
Submitted:
11 December 2025
Posted:
14 December 2025
You are already at the latest version
Abstract
Insectary plants are used to attract and boost the multiplication of beneficial arthropods, improving biological control in greenhouses. Three insectary plants were selected for this study: alyssum (Lobularia maritima (L.) Desv.), yarrow (Achillea millefolium L.), and dill (Anethum graveolens L.). This study was performed in two years, 2021 and 2025, and focused on Orius laevigatus (Fieber) (Hemiptera, Anthocoridae), one of the most im-portant predators of Thysanoptera pests in greenhouse crops. Four ornamental crops were included to analyse the movement and installation of the predator. Alyssum and yarrow housed O. laevigatus in both years (with total mean values per sampling date of 3.0±1.3 and 2.7±1.0 on alyssum and 7.0±2.8 and 1.8±0.8 on yarrow in 2021 and 2025 re-spectively), increasing its population in the greenhouse. In contrast, dill was unsuitable to settle populations of the predator but attracted other potential pests, with the addi-tional disadvantage of its short blooming period and quick decline. Orius laevigatus adults did not show great mobility during the study, and it had small populations among the ornamental crops in the greenhouse. The ornamental plant statice (Limonium sinuatum (L.) Mill.) had the highest population of the predator.
Keywords:
insectary plants
; greenhouse
; Lobularia maritima
; Achillea millefolium
; Anethum graveolens
; Orius laevigatus
; ornamental crops
1. Introduction
Pest control strategies increasingly rely on the use of biological control, particularly in greenhouses [1]. The classical strategy for implementing biological control in greenhouses is to introduce beneficial arthropods directly into the crop, with or without the presence of the pest, but other strategies can be used [2]. One of the most interesting is the use of plants that can harbour (or attract) beneficial arthropods, even before planting the crop [3]. This strategy is part of a conservation biological control approach, which aims to improve the presence and impact of natural enemies by manipulating plant-based resources in the landscape [4]. Meta-analysis has shown a general increase in natural enemies and a reduction of pest damage in crops due to diversification strategies [5]. This diversification strategy requires the selection of plants that provide resources, such as shelter, nectar, alternative prey/hosts, and pollen, and the establishment of these plants or plant communities in the managed system [6,7].
Amoabeng et al. [8] highlighted that most plants used in habitat management belong to three families, Apiaceae, Asteraceae, and Lamiaceae, but two other families, Brassicaceae and Fabaceae, also contain several species of interest in habitat manipulation studies. The screening of plants useful for this purpose has been reported in several reviews, which have generally identified the arthropod fauna associated with them [4,9,10], and a meta-analysis review has identified several flowering insectary plants (and plant families) that improve the biological fitness of important predators and parasitoids [11]. Plants native to a region may be equally or even more attractive to local natural enemies than commonly recommended exotic plants [4], and native plants attractive to important natural enemies have been determined in several crops in Spain [12,13,14,15].
The plants investigated in the present study are included in the category of insectary plants because they have a flower that provides nectar and pollen to the natural enemies we are interested in, but they are also considered “banker plants”, as they can have several types of arthropods that serve as prey (or host) to different predators (or parasitoids) [2,3,16]. In the management of banker plants, biological control agents are released onto them, and as they reproduce and increase in number, they spread into the rest of the greenhouse. This represents a mini-rearing system for natural enemies and can serve as a substitute for current augmentative releases of natural enemies in greenhouses [2,3] because the production and release of natural enemies can be continuous.
Alyssum (Lobularia maritima (L.) Desv.), belonging to the Brassicaceae family, has been the object of many studies, highlighting its attractiveness. Syrphidae (Diptera) is the most prominent family of this order associated with alyssum. Previous studies have classified it as one of the most attractive plants for these dipterans [2,9,17,18], ], whose adults feed on its nectar and pollen, and larvae can be voracious predators of aphids. Another family found on this plant is Anthocoridae (Hemiptera), especially those of the genus Orius, as they are found in greater numbers on alyssum than on other plants [2,9,10,19,20]. Alyssum represents a vital resource for a wide variety of Hymenoptera parasitoids, increasing the longevity and fertility of species in the Braconidae, Ichneumonidae, Eulophidae, and Trichogrammatidae families [17,21,22].
Yarrow (Achillea millefolium L.), which belongs to the Asteraceae family, has received less attention as an insectary plant than alyssum. However, previous studies have highlighted its effectiveness in attracting Hymenoptera parasitoids compared to other plants or spontaneous vegetation [23,24]. Other arthropod fauna of interest, including Syrphidae, have been studied on this plant, but the results have been inconsistent. For example, Colley and Luna [18] reported a preference for this plant species. However, other studies [23] have not shown that there is a particular interest for yarrow when other plants were present. A previous study has indicated that anthocorids are attracted to yarrow [23].
Dill (Anethum graveolens L.), which belongs to the Apiaceae family, has also attracted interest in insectary plant research. However, it is much less important than alyssum or yarrow. Sirphydae adults are strongly attracted to dill and feed on its nectar and pollen [25], and Chrysopidae (Neuroptera) species visit and utilise dill resources, enabling their survival and reproduction [6,26]. Even Coccinellidae (Coleoptera) species visit dill flowers [6], but the scarce information available indicates that Anthocoridae are not common visitors of dill [27].
One of the most important pest groups in greenhouses is the order Thysanoptera from Insecta, with species, such as Frankliniella occidentalis Pergande or Thrips tabaci Lindeman, having special relevance, particularly in greenhouses in southern Spain [28,29], although their presence and damage extends worldwide [30]. They cause damage to flowers, fruit, and leaves and can be vectors of different viruses [31]. They are very polyphagous and have been reported in many horticultural, fruit, and ornamental crops because adults prefer flowers, where they feed and lay their eggs [31].
Thysanoptera have several natural enemies that help control their populations, and they are widely used in augmentative biological control strategies. Some of these natural enemies are mainly predators, such as the acari of the Phytoseiidae family [32] and Hemiptera Anthocoridae, such as Orius laevigatus (Fieber) and other Orius species [1,33,34,35].
Studies on insectary plants have been conducted mainly in open field crops or in outdoor locations, and very few have been conducted in a greenhouse [36]. The present study used three insectary plants in a greenhouse environment over two years and tested the hypothesis that the three species were able to equally house and foster populations of O. laevigatus. Secondarily, it was tested the hypothesis that O. laevigatus moved to and colonised in the same degree different ornamental crops present in the greenhouse. As results of this study, Orius laevigatus settled and reproduced on alyssum and yarrow, whereas dill was unsuitable for it. Additionally, colonisation of the ornamental crops by O. laevigatus in the same greenhouse was very limited, although differences between them were found.
2. Materials and Methods
2.1. Location
The experiments were conducted in an experimental greenhouse (240 m2) in the facilities of Escuela Técnica Superior de Ingeniería Agronómica (ETSIA) at Universidad de Sevilla (Seville, Spain). The geographical coordinates of this location are 37°35’19.84”N and −5°93’81.65”S. The experiment was conducted in 2021 and 2025.
2.2. Experimental Design
In one-quarter of the greenhouse, three parallel growing beds were set up to cultivate ornamental plants. Each bed measured 10 m long by 1 m wide, with an elevation of 10 cm above the aisles, which were 1 m wide (Figure 1).
The insectary plants investigated in this study were alyssum (L. maritima), yarrow (Ac. millefolium), and dill (An. graveolens), which were obtained from seeds (Cantueso Natural Seeds, https://cantuesoseeds.com/). In both experiments, the seeds of the three species were planted in a seed bed in mid-September 2020 and 2024, and when they reached about 10 cm (approximately 1 month later), they were transplanted into 1-L pots containing a mix of horticultural substrate and sand (3:1) and placed in the experimental greenhouse.
In the 2021 experiment, each growing bed was divided into three zones and planted with three carnation (Dianthus caryophyllus L.) cultivars, which were assigned randomly. Another ornamental plant, gerbera daisy (Gerbera sp. L.), was planted in 15-L pots (with the same previously described substrate mixture and 4 g of Osmocote®) in one-quarter of the greenhouse (Figure 1a). Both ornamental plant species were transplanted into the greenhouse at the end of October 2020. Beds and pots were drip-irrigated. The insectary plants were transplanted from the 1-L pots to the ground at the beginning of November 2020 and placed in the beds, as shown in Figure 1a.
In the 2025 experiment, the same three growing beds were planted with three ornamental plants, namely carnation (D. caryophyllus L.), sweet William (Dianthus barbatus L.), and statice (Limonium sinuatum (L.) Mill.) (Figure 1b). In another one-quarter of the greenhouse, gerbera daisy (Gerbera sp.) was planted, with some in 15-L pots (with the same mixture as in 2021) and some planted in three rows in the ground (Figure 1b). Ornamental plant species were transplanted in the greenhouse (in beds, the ground, and pots) in mid-October 2024. Insectary plants were transplanted at the beginning of December 2024 from 1-L pots to five 15-L pots (the same mixtures as used for gerbera daisy) per insectary plant and distributed randomly around the area of the beds and near the gerbera daisies, as indicated in Figure 1b.
2.3. Sampling Procedure
The insectary plants were sampled on 9 dates in the 2021 experiment: February 18, March 2, 9, and 23, April 13, May 4, 11, and 21, and June 22. In the 2025 experiment, there were 11 sampling dates: January 31, February 13 and 27, March 13 and 27, April 9 and 24, May 6 and 22, and June 6 and 18. The phenology of the insectary plants was recorded on each sampling date and is presented in Appendix A (Table A1).
