Submitted:
12 December 2025
Posted:
14 December 2025
You are already at the latest version
Abstract
Legumes are among the most important crop species in the world. They are the foundation of global food security and, in addition to providing protein, improve soil fer-tility and sustainable agriculture. However, they are sensitive to climate change and une-ven rainfall distribution. Three two-factor pot experiments were conducted in MI-CRO-CLIMA phytotrons. The objects of the study were three legume species: faba bean, pea and soybean. The first factor was the superabsorbent (SAP) rate (0, 2, 4, 6 g·kg-1 of sub-strate), while the second factor was the watering frequency (the subjects were watered every 1, 3, 6, 9 days). The scope of the research included measurements of chlorophyll flu-orescence (Fv/Fm, PI), leaf greenness index (SPAD) and biometric measurements. The study showed a significant effect of superabsorbent application on biometric parameters of legumes. Faba bean and pea plants were significantly taller after superabsorbent appli-cation and developed greater root mass. The highest SAP dose reduced the Fv/Fm index in soybean and the PI index in faba bean and soybean. The lowest SPAD index in pea was recorded in the control treatment (without SAP), while the highest SAP dose in soybean resulted in a decrease in relative leaf chlorophyll content compared to the other treat-ments. Plants watered daily were significantly taller, developed a greater number of nodes, and had higher dry mass of both above-ground and underground parts compared to plants watered every 3, 6, and 9 days (only in soybeans the dry mass of the under-ground part was significantly higher in the treatment watered least frequently). Higher values of the Fv/Fm index were observed in soybean, and higher values of the PI index were observed in faba bean, pea, and soybean in the least frequently watered treatments. In all species studied, SPAD index was higher under conditions of the greatest water defi-cit (watering every 9 days) compared to plants watered every 1, 3, and 6 days.
Keywords:
SAP
; superabsorbent polymer
; pea
; faba bean
; soybean
; chlorophyll fluorescence
; leaf greenness index
; watering
; drought
1. Introduction
One of the most important global challenges of the modern world is food security, and the greatest threat to it is the observed increase in air temperature on the Earth surface and its consequences in the form of climate change [1,2]. According to long-term forecasts, by the end of the 21st century, the average air temperature will increase by about 3°C [3]. One of the sectors of the economy that is particularly sensitive to climate change is agriculture [4]. Extreme weather events, including increasingly frequent droughts and heavy rainfall, have a significant impact on crop yields and food production stability [5,6]. Agricultural productivity is also affected by changing soil conditions, disease and pest pressure, and changing crop and land management practices [7,8,9]. These changes pose a serious threat to food security and require adaptation measures and the search for new solutions, such as the selection of resistant species and cultivars, the use of water-saving technologies and the introduction of regenerative agriculture practices [10,11]. An example of water-saving solutions is the use of soil additives that retain water, known as hydrogels or superabsorbents (SAP). Superabsorbents are synthetic or bio-derived polymers capable of storing large amounts of water in relation to their own weight. When applied to the soil, these polymers swell, increasing their volume, thereby improving soil structure, increasing its porosity and aggregation, reducing losses caused by water seepage, and thus increasing water capacity [12,13]. As a result, they help to mitigate the effects of water scarcity on plants. Thanks to the application of SAP, the root system is better developed and allows for the uptake of water and nutrients from deeper soil layers. In addition, water retention within the root ball allows plants to draw on water reserves during dry periods, which reduces stress, and proper soil moisture translates into better turgor and photosynthetic efficiency of plants, which in turn increases crop yields [14,15,16].
In an era of climate change and global population growth, ensuring sufficient food supplies while caring for natural resources is becoming an important element of the global economy. Legumes play a key role in ensuring food security, as they are a source of feed protein and also support a balanced human diet. In addition, these plants are an important source of fibre, vitamins and minerals [17]. Legumes are very important for sustainable agriculture due to their ability to biological nitrogen fixation and improve soil structure, thereby increasing soil fertility and improving water and air conditions [18].
However, the water requirements of legumes are high and varied. This group is very sensitive to uneven rainfall distribution and water shortages during critical periods, which occur during the germination, flowering, pod formation and seed filling. Drought during these periods can significantly reduce seed yield [19].
Water stress leads to disturbances in the structure and functioning of plant cells through numerous biochemical and physiological changes in plants [20,21]. The main factors determining the impact of soil water deficiency on plant physiology are the duration, frequency and intensity of drought. Moderate stress causes changes at the stomatal level, while prolonged and severe water shortages contribute to metabolic and structural changes [22,23]. In legumes, stress associated with soil water deficiency limits basic life processes (photosynthesis and transpiration), which adversely affects growth and development and, as a consequence, reduces seed yield and quality, often through changes in protein content and anti-nutritional substances [19,24]. The impact of drought stress on the photosynthetic apparatus of plants is well known [19,24,25]. Photosystem II (PSII) is more sensitive to water deficiency than photosystem I (PSI), and severe stress caused by limited soil water availability can contribute to its damage [23,27]. One method of studying the effect of stress factors on the functioning of the photosynthetic apparatus is to measure chlorophyll fluorescence. Analysis of individual parameters provides a wealth of information about the functioning of PSII in plants exposed to biotic and abiotic stress factors [23,28,29]. Chlorophyll pigments are an indicator of plant vitality and resistance to environmental stresses [30]. Their content in leaves depends on habitat and weather conditions, but above all it is a genetic trait [31]. Under conditions of water deficit, cells in the leaves shrink and become denser, which leads to an increase in the concentration of small and large molecules in the cell, including chlorophyll [32]. However, other authors report a decrease in chlorophyll content under water shortage conditions, which may be caused by excessive production of reactive oxygen species that damage chloroplasts and accelerate chlorophyll breakdown under water deficit [19,20].
The aim of the study was to assess the effect of superabsorbent on the condition of selected legume species grown with different watering frequencies.
2. Materials and Methods
Three, two-factor pot experiments were conducted in MICRO-CLIMA phytotrons from SNIJDERS LABS. The objects of the study were three legume species: faba bean (Vicia faba L.), pea (Pisum sativum L.) and soybean (Glycine max (L.) Merrill). The first factor was the superabsorbent (SAP) rate - 0, 2, 4, 6 g·kg-1 of substrate (SAP0, SAP2, SAP4, SAP6), while the second factor was the watering frequency - the subjects were watered every 1, 3, 6, 9 days (W1, W3, W6, W9). The experiment was conducted using a completely randomized design, with 4 replicates. The substrate, dried for 72 hours at 50ºC, was a mixture of horticultural soil for vegetables and sand in a ratio of 5:2. Pots with a capacity of 0.5 l were filled with substrate (300 g) mixed with an appropriate rate of granulated hydrogel (Aqua Terra Hydrogel). The seeds of faba bean (Granit cultivar), pea (Hubal cultivar) and soybean (Aldana cultivar) were treated with an antifungal dressing (Funaben) before sowing and then sown in moist substrate (3 seeds per pot). The faba bean were sown at a depth of 4-5 cm, the pea at 3-4 cm and the soybean at 1.5 cm. The pots were placed in a phytotron chamber. The conditions in the phytotron are presented in Table 1.
For each hydrogel rate, the maximum water capacity of the soil was determined using Vanschafi cylinders. In the 3-leaf stage of pea (BBCH 13) and faba bean (BBCH 13) and the 3-leaf stage of soybean (BBCH 13), different watering frequencies were introduced (every 1, 3, 6 and 9 days). All pots were weighed and watered to optimal conditions (60% of field water capacity).
