
Submitted:
13 November 2025
Posted:
14 November 2025
You are already at the latest version
Abstract
The fruits of Solanum lycopersicum L. cultivar “Microtom” are a powerful source of antioxidants. We investigated whether two light-quality regimes, i.e., fluorescent white (FL) and red-blue (RB), influenced the antioxidant composition in such fruits, and assessed the potential radioprotective properties of their extracts on normal human cells exposed to clinical photons as used in cancer radiotherapy (RT). Increasing normal-tissue tolerance to radiation is critical for reducing the risk of RT-associated sequelae. Biochemical characterization showed that RB enhanced the content of antioxidant phytochemicals (i.e., polyphenols, flavonoids, total carotenoids, lycopene) while FL promoted ascorbic acid synthesis. Initially tested at 200 µg ml-1, RB-derived extracts decreased radiation-induced DNA damage as measured by the cytokinesis-block micronucleus (CBMN) assay in epidermal HaCaT cells. Both RB and FL regimes were subsequently studied in MCF-10A breast cancer (BC) cells, a model of normal-tissue radioresponse in BC RT, using extracts at 100 and 200 µg ml-1 also evaluating oxidative stress by a ROS detection assay. Both FL and RB afforded radioprotection. However, RB suppressed radiation-induced MN formation and oxidative stress to a greater extent compared to FL. Therefore, modulation of light-quality regimes represents an innovative approach for developing radionutraceuticals with potential benefits for RT patients.
Keywords:
antioxidant molecules
; ionizing radiation
; DNA damage
; cancer radiotherapy
; normal-tissue radioprotection
; plant growth
; light quality
; bioprospecting
; nutraceuticals
; functional foods
1. Introduction
Tomatoes (Solanum lycopersicum L.), archetypal example of the Mediterranean diet where they are widely consumed as vegetables, play an essential role in human nutrition due to their well-recognized health benefits [1]. They are rich in vitamins, minerals, fibers, proteins, essential amino acids, monounsaturated fatty acids, carotenoids, and phytosterols [2,3,4]. Moreover, tomatoes are an excellent source of bioactive compounds with natural antioxidant properties, such as carotenoids (β-carotenoids and lycopene), ascorbic acid (vitamin C), tocopherol (vitamin E), and are highly abundant in secondary metabolites, like phenolic compounds (quercetin, kaempferol, naringenin, and lutein, as well as caffeic, ferulic, and chlorogenic acids). These compounds are valuable in preventing human pathologies, such as cardiovascular disease (CVD) and neurodegenerative impairment, also exerting an anti-inflammatory action that may reduce cancer risk [5,6,7]. Therefore, the regular consumption of tomatoes may help alleviate symptoms or prevent the onset of several diseases, particularly those feeding on chronic inflammatory conditions [8]. At the cellular level, such beneficial health properties are likely due to a concentration-dependent protection that these molecules may afford to normal cells [9] by scavenging free radicals and neutralizing Reactive Oxygen Species (ROS), suppressing apoptosis and metal chelation, aiding DNA damage repair, and modulating enzyme activities, cytokine expression and signal transduction pathways [10,11,12].
Among tomato cultivars, “Microtom” presents peculiar traits: parthenocarpy[13,14], dwarf size allowing growth at high plant density in small indoor systems (up to 1357 plants m−2), and a short life cycle (70–90 days from sowing to fruit ripening)[14]. Moreover, its intrinsic richness in bioactive compounds can be potentially enhanced through targeted light manipulation [15,16]. In fact, although Microtom already boasts high levels of antioxidants and other health-promoting phytochemicals, recent studies under controlled environments have demonstrated that the application of specific light-quality regimes, using the Light Emitting Diode (LED) technology, can significantly boost the synthesis and accumulation of these molecules. This makes Microtom an ideal candidate for producing nutrient-rich fruits and specialized extracts tailored for functional foods and nutraceuticals [17,18].
Recent studies have gone a step further, leveraging Microtom’s unique characteristics to develop a kind of ‘super-extracts’. For instance, research by Vitale et al. [15] and Hay Mele et al. [19] has shown that by optimizing light quality during cultivation, it is possible to obtain Microtom extracts with exceptionally high concentrations of antioxidants and phenolic compounds. These ‘super-extracts’ exhibit enhanced biological activity, making them highly attractive for several applications. Similarly, Paolillo et al. [20] have implemented comparable strategies in Brassica species, underscoring the versatility and scalability of this approach. Notably, the methodology is not limited to tomatoes or brassica: Costanzo et al. [21] have successfully translated these techniques to other crops such as mandarin, suggesting broad applicability across diverse plant systems.
The implications of these innovations are far-reaching. The production of highly enriched plant extracts opens avenues for research and development in multiple sectors. Besides dietary supplementation, pharmaceutical and cosmetics, Microtom-enriched extracts may lend themselves to salutary applications in medicine, providing a natural radioprotection approach to mitigate cellular damage caused by ionizing radiation (IR) in patients undergoing cancer radiotherapy (RT). RT has arguably proven to be a highly successful treatment modality, either alone or in combination with chemotherapy, surgery, immunotherapy and other novel anti-cancer strategies [22,23,24,25,26,27,28,29]. However, RT very often entails irradiation of healthy tissues and organs at risk, especially in the case of deep-seated solid tumours. The radiosensitivity of these tissues is the factor that limits the maximum dose that can be safely administered to achieve successful tumour control without increasing the occurrence of acute adverse reactions, such as skin erythema, or late complications, such as fibrosis or RT-induced secondary cancers [30]. Consequently, there is a strong need for radioprotectors to effectively counteract such deleterious effects. Plant-derived polyphenols have attracted attention due to their natural origin and expected lower intrinsic cytotoxicity [31]. To act as viable radiomitigators, these compounds must disrupt the molecular pathways responsible for the generation of highly reactive species following IR exposure. Via sustained levels of oxidative stress, these mechanisms can perpetuate the initial damage inflicted to DNA by radiation, leading to long-term genetic instability in the progeny of sublethally damaged normal cells/tissues [32]. ROS and reactive nitrogen species reinforce IR-induced geno- and cyto-toxicity, thereby increasing the risk of sequalae [33]. By counteracting oxidative stress, it is possible to prevent structural and functional alterations of proteins and lipids, too [33]. Persistent oxidative stress, e.g., in the form of elevated ROS levels, may impair lipid metabolism, whose dysregulation is implicated in the alteration of the heterogeneous tumor microenvironment by promoting cancer cell proliferation and perturbing the crosstalk between the redox system and lipid metabolic reprogramming [34].
In the rapidly evolving field of nutraceuticals, natural compounds may therefore combine their ability of protecting normal tissue during curative cancer RT, while offering additional benefits when integrated into the diet of oncological patients or the general population to reduce the risk of cancer and CVD. This work evaluated whether different light-quality regimes, namely fluorescent light (FL) and red-blue (RB), under which tomato fruits were grown, may strengthen the antioxidant profile of Microtom fruits and increase the concentration of polyphenols in tomato extracts. We also investigated in vitro if such polyphenolic extracts may confer radioprotection to IR-exposed normal human cell lines. To this end, we used pre-clinically relevant radiobiological assays to quantify the putative extract-induced reduction in DNA damage and oxidative stress levels following doses of high-energy photons as employed in RT. Preliminary tests conducted on an immortalized human skin cell line showed a significant reduction in radiation-induced micronuclei (MN) frequency when treated with extracts from RB-grown tomatoes at a specific concentration. Encouraged by these results, we extended our analysis to both light conditions, testing two concentrations for each condition on a breast epithelial cell line widely considered as a reliable in-vitro model for normal-tissue radioresponse in breast cancer (BC) [35,36].
2. Materials and Methods
2.1. Plant Growth Conditions and Experimental Design
Dry seeds of Solanum lycopersicum L. cultivar “Microtom”, provided by Holland Online Vof (Amsterdam, The Netherlands), from the same batch, were sown in 1.2 l pots containing potting soil composed of 86% peat, 9% sand, 3% quartz sand, 2% perlite. These pots were kept in darkness until germination. Fifteen days after sowing (DAS), once seedlings had emerged, they were transferred to a growth chamber and exposed to two different light-quality regimes, with five pots per light treatment: fluorescent white (FL) and red-blue (RB). These light regimes were chosen based on previous studies demonstrating enhanced photosynthesis and fruit quality in this cultivar [16,37]. The FL regime was achieved by means of fluorescent tubes (Lumilux L36W/640 and L36W/830, Osram, München, Germany), while RB consisted of a combination of red (R: 60%) and blue (B: 40%) light-emitting diode (LED) arrangements (LedMarket Ltd., Plovdiv, Bulgaria). The spectral profiles of the light treatments (Figure 1) were measured using a SR- 3000A spectroradiometer with 10 nm resolution (Macam Photometrics Ltd., Livingston, Scotland, UK). The Photosynthetic photon flux density in the growth chamber was maintained at 360 μmol photons m-2 s-1 at canopy level for both light conditions, with a 14-h photoperiod. Environmental conditions were controlled at 60-70% relative humidity and day/night air temperature of 25/20 °C ± 2 °C. Plants were watered weekly to field capacity to compensate for evapotranspiration losses and fertilized weekly with half-strength Hoagland's solution. Plant growth was monitored up to 120 DAS until fruit ripening. After being collected, fruits were powdered with liquid nitrogen by a mortar and a pestle, and stored at -80 °C in the Ecology Laboratory (University of Naples Federico II), until further biochemical analyses and extract preparation for radiobiological in-vitro tests on cell cultures.
2.2. Determination of Total Polyphenol, Flavonoid and Anthocyanin Content
Samples (0.200 g) were extracted in methanol. The extracts were kept for 1 h at 4 °C and then centrifuged at 11,000 rpm for 5 min.
To assess the total polyphenol content, methanolic extracts were mixed with 1:1 (v:v) 10% Folin–Ciocâlteu reagent and shaken. After 3 min, the mixture was complemented with 700 mM Na2CO3 solution in the ratio 5:1 (v:v). Samples were incubated for 2 h in the dark. The absorbance was read at 765 nm with a spectrophotometer (UV-VIS Cary 100, Agilent Technologies, Palo Alto, CA, USA) and the polyphenol concentration was estimated calculated using a gallic acid standard curve and expressed as milligrams of gallic acid equivalents per gram of fresh weight (mg GAE g-1 FW)[38].
Total flavonoid content was estimated following the procedures by Moulehi et al.[39] and Sun et al. [40]. The methanolic extracts were mixed with 5% NaNO2 solution and, after 6 min, with 10% AlCl3 and 1 M NaOH solutions. The volume of the mixture was adjusted with distilled water, and the absorbance was read at 510 nm. Total flavonoid content was calculated with a standard catechin (CE) curve and expressed as milligrams of catechin equivalent per gram of fresh weight (mg CE g-1 FW).
The anthocyanin concentration was quantified as reported by Mancinelli et al.[41] and Chung et al. [42]. Samples (0.250 g) were extracted in the dark for 24 h using acidified methanol (1% HCl) at 4 °C. Once centrifuged, the absorbance of supernatants was read at 530 and 657 nm. The anthocyanin content was quantified by the following equation: (µmoL g-1) = [(A530 – 0.33 x A657)/31.6] x [volume (ml)/weight (g)], where the extinction coefficient is 31.6 M-1 cm-1.
2.3. Determination of Total Carotenoid, Lycopene and Ascorbic Acid Content
Total carotenoid content was quantified according to Lichtenthaler [43]. Samples (0.100 g) were extracted with cold 100% acetone and centrifuged (Labofuge GL, Heraeus Sepatech, Hanau, Germany) at 5,000 rpm for 5 min. Absorbance was measured with a spectrophotometer (Cary 100 UV-VIS, Agilent Technologies, Santa Clara, CA, USA) at 470, 645 and 662 nm wavelengths, and carotenoid concentration was expressed as milligrams per gram of fresh weight (mg g-1 FW).
