
Submitted:
10 December 2025
Posted:
10 December 2025
You are already at the latest version
Abstract
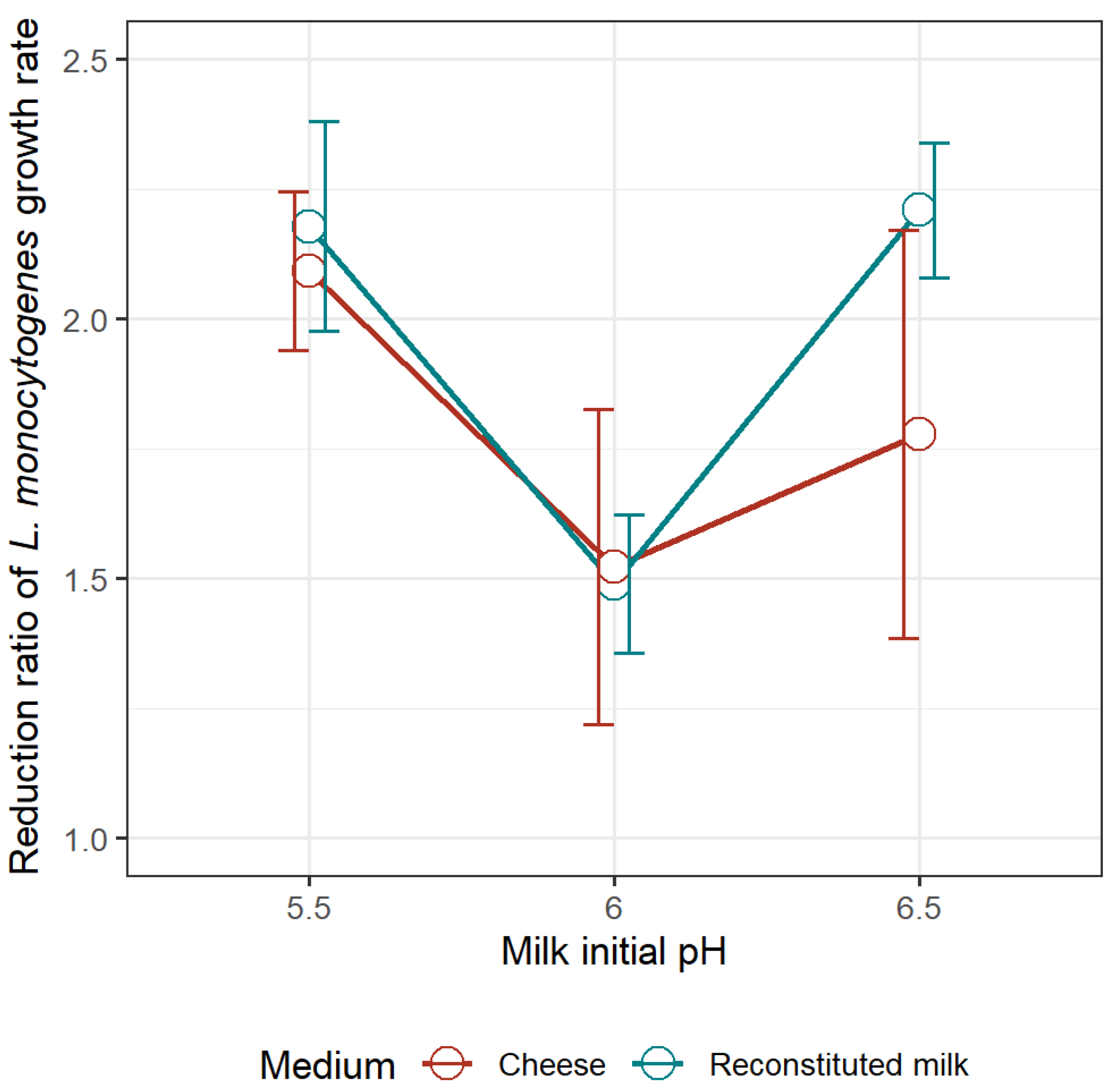
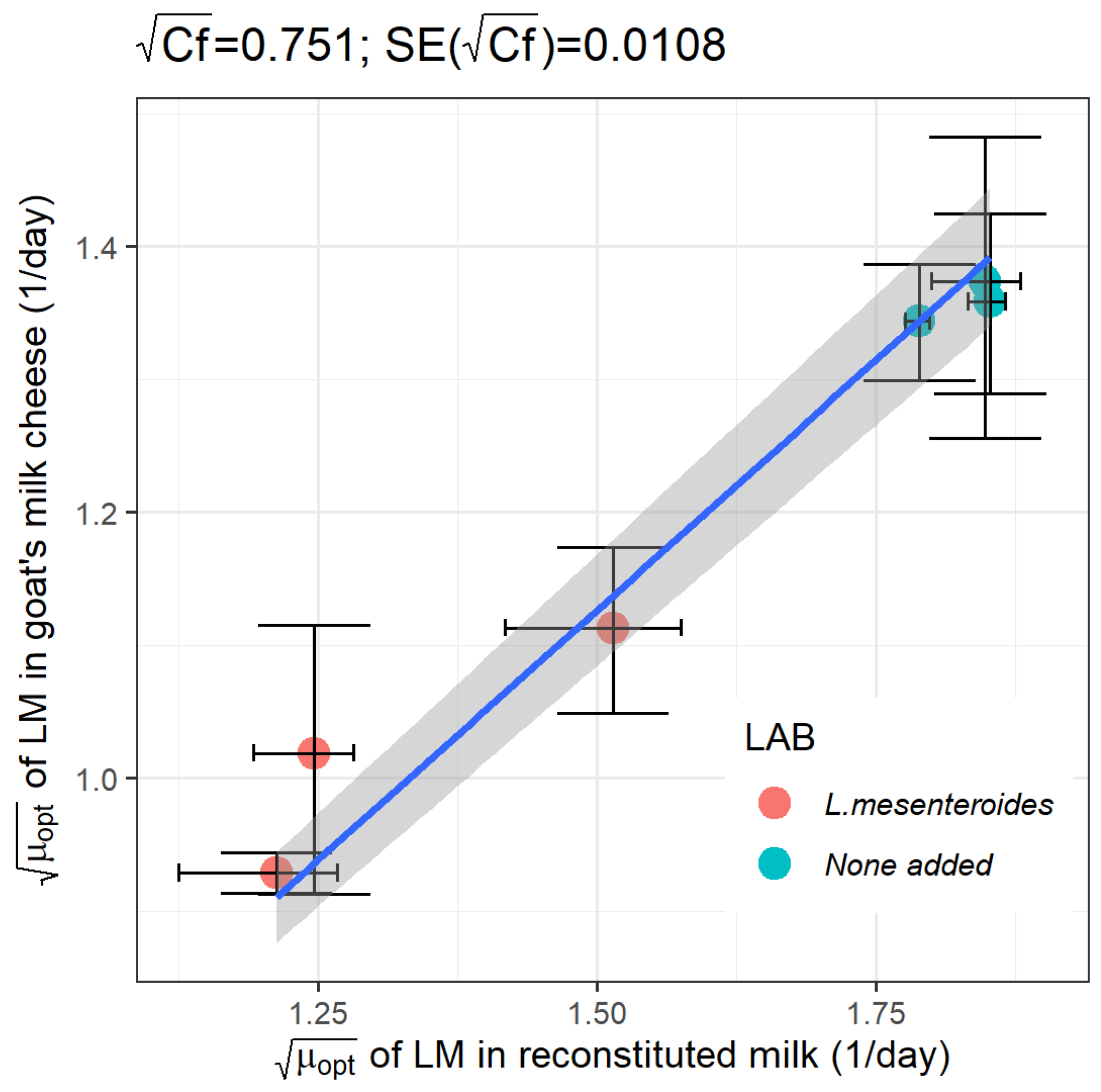
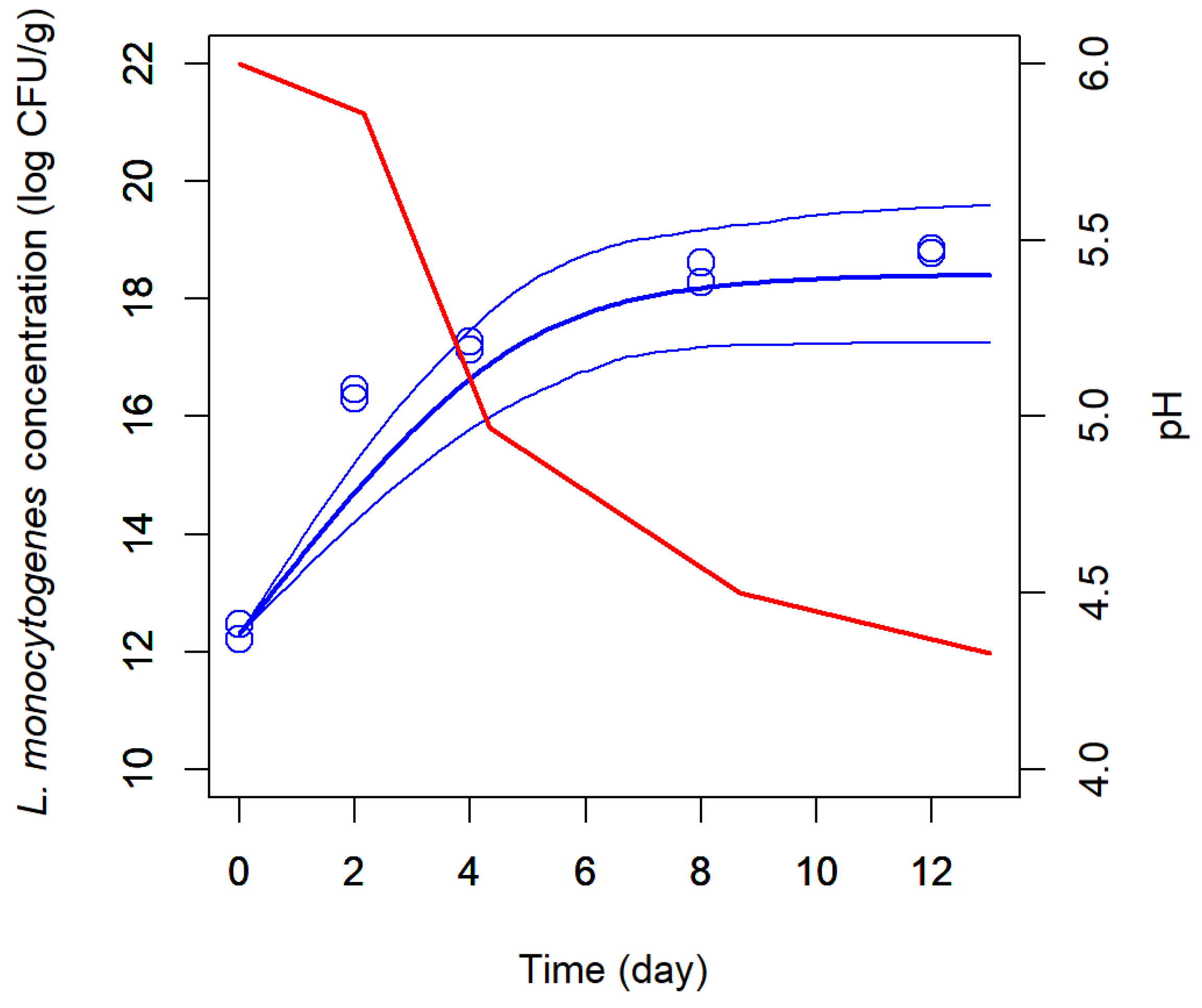
Previous research showed that a strain of Leuconostoc mesenteroides, isolated from goat’s raw milk cheese, was effective in slowing down the growth and reducing the maximum concentration of L. monocytogenes when evaluated in a milk model; and, furthermore, that the extent of inhibition was dependent on the milk initial pH. The objectives of this study were: (1) to determine whether the growth of L. monocytogenes in goat’s pasteurized milk cheese during maturation could be approximated from growth data obtained in the milk model medium, either in monoculture or in coculture with L. mesenteroides; and if so; (2) to model a milk-to-cheese conversion factor (Cf) for L. monocytogenes growth rate. Challenge tests were conducted by inoculating L. monocytogenes in monoculture and in coculture with L. mesenteroides in goat’s pasteurized milk adjusted at initial pH levels of 5.5, 6.0 and 6.5. The process of cheesemaking went on, and cheeses were ripened at 12 ºC during 12 days. Each experimental growth curve was adjusted to a pH-driven dynamic model where the microbial maximum growth rate is a function of pH. As observed in the milk model medium, in coculture with L. mesenteroides, the optimum growth rate (μopt) of L. monocytogenes in maturing cheese was affected by the initial pH of milk: the lowest rate of 0.863 ± 0.042 day-1 was obtained at the initial pH 5.5, in comparison to 1.239 ± 0.208 and 1.038 ± 0.308 day-1 at pH 6.0 and 6.5, respectively. Regardless of the milk initial pH, L. mesenteroides did not reduce the maximum load of L. monocytogenes in maturing cheeses, as it did in the milk medium. By