2.1. Measurement Methods and Laboratory Analyses
During the experiments, measurements of chlorophyll fluorescence and the leaf greenness index (SPAD) were performed. Direct chlorophyll a fluorescence measurements were performed using a non-invasive (in vivo) method with a PocketPEA fluorometer (Hansatech Instruments – WB). Two indices were assessed: Fv/Fm, determining the maximum quantum efficiency of photosystem II (PSII), and PI, the PSII performance index. Chlorophyll fluorescence indices were used to determine the efficiency of the photosynthetic apparatus and to assess the physiological condition of the plants. SPAD measurements were performed using a SPAD – HYDRO N-TesterTM chlorophyll meter. The device measures the differences between light absorption by the leaf at two wavelengths (650 and 950 nm), and the quotient of these differences is the leaf greenness index, or relative chlorophyll content. Measurements were taken for each treatment one day before the planned watering date. Four measurements were taken for treatments watered every 1, 3 and 6 days, and three measurements for treatments watered every 9 days. The results presented are the average of all measurements for a given watering frequency. Chlorophyll fluorescence measurements were performed in 9 replicates, and the SPAD index in 3 replicates (one replicate as the average of 30 measurements). In the BBCH 50 phase (first flower buds visible), the plants were cut and the following measurements were taken: height, number of nodes, dry weight of the aboveground part (shoot) and underground part (root).
The collected results were statistically analyzed for a completely randomized design using analysis of variance (ANOVA) in the Statgraphic Centurion XVI programme. To compare the differences between the means for the main factors and interactions, a multiple confidence interval test (Tukey's test) was used at a significance level of p≤0.05.
3. Results
3.1. Faba Bean
The study showed that the SAP rate significantly affected the height of the faba bean. The lowest plants were recorded in the control group, while significantly higher plants were recorded when superabsorbent was applied in rates of 2 and 4 g·kg-1 (by 16.8 and 13.3%, respectively) (Table 2). The rate of superabsorbent used did not significantly differentiate the number of nodes on the plants or the dry weight of the aboveground part of the faba bean, but it did affect the dry weight of the underground part. The use of hydrogel at a rate of 6 g·kg-1 significantly increased the dry weight of the root (by an average of 56.8%) compared to the control treatment.
Taking into account the frequency of watering, the tallest plants were recorded in the control group watered daily, while with decreasing frequency the plants were significantly shorter, by 8.9% when watered every 3 days, by 18.2% when watered every 6 days, and by 23.3% when watered every 9 days, compared to the control treatment. The number of nodes per plant was also significantly lower when watered every 9 days compared to the treatment watered every 3 days. The frequency of watering significantly affected the dry weight of the aboveground part. With daily watering, the dry weight of the shoot was significantly higher compared to the plants watered every 6 and 9 days (by 24.7% and 29.1%, respectively).
Considering the efficiency of the photosynthetic apparatus, the SAP rate in faba beans significantly affected only the PSII functioning index (Table 3). The PI index was significantly higher in the SAP4 plot compared to the plot where the highest (SAP6) rate was applied (by 8.5%).
The frequency of watering significantly affected all the parameters tested in faba beans. The maximum PSII efficiency index was highest on the plot watered daily, and its value decreased with decreasing watering frequency. The PI index was significantly higher on the plot watered least frequently compared to the other plots. The leaf greenness index increased with decreasing watering frequency. At treatments W6 and W9, the leaf greenness index was significantly higher compared to treatments watered daily and every 3 days.
Interactions between the factors of the experiment and their impact on chlorophyll fluorescence indices were demonstrated (Figure 1). The Fv/Fm index decreased with daily watering as the SAP rate increased. Daily watering significantly increased the maximum PSII efficiency index compared to the treatment watered every 9 days at all SAP rates used. In contrast, the PI index value for watering every 9 days was highest after the application of SAP (regardless of the rate).
3.2. Pea
In peas, the rate of superabsorbent significantly affected plant height, but did not differentiate the number of nodes per plant (Table 4). When hydrogel was applied at a rate of 2 g·kg-1, significantly taller plants were observed compared to the control (by 10.9%). The SAP rate significantly affected the dry weight of the above-ground and underground parts of peas. The dry weight of the shoot was significantly higher in the SAP2 compared to the control (by 33.9%). The application of the highest rate of SAP significantly increased the dry weight of the root compared to the control treatment and the SAP2 rate (by 85.9 and 41.9%, respectively). Considering the watering frequency, the tallest plants were recorded at the treatment watered daily, and significantly shorter ones at the treatments watered every 6 and 9 days (by 9.2 and 27.2%, respectively). The lowest number of nodes was found at the treatment watered every 9 days compared to the other treatments. Pea plants watered daily developed significantly higher dry weight of the aboveground part compared to plants watered every 6 and 9 days (by 32.1 and 84.7%, respectively). The dry weight of the underground parts was also highest in the test watered daily. Significant differences were noted in comparison with the test watered every 6 and 9 days (by 43.7 and 36.7%, respectively).
A significant interaction between the factors of the experiment and its effect on the dry weight of the underground part of peas was demonstrated. Plants watered daily with a rate of SAP6 developed a significantly higher weight of the underground part compared to plants watered every 6 days (by 182.8%).
The tested factors significantly affected the efficiency of the photosynthetic apparatus in peas (Table 5). A rate of 2 g of superabsorbent significantly increased the PI index compared to the control (by 12.9%). The SPAD index value was significantly higher after the application of SAP (regardless of the rate) compared to the control treatment. The frequency of watering significantly affected all examined parameters of photosynthetic efficiency in peas. With daily watering, the Fv/Fm index was significantly higher compared to the treatments watered least frequently (by 0.4%). Restricting watering of peas (regardless of frequency) significantly increased the PI index compared to the treatment watered daily. The highest PI values were recorded in plants watered every 6 days. The frequency of watering significantly differentiated the leaf greenness index, which increased with its reduction. Significant differences were found between the plants watered daily and other treatments.
A significant interaction between the factors of the experiment and its effect on the PI index and SPAD index in peas was demonstrated (Figure 2). With an increase in the SAP rate, the PI index decreased with daily watering. The highest PI value was recorded with watering every 6 days at the SAP2 rate. When using SAP (regardless of the rate), the PI and SPAD index values were lower in the treatments watered daily compared to the other treatments (watered less frequently).
3.3. Soybean
The superabsorbent rate did not significantly affect the tested parameters of the plant structure in soybeans (Table 6). The second test factor (watering frequency), on the other hand, significantly differentiated all the tested parameters. Plants watered daily were significantly taller than plants watered every 3, 6 and 9 days (by 26.5, 51.5 and 60.9%, respectively). The number of nodes per plant was also significantly higher with daily watering compared to the other treatments. A significantly higher dry shoot weight was recorded with daily watering compared to the treatment watered every 6 and 9 days (by 60.6 and 42.6%, respectively). In contrast, the dry root weight was significantly higher with watering every 9 days compared to the treatments watered every 3 and 6 days (by 52.5 and 69.1%, respectively).
