Submitted:
12 November 2025
Posted:
13 November 2025
Read the latest preprint version here
Abstract
RAPID ALKALINIZATION FACTORS (RALFs) are a family of plant peptide hormones involved in development, reproduction, and response to stresses. These versatile peptides are found throughout land plants, but their molecular mechanisms of action are best understood in the model plant Arabidopsis thaliana. Known to science for more than 20 years, RALFs were initially viewed as apoplastic signaling molecules that elicit intracellular responses through their canonical membrane receptors, the Catharanthus roseus RECEPTOR-LIKE KINASE 1-LIKE (CrRLK1L) family. Recently, it was shown that RALF peptides also have structural roles by binding to LEUCINE-RICH REPEAT EXTENSINS (LRXs) and pectin, forming cell wall-associated complexes. Currently the focus of state-of-the-art science, RALF peptides' central influence on plants still leaves unanswered questions. This work is a detailed review of RALF peptides in A. thaliana, but it also encompasses the literature on other species. As new discoveries in the field are published, this review will be updated.
Keywords:
I. Introduction
II. Structure
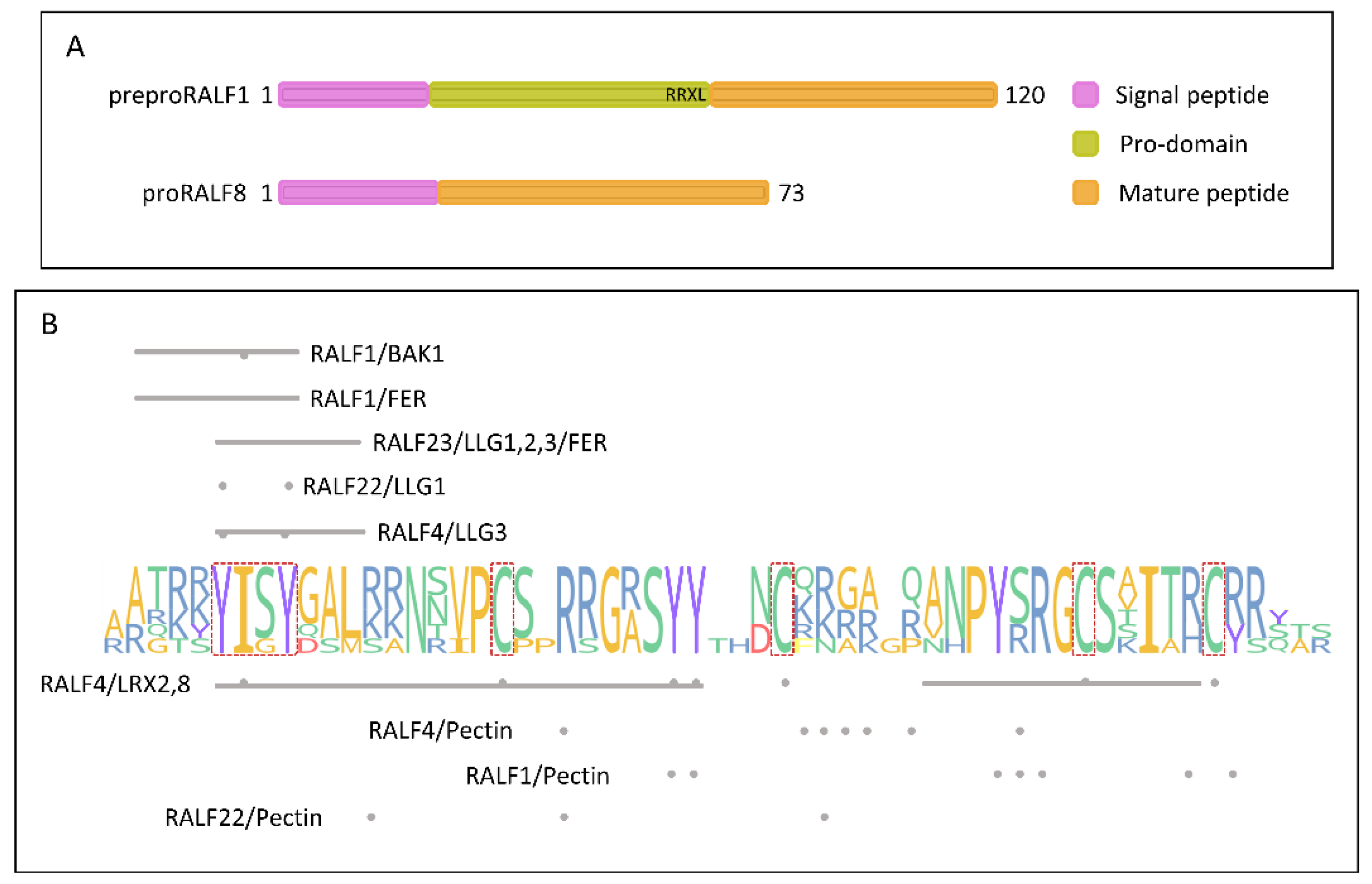
III. Receptors
IV. Rapid Cellular Responses
V. Plant Development
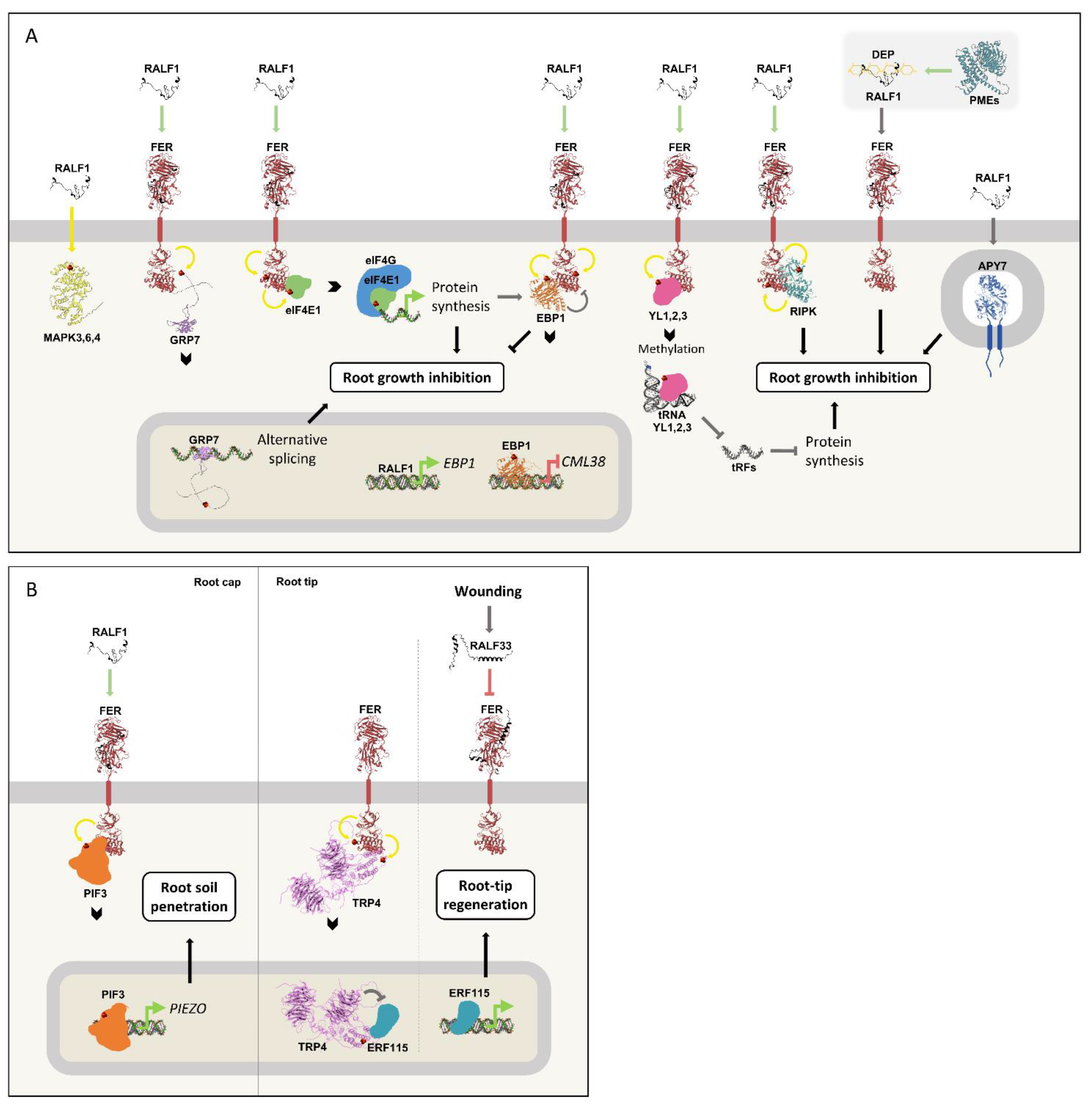
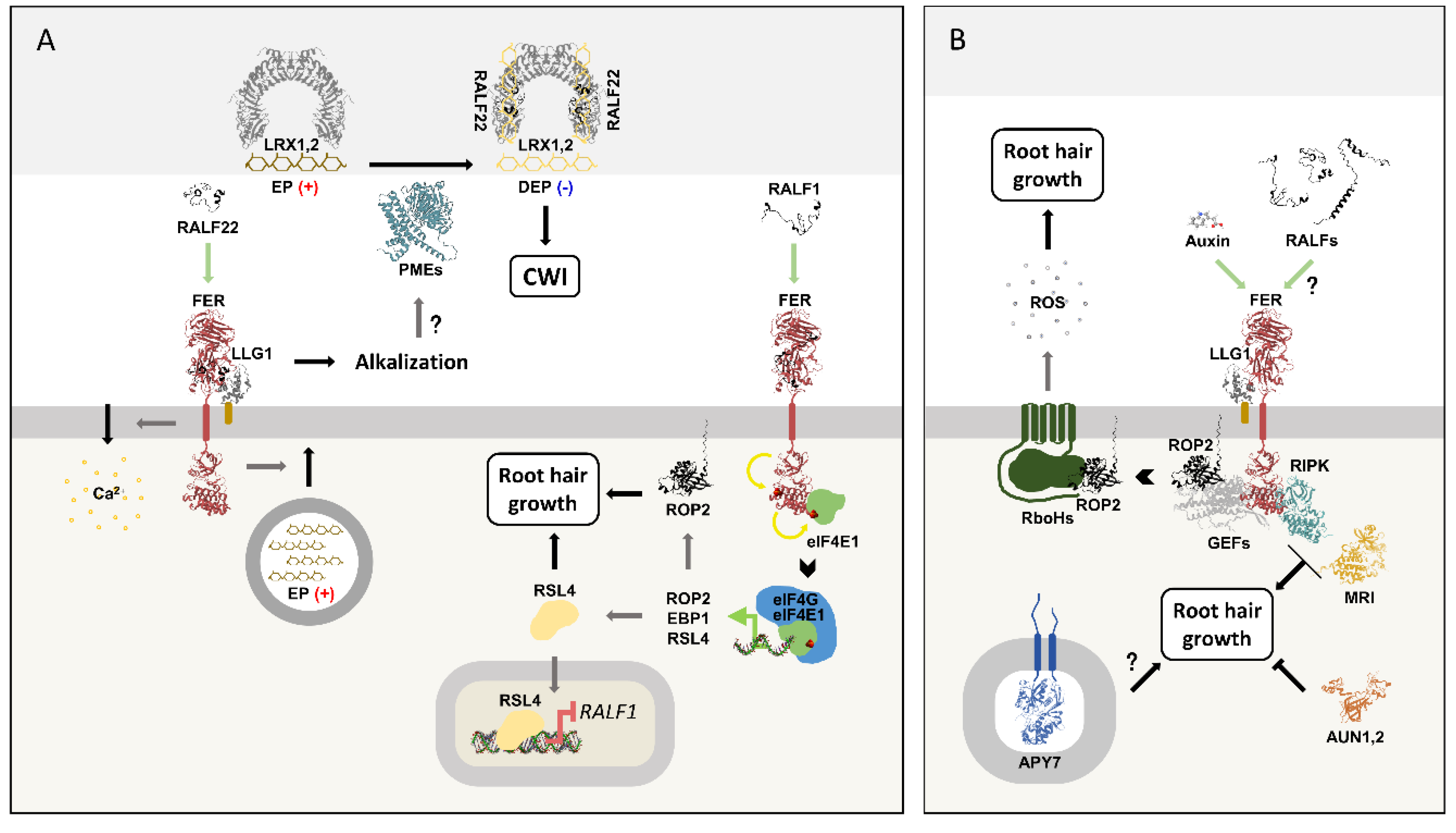
VI. Membrane Dynamics
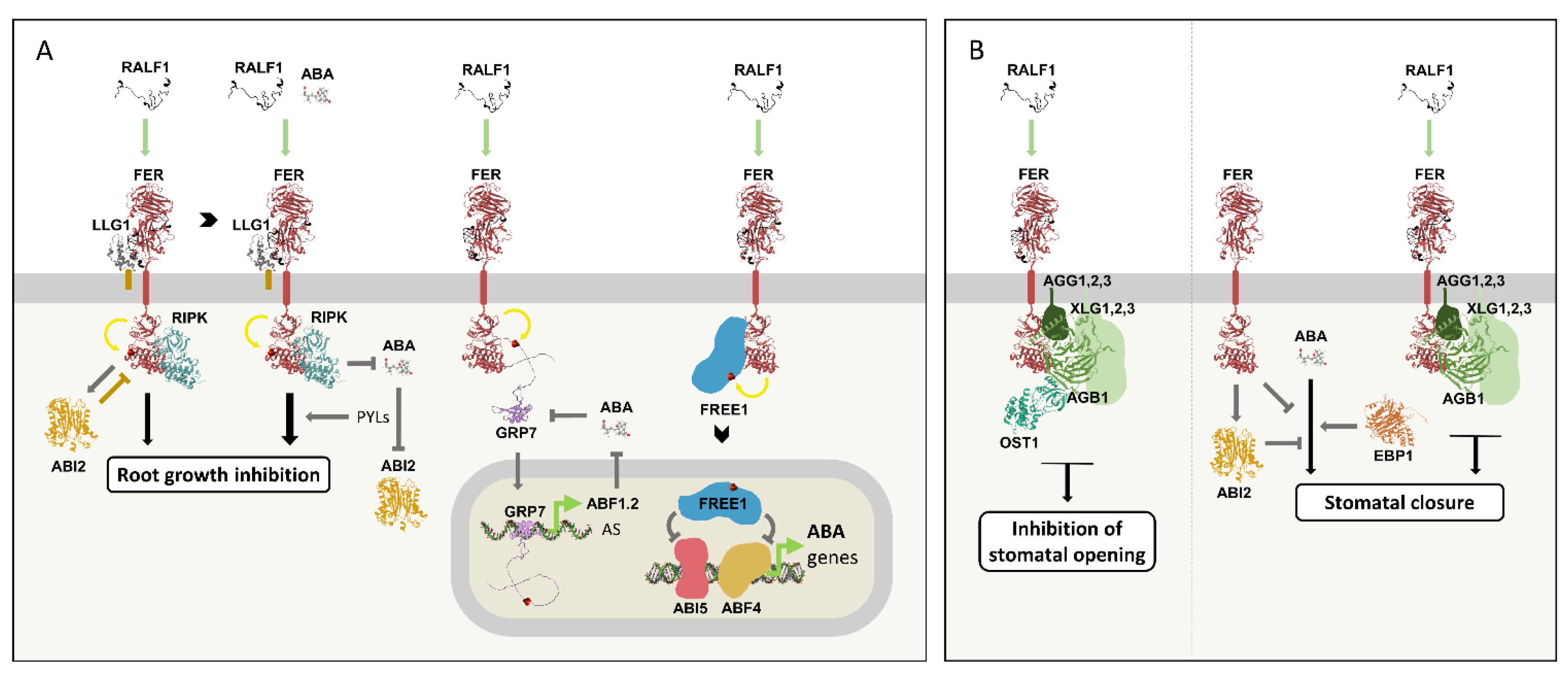
VII. RALF-ABA Crosstalk
VIII. Abiotic Stresses
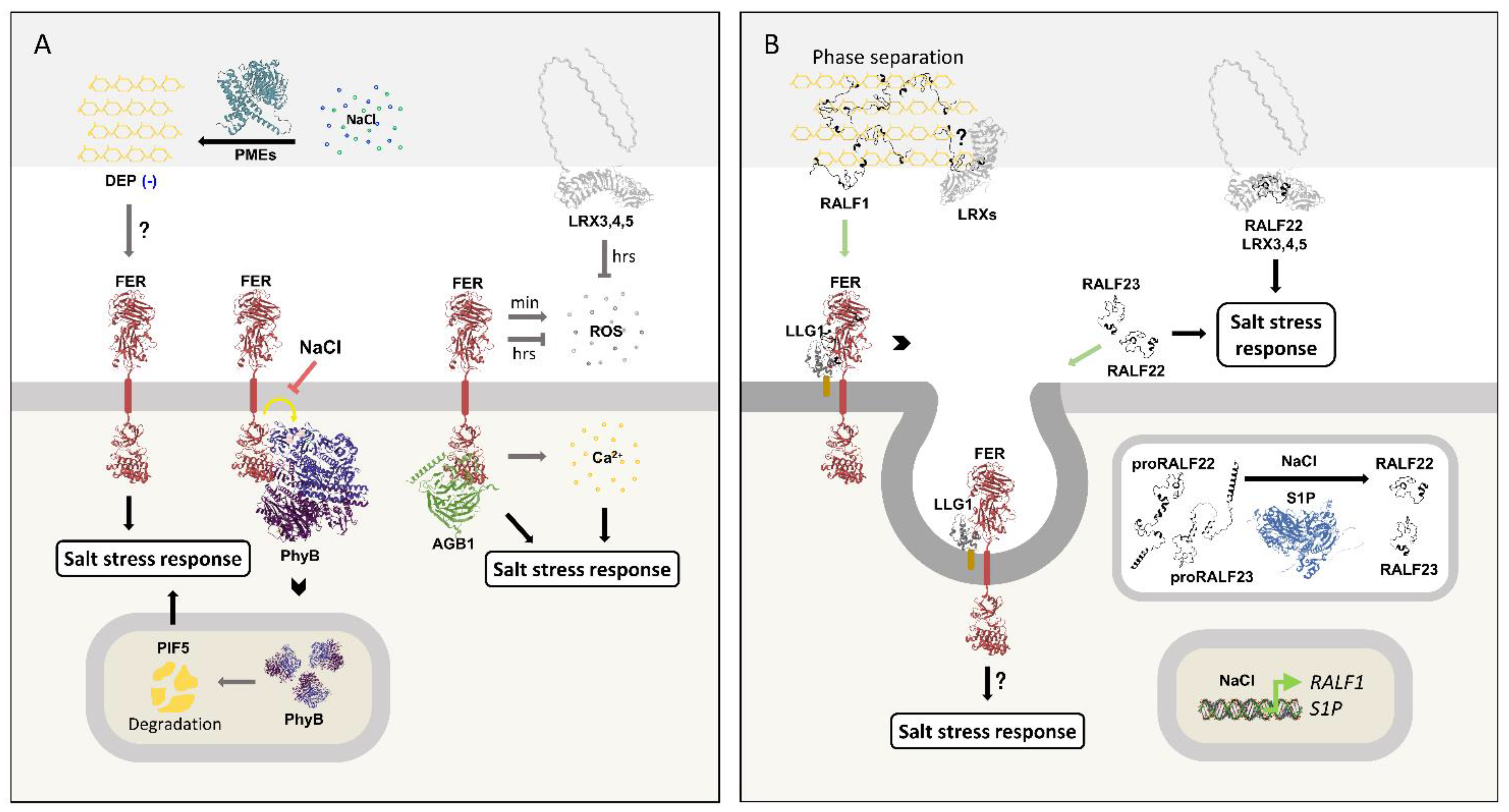
IX. Biotic Stresses
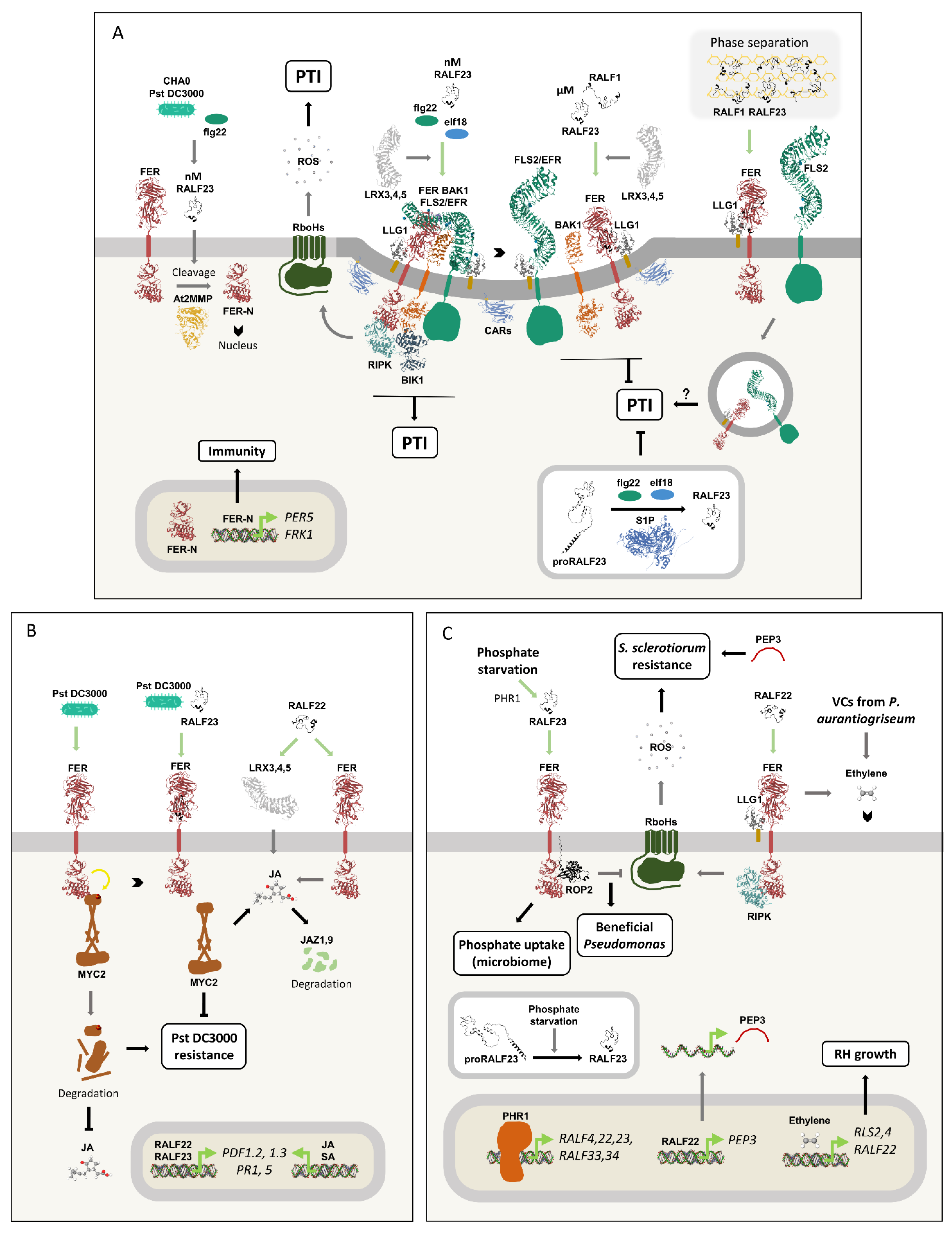
X. Reproduction
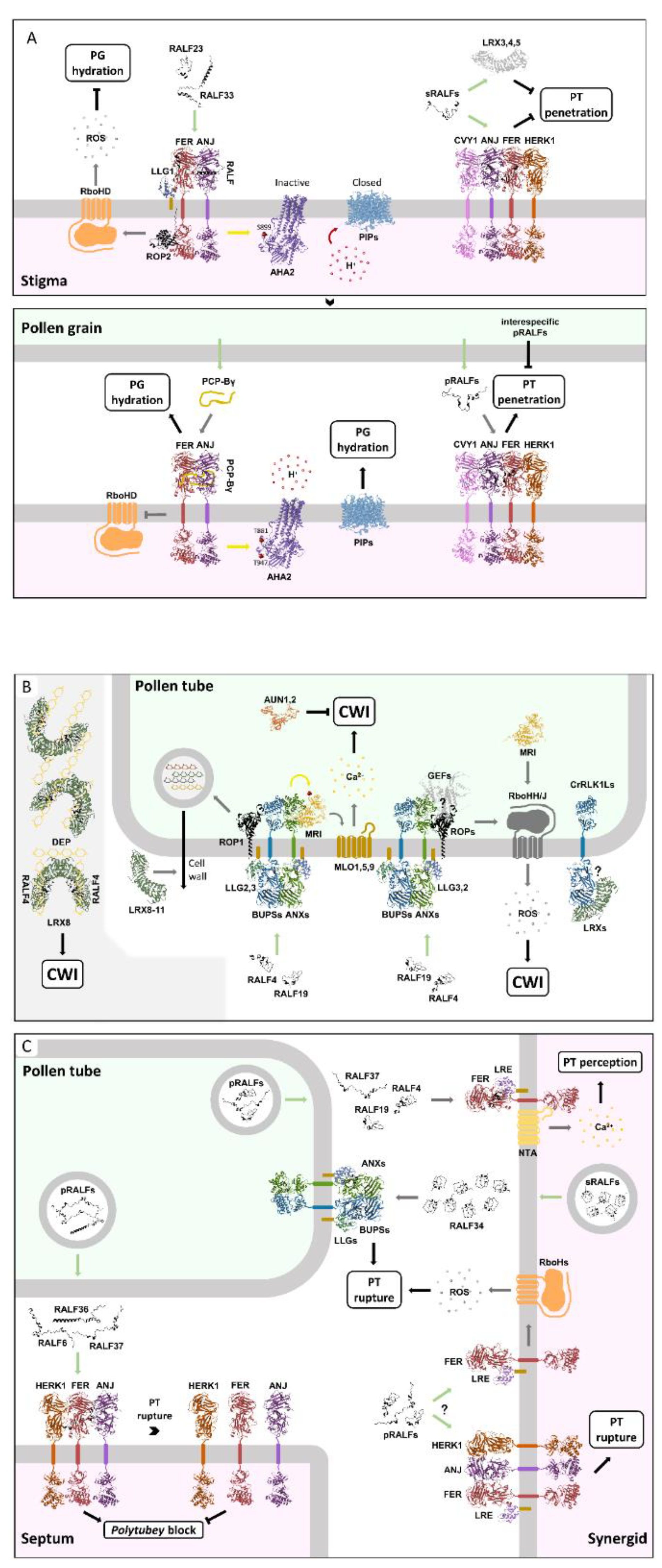
XI. Diversity
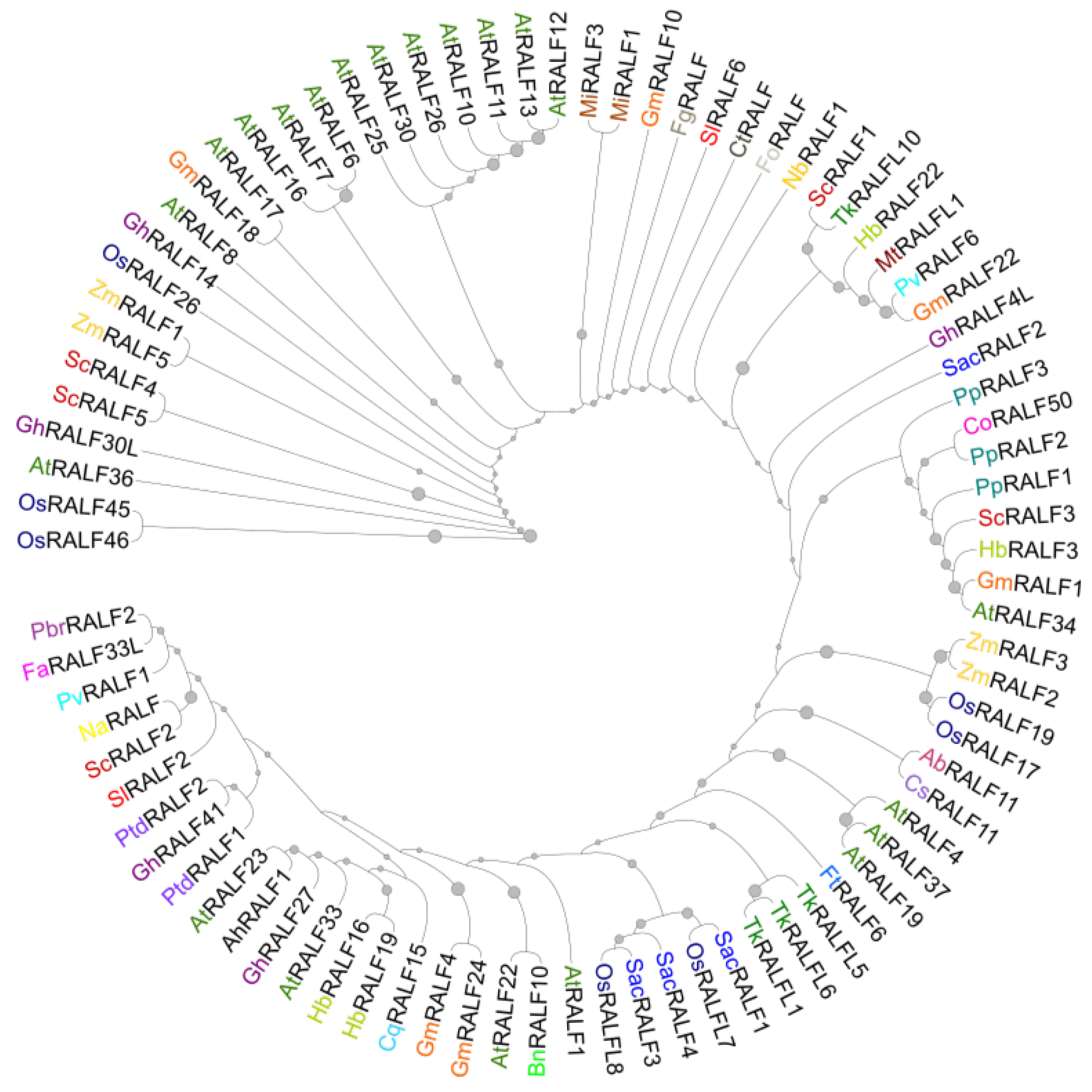
XII. Concluding Remarks
References
- Kende, H.; Zeevaart, J. The Five “Classical” Plant Hormones. Plant Cell. 1997, 9, 1197–1210. [Google Scholar] [CrossRef]
