
Submitted:
04 February 2025
Posted:
05 February 2025
You are already at the latest version
Abstract
Phytotelmata are aquatic microenvironments formed by the accumulation of water and organic matter in cavities of plants. These microenvironments serve as breeding sites for various species of mosquitoes, including some of epidemiological importance. Our objective was to identify the mosquito fauna in these microenvironments and to analyze variations in mosquito fauna diversity between bromeliads, tree holes and bamboo internodes in Cantareira State Park, São Paulo, Brazil, where there have been reports of yellow-fever epizootics in non-human primates and circulation of plasmodia. Collections were carried out monthly from February 2015 to April 2017. Bromeliads showed greater mosquito species richness and diversity than tree holes and bamboo internodes, as well as a very different composition. Of the 11 genera collected and 49 taxa identified, Culex (Carrolia) iridescens, Cx. ocellatus, Cx. (Microculex) imitator and Anopheles (Kerteszia) cruzii were the most abundant. The phytotelmata in CSP were found to allow a diverse mosquito fauna to develop, including An. cruzii and the sylvatic yellow fever virus vector Haemagogus leucocelaenus. The finding of these epidemiologically important species highlights the key role played by phytotelma breeding sites as places of refuge and species maintenance for these vectors in green areas close to urban centers.
Keywords:
Phytotelmata
; mosquitoes
; biodiversity
; mosquito-borne pathogens
; Cantareira State Park
1. Introduction
Ecosystems have a complex plant physiognomy because of the biodiversity of species that comprise the flora and fauna in different biomes. Among the wide range of existing habitats and microhabitats, phytotelmata (from the Greek phyton + telm = plant + pond) are aquatic microenvironments formed by the retention of water in any part of the plant body, such as leaf axils, modified leaves, flowers, holes in stems or fallen vegetative parts, including bracts, fruit peels, flowers and inflorescences [1]. The water that accumulates in phytotelmata can originate from rain or metabolic processes, as observed in bamboo, resulting in a microenvironment with physical and chemical variations that can harbor different compositions of organisms [2,3]. These microenvironments are easy to handle, allowing studies and experiments to be carried out under natural conditions [4,5], and act as biotopes for various organisms, including terrestrial arthropods that have immature aquatic stages, such as mosquitoes (Diptera: Culicidae) [6,7]. By providing refuge for native and exotic populations of mosquitoes of epidemiological importance that act as vectors of pathogens, phytotelmata put the health of those handling them at risk [8,9].
Forming a taxonomic group of considerable biodiversity with more than 3700 species described and validated to date, mosquitoes have adapted to the most diverse environments, from natural forests to highly urbanized areas [10]. Colonization in phytotelmata is selective and requires adaptations of the mosquito species that occupy them for oviposition and development of immature stages. Among the mosquito species found in these microenvironments, we highlight those of genera Haemagogus, Trichoprosopon and Sabethes associated with tree holes and bamboo internodes [11]. In addition to occupying these two phytotelmata, some species of Aedes also appear to occupy bromeliads [12]. Mosquito species of subgenera Kerteszia of Anopheles and Microculex of Culex, as well as most species in genus Wyeomyia, have greater affinity for bromeliads [13,14].
Most of the green areas in major cities have been converted to urban parks and environmental protective areas in order to preserve natural habitats and biodiversity and provide the population with leisure alternatives, space for physical activities and the opportunity to have contact with nature. An example of these urban parks, Cantareira State Park (CSP), situated in the city of São Paulo, the most populous city in the southern hemisphere [15], provides important ecological services for greater São Paulo, such as the conservation of biodiversity and protection of aquatic environments by vegetation, as well as providing water for this region from springs in the park [16]. CSP also serves as a refuge for native and exotic fauna and flora, and there have been reports of enzootic cycles of vector-borne pathogens and epizootics of vector-borne diseases such as malaria and yellow fever (YF) involving mosquitoes and non-human primates in the park [17,18]. Both YF virus and the malaria-causing species in the genus Plasmodium that circulate in the Atlantic Forest are carried mainly by phytotelma mosquitoes of genera Haemagogus and Anopheles, respectively [19,20]. Recently, CSP was affected by yellow-fever epizootics which infected primarily the howler monkey population but also resulted in human cases [18,21].
Some studies have been investigated the mosquito fauna of CSP [22,23,24,25,26,27]. A previous study conducted by our group aimed to provide an updated list of mosquito fauna in this park by employing various techniques to collect both adult and immature mosquitoes [25]. The results demonstrated that incorporating multiple mosquito capture methods led to an increase in the number of taxa collected, highlighting the importance of using diverse strategies to comprehensively sample the local mosquito fauna and explore a broader range of ecotopes [25]. Herein, we have focused the analyzes taking in account the great the importance of phytotelmata in the life cycle of mosquitoes. Therefore, the present work sought to identify the mosquito species found in phytotelmata in CSP and to analyze the variations in diversity of these insects according to the collection area and type of breeding site investigated.
2. Materials and Methods
2.1. Characterization of the Study Area: Cantareira State Park
With a total area of 7,916.52 hectares, CSP is located in the northern region of the municipality of São Paulo between latitudes 23º35'/23º45'S and 46º70'/46º48'W and extends into the municipalities of Mairiporã, Caieiras and Guarulhos [15]. Considering the different combinations of major classes of landscape elements (urbanized, forest, transition area), three collection areas were selected to reflect the environment gradient in the park (from urbanized to wild) (Figure 1).
Area 1 (Human-impacted Environment) – Bica Trail (23º27.237'S, 46º38.089'W): This location was selected because of its proximity to dwellings close to a densely urbanized region with few green spaces. The trail is situated in an area featuring a uniformly structured, medium-height-to-tall forest canopy which has experienced minimal man-made changes despite being in a zone affected by human activities and invasion by domestic animals. The phytotelmata in this area consist of epiphytic bromeliads and some bamboo clumps.
Area 2 (Transition Area) – Administration Area (23º27.062'S, 46º38.143'W): Located deeper within the park, approximately 400 meters from the interface between the forested area and the highly urbanized zone, the Administration Area has experienced significant changes. It features medium-height-to-tall trees with an uneven canopy structure, a grassy garden, two ponds, some cultivated plants and trees with bromeliads and is surrounded by native forest in the process of restoration.
Area 3 (Wild Environment) – Pinheirinho Trail (23º24.624'S, 46º37.205'W): This is a forested area with tall trees in an advanced stage of regeneration and an uneven canopy structure. The area has not undergone many changes, and epiphytic bromeliads, tree holes and bamboo internodes are still found in abundance. There is also a residential complex and country houses.
2.2. Mosquito Collection
Fieldwork was performed monthly from February 2015 to April 2017. To collect immatures in the phytotelmata (soil bromeliads, epiphytic bromeliads, exposed bamboo internodes, bamboo internodes with lateral holes and tree holes), the water was removed with manual suction pumps [26] and transferred to plastic containers from which Culicidae larvae were collected with the aid of Pasteur-type plastic pipettes. These immatures were transported to the Entomology Laboratory at the School of Public Health, University of São Paulo (LESP/FSP/USP), where they were kept until the emergence of the adults for morphological identification.
2.3. Identification and Cataloging
Species identification and cataloging was carried out at the LESP/FSP/USP. Species were identified morphologically with the taxonomic keys of Lane [27], Galindo et al. [28], Corrêa and Ramalho [29], Arnell [30], Sirivanakarn [31], Consoli and Lourenço-de-Oliveira [32] and Forattini [33]. The abbreviations for genera and subgenera followed the standardization proposed by Reinert [34].
2.4. Statistical Analysis
Richness and diversity were compared between the study areas and between the different types of phytotelma breeding sites (bamboo, bromeliads and tree holes) with the aid of rarefaction and extrapolation curves. To calculate the richness and diversity indices and construct the corresponding curves, the models proposed by Chao et al. [35] based on Hill's numbers [36] were used. The diversity index used was Simpson’s reciprocal diversity index (1/D), and 95% confidence intervals for the estimates were calculated by the bootstrap method with 100 replicates. To reduce the effect of differences in sampling effort (the number of phytotelma breeding sites were different in the three areas), the rarefaction/extrapolation curves were based on the number of individuals rather than samples. To estimate and compare species richness between areas and between breeding-site types, the curves were extrapolated up to the number of individuals observed in the area or breeding-site type with the largest number of individuals, respectively. Similarly, to estimate Simpson’s reciprocal diversity index, the rarefaction curves were used to compare the different collection areas and breeding-site types. In this case, the curves were estimated up to the number of individuals observed in the collection area or breeding-site type with the lowest mosquito abundance.
