
Submitted:
13 November 2025
Posted:
17 November 2025
You are already at the latest version
Abstract
Quinoa (Chenopodium quinoa, 2n = 4x = 36, AABB subgenomes) is a highly nutritious crop with potential to diversify global diets and alleviate malnutrition. It is also adaptable for production in soils increasingly affected by salinization and water scarcity. Quinoa was domesticated and artificially selected as a crop within the Andes Mountains, the geographically isolated Mediterranean climate zone of coastal Chile, and along the northwestern fringe of the Argentine dry Pampas. In addition, there is now abundant information regarding the wild species that were its immediate ancestors and which should be viewed as its secondary and tertiary breeding gene pools. These same ancestors contributed to independent domestications of the other forms of “quinoa” in ancient Mesoamerica and eastern North America from a common AABB ancestor-species, C. berlandieri, known commonly as pitseed goosefoot (PG). This review explores the biogeography of the diploid and polyploid relatives of the AABB allotetraploid goosefoot complex (ATGC). The seven or more ecotypes of PG, including the South American taxon C. hircinum or avian goosefoot, contain broad genetic variability and some can be used directly as crossing partners in making quinoa breeding populations. Of the extant diploid relatives, C. subglabrum is most closely related to the original maternal subgenome A of PG, while C. suecicum or C. ficifolium are most closely related to paternal subgenome B. These and the other AA and BB diploids are valuable model organisms for locating and modifying genes of interest and their expression, the ultimate goals being to increase quinoa’s yield potential, improve its nutritional attributes, explore value-adding industrial uses, and enhance quinoa's already formidable mechanisms to resist environmental stresses.
Keywords:
polyploidization
; Chenopodium
; crop breeding
; crop genetic resources
; pitseed goosefoot
1. Introduction
Many important crops are tetraploid, including cotton, durum wheat, potato, sweet potato, peanut, coffee, and quinoa [1]. Allotetraploids, which carry two distinct subgenomes derived via hybridization between different diploids, can be interesting models for studying genome evolution, especially subgenome interaction and divergence. In essence, heterosis due to overdominance is fixed in an allotetraploid and whole-genome duplication can eventually result in various fates such as loss of function (pseudogenization), subfunctionalization – temporal- or tissue-specific patterns of gene expression – and neofunctionalization, or the mutation-derived acquisition of new gene functions [2,3]. Whole-genome duplication also buffers the organism from the effects of deleterious mutations, while fixed heterosis facilitates flexibility for adapting to diverse, and especially disturbed, environments [4,5,6]. Subgenomes may show uneven rates of protein-sequence evolution, because they may accumulate mutations at different rates: a phenomenon known as biased fractionation, which in turn leads to subfunctionalization and neofunctionalization [7]. Domestication, geographic dispersal, and length of time since the hybridization event also influence the rate of subgenome evolution [1].
Quinoa (Chenopodium quinoa Willd 2n = 4x = 36, AABB genome formula) belongs to a New World allotetraploid goosefoot complex (ATGC) that includes North American pitseed goosefoot (C. berlandieri Moq; hereafter abbreviated PG) and South American avian goosefoot (C. hircinum Schreb) [8]. The hybridization event for PG was estimated at 3.3–6.3 Mya [9]. Quinoa was deemed to be a relatively recent domesticate, as evidenced by its high level of linkage disequilibrium although this could also be due to a history of strong selective pressure for unique local production environments (in quinoa's case, at extreme altitudes in the arid/saline Andean Altiplano) and/or limited intercrossing [10]. In addition to Andean quinoa, other domesticates from the ATGC include a Chilean coastal form that could be derived from geographic translocation of the montane ancestor [9,11]; Mesoamerican vegetable and seed crops of PG subsp. nuttaliae [8]; and at least one well-documented eastern North American domesticate, possibly from PG var. bushianum but classified by archaeologists as C. berlandieri subsp. jonesianum, that was extinct at or prior to the Conquest [12,13]. The ATGC is recognized as a monophyletic crop/weed/wild plant system [8,14,15].
In recent decades, quinoa has become a focus of international attention as a potential crop to diversify the world’s diet [16]. Since successful breeding depends upon genetic variation – especially when the goal is to readapt a crop for global use that was domesticated in specific environments like the Andean Altiplano and the Chilean Mediterranean-climate coastal zone – characterization of genetic resources for quinoa improvement is a crucial prerequisite [17]. The ATGC may be expected to encompass quinoa’s primary and secondary breeding pools, while AA and BB diploids are likely tertiary breeding resources. Quinoa’s wild relatives are threatened by habitat loss and climate change, including in the Andean region [18]. The proposed BB progenitors, the interfertile C. ficifolium and C. suecicum, are relatively common disturbance weeds in Eurasia and thus are not threatened [19]. Systematic, morphology-based assessment of Chenopodium AA and BB diploids is an ongoing process and has resulted in the designation of more than a dozen new species within the last decade [20,21,22,23]. Characterization of the diploid ancestors and wild relatives of quinoa is shedding light on the ATGC polyploidization and domestication events and is helping scientists formulate quinoa improvement strategies via interspecific crossing, particularly within the primary and secondary breeding pools (the ATGC) [8].
The purpose of this paper is to review what we now know regarding the status of quinoa’s primary, secondary, and tertiary gene pools (Figure 1). Since the landmark report of Fuentes-Bazan et al. [24] subdivided the old polyphyletic Chenopodium sensu lato into six genera, the current review will only focus on species of Chenopodium sensu stricto (also known as the C. album aggregate).
2. The Primary Gene Pool of C. quinoa
Although Gandarillas [25] attempted to classify quinoa into 17 Andean cultivated phenotypic races, the majority in the quinoa community currently recognize the existence of four ecotypes: Salares, Altiplano, Valley, and Sea Level or Coastal; a fifth proposed ecotype, Subtropical or Yungas, has not been recognized outside the mid-elevation valleys of the humid eastern slope of the Bolivian, and possibly Peruvian, Andes [26,27] and germplasm from this Yungas pool is unavailable for international quinoa breeders. However, a 1988 isozyme- and morphology-based diversity study of 98 samples (including one from the Yungas ecotype) by Wilson [28] concluded that there were only two distinct genetic groups: Andean, including Salares, Altiplano, Valley, and Yungas (a single sample) types; and Coastal or Sea Level from Chile. This clear bifurcation of quinoa into two basic clades has been reaffirmed by microsatellite- [29,30,31] and SNP-based marker analyses [11,32], along with limited evidence for independent domestications of the Highland and Coastal gene pools [8,11]. Following this trend, a number of AMOVA studies have demonstrated higher levels of within- versus among-group genetic variance in quinoa [33,34,35,36].
In contrast to these findings, an extensive study of 35 quinoa landrace populations from Northwest Argentina using a 22-set microsatellite panel found comparatively greater among-group genetic variance values, along with rich within-population genetic diversity [37]. They reported strong genetic differentiation of Argentine quinoas into four distinct geographic groups: Puna, Dry Valleys, Precordillera (Transition Zone), and Eastern Humid Valleys. This high degree of genetic diversity may reflect its pronounced environmental heterogeneity for elevation, temperature, and precipitation, as well as sympatry with highly variable populations of interfertile, wild C. hircinum [38]. These 35 Argentine accessions are not present in expatriate collections and may more accurately reflect genotypic stratification that once existed within the fragmented Andean sub-environments, especially prior to the widespread translocation of quinoas throughout the Inca Empire and its predecessors. It seems reasonable to assume a much higher degree of regional germplasm exchange in pre-Conquest times would have occurred within the relatively uniform, and highly populated, Altiplano region – the heartland of the Tiwanakota/Wari civilizations – and from there extending north to the Inca heartland around Cusco – as analysis of archaeological samples indicated may have happened at least twice in the last 18 centuries in the Antofagasta de la Sierra region of the Argentine northwest [39] Additionally, or alternatively, these data may indicate that quinoa's original Center of Diversity was not in the Lake Titicaca Basin, but rather farther south and east in what is now northwestern Argentina [37].
Breeding-based improvement of quinoa’s heat tolerance in lowland environments has largely focused on commercialization of, and hybridization with, germplasm adapted to the hot Chilean Central Valley. This heat-tolerant germplasm is distinct from the Salares-type quinoas grown in the far northern corner of the Chilean Altiplano [31] and the growing conditions also differ markedly from those found just over the coastal ranges along the cool Chilean Coast [40]. The presence of a large pericentric inversion comprising 71.7% (~52 Mb) of chromosome 3B [denoted In(3B)] in approximately 20% of Chilean germplasm, and first discovered in the QQ74 whole genome assembly, could complicate such hybridization-based breeding efforts, especially if significant genes for low-elevation adaptation are located within the inversion loop [9,11,41]. Single within-loop crossovers in inversion heterozygotes do occur at appreciable frequencies in model eukaryotes and produce approximately 50% duplicate-deficient gametes, thus creating a single haploblock spanning the inversion and potentially decreasing hybrid fertility [42]. This inversion is largely absent from highland gene pools [8,9,11,40]. While conserved inversions are often associated with adaptive gene blocks [43], the evolutionary significance of the 3B inversion is yet to be determined. However, the presence of an extensive pericentromeric area of reduced recombination, including on quinoa chromosome 3B, suggests that even a large pericentric inversion like Inv-3B would cause only a modest decrease in fertility in quinoa hybrids heterozygous for this rearrangement [8].
Quinoa breeding has traditionally been focused on mass selection of existing variability from among heterogeneous landrace populations. Intentional hybridization is a relatively new breeding approach in this crop. For instance, the commercial landrace Salares variety ‘Real’ has been the source of at least 22 selections, among them Ollagüe, Mañiqueña, Huallata, Toledo, Mok'o Rosado, Tres Hermanas, K'ellu, Canchis Anaranjado, Pisankalla, Real Blanca, Pandela Rosada, Perlasa, Achachino, Hilo, Rosa Blanca, Mok’o, Timsa, Lipeña, Chillpi Amapola, Utusaya, and Canchis Rosado [44]. The 2015 INIA Catalogue of Commercial Varieties of Quinoa in Peru listed 20 varieties, of which 11 were registered as cultivars and three (INIA 431-Altiplano, Illpa INIA, Salcedo INIA) were derived from intentional hybridization between selected parent [45]. Additional breeding efforts in Europe by companies like Quinoa Quality has resulted in the release and distribution of internationally accessible improved varieties including Puno, Titicaca, Vikinga, Carmen, Pasto, and Atlas [46] and, more recently, Zeno and low-saponin varieties Dagr and Freyja [47]. Crossing-based quinoa breeding has also accelerated in China, resulting in the release of at least nine registered varieties for local production [48]. Recently, an intentional crossing program between the Universidad Nacional del Altiplano-Puno and the University of Hohenheim, employing a machine-learning pipeline for optimizing panicle selection, has resulted in the release of three new varieties [49,50].
Some quinoa breeders, particularly in Peru and Mexico, have looked to mutagenesis as a means of enhancing natural variation within the primary gene pool [51]. One of the significant challenges with this approach is the allotetraploid nature of the quinoa genome, which makes identification of recessive mutations extremely difficult due to genetic buffering. An additional challenge with radiation-based mutagenesis is the induction of chromosome SV, which can at least partly diminish the benefits of yield- or quality-improving mutations by reducing recombination and increasing sterility when crossing mutagenized lines with strains lacking the novel SV [52]. Nevertheless, chemically-induced point mutations have been successfully used to characterize quinoa genes for betalain pigment synthesis [53], herbicide resistance [54], and epidermal bladder formation [55].
