
Submitted:
06 May 2025
Posted:
08 May 2025
You are already at the latest version
Abstract
The invasive Delottococcus aberiae is one of the most harmful mealybugs on citrus in Spain, since it causes deformations and the premature falling of fruits. To improve control strategies, this study evaluates its population above and belowground, distribution in the soil and identify natural enemies emerging from belowground of citrus orchards. The distribution in the soil of emerged males varies significantly with the distance to the tree trunks with an annual average number of 0.95 males/trap/day at 0.5 m, and ranging between 0.25-0.32 at 1 m, 1.5 m and 2 m. There is a constant emergence of males throughout the year, both aboveground and belowground, with four distinct peaks, the first two occurring in February and April. This observation is essential to manage fruit damages that follows between March and June. Among the natural enemies detected belowground the following are highlighted: Bdellidae mites (52%), Hymenoptera parasitoids (16%) and spiders (11%). When implementing IPM strategies, emphasis should be place on adequate soil management (crops, mulching, or the release of natural enemies) and to the appearance of first generation. We propose a belowground population sampling method as a simple and practical tool to quantify this pest’s the winter and spring populations, which remain undetectable using current methodologies.
Keywords:
sampling methodologies
; population dynamics
; natural enemies
; soil distribution
; male captures
; Coccoidea
1. Introduction
On a global scale, invasive insects are one of the most important threats facing an agriculture. As a result, there is a serious risk of economic losses to crops and a challenge to human food security [1]. Due to a lack of effective natural enemies, a foreign insect will spread very quickly when it enters a new agricultural area. An example of this occurred in 2009 in Spain with the arrival of the Delottococcus aberiae De Lotto (Hemiptera: Pseudococcidae) [2]. This insect, responsible for fruit deformities and significant economic losses in citrus crops, originates from South Africa, where it is not considered a pest [2]. It was first detected near Valencia (eastern Spain), and continues to spread. In 2023 (14 years after it was identified), it was present in almost all citrus-producing areas of eastern Spain (about 200,000 hectares) from Catalonia in the north to Murcia region in the south [3]. According to a recent study conducted by Gavara et al. [3] this mealybug is in full expansion and will eventually affect all citrus-growing areas in Spain. Its presence has also been reported in other crops such as persimmons, medlar, pomegranate, and olive trees [4]. The damage provoked by this insect makes it one of the most harmful cushion scales currently found in Spain. The feeding behaviour of this species leads to direct injury to fruits, distorting their shape, producing premature falling, and/or causing a reduction in their size. Thus, two types of deteriorations have been described: small-sized fruits that do not grow and remain on the tree (aberrant, dwarf, and totally deformed fruits), and fruits that do reach the proper size but are deformed in the areas that have been stung by the insect, making them commercially unviable [3].
To control this pest, an Integrated Pest Management (IPM) program is being implemented, combining mass trapping pheromones and biological, chemical and cultural controls. Chemical control products such as acetamiprid, paraffin oil, and spirotetramat are recommended. However, this application often fails to effectively control D. aberiae due to their resistance to insecticides, influenced by, among other reasons, its protective waxy coating and its reproduction rate. In addition, this insect can find hidden places in plants where it can take refuge [3,5]. Since 2020, classical biological control programs have been developed in Spain by introducing a parasitoid insect from South Africa, Anagyrus aberiae Guerrieri [5]. Previously, it was demonstrated that the Spanish native and naturalized parasitoids failed to control this mealybug due to high immature mortality, encapsulation, and its defensive mechanisms [6]. For this reason, this type of control is combined with a summer release of the predator Cryptolaemus montrouzieri (Mulsant) with the aim of reducing populations, when they reach their peak. A great breakthrough was the discovery of the sex pheromone of D. aberiae [7], which it is now commercially available for mass capture using the "Attract and Kill" technique (Vynyty® Citrus* BAYER). In addition, aeration pruning is recommended as a cultural control.
Despite all the control strategies currently in place, this pest continues to account for 30-70% crop loss [3]. The severity of the problem is such that government authorities provide subsidies to producers to compensate for losses. [8]. Furthermore, as mentioned above, its expansion in the Mediterranean region is likely to continue, as it has since its arrival in Spain, potentially affecting other citrus-growing regions. Therefore, on-going research is fundamental to improve control strategies, focusing on the pest’s biology and establishing fast and efficient sampling strategies to aid in decision-making.
Its biology, population dynamics [3] and distribution on the tree are well understood [9,10], but little is known about the D. aberiae population that may exist belowground, as occurs in other mealybugs [11,12]. These studies showed that there is movement of females and immature insects between aboveground (canopy) and belowground (ground) in both directions, although the belowground population dynamics are completely unknown. It is also unclear how this belowground population compares to the aboveground population or how it is distributed in the soil. Addressing these questions is the aim of this work, as it will allow to improve sampling methods and implement new management strategies. Additionally, natural enemies emerging from the soil are studied, along with their potential role as biological control agents for D. aberiae.
2. Materials and Methods
2.1. Study area and citrus varieties
The field experiments were carried out on a citrus farm of 20 hectares located in the Valencia Region (eastern Spain, 39°64'N 0°42'W), the most important citrus producer in Spain. The farm belongs to the Cooperative “Sant Vicent Ferrer de Benaguasil”, which is responsible for the management of these fields. The use of pesticides is minimized since it is under agroecological transition. The farm is divided into orchards (5 to 8-year-old trees), growing different varieties of citrus. The sampling was conducted on two Lanetale variety citrus orchards of 0.6 ha and 1.8 ha (Figure 1).
The mealybug D. aberiae invaded the farm in 2020, rapidly spreading, displacing previously existing mealybugs and becoming the dominate pest. Pest management in 2023 and 2024 was carried out following the principles of IPM, combining the use of pheromone “attack and kill” (Vynyty® Citrus* BAYER), with one treatment with Acetamiprid in April and inoculative releases of D. aberiae natural enemies in July (C. montrouzieri and A. aberiae).
2.2. Above and belowground sampling of D. aberiae and natural enemies
To sample the belowground population, the methodology developed by Vercher et al. [12], for Planococcus citri (Risso) in persimmon, was used. This consists of capturing males of D. aberiae emerging from the ground using traps with sexual pheromone. To do this, hard plastic trays (50 × 30 × 10 cm, 0.15 m2), were placed upside-down on the soil surface after removing leaf litter and grass. A 10 × 25 cm yellow sticky plastic trap (Econex, Murcia, Spain) was placed on the upper interior surface of each tray and baited with a red rubber septum loaded with 250 μg of synthetic D. aberiae sex pheromone (Zentinel® DAB, EPA SL, Carlet, Spain) The septa were replaced every two months, as a period of efficacy ensured by the manufacturer. Plastic trays were arranged side by side on the ground, forming a continuous line of 8 trays that span the gap between two opposite trees in adjacent rows. To prevent insects from entering laterally, the borders of the trays were covered and sealed with loose soil (Figure 1). The experiment was conducted in the central part of the orchard, and was replicated in the two orchards of the farm. Therefore, for each repetition, there were 2 trays at each of the following distances from the tree: 0.5 m, 1 m, 1.5 m and 2 m. In each sample, there were 4 repetitions of each of the indicate distances, 396 traps all in total. To follow aboveground population, Delta trap with pheromone was used on each orchard. They were placed in the canopy of the trees, in the inner branches (at 150–190 cm above the ground) representing 62 traps in total. From July 2023 until June 2024, above and belowground traps were collected and replaced every fourteen days.
In the laboratory, the D. aberiae specimens as well as all the natural enemies (down to the genus or family level, depending on the type of arthropod) were identified and counted using a stereomicroscope (Nikon SMZ745). This task was carried out by experts from the Mediterranean Agroforestry Institute (IAM-UPV). In some cases, it was not possible to identify the genus or family level because the arthropods were in poor condition.
To check the movement between above ground and below, corrugated cardboard bands were placed around tree trunks, since females and immature male instars take refuge to lay their eggs or to make cocoons [13]. The cardboard bands (approximately 40 cm wide each) were wrapped around the trunks of the trees and replaced fortnightly. The sampling period was March and April 2024. This period was selected because it had previously been shown that during these months there was an initial peak of scale insects climbing up and down the trunk [9,10]. In the laboratory, the corrugated cardboard boxes were opened, and the adult females and immature individuals were counted.