To calculate the variation in species composition, two strategies were used. The first was to measure species dissimilarity between pairs of collection areas and breeding-site types using Sorensen's qualitative dissimilarity index. The second strategy was to measure the total dissimilarity in mosquito composition and partition this dissimilarity into nestedness (species loss or gain) and turnover (species replacement) [37]. Nestedness occurs when the species composition of an area is formed by a subset of the species composition of another area with greater richness, reflecting a non-random process of species loss or gain. Turnover occurs when species are replaced between environments, reflecting random processes or local characteristics that favor the colonization, permanence or exclusion of certain species in each area. These two properties of beta diversity can be measured from the partitioning of Sorensen's total dissimilarity index [38]. The analyses were performed in R [38] with the iNEXT package [39].
3. Results
A total of 3,124 immature Culicidae specimens belonging to 49 taxa distributed in 11 genera (Aedes, Anopheles, Culex, Haemagogus, Lutzia, Sabethes, Toxorhynchites, Trichoprosopon, Shannoniana, Runchomyia and Wyeomyia) (Table S1) were collected. Of these, the genus Culex (2,048 specimens, 65.56% of the specimens collected) was the most representative, comprising 22 taxa (corresponding to 44.90% of the taxa sampled) in 4 subgenera, of which the subgenus Microculex was the richest (14 taxa), followed by genera Wyeomyia (11 taxa) and Toxorhynchites (7 taxa).
Among the most abundant species collected, we can highlight Cx. (Car.) iridescens (547), Cx. ocellatus (308), Cx. (Mcx.) imitator (301), An. (Ker.) cruzii (257), Cx. (Mcx.) pleuristriatus (191), Wy. (Pho.) theobaldi (204), Cx. (Mcx.) worontzowi (155), Sh. fluviatile (145), Cx. (Mcx.) albipes (124), Wy. (Pho.) davisi (123) and Cx. (Mcx.) sp. (116), which together account for about 79.10% of the total number of specimens collected (Table S1).
The following species occupied all three environments explored: An. (Ker.) cruzii, Cx. (Mcx.) albipes, Cx. (Mcx.) imitator, Cx. (Mcx.) pleuristriatus, Cx. (Mcx.) worontzowi, Cx. ocellatus, Wy. (Pho.) davisi and Wy. (Pho.) theobaldi. Only Cx. ocellatus was found in all three types of phytotelma explored. Also noteworthy is the occurrence of species of epidemiological importance, such as Aedes aegypti, Ae. albopictus, An. cruzii and Hg. leucocelaenus.
When the temporal distribution of these specimens by collection date was analyzed (Table S2), it was observed that some species, such as An. cruzii, Cx. iridescens, Cx. imitator, Cx. worontzowi, Cx. ocellatus, Wy. davisi and Wy. theobaldi, were quite frequent in terms of the number of times they were collected during the study even though they were not among the most abundant species, while others were infrequent or extremely infrequent.
Of the areas investigated, Pinheirinho Trail yielded the highest number of specimens collected (2,050) and the highest number of taxa (42), followed by the Administration Area (844 specimens and 28 taxa). The richness estimate suggests that with an increased sampling effort, more species could have been collected in the Administration Area and Pinheirinho Trail. Simpson’s reciprocal diversity index varied little between the environments and was higher in the Administration Area and lower in the Bica Trail, but with overlapping confidence intervals (Table 1 and Figure 2).
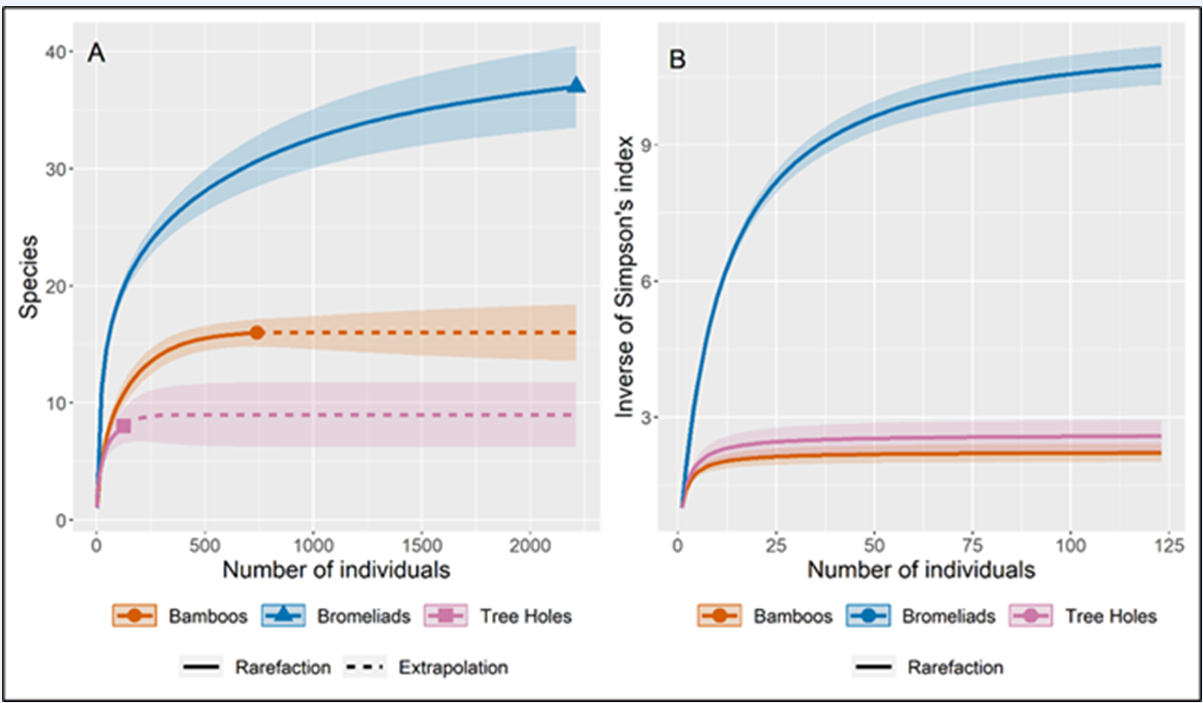
Our results show that bromeliads had the highest numbers of both specimens and species, followed by bamboo internodes. The richness estimates suggest that with an increase in sampling effort more species could have been collected in bromeliads and tree holes. Simpson's reciprocal diversity index varied greatly between the different types of breeding sites and was high for bromeliads and relatively low for bamboo and tree holes (Table 2 and Figure 3).
Dissimilarity in species composition between collection areas had an intermediate value (0.53), while the corresponding figure between breeding-site types was relatively high (0.79) (Table S3). The greatest dissimilarity between locations was between the Bica Trail and the other areas (> 0.5) (Table S4), while the dissimilarity between breeding-site types was quite high between bromeliads and tree holes (0.82) and bromeliads and bamboo (0.77) (Table S5). By partitioning the dissimilarity between nestedness and turnover, it was observed that whereas the variation between study areas had a similar contribution from each of these processes, the variation between breeding-site type was mainly due to species turnover (Table S3).
4. Discussion
Phytotelmata are excellent breeding sites for mosquitoes, which exhibit notorious plasticity in their colonizing habits and coexistence with other animal groups in these microenvironments [6,7,40]. This is in agreement with our results, which reveal a high diversity of Culicidae breeding in these natural ‘containers’. Among the mosquito species found in the phytotelmata in CSP were some of major epidemiological importance, such as Hg. leucocelaenus, An. cruzii and Ae. aegypti. Our analyses show that the composition of species in these microhabitats varies more between the different types of breeding sites than between the environments surveyed. Bromeliad breeding sites contributed the most to species richness and exhibited significantly higher diversity indices than bamboo and tree holes. Additionally, species composition in bromeliads differed markedly from that of tree holes and bamboo internodes. Although the Simpson’s reciprocal diversity index did not vary significantly between the study areas, the collection area with the highest species richness was the most preserved of the three, and the area with the lowest number of species was the most urbanized, corroborating the results of previous studies [25,41,42].