contrary, at the milk initial pH of 5.5, 6.0, and 6.5, L. mesenteroides was able to decrease, on average, 2.2-fold, 1.5-fold and 1.9-fold the μopt of L. monocytogenes in both milk medium and cheese, without significant differences between matrices. Following such validation in goat’s cheese, the square-root of milk-to-cheese Cf for L. monocytogenes was estimated as 0.751 (SE=0.0108), and type of culture (monoculture, coculture) was not found to affect Cf (p=0.320). In conclusion, this work validated pre-acidification of milk as an efficient strategy that, when combined with the use of a protective culture, can synergically enhance the control of L. monocytogenes in cheese.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Preparation
2.2. Manufacture of the Cheese Matrix: Bacterial Strains Inoculation in Milk
2.3. Microbiological and Physicochemical Analyses
2.4. Modelling Approach
2.4.1. Modelling the Growth of L. monocytogenes in Goat’s Milk Cheese During Maturation, and as Affected by L. mesenteroides
2.4.2. Prediction of the Growth of L. monocytogenes in Goat’s Milk Cheese from a Milk Model Medium
3. Results and Discussion
3.1. Listeria Monocytogenes Growth in Cheese During Ripening
3.2. Prediction of the Growth of L. monocytogenes in Goat’s Pasteurized Milk Cheese from a Milk Model Medium
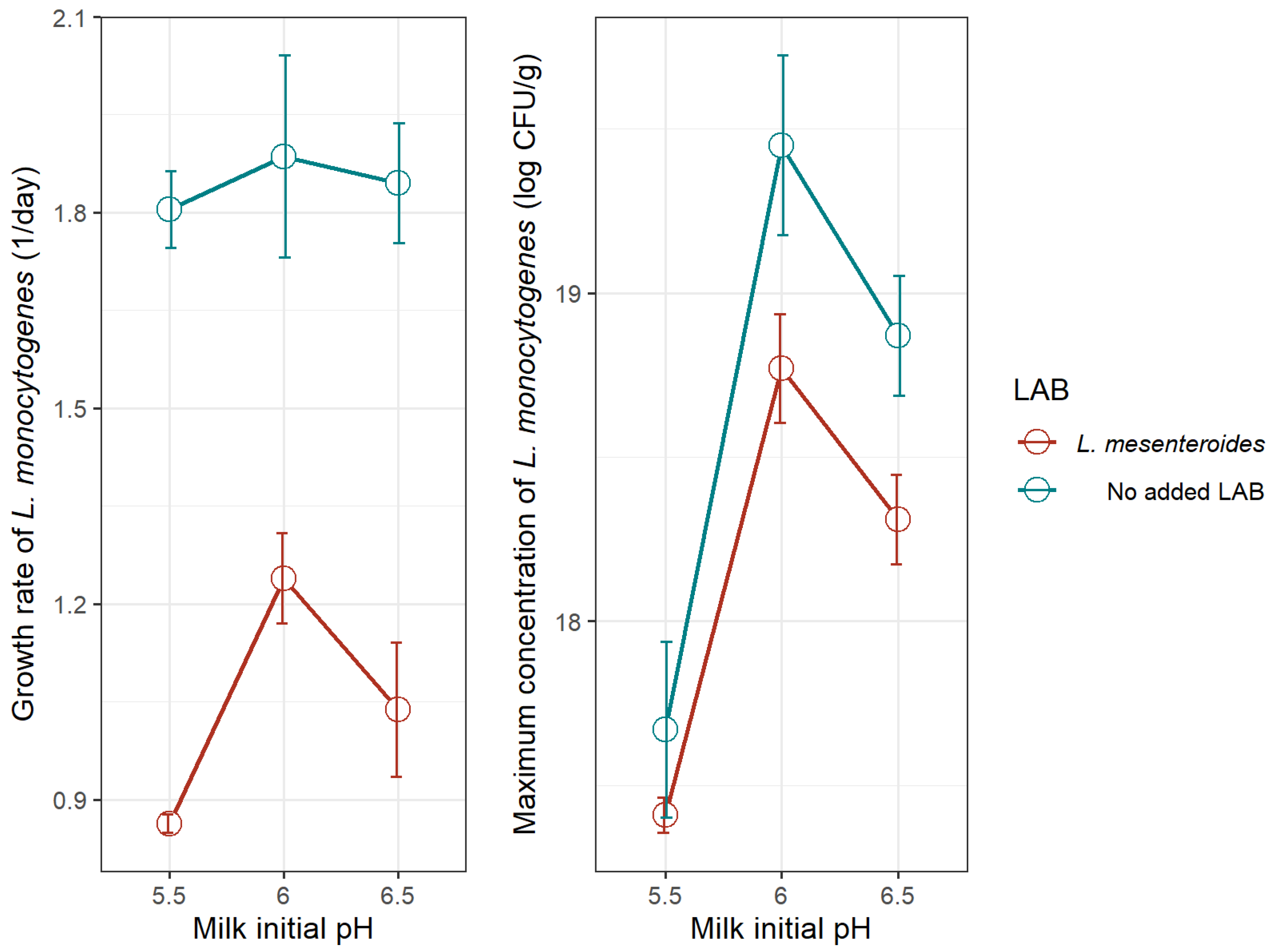
3.2.1. Comparison of L. monocytogenes Growth Kinetics in the Milk Model and Goat’s Pasteurized Milk Cheese in Monoculture and Coculture
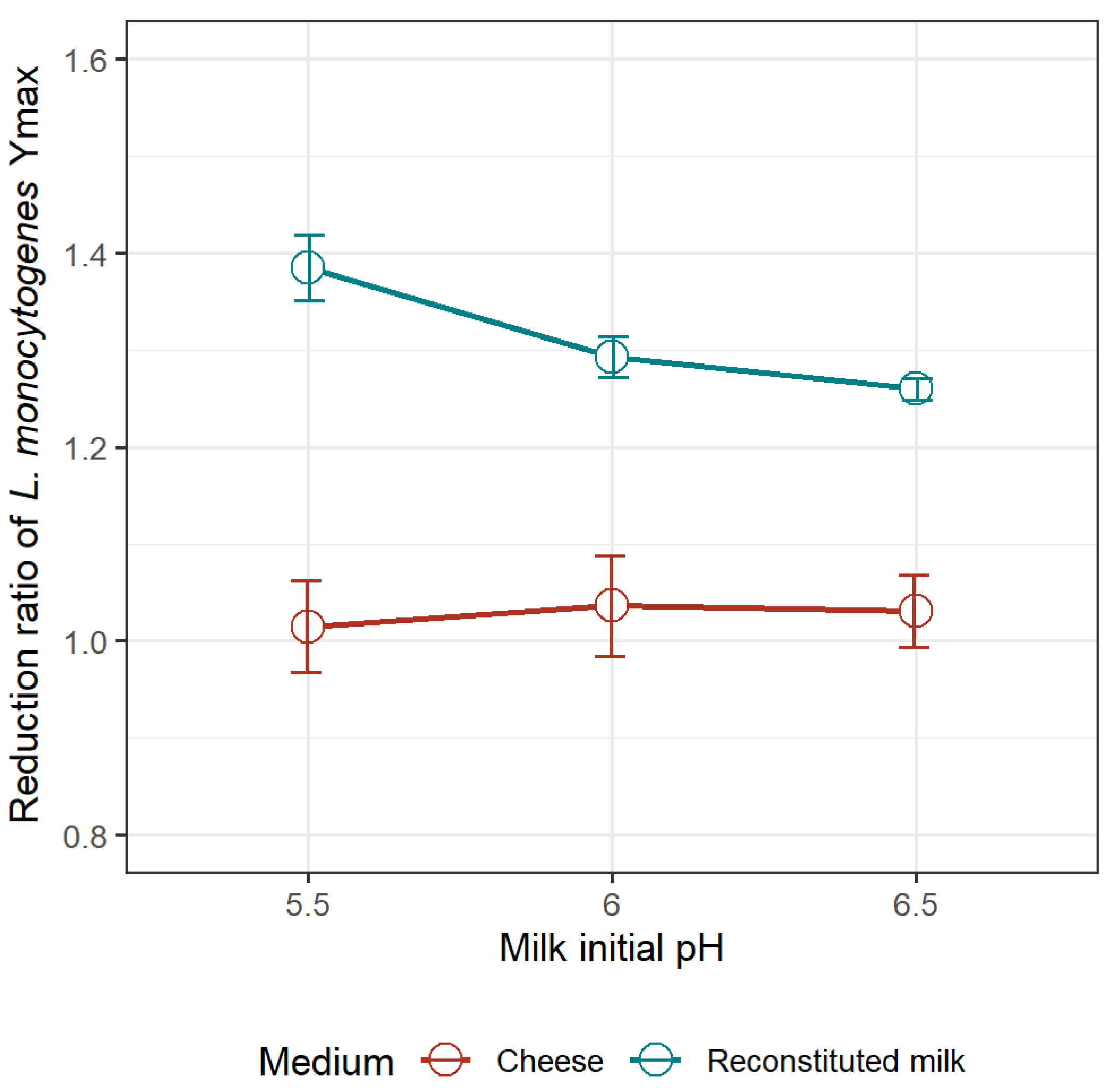
3.2.2. Reduction Ratio of the Optimum Growth Rate of L. monocytogenes Due to the Presence of L. mesenteroides in the Milk Model and in Pasteurized Goat’s Milk Cheese
