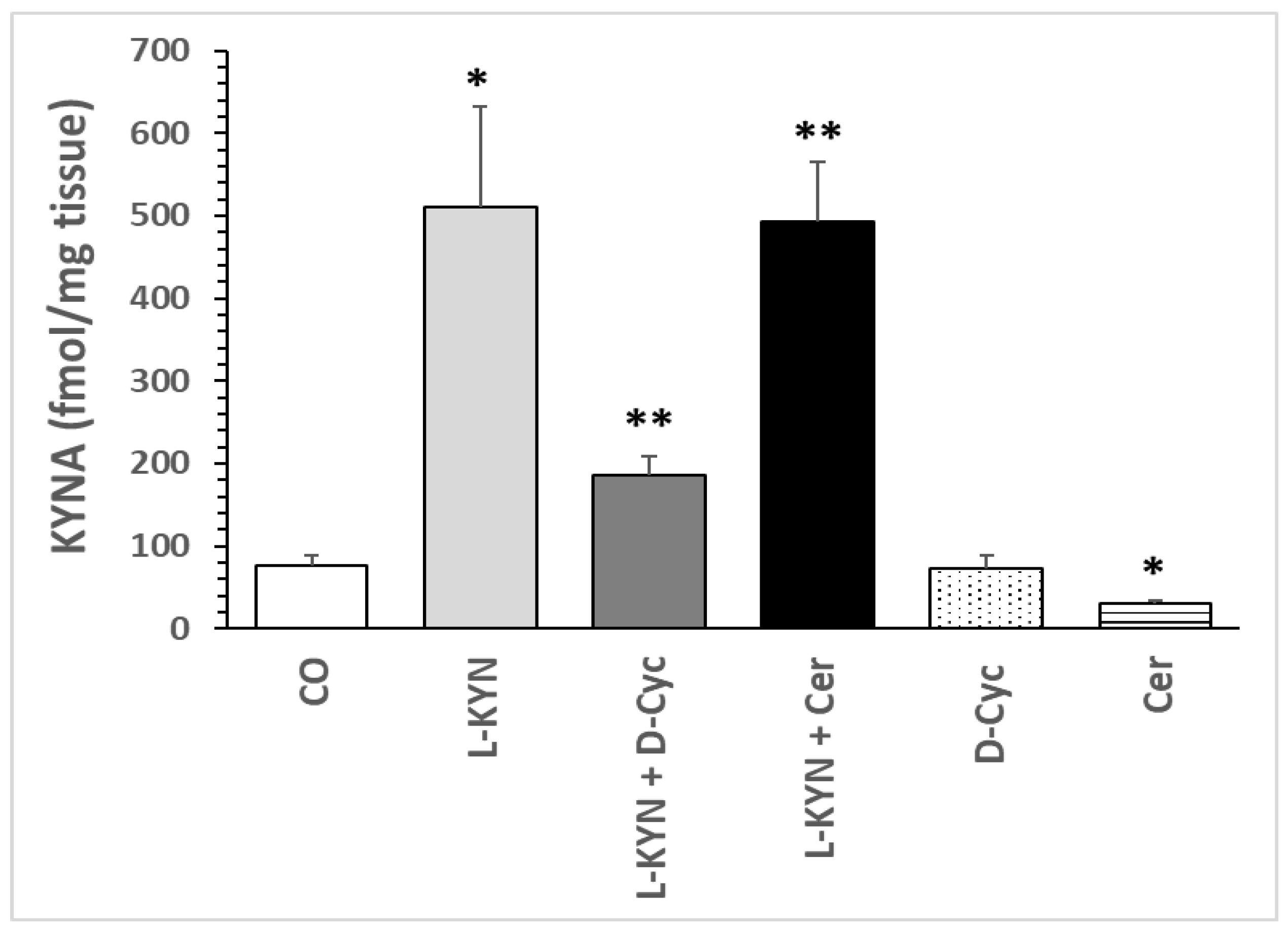
7. Effect on KYNA Formation in an In Vivo Study
7.1. Effect of Dose–Response of L-Kynurenine, Cerebrolysin and D-Cycloserine on KYNA Formation In Vivo
7.1.1. Effect of L-Kynurenine
7.1.2. Effect of Cerebrolysin
7.1.3. Effect of D-Cycloserine
8. Pharmacological Treatment of Snails
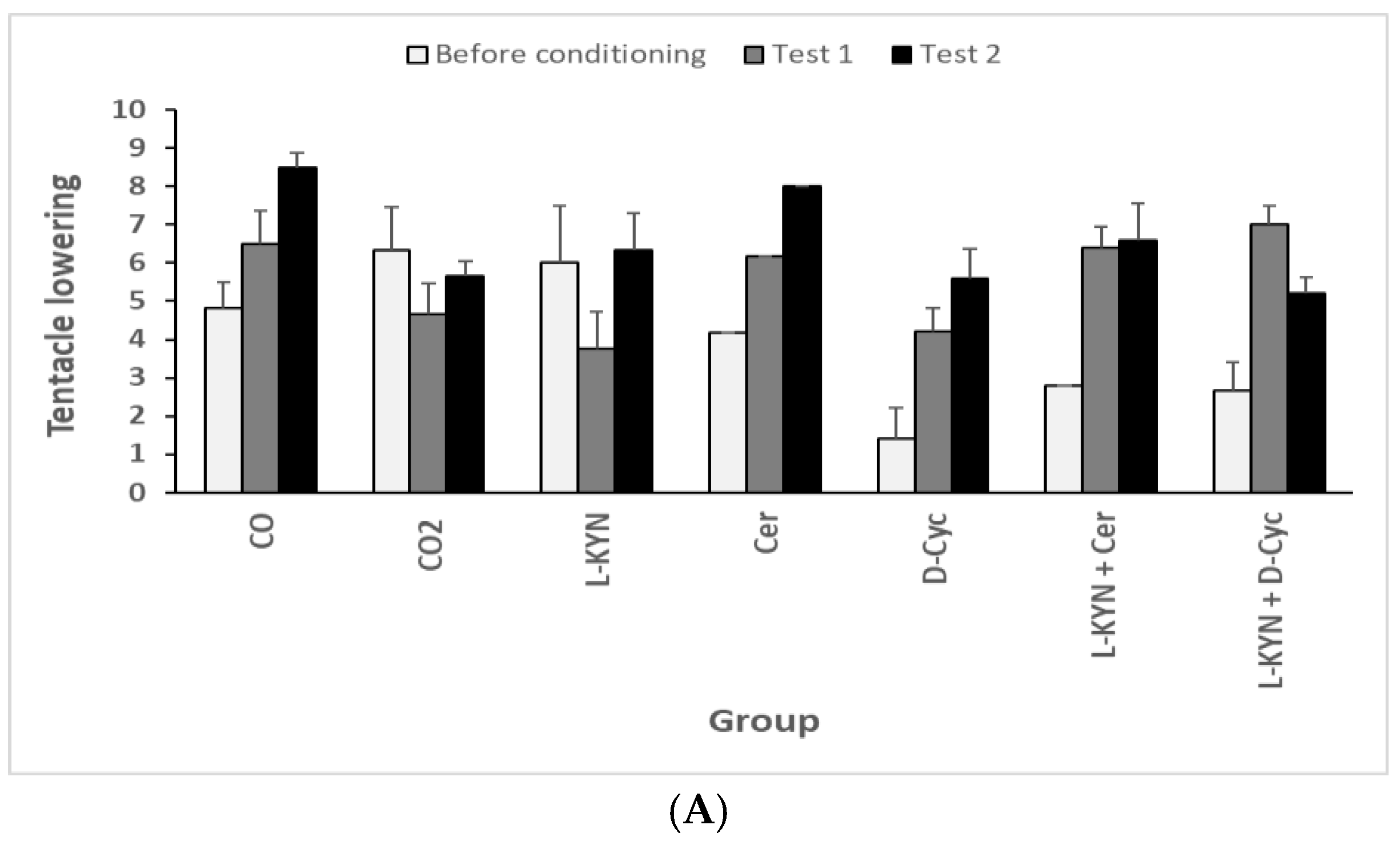
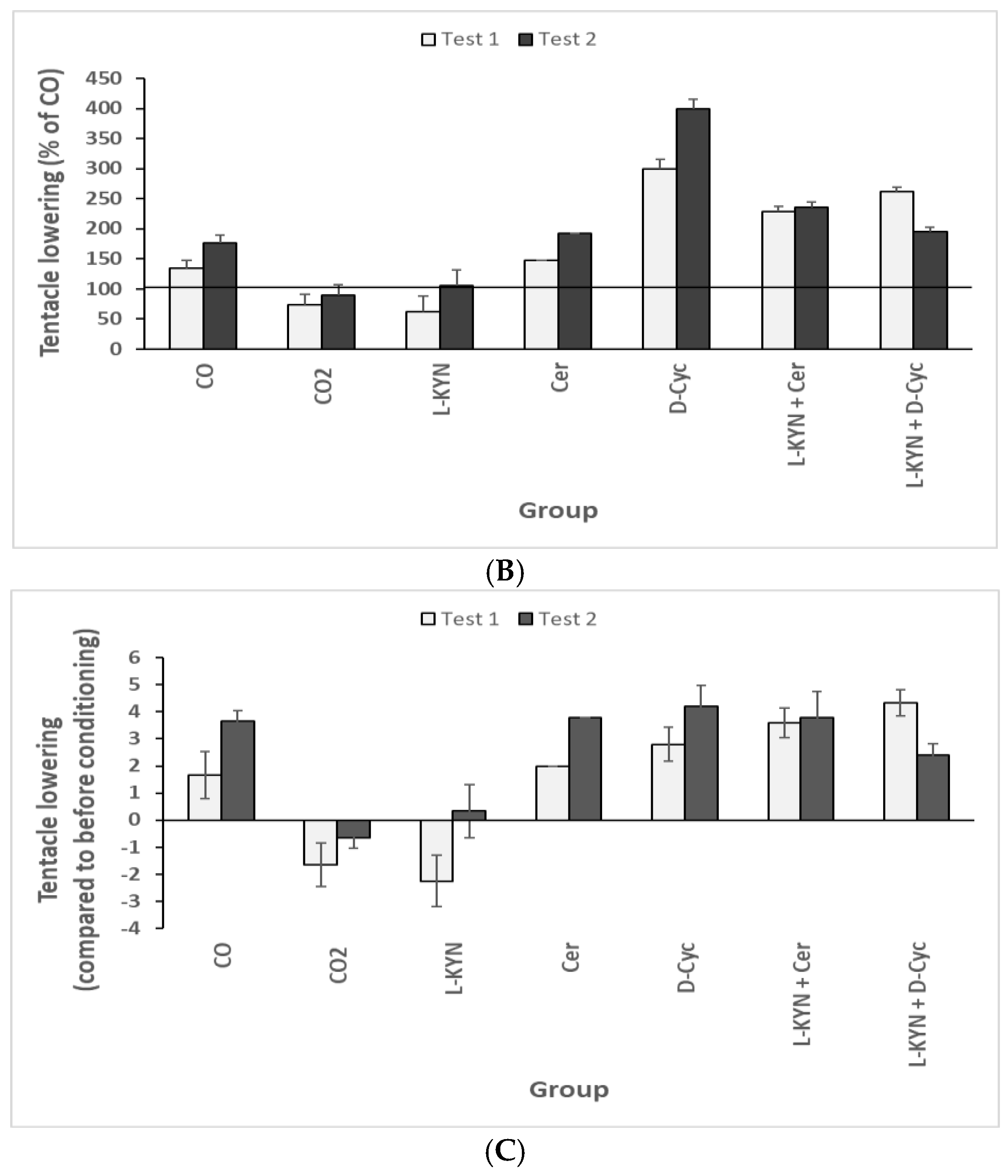
8.1. Effect of L-Kynurenine and an Anti-Dementia Drug on KYNA Formation and Tentacle Behaviour
8.1.1. Changes in KYNA
8.1.2. Behaviours-Alteration of Tentacle Lowering
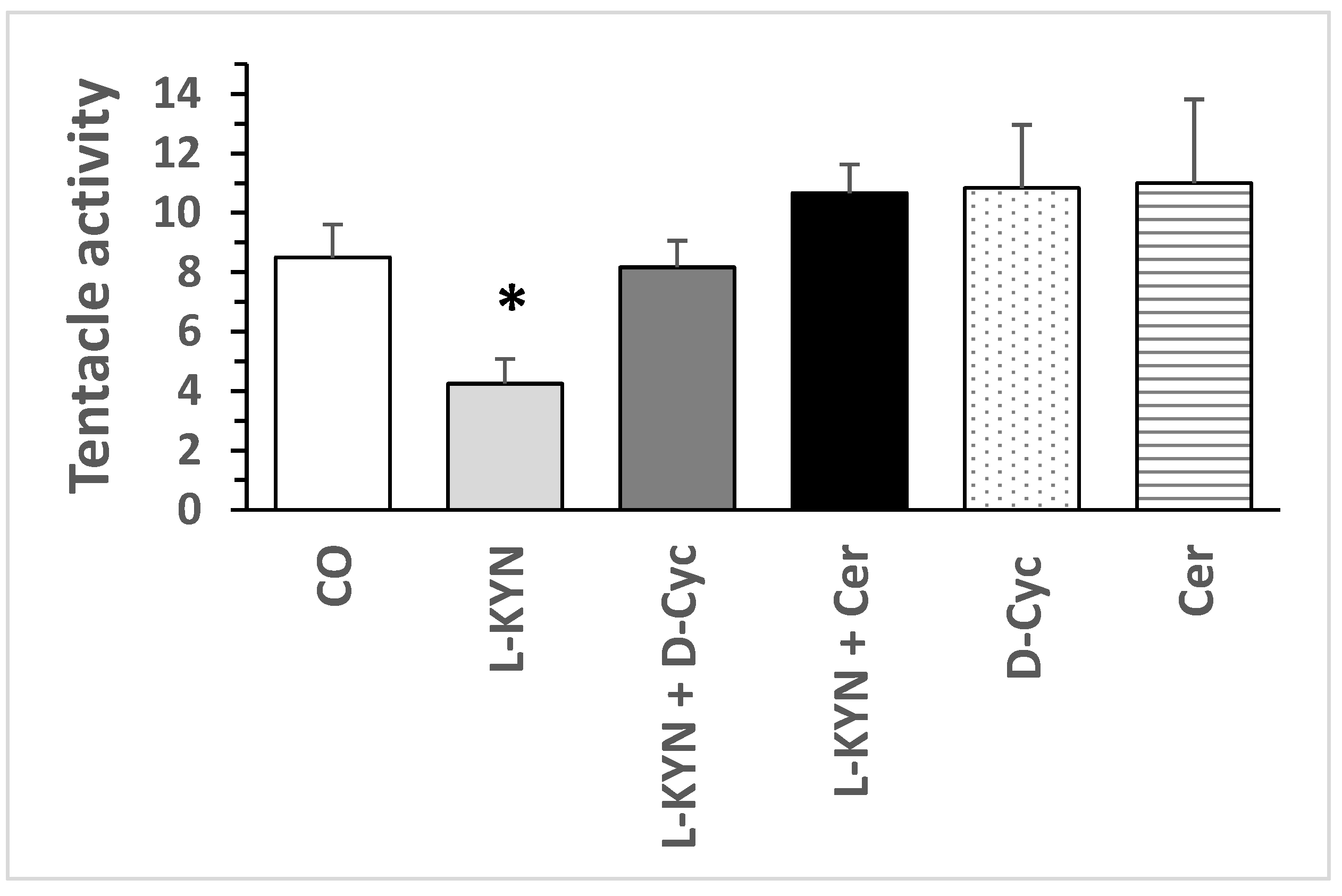
8.2. Effect of Pharmacological Treatment on Helix pomatia Snail Behaviour
9. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Baran, H.; Jellinger, K.; Deecke, L. Kynurenine metabolism in Alzheimer’s disease. J Neural Transm. 1999, 106, 165–181. [Google Scholar] [CrossRef]
- González-Sánchez, M.; Jiménez, J.; Narváez, A.; Antequera, D.; Llamas-Velasco, S.; Martín, A.H.-S.; Molina Arjona, J.A.; López de Munain, A.; Lleó Bisa, A.; Marco, M.-P.; Rodríguez-Núñez, M.; Pérez-Martínez, D.A.; Villarejo-Galende, A.; Bartolome, F.; Domínguez, E.; Carro, E. Kynurenic Acid Levels are Increased in the CSF of Alzheimer’s Disease Patients. Biomolecules 2020, 10, 571. [Google Scholar] [CrossRef]
- Knapskog, A.-B.; Aksnes, M.; Edwin, T.H.; Ueland, P.M.; Ulvik, A.; Fang, E.F.; Eldholm, R.S.; Halaas, N.B.; Saltvedt, I.; Giil, L.M.; Watne, L.O. Higher concentrations of kynurenic acid in CSF are associated with the slower clinical progression of Alzheimer’s disease. Alzheimer’s & Dementia 2023, 19(12), 5573–5582. [Google Scholar]
