
Submitted:
23 December 2024
Posted:
24 December 2024
You are already at the latest version
Abstract
Plant inflorescence is not only the source of nectar and pollen to attract beneficial arthropods but also acts as the major tool for conservation of biodiversity in an agro-ecosystem. The present study aimed to quantify the ecosystem service value of arthropod predators and pollinators through enhanced plant biodiversity and mass flowering. The experiment consisted of a direct comparison between wheat monoculture crop versus wheat crop embedded with one strip each of three floral crops [singhra (Raphanus raphanistrum L.), coriander (Coriandrum sativum L.) and linseed (Linum usitatissimum L.)]. Thirty beneficial insect species including pollinators and predators belonging to 14 families from six arthropod orders were recorded in the strip planted field. Dominant pollinators' activity increased linearly with the blooming of R. raphanistrum, L. usitatissimum and C. sativum, while predator populations were directly proportional to aphid populations. The maximum foraging activity of dominant pollinators was recorded during afternoon hours (1200hr) than in the morning and evening hours. Flower initiation, duration of blooming, uniformity of flowering and flower morphometric variation directly affected the population activity of pollinators, where predator occurrences were dependent on the abundance and activity of herbivores, Rhopalosiphum padi (bird cherry-oat aphid) and Sitobion avenae (grain aphid), on wheat. Both pollinator and predator population dynamics positively correlated with ambient temperature, whereas the relative humidity and rainfall negatively impacted beneficial arthropod activities. Therefore, the selection and appropriate mix of suitable crops as intercrop, border, cover or bunker crops will provide an opportunity for enhanced habitat diversity thereby increasing pollinator and predator activities and lowering the risk of pest-induced yield losses.
Keywords:
Flowering strips
; conservation
; predators
; pollinators
; intercropping
; sustainability
1. Introduction
Wheat (Triticum aestivum L.) being the most important cereal crop is cultivated for its excellent food quality worldwide. Globally, wheat production ranks second after rice contributing 35% of total food grain production [1,2]. Owing to its nutrition and affordable price, wheat is the preferred grain food and is consumed by 35% of the world's population [3]. However, wheat crops are attacked by various herbivorous arthropods under field conditions, causing 20-37% grain yield loss [4]. Over 100 herbivorous arthropods cause damage to the crop at various growth stages and a dozen of them attain major pest status from seedling to maturity of wheat [3]. Of these, the aphids, Rhopalosiphum maidis (Fitch), Rhopalosiphum padi L., Sitobion miscanthi (Takahashi), Sitobion avenae (F.), Schizaphis graminurn (Rondani), Metopoliphiurn dirhodum (Walker) and Diuraphis noxia (Mordvilko ex Kurdjumov), are the major pests that attack during the reproductive stage of the crop in northwestern plains of India [5,6]. These aphids can cause yield losses between 20 to 30% through direct feeding [7] or transmitting viral diseases as a vector [8].
Several chemical pesticides are used frequently to control the aphids infesting wheat to maximize the grain yield [9]. However, the indiscriminate use of insecticides directly affects the soil and water quality, threatens the diversity of beneficial natural enemies such as predators, parasitoids and pollinators, contributes to the development of pest resistance, and poses risks to human health and the environment [10,11]. Consequently, the loss of major ecosystem services such as pollination, biological pest control, water and fertility regeneration and nutrient recycling follows [12]. Therefore, a more ecosystem-based pest management approach must be formulated to lower the pest pressure below the economic threshold level while enhancing the ecosystem services [13,14]. This approach includes habitat manipulation that promotes the restoration of functional agro-biodiversity in the agricultural landscape [15]. This strategy involves conserving beneficial insects by providing them with diverse vegetation, alternative prey, and food resources that enhance fecundity and longevity and withstand adverse environmental conditions for increased performance [16,17,18]. Nevertheless, the recent intensification of agriculture and urbanization has drastically impacted the diversity of predators and bee pollinators [19]. Therefore, it is a pressing need to offer suitable habitats for their conservation in the ecosystem.
Habitat manipulation is a modified concept of cultural methods wherein cultivating crops in a piece of land with high pollen and nectar are available in flowers to facilitate colonization and conservation of beneficial insect fauna, such as predators and pollinators [17,18,20,21]. The use of multi-floral strips in a cultivated field to enhance plant diversity has increased the activity of beneficial arthropod diversity and biological pest control in the agro-ecosystem [22,23]. The success of this strategy largely depends on several factors such as flowering time, synchronization of beneficial insect fauna and pests, nectar and pollen production [15,24,25], and flower shape and colour [26,27] and should be explored for an effective implementation of this strategy. These factors affect the reproductive fitness of beneficial arthropods like pollinators and predators in the ecosystem [17,28] which, in turn, would render the effectiveness of the beneficial arthropods. Therefore, habitat management is a potential area that needs to be explored to create a more favourable environment for locally occurring beneficial arthropods and as an alternative or complementary option for chemical control [29,30]. Thus, the present study aimed to determine whether the activity of aphidophagous predators and diverse pollinators enhanced in a wheat cropping system through incorporating insectary crops. Specific objectives of the study were to quantify i) the effect of flowering period and flower color on the activity of predators and pollinators, ii) the level of predator colonization and enhancement in intercropped system, and iii) the value of intercropping pattern in promoting biodiversity restoration.
2. Materials and Methods
All the growth stages of wheat were closely inspected and the presence and abundance of pests as well as pollinators and predators infesting in sole wheat crop were recorded. The arthropod pests, pollinators, and predators were also counted and recorded for their presence and abundance in the singhra (R. raphanistrum), coriander (C. sativum) and linseed (L. usitatissimum) strips intercropped within the wheat field as well as on sole wheat crop. The collection began in the last week of January, approximately four weeks after sowing each crop to the end of March. The insects visited different flowering crops grown as an intercrop between wheat and were observed and collected twice per week. The abundance of the insect pollinators and predators was recorded during morning, afternoon and evening hours for five minutes each on the randomly selected three plants in an area of 1 × 1 m2 in the intercropped flower strips as well as from wheat. In addition, the aphid population was also counted from five randomly selected plants from both treatments. The collected arthropods were categorized by insect orders including Hymenoptera, Diptera, Lepidoptera, Coleoptera, and Hemiptera. These collected pollinators, predators and pests were preserved in 70% alcohol for further study. The identity of insect species was confirmed by various insect taxonomists and using the available literature. Weather data including temperature, relative humidity, and rainfall were also recorded during the cropping season from the meteorological observatory located at Department of Agronomy, Agriculture Research Farm, School of Agriculture, LPU, Phagwara, Punjab.
Morphological Variation in Strip Flowers
Floral traits of linseed, singhra and coriander were examined by picking ten flowers for each flowering plant grown in the floral strips. Floral traits such as corolla length, diameter of flower petals, and the number of flowers per plant were recorded in the morphometric study [31]. Pollen characteristics of each flower were also observed for their shape and size under the microscope.
Statistical Analysis
All the analyses were performed using SPSS (22 version). The student t-test was used to compare the population of predators and pollinators observed in the flowering strips. Pearson’s correlation was performed to estimate the effect of weather parameters on the population of aphids, predators and pollinators that occurred in wheat as well as in the floral strip. The data obtained on the morphometrics including flower densities, corolla length and flower size of floral plants were subjected to descriptive analysis.
