
Submitted:
02 December 2025
Posted:
03 December 2025
You are already at the latest version
Abstract
Pasture-based ruminant systems link nitrogen (N) nutrition with ecosystem N cycling. Grazing ruminants convert fibrous forages into milk and meat but excrete 65 to 80% of ingested N, creating excreta hotspots that drive ammonia volatilization, nitrate leaching, and nitrous oxide (N₂O) emissions. This review synthesizes ecological and rumen-nutrition evidence on N flows, emphasizing microbial processes, biological N₂ fixation, plant diversity, and urine patch biogeochemistry, and evaluates strategies to improve N use efficiency (NUE). We examine rumen N metabolism in relation to microbial protein synthesis, urea recycling, and dietary factors including crude protein concentration, energy supply, forage composition, and plant secondary compounds that modulate protein degradability and microbial N capture, thereby influencing N partitioning among animal products, urine, and feces, as reflected in milk and blood urea N. Also, examine how grazing patterns and excreta distribution, assessed with sensor technologies, modify N flows. Evidence indicates that integrated management combining dietary manipulation, forage diversity, targeted grazing, and decision tools can increase farm-gate NUE from 20-25% to over 30% while sustaining performance. Framing these processes within the global N cycle positions pasture-based ruminant systems as critical leverage points for aligning ruminant production with environmental and climate sustainability goals.
Keywords:
nitrogen use efficiency
; grazing management
; urine patches
; condensed tannins
; forage diversity
; nitrous oxide
1. Introduction
Globally, human activities have more than doubled flows of reactive nitrogen (Nr), largely through synthetic fertilizer production and expansion of livestock systems, with significant consequences for water quality, biodiversity, and climate forcing via nitrous oxide (N2O) emissions [1,2]. Livestock accounts for a major share of anthropogenic Nr and contributes substantially to the exceedance of safe planetary boundaries for Nr [3]. Within this global context, pasture-based grazing systems are critical interfaces between Nr inputs and the wider environment. Nitrogen underpins plant productivity, forage nutritive value, microbial protein synthesis, and animal performance, yet mismanagement generates substantial N losses to air and water. In grazing dairy and beef systems, nitrogen use efficiency (NUE)- the proportion of N intake recovered in milk and meat typically ranges from 10 to 35%, with the remainder excreted and lost as ammonia (NH3), nitrate (NO3−), and N2O [4,5]. This low NUE reflects imbalances between N inputs and productive demand arising from interactions among soil, plant, microbial, and ruminant processes [6]. Legume-based swards can enhance forage protein and reduce reliance on fertilizer N [4,7], but when total N inputs exceed removal in animal products and plant biomass, surpluses and losses intensify, particularly under high stocking densities and uneven excreta deposition [8,9,10] Grazing returns N in urine and dung, accelerates N cycling, and creates excreta hotspots associated with disproportionately high N2O emissions [11,12].
In ruminants, dietary N moves through tightly coupled microbial–host pathways. Rumen microbes convert degradable protein into microbial protein and ammonia; when protein supply exceeds fermentable energy, excess ammonia is converted to urea and excreted [12,13]. Balancing crude protein with energy and synchronizing rumen-degradable protein with fermentable carbohydrate increases microbial ammonia capture and N retention, and reduces urinary N losses [14,15,16]. Forage composition further modulates these flows. Plant secondary compounds, particularly condensed tannins, can reduce ruminal proteolysis, shift N excretion from urine to feces, and mitigate NH3 volatilization and N2O emissions [16,17,18,19]. Milk urea nitrogen (MUN) and blood urea nitrogen (BUN) integrate these processes and provide practical indicators of herd-level NUE [20,21,22]. Once excreted, urine and dung generate fine-scale heterogeneity in nutrient availability and soil N processes through rapid urea hydrolysis, nitrification, and denitrification, as well as slower organic matter decomposition [23,24,25]. Excreta distribution is shaped by grazing–resting cycles, terrain, and attraction to shade, water, and supplements, and emerging tracking tools (GPS, accelerometers, biologgers) now link animal movement and behavior to N return patterns and emission hotspots [26]. Similar mismatches between N supply, animal demand, and spatial N return occur across mixed crop–livestock and pastoral systems, including low-input systems in sub-Saharan Africa [27,30,31,32].
Despite extensive research, ecological and nutritional work on N in grazing systems remains fragmented. Ecological studies emphasize soil–plant–microbe dynamics and excreta hotspots, whereas ruminant nutrition studies focus on rumen function, diet formulation, and indicators such as MUN and BUN [28,29,30]. Global assessments indicate that single agronomic, nutritional, or behavioral interventions are unlikely to achieve the reductions in N losses needed to remain within planetary boundaries [3,31,32]. This review synthesizes ecological and ruminant nutrition perspectives on N dynamics and use efficiency in pasture-based grazing systems by linking soil microbial processes, forage chemistry, rumen and post-ruminal N metabolism, animal behavior, spatial excreta patterns, and farm-scale N balances, with particular emphasis on how rumen N metabolism, grazing behavior, and spatial N returns jointly influence NUE. The overarching objective is to clarify the mechanisms underlying low NUE, evaluate strategies to enhance N retention and reduce environmental N losses, and identify research priorities for pasture-based grazing systems that sustain animal performance while aligning ruminant production with environmental and climate sustainability goals.
2. Discussion
2.1. Nitrogen Flows in Pasture-Based Grazing Systems
Pasture-based grazing systems exhibit nitrogen (N) flows that span interconnected biological, ecological, and behavioral processes. Nitrogen inputs derived from biological fixation, synthetic fertilizer, imported feeds, and atmospheric deposition enter a dynamic continuum of plant uptake, soil microbial transformations, and animal ingestion and excretion. Losses through volatilization, leaching, and N2O emissions emerge from these processes and are strongly modulated by diet composition, grazing behavior, and the spatial distribution of excreta. Synthesizing N dynamics across soil, plant, animal, and behavioral scales is therefore essential to evaluate and improve nitrogen use efficiency (NUE) and environmental performance in grazing systems. These multi-scale nitrogen flows are illustrated in Figure 1, which summarizes farm inputs, internal cycling, product exports, and loss pathways in a pasture-based grazing system.
2.1.1. System Boundaries and Nitrogen Use Efficiency
Nitrogen flows in grazing systems operate simultaneously at whole-farm, paddock, and animal scales, and inefficiencies at one level propagate to others. At the farm gate, inputs include imported feeds, mineral N fertilizer, biological N fixation (BNF), atmospheric deposition, and sometimes bedding or imported manure. In contrast, outputs comprise milk, meat, live animals, and conserved forage. Internal flows connect plant uptake, soil organic N dynamics, excreta deposition, and gaseous or leaching losses, indicating that NUE emerges from coupled ecological and nutritional processes rather than isolated components. Across intensive grazing dairy systems, farm-gate NUE typically ranges from 20 to 35%, with the remainder of N accumulating in soil, lost as gases, or exported in leachate [33,34].
At the animal level, biological retention is similarly low. Grazing dairy cows retain only 18 to 26% of ingested N in milk, and young heifers retain about 20 to 30% of dietary N [35,36], implying that roughly 60 to 80% of N intake is excreted [37,38]. Small physiological shifts, such as selecting cows with lower milk urea nitrogen (MUN) breeding values, can increase NUE by 3- [39], demonstrating that both genetics and nutrition influence N flows. However, NUE estimates vary widely depending on system boundaries. Schils et al. [40] show that Dutch dairy farm-gate NUE ranged from 14 to 44%, primarily driven by fertilizer rates, stocking density, and feed imports rather than by inherent biological limits. Powell et al. [41] noted that low NUE may still coincide with productive soil N accumulation, whereas Oenema et al. [42] emphasized that even moderate NUE (25 to 30%) can be associated with high absolute N surpluses. Thus, efficiency ratios alone cannot diagnose sustainability unless absolute surplus N and the fate of accumulated N are also considered.
Reducing dietary crude protein (CP) can improve animal-level NUE and reduce excretory losses when CP exceeds requirements. For example, decreasing CP from 17% to 15% reduced urinary N excretion by about 20% without compromising milk yield [43], whereas further reductions below physiological thresholds decreased performance [44]. Genetic gains in NUE are possible [45], but remain smaller in magnitude than nutritional and management interventions [46]. Collectively, these findings indicate that improving NUE requires coordinated genetic, dietary, and management strategies, and that interpretations must be explicit about the spatial and temporal boundaries used to calculate NUE.
2.1.2. External Farm Inputs and Internal N Cycling
External N inputs and internal cycling together determine whether pasture-based systems function as N sources or sinks. At the farm and paddock scales, BNF from legumes, mineral fertilizer, imported feeds, and atmospheric deposition all contribute to the soil–plant–animal continuum. The relative contribution of these sources is strongly management-dependent. For example, grass–clover swards can supply N via Rhizobium-driven BNF at rates comparable to fertilized grass monocultures, but clover content and N inputs interact with stocking density, grazing pressure, and fertilizer strategy to shape long-term N balances [47,48].
Within the soil, mineralization processes regulated by temperature, moisture, C: N ratio, and microbial communities interact with plant uptake and root turnover to determine whether N is retained in soil organic pools, captured in above- and below-ground biomass, or left vulnerable to loss [49,50,51]. Integrated crop–livestock systems can improve N capture by distributing excreta more evenly and by using cover crops to intercept nitrate and recycle N back into the system [52]. However, long-term accumulation of 200 to 400 kg N ha−1 in soil [53] may either support climate mitigation or increase emissions and leaching, depending on how internal cycling is managed [54]. These patterns highlight that external inputs must be interpreted through the lens of internal cycling dynamics and system goals.
2.1.3. Loss Pathways: Leaching, Volatilization, and N2O Emissions
Loss pathways from grazing systems arise when N inputs and internal cycling exceed the capacity of soils and plants to retain reactive N. Under these conditions, losses
occur primarily via ammonia (NH3) volatilization, nitrate (NO3−) leaching, and gaseous emissions produced by nitrification and denitrification, with nitrous oxide (N2O) and nitric oxide (NO) representing key reactive intermediates and dinitrogen gas (N2) representing the main terminal form of N returned to the atmosphere. Mechanistically, urine urea hydrolyses rapidly to NH4+ via urease, elevating soil pH and promoting NH3 loss. Nitrifiers—ammonia-oxidizing bacteria and archaea—oxidize NH4+ to NO2− and NO3− under aerobic conditions, while denitrifiers reduce NO3− to gaseous forms under anaerobic or intermittently wet conditions[55]; [56]. The balance between retention and loss is therefore strongly governed by soil moisture, oxygen availability, labile carbon, and the spatial distribution and intensity of excreta deposition. In addition, anaerobic ammonium oxidation (anammox) and dissimilatory nitrate reduction to ammonium (DNRA) may operate in waterlogged or compellingly reducing soil microsites beneath excreta patches. However, their quantitative contribution in temperate grazed pastures remains less well constrained than that of nitrification and denitrification.
Meta-analyses show that N2O emission factors from cattle excreta are generally lower than the IPCC default but highly variable, with a small number of high-flux events contributing disproportionately to annual emissions [57,58,59]. Patch-scale studies further reveal that concentrated urine loads create hotspots of nitrification and denitrification, while on free-draining soils NO3− leaching dominates during periods of high drainage and low plant [57], 60-62]. The NH3 volatilization from urine is influenced by temperature, wind, pH, and soil cation exchange capacity, which in turn is shaped by soil texture and clay mineralogy, and can be moderated by botanical composition. Foresentence, multispecies swards containing plantain and chicory, whose deeper rooting, lower herbage N concentrations, and bioactive secondary compounds (i.e., tannins) dilute urinary N, enhance plant N uptake, and thereby reduce NH3 losses [63,64,65,66]. Regionally, nitrate leaching tends to dominate in humid climates, NH3 volatilization in warm alkaline soils, and N2O remains the most climate-intensive loss pathway [56,67]. In the following sections, we examine these pathways at patch, paddock, and landscape scales and discuss how nutrition, plant composition, and grazing behavior can be leveraged to reduce N losses while maintaining productivity.
2.2. Ecological Perspectives on Nitrogen Cycling at the Pasture scale
At the pasture scale, nitrogen cycling emerges from interactions among soil microbes, plant functional groups, and the spatial patterning of excreta. Soil microorganisms mediate mineralization, nitrification, and denitrification, thereby regulating the balance between plant-available N, soil organic N storage, and gaseous losses. Plant functional composition and diversity, particularly the presence of legumes and deep-rooted forbs, determine how effectively these mineral N pools are captured and recycled within the sward [68]. Superimposed on these processes, excreta patches introduce localized N surpluses, creating biogeochemical hotspots that can either enhance productivity or intensify N losses, depending on soil conditions and management. Collectively, these ecological processes shape whether grazed pastures act as N sinks or sources and provide the mechanistic context for interpreting farm- and animal-scale NUE.
2.2.1. Soil Microbial Processes and Nitrogen Transformations
Soil microbes regulate N transformations in grazed systems through tightly coupled mineralization, nitrification, and denitrification pathways. For clarity, we refer to mineral nitrogen primarily as ammonium (NH4+) and nitrate (NO3−), and to gaseous losses as ammonia (NH3) and nitrous oxide (N2O). During mineralization, also referred to as ammonification, heterotrophic microbes decompose organic matter and convert organic N into ammonium (NH4+), with rates governed by substrate quality (e.g., C: N ratio and lignin or polyphenol content) and oil temperature and moisture [49]. Nitrification oxidizes NH4+ to nitrate (NO3−) via ammonia-oxidizing bacteria and archaea (e.g., Nitrosomonas) and nitrite-oxidizing bacteria (e.g., Nitrobacter), with process rates strongly modulated by soil pH, oxygen availability, and temperature [56]. Denitrification reduces NO3− to N2O and dinitrogen (N2) under anaerobic or intermittently saturated conditions through facultative anaerobes [74,75], such as Pseudomonas, with the N2O: N2 ratio controlled by oxygen status, the availability of labile carbon, and nitrate concentration [69]. Other nitrogen transformation pathways, such as anaerobic ammonium oxidation (anammox) and dissimilatory nitrate reduction to ammonium (DNRA), may also occur in saturated or strongly reducing soil microsites. Still, their relative importance in managed pastures is less well quantified than that of classical nitrification and denitrification.
In grazed pastures, these pathways are strongly shaped by spatial and temporal variability in soil moisture and carbon inputs. In particular, soil moisture modulates both mineralization and denitrification by controlling oxygen availability and substrate diffusion. Moisture fluctuations alter pathway dominance. Wrage-Mönnig et al. [70] reported that transient wetting events can shift N2O production from coupled nitrification–denitrification toward classical heterotrophic denitrification, particularly where nitrate and carbon accumulate. Integrated crop–livestock systems further modify microbial N dynamics by adding plant residues and cover crops with contrasting C: N ratios and residue chemistries, key components of substrate quality, thereby enhancing soil organic matter inputs, moderating the mineralization–immobilization balance, and synchronizing N release with forage uptake [52,71]. Over time, these microbial processes determine whether grazing systems accumulate soil organic N or leak reactive N through gaseous and leaching pathways [50,51] thereby linking pasture-scale ecology directly to whole-farm NUE outcomes.