One of the curiosities related to quinoa is its association with free-living ajara types. Genetic sequencing of ajara types from Puno (BYU 1777 and 1789), Arequipa (BYU 1904), and Tarapacá (BYU 566) showed they cluster with domesticated quinoas and not with three free-living C. hircinum samples from outside the Andean Cordillera [8]. Although C. hircinum populations, as recognized by their characteristic odor of trimethylamine (TMA) and smaller seeds, exist well into the Andes up to at least 3000 masl [38], they are rare to absent in the Altiplano. Consequently, the appearance of ajara-type quinoas may be evidence of endoferalization due to spontaneous mutation, transposable elements, and/or epigenetic repatterning, rather than exoferalization via wild admixture [56,57,58].
Within the genome of C. quinoa, genome-wide association studies (GWAS) have found agronomically important quantitative trait loci (QTL) or marker-trait associations (MTA) for days to 50% flowering, thousand-grain weight, panicle length, betalain (pigment) biosynthesis, and saponin content [10,59,60]. The saponin trait is relevant because it affects processing time/complexity and may also affect tolerance to biotic stressors. Major maturity QTLs were found on chromosomes 4B, 5B, 6A, 7A, and 8B and were mostly associated with meristem regulatory genes, stress responses, flowering regulatory genes, transcriptional regulatory genes, and male fertility [59]. A seed weight candidate gene was found on 1B and was related to the timing of flowering transition [10]. The major saponin MTA was on 5B, presumably corresponding to the main BHLH25 saponin pathway regulatory locus previously identified in the v. 1 report of the quinoa whole-genome sequence [9,59,60]. Similarly, Patirange et al. [10] reported putative homology between the betalain pigmentation MTA on 1B and a pair of underlying betalain pathway genes previously identified in quinoa’s relative Beta vulgaris. The panicle length candidate genes were located on 3A and 7B and were associated with hormone regulation [59]. Interestingly, a well conducted GWAS failed to identify significant major QTL or MTA for resistance to the downy mildew pathogen, Peronospora variabilis, despite measuring relatively high heritabilities (0.72-0.81) for resistance to this most important of quinoa diseases [61].
Having a whole-genome assembly of the quinoa genome [9] allowed for an in-depth sequence comparison between subgenomes A and B. This analysis showed that the B subgenome comprises 55.8% of the QQ74 genome and is more dynamic structurally than the A [11], with a dramatic expansion of Gypsy long-terminal repeat (LTR) transposons [62]. Subgenome A is the maternal genome donor due to the cytoplasmic-maternal inheritance within the species and the greater similarity between cpDNA and mtDNA of quinoa with the existing AA diploids [63]. Subgenome A was shown to have the highest percentage of Copia LTR elements of all the known Chenopodium genomes [62].
3. The Secondary Gene Pool of C. quinoa
The secondary gene pool for quinoa breeding consists of free-living and domesticated forms of PG as well as wild-weedy South American C. hircinum, which together constitute the ATGC along with quinoa (Table 1). Spontaneous outcrossing in the field between quinoa and PG in North America was previously noted [64]. Early intentional intertaxa crosses between quinoa and PG resulted in numerous hybrid families with 1-8% stainable pollen in the F1 and universal seed set upon backcrossing with the pollen (usually wild) parent [65]. Previously, fertile hybrids between the Mexican domesticate PG subsp. nuttaliae and quinoa had also been reported [66].
Quinoa secondary genetic resources include a rich and highly diverse array of germplasm containing adaptation genes for expanding quinoa production into warm-season temperate, humid subtropical, and even lowland tropical environments. A genome-wide SNP-based phylogenetic analysis of 90 ATGC accessions, supplemented by the addition of eight newly sequenced quinoa strains [41] as well as a free-living PG strain from the prairies of Manitoba [67], has revealed a New World genetic landscape characterized by a relatively complex and diverse North American ecotypic distribution [8]. While South America is the center of domestication of C. quinoa, our results thus far indicate only C. hircinum, or avian goosefoot, is present within the region where quinoa was most likely domesticated [8].
There is considerable confusion as to the taxonomic separation of free-living South American feral ajara quinoas and avian goosefoot. The relatively recent Flora of Argentina differentiates between free-living quinoa (subsp. melanospermum) and avian goosefoot based on the presence/absence of two principal characters: alveoli-separating micro-canals on the pericarp surface of their pitted exoderm and basal leaf lobes [68]. The presence of both traits is supposedly diagnostic for avian goosefoot. However, neither character appears to be definitively diagnostic, as illustrated in Figure 2 for pericarp patterning and by Curti et al. [69] for basal leaf lobes. Regardless, free-living South American ATGC members not associated with current quinoa cultivation have been found from north of the Tropic of Capricorn in the irrigated Pacific deserts (Tarapacá Province, Chile) and Quebrada de Humahuaca (Jujuy Province, Argentina), southward to at least the 40th Parallel, and eastward to at least metropolitan Buenos Aires [38]. While the presence of malodorous trimethylamine (TMA) is a common characteristic of C. hircinum foliage and fruits, we have noted this character is less prominent above 2000 meters elevation [69].
Another question is whether avian goosefoot or PG are present in the northern Andes. The presence of a free-living quinoa ancestor there was inferred due to a progressive increase in quinoa null isozyme variants moving southward along the Andes [14]. Over two decades later, in 2012, a personal observation of one of the authors confirmed the presence of a free-living ATGC plant, with pitted pericarp and most closely resembling PG, growing in a disturbed site at the Agrosavia Mosquera campus outside Bogotá (Figure 3).
3.1. The Mesoamerican ATGC Gene Pool
The Mesoamerican ancestry and domestication of PG as subsp. nuttaliae in its panicle-vegetable (huauzontle), pseudocereal (chia roja), and leafy vegetable (quelite) forms has not been studied in depth. Huauzontle and its sympatric complex of PG weeds are found throughout the south-central highlands of Mexico within the area delimited by the Federal District and states of Tlaxcala, México, Puebla, Morelos, Jalisco, Colima, Michoacán, and Guerrero [70]. Distribution of chia roja is restricted to the Lake Pátzcuaro region of Michoacán and the leafy vegetable form, quelite de cenizo, is reported to have disappeared in at least part of its original range [14]. One curiosity raised by Wilson [14] is that while spontaneous outcrossing in Mexico between huauzontle and its nearby free-living relative PG (presumably var. sinuatum) is apparently common, intermediate semi-domesticated forms and black-seeded morphs that mimic the domesticated phenotype are either rare or non-existent. This contrasts with the situation in the Andes, where intermediate quinua-ajara forms are reportedly common [56]. This observation suggests that Mesoamerican huauzontle-PG hybrids sharply segregate toward the parental phenotypes – a phenomenon possibly attributable to chromosome structural differentiation, which would also lead to significant reductions in hybrid seed-set. In addition, or alternatively, the Andean ajaras might be arising from a gradual endoferalization process due to either transposon-mediated genome instability or epigenetic silencing (of domestication genes) and/or awakening (of atavistic free-living genes) [57].
The Mesoamerican nuttaliae gene pool includes variation for several traits of potential interest for quinoa improvement. Chia roja – the pseudocereal specialty domesticate from Michoacán - possesses saponin-free seeds and dense concentrations of deep red betacyanin pigments [14]. Additionally, Brown et al. [71] and Cepeda-Cornejo et al. [72] studied A- and B-genome allelic diversity at the GBSSI locus controlling seed amylose production and found that amylose-free, or waxy, starch is a common characteristic in huauzontle. At the molecular level, this phenotype is due to the presence of a homozygous 78-base deletion mutation (gbssib-Δ) toward the 5'-end of the B-genome locus, together with a homozygous mutation (gbssia-tp) within the leader (amyloplast-targeting) sequence in the A-genome locus [64]. Of the six quinoa accessions examined (three Highland, three Coastal), none had the waxy phenotype and only Coastal strain G-205 possessed (homozygous) a null mutation, gbssib-t, with a premature termination mutation at codon 129 [71] in subgenome B. This genotype was homozygous for the wild-type (GBSSIa-1) allele in subgenome A.
A whole-genome assembly for huauzontle (PI 433231 from Puebla, Mexico), a waxy strain, was published and compared with the genome of quinoa strain QQ74 (from Maule, Chile) [8]. Interestingly, genetic analysis of a set of 90 ATGC accessions resequenced for this study revealed the existence of a Peruvian germplasm, designated ‘0654’, harboring significant components of both Andean quinoa and Mesoamerican huauzontle genomes – an apparent result of outcrossing. Line 0654 is late maturing, with a semi-compact panicle and deep magenta pigmentation. Chromosomal collinearity analysis revealed the existence of chromosome 4B and 5A inversions and a 4A-6B reciprocal translocation in huauzontle [8].
3.2. Crop-Weed Complexes in Colorado
An interesting example of a Chenopodium crop-weed complex that formed outside an area of quinoa domestication comes from the San Luis Valley. This was first noted by Wilson [14] and he suggested the appearance of free-living types in and around these cultivated quinoa fields was more likely due to unintentional importation of ajara-types with contaminated seed quinoa, as opposed to outcrossing with local C. berlandieri. Quinoa was introduced to this high, arid valley straddling the Colorado-New Mexico border approximately 40 years ago. A 2019 quinoa field survey at White Mountain Quinoa Farm in Mosca, Colorado, sampled 15 fruiting plants representing an array of intermediate panicle, whole-plant, and seed phenotypes (Figure 4) and subsequent analysis of 16,916 SNP markers confirmed the presence of varying amounts of PG DNA within these lines (2nd International Quinoa Research Symposium, Aug 17-19, 2019, https://www.quinoasymposium.com/).
3.3. Quinoa Domestication Traits
Seed domestication in the ATGC, including quinoa, was likely a complex process involving interactions between migrating herds of seed-dispersing, post-Pleistocene ruminant megafauna (e.g., camelids in South America and bison in North America), prairie and woodland fires (both wildfires and anthropogenic), seasonal riparian flooding, and transhumance of hunter-gatherer groups [73,74]. Defining a domestication syndrome, or domestication traits, in quinoa is surprisingly complicated for characters that would likely be considered domestication signatures in other crops. The presence of a thin, non-lignified testa or epiderm that readily allows for imbibition of water would seem to be a necessity in most seed-propagated crops, and especially in quinoa, where the small seed size would make scarification difficult. However, some degree of physical and/or chemical dormancy is essential to prevent preharvest sprouting, especially as quinoa production expands into more humid regions like China’s Chengdu Plain [75], and to improve storage life of quinoa seeds. Additionally, the thickened seed coat in varieties like ‘Pasankalla’ may contribute to this variety’s ability to be popped. In addition, Belcher et al. [76] provided evidence that the thin-testa domestication trait, at least in PG, is expressed more abundantly under artificial garden conditions, suggesting that phenotypic plasticity may have played an important role in seed domestication.
Seed retention is another surprisingly complex trait. Quinoa would reasonably be expected to retain its seed for uniform harvest due to artificial selection through the domestication process, while free-living avian and PG would be likely to spontaneously disperse their seeds. However, the authors were able to collect abundant seeds off non-shattered plants of PG var. boscianum along the northern Gulf of Mexico Coast in late November of 2014 – at the end of an exceptionally mild tropical cyclone season that didn’t experience a single storm in the Gulf north of the 26th Parallel. Halwas and Worley [77] likewise reported prolonged retention of seeds on PG plants cultivated in their common garden experiments. This suggests that seed dispersal in at least some free-living populations is not spontaneous, but rather dependent on weather and other environmental disturbances.