3. Results
3.1. Collection of Beneficial Arthropods in Floral Strips
A total of 30 beneficial insect species including pollinators and predators belonging to 13 families of the six insect orders were recorded (Table 1). In floral strips, 25 species of pollinators and five species of predators were recorded. Pollinators visiting floral strips belonged to orders Hymenoptera, Diptera, Lepidoptera and Hemiptera. In Hymenoptera, the bee species Apis mellifera L., Apis cerana F., Apis florea F., and Apis dorsata F. were abundant and occurred frequently. Seven Syrphidae species namely, Ceriana sp., Eristalis tenax L., Eristalinus tabanoides Jaenicke, Eristalinus obscuritarsus Meijere, Episyrphus balteatus De Geer,Eupeodus bucculatus and Metasyrphus corolla F. were recorded as dipteran pollinators (Table 1). Five species of aphidophagous coccinellids in the order Coleoptera were recorded preying on aphids Rhopalosiphum padi (L.) and Sitobion avenae (F.) in wheat. However, Coccinella septempunctata and Cheilomenes sexmaculata were observed with high intensity in both the sole wheat and intercropped wheat (Table 1).
3.2. Effect of Floral Strips on Predators and Pollinators
Predator densities were considerably higher in singhra (df= 24, t=2.603; p=0.016), whereas significantly lower numbers were recorded in coriander (df=24, t=3.177; p=0.004) compared to that in linseed (Figure 1). At the same time, densities of pollinators did not significantly vary between singhra and linseed where bluish colour flowers with oval and spherical pollen, respectively, appeared more preferred as indicated by greater abundance. However, white colour flower with rod shape pollen in coriander were significantly less preferred by pollinators compared to that on linseed (df=17, t=2.244; p=0.038) and singhra (P>0.05) (Figure 1). The activity of pollinators was greater during noon hours (1200hr) as compared to morning (0900hr) and evening (1700 hr) hours on the floral strips (Figure 2). The most common pollinator taxa found on floral strips were Apidae, Syrphidae and Pieridae and predator Coccinellidae (Table 1). The combination of purple and white flowers of singhra bloomed for 49 days and attracted 2895 pollinators followed by coriander (1324 individuals) with asymmetrical white flowers. The fewest numbers of pollinators were attracted to linseed (1185 individuals) having bluish flowers with oval and spherical pollen. Similarly, higher densities of predators were obtained on coriander (704 individuals) with blooming period of 33 days and lowest densities observed in linseed (310 individuals) where flowers bloomed for up to 28 days (Table 2).
3.3. Population Dynamics of Predators and Pollinators on Floral Strips
The population density of beneficial insects gradually increased with flower blooming time and significantly varied between the floral strips (Figure 3). The activity of beneficial insects in R. raphanistrum strip was observed between 3rd standard meteorological week (SMW) to 10th SMW, with peak activity during 8th SMW (4.21/m2 of Apidae, 5.18/m2 of Syrphidae and 1.18 / m2 of Pieridae) coinciding with flower blooming period (Figure 3a). The Coccinellidae population peaked at 8th SMW synchronizing with the aphid population (Figure 3a). However, the peak activity of pollinators was recorded during 9th SMW (2.36 /m2 of Apidae and 4,68/ m2 of Syrphidae) during full blooming of crops and coccinellids reached at 12th SMW (0.58/ m2) (Figure 3b). The activity of beneficial insects in coriander began with the initiation of blooming at 8th SMW and continued till crop maturity (13th SMW) with the peak density occurring at 11th SMW (Figure 3c).
3.4. Impact of Abiotic Factors on Predators and Pollinators
Temperature influenced the predator and pollinator densities in the floral strips with a strong positive correlation between temperature and arthropod densities, whereas relative humidity and rainfall correlated negatively with beneficial arthropod densities on flower strips. In coriander strip, the predator density positively correlated with both maximum (r = 0.72) and minimum (r = 0.89) temperature and the predator density improved with increase in temperature. The minimum temperature also correlated positively with pollinator densities in coriander strip. Aphid population on wheat was negatively impacted by rainfall (r = -0.83) and relative humidity (Table 3).
3.5. Interaction of Pest and Predator in Relation to Crop Growth Stages
The wheat crop reached the milk stage at 7th SMW which coincided with the occurrence of aphid population on both strip-cropped and sole wheat plots. Aphid population gradually increased from 7th SMW up to 11th SMW and then declined during 13th SMW where the crop attained the dough stage (Figure 4). Aphid densities varied between 67.8-125.3 and 85.8- 212.2 aphids per five plants in wheat with intercropped flowering strips and sole wheat crop, respectively. The average population of aphid was significantly lower in intercropped wheat than in sole wheat crop (df= 10, t=-2.262; p=0.047), (Figure 4). As expected, predator population was significantly greater in wheat with strip-cropping of singhra, linseed and coriander compared with that in the sole wheat crop (df= 10, t=2.395; p <0.005). The likely scenario for this predator enhancement in strip-cropped wheat is the abundance of flower nectar and pollen supporting predator development together with sufficient availability of aphid population (Figure 4). Numerically, the highest average predator population (22.20 ±6.57) and lowest aphid population (103.17 ±18.99) were observed in flowering strip incorporated field compared to that in sole wheat crop where predator actively suppressed the aphid population. It was also revealed that the pest and defender (predator) ratio (P:D ratio) was 5:1 in strip-cropped plots while the ratio was 10:1 in sole-planted wheat plots.
4. Discussion
4.1. Collection of Beneficial Arthropods in Floral Strips
The use of floral strips in cultivated crops has been gaining importance in conservation biological control strategies by enhancing the diversity of beneficial arthropods [15,18]. This type of habitat manipulation helps to conserve beneficial insect fauna and provide benefits if adopted after multiple years of field testing [15,22,32,33]. In this study, the deployment of floral strips enhanced the diversity and abundance of beneficial arthropods in wheat. It has been found that the activity of beneficial insects such as predators and pollinators was higher in the vicinity of floral strips compared to the sole wheat crop. The higher population of predators and pollinators in strip-cropped wheat can be attributed to the high mobility towards floral strips due to ample availability of pollen and nectar of flowering plants and move to adjacent wheat due to their proximity (predators) as well as for aphid prey on wheat ([34,35,36]. In this study, we recorded 30 species of beneficial arthropods, of which predators and pollinators were the major arthropod groups found on floral strips. Furthermore, higher densities of Apidae, Coccinellidae, Syrphidae and Pieridae were observed in all three floral strips examined in our study. These findings are in agreement with earlier studies where predator and pollinator populations were augmented in the presence of floral strips [35,37,38,39]. It is well-known that the increase in landscape heterogeneity in agricultural land promotes predators and pollinators [28,33,36]. In this study, floral strips attracted and enhanced various species of bees, seven species of syrphids and five species of aphidophagous coccinellids. Previous studies have also reported augmentation of bees, coccinellids and syrphids on the landscape planted with flowering plants [15,37,40].