The quantification of lycopene concentration was performed as reported by Fish et al.[44] and Davis et al. [45] with slight modifications. Samples (0.500 g) were mixed in a beaker with hexane: ethanol: 0.05% BHT in acetone (v:v:v, 2:1:1), preventively preparing the solution of butylated hydroxytoluene (BHT) in acetone at the final concentration of 0.05% (w:v). The mixture-containing beaker was then placed on a stirring plate for ~15 min at room temperature. After shaking, deionized water was added to each sample, and the shaking step was repeated for 15 min. The sample was then left at room temperature for 5 min to allow phase separation. The upper hexane layer was transferred into a test tube, and the absorbance of this fraction was read in a 1-cm path length quartz cuvette at 503 nm. The extinction coefficient of lycopene in hexane (17.2 x 104 M-1 cm-1) and the lycopene molecular weight (536.9 g mol-1) were used to convert the absorbance into lycopene concentration expressed as milligrams per gram of fresh weight (mg g-1 FW).
The ascorbic acid (AsA) concentration was determined using the AsA Assay Kit (MAK074, Sigma-Aldrich, St. Louis, MO, USA), following the procedure reported by Costanzo et al. [46]. The sample (10 mg) was homogenized in 4 volumes of cold AsA buffer and then centrifuged at 13,000 rpm for 10 min at 4 °C. The volume of the liquid fraction was adjusted with the AsA assay buffer and mixed with the kit reagents to start the colorimetric coupled enzyme reaction. The colored product developed from the reaction is proportional to the amount of AsA contained in the sample. The absorbance was read at 570 nm and the AsA concentration was calculated using a standard curve and expressed in ng µl-1.
2.4. Antioxidant Capacity (FRAP assay) and Free Radical Scavenging Activity (DPPH Assay)
The antioxidant capacity was assessed through the Ferric Reducing Antioxidant Power (FRAP) assay, according to George et al. [47] and modified by Costanzo et al. [46]. Samples (0.250 g) were extracted in methanol: water solution (60:40, v:v) for 1 h and centrifuged for 15 min at 14,000 rpm at 4 °C. The extracts were mixed with the FRAP reagents and incubated in the dark for 1 h. After the reaction, the absorbance was read at 593 nm. The antioxidant capacity was determined using a Trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid) standard curve and quantified as µmol Trolox equivalents per gram of fresh weight (µmol TE g−1 FW).
The free radical scavenging activity of tomato fruit extracts was evaluated using the 1,1-diphenyl-2-picrylhydrazyl (DPPH) assay, according to Dudonné et al. [48] and modified by Costanzo et al. [46]. An aliquot (67 μl) of methanolic extract was added to 2 ml of 6 × 10-5 M DPPH methanolic solution. The mixture was stirred and incubated at 37 °C for 20 min. The absorbance of the yielded sample was read at 515 nm and converted to DPPH radical inhibition percentage using the following equation:
where Abblank is the absorbance of the blank, and Absample is the absorbance of the tested methanolic extract. Trolox was used as the positive control.
2.5. Cell Cultures
Two cell lines were used in this work. They were grown and serially sub-cultivated according to conventional cell culture techniques. The spontaneously immortalized HaCaT cell line, derived from adult human epidermal cells and retaining the capacity to proliferate and differentiate in a manner similar to basal keratinocytes in vivo, was purchased from the Istituto Zooprofilattico Sperimentale della Lombardia e dell’Emilia Romagna (IZS-LER), Brescia, Italy. HaCaT cells were grown in DMEM (Dulbecco's Modified Eagle Medium), supplemented with 10% fetal bovine serum, 1% penicillin/streptomycin, 1% L-glutamine, and 1% sodium pyruvate (Gibco, ThermoFisher Scientific, US). Non-tumorigenic breast epithelial MCF-10A cells were kindly donated to the Radiation Biophysics Laboratory, University of Naples Federico II, by Dr P. Chaudhary, Queen’s University Belfast, UK. As described in detail by Debnath et al. [49], two DMEM/F12-based culture media (Gibco, ThermoFisher Scientific, US) are required for maintenance of these cells: one formulation is intended for optimal growth, and is enriched with 5% horse serum, endothelial growth factor (20 ng ml-1), hydrocortisone (0.5 µg ml-1), insulin (10 µg ml-1), and cholera toxin (100 ng ml-1), all from Sigma-Aldrich, Merck, Germany; the other, devoid of all supplements but rich in horse serum (20%), is used for routine passaging, i.e. for trypsin quenching and dilutions for cell counting prior to plating. Both cell lines were grown in standard tissue T75 culture flasks placed at 37 °C in a humidified atmosphere with 5% CO2.
2.6. Extract Treatment of Cell Cultures
To evaluate the putative radioprotective properties of tomato fruits grown under FL and RB light regimes, the phenolic extracts were preventively dried at a temperature of 50°C for 30 min in a Rotavapor RII (BÜCHI Labortechnik AG ®, Flawil, Switzerland). The dry material was weighed and solubilized in methanol to reach the concentration of 1000 mg ml-1 (stock solution). The phenolic extracts were then tested at two final concentrations, 100 and 200 mg ml-1, by adding them to the medium of exponentially growing cells in either slide flasks (Nunclon, ThermoFisher Scientific, US, 9 cm-2 growth surface) for DNA damage evaluation by the cytokinesis-block micronucleus (CBMN) assay or in the T25 tissue culture flasks for the ROS assay. Cultures were thus incubated for 24 h and then exposed to radiation in the presence of the extracts. Immediately after irradiation, cells were rinsed thoroughly by phosphate-buffered saline (PBS) solution and subjected to the assay procedure. Cultures irradiated in the absence of the extracts served as controls.
2.7. Cell Exposure to Clinical Photons
Irradiations of cell cultures prepared as above described were performed at Istituto Nazionale Tumori, IRCCS Fondazione G. Pascale, Naples, Italy, by a Synergy Agility LINAC (Elekta, Sweden). Such an accelerator is conceived to deliver RT doses for treatments of oncological patients; therefore, routine quality control checks are performed to make sure the dose delivered adheres to the prescribed one as computed by a Treatment Planning System (TPS). Three-dimensional conformal radiation therapy (3D-CRT) treatment plans were designed specifically for these experiments by means of a dedicated software (Monaco v5.11.03 TPS, Elekta, Sweden). Flasks were imaged by a Computed Treatment (CT) scan in the exact position they were treated.
By means of the CT-image based TPS, the volume to be treated was thus contoured and the calculation of the prescribed radiation dose to be administered was performed. To achieve a homogeneous dose distribution within the irradiated field, the geometric setup consisted of two opposing fields at 0° and 180°. Specifically, two 6-MeV squared photon beams covering a surface of 20 x 20 cm2 were used to deliver doses of 0.5, 2 and 4 Gy to the cells. To ensure adequate build-up during dose delivery by the LINAC rotating gantry, flasks were placed between layers of solid-water slabs of appropriate thickness, namely 2.8 cm for the upper slab and 5 cm for the lower slab. The associated dose uncertainty was estimated to be less than 3%, in line with standard tolerances routinely adopted in RT.
2.8. Measurement of Radiation-Induced DNA Damage
The well-established CBMN genotoxicity assay [50] was performed to evaluate the putative radioprotective effect exerted on DNA by the tomato extracts from plants grown under FL and RB light-quality regimes. In the CBMN assay, DNA damage manifests itself in the form of round portions of DNA, called micronuclei (MN), originated from lagging chromosomes or acentric chromosome fragments at anaphase [51,52] that fail to be incorporated into either of the two daughter nuclei. Based on this, because MN can be associated to the daughter nuclei before cytokinesis ends the telophase stage, they must be scored in binucleated (BN) cells. The amount of IR-induced damage is quantified by the presence of those MN that meet well-defined morphological criteria as described by Fenech et al. [50]. The actin-destroying agent Cytochalasin B (CytB), which inhibits cleavage of the cytoplasmic furrow during cytokinesis, was used for arresting cells at the BN stage. Measurement of the MN frequency in BN HaCaT and MCF-10A cells allowed the evaluation of eventual mitigation of radiation-induced genotoxicity following treatment with the tomato extracts.
2.8.1. Micronucleus Frequency Measurement
After photon irradiation (0, 0.5, 2 and 4 Gy), cells were treated for 24 h with 2.0 μg ml-1 of CytB. These were the optimal treatment duration and CytB concentration as determined by preliminary tests as to yield the highest proportion of BN cells compatibly with the lowest CytB-induced baseline genotoxicity. Subsequently, the CytB-containing medium was removed, cells were rinsed in PBS and fixed for 20 min at -20°C with freshly prepared 4:1 Carnoy's solution (methanol: acetic acid) that had been pre-cooled at -20°C for 30 min. After removal of the fixative solution, slides were air-dried for 24 h and then stained with 14 μl of ProLong Gold DAPI in antifade (ThermoFisher Scientific, US). Controls samples were subjected to the same procedure. The analysis was then manually performed with an Axioplan 2 epi-fluorescence imaging microscope (Carl Zeiss AG, Germany), using a 40X magnification objective. MN frequency was determined in BN cells according to the formula:
where MNn is the number of BN cells with n MN, and BN is the total number of BN cells scored. Between 500 and 1,500 BN cells were scored for each sample, depending on the radiation dose, for statistical robustness.
2.9. Determination of Radiation-Induced Oxidative Stress
The level of oxidative stress was determined in MCF-10A breast epithelial cells to evaluate the antioxidant action exerted by the tomato extracts from plants grown under FL and RB light-quality regimes. Cells were irradiated with the same doses employed for the CBMN assay, i.e., 0, 0.5, 2 and 4 Gy. Oxidative stress was measured by means of a flow cytometer-based assay (Invitrogen Attune NxT Flow, ThermoFisher Scientific, US) using a commercially available kit composed of the fluorogenic CellROX™ Green Reagent for measurement of cellular oxidative stress in live cells, and the SYTOX™ AADvanced™ Dead Cell Stain Kit (both from ThermoFisher Scientific) to discriminate live cells against the dead cell population. Upon oxidation by ROS and subsequent binding to DNA, the cell-permeant CellROX dye, weakly fluorescent while in a reduced state, emits bright green, fluorescent light, with absorption/emission maxima of 485/520 nm.
2.9.1. ROS Level Measurement
Cells were treated 1 h after irradiation. Firstly, they were harvested by trypsinization and centrifuged (200 g for 5 min). The pellet was resuspended in medium using Eppendorf tubes to a concentration of about 2·105 cells ml-1. The staining procedure by the CellROX green (250mM) was carried out for 30 min in the incubator at 37°C, using 0.5 ml of cell suspension containing a final number of 1·10⁵ cells in accordance with the manufacturer’s guidelines; afterwards, the Sytox AADvanced stain (1µl per 1ml of cell suspension) was added for 5 min at room temperature. Flow cytometry analysis followed immediately. “Blank” samples, i.e. without fluorescent probes, were used to set the mean cellular autofluorescence level as to avoid overestimating the presence of intracellular ROS and, hence, to position the quadrants to discriminate between alive non-stressed cells, alive stressed cells, false double positives and dead cells. Cells not treated with the tomato extracts were used as controls and underwent the same experimental procedure.
2.9. Statistical Analysis
Statistical analysis and data representation were performed using Sigma Plot 12.0 (Jandel Scientific, San Rafael, CA, USA). Concerning biochemical investigations, statistically significant differences between FL and RB samples were checked by T-test, based on a significance level of p < 0.05. All data were expressed as means ± SE (n = 8). As regards the radioprotective effect of tomato extracts, within each radiation dose (0, 0.5, 2 and 4 Gy) and for each extract concentration (100 and 200 mg ml-1), FL and RB-treated cell lines were compared to controls by a T-test based on a significance level of p<0.05. The normal distribution of data was verified by Shapiro–Wilk test. Error on the frequency f of MN and on that of ROS-positive cells was assumed to follow a binomial distribution per each individual experiment according to formula 3:
For both CBMN and ROS assays, results were then plotted as the weighted means (formula 4) with relative SE (formula 5) from at least three separate experiments per each cell line, extract concentration, light-regime quality, and radiation dose, the individual weights wi being the inverse of the squared SE:
3. Results
3.1. Total Phenolic Compounds
Different light-quality regimes significantly influenced the total content of polyphenols, flavonoids, and anthocyanins in tomato fruits (Figure 2).
The greatest amount of polyphenols was found in tomato fruits treated with RB light regime (26.3 ± 0.3 mg GAE g−1 FW) compared to FL (23.9 ± 0.3 mg GAE g−1 FW) (Figure 2A). Total flavonoid content varied significantly among RB and FL light regimes, showing higher concentration in RB tomatoes (78 ± 7 mg GAE g−1 FW) than in FL (51 ± 4 mg GAE g−1 FW) (Figure 2B). Conversely, total anthocyanin content increased in tomatoes treated with RB light regime (11.7 ± 0.6 μmol g-1 FW) compared to FL (10.2 ± 0.4 μmol g-1 FW) (Figure 2C).