- Fernandes, B.S.; Inam, M.E.; Enduru, N.; Quevedo, J.; Zhao, Z. The kynurenine pathway in Alzheimer’s disease: a meta-analysis of central and peripheral levels. Braz. J. Psychiatry 2023, 45, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Choe, K.; Bakker, L.; van den Hove, D.L.A.; Eussen, S.J.P.M.; Kenis, G.; Ramakers, I.H.G.B.; Verhey, F.R.J.; Rutten, B.P.F.; Köhler, S. Kynurenine pathway dysregulation in cognitive impairment and dementia: a systematic review and meta-analysis. GeroScience. 2025. REVIEW. [Google Scholar] [CrossRef]
- Stone, T.W. Neuropharmacology of quinolinic and kynurenic acids. Pharmacol. Rev. 1993, 45, 309–379. [Google Scholar] [CrossRef]
- Birch, P.J.; Grossman, C.J.; Hayes, A.G. Kynurenic acid antagonizes responses to NMDA via an action at the strychnine-insensitive glycine receptor. Eur. J. Pharmacology. 1988, 154, 85–87. [Google Scholar] [CrossRef]
- Hilmas, C.; Pereira, E.F.R.; Alkondon, M.; Rassoulpour, A.; Schwarcz, R.; Albuquerque, E.X. The brain metabolite kynurenic acid inhibits alpha7 nicotinic receptor activity and increases non-alpha7 nicotinic receptor expression: physiopathological implications. J Neurosci. 2001, 21, 7463–7473. [Google Scholar] [CrossRef]
- Wang, J.; Simonavicius, N.; Wu, X.; Swaminath, G.; Reagan, J.; Tian, H.; Ling, L. Kynurenic acid as a ligand for orphan G protein-coupled receptor GPR35. Biol J Chem. 2006, 281(31), 22021–22028. [Google Scholar] [CrossRef]
- Stone, T.W.; Stoy, N.; Darlington, L.G. An Expanding range of targets for kynurenine metabolism of tryptophan. Trends in Paharmacological Science 2013, 34, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.C.; Vezzani, A.; French, E.D.; Schwarcz, R. Kynurenic acid blocks neurotoxicity and seizures induced in rats by the related brain metabolite quinolinic acid. Neurosci Lett. 1984, 48, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Bratek-Gerej, E.; Ziembowicz, A.; Godlewski, J.; Salinska, E. The Mechanism of the Neuroprotective Effect of KynurenicAcid in the Experimental Model of Neonatal Hypoxia–Ischemia: The Link to Oxidative Stress. Antioxidants 2021, 10, 1775. [Google Scholar] [CrossRef]