The tested factors significantly affected the efficiency of the photosynthetic apparatus in soybeans (Table 7). The SAP6 treatment showed significantly lower values of Fv/Fm, PI and SPAD index compared to the other SAP rates. The values of all tested parameters decreased with increasing superabsorbent rate. Watering frequency also differentiated the tested parameters in soybeans. Plants watered every 9 days were characterized by significantly higher values of the PSII index, and SPAD index compared to plants watered more frequently. Plants watered least frequently had a higher Fv/Fm index by 0.7%, PI index by 31.8% and leaf greenness index by 8.7% compared to plants watered daily.
An interaction between the experimental factors and its effect on chlorophyll fluorescence parameters and leaf greenness index in soybeans was demonstrated (Figure 3). With increasing SAP rate, the values of chlorophyll fluorescence indices (Fv/Fm and PI) decreased when watering every 6 days. The PI index value was highest at all SAP rates when watering every 9 days. The leaf greenness index when watering every 6 days was lower at higher SAP rates (SAP4 and SAP6) compared to SAP0 and SAP2.
4. Discussion
The climate changes observed in recent years and the associated irregular rainfall, prolonged droughts and sudden violent storms causing rapid runoff of rainwater from fields indicate the need for rational water management in agriculture. The use of artificial irrigation systems is often uneconomical and/or impossible for technical and environmental reasons. Areas with light, permeable soils with low water retention are most sensitive to lack of rainfall. Legumes are among the species most sensitive to water stress, but the requirements of individual species vary. This group is particularly sensitive to uneven rainfall distribution and water shortages during critical periods, which occur during germination, flowering, pod formation, and seed development. Drought during these periods affects morphological traits and physiological processes and can significantly reduce yields [19,33,34]. The use of a soil additive in the form of a superabsorbent can be an effective way to retain water in the soil. However, for the hydrogel to work effectively, regular rainfall is required to allow water to be absorbed and then made available to plants.
The growth of cells and organs in plants requires large amounts of energy and water availability. In order for cells to increase in size, water from the intercellular space must enter them. During water shortage, the plant limits its growth by inhibiting cell division and cell growth [35]. Restricting the growth of the aboveground part of the plant is one of the defense mechanisms against water stress. The results of our research showed a significant impact of the superabsorbent on the biometric parameters of legumes. Faba bean and pea plants were significantly taller after the application of hydrogel. In addition, the application of SAP at a rate of 6 g·kg-1 increased the dry weight of the underground parts of faba beans and peas (by 56.8% and 85.9%, respectively) compared to the control. Regular water supply caused the superabsorbent to retain water in the soil and make it available to plants as needed, so that the plants did not experience drought stress. This enabled the proper development of aboveground and underground biomass. In addition, the superabsorbent improved soil structure and enabled better root system development. Ryan et al. [36] demonstrated a significant effect of hydrogel on soybean biometric parameters under field conditions. According to the authors, the most effective was the application of SAP at a rate of 7.5 kg·ha-1, which contributed to an increase in plant height by 35.6%, leaf area by 93.4%, number of pods per plant by 99.3% and dry plant mass by 39.8% compared to the control treatment. In field studies conducted by Shankarappa et al. [37], the use of superabsorbent in the cultivation of lentils at a rate of 5 kg·ha-1 resulted in a 22.0% increase in the number of pods per plant, and the plants were 16.6% taller compared to the control. Youssef et al. [38], based on studies conducted in Egypt, showed that the application of superabsorbent at a rate of 0.7% of soil weight significantly increased the parameters of vegetative growth of peas, i.e. plant height, number of leaves per plant, dry weight of above-ground and underground parts, and root length compared to the control where SAP was not applied. Norodinvand et al. [39] demonstrated a significant effect of hydrogel on dry matter accumulation in common peas. The application of SAP at a rate of 0.5 and 1.0% of soil mass significantly increased the dry matter of plants. In contrast, Akhter et al. [40] obtained different results, showing no effect of hydrogel at rates of 0.1, 0.2 and 0.3% of soil weight on the length, fresh weight and dry weight of chickpea shoots.
Our experiment demonstrated the effect of watering frequency on the biometric characteristics of legumes. Plants watered daily were significantly taller and developed a greater number of nodes compared to plants watered every 3, 6 and 9 days. They also had significantly higher dry weight of the above-ground and below-ground parts. Only in soybeans was the dry weight of the underground parts significantly higher in the treatment watered least frequently (every 9 days) compared to treatments watered more frequently. Differences in response to stress result from species differences and strategies for adapting to water deficit. Field beans and peas are among the species that are more sensitive to periodic water shortages. Under optimal conditions, they are characterized by rapid shoot and root growth, but under conditions of water shortage, they limit biomass development. Soybeans, on the other hand, originate from a warmer climate and have developed certain adaptations to dry conditions. Under conditions of soil water shortage, this species limits the development of its above-ground parts and at the same time intensively develops its root system in order to reach deeper for water [41]. This is confirmed by the research of Desclaux et al. [42], who showed that soybean plants subjected to water shortages were shorter because they developed fewer nodes per plant and shorter internodes. Sadeghipour and Abbasi [43] showed that water deficit in soybean cultivation stimulated defense mechanisms and caused flower and pod shedding, which reduced the number of pods per plant, the number of seeds per pod and seed weight. In turn, in the study by Ohashi et al. [44], drought stress limited the growth and development of pods and reduced the dry weight of the vegetative part of soybeans. Reductions in growth parameters in peas under water deficit conditions were also demonstrated by Bodah et al. [45] and Osman [46].
One way to determine the efficiency of the photosynthetic apparatus is to measure the chlorophyll fluorescence emitted from chloroplasts. This is a quick, non-invasive method of assessing the physiological condition of plants growing in different environmental conditions. Chlorophyll fluorescence indices are used as one of the most effective parameters for estimating photosynthetic activity [47]. A decrease in photosynthetic activity under water deficit conditions limits the growth and productivity of crops [48]. The Fv/Fm index determines the maximum quantum efficiency of photosystem II and can be used to determine the condition of plants under stress conditions [49]. The results of our research did not show a significant effect of superabsorbent on the Fv/Fm index in faba beans and peas. In soybeans, the application of the highest rate of superabsorbent significantly reduced the Fv/Fm index value compared to the other treatments. This could have been caused by an excessive rate of SAP, which led to poorer soil aeration and root hypoxia, which in turn limited nutrient uptake and PSII efficiency.
The effect of watering frequency on the chlorophyll fluorescence index was demonstrated. In faba beans and peas, the highest values of this index were recorded with daily watering compared to treatments watered less frequently. In contrast, the response was reversed in soybeans, with plants watered every 9 days showing higher PSII efficiency compared to those watered more frequently. The different response of soybeans to moisture conditions can be explained by the genetic adaptation of this species to periodic water shortages and high temperatures. In studies conducted by Abid et al. [48], drought stress did not significantly affect the Fv/Fm index in tolerant cultivars of faba beans, while in a cultivar sensitive to water shortage, a significant decrease in Fv/Fm was observed at 30% relative humidity, which may suggest disturbances in PSII functioning. In turn, Allahmoradi et al. [50] showed a decrease in the maximum efficiency of PSII (Fv/Fm) in beans under water deficit conditions during the vegetative phase, compared to plants growing under optimal conditions. Ali et al. [51] demonstrated an 85.7% reduction in the Fv/Fm index under water deficit conditions in two soybean cultivars compared to optimal conditions in a greenhouse experiment.