The sampling unit in 2021 was a bunch of flowers with a diameter of approximately 15–20 cm for alyssum, a similar bunch of flowers of 15–20 cm for yarrow, and 2 or 3 inflorescences for dill. Three sampling units from each insectary plant species were randomly selected from the three beds on each sampling date. The sampling unit was agitated/tapped vigorously three times on a white pan of 50 × 30 cm, and the arthropods removed from the plant were counted in situ. During this period, specimens of Thysanoptera were collected in vials with 70% ethanol to determine the species in the laboratory with the help of an optical microscope, and using specific keys for thrips identification [37,38].
The sampling unit in 2025 was similar for alyssum. There were no yarrow flowers during part of the sampling period, and a 15–20 cm bunch of leaves was collected as the sampling unit until flowers were available. The sampling unit for dill was one inflorescence. One sampling unit per insectary plant species was taken from three randomly selected pots where the insectary plants were on each sampling date. The process of extracting the arthropods was the same as that in 2021. As in 2021, Thysanoptera specimens were collected in vials with 70% ethanol to determine the species in the laboratory with the same procedure.
Ornamental plants were also sampled in 2021; between three and six flowers of each carnation cultivar were collected and opened in the laboratory under a stereo binocular (45x) to count thrips and other arthropods, especially O. laevigatus. Sampling of ornamentals in 2025 was carried out in the greenhouse using the same methodology as that used for the insectary plants. A sampling unit was selected (one open flower in carnation, one inflorescence in sweet William, a plant inflorescence in statice, and one flower in gerbera daisy) and tapped against a white pan, and the arthropods were counted in situ. Six sampling units were collected per ornamental species on each sampling date.
In 2025, the number of sampling units available in each pot of the insectary plants selected for sampling was also counted. The quantities of sampling units were also obtained for the ornamental plants by counting the flowers and inflorescences on each sampling date. The absolute arthropod population present in the insectary plants (mean value of arthropods per sampling unit in each sampling date by the average sampled units per pot by five pots) and ornamental plants (mean value of arthropods per sampling unit on each sampling date by the number of sampled units observed on each ornamental plant) was estimated using these figures. The number of sampling units for insectary and ornamental plants are given in Appendix A (Table A2).
Additionally, four yellow sticky traps were situated in the half of the greenhouse in which the study was carried out (Figure 1). They were changed after one week, and all specimens trapped in the laboratory were identified using a a stereo binocular (45 x) and a general key [39]. Fourteen samples were collected from these traps in 2021 (from October 2020 to May 2021) and nine samples in 2025 (from February to June).
2.4. Release of Orius laevigatus
Insects were provided by Biosur (Vícar (Almería), Spain, https://www.biosur.es) with its Orius Biosur product. There were 2 releases in 2021, on February 21 and April 30, with 1000 adults distributed homogenously over the insectary plants on each date. In 2025, there were also 2 releases, on February 26 and April 30, with 1500 adults distributed homogenously over the insectary plants on each date.
2.5. Statistical Analyses
Data obtained from the sampling dates were analysed separately by year with a repeated-measures analysis of variance (ANOVA) (SPSSv15.0 for Windows), with the factors date and plant species. Statistical analyses were performed in two steps: first, with the complete set of data for alyssum (L. maritima) and yarrow (Ac. millefolium) (9 sampling dates in 2021 and 11 sampling dates in 2025), and after using the three insectary plants (including dill (An. graveolens)) but only with the sampling dates in which all three plants were available (5 sampling dates in 2021 and 8 sampling dates in 2025). Data were not transformed.
3. Results
Alyssum (L. maritima) and yarrow (Ac. millefolium) showed the capacity to harbour and promote the presence of O. laevigatus over the two years of this study, whereas dill (An. graveolens) did not (Table 1). Orius laevigatus was found in consistent numbers in the flowers of alyssum (81 individuals in 2021 and 88 individuals in 2025) and yarrow (188 individuals in 2021 and 59 individuals in 2025). The most frequent arthropod group consistently found in the flowers of the three insectary plants was Thysanoptera, with a total population of 1792 individuals (1072 and 720 individuals in 2021 and 2025, respectively; Table 1). The species identified in the two seasons was almost exclusively Thrips tabaci, with 69 individuals in 2021 (only 1 individual was of other species) and 46 individuals in 2025 examined in the laboratory.
The second most abundant arthropod group in the insectary plants was Collembola, with 1442 individuals, which occurred frequently in alyssum (532 and 242 individuals in 2021 and 2025, respectively) and yarrow (376 and 248 in 2021 and 2025, respectively) but was almost absent in dill (Table 1). Other arthropods present in the insectary plants included Aphididae, Formicidae, Hymenoptera parasitoids, Araneae, and Acari. Aphididae was very abundant in dill in 2021 (2660 individuals were counted), but almost no presence was observed in the three insectary species in 2025 (Table 1).
Captures with the yellow traps indicated the high presence of certain groups (Table 2), particularly Thysanoptera in both years (2591 and 6607 individuals in 2021 and 2025, respectively), with other groups that varied between years. Aphididae was the most frequent in the traps in 2021 (with 4146 individuals) but not in 2025 (with 129 individuals), and Diptera was very abundant in 2025 (with 5531 individuals) but not in 2021 (with 837 individuals). There were few captures of O. laevigatus in both years, with 12 individuals in 2021 and 7 individuals in 2025.
Significant differences were detected in the adult population of O. laevigatus in 2021 between alyssum and yarrow (0.6 ± 0.2 and 3.6 ± 1.4 individuals per sampled unit and date (ISUD), respectively, F = 200.0, p < 0.001, Table 3), but no differences were found in the nymphs (2.4 ± 1.1 and 3.4 ± 1.6 ISUD, respectively, F = 4.1, p = 0.121, Table 3). In contrast, no significant differences were found in 2025 between the alyssum and yarrow populations of either adults or nymphs of O. laevigatus found in their flowers, with p > 0.05 in all cases (Table 3). The total O. laevigatus population was very similar in alyssum in both years (3.0 ± 1.3 and 2.7 ± 1.0 ISUD in 2021 and 2025, respectively), but there were differences between the years in yarrow (7.0 ± 2.8 and 1.8 ± 0.8 ISUD in 2021 and 2025, respectively), possibly due to the different phenology of the plant in each year, which could have been induced by the differences in the cultivation methods.
Thysanoptera was present in significant densities in the three insectary plants, ranging from 4.9 ± 1.2 to 20.6 ± 10.4 ISUD (Table 3). In general, the densities of the three insectary plants showed near-significant differences in 2021 (F = 5.1, p = 0.052, Table 3) and significant differences in 2025 (F = 7.6, p = 0.023, Table 3), when dill had the highest density. With the two plants that lasted longer in the experiments, alyssum and yarrow, the 2021 results showed low significance (F = 8.0, p = 0.048, Table 3), with similar results in 2025 (F = 6.7, p = 0.060, Table 3). In both years, yarrow showed higher densities but with high variability (20.6 ± 10.4 and 8.2 ± 4.0 ISUD in 2021 and 2025, respectively, Table 3).
Collembola was the second most abundant arthropod group, with remarkably similar densities in alyssum and yarrow in each year (19.7 ± 8.4 and 13.9 ± 5.4 ISUD, respectively, in 2021; 7.3 ± 1.8 and 7.5 ± 2.8 ISUD, respectively, in 2025, Table 3) and no statistical differences when they were compared (F = 4.0, p = 0.115 in 2021; F = 0.0, p = 0.910 in 2025, Table 3). In contrast, dill had very low densities in both years (1.6 ± 0.8 and 0.8 ± 0.8 ISUD in 2021 and 2025, respectively), and significant differences were observed when comparing the three insectary plants in both years (F = 31.3, p = 0.001 in 2021; F = 17.8, p = 0.003 in 2025, Table 3).
Aphididae was only present in 2021 with very high densities in dill (177.3 ± 124.9 ISUD) but only on a few dates. It was almost absent in the other two plants (alyssum and yarrow). Therefore, the population comparison with the three insectary plants was significant (F = 9.6, p = 0.014, Table 3), but the comparison between alyssum and yarrow was not (F = 1.1, p = 0.349, Table 3). In 2025, almost no Aphididae were found in the three insectary plants (Table 3).
In general, the mean densities in the other arthropod groups were very low, with significant differences between the insectary plants for Araneae and Acari in 2021 and Formicidae in 2025 (Table 3).
The total number of arthropods registered in each year (Table 3) showed similarity between alyssum and yarrow in 2021 (45.4 ± 8.0 and 44.4 ± 9.6 ISUD, respectively) and 2025 (19.7 ± 2.3 and 19.9 ± 5.7 ISUD, respectively); thus, there were no significant differences in the statistical comparison (p > 0.05 in both years, Table 3). Dill had a very high presence of Aphididae in 2021, which increased the mean number of arthropods (190.9 ± 122.4 ISUD), but no significant differences were observed between the three insectary plants (F = 5.2, p = 0.050, Table 3). In 2025, dill had a more similar population to the other two insectary plants (13.9 ± 4.7 ISUD), but a significant difference was found between them in this year (F = 5.7, p = 0.042, Table 3).