Panicle and stem morphology are other ambiguous domestication traits. Four general panicle types are found in domesticated quinoa; glomerulate, consisting of compound spherical flower clusters arising from secondary and tertiary panicle branchlets and comprising a compact to semi-compact panicle; amaranthiform, composed of clusters of upright floral stalks arising from a central inflorescence stem; intermediate, with shortened upright stalks holding glomerules; and lax [50,78]. Lozano-Isla et al. [50] showed that crosses between varieties with amaranthiform and glomerulate panicles segregate bimodally toward the two parental forms. Though free-living ATGC members usually have lax panicles, considered to be the extreme wild-type variant of the glomerulate panicle present in most quinoa varieties [78], lax panicles are also produced by some domesticated quinoas, either constitutively or as a phenotypic plasticity response to heat stress [78,79]. Interestingly, Smith [80] reported that seed yields of wild PG populations from eastern North America could approach those of cultivated quinoa when grown in tended garden plots, despite their lax panicles and profuse stem branching, and that PG’s nutritional profile is comparable to quinoa’s [77]. Amaranthiform ‘Real’-type quinoas from the Bolivian-Chilean Salares region are well known for their profuse lateral stem branching in response to increased plant spacing, ambient moisture, and fertility [40,68].
Saponins are highly heterogeneous, amphipathic, triterpenoid glucoside compounds produced by quinoa and deposited in various tissues, most notably in the pericarp surrounding the seed [81]. They are also considered to be antinutritional and unpalatable, thus necessitating their removal via washing or pearling prior to seed consumption. However, the retention of saponins in most domesticated quinoa and huauzontle varieties, despite an easily selectable single-gene control mechanism with naturally occurring null alleles [9,82], suggests these secondary metabolites may have been retained for their biotic stress tolerance (including against avian predation), anti-inflammatory properties, and/or maintenance of seed viability [83]. Various studies have established potential commercial and industrial applications of quinoa saponins [84], while others have demonstrated that alteration of production parameters like soil salinity and irrigation rate can modulate pericarp saponin production [85].
Three less-studied metabolites that are also present in members of the ATGC are trimethylamine (TMA), its oxidized form trimethylamine oxide (TMAO), and geosmin. The fishy-smelling TMA and non-odorous TMAO, which are by-products of choline and betaine degradation, are apparently functional in abiotic stress tolerance [86] and TMA is notably present throughout the vegetative and reproductive structures in many or all genotypes of PG var. boscianum, PG var. berlandieri, and avian goosefoot, as well as in some populations of PG var. zschackei (especially from Oklahoma) and PG var. sinuatum [87]. Geosmin is an aromatic terpene that smells and tastes “earthy” and has been shown to be endogenously synthesized in quinoa’s close relative beet (Beta vulgaris) [88,89] and, presumably, also in the ATGC and its earthy-smelling A-genome diploid relatives like C. fremontii. These odorous chemicals have been hypothesized to function as herbivore repellants or, possibly, pollinator attractants [90]. The TMA odor is strongly expressed in F1 hybrids from quinoa x TMA-odor PG and quinoa x avian goosefoot crosses [87], suggesting the presence of a dominant inhibitor of the TMA --> TMAO oxidation step, which is universally catalyzed by a flavin monoamine oxidase enzyme [91].
3.4. Secondary Germplasm Collection and Characterization Looking into the Future
There remains a tremendous amount of research to better comprehend ATGC secondary genetic resources in the Americas. Ongoing work in Argentina is characterizing native secondary genetic resources for quinoa improvement and shedding light on their ancestral populations’ roles in quinoa domestication [37,38,69,92,93,94]. Since Andean Colombia and southeastern Bolivia also feature prominently in discussions of potential key locations in quinoa domestication, they should be targeted as sites to study cultivated, feral, and free-living ATGC populations.
Lowland subtropical and tropical areas of Mexico may have provided refugia for PG and other Chenopodium species during Pleistocene glacial expansions and their resultant dynamic disturbances in local climates, watersheds, and vegetation, not to mention immigration of human populations from the north during the last ~20,000 years [95,96]. For example, how far south do the sinuatum, zschackei, berlandieri, and boscianum ecotypes extend? Mesoamerican ATGC populations, extending at least as far south as highland areas of Guatemala like the Sierra de los Cuchumatanes, should therefore be prospected and studied to characterize the potentially complex ecotype structure of PG and clarify the picture of huauzontle domestication in Mexico.
4. Tertiary Gene Pool of C. quinoa
4.1. The AA Diploids
American diploids from both North and South America commonly possess the A genome [97,98,99]. Mandák et al. [100] provided phylogenetic evidence supporting C. bryoniifolium as the lone Eurasian A-diploid relic of a Eurasian-American species isolated by glacial expansion during the Northern Hemisphere Quaternary cooling period 2-3 Mya. In contrast, the American remnant diversified into ~45 free-living taxa, the majority in North America (Table 2 and Figure 5) [15]. South American members comprise ~10 taxa, among which is a single cultigen, Andean C. pallidicaule or cañahua [101,102]. The A-genome group have genome sizes of approximately 550-650 Mb [15,103]. It is also evident that the AA ancestor was the maternal parent in the original hybridization event that formed the tetraploid, based on greater sequence similarity of the ATGC mtDNA and cpDNA to those of the AA diploids [63].
The lone source in the literature regarding molecular phylogenetic relationships in a large set of AA diploids is Young et al. [15], who employed 10,588 filtered SNPs to produce a maximum likelihood tree with 42 diploid sample accessions along with extracted AA subgenomes from two quinoas and one PG (BYU 937 from Texas). Their analysis included samples of South American accessions that at the time were tentatively classified (and later determined, based on subsequent scrutiny of the recent Flora of Argentina [68] key) as C. cordobense, C. pallidicaule, C. petiolare (a low-growing ecotype of C. cordobense, BYU 1723), C. carnosolum (sample BYU 562, actually C. petiolare), and C. ruiz-lealii (actually, C. pilcomayense, BYU 1749) and noted they all formed a single clade (Group I), distinct from the multiple clades of North American species. Interestingly, the South Texas species C. albescens (BYU 1816-2) also fell within this clade, sister to the sample of C. pilcomayense, and both come from similar subtropical scrub-woodland habitats: mesquite (Prosopis) scrub in the former and semi-arid Pampas-Chaco habitat in the latter case, while their similar fruit morphology suggests these two species might be related through long-range northward dispersal. One of the most widespread taxa within the Andean region, C. petiolare, is the only perennial AA diploid and is found in both the winter-fog zone (lomas) of the hyper-arid Peruvian-Atacama coastal deserts and at altitudes above 2000 masl in more arid parts of the Andean Cordillera (unpublished observations of the authors). Other putative Andean diploids include C. carnosolum – which is also widespread in far southern Patagonia and Tierra del Fuego – and C. philippianum. Other Pampas species include C. cordobense, C. obscurum, C. papulosum, and C. ruiz-lealii, along with two taxa native to North America: C. desiccatum and C. pratericola, and Young et al. [15] reported a C. desiccatum sample, BYU 1755, from the desert in San Juan Province, Argentina (erroneously labeled as C. papulosum) as grouping with other C. desiccatum and C. pratericola samples from North America in their Group II. One additional species, C. scabricaule, resides in the Patagonian Desert and southern Andean foothills. An additional taxon described in the Flora of Argentina [68], C. frigidum, has vertically positioned fruits and therefore most likely should be included in a different genus, possibly Blitum (unpublished observations of the authors).
The free-living North American AA diploids are still in a process of taxonomic revision and are variously distributed, albeit mostly west of -97° W longitude in areas receiving <600mm annual precipitation [87]. The three main exceptions to this are C. foggii, a very rare and apparently declining taxon inhabiting granitic soils in the northern Appalachian Mountains; C. standleyanum, a widespread understory species of the Oak-Hickory Woodlands of East-Central North America; and the very rare C. pallescens of the Tallgrass Prairie zone [104]
Young et al. [15] reported in their SNP-based phylogenetic analysis of a subset of AA diploids that the North American taxa can be subdivided genetically into seven subclades, apart from South American Group I noted above. Group II comprised C. desiccatum, C. pratericola, and the rare northern California taxon C. howellii. Group III included some California C. hians-complex taxa as well as C. atrovirens and C. littoreum. Group IV contained C. fremontii. Group V was most closely related to subgenome A in the ATGC and included C. subglabrum, C. cycloides, C. standleyanum, C. pallescens, and C. nitens. Group VI included C. neomexicanum-complex taxa and C. watsonii. Group VII consisted of C. hians and Group VIII comprised C. nevadense [15].
The AA diploids are mostly adapted to well-drained soils in disturbed locations; often, multiple taxa will be found in mixed populations, including with PG in western North America. These include multiple members of the C. hians group in the Sierra Nevada, Transverse, and Peninsular Ranges of California [22] and members of the C. neomexicanum group in the southwestern United States [21]. Three of the psammophytes with limited distributions and/or disappearing habitats are C. littoreum on the California coast [20] and C. subglabrum and C. cycloides in the northern and southern Great Plains, respectively [15,105]. We suspect the decline of the latter two species, as well as C. pallescens in the Tallgrass Prairie, may have been precipitated by disruption of the annual pattern of seasonal disturbance across the Great Plains when the massive populations of American bison and other seasonally migrating megafauna were removed in the 19th Century [73,106].
Mangelsen et al. [79] and Young et al. [15] provided whole-genome, A-diploid assemblies, the former of the Andean crop cañahua (C. pallidicaule) and the latter of wild Watson’s stinking goosefoot (C. watsonii) from Southwest North America. Cañahua is an important food-security pseudocereal at elevations above 3500 meters in the Andes Mountains, is saponin-free, and has a free-threshing utriculate pericarp. Cañahua was presumably domesticated in the Andes from the free-living form of C. pallidicaule. Watson’s stinking goosefoot was selected for whole-genome sequencing based on its placement in the Foveosa series of Chenopodium Subsection Cellulata by Aellen [107] and preliminary isozyme [108] and molecular work [87] suggesting it might be one of the closer extant relatives to subgenome A in the ATGC – a presumption subsequently shown to be erroneous [15].
4.2. The BB Diploids
Three of the Eastern Hemisphere diploid species possess genome B: C. ficifolium, C. suecicum, and C. ucrainicum. The BB diploids also figured prominently in the evolution of the Eurasian complex of free-living and domesticated forms of BBCCDD C. album; BBCC C. ugandae; BBDD C. acerifolium and C. oahuense; BBEE C. karoi and C. jenissejense; and BBDDFF C. opulifolium [19].
Chenopodium ficifolium (figleaf goosefoot) is a fairly common temperate Eurasian weed that has spread to the Americas and was recently sequenced, with a genome size of approximately 730 Mb [109]. In Europe, it is found in disturbed areas, mostly between the 40th and 55th Parallels [100,110]. In the United States and Canada, figleaf goosefoot has been observed as a summer weed in New England, Quebec, and the Maritime Provinces, with additional spring-flowering populations in southern Arizona and Florida down to at least the 30th Parallel. Ludwig et al. [109] and Subedi et al. [111] discussed the suitability of C. ficifolium as a model species for quinoa genetics due to its smaller, diploid genome; relative ease of crossing; relatively short growth cycle; and its relative fecundity.