2.3. D. aberiae population on fruits
During the period of fruit set and development in citrus, like other mealybugs, D. aberiae tends to aggregate and concentrate on fruits. Therefore, the most appropriate sampling method for assessing its aerial females and immatures population (aboveground) is the direct evaluation on fruits [9,10]. Fortnightly, from July 2023 and June 2024, ten trees were randomly selected from the central area of each orchard. From each tree, six fruits were collected from five canopy positions: North, East, South, West, and the centre. In total, 3 repetitions were carried out, two in the largest orchard (1.8 ha), and one in the smallest (0.6 ha). Therefore, a total of 180 fruits were picked (60 fruits per replication), and examined in the laboratory under the stereomicroscope (this pest prefers to shelter in the internal area of the fruit calyx) [9]. The number of fruits with presence of females and immatures of D. aberiae was counted.
2.4. Fruit Damage caused by D. aberie
To assess the damage caused by D. aberiae on the fruits, the counts were conducted at the end of their growing season (October 2023 and 2024) since D. aberiae cause damages to the fruit between April and June, and the damage is not fully visible until the fruit is completely developed [3]. A visual sampling of fruits in the orchard was carried out. To this, 20 fruit (4 fruits per each orientation North, East, West, South and central part of the canopy) were inspected, out of a total of 23 trees previously randomly selected and marked on each orchard. Therefore, 460 fruits were counted each year, which were categorized into four categories, according to Gavara et al. [3], considering 0—healthy fruit; 1—slight fruit deformation; 2—fruit deformation with a clear loss of symmetry; and 3—aberrant fruits.
2.5. Data analysis
To standardize the results, the number of D. aberiae males captured in all traps (at the tree canopy and ground levels) was divided by the number of days each trap was in the field, to obtain the average number of mealybugs captured daily. In ground traps, the males per m² were calculated, considering that each trap at ground level covered an area of 0.15 m². In the case of natural enemies, the average was estimated as number of arthropods/trap/week.
The Statgraphics Centurion XIX.64 package (Fisher LSD; α = 0.05) was used to evaluate the effect of: 1. The distance (from the trunk) on the variable "number of males captured/day" through a one-way-ANOVA; 2. The month on the aboveground and belowground male population and on the percentage of fruits with D. aberiae through a one-way-ANOVA; 3. The month and the distance (from the trunk) on the abundance of natural enemies (arthropods/trap/week) by means of a multifactor ANOVA. Data on percentages were expressed as arcsine square-root-transformed to stabilise the variance before the ANOVAs. In the case of the daily or weekly number of arthropods per trap, the data was log-transformed (ln[captures + 1]) when necessary, to normalize residual data distribution and to homogenize the variance.
To assess the differences in the percentage of fruit damage across severity levels and percentage of marketable fruits, between the years 2023 and 2024, a paired comparative analysis was performed. For each damage level, the distribution of data was first assessed for normality using the Shapiro–Wilk test, applied separately to the 2023 and 2024 datasets. Given that several groups showed non-normal distributions (p < 0.05), the Wilcoxon signed-rank test, a non-parametric alternative to the paired t-test, was used to determine significant differences between years.
3. Results
3.1. Soil distribution of D. aberiae males
The average values of the captures of D. aberiae males in pheromone traps placed in trays on the ground at different distances (0.5, 1, 1.5, and 2 meters) from the citrus tree trunks are shown in Figure 2. The distribution of emerged males from the soil showed a significant difference in relation to the distance to the tree trunks, (F3, 393=2.91, P=0.034), between the traps placed near the trunk (0.5 m) and the traps placed at the rest of the distances (1 m, 1.5 m and 2 m). The annual average number of males was 0.95 /trap/day in trays located next to the trunk at 0.5 m, and between 3 and 4 times lower at the other distances, with values ranging between 0.25-0.32/trap/day.
3.2. Above and belowground D. aberiae seasonal evolution
The annual average of D. aberiae males was higher in the aerial traps (5.5±0.73 males/traps/day) than in the ground traps (0.4±0.10 males/traps/day, considering all distances). In Figure 3, aboveground males captures are compared with those from belowground, using only data from the trays located closest to the tree (0.5m), since at this distance the highest values were obtained on the ground. It is observed that there is a constant emergence of males throughout the year, both aboveground and belowground. The population peak in the soil traps occurs in July (3.25 ± 2.28 males/traps/day), with no significant differences between months (F11,99=0.95; P=0.497). The minimum levels on the tree canopy (1.72 ± 0.35 males/traps/day) occurred in January, while on the ground, were observed also in January (0.06 ± 0.022 males/traps/day) and in October (0.01 ± 0.008 males/traps/day). Expressing the number of emerging males per day and per square meter, there were 21.67 males/day/m² in July belowground. In the canopy traps, a significant effect is observed only in August compared to the other months (F11,50=3.18; P=0.025), with a mean value of 15.28 males/traps/day.
When analysing the population in fruits, expressed as the % of fruits with D. aberiae stages (red line in Figure 2), significant differences were observed (F6,26=13.60; P=0.000) between months, with July, August, and September being significantly higher than October and November. The maximum values were in July (44%) and the minimum in November (0%), close to the harvesting date (January). In the spring of 2024, the population increases starting in May, reaching 7% of fruits occupied by June. Additionally, with the cardboard method, the average number of females and immature males observed was the same in March and April, with a value of 0.14 ±0.04 mealybugs/day.
3.3. Male distribution of D. aberiae over the year
The distribution of D. aberiae males throughout the months (% of males in each month relative to the annual total captures) belowground (average of all distances) and aboveground is shown in Figure 4. Four peaks can be observed throughout the year, both belowground and aboveground. Belowground, the population peaks occur in February (11.4 % of total captures), in April (9.3 % of total), in July (23.3 % of total), and in November (15.4 % of total). Aboveground, the population peaks occur in February (5.4 % of total captures), in April (6.4 % of total), in August (25.5% of total), and in October (12.2 % of total).
3.4. Fruit damage level
The percentage of fruit with damage obtained at the end of the season 2023 and 2024 is shoed in Table 1. Data were first tested for normality using the Shapiro–Wilk test on each year's dataset. Since several variables violated the assumption of normality (p < 0.05), the Wilcoxon signed-rank test was used for all paired comparisons. Overall, results indicate that in 2024 there was a higher proportion of asymptomatic fruits (55.78 ± 4.21) compared to 2023 (31.75 ± 5.38). Likewise, in 2024 the incidence of fruits with level 1 (slight fruit deformation) was 40.31 ± 4.50 with level 2 (deformation with clear loss of symmetry) was 1.48 ± 0.48, which was lower than in 2023 (57.92 ± 4.70 and 8.92 ± 2.30, respectively). However, for aberrant fruits (level 3), no significant differences were observed between the two years with percentages of 1.42 ± 0.61 in 2023 and 2.42 ± 1.11 in 2024.
3.5. Presence and abundance of natural enemies belowground
The total number of natural enemies identified in the yellow sticky traps on the ground, as a function of the distance from the trunk are shown in Table 2. The results revel that the most common are from the class Arachnida (70%), compared to 30% represented by insects. Bdellidae mites represent 52%, spiders 11%, and Pseudoscorpionida 6% of the total. Among the insects, the most abundant are Hymenoptera parasitoids (16% of the total), followed by Diptera (11%) and the Staphylinidae beetles (3%). Among the Hymenoptera parasitoids, the families Mymaridae, Ceraphronidae, and Scelionidae are the most common, representing 25%, 16%, and 13% of the total parasitoids, respectively. The superfamily Ichneumonoidea was also important, representing 20% of the total, but only a few specimens could be classified to the family level, as the wings of many could not be clearly distinguished. Among the Diptera, the vast majority was Cecidomyiidae.