Seasonal changes in O. laevigatus density (Figure 2) indicate that its presence in the two insectary plants (alyssum and yarrow) that harboured it were generally low after the first release in the two years of the study (at the end of February). Just after release, a low number of adults were detected in both years, but very few nymphs were observed after a period of 2 months (Figure 2). After the second release (April 30 th in both years), the number of adults and, especially, nymphs increased in 2021, with significant values in the date × species effect for adults and nymphs (F = 6.0, p = 0.027 and F = 4.6, p = 0.028, respectively; Table 3). Similarly, this occurred in 2025, but the date × species effect was not significant (F = 1.0, p = 0.417 and F = 1.8, p = 0.217 for adults and nymphs, respectively; Table 3). The number of nymphs and adults remained at high levels for several weeks and declined quickly at the end of June in 2021 (plants started to senescent) and moderately in 2025. Yarrow and alyssum showed a similar number of nymphs (Figure 2b), with a peak of approximately 10 individuals per sampling unit (ISU) in May 2021 and a peak of 4–6 ISU in 2025 (Figure 2d). In 2021 (Figure 2a), yarrow had a significantly higher population of O. laevigatus adults than alyssum, with a peak of about 10 ISU at the beginning of May, whereas that for alyssum was near 2 ISU. This pattern was not observed in 2025, when adults were not as abundant (reaching 2–3 ISU), but similar results were obtained for both plant species (Figure 2c).
Thysanoptera was present in the three insectary plants in the two years, with peaks in April (2021, Figure 3a) and May (2025, Figure 3c), particularly in yarrow in 2021 when it reached a peak of 100 ISU. Although dill showed a quick decline in both years, it was able to reach similar densities as yarrow in 2025 (Figure 3c), and alyssum showed intermediate densities in general. The date × species effect was not significant in either year (Table 3).
Collembola was present in alyssum and yarrow in 2021 and 2025 and almost absent in dill (Figure 3b and Figure 3d). It was more abundant in 2021, with peaks of approximately 80 ISU in alyssum, but its density declined quickly in April and May. In contrast, in 2025, the density was not as high (peaks of 20–30 ISU) but declined equally towards the end of the sampling period. The date × species effect was not significant in either year (Table 3).
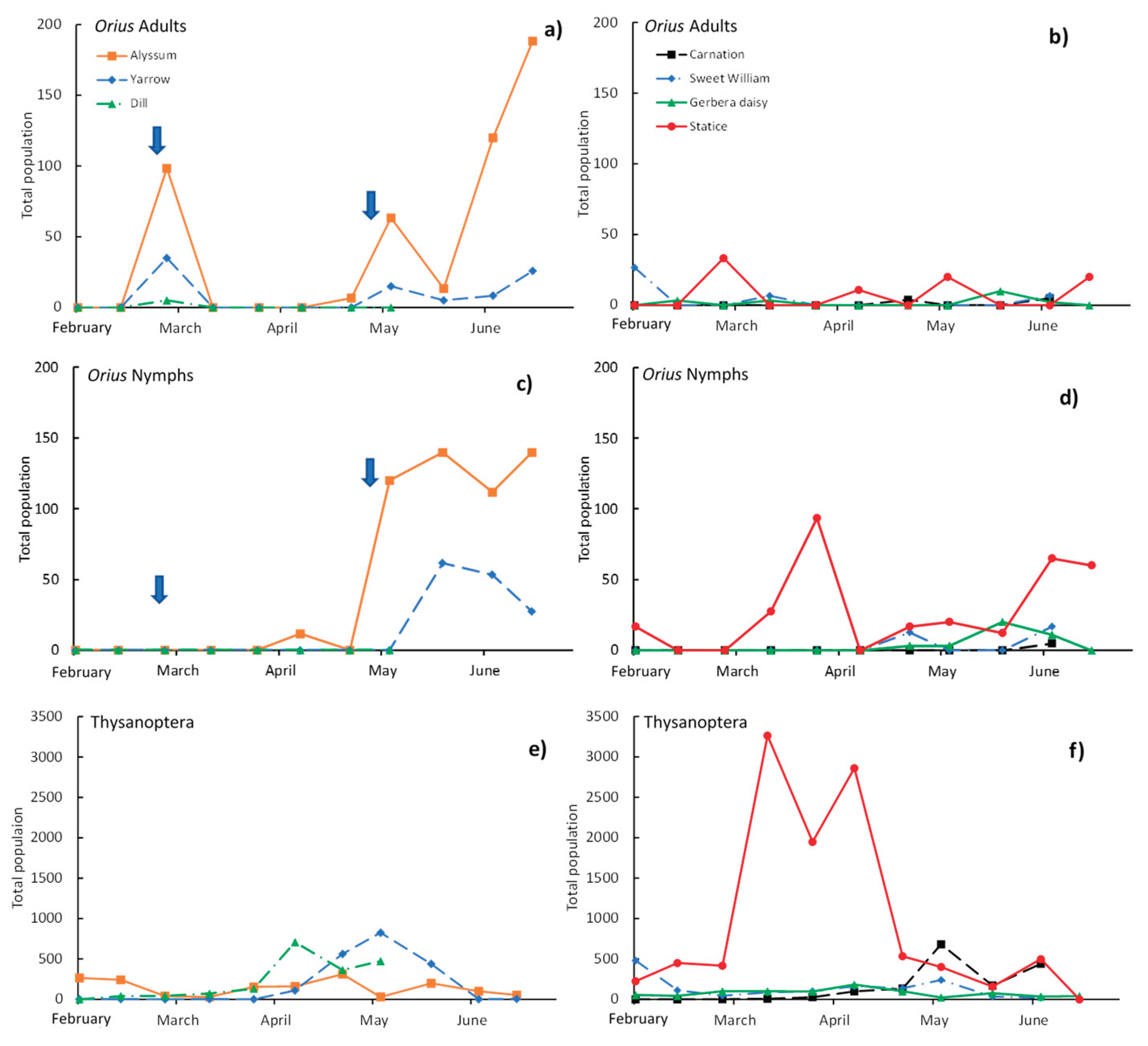
The absolute population of O. laevigatus estimated in 2025 (Figure 3) showed the potential of the insectary plants to house the predator and how it can be present in ornamental plants in the same greenhouse. The results from 2021 are not presented because carnation (with three cultivars) was the only ornamental plant used, and no O. laevigatus individuals were detected in the flowers during this study. Alyssum had the highest potential in 2025 because of the bloom, which occurred for a long period of time, even when the plants were in pots and not in the ground (Table A1 and Table A2). At the end of the sampling period, an estimated population of around 200 adults and 150 nymphs were present in alyssum (Figure 3a and Figure 3c), much more than that in yarrow, which with its scarce bloom could not house a high number of predators (Table A1 and Table A2). In general, the O. laevigatus population observed in the ornamental plants (Figure 3b and Figure 3d) was not paired with the population estimated in the insectary plants, especially when considering adults. Only statice presented a very low estimated adult population during this period, but a higher estimated nymph population (around 100 individuals) were identified after the first release of the predator (February 26 th), after the second release (April 30 th), and at the end of the sampling period (50–60 individuals), which could be related to the abundant and permanent blossom of the ornamental plant and constant presence of thrips (Figure 3d). Gerbera daisy also had a low O. laevigatus (nymph) population at the end of the sampling period, but it was negligible in carnation and sweet William.
Figure 3.
Seasonal changes in the mean population of Thysanoptera and Collembola per sampling unit in the insectary plants. a) and b) in 2021, c) and d) in 2025. Vertical bars are standard errors of the mean.
Figure 3.
Seasonal changes in the mean population of Thysanoptera and Collembola per sampling unit in the insectary plants. a) and b) in 2021, c) and d) in 2025. Vertical bars are standard errors of the mean.