3.2. Total Carotenoids, Lycopene and Ascorbic Acid
Total carotenoid content varied among different light regimes. RB fruits exhibited a higher concentration of carotenoids (0.110 ± 0.001 µg g-1 FW) than those derived from FL (0.084 ± 0.001 µg g-1FW) (Figure 3A). Total lycopene content varied among different light regimes too. RB fruits exhibited a higher concentration of lycopene (30.4 ± 0.05 µg g-1 FW) than those from FL (26.1 ± 0.1 µg g-1 FW) (Figure 3B). Finally, light quality significantly affected the ascorbic acid (AsA) content (Figure 3C). More specifically, FL regime (1.95 ± 0.17 ng μl-1) showed a higher amount of ascorbic acid compared to RB light (1.40 ± 0.09 ng μl-1).
3.3. Sample Antioxidant Activity
Total antioxidant capacity evaluated by the FRAP assay significantly varied between RB and FL light regimes (Figure 4). Similarly to the case of total polyphenols, the highest value was observed in tomatoes treated with RB light (4.9 ± 0.3 μmol Trolox eq-1 FW) compared to FL (4.1 ± 0.1 μmol Trolox eq g-1 FW). The DPPH radical scavenging assay also confirmed that the antioxidant activity changed according to the different light-regime treatments. More specifically, the highest value was found in tomatoes treated with the RB regime (45.3% of inhibition) compared to FL (38.8% of inhibition).
3.4. Radiation-Induced Genotoxicity
A preliminary series of experiments was carried out in the human keratinocyte HaCaT cell line with the aim to pre-screen a possible radioprotective action by the tomato extracts biochemically characterized as described above. To this end, the anti-genotoxic activity of the RB extract (200 µg ml⁻¹) was tested using the CBMN assay, as shown in Figure 5. Data clearly show that the extract did not increase the baseline MN frequency in unirradiated HaCaT cells (0 Gy), indicating that the RB extract does not cause genotoxicity per se. At all radiation doses, the presence of the RB-grown tomato extract significantly reduced MN frequency compared to cells irradiated without extract. At the lowest doses used, i.e., 2 and 0.5 Gy, RB-treated cells showed a marked reduction in MN frequency, between 30% and 25%, respectively. Although the protective effect appeared to diminish at the highest dose of 4 Gy, the extract-associated reduction in MN frequency remained statistically significant (p<0.05).
Investigation into whether the radioprotective effect observed for the RB-derived extract at the 200 mg ml-1 concentration in the HaCaT cell line could also be detected at lower concentrations, and whether such radioprotection could be influenced by plant growth light regimes, was extended to the breast epithelial cell line MCF-10A, due to its pre-clinical relevance for healthy tissue radioprotection in breast cancer (BC) patients. Modulation of cellular radioresponse by the two concentrations of the extracts derived from fruits grown under FL- and RB-light regimes is presented in Figure 6. Similar to what had been found in HaCaT cells, the extract, irrespective of the light regimes, did not affect the baseline MN frequency in MCF-10A cells. Except for 100 mg ml-1 FL-derived extracts following exposure to 0.5 and 2 Gy of clinical photons (Figure 6A), both extract concentrations and light regimes demonstrated a significant radioprotective effect on cellular DNA compared to extract-untreated irradiated cells (Figure 6). Interestingly, the RB light regime resulted in concentration-dependent radioprotection, which exceeded the magnitude of the FL extract protection at each radiation dose. In detail, at the highest concentration (200 µg ml-1) under RB light, the reduction in mean MN frequency ranged from 30 to 40% relative to untreated controls, consistently outdoing the effect observed at 100 mg ml-1 (Figure 6). For instance, after 0.5 Gy, a dose close to those affecting the healthy tissue in RT, 100 µg ml-1 of RB extract decreased MN frequency by 20% compared to the untreated sample, while at the same radiation dose, a concentration of 200 µg ml-1 achieved a reduction of 39% (Figure 6A vs. Figure 6B). Notably, at the highest tested radiation dose (4 Gy), the MCF-10A cells did not exhibit the decrease in radioprotection measured in the HaCaT cell line (Figure 5).
Figure 7 portrays examples of scored MN in 0.5 Gy-irradiated BN MCF-10A cells treated with 200 µg ml-1 RB and FL extracts: note the presence of multiple MN in extract-untreated BN cells (Figure 7D) and the extract-mediated attenuation of damage in RB- and FL-treated ones as shown Figure 7B,C, respectively.
3.5. Oxidative Stress Levels
The elevation of ROS-associated oxidative stress following exposure of MCF-10A cells to clinical photons was evaluated by the flow cytometry-based CellROX™ assay. Both FL and RB extracts were tested at a concentration of 200 µg ml-1 since this was found by the CBMN assay to provide the greatest reduction in radiation-induced DNA damage (Figure 6). As shown in Figure 8, radiation exposure led to a marked dose-dependent increase in the baseline level of intracellular oxidative stress in extract-untreated samples as measured 1h post irradiation. Importantly, however, tomato extracts were able to significantly reduce such stress levels in both unirradiated controls and irradiated cells. This antioxidant effect was observed for both light regimes although RB generally demonstrated greater efficacy, particularly at higher doses (i.e., at 2 and 4 Gy). Moreover, treatment with RB nearly restored ROS levels in cells exposed to 0.5 and 2 Gy to those measured in unirradiated controls (0 Gy). Specifically, the RB extract reduced ROS levels by approximately 43% at 0.5 Gy and 55% at 2 Gy compared to irradiated cells without treatment.
4. Discussion
This study demonstrates that different light-quality regimes can influence the concentration of bioactive molecules and the antioxidant capacity of Solanum lycopersicum L. cultivar “Microtom” tomato fruits, confirming that modulation of the light spectrum is a valuable tool for enhancing fruit nutritional and functional properties [53]. Notably, for the first time, we provide in vitro evidence that extracts from these tomatoes exhibit pronounced radioprotective activity in irradiated non-cancer cells. This may have important implications for the welfare of cancer patients undergoing radiotherapy (RT). Mitigation of radiation-induced DNA damage and intracellular oxidative stress was proven by a significant reduction in the frequency of micronuclei (MN) and by lowering of the ROS levels by 100 and 200 µg ml-1 of extracts from fruits grown under fluorescence (FL) and red-blue (RB) light regimes.
The total antioxidant capacity of tomato fruits primarily arises from ascorbic acid (AsA), carotenoids, and polyphenols. FL light, for example, enhances AsA content in tomato fruits due to its broader spectrum, including far-red and UV wavelengths that can activate additional photoreceptors and signaling pathways involved in ascorbate biosynthesis. As demonstrated by Hay Mele et al. [19], tomatoes grown under fluorescent or full-spectrum light exhibited significantly higher AsA levels than those cultivated under RB light, underscoring the importance of specific wavelength ranges in modulating antioxidant metabolism. While exposure to blue light is known to increase AsA content in detached tomato fruits compared to white, red, or green light [54,55,56,57,58,59,60,61], our study found that the enhanced antioxidant activity in fruits exposed to RB light was not due to increased AsA. Instead, it was attributed to elevated carotenoids and phenolic compounds [19,38,55]. This aligns with previous observations in the same cultivar, where an increase in total antioxidant capacity under RB regime was linked to a concomitant rise in polyphenols, including flavonoids, anthocyanins, and lycopene among carotenoids, particularly stimulated by red and blue wavelengths [19,38,54,62]. These bioactive compounds enhance the nutritional value of fruits and vegetables and contribute to multiple health benefits, mainly through their antioxidant and anti-inflammatory roles. In tomatoes, the synthesis of polyphenols is strongly modulated by blue and red light, leading to increased antioxidant capacity and ROS scavenging potential. A recent study has demonstrated that tomatoes grown under RB light produce fruits richer in quercitin and derivates, like quercitin rutinoside, quercitin 3-O-Galactoside, and quercitin acetylhexoside [19,38]. These compounds are known for their free radical scavenging activity, modulation of pro-inflammatory cytokines, and inhibition of cancer cell proliferation. Notably, rutin and quercitin rutinoside have been associated with protective effects against cardiovascular diseases [19,63,64]. Additionally, RB tomatoes extracts enriched in chlorogenic acid and caffeoylquinic acid derivates, apigenin and apigenin-8-c-Glucoside, recognized for their anticancer and anti-inflammatory properties [65,66,67,68]. Therefore, the regular intake in daily diet of such antioxidant compounds through fruits and vegetables can counteract ROS, promote human health [69,70], exert antiproliferative and pro-oxidant effects on tumor cells [71], while providing anti-inflammatory activity in normal cells [72].
Despite the encouraging properties against oxidative stress-induced genotoxicity [20], scant data exists on the protective effects of tomato fruit-derived compounds on radiation-induced cytogenotoxicity in the therapeutic context of counteracting radiotoxicity of healthy tissues [73,74]. To our knowledge, this is the first report linking light-dependent metabolic modulation in tomatoes to radioprotective efficacy in normal human cells.The chosen radiobiological endpoints (MN induction and ROS levels) hold valuable pre-clinical relevance. The presence of MN in irradiated cells reflect chromosomal damage and mis-segregation that, if sublethal to the surviving proliferating cell, can be transmitted through cell division, compromising genomic stability and potentially inactivating the newly discovered cGAS-Sting pathway that stimulates systemic immune response [75,76,77]. Similarly, the perpetuation of oxidative stress is a known risk factor for RT-related secondary cancers [60,78].
The varying degrees of radioprotection observed in our study, directly influenced by the modulation of light regimes, indicate that extracts from plants grown under an RB light regime are more effective than those from FL conditions in counteracting radiation-induced genototoxicity (Figure 5 and Figure 6) and oxidative stress (Figure 8). The enhanced efficacy of RB-grown tomatoes appears to correlate to a marked increase in key antioxidant compounds, especially polyphenols. This enrichment is not merely a quantitative change; RB lighting triggers specific photoreceptors and transcriptional programs that enhance the biosynthesis of these molecules, effectively ‘priming’ the fruit to produce higher levels of defensive secondary metabolites, well known for their ability to scavenge radiation-induced ROS, stabilize cellular structures, and mitigate DNA damage in irradiated cells.
In this context, natural compounds can reduce the adverse effects of RT and improve the tolerance to radiation of healthy cells [79]. Antioxidant compounds, as secondary metabolites, play a key role in plants, especially as a defense strategy in response to environmental stressors. Among these, light represents one of the most powerful environmental cues modulating plant metabolism. Different wavelengths can activate transcriptional pathways that lead to the synthesis and accumulation of antioxidant molecules [80,81].
In humans, through the ingestion of fruits and vegetables, these compounds act as antioxidants by counteracting ROS and promoting human welfare [82,83]. Therefore, the modulation of light during plant growth does not only influence agricultural traits but directly impacts on the nutraceutical profile of the harvested fruits, ultimately determining their potential health-promoting properties. Food-isolatable nutraceuticals demonstrate antiproliferative and pro-oxidant effects on tumors [84,85], alongside antioxidant and anti-inflammatory activity in normal cells [86]. Indeed, the attractive double-edged potential of pure polyphenols or polyphenol-enriched extracts could be used to widen the therapeutic window in RT [87] .
At least as regards the radioprotective capacity of tomato fruits, preliminary experiments revealed that extracts obtained from tomatoes grown under RB light regimes at a concentration of 200 µg ml⁻¹, significantly reduced radiation-induced DNA damage in human HaCaT keratinocytes (Figure 5). This prompted us to focus our study on the MCF-10A cell line using the extracts from both growth light regimes. Not only did we confirm their DNA-protecting action against radiation-induced chromosomal damage, but we also found a pronounced ability to reduce radiation-elevated ROS levels. Since such results were obtained following exposure to photons as used in RT in the MCF-10A cell line, which is widely regarded as an in-vitro model for normal-tissue reaction in breast cancer (BC) patients in vivo [35,36], they hold pre-clinical importance. Hence, the extract-induced radioresistance supports the possible therapeutic use of such extracts as bioactive compounds capable of mitigating the adverse effects of radiation. The radiosensitivity of healthy tissues and organs is the main dose-limiting factor in curative RT because of late-occurring consequences such fibrosis and chronic inflammation. Indeed, this aspect is even more critical for BC, where mitigation of radiation-associated cardiotoxicity, often fueled by pro-inflammatory and pro-oxidant responses [88] has emerged as a priority for treatment of this malignancy [89].