3.2.3. Model Validation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kontodimos, I.; Chatzimanoli, E.; Kasapidou, E.; Basdagianni, Z.; Karatzia, M.-A.; Amanatidis, M.; Margaritis, N. Characterization of Bioactive Compounds and Element Content in Goat Milk and Cheese Products. Biol. Life Sci. Forum 2023, 26, 98. [CrossRef]
- Tilocca, B.; Soggiu, A.; Iavarone, F.; Greco, V.; Putignani, L.; Ristori, M.V.; Macari, G.; Spina, A.A.; Morittu, V.M.; Ceniti, C.; et al. The Functional Characteristics of Goat Cheese Microbiota from a One-Health Perspective. Int. J. Mol. Sci. 2022, 23, 14131. [CrossRef]
- Quigley, L.; O’Sullivan, O.; Stanton, C.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. The Complex Microbiota of Raw Milk. FEMS Microbiol. Rev. 2013, 37, 664–698. [CrossRef]
- Lomonaco, S.; Decastelli, L.; Nucera, D.; Gallina, S.; Manila Bianchi, D.; Civera, T. Listeria monocytogenes in Gorgonzola: Subtypes, Diversity and Persistence over Time. Int. J. Food Microbiol. 2009, 128, 516–520. [CrossRef]
- Martinez-Rios, V.; Gkogka, E.; Dalgaard, P. Predicting Growth of Listeria monocytogenes at Dynamic Conditions during Manufacturing, Ripening and Storage of Cheeses – Evaluation and Application of Models. Food Microbiol. 2020, 92, 103578. [CrossRef]
- Gonzales-Barron, U.; Cadavez, V.; Guillier, L.; Sanaa, M. A Critical Review of Risk Assessment Models for Listeria monocytogenes in Dairy Products. Foods 2023, 12, 4436. [CrossRef]
- Bashiry, M.; Javanmardi, F.; Taslikh, M.; Sheidaei, Z.; Sadeghi, E.; Abedi, A.-S.; Mirza Alizadeh, A.; Hashempour-Baltork, F.; Beikzadeh, S.; Riahi, S.M.; et al. Listeria monocytogenes in Dairy Products of the Middle East Region: A Systematic Review, Meta-Analysis, and Meta-Regression Study. Iran. J. Public Health 2022, 51, 292–305. [CrossRef]
- Morandi, S.; Silvetti, T.; Battelli, G.; Brasca, M. Can Lactic Acid Bacteria Be an Efficient Tool for Controlling Listeria monocytogenes Contamination on Cheese Surface? The Case of Gorgonzola Cheese. Food Control 2019, 96, 499–507. [CrossRef]