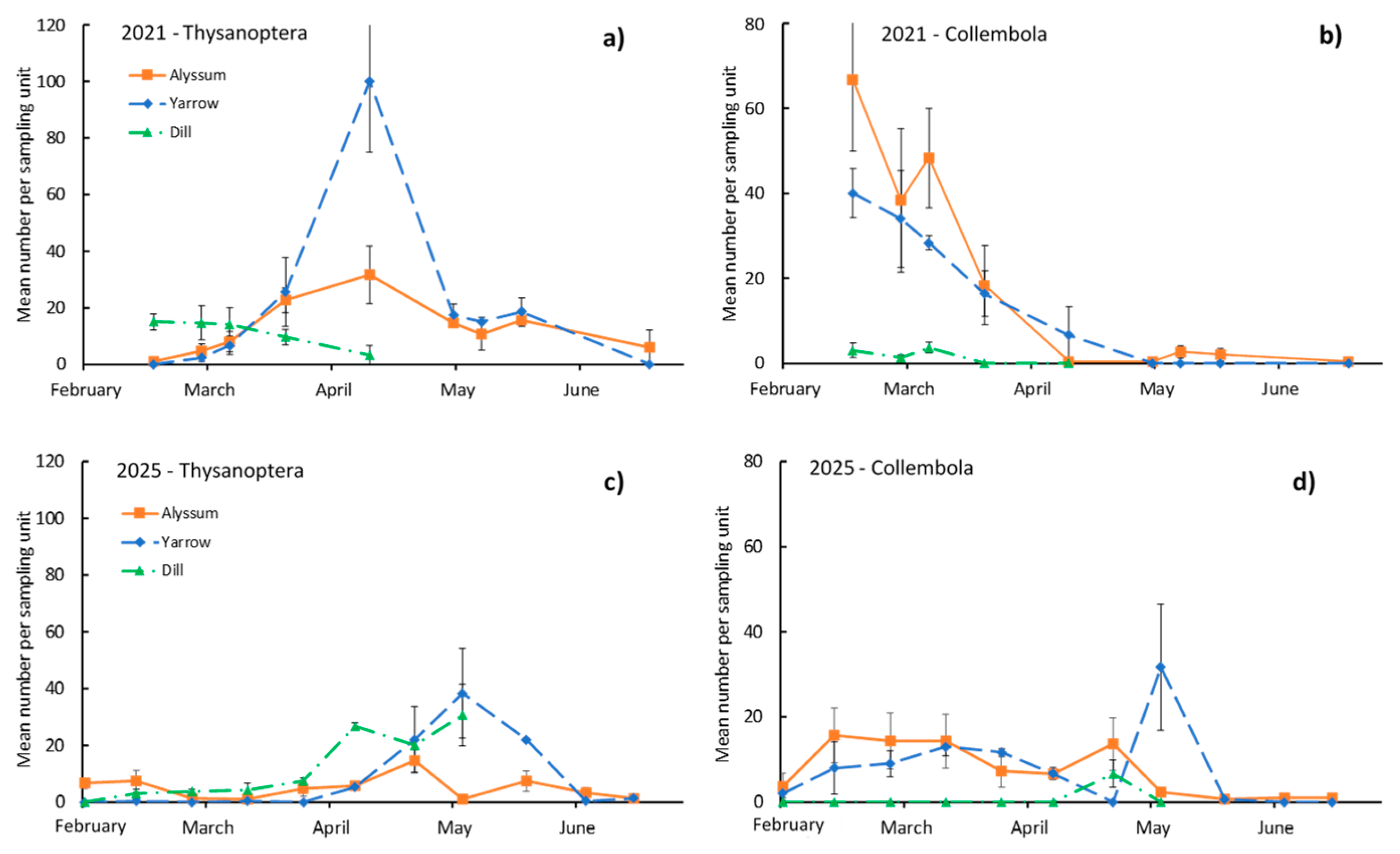
Figure 4.
Seasonal changes in the the estimated total population of Orius laevigatus (adults and nymphs) and Thysanoptera in the insectary plants and in the ornamental plants in 2025. a), c) and e) are the insectary plants; b), d) and f) are the ornamental plants. Arrows indicate the moment O. laevigatus adults were released in the insectary plants.
Figure 4.
Seasonal changes in the the estimated total population of Orius laevigatus (adults and nymphs) and Thysanoptera in the insectary plants and in the ornamental plants in 2025. a), c) and e) are the insectary plants; b), d) and f) are the ornamental plants. Arrows indicate the moment O. laevigatus adults were released in the insectary plants.
4. Discussion
Two of the insectary plants studied, namely alyssum (L. maritima) and yarrow (Ac. millefolium), showed a capacity to house and promote the populations of the predator O. laevigatus in a greenhouse environment in two seasons. In contrast, the other insectary plant, dill (An. graveolens), was unable to house this insect in either year, confirming the poor records of its presence on this plant [27]. The similarities in the results in two separate years highlights the robustness of the results regarding the use of these insectary plants in a greenhouse.
The two years of study showed some differences, as there was a greater number of O. laevigatus in 2021 than in 2025. Yarrow had higher O. laevigatus population than alyssum in 2021, whereas the opposite was observed in 2025. This difference between years can be attributed to the different ways in which the insectary plants were cultivated. In 2021, they were in the soil and grew very vigorously with a great production of blooms in both plants, whereas in 2025, they were in pots, growth was not as vigorous, and they produced less flowers, particularly in yarrow.
Alyssum is the most studied insectary plant among the three species used in this research. Hogg et al. [9] found that alyssum had the highest population of Anthocoridae (in which Orius spp. are included) compared to the other nine plants, and it was also the most abundant Hemiptera group on this plant, together with Lygaeidae. They found no Thysanoptera on alyssum in their study, but this must be due to the methodology used (vacuum sampling). Other studies have also highlighted the presence of anthocorids on this plant [2,4,10], and our results confirm that alyssum is a suitable insectary plant for O. laevigatus also in greenhouses.
The capacity of alyssum to house and promote different groups of natural enemies of pests (as indicated in the Introduction) has led to its implementation in different crops to improve the effectiveness of biological control in vineyards [22], strawberry [19], peach [23], apple [24], brassicas [40,41], and lettuce [10].
The other plant to house O. laevigatus in this study was yarrow, which housed this predator in similar, or even higher, densities than alyssum in the two years of the study, with the stable presence of adults and nymphs. Yarrow has been studied less than alyssum, but Aparicio et al. [23] also found that yarrow was able to house anthocorids with numbers very similar to alyssum. The results presented in our work confirm this aspect and make yarrow another plant with interest as insectary plant for O. laevigatus in greenhouses.
The presence of Thysanoptera was generally observed in the three insectary plants in our study. The main (and only) identified species was T. tabaci, an important pest in greenhouses and outdoor crops. Its presence can be a threat to crops because of its ability to colonise many plants. Although this can be a drawback to using insectary plants in greenhouses, the presence of thrips can be a great advantage for using predators on some of these plants, as occurred in with alyssum and yarrow, where O. laevigatus and other anthocorids can find shelter, prey (thrips), and alternative food (pollen) and have been utilised with success [42]. In contrast, dill housed thrips (a result highlighted also in [27]), but not anthocorids, which is another reason why this plant would not be considered suitable for use in greenhouses.
Aphididae were very scarce in alyssum and yarrow in both years, which is interesting because aphids (with different species) are important pests in greenhouse crops. Dill showed a different pattern, at least in 2021, when it had a heavy aphid infestation (observed also in the results of [27]), although the species was not identified. Another abundant group of arthropods in the insectary plants was Collembola, whose populations in alyssum and yarrow were similar in both plants in both years, but significantly lower in dill in both years. Other arthropod groups were less abundant, although their presence could be of interest, as with Araneae.
The duration of the flowering period is important for ensuring that natural enemies have long-term access to resources. Of the three insectary species in our study, alyssum showed the longest flowering period, from the start (and normally before) of the sampling period in February until the last sampling date, in the middle of June (but could be longer), with abundant and continuous flower production. This long flowering period has also been highlighted in another study [43], as it is one of the reasons for including this plant in this type of study and explains its attraction of different insects (syrphids, hymenopteran parasitoids, and others) [9,10,20].
Yarrow presented a more reduced flowering period in our study, although it was dependent on the year of study because it produced abundant flowers in 2021. In general, its flowering period is shorter than that of alyssum, starting at the middle-end of March (or later), which synchronised with the second release of the predator (end of April), and producing abundant flowers until the end of June (and even later). Some studies have included yarrow for its late-season flowering outdoors [18]. Dill flowered early, but during a short period, and at the beginning of May the plants had no flowers and died soon after. Compared to the other two insectary plants, the short flowering period observed in our study, which other authors have confirmed as being as short as seven days [25], limits its usefulness as an insectary plant inside a greenhouse.
Orius laevigatus was released on two occasions in both years, with very similar dates in each year, but installation and reproduction of the predator was observed in alyssum and yarrow only after the second release. In different species of the genus Orius, reproductive diapause is induced by short photoperiods and low temperatures [44,45]. Orius laevigatus showed a reduced response to daylight but was more affected by temperature [46], which can affect its reproductive performance. Temperatures during the first release (end of February in both years) were lower than those during the second release (end of April in both years). This could have caused difficulties in reproduction, as was observed in both seasons, with almost no nymphs present in the insectary plants after the first release. This contrasts with the second release, when the presence of nymphs was clear and, in some cases, abundant on the sample day after the release (as in 2021).
Orius laevigatus was evident in two of the insectary plants in the last part of the sampling period in both years, with significant densities in the sampling units and abundant total populations, particularly in alyssum due to its abundant blooming. However, its presence in ornamental plants was not as evident, particularly in carnation, in which almost no O. laevigatus adults or nymphs were observed in the flowers during the two years of the study, which may be explained by low attraction to this crop, as observed by other authors [47]. ]. In contrast, the ornamental plant statice presented had a low density of O. laevigatus in its flowers in 2025 but, with its very abundant blooming, housed a relatively important population, which was probably attracted by the constant presence of thrips. Gerbera daisy also had low densities of O. laevigatus (mainly nymphs), but its population was low due to the low number of flowers present throughout the study. Sweet William also had very low densities of O. laevigatus throughout the study. Although it produced abundant flowers, it had a small population due to the final decline of the crop at the end of the study. The movement of predators from the insectary plants to the crop plants (the ornamental species in our case) was low, as observed by other authors [10].
Our results showed very low captures of O. laevigatus in the yellow traps, indicating the low mobility of adults, or that they are not very attracted to the colour yellow. Other groups of beneficial insects, such as hymenopteran parasitoids and lacewings (Neuroptera, Chrysopidae), are attracted by yellow sticky traps [48,49], but this could be a drawback for its performance in outdoor and indoor crops.