- Albuquerque, E.X.; Alkondon, M.; Pereira, E.F.R.; Castro, N.G.; Schrattenholz, A.; Barbosa, C.T.F.; Bonfante-Carbarcas, R.; Aracava, Y.; Eisenberg, H.M.; Maelike, A. Properties of neuronal nicotinic acetylcholine receptors: pharmacological characterization and modulation of synaptic function. J Pharmacol Exp Ther. 1997, 280, 1117–1136. [Google Scholar] [CrossRef]
- Albuquerque, E.X.; Pereira, E.F.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic receptors: from structure to function. Physiol Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef]
- Greenamyre, J.T.; Maragos, W.F.; Albin, R.L.; Penney, J.B.; Young, A.B. Glutamate transmission and toxicity in Alzheimer’s disease. Review. Prog Neuropsychopharmacol Biol Psychiatry 1988, 12, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Baran, H.; Jan Pietryja, M.; Kepplinger, B. Importance of Modulating Kynurenic Acid Metabolism—Approaches for the Treatment of Dementia. Biomolecules. 2025, 15, 74. [Google Scholar] [CrossRef] [PubMed]
- Kepplinger, B.; Baran, H.; Kainz, A.; Ferraz-Leite, H.; Newcombe, J.; Kalina, P. Age-related increase of kynurenic acid in human cerebrospinal fluid: Positive correlation with IgG and beta2-microglobulin changes. Neurosignals. 2005, 14, 126–135. [Google Scholar] [CrossRef]
- Chess, A.C.; Simoni, M.K.; Alling, T.E.; Bucci, D.J. Elevations of kynurenic acid produce working memory deficits. Schizophr Bull. 2007, 33, 797–804. [Google Scholar] [CrossRef]