When observing their abundance based on the distance from the tree, a gradient was shown for some arthropods. For example, Bedillae and Cecidomyiidae were more abundant in the area farthest from the tree, while Pseudoscorpionida were more abundant near the tree. In other groups, such as Hemiptera and Staphylinidae, no gradient was observed.
The multifactor ANOVA (considering months and distance to the trunk; Table 3) of the most abundant arthropods before mentioned (arthropods/week/ trap), indicates no significant differences were observed in any case in relation to the distances. However, in all cases, there was a month effect, with the highest values observed in the summer period. This value was statistically significant for Staphylinidae (September), Alaptus spp. (September), Bdellidae (May and June), and Pseudoscorpionida (June).
4. Discussion
This study shows for the first time that there is a constant population of D. aberiae belowground, as males were detected and captured when emerging from the soil throughout the year. This discovery is of great importance because it reveals that there is a permanent pest population in the soil which has never been managed. This may explain the considerable difficulty in controlling this pest, which continues to cause significant damage despite the implementation of various management strategies. Previous studies using different methodologies also indicated the presence of this pest in the soil, locating nymphs and adult females belowground from February to September, although at very low levels, and only in the area near the trunk [10]. Our study demonstrates that there is a constant population, not only near the tree (where the concentration is highest), but also across the entire cultivation area between tree rows.
Observing the population levels of aboveground and belowground, the results are not directly comparable, as the soil traps cover an area of 0.15 m², while the aerial traps influence a much wider area. Therefore, some of the insects captured in the tree canopy may come from the emerging underground population. It should also be considered that the moisture that accumulates over time in the soil traps can reduce their capture capacity. Thus, the actual belowground population may be larger than what was observed in this study. Even so, these belowground captures at 0.5 m represent up to 33% of the total (above and belowground) in July and 24% in November. Expressing the number of emerging males per day and per square meter, there were 21.67 males/day/m2 in July belowground at 0.5 m from the trunk and 6.4 males/day/m2 considering all the distances observed herein. The sex ratio of mealybugs is usually 1:1 [14,15], this means, for each hectare of citrus 64,000 males can emerge from the soil in July resulting in a population of 128,000 adult mealybugs/ha. This suggests the importance of the belowground D. aberiae population.
Researchers who developed the D. aberie pheromone stated that “…it is possible to capture this scale insect at distances greater than 3 m” (V. Navarro, personal communication). Taking this into account (but being conservative and considering that each trap attracts males located within a 3 m radius), it could be said that each aerial trap attracts males within a surface area of 28.26 m2 (area of the 3 m radius circle). If these surfaces are considered, then the ground population becomes much more important, increasing from 10% of total captures to 90%. These results are approximate, and it will be interesting to conduct future specific studies that will allow us to more precisely compare these populations.
When studying the population dynamics referring to the number of adult males, a high peak is observed only in the summer season, with no other generations appearing, as shown by our data and confirmed in a 4-year study reported by Gavara et al. [3]. However, when the data are analysed as monthly male percentages relative to the annual total, it shows that there are 4 population peaks both aboveground and belowground. Belowground, the population peaks occur in February, April, July, and November. Aboveground, the first two peaks are also in February and April; however, the third peak occurs in August and the fourth in October. These variations are likely related to temperature differences belowground and aboveground. In the soil, temperatures are more stable, not undergoing the wide variations that occur aboveground, but neither do they reach the thermal peaks that occur in the Mediterranean during the warmer months [16].
To date, the number of generations of D. aberiae has not been determined because there is a mix of developmental stages starting in the summer [9]. It is common not to have a clear understanding of the generations of mealybugs in the Mediterranean, as many species show a similar pattern [12]. In the study conducted by Martínez-Blay et al. [9,10] only on aboveground females and immatures, two generations are clearly marked, the first in spring and the second in summer, coinciding with our second and third generations. These authors highlighted at least three more generations: one between January and February, another between August and October, and one more between October and December. Our results coincide with the February and October (aboveground) – November (belowground) generations, but no generation is observed between August and October. It is essential to identify the precise timing of generation peaks, as this enables pest management to be optimized for the most vulnerable stages. This is crucial for mealybugs, as adults are resistant to pesticides, and only those in their early stages are the most vulnerable [17]. Since most of fruit deformities occur in March-April-May-June [9,10], when aboveground populations are very low, controlling the first generation that occurs in winter, (which represents 12% of the total annual males) may be the key to improving pest management. Second generation, in April (9% of total), can also be important, as damage continues to occur until the end June. Gavara et al. [3] indicate that fruit injuries were not directly related to the maximum annual male catches, and the damage caused, and thus the economic losses, are related to the population levels in spring (April–June). This present research confirms these results and suggests that controlling the winter generation can be even more important to reduce fruit damage.
In general, aboveground mealybugs are difficult to detect during winter or at the beginning of the season due to their morphology, cryptic behaviour, and clumped distribution [18]. This is the case with D. aberiae, which is particularly hard to locate using traditional sampling methods (aboveground visual inspections searching for individuals or sooty mold resulting from honeydew residues) during the winter and spring [9,10]. Therefore, we recommend the method used in this study [12], which focuses on the 0.5 m area surrounding the trunk, together with pheromone traps placed in the canopy as a quick and simple way to sample the initial annual populations, which could be responsible for the most significant fruit damage. Since thresholds for this pest are still unknown, this method provides a valuable tool for future development [3]. Although this behaviour is observed for the first time in D. aberie, it has already been described in other species of Pseudococcidae [11]. Similar results were obtained by studying emerging males from the soil of Planococcus citri in persimmons [12] and citrus (unpublished data). Also, Planococcus ficus (Signoret), the vine mealybug, feeds on the vine roots, trunk, cordon, canes, leaves, and fruit [19]. Planococcus sp., P. citri, and Planococcus lilacinus (Cockerell) were found infesting the roots and basal region of the stem of black pepper vines and affecting the aerial parts such as the tender shoots, leaves, and berries [20]. Xu et al., [21] indicate that the mealybugs Saccharicoccus sacchari (Cockerell) and Heliococcus summervillei (Brookes) feed on sugarcane aboveground, on leaves, and belowground, on roots. There are also some mealybugs that live exclusively belowground and feed on sucking sap from plant rootlets, like the family Rhizoecidae, previously included in Pseudococcidae, and recently separated [22].
Soil and litter fauna play a primary role in ecosystem functioning, but their biodiversity and ecological relationships remain largely unknown [23]. In this study, we have identified the predators and parasitoids captured in soil traps. They are mostly predators, arachnids (Bdellidae mites, pseudoscorpions, and spiders), as well as Cecidomyiidae and Staphylinidae, all of which are common inhabitants of soils [24]. Many of these predators are generalists and can feed on mealybugs. Bdellidae are considered active predators of small arthropods such as soft-bodied insects, collembolans, and mites [25]. Cecidomyiidae are some of the most common natural enemies associated with mealybugs [26], such as Diadiplosis saccharum sp. nov., which is described as a predator of S. sacchari, which also lives in the roots [27]. Additionally, 16% of parasitoid hymenopterans have also been identified. Among them, the most common are Myrmaridae, Scelionidae, and Ceraphronidae. Scelionids are relevant for being egg parasitoids of arthropods and are observed in the soil [28]. ºare commonly collected in traps associated with soil and litter, and their host range is extensive, including Hemiptera, especially aphids and mealybugs [29]. All Myrmaridae are internal parasitoids of insect eggs and prefer hosts whose eggs are concealed within plant tissues, under bark, and in soil [30]. Most recorded hosts are Sternorrhyncha (Hemiptera), but the eggs of other Hemiptera, Coleoptera, and Psocoptera are also commonly attacked [31]. We also found some parasitoids from other families of Chalcidoidea, but in low numbers. Other studies report similar results, for example, [24], researching soil and litter, identified parasitoids of Eulophidae, Mymaridae, and Pteromalidae, representing only 0.5% of the total collected fauna. Lotfalizadeh et al. [32] also identified several Chalcidoidea in the soil, observing that the ecological role of this Superfamily in the soil is unknown. It is possible that some of these natural enemies feed on D. citri in the soil, but more specific studies are needed to understand the impact and role of this soil entomofauna in the control of mealybugs.