2.2.2. Plant Functional Diversity, Legumes, and Biological Nitrogen Fixation
Plant functional composition shapes N cycling by determining biomass production, root distribution, litter quality, and the timing of N uptake. Legumes introduce new N to the system via symbiotic associations with Rhizobium, which reduces atmospheric N2 to ammonia through nitrogenase activity in root nodules. This biologically fixed N is incorporated into legume biomass and enters the soil N cycle through root exudation, senesced leaves, and root turnover, with fixation rates influenced by legume proportion, defoliation intensity, and soil mineral N levels [47,48]. In addition to symbiotic Rhizobium–legume associations, free-living and associative diazotrophs in the rhizosphere and soil surface can also contribute to N2 fixation. However, their quantitative importance in managed pastures is less well constrained. Deep-rooted forbs and grasses complement legumes by capturing NO3− and water from deeper soil layers, thereby reducing NO3− leaching risk and stabilizing seasonal forage production under variable rainfall [72,73].
In temperate grass–clover swards, biological N fixation can supply approximately 150–250 kg N ha−1 yr−1 when clover comprises about 20–30% of the sward and fertilizer N inputs remain low[48,74,75]. Under these conditions, mixtures can match or surpass the productivity of high-N fertilized grass monocultures receiving 200–250 kg N ha−1 yr−1 [76,77,78]. However, the ecological benefits of legumes are highly sensitive to management. High mineral N rates above about 100 kg N ha−1 yr−1 suppress clover, reduce nodule activity, and promote grass dominance, thereby favoring non-fixing species, diminishing biological N fixation (including both symbiotic and free-living diazotrophs), and increasing reliance on external N inputs [79]. At the same time, stocking density and grazing pressure alter legume persistence[80] while selective grazing, trampling, and competition fueled by urine-derived mineral N can reduce clover content and destabilize mixtures [81]; [82].
Modern grass–clover mixtures and multispecies swards offer management options to exploit functional diversity while moderating N surpluses. Cultivars with high stolon density and persistence can maintain 25–35% clover under rotational grazing and modest fertilizer N inputs (<50 kg ha−1 yr−1), thereby sustaining biological N fixation and limiting external N requirements [83]. Tactical spring fertilizer applications of 40–60 kg N ha−1 can boost early-season grass growth without permanently depressing clover or biological N fixation later in the season [84]. Moderate fertilizer strategies around 80 kg N ha−1 yr−1 may reduce clover-derived N by 30–40% but still increase total N supply by 15–20% [85], illustrating the trade-off between maximizing biological inputs and stabilizing forage yield. Economically, modern grass–clover mixtures can lower synthetic N fertilizer expenditure while maintaining similar or higher forage yields than fertilized grass monocultures because biological N fixation from clover can increase total N yield by up to 57% [86,87]. However, these gains come at the cost of more complex management to maintain species balance and control weeds, which can increase labor and knowledge requirements [88], and moderate mineral N strategies may reduce clover-derived N inputs by 30–40% [89]. Thus, economic viability depends on whether savings in fertilizer and improved environmental compliance outweigh these additional management demands, and is further shaped by market prices and policy incentives, such as area payments, that support grassland use [90]. Overall, plant functional diversity and legume-based biological N fixation can significantly enhance internal N cycling and soil N accumulation. Still, the net environmental and economic benefits ultimately depend on total system N loading, grazing intensity, and the persistence of legumes within the sward.
2.2.3. Excreta Patches as Hotspots and Their Spatial Patterning
Excreta patches create extreme nutrient heterogeneity in grazed pastures by concentrating N, carbon, and other nutrients into small areas relative to the paddock. Urine deposition can locally apply the equivalent ofmore than 1000 kg N ha−1, primarily as urea, to a small proportion of the pasture surface, while dung adds organic N and carbon with slower release dynamics [60]; [91] Following deposition, urea in urine is rapidly hydrolyzed to NH4+ by urease, causing short-lived spikes in soil pH and NH3 concentration. Subsequent nitrification converts NH4+ to NO3−, and under moist or anaerobic microsites, denitrification reduces NO3− to N2O and N2 [55,56]. These conditions make urine patches disproportionate contributors to N2O emissions and nitrate leaching at the paddock scale. Meta-analyses report N2O emission factors from cattle urine and dung generally in the range of 0.3–1.0% of excreted N, but values vary from about 0.07 to 5.9% depending on drainage, rainfall, and urine N load [92,99,100]
The balance between gaseous and leaching losses from patches depends strongly on soil physical conditions. On free-draining soils, up to 20–40% of urine N can be leached as NO3− during periods of high drainage and low plant uptake [60,93,94]. Where soils are wetter or poorly drained, denitrification processes dominate, increasing the fraction of N lost as N2O and N2 [56]. Ammonia volatilization from urine patches typically accounts for about 10–25% of excreted N and is influenced by temperature, wind speed, soil pH, and cation exchange capacity [63,64]. Botanical composition can mitigate these losses: multispecies swards containing plantain (Plantago lanceolata) and chicory (Cichorium intybus) dilute urine N concentration, alter urine chemistry, and reduce NH3 volatilization by approximately 30–50%, while also enhancing NO3− capture through deeper rooting and extended growth periods [95].
At the paddock scale, the cumulative effect of many urine and dung patches is a mosaic of nutrient-rich and nutrient-poor zones, with a relatively small proportion of the area receiving a large share of total N inputs (Figure 2). Overlapping urine and dung deposits can intensify N2O emissions and NO3− leaching by stacking labile carbon and high N in the same microsites [96,97]. Although the exact spatial arrangement of patches is strongly influenced by animal behavior, resting sites, and landscape features (considered in Section 2.4), excreta hotspots themselves represent the primary ecological interface where animal-derived N either supports plant productivity and soil organic matter formation or is lost to the broader environment. Understanding patch-scale biogeochemistry is therefore critical for designing grazing and pasture management strategies that improve paddock-scale N retention and reduce N2O, NH3, and NO3− losses. The amount and chemical form of N entering these hotspots are, in turn, governed by animal-level intake and partitioning, which are examined in Section 2.3.
2.3. Animal Nutrition Perspective on Nitrogen Partitioning and Use
The ecological processes described in Section 2.2 determine how N is transformed and retained within soil and plant pools. But the amount and chemical form of N entering excreta hotspots ultimately depend on animal-level intake, metabolism, and partitioning. From a nutritional perspective, N use efficiency is governed by the synchrony between rumen N supply and fermentable energy, the balance between dietary crude protein (CP) and metabolizable energy, and the regulation of hepatic urea production reflected in milk and blood urea nitrogen (Figure 3 here). This section examines how rumen N metabolism, diet composition, and urea-based indicators together shape the proportions of N retained in animal products versus excreted in urine and feces, thereby controlling the N loads delivered to the patches described in Section 2.2.3.
2.3.1. Rumen Nitrogen Metabolism and Synchrony
Interactions between dietary protein degradability, microbial capture capacity, and fermentable energy supply govern rumen N metabolism. Dietary protein and non-protein N are hydrolyzed to peptides, amino acids, and NH3-N by microbial proteases and deaminases [12]. Microbes assimilate NH3 using energy derived from carbohydrate fermentation to synthesize microbial protein, which is the primary amino acid source for the host [13]. Low NUE arises when NH3 production exceeds microbial demand, particularly in high-CP pastures where soluble protein outpaces available fermentable energy. Excess NH3 is absorbed across the rumen wall, converted to urea in the liver, and partitioned between recycling to the gut and urinary excretion [98].
Typical temperate pastures contain CP levels of 18–24%, frequently exceeding animal requirements. Keim and Anrique [99] showed that such surpluses elevate urinary N and depress NUE, especially when fermentable energy is limiting. Microbial growth plateaus at rumen NH3 concentrations of 5–8 mg dL−1; additional NH3 does not enhance microbial protein synthesis but increases urea production [100]. Conversely, NH3 < 2 mg dL−1 constrains microbial growth and fiber digestion, indicating a narrow optimal range that depends on diet type and microbial community composition [14]. Protein–energy synchrony improves microbial capture and reduces urinary N. Supplementing high-CP pasture with starch-rich feeds consistently reduces rumen NH3 accumulation and urinary N excretion [101,102]. Voglmeier et al. [95] demonstrated that inclusion of maize silage lowered excreta N by 18% and reduced N2O emission factors from 1.0% to 0.74–0.83%, linking rumen synchrony directly to downstream loss pathways. Timing further modifies outcomes: Ferreira et al. [103] found that split starch feeding, defined as dividing the daily starch supplement into two meals offered at different times of the day so that starch intake peaks coincide with peaks in rumen NH3 release, aligned peak NH3 availability with fermentable energy supply and reduced urinary N by 22% compared with providing the same starch dose all at once. Genetic variation also modulates responses. Cantalapiedra-Hijar et al.[104] showed that high-efficiency cows produced less NH3 and greater microbial protein per unit N intake on identical diets. Thus, rumen N metabolism depends on synchronizing protein degradability with fermentable energy, with dietary composition, feeding schedule, and host–microbiome interactions collectively determining how much N is captured in microbial protein versus excreted as urea that fuels the excreta hotspots described in Section 2.2.3.
2.3.2. Diet Composition, Intake, and Nitrogen Partitioning
Diet composition governs N intake and excretion by determining CP concentration, protein degradability, carbohydrate type, and amino acid supply. Reducing dietary CP from 18–20% to 14–16% typically lowers urinary N excretion by 15–30% without compromising productivity, provided energy supply and metabolizable protein meet requirements [37,45]. Doran et al. [106] observed that reducing concentrate CP from 18% to 15% lowered MUN by 25% and urinary N by 20% without affecting milk yield. Multiple well-designed trials report that moderate CP diets (~14.5%) can sustain milk yield and composition while reducing nitrogen excretion, especially urinary N, thereby improving NUE [114,115,116]. However, CP reductions have limits. Huhtanen and Hristov [45] reported a curvilinear response: NUE increases sharply as CP declines from 19% to 16%, but plateaus below 15%, where an insufficient total amino acid supply constrains milk protein yield. Lee et al. [107] documented that high-yielding cows (>35 kg d−1 milk) fed CP <15% experienced reduced milk protein and body condition loss, even when supplemented with rumen-protected amino acids, demonstrating that CP reductions below optimal thresholds compromise metabolizable protein supply rather than simply reducing surplus N (Figure 4).
Botanical composition also modifies N partitioning by altering forage quality and rumen fermentability. Diverse legume–forb swards generally enhance microbial protein synthesis and N retention. Dodd et al. [108] reported that cows grazing diverse swards increased milk N retention from 0.26 to 0.30 of N intake and produced 8–12% more milk than those on grass monoculture. However, these benefits diminish when pasture CP exceeds ~20% due to excessive N intake and elevated urea production [109]. Carbohydrate profile is equally influential. Hall and Huntington [110] showed that maize silage increases microbial protein flow by 15–20% compared with grass silage due to greater starch fermentation. Conversely, Ellis et al. [121] cautioned that very high starch levels (>30% of dry matter) depress fiber digestion and reduce overall N efficiency. Thus, optimal N partitioning requires integrating CP concentration, protein degradability, fermentable carbohydrate supply, and botanical diversity to reduce urinary N while maintaining milk and growth performance (Figure 4).
2.3.3. Indicators of Nitrogen Status: Milk and Blood Urea Nitrogen
Milk urea nitrogen (MUN) and blood urea nitrogen (BUN) reflect hepatic urea production and serve as integrative indicators of dietary protein–energy balance and urinary N excretion. Mechanistically, elevated rumen NH3 from excess degradable protein is absorbed, converted to urea in the liver, and equilibrates across blood, milk, and urine [99]. Kohn et al. ([111] demonstrated strong quantitative relationships, with BUN accounting for 50–70% of the variation in urinary N excretion across diets.
Genetic variation in urea metabolism offers opportunities for long-term improvement in NUE. Correa-Luna et al. [40] showed that cows selected for low MUN breeding values excreted 12% less urinary N and achieved 3–4% point improvements in NUE while consuming identical diets. Beatson et al. [112] similarly found heritable variation in MUN linked to rumen NH3 production and urea recycling efficiency. However, Richardson et al. [47] cautioned that reductions in MUN must be interpreted contextually; excessively low MUN may denote insufficient protein supply rather than improved efficiency, and environmental gains (5–8% reductions in urinary N) are generally smaller than those achieved through dietary manipulation (15–25%).
MUN threshold ranges are widely used to monitor herd-level protein balance. Zhao et al. [113] recommended 10–14 mg dL−1 for grazing dairy cows to balance production with environmental objectives. Conversely, Spek et al. [22] demonstrated that MUN–urinary N relationships vary with water intake, body weight, milk yield, and sampling time, explaining only 40–60% of the individual variation. Thus, MUN is most reliable for herd-level diagnostics rather than individual prediction. Sensor-based MUN monitoring is emerging as a tool for precision feeding. Lavery and Ferris [114] showed that inline MUN sensors reduced herd-average MUN by 15% and urinary N by 12% via dynamic CP adjustments. However, the environmental benefits of these nutritional adjustments depend on how the reduced urinary N is redistributed and transformed in the field through the excreta hotspots and soil processes [115,116,117] described in Section 2.2 and 2.4 Prevalent, MUN, and BUN provide valuable indicators of N status and support precision feeding, but they must be integrated with dietary, spatial, and soil-management strategies to achieve meaningful improvements in whole-system NUE.
2.4. Behavioral and Spatial Drivers of Nitrogen Return
The patterns of N partitioning described in Section 2.3 determine how much N is excreted in urine and feces, while animal behavior governs where this N is deposited across the landscape. Grazing circuits, resting-site preferences, and multispecies assemblages redistribute N from foraging areas to preferred lying, shading, and watering sites, thereby shaping the spatial arrangement of excreta hotspots characterized in Section 2.2.3. These behavioral and spatial processes control whether excreted N reinforces productive zones, creates chronic nutrient surpluses, or is lost via leaching and gaseous emissions.
2.4.1. Grazing Patterns, Resting Sites, and Excreta Distribution
Excreta deposition is mainly shaped by grazing behavior, with animals concentrating urine and dung in predictable zones influenced by diurnal rhythms, resource distribution, and social interactions. Cattle typically graze during early morning and late afternoon, then rest and ruminate near shade, water, or fence lines, depositing a disproportionate share of excreta in these areas [116,118]. This behavior creates persistent nutrient hotspots that elevate nitrate leaching and N2O emissions when N loads exceed the soil’s capacity to retain and transform reactive N. Betteridge et al. [119] showed that urination peaks during transitions between grazing and resting periods, amplifying deposition near water troughs and milking areas. Shade structures intensify spatial clustering. Carnevalli et al. [120] found that dairy heifers deposited 45% of dung in only 20% of the paddock area under silvopastoral shade. Carpinelli et al. [121] demonstrated that these concentrated dung zones increased subsequent crop N uptake by 15–25%, indicating potential agronomic benefits where hotspots coincide with target crop areas. However, White et al. [118] cautioned that N loading near water or shade often exceeds 500 kg N ha−1 yr−1, creating high-risk nitrate leaching and N2O hotspots that can offset productivity gains.