The ornamental crops more receptive to O. laevigatus (particularly statice) arrived at the end of their productive life at the end of June, and so there was little time to show the practical effect of the insectary plants. The implementation of insectary plants in greenhouse crops to improve the installation of O. laevigatus has been performed with success but in other types of crops, such as sweet pepper [36], which is started in August in greenhouses in the south of Spain. Although the low movement of predators from insectary plants to crops could be an obstacle to using such plants, their presence has shown a good effect in the increment of parasitism of different pests by hymenopterans [21,50,51,52] due different mechanisms [2], which could be enough of a reason to consider its utilisation in both outdoor and indoor crops.
5. Conclusions
Alyssum (L. maritima) and yarrow (Ac. millefolium) housed O. laevigatus adults and nymphs, increasing its population in a greenhouse. This result was confirmed in two years. Alyssum presented long flowering and vegetation periods, which are crucial for insectary plants. Yarrow showed a more variable phenology between the two years of the study, and the cultivation method should be considered. Dill (An. graveolens) was neither attractive to O. laevigatus nor useful as an insectary plant in the greenhouse, mainly because of its limited flowering and vegetative periods, which was consistent in both years. In addition, dill was attractive to thrips and a reservoir of a great number of Aphididae in one year of the study. Colonisation of the ornamental plants cultivated in the greenhouse by O. laevigatus from the insectary plants was not remarkable, being almost null in the case of carnation (D. carophyllus), very low in gerbera daisy (Gerbera sp.) and sweet William (D. barbatus), and of some importance in statice (L. sinuatum).
Author Contributions
Conceptualization, M.C. and J.E.G.Z.; methodology, M.A.M. and J.E.G.Z.; validation, J.E.G.Z.; formal analysis, E.L. M.F. and J.E.G.Z. ; investigation, E.L. M.F., M.A.M. and J.E.G.Z.; resources, J.E.G.Z.; data curation, E.L. M.F. and J.E.G.Z.; writing—original draft preparation, J.E.G.Z.; writing—review and editing, M.C., M.A.M. and J.E.G.Z.; visualization, E.L., M.F. and J.E.G.Z.; supervision, J.E.G.Z.; project administration, M.C. and J.E.G.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The full raw data from the research are available in the public repository of the corresponding author’s institution: (in process).
Acknowledgments
The authors would like to thank the technical staff at the ETSIA experimental farm for their help with the study.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A

Table A1.
Phenology state of the insectary plants in the two years of study during the sampling period.
Table A1.
Phenology state of the insectary plants in the two years of study during the sampling period.
| Alyssum | Yarrow | Dill | |
| 2021 | |||
| 18-Feb | Flowering | Vegetative | Vegetative - Initial flowering |
| 2-Mar | Flowering | Vegetative – First inflorescences stems | Flowering |
| 9-Mar | Flowering | Initial flowering | Flowering |
| 23-Mar | Flowering | Flowering | Flowering + Fructification |
| 13-Apr | Flowering | Flowering | Final flowering and fructification |
| 4-May | Flowering + Fructification | Flowering | Senescent |
| 11-May | Flowering + Fructification | Flowering | Senescent |
| 21-May | Flowering + Fructification | Flowering +Fructification | Senescent |
| 22-Jun | Flowering + Fructification | Few flowering + Fructification | Senescent |
| 2025 | |||
| 31-Jan | Flowering | Vegetative | Initial flowering |
| 13-Feb | Flowering | Vegetative | Flowering |
| 27-Feb | Flowering | Vegetative | Flowering |
| 13-Mar | Flowering | Vegetative | Flowering |
| 27-Mar | Flowering + Fructification | Vegetative – First inflorescences stems | Flowering |
| 9-Apr | Flowering + Fructification | Vegetative + Initial flowering | Flowering + Fructification |
| 24-Apr | Flowering + Fructification | Initial flowering | Flowering + Fructification |
| 6-May | Flowering + Fructification | Flowering | Few flowering + Fructification |
| 22-May | Flowering + Fructification | Flowering | Senescent |
| 6-Jun | Flowering + Fructification | Flowering | Senescent |
| 18-Jun | Few flowering + Fructification | Few flowering + Fructification | Senescent |
Table A2.
Number of sampling units on each sampling date in 2025, used to estimate the absolute populations in the insectary plants and ornamental crops.
Table A2.
Number of sampling units on each sampling date in 2025, used to estimate the absolute populations in the insectary plants and ornamental crops.
| 31/1/2025 | 13/2/2025 | 27/2/2025 | 13/3/2025 | 27/3/2025 | 9/4/2025 | 24/4/2025 | 6/5/2025 | 22/5/2025 | 6/6/2025 | 18/6/2025 | |
| Insectary plants1 | |||||||||||
| Alyssum | 7.7 | 6.3 | 6.0 | 5.3 | 6.3 | 5.3 | 4.3 | 4.7 | 6.7 | 6.3 | 9.7 |
| Yarrow | 5.0 | 3.0 | 3.0 | 3.3 | 5.3 | 4.3 | 5.3 | 4.3 | 4.0 | 1.3 | 2.2 |
| Dill | 13.0 | 1.7 | 2.3 | 3.7 | 3.7 | 5.3 | 3.7 | 2.7 | - | - | - |
| Ornamentals2 | |||||||||||
| Carnation | 0 | 0 | 4 | 3 | 10 | 19 | 23 | 67 | 33 | 15 | - |
| Sweet William | 80 | 45 | 55 | 40 | 120 | 75 | 75 | 90 | 15 | 10 | - |
| Gerbera daisy | 20 | 20 | 20 | 20 | 20 | 20 | 20 | 20 | 20 | 13 | 24 |
| Statice | 50 | 50 | 50 | 55 | 70 | 65 | 100 | 120 | 73 | 65 | 60 |
1 Figures are the average number of sample units per pot (three pots were sampled in each date). Average population in the sample unit is then multiplied by the factor in the table and by 5 (the number of pots with insectary plants) to estimate the absolute population. 2 Figures are the number of sample units in each date. Average population in the sample unit is then multiplied by the factor in the table to estimate the absolute population.
References
- van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef]
- Pijnakker, J.; Vangansbeke, D.; Duarte, M.; Moerkens, R.; Wäckers, F.L. Predators and Parasitoids-in-First: From Inundative Releases to Preventative Biological Control in Greenhouse Crops. Front. Sustain. Food Syst. 2020, 4, 595630. [Google Scholar] [CrossRef]
- Parolin, P.; Bresch, C.; Desneux, N.; Brun, R.; Bout, A.; Boll, R.; Poncet, C. Secondary plants used in biological control: A review. Int. J. Pest Manag. 2012, 58, 91–100. [Google Scholar] [CrossRef]
- Fiedler, A.K.; Landis, D.A.; Wratten, S.D. Maximizing ecosystem services from conservation biological control: The role of habitat management. Biol. Control 2008, 45, 254–271. [Google Scholar] [CrossRef]
- Letourneau, D.K.; Armbrecht, I.; Rivera, B.S.; Lerma, J.M.; Carmona, E.J.; Daza, M.C.; Escobar, S.; Galindo, V.; Gutiérrez, C.; López, S.D.; et al. Does plant diversity benefit agroecosystems? A synthetic review. Ecol. Appl. 2011, 21, 9–21. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. HABITAT MANAGEMENT TO CONSERVE NATURAL ENEMIES OF ARTHROPOD PESTS IN AGRICULTURE. Annu. Rev. Entomol 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M. Habitat Management to Suppress Pest Populations: Progress and Prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef]
- Amoabeng, B.W.; Johnson, A.C.; Gurr, G.M. Natural enemy enhancement and botanical insecticide source: a review of dual use companion plants. Appl. Entomol. Zool. 2019, 54, 1–19. [Google Scholar] [CrossRef]
- Hogg, B.N.; Bugg, R.L.; Daane, K.M. Attractiveness of common insectary and harvestable floral resources to beneficial insects. Biol. Control 2011, 56, 76–84. [Google Scholar] [CrossRef]
- Hogg, B.N.; Nelson, E.H.; Daane, K.M. A comparison of candidate banker plants for management of pests in lettuce. Environ. Entomol. 2023, 52, 379–390. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Kiær, L.P.; Jensen, P.M.; Sigsgaard, L. The effect of floral resources on predator longevity and fecundity: A systematic review and meta-analysis. Biol. Control 2021, 153, 104476. [Google Scholar] [CrossRef]
- Alcalá Herrera, R.; Ruano, F.; Gálvez Ramírez, C.; Frischie, S.; Campos, M.; Alcala Herrera, R.; Ruano, F.; Gálvez, C.; Frischie, S. Attraction of green lacewings ( Neuroptera : Chrysopidae ) to native plants used as ground cover in woody Mediterranean agroecosystems. Biol. Control 2019, 139, 104066. [Google Scholar] [CrossRef]
- Alcalá Herrera, R.; Castro-Rodríguez, J.; Fernández-Sierra, M.L.; Campos, M.; Alcala Herrera, R.; Castro-Rodríguez, J.; Fernández-Sierra, M.L.; Campos, M. Dittrichia viscosa (Asterales: Asteraceae) as an Arthropod Reservoir in Olive Groves. Front. Sustain. Food Syst. 2019, 3, 1–8. [Google Scholar] [CrossRef]