- Webb, L.; Ma, L.; Lu, X. Impact of Lactic Acid Bacteria on the Control of Listeria monocytogenes in Ready-to-Eat Foods. Food Qual. Saf. 2022, 6, fyac045. [CrossRef]
- Morgan, F.; Bonnin, V.; Mallereau, M.-P.; Perrin, G. Survival of Listeria monocytogenes during Manufacture, Ripening and Storage of Soft Lactic Cheese Made from Raw Goat Milk. Int. J. Food Microbiol. 2001, 64, 217–221. [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC) The European Union One Health 2022 Zoonoses Report. EFSA J. 2023, 21, e8442. [CrossRef]
- Jordan, K.; Hunt, K.; Lourenco, A.; Pennone, V. Listeria monocytogenes in the Food Processing Environment. Curr. Clin. Microbiol. Rep. 2018, 5, 106–119. [CrossRef]
- Saraoui, T.; Leroi, F.; Chevalier, F.; Cappelier, J.-M.; Passerini, D.; Pilet, M.-F. Bioprotective Effect of Lactococcus piscium CNCM I-4031 Against Listeria monocytogenes Growth and Virulence. Front. Microbiol. 2018, 9, 1564. [CrossRef]
- Morandi, S.; Silvetti, T.; Vezzini, V.; Morozzo, E.; Brasca, M. How We Can Improve the Antimicrobial Performances of Lactic Acid Bacteria? A New Strategy to Control Listeria monocytogenes in Gorgonzola Cheese. Food Microbiol. 2020, 90. [CrossRef]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A Review of Listeria monocytogenes: An Update on Outbreaks, Virulence, Dose-Response, Ecology, and Risk Assessments. Food Control 2017, 75, 1–13. [CrossRef]
- Liang, J.-J.; He, X.-Y.; Ye, H. Rhombencephalitis Caused by Listeria monocytogenes with Hydrocephalus and Intracranial Hemorrhage: A Case Report and Review of the Literature. WORLD J. Clin. CASES 2019, 7, 538–546. [CrossRef]
- Commission Regulation (EC) No 2073/2005 COMMISSION REGULATION (EC) No 2073/2005 of 15 November 2005 on Microbiological Criteria for Foodstuffs 2005.