Urine–dung overlap further amplifies emissions. Auerswald et al. [122] found that overlapping patches produced 30–50% greater N2O emissions than separate patches because dung-supplied labile C enhances denitrification of urine-derived NO3−. Strategic resource placement can mitigate hotspot formation. Hirata et al. [123] reported that shifting water and shade redistributed resting areas by up to 50 m, reducing N deposition near streams by 20–30%. Nonetheless, behavioral inertia constrains effectiveness. Bailey et al. [124] observed that cattle continued to use familiar sites despite the availability of alternative shade, thereby limiting redistribution. Stocking density also matters; Schoenbach et al. [125] found that in small paddocks (<2 ha), high grazing pressure homogenized deposition regardless of resource placement, reducing the scope for spatial steering. Thus, excreta patterns result from behaviorally driven clustering around preferred resting sites, producing chronic nutrient hotspots superimposed on the soil and plant processes described in Section 2.2. Strategic design of shade, water, and grazing sequences can redistribute 10–30% of excreta and partially decouple resting sites from sensitive areas. Still, long-term effectiveness is constrained by herd familiarity, stocking density, and paddock structure.
2.4.2. Multispecies Grazing and Nitrogen Translocation
Grazing animals redistribute N across landscapes by consuming forage in nutrient-rich zones and excreting in preferred resting areas, creating cross-scale nutrient translocation patterns that influence vegetation structure and soil fertility. In mountain pastures, Svensk et al. [126] observed that Highland cattle grazed high-quality grass patches for 60–70% of their grazing time but deposited 30–40% of their excreta in Alnus viridis thickets used as shade, accelerating shrub expansion through localized nutrient enrichment. Riesch et al. [127] confirmed similar dynamics across multiple herbivore systems, with translocation distances from grazing to resting zones often exceeding 100 m and generating fertility gradients that shape plant community composition. Integrated crop–livestock systems may benefit from this redistribution. Lemaire et al. [72] and Carvalho et al. [53] demonstrated that manure-enriched resting zones increased soil organic matter and enhanced subsoil N retention. Carpinelli et al. [121] found 15–25% greater crop N uptake in areas previously used as cattle resting sites, suggesting the potential to use controlled resting patterns to transfer N intentionally. However, uncontrolled redistribution carries ecological risks. Schoenbach et al. [125] reported that nutrient-enriched resting sites became dominated by nitrophilous weeds, reducing pasture quality and grazing capacity. Environmental impact depends on the characteristics of receiving areas: translocation to well-drained soils increases leaching risk, whereas deposition in wetter, organic-rich areas may enhance denitrification and shift the balance of gaseous losses [127].
Multispecies grazing modifies spatial patterns through complementary dietary and resting behaviors. Dumont et al. [128] showed that mixed sheep–cattle grazing increased herbage utilization by 10–15% through niche partitioning but did not quantify N losses. Rook et al. [129] reported that mixed grazing reduced patchiness and improved sward uniformity, indicating potential for more even nutrient return. However, differences in excreta N concentration between species (often 20–40% variation) complicate predictions of system-level N fluxes. Thus, multispecies systems may enhance spatial nutrient capture and dilute hotspots but require careful stocking rate calibration and spatial planning to avoid unintended nutrient accumulation. Landscape-scale N translocation remains a significant uncertainty in N budgeting, and integrating GPS tracking, behavioral observation, and soil N flux measurements is needed to quantify redistribution pathways and to design spatially explicit N management strategies that align animal behavior with the ecological and nutritional principles outlined in Section 2.1–2.3.
2.5. Management Strategies to Enhance Nitrogen Use Efficiency and Reduce Losses
Building on the ecological, nutritional, and behavioral mechanisms outlined in Section 2.1–2.4, this section evaluates management strategies and tools to enhance NUE and reduce N losses in pasture-based ruminant systems.
2.5.1. Pasture Botanical Composition: Legumes and Multispecies Swards
Pasture botanical composition influences N cycling by altering N inputs, rooting depth distribution, forage nutritive quality, and animal N partitioning. Legume-based swards supply biologically fixed N via Rhizobium symbiosis, reduce reliance on synthetic fertilizer, and enhance forage protein [49]. Egan et al. [130] showed that grass–clover mixtures receiving ~150 kg N ha−1 yr−1 matched the yields of grass monocultures fertilized at 250 kg N ha−1 yr−1, while reducing N surpluses and nitrate leaching. Lüscher et al. [49] synthesized evidence showing reductions of 30–60% in fertilizer N requirements without compromising production. However, environmental outcomes depend on total system N loading. Ledgard et al. [131] documented that high-clover dairy systems exhibited leaching comparable to fertilized grass systems because total N inputs (BNF + excreta + deposition) exceeded removal. Eriksen et al. [132] similarly reported higher leaching in organic clover systems due to poor temporal synchrony between N release and crop demand. Legume persistence poses further challenges. Parsons et al. [133] showed that clover declined rapidly under high stocking rates due to selective grazing and shading, reducing long-term BNF benefits.
Multispecies swards incorporating plantain and chicory offer additional benefits. Jezequel et al. [134] found that diversifying swards while reducing fertilizer N by 25–50% maintained milk yield, improved herbage NUE from 35 to 50 kg herbage N per kg applied N, and reduced N surplus by 30–40 kg N ha−1 yr−1. Plantain may reduce urine N concentration via altered water intake and renal N handling [135]. However, Gardiner et al. [136] found no change in urine volume, suggesting a physiological rather than behavioral mechanism. Minneé et al. [154] observed reduced urinary N (37 -38%) and increased fecal N using 15N tracing, consistent with protein-sparing effects. However, Suter et al. [87] highlighted high variability in agronomic performance: multispecies swards underperformed when legume establishment failed, or forb palatability declined. Thus, botanical composition is a powerful but context-dependent lever requiring active management to maintain species persistence, synchronize N supply with demand, and integrate with grazing and dietary strategies for maximum NUE benefits.
2.5.2. Precision Feeding and Nitrogen Management
Precision feeding aligns dietary CP supply with animal requirements to reduce rumen NH3 accumulation, enhance microbial protein synthesis, and minimize urinary N losses. Reductions of 1–2% units of dietary CP consistently lower urinary N excretion by 8–15% without affecting milk yield, provided metabolizable protein and energy needs are met [45,102]. Keim and Anrique [36] emphasized combining high-protein pasture with high-energy/low-CP supplements, particularly maize silage, to improve protein–energy synchrony. Voglmeier et al. [96] demonstrated that combining high-protein grass (22–24% CP) with maize silage reduced excreta N by 18% and N2O emission factors from 1.0% to 0.74–0.83%. Yang et al. [137] showed that temporal feeding strategies modify metabolic responses, providing starch 4–6 hours after grazing improved synchrony between NH3 release and fermentable energy availability, reducing urinary N by 22% compared with 14% for simultaneous feeding. These findings highlight that both diet composition and timing influence N retention.
Precision tools use real-time indicators, such as MUN and BUN, to dynamically adjust diets. Studies have reported that sensor-guided dietary interventions achieved reductions in urinary nitrogen excretion ranging from 7% to over 40%, often linked to crude protein management [138,139]. However, practical adoption remains limited by infrastructure requirements, especially in grazing systems. Moreover, dietary improvements alone cannot address concentrated excreta deposition near shade and water; even with lower urine N concentration, hotspot formation may persist [16]. Thus, precision feeding reduces urinary N by 10–25% and N2O emissions by 10–20% through improved protein–energy balance but must be combined with pasture composition and spatial management to achieve whole-system gains. Future applications require integrating precision feeding with GPS-based excreta mapping and soil N models to capture interactions among diet, behavior, and spatial nutrient cycling.
2.5.3. Phytochemicals as Nitrogen Management Tools
Plant secondary metabolites, particularly condensed and hydrolysable tannins, offer a biochemical mechanism for improving nitrogen use by modulating rumen protein degradation, ammonia production, and N partitioning. Tannins form reversible protein–tannin complexes stable at rumen pH but dissociable in the acidic abomasum, reducing proteolysis and NH3 formation while increasing post-ruminal amino acid flow [17,18]. These shifts reduce ruminal NH3 surplus, lower hepatic urea synthesis, and shift N excretion from urine to feces, where N is less volatile and less susceptible to rapid nitrification and N2O formation [14]. Moderate tannin inclusion (2–4% DM) yields consistent reductions in urinary N. Zhang et al. [140] observed that quebracho and chestnut tannins lowered urinary N by 15–20% and increased fecal N by 10–15% without reducing milk yield. Aboagye et al. [141] found that hydrolysable tannins reduced methane by 12–18% and urinary N by 10–15% in beef cattle, indicating synergistic benefits for greenhouse gas mitigation. Thompson et al. [142] reported that ewes offered CT-rich willow (Salix Beagle) had 19% lower urinary N and 19% higher fecal N excretion than controls, clearly demonstrating a shift from urinary to fecal N. However, effects are strongly dose- and source-dependent. Excessive tannin levels (>5% DM) depress intake, fiber digestion, and microbial protein synthesis due to excessive protein binding. Da Silva Aguiar et al. (2023) emphasized the high variability across tannin types, dietary compositions, and animal species, which limits generalization. Soil-level responses remain largely unresolved. Carulla et al. [143] found fecal N from tannin-fed animals mineralized 20–30% more slowly, potentially reducing short-term N losses but also delaying plant N availability. Critically, only one study has directly linked tannin-induced urine chemistry changes to N2O fluxes: Luo et al. [144] reported a 25% reduction in N2O emissions from urine patches of tannin-supplemented cattle. However, high spatial variability obscured consistency, and no reduction in leaching was observed. Thus, tannins can reduce urinary N by 10–20% and alter excreta composition, potentially mitigating field-scale losses. However, high dose sensitivity, variable responses across systems, and limited soil-level evidence highlight the need for integrated pasture-scale validation linking rumen processes to soil N transformations.
2.5.4. Spatial Management: Shade, Water, and Grazing Design
Spatial management modifies animal movement and resting behavior to redistribute excreta and mitigate localized nitrogen hotspots. Since cattle preferentially rest near shade, water, and high-traffic corridors, manipulating the spatial configuration of these resources can influence where urine and dung accumulate [145,156,157,158]. Shade structures are particularly influential. Carnevalli et al. [131] reported that 45% of dung deposition occurred in only 20% of the paddock area under trees. This spatial clustering can be beneficial where additional fertility is desired—for example, in integrated crop–livestock rotations. Carpinelli et al. [121] showed that prior dung hotspots increased soybean N uptake by 15–25%, demonstrating the potential to use a resting-zone design as a nutrient-delivery tool. However, concentrated deposition near water or shade often produces excessive N loading (>500 kg N ha−1 yr−1), increasing nitrate leaching and N2O emissions [118]. Overlapping urine and dung deposition increases N2O emissions by 30–50% due to enhanced denitrification from combined labile C and a high inorganic N supply [157]
Strategic placement of water troughs or shade can shift resting areas by 20–30%, reducing N loading in sensitive zones such as riparian buffers [123]. Conversely, cattle exhibit strong site fidelity. Bailey et al. [146] found that herds repeatedly returned to familiar resting sites despite the presence of new shade or water, limiting the potential for redistribution. The grazing method also influences excreta patterns. Rotational and strip grazing reduce time spent near shade relative to the grazed area, thereby flattening spatial gradients. Betteridge et al. [119] documented reductions of 15–25% in excreta clustering under short-interval rotations. However, total N inputs remain unchanged, and hotspots can shift to gateways and fence lines. Implementation barriers include the costs of infrastructure and labor, as well as variable success across landscapes. Overall, spatial management can redistribute 10–30% of excreta and mitigate high-risk hotspots, but behavioral inertia, paddock structure, and logistical constraints limit efficiency. Integrating spatial design with dietary strategies and pasture composition is essential for meaningful gains in N cycling at the paddock scale.
2.5.5. Technologies and Decision-Support Tools
Sensor technologies and decision-support tools enable integration of animal behavior, pasture dynamics, and nitrogen fluxes to improve real-time N management. GPS tracking and accelerometers quantify spatial use, grazing–resting cycles [147] and inferred urination and defecation events, allowing construction of spatially explicit N-return maps [27,148]. Hassan-Vásquez et al. [27] showed that in a Mediterranean dehesa paddock, two dung “hotspot” plots representing only 2.2% of the sampled area accumulated 24.3% of droppings, and 42.6% of feces were deposited within 8.7% of the paddock near the water trough, spatial clustering that models must represent to predict N losses accurately. Decision-support systems linking intake, N excretion, soil processes, and management variables provide practical tools. Overseer, Kankanamge et al. [149] and Giakoumatos et al [164] recommended integrating animal N intake, excreta N, paddock-scale deposition assumptions, and soil N transformations to predict nitrate leaching. However, Cichota et al. [150] demonstrated that assuming a uniform distribution of excreta can underestimate losses because real-world deposition is highly clustered.
More advanced modelling approaches combine GPS-derived grazing intensity and soil moisture data with process-based models (e.g., APSIM). Studies showed that incorporating spatial grazing patterns improved predictions of animal grazing [151] pasture growth [152] and N leaching [27]. Inline milk MUN sensors enable adaptive dietary adjustments [114]. Integrating MUN monitoring with GPS excreta maps could link protein–energy balance with spatial N-return risk, but such integration remains experimental. Barriers include high cost, complex data processing, paddock-specific calibration requirements, and the need for secure digital infrastructure. Policy-driven adoption of decision-support tools is expanding. In New Zealand, regulatory N caps (30–50 kg N ha−1 yr−1) prompted adoption of modelling tools and stocking-rate adjustments, resulting in 15–30% reductions in nitrate leaching within five years [153]. However, compliance costs raise equity concerns for smaller farms. Collectively, technologies can improve representation of spatial N cycling and inform targeted interventions, but complexity, costs, and scalability limitations constrain widespread use. Simplified, affordable systems integrating GPS, MUN, and soil data remain a critical research frontier for operationalizing precision N management in grazing systems.
2.6. Integrating Ecological and Animal Science Perspectives
Integrating ecological and animal-nutrition perspectives requires viewing grazing animals simultaneously as metabolically constrained organisms and as mobile vectors of N across heterogeneous landscapes. From the animal side, rumen N metabolism, dietary CP concentration, protein–energy synchrony, and amino acid balance determine N intake, NUE, and the urine: feces N ratio [13,20]. From an ecological perspective, soil texture, hydrology, plant functional composition, and excreta patch dynamics govern how returned N is transformed, retained, or lost, with urine patches acting as transient hotspots that drive N2O emissions and nitrate leaching [10,12]. Legume-based and multispecies swards exemplify this coupling. Biological N fixation alters N inputs, while changes in forage quality and plant secondary metabolite profiles affect intake, N metabolism, and excreta composition, feeding back to soil microbial processes and N losses [49,134]. Plantain-rich swards can reduce effective CP degradability, lower urinary N concentration, and dilute individual urine patches via altered urination patterns, potentially decreasing N2O emissions per patch [135,154], although net effects depend on total patch area and compensatory mineralization dynamics that remain poorly resolved [136]. Similarly, tannin-rich shrubs or trees influence both internal N partitioning and external spatial patterns by modifying grazing behavior, shade use, and resting sites, concentrating excreta under canopies and enhancing tree growth but potentially increasing leaching risks near waterways [120,126].