Another important chlorophyll fluorescence index that describes the amount of effective energy converted by photosystem II is the PSII vitality index (PI). This index expresses the plant's ability to defend itself against stress [23]. The results of our research showed that the use of hydrogel at the highest rate of 6 g ka-1 of substrate significantly reduced the PI index compared to other rates and the control treatment in soybeans and the SAP rate of 4 g·kg-1 of substrate in field beans. In peas, however, the addition of hydrogel increased the PI index compared to the control. Furthermore, the highest values of the PSII functioning index in all legume species studied were observed in the treatments watered least frequently (every 9 days), and the PI value decreased with increasing watering frequency. In studies conducted by Allahmoradi et al. [50], drought stress during the vegetative growth phase reduced the PI index in mungbean (Vigna radiata L.) by 76.4% compared to the control treatment and by 75.9% compared to the treatment on which stress was initiated during the generative growth phase.
Drought conditions alter the content of photosynthetic pigments. Damage to thylakoid membranes and inhibition of biosynthesis lead to chlorophyll degradation. This limits the plant's ability to absorb light, leading to reduced photosynthetic intensity [52,53]. Studies have shown a variation in the relative chlorophyll content in leaves depending on the rate of hydrogel applied. In peas, the lowest SPAD index value was recorded in the control treatment, where no hydrogel was used, while in soybeans, the use of the highest rate of SAP reduced the relative chlorophyll content in the leaves compared to the other treatments. The response of individual species differed due to species differences in water management strategies in peas and soybeans. In peas, higher SAP rates improved soil moisture conditions, which resulted in higher chlorophyll content in the leaves. In soybeans, on the other hand, the highest rate of SAP may have caused root hypoxia, which limited nutrient uptake and disrupted chlorophyll metabolism. Compared to field peas, soybeans require better aerated soil due to their taproot system. Ahmed et al. [54] showed that the leaf greenness index in green beans increased after the application of superabsorbent (from 0.1 to 0.9% of soil mass) compared to the control field (without SAP). A significant increase in the SPAD index was also noted in peas after the application of superabsorbent at a dose of 0.7% of soil mass in the plots, compared to the treatment without SAP [38].
The results of our research showed that in all plant species studied, the leaf greenness index was higher under conditions of maximum water shortage (watering every 9 days) compared to plants watered every 1, 3 and 6 days. Drought caused water loss and reduced cell turgor, resulting in thinner leaves and increased chlorophyll density per unit leaf area [32]. In studies by Rahbarian et al. [33], the total chlorophyll content in chickpea (Cicer arietinum L.) leaves decreased under conditions of limited soil moisture (25% ppw) by 12.4% in the seedling stage and by 57.5% at the beginning of flowering, while the opposite relationship was observed in the pod formation stage. Higher chlorophyll content in leaves was observed under water deficit conditions compared to control conditions (by 27.2%). A decrease in the SPAD index under water deficit was also observed in peas [38,55].
5. Conclusions
The study showed that the SAP dose and watering frequency significantly influenced the studied parameters of development and condition of the photosynthetic apparatus of legumes. The superabsorbent significantly affected the biometric parameters. Plants were taller and had greater aboveground and underground mass after superabsorbent application. In soybean, the highest SAP dose significantly reduced the chlorophyll fluorescence parameters, while in pea, SAP application increased the PI and SPAD indexes. Plants watered most frequently developed the highest vegetative mass. Root dry mass was highest in faba bean and pea in the daily-watered treatments, while in soybean, it was the least frequently watered treatment. Drought reduced the chlorophyll fluorescence values in the studied species. The response of the legumes to the experimental factors was varied. Soybean responded differently to both the superabsorbent dose and watering frequency, which indicates different climatic requirements related to the origin of this species.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Fv/Fm | maximum quantum efficiency of PSII |
| PI | PSII Performance Index |
| PH | Plant height |
| NN | number of nodes |
| DWA | dry weight of the aboveground |
| DWU | dry weight of the underground |
References
- Wheeler, T.; Von Braun, J. Climate change impacts on global food security. Sci. 2013, 341, 508–513. [Google Scholar] [CrossRef]
- Schmidhuber, J.; Tubiello, F.N. Global food security under climate change. Proc. Natl. Acad. Sci. 2007, 104(50), 19703–19708. [Google Scholar] [CrossRef]
- Nazarenko, L. S.; Tausnev, N.; Russell, G. L.; Rind, D.; Miller, R. L.; Schmidt, G. A.; Bauer, S. E.; Kelley, M.; Ruedy, R.; Ackerman, A. S.; Aleinov, I.; Bauer, M.; Bleck, R.; Canuto, V.; Cesana, G.; Cheng, Y.; Clune, T. L.; Cook, B. I.; Cruz, C. A.; Del Genio, A. D.; Elsaesser, G. S.; Faluvegi, G.; Kiang, N. Y.; Kim, D.; Lacis, A. A.; Leboissetier, A.; LeGrande, A. N.; Lo, K. K.; Marshall, J.; Matthews, E. E.; McDermid, S.; Mezuman, K.; Murray, L. T.; Oinas, V.; Orbe, C.; Pérez García-Pando, C.; Perlwitz, J. P.; Puma, M. J.; Romanou, A.; Shindell, D. T.; Sun, S.; Tsigaridis, K.; Tselioudis, G.; Weng, E.; Wu, J.; Yao, M. S. Future climate change under SSP emission scenarios with GISS-E2. 1. J. Adv. Model. Earth Syst. 2022, 14(7), e2021MS002871. [Google Scholar] [CrossRef]
- Anderson, R.; Bayer, P. E.; Edwards, D. Climate change and the need for agricultural adaptation. Opin. Plant Biol. 2020, 56, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Spinoni, J.; Barbosa, P.; De Jager, A.; McCormick, N.; Naumann, G.; Vogt, J. V.; Magni, D.; Masante, D.; Mazzeschi, M. A new global database of meteorological drought events from 1951 to 2016. J. Hydrol. Reg. Stud. 2019, 22, 100593. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Coumou, D.; Frieler, K. Increased record-breaking precipitation events under global warming. Clim. Change 2015, 132(4), 501–515. [Google Scholar] [CrossRef]