It is important to note that even under baseline conditions, that is when cell cultures were not exposed to radiation, the extracts protected cells by lowering physiologically oxidative stress level. Moreover, the extent of protection granted by the extracts towards radiation-induced effects depended on the specific light regime the plants from which they derived had been grown. Thus, FL reduced the percentage of intracellular ROS, albeit less efficiently than RB. The difference between the extracts is significant for doses of 0, 2, and 4 Gy, while it is comparable at 0.5 Gy. Examining the data at 0 Gy in Figure 6 A,B, we notice that the frequency of MN from tomato extracts is comparable to that of the control. While one might expect a similar trend for oxidative stress, this was not the case. This apparent discrepancy can be explained by the fact that the mechanisms underlying MN formation are not exclusively driven by oxidative damage. MN originating from acentric chromosomal fragments are typically linked to direct DNA double-strand break mis-repair, whereas those containing whole chromosomes result from mitotic mis-segregation. Therefore, oxidative stress is only one of several factors contributing to MN induction. Additionally, it ought to be considered that the probe used in the oxidative stress assay simultaneously assesses the presence of ROS in both the mitochondria and the nucleus. At a baseline level, nuclear ROS levels may remain unchanged, while mitochondrial ROS could be more responsive to the antioxidant activity of the extracts, as suggested by previous reports on compartmentalized ROS signaling [90]. This compartmentalization might thus account for the observed divergence between MN and ROS data at 0 Gy.
In conclusion, our results hold significant implications in BC RT scenarios because, by reducing late normal-tissue complications, particularly those linked with RT-induced cardiovascular disease CVD), through a toxicity-free nutraceutical approach, the therapeutic window for BC RT may be enlarged. Furthermore, such an approach may be interesting for the radioprotection of manned space exploration crews, who are exposed to a peculiar radiation field and physiological conditions [91,92,93], and share similar serious long-term health hazards with RT patients, such as increased risk of radiation-induced CVD and cancer, which also invoke bioactive countermeasures [94,95].
These considerations collectively highlight the novelty of our results and justifies further studies on the radiomodulatory properties of Solanum lycopersicum L. cultivar “Microtom” extracts from fruits grown under different light regimes.
5. Conclusions
For the first time this study evaluated in vitro whether phenolic extracts of “Microtom” fruits, grown under fluorescent white (FL) and red-blue (RB) light regimes, may provide radioprotection on normal human cell lines. Our findings suggest that light spectrum quality can profoundly shape the biochemical profile of tomato fruits. The tomato extracts, administered at 100 and 200 µg ml-1, effectively mitigated normal-tissue radiotoxicity in normal breast cells exposed to therapeutic photon beams. Notably, the RB extract at a concentration of 200 µg ml-1, provided the greatest degree of radioprotection. This is in keeping with our findings that tomatoes grown under RB lighting were richer in polyphenols, flavonoids, carotenoids, and lycopene, thereby resulting in higher antioxidant capacity compared to those grown under FL light. These findings underpin the potential of targeted light regimes as powerful tools to produce healthier nutrient-enriched food and their possible usefulness for breast cancer patients undergoing radiotherapy. Overall, our results may represent a starting point for further research aimed at:
- investigating the molecular pathways by which RB light enhances the synthesis of radioprotective polyphenols in tomato fruits;
- testing whether similar light-induced biochemical profiles can be obtained in other edible plant species;
- developing standardized tomato-derived extracts or formulations to reduce radiotherapy-induced normal tissue toxicity.
Author Contributions
Conceptualization, C.A. and L.M.; methodology, F.V., V.C.E. and E.V.; software, F.V., V.C.E., E.V, F.F. and E.F.; formal analysis, C.A., L.M. A.G.; investigation, F.V., V.C.E., G.A., E.V., M.S., P.M. and V.A.; resources, C.A., P.M. and L.M.; data curation, F.V., V.C.E., E.V., E.F and F.F.; writing—original draft preparation, F.V., V.C.E. and E.V.; writing—review and editing, A.G., C.A. and L.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available at request to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest
References
- Salehi, B.; Sharifi-Rad, R.; Sharopov, F.; Namiesnik, J.; Roointan, A.; Kamle, M.; Kumar, P.; Martins, N.; Sharifi-Rad, J. Beneficial Effects and Potential Risks of Tomato Consumption for Human Health: An Overview. Nutrition 2019, 62, 201–208. [Google Scholar] [CrossRef]
- Elbadrawy, E.; Sello, A. Evaluation of Nutritional Value and Antioxidant Activity of Tomato Peel Extracts. Arabian Journal of Chemistry 2016, 9, S1010–S1018. [Google Scholar] [CrossRef]
- Abdullahi, I.I.; Abdullahi, N.; Abdu, A.M.; Ibrahim, A.S. Proximate, Mineral and Vitamin Analysis of Fresh and Canned Tomato. Biosci Biotechnol Res Asia 2016, 13, 1163–1169. [Google Scholar] [CrossRef]
- Chaudhary, P.; Sharma, A.; Singh, B.; Nagpal, A.K. Bioactivities of Phytochemicals Present in Tomato. J Food Sci Technol 2018, 55, 2833–2849. [Google Scholar] [CrossRef]
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.W.; Siervo, M.; Lara, J. Lycopene and Tomato and Risk of Cardiovascular Diseases: A Systematic Review and Meta-Analysis of Epidemiological Evidence. Crit Rev Food Sci Nutr 2019, 59, 141–158. [Google Scholar] [CrossRef]
- Li, N.; Wu, X.; Zhuang, W.; Xia, L.; Chen, Y.; Wu, C.; Rao, Z.; Du, L.; Zhao, R.; Yi, M.; et al. Tomato and Lycopene and Multiple Health Outcomes: Umbrella Review. Food Chem 2021, 343, 128396. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-A.; Hayden, M.M.; Bannerman, S.; Jansen, J.; Crowe-White, K.M. Anti-Apoptotic Effects of Carotenoids in Neurodegeneration. Molecules 2020, 25, 3453. [Google Scholar] [CrossRef]
- Navarro-González, I.; García-Alonso, J.; Periago, M.J. Bioactive Compounds of Tomato: Cancer Chemopreventive Effects and Influence on the Transcriptome in Hepatocytes. J Funct Foods 2018, 42, 271–280. [Google Scholar] [CrossRef]
- Faramarzi, S.; Piccolella, S.; Manti, L.; Pacifico, S. Could Polyphenols Really Be a Good Radioprotective Strategy? Molecules 2021, 26, 4969. [Google Scholar] [CrossRef]
- Dutta Gupta, S.; Jatothu, B. Fundamentals and Applications of Light-Emitting Diodes (LEDs) in in Vitro Plant Growth and Morphogenesis. Plant Biotechnol Rep 2013, 7, 211–220. [Google Scholar] [CrossRef]
- van Iersel, M.W. Optimizing LED Lighting in Controlled Environment Agriculture. In Light Emitting Diodes for Agriculture. In Light Emitting Diodes for Agriculture; Springer Singapore: Singapore, 2017; pp. 59–80. [Google Scholar]
- Hossen, Md.S.; Ali, Md.Y.; Jahurul, M.H.A.; Abdel-Daim, M.M.; Gan, S.H.; Khalil, Md.I. Beneficial Roles of Honey Polyphenols against Some Human Degenerative Diseases: A Review. Pharmacological Reports 2017, 69, 1194–1205. [Google Scholar] [CrossRef]
- Okazaki, M.; Ezura, H. Profiling of Melatonin in the Model Tomato (Solanum Lycopersicum L.) Cultivar Micro-Tom. J Pineal Res 2009, 46, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; Vitale, E.; Hay Mele, B.; Cataletto, P.R.; Turano, M.; Simoniello, P.; De Micco, V. Suitability of Solanum Lycopersicum L. ‘Microtom’ for Growth in Bioregenerative Life Support Systems: Exploring the Effect of High- LET Ionising Radiation on Photosynthesis, Leaf Structure and Fruit Traits. Plant Biol 2019, 21, 615–626. [Google Scholar] [CrossRef]
- Vitale, E.; Velikova, V.; Tsonev, T.; Costanzo, G.; Paradiso, R.; Arena, C. Manipulation of Light Quality Is an Effective Tool to Regulate Photosynthetic Capacity and Fruit Antioxidant Properties of Solanum Lycopersicum L. Cv. ‘Microtom’ in a Controlled Environment. PeerJ 2022, 10, e13677. [Google Scholar] [CrossRef]
- Vitale, E.; Vitale, L.; Costanzo, G.; Velikova, V.; Tsonev, T.; Simoniello, P.; De Micco, V.; Arena, C. Light Spectral Composition Influences Structural and Eco-Physiological Traits of Solanum Lycopersicum L. Cv. ‘Microtom’ in Response to High-LET Ionizing Radiation. Plants 2021, 10, 1752. [Google Scholar] [CrossRef]
- Dutta Gupta, S.; Jatothu, B. Fundamentals and Applications of Light-Emitting Diodes (LEDs) in in Vitro Plant Growth and Morphogenesis. Plant Biotechnol Rep 2013, 7, 211–220. [Google Scholar] [CrossRef]
- van Iersel, M.W. Optimizing LED Lighting in Controlled Environment Agriculture. In Light Emitting Diodes for Agriculture. In Light Emitting Diodes for Agriculture; Springer Singapore: Singapore, 2017; pp. 59–80. [Google Scholar]