- Organization, W.H. Risk Assessment of Listeria monocytogenes in Ready-to-Eat Foods: Technical Report; Microbiological Risk Assessment Series No. 5; World Health Organization: Geneva, 2004; ISBN 978-92-4-156262-1.
- Engstrom, S.K.; Cheng, C.; Seman, D.; Glass, K.A. Growth of Listeria monocytogenes in a Model High-Moisture Cheese on the Basis of pH, Moisture, and Acid Type. J. Food Prot. 2020, 83, 1335–1344. [CrossRef]
- Bansal, V.; Veena, N. Understanding the Role of pH in Cheese Manufacturing: General Aspects of Cheese Quality and Safety. J. Food Sci. Technol. 2024, 61, 16–26. [CrossRef]
- Ho, V.T.T.; Lo, R.; Bansal, N.; Turner, M.S. Characterisation of Lactococcus lactis Isolates from Herbs, Fruits and Vegetables for Use as Biopreservatives against Listeria monocytogenes in Cheese. Food Control 2018, 85, 472–483. [CrossRef]
- Le Marc, Y.; Huchet, V.; Bourgeois, C.M.; Guyonnet, J.P.; Mafart, P.; Thuault, D. Modelling the Growth Kinetics of Listeria as a Function of Temperature, pH and Organic Acid Concentration. Int. J. Food Microbiol. 2002, 73, 219–237. [CrossRef]
- Tirloni, E.; Stella, S.; Bernardi, C.; Dalgaard, P.; Rosshaug, P.S. Predicting Growth of Listeria monocytogenes in Fresh Ricotta. Food Microbiol. 2019, 78, 123–133. [CrossRef]
- Yap, P.-C.; MatRahim, N.-A.; AbuBakar, S.; Lee, H.Y. Antilisterial Potential of Lactic Acid Bacteria in Eliminating Listeria monocytogenes in Host and Ready-to-Eat Food Application. Microbiol. Res. 2021, 12, 234–257. [CrossRef]
- Martín, I.; Rodríguez, A.; Delgado, J.; Córdoba, J.J. Strategies for Biocontrol of Listeria monocytogenes Using Lactic Acid Bacteria and Their Metabolites in Ready-to-Eat Meat- and Dairy-Ripened Products. Foods 2022, 11, 542. [CrossRef]
- Pessione, E. Lactic Acid Bacteria Contribution to Gut Microbiota Complexity: Lights and Shadows. Front. Cell. Infect. Microbiol. 2012, 2, 86. [CrossRef]
- European Food Safety Authority (EFSA). Introduction of a Qualified Presumption of Safety (QPS) Approach for Assessment of Selected Microorganisms Referred to EFSA - Opinion of the Scientific Committee. EFSA J. 2007, 5, 587. [CrossRef]
- Kasra-Kermanshahi, R.; Mobarak-Qamsari, E. Inhibition Effect of Lactic Acid Bacteria against Food Born Pathogen, Listeria monocytogenes. Appl. Food Biotechnol. 2015, 2, 11–19. [CrossRef]
- Castellano, P.; Pérez Ibarreche, M.; Blanco Massani, M.; Fontana, C.; Vignolo, G.M. Strategies for Pathogen Biocontrol Using Lactic Acid Bacteria and Their Metabolites: A Focus on Meat Ecosystems and Industrial Environments. Microorganisms 2017, 5. [CrossRef]
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C.G. Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis to Health-Promoting Properties. Foods 2022, 11, 156. [CrossRef]
- Martín, I.; Rodríguez, A.; Alía, A.; Martínez-Blanco, M.; Lozano-Ojalvo, D.; Córdoba, J.J. Control of Listeria monocytogenes Growth and Virulence in a Traditional Soft Cheese Model System Based on Lactic Acid Bacteria and a Whey Protein Hydrolysate with Antimicrobial Activity. Int. J. Food Microbiol. 2022, 361. [CrossRef]
- Ahansaz, N.; Tarrah, A.; Pakroo, S.; Corich, V.; Giacomini, A. Lactic Acid Bacteria in Dairy Foods: Prime Sources of Antimicrobial Compounds. Fermentation 2023, 9, 964. [CrossRef]
- Egan, K.; Field, D.; Rea, M.C.; Ross, R.P.; Hill, C.; Cotter, P.D. Bacteriocins: Novel Solutions to Age Old Spore-Related Problems? Front. Microbiol. 2016, 7. [CrossRef]
- Lund, P.A.; De Biase, D.; Liran, O.; Scheler, O.; Mira, N.P.; Cetecioglu, Z.; Fernández, E.N.; Bover-Cid, S.; Hall, R.; Sauer, M.; et al. Understanding How Microorganisms Respond to Acid pH Is Central to Their Control and Successful Exploitation. Front. Microbiol. 2020, 11, 556140. [CrossRef]
- Loforte, Y.; Zanzan, M.; Cadavez, V.; Gonzales-Barron, U. Dynamic Modelling of Listeria monocytogenes Growth in a Milk Model Medium as Affected by pH and Selected Lactic Acid Bacteria Strains. Foods 2025, 14, 3999. [CrossRef]
- Silva, B.N.; Faria, A.S.; Cadavez, V.; Teixeira, J.A.; Gonzales-Barron, U. Technological Potential of Lactic Acid Bacteria Isolated from Portuguese Goat’s Raw Milk Cheeses. In Proceedings of the Foods 2021; MDPI, October 14 2021; p. 9.