- Singh, B. K.; Delgado-Baquerizo, M.; Egidi, E.; Guirado, E.; Leach, J. E.; Liu, H.; Trivedi, P. Climate change impacts on plant pathogens, food security and paths forward. Nat. Rev. Microbiol. 2023, 21(10), 640–656. [Google Scholar] [CrossRef] [PubMed]
- Ray, D. K.; Gerber, J. S.; MacDonald, G. K.; West, P. C. Climate variation explains a third of global crop yield variability. Nat. Commun. 2015, 6(1), 5989. [Google Scholar] [CrossRef]
- Challinor, A. J.; Watson, J.; Lobell, D. B.; Howden, S. M.; Smith, D. R.; Chhetri, N. A meta-analysis of crop yield under climate change and adaptation. Nat. Clim. Change 2014, 4(4), 287–291. [Google Scholar] [CrossRef]
- Oiganji, E.; Igbadun, H.; Amaza, P. S.; Lenka, R. Z. Innovative technologies for improved water productivity and climate change mitigation, adaptation, and resilience: a review. J. Appl. Sci. Environ. Manage. 2025, 29(1), 123–136. [Google Scholar] [CrossRef]
- Garg, S.; Rumjit, N. P.; Arora, P. Addressing climate change impacts through sustainable agricultural solutions: a review. Environ. Dev. Sustain. 2024, 1–27. [Google Scholar] [CrossRef]
- Malik, S.; Chaudhary, K.; Malik, A.; Punia, H.; Sewhag, M.; Berkesia, N.; Nagora, M.; Kalia, S.; Malik, K.; Kumar, D.; Kumar, P.; Kamboj, E.; Ahlawat, V.; Kumar, A.; Boora, K. Superabsorbent polymers as a soil amendment for increasing agriculture production with reducing water losses under water stress condition. Polymers 2022, 15(1), 161. [Google Scholar] [CrossRef]
- Abdelghafar, R.; Abdelfattah, A.; Mostafa, H. Effect of super absorbent hydrogel on hydro-physical properties of soil under deficit irrigation. Sci. Rep. 2024, 14(1), 7655. [Google Scholar] [CrossRef] [PubMed]
- AbdAllah, A. M.; Mashaheet, A. M.; Burkey, K. O. Super absorbent polymers mitigate drought stress in corn (Zea mays L.) grown under rainfed conditions. Agric. Water Manag. 2021, 254, 106946. [Google Scholar] [CrossRef]
- Zheng, H.; Mei, P.; Wang, W.; Yin, Y.; Li, H.; Zheng, M.; Ou, X.; Cui, Z. Effects of super absorbent polymer on crop yield, water productivity and soil properties: A global meta-analysis. Agric. Water Manag. 2023, 282, 108290. [Google Scholar] [CrossRef]
- Sroka, K.; Sroka, P. Superabsorbent hydrogels in the agriculture and reclamation of degraded areas. Sustainability 2024, 16(7), 2945. [Google Scholar] [CrossRef]
- Acheampong, R.; Tutu, C. O.; Owusu-Bempah, J.; Kumador, D. K.; Mahama, S.; Kortei, N. K.; Wiafe-Kwagyan, M.; Akonor, P.T.; Ayim-Akonor, M. Underutilised legumes in regenerative agriculture: Implications for food and nutritional security–A review. Appl. Food Res. 2025, 5(2), 101313. [Google Scholar] [CrossRef]
- Cimarelli, S.; Goglio, P.; Serpa, D; Quagliolo, C.; Dorca-Preda, T.; Sadhu, A.; Rai, K.; Roebeling, P.; Nemecek, T.; Cipolla, A.M.; Achneider, A.; Smetana, S.; Vasconcelos, M.; Kartal, U.; Joensuu, K.; Petrusan, J-I.; Dauguet, S.; Falchetti-Cartier, A.; Wilkinson, T.; Iannetta, P. Integrating ecosystem services provided by legumes in agricultural life cycle assessment (LCA): A review of methodologies. Ecol. Indic. 2025, 181, 114462. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research progress and perspective on drought stress in legumes: A review. Int. J. Mol. Sci. 2019, 20(10), 2541. [Google Scholar] [CrossRef]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S. S.; Siddique, K. H. Drought stress in grain legumes during reproduction and grain filling. J. Agron. Crop Sci. 2017, 203(2), 81–102. [Google Scholar] [CrossRef]
- Fazeli, F.; Ghorbanli, M.; Niknam, V. Effect of drought on biomass, protein content, lipid peroxidation and antioxidant enzymes in two sesame cultivars. Biol. Plant. 2007, 51(1), 98–103. [Google Scholar] [CrossRef]
- Jedmowski, C.; Ashoub, A.; Brüggemann, W. Reactions of Egyptian landraces of Hordeum vulgare and Sorghum bicolor to drought stress, evaluated by the OJIP fluorescence transient analysis. Acta Physiol. Plant. 2013, 35(2), 345–354. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]
- Khatun, M.; Sarkar, S.; Era, F. M.; Islam, A. M.; Anwar, M. P.; Fahad, S.; Datta, R.; Islam, A. A. Drought stress in grain legumes: Effects, tolerance mechanisms and management. Agronomy 2021, 11(12), 2374. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103(4), 551–560. [Google Scholar] [CrossRef]
- Zargar, S.M.; Gupta, N.; Nazir, M.; Mahajan, R.; Malik, F.A.; Sofi, N.R.; Shikari, A.B.; Salgotra, R.K. Impact of drought on photosynthesis: molecular perspective. Plant Gene 2017, 11, 154–159. [Google Scholar] [CrossRef]
- Lauriano, J.A.; Ramalho, J.C.; Lidon, F.C.; do Céu Matos, M. Mechanisms of energy dissipation in peanut under water stress. Photosynthetica 2006, 44(3), 404-410 154-159. [Google Scholar] [CrossRef]
- Moustakas, M.; Calatayud, Á.; Guidi, L. Chlorophyll fluorescence imaging analysis in biotic and abiotic stress. Front. Plant Sci. 2021, 12, 658500. [Google Scholar] [CrossRef]
- Staniak, M.; Baca, E. The effect of drought stress on chlorophyll fluorescence indicators in alfalfa (Medicago × varia Martyn), red clover (Trifolium pratense L.) and white clover (Trifolium repens L.). Grassl, Sci. Pol. 2018, 21, 127–138. [Google Scholar]
- Staniak, M. Dry matter and protein yields and leaf greenness (SPAD) of alfalfa and festulolium depending on cultivation method and soil moisture level. J. Res. Appl. Agric. Eng. 2018, 63(3), 92–97. [Google Scholar]
- Todorova, M.; Stoilova, T.; Gerdzhikova, M.; Rangelova, H.; Zherkova, Z.; Grozeva, N.; Tzanova, M. Relationships of NDVI and chlorophyll content with yield components of different accessions of Phaseolus vulgaris L. and vigna (Vigna unguiculata L. Walp.). Sci. Pap. Ser. A Agron. 2025, 68(1). [Google Scholar]
- Jones, M.B.; Leafe, E.L.; Stilles, W. Water stress in field-grown perennial ryegrass. II. Its effects on leaf water status, stomatal resistance and leafy morphology. Ann. Appl. Biol. 1980, 96, 103–110. [Google Scholar] [CrossRef]