Chenopodium suecicum (Swedish goosefoot) is a Eurasian boreal weed, though its exact distribution may be muddled by confounding of Swedish goosefoot with lambsquarters (C. album) due to their morphological similarities. In places where Swedish goosefoot is sympatric with figleaf goosefoot hybridization between the two taxa may be common and their fertile and heterotic hybrids are capable of backcrossing with either parent [19]. It is also either native to, or naturalized in, Alaska, where it is a common weed based on the personal observations of the authors (Figure 6). The genome of C. suecicum has also been partially sequenced [9,87].
4.3. Breeding value of AA and BB diploids
The value of A- and B-genome diploids for quinoa improvement will depend upon the presence of post-zygotic barriers restricting chromosome pairing and recombination. The most significant post-zygotic barriers we expect to encounter are SV’s, based on what we have observed in reference-scale assemblies of AA diploids C. pallidicaule [101] and C. watsonii [15] and BB diploids C. suecicum [62] and C. ficifolium [109], in comparison with chromosomes of sequenced strains from the ATGC. These included reciprocal translocations Cq1B-Cq2B, Cq7B-Cq9B, Cq2B-Cq4A, and Cq6A-Cq6B [11,15], as well as large duplications involving chromosomes Cq1A, Cq2A, Cq2B, and Cq4B [11]. The data show that since formation of the ATGC polyploid complex over 11% and 14% of A- and B-genome genes, respectively, have been involved in structural rearrangements. These represent 0.63% and 5.93% of the A- and B-subgenomes, respectively [11].
5. Discussion
Within South America, quinoa breeders have at their disposal not only the extensive variation present in Andean and coastal Chilean quinoas, but also secondary genetic resources in the form of avian goosefoot (C. hircinum) and at least 12 native or naturalized AA diploids in the tertiary gene pool. Quinoa breeders outside of South America have limited access to the full range of quinoa genetic variation in the Andes and Chilean Coast due to international germplasm exchange restrictions and bureaucratic regulations. Nevertheless, a tremendously diverse array of secondary germplasm in PG is available through the USDA-NPGS gene bank. Both passive and intentional intercrossing of the two sister-taxa has occurred, with new quinoa strains being selectively bred for release to producers worldwide. These intertaxa hybrid-derived lines are being distributed to breeders and tested globally. The main challenge in using these populations is sifting through the massive amount of segregating variation. However, the potential payoff is huge. For example, genes for adaptation of quinoa to lowland, dry-season rice production regions affected by salinization due to sea-level rise could play an important role in not only ensuring food security, but also improving human and livestock nutrition, in these areas. The same may be true for dry-season quinoa production in upland tropical savanna and semi-arid climates.
In the long run, quinoa’s tertiary gene pool of AA- and BB-diploid species may begin to be exploited for introgression into quinoa of genes controlling their unusual traits and broad range of environmental adaptation, either via wide hybridization with embryo rescue (presumably) or by cisgenesis. One of the more agronomically interesting species in this group is figleaf goosefoot with its very short growth cycle, short stature, ease of hybridization, and high seed fecundity [111]. Nevada goosefoot (C. nevadense) stands out for its unique adaptation to heavy, poorly drained sodic clay soils. A large number of species are well adapted to thermic, xeric desert conditions, most notably C. desiccatum and C. incanum. In addition, there is the large set of AA-diploids with free-threshing pericarps (Figure 5b), most notably quinoa’s close relative smooth goosefoot (C. subglabrum); incorporation of this trait into quinoa could potentially allow the crop to retain whatever biotic-resistance protection saponins provide, while also making removal of the pericarp – where the majority of the fruit’s saponin appears to reside – easier. Having a free-threshing, utriculate pericarp would also facilitate the breeding and processing of high-saponin industrial quinoas.
The extant species most closely related to the original donors of the A and B subgenomes in the ATGC appear to be C. subglabrum and either C. suecicum or C. ficifolium, respectively (Figure 2). Although we have not attempted to create allotetraploids through hybridization, hybrid recovery, and chromosome doubling, that would be an interesting experiment. It would not only allow for comparison of the phenotype of the synthetic AABB tetraploid with PG, but also the breeding behavior (chromosome pairing, hybrid viability and fertility) in crosses with quinoa. We are awaiting the results of whole-genome DNA sequencing and assembly studies of these and other potential synthetic AABB parents to assess their genetic diversity and identify SV that could present obstacles to introgression of favorable genes into quinoa.
Author Contributions
KBS: SCT, and ENJ contributed heavily to writing of the manuscript; RLR contributed to evaluation and formatting of the references; DEJ and PJM were primarily involved in manuscript editing, review, and interpretation/contextualization of studies cited in the manuscript.
Funding
Since the article does not contain original data, no external research funding was involved in generating the data. SCT was supported by a Fulbright Fellowship to BYU.
Data Availability Statement
None of the data presented are original and everything to our knowledge lies within the public domain.
Acknowledgments
The authors gratefully acknowledge assistance and contributions of numerous quinoa scientists and producers in the Global South, particularly in the Andean region of Peru-Bolivia-Chile-Argentina; of Chenopodium taxonomic experts like S. Mosyakin, B. Mandák, P. Uotila, and N. Benet-Pierce; and D. Brenner at the Chenopodium germplasm repository NC-7 of USDA-NPGS at Iowa State University. The authors are also grateful for helpful conversations with Argentine scientists H.D. Bertero and R. Curti.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ATGC | Allotetraploid goosefoot complex |
| FISH | Fluorescence in situ hybridization |
| GWAS | Genome-wide association study |
| HIAN | C. hians taxonomic group |
| ITS | Internal transcribed sequence (rRNA) |
| LTR | Long-terminal repeat transposable element |
| MTA | Marker-trait association |
| NEOM | C. neomexicanum taxonomic group |
| PG | Pitseed goosefoot |
| QTL | Quantitative trait locus |
| SV | Structural variant |
| TMA | Trimethylamine |
| TMAO | Trimethylamine oxide |
References
- Sharbrough, J.; Conover, J.L.; Fernandes Gyorfy, M.; Grover, C.E.; Miller, E.R.; Wendel, J.F.; Sloan, D.B. Global Patterns of Subgenome Evolution in Organelle-Targeted Genes of Six Allotetraploid Angiosperms. Mol. Biol. Evol. 2022, 39, msac074. [CrossRef]
- Soltis, P.S.; Soltis, D.E. The Role of Genetic and Genomic Attributes in the Success of Polyploids. Proc. Natl. Acad. Sci. 2000, 97, 7051–7057. [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.-H. Evolution of Gene Duplication in Plants. Plant Physiol. 2016, 171, 2294–2316. [CrossRef]
- Coughlan, J.M.; Han, S.; Stefanović, S.; Dickinson, T.A. Widespread Generalist Clones Are Associated with Range and Niche Expansion in Allopolyploids of Pacific Northwest Hawthorns (Crataegus L.). Mol. Ecol. 2017, 26, 5484–5499. [CrossRef]
- te Beest, M.; Le Roux, J.J.; Richardson, D.M.; Brysting, A.K.; Suda, J.; Kubešová, M.; Pyšek, P. The More the Better? The Role of Polyploidy in Facilitating Plant Invasions. Ann. Bot. 2012, 109, 19–45. [CrossRef]
- Griffiths, A.G.; Moraga, R.; Tausen, M.; Gupta, V.; Bilton, T.P.; Campbell, M.A.; Ashby, R.; Nagy, I.; Khan, A.; Larking, A.; et al. Breaking Free: The Genomics of Allopolyploidy-Facilitated Niche Expansion in White Clover. Plant Cell 2019, 31, 1466–1487. [CrossRef]
- Schiavinato, M.; Bodrug-Schepers, A.; Dohm, J.C.; Himmelbauer, H. Subgenome Evolution in Allotetraploid Plants. Plant J. 2021, 106, 672–688. [CrossRef]
- Maughan, P.J.; Jarvis, D.E.; de la Cruz-Torres, E.; Jaggi, K.E.; Warner, H.C.; Marcheschi, A.K.; Bertero, H.D.; Gomez-Pando, L.; Fuentes, F.; Mayta-Anco, M.E.; et al. North American Pitseed Goosefoot (Chenopodium Berlandieri) Is a Genetic Resource to Improve Andean Quinoa (C. Quinoa). Sci. Rep. 2024, 14, 12345. [CrossRef]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmöckel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The Genome of Chenopodium Quinoa. Nature 2017, 542, 307–312. [CrossRef]
- Patiranage, D.S.; Rey, E.; Emrani, N.; Wellman, G.; Schmid, K.; Schmöckel, S.M.; Tester, M.; Jung, C. Genome-Wide Association Study in Quinoa Reveals Selection Pattern Typical for Crops with a Short Breeding History. eLife 2022, 11, e66873. [CrossRef]
- Rey, E.; Maughan, P.J.; Maumus, F.; Lewis, D.; Wilson, L.; Fuller, J.; Schmöckel, S.M.; Jellen, E.N.; Tester, M.; Jarvis, D.E. A Chromosome-Scale Assembly of the Quinoa Genome Provides Insights into the Structure and Dynamics of Its Subgenomes. Commun. Biol. 2023, 6, 1263. [CrossRef]
- Smith, B.D.; Yarnell, R.A. Initial Formation of an Indigenous Crop Complex in Eastern North America at 3800 B.P. Proc. Natl. Acad. Sci. 2009, 106, 6561–6566. [CrossRef]
- Kistler, L.; Shapiro, B. Ancient DNA Confirms a Local Origin of Domesticated Chenopod in Eastern North America. J. Archaeol. Sci. 2011, 38, 3549–3554. [CrossRef]
- Wilson, H.D. Quinua and Relatives (Chenopodium Sect.Chenopodium Subsect.Celluloid). Econ. Bot. 1990, 44, 92–110. [CrossRef]
- Young, L.A.; Maughan, P.J.; Jarvis, D.E.; Hunt, S.P.; Warner, H.C.; Durrant, K.K.; Kohlert, T.; Curti, R.N.; Bertero, D.; Filippi, G.A.; et al. A Chromosome-Scale Reference of Chenopodium Watsonii Helps Elucidate Relationships within the North American A-Genome Chenopodium Species and with Quinoa. Plant Genome 2023, 16, e20349. [CrossRef]
- Bazile, D. Global Trends in the Worldwide Expansion of Quinoa Cultivation. Biol. Life Sci. Forum 2023, 25, 13. [CrossRef]
- Khoury, C.K.; Carver, D.; Greene, S.L.; Williams, K.A.; Achicanoy, H.A.; Schori, M.; León, B.; Wiersema, J.H.; Frances, A. Crop Wild Relatives of the United States Require Urgent Conservation Action. Proc. Natl. Acad. Sci. 2020, 117, 33351–33357. [CrossRef]
- Fundación PROINPA; Bonifacio, A. Improvement of Quinoa (Chenopodium Quinoa Willd.) and Qañawa (Chenopodium Pallidicaule Aellen) in the Context of Climate Change in the High Andes. Cienc. E Investig. Agrar. 2019, 46, 113–124. [CrossRef]
- Hodková, E.; Mandák, B. An Overlooked Hybrid between the Two Diploid Chenopodium Species in Central Europe Determined by Microsatellite and Morphological Analysis. Plant Syst. Evol. 2018, 304, 295–312. [CrossRef]
- Benet-Pierce, N.; Simpson, M.G. Chenopodium Littoreum (Chenopodiaceae), a New Goosefoot from Dunes of South-Central Coastal California. Madroño 2010, 57, 64–72. [CrossRef]
- Benet-Pierce, N.; Simpson, M.G. Taxonomic Recovery of the Species in the Chenopodium Neomexicanum (Chenopodiaceae) Complex and Description of Chenopodium Sonorense Sp. Nov.1. J. Torrey Bot. Soc. 2017, 144, 339–356. [CrossRef]
- Benet-Pierce, N.; Simpson, M.G. THE TAXONOMY OF CHENOPODIUM HIANS, C. INCOGNITUM, AND TEN NEW TAXA WITHIN THE NARROW-LEAVED CHENOPODIUM GROUP IN WESTERN NORTH AMERICA, WITH SPECIAL ATTENTION TO CALIFORNIA. Madroño 2019, 66, 56–75. [CrossRef]
- S.l, M.; B, M. Chenopodium Ucrainicum (Chenopodiaceae / Amaranthaceae Sensu APG), a New Diploid Species: A Morphological Description and Pictorial Guide. Ukr. Bot. J. 2020, 77, 237–248.