- Pearce, G.; Strydom, D.; Johnson, S.; Ryan, C.A. A Polypeptide from Tomato Leaves Induces Wound-Inducible Proteinase Inhibitor Proteins. Science 1991, 253, 895–897. [Google Scholar] [CrossRef]
- Gancheva, M.S.; Malovichko, Y.V.; Poliushkevich, L.O.; Dodueva, I.E.; Lutova, L.A. Plant Peptide Hormones. Russ. J. Plant Physiol. 2019, 66, 171–189. [Google Scholar] [CrossRef]
- Campbell, L.; Turner, S.R. A Comprehensive Analysis of RALF Proteins in Green Plants Suggests There Are Two Distinct Functional Groups. Front. Plant Sci. 2017, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Abarca, A.; Franck, C.M.; Zipfel, C. Family-wide evaluation of RAPID ALKALINIZATION FACTOR peptides. Plant Physiol. 2021, 187, 996–1010. [Google Scholar] [CrossRef]
- Blackburn, M.R.; Haruta, M.; Moura, D.S. Twenty Years of Progress in Physiological and Biochemical Investigation of RALF Peptides. Plant Physiol. 2020, 182, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Shi, F. Evolution of the RALF Gene Family in Plants: Gene Duplication and Selection Patterns. Evol. Bioinform. 2012, 8, 271–292. [Google Scholar] [CrossRef]