- Vohra, M.; Lemieux, G.A.; Lin, L.; Ashrafi, K. The beneficial effects of dietary restriction on learning are distinct from its effects on longevity and mediated by depletion of a neuroinhibitory metabolite. PLoS Biol 2017, 15, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.C.; Elmer, G.I.; Bergeron, R.; Albuquerque, E.X.; Guidetti, P.; Wu, H.Q.; Schwarcz, R. Reduction of endogenous kynurenic acid formation enhances extracellular glutamate, hippocampal plasticity, and cognitive behavior. Neuropsychopharmacol. 2010, 35, 1734–1742. [Google Scholar] [CrossRef]
- Kozak, R.; Campbell, B.M.; Strick, C.A.; Horner, W.; Hoffmann, W.E.; Kiss, T.; Chapin, D.S.; McGinnis, D.; Abbott, A.L.; Roberts, B.M.; Fonseca, K.; Guanowsky, V.; Young, D.A.; Seymour, P.A.; Dounay, A.; Hajos, M.; Williams, G.V.; Castner, S.A. Reduction of Brain Kynurenic Acid Improves Cognitive Function. J. Neurosci. 2014, 34, 10592–10602. [Google Scholar] [CrossRef]
- Stone, T.W. Does Kynurenic Acid Act on Nicotinic Receptors? An Assessment of the Evidence. J. Neurochem. 2020, 152, 627–649. [Google Scholar] [CrossRef]
- Ishikawa, T.; Okuno, E.; Tsujimoto, M.; Nakamura, M.; Kido, R. Kynurenine-pyruvate Aminotransferase in Rat Kidney and Brain. Adv. Exp. Med. Biol. 1991, 294, 567–572. [Google Scholar] [PubMed]
- Kido, R. Kynurenate Forming Enzymes in Liver, Kidney and Brain. Kynurenine and Serotonin Pathways. Adv. Exp. Med. Biol. 1991, 294, 201–205. [Google Scholar] [PubMed]
- Okuno, E.; Nakamura, M.; Schwarcz, R. Two kynurenine aminotransferases in human brain. Brain Res. 1991, 542, 307–312. [Google Scholar] [CrossRef]
- Baran, H.; Kepplinger, B. Cerebrolysin Lowers Kynurenic Acid Formation - An In Vitro Study. Eur. Neuropsychopharmacol. 2009, 19, 161–168. [Google Scholar] [CrossRef]