- Fuentes-Bazan, S.; Uotila, P.; Borsch, T. A Novel Phylogeny-Based Generic Classification for Chenopodium Sensu Lato, and a Tribal Rearrangement of Chenopodioideae (Chenopodiaceae). Willdenowia 2012, 42, 5–24. [CrossRef]
- Gandarillas, H. Razas de Quinua; Bull; Instituto Boliviano de Cultivos Andinos, División de Investigaciones Agrícolas, Ministerio de Agricultura: La Paz, Bolivia, 1968;
- Tapia, M.E. Origen, Distribución Geográfica y Sistemas de Producción de La Quinua. In; Proceedings of the Reunión Sobre Genética y Fitomejoramiento de la Quinua: Puno, Peru, 1980.
- Risi, J. C.; Galwey, N. W. The Chenopodium Grains of the Andes: Inca Crops for Modern Agriculture. In; Advances in Applied Biology; Academic Press, 1984; pp. 145–215.
- Quinua Biosystematics I: Domesticated Populations on JSTOR Available online: https://www.jstor.org/stable/4255110?seq=1 (accessed on 31 October 2025).
- Mason, S.L.; Stevens, M.R.; Jellen, E.N.; Bonifacio, A.; Fairbanks, D.J.; Coleman, C.E.; McCarty, R.R.; Rasmussen, A.G.; Maughan, P.J. Development and Use of Microsatellite Markers for Germplasm Characterization in Quinoa (Chenopodium Quinoa Willd.). Crop Sci. 2005, 45, 1618–1630. [CrossRef]
- Christensen, S.A.; Pratt, D.B.; Pratt, C.; Nelson, P.T.; Stevens, M.R.; Jellen, E.N.; Coleman, C.E.; Fairbanks, D.J.; Bonifacio, A.; Maughan, P.J. Assessment of Genetic Diversity in the USDA and CIP-FAO International Nursery Collections of Quinoa (Chenopodium Quinoa Willd.) Using Microsatellite Markers [Abstract].
- Fuentes, F.F.; Martinez, E.A.; Hinrichsen, P.V.; Jellen, E.N.; Maughan, P.J. Assessment of Genetic Diversity Patterns in Chilean Quinoa (Chenopodium Quinoa Willd.) Germplasm Using Multiplex Fluorescent Microsatellite Markers. Conserv. Genet. 2009, 10, 369–377. [CrossRef]
- Maughan, P.J.; Smith, S.M.; Rojas-Beltrán, J.A.; Elzinga, D.; Raney, J.A.; Jellen, E.N.; Bonifacio, A.; Udall, J.A.; Fairbanks, D.J. Single Nucleotide Polymorphism Identification, Characterization, and Linkage Mapping in Quinoa. Plant Genome 2012, 5. [CrossRef]
- Genetic Identity Based on Simple Sequence Repeat (SSR) Markers for Quinoa (Chenopodium Quinoa Willd.) | International Journal of Agriculture and Natural Resources Available online: https://ojs.uc.cl/index.php/ijanr/article/view/28571 (accessed on 31 October 2025).
- Morillo, A.C.; Manjarres, E.H.; Reyes, W.L.; Morillo, Y. Research Article Molecular Characterization of Intrapopulation Genetic Diversity in Chenopodium Quinoa (Chenopodiaceae). Genet. Mol. Res. 2020, 19. [CrossRef]
- EL-Harty, E.H.; Ghazy, A.; Alateeq, T.K.; Al-Faifi, S.A.; Khan, M.A.; Afzal, M.; Alghamdi, S.S.; Migdadi, H.M. Morphological and Molecular Characterization of Quinoa Genotypes. Agriculture 2021, 11, 286. [CrossRef]
- Manjarres-Hernández, E.H.; Morillo-Coronado, A.C.; Morillo-Coronado, Y. Characterization of the Genetic Diversity of Chenopodium Quinoa from the Department of Boyacá-Colombia Using Microsatellite Markers. Biodiversitas J. Biol. Divers. 2025, 26. [CrossRef]
- Costa Tártara, S.M.; Manifesto, M.M.; Bramardi, S.J.; Bertero, H.D. Genetic Structure in Cultivated Quinoa (Chenopodium Quinoa Willd.), a Reflection of Landscape Structure in Northwest Argentina. Conserv. Genet. 2012, 13, 1027–1038. [CrossRef]
- Rodriguez, J.; Quipildor, V.B.; Giamminola, E.M.; Bramardi, S.J.; Jarvis, D.; Maughan, J.; Xu, J.; Farooq, H.U.; Ortega-Baes, P.; Jellen, E.; et al. Climate Links Leaf Shape Variation and Functional Strategies in Quinoa’s Wild Ancestor. AoB PLANTS 2025, 17, plaf049. [CrossRef]
- Winkel, T.; Aguirre, M.G.; Arizio, C.M.; Aschero, C.A.; Babot, M. del P.; Benoit, L.; Burgarella, C.; Costa-Tártara, S.; Dubois, M.-P.; Gay, L.; et al. Discontinuities in Quinoa Biodiversity in the Dry Andes: An 18-Century Perspective Based on Allelic Genotyping. PLOS ONE 2018, 13, e0207519. [CrossRef]
- Diversity of Quinoa in a Biogeographical Island: A Review of Constraints and Potential from Arid to Temperate Regions of Chile | Notulae Botanicae Horti Agrobotanici Cluj-Napoca Available online: https://www.notulaebotanicae.ro/index.php/nbha/article/view/9733 (accessed on 31 October 2025).
- Rey, E.; Abrouk, M.; Dufau, I.; Rodde, N.; Saber, N.; Cizkova, J.; Fiene, G.; Stanschewski, C.; Jarvis, D.E.; Jellen, E.N.; et al. Genome Assembly of a Diversity Panel of Chenopodium Quinoa. Sci. Data 2024, 11, 1366. [CrossRef]
- X Chromosome Inversions and Meiosis in Drosophila Melanogaster | PNAS Available online: https://www.pnas.org/doi/abs/10.1073/pnas.21.6.384 (accessed on 31 October 2025).
- Schaal, S.M.; Haller, B.C.; Lotterhos, K.E. Inversion Invasions: When the Genetic Basis of Local Adaptation Is Concentrated within Inversions in the Face of Gene Flow. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 20210200. [CrossRef]
- Aroni, J. C.; Aroni, G.; Quispe, R.; Bonifacio, A. Quinua Real: Catálogo; Fundación McKnight; Fundación PROINPA: La Paz, Bolivia, 2003;
- Apaza, V.; Cáceres, G.; Estrada, R.; Pinedo, R. Catalogue of Commercial Varieties of Quinoa in Peru; UN-FAO; INIA: Lima, Peru, 2015; p. 74;.
- Yadav, R.; Gore, P.G.; Gupta, V.; Saurabh; Siddique, K.H.M. Quinoa (Chenopodium Quinoa Willd.)—a Smart Crop for Food and Nutritional Security. In Neglected and Underutilized Crops; Farooq, M., Siddique, K.H.M., Eds.; Academic Press, 2023; pp. 23–43 ISBN 978-0-323-90537-4.
- Quinoa Seeds Wholesale | Quinoa Quality Available online: https://www.quinoaquality.com (accessed on 31 October 2025).
- Institute of Crop Science; Xiu-shi, Y.; Pei-you, Q.; Quinoa Committee of the Crop Science Society of China; Hui-min, G.; Gui-xing, R. Quinoa Industry Development in China. Cienc. E Investig. Agrar. 2019, 46, 208–219. [CrossRef]
- Lozano-Isla, F.; Apaza, J.-D.; Mujica Sanchez, A.; Blas Sevillano, R.; Haussmann, B.I.G.; Schmid, K. Enhancing Quinoa Cultivation in the Andean Highlands of Peru: A Breeding Strategy for Improved Yield and Early Maturity Adaptation to Climate Change Using Traditional Cultivars. Euphytica 2023, 219, 26. [CrossRef]
- Lozano-Isla, F.; Kienbaum, L.; Haussmann, B.I.G.; Schmid, K. A High-Throughput Phenotyping Pipeline for Quinoa (Chenopodium Quinoa) Panicles Using Image Analysis With Convolutional Neural Networks. Plant Breed. n/a. [CrossRef]
- Gomez-Pando, L.R.; Barra, A.E. la Developing Genetic Variability of Quinoa (Chenopodium Quinoa Willd.) with Gamma Radiation for Use in Breeding Programs. Am. J. Plant Sci. 2013, 4, 349–355. [CrossRef]
- Heddle, J.A.; Whissell, D.; Bodycote, D.J. Changes in Chromosome Structure Induced by Radiations: A Test of the Two Chief Hypotheses. Nature 1969, 221, 1158–1160. [CrossRef]
- Imamura, T.; Takagi, H.; Miyazato, A.; Ohki, S.; Mizukoshi, H.; Mori, M. Isolation and Characterization of the Betalain Biosynthesis Gene Involved in Hypocotyl Pigmentation of the Allotetraploid Chenopodium Quinoa. Biochem. Biophys. Res. Commun. 2018, 496, 280–286. [CrossRef]
- Mestanza, C.; Riegel, R.; Vásquez, S.C.; Veliz, D.; Cruz-Rosero, N.; Canchignia, H.; Silva, H. Discovery of Mutations in Chenopodium Quinoa Willd through EMS Mutagenesis and Mutation Screening Using Pre-Selection Phenotypic Data and next-Generation Sequencing. J. Agric. Sci. 2018, 156, 1196–1204. [CrossRef]
- Moog, M.W.; Trinh, M.D.L.; Nørrevang, A.F.; Bendtsen, A.K.; Wang, C.; Østerberg, J.T.; Shabala, S.; Hedrich, R.; Wendt, T.; Palmgren, M. The Epidermal Bladder Cell-Free Mutant of the Salt-Tolerant Quinoa Challenges Our Understanding of Halophyte Crop Salinity Tolerance. New Phytol. 2022, 236, 1409–1421. [CrossRef]
- Wilson, H.D. Quinua Biosystematics II: Free-Living Populations. Econ. Bot. 1988, 42, 478–494. [CrossRef]
- Mabry, M.E.; Rowan, T.N.; Pires, J.C.; Decker, J.E. Feralization: Confronting the Complexity of Domestication and Evolution. Trends Genet. 2021, 37, 302–305. [CrossRef]
- Ellstrand, N.C.; Heredia, S.M.; Leak-Garcia, J.A.; Heraty, J.M.; Burger, J.C.; Yao, L.; Nohzadeh-Malakshah, S.; Ridley, C.E. Crops Gone Wild: Evolution of Weeds and Invasives from Domesticated Ancestors. Evol. Appl. 2010, 3, 494–504. [CrossRef]
- Curti, R.N.; Rodriguez, J.; Ortega-Baes, P.; Bramardi, S.J.; Jellen, E.N.; Jarvis, D.E.; Maughan, P.J.; Tester, M.; Bertero, H.D. Leaf Shape of Quinoa’s Wild Ancestor Chenopodium Hircinum (Amaranthaceae) in a Geographic Context. Bot. J. Linn. Soc. 2025, boaf040. [CrossRef]
- Anton, A. M.; Zuloaga, F. O. Flora Argentina: Vascular Flora of the Argentine Republic. Dicotyledonae: Caryophyllales, Ericales, Gentianales. Chenopodium L.; Flora Argentina; IBODA, CONICET: San Isidro, Buenos Aires, Argentina, 2022; Vol. 19;.