- Hay Mele, B.; Vitale, E.; Velikova, V.; Tsonev, T.; Fontanarosa, C.; Spinelli, M.; Amoresano, A.; Arena, C. Harnessing Light Wavelengths to Enrich Health-Promoting Molecules in Tomato Fruits. Int J Mol Sci 2025, 26, 5712. [Google Scholar] [CrossRef] [PubMed]
- Paolillo, I.; Costanzo, G.; Delicato, A.; Villano, F.; Arena, C.; Calabrò, V. Light Quality Potentiates the Antioxidant Properties of Brassica Rapa Microgreen Extracts against Oxidative Stress and DNA Damage in Human Cells. Antioxidants 2023, 12, 1895. [Google Scholar] [CrossRef]
- Costanzo, G.; Vitale, E.; Iesce, M.R.; Spinelli, M.; Fontanarosa, C.; Paradiso, R.; Amoresano, A.; Arena, C. Modulation of Antioxidant Compounds in Fruits of Citrus Reticulata Blanco Using Postharvest LED Irradiation. Biology (Basel) 2023, 12, 1029. [Google Scholar] [CrossRef]
- Abdel-Wahab, M.; Gondhowiardjo, S.S.; Rosa, A.A.; Lievens, Y.; El-Haj, N.; Polo Rubio, J.A.; Prajogi, G. Ben; Helgadottir, H.; Zubizarreta, E.; Meghzifene, A.; et al. Global Radiotherapy: Current Status and Future Directions—White Paper. JCO Glob Oncol 2021, 827–842. [Google Scholar] [CrossRef]
- Elia, V.C.; Fede, F.; Bortolussi, S.; Cansolino, L.; Ferrari, C.; Formicola, E.; Postuma, I.; Manti, L. Fluorescence In Situ Hybridization-Based Chromosome Aberration Analysis Unveils the Mechanistic Basis for Boron-Neutron Capture Therapy’s Radiobiological Effectiveness. Applied Sciences 2024, 14, 1171. [Google Scholar] [CrossRef]
- Schillaci, F.; Anzalone, A.; Cirrone, G.A.P.; Carpinelli, M.; Cuttone, G.; Cutroneo, M.; De Martinis, C.; Giove, D.; Korn, G.; Maggiore, M.; et al. ELIMED, MEDical and Multidisciplinary Applications at ELI-Beamlines. J Phys Conf Ser 2014, 508, 012010. [Google Scholar] [CrossRef]
- Bláha, P.; Feoli, C.; Agosteo, S.; Calvaruso, M.; Cammarata, F.P.; Catalano, R.; Ciocca, M.; Cirrone, G.A.P.; Conte, V.; Cuttone, G.; et al. The Proton-Boron Reaction Increases the Radiobiological Effectiveness of Clinical Low- and High-Energy Proton Beams: Novel Experimental Evidence and Perspectives. Front Oncol 2021, 11. [Google Scholar] [CrossRef]
- Cardamuro, V.; Faramarzi, B.; Moggio, M.; Elia, V.C.; Portaccio, M.; Diano, N.; Manti, L.; Lepore, M. Analysis of the X-Ray Induced Changes in Lipids Extracted from Hepatocarcinoma Cells by Means of ATR-FTIR Spectroscopy. Vib Spectrosc 2024, 132, 103697. [Google Scholar] [CrossRef]
- Turchan, W.T.; Pitroda, S.P.; Weichselbaum, R.R. Treatment of Cancer with Radio-Immunotherapy: What We Currently Know and What the Future May Hold. Int J Mol Sci 2021, 22, 9573. [Google Scholar] [CrossRef] [PubMed]
- Sala, C.; Tarozzi, M.; Simonetti, G.; Pazzaglia, M.; Cammarata, F.P.; Russo, G.; Acquaviva, R.; Cirrone, G.A.P.; Petringa, G.; Catalano, R.; et al. Impact on the Transcriptome of Proton Beam Irradiation Targeted at Healthy Cardiac Tissue of Mice. Cancers (Basel) 2024, 16, 1471. [Google Scholar] [CrossRef]
- Tessonnier, T.; Filosa, D.I.; Karle, C.; Baltazar, F.; Manti, L.; Glimelius, L.; Haberer, T.; Abdollahi, A.; Debus, J.; Mein, S.; et al. First Dosimetric and Biological Verification for Spot-Scanning Hadron Arc Radiation Therapy With Carbon Ions. Adv Radiat Oncol 2024, 9, 101611. [Google Scholar] [CrossRef]
- Kim, J.H.; Jenrow, K.A.; Brown, S.L. Mechanisms of Radiation-Induced Normal Tissue Toxicity and Implications for Future Clinical Trials. Radiat Oncol J 2014, 32, 103. [Google Scholar] [CrossRef]
- Patyar, R.R.; Patyar, S. Role of Drugs in the Prevention and Amelioration of Radiation Induced Toxic Effects. Eur J Pharmacol 2018, 819, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; Jay-Gerin, J.-P.; Pain, D. Ionizing Radiation-Induced Metabolic Oxidative Stress and Prolonged Cell Injury. Cancer Lett 2012, 327, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Sueiro-Benavides, R.A.; Leiro-Vidal, J.M.; Salas-Sánchez, A.Á.; Rodríguez-González, J.A.; Ares-Pena, F.J.; López-Martín, M.E. Radiofrequency at 2.45 GHz Increases Toxicity, pro-Inflammatory and Pre-Apoptotic Activity Caused by Black Carbon in the RAW 264.7 Macrophage Cell Line. Science of The Total Environment 2021, 765, 142681. [Google Scholar] [CrossRef]
- Li, S.; Yuan, H.; Li, L.; Li, Q.; Lin, P.; Li, K. Oxidative Stress and Reprogramming of Lipid Metabolism in Cancers. Antioxidants 2025, 14, 201. [Google Scholar] [CrossRef] [PubMed]
- Bravatà, V.; Minafra, L.; Cammarata, F.P.; Pisciotta, P.; Lamia, D.; Marchese, V.; Petringa, G.; Manti, L.; Cirrone, G.A.; Gilardi, M.C.; et al. Gene Expression Profiling of Breast Cancer Cell Lines Treated with Proton and Electron Radiations. Br J Radiol 2018, 20170934. [Google Scholar] [CrossRef] [PubMed]
- Bravatà, V.; Cammarata, F.P.; Minafra, L.; Pisciotta, P.; Scazzone, C.; Manti, L.; Savoca, G.; Petringa, G.; Cirrone, G.A.P.; Cuttone, G.; et al. Proton-Irradiated Breast Cells: Molecular Points of View. J Radiat Res 2019, 60, 451–465. [Google Scholar] [CrossRef]
- Vitale, E.; Velikova, V.; Tsonev, T.; Costanzo, G.; Paradiso, R.; Arena, C. Manipulation of Light Quality Is an Effective Tool to Regulate Photosynthetic Capacity and Fruit Antioxidant Properties of Solanum Lycopersicum L. Cv. ‘Microtom’ in a Controlled Environment. PeerJ 2022, 10, e13677. [Google Scholar] [CrossRef]
- Vitale, E.; Velikova, V.; Tsonev, T.; Costanzo, G.; Paradiso, R.; Arena, C. Manipulation of Light Quality Is an Effective Tool to Regulate Photosynthetic Capacity and Fruit Antioxidant Properties of Solanum Lycopersicum L. Cv. ‘Microtom’ in a Controlled Environment. PeerJ 2022, 10, e13677. [Google Scholar] [CrossRef]
- Moulehi, I.; Bourgou, S.; Ourghemmi, I.; Tounsi, M.S. Variety and Ripening Impact on Phenolic Composition and Antioxidant Activity of Mandarin (Citrus Reticulate Blanco) and Bitter Orange (Citrus Aurantium L.) Seeds Extracts. Ind Crops Prod 2012, 39, 74–80. [Google Scholar] [CrossRef]
- Sun, B.; Ricardo-da-Silva, J.M.; Spranger, I. Critical Factors of Vanillin Assay for Catechins and Proanthocyanidins. J Agric Food Chem 1998, 46, 4267–4274. [Google Scholar] [CrossRef]
- Mancinelli, A.L.; Rabino, I. Photocontrol of Anthocyanin Synthesis. Plant Physiol 1975, 56, 351–355. [Google Scholar] [CrossRef]
- Chung, Y.-C.; Chen, S.-J.; Hsu, C.-K.; Chang, C.-T.; Chou, S.-T. Studies on the Antioxidative Activity of Graptopetalum Paraguayense E. Walther. Food Chem 2005, 91, 419–424. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In; 1987; pp. 350–382.
- Fish, W.W.; Perkins-Veazie, P.; Collins, J.K. A Quantitative Assay for Lycopene That Utilizes Reduced Volumes of Organic Solvents. Journal of Food Composition and Analysis 2002, 15, 309–317. [Google Scholar] [CrossRef]
- Davis, A.R.; Fish, W.W.; Perkins-Veazie, P. A Rapid Spectrophotometric Method for Analyzing Lycopene Content in Tomato and Tomato Products. Postharvest Biol Technol 2003, 28, 425–430. [Google Scholar] [CrossRef]
- Costanzo, G.; Vitale, E.; Iesce, M.R.; Naviglio, D.; Amoresano, A.; Fontanarosa, C.; Spinelli, M.; Ciaravolo, M.; Arena, C. Antioxidant Properties of Pulp, Peel and Seeds of Phlegrean Mandarin (Citrus Reticulata Blanco) at Different Stages of Fruit Ripening. Antioxidants 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- George, B.; Kaur, C.; Khurdiya, D.S.; Kapoor, H.C. Antioxidants in Tomato (Lycopersium Esculentum) as a Function of Genotype. Food Chem 2004, 84, 45–51. [Google Scholar] [CrossRef]
- Dudonné, S.; Vitrac, X.; Coutiére, P.; Woillez, M.; Mérillon, J.M. Comparative Study of Antioxidant Properties and Total Phenolic Content of 30 Plant Extracts of Industrial Interest Using DPPH, ABTS, FRAP, SOD, and ORAC Assays. J Agric Food Chem 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Debnath, J.; Muthuswamy, S.K.; Brugge, J.S. Morphogenesis and Oncogenesis of MCF-10A Mammary Epithelial Acini Grown in Three-Dimensional Basement Membrane Cultures. Methods 2003, 30, 256–268. [Google Scholar] [CrossRef]
- Fenech, M.; Chang, W.P.; Kirsch-Volders, M.; Holland, N.; Bonassi, S.; Zeiger, E. HUMN Project: Detailed Description of the Scoring Criteria for the Cytokinesis-Block Micronucleus Assay Using Isolated Human Lymphocyte Cultures; 2003; Vol. 534;
- Fenech, M. The Cytokinesis-Block Micronucleus Technique and Its Application to Genotoxicity Studies in Human Populations; 1993; Vol. 101;
- Fenech, M. The in Vitro Micronucleus Technique; 2000; Vol. 455;
- Njus, D.; Kelley, P.M.; Tu, Y.-J.; Schlegel, H.B. Ascorbic Acid: The Chemistry Underlying Its Antioxidant Properties. Free Radic Biol Med 2020, 159, 37–43. [Google Scholar] [CrossRef]
- Ntagkas, N.; Woltering, E.; Nicole, C.; Labrie, C.; Marcelis, L.F.M. Light Regulation of Vitamin C in Tomato Fruit Is Mediated through Photosynthesis. Environ Exp Bot 2019, 158, 180–188. [Google Scholar] [CrossRef]
- Castagna, A.; Dall’Asta, C.; Chiavaro, E.; Galaverna, G.; Ranieri, A. Effect of Post-Harvest UV-B Irradiation on Polyphenol Profile and Antioxidant Activity in Flesh and Peel of Tomato Fruits. Food Bioproc Tech 2014, 7, 2241–2250. [Google Scholar] [CrossRef]
- Xie, B.; Liu, H.; Sun, G.; Chen, R. Effects of Light Quality on The Quality Formation of Tomato Fruits; 2016.