- Rahbarian, R.; Khavari-Nejad, R.; Ganjeali, A.; Bagheri, A.; Najafi, F. Drought stress effects on photosynthesis, chlorophyll fluorescence and water relations in tolerant and susceptible chickpea (Cicer arietinum L.) genotypes. Acta Biol. Cracov. Bot. 2011, 53, 47–56. [Google Scholar] [CrossRef]
- Magyar-Tábori, K.; Mendler-Drienyovszki, N.; Dobránszki, J. Models and tools for studying drought stress responses in peas. OMICS 2011, 15, 829–838. [Google Scholar] [CrossRef]
- Forouzesh, E.; Goel, A.; Mackenzie, S.A; Turner, J.A. In vivo extraction of Arabidopsis cell turgor pressure using nanoindentation in conjunction with finite element modeling. Plant J. 2013, 73, 509–520. [Google Scholar] [CrossRef]
- Ryan, Q.; Geetha, K.N.; Hashimi, R.; Atif, R.; Habimana, S. Growth and yield of soybean [Glycine max (L.) Merrill] as influenced by organic manures and superabsorbent polymers. J. Exp. Agric. Int. 2020, 42, 77–85. [Google Scholar] [CrossRef]
- Shankarappa, S.K.; Muniyandi, S.J.; Chandrasheka, A.B.; Singh, A.K.; Nagabhushanaradhya, P.; Shivashankar, B.; El-Ansary, D.O.; Wani, S.H.; Elansary, H.O. Standardizing the hydrogel application rates and foliar nutrition for enhancing yield of lentil (Lens culinaris). Processes 2020, 8, 420. [Google Scholar] [CrossRef]
- Youssef, S.; Riad, G.; Abu El-Azm, N.A.I.; Ahmed, E. Amending sandy soil with biochar or/and superabsorbent polymer mitigates the adverse effects of drought stress on green pea. Egypt. J. Hort. 2018, 45, 169–183. [Google Scholar] [CrossRef]
- Norodinvand, F.A.; Dehkordi, D.K.; Egdernezhad, A. Evaluation of the superabsorbent effect on the yield and some growth factors of Pisum sativum L. under drought stress conditions. Recent Pat. Food Nutr. Agric. 2019, 10(2), 106–114. [Google Scholar] [CrossRef]
- Akhter, J.; Mahmood, K.; Malik, K.A.; Mardan, A.; Ahmad, M.; Iqbal, M.M. Effects of hydrogel amendment on water storage of sandy loam and loam soils and seedling growth of barley, wheat and chickpea. Plant Soil Environ. 2004, 50(10), 463–469. [Google Scholar] [CrossRef]
- Kunert, K. J.; Vorster, B. J.; Fenta, B. A.; Kibido, T.; Dionisio, G.; Foyer, C. H. Drought stress responses in soybean roots and nodules. Front. Plant Sci. 2016, 7, 1015. [Google Scholar] [CrossRef]
- Desclaux, D.; Huynh, T. T.; Roumet, P. Identification of soybean plant characteristics that indicate the timing of drought stress. Crop Sci. 2000, 40, 716–722. [Google Scholar] [CrossRef]
- Sadeghipour, O.; Abbasi, S. Soybean response to drought and seed inoculation. World Appl. Sci. J. 2012, 17(1), 55–60. [Google Scholar]
- Ohashi, Y.; Nakayama, N.; Saneoka, H.; Mohapatra; Fujita, P.K. Differences in the responses of stem diameter and pod thickness to drought stress during the grain filling stage in soybean plants. Acta Physiol. Plant. 2009, 31, 271–277. [Google Scholar] [CrossRef]
- Bodah, E.T.; Braunwart, K.; Bodah, B.W.; Alcindo Neckel, A. A rapid morphological screening procedure for pea (Pisum sativum L.) under drought stress in greenhouse settings. Am. J. Exp. Agric. 2015, 8(1), 68–74. [Google Scholar] [CrossRef]
- Osman, H.S. Enhancing antioxidant-yield relationship of pea plant under drought at different growth stages by exogenously applied glycine betaine and proline. Ann. Agric. Sci. 2015, 60(2), 389–402. [Google Scholar] [CrossRef]
- Ptushenko, V.V.; Ptushenko, O.S.; Tikhonov, A.N. Chlorophyll fluorescence induction, chlorophyll content, and chromaticity characteristics of leaves as indicators of photosynthetic apparatus senescence in arboreous plants. Biochem. (Mosc.) 2014, 79, 260–272. [Google Scholar] [CrossRef] [PubMed]
- Abid, G.; M’hamdi, M.; Mingeot, D.; Aouida, M.; Aroua, I.; Muhovski, Y.; Sassi, K.; Souissi, F.; Mannai, K.; Jebara, M. Effect of drought stress on chlorophyll fluorescence, antioxidant enzyme activities and gene expression patterns in faba bean (Vicia faba L.). ARCH AGRON SOIL SCI 2016, 63(4), 536–552. [Google Scholar] [CrossRef]
- Ahmed, S.; Nawata, E.; Hosokawa, M.; Domae, Y.; Sakuratani, T. Alterations in photosynthesis and some antioxidant enzymatic activities of mungbean subjected to waterlogging. Plant Sci 2002, 163, 117–123. [Google Scholar] [CrossRef]
- Allahmoradi, P.; Ghobadi, M.; Taherabadi, S.; Taherabadi, S. Physiological aspects of mungbean (Vigna radiata L.) in response to drought stress. ICFEB 2011, 9, 272–275. [Google Scholar]
- Ali, F.; Umar, M.; Siddiqui, Z.S. Comparative physiological assessment of some edible oil-seed crops under drought stress environment using fluorescence and IR imaging techniques. Pak. J. Bot. 2021, 53(4), 1183–1192. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Qiao, M.; Hong, C.; Jiao, Y.; Hou, S.; Gao, H. Impacts of drought on photosynthesis in major food crops and the related mechanisms of plant responses to drought. Plants 2024, 13(13), 1808. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.M.; El-Tohamy, W.A.; El-Abagy, H.M.H.; Aggor, F.S.; Nada, S.S. Response of snap bean plants to super absorbent hydrogel treatments under drought stress conditions. Curr. Sci. Int. 2015, 4, 467–472. [Google Scholar]
- Shinde, B.P.; Thakur, J. Influence of arbuscular mycorrhizal fungi on chlorophyll, proteins, proline and total carbohydrates content of the pea plant under water stress condition. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4(1), 809–821. [Google Scholar]
Figure 1.
Interaction of the SAP dose and watering frequency and its influence on the value of chlorophyll fluorescence indices (Fv/Fm and PI) in faba bean (means followed by different letters are significantly different).
Figure 1.
Interaction of the SAP dose and watering frequency and its influence on the value of chlorophyll fluorescence indices (Fv/Fm and PI) in faba bean (means followed by different letters are significantly different).

Figure 2.
Interaction of the dose of SAP and watering frequency its influence on the value of photosystem II performance index (PI) and SPAD index in pea (means followed by different letters are significantly different).
Figure 2.
Interaction of the dose of SAP and watering frequency its influence on the value of photosystem II performance index (PI) and SPAD index in pea (means followed by different letters are significantly different).