An integrated framework that couples (i) animal NUE models predicting N partitioning from diet and animal traits [13,103]; (ii) pasture growth and N cycling models capturing BNF, mineralization, and loss pathways [155] and (iii) spatial behavior models predicting excreta deposition patterns from movement and resource distribution [156] is required. Classical pasture N-cycling models, such as that of Scholefield et al. [157], showed that matching fertilizer N inputs with soil mineralization and site characteristics is critical for limiting NH3 volatilization, denitrification, and leaching while improving N use efficiency in grazed beef systems. Gregorini et al. [156], together with their mechanistic and dynamic model of a grazing dairy cow (MINDY), further emphasized that explicitly representing diurnal feeding motivation, sward structure, and spatial–temporal patterns of grazing and urination is essential for linking internal metabolic state and individual foraging decisions to paddock-scale N loading, leaching, and gaseous losses. Coupled models can evaluate scenarios such as adding clover and plantain, lowering supplement CP, relocating shade, or integrating tannin-rich forages, and quantify consequences for animal performance, N surpluses, and emissions. However, they require extensive calibration data and must capture feedback among stocking density, botanical composition, dietary CP, and excreta N loading, which can drive rapid system change if mismanaged [8,38,132]. Developing and simplifying such integrated models for practical use remains a major research priority.
2.7. Research Gaps and Future Directions
Key knowledge gaps continue to limit progress toward nitrogen-efficient pasture-based ruminant systems. A significant gap concerns how individual-animal variability scales to paddock- and farm-level N dynamics. Grazing animals differ in behavior, rumen microbiota, N metabolism, and MUN/BUN, but how these differences translate into spatial excreta patterns and N fluxes under real grazing conditions remains poorly quantified. Although genetic variation in MUN explains part of the variation in urinary N excretion [39], it is unknown whether low-MUN animals also differ in excreta spatial distribution or diurnal deposition patterns. Integrative studies linking individual-level monitoring (GPS, activity sensors, MUN/BUN, intake) with excreta mapping and soil N measurements are needed to determine how animal heterogeneity shapes whole-system N losses. The long-term ecosystem effects of tannins and other phytochemicals also remain uncertain. Short-term experiments show that tannins shift N from urine to feces and reduce N2O emissions [112]; [141], but multi-year impacts on soil organic N pools, microbial communities, and botanical composition are largely unmeasured. Only one study has quantified N2O emissions from tannin-modified urine patches [158], and no multi-year datasets exist to determine whether tannin-based strategies produce durable reductions in N losses or merely delay mineralization.
Scaling N processes across patch, paddock, farm, and watershed levels is another constraint. Patch-scale studies (<1 m2) provide mechanistic insights [9], but management requires validated spatially explicit N budgets. Integrating patch-scale fluxes with GPS-derived behavior, eddy-covariance N2O measurements, and hydrological models [159]; [160] demands multi-scale measurement campaigns across soil types and climates. Quantitative evidence for N cycling in multispecies grazing systems is also scarce. While complementary spatial use by cattle, sheep, and goats is documented [126,127], species-specific effects on excreta N composition, deposition patterns, and soil N transformations are poorly resolved. Finally, decision-support tools that integrate animal, pasture, and spatial processes remain data-intensive and are primarily used in research settings [154]. Simplifying model inputs, validating outputs under diverse grazing systems, and aligning tools with policy frameworks is likely to be essential for broader adoption and measurable reductions in N losses [152]
3. Conclusions
Pasture-based ruminant systems tightly couple animal N metabolism with soil–plant–microbial N cycling, and inefficiencies in these linked processes propagate as elevated reactive N losses. This synthesis indicates that low nitrogen use efficiency (NUE) arises mainly from crude protein supply exceeding fermentable energy, which increases ruminal NH3 production, hepatic urea synthesis, and urinary N excretion; from behaviorally driven excreta clustering that creates biogeochemical hotspots responsible for a disproportionate share of nitrate leaching and N2O emissions; and from temporal and spatial asynchrony between N inputs (fertilizer, biological N fixation, excreta) and plant uptake, which enhances soil mineral N accumulation and loss. Integrated management that combines precision feeding, legume- and forb-rich multispecies swards, targeted use of plant secondary metabolites such as tannins, and spatial manipulation of shade, water, and grazing allocation can increase NUE from approximately 20–25% to above 30% without reducing animal performance. Further research is recommended on multi-year, multi-scale experiments that jointly quantify rumen N dynamics, excreta chemistry, spatial deposition patterns, and soil N fluxes under realistic grazing conditions, with individual-animal monitoring (intake, MUN/BUN, microbiome, GPS-based behavior) explicitly linked to paddock- and farm-scale N budgets. Moreover, long-term field trials are needed to evaluate whether tannin- and phytochemical-based interventions produce sustained reductions in urinary N and N2O emissions while maintaining or enhancing soil organic N pools, microbial community function, and pasture botanical composition. Additional studies are needed to develop, calibrate, and simplify spatially explicit models that couple animal nutrition, pasture growth, and hydrological N transport into decision-support tools that are technically and economically accessible to small- and medium-scale grazing enterprises.
Funding
This research received no external funding.
Acknowledgments
The author gratefully acknowledges Fred Provenza for his thorough review of the manuscript and for providing valuable insights that strengthened the conceptual framework. Sincere appreciation is also extended to Chandra Man Rai and Goodluck Jonas for their insightful discussions and constructive suggestions during the development of this work.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Abbreviation | Full name / description |
| APSIM | Agricultural Production Systems simulator (crop–soil simulation model) |
| BNF | Biological nitrogen fixation |
| BUN | Blood urea nitrogen |
| CAB | Commonwealth Agricultural Bureaux (historical name associated with CAB International) |
| CABI | Centre for Agriculture and Bioscience International |
| CP | Crude protein |
| CT | Condensed tannins |
| DM | Dry matter |
| DNRA | Dissimilatory nitrate reduction to ammonium |
| DVE | Truly digestible protein in the small intestine (DVE protein evaluation system) |
| GHG | Greenhouse gas(es) |
| GPS | Global Positioning System |
| IPCC | Intergovernmental Panel on Climate Change |
| MINDY | Mechanistic, dynamic model of a grazing dairy cow |
| MUN | Milk urea nitrogen |
| N | Nitrogen |
| Nr | Reactive nitrogen |
| N2 | Dinitrogen (molecular nitrogen gas) |
| N2O | Nitrous oxide |
| NH3 | Ammonia |
| NH4+ | Ammonium |
| NO | Nitric oxide |
| NO2− | Nitrite |
| NO3− | Nitrate |
| NUE | Nitrogen use efficiency |
| OEB2010 | Degraded protein balance in the rumen (2010 revision of the Dutch OEB system) |
References
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the Nitrogen Cycle: Recent Trends, Questions, and Potential Solutions. Science (1979) 2008, 320, 889–892.
- Sutton, M. European Nitrogen Assessment. 2010.
- Uwizeye, N.F.; Boer, A.; Opio, I.J.M.; Schulte, C.; Falcucci, R.P.O.;; Et Al, A. Nitrogen Emissions along Global Livestock Supply Chains.. [CrossRef]
- Haynes, R.J.; Williams, P.H. Nutrient Cycling and Soil Fertility in the Grazed Pasture Ecosystem. Advances in Agronomy 1993, 49, 119–199. [CrossRef]
- Powell, J.M.; Gourley, C.J.P.; Rotz, C.A.; Weaver, D.M. Nitrogen Use Efficiency: A Potential Performance Indicator and Policy Tool for Dairy Farms. Environ Sci Policy 2010, 13, 217–228. [CrossRef]
- Liu, J.; You, L.; Amini, M.; Obersteiner, M.; Herrero, M.; Zehnder, A.J.B.; Yang, H. A High-Resolution Assessment on Global Nitrogen Flows in Cropland. Proc Natl Acad Sci U S A 2010, 107, 8035–8040.
- Guy, C.; Hennessy, D.; Gilliland, T.J.; Coughlan, F.; McCarthy, B. Growth, Morphology and Biological Nitrogen Fixation Potential of Perennial Ryegrass-White Clover Swards throughout the Grazing Season. J Agric Sci 2018, 156, 188–199. [CrossRef]
- Schils, R.L.M.; Verhagen, A.; Aarts, H.F.M.; Šebek, L.B.J. A Farm Level Approach to Define Successful Mitigation Strategies for GHG Emissions from Ruminant Livestock Systems. Nutr Cycl Agroecosyst 2005, 71, 163–175.
- Nitrogen in Current European Policies Available online: https://elibrary.ru/item.asp?id=22420754 (accessed on 28 November 2025).
- Selbie, D.R.; Buckthought, L.E.; Shepherd, M.A. The Challenge of the Urine Patch for Managing Nitrogen in Grazed Pasture Systems. Advances in Agronomy 2015, 129, 229–292. [CrossRef]
- Bouwman, L.; Goldewijk, K.K.; Van Der Hoek, K.W.; Beusen, A.H.W.; Van Vuuren, D.P.; Willems, J.; Rufino, M.C.; Stehfest, E. Exploring Global Changes in Nitrogen and Phosphorus Cycles in Agriculture Induced by Livestock Production over the 1900-2050 Period. Proc Natl Acad Sci U S A 2013, 110, 20882–20887. [CrossRef]
- Marsden, K.A.; Jones, D.L.; Chadwick, D.R. The Urine Patch Diffusional Area: An Important N2O Source? Soil Biol Biochem 2016, 92, 161–170. [CrossRef]
- Dijkstra, J.; France, J.; Ellis, J.L.; Strathe, A.B.; Kebreab, E.; Bannink, A. Production Efficiency of Ruminants: Feed, Nitrogen and Methane. Sustainable animal agriculture 2013, 10–25. [CrossRef]
- Hristov, A.N.; Bannink, A.; Crompton, L.A.; Huhtanen, P.; Kreuzer, M.; McGee, M.; Nozière, P.; Reynolds, C.K.; Bayat, A.R.; Yáñez-Ruiz, D.R.; et al. Invited Review: Nitrogen in Ruminant Nutrition: A Review of Measurement Techniques. J Dairy Sci 2019, 102, 5811–5852. [CrossRef]
- Chen, P.; Li, Y.; Wang, M.; Shen, Y.; Liu, M.; Xu, H.; Ma, N.; Cao, Y.; Li, Q.; Abdelsattar, M.M.; et al. Optimizing Dietary Rumen-Degradable Starch to Rumen-Degradable Protein Ratio Improves Lactation Performance and Nitrogen Utilization Efficiency in Mid-Lactating Holstein Dairy Cows. Front Vet Sci 2024, 11, 1330876.
- Dijkstra, J.; Reynolds, C.K.; Kebreab, E.; Bannink, A.; Ellis, J.L.; France, J.; van Vuuren, A.M. Challenges in Ruminant Nutrition: Towards Minimal Nitrogen Losses in Cattle. Energy and protein metabolism and nutrition in sustainable animal production 2013, 47–58. [CrossRef]
- Min, B.R.; Barry, T.N.; Attwood, G.T.; McNabb, W.C. The Effect of Condensed Tannins on the Nutrition and Health of Ruminants Fed Fresh Temperate Forages: A Review. Anim Feed Sci Technol 2003, 106, 3–19. [CrossRef]
- Waghorn, G. Beneficial and Detrimental Effects of Dietary Condensed Tannins for Sustainable Sheep and Goat Production—Progress and Challenges. Anim Feed Sci Technol 2008, 147, 116–139. [CrossRef]
- Baker, S.; Lynch, M.B.; Godwin, F.; Boland, T.M.; Evans, A.C.O.; Murphy, P.N.C.; Kelly, A.K.; Sheridan, H. A Comparison of the Nutritive Value of Perennial Ryegrass, Perennial Ryegrass and White Clover and Multispecies-Based Farmlet Systems. Grassland Research 2023, 2, 129–139. [CrossRef]
- Hristov, A.N.; Bannink, A.; Crompton, L.A.; Huhtanen, P.; Kreuzer, M.; McGee, M.; Nozière, P.; Reynolds, C.K.; Bayat, A.R.; Yáñez-Ruiz, D.R.; et al. Invited Review: Nitrogen in Ruminant Nutrition: A Review of Measurement Techniques Nure N Emissions in the Context of Feed Composition and Ruminant N Metabolism. J Dairy Sci 2019, 102, 5811–5852. [CrossRef]
- Jonker, J.S.; Kohn, R.A.; Erdman, R.A. Using Milk Urea Nitrogen to Predict Nitrogen Excretion and Utilization Efficiency in Lactating Dairy Cows. J Dairy Sci 1998, 81, 2681–2692. [CrossRef]
- Spek, J.W.; Dijkstra, J.; Van Duinkerken, G.; Bannink, A. A Review of Factors Influencing Milk Urea Concentration and Its Relationship with Urinary Urea Excretion in Lactating Dairy Cattle. J Agric Sci 2013, 151, 407–423. [CrossRef]
- Lavery, A.; Ferris, C.P. Proxy Measures and Novel Strategies for Estimating Nitrogen Utilisation Efficiency in Dairy Cattle. Animals 2021, Vol. 11, Page 343 2021, 11, 343. [CrossRef]
- Selbie, D.R.; Buckthought, L.E.; Shepherd, M.A. The Challenge of the Urine Patch for Managing Nitrogen in Grazed Pasture Systems. Advances in Agronomy 2015, 129, 229–292. [CrossRef]
- Orwin, K.H.; Bertram, J.E.; Clough, T.J.; Condron, L.M.; Sherlock, R.R.; O’Callaghan, M. Short-Term Consequences of Spatial Heterogeneity in Soil Nitrogen Concentrations Caused by Urine Patches of Different Sizes. Applied Soil Ecology 2009, 42, 271–278. [CrossRef]
- Holatko, J.; Hammerschmiedt, T.; Kintl, A.; Mustafa, A.; Naveed, M.; Baltazar, T.; Latal, O.; Skarpa, P.; Ryant, P.; Brtnicky, M. Co-Composting of Cattle Manure with Biochar and Elemental Sulphur and Its Effects on Manure Quality, Plant Biomass and Microbiological Characteristics of Post-Harvest Soil. Front Plant Sci 2022, 13, 1004879.
- Hassan-Vásquez, J.A.; Maroto-Molina, F.; Guerrero-Ginel, J.E. GPS Tracking to Monitor the Spatiotemporal Dynamics of Cattle Behavior and Their Relationship with Feces Distribution. Animals 2022, 12, 2383.
- Valbuena, D.; Erenstein, O.; Homann-Kee Tui, S.; Abdoulaye, T.; Claessens, L.; Duncan, A.J.; Gérard, B.; Rufino, M.C.; Teufel, N.; van Rooyen, A.; et al. Conservation Agriculture in Mixed Crop–Livestock Systems: Scoping Crop Residue Trade-Offs in Sub-Saharan Africa and South Asia. Field Crops Res 2012, 132, 175–184. [CrossRef]
- Rivera, J.E.; Chará, J. CH4 and N2O Emissions From Cattle Excreta: A Review of Main Drivers and Mitigation Strategies in Grazing Systems. Front Sustain Food Syst 2021, 5, 657936.