- Silva, B.N.; Fernandes, N.; Carvalho, L.; Faria, A.S.; Teixeira, J.A.; Rodrigues, C.; Gonzales-Barron, U.; Cadavez, V. Lactic Acid Bacteria from Artisanal Raw Goat Milk Cheeses: Technological Properties and Antimicrobial Potential. Ital. J. Food Saf. 2023, 12, 11559. [CrossRef]
- Schvartzman, M.S.; Belessi, X.; Butler, F.; Skandamis, P.; Jordan, K. Comparison of Growth Limits of Listeria monocytogenes in Milk, Broth and Cheese: Comparison of Growth Limits of L. monocytogenes. J. Appl. Microbiol. 2010, no-no. [CrossRef]
- Silva, B.N.; Coelho-Fernandes, S.; Teixeira, J.A.; Cadavez, V.; Gonzales-Barron, U. Dynamic Modelling to Describe the Effect of Plant Extracts and Customised Starter Culture on Staphylococcus aureus Survival in Goat’s Raw Milk Soft Cheese. Foods 2023, 12, 2683. [CrossRef]
- ISO 11290-1:2017; ISO 11290-1: Microbiology of the Food Chain — Horizontal Method for the Detection and Enumeration of Listeria monocytogenes and of Listeria Spp. — Part 1: Detection Method. International Organization for Standardization (ISO): Geneva, Switzerland, 2017.
- ISO 15214:1998; ISO 15214: 4: Microbiology of Food and Animal Feeding Stuffs — Horizontal Method for the Emumeration of Mesophilic Lactic Acid Bacteria — Colony-Count Technique at 30 °C. International Organization for Standardization (ISO): Genève, Switzerland, 1998.
- Campagnollo, F.B.; Gonzales-Barron, U.; Pilão Cadavez, V.A.; Sant’Ana, A.S.; Schaffner, D.W. Quantitative Risk Assessment of Listeria monocytogenes in Traditional Minas Cheeses: The Cases of Artisanal Semi-Hard and Fresh Soft Cheeses. Food Control 2018, 92, 370–379. [CrossRef]
- Lau, S.; Trmcic, A.; Martin, N.H.; Wiedmann, M.; Murphy, S.I. Development of a Monte Carlo Simulation Model to Predict Pasteurized Fluid Milk Spoilage Due to Post-Pasteurization Contamination with Gram-Negative Bacteria. J. Dairy Sci. 2022, 105, 1978–1998. [CrossRef]
- R: The R Project for Statistical Computing Available online: https://www.r-project.org/ (accessed on 1 October 2025).
- Rosso, L.; Lobry, J.R.; Bajard, S.; Flandrois, J.P. Convenient Model To Describe the Combined Effects of Temperature and pH on Microbial Growth. Appl. Environ. Microbiol. 1995, 61, 610–616. [CrossRef]
- Listeria Available online: https://www.asae.gov.pt/seguranca-alimentar/riscos-biologicos/listeria-monocytogenes.aspx (accessed on 16 November 2025).
- Faezi-Ghasemi, M.; Kazemi, S. Effect of Sub-Lethal Environmental Stresses on the Cell Survival and Antibacterial Susceptibility of Listeria monocytogenes PTCC1297; Zahedan Journal of Research in Medical Sciences, 2013.
- Jin, Q.; Kirk, M.F. pH as a Primary Control in Environmental Microbiology: 1. Thermodynamic Perspective. Front. Environ. Sci. 2018, 6. [CrossRef]
- Wiktorczyk-Kapischke, N.; Skowron, K.; Grudlewska-Buda, K.; Wałecka-Zacharska, E.; Korkus, J.; Gospodarek-Komkowska, E. Adaptive Response of Listeria monocytogenes to the Stress Factors in the Food Processing Environment. Front. Microbiol. 2021, 12. [CrossRef]
- Martín, I.; Córdoba, J.J.; Rodríguez, A. Effect of Acidic Conditions on the Growth and Expression of Two Virulence Genes of Listeria monocytogenes Serotype 4b. Res. Microbiol. 2023, 174, 104042. [CrossRef]
- Bergis, H.; Bonanno, L.; Asséré, A.; Lombard, B. On Challenge Tests and Durability Studies for Assessing Shelf-Life of Ready-to-Eat Foods Related to Listeria monocytogenes. 2021.
- Kapetanakou, A.E.; Gkerekou, M.A.; Vitzilaiou, E.S.; Skandamis, P.N. Assessing the Capacity of Growth, Survival, and Acid Adaptive Response of Listeria monocytogenes during Storage of Various Cheeses and Subsequent Simulated Gastric Digestion. Int. J. Food Microbiol. 2017, 246, 50–63. [CrossRef]
- Leong, W.M.; Geier, R.; Engstrom, S.; Ingham, S.; Ingham, B.; Smukowski, M. Growth of Listeria monocytogenes, Salmonella spp., Escherichia Coli O157:H7, and Staphylococcus Aureus on Cheese during Extended Storage at 25 Degrees C. J. Food Prot. 2014, 77, 1275–1288. [CrossRef]
- Ariahu, C.C.; Igyor, M.A.; Umeh, E.U. Growth kinetics of Listeria monocytogenes in soymilk of varying initial pH and Sugar Concentrations. J. Food Qual. 2010, 33, 545–558. [CrossRef]
- Gonzales-Barron, U.; Campagnollo, F.B.; Schaffner, D.W.; Sant’Ana, A.S.; Cadavez, V.A.P. Behavior of Listeria monocytogenes in the Presence or Not of Intentionally-Added Lactic Acid Bacteria during Ripening of Artisanal Minas Semi-Hard Cheese. Food Microbiol. 2020, 91. [CrossRef]
- Cadavez, V.A.P.; Campagnollo, F.B.; Silva, R.A.; Duffner, C.M.; Schaffner, D.W.; Sant’Ana, A.S.; Gonzales-Barron, U. A Comparison of Dynamic Tertiary and Competition Models for Describing the Fate of Listeria monocytogenes in Minas Fresh Cheese during Refrigerated Storage. Food Microbiol. 2019, 79, 48–60. [CrossRef]
- Mejlholm, O.; Dalgaard, P. Modelling and Predicting the Simultaneous Growth of Listeria monocytogenes and Psychrotolerant Lactic Acid Bacteria in Processed Seafood and Mayonnaise-Based Seafood Salads. Food Microbiol. 2015, 46, 1–14. [CrossRef]
- Lambert, R.J.W. A New Model for the Effect of pH on Microbial Growth: An Extension of the Gamma Hypothesis. J. Appl. Microbiol. 2011, 110, 61–68. [CrossRef]