4.2. Effect of Floral Strip on Predators and Pollinators
Floral recourses like nectar and pollen can be a significant source of protein and sucrose for pollinators and predators for successive reproduction [41,42]. A synchronized sowing time of insectary crops, density and type of crops (nectar and pollen source) and floral diversity are important factors to enhance predator and parasitoid activity in an agro-ecosystem [43,44,45,46,47]. Along with a phenotypic character like inflorescence height and display size, flower colour was partly responsible for attraction of pollinators and predators [42,48]. In this study, we found that the foraging activities of pollinators like Syrphidae and Apidae in floral strips (singhra and coriander) were more pronounced during afternoon hours because of the intensity of daylight such that the flower petals clusters help to recognize pollinators. Similar results were reported by previous researchers in that a predominant pollinator A. melifera foraging activity peaked after 1200 hours on Brassica napus [49,50,51]. However, the insect visitor’s activity after 1100 hours was reduced in linseed as most petals were shed by noon and thereafter insect visitors were not able to get support of petals during foraging [52].
Foraging activity of prominent pollinators depends on the duration of blooming period. Our work demonstrated that the maximum mass flowering period of up to 49 days in singhra was directly proportional to pollinator and predator activities compared to that in coriander and linseed. Floral traits of coriander plants were more attractive for several natural enemies like syrphids, contributing to a successful strategy of predator conservation [53,54,55,56]. Morphometric variations in plants influenced the density and activity of beneficial arthropods. Experts in the field suggest that the flower with a short corolla facilitates the access to nectar and pollen, whereas white flowers stimulated feeding in hoverflies [26,53,57]. Therefore, the flower strip and landscape characteristics are significant and integral components of an agroecosytem that contribute to the enhancement of ecological services and the promotion of conservation programs [58,59].
4.3. Population Dynamics of Predators and Pollinators on Floral Strips
The presence of flowers that contain nectar and pollen is a variable that influences the behavior of pollinators. This study showed that the populations of pollinators including bee and non-bee species were directly correlated with blooming time of flowers. These findings are consistent with [60] who stated that the abundance of pollinators attracted at full blooming period increased the quality and quantity of seed yield in mustard and radish [61]. The floral scent can be a significant factor for successful pollinators where the actual number of visits significantly decreased or increased over time [62]. Therefore, pollinator visits were highly impacted by chemical properties of nectar, quantity of pollen grains and floral canopy in crops [63].
Predators are important biocontrol agents for suppressing the soft-body pest insects. Our study clearly demonstrated the synchronization of predator-prey population dynamics in both monoculture wheat and intercropped wheat, except that the intercropped wheat attracted significantly greater densities. In several other studies, primary predators such as coccinellids and syrphids increased when wheat was intercropped with Brassica and pulse crops [64,65,66,67]. The increase in predator abundance in intercropped system is attributed to both the early colonization of prey due to habitat heterogeneity and availability of sufficient amount of pollen and nectar to support predators even when the prey population is insignificant [69]. As with singhra and linseed, the coriander supported a large diversity of coccinellids. It is possible that these coccinellids migrate from the wheat crop to coriander to oviposit and pupate. Also, the volatile substance released from the coriander crop may be partly responsible for increased attraction of the adult aphidophagous Coccinellidae [70]. Furthermore, the consumption of pollen may have been a contributing factor in the diversification of Coccinellidae family of predators [71].
5. Conclusions
Our results support that the existing landscape composite can be modified to diversify the agroecosystem to manage soft-bodied insect in wheat via planting of flower strips of R. raphanistrum, C. sativum and L. usitatissimum. The flower strips provided nectar and pollen in direct proximity to the predator and pollinator. Aside from pollen and nectar, additional elements such as flower color, blooming period, and weather parameters all played significant influence on pollinator and predator colonization and aggregation. Therefore, we emphasize the need for additional research into the interaction between predator/pollinators and flowering strips to further characterize habitat heterogeneity and conservation biological control across various cropping systems.
Author Contributions
Conceptualization- A.M.R and A.N.B.; methodology- A.S., A.M.R. and A.N.B.; investigation- A.S. and A.M.R.; writing-original draft preparation- A.M.R. and A.N.B..; data visualization- A.M.R, O.N., M.N.P., A.H. and E.F.A.; writing-review and editing, A.M.R., A.N.B., O.N., M.N.P; A.H., K.F.A. and E.F.A; funding acquisition: A.H., and E.F.A. All authors have read and agreed to the published version of the manuscript.
Funding
The authors would like to extend their sincere appreciation to the Researchers Supporting Project Number (RSP2025R134), King Saud University, Riyadh, Saudi Arabia.
Data Availability Statement
Data is contained within the article in the form of tables and figures.
Acknowledgments
The authors are grateful to the Administration of Lovely Professional University for providing research field site and lab facilities. Also, authors would like to extend their sincere appreciation to the Researchers Supporting Project Number (RSP2025R134), King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Miransari, M.; Smith, D. Sustainable wheat (Triticum aestivum L. ) production in saline fields: a review, Crit. Rev. Biotechnol. 2019, 39, 999–1014. [Google Scholar] [CrossRef]
- Anonymous. These are the top 10 countries that produce the most wheat. Word Economic Forum. 2022. Available online: https://www.weforum.org/agenda/2022/08/top-10-countries-produce-most-wheat/ (accessed on day month year).
- Gaur, N.; Mogalapu, S. 2018. Pests of wheat. In. Omkar (eds) Pests and their management. Springer, Singapore. [CrossRef]
- Pimentel, D.; Houser, J.; Preiss, E.; White, O.; Fang, H.; Mesnick, L.; Barsky, T.; Tariche, S.; Schreck, J.; Alpert, S. Water resources: agriculture, the environment, and society. Biomed. Sci. 1997, 47, 97–106. [Google Scholar] [CrossRef]