The largest numbers of species found were in the genera Culex (mainly in the subgenus Microculex) and Wyeomyia, which are commonly recorded in surveys of mosquito fauna in bromeliads, tree holes and bamboo internodes, supporting the findings of previously published works [40,43,44,45]. Culex is the genus with the largest number of species described to date; 819 species distributed in 27 subgenera have been identified [10], partly explaining the species richness for this genus observed in the present study. The genus is cosmopolitan and generally colonizes permanent underground water bodies. However, many Culex species colonize holes in rocks, holes made by crabs, phytotelmata or even artificial containers [10,46].
The presence of Cx. (Car.) iridescens, Cx. ocellatus, Cx. (Mcx.) imitator, Cx. (Mcx.) pleuristriatus and Cx. (Mcx.) albipes among the species collected in this work, which was conducted in an area recognized as one of the largest urban forests in the world, can be attributed to the fact that these mosquito species are highly adapted to wild environments [47,48], although certain species in the Microculex subgenus are also found in environments where vegetation loss is significant [48].
The finding of species of subgenus Carrollia is not surprising because this subgenus includes 18 wild species found in Neotropical forests in southeastern Mexico, Brazil and northeastern Argentina which are considered to have high ecological plasticity and to be associated with several breeding sites, including broken and cut bamboo [49,50,51].
Culex ocellatus and Culex (Microculex) species recorded here are highly dependent on bromeliads for larval habitat [9,52,53]. The former is abundant in bromeliads in both floodplain areas and mountains in the Atlantic Forest in less sunny areas [9,44,54]. In the present study, Cx. ocellatus was observed in all three environments and in all the phytotelmata explored. Although species of subgenus Microculex are usually found in preserved environments [43], some species of this subgenus collected in this study, such as Cx. (Mcx.) pleuristriatus and Cx. (Mcx.) albipes, exhibit ecological plasticity and can use artificial breeding sites in urban and periurban areas [40,48].
The high number of specimens of An. (Ker.) cruzii collected in the present study can be attributed to the fact that epiphytic bromeliads were the most abundant phytotelma in the areas surveyed. These plants serve as natural breeding sites for this species of mosquito, which is involved in the transmission of the etiological agents of human and simian malaria in the South and Southeast regions of Brazil, where bromeliaceae are abundant, especially in primary forests [55,56,57]. In contrast, An. (Ker.) bellator was rare, and only one specimen was found during the entire collection period. The finding of An. cruzii in the three areas explored can be explained by the fact that subgenus Kerteszia is considered a bioindicator of preserved environments [58]. However, members of this subgenus have highly anthropophilic behavior, and the finding of these mosquitoes in human households has long been reported [59], which explains their occurrence in the Bica Trail, an environment in close proximity to human dwellings.
Immature stages of Wy. (Pho.) davisi and Wy. (Pho.) theobaldi are commonly found in bromeliads, and adults are usually found in humid forests close to larval breeding sites. Adult females have been observed feeding on humans as well as other animals and were also collected in traps using birds as bait [60,61]. Although they bite humans, there are no reports of these species participating in viral transmission cycles. With the exception of Wy. (Pho.) trinidadensis, all the other species of subgenus Phoniomyia occur in Brazil, and only six species of this group have been observed outside Brazil [10].
Our finding of species of Cx. (Car.) and Cx. (Mcx.), Wyeomyia, Shannoniana and Toxorhynchites attests to the good state of preservation of the environments explored in CSP. These Culicidae are considered to belong to the natural mosquito fauna in the state of São Paulo and are better adapted to wild environments [40,62,63].
Immature stages of Ae. aegypti were collected in the three types of phytotelmata, while specimens of Ae. albopictus were collected only in bromeliads and bamboo and in smaller numbers. Aedes aegypti is considered highly anthropophilic and synanthropic, and is very dependent on artificial containers for breeding [64]. Aedes albopictus, in turn, is more associated with wild environments and occupies different forest habitats, but also rural and/or periurban human-impacted environments, where it has a tendency to explore peridomestic environments during the day [65]. While immature forms of Ae. aegypti and Ae. albopictus have long been recorded in bromeliads, in particular in environments changed as a result of human activities [54,66], they also occur in preserved environments but always in low density in relation to other Culicidae species, such as Culex belonging to the Microculex and Ocellatus group and species of Phoniomyia of Wyeomyia that appear to be closely associated with bromeliads in the Neotropics [54]. Thus, it seems that the interspecific competition with mosquitoes specialized in bromeliads makes these habitats less favorable to generalist species that use artificial breeding sites such as Ae. albopictus and Ae. aegypti, corroborating the results of previous studies [54,67,68].
The finding of Hg. leucocealeanus in CSP corroborates the results reported by Mucci et al. [24] and Wilk-da-Silva et al. [69,70]. This species typically colonizes tree holes, although it is also found in artificial breeding sites [69,70,71,72,73].
Our results show fewer taxa in tree holes and bamboo than in bromeliads. Tree holes and bamboo are among the oldest and most specialized breeding sites for mosquito larvae [14]. They are also the primary breeding sites for Aedes mosquito species such as Ae. aegypti in Africa, Ae. albopictus in Asia and Ae. serratus, all of which are implicated in the transmission of the YF virus [73,74,75,76], as well as other important arboviruses such as dengue virus (DENV), Chikungunya virus (CHIKV) and Zika virus (ZIKV) [78].
Bamboo is used for landscaping purposes in parks but also occurs naturally, and one of the largest native bamboo forests on the planet is in the southern Amazon [79]. Their internodes can be used as breeding sites when the stem breaks transversely and the internode is exposed to rainwater, making it a suitable habitat for oviposition and development of immature mosquitoes [14]. In addition, wild insects such as some coleoptera can drill laterally into the stems of these plants, allowing rainwater and physiological water to accumulate in the internodes and become potential habitats for mosquito larvae [3,14,80]. Mosquito species found in bamboo culms are considered wild and develop their own strategies to exploit these specialized environments [14,80].
Our results show that bromeliads were the most important phytotelmata in the study, with greater richness of species collected compared with bamboo and tree holes, as they constitute a permanent source of water and micronutrients and are therefore productive breeding grounds for mosquitoes [9]. These results disagree with those reported by Muller et al. [81], who found that bamboos were richer in immature forms than other phytotelmata. In previous studies by our group carried out in fragments of the Atlantic Forest in urban parks in São Paulo, we observed lower mosquito richness in naturally perforated bamboo than in other natural breeding sites such as bromeliads and lakes [2]. Our results also corroborate the results of Bastos et al. [82], who observed greater species richness in bromeliads than in bamboos and lakes.
Of all the Bromeliacea species, 40% are found in Brazil, mostly in the Atlantic Forest, where they are considered one of the main breeding sites for wild species of Culicidae. More than 30 species of this family have been observed in water accumulated in the leaf axils of these plants [44,81,83], and in the present study we found 37 species of Bromeliacea in CSP alone. Bromeliads are also widely used for ornamentation in public and private gardens and can be observed in the natural landscapes of ecologically preserved areas as well as in residential gardens and urban parks in the municipalities within the Atlantic Forest biome [9,62].
Phytotelmata are important natural breeding sites capable of retaining water and organic matter and maintaining ecological relationships between organisms that inhabit these microenvironments. They therefore can serve as shelters and refuges for some autochthonous and exotic species of mosquito fauna which have been shown to transmit etiological agents of disease to humans and other animals. The presence of Ae. aegypti and Hg. leucocelaenus in CSP points to a permanent scenario involving a risk of spillover of the YF virus from the wild to the urban environment, whereas the occurrence of An. cruzii is closely associated with the maintenance of simian-malaria transmission within the park.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Culicidae immatures collected in phytotelmata in Cantareira State Park, São Paulo, Brazil from February 2015 to April 2017; Table S2. Temporal distribution of Culicidae immature forms collected in phytotelmata in the Cantareira State Park, São Paulo, Brazil, according to the month of collection, from February 2015 to April 2017. Table S3: Total and partitioned dissimilarity index (turnover and nestedness) for phytotelma mosquito composition for the different study areas (Administration Area, Pinheirinho Trail and Bica Trail) and different types of breeding sites (bromeliads, bamboo and tree holes); Table S4.: Dissimilarity of phytotelma Culicidae species composition in the three CSP collection areas measured with the Sorensen index; Table S5. Dissimilarity of phytotelma Culicidae species composition for the three breeding-site types measured with the Sorensen index.