- Cano, M.; Vila, E.; Janssen, D.; Bretones, G.; Salvador, E.; Lara, L.; Téllez, M. Selection of refuges for Nesidiocoris tenuis (Het.: Miridae) and Orius laevigatus (Het.: Anthocoridae): virus reservoir risk assessment. IOBC/OILB, wprs/srop Bull. 2009, 49, 281–286. [Google Scholar]
- Alomar, O.; Gabarra, R.; González, O.; Arnó, J. Selection of insectary plants for ecological infrastructure in Mediterranean vegetable crops; International Organization for Biological and Integrated Control of Noxious Animals and Plants (OIBC/OILB), West Palaearctic Regional Section (WPRS/SROP): Dijon, 2006; Vol. 29, pp. 5–8. [Google Scholar]
- Parolin, P.; Bresch, C.; Poncet, C.; Desneux, N. Functional characteristics of secondary plants for increased pest management. Int. J. Pest Manag. 2012, 58, 369–377. [Google Scholar] [CrossRef]
- Badenes-Pérez, F.R. Trap Crops and Insectary Plants in the Order Brassicales. Ann. Entomol. Soc. Am. 2019, 112, 318–329. [Google Scholar] [CrossRef]
- Colley, M.R.; Luna, J.M. Relative Attractiveness of Potential Beneficial Insectary Plants to Aphidophagous Hoverflies (Diptera: Syrphidae). Environ. Entomol 2000, 29, 1054–1059. [Google Scholar] [CrossRef]
- Bennison, J.; Pope, T.; Maulden, K. The potential use of flowering alyssum as a “banker” plant to support the establishment of Orius laevigatus in everbearer strawberry for improved biological control of western flower thrips. IOBC/WPRS Bull. 2011, 68 PP-D, 15–18. [Google Scholar]
- Pumariño, L.; Alomar, O. The role of omnivory in the conservation of predators: Orius majusculus (Heteroptera: Anthocoridae) on sweet alyssum. Biol. Control 2012, 62, 24–28. [Google Scholar] [CrossRef]
- Berndt, L.A.; Wratten, S.D. Effects of alyssum flowers on the longevity, fecundity, and sex ratio of the leafroller parasitoid Dolichogenidea tasmanica. Biol. Control 2005, 32, 65–69. [Google Scholar] [CrossRef]
- BEGUM, M.; GURR, G.M.; WRATTEN, S.D.; HEDBERG, P.R.; NICOL, H.I. Using selective food plants to maximize biological control of vineyard pests. J. Appl. Ecol. 2006, 43, 547–554. [Google Scholar] [CrossRef]
- Aparicio, Y.; Riudavets, J.; Gabarra, R.; Agustí, N.; Rodríguez-Gasol, N.; Alins, G.; Blasco-Moreno, A.; Arnó, J. Can Insectary Plants Enhance the Presence of Natural Enemies of the Green Peach Aphid (Hemiptera: Aphididae) in Mediterranean Peach Orchards? J. Econ. Entomol. 2021, 114, 784–793. [Google Scholar] [CrossRef]
- Rodríguez-Gasol, N.; Avilla, J.; Aparicio, Y.; Arnó, J.; Gabarra, R.; Riudavets, J.; Alegre, S.; Lordan, J.; Alins, G. The Contribution of Surrounding Margins in the Promotion of Natural Enemies in Mediterranean Apple Orchards. Insects 2019, 10, 148. [Google Scholar] [CrossRef] [PubMed]
- Tavares, J.; Wang, K.-H.; Hooks, C.R.R. An evaluation of insectary plants for management of insect pests in a hydroponic cropping system. Biol. Control 2015, 91, 1–9. [Google Scholar] [CrossRef]
- Resende, A.L.S.; Souza, B.; Ferreira, R.B.; Aguiar-Menezes, E.L. Flowers of Apiaceous species as sources of pollen for adults of Chrysoperla externa (Hagen) (Neuroptera). Biol. Control 2017, 106, 40–44. [Google Scholar] [CrossRef]
- Jeddi, K.; Abbes, K.; Lassoued, M.; Jeddi, K.; Hessini, K.; Siddique, K.H.M.; Chermiti, B. Attractiveness of mediterranean native plants to arthropod natural enemies and herbivores. Arthropod. Plant. Interact. 2025, 19, 98. [Google Scholar] [CrossRef]
- Calvo, F.J. Evolución del control de plagas en la horticultura española: papel del control biológico aumentativo. Phytoma España 2019, 96–100. [Google Scholar]
- Rodríguez, E.; González, M.; Paredes, D.; Campos, M.; Benítez, E. Selecting native perennial plants for ecological intensification in Mediterranean greenhouse horticulture. Bull. Entomol. Res. 2018, 108, 694–704. [Google Scholar] [CrossRef]
- He, Z.; Guo, J.-F.; Reitz, S.R.; Lei, Z.-R.; Wu, S.-Y. A global invasion by the thrip, Frankliniella occidentalis: Current virus vector status and its management. Insect Sci. 2020, 27, 626–645. [Google Scholar] [CrossRef]
- EPPO Data sheets on Quarantine Pests: Frankliniella occidentalis; 1989.
- Calvo, F.J.; Knapp, M.; van Houten, Y.M.; Hoogerbrugge, H.; Belda, J.E. Amblyseius swirskii: What made this predatory mite such a successful biocontrol agent? Exp. Appl. Acarol. 2015, 65, 419–433. [Google Scholar] [CrossRef]
- Brodsgaard, H. Biological control of thrips on ornamental crops. In Biocontrol in Protected Culture; Heinz, K., Van Driesche, R., Parrella, M., Eds.; Ball Publishing: Batavia, IL, 2004; pp. 253–264. [Google Scholar]
- Sanchez, J.; Alcázar, A.; Lacasa, A.; Llamas, A.; Bielza, P. Integrated pest management strategies in sweet pepper plastic houses in the southeast of Spain. IOBC/WPRS Bull 2000, 23, 21–30. [Google Scholar]
- Chow, A.; Chau, A.; Heinz, K. Control of Frankliniella occidentalis on greenhouse roses with Amblyseius (Typhlodromips) swirskii and Orius insidiosus. IOBC/WPRS Bull 2008, 32, 45–48. [Google Scholar]
- Cano, M.; Salvador, E.; Janssen, D.; Lara, L.; Tellez, M. Utilización de Mentha suaveolens Ehrh y Ocimum basilicum Linnaeus como plantas refugio para adelantar la instalación de Orius laevigatus Fieber ( Hemiptera : Anthocoridae ) en cultivo de pimiento. Bol. San. Veg. Plagas 2012, 38, 311–319. [Google Scholar]
- Mound, L.A.; Morison, G.D.; Pitkin, B.R.; Palmer, J.M. Handbooks for the identification of British insects . In part 11. Thysanoptera; Royal Entomological Society of London.: London, 1976; Vol. 1. [Google Scholar]
- Palmer, J.M.; Mound, L.A.; Heaume, G.J. CIE guides to insects of importance to man. 2. Thysanoptera.; CAB International: Wallingford, Oxon, 1989; ISBN 9780851986340. [Google Scholar]
- Curso práctico de entomología; Barrientos, A.J., Barrientos, J.A., Eds.; Manuals: Alicante, Bellaterra; 41; Asociación Española de Entomología; CIBIO; Universidad Autónoma de Barcelona, 2004; ISSN ISBN 8449023831. [Google Scholar]
- Ribeiro, A.L.; Gontijo, L.M. Alyssum flowers promote biological control of collard pests. BioControl 2017, 62, 185–196. [Google Scholar] [CrossRef]
- Tiwari, S.; Sharma, S.; Wratten, S.D. Flowering alyssum (Lobularia maritima) promote arthropod diversity and biological control of Myzus persicae. J. Asia. Pac. Entomol. 2020, 23, 634–640. [Google Scholar] [CrossRef]
- Pumariño, L.; Alomar, O. Assessing the use of Lobularia maritima as an insectary plant for the conservation of Orius majusculus and biological control of Frankliniella occidentalis. IOBC/WPRS Bull. 2014, 100, 113–116. [Google Scholar]
- Picó, F.X.; Retana, J. The flowering pattern of the perennial herb Lobularia maritima: an unusual case in the Mediterranean basin. Acta Oecologica 2001, 22, 209–217. [Google Scholar] [CrossRef]
- Bahşi, Ş.Ü.; Tunç, İ. Development, survival and reproduction of Orius niger (Hemiptera:Anthocoridae) under different photoperiod and temperature regimes. Biocontrol Sci. Technol. 2008, 18, 767–778. [Google Scholar] [CrossRef]
- van den Meiracker, R.A.F. Induction and termination of diapause in Orius predatory bugs. Entomol. Exp. Appl. 1994, 73, 127–137. [Google Scholar] [CrossRef]
- Tommasini, M.G.; Van Lenteren, J.C. Occurrence of diapause in orius laevigatus. Bull. Insectology 2003, 56, 225–251. [Google Scholar]
- Riudavets, J.; Castañé, C.; Gabarra, R. Native Predators of Western Flower Thrips in Horticultural Crops. In Thrips Biology and Management; Parker, B.L., Skinner, M., Lewis, T., Eds.; Springer US: Boston, MA, 1995; pp. 255–258. ISBN 978-1-4899-1409-5. [Google Scholar]
- Sciarretta, A.; Travaglini, T.; Kfoury, L.; Ksentini, I.; Yousef-Yousef, M.; Sotiras, M.-I.; El Bitar, A.; Ksantini, M.; Quesada-Moraga, E.; Perdikis, D. Comparison of different trapping devices for the capture of Bactrocera oleae (Rossi) and other non-target insects in the Mediterranean basin. J. Entomol. Acarol. Res. 2024, 56. [Google Scholar] [CrossRef]
- Moreno-Alcaide, F.; Quesada-Moraga, E.; Valverde-García, P.; Yousef-Yousef, M. Optimizing decision-making potential, cost, and environmental impact of traps for monitoring olive fruit fly Bactrocera oleae (Rossi) (Diptera: Tephritidae). J. Econ. Entomol. 2025. [Google Scholar] [CrossRef]
- Araj, S.-E.; Shields, M.W.; Wratten, S.D. Weed floral resources and commonly used insectary plants to increase the efficacy of a whitefly parasitoid. BioControl 2019, 64, 553–561. [Google Scholar] [CrossRef]
- Araj, S.-E.; Wratten, S.D. Comparing existing weeds and commonly used insectary plants as floral resources for a parasitoid. Biol. Control 2015, 81, 15–20. [Google Scholar] [CrossRef]
- Arnó, J.; Oveja, M.F.; Gabarra, R. Selection of flowering plants to enhance the biological control of Tuta absoluta using parasitoids. Biol. Control 2018, 122, 41–50. [Google Scholar] [CrossRef]
Figure 1.