- Deol, G.S.; Gill, K.S.; Brar, J.S. Aphid outbreak on wheat and barley in Punjab. Newsletter Aphid Soc. India. 1987, 6, 7–9. [Google Scholar]
- Farook, U.B.; Khan, Z.H.; Ahad, I.; Maqbool, S.; Yaqoob, M.; Rafieq, I.; Sultan, N. A review on insect pest complex of wheat (Triticum aestivum L. ). J. Entomol. Zool. Studies. 2019, 7, 1292–1298. [Google Scholar]
- Kolbe, W.; Linke, W. Studies of cereal aphids; their occurrence, effect on yield in relation to density levels and their control. Ann. Appl. Biol. 1974, 77, 85–87. [Google Scholar] [CrossRef]
- Liu, S.Y.; Stoltz, R.L.; Ni, X.Z. Damage to wheat by Macrosiphum avenae (F. ) (Homoptera: Aphididae) in northwest China. J. Econ. Entomol. 1986, 79, 1688–1691. [Google Scholar]
- Tanguy, S.; Dedryver, C.A. Reduced BYDV–PAV transmission by the grain aphid in a Triticum monococcum line. European J. Plant Pathol. 2009, 123, 281–289. [Google Scholar] [CrossRef]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Foster, S.P.; Paul, V.L.; Slater, R.; Warren, A.; Denholm, I.; Field, L.M.; Williamson, M.S. A mutation (L1014F) in the voltage gated sodium channel of the grain aphid, Sitobion avenae, is associated with resistance to pyrethroid insecticides. Pest Manage. Sci. 2014, 70, 1249–1253. [Google Scholar] [CrossRef] [PubMed]
- Steffen, W.; Richardson, K.; Rockström, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; De Vries, W.; De Wit, C.A.; Folke, C. Planetary boundaries: Guiding human development on a changing planet. Science. 2015, 347, 6223. [Google Scholar] [CrossRef]
- Tittonell, P. Ecological intensification of agriculture-sustainable by nature. Curr. Opin. Environ. Sustain. 2014, 8, 53–61. [Google Scholar] [CrossRef]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; Lamichhane, J.R. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Alcalá Herrera, R.; Cotes, B. ; Agustí, N; Tasin, M. ; Porcel, M. Using flower strips to promote green lacewings to control cabbage insect pests. J. Pest Sci. 2022, 95, 669–683. [Google Scholar] [CrossRef]
- Wratten, S.; Lavandero, B.; Scarratt, S.; Vattala, D. Conservation biological control of insect pests at the landscape scale. IOBC WPRS Bull. 2003, 26, 215–220. [Google Scholar]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M. Habitat management to suppress pest populations: progress and prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Ayers, A.C.; Rehan, S.M. Supporting bees in cities: how bees are influenced by local and landscape features. Insects. 2021, 12, 128. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.X.; Zhu, P.Y.; Gurr, G.M.; Zheng, X.S.; Read, D.M.; Heong, K.L.; Xu, H.X. Mechanisms for flowering plants to benefit arthropod natural enemies of insect pests: Prospects for enhanced use in agriculture. Insect Sci. 2014, 21, 1–12. [Google Scholar] [CrossRef]
- Butters, J.; Murrell, E.; Spiesman, B.J.; Kim, T.N. Native flowering border crops attract high pollinator abundance and diversity, providing growers the opportunity to enhance pollination services. Environ. Entomol. 2022, 51, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Wratten, S.D.; Gillespie, M.; Decourtye, A.; Mader, E.; Desneux, N. Pollinator habitat enhancement: benefits to other ecosystem services. Agric. Ecosyst. Environ. 2012, 159, 112–122. [Google Scholar] [CrossRef]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; Ganser, D. , The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: a quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- Fiedler, A.K.; Landis, D.A. Plant characteristics associated with natural enemy abundance at Michigan native plants. Environ. Entomol. 2007, 36, 878–886. [Google Scholar] [CrossRef]
- Tscharntke, T.; Karp, D.S.; Chaplin-Kramer, R.; Batáry, P.; DeClerck, F.; Gratton, C.; Hunt, L.; Ives, A.; Jonsson, M.; Larsen, A.; Martin, E.A. , When natural habitat fails to enhance biological pest control – Five hypotheses. Biol. Conserv. 2016, 204, 449–458. [Google Scholar] [CrossRef]
- Vattala, H.D.; Wratten, S.D.; Phillips, C.B.; Wäckers, F.L. The influence of flower morphology and nectar quality on the longevity of a parasitoid biological control agent. Biol. Control. 2006, 39, 179–185. [Google Scholar] [CrossRef]
- Nicolson, S.W.; Thornburg, R.W. 2007. Nectar chemistry. In Nectaries and nectar (pp. 215-264). Springer, Dordrecht.
- Yan, J.; Wang, G.; Sui, Y.; Wang, M.; Zhang, L. Pollinator responses to floral colour change, nectar and scent promote reproductive fitness in Quisqualisindica (Combretaceae). Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Balmer, O.; Géneau, C.E.; Belz, E.; Weishaupt, B.; Förderer, G.; Moos, S.; Ditner, N.; Juric, I.; Luka, H. Wildflower companion plants increase pest parasitation and yield in cabbage fields: Experimental demonstration and call for caution. Biol. Control. 2014, 76, 19–27. [Google Scholar] [CrossRef]
- Nilsson, U.; Porcel, M.; Świergiel, W.; Wivstad, M. 2016. Habitat manipulation– as a pest management tool in vegetable and fruit cropping systems, with the focus on insects and mites. SLU, EPOK – Centre for Organic Food & Farming, Uppsala. https://orgprints.org/id/eprint/30032/1/biokontrollsyntes_web.
- Van der Niet, T.; Jürgens, A.; Johnson, S.D. Pollinators, floral morphology and scent chemistry in the southern African orchid genus Schizochilus. S. Afr. J. Bot. 2010, 76, 726–738. [Google Scholar] [CrossRef]
- Parajulee, M.N.; Montandon, R.; Slosser, J.E. Relay intercropping to enhance predators of the cotton aphids in Texas cotton. Int. J. Pest Manage. 1997, 43, 227–232. [Google Scholar] [CrossRef]
- Parajulee, M.N.; Slosser, J.E. Evaluation of potential relay strip crops for predator enhancement in Texas cotton. Int. J. Pest Manage. 1999, 45, 275–286. [Google Scholar] [CrossRef]
- Feltham, H.; Park, K.; Minderman, J.; Goulson, D. Experimental evidence that wildflower strips increase pollinator visits to crops. Ecol. Evol. 2015, 5, 3523–3530. [Google Scholar] [CrossRef]
- Middleton, E.G.; MacRae, I.V.; Philips, C.R. Floral plantings in large-scale commercial agroecosystems support both pollinators and arthropod predators. Insects. 2021, 12, 91. [Google Scholar] [CrossRef] [PubMed]
- Serée, L.; Chiron, F.; Valantin-Morison, M.; Barbottin, A.; Gardarin, A. Flower strips, crop management and landscape composition effects on two aphid species and their natural enemies in faba bean. Agric. Ecosyst. Environ. 2022, 331, 107902. [Google Scholar] [CrossRef]
- Kunjwal, N.; Kumar, Y.; Khan, M.S. Flower-visiting insect pollinators of Brown Mustard, Brassica juncea (L. ) Czern and Coss and their foraging behaviour under caged and open pollination. Afr. J. Agric. Res. 2014, 9, 1278–1286. [Google Scholar]
- Kamel, S.M.; Mahfouz, H.M.; Blal, E.F.A.H.; Said, M.; Mahmoud, M.F. Diversity of insect pollinators with reference to their impact on yield production of canola (Brassica napus L. ) in Ismailia, Egypt. Pesticidi i fitomedicina. 2015, 30, 161–168. [Google Scholar] [CrossRef]