Author Contributions
Conceptualization, W.C.J., A.R.M.S., R.O.C. and M.T.M.; methodology, W.C.J., A.R.M.S. and R.O.C.; formal analysis, W.C.J., A.R.M.S.; investigation, W.C.J., A.R.M.S., E.E., K.M.B.N., R.W.S. and R.O.C.; resources, M.T.M.; data curation, A.R.M.S., M.B.P., K.M.B.N., R.W.S. and R.O.C.; writing—original draft preparation, W.C.J., A.R.M.S., R.O.C. and M.T.M; writing—review and editing, W.C.J., A.R.M.S., M.B.P., K.M.B.N., R.W.S., R.O.C. and M.T.M; supervision, A.R.M.S, R.O.C. and M.T.M.; project administration, M.T.M.; funding acquisition, A.R.M.S, R.O.C., M.T.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the State of São Paulo Research Foundation (FAPESP Grants Nos. 2014/50444-5, 2014/10919-4, 2018/18751-6 and 2021/14677-9) and the National Council for Scientific and Technological Development, Brazil (CNPq 309872/2021-9).
Data Availability Statement
All data are available in the article.
Acknowledgments
We are grateful to Vladimir Arrais and Aline Taminato at the Institute of Forestry for logistics support and for granting permission to work in the park as well as to the Institute of Forestry for hosting the researchers and granting a license for the project (SMA 260108-008.481/2014). We would also like to express our gratitude to the field and laboratory teams at the Department for the Control of Endemic Diseases, São Paulo Zoonosis Control Center, and the School of Public Health, University of São Paulo: João Carlos do Nascimento, Paulo Frugoli dos Santos, Luis Milton Bonafé, Antônio Waldomiro de Oliveira, Luis Filipe Mucci, Ana Maria Ribeiro de Castro Duarte, Laércio Molinari, Gabriel Marcelino Neto, Luiz Sposito Jr, Renildo Souza Teixeira, Aristides Fernandes, Daniel Pagotto Vendrami, Gabriela Cristina de Carvalho and Amanda Alves Camargo.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Maguirre, B. Phytotelmata: Biota and community structure determination in plant-held waters. Annu. Ver. Ecol. Evol. Syst. 1971, 2, 439–464. [Google Scholar] [CrossRef]
- Medeiros-Sousa, A.R.; OLiveira-Christe, R.; Camargo, A.A.; Milani, G.M.; Urbinatti, P.R.; Natal, D.; Ceretti-Junior, W.; Marrelli, M.T. Influence of water's physical and chemical parameters on mosquito (Diptera: Culicidae) assemblages in larval habitats in urban parks of São Paulo, Brazil. Acta Trop. 2020, 205, 105394. [Google Scholar] [CrossRef] [PubMed]
- Fang, D.; Mei, T.; Röll, A.; Hölscher, D. Water transfer between bamboo culms in the period of sprouting. Front. Plant Sci. 2019, 10, 786. [Google Scholar] [CrossRef] [PubMed]
- Campos Raúl, E.; Spinelli, G.; Mogi, M. Culicidae and ceratopogonidae (diptera: nematocera) inhabiting phytotelmata in Iguazú National Park, Misiones Province, subtropical Argentina. Rev. Soc. Entomol. Argent. 2011, 70, 111–118. [Google Scholar]
- Noutcha, M.A.E.; Harry, O.T.; Isang, K.O.; Okiwelu, S.N. Arthropod communities in Phytotelmata of the Musacae, Lauraceae and Burseraceae. Public Health Res. 2018, 8, 31–34. [Google Scholar]
- Wagner, R.; Barták, M.; Borkent, A.; Courtney, G.; Goddeeris, B.; Haenni, J.; Knutson, L.; Pont, A.; Rotheray, G.E.; Roskosny, R.; Sinclair, B.; Woodley, N.; Zatwarnicki, T.; Zwick, P. Global diversity of dipteran families (Insecta Diptera) in freshwater (excluding Simulidae, Culicidae, Chironomidae, Tipulidae and Tabanidae). Hydrobiologia 2008, 595, 489–519. [Google Scholar] [CrossRef]
- Mangudo, C.; Aparicio, J.P.; Rossi, G.C.; Gleiser, R.M. Tree hole mosquito species composition and relative abundances differ between urban and adjacent forest habitats in northwestern Argentina. Bull. Entomol. Res. 2018, 108, 203–212.
- Frank, J.H., Lounibos. Insects and allies associated with bromeliads: a review. Terr. Arthropod. Rev. 2009, 1, 125–153. [Google Scholar]
- Marques, G.R.A.; ForattiniI, O.P. Culicidae in bromeliads: diversity of species by anthropic environments, coastal area of Southeastern Brazil. Rev. Saúde Pública 2008, 42, 979–85. [Google Scholar] [CrossRef]
- Harbach, R.E. Mosquito Taxonomic Inventory. 2024. https://mosquito-taxonomic-inventory.myspecies.info/valid-species-list (Access Feb 22 2024).
- Bastos, A.Q.; Mello, C.F.; Santos Silva, J.; Gil-Santana, H.R.; Silva, S.O.F.; Alencar, J. Diversity of Mosquitoes (Diptera: Culicidae) in the Bom Retiro Private Natural Heritage Reserve, Rio de Janeiro State, Brazil. J. Med. Entomol. 2022, 59, 446–453. [Google Scholar] [CrossRef]
- Lounibos 2009.
- Lourenço-de-Oliveira, R.; Heyden, R.; Silva, T.F. Alguns aspectos da ecologia dos mosquitos (Diptera, Culicidae) de uma área de barreiras (granjas Calábria), em Jacarepaguá, Rio de Janeiro: V. Criadouros. Mem. Inst. Oswaldo Cruz 1986, 81, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Zequi, J.A.; Lopes, J. Culicideofauna (Diptera) encontrada em entrenós de taquara de uma mata residual na área urbana de Londrina, Paraná, Brasil. Rev. Bras. Zool. 2001, 18, 429–438. [Google Scholar] [CrossRef]
- Governo do Estado de São Paulo; Sistema Ambiental Paulista. Plano de manejo do Parque Estadual da Cantareira. 2009. Available at: http://www3.ambiente.sp.gov.br. (Access December 06 2024).
- Rares, C.D., Brandimarte. O desafio da conservação de ambientes aquáticos e manutenção de serviços ambientais em áreas verdes urbanas: o caso do Parque Estadual da Cantareira. Ambiente Soc. 2014, 17, 111–128. [Google Scholar] [CrossRef]
- Deane, L.M.; Deane, M.P.; Neto, J.A.; Almeida, F.B. On the transmission of simian malaria in Brazil. Rev. Inst. Med.Trop. 1971, 13, 311–319. [Google Scholar]
- Cunha, M.S.; Costa, A.C.; Fernandes, N.C.; Guerra, J.M.; Santos, F.C.; Nogueira, J.S.; D’Agostino, L.G.; Komninakis, S.V.; Witkin, S.S.; Ressio, R.A.; et al. Epizootics due to Yellow Fever Virus in São Paulo State, Brazil: viral dissemination to new areas (2016–2017). Sci. Rep. 2019, 9, 5474. [Google Scholar] [CrossRef]
- Buery, J.C.; Alencar, F.E.C.; Duarte, A.M.R.; Loss, A.C.; Vicente, C.R.; Ferreira, L.M.; Fux, B.; Medeiros, M.M.; Cravo, P.; Arez, A.P.; et al. Atlantic Forest Malaria: A Review of more than 20 years of epidemiological investigation. Microorganisms 2021, 9, 132. [Google Scholar] [CrossRef]
- Abreu, F.V.S., Ribeiro; et al. Haemagogus leucocelaenus and Haemagogus janthinomys are the primary vectors in the major yellow fever outbreak in Brazil, 2016–2018. Emerg. Microbes Infect 2019, 8, 218–231. [Google Scholar] [CrossRef]
- Governo do Estado de São Paulo. Secretaria de Estado da Saúde - Centro de Vigilância Epidemiológica Prof. Alexandre Vranjac. Boletim Epidemiológico Febre Amarela. 2018. Available at: http://www.saude.sp.gov.br/resources/cvecentro-de-vigilancia-epidemiologica/areas-de-vigilancia/doencas-de-transmissaopor-vetores-ezoonoses/doc/famarela/fa18_boletim_epid_0207.pdf (Access December 20 2024).