- Racchi, M. Antioxidant Defenses in Plants with Attention to Prunus and Citrus Spp. Antioxidants 2013, 2, 340–369. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef]
- Xu, F.; Shi, L.; Chen, W.; Cao, S.; Su, X.; Yang, Z. Effect of Blue Light Treatment on Fruit Quality, Antioxidant Enzymes and Radical-Scavenging Activity in Strawberry Fruit. Sci Hortic 2014, 175, 181–186. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of Supplemental Light Quality on Growth and Phytochemicals of Baby Leaf Lettuce. Environ Exp Bot 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.; Setiawan, C.K.; Yamawaki, K.; Asai, T.; Nishikawa, F.; Maezawa, S.; Sato, H.; Kanemitsu, N.; Kato, M. Effect of Red and Blue LED Light Irradiation on Ascorbate Content and Expression of Genes Related to Ascorbate Metabolism in Postharvest Broccoli. Postharvest Biol Technol 2014, 94, 97–103. [Google Scholar] [CrossRef]
- Giliberto, L.; Perrotta, G.; Pallara, P.; Weller, J.L.; Fraser, P.D.; Bramley, P.M.; Fiore, A.; Tavazza, M.; Giuliano, G. Manipulation of the Blue Light Photoreceptor Cryptochrome 2 in Tomato Affects Vegetative Development, Flowering Time, and Fruit Antioxidant Content. Plant Physiol 2005, 137, 199–208. [Google Scholar] [CrossRef]
- Ohashi-Kaneko, K.; Takase, M.; Kon, N.; Fujiwara, K.; Kurata, K. Effect of Light Quality on Growth and Vegetable Quality in Leaf Lettuce, Spinach and Komatsuna. Environment Control in Biology 2007, 45, 189–198. [Google Scholar] [CrossRef]
- Olaleye, M.; Crown, O.; Akinmoladun, A.; Akindahunsi, A. Rutin and Quercetin Show Greater Efficacy than Nifedipin in Ameliorating Hemodynamic, Redox, and Metabolite Imbalances in Sodium Chloride-Induced Hypertensive Rats. Hum Exp Toxicol 2014, 33, 602–608. [Google Scholar] [CrossRef] [PubMed]
- da Rocha, E.V.; Falchetti, F.; Pernomian, L.; de Mello, M.M.B.; Parente, J.M.; Nogueira, R.C.; Gomes, B.Q.; Bertozi, G.; Sanches-Lopes, J.M.; Tanus-Santos, J.E.; et al. Quercetin Decreases Cardiac Hypertrophic Mediators and Maladaptive Coronary Arterial Remodeling in Renovascular Hypertensive Rats without Improving Cardiac Function. Naunyn Schmiedebergs Arch Pharmacol 2023, 396, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Bernatova, I. Biological Activities of (−)-Epicatechin and (−)-Epicatechin-Containing Foods: Focus on Cardiovascular and Neuropsychological Health. Biotechnol Adv 2018, 36, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.H.; Alsahli, M.A.; Almatroudi, A.; Almogbel, M.A.; Khan, A.A.; Anwar, S.; Almatroodi, S.A. The Potential Role of Apigenin in Cancer Prevention and Treatment. Molecules 2022, 27, 6051. [Google Scholar] [CrossRef]
- Williamson, G. Protection against Developing Type 2 Diabetes by Coffee Consumption: Assessment of the Role of Chlorogenic Acid and Metabolites on Glycaemic Responses. Food Funct 2020, 11, 4826–4833. [Google Scholar] [CrossRef]
- Sharma, S.; Ali, A.; Ali, J.; Sahni, J.K.; Baboota, S. Rutin: Therapeutic Potential and Recent Advances in Drug Delivery. Expert Opin Investig Drugs 2013, 22, 1063–1079. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, Y.B.; Li, X.; Choi, S.R.; Park, S.; Park, J.S.; Lim, Y.P.; Park, S.U. Accumulation of Phenylpropanoids by White, Blue, and Red Light Irradiation and Their Organ-Specific Distribution in Chinese Cabbage (Brassica Rapa Ssp. Pekinensis). J Agric Food Chem 2015, 63, 6772–6778. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.-R.; Liu, J.; Fu, L.; Shu, T.; Yang, L.; Zhang, X.; Jiang, Z.-H.; Bai, L.-P. Anti-Entry Activity of Natural Flavonoids against SARS-CoV-2 by Targeting Spike RBD. Viruses 2023, 15, 160. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.; Iesce, M.R.; Naviglio, D.; Ciaravolo, M.; Vitale, E.; Arena, C. Comparative Studies on Different Citrus Cultivars: A Revaluation of Waste Mandarin Components. Antioxidants 2020, 9, 517. [Google Scholar] [CrossRef]
- Nakamura, A.; Itaki, C.; Saito, A.; Yonezawa, T.; Aizawa, K.; Hirai, A.; Suganuma, H.; Miura, T.; Mariya, Y.; Haghdoost, S. Possible Benefits of Tomato Juice Consumption: A Pilot Study on Irradiated Human Lymphocytes from Healthy Donors. Nutr J 2017, 16, 27. [Google Scholar] [CrossRef] [PubMed]
- De Ruysscher, D.; Niedermann, G.; Burnet, N.G.; Siva, S.; Lee, A.W.M.; Hegi-Johnson, F. Author Correction: Radiotherapy Toxicity. Nat Rev Dis Primers 2019, 5, 15. [Google Scholar] [CrossRef]
- Manti, L.; Durante, M.; Grossi, G.; Ortenzia, O.; Pugliese, M.; Scampoli, P.; Gialanella, G. Measurements of Metaphase and Interphase Chromosome Aberrations Transmitted through Early Cell Replication Rounds in Human Lymphocytes Exposed to Low-LET Protons and High-LET 12C Ions. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2006, 596, 151–165. [Google Scholar] [CrossRef]
- Manti, L.; Jamali, M.; Prise, K.M.; Michael, B.D.; Trott, K.R. Genomic Instability in Chinese Hamster Cells after Exposure to X Rays or Alpha Particles of Different Mean Linear Energy Transfer. Radiat Res 1997, 147, 22–28. [Google Scholar] [CrossRef]
- Takaki, T.; Millar, R.; Hiley, C.T.; Boulton, S.J. Micronuclei Induced by Radiation, Replication Stress, or Chromosome Segregation Errors Do Not Activate CGAS-STING. Mol Cell 2024, 84, 2203–2213.e5. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, S.; Yang, L.; Song, P.; Liu, Z.; Liu, X.; Yan, X.; Dong, Q. Roles of Reactive Oxygen Species in Inflammation and Cancer. MedComm (Beijing) 2024, 5. [Google Scholar] [CrossRef]
- Faramarzi, S.; Piccolella, S.; Manti, L.; Pacifico, S. Could Polyphenols Really Be a Good Radioprotective Strategy? Molecules 2021, 26, 4969. [Google Scholar] [CrossRef]
- Thongtip, A.; Mosaleeyanon, K.; Janta, S.; Wanichananan, P.; Chutimanukul, P.; Thepsilvisut, O.; Chutimanukul, P. Assessing Light Spectrum Impact on Growth and Antioxidant Properties of Basil Family Microgreens. Sci Rep 2024, 14, 27875. [Google Scholar] [CrossRef] [PubMed]
- Borbély, P.; Gasperl, A.; Pálmai, T.; Ahres, M.; Asghar, M.A.; Galiba, G.; Müller, M.; Kocsy, G. Light Intensity- and Spectrum-Dependent Redox Regulation of Plant Metabolism. Antioxidants 2022, 11, 1311. [Google Scholar] [CrossRef] [PubMed]
- Lugasi, A.; Hóvári, J. Antioxidant Properties of Commercial Alcoholic and Nonalcoholic Beverages. Nahrung/Food 2003, 47, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Marie, A.; Hortresearch, C.; Stephens, M.J.; Hall, H.K.; Alspach, P.A. Variation and Heritabilities of Antioxidant Activity and Total Phenolic Content Estimated from a Red Raspberry Factorial Experiment; 2005; Vol. 130;
- Cárdeno, A.; Sánchez-Hidalgo, M.; Rosillo, M.A.; De La Lastra, C.A. Oleuropein, a Secoiridoid Derived from Olive Tree, Inhibits the Proliferation of Human Colorectal Cancer Cell through Downregulation of HIF-1α. Nutr Cancer 2013, 65, 147–156. [Google Scholar] [CrossRef]
- Benot-Dominguez, R.; Tupone, M.G.; Castelli, V.; d’Angelo, M.; Benedetti, E.; Quintiliani, M.; Cinque, B.; Forte, I.M.; Cifone, M.G.; Ippoliti, R.; et al. Olive Leaf Extract Impairs Mitochondria by Pro-Oxidant Activity in MDA-MB-231 and OVCAR-3 Cancer Cells. Biomedicine and Pharmacotherapy 2021, 134. [Google Scholar] [CrossRef]
- Hahn, D.; Shin, S.H.; Bae, J.S. Natural Antioxidant and Anti-Inflammatory Compounds in Foodstuff or Medicinal Herbs Inducing Heme Oxygenase-1 Expression. Antioxidants 2020, 9, 1–40. [Google Scholar] [CrossRef]
- Pacifico, S.; Bláha, P.; Faramarzi, S.; Fede, F.; Michaličková, K.; Piccolella, S.; Ricciardi, V.; Manti, L. Differential Radiomodulating Action of Olea Europaea L. Cv. Caiazzana Leaf Extract on Human Normal and Cancer Cells: A Joint Chemical and Radiobiological Approach. Antioxidants 2022, 11, 1603. [Google Scholar] [CrossRef]
- Vasbinder, A.; Cheng, R.K.; Heckbert, S.R.; Thompson, H.; Zaslavksy, O.; Chlebowski, R.T.; Shadyab, A.H.; Johnson, L.; Wactawski-Wende, J.; Wells, G.; et al. Chronic Oxidative Stress as a Marker of Long-Term Radiation-Induced Cardiovascular Outcomes in Breast Cancer. J Cardiovasc Transl Res 2023, 16, 403–413. [Google Scholar] [CrossRef]
- Valiyaveettil, D.; Joseph, D.; Malik, M. Cardiotoxicity in Breast Cancer Treatment: Causes and Mitigation. Cancer Treat Res Commun 2023, 37, 100760. [Google Scholar] [CrossRef]
- Murphy, M.P. How Mitochondria Produce Reactive Oxygen Species. Biochemical Journal 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Manti, L.; Durante, M.; Cirrone, G.; Grossi, G.; Lattuada, M.; Pugliese, M.; Sabini, M.; Scampoli, P.; Valastro, L.; Gialanella, G. Modelled Microgravity Does Not Modify the Yield of Chromosome Aberrations Induced by High-Energy Protons in Human Lymphocytes. Int J Radiat Biol 2005, 81, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Manti, L. Does Reduced Gravity Alter Cellular Response to Ionizing Radiation? Radiat Environ Biophys 2006, 45, 1–8. [Google Scholar] [CrossRef]
- Bertucci, A.; Durante, M.; Gialanella, G.; Grossi, G.; Manti, L.; Pugliese, M.; Scampoli, P.; Mancusi, D.; Sihver, L.; Rusek, A. Shielding of Relativistic Protons. Radiat Environ Biophys 2007, 46, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Sishc, B.J.; Zawaski, J.; Saha, J.; Carnell, L.S.; Fabre, K.M.; Elgart, S.R. The Need for Biological Countermeasures to Mitigate the Risk of Space Radiation-Induced Carcinogenesis, Cardiovascular Disease, and Central Nervous System Deficiencies. Life Sci Space Res (Amst) 2022, 35, 4–8. [Google Scholar] [CrossRef]
- Patel, S. The Effects of Microgravity and Space Radiation on Cardiovascular Health: From Low-Earth Orbit and Beyond. IJC Heart & Vasculature 2020, 30, 100595. [Google Scholar] [CrossRef]
Figure 1.
Light spectra used in the experiment. FL, white fluorescent tubes; RB, red-blue, LED. Photon flux density: 360 µmol photons m−2s−1. Irradiance Range: 350–800 nm.
Figure 1.
Light spectra used in the experiment. FL, white fluorescent tubes; RB, red-blue, LED. Photon flux density: 360 µmol photons m−2s−1. Irradiance Range: 350–800 nm.
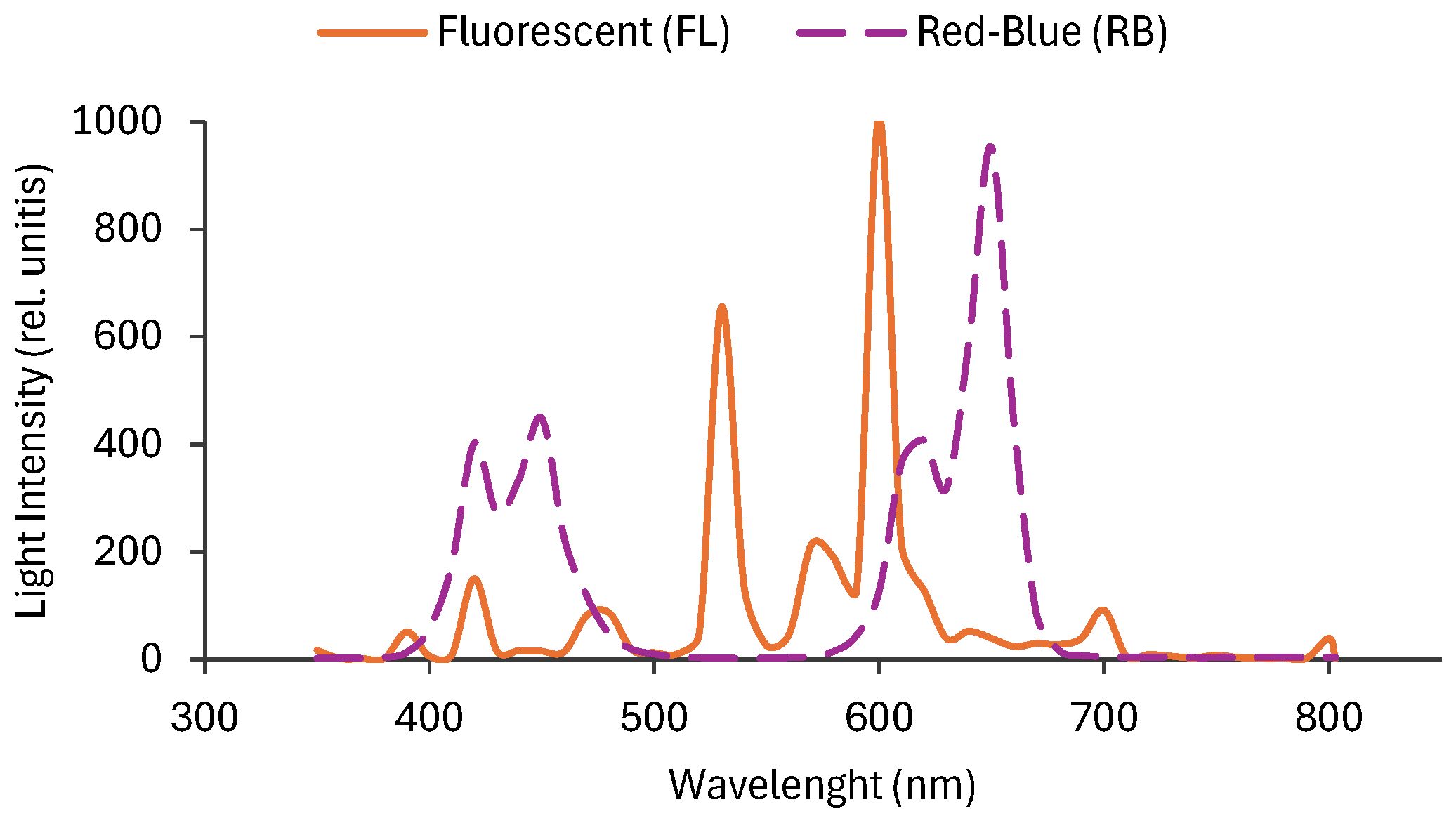
Figure 2.