- Usman, M.; Amin, F.; Sohail, K.; Shah, S.F.; Aziz, A. Incidence of different insect visitors and their relative abundance associated with coriander (Coriandrum sativum) in district Charsadda. Pure Appl. Biol. 2018, 7, 539–546. [Google Scholar] [CrossRef]
- Amy, C.; Noël, G.; Hatt, S.; Uyttenbroeck, R.; Van de Meutter, F.; Genoud, D.; Francis, F. Flower strips in wheat intercropping system: effect on pollinator abundance and diversity in Belgium. Insects. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Raderschall, C.A.; Lundin, O.; Lindström, S.A.; Bommarco, R. Annual flower strips and honeybee hive supplementation differently affect arthropod guilds and ecosystem services in a mass-flowering crop. Agric. Ecosyst. Environ. 2022, 326, 107754. [Google Scholar] [CrossRef]
- Trunschke, J.; Lunau, K.; Pyke, G.H.; Ren, Z.X.; Wang, H. Flower color evolution and the evidence of pollinator-mediated selection. Front. Plant Sci. 2021, 12, 617851. [Google Scholar] [CrossRef]
- Brennan, E.B. Agronomic aspects of strip intercropping lettuce with alyssum for biological control of aphids. Biol. Control. 2013, 65, 302–311. [Google Scholar] [CrossRef]
- Saeed, N.; Mori, N.; Battisti, A.; Ashraf, M. Effect of Brassica napus, Medicago sativa, Trifolium alexandrinum and Allium sativum strips on the population dynamics of Sitobean avenae and predators in wheat ecosystem. J. Entomol. Zool. Stud. 2016, 4, 178–182. [Google Scholar]
- Amala, U.; Shivalingaswamy, T.M. Effect of intercrops and border crops on the diversity of parasitoids and predators in agroecosystem. Egyptian J. Biol. Pest Control. 2018, 28, 1–4. [Google Scholar] [CrossRef]
- Bybee-Finley, K.; Ryan, M.R. Advancing intercropping research and practices in industrialized agricultural landscapes. Agriculture 2018, 8, 80. [Google Scholar] [CrossRef]
- Puliga, G.A.; Arlotti, D. Wheat-pea intercrop affects activity density and bio control potential of generalist predators. Ann. Appl. Biol. 2021, 1, 1–11. [Google Scholar]
- Haussler, J.; Sahlin, U.; Baey, C.; Smith, H.G.; Clough, Y. Pollinator population size and pollination ecosystem service responses to enhancing floral and nesting resources. Ecol. Evol. 2017, 7, 1898–1908. [Google Scholar] [CrossRef]
- Ali, M.; Saeed, S.; Sajjad, A.; Whittington, A. In search of the best pollinators for canola (Brassica napus L. ) production in Pakistan. Appl. Entomol. Zool. 2011, 46, 353–361. [Google Scholar] [CrossRef]
- Goswami, V.; Khan, M.S. Impact of honey bee pollination on pod set of mustard (Brassica juncea L. : Cruciferae) at Pantnagar. The Bioscan 2014, 9, 75–78. [Google Scholar]
- Akhtar, T.; Aziz, M.A.; Naeem, M.; Ahmed, M.S.; Bodlah, I. Diversity and relative abundance of pollinator fauna of canola (Brassica napus L. Var Chakwal Sarsoon) with managed Apis mellifera L. in Pothwar Region, Gujar Khan, Pakistan. Pakistan J. Zool. 2018, 50, 567–577. [Google Scholar] [CrossRef]
- Gubin, A.F. Cross pollination of fibre flax. Bee world. 1945, 26, 30–31. [Google Scholar] [CrossRef]
- Colley, M.R.; Luna, J.M. Relative attractiveness of potential beneficial insectary plants to aphidophagous hoverflies (Diptera: Syrphidae). Environ. Entomol. 2000, 29, 1054–1059. [Google Scholar] [CrossRef]
- Ambrosino, M.D.; Luna, J.M.; Jepson, P.C.; Wratten, S.D. Relative frequencies of visits to selected insectary plants by predatory hoverflies (Diptera: Syrphidae), other beneficial insects, and herbivores. Environ. Entomol. 2006, 35, 394–400. [Google Scholar] [CrossRef]
- Hogg, B.N.; Bugg, R.L.; Daane, K.M. Attractiveness of common insectary and harvestable floral resources to beneficial insects. Biol. Control. 2011, 56, 76–84. [Google Scholar] [CrossRef]
- Amorós-Jiménez, R.; Pineda, A.; Fereres, A.; Marcos-García, M.Á. Feeding preferences of the aphidophagous hoverfly Sphaerophoria rueppellii affect the performance of its offspring. Biol. Control. 2014, 59, 427–435. [Google Scholar] [CrossRef]
- Branquart, E.; Hemptinne, J.L. Selectivity in the exploitation of floral resources by hoverflies (Diptera: Syrphinae). Ecography. 2000, 23, 732–742. [Google Scholar] [CrossRef]
- Schoch, K.; Tschumi, M.; Lutter, S.; Ramseier, H.; Zingg, S. Competition and facilitation effects of semi-natural habitats drive total insect and pollinator abundance in flower strips. Front. Ecol. Evol. 2022, 10. [Google Scholar] [CrossRef]
- Barda, M.; Karamaouna, F.; Kati, V.; Perdikis, D. Do patches of flowering plants enhance insect pollinators in apple orchards? . Insects 2023, 14, 208. [Google Scholar] [CrossRef] [PubMed]
- Devi, M.; Sharma, H.K.; Thakur, R.K.; Bhardwaj, S.K.; Rana, K.; Thakur, M.; Ram, B. Diversity of insect pollinators in reference to seed set of mustard (Brassica juncea L. ). Int. J. Curr. Microbiol. App. Sci. 2017, 6, 2131–2144. [Google Scholar] [CrossRef]
- Divija, S.D.; Jayanthi, P.K.; Varun, Y.B.; Kumar, P.S.; Krishnarao, G.; Nisarga, G.S. Diversity, abundance and foraging behaviour of insect pollinators in Radish (Raphanus raphanistrum subsp. sativus L.). J. Asia-Pacific Entomol. 2022, 25, 101909. [Google Scholar] [CrossRef]
- Van Doorn, W.G. Effects of pollination on floral attraction and longevity. J. Exp. Bot. 1997, 48, 1615–1622. [Google Scholar] [CrossRef]
- Shakeel, M.; Ali, H.; Ahmad, S.; Said, F.; Khan, K.A.; Bashir, M.A.; Ali, H. Insect pollinators diversity and abundance in Eruca sativa Mill. (Arugula) and Brassica rapa L.(Field mustard) crops. Saudi J. Biol. Sci. 2019, 26, 1704–1709. [Google Scholar] [CrossRef]
- Xie, H.C.; Chen, J.L.; Cheng, D.F.; Zhou, H.B.; Sun, J.R.; Liu, Y.; Francis, F. Impact of wheat–mung bean intercropping on English grain aphid (Hemiptera: Aphididae) populations and its natural enemy. J. Econ. Entomol. 2012, 105, 854–859. [Google Scholar] [CrossRef]
- Zhou, H. 2012. Use of intercropping and infochemical releasers to control aphids in wheat (Doctoral dissertation, ULiège. GxABT-Liège Université. Gembloux Agro-Bio Tech).
- Wang, W.L.; Liu, Y.; Ji, X.L.; Wang, G.; Zhou, H.B. Effects of wheat-oilseed rape or wheat-garlic intercropping on the population dynamics of Sitobion avenae and its main natural enemies. J. Appl. Ecol. 2008, 19, 1331–1336. [Google Scholar]
- Hayat, K.; Chuhan, M.A.; Rasul, A.; Arshad, I. Intercropping of wheat and oilseed crops reduces wheat aphid, Sitobion avenae (Fabricius) (Hemiptera: Aphididae) incidences: a field study. J. Agric. Res. 2018, 56, 181–185. [Google Scholar]
- Shrestha.
- Togni, P.H.; Venzon, M.; Muniz, C.A.; Martins, E.F.; Pallini, A.; Sujii, E.R. Mechanisms underlying the innate attraction of an aphidophagous coccinellid to coriander plants: Implications for conservation biological control. Bio. Control. 2016, 92, 77–84. [Google Scholar] [CrossRef]
- Giorgi, J.A.; Vandenberg, N.J.; McHugh, J.V.; Forrester, J.A.; Ślipiński, S.A.; Miller, K.B.; Shapiro, L.R.; Whiting, M.F. The evolution of food preferences in Coccinellidae. Bio. Control. 2009, 51, 215–231. [Google Scholar] [CrossRef]
Figure 1.