- Deane, L.M. Monkey malaria in Brasil. A summary of studies performed in 1964–1966. Rev. Brasil. Biol. 1967, 27, 213–228. [Google Scholar]
- Montes, J. Culicidae fauna of Serra da Cantareira, São Paulo, Brazil. Rev. Saúde Pública 2005, 39, 578–584. [Google Scholar] [CrossRef]
- Mucci, L.F.; Medeiros-Sousa, A.R.; Ceretti-Júnior, W.; Fernandes, A.; Camargo, A.A.; Evangelista, E.; Oliveira-Christe, R.; Montes, J.; Teixeira, R.S.; Marrelli, M.T. Haemagogus leucocelaenus and other mosquitoes potentially associated with sylvatic yellow fever in Cantareira State Park in the São Paulo metropolitan area, Brazil. J. Am. Mosq. Control Assoc. 2016, 32, 329–332. [Google Scholar] [CrossRef]
- Ceretti-Junior, W.; Oliveira-Christe, R.; Wilk-da-Silva, R.; Mucci, L.F.; Duarte, A.M.; Fernandes, A.; Barrio-Nuevo, K.M.; Carvalho, M.P.; Marrelli, M.T.; Medeiros-Sousa, A.R. Diversity analysis and an updated list of mosquitoes (Diptera: Culicidae) found in Cantareira State Park, São Paulo, Brazil. Acta Trop. 2020, 212, 105669. [Google Scholar] [CrossRef] [PubMed]
- Lozovei, A.L.; Silva, M.A.N. Análise comparativa entre métodos alternativo e convencional para amostras de mosquitos obtidos a partir de habitats fitotélmicos (Bromeliaceae) na Floresta Atlântica, Serra do Mar, Paraná, Brasil. Rev. Bras. Zool. 1999, 16, 957–66. [Google Scholar] [CrossRef]
- Lane, J. Neotropical Culicidae. 1953. Universidade de São Paulo, São Paulo. Vol 2.
- Galindo, P.; Blanton, F.S.; Peyton, E.L. A revision of the Uranotaenia of Panama with notes on other American species of the genus (Diptera, Culicidae). Ann. Entomol. Soc. Am. 1954, 47, 107–177. [Google Scholar] [CrossRef]
- Correa, R.R., Ramalho. Revision of the Genus Phoniomyia. Folia Clin. Biol. 1956, 25, 1–176. [Google Scholar]
- Arnell, J.H. Mosquito Studies (Diptera: Culicidae). XXXIII. A revision of the Scapularis Group of Aedes (Ochlerotatus). Contr. Am. Ent. Inst. 1976, 13, 1–44. [Google Scholar]
- Sirivanakarn, S. A review of the systematics and a proposed scheme of internal classification of the New World subgenus Melanoconion of Culex (Diptera, Culicidae). 1982. Smithsonian Institution, Dept of Entomology, Washington DC.
- Consoli, R.A. Lourenço-de-Oliveira, R. Principais mosquitos de importância sanitária do Brasil. 1994. FIOCRUZ, Rio de Janeiro Ed.
- Forattini, O.P. Culicidologia Médica: Identificação, Biologia, Epidemiologia 2. 2002. Edusp, São Paulo.
- Reinert, J.F. List of abbreviations for currently valid generic-level taxa in Family Culicidae (Diptera). Eur. Mosq. Bull. 2009, 27, 68–76. [Google Scholar]
- Chao, A.; Gotelli, N.J.; Hsieh, T.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and evenness: a unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- R Development Core Team, 2020. R: A language and environment for statistical computing. R Foundation for Statistical Comput URL. https://www.r-project.org/ (accessed: Mar. 04 2024).
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Meth. Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Ceretti-Junior, W.; Oliveira Christe, R.; Rizzo, M.; Strobel, R.C.; Matos Junior, M.O.; Mello, M.H.; Fernandes, A.; Medeiros-Sousa, A.R.; de Carvalho, G.C.; Marrelli, M.T. Species composition and ecological aspects of immature mosquitoes (Diptera: Culicidae) in bromeliads in urban parks in the city of São Paulo, Brazil. J. Arthropod. Borne Dis. 2016, 10, 102–112. [Google Scholar]
- Medeiros-Sousa, A.R.; Fernandes, A.; Ceretti-Junior, W.; Wilke, A.B.B.; Marrelli, M.T. Mosquitoes in urban green spaces: using an island biogeographic approach to identify drivers of species richness and composition. Sci. Rep. 2017, 7, 17826. [Google Scholar] [CrossRef] [PubMed]
- Orta-Pineda, G.; Abella-Medrano, C.A.; Suzán, G.; Serrano-Villagrana, A.; Ojeda-Flores, R. Effects of landscape anthropization on sylvatic mosquito assemblages in a rainforest in Chiapas, Mexico. Acta Trop. 2021, 216, 105849.
- Lozovei, A.L. Microhabitats de mosquitos (Diptera, Culicidae) em internódios de taquara na mata atlântica, Paraná, Brasil [Microhabitats of mosquitoes (Diptera, Culicidae) in internodes of bamboo in the rainforest, Paraná, Brazil]. Iheringia Sér. Zool. 2001, 90, 3–13. [Google Scholar] [CrossRef]
- Cardoso, A.A.; Lourenço-de-Oliveira, R.; Codeço, C.T.; Motta, M.A. Mosquitoes in bromeliads at ground level of the Brazilian Atlantic Forest: the relationship between mosquito fauna, water volume, and plant type. Ann. Entomol. Soc. Am. 2015, 108, 449–458. [Google Scholar] [CrossRef]
- Marteis, L.S.; Natal, D.; Sallum, M.A.M.; Medeiros-Sousa, A.R.; La Corte, R. Mosquitos da Caatinga: 2. Species from periodic sampling of bromeliads and tree holes in a dry Brazilian forest. Acta Trop. 2017, 171, 114–123. [Google Scholar] [CrossRef]
- Srisuka, W.; Sulin, C.; Sommitr, W.; Rattanarithikul, R.; Aupalee, K.; Harbach, R. Mosquito (Diptera: Culicidae) diversity and community in Doi Inthanon National Park, Northern Thailand. Insects 2022, 13, 814. [Google Scholar] [CrossRef]
- Harbach, R.E. Classification within the cosmopolitan genus Culex (Diptera: Culicidae): The foundation for molecular systematics and phylogenetic research. Acta Trop. 2011, 120, 1–14. [Google Scholar] [CrossRef]
- Oliveira-Christe, R.; Medeiros-Sousa, A.R.; Fernandes, A.; Ceretti-Junior, W.; Marrelli, M.T. Distribution of Culex (Microculex) (Diptera: Culicidae) in forest cover gradients. Acta Trop. 2020, 105264. [Google Scholar] [CrossRef]
- Valencia, J.D. Mosquito studies (Diptera, Culicidae) XXXI. A revision of the subgenus Carrollia of Culex. Contrib. Am. Entomol. Inst. 1973, 9, 1–173. [Google Scholar]
- Pecor, J.E.; Jones, J.; Turell, J.; Fernández, R. Annotated checklist of the mosquito species encountered during arboviral studies in Iquitos, Peru (Diptera: Culicidae). J. Am. Mosq. Control. Assoc. 2000, 16, 210–218. [Google Scholar] [PubMed]
- Ayala-Sulca, Y.; Carrasco-Badajoz, C.; Huicho-Yanasupo, N.; Zamalloa-Vilca, C.; Arque-Chunga, W.; Ortega-Morales, A.I.; Ramírez, R.; Fernadez-Salas, I. First National Record for Culex iridescens in Peru. J. Am. Mosq. Control Assoc. 2021, 37, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.A.; Marcondes, C.B. Bromeliad-associated mosquitoes from Atlantic forest in Santa Catarina Island, southern Brazil (Diptera, Culicidae), with new records for the State of Santa Catarina. Iheringia Sér. Zool. 2006, 96, 315–319. [Google Scholar] [CrossRef]
- Marques, T.C.; Bourke, B.P.; Laporta, G.Z.; Sallum, M.A.M. Mosquito (Diptera: Culicidae) assemblages associated with Nidularium and Vriesea bromeliads in Serra do Mar, Atlantic Forest, Brazil. Parasites & vectors 5, 1–9.