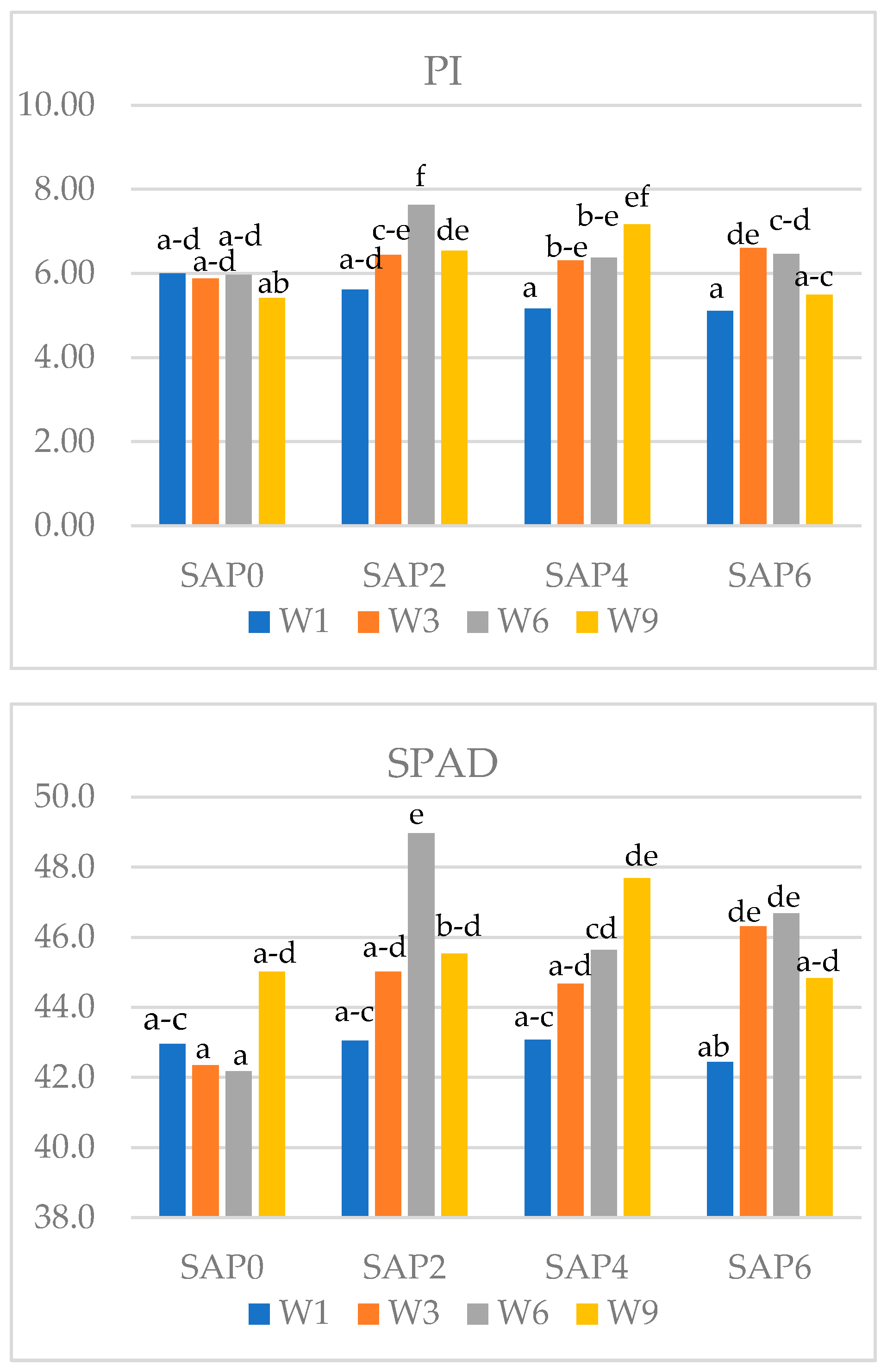
Figure 3.
Interaction of the dose of SAP and watering frequency its influence on the value of chlorophyll fluorescence indices (Fv/Fm and PI) and SPAD index in soybean (Means followed by different letters are significantly different).
Figure 3.
Interaction of the dose of SAP and watering frequency its influence on the value of chlorophyll fluorescence indices (Fv/Fm and PI) and SPAD index in soybean (Means followed by different letters are significantly different).
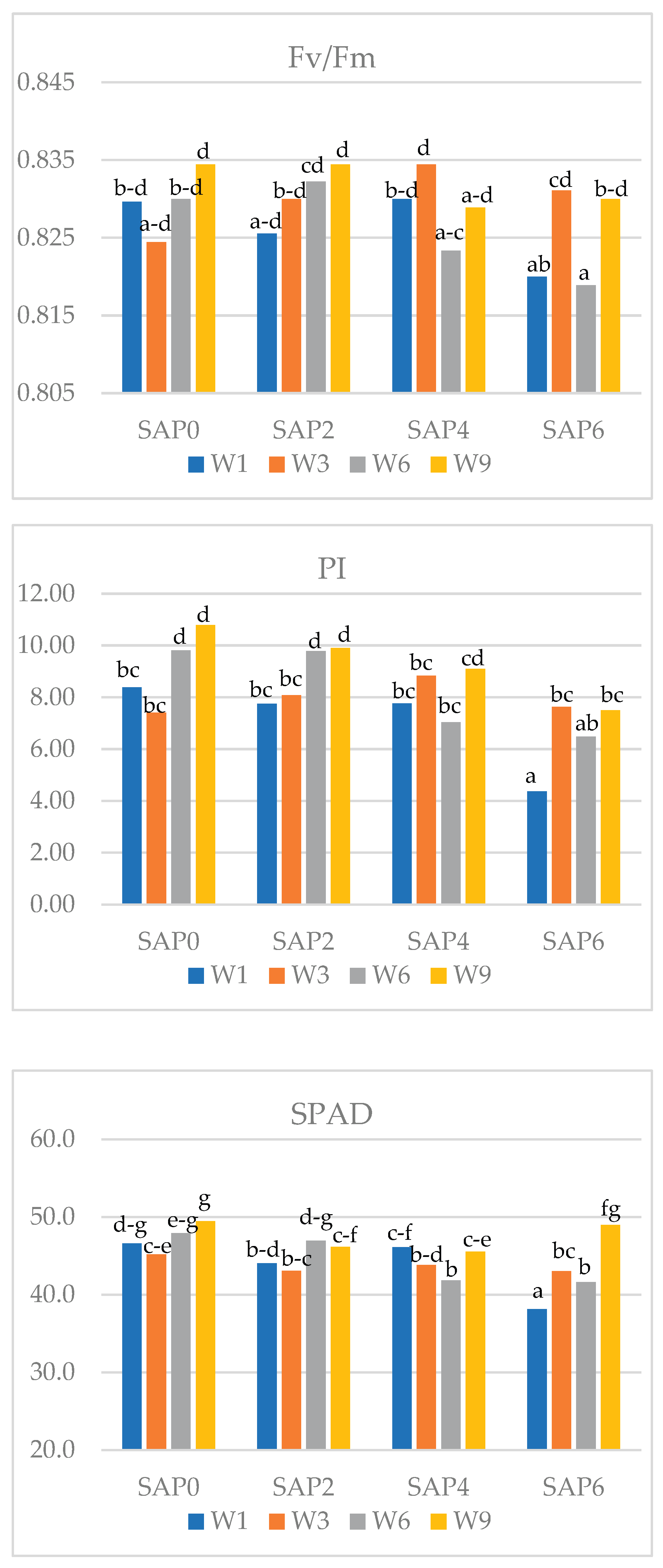

Table 1.
Conditions in the phytotron for individual species.
| Parameters | Faba bean | Pea | Soybean | |
| Temperature (°C) | day | 15 | 15 | 20 |
| night | 10 | 10 | 15 | |
| Day length (h) | day | 14 | 14 | 16 |
| night | 10 | 10 | 8 | |
| Air humidity (%) | 50 | 50 | 50 | |
Table 2.
Effects of the dose of SAP and watering frequency on the plant height (PH), number of nodes (NN), dry weight of the aboveground (DWA), dry weight of the underground (DWU) in faba bean.
Table 2.
Effects of the dose of SAP and watering frequency on the plant height (PH), number of nodes (NN), dry weight of the aboveground (DWA), dry weight of the underground (DWU) in faba bean.
| Factor | Source of variation | PH (cm) | NN | DWA | DWU | |
|---|---|---|---|---|---|---|
| Dose of SAP (S) | 0 | 51.8a | 11.9a | 1.73a | 0.491a | |
| 2 | 60.5b | 11.9a | 2.01a | 0.562ab | ||
| 4 | 58.7b | 11.8a | 1.94a | 0.633ab | ||
| 6 | 55.9ab | 12.0a | 1.92a | 0.773b | ||
| p-value | *** | ns | ns | ** | ||
| Watering frequency (W) | 1 | 64.9c | 11.8a | 2.27b | 0.718a | |
| 3 | 59.1b | 12.5b | 2.02ab | 0.511a | ||
| 6 | 53.1a | 12.0ab | 1.71a | 0.568a | ||
| 9 | 49.8a | 11.2a | 1.61a | 0.661a | ||
| p-value | *** | ** | ** | ns | ||
| S x W | p-value | ns | ns | ns | ns |
* Means followed by different letters are significantly different. The level of significance: ***p≤0.001. ** p≤0.01. * p≤0.05, ns – not significant at p≤0.05 (Tukey Test);
Table 3.