Total polyphenols (A), total flavonoids (B), total anthocyanins (C) in tomato fruits treated with different light regimes (FL and RB). GAE: gallic acid equivalents; CE: catechin equivalents; FW: fresh weight. Each bar represents the mean ± SE (n = 8). Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001), according to T-test.
Figure 2.
Total polyphenols (A), total flavonoids (B), total anthocyanins (C) in tomato fruits treated with different light regimes (FL and RB). GAE: gallic acid equivalents; CE: catechin equivalents; FW: fresh weight. Each bar represents the mean ± SE (n = 8). Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001), according to T-test.
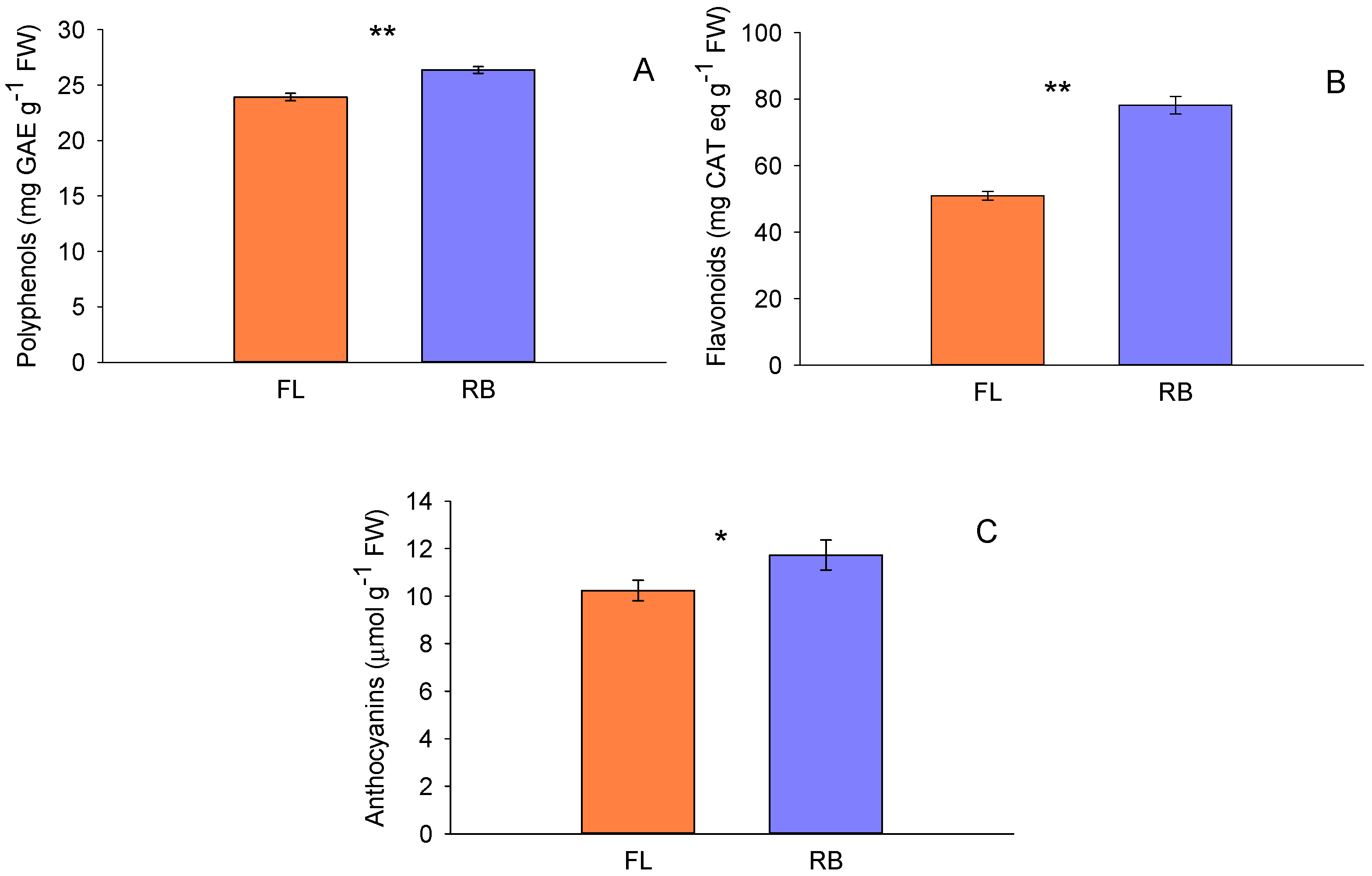
Figure 3.
Total carotenoids (A), total lycopene content (B) and total ascorbic acid (C), in tomato fruits treated with different light regimes (FL and RB). Each bar represents the mean ± SE (n = 8). Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001), according to T-test.
Figure 3.
Total carotenoids (A), total lycopene content (B) and total ascorbic acid (C), in tomato fruits treated with different light regimes (FL and RB). Each bar represents the mean ± SE (n = 8). Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001), according to T-test.
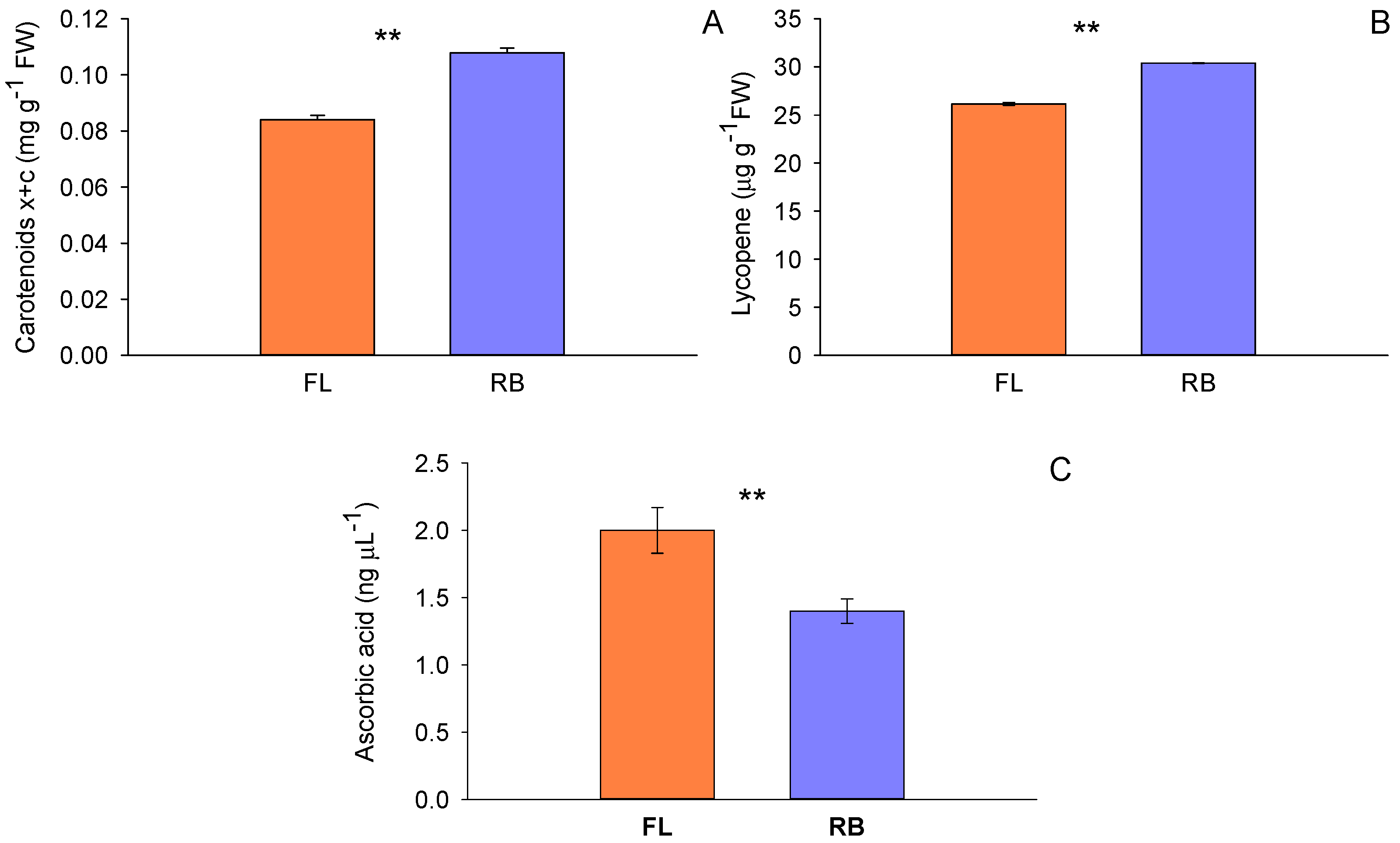
Figure 4.
Total soluble antioxidant capacity (A) and DPPH (1,1-diphenyl-2-picrylhydrazyl) scavenging activity (B) in tomato fruits treated with different light regimes (FL and RB). Each bar represents the mean ± SE (n = 8). TE: Trolox equivalents; FW: fresh weight. Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001), according to T-test.
Figure 4.
Total soluble antioxidant capacity (A) and DPPH (1,1-diphenyl-2-picrylhydrazyl) scavenging activity (B) in tomato fruits treated with different light regimes (FL and RB). Each bar represents the mean ± SE (n = 8). TE: Trolox equivalents; FW: fresh weight. Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001), according to T-test.
Figure 5.
Dose-dependent radiation-induced MN frequency in the human keratinocyte HaCaT cell line treated with RB extract at 200 µg ml-1. The results represent weighted mean values and relative SE from three independent experiments for each radiation dose, significance T-test levels being: *p < 0.05, **p < 0.01, ***p < 0.001. At low and intermediate doses (0.5 and 2 Gy) RB extract showed the highest protection from radiation-induced genotoxicity (p<0.001).
Figure 5.
Dose-dependent radiation-induced MN frequency in the human keratinocyte HaCaT cell line treated with RB extract at 200 µg ml-1. The results represent weighted mean values and relative SE from three independent experiments for each radiation dose, significance T-test levels being: *p < 0.05, **p < 0.01, ***p < 0.001. At low and intermediate doses (0.5 and 2 Gy) RB extract showed the highest protection from radiation-induced genotoxicity (p<0.001).