Comparison of predators and pollinators within flowering strips.
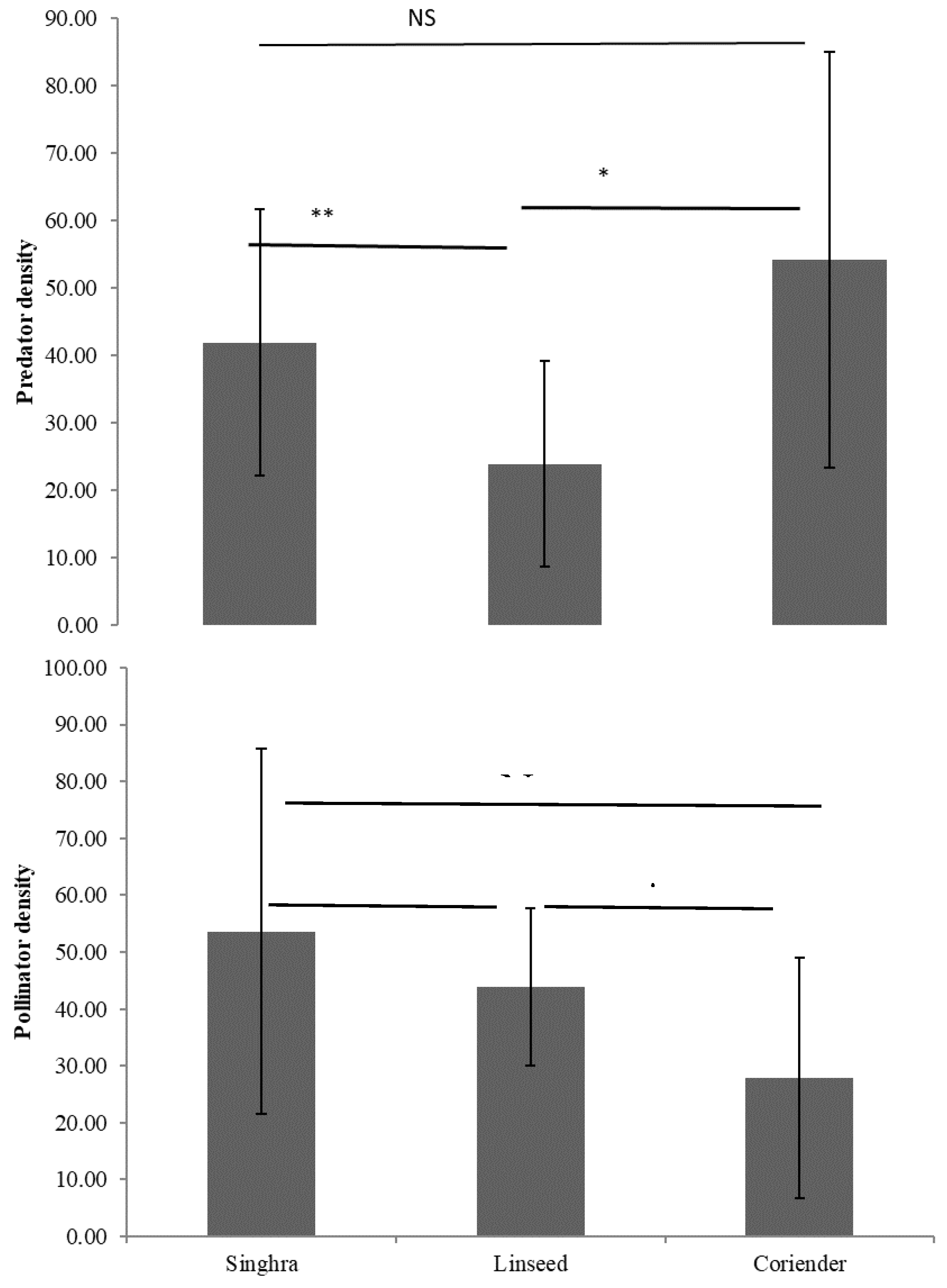
Figure 2.
Temporal patterns of diurnal pollinator activity on flowering strips.
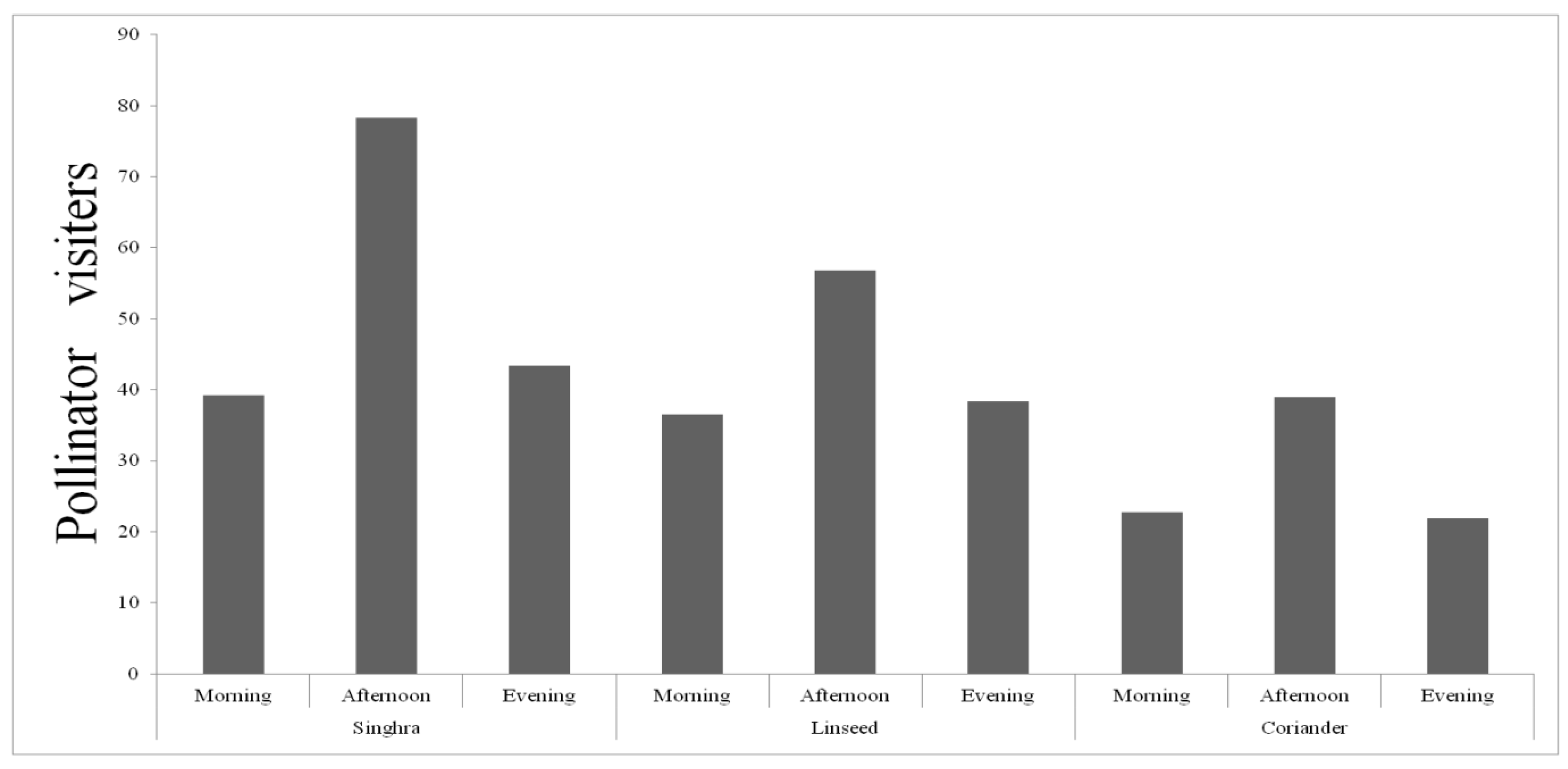
Figure 3.