Effects of the SAP dose and watering frequency on the Fv/Fm (maximum quantum efficiency of photosystem II) and PI (photosystem II performance index) and SPAD (leaf greenness index) in faba bean.
Table 3.
Effects of the SAP dose and watering frequency on the Fv/Fm (maximum quantum efficiency of photosystem II) and PI (photosystem II performance index) and SPAD (leaf greenness index) in faba bean.
| Factor | Source of variation | Fv/Fm | PI | SPAD |
|---|---|---|---|---|
| Dose of SAP (S) | 0 | 0.827a | 5.47ab | 39.4a |
| 2 | 0.828a | 5.60ab | 38.5a | |
| 4 | 0.827a | 5.66b | 39.0a | |
| 6 | 0.826a | 5.18a | 39.7a | |
| p-value | ns | * | ns | |
| Watering frequency (W) | 1 | 0.833c | 5.43a | 36.6a |
| 3 | 0.827b | 5.10a | 37.6a | |
| 6 | 0.826b | 5.40a | 40.0b | |
| 9 | 0.822a | 5.97b | 42.4c | |
| p-value | *** | *** | *** | |
| S x W | p-value | *** | * | ns |
* Means followed by different letters are significantly different. The level of significance: ***p≤0.001. ** p≤0.01. * p≤0.05, ns – not significant at p≤0.05 (Tukey test);.
Table 4.
Effects of the SAP dose and watering frequency on the plant height (PH), number of nodes (NN), dry weight of the aboveground mass (DWA), dry weight of the underground mass (DWU) in pea.
Table 4.
Effects of the SAP dose and watering frequency on the plant height (PH), number of nodes (NN), dry weight of the aboveground mass (DWA), dry weight of the underground mass (DWU) in pea.
| Factor | Source of variation | PH (cm) | NN | DWA | DWU |
|---|---|---|---|---|---|
| Dose of SAP (S) | 0 | 54.1a | 15.9a | 1.24a | 0.319a |
| 2 | 60.0b | 17.3a | 1.66b | 0.418ab | |
| 4 | 55.5ab | 17.0a | 1.32ab | 0.465bc | |
| 6 | 55.7ab | 17.1a | 1.47ab | 0.593c | |
| p-value | * | ns | * | *** | |
| Watering frequency (W) | 1 | 62.9c | 18.2b | 1.81c | 0.562b |
| 3 | 59.5bc | 16.9b | 1.54bc | 0.432ab | |
| 6 | 57.1b | 18.0b | 1.37ab | 0.391a | |
| 9 | 45.8a | 14.2a | 0.98a | 0.411a | |
| p-value | *** | *** | *** | ** | |
| S x W | p-value | ns | ns | ns | ** |
* Means followed by different letters are significantly different. The level of significance: ***p≤0.001. ** p≤0.01. * p≤0.05, ns – not significant at p≤0.05 (Tukey test);.
Table 5.
Effects of the SAP dose and watering frequency on the Fv/Fm (maximum quantum efficiency of photosystem II) and PI (photosystem II performance index) and SPAD (leaf greenness index) in pea.
Table 5.
Effects of the SAP dose and watering frequency on the Fv/Fm (maximum quantum efficiency of photosystem II) and PI (photosystem II performance index) and SPAD (leaf greenness index) in pea.
| Factor | Source of variation | Fv/Fm | PI | SPAD |
|---|---|---|---|---|
| Dose of SAP (S) | 0 | 0.831a | 5.81a | 43.1a |
| 2 | 0.828a | 6.56c | 45.6b | |
| 4 | 0.829a | 6.25bc | 45.3b | |
| 6 | 0.831a | 5.92ab | 45.1b | |
| p-value | ns | *** | *** | |
| Watering frequency (W) | 1 | 0.831b | 5.47a | 42.9a |
| 3 | 0.831ab | 6.31bc | 44.6b | |
| 6 | 0.829ab | 6.61c | 45.9c | |
| 9 | 0.828a | 6.15b | 45.8c | |
| p-value | ** | *** | *** | |
| S x W | p-value | ns | *** | *** |
* Means followed by different letters are significantly different. The level of significance: ***p≤0.001, ** p≤0.01, * p≤0.05, ns – not significant at p≤0.05 (Tukey test);
Table 6.
Effects of the SAP dose and watering frequency on the plant height (PH), number of nodes (NN), dry weight of the aboveground (DWA), dry weight of the underground (DWU) in soybean.
Table 6.
Effects of the SAP dose and watering frequency on the plant height (PH), number of nodes (NN), dry weight of the aboveground (DWA), dry weight of the underground (DWU) in soybean.
| Factor | Source of variation | PH (cm) | NN | DWA | DWU |
|---|---|---|---|---|---|
| Dose of SAP (S) | 0 | 39.2a | 6.88a | 2.44a | 0.420a |
| 2 | 45.9a | 7.44a | 2.65a | 0.507a | |
| 4 | 45.6a | 7.06a | 2.70a | 0.561a | |
| 6 | 43.9a | 6.56a | 2.04a | 0.539a | |
| p-value | ns | ns | ns | ns | |
| Watering frequency (W) | 1 | 56.8c | 7.88b | 3.08b | 0.526ab |
| 3 | 44.9b | 6.50a | 2.41ab | 0.438a | |
| 6 | 37.5a | 6.88a | 2.19b | 0.395a | |
| 9 | 35.3a | 6.69a | 2.16b | 0.668b | |
| p-value | *** | *** | ** | ** | |
| S x W | p-value | ns | ns | ns | * |
* Means followed by different letters are significantly different. The level of significance: ***p≤0.001. ** p≤0.01. * p≤0.05, ns – not significant at p≤0.05 (Tukey test);.
Table 7.
Effects of the SAP dose and watering frequency on the Fv/Fm (maximum quantum efficiency of photosystem II) and PI (photosystem II performance index) and SPAD (leaf greenness index) in soybean.
Table 7.
Effects of the SAP dose and watering frequency on the Fv/Fm (maximum quantum efficiency of photosystem II) and PI (photosystem II performance index) and SPAD (leaf greenness index) in soybean.
| Factor | Source of variation | Fv/Fm | PI | SPAD |
|---|---|---|---|---|
| Dose of SAP (S) | 0 | 0.830b | 9.10c | 47.3c |
| 2 | 0.831b | 8.88bc | 45.1b | |
| 4 | 0.829b | 8.19b | 44.3b | |
| 6 | 0.825a | 6.50a | 42.9a | |
| p-value | ** | *** | *** | |
| Watering frequency (W) | 1 | 0.826a | 7.07a | 43.7a |
| 3 | 0.830ab | 7.99b | 43.8a | |
| 6 | 0.826a | 8.28b | 44.6a | |
| 9 | 0.832b | 9.32c | 47.5b | |
| p-value | *** | *** | *** | |
| S x W | p-value | *** | *** | *** |
* Means followed by different letters are significantly different. The level of significance: ***p≤0.001. ** p≤0.01. * p≤0.05, ns – not significant at p≤0.05 (Tukey test);
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