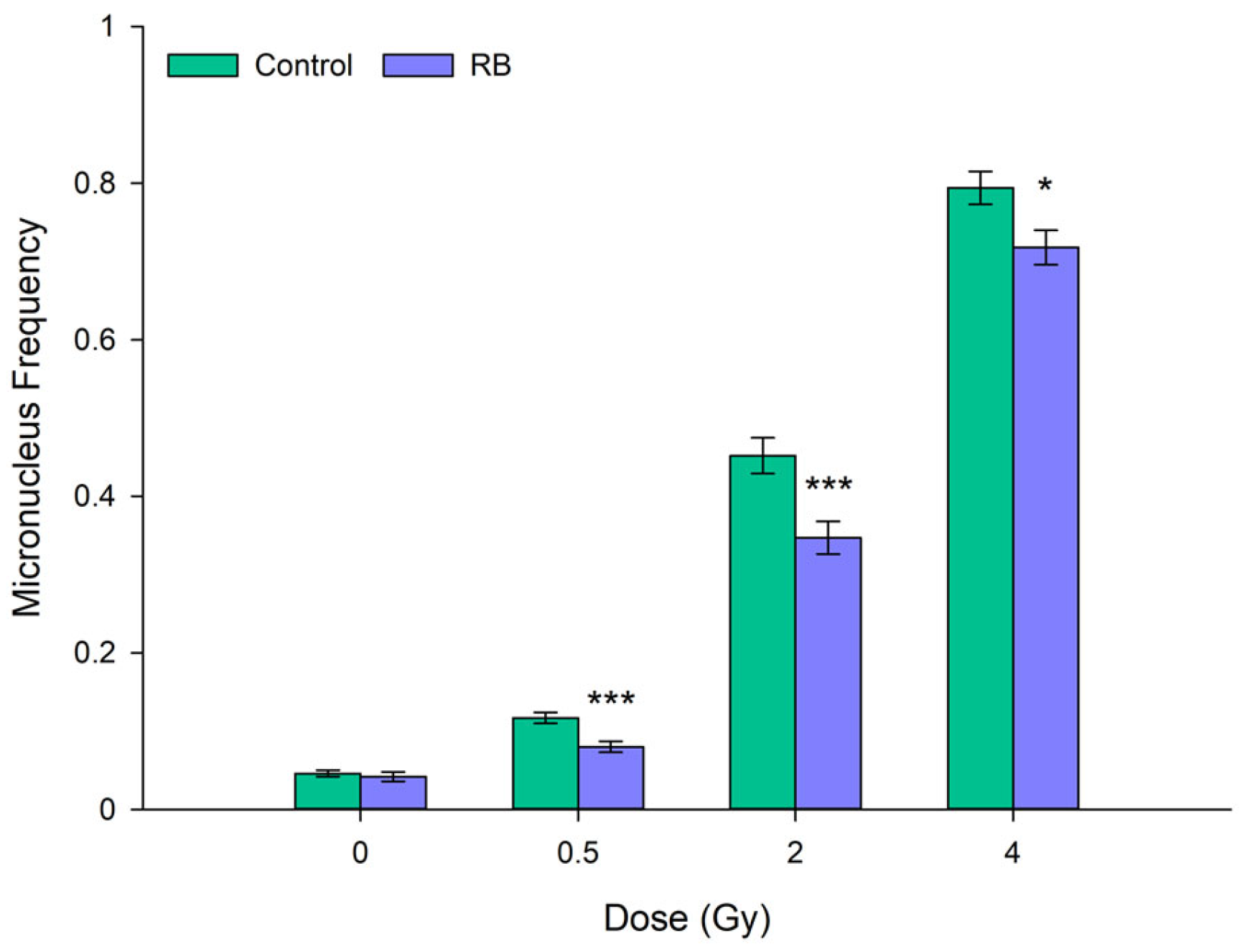
Figure 6.
Dose-dependent MN frequency in the normal breast epithelial MCF-10 cell line, comparing cells untreated (controls) and cells treated with extracts obtained from tomatoes grown under FL and RB light regimes, and tested at two different concentrations, i.e., 100 μg ml-1 (A) and 200 μg ml-1 (B). Results represent weighted averages and relative SE of three independent experiments for each concentration and light regimen. MN frequency values were compared between extract-treated samples and their own non-irradiated controls. Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001).
Figure 6.
Dose-dependent MN frequency in the normal breast epithelial MCF-10 cell line, comparing cells untreated (controls) and cells treated with extracts obtained from tomatoes grown under FL and RB light regimes, and tested at two different concentrations, i.e., 100 μg ml-1 (A) and 200 μg ml-1 (B). Results represent weighted averages and relative SE of three independent experiments for each concentration and light regimen. MN frequency values were compared between extract-treated samples and their own non-irradiated controls. Asterisks indicate statistically significant differences (*p < 0.05, **p < 0.01, ***p < 0.001).
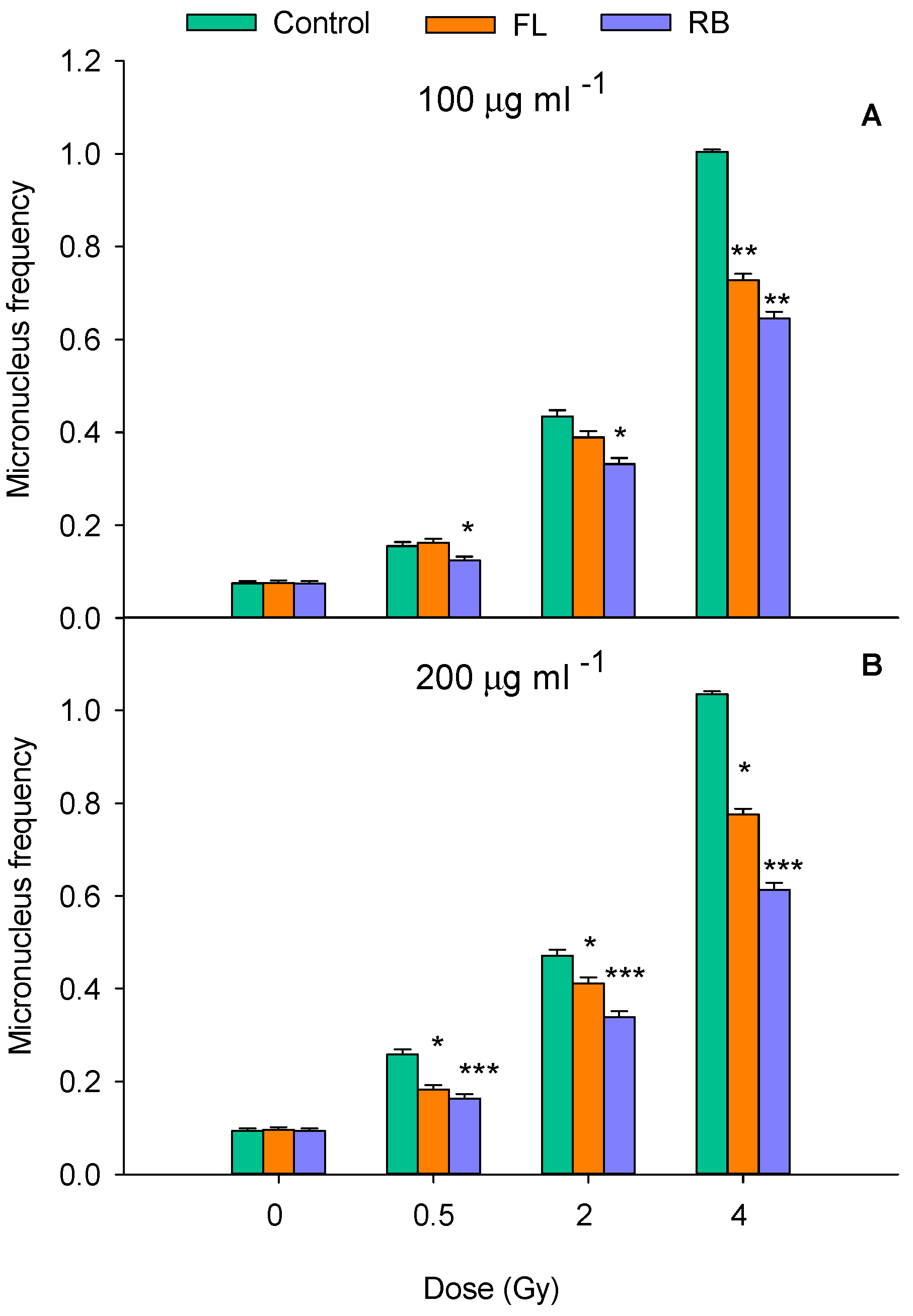
Figure 7.
40× magnification micrograph showing radiation-induced DNA damage in the form of MN in BN MCF-10A cells.
Figure 7.
40× magnification micrograph showing radiation-induced DNA damage in the form of MN in BN MCF-10A cells.
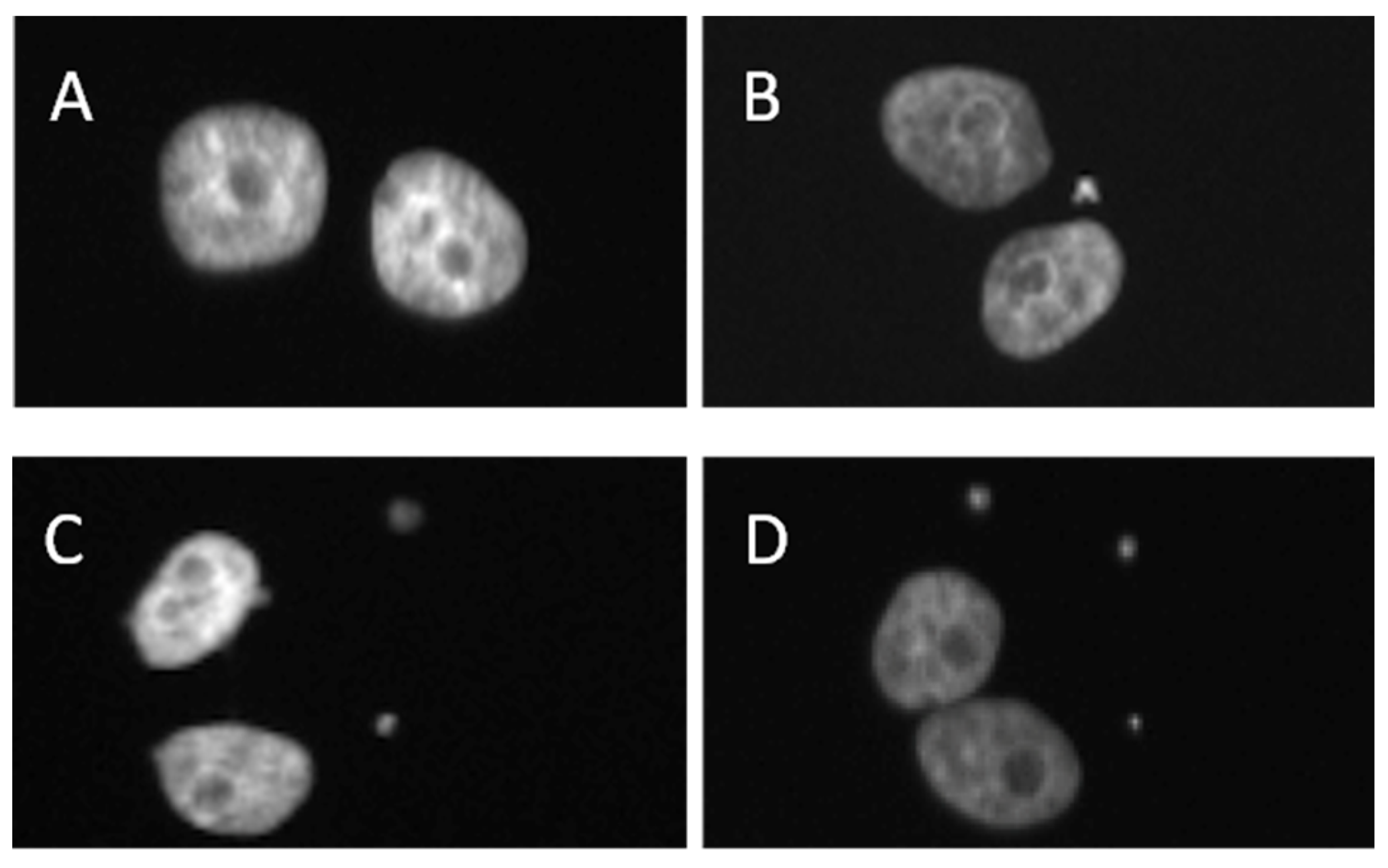
Figure 8.
Oxidative stress expression in MCF-10A cells, either untreated (Control) or treated with extracts obtained from tomatoes grown under FL and RB light regimes at the concentration of 200 mg ml-1 of FL and RB extracts. Results represent weighted averages and relative SE of three independent experiments Here, “***” means a statistically significant difference (p < 0.001) between control and substance; “ ••• ” means a statistically significant difference (p < 0.001) between FL and RB while “ • ”indicates a significance level of <0.05 as reported in previous figure legends.
Figure 8.
Oxidative stress expression in MCF-10A cells, either untreated (Control) or treated with extracts obtained from tomatoes grown under FL and RB light regimes at the concentration of 200 mg ml-1 of FL and RB extracts. Results represent weighted averages and relative SE of three independent experiments Here, “***” means a statistically significant difference (p < 0.001) between control and substance; “ ••• ” means a statistically significant difference (p < 0.001) between FL and RB while “ • ”indicates a significance level of <0.05 as reported in previous figure legends.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