To manage this soil population, we propose biological control strategies through conservation (cover crops or mulching) and enhancement (soil arthropods and entomopathogens). Agroecological practices such as the implementation of cover crops are known to contribute to the maintenance of local biodiversity in farming systems [33] and increase natural enemies in the agroecosystem [34]. Several authors indicate that a rich cover crop increases the biodiversity of ground-dwelling arthropods. De Pedro et al. [28] demonstrate that in pear orchards, the ground cover increases the abundance of several families of spiders, beetles (Carabidae, Staphylinidae), and hymenopterans (Scelionidae). Sommaggio et al. [35] found a significantly higher activity and density of staphylinids and carabids on the soil surface of a vineyard with cover crops, compared to the control (without cover crops). In a study carried out by our team (unpublished data), we found that the captures of male D. aberiae on citrus trees were significantly lower when there was vegetative cover beneath the trees and between rows compared to when there was only vegetation between rows. Also, soil predatory mites in citrus could be enhanced in conservation biological control by adding compost [36] or through the application of mulch [37]. As an augmentative strategy, soil predatory mites can be used to reduce soil mealybugs population, since according to Pérez-Rodríguez et al. [38] they are potential biological control agents of D. aberiae. In fact, Gaeolaelaps aculeifer (Canestrini) and Stratiolaelaps miles (Berlese), have been successfully released for the augmentative biological control of different pests [39]. Regarding entomopathogens, currently there is interest in using Entomopathogenic nematodes (EPNs) on soil within an IPM scheme to reduce chemical pesticides, since EPNs is generally more suited to control soil-dwelling insect stages than aboveground insect pests [40]. Le Vieux and Malan [41] shown that EPNs, and specifically Steinernema yirgalemense Nguyen, Tesfamariam, Gozel, Gaugler and Adams (Rhabditida: Steinernematidae) show great potential as biological control agents for the control of P. ficus soil populations. Another alternative is use of Entomopathogenic fungi (EPF) that has also many advantages, such as specificity against pests and no effect on the environment or mammalian health and does not cause serious insect resistance [42]. The species most frequently isolated from soils in temperate regions belong to the genera Beauveria, Isaria (Cordycipitaceae) and Metarhizium (Clavicipitaceae) [43]. They have been shown to be effective in controlling aboveground mealybugs such as Paracoccus marginatus (Williams and Granara de Willink) [44] and Pseudococcus viburni Signoret [45]. Soil application of EPFs in field or semi-field conditions has successfully reduced the population of pests in their soil-dwelling stage, such as the beetles, thrips, and mites lepidopteran larvae [46].
5. Conclusions
The present study shows that a permanent population of D. aberie exists belowground in citrus orchards, primarily concentrated within the first 50 cm around the trunk and distributed throughout the soil between the tree rows. This belowground population can be widespread and should be considered when implementing IPM strategies. It would be interesting to conduct specific studies on the effectiveness of implementing cover crops, mulching, or the increased releases of soil arthropods or entomopathogens to reduce the significant damage caused to fruits by this invasive pest. This study suggests that the number of generations of this species is four, both aboveground and belowground. Furthermore, the importance of managing that first generation, which occurs belowground in winter and will later attack the fruits in March and April, is emphasized. To this end, we propose our belowground population sampling method as a simple and effective tool for detecting and quantifying the winter and spring populations of this pest, which are otherwise undetectable using conventional sampling methods.
Author Contributions
Conceptualization, R.V. and I.E.; methodology, R.V.; formal analysis, A.S.D.; investigation, R.V.; resources, R.V and I.E..; data curation, R.V. and A.S.D; writing—original draft preparation, R.V and I.E.; writing—review and editing, R.V and I.E.; project administration, I.E.; funding acquisition, I.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Next Generation European Union and the Plan de Recuperación, Transformación y Resiliencia of the Spanish Government, with the support of Generalitat Valenciana (project AGROALNEXT/2022/043). The authors also thank the project “Ruralfruit-AgroEco” subsidized by the PDR-CV 2014-2020.
Data Availability Statement
All data supporting the findings of this study are available from the corresponding authors upon reasonable request, subject to the restrictions imposed by the project’s funding agency.
Acknowledgments
The authors would like to thank “Cooperative Sant Vicent Ferrer de Benaguasil (Valencia)” for providing the orchards necessary to carry out this study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| IPM | Integrated Pest-Management |
References
- Skendžić, S.; Zovko, M.; Pajač Živković, I.; Lešić, V.; Lemić, D. Effect of Climate Change on Introduced and Native Agricultural Invasive Insect Pests in Europe. Insects 2021, 12, 985. [Google Scholar] [CrossRef] [PubMed]
- Beltrà, A.; Garcia-Marí, A.; Soto, A. El cotonet de Les Valls, Delottococcus aberiae, nueva plaga de los cítricos. Levante Agrícola 2013, 419, 348–352. Available online: https://www.researchgate.net/profile/Aleixandre-Beltra/publication/259621470_El_cotonet_de_Les_Valls_Delottococcus_aberiae_nueva_plaga_de_los_citricos/links/02e7e535a1553691e6000000/El-cotonet-de-Les-Valls-Delottococcus-aberiae-nueva-plaga-de-los-citricos.pdf (accessed on 21 April 2025).
- Gavara, A.; Vacas, S.; Navarro-Llopis, V. Geographic Location, Population Dynamics, and Fruit Damage of an Invasive Citrus Mealybug: The Case of Delottococcus aberiae De Lotto in Eastern Spain. Insects 2024, 15, 659. [Google Scholar] [CrossRef] [PubMed]
- MAPA (Ministerio de Agricultura, Pesca y Alimentación). Plan de Acción de Delottococcus aberiae (De Lotto). Available online: https://www.mapa.gob.es/es/agricultura/temas/sanidad-vegetal/plan_accion_d_aberiae_mayo2024_tcm30-684109.pdf (accessed on 21 April 2025).
- Soto, A.; Benito, M.; Puig Bargués, J.; Mocholí, S.; Martínez-Blay, V. Avances en la aplicación del control biológico del cotonet Delottococcus aberiae (De Lotto) (Hemiptera: Pseudococcidae). Phytoma España 2020, 318, 26–30. Available online: https://dialnet.unirioja.es/servlet/articulo?codigo=8412618 (accessed on 23 April 2025).
- Tena, A.; García-Bellón, J.; Urbaneja, A. Native and naturalized mealybug parasitoids fail to control the new citrus mealybug pest Delottococcus aberiae. J Pest Sci. 2017, 90, 659–667. [Google Scholar] [CrossRef]
- Vacas, S.; Navarro, I.; Marzo, J.; Navarro-Llopis, V.; Primo, J. Sex pheromone of the invasive mealybug citrus pest, Delottococcus aberiae (Hemiptera: Pseudococcidae). A new monoterpenoid with a necrodane skeleton. J. Agric. Food Chem. 2019, 67, 9441–9449. [Google Scholar] [CrossRef]
- DOGV (Diari oficial de la Generalitat Valenciana). Núm. 9961, del 21.10.2024. RESOLUCIÓN de 23 de septiembre de 2024, de la Direcció General de Producción Agrícola y Ganadera, de solicitudes aprobadas (lote número 2) en las ayudas convocadas por la Resolución de 8 de julio de 2022, del director general de Agricultura, Ganadería y Pesca, por la que se convocan las ayudas de mínimis destinadas a compensar las pérdidas económicas de las explotaciones agrícolas de cítricos y de caqui durante la campaña 2021-2022 como consecuencia de la afección de la plaga de Delottococcus aberiae (De Lotto) y de otros cotonets. 2024, 1-3. Available online: https://dogv.gva.es/datos/2024/10/21/pdf/2024_10030_es.pdf (accessed on 23 April 2025).