- Rahman, H.; Vikram, P.; Hu, Y.; Asthana, S.; Tanaji, A.; Suryanarayanan, P.; Quadros, C.; Mehta, L.; Shahid, M.; Gkanogiannis, A.; et al. Mining Genomic Regions Associated with Agronomic and Biochemical Traits in Quinoa through GWAS. Sci. Rep. 2024, 14, 9205. [CrossRef]
- Colque-Little, C.; Abondano, M.C.; Lund, O.S.; Amby, D.B.; Piepho, H.-P.; Andreasen, C.; Schmöckel, S.; Schmid, K. Genetic Variation for Tolerance to the Downy Mildew Pathogen Peronospora Variabilis in Genetic Resources of Quinoa (Chenopodium Quinoa). BMC Plant Biol. 2021, 21, 41. [CrossRef]
- Trinh, M.D.L.; Visintainer, D.; Günther, J.; Østerberg, J.T.; da Fonseca, R.R.; Fondevilla, S.; Moog, M.W.; Luo, G.; Nørrevang, A.F.; Crocoll, C.; et al. Site-Directed Genotype Screening for Elimination of Antinutritional Saponins in Quinoa Seeds Identifies TSARL1 as a Master Controller of Saponin Biosynthesis Selectively in Seeds. Plant Biotechnol. J. 2024, 22, 2216–2234. [CrossRef]
- Maughan, P.J.; Chaney, L.; Lightfoot, D.J.; Cox, B.J.; Tester, M.; Jellen, E.N.; Jarvis, D.E. Mitochondrial and Chloroplast Genomes Provide Insights into the Evolutionary Origins of Quinoa (Chenopodium Quinoa Willd.). Sci. Rep. 2019, 9, 185. [CrossRef]
- Jacobsen, S.-E. The Worldwide Potential for Quinoa (Chenopodium quinoaWilld.). Food Rev. Int. 2003, 19, 167–177. [CrossRef]
- Mandák, B.; Krak, K.; Vít, P.; Pavlíková, Z.; Lomonosova, M.N.; Habibi, F.; Wang, L.; Jellen, E.N.; Douda, J. How Genome Size Variation Is Linked with Evolution within Chenopodium Sensu Lato. Perspect. Plant Ecol. Evol. Syst. 2016, 23, 18–32. [CrossRef]
- Mandák, B.; Krak, K.; Vít, P.; Lomonosova, M.N.; Belyayev, A.; Habibi, F.; Wang, L.; Douda, J.; Štorchová, H. Hybridization and Polyploidization within the Chenopodium Album Aggregate Analysed by Means of Cytological and Molecular Markers. Mol. Phylogenet. Evol. 2018, 129, 189–201. [CrossRef]
- Štorchová, H.; Drabešová, J.; Cháb, D.; Kolář, J.; Jellen, E.N. The Introns in FLOWERING LOCUS T-LIKE (FTL) Genes Are Useful Markers for Tracking Paternity in Tetraploid Chenopodium Quinoa Willd. Genet. Resour. Crop Evol. 2015, 62, 913–925. [CrossRef]
- Walsh, B.M.; Adhikary, D.; Maughan, P.J.; Emshwiller, E.; Jellen, E.N. Chenopodium Polyploidy Inferences from Salt Overly Sensitive 1 (SOS1) Data. Am. J. Bot. 2015, 102, 533–543. [CrossRef]
- Kolano, B.; McCann, J.; Orzechowska, M.; Siwinska, D.; Temsch, E.; Weiss-Schneeweiss, H. Molecular and Cytogenetic Evidence for an Allotetraploid Origin of Chenopodium Quinoa and C. Berlandieri (Amaranthaceae). Mol. Phylogenet. Evol. 2016, 100, 109–123. [CrossRef]
- Hinojosa, L.; Leguizamo, A.; Carpio, C.; Muñoz, D.; Mestanza, C.; Ochoa, J.; Castillo, C.; Murillo, A.; Villacréz, E.; Monar, C.; et al. Quinoa in Ecuador: Recent Advances under Global Expansion. Plants 2021, 10, 298. [CrossRef]
- Wilson, H.; Manhart, J. Crop/Weed Gene Flow:Chenopodium Quinoa Willd. andC. Berlandieri Moq. Theor. Appl. Genet. 1993, 86, 642–648. [CrossRef]
- Artificial Hybridization Among Species of Chenopodium Sect. Chenopodium on JSTOR Available online: https://www.jstor.org/stable/2418372?origin=crossref&seq=1 (accessed on 31 October 2025).
- Heiser, C.B.; Nelson, D.C. On the Origin of the Cultivated Chenopods (Chenopodium). Genetics 1974, 78, 503–505. [CrossRef]
- Bazile, D. Le Quinoa, Les Enjeux d’une Conquête. 2015, 1–112.
- Rodriguez, J.P.; Bonifacio, A.; Gómez-Pando, L.R.; Mujica, A.; Sørensen, M. Cañahua (Chenopodium Pallidicaule Aellen). In Neglected and Underutilized Crops; Farooq, M., Siddique, K.H.M., Eds.; Academic Press, 2023; pp. 45–93 ISBN 978-0-323-90537-4.
- Samuels, M.E.; Lapointe, C.; Halwas, S.; Worley, A.C. Genomic Sequence of Canadian Chenopodium Berlandieri: A North American Wild Relative of Quinoa. Plants 2023, 12, 467. [CrossRef]
- Jellen, E.N.; Jarvis, D.E.; Hunt, S.P.; Mangelsen, H.H.; Maughan, P.J. New Seed Collections of North American Pitseed Goosefoot (Chenopodium Berlandieri) and Efforts to Identify Its Diploid Ancestors through Whole-Genome Sequencing. Int. J. Agric. Nat. Resour. 2019, 46, 187–196. [CrossRef]
- Mangelson, H.; Jarvis, D.E.; Mollinedo, P.; Rollano-Penaloza, O.M.; Palma-Encinas, V.D.; Gomez-Pando, L.R.; Jellen, E.N.; Maughan, P.J. The Genome of Chenopodium Pallidicaule: An Emerging Andean Super Grain. Appl. Plant Sci. 2019, 7, e11300. [CrossRef]
- Habibi, F.; Mosyakin, S.L.; Shynder, O.I.; Krak, K.; Čortan, D.; Filippi, G.A.; Mandák, B. Chenopodium Ucrainicum (Amaranthaceae), a New ‘BB’ Genome Diploid Species: Karyological, Cytological, and Molecular Evidence. Bot. J. Linn. Soc. 2023, 203, 401–410. [CrossRef]
- Curti, R. n.; Andrade, A. j.; Bramardi, S.; Velásquez, B.; Daniel Bertero, H. Ecogeographic Structure of Phenotypic Diversity in Cultivated Populations of Quinoa from Northwest Argentina. Ann. Appl. Biol. 2012, 160, 114–125. [CrossRef]
- Curti, R.N.; de la Vega, A.J.; Andrade, A.J.; Bramardi, S.J.; Bertero, H.D. Multi-Environmental Evaluation for Grain Yield and Its Physiological Determinants of Quinoa Genotypes across Northwest Argentina. Field Crops Res. 2014, 166, 46–57. [CrossRef]
- Curti, R.N.; de la Vega, A.J.; Andrade, A.J.; Bramardi, S.J.; Bertero, H.D. Adaptive Responses of Quinoa to Diverse Agro-Ecological Environments along an Altitudinal Gradient in North West Argentina. Field Crops Res. 2016, 189, 10–18. [CrossRef]
- Subedi, M.; Neff, E.; Davis, T.M. Developing Chenopodium Ficifolium as a Potential B Genome Diploid Model System for Genetic Characterization and Improvement of Allotetraploid Quinoa (Chenopodium Quinoa). BMC Plant Biol. 2021, 21, 490. [CrossRef]
- Ludwig, C.D.; Maughan, P.J.; Jellen, E.N.; Davis, T.M. The Genome of Chenopodium Ficifolium: Developing Genetic Resources and a Diploid Model System for Allotetraploid Quinoa. G3 GenesGenomesGenetics 2025, 15, jkaf162. [CrossRef]
- Jarvis, D.E.; Sproul, J.S.; Navarro-Domínguez, B.; Krak, K.; Jaggi, K.; Huang, Y.-F.; Huang, T.-Y.; Lin, T.C.; Jellen, E.N.; Maughan, P.J. Chromosome-Scale Genome Assembly of the Hexaploid Taiwanese Goosefoot “Djulis” (Chenopodium Formosanum). Genome Biol. Evol. 2022, 14, evac120. [CrossRef]
- Wilson, H.D. Quinua and Relatives (Chenopodium Sect. Chenopodium Subsect. Cellulata). Econ. Bot. 1990, 44, 92–110.
- Wilson, H.D.; Heiser, C.B. The Origin and Evolutionary Relationships of ‘Huauzontle’ (Chenopodium Nuttaliae Safford), Domesticated Chenopod of Mexico. Am. J. Bot. 1979, 66, 198–206.
- Spengler, R.N.; Mueller, N.G. Grazing Animals Drove Domestication of Grain Crops. Nat. Plants 2019, 5, 656–662. [CrossRef]
- Langlie, B.S.; Hastorf, C.A.; Bruno, M.C.; Bermann, M.; Bonzani, R.M.; Condarco, W.C. Diversity In Andean Chenopodium Domestication: Describing A New Morphological Type From La Barca, Bolivia 1300-1250 B.C. J. Ethnobiol. 2011, 31, 72–88. [CrossRef]
- Mueller, N.G.; SpenglerIII, R.N.; Glenn, A.; Lama, K. Bison, Anthropogenic Fire, and the Origins of Agriculture in Eastern North America. Anthr. Rev. 2021, 8, 141–158. [CrossRef]
- Belcher, M.E.; Williams, D.; Mueller, N.G. Turning Over a New Leaf: Experimental Investigations into the Role of Developmental Plasticity in the Domestication of Goosefoot (Chenopodium Berlandieri) in Eastern North America. Am. Antiq. 2023, 88, 554–569. [CrossRef]
- Pan, X.; Gao, Y.; Zeng, F.; Zheng, C.; Ge, W.; Wan, Y.; Sun, Y.; Wu, X. Evaluation of Pre-Harvest Sprouting (PHS) Resistance and Screening of High-Quality Varieties from Thirty-Seven Quinoa (Chenopodium Quinoa Willd.) Resources in Chengdu Plain. Phyton-Int. J. Exp. Bot. 2023, 92, 2921–2936. [CrossRef]
- Craine, E.B.; Davies, A.; Packer, D.; Miller, N.D.; Schmöckel, S.M.; Spalding, E.P.; Tester, M.; Murphy, K.M. A Comprehensive Characterization of Agronomic and End-Use Quality Phenotypes across a Quinoa World Core Collection. Front. Plant Sci. 2023, 14. [CrossRef]
- Tovar, J.C.; Quillatupa, C.; Callen, S.T.; Castillo, S.E.; Pearson, P.; Shamin, A.; Schuhl, H.; Fahlgren, N.; Gehan, M.A. Heating Quinoa Shoots Results in Yield Loss by Inhibiting Fruit Production and Delaying Maturity. Plant J. 2020, 102, 1058–1073. [CrossRef]
- Halwas, S.; Worley, A.C. Incorporating Chenopodium Berlandieri into a Seasonal Subsistence Pattern: Implications of Biological Traits for Cultural Choices. J. Ethnobiol. 2019, 39, 510–529. [CrossRef]
- Smith, B.D. The Economic Potential of Chenopodium Berlandieri in Prehistoric Eastern North America. J. Ethnobiol. 1987, 7, 29–54.