- Thynne, E.; Saur, I.M.L.; Simbaqueba, J.; Ogilvie, H.A.; Gonzalez-Cendales, Y.; Mead, O.; Taranto, A.; Catanzariti, A.; McDonald, M.C.; Schwessinger, B.; et al. Fungal phytopathogens encode functional homologues of plant rapid alkalinization factor (RALF) peptides. Mol. Plant Pathol. 2016, 18, 811–824. [Google Scholar] [CrossRef]
- Zhang, X.; Peng, H.; Zhu, S.; Xing, J.; Li, X.; Zhu, Z.; Zheng, J.; Wang, L.; Wang, B.; Chen, J.; et al. Nematode-Encoded RALF Peptide Mimics Facilitate Parasitism of Plants through the FERONIA Receptor Kinase. Mol. Plant 2020, 13, 1434–1454. [Google Scholar] [CrossRef]
- Pearce, G.; Moura, D.S.; Stratmann, J.; Ryan, C.A. RALF, a 5-kDa ubiquitous polypeptide in plants, arrests root growth and development. Proc. Natl. Acad. Sci. 2001, 98, 12843–12847. [Google Scholar] [CrossRef]
- Wang, L.; Yang, T.; Wang, B.; Lin, Q.; Zhu, S.; Li, C.; Ma, Y.; Tang, J.; Xing, J.; Li, X.; et al. RALF1-FERONIA complex affects splicing dynamics to modulate stress responses and growth in plants. Sci. Adv. 2020, 6, eaaz1622. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Estévez, J.M.; Liao, H.; Zhu, Y.; Yang, T.; Li, C.; Wang, Y.; Li, L.; Liu, X.; Pacheco, J.M.; et al. The RALF1–FERONIA Complex Phosphorylates eIF4E1 to Promote Protein Synthesis and Polar Root Hair Growth. Mol. Plant 2020, 13, 698–716. [Google Scholar] [CrossRef]
- Chen, J.; Xu, F.; Qiang, X.; Liu, H.; Wang, L.; Jiang, L.; Li, C.; Wang, B.; Luan, S.; Wu, D.; et al. Regulated cleavage and translocation of FERONIA control immunity in Arabidopsis roots. Nat. Plants 2024, 10, 1761–1774. [Google Scholar] [CrossRef]
- Matos, J.L.; Fiori, C.S.; Silva-Filho, M.C.; Moura, D.S. A conserved dibasic site is essential for correct processing of the peptide hormone AtRALF1 in Arabidopsis thaliana. FEBS Lett. 2008, 582, 3343–3347. [Google Scholar] [CrossRef]