- Baran, H.; Kepplinger, B. D-Cycloserine Lowers Kynurenic Acid Formation - New Mechanism of action. Eur. Neuropsychopharmacol. 2014, 24, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Baran, H.; Kepplinger, B. Porcine tissues influence kynurenic acid formation. Parkinsonism Relat. Disord. 2006, 12 Suppl.1, 36. [Google Scholar] [CrossRef]
- Baran, H.; Pietryja, M.J.; Kronsteiner, C.; Kepplinger, B. Jerusalem Balsam Lowers Kynurenic Acid Formation: An In Vitro Study. J. Tradit. Med. Clin. Naturopat. 2017, 3, 1–5. [Google Scholar] [CrossRef]
- Kepplinger, B.; Kronsteiner, C.; Więcek, M.; Baran, H. Hawthorn berry extract lowers kynurenic acid and anthranilic acid formation. 15th ANA Meeting at the IST Austria, 2017, September 24th-26th; Abstract. [Google Scholar]
- Radzki, R.P.; Bieńko, M.; Polak, P.; Szkucik, K.; Ziomek, M.M.; Ostapiuk, M.; Bieniaś, J. Is the consumption of snail meat actually healthy? An analysis of the osteotropic influence of snail meat as a sole source of protein in growing rats. J. Anim. Physiol. Anim. Nutr. 2018, 102, 885–891. [Google Scholar] [CrossRef]
- Acebes, F.; Solar, P.; Moris, J.; Loy, I. Associative learning phenomena in the snail (Helix aspersa): Conditioned inhibition. Learn. Behav. 2012, 40, 34–41. [Google Scholar] [CrossRef]
- Crow, T. Pavlovian Conditioning of Hermissenda: Current Cellular, Molecular, and Circuit. Review. Learn. Mem. 2004, 11, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Ungless, M.A. Dissociation of food-finding and tentacle-lowering, following food-attraction conditioning in the snail, Helix aspersa. In Behavioural processes; Elsevier, 2001. [Google Scholar]
- Kronsteiner, C.; Baran, H.; Kepplinger, B. Kynurenic acid levels and kynurenine aminotransferase I, II and III activities in ganglia, heart and liver of snail Helix Pomatia. Cell Physiol. Biochem. 2023, 18(57), 279–297. [Google Scholar] [CrossRef]