Populating fluctuation of predators and pollinators in floral strips.
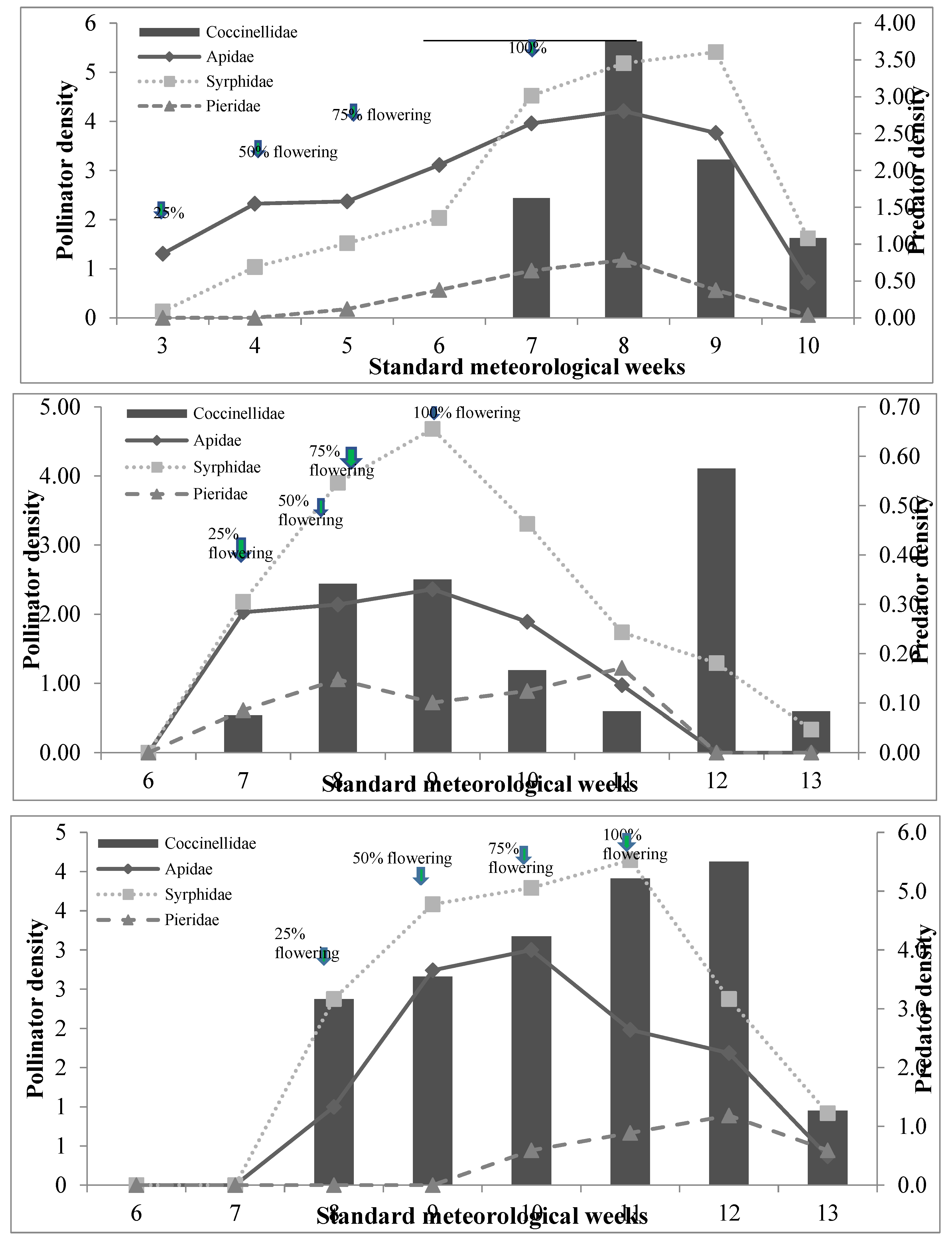
Figure 4.
Pest predator relations with critical growth stage of wheat crop.
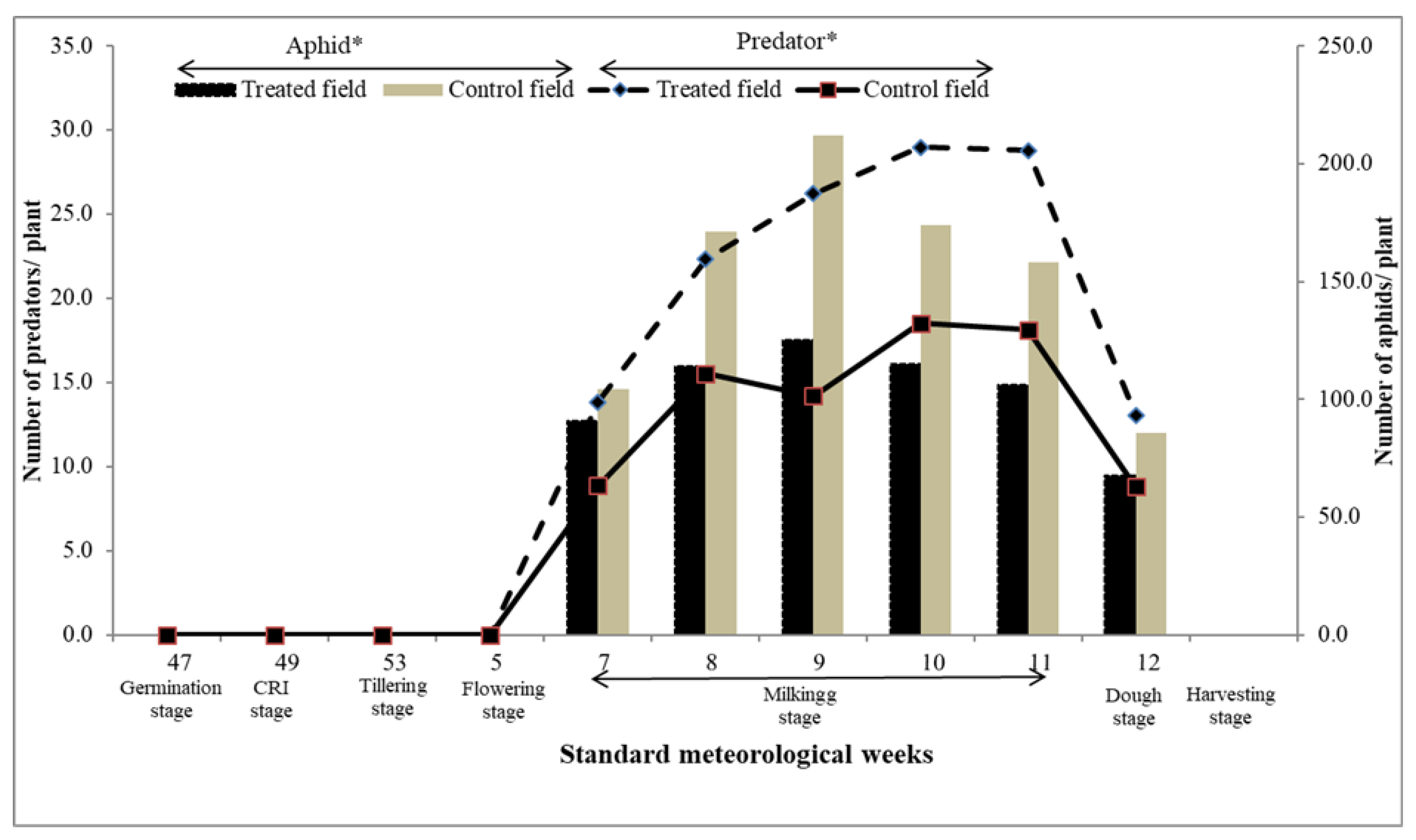

Table 1.