- Mocellin, M.G.; Simões, T.C.; Silva-do-Nascimento, T.F.; Teixeira, M.L.F.; Lounibos, L.P.; Lourenço-de-Oliveira, R. Bromeliad-inhabiting mosquitoes in an urban botanical garden of dengue endemic Rio de Janeiro - Are bromeliads productive habitats for the invasive vectors Aedes aegypti and Aedes albopictus? Mem. Inst. Oswaldo Cruz 2009, 104, 1171–1176. [Google Scholar] [CrossRef]
- Forattini, O.P.; Kakitani, I.; Massad, E.; Marucci, D. Studies on mosquitoes (Diptera: Culicidae) and anthropic environment: 11-Biting activity and blood-seeking parity of Anopheles (Kerteszia) in South-Eastern Brazil. Rev. Saúde Pública 1996, 30, 107–114. [Google Scholar] [CrossRef]
- Branquinho, M.S.; Marrelli, M.T.; Curado, I.; Natal, D.; Barata, J.M.S.; Tubaki, R.M.; Carréri-Bruno, G.C.; Menezes, R.T.; Kloetzel, J.K. Infecção do Anopheles (Kerteszia) cruzii por Plasmodium vivax e Plasmodium vivax variante VK247 nos municípios de São Vicente e Juquitiba, São Paulo. Pan. Am. J. Public Health 1997, 2, 189–193. [Google Scholar] [CrossRef]
- Bona, A.C.; Navarro-Silva, M.A. A. Diversidade de Culicidae durante os períodos crepusculares em bioma de Floresta Atlântica e paridade de Anopheles cruzii (Diptera: Culicidae). Rev. Bras. Zool. 2008, 25, 40–48. [Google Scholar] [CrossRef]
- Dorvillé, L.F.M. Mosquitoes as bioindicators of forest degradation in southeastern Brazil, a statistical evaluation of published data in the literature. Stud. Neotrop. Fauna Environm. 1996, 31, 68–78. [Google Scholar] [CrossRef]
- Forattini, O.P.; Kakitani, I.; Santos, R.L.; Ueno, H.M.; Kobayashi, K.M. Role of Anopheles (Kerteszia) bellator as malaria vector in Southern Brazil (Diptera: Culicidae). Mem. Inst. Oswaldo Cruz 1999, 94, 715–718. [Google Scholar] [CrossRef]
- Guimarães, A.E.; Mello, R.P.D.; Lopes, C.M.; Gentile, C. Ecology of mosquitoes (Diptera: Culicidae) in areas of Serra do Mar State Park, State of São Paulo, Brazil. I-monthly frequency and climatic factors. Mem. Inst. Oswaldo Cruz 2000, 95, 01–16. [Google Scholar] [CrossRef] [PubMed]
- Paterno, U., Marcondes. Mosquitos antropofílicos de atividade matutina em Mata Atlântica, Florianópolis, SC. Rev. Saúde Pública 2004, 38, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Ceretti-Júnior, W.; Medeiros-Sousa, A.R.; Multini, L.C; Urbinatti, P.R.; Vendrami, D.P.; Natal, D.; Marques, S.; Fernandes, A.; Ogata, H.; Marrelli, M.T. Immature mosquitoes in bamboo internodes in municipal parks, city of São Paulo, Brazil. J. Am. Mosq. Control Assoc. 2014, 30, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Medeiros-Sousa, A.R., Ceretti-Júnior, W., de Carvalho, G.C., Nardi, M.S., Araujo, A.B., Vendrami, D.P., Marrelli, M.T. Diversity and abundance of mosquitoes (Diptera: Culicidae) in an urban park: Larval habitats and temporal variation. Acta Trop. 2015, 150, 200–209. [CrossRef]
- Natal, D. Bioecologia do Aedes aegypti. Biológico 2002, 64, 205–207. [Google Scholar]
- Albuquerque, C.M.R., Melo-Santos, M.A.V., Bezerra, M.A.S., Barbosa, R.M.R., Silva, D.F., Silva, E. Primeiro registro de Aedes albopictus em área de Mata Atlântica, Recife, PE, Brasil. Rev. Saúde Pública 2000, 34, 314–315. [CrossRef]
- Fouque, F., Garinci, R., Gaborit, P. Epidemiological and entomological surveillance of the co-circulation of DEN-1, DEN-2 and DEN-4 viruses in French Guiana. Trop. Med. Int. Health 2004, 9, 41–46. [CrossRef]
- Cunha, S.P., Carreira Alves, J R., Lima, M.M., Duarte, J.R., Barros, L.C., Silva, J.L., Gammaro, A.T., Monteiro Filho, O.S., Wanzeler, A.R. Presença de Aedes aegypti em Bromeliaceae e depósitos com plantas no município do Rio de Janeiro, RJ. Rev. Saúde Pública 2002, 6.
- Maciel-de-Freitas, R., Marques, W.A., Peres, R.C., Cunha, S.P., Lourenço-de-Oliveira, R. Variation in Aedes aegypti (Diptera: Culicidae) container productivity in a slum and a suburban district of Rio de Janeiro during dry and wet seasons. Mem. Inst. Oswaldo Cruz 2007, 102, 489–496. [CrossRef]
- Wilk-da-Silva, R., Mucci, L.F., Ceretti-Junior, W., Duarte, A.M.R., Marrelli, M.T., Medeiros-Sousa, A.R. Influence of landscape composition and configuration on the richness and abundance of potential sylvatic yellow fever vectors in a remnant of Atlantic Forest in the city of São Paulo, Brazil. Acta Trop. 2020, 204, 105385.
- Wilk-da-Silva, R. Wilk-da-Silva, R., Medeiros-Sousa, A.R., Mucci, L.F., Alonso, D.P., Alvarez, M.V., Ribolla, P.E.M., Marrelli, M.T. Genetic structuring of one of the main vectors of sylvatic yellow fever: Haemagogus (Conopostegus) leucocelaenus (Diptera: Culicidae). Genes 2023, 14, 1671. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J. Ecologia de mosquitos (Diptera: Culicidae) em criadouros naturais e artificiais de área rural do Norte do Estado do Paraná, Brasil. V. Coleta de larvas em recipientes artificiais instalados em mata ciliar. Rev. Saúde Pública 1997, 31, 370–377. [Google Scholar]
- Freitas-Silva, S.O., Mello, C.F., Machado, S.L., Leite, P.J., Alencar, J. Interaction of Haemagogus leucocelaenus (Diptera: Culicidae) and other mosquito vectors in a Forested Area, Rio de Janeiro, Brazil. Trop. Med. Infect Dis. 2022, 7, 94. [CrossRef] [PubMed]
- Silva, S.O., Ferreira de Mello, C., Figueiró, R., Aguiar Maia, D., Alencar, J. Distribution of the mosquito communities (Diptera: Culicidae) in oviposition traps introduced into the Atlantic Forest in the State of Rio de Janeiro, Brazil. Vector Borne Zoonotic Dis. 2018, 18, 214–221. [CrossRef]
- Crovello, T.J.; Hacker, C.S. Evolutionary strategies in life table characteristics among feral and urban strains of Aedes aegypti (L.). Evolution 1972, 26, 185–196. [Google Scholar] [CrossRef]
- Barrera, R. Competition and resistance to starvation in larvae of container-inhabiting Aedes mosquitoes. Ecol. Entomol. 1996, 21, 117–127. [Google Scholar] [CrossRef]
- Juliano, S.A. Species introduction and replacement among mosquitoes: interspecific resource competition or apparent competition? Ecology 1998, 79, 255–268. [Google Scholar] [CrossRef]
- Cardoso, J.C., Almeida, M.A.B., Santos, E., Fonseca, D.F., Sallum, M.A., Noll, C.A., Monteiro, H.A., Cruz, A.C., Carvalho, V.L., Pinto, E.V., Castro, F.C., Nunes Neto, J. P., Segura, M.N., Vasconcelos, P.F.C. Yellow fever virus in Haemagogus leucocelaenus and Aedes serratus mosquitoes, southern Brazil, 2008. Emerg. Infect. Dis. 2010, 16, 1918.
- Donalisio, M.R.; Freitas, A.R.R.; Zuben, A.P. Arboviroses emergentes no Brasil: desafios para a clínica e implicações para a saúde pública. Rev. Saúde Pública 2017, 51, 30. [Google Scholar]
- Drumond, P.M., Wiedman, G., Patrícia Maria Drumond, C.P., Wiedman, G. Bambus no Brasil: da biologia à tecnologia. 2017. Rio de Janeiro: Instituto Ciência Hoje, 2017. 655 p.
- Lozovei, A.L. Mosquitos dendrícolas (Diptera, Culicidae) em internódios de taquara da Floresta Atlântica, serra do mar e do primeiro planalto, Paraná, Brasil. Braz. Arch. Biol. Technol. 1998, 41, 501–510. [Google Scholar] [CrossRef]
- Müller, G.A., Mello, C.F., Bueno, A.S., de Alcantara Azevedo, W., Alencar, J. Little noticed, but very important: The role of breeding sites formed by bamboos in maintaining the diversity of mosquitoes (Diptera: Culicidae) in the Atlantic Forest biome. PloS one 2022, 17, e0273774.