- Rojas, W. Multivariate Analysis of Genetic Diversity of Bolivian Quinoa Germplasm. Food Rev. Int. 2003, 19, 9–23.
- Otterbach, S.; Wellman, G.; Schmöckel, S.M. Saponins of Quinoa: Structure, Function and Opportunities. In The Quinoa Genome; Schmöckel, S.M., Ed.; Springer International Publishing: Cham, 2021; pp. 119–138 ISBN 978-3-030-65237-1.
- Kollmar, M.; Böndel, K.B.; John, L.; Arold, S.; Schmid, K.; Jarvis, D.; Schmöckel, S.M.; Otterbach, S.L. Investigating the bHLH Transcription Factor TSARL1 as Marker and Regulator of Saponin Biosynthesis in Chenopodium Quinoa. J. Sci. Food Agric. 2025, 105, 7329–7339. [CrossRef]
- El Hazzam, K.; Hafsa, J.; Sobeh, M.; Mhada, M.; Taourirte, M.; EL Kacimi, K.; Yasri, A. An Insight into Saponins from Quinoa (Chenopodium Quinoa Willd): A Review. Molecules 2020, 25, 1059. [CrossRef]
- Bustos, K.A.G.; Muñoz, S.S.; da Silva, S.S.; Alarcon, M.A.D.F.; dos Santos, J.C.; Andrade, G.J.C.; Hilares, R.T. Saponin Molecules from Quinoa Residues: Exploring Their Surfactant, Emulsifying, and Detergent Properties. Molecules 2024, 29, 4928. [CrossRef]
- Gómez-Caravaca, A.M.; Iafelice, G.; Lavini, A.; Pulvento, C.; Caboni, M.F.; Marconi, E. Phenolic Compounds and Saponins in Quinoa Samples (Chenopodium Quinoa Willd.) Grown under Different Saline and Nonsaline Irrigation Regimens. J. Agric. Food Chem. 2012, 60, 4620–4627. [CrossRef]
- FNA: Chenopodium Desiccatum vs. Chenopodium Simplex Available online: https://nwwildflowers.com/compare/?t=Chenopodium+desiccatum,+Chenopodium+simplex (accessed on 5 November 2025).
- Catalá, R.; López-Cobollo, R.; Berbís, M.Á.; Jiménez-Barbero, J.; Salinas, J. Trimethylamine N-Oxide Is a New Plant Molecule That Promotes Abiotic Stress Tolerance. Sci. Adv. 2021, 7, eabd9296. [CrossRef]
- Jellen, E.N.; Jarvis, D.E.; Hunt, S.P.; Mangelsen, H.H.; Maughan, P.J. New Seed Collections of North American Pitseed Goosefoot (Chenopodium Berlandieri) and Efforts to Identify Its Diploid Ancestors through Whole-Genome Sequencing. Int. J. Agric. Nat. Resour. 2019, 46, 187–196. [CrossRef]
- Lu, G.; Edwards, C.G.; Fellman, J.K.; Mattinson, D.S.; Navazio, J. Biosynthetic Origin of Geosmin in Red Beets (Beta Vulgaris L.). J. Agric. Food Chem. 2003, 51, 1026–1029. [CrossRef]
- Freidig, A.K.; Goldman, I.L. Geosmin (2β,6α-Dimethylbicyclo[4.4.0]Decan-1β-Ol) Production Associated with Beta Vulgaris Ssp. Vulgaris Is Cultivar Specific. J. Agric. Food Chem. 2014, 62, 2031–2036. [CrossRef]
- Zaroubi, L.; Ozugergin, I.; Mastronardi, K.; Imfeld, A.; Law, C.; Gélinas, Y.; Piekny, A.; Findlay, B.L. The Ubiquitous Soil Terpene Geosmin Acts as a Warning Chemical. Appl. Environ. Microbiol. 2022, 88, e00093-22. [CrossRef]
- Tararina, M.A.; Allen, K.N. Bioinformatic Analysis of the Flavin-Dependent Amine Oxidase Superfamily: Adaptations for Substrate Specificity and Catalytic Diversity. J. Mol. Biol. 2020, 432, 3269–3288. [CrossRef]
- Domínguez-Vázquez, G.; Osuna-Vallejo, V.; Castro-López, V.; Israde-Alcántara, I.; Bischoff, J.A. Changes in Vegetation Structure during the Pleistocene–Holocene Transition in Guanajuato, Central Mexico. Veg. Hist. Archaeobotany 2019, 28, 81–91. [CrossRef]
- Caballero, M.; Ortega, B.; Lozano-García, S.; Montero, D.; Torres, E.; Soler, A.M. Environmental Changes in Central Mesoamerica in the Archaic and Formative Periods. Veg. Hist. Archaeobotany 2025, 34, 669–684. [CrossRef]
- Aellen, P. Illustrierte Flora von Mitteleuropa; 2nd ed.; Hanser: Munich, Germany, 1960; Vol. 3;.
Figure 1.
Illustration of the primary, secondary, and tertiary gene pools for breeding quinoa. (a) South American species distribution: quinoa (yellow, primary gene pool); avian goosefoot (black outline, secondary gene pool); tertiary gene pool diploids cañahua (teal), C. pilcomayense (green), C. carnosolum (orange), and mixed ranges of other South American diploids (red). (b) North American species distribution: pitseed goosefoot (black outline, secondary gene pool); main tertiary gene pool diploids C. neomexicanum complex (orange), C. hians complex (teal), C. leptophyllum-subglabrum-pratericola-desiccatum narrowleaf complex (green), C. standleyanum (yellow).
Figure 1.
Illustration of the primary, secondary, and tertiary gene pools for breeding quinoa. (a) South American species distribution: quinoa (yellow, primary gene pool); avian goosefoot (black outline, secondary gene pool); tertiary gene pool diploids cañahua (teal), C. pilcomayense (green), C. carnosolum (orange), and mixed ranges of other South American diploids (red). (b) North American species distribution: pitseed goosefoot (black outline, secondary gene pool); main tertiary gene pool diploids C. neomexicanum complex (orange), C. hians complex (teal), C. leptophyllum-subglabrum-pratericola-desiccatum narrowleaf complex (green), C. standleyanum (yellow).
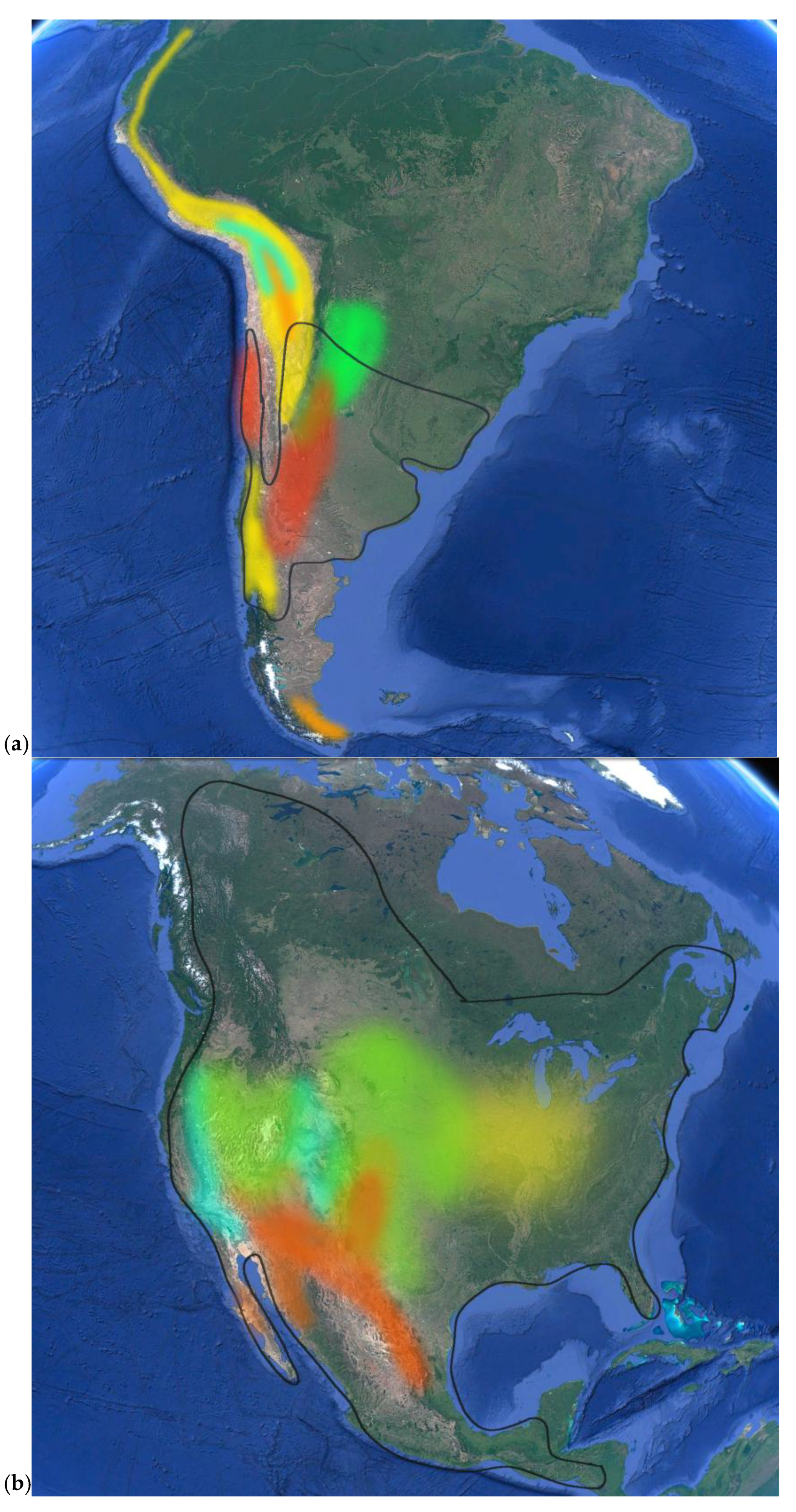
Figure 2.