- Srivastava, R.; Liu, J.; Guo, H.; Yin, Y.; Howell, S.H. Regulation and processing of a plant peptide hormone, AtRALF23, in Arabidopsis. Plant J. 2009, 59, 930–939. [Google Scholar] [CrossRef]
- Stegmann, M.; Monaghan, J.; Smakowska-Luzan, E.; Rovenich, H.; Lehner, A.; Holton, N.; Belkhadir, Y.; Zipfel, C. The receptor kinase FER is a RALF-regulated scaffold controlling plant immune signaling. Science 2017, 355, 287–289. [Google Scholar] [CrossRef]
- Zhao, C.; Zayed, O.; Yu, Z.; Jiang, W.; Zhu, P.; Hsu, C.-C.; Zhang, L.; Tao, W.A.; Lozano-Durán, R.; Zhu, J.-K. Leucine-rich repeat extensin proteins regulate plant salt tolerance in Arabidopsis. Proc. Natl. Acad. Sci. 2018, 115, 13123–13128. [Google Scholar] [CrossRef]
- Xiao, Y.; Stegmann, M.; Han, Z.; DeFalco, T.A.; Parys, K.; Xu, L.; Belkhadir, Y.; Zipfel, C.; Chai, J. Mechanisms of RALF peptide perception by a heterotypic receptor complex. Nature 2019, 572, 270–274. [Google Scholar] [CrossRef]