- Bastos, A.Q., Leite, P.J., Mello, C.F., Maia, D.A., Machado, S.L., Gil-Santana, H.R., Silva, S.O.F., Santos-Mallet, J.R., Alencar, J. Bionomy of Mosquitoes in Bamboo Internodes in an Atlantic Forest Remnant of the State of Rio De Janeiro, Brazil. J. Am. Mosq. Control Assoc. 2021, 37, 208–215. [CrossRef] [PubMed]
- Givnish, T.J., Barffus, M.H.J., Van, E.B., Riina, R., Schulte, K., Horres, R., Gonsiska, P.A., Jabaily, R.S., Crayn, D. M., Smith, J.A.C., Inverno, K., Brown, G.K., Evans, T.M., Holst, B.K., Luther, H., Até, W., Zizka, G., Barry, P.E., Sytsma, K.J. Phylogeny, adaptative radiation, and historical biogeography in bromeliaceae: insights from an eight-locus plastid phylogeny. Am. J. Bothany 2011, 98, 872–895.
Figure 1.
Location of the sampled areas in Cantareira State Park; plant cover is highlighted in light green, while urban areas are highlighted in grey. The map was constructed using QGIS v3.4.12 (http://www.qgis.org). Source: Ceretti-Junior et al. [26].
Figure 1.
Location of the sampled areas in Cantareira State Park; plant cover is highlighted in light green, while urban areas are highlighted in grey. The map was constructed using QGIS v3.4.12 (http://www.qgis.org). Source: Ceretti-Junior et al. [26].
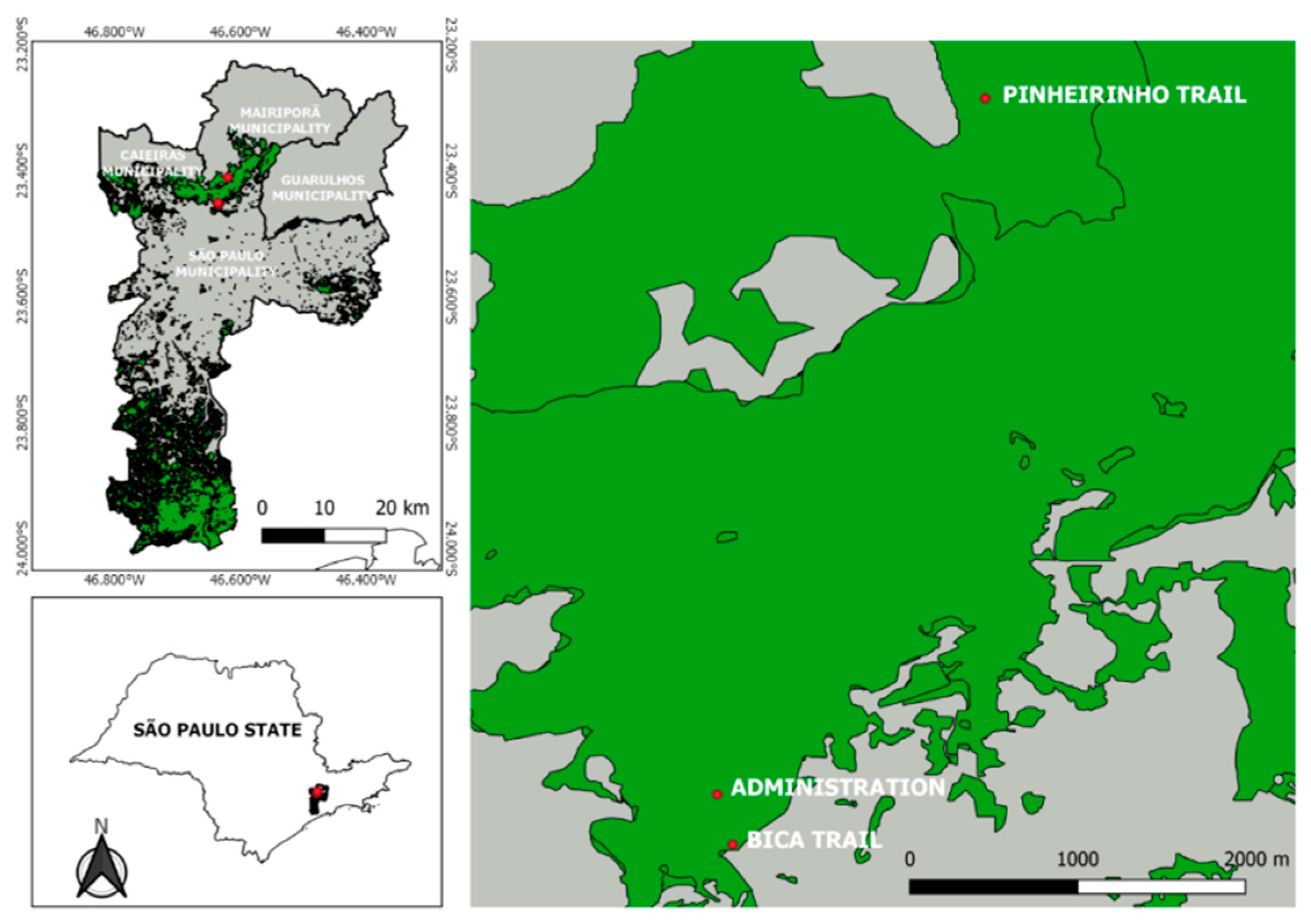
Figure 2.
Rarefaction and extrapolation curves (with 95% CI) of (A) species richness, and (B) Simpson’s reciprocal diversity index (1/D) for mosquitoes collected in phytotelmata at three Cantareira State Park collection areas. The shaded areas represent the 95% confidence interval for the mean estimate.
Figure 2.
Rarefaction and extrapolation curves (with 95% CI) of (A) species richness, and (B) Simpson’s reciprocal diversity index (1/D) for mosquitoes collected in phytotelmata at three Cantareira State Park collection areas. The shaded areas represent the 95% confidence interval for the mean estimate.
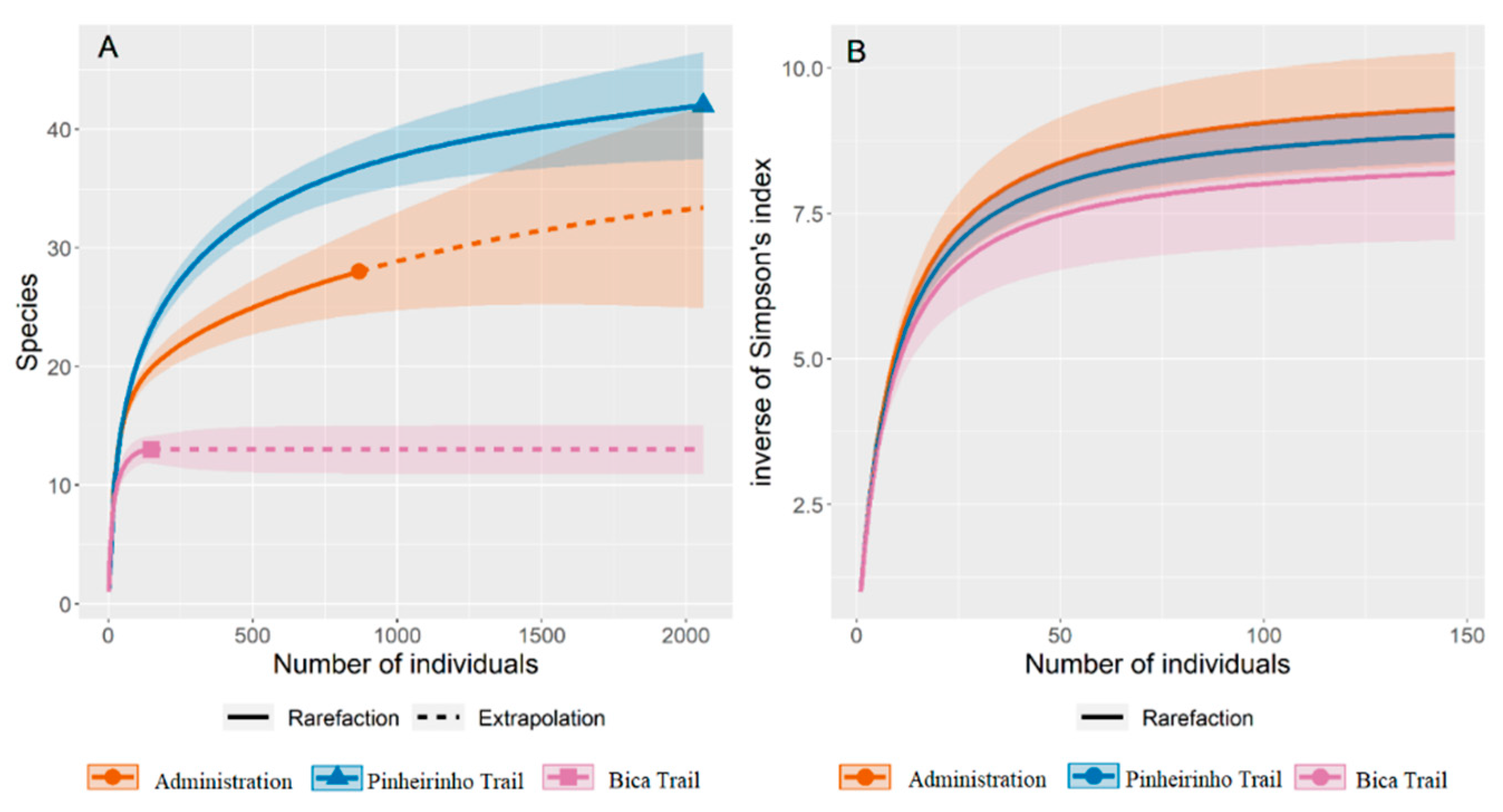
Figure 3.