- Nunes Silva, B.; Cadavez, V.; Teixeira, J.A.; Ellouze, M.; Gonzales-Barron, U. Cardinal Parameter Meta-Regression Models Describing Listeria monocytogenes Growth in Broth. Food Res. Int. 2020, 136, 109476. [CrossRef]
- Loforte, Y.; Fernandes, N.; de Almeida, A.M.; Cadavez, V.; Gonzales-Barron, U. A Meta-Analysis on the In Vitro Antagonistic Effects of Lactic Acid Bacteria from Dairy Products on Foodborne Pathogens. Foods 2025, 14, 907. [CrossRef]
- Augustin, J.-C.; Czarnecka-Kwasiborski, A. Single-Cell Growth Probability of Listeria monocytogenes at Suboptimal Temperature, pH, and Water Activity. Front. Microbiol. 2012, 3. [CrossRef]
- Barbosa, J.; Borges, S.; Teixeira, P. Influence of Sub-Lethal Stresses on the Survival of Lactic Acid Bacteria after Spray-Drying in Orange Juice. Food Microbiol. 2015, 52, 77–83. [CrossRef]
- Coelho, M.C.; Malcata, F.X.; Silva, C.C.G. Lactic Acid Bacteria in Raw-Milk Cheeses: From Starter Cultures to Probiotic Functions. Foods 2022, 11, 2276. [CrossRef]
- Hemme, D.; Foucaud-Scheunemann, C. Leuconostoc, characteristics, use in dairy technology and prospects in functional foods. Int. Dairy J. 2004, 14, 467–494. [CrossRef]
- Wadhawan, K.; Steinberger, A.; Rankin, S.; Suen, G.; Czuprynski, C. Inhibition of Listeria monocytogenes by Broth Cultures of Surface Microbiota of Wooden Boards Used in Cheese Ripening. Appl. Sci. 2023, 13, 5872. [CrossRef]
- Chi, H. Identification and Characterization of a Bacteriocin-Like Substance, Produced by Leuconostoc mesenteroides as a Bio-Preservative Against Listeria monocytogenes. Int. J. Nutr. Food Sci. 2017, 6, 167. [CrossRef]
- Héchard, Y.; Dérijard, B.; Letellier, F.; Cenatiempo, Y. Characterization and Purification of Mesentericin Y105, an Anti-Listeria Bacteriocin from Leuconostoc mesenteroides. Microbiology 1992, 138, 2725–2731. [CrossRef]
- Shi, F.; Wang, Y.; Li, Y.; Wang, X. Mode of Action of Leucocin K7 Produced by Leuconostoc mesenteroides K7 against Listeria monocytogenes and Its Potential in Milk Preservation. Biotechnol. Lett. 2016, 38, 1551–1557. [CrossRef]
- Borges, D.O.; Matsuo, M.M.; Bogsan, C.S.B.; Silva, T.F. da; Casarotti, S.N.; Penna, A.L.B. Leuconostoc mesenteroides subsp. mesenteroides SJRP55 Reduces Listeria monocytogenes Growth and Impacts on Fatty Acids Profile and Conjugated Linoleic Acid Content in Fermented Cream. LWT 2019, 107, 264–271. [CrossRef]
- Silva, B.N.; Coelho-Fernandes, S.; Teixeira, J.A.; Cadavez, V.; Gonzales-Barron, U. Dynamic Modelling to Describe the Effect of Plant Extracts and Customised Starter Culture on Staphylococcus aureus Survival in Goat’s Raw Milk Soft Cheese. Foods 2023, 12, 2683. [CrossRef]
- Schvartzman, M.S.; Belessi, X.; Butler, F.; Skandamis, P.; Jordan, K. Comparison of Growth Limits of Listeria monocytogenes in Milk, Broth and Cheese: Comparison of Growth Limits of L. monocytogenes. J. Appl. Microbiol. 2010, no-no. [CrossRef]
- Schvartzman, M.S.; Belessi, C.; Butler, F.; Skandamis, P.N.; Jordan, K.N. Effect of pH and Water Activity on the Growth Limits of Listeria monocytogenes in a Cheese Matrix at Two Contamination Levels. J. Food Prot. 2011, 74, 1805–1813. [CrossRef]
- Nuñez, M.; Calzada, J.; Olmo, A. del. High Pressure Processing of Cheese: Lights, Shadows and Prospects. Int. Dairy J. 2020, 100, 104558. [CrossRef]
- Murphy, P.M.; Rea, M.C.; Harrington, O. Development of a Predictive Model for Growth of Listeria monocytogenes in a Skim Milk Medium and Validation Studies in a Range of Dairy Products. J. Appl. Bacteriol. 1996, 80, 557–564. [CrossRef]
- Lund, P.; Tramonti, A.; De Biase, D. Coping with Low pH: Molecular Strategies in Neutralophilic Bacteria. FEMS Microbiol. Rev. 2014, 38, 1091–1125. [CrossRef]
- Cotter, P.D.; Ryan, S.; Gahan, C.G.M.; Hill, C. Presence of GadD1 glutamate decarboxylase in selected Listeria monocytogenes strains is associated with an ability to grow at low pH. Appl. Environ. Microbiol. 2005, 71, 2832–2839. [CrossRef]
- Cotter, P.D.; O’reilly, K.; Hill, C. Role of the glutamate decarboxylase acid resistance system in the survival of Listeria monocytogenes LO28 in low pH foods. J. Food Prot. 2001, 64, 1362–1368. [CrossRef]
- Cotter, P.D.; Gahan, C.G.M.; Hill, C. Analysis of the role of the Listeria monocytogenes F0F1-ATPase operon in the acid tolerance response. Int. J. Food Microbiol. 2000, 60, 137–146. [CrossRef]
- Maloney, P.C. Microbes and Membrane Biology. FEMS Microbiol. Lett. 1990, 87, 91–102. [CrossRef]
- Scatassa, M.L.; Gaglio, R.; Cardamone, C.; Macaluso, G.; Arcuri, L.; Todaro, M.; Mancuso, I. Anti-Listeria Activity of Lactic Acid Bacteria in Two Traditional Sicilian Cheeses. Ital. J. Food Saf. 2017, 6, 6191. [CrossRef]
- Schvartzman, M.S.; Gonzalez-Barron, U.; Butler, F.; Jordan, K. Modeling the Growth of Listeria monocytogenes on the Surface of Smear- or Mold-Ripened Cheese. Front. Cell. Infect. Microbiol. 2014, 4. [CrossRef]
- Kongo, J.M. Lactic Acid Bacteria as Starter-Cultures for Cheese Processing: Past, Present and Future Developments. In Lactic Acid Bacteria - R & D for Food, Health and Livestock Purposes; IntechOpen, 2013 ISBN 978-953-51-0955-6.