- Moroni, F.; Russi, P.; Lombardi, G.; Beni, M.; Carlà, V. Presence of Kynurenic Acid in the Mammalian Brain. J. Neurochem. 1988, 51, 177–180. [Google Scholar] [CrossRef]
- Fukui, S.; Schwarcz, R.; Rapoport, S.I.; Takada, Y.; Smith, Q.R. Blood-brain barrier transport of kynurenines: implications for brain synthesis and metabolism. J. Neurochem. 1991, 56, 2007–2017. [Google Scholar] [CrossRef]
- Turski, W.A.; Gramsbergen, J.B.P.; Traitler, H.; Schwarcz, R. Rat brain slices produce and liberate kynurenic acid upon expose to L-kynurenine. J Neurochem. 1989, 52, 1629–1636. [Google Scholar] [CrossRef]
- Swartz, K.J.; Matson, W.R.; MacGarvey, U.; Ryan, E.A.; Beal, M.F. Measurement of Kynurenic Acid in Mammalian Brain Extracts and Cerebrospinal Fluid by High-performance Liquid Chromatography with Fluorometric and Coulometric Electrode Array Detection. Anal. Biochem. 1990, 185, 363–376. [Google Scholar] [CrossRef]
- Ungless, M.A. A Pavlovian analysis of food-attraction conditioning in the snail Helix aspersa. Animal Learning & Behavior. 1998, 26, 15–19. [Google Scholar]
- Acebes, F.; Solar, P.; Carnero, S.; Loy, I. Blocking of conditioning of tentacle lowering in the snail (Helix asparsa). The Qarterly Journal of experimental Psychology 2009, 62, 1315–1327. [Google Scholar] [CrossRef] [PubMed]
- Goff, D.C. D-Cycloserine: An evolving role in learning and neuroplasicity in Schizophrenia. Schizophrenia Bulletin 2012, 38(5), 936–941. [Google Scholar] [CrossRef]
- Goff, D.C. D-cycloserine in Schizophrenia: New Strategies for Improving Clinical Outcomes by Enhancing Plasticity. Curr. Neuropharmacol. 2017, 15, 21–34. [Google Scholar] [CrossRef]