Distribution of insectary plants and ornamental crops in the greenhouse during the two years of the study. (a) 2021, (b) 2025.
Figure 1.
Distribution of insectary plants and ornamental crops in the greenhouse during the two years of the study. (a) 2021, (b) 2025.
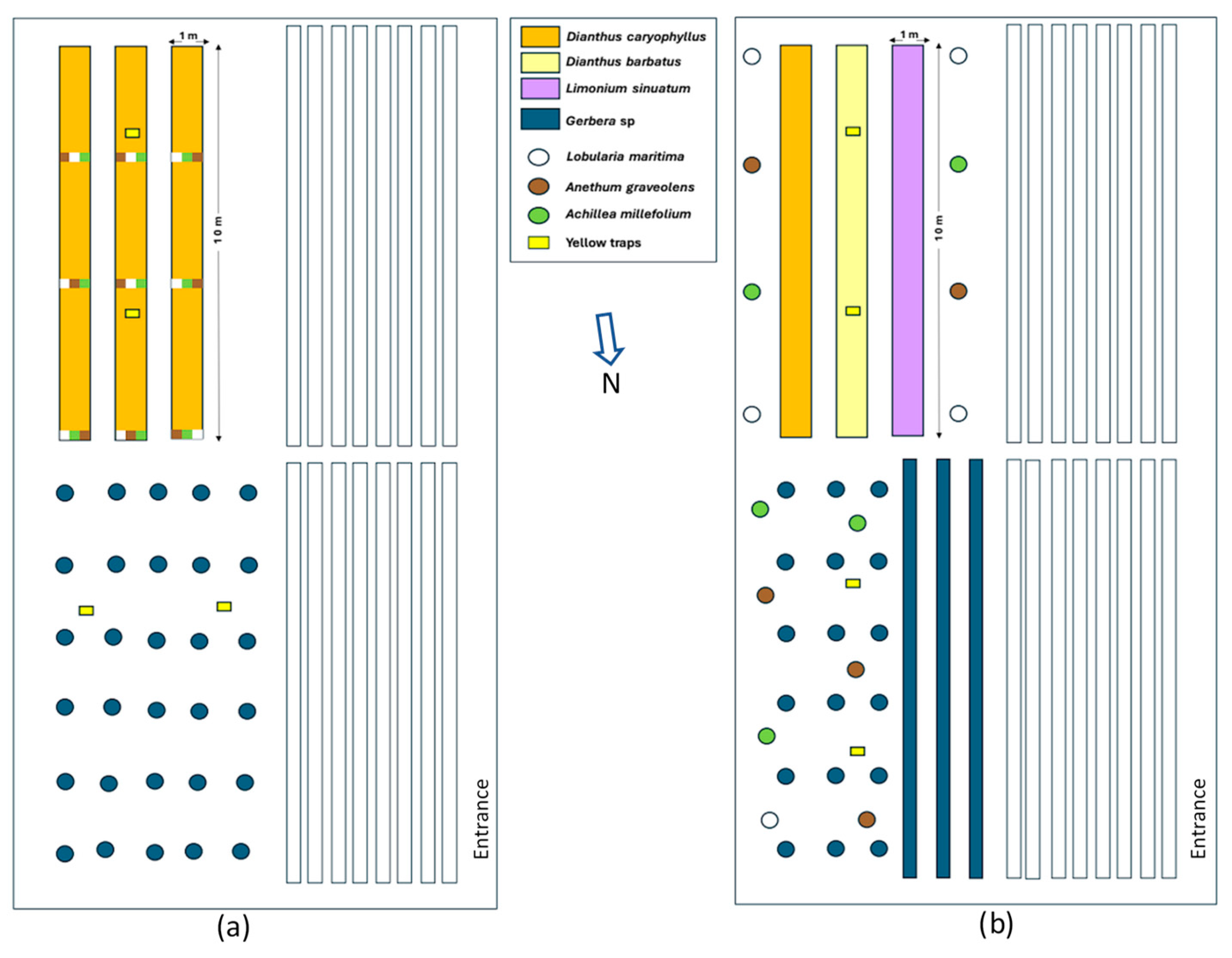
Figure 2.
Seasonal changes in the mean population of Orius laevigatus (adults and nymphs) per sampling unit in the insectary plants. a) and b) in 2021, c) and d) in 2025. Arrows indicate the moment O. laevigatus adults were released in the insectary plants. Vertical bars are standard errors of the mean.
Figure 2.
Seasonal changes in the mean population of Orius laevigatus (adults and nymphs) per sampling unit in the insectary plants. a) and b) in 2021, c) and d) in 2025. Arrows indicate the moment O. laevigatus adults were released in the insectary plants. Vertical bars are standard errors of the mean.
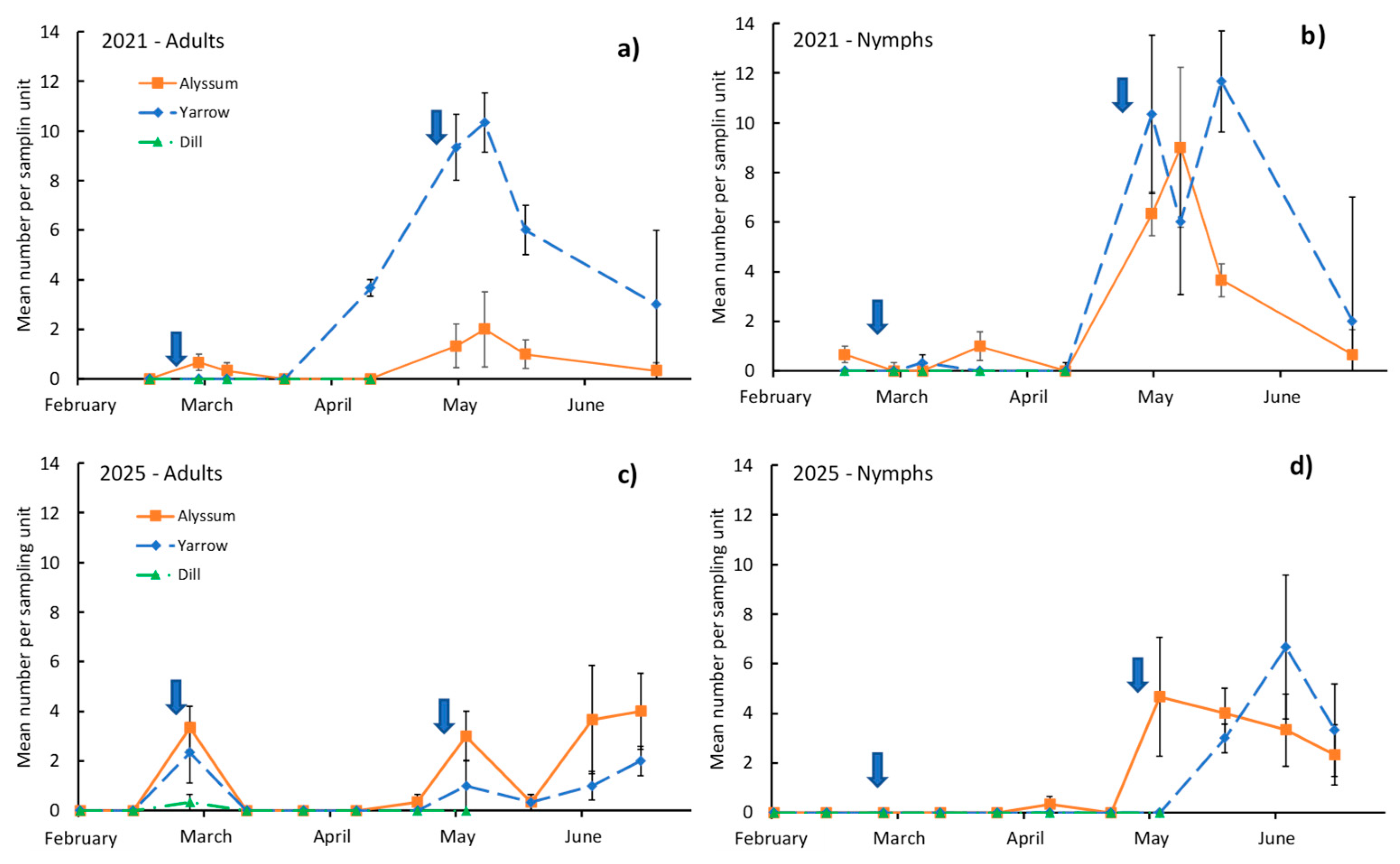
Table 1.