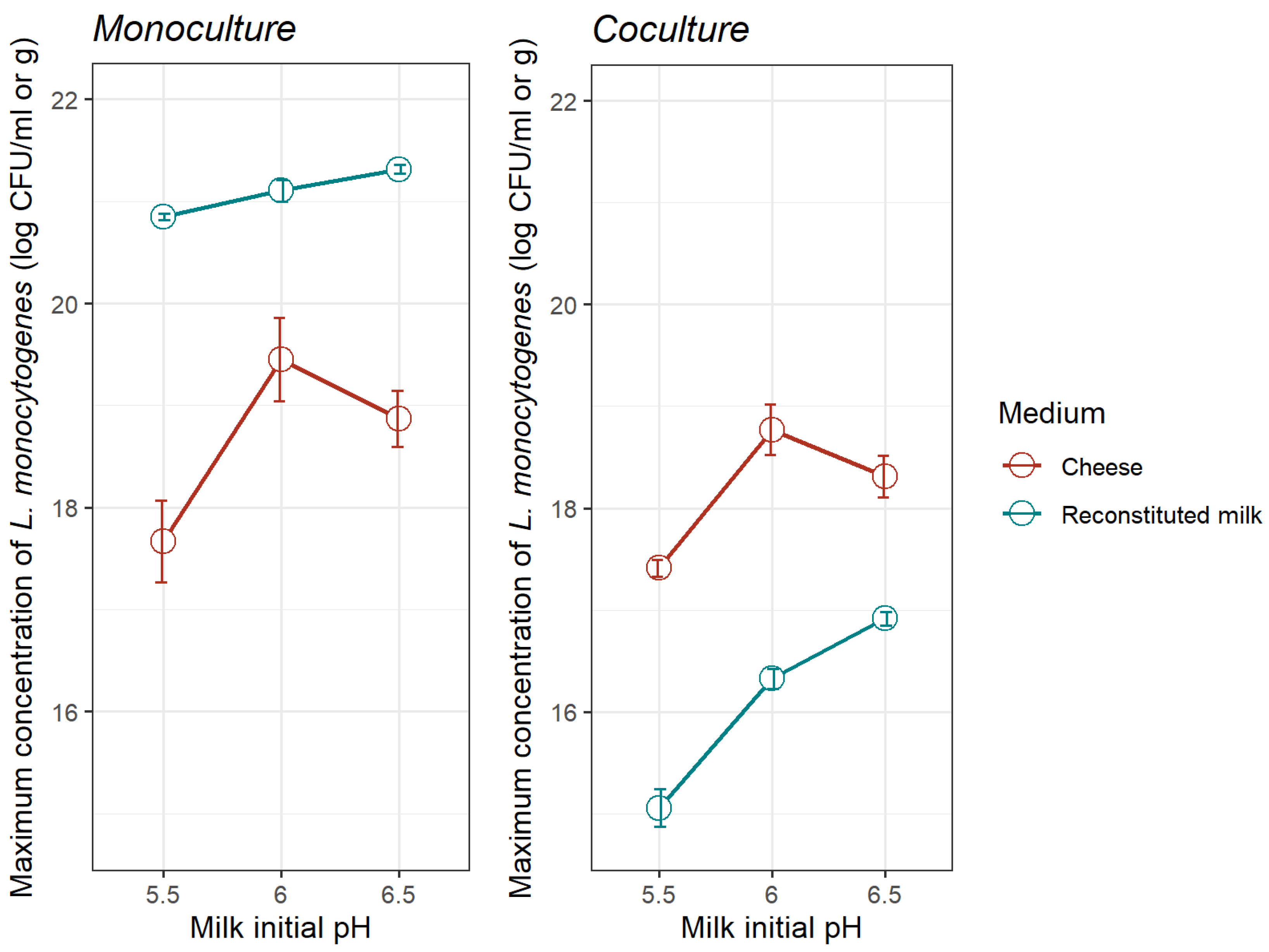

| Milk initial pH | Parameters | Monoculture | Coculture |
|---|---|---|---|
| 5.5 | μopt | 3.201 (0.060) | 1.469 (0.205) |
| Ymax | 20.85 (0.060) | 15.05 (0.367) | |
| 6.0 | μopt | 3.416 (0.177) | 2.293 (0.284) |
| Ymax | 21.10 (0.212) | 16.32 (0.204) | |
| 6.5 | μopt | 3.432 (0.073) | 1.552 (0.132) |
| Ymax | 21.31 (0.085) | 16.91 (0.132) |
| Initial pH of milk | L. monocytogenes | Fit quality | ||
|---|---|---|---|---|
| Parameters | Mean (SE) | Pr > |t| | ||
| 5.5 | Y0 | 13.07 (1.132) | 0.055 | σ²: 0.4268 |
| μopt | 1.805 (0.177) | 0.040 | RMSE: 0.5658 | |
| Ymax | 17.67 (0.800) | 0.106 | MAE: 0.4002 | |
| 6.0 | Y0 | 12.72 (1.163) | 0.048 | σ²: 0.4495 |
| μopt | 1.886 (0.465) | 0.002 | RMSE: 0.5806 | |
| Ymax | 19.45 (0.821) | 0.027 | MAE: 0.4109 | |
| 6.5 | Y0 | 12.73 (0.771) | 0.038 | σ²: 0.1976 |
| μopt | 1.845 (0.275) | 0.024 | RMSE: 0.3851 | |
| Ymax | 18.87 (0.545) | 0.018 | MAE: 0.2724 | |
| Initial pH of milk | L. monocytogenes | L. mesenteroides | |||
|---|---|---|---|---|---|
| Parameters | Mean (SE) | Pr > |t| | Mean (SE) | Pr > |t| | |
| 5.5 | Y0 | 13.08 (0.049) | <0.001 | 16.88 (0.205) | <0.001 |
| μopt | 0.863 (0.042) | <0.001 | 0.646 (0.087) | 0.005 | |
| Ymax | 17.41 (0.163) | <0.001 | 20.49 (0.231) | <0.001 | |
| Fit quality | |||||
| σ² | 0.0012 | 0.0259 | |||
| RMSE | 0.0312 | 0.1468 | |||
| MAE | 0.0239 | 0.1280 | |||
| 6.0 | Y0 | 12.45 (0.479) | <0.001 | 15.91 (0.525) | <0.001 |
| μopt | 1.239 (0.208) | <0.001 | 0.713 (0.082) | 0.030 | |
| Ymax | 18.77 (0.497) | <0.001 | 22.00 (3.964) | 0.011 | |
| Fit quality | |||||
| σ² | 0.1390 | 0.1709 | |||
| RMSE | 0.3404 | 0.3780 | |||
| MAE | 0.2468 | 0.3446 | |||
| 6.5 | Y0 | 12.56 (0.615) | <0.001 | 15.54 (0.268) | <0.001 |
| μopt | 1.038 (0.308) | 0.043 | 1.009 (0.091) | <0.001 | |
| Ymax | 18.31 (0.407) | <0.001 | 21.57 (0.187) | <0.001 | |
| Fit quality | |||||
| σ² | 0.2256 | 0.0636 | |||
| RMSE | 0.4335 | 0.2336 | |||
| MAE | 0.3697 | 0.1780 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