- Martínez-Blay, V.; Perez-Rodriguez, J.; Tena, A.; Soto, A. Density and phenology of the invasive mealybug Delottococcus aberiae on citrus: implications for integrated pest management. J. Pest. Sci. 2018, 91, 625–637. [Google Scholar] [CrossRef]
- Martínez-Blay, V.; Pérez-Rodríguez, J.; Tena, A., Soto, A. Seasonal Distribution and Movement of the Invasive Pest Delottococcus aberiae (Hemiptera: Pseudococcidae) Within Citrus Tree: Implications for Its Integrated Management. Econ. Entomol. 2018, 111, 6, 2684–2692. [CrossRef]
- Franco, J.C.; Suma, P.; Silva, E.; Blumberg, D.; Mendel, Z. (2004). Management strategies of mealybug pests of citrus in Mediterranean countries. Phytoparasitica 2004, 32, 507–522. [Google Scholar] [CrossRef]
- Vercher, R.; González, S.; Sánchez-Domingo, A.; Sorribas, J. A Novel Insect Overwintering Strategy: The Case of Mealybugs. Insects 2023, 14, 481. [Google Scholar] [CrossRef]
- Goolsby, J.; Kirk, A.; Meyerdirk, D.E. Seasonal phenology and natural enemies of Maconellicoccus hirsutus (Hemiptera: Pseudococcidae) in Australia. Fla Entomol. 2002. 85, 494- 498. [CrossRef]
- James, H.C. Sex ratios and the status of the male in Pseudococcinae (Hem. Coccidae). Bull. Entomol. Res. 1937, 28, 429–461. [Google Scholar] [CrossRef]
- Tong, H.; Li, Z.; Ye, W.; Wang, Y.; Omar, M.A.A.; Ao, Y.; …;Jiang, M. Male mating and female postmating performances in cotton mealybug (Hemiptera: Pseudococcidae): effects of female density. J. Econ. Entomol. 2019, 112, 1145–1150. [CrossRef]
- Muñoz-Romero, V.; López-Bellido, L.; Lopez-Bellido, R.J. Effect of tillage system on soil temperature in a rainfed Mediterranean Vertisol. Int. Agrophys. 2015, 29, 467–473. [Google Scholar] [CrossRef]
- Venkatesan, T.; Jalali, S.K.; Ramya, S.L.; Prathibha, M. Insecticide resistance and its management. In mealybugs and their Management. Agricultural and Horticultural Crops. Springer, New Delhi, India, 2016; pp. 223–229.
- Millar, J.G.; Daane, K.M.; Steven Mcelfresh, J.; Moreira, J.A.; Malakar-Kuenen, R.; Guillén, M.; Bentley, W.J. Development and optimization of methods for using sex pheromone for monitoring the mealybug Planococcus ficus (Homoptera: Pseudococcidae) in California vineyards. J. Econ. Entomol. 2002, 95, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.E.; Daane, K.M.; Bentley, W.J.; Gill, R.J.; Malakar-Kuenen, R. Mealybugs in California vineyards (Publication 21612); University of California, Agriculture & Natural Resources: Oakland, CA, USA, 2002. Available online: https://books.google.es/books?hl=en&lr=&id=lY-GA8inrnYC&oi=fnd&pg=PA2&dq=17.%09Godfrey,+K.E.%3B+Daane,+K.M.%3B+Bentley,+W.J.%3B+Gill,+R.J.%3B+Malakar-Kuenen,+R.+Mealybugs+in+California+vineyards+&ots=37LfnCv_bm&sig=s8U1-_t5kB0ElIYcqgPfN5oeUW8&redir_esc=y#v=onepage&q&f=false (accessed on 21 April 2025).
- Mani, M.; Smitha, M.S.; Najitha, U. Root mealybugs and their management in horticultural crops in India. PMHE. 2016, 22, 103–113. Available online: https://www.indianjournals.com/ijor.aspx?target=ijor:pmhe&volume=22&issue=2&article=001 (accessed on 21 April 2025).
- Xu, H.; Humpal, J.A.; Wilson, B.A.L.; Ash, G.J.; Powell, K.S. Mealybug Population Dynamics: A Comparative Analysis of Sampling Methods for Saccharicoccus sacchari and Heliococcus summervillei in Sugarcane (Saccharum sp. Hybrids). Insects 2024, 15, 492. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, C.J. Comparison of the morphology of the adult males of the Rhizoecine, Phenacoccine and Pseudococcine mealybugs (Hemiptera: Sternorrhyncha: Coccoidea) with the recognition of the family Rhizoecidae Williams. Zootaxa 2012, 3291, 1–79. Available online: https://www.mapress.com/zootaxa/2012/f/z03291p079f.pdf (accessed on 21 April 2025).
- Wall, D.H.; Bardgett, R.D.; Behan-Pelletier, V.; Jones, T.H.; Herrick, J.E. Soil Ecology and Ecosystem Services. Oxford University Press, Oxford, UK. 2012. Available online: https://books.google.es/books?hl=en&lr=&id=J_HTBgrYQdwC&oi=fnd&pg=PP2&dq=21.%09Wall,+D.H.,+Bardgett,+R.D.,+Behan-Pelletier,+V.,+Jones,+T.H.,+Herrick,+J.E.+Soil+Ecology+and+Ecosystem+Services.+ (accessed on 21 April 2025).
- Martínez-Falcón, A.P.; Moreno, C.E.; Pavón, N.P. Litter fauna communities and litter decomposition in a selectively logged and an unmanaged pine-oak forest in Mexico. Bosque 2015, 36, 81–93. Available online: https://revistabosque.org/index.php/bosque/article/view/558 (accessed on 21 April 2025). [CrossRef]
- Hernandes, F.A.; Skvarla, M.J., Fisher, J.R., Dowling, A.P.G., Ochoa, R., Ueckermann, E.A., Bauchan, G.R. Catalogue of snout mites (Acariformes: Bdellidae) of the world. Zootaxa 2016, 152 (1), 001-083. [CrossRef]
- Hayon, I.; Mendel, Z.; Dorchin, N. Predatory gall midges on mealybug pests -diversity, life history, and feeding behavior in diverse agricultural settings. Biol. Control 2016, 99, 19–27. [Google Scholar] [CrossRef]
- Urso-Guimarães, M.V.; Cruz, M.A.; Martinelli, N.M.; Peronti, A.L.G.B. Description of a new species of cecidomyiid (Diptera: Cecidomyiidae) predator of mealybugs (Hemiptera: Pseudococcidae) on sugarcane. Pap. Avulsos Zool. 2020, 60, e20206041. [Google Scholar] [CrossRef]
- De Pedro, L., Tormos, J., Harbi, A., Ferrara, F., Sabater-Munoz, B., Asís, J.D., & Beitia, F.J.. Acción conjunta de los parasitoides Aganaspis daci y Diachasmimorpha longicaudata para el control de la mosca mediterránea de la fruta, Ceratitis capitata: ¿Una estrategia recomendable?. Levante Agrícola 2020, 450, 5–13. Available online: https://redivia.gva.es/bitstream/handle/20.500.11939/6404/2020_de-Pedro_Acci%c3%b3n.pdf?sequence=3&isAllowed=y (accessed on 21 April 2025).
- Moser, M.; Salden, T.; Mikó, I.; Krogmann, L. Synthesis of the host associations of Ceraphronoidea (Hymenoptera): a key to illuminating a dark taxon. ISD, 2024. 8 (6), 6. [CrossRef]
- Noyes, J.S.; Valentine, E.W. Mymaridae (Insecta: Hymenoptera) – introduction, and review of genera. Fauna N. Z. 1989, 17. [Google Scholar] [CrossRef]
- Huber, J.T. Systematics, biology, and hosts of the Mymaridae and Mymarommatidae (Insecta: Hymenoptera):1758-1984. Entomography 1986, 4, 185–243. Available online: https://www.researchgate.net/profile/John-Huber-2/publication/278016087_Premiere_mention_en_Suisse_de_la_famille_Mymarommatidae_Hymenoptera/links/5745abb108ae9f741b42d971/Premiere-mention-en-Suisse-de-la-famille-Mymarommatidae-Hymenoptera.pdf (accessed on 23 April 2025).
- Lotfalizadeh, H.; Iranpoor, A.; Mohammadi-Khoramabadi, A. First reports of temporally soil-dwelling Chalcidoidea (Hymenoptera). Biharean Biologist. 2019. 13, 89-93. Available online: http://biozoojournals.ro/bihbiol/index.html (accessed on 21 April 2025).