- Oxenkrug, G.; van der Hart, M.; Roeser, J.; Summergrad, P. Anthranilic acid: a potential biomarker and treatment target for schizophrenia. Ann. Psychiatry Ment. Health 2016, 4, 1059. [Google Scholar]
- Kepplinger, B; Sedlnitzky-Semler, B.; Eigner, S.; Kalina, P.; Berger, P.; Baran, H. Stroke Patients after repetitive Transcranial Magnetic Stimulation (rTMS) –Alterations of Tryptophan Metabolites in the Serum. Int. J. Neurorehabilitation 2014, 1, 128. [Google Scholar] [CrossRef]
- Kepplinger, B.; Baran, H.; Sedlnitzky-Semler, B.; Badawi, N.R.; Erhart, H. Stochastic resonance activity influences serum tryptophan metabolism in healthy human subjects. Int. J. Tryptophan Res. 2011, 4, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Baran, H.; Staniek, K.; Bertignol-Spörr, M.; Attam, M.; Kronsteiner, C.; Kepplinger, B. Effects of various kynurenine metabolites on respiratory parameters of rat brain, liver and heart mitochondria. Int. J. Tryptophan. Res. 2016, 9, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, G.J.; Wang, L.; Brew, B.J. Quinolinic acid selectively induces apoptosis of human astrocytes: potential role in AIDS dementia complex. J Neuroinflammation 2005, 26(2), 16. [Google Scholar] [CrossRef]
- Heyes, M.P.; Brew, B.J.; Saito, K.; Quearry, B.J.; Price, R.W.; Lee, K.; Bhalla, R.B.; Der, M.; Markey, S.P. Inter-relationships between quinolinic acid, neuroactive kynurenines, neopterin and beta2-microglobulin in cerebrospinal fluid and serum of HIV-1-infected patients. J. Neuroimmunol. 1992, 40, 71–80. [Google Scholar] [CrossRef]
- Baran, H.; Hainfellner, J.A.; Kepplinger, B. Kynurenic acid metabolism in various types of brain pathology in HIV-1 infected patients. Int J Tryptophan Res. 2012, 5, 49–64. [Google Scholar] [CrossRef]