Rarefaction and extrapolation curves (with 95% CI) of (A) species richness and (B) Simpson’s reciprocal diversity index (1/D) for Culicidae collected in three types of breeding sites (phytotelmata) in Cantareira State Park (CSP). The shaded areas represent the 95% confidence interval for the estimated mean.
Figure 3.
Rarefaction and extrapolation curves (with 95% CI) of (A) species richness and (B) Simpson’s reciprocal diversity index (1/D) for Culicidae collected in three types of breeding sites (phytotelmata) in Cantareira State Park (CSP). The shaded areas represent the 95% confidence interval for the estimated mean.
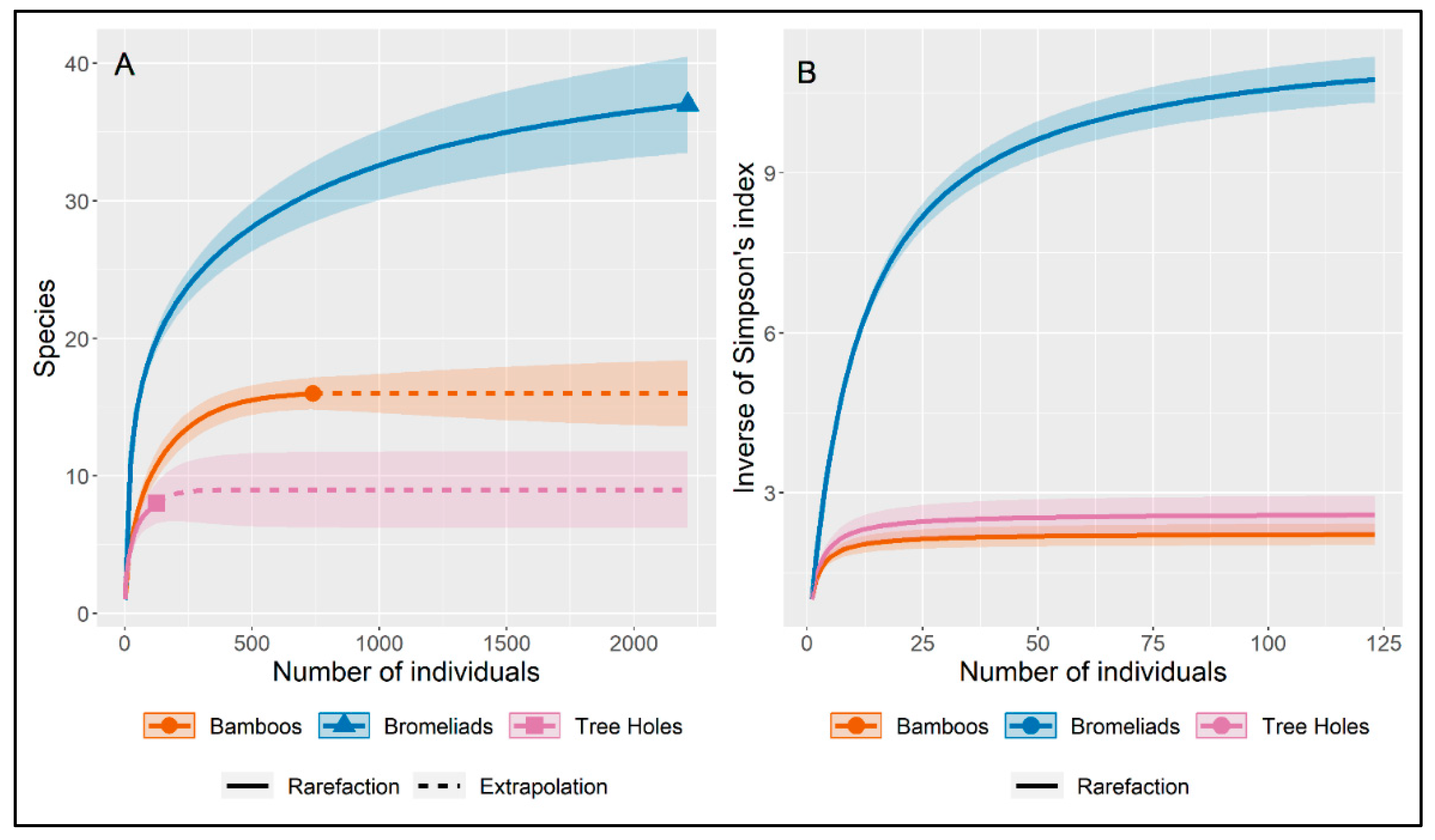

Table 1.
Observed and estimated values of species richness and Simpson’s reciprocal diversity index (1/D) for phytotelma Culicidae collected in the three study areas in Cantareira State Park (CSP), including upper and lower estimates for 95% CI.
Table 1.
Observed and estimated values of species richness and Simpson’s reciprocal diversity index (1/D) for phytotelma Culicidae collected in the three study areas in Cantareira State Park (CSP), including upper and lower estimates for 95% CI.
| Diversity index | CSP Collection Area | ||
|---|---|---|---|
| Administration Area | Pinheirinho Trail | Bica Trail | |
| Individuals | 868 | 2059 | 148 |
| Observed richness | 28 | 42 | 13 |
| Estimated richness | 37 | 60 | 13 |
| Upper Est_richness | 58 | 82 | 15 |
| Lower Est_richness | 28 | 42 | 13 |
| Observed Simpson | 9.76 | 9.31 | 8.20 |
| Estimated Simpson | 9.86 | 9.34 | 8.62 |
| Upper Est_Simpson | 10.81 | 9.92 | 9.83 |
| Lower Est_Simpson | 8.91 | 8.77 | 7.41 |
Table 2.
Observed and estimated values for species richness and Simpson’s reciprocal diversity index (1/D) for Culicidae collected in three types of breeding sites (phytotelmata) in Cantareira State Park (CSP), including upper and lower estimates for 95% CI.
Table 2.
Observed and estimated values for species richness and Simpson’s reciprocal diversity index (1/D) for Culicidae collected in three types of breeding sites (phytotelmata) in Cantareira State Park (CSP), including upper and lower estimates for 95% CI.
| Diversity index | Breeding-site type | ||
|---|---|---|---|
| Bamboo | Bromeliads | Tree Holes | |
| Individuals | 739 | 2212 | 124 |
| Observed richness | 16 | 37 | 8 |
| Estimated richness | 16 | 43 | 9 |
| Upper Est_richness | 20 | 57 | 12 |
| Lower Est_richness | 16 | 37 | 8 |
| Observed Simpson | 2.24 | 11.63 | 2.59 |
| Estimated Simpson | 2.24 | 11.69 | 2.62 |
| Upper Est_Simpson | 2.43 | 12.18 | 3.07 |
| Lower Est-Simpson | 2.06 | 11.19 | 2.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