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecology letters. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Sorribas, J.; González, S.; Domínguez-Gento, A.; Vercher, R. Abundance, movements and biodiversity of flying predatory insects in crop and non-crop agroecosystems. Agron. Sustain. Dev. 2016, 36, 1–9. [Google Scholar] [CrossRef]
- Sommaggio, D.; Peretti, E.; Burgio, G. The effect of cover plants management on soil invertebrate fauna in vineyard in Northern Italy. BioControl 2018, 63, 795–806. [Google Scholar] [CrossRef]
- Navarro-Campos, C.; Pekas, A.; Aguilar, A.; Garcia-Marí, F. Factors influencing citrus fruit scarring caused by Pezothrips kellyanus. J Pest Sci. 2013, 86, 459–467. [Google Scholar] [CrossRef]
- Parr, M.; Grossman, J.M.; Reberg-Horton, S.C.; Brinton, C.; Crozier, C. Nitrogen delivery from legume cover crops in no-till organic corn production. Agron. J. 2011. 103, 1578-1590. [CrossRef]
- Pérez-Rodríguez, J.: Calvo, J.; Urbaneja, A.; Tena, A. The soil mite Gaeolaelaps (Hypoaspis) aculeifer (Canestrini)(Acari: Laelapidae) as a predator of the invasive citrus mealybug Delottococcus aberiae (De Lotto)(Hemiptera: Pseudococcidae): Implications for biological control. Biol. Control 2018, 127, 64–69. [CrossRef]
- Navarro-Campos, C.; Pekas, A.; Moraza, M.L.; Aguilar, A.; Garcia-Marí, F. Soil-dwelling predatory mites in citrus: Their potential as natural enemies of thrips with special reference to Pezothrips kellyanus (Thysanoptera: Thripidae). Biol. Control 2012, 63, 201–209. [Google Scholar] [CrossRef]
- Wilson, M.J.; Gaugler. R. Factors limiting short-term persistence of entomopathogenic nematodes. J. Appl. Entomol. 2004, 128, 250–253. [CrossRef]
- Le Vieux, P.D.; Malan, A.P. Prospects for using entomopathogenic nematodes to control the vine mealybug, Planococcus ficus, in South African vineyards. SAJEV. 2015, 36, 59–70. Available online: https://www.scielo.org.za/pdf/sajev/v36n1/12.pdf (accessed on 21 April 2025). [CrossRef]
- Khan, S., Guo, L., Maimaiti, Y., Mijit, M., & Qiu, D. (2012). Entomopathogenic fungi as microbial biocontrol agent. Mol. Plant Breed. 2012, 3, 63–79. Available online: http://mpb.sophiapublisher.com (accessed on 21 April 2025).
- Meyling, N.V.; Eilenberg, J. Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: potential for conservation biological control. Biol. Control. 2007, 43, 145–155. [Google Scholar] [CrossRef]
- Amutha, M.; Gulsar Banu, J. Variation in Mycosis of Entomopathogenic Fungi on Mealybug, Paracoccus marginatus (Homoptera: Pseudococcidae). Proceedings of the National Academy of Sciences, India Section B: Biological Sciences. 2015, 87, 343–349. [Google Scholar] [CrossRef]
- Mathulwe, L.L., Malan, A.P., & Stokwe, N.F. Laboratory screening of entomopathogenic fungi and nematodes for pathogenicity against the obscure mealybug, Pseudococcus viburni (Hemiptera: Pseudococcidae). Biocontrol Sci. Technol. 2021. 32, 397–417. [CrossRef]
- Jeon, I.; Kim, J.S. Soil treatment with Beauveria and Metarhizium to control fall armyworm, Spodoptera frugiperda, during the soil-dwelling stage. J. Asia. Pac. Entomol. 2024, 27, 102193. [Google Scholar] [CrossRef]
Figure 1.
Maps showing the geographic location of the study, some damages caused by D. aberiae, and soil and trunk sampling methodologies used to conduct this study.
Figure 1.
Maps showing the geographic location of the study, some damages caused by D. aberiae, and soil and trunk sampling methodologies used to conduct this study.

Figure 2.
Captures of D. aberiae males in pheromone traps placed in trays on the ground at different distances (0.5, 1, 1.5, and 2 meters) from the citrus tree trunks. The trees were in 2 citrus orchards (in Valencia region, eastern Spain). From July 2023 to June 2024 sampling was replicated four times fortnightly. Data are expressed as average per day (n=394) and standard error (SE). Values followed by the same letter do not differ significantly (Fisher LSD test, p< 0.05).
Figure 2.
Captures of D. aberiae males in pheromone traps placed in trays on the ground at different distances (0.5, 1, 1.5, and 2 meters) from the citrus tree trunks. The trees were in 2 citrus orchards (in Valencia region, eastern Spain). From July 2023 to June 2024 sampling was replicated four times fortnightly. Data are expressed as average per day (n=394) and standard error (SE). Values followed by the same letter do not differ significantly (Fisher LSD test, p< 0.05).
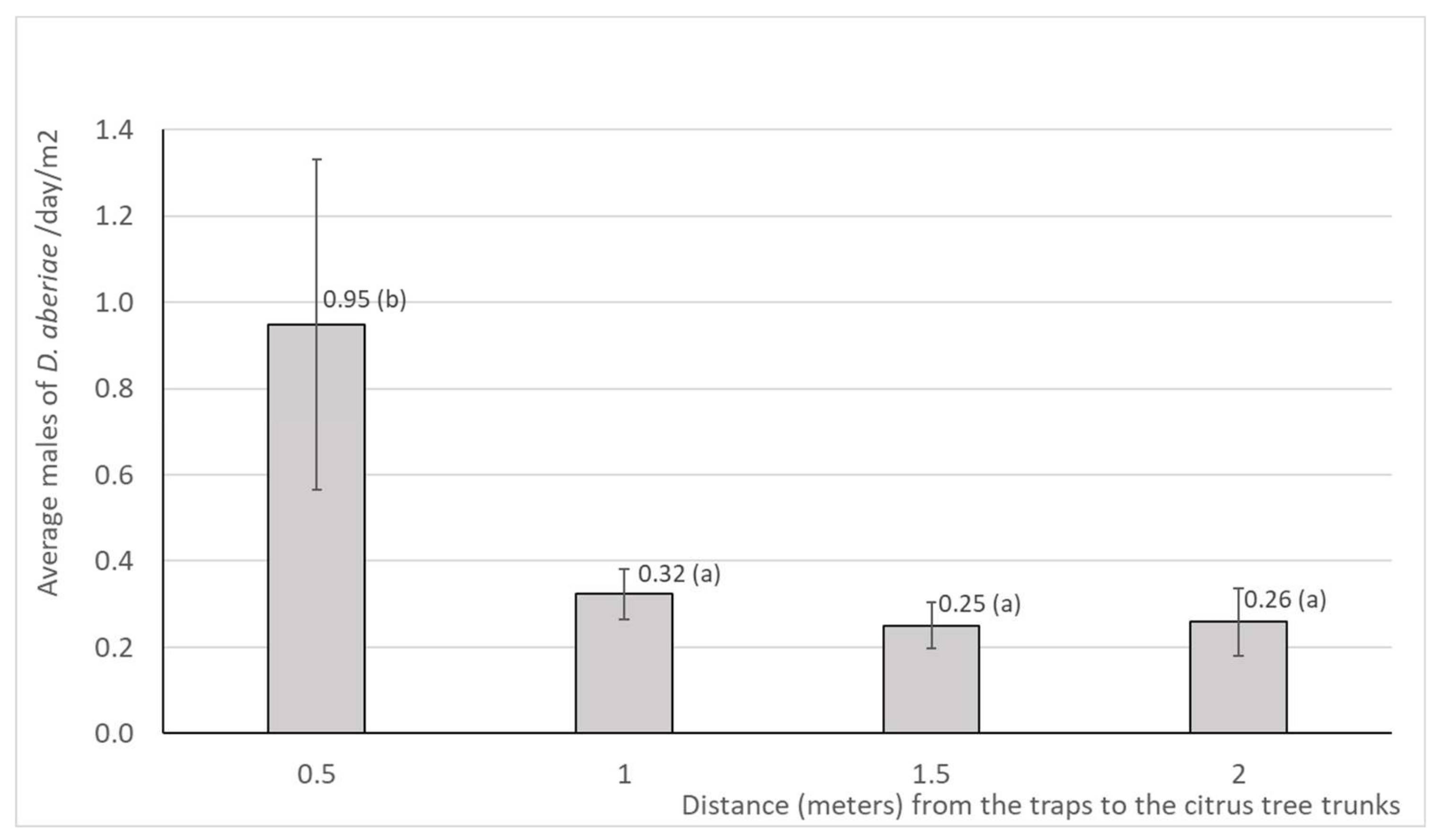
Figure 3.