Number of arthropods observed in the insectary plants in the two years.
| Thysanoptera | Orius total | Orius adults | Orius nymphs | Aphididae | Formicidae | Hymenoptera parasitoids | Araneae | Acari | Collembola | TOTAL1 | |
| 2021 | |||||||||||
| Alyssum | 345 | 81 | 17 | 64 | 32 | 56 | 3 | 44 | 101 | 532 | 1226 |
| yarrow | 557 | 188 | 97 | 91 | 10 | 10 | 4 | 39 | 9 | 376 | 1199 |
| Dill | 170 | 0 | 0 | 0 | 2660 | 0 | 1 | 6 | 0 | 24 | 2863 |
| Total | 1072 | 269 | 114 | 155 | 2702 | 66 | 8 | 89 | 110 | 932 | 5288 |
| 2025 | |||||||||||
| Alyssum | 163 | 88 | 44 | 44 | 0 | 43 | 4 | 23 | 9 | 242 | 649 |
| yarrow | 270 | 59 | 20 | 39 | 3 | 5 | 10 | 32 | 10 | 248 | 657 |
| Dill | 287 | 1 | 1 | 0 | 0 | 6 | 0 | 9 | 1 | 20 | 333 |
| Total | 720 | 148 | 65 | 83 | 3 | 54 | 14 | 64 | 20 | 510 | 1639 |
| TOTAL (two years) | 1792 | 417 | 179 | 238 | 2705 | 120 | 22 | 153 | 130 | 1442 | 6927 |
1 All the arthropods observed in each plant are included.
Table 2.
Number of arthropods observed in the yellow sticky traps in the two years.
| Thysanoptera | Orius | Diptera | Aphididae | Aleyrodidae | Araneae | Coleoptera | Hymenoptera | TOTAL1 | |
| 2021 | 2591 | 12 | 837 | 4146 | 109 | 43 | 22 | 482 | 8371 |
| 2025 | 6607 | 7 | 5531 | 129 | 0 | 82 | 35 | 41 | 12500 |
| TOTAL | 9198 | 19 | 6368 | 4275 | 109 | 125 | 57 | 523 | 20871 |
1 All the arthropods observed are included.
Table 3.
Mean values (with standard error) per sampling date and sampling unit of the principal arthropods found in the insectary plants in 2021 and 2025. The repeated measures ANOVA was used to compare two plant species (alyssum and yarrow) and three plant species (alyssum, yarrow, and dill).
Table 3.
Mean values (with standard error) per sampling date and sampling unit of the principal arthropods found in the insectary plants in 2021 and 2025. The repeated measures ANOVA was used to compare two plant species (alyssum and yarrow) and three plant species (alyssum, yarrow, and dill).
| Plant species | Comparation of two plant species1 | Comparation of three plant species2 | ||||||||||||||||||
| Alyssum | Yarrow | Dill | Plant species | Date x Plant | Plant species | Date x Plant | ||||||||||||||
| Mean3 | s.e. | Mean3 | s.e. | Mean4 | s.e. | F5 | p | F6 | p | F7 | p | F6 | p | |||||||
| 2021 | ||||||||||||||||||||
| Thysanoptera | 12.8 | 3.2 | 20.6 | 10.4 | 11.3 | 2.2 | 8.0 | 0.048 (*) | 8.0 | 0.066 | 5.1 | 0.052 | 8.4 | 0.006(*) | ||||||
| Orius total | 3.0 | 1.3 | 7.0 | 2.8 | 0.0 | 0.0 | 44.7 | 0.003(*) | 4.0 | 0.056 | 11.2 | 0.009(*) | 19.4 | <0.001(*) | ||||||
| Orius adults | 0.6 | 0.2 | 3.6 | 1.4 | 0.0 | 0.0 | 200.0 | <0.001(*) | 6.0 | 0.027(*) | 97.0 | <0.001(*) | 38.1 | <0.001(*) | ||||||
| Orius nymphs | 2.4 | 1.1 | 3.4 | 1.6 | 0.0 | 0.0 | 4.1 | 0.112 | 4.6 | 0.028((*) | 2.6 | 0.152 | 2.9 | 0.087 | ||||||
| Aphididae | 1.2 | 0.5 | 0.4 | 0.2 | 177.3 | 124.9 | 1.1 | 0.349 | 0.7 | 0.483 | 9.6 | 0.014(*) | 3.0 | 0.124 | ||||||
| Formicidae | 2.1 | 0.5 | 0.4 | 0.3 | 0.0 | 0.0 | 4.1 | 0.114 | 0.8 | 0.470 | 2.4 | 0.170 | 0.7 | 0.540 | ||||||
| Hymenoptera | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.0 | 0.851 | 1.2 | 0.359 | 0.7 | 0.533 | 1.1 | 0.403 | ||||||
| Araneae | 1.6 | 0.6 | 1.4 | 0.6 | 0.4 | 0.1 | 0.3 | 0.597 | 2.9 | 0.107 | 25.8 | 0.001(*) | 1.3 | 0.331 | ||||||
| Acari | 3.7 | 2.3 | 0.3 | 0.2 | 0.0 | 0.0 | 9.2 | 0.038(*) | 1.1 | 0.347 | 10.2 | 0.012(*) | 0.9 | 0.452 | ||||||
| Collembola | 19.7 | 8.4 | 13.9 | 5.4 | 1.6 | 0.8 | 4.0 | 0.115 | 0.9 | 0.423 | 31.3 | 0.001(*) | 2.1 | 0.174 | ||||||
| Total arthropods | 45.4 | 8.0 | 44.4 | 9.6 | 190.9 | 122.4 | 0.0 | 0.871 | 2.7 | 0.130 | 5.2 | 0.050 | 2.7 | 0.142 | ||||||
| 2025 | ||||||||||||||||||||
| Thysanoptera | 4.9 | 1.2 | 8.2 | 4.0 | 12.0 | 4.2 | 6.7 | 0.060 | 3.8 | 0.106 | 7.6 | 0.02(*) | 2.9 | 0.114 | ||||||
| Orius total | 2.7 | 1.0 | 1.8 | 0.8 | 0.0 | 0.0 | 2.6 | 0.182 | 1.5 | 0.273 | 14.1 | 0.005(*) | 7.6 | <0.001(*) | ||||||
| Orius adults | 1.3 | 0.5 | 0.6 | 0.3 | 0.0 | 0.0 | 3.0 | 0.157 | 1.0 | 0.417 | 8.2 | 0.019(*) | 2.6 | 0.009(*) | ||||||
| Orius nymphs | 1.3 | 0.6 | 1.2 | 0.7 | 0.0 | 0.0 | 0.1 | 0.782 | 1.8 | 0.217 | 3.6 | 0.095 | 3.7 | <0.001(*) | ||||||
| Aphididae | 0.0 | 0.0 | 0.1 | 0.1 | 0.0 | 0.0 | - | - | - | - | - | - | - | - | ||||||
| Formicidae | 1.3 | 0.4 | 0.2 | 0.1 | 0.3 | 0.2 | 32.9 | 0.005(*) | 0.6 | 0.577 | 15.2 | 0.004(*) | 0.4 | 0.788 | ||||||
| Hymenoptera | 0.1 | 0.1 | 0.3 | 0.2 | 0.0 | 0.0 | 3.3 | 0.145 | 0.9 | 0.412 | 2.4 | 0.174 | 1.3 | 0.335 | ||||||
| Araneae | 0.7 | 0.2 | 1.0 | 0.3 | 0.4 | 0.1 | 1.4 | 0.295 | 1.4 | 0.299 | 0.9 | 0.465 | 1.0 | 0.466 | ||||||
| Acari | 0.3 | 0.2 | 0.3 | 0.2 | 0.0 | 0.0 | 0.0 | 0.904 | 1.8 | 0.223 | 1.2 | 0.370 | 1.9 | 0.182 | ||||||
| Collembola | 7.3 | 1.8 | 7.5 | 2.8 | 0.8 | 0.8 | 0.0 | 0.910 | 2.6 | 0.123 | 17.8 | 0.003(*) | 2.5 | 0.011(*) | ||||||
| Total arthropods | 19.7 | 2.3 | 19.9 | 5.7 | 13.9 | 4.7 | 0.0 | 0.919 | 4.3 | 0.045(*) | 5.7 | 0.042(*) | 3.5 | 0.001(*) | ||||||
1 Statistical comparations made with alyssum and yarrow, using the total sampling dates in each year: 9 sampling dates in 2021, and 11 sampling dates in 2025. 2 Statistical comparations made with alyssum, yarrow, and dill using the sampling dates in which the three plants were present simultaneously: 5 sampling dates in 2021, and 8 sampling dates in 2025. 3 Mean value obtained with the total number of sampling dates made in 2021 (9 sampling dates) and in 2025 (11 sampling dates). 4 Mean value obtained with the number of sampling dates available in 2021 (5 sampling dates) and in 2025 (8 sampling dates). 5 Degrees of freedom of the F test: 1, 4. 6 Degrees of freedom varied in each year, and also depending if the sphericity test of Mauchly was significant, in which case the Greenhouse-Geiser correction was used. 7 Degrees of freedom of the F test: 2, 6. (*) Significant, with p < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