- Kronsteiner, C. Tryptophanmetabolismus bei Steinadlern (Aquila chrysaetos)—Untersuchung des Alterungsprozesses. Ph.D./Mater’s Thesis, Julius-Maximilians-Universität Würzburg Julius Maximilian University of Würzburg, Würzburg, Germany, 2014. [Google Scholar]
- Oxenkrug, G. Anthranilic Acid–G-Protein coupled receptor109A–Cytosolic Phospholipase A2–Myelin–Cognition Cascade: A New Target for the Treatment/Prevention of Cognitive Impairment in Schizophrenia, Dementia, and Aging. Int. J. Mol. Sci. 2024, 25, 13269. [Google Scholar] [CrossRef]
- Berlinguer-Palmini, R.; Masi, A.; Narducci, R.; Cavone, L.; Maratea, D.; Cozzi, A.; Sili, M.; Moroni, F.; Mannaioni, G. GPR35 Activation Reduces Ca2+Transients and Contributes to the Kynurenic Acid-Dependent Reduction of Synaptic Activity at CA3-CA1 Synapses. PLoS ONE 2013, 8, e82180. [Google Scholar] [CrossRef]
- Deng, F.; Yang, D.; Qing, L.; Chen, Y.; Zou, J.; Jia, M.; Wang, Q.; Jiang, R.; Huang, L. Exploring the interaction between the gut microbiota and cyclic adenosine monophosphate-protein kinase A signaling pathway: a potential therapeutic approach for neurodegenerative diseases. Neural Regen Res. 2025, 20, 3095–3112. [Google Scholar] [CrossRef]
- Ted Abel, T.; Nguyen, P.V. Regulation of hippocampus-dependent memory by cyclic AMP-dependent protein kinase. Prog. Brain Res. 2008, 169, 97–115. [Google Scholar] [CrossRef]
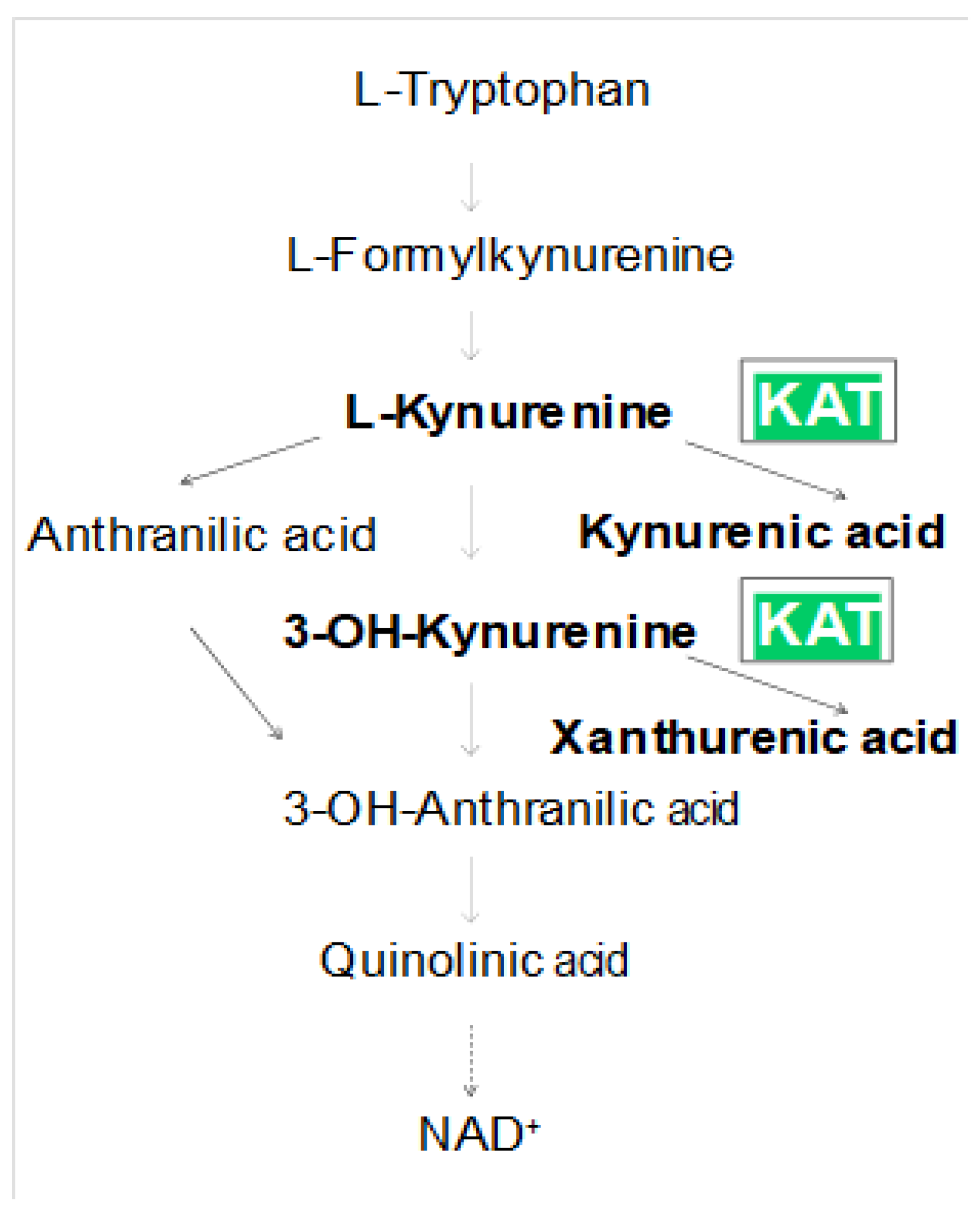
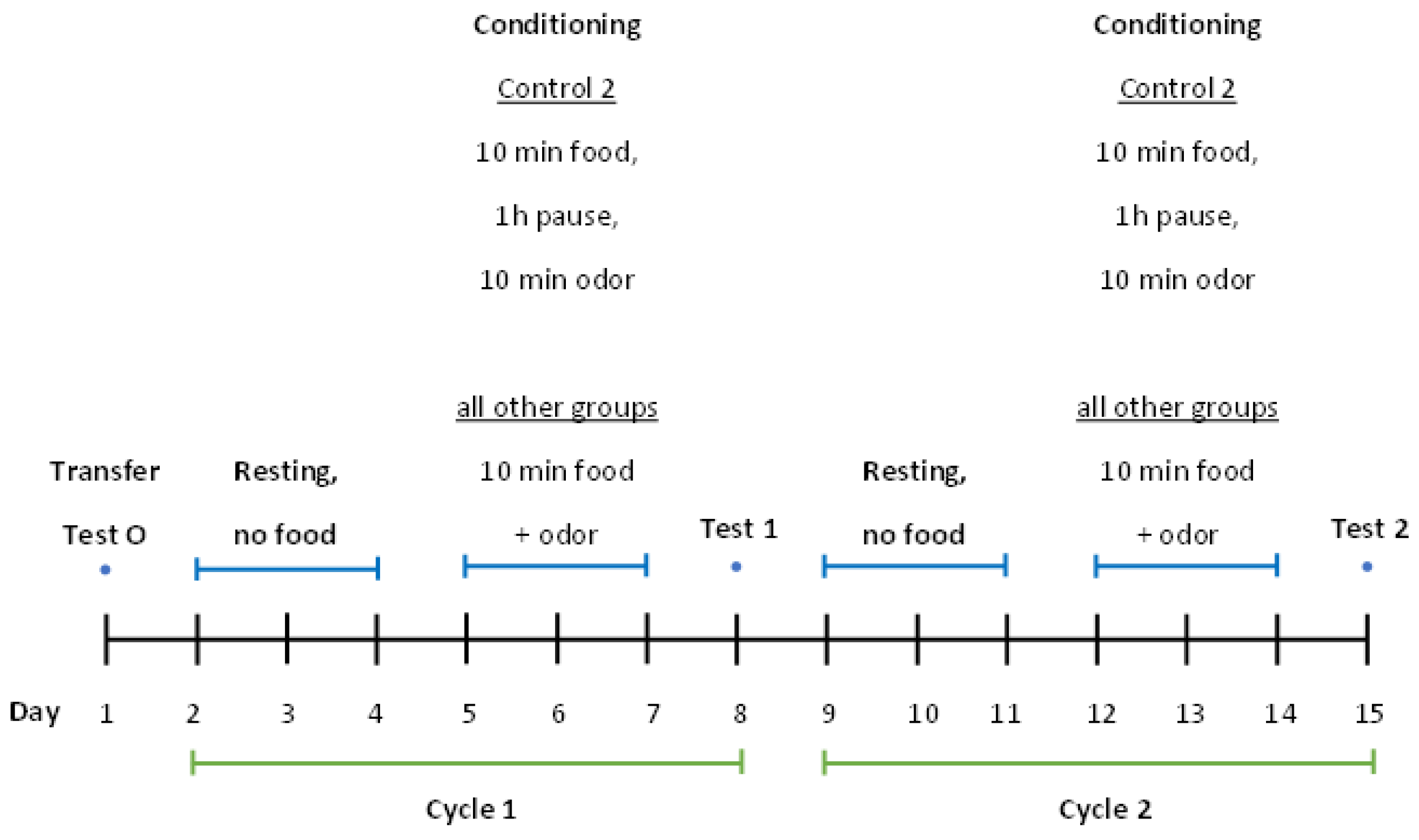

| Number of Groups/Name | Treatment |
| Group 1: CO1 | Water |
| Group 2: CO2 | Water |
| Group 3: L-KYN | L-kynurenine |
| Group 4: D-CYC | D-cycloserine |
| Group 5: CER | Cerebrolysin |
| Group 6: L-KYN/D-CYC | L-kynurenine + D-cycloserine |
| Group 7: L-KYN/CER | L-kynureniene + cerebrolysin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