Captures of D. aberiae males in pheromone traps placed on the branches (aboveground sampling), on the ground (belowground sampling), and percentages of fruits with females and immatures, in a citrus orchard in Valencia region (eastern Spain). Sampling was carried out monthly from July 2023 to Jun 2024. Data are expressed as average per day and standard error (SE). Values followed by the same letter do not differ significantly (Fisher LSD test, p< 0.05).
Figure 3.
Captures of D. aberiae males in pheromone traps placed on the branches (aboveground sampling), on the ground (belowground sampling), and percentages of fruits with females and immatures, in a citrus orchard in Valencia region (eastern Spain). Sampling was carried out monthly from July 2023 to Jun 2024. Data are expressed as average per day and standard error (SE). Values followed by the same letter do not differ significantly (Fisher LSD test, p< 0.05).
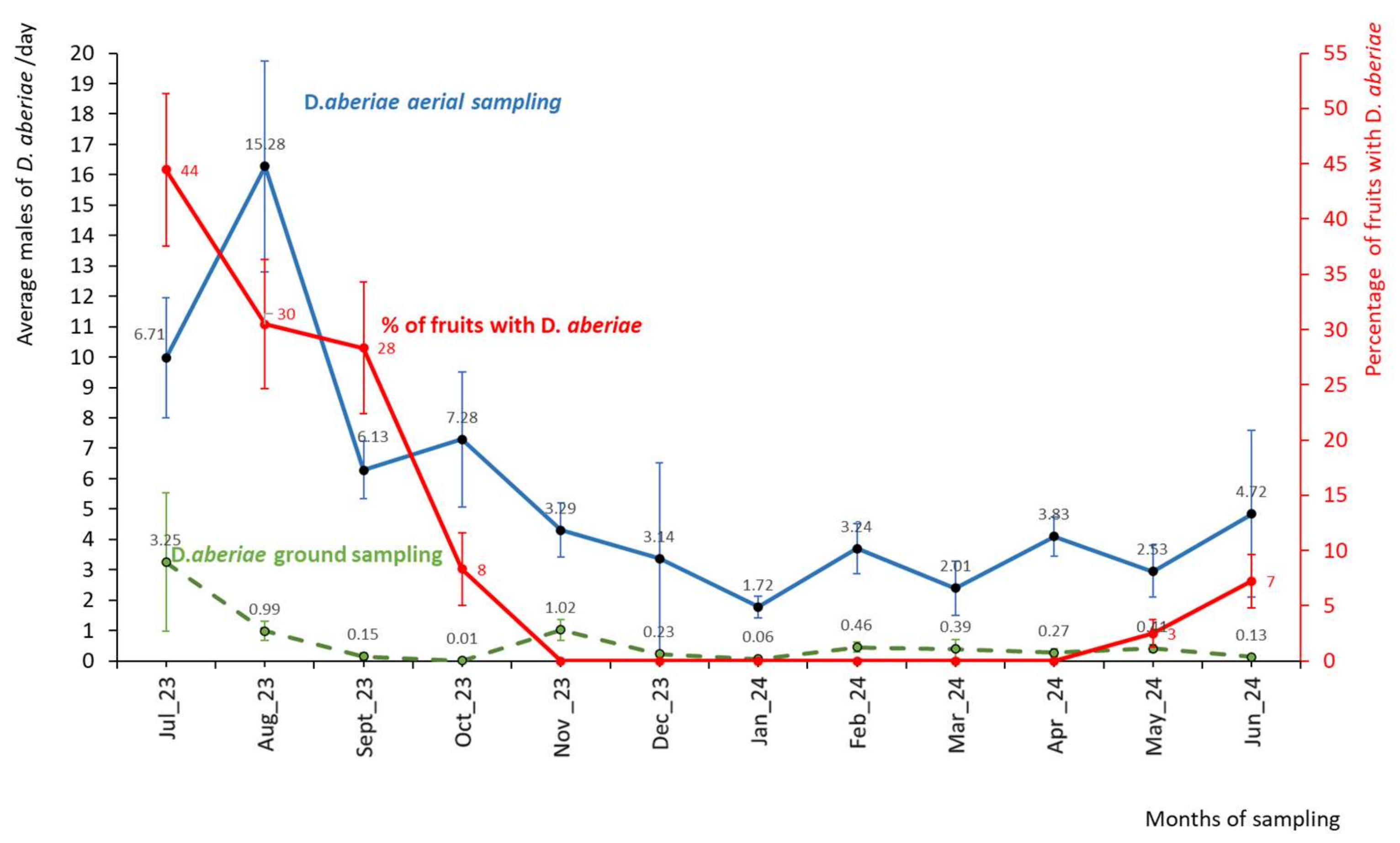
Figure 4.
Captures of D. aberiae males in pheromone traps placed on the branches (aboveground sampling) and on the ground (belowground sampling) in a citrus orchard in Valencia region (eastern Spain). Sampling was carried out monthly from July 2023 to June 2024. Data are expressed as % of monthly catches with respect to the annual total.
Figure 4.
Captures of D. aberiae males in pheromone traps placed on the branches (aboveground sampling) and on the ground (belowground sampling) in a citrus orchard in Valencia region (eastern Spain). Sampling was carried out monthly from July 2023 to June 2024. Data are expressed as % of monthly catches with respect to the annual total.
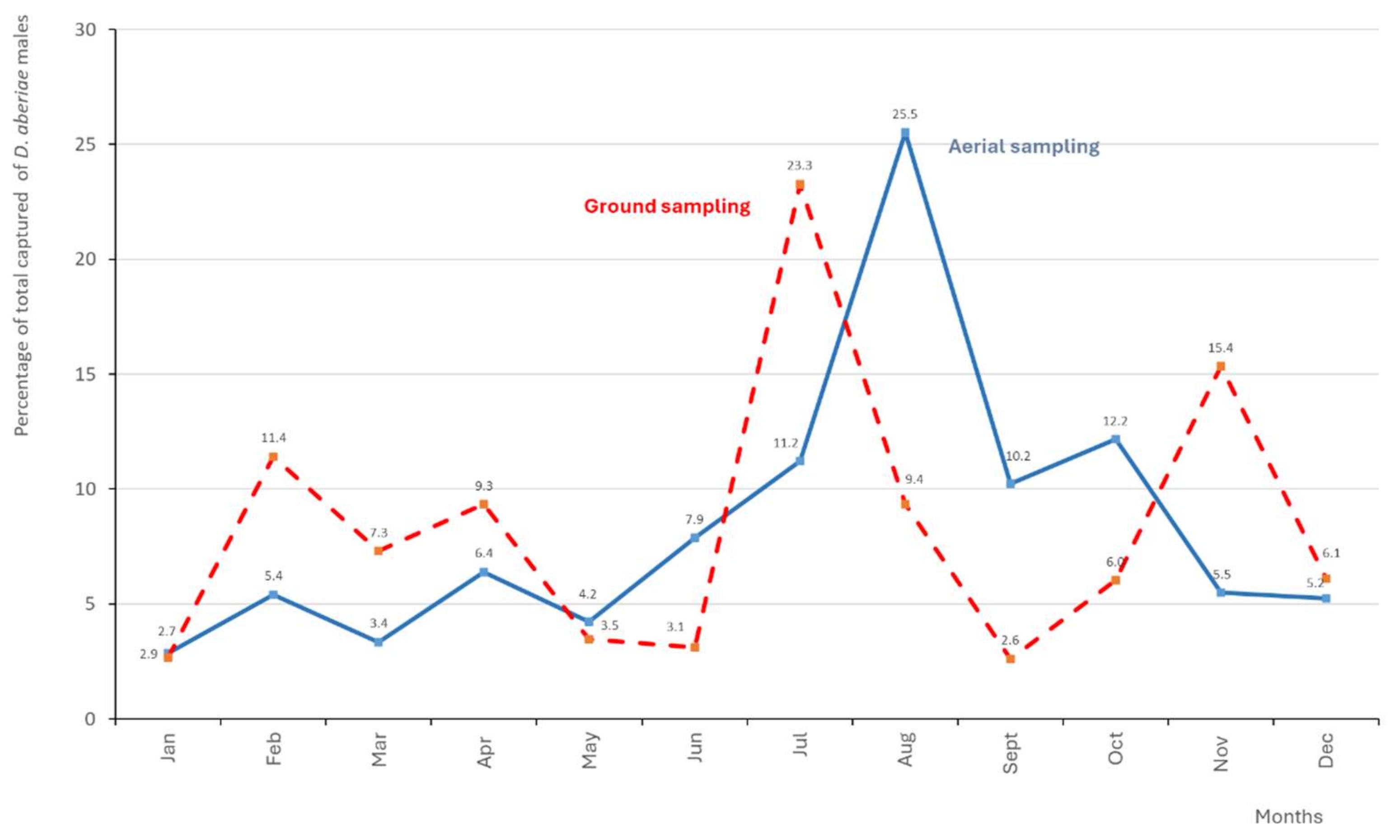

Table 1.