- Rawal, S.; Kaur, H.; Bhathan, S.; Mittal, D.; Kaur, G.; Ali, S.A. Ruminant Gut Microbiota: Interplay, Implications, and Innovations for Sustainable Livestock Production. 2024, 205–228. [CrossRef]
- de Queiroz, G.M.; Moreira, T.S.; Salgado, H.; Moreira, R.R.D.; Utrera, S.H.; Martins, C.H.G.; Pietro, R. Nitrogen Metabolism and Excretion in Holstein Dairy Heifers Fed Different Levels of Metabolizable Protein. 2024. [CrossRef]
- De Vries, W.; Kros, J.; Kroeze, C.; Seitzinger, S.P. Assessing Planetary and Regional Nitrogen Boundaries Related to Food Security and Adverse Environmental Impacts. Curr Opin Environ Sustain 2013, 5, 392–402. [CrossRef]
- Schulte-Uebbing, L.F.; Beusen, A.H.W.; Bouwman, A.F.; de Vries, W. From Planetary to Regional Boundaries for Agricultural Nitrogen Pollution. Nature 2022 610:7932 2022, 610, 507–512. [CrossRef]
- Romera, A.J.; Cichota, R.; Beukes, P.C.; Gregorini, P.; Snow, V.O.; Vogeler, I. Combining Restricted Grazing and Nitrification Inhibitors to Reduce Nitrogen Leaching on New Zealand Dairy Farms. J Environ Qual 2017, 46, 72–79. [CrossRef]
- Akert, F.S.; Dorn, K.; Frey, H.; Hofstetter, P.; Berard, J.; Kreuzer, M.; Reidy, B. Farm-Gate Nutrient Balances of Grassland-Based Milk Production Systems with Full- or Part-Time Grazing and Fresh Herbage Indoor Feeding at Variable Concentrate Levels. Nutr Cycl Agroecosyst 2020, 117, 383–400.
- Keim, J.P.; Anrique, R. O NUTRITIONAL STRATEGIES TO IMPROVE NITROGEN USE EFFICIENCY BY GRAZING DAIRY COWS. Chil J Agric Res 71.
- Beltran, I.E.; Tellez, D.; Cabanilla, J.; Balocchi, O.; Arias, R.; Keim, J.P. Nitrogen Use Efficiency and Partitioning of Dairy Heifers Grazing Perennial Ryegrass (Lolium Perenne L.) or Pasture Brome (Bromus Valdivianus Phil.) Swards during Spring. Agronomy 2022, Vol. 12, Page 1953 2022, 12, 1953. [CrossRef]
- Castillo, A.R.; Kebreab, E.; Beever, D.E.; France, J. A Review of Efficiency of Nitrogen Utilisation in lactating Dairy Cows and Its Relationship with environmental Pollution. J Anim Feed Sci 2000, 9, 1–32. [CrossRef]
- Dijkstra, J.; Oenema, O.; van Groenigen, J.W.; Spek, J.W.; van Vuuren, A.M.; Bannink, A. Diet Effects on Urine Composition of Cattle and N2O Emissions. animal 2013, 7, 292–302. [CrossRef]
- Correa-Luna, M.; Donaghy, D.; Kemp, P.; Schutz, M.; López-Villalobos, N. Nitrogen Use Efficiency and Excretion in Grazing Cows with High and Low Milk Urea Nitrogen Breeding Values. Sustainability 2021, Vol. 13, Page 9827 2021, 13, 9827.
- Schils, R.L.M.; Verhagen, A.; Aarts, H.F.M.; Šebek, L.B.J. A Farm Level Approach to Define Successful Mitigation Strategies for GHG Emissions from Ruminant Livestock Systems. Nutr Cycl Agroecosyst 2005, 71, 163–175.
- Powell, J.M.; Gourley, C.J.P.; Rotz, C.A.; Weaver, D.M. Nitrogen Use Efficiency: A Potential Performance Indicator and Policy Tool for Dairy Farms. Environ Sci Policy 2010, 13, 217–228. [CrossRef]
- Nitrogen in Current European Policies Available online: https://www.elibrary.ru/item.asp?id=22420754 (accessed on 28 November 2025).
- Van Duinkerken, G.; Blok, M.C.; Bannink, A.; Cone, J.W.; Dijkstra, J.; Van Vuuren, A.M.; Tamminga, S. Update of the Dutch Protein Evaluation System for Ruminants: The DVE/OEB2010 System. J Agric Sci 2011, 149, 351–367. [CrossRef]
- Huhtanen, P.; Hristov, A.N. A Meta-Analysis of the Effects of Dietary Protein Concentration and Degradability on Milk Protein Yield and Milk N Efficiency in Dairy Cows. J Dairy Sci 2009, 92, 3222–3232. [CrossRef]
- Beatson, P.R.; Meier, S.; Cullen, N.G.; Eding, H. Genetic Variation in Milk Urea Nitrogen Concentration of Dairy Cattle and Its Implications for Reducing Urinary Nitrogen Excretion. animal 2019, 13, 2164–2171. [CrossRef]
- Richardson, C.M.; Amer, P.R.; Quinton, C.; Crowley, J.; Hely, F.S.; van den Berg, I.; Pryce, J.E. Reducing Greenhouse Gas Emissions through Genetic Selection in the Australian Dairy Industry. J Dairy Sci 2022, 105, 4272–4288. [CrossRef]
- Ledgard, S.F.; Sprosen, M.S.; Penno, J.W.; Rajendram, G.S. Nitrogen Fixation by White Clover in Pastures Grazed by Dairy Cows: Temporal Variation and Effects of Nitrogen Fertilization. Plant Soil 2001, 229, 177–187.
- Lüscher, A.; Mueller-Harvey, I.; Soussana, J.F.; Rees, R.M.; Peyraud, J.L. Potential of Legume-Based Grassland–Livestock Systems in Europe: A Review. Grass and Forage Science 2014, 69, 206–228. [CrossRef]
- Booth, M.S.; Stark, J.M.; Rastetter, E. CONTROLS ON NITROGEN CYCLING IN TERRESTRIAL ECOSYSTEMS: A SYNTHETIC ANALYSIS OF LITERATURE DATA. Ecol Monogr 2005, 75, 139–157. [CrossRef]
- Conant, R.T.; Cerri, C.E.P.; Osborne, B.B.; Paustian, K. Grassland Management Impacts on Soil Carbon Stocks: A New Synthesis. Ecological Applications 2017, 27, 662–668. [CrossRef]
- Soussana, J.F.; Lemaire, G. Coupling Carbon and Nitrogen Cycles for Environmentally Sustainable Intensification of Grasslands and Crop-Livestock Systems. Agric Ecosyst Environ 2014, 190, 9–17. [CrossRef]
- de Faccio Carvalho, P.C.; Anghinoni, I.; de Moraes, A.; de Souza, E.D.; Sulc, R.M.; Lang, C.R.; Flores, J.P.C.; Terra Lopes, M.L.; da Silva, J.L.S.; Conte, O.; et al. Managing Grazing Animals to Achieve Nutrient Cycling and Soil Improvement in No-till Integrated Systems. Nutr Cycl Agroecosyst 2010, 88, 259–273.
- Burchill, W.; Li, D.; Lanigan, G.J.; Williams, M.; Humphreys, J. Interannual Variation in Nitrous Oxide Emissions from Perennial Ryegrass/White Clover Grassland Used for Dairy Production. Glob Chang Biol 2014, 20, 3137–3146. [CrossRef]
- Mazzetto, A.M.; Barneze, A.S.; Feigl, B.J.; Van Groenigen, J.W.; Oenema, O.; Cerri, C.C. Temperature and Moisture Affect Methane and Nitrous Oxide Emission from Bovine Manure Patches in Tropical Conditions. Soil Biol Biochem 2014, 76, 242–248. [CrossRef]
- WHITEHEAD, D.C.; RAISTRICK, N. Ammonia Volatilization from Five Nitrogen Compounds Used as Fertilizers Following Surface Application to Soils. Journal of Soil Science 1990, 41, 387–394. [CrossRef]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous Oxide Emissions from Soils: How Well Do We Understand the Processes and Their Controls? Philosophical Transactions of the Royal Society B: Biological Sciences 2013, 368. [CrossRef]
- Shakoor, A.; Shahzad, S.M.; Chatterjee, N.; Arif, M.S.; Farooq, T.H.; Altaf, M.M.; Tufail, M.A.; Dar, A.A.; Mehmood, T. Nitrous Oxide Emission from Agricultural Soils: Application of Animal Manure or Biochar? A Global Meta-Analysis. J Environ Manage 2021, 285, 112170. [CrossRef]
- Purath, E.; Sajeev, M.; Winiwarter, W.; Amon, B. Greenhouse Gas and Ammonia Emissions from Different Stages of Liquid Manure Management Chains: Abatement Options and Emission Interactions.. [CrossRef]
- Soares, J.R.; Souza, B.R.; Mazzetto, A.M.; Galdos, M. V; Chadwick, D.R.; Campbell, E.E.; Deepak, ·; · J.; Oliveira, J.C.; Monteiro, L.A.; et al. Mitigation of Nitrous Oxide Emissions in Grazing Systems through Nitrification Inhibitors: A Meta-Analysis. Nutr Cycl Agroecosyst 2023, 125, 359–377. [CrossRef]
- Di, H.J.; Cameron, K.C. Nitrate Leaching in Temperate Agroecosystems: Sources, Factors and Mitigating Strategies. Nutr Cycl Agroecosyst 2002, 64, 237–256.
- Selbie, D. The Fate of Nitrogen in an Animal Urine Patch as Affected by Urine Nitrogen Loading Rate and the Nitrification Inhibitor Dicyandiamide 2014.
- Cameron, K.C.; Di, H.J.; Moir, J.L. Nitrogen Losses from the Soil/Plant System: A Review. Annals of Applied Biology 2013, 162, 145–173. [CrossRef]
- Sommer, S.G.; Schjoerring, J.K.; Denmead, O.T. Ammonia Emission from Mineral Fertilizers and Fertilized Crops. Advances in Agronomy 2001, 82, 557–622. [CrossRef]
- Misselbrook, T.H.; Powell, J.M.; Broderick, G.A.; Grabber, J.H. Dietary Manipulation in Dairy Cattle: Laboratory Experiments to Assess the Influence on Ammonia Emissions. J Dairy Sci 2005, 88, 1765–1777. [CrossRef]
- Zhu, Z.; Dong, H.; Zhou, Z. Ammonia and Greenhouse Gas Emissions from a Dairy Cattle Barn with a Daily Manure Collection System. Trans ASABE 2012, 55, 1959–1964. [CrossRef]
- D’Urso, P.R.; Arcidiacono, C.; Valenti, F.; Cascone, G. Assessing Influence Factors on Daily Ammonia and Greenhouse Gas Concentrations from an Open-Sided Cubicle Barn in Hot Mediterranean Climate. Animals 2021, Vol. 11, Page 1400 2021, 11, 1400. [CrossRef]
- Bouwman, A.F.; Boumans, L.J.M.; Batjes, N.H. Emissions of N2O and NO from Fertilized Fields: Summary of Available Measurement Data. Global Biogeochem Cycles 2002, 16, 6–1. [CrossRef]
- Aljazairi, S.; Ribas, A.; Llurba, R.; Ferrio, J.P.; Voltas, J.; Nogués, S.; Sebastiá, M.T. Sown Diversity Effects on the C and N Cycle and Interactions with Fertilization. Agronomy 2025, 15, 287.
- Arat, S.; Bullerjahn, G.S.; Laubenbacher, R. A Network Biology Approach to Denitrification in Pseudomonas Aeruginosa. PLoS One 2015, 10, e0118235. [CrossRef]
- Wrage-Mönnig, N.; Horn, M.A.; Well, R.; Müller, C.; Velthof, G.; Oenema, O. The Role of Nitrifier Denitrification in the Production of Nitrous Oxide Revisited. Soil Biol Biochem 2018, 123, A3–A16. [CrossRef]
- Lemaire, G.; Franzluebbers, A.; Carvalho, P.C. de F.; Dedieu, B. Integrated Crop–Livestock Systems: Strategies to Achieve Synergy between Agricultural Production and Environmental Quality. Agric Ecosyst Environ 2014, 190, 4–8. [CrossRef]
- Stanciu, A. Ștefania Sustainable Forage Yield Improvement through Perennial Grass–Legume Intercropping in Degraded Grasslands. Turkish Journal of Agriculture and Forestry 2025, 49, 215–241. [CrossRef]
- Hayes, R.C.; Ara, I.; Badgery, W.B.; Culvenor, R.A.; Haling, R.E.; Harris, C.A.; Li, G.D.; Norton, M.R.; Orgill, S.E.; Penrose, B.; et al. Prospects for Improving Perennial Legume Persistence in Mixed Grazed Pastures of South-Eastern Australia, with Particular Reference to White Clover. Crop & Pasture Science 2019, 70, 1141–1162. [CrossRef]
- Oberson, A.; Frossard, E.; Bühlmann, C.; Mayer, J.; Mäder, P.; Lüscher, A. Nitrogen Fixation and Transfer in Grass-Clover Leys under Organic and Conventional Cropping Systems. Plant Soil 2013, 371, 237–255.
- Tzanakakis, V.; Sturite, I.; Dörsch, P. Biological Nitrogen Fixation and Transfer in a High Latitude Grass-Clover Grassland under Different Management Practices. Plant Soil 2017, 421, 107–122.
- Munkaila, Mohammed. Yield, Quality, and Economic Returns of Alfalfa-Grass Mixtures Planted at Different Configurations under Full and Deficit Irrigations. 2023, 133.
- Corrall, A.J.; Fenlon, J.S. A Comparative Method for Describing the Seasonal Distribution of Production from Grasses. J Agric Sci 1978, 91, 61–67. [CrossRef]
- Känkänen, H. Undersowing in a Northern Climate: Effects on Spring Cereal Yield and Risk of Nitrate Leaching. 2010.
- Ledgard, S.F.; Steele, K.W. Biological Nitrogen Fixation in Mixed Legume/Grass Pastures. Plant Soil 1992, 141, 137–153.
- Zegler, C.H.; Brink, G.E.; Renz, M.J.; Ruark, M.D.; Casler, M.D. Management Effects on Forage Productivity, Nutritive Value, and Legume Persistence in Rotationally Grazed Pastures. Crop Sci 2018, 58, 2657–2664. [CrossRef]
- Cluzeau, D.; Binet, F.; Vertes, F.; Simon, J.C.; Riviere, J.M.; Trehen, P. Effects of Intensive Cattle Trampling on Soil-Plant-Earthworms System in Two Grassland Types. Soil Biol Biochem 1992, 24, 1661–1665. [CrossRef]
- Sanderson, M.A.; Skinner, R.H.; Barker, D.J.; Edwards, G.R.; Tracy, B.F.; Wedin, D.A. Plant Species Diversity and Management of Temperate Forage and Grazing Land Ecosystems. Crop Sci 2004, 44, 1132–1144. [CrossRef]
- Suter, M.; Connolly, J.; Finn, J.A.; Loges, R.; Kirwan, L.; Sebastià, M.T.; Lüscher, A. Nitrogen Yield Advantage from Grass–Legume Mixtures Is Robust over a Wide Range of Legume Proportions and Environmental Conditions. Glob Chang Biol 2015, 21, 2424–2438. [CrossRef]
- Humphreys, J.; Casey, I.A.; Laidlaw, A.S. Comparison of Milk Production from Clover-Based and Fertilizer-N-Based Grassland on a Clay-Loam Soil under Moist Temperate Climatic Conditions. Journal of Agricultural and Food Research 2009, 48, 189–207.