Near-identical fruit/seed morphology is a characteristic of the three free-living members of the ATGC. C. (a) Five samples of Chenopodium hircinum: accession 1703 from Malbrán, Santiago del Estero, Argentina; 17105 from Chicureo, Santiago, Chile; 2301 from Gualeguay, Entre Ríos, Argentina; 2318 from Campo Quijano, Salta, Argentina; and 2327 from Humahuaca, Jujuy, Argentina. (b) Five samples of C. quinoa subsp. melanospermum: accession 1775 from Paucarcolla, Puno, Perú; 1904 from Hunter District, Arequipa, Perú; 23143 (CHEN 413) from Cachi Adentro, Salta, Argentina; 23242 from El Pedregal, Arequipa, Perú; and 25343 from Cota Cota District, La Paz, Bolivia. (c) Five samples of different ecotypes of C. berlandieri: accession 803 (macrocalycium) from York County, Maine, USA; 1486 (nuttaliae) from Atlacomulco, Mexico State, Mexico; 1840 (sinuatum) from Malibu, California, USA; 19170 (zschackei) from Lincoln County, Colorado, USA; 23176 (bushianum) from Scioto, Ohio, USA. (d) Closeup of pericarp surfaces of C. hircinum (top), C. quinoa melanospermum (middle), and C. berlandieri (bottom) showing a lack of inter-alveolar canals in all three taxa.
Figure 2.
Near-identical fruit/seed morphology is a characteristic of the three free-living members of the ATGC. C. (a) Five samples of Chenopodium hircinum: accession 1703 from Malbrán, Santiago del Estero, Argentina; 17105 from Chicureo, Santiago, Chile; 2301 from Gualeguay, Entre Ríos, Argentina; 2318 from Campo Quijano, Salta, Argentina; and 2327 from Humahuaca, Jujuy, Argentina. (b) Five samples of C. quinoa subsp. melanospermum: accession 1775 from Paucarcolla, Puno, Perú; 1904 from Hunter District, Arequipa, Perú; 23143 (CHEN 413) from Cachi Adentro, Salta, Argentina; 23242 from El Pedregal, Arequipa, Perú; and 25343 from Cota Cota District, La Paz, Bolivia. (c) Five samples of different ecotypes of C. berlandieri: accession 803 (macrocalycium) from York County, Maine, USA; 1486 (nuttaliae) from Atlacomulco, Mexico State, Mexico; 1840 (sinuatum) from Malibu, California, USA; 19170 (zschackei) from Lincoln County, Colorado, USA; 23176 (bushianum) from Scioto, Ohio, USA. (d) Closeup of pericarp surfaces of C. hircinum (top), C. quinoa melanospermum (middle), and C. berlandieri (bottom) showing a lack of inter-alveolar canals in all three taxa.


Figure 3.
A free-living Chenopodium plant growing on an untended site at the Agrosavia campus in Mosquera, Cundinamarca, Colombia. Fruits were inspected by one of the authors and confirmed to be pitted and therefore not the relatively common C. album. The plant morphology was most similar to C. berlandieri. Photo was taken by E. Jellen in December, 2012.
Figure 3.
A free-living Chenopodium plant growing on an untended site at the Agrosavia campus in Mosquera, Cundinamarca, Colombia. Fruits were inspected by one of the authors and confirmed to be pitted and therefore not the relatively common C. album. The plant morphology was most similar to C. berlandieri. Photo was taken by E. Jellen in December, 2012.

Figure 4.
Wild, intermediate, and domesticated phenotypes present in a quinoa production field in Mosca, Colorado, in September, 2019. (a) Variation for panicle morphology. (b) Variation for seed/fruit morphology. Accession 2127 (top left corner in each panel) is a native San Luis Valley C. berlandieri var. zschackei population several miles from the quinoa production area. Selection 19214 (bottom right corner in each panel) is representative of the cultivated ‘Blanca’ and ‘Medano’ phenotypes.
Figure 4.
Wild, intermediate, and domesticated phenotypes present in a quinoa production field in Mosca, Colorado, in September, 2019. (a) Variation for panicle morphology. (b) Variation for seed/fruit morphology. Accession 2127 (top left corner in each panel) is a native San Luis Valley C. berlandieri var. zschackei population several miles from the quinoa production area. Selection 19214 (bottom right corner in each panel) is representative of the cultivated ‘Blanca’ and ‘Medano’ phenotypes.

Figure 5.
Plant and fruit morphology of some Chenopodium AA diploids. (a) Variation for whole plant morphology: C. littoreum, C. arizonicum, C. desiccatum, C. subglabrum. (b) Variation for seed/fruit morphology. Left two columns, achenes with adhering pericarps; right two columns, utricles with free-threshing pericarps. Top to bottom, starting with the left column: C. luteum, C. hians, C. cycloides, C. nevadense, C. palmeri, C. arizonicum, C. lenticulare, C. pallescens, C. watsonii, C. neomexicanum, C. littoreum, C. papulosum, C. ruiz-lealii, C. fremontii, C. incanum, C. standleyanum, C. pilcomayense, C. pallidicaule, C. subglabrum, C. pratericola.
Figure 5.
Plant and fruit morphology of some Chenopodium AA diploids. (a) Variation for whole plant morphology: C. littoreum, C. arizonicum, C. desiccatum, C. subglabrum. (b) Variation for seed/fruit morphology. Left two columns, achenes with adhering pericarps; right two columns, utricles with free-threshing pericarps. Top to bottom, starting with the left column: C. luteum, C. hians, C. cycloides, C. nevadense, C. palmeri, C. arizonicum, C. lenticulare, C. pallescens, C. watsonii, C. neomexicanum, C. littoreum, C. papulosum, C. ruiz-lealii, C. fremontii, C. incanum, C. standleyanum, C. pilcomayense, C. pallidicaule, C. subglabrum, C. pratericola.

Figure 6.
Plant and fruit morphology of Chenopodium BB diploids. (a) Variation for whole plant morphology: C. ucrainicum, C. suecicum, C. ficifolium. (b) Variation for seed/fruit morphology: C. ucrainicum (top); left column: C. ficifolium (Czechia, top), C. ficifolium (Arizona, bottom); right column: C. suecicum (Czechia, top), C. suecicum (Alaska, bottom).
Figure 6.
Plant and fruit morphology of Chenopodium BB diploids. (a) Variation for whole plant morphology: C. ucrainicum, C. suecicum, C. ficifolium. (b) Variation for seed/fruit morphology: C. ucrainicum (top); left column: C. ficifolium (Czechia, top), C. ficifolium (Arizona, bottom); right column: C. suecicum (Czechia, top), C. suecicum (Alaska, bottom).



Table 1.
Accepted and proposed taxa in the Allotetraploid Goosefoot Complex (ATGC, 2n = 4x = 36, AABB genome), comprising the primary and secondary gene pools for quinoa improvement.
Table 1.
Accepted and proposed taxa in the Allotetraploid Goosefoot Complex (ATGC, 2n = 4x = 36, AABB genome), comprising the primary and secondary gene pools for quinoa improvement.
| Chenopodium Taxon | Common Name(s) | Distribution |
|---|---|---|
| berlandieri subsp. berlandieri | Pitseed goosefoot | |
| var. berlandieri | South Texas | |
| var. boscianum | Gulf of Mexico Coast | |
| var. bushianum | Eastern North America | |
| var. macrocalycium | New England Coast | |
| var. sinuatum | Mojave and Sonoran Deserts | |
| var. wilsonii1 | Eastern Oklahoma | |
| var. zschackei | Great Basin, Rocky Mountains, western Great Plains | |
| berlandieri subsp. nuttaliae | ||
| var. nuttaliae | Huauzontle, chia | South-central Mexico |
| var. pueblense1 | quelite | South-central Mexico |
| hircinum | Avian goosefoot | Southern South America |
| quinoa subsp. melanospermum | Ajara, ayara | Quinoa fields |
| quinoa subsp. milleanum | Ajara, ayara | Quinoa fields |
| quinoa subsp. quinoa | Quinoa, quinua, dzawe, jirwa | Andes, South-central Chile |
1 Proposed based on phylogenetic information in [8].
Table 2.
Accepted and proposed AA and BB diploid taxa (2n = 2x = 18), comprising most of the tertiary gene pool for quinoa improvement. Taxonomic subgroups are indicated based on classifications of [21] (NEOM = C. neomexicanum group) and [22] (HIAN = C. hians group) and BB species are noted.
| Chenopodium Taxon | Subgroup | Distribution |
|---|---|---|
| albescens Small | Texas; South America (?) | |
| arizonicum Standl. | NEOM | Arizona, New Mexico |
| atrovirens Rydb. | Great Basin, Rocky Mts. | |
| aureum Benet-Pierce | HIAN | Sierra Nevada Mts. |
| brandegeeae Benet-Pierce | HIAN | S California |
| bryoniifolium | NE Asia | |
| carnosolum Moq | S Patagonia, Andes Mts. | |
| cordobense Aellen | W Argentina | |
| cycloides A. Nelson | Southern Great Plains | |
| desiccatum A. Nelson | W North America, South America | |
| eastwoodiae Benet-Pierce | HIAN | Sierra Nevada Mts. |
| ficifolium Sm. | BB | Eurasia; global weed |
| flabellifolium Standl. | NEOM | Isla San Martin (Mexico) |
| foggii Wahl | Appalachian Mts. | |
| fremontii S. Wats. | W North America | |
| hians Wahl | HIAN | W North America |
| howellii Benet-Pierce | HIAN | Sierra Nevada Mts. |
| incanum A. Heller | Great Basin, Arizona, Great Plains | |
| incognitum Wahl | HIAN | Great Basin, Rocky Mts. |
| lenticulare Aellen | NEOM | Chihuahuan Desert |
| leptophyllum (Nutt ex Moq) B.D. Jacks | W North America | |
| lineatum Benet-Pierce | HIAN | Sierra Nevada Mts. |
| littoreum Benet-Pierce & M.G. Simpson | Central California Coast | |
| luteum Benet-Pierce | HIAN | Sierra Nevada Mts. |
| neomexicanum Standl. | NEOM | Arizona, New Mexico |
| nevadense Standl. | W Great Basin | |
| nitens Benet-Pierce & M.G. Simpson | W North America | |
| obscurum Aellen | W Argentina | |
| pallescens Standl. | Great Plains | |
| pallidicaule Aellen | Andes Mts.; incl. cultigens (cañahua) | |
| palmeri Standl. | NEOM | Sonoran Desert |
| papulosum Moq | W Argentina | |
| parryi Standl. | NEOM | Sierra Madre Oriental Mts. |
| petiolare Kunth | Andes Mts., Peruvian Coast | |
| philippianum Aellen | Andes Mts. | |
| pilcomayense Aellen | N Pampas, Gran Chaco | |
| pratericola Rydb. | W North America, South America | |
| ruiz-lealii Aellen | W Argentina | |
| sandersii Benet-Pierce | HIAN | S California |
| scabricaule Speg. | Patagonia | |
| simpsonii Benet-Pierce | HIAN | S California |
| sonorense Benet-Pierce & M.G. Simpson | NEOM | Sonoran Desert |
| standleyanum Aellen | E North America | |
| subglabrum (S. Watson) A. Nelson | Great Plains, Great Basin | |
| suecicum Murr | BB | Alaska, Eurasia; global weed |
| twisselmannii Benet-Pierce | HIAN | Sierra Nevada Mts. |
| ucrainicum Mosyakin & Mandák | BB | E Europe |
| wahlii Benet-Pierce | HIAN | S California |
| watsonii A. Nelson | NEOM | Colorado Plateau, Great Plains |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