Percentage of damaged fruits at the end of the years 2023 and 2024 considering different levels: (0—healthy fruit; 1—slight fruit deformation; 2—fruit deformation with a clear loss of asymmetry; and 3—aberrant fruits). Marketable fruits refer to fruit with level 0 and 1. All values are reported as means ± standard errors (SE). The analysis was designed to evaluate year-to-year shifts in fruit quality and damage severity under field conditions. Normality Shapiro-Wilk test indicated that non-parametric methods such as the Wilcoxon signed-rank test were appropriate for most cases. Statistical significance was determined at the 0.05 level.
Table 1.
Percentage of damaged fruits at the end of the years 2023 and 2024 considering different levels: (0—healthy fruit; 1—slight fruit deformation; 2—fruit deformation with a clear loss of asymmetry; and 3—aberrant fruits). Marketable fruits refer to fruit with level 0 and 1. All values are reported as means ± standard errors (SE). The analysis was designed to evaluate year-to-year shifts in fruit quality and damage severity under field conditions. Normality Shapiro-Wilk test indicated that non-parametric methods such as the Wilcoxon signed-rank test were appropriate for most cases. Statistical significance was determined at the 0.05 level.
| FRUIT DAMAGE LEVEL (%) | ||||
|---|---|---|---|---|
| YEAR | 0 | 1 | 2 | 3 |
| 2023 | 31.75 ± 5.38 | 57.92 ± 4.70 | 8.92 ± 2.30 | 1.42 ± 0.61 |
| 2024 | 55.78 ± 4.21 | 40.31 ± 4.50 | 1.48 ± 0.48 | 2.42 ± 1.11 |
| Shapiro p (2023) | 0.049 | 0.623 | 0.003 | 0.000 |
| Shapiro p (2024) | 0.151 | 0.028 | 0.000 | 0.000 |
|
Wilcoxon signed-rank test |
W=22.5 p= 0.007 |
W=36.5 p= 0.003 |
W=5.0 p= 0.005 |
W=19 p= 0.68 |
| Significant (α = 0.05) | Yes | Yes | Yes | No |
Table 2.
Total number of natural enemies identified in soil traps in relation to the distance to the trunk (0.5 m, 1 m, 1.5 m y 2 m). The experiment was replicated 4 times for each distance every 14 days from July 2023 – June 2024 in a citrus orchard in Valencia (eastern Spain).
Table 2.
Total number of natural enemies identified in soil traps in relation to the distance to the trunk (0.5 m, 1 m, 1.5 m y 2 m). The experiment was replicated 4 times for each distance every 14 days from July 2023 – June 2024 in a citrus orchard in Valencia (eastern Spain).
| Arthropods | Distance to trunk (m) | |||||||
|---|---|---|---|---|---|---|---|---|
| Class and Order | Superfamily | Family | Genus | 0.5 | 1 | 1.5 | 2 | Total |
| Insecta | 46 | 42 | 36 | 49 | 173 | |||
| Diptera | 8 | 12 | 14 | 30 | 64 | |||
| Sciaroidea | Cecidomyiidae | 6 | 12 | 14 | 29 | 61 | ||
| Empidoidea | Hybotidae | Platypalpus spp. | 2 | 0 | 0 | 1 | 3 | |
| Coleoptera | Staphylinoidea | Staphylinidae | 5 | 2 | 5 | 4 | 16 | |
| Hymenoptera | 33 | 28 | 17 | 15 | 93 | |||
| Ceraphronoidea | Ceraphronidae | 4 | 5 | 3 | 3 | 15 | ||
| Megaspilidae | 0 | 0 | 1 | 0 | 1 | |||
| Chalcidoidea | Encyrtidae | 1 | 0 | 0 | 0 | 1 | ||
| Eulophidae | 1 | 0 | 0 | 0 | 1 | |||
| Mymaridae | Alaptus spp. | 4 | 5 | 3 | 5 | 17 | ||
| Camptoptera spp | 0 | 0 | 1 | 0 | 1 | |||
| Others | 3 | 1 | 1 | 0 | 5 | |||
| Others | 3 | 1 | 0 | 1 | 5 | |||
| Cynipoidea | Cynipidae | 5 | 1 | 0 | 0 | 6 | ||
| Ichneumonoidea | Ichneumonidae | 3 | 0 | 2 | 1 | 6 | ||
| unidentiffied | 0 | 10 | 3 | 2 | 15 | |||
| Platygastroidea | Scelionidae | 5 | 3 | 1 | 3 | 12 | ||
| Others | 4 | 2 | 2 | 0 | 8 | |||
| Arachnida | 91 | 88 | 72 | 150 | 401 | |||
| Araneae | 13 | 17 | 15 | 20 | 65 | |||
| Trombidiformes | Bdelloidea | Bdellidae | 66 | 57 | 54 | 124 | 301 | |
| Pseudoscorpionida | 12 | 14 | 3 | 6 | 35 | |||
Table 3.
Multifactor ANOVA (months and distance to the trunk) for the more abundant natural enemies identified in soil traps (Fisher LSD test, p< 0.05) from captures/week/trap.
Table 3.
Multifactor ANOVA (months and distance to the trunk) for the more abundant natural enemies identified in soil traps (Fisher LSD test, p< 0.05) from captures/week/trap.
| Order | Superfamily | Family | Genus | ANOVA (Fratio; Pvalue) | ||
|---|---|---|---|---|---|---|
| Moth (M) | Distance (D) | MxD | ||||
| Diptera | Sciaroidea | Cecidomyiidae | 0.94 (0.503) | 0.90 (0.441) | 1.22 (0.193) | |
| Coleoptera | Staphylinoidea | Staphylinidae | 6.05 (0.000)* | 0.47 (0.701) | 0.82 (0.758) | |
| Hymenoptera | Ceraphronoidea | Ceraphronidae | 1.53 (0.119) | 0.14 (0.937) | - | |
| Mymaridae | Alaptus | 2.28(0.011)* | 0.13 (0.944) | 0.28 (1.00) | ||
| Platygastroidea | Scelionidae | 0.98 (0.463) | 0.37 (0.774) | 0.79 (0.792) | ||
| Trombidiformes | Bdelloidea | Bdellidae | 13.97(0.000)* | 2.08 (0.103) | 1.00 (0.474) | |
| Pseudoscorpionida | 7.62 (0.000)* | 0.82 (0.482) | 0.52 (0.989) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