- Durand, J.L.; Gonzalez-Dugo, V.; Gastal, F. How Much Do Water Deficits Alter the Nitrogen Nutrition Status of Forage Crops? Nutr Cycl Agroecosyst 2010, 88, 231–243. [CrossRef]
- Suter, A.:; Connolly, M.; Finn, J.; Loges, J.A.; Kirwan, R. TITLE: Nitrogen Yield Advantage from Grass-Legume Mixtures Is Robust over a Wide Range of Legume Proportions and Environmental Conditions.. [CrossRef]
- Suter, M.; Finn, J.A.; Connolly, J.; Loges, R.; Lüscher, A. Gain in Nitrogen Yield from Grass-Legume Mixtures Is Robust Over a Wide Range of Legume Proportions and Environmental Conditions. Procedia Environ Sci 2015, 29, 187–188. [CrossRef]
- Jaramillo, D.M.; Sheridan, H.; Soder, K.; Dubeux, J.C.B.; Jaramillo, D.M.; Sheridan, H.; Soder, K.; Dubeux, J.C.B.; Villalba, J.J.; Macadam, J. Enhancing the Sustainability of Temperate Pasture Systems through More Diverse Swards. Agronomy 2021, Vol. 11, Page 1912 2021, 11, 1912. [CrossRef]
- Huguenin-Elie; Nyfeler, D.; Suter, M.; Frossard, E.; Lüscher, A. Positive Mischungseffekte Auf Ertrag Und Stickstoffversorgung in Klee-Gras-Mischungen Einleitung Und Zielsetzung.
- Peyraud, J.-L.; Van Den Pol-Van Dasselaar, A.; Collins, R.P.; Huguenin-Elie, O.; Dillon, P.; Peeters, A. Multi-Species Swards and Multi Scale Strategies for Multifunctional Grassland-Base Ruminant Production Systems: An Overview of the FP7-MultiSward Project. Multi-Species Swards and Multi Scale Strategies for Multifunctional Grassland-Base Ruminant pro-Duction Systems: An Overview of the FP7-MultiSward Project. 2014, 19.
- Chadwick, D.R.; Cardenas, L.; Misselbrook, T.H.; Smith, K.A.; Rees, R.M.; Watson, C.J.; Mcgeough, K.L.; Williams, J.R.; Cloy, J.M.; Thorman, R.E.; et al. Optimizing Chamber Methods for Measuring Nitrous Oxide Emissions from Plot-Based Agricultural Experiments. Eur J Soil Sci 2014, 65, 295–307. [CrossRef]
- Selbie, D.R.; Buckthought, L.E.; Shepherd, M.A. The Challenge of the Urine Patch for Managing Nitrogen in Grazed Pasture Systems. Advances in Agronomy 2015, 129, 229–292. [CrossRef]
- Eriksen, J.; Askegaard, M.; Rasmussen, J.; Søegaard, K. Nitrate Leaching and Residual Effect in Dairy Crop Rotations with Grass–Clover Leys as Influenced by Sward Age, Grazing, Cutting and Fertilizer Regimes. Agric Ecosyst Environ 2015, 212, 75–84. [CrossRef]
- SCHOLEFIELD, D.; TYSON, K.C.; GARWOOD, E.A.; ARMSTRONG, A.C.; HAWKINS, J.; STONE, A.C. Nitrate Leaching from Grazed Grassland Lysimeters: Effects of Fertilizer Input, Field Drainage, Age of Sward and Patterns of Weather. Journal of Soil Science 1993, 44, 601–613. [CrossRef]
- Voglmeier, K.; Jocher, M.; Häni, C.; Ammann, C. Ammonia Emission Measurements of an Intensively Grazed Pasture. Biogeosciences 2018, 15, 4593–4608. [CrossRef]
- Lombardi, B.; Alvarado, P.I.; Ricci, P.; Buraschi, L.M.; Viduzzi, G.; Palladino, R.A.; Gonda, H.L.; Juliarena, M.P. Combination of Cattle Urine and Dung Patches Synergically Increased Nitrous Oxide Emissions from a Temperate Grassland under Wet Conditions. Agric Ecosyst Environ 2022, 340, 108147. [CrossRef]
- Zhu, Y.; Butterbach-Bahl, K.; Merbold, L.; Leitner, S.; Pelster, D.E. Nitrous Oxide Emission Factors for Cattle Dung and Urine Deposited onto Tropical Pastures: A Review of Field-Based Studies. Agric Ecosyst Environ 2021, 322, 107637. [CrossRef]
- Reynolds, C.K.; Kristensen, N.B. Nitrogen Recycling through the Gut and the Nitrogen Economy of Ruminants: An Asynchronous Symbiosis. J Anim Sci 2008, 86, E293–E305. [CrossRef]
- Keim, J.P.; Anrique, R. Nutritional Strategies to Improve Nitrogen Use Efficiency by Grazing Dairy Cows. Chil J Agric Res 2011, 71, 623–633. [CrossRef]
- Dijkstra, J.; Reynolds, C.K.; Kebreab, E.; Bannink, A.; Ellis, J.L.; France, J.; van Vuuren, A.M. Challenges in Ruminant Nutrition: Towards Minimal Nitrogen Losses in Cattle. Energy and protein metabolism and nutrition in sustainable animal production 2013, 47–58. [CrossRef]
- Castillo, A.R.; Kebreab, E.; Beever, D.E.; France, J. A Review of Efficiency of Nitrogen Utilisation in lactating Dairy Cows and Its Relationship with environmental Pollution. J Anim Feed Sci 2000, 9, 1–32. [CrossRef]
- Kebreab, E.; France, J.; Beever, D.E.; Castillo, A.R. Nitrogen Pollution by Dairy Cows and Its Mitigation by Dietary Manipulation. Nutr Cycl Agroecosyst 2001, 60, 275–285.
- Fotoran, W.L.; Silva, J.R.D.; Glitz, C.; Ferreira, L.C.D.S.; Wunderlich, G. Establishment of an Antiplasmodial Vaccine Based on PfRH5-Encoding RNA Replicons Stabilized by Cationic Liposomes. Pharmaceutics 2023, 15. [CrossRef]
- Cantalapiedra-Hijar, G.; Abo-Ismail, M.; Carstens, G.E.; Guan, L.L.; Hegarty, R.; Kenny, D.A.; Mcgee, M.; Plastow, G.; Relling, A.; Ortigues-Marty, I. Review: Biological Determinants of between-Animal Variation in Feed Efficiency of Growing Beef Cattle. Animal 2018, 12, S321–S335.
- Doran, M.J.; Mulligan, F.J.; Lynch, M.B.; Fahey, A.G.; Ryan, N.J.; McDonnell, C.; McCabe, S.; Pierce, K.M. Effect of Supplement Crude Protein Concentration on Milk Production over the Main Grazing Season and on Nitrogen Excretion in Late-Lactation Grazing Dairy Cows. J Dairy Sci 2022, 105, 347–360.
- Lee, C.; Hristov, A.N.; Cassidy, T.W.; Heyler, K.S.; Lapierre, H.; Varga, G.A.; de Veth, M.J.; Patton, R.A.; Parys, C. Rumen-Protected Lysine, Methionine, and Histidine Increase Milk Protein Yield in Dairy Cows Fed a Metabolizable Protein-Deficient Diet. J Dairy Sci 2012, 95, 6042–6056. [CrossRef]
- Dodd, M.; Dalley, D.; Wims, C.; Elliott, D.; Griffin, A. A Comparison of Temperate Pasture Species Mixtures Selected to Increase Dairy Cow Production and Reduce Urinary Nitrogen Excretion. New Zealand Journal of Agricultural Research 2018, 62, 504–527. [CrossRef]
- Pembleton, K.G.; Tozer, K.N.; Edwards, G.R.; Jacobs, J.L.; Turner, L.R. Simple versus Diverse Pastures: Opportunities and Challenges in Dairy Systems. Anim Prod Sci 2015, 55, 893–901. [CrossRef]
- Hall, M.B.; Huntington, G.B. Nutrient Synchrony: Sound in Theory, Elusive in Practice. J Anim Sci 2008, 86, E287–E292. [CrossRef]
- Kohn, D.D.; Walsh, D.M. Plant Species Richness--The Effect of Island Size and Habitat Diversity. J Ecol 1994, 82, 367. [CrossRef]
- Beatson, P.R.; Meier, S.; Cullen, N.G.; Eding, H. Genetic Variation in Milk Urea Nitrogen Concentration of Dairy Cattle and Its Implications for Reducing Urinary Nitrogen Excretion. animal 2019, 13, 2164–2171. [CrossRef]
- Zhao, X.; Zang, C.; Zhao, S.; Zheng, N.; Zhang, Y.; Wang, J. Assessing Milk Urea Nitrogen as an Indicator of Protein Nutrition and Nitrogen Utilization Efficiency: A Meta-Analysis. J Dairy Sci 2025, 108, 4851–4862.
- Lavery, A.; Ferris, C.P. Proxy Measures and Novel Strategies for Estimating Nitrogen Utilisation Efficiency in Dairy Cattle. Animals 2021, Vol. 11, Page 343 2021, 11, 343. [CrossRef]
- Aliahmad, A.; Simha, P.; Vinnerås, B.; McConville, J. Comparative Environmental Assessment of Three Urine Recycling Scenarios: Influence of Treatment Configurations and Life Cycle Modeling Approaches. Environ Sci Technol 2025, 59, 21160–21173.
- Rivera, J.E.; Chará, J. CH4 and N2O Emissions From Cattle Excreta: A Review of Main Drivers and Mitigation Strategies in Grazing Systems. Front Sustain Food Syst 2021, 5, 657936.
- Dijkstra, J.; Reynolds, C.K.; Kebreab, E.; Bannink, A.; Ellis, J.L.; France, J.; van Vuuren, A.M. Challenges in Ruminant Nutrition: Towards Minimal Nitrogen Losses in Cattle. Energy and protein metabolism and nutrition in sustainable animal production 2013, 47–58. [CrossRef]
- White, S.L.; Sheffield, R.E.; Washburn, S.P.; King, L.D.; Green, J.T. Spatial and Time Distribution of Dairy Cattle Excreta in an Intensive Pasture System. J Environ Qual 2001, 30, 2180–2187. [CrossRef]
- Betteridge, K.; Costall, D.A.; Li, F.Y.; Luo, D.; Ganesh, S. Why We Need to Know What and Where Cows Are Urinating - a Urine Sensor to Improve Nitrogen Models. Proceedings of the New Zealand Grassland Association 2013, 119–124. [CrossRef]
- Carnevalli, R.A.; De Mello, A.C.T.; Shozo, L.; Crestani, S.; Coletti, A.J.; Eckstein, C. Spatial Distribution of Dairy Heifers’ Dung in Silvopastoral Systems. Ciência Rural 2019, 49, e20180796. [CrossRef]
- Carpinelli, S.; Da Fonseca, A.F.; Neto, P.H.W.; Dias, S.H.B.; Da Silveira Pontes, L. Spatial and Temporal Distribution of Cattle Dung and Nutrient Cycling in Integrated Crop–Livestock Systems. Agronomy 2020, Vol. 10, Page 672 2020, 10, 672. [CrossRef]
- Auerswald, K.; Mayer, F.; Schnyder, H. Coupling of Spatial and Temporal Pattern of Cattle Excreta Patches on a Low Intensity Pasture. Nutr Cycl Agroecosyst 2010, 88, 275–288.
- Hirata, M.; Ogura, S. The Effect of Progressive Grazing of a Pasture on the Spatial Distribution of Herbage Mass and Utilization by Cattle. IGC Proceedings (1985-2023) 2021.
- Bailey, D.W.; Stephenson, M.B.; Pittarello, M. Effect of Terrain Heterogeneity on Feeding Site Selection and Livestock Movement Patterns. Anim Prod Sci 2015, 55, 298–308. [CrossRef]
- Schönbach, P.; Wan, H.; Schiborra, A.; Gierus, M.; Bai, Y.; Müller, K.; Glindemann, T.; Wang, C.; Susenbeth, A.; Taube, F. Short-Term Management and Stocking Rate Effects of Grazing Sheep on Herbage Quality and Productivity of Inner Mongolia Steppe. Crop & Pasture Science 2009, 60, 963–974. [CrossRef]
- Svensk, M.; Pittarello, M.; Mariotte, P.; Nota, G.; Schneider, M.K.; Frund, D.; Dubois, S.; Allan, E.; Probo, M. Nitrogen Translocation by Highland Cattle Grazing in Alnus Viridis-Encroached Pastures. Nutr Cycl Agroecosyst 2023, 126, 127–141.