Beneficial arthropod visitors in intercropped with floral strips.
| Common name | Scientific name | Intensity | Family | Order | Type |
|---|---|---|---|---|---|
| Western honey bee | Apis mellifera Linnaeus | High | Apidae | Hymenoptera | Pollinators |
| Indian bee | Apis cerana Fabricius | High | Apidae | Hymenoptera | Pollinators |
| Little honey bee | Apis florae Fabricius | Medium | Apidae | Hymenoptera | Pollinators |
| Rock bee | Apis dorsata Fabricius | Medium | Apidae | Hymenoptera | Pollinators |
| Carpenter bee | Xylocopa sp. | Medium | Apidae | Hymenoptera | Pollinators |
| Sweet bee | Halictus sp.1 | Low | Halictidae | Hymenoptera | Pollinators |
| Sweet bee | Halictussp.2 | Low | Halictidae | Hymenoptera | Pollinators |
| Hover fly | Ceriana sp. | Low | Syrphidae | Diptera | Pollinators |
| Drone fly | Eristalistenax Linnaeus | Medium | Syrphidae | Diptera | Pollinators |
| Hover fly | Eristalinustabanoides(Jaenicke) | Low | Syrphidae | Diptera | Pollinators/ predator |
| Lagoon Flies | Eristalinusobscuritarsus(Meijere) | Medium | Syrphidae | Diptera | Pollinators |
| Marmalade hoverfly | Episyrphusbalteatus(De geer) | High | Syrphidae | Diptera | Pollinators |
| Hoverfly | Eupeodes sp. | High | Syrphidae | Diptera | Pollinators/ predator |
| European hoverfly | Metasyrphus corolla (Fabricius) | Medium | Syrphidae | Diptera | Pollinators |
| Snout fly | Stomorhinasp. | Low | Rhiniidae | Diptera | Pollinators |
| Housefly | Musca sp. | Low | Muscidae | Diptera | Pollinators |
| Plain tiger | Danauschrysippus | Medium | Nymphalidae | Lepidoptera | Pollinators |
| Painted lady | Venessacardui(Linnaeus) | Low | Nymphalidae | Lepidoptera | Pollinators |
| Cabbage butterfly | Pieris brassicae | High | Pieridae | Lepidoptera | Pollinators |
| African clouded yellow | Coliaselecto(Linnaeus) | Low | Pieridae | Lepidoptera | Pollinators |
| Howk moth | Macroglossumstellatarum | Low | Sphingidae | Lepidoptera | Pollinators |
| Seed bug | Graptostethusservus(Fabricius) | Low | Lygaeidae | Hemiptera | Pollinators |
| Shield bug | Dolycoris indicus Stal. | Low | Pentatomidae | Hemiptera | Pollinators |
| Lychee Shield bug | Chrysocorispatricius (Fabricius) | Medium | Scutelleridae | Hemiptera | Pollinators |
| Red pumpkin beetal | Raphidopalpafoveicollis | Low | Chrysomelidae | Coleoptera | Pollinators |
| Seven spotted ladybird beetle | Coccinella septempunctata Linnaeus | High | Coccinellidae | Coleoptera | Predator |
| Six-spotted zigzag ladybird | Cheilomenes sexmaculata (Fabricius) | High | Coccinellidae | Coleoptera | Predator |
| Transverse ladybird | Coccinella transversalis (Fabricius) | Medium | Coccinellidae | Coleoptera | Predator |
| Spotted amber ladybeetle | Hippodamiavariegata(Goeze) sp.1 | Low | Coccinellidae | Coleoptera | Predator |
| Harmonia ladybeetle | Harmonia dimidiate (Fabricius) | Medium | Coccinellidae | Coleoptera | Predator |
Table 2.
Morphometric variation of flowering strips.
| Crop name | Flower type | Pollination type | Flower colour | Flowers per plant | Total flowers (Number) |
Diameter of flower (cm) | Ovary length (cm) | Type of pollen | Blooming period (days) | Total pollinators (Number) |
Total Predators (Number) |
|||
| 25% flowering | 50% flowering | 75% flowering | 100% flowering | |||||||||||
| Singhra | Cross Shaped | Cross | Purple and white combination | 14 ± 3# | 24 ± 4 | 38 ± 8 | 48 ± 8 | 124 | 3.2 ± 0.19 | 1.6 | Oval | 49 | 2895 | 544 |
| Linseed | Disk Shape | Cross | Blue | 6 ± 1 | 8 ± 1 | 8 ± 1 | 9 ± 1 | 32 | 2.4±0.16 | 0.8 | Spherical | 28 | 1185 | 310 |
| Coriander | Asymmetrical | Cross | White | 9 ± 1 | 14 ± 3 | 22 ± 2 | 31 ± 2 | 76 | 1.2 ± 0.11 | 0.2 | Rod | 33 | 836 | 704 |
#value represent as ±SD (standard deviation) from randomly selected ten plants .
Table 3.
Effect of environmental factors on predator and pollinators population in inter.cropped field.
Table 3.
Effect of environmental factors on predator and pollinators population in inter.cropped field.
| Factors | Aphid population | Pollinator population | Predator population | ||||
| Singhra | Linseed | Coriander | Singhra | Linseed | Coriander | ||
| Maximum Temp (0C) | 0.279 | 0.570 | 0.134 | 0.644 | 0.395 | 0.152 | 0.721* |
| Minimum Temp (0C) | 0.392 | 0.356 | 0.516 | 0.943** | 0.187 | 0.162 | 0.889** |
| Maximum RH (%) | -0.049 | -0.020 | 0.168 | -0.498 | -0.008 | 0.085 | -0.618 |
| Minimum RH (%) | -0.010 | 0.045 | 0.265 | -0.499 | 0.236 | 0.131 | -0.621 |
| Rainfall (mm) | -0.834* | -- | -0.467 | 0.138 | -- | 0.125 | 0.381 |
*Correlation is significant at the 0.05 level (2-tailed).**. Correlation is significant at the 0.01 level (2-tailed).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