- Riesch, F.; Isselstein, J.; Balkenhol, N.; Beckmann, J.; Katarzyna Bojarska, ·; Gerber, N.; Herzog, S.; Jarmer, E.; Meißner, M.; Raab, C.; et al. From Here to There: Free-Ranging Large Herbivores Redistribute Nutrients from Grassland to Forest Soil. Landscape Ecology 2025 40:5 2025, 40, 96-. [CrossRef]
- Dumont, B.; Farruggia, A.; Garel, J.P.; Bachelard, P.; Boitier, E.; Frain, M. How Does Grazing Intensity Influence the Diversity of Plants and Insects in a Species-Rich Upland Grassland on Basalt Soils? Grass and Forage Science 2009, 64, 92–105. [CrossRef]
- Rook, A.J.; Dumont, B.; Isselstein, J.; Osoro, K.; WallisDeVries, M.F.; Parente, G.; Mills, J. Matching Type of Livestock to Desired Biodiversity Outcomes in Pastures – a Review. Biol Conserv 2004, 119, 137–150. [CrossRef]
- Egan, M.; Galvin, N.; Hennessy, D. Incorporating White Clover (Trifolium Repens L.) into Perennial Ryegrass (Lolium Perenne L.) Swards Receiving Varying Levels of Nitrogen Fertilizer: Effects on Milk and Herbage Production. J Dairy Sci 2018, 101, 3412–3427. [CrossRef]
- Ledgard, S.F.; Penno, J.W.; Sprosen, M.S. Nitrogen Inputs and Losses from Clover/Grass Pastures Grazed by Dairy Cows, as Affected by Nitrogen Fertilizer Application. J Agric Sci 1999, 132, 215–225. [CrossRef]
- Eriksen, J.; Askegaard, M.; Kristensen, K. Nitrate Leaching from an Organic Dairy Crop Rotation: The Effects of Manure Type, Nitrogen Input and Improved Crop Rotation. Soil Use Manag 2004, 20, 48–54. [CrossRef]
- Parsons, A.J.; Edwards, G.R.; Newton, P.C.D.; Chapman, D.F.; Caradus, J.R.; Rasmussen, S.; Rowarth, J.S. Past Lessons and Future Prospects: Plant Breeding for Yield and Persistence in Cool-Temperate Pastures. Grass and Forage Science 2011, 66, 153–172. [CrossRef]
- Jezequel, A.; Delaby, L.; McKay, Z.C.; Fleming, C.; Horan, B. Effect of Sward Species Diversity Combined with a Reduction in Nitrogen Fertilizer on the Performances of Spring-Calving Grazing Dairy Cows. J Dairy Sci 2024, 107, 11104–11116. [CrossRef]
- Box, L.A.; Edwards, G.R.; Bryant, R.H. Milk Production and Urinary Nitrogen Excretion of Dairy Cows Grazing Plantain in Early and Late Lactation. New Zealand Journal of Agricultural Research 2017, 60, 470–482. [CrossRef]
- Gardiner, C.A.; Clough, T.J.; Cameron, K.C.; Di, H.J.; Edwards, G.R.; de Klein, C.A.M. Potential for Forage Diet Manipulation in New Zealand Pasture Ecosystems to Mitigate Ruminant Urine Derived N2O Emissions: A Review. New Zealand Journal of Agricultural Research 2016, 59, 301–317. [CrossRef]
- Yang, C.T.; Ferris, C.P.; Yan, T. Effects of Dietary Crude Protein Concentration on Animal Performance and Nitrogen Utilisation Efficiency at Different Stages of Lactation in Holstein-Friesian Dairy Cows. animal 2022, 16, 100562. [CrossRef]
- Mangwe, M.C.; Mason, W.A.; Reed, C.B.; Spaans, O.K.; Pacheco, D.; Bryant, R.H. A Systematic Review and Meta-Analysis of Cow-Level Factors Affecting Milk Urea Nitrogen and Urinary Nitrogen Output under Pasture-Based Diets. J Dairy Sci 2025, 108, 579–596. [CrossRef]
- Herremans, S.; Vanwindekens, F.; Decruyenaere, V.; Beckers, Y.; Froidmont, E. Effect of Dietary Tannins on Milk Yield and Composition, Nitrogen Partitioning and Nitrogen Use Efficiency of Lactating Dairy Cows: A Meta-Analysis. J Anim Physiol Anim Nutr (Berl) 2020, 104, 1209–1218. [CrossRef]
- Zhang, J.; Xu, X.; Cao, Z.; Wang, Y.; Yang, H.; Azarfar, A.; Li, S. Effect of Different Tannin Sources on Nutrient Intake, Digestibility, Performance, Nitrogen Utilization, and Blood Parameters in Dairy Cows. Animals 2019, Vol. 9, Page 507 2019, 9, 507. [CrossRef]
- Aboagye, I.A.; Lynch, J.P.; Church, J.S.; Baah, J.; Beauchemin, K.A. Digestibility and Growth Performance of Sheep Fed Alfalfa Hay Treated with Fibrolytic Enzymes and a Ferulic Acid Esterase Producing Bacterial Additive. Anim Feed Sci Technol 2015, 203, 53–66. [CrossRef]
- Thompson, J.P.; Cristobal-Carballo, O.; Yan, T.; Zeller, W.E.; Huws, S.; Safoi, L.; Southam, A.D.; Ludwig, C.; Lloyd, G.R.; Stergiadis, S.; et al. Supplementation with Willow Containing Condensed Tannins Shifted Nitrogen Excretion from Urine to Faeces in Yearling Ewes. animal 2025, 19, 101698. [CrossRef]
- Carulla, J.E.; Kreuzer, M.; Machmüller, A.; Hess, H.D. Supplementation of Acacia Mearnsii Tannins Decreases Methanogenesis and Urinary Nitrogen in Forage-Fed Sheep. Aust J Agric Res 2005, 56, 961–970. [CrossRef]
- Luo, J.; Ledgard, S.F.; De Klein, C.A.M.; Lindsey, S.B.; Kear, M. Effects of Dairy Farming Intensification on Nitrous Oxide Emissions. Plant Soil 2008, 309, 227–237.
- White, S.L.; Sheffield, R.E.; Washburn, S.P.; King, L.D.; Green, J.T. Spatial and Time Distribution of Dairy Cattle Excreta in an Intensive Pasture System. J Environ Qual 2001, 30, 2180–2187. [CrossRef]
- Bailey, D.W.; Provenza, F.D. Mechanisms Determining Large-Herbivore Distribution. Resource Ecology 2008, 7–28. [CrossRef]
- Muzzo, B.I.; Bladen, K.; Perea, A.; Nyamuryekung’e, S.; Villalba, J.J. Multi-Sensor Integration and Machine Learning for High-Resolution Classification of Herbivore Foraging Behavior. Animals 2025, Vol. 15, Page 913 2025, 15, 913. [CrossRef]
- Florence, M.; Defourneaux, M. The Impact of Spatio-Temporal Shifts in Vertebrate Herbivore Communities on the Functioning the Icelandic Tundra; 2024; ISBN 978-9935-512-51-2.
- Kalehe Kankanamge, E.; Ramilan, T.; Tozer, P.R.; de Klein, C.; Romera, A.; Pieralli, S. Greenhouse Gas Mitigation in Pasture-Based Dairy Production Systems in New Zealand: A Review of Mitigation Options and Their Interactions. Climate Smart Agriculture 2025, 2, 100071. [CrossRef]
- Cichota, R.; Snow, V.O.; Vogeler, I.; Wheeler, D.M.; Shepherd, M.A. Describing N Leaching from Urine Patches Deposited at Different Times of the Year with a Transfer Function. Soil Research 2013, 50, 694–707. [CrossRef]
- Muzzo, B.I.; Ramsey, R.D.; Bladen, K.; Villalba, J.J. Tannin Supplementation Alters Foraging Behavior and Spatial Distribution in Beef Cattle. Sustainability 2025, Vol. 17, Page 10611 2025, 17, 10611. [CrossRef]
- Woodward, S.J.R.; Neal, M.B.; Cross, P.S. Preliminary Investigation into the Feasibility of Combining Satellite and GPS Data to Identify Pasture Growth and Grazing. Journal of New Zealand Grasslands 2019, 81, 47–54. [CrossRef]
- Monaghan, R.M.; Hedley, M.J.; Di, H.J.; McDowell, R.W.; Cameron, K.C.; Ledgard, S.F. Nutrient Management in New Zealand Pastures— Recent Developments and Future Issues. New Zealand Journal of Agricultural Research 2007, 50, 181–201. [CrossRef]
- Minneé, E.M.K.; Waghorn, G.C.; Lee, J.M.; Clark, C.E.F. Including Chicory or Plantain in a Perennial Ryegrass/White Clover-Based Diet of Dairy Cattle in Late Lactation: Feed Intake, Milk Production and Rumen Digestion. Anim Feed Sci Technol 2017, 227, 52–61. [CrossRef]
- Beukes, P.C.; Gregorini, P.; Romera, A.J.; Levy, G.; Waghorn, G.C. Improving Production Efficiency as a Strategy to Mitigate Greenhouse Gas Emissions on Pastoral Dairy Farms in New Zealand. Agric Ecosyst Environ 2010, 136, 358–365. [CrossRef]
- Cichota, R.; Snow, V.O.; Vogeler, I.; Wheeler, D.M.; Shepherd, M.A. Describing N Leaching from Urine Patches Deposited at Different Times of the Year with a Transfer Function. Soil Research 2013, 50, 694–707. [CrossRef]
- Gregorini, P.; Villalba B, J.J.; Chilibroste, P.; Provenza, F.D. Grazing Management: Setting the Table, Designing the Menu and Influencing the Diner.. [CrossRef]
- Scholefield, D.; Lockyer, D.R.; Whitehead, D.C.; Tyson, K.C. A Model to Predict Transformations and Losses of Nitrogen in UK Pastures Grazed by Beef Cattle. Plant Soil 1991, 132, 165–177.
- Luo, X.; Zhang, M.; Ni, Y.; Shen, G. Mitigation Strategies for NH3 and N2O Emissions in Greenhouse Agriculture: Insights into Fertilizer Management and Nitrogen Emission Mechanisms. Environ Technol Innov 2025, 37, 103995. [CrossRef]
- Laubach, J.; Taghizadeh-Toosi, A.; Gibbs, S.J.; Sherlock, R.R.; Kelliher, F.M.; Grover, S.P.P. Ammonia Emissions from Cattle Urine and Dung Excreted on Pasture. Biogeosciences 2013, 10, 327–338. [CrossRef]
- Gregorini, P.; Villalba, J.J.; Chilibroste, P.; Provenza, F.D. Grazing Management: Setting the Table, Designing the Menu and Influencing the Diner. Anim Prod Sci 2017, 57, 1248–1268.
Figure 1.
Conceptual representation of multi-scale nitrogen (N) flows and nitrogen use efficiency (NUE) in pasture-based grazing systems. At the farm-gate scale, N enters as fertilizer, biological N2 fixation, imported feed, and atmospheric deposition and leaves in animal products and conserved forage, with losses as NH3, N2O, and NO3−. At the paddock/soil–plant scale, N cycles through plant uptake, soil processes, and excreta patches. At the animal scale, dietary N is partitioned through rumen N metabolism into product N and excreted N. NUE emerges from the coupled interactions among these scales rather than from any single component.
Figure 1.
Conceptual representation of multi-scale nitrogen (N) flows and nitrogen use efficiency (NUE) in pasture-based grazing systems. At the farm-gate scale, N enters as fertilizer, biological N2 fixation, imported feed, and atmospheric deposition and leaves in animal products and conserved forage, with losses as NH3, N2O, and NO3−. At the paddock/soil–plant scale, N cycles through plant uptake, soil processes, and excreta patches. At the animal scale, dietary N is partitioned through rumen N metabolism into product N and excreted N. NUE emerges from the coupled interactions among these scales rather than from any single component.
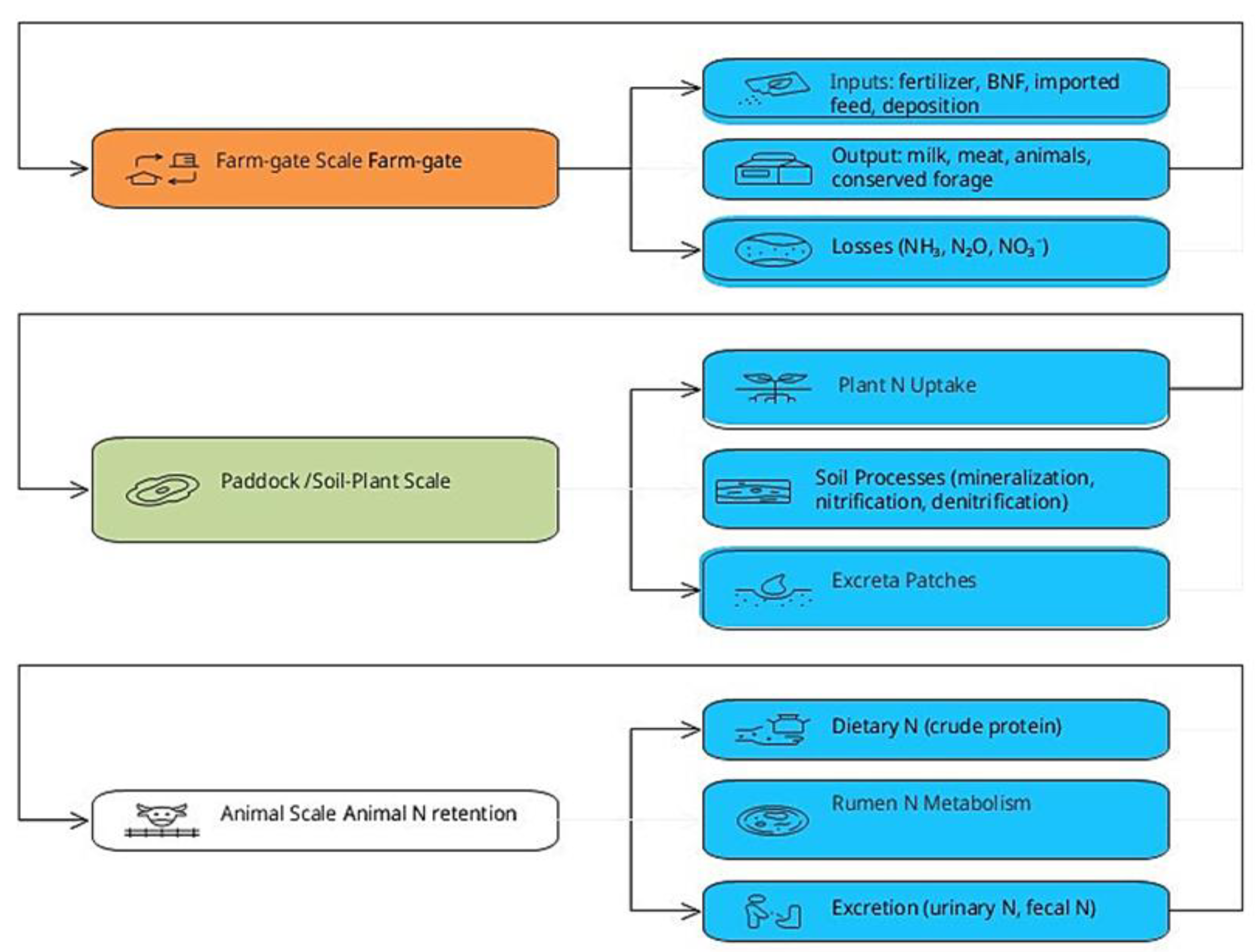
Figure 2.
Patch-scale nitrogen (N) pathways from urine and dung patches in grazed pastures. A grazing cow deposits urine and dung that form distinct patches on the soil surface. In the urine patch (left), urea in urine is rapidly hydrolyzed to ammonium (NH4+) by urease, with a fraction volatilized to the atmosphere as ammonia (NH3). Remaining NH4+ is oxidized to nitrite (NO2−) and nitrate (NO3−) via nitrification, and NO3− can either be taken up by plant roots or leach below the root zone. In the dung patch (right), organic N, carbon, and other nutrients enter the soil more slowly through decomposition, contributing to soil organic N and labile carbon pools. Within the soil, microsites with aerobic and anaerobic zones regulate the balance between N retention and loss, including denitrification of NO3− to nitrous oxide (N2O), nitric oxide (NO), and dinitrogen (N2). Plants integrate these processes by taking up inorganic N (NH4+, NO3−) from the soil. The magnitude and direction of these fluxes are modulated by controlling factors such as soil moisture and oxygen, temperature, labile carbon supply, and botanical composition (e.g., plantain, chicory, multispecies swards).
Figure 2.
Patch-scale nitrogen (N) pathways from urine and dung patches in grazed pastures. A grazing cow deposits urine and dung that form distinct patches on the soil surface. In the urine patch (left), urea in urine is rapidly hydrolyzed to ammonium (NH4+) by urease, with a fraction volatilized to the atmosphere as ammonia (NH3). Remaining NH4+ is oxidized to nitrite (NO2−) and nitrate (NO3−) via nitrification, and NO3− can either be taken up by plant roots or leach below the root zone. In the dung patch (right), organic N, carbon, and other nutrients enter the soil more slowly through decomposition, contributing to soil organic N and labile carbon pools. Within the soil, microsites with aerobic and anaerobic zones regulate the balance between N retention and loss, including denitrification of NO3− to nitrous oxide (N2O), nitric oxide (NO), and dinitrogen (N2). Plants integrate these processes by taking up inorganic N (NH4+, NO3−) from the soil. The magnitude and direction of these fluxes are modulated by controlling factors such as soil moisture and oxygen, temperature, labile carbon supply, and botanical composition (e.g., plantain, chicory, multispecies swards).
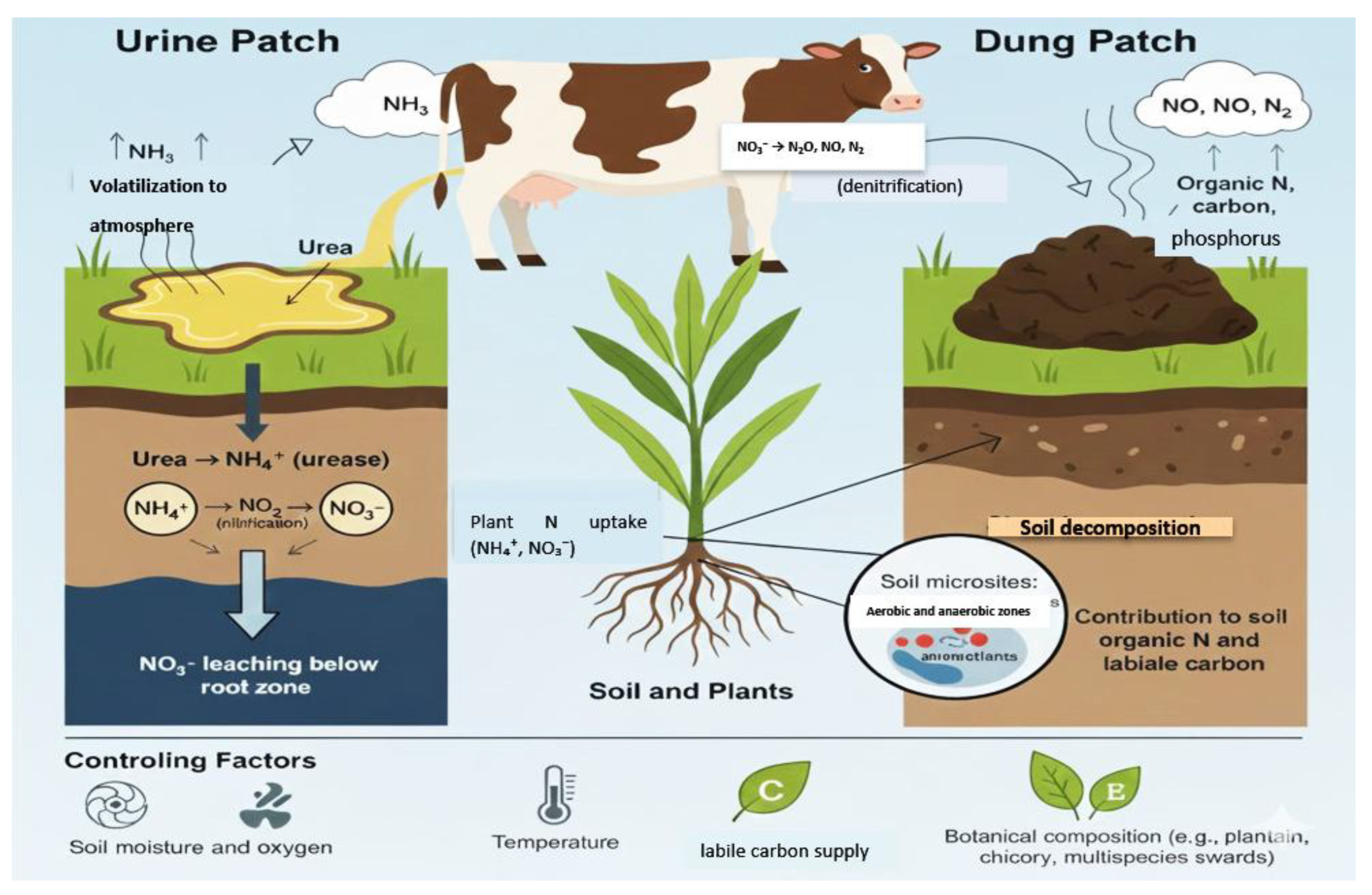
Figure 3.
This conceptual diagram illustrates how dietary crude protein (CP) and fermentable energy regulate rumen N metabolism, microbial protein synthesis, and the partitioning of N into milk/meat, feces, and urine. Degradable protein and non-protein N are hydrolyzed in the rumen to ammonia (NH3), which is incorporated into microbial protein when fermentable energy is adequate. Excess NH3 is absorbed into the bloodstream and converted to urea in the liver, with urea partitioned between urinary excretion and recycling back to the gut. Undigested protein contributes to fecal N, whereas microbial protein and undegraded protein supply amino acids that support milk and meat N deposition. Milk urea nitrogen (MUN) and blood urea nitrogen (BUN) reflect hepatic urea production and serve as indicators of dietary protein–energy balance. The diagram highlights how synchronized diets (balanced CP and energy) shift N toward productive pathways, whereas excess CP with insufficient energy increases urinary N losses.
Figure 3.
This conceptual diagram illustrates how dietary crude protein (CP) and fermentable energy regulate rumen N metabolism, microbial protein synthesis, and the partitioning of N into milk/meat, feces, and urine. Degradable protein and non-protein N are hydrolyzed in the rumen to ammonia (NH3), which is incorporated into microbial protein when fermentable energy is adequate. Excess NH3 is absorbed into the bloodstream and converted to urea in the liver, with urea partitioned between urinary excretion and recycling back to the gut. Undigested protein contributes to fecal N, whereas microbial protein and undegraded protein supply amino acids that support milk and meat N deposition. Milk urea nitrogen (MUN) and blood urea nitrogen (BUN) reflect hepatic urea production and serve as indicators of dietary protein–energy balance. The diagram highlights how synchronized diets (balanced CP and energy) shift N toward productive pathways, whereas excess CP with insufficient energy increases urinary N losses.
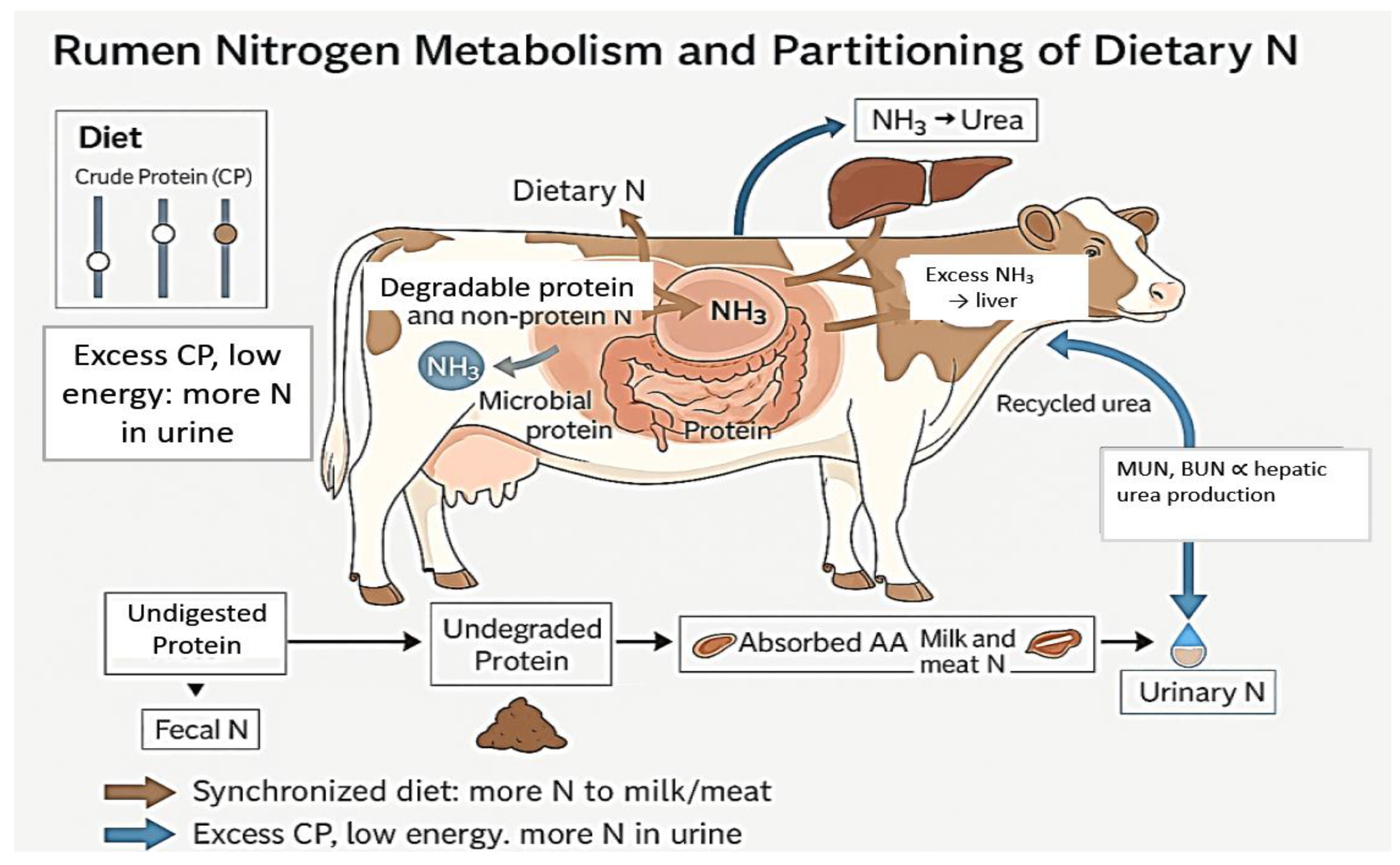
Figure 4.
Conceptual comparison of nitrogen partitioning in cows fed (left) a synchronized diet where crude protein (CP) supply is matched with fermentable energy, (middle) a high-CP, low-energy diet where rumen NH3 production exceeds microbial demand, and (right) a tannin/plant secondary metabolite (PSM)-supplemented diet. In the synchronized diet, balanced rumen NH3 and high microbial protein increase N retained in milk and meat and reduce urinary N, yielding higher nitrogen use efficiency (NUE). Under excess CP and low energy, NH3 surplus is converted to urea in the liver, increasing urinary N losses and lowering NUE. In tannin/PSM-supplemented diets, moderate condensed/hydrolysable tannins reduce rumen protein degradation and NH3 concentration, shifting N excretion toward more stable fecal N while maintaining milk and meat N at similar or slightly higher levels, thereby improving partitioning and potentially lowering field N losses.
Figure 4.
Conceptual comparison of nitrogen partitioning in cows fed (left) a synchronized diet where crude protein (CP) supply is matched with fermentable energy, (middle) a high-CP, low-energy diet where rumen NH3 production exceeds microbial demand, and (right) a tannin/plant secondary metabolite (PSM)-supplemented diet. In the synchronized diet, balanced rumen NH3 and high microbial protein increase N retained in milk and meat and reduce urinary N, yielding higher nitrogen use efficiency (NUE). Under excess CP and low energy, NH3 surplus is converted to urea in the liver, increasing urinary N losses and lowering NUE. In tannin/PSM-supplemented diets, moderate condensed/hydrolysable tannins reduce rumen protein degradation and NH3 concentration, shifting N excretion toward more stable fecal N while maintaining milk and meat N at similar or slightly higher levels, thereby improving partitioning and potentially lowering field N losses.
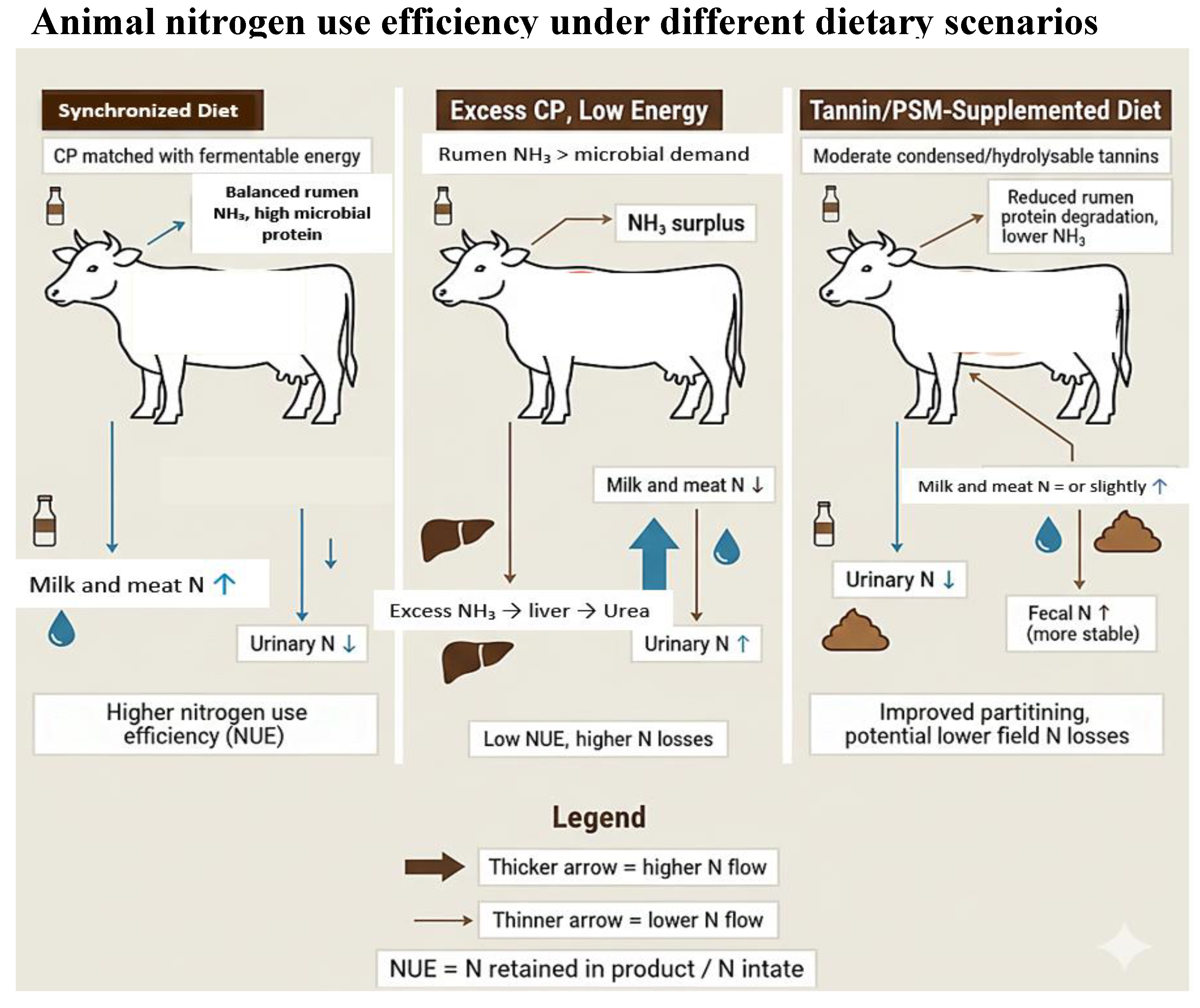
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