- Dressano, K.; O Ceciliato, P.H.; Silva, A.L.; Guerrero-Abad, J.C.; Bergonci, T.; Ortiz-Morea, F.A.; Bürger, M.; Silva-Filho, M.C.; Moura, D.S. BAK1 is involved in AtRALF1-induced inhibition of root cell expansion. PLOS Genet. 2017, 13, e1007053. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.; Zhao, Y.; Liu, M.-C.; Zhou, L.-Z.; Wang, L.; Zhong, S.; Hou, S.; Jiang, J.; Liu, T.; Huang, Q.; et al. LLG2/3 Are Co-receptors in BUPS/ANX-RALF Signaling to Regulate Arabidopsis Pollen Tube Integrity. Curr. Biol. 2019, 29, 3256–3265.e5. [Google Scholar] [CrossRef] [PubMed]
- Haruta, M.; Sabat, G.; Stecker, K.; Minkoff, B.B.; Sussman, M.R. A Peptide Hormone and Its Receptor Protein Kinase Regulate Plant Cell Expansion. Science 2014, 343, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Moussu, S.; Broyart, C.; Santos-Fernandez, G.; Augustin, S.; Wehrle, S.; Grossniklaus, U.; Santiago, J. Structural basis for recognition of RALF peptides by LRX proteins during pollen tube growth. Proc. Natl. Acad. Sci. 2020, 117, 7494–7503. [Google Scholar] [CrossRef]
- Gao, Q.; Wang, C.; Xi, Y.; Shao, Q.; Li, L.; Luan, S. A receptor–channel trio conducts Ca2+ signalling for pollen tube reception. Nature 2022, 607, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Lan, Z.; Song, Z.; Wang, Z.; Li, L.; Liu, Y.; Zhi, S.; Wang, R.; Wang, J.; Li, Q.; Bleckmann, A.; et al. Antagonistic RALF peptides control an intergeneric hybridization barrier on Brassicaceae stigmas. Cell 2023, 186, 4773–4787.e12. [Google Scholar] [CrossRef]
- Zhong, S.; Li, L.; Wang, Z.; Ge, Z.; Li, Q.; Bleckmann, A.; Wang, J.; Song, Z.; Shi, Y.; Liu, T.; et al. RALF peptide signaling controls the polytubey block in Arabidopsis. Science 2022, 375, 290–296. [Google Scholar] [CrossRef]
- Bhalla, H.; Sudarsanam, K.; Srivastava, A.; Sankaranarayanan, S. Structural insights into the recognition of RALF peptides by FERONIA receptor kinase during Brassicaceae pollination. Plant Mol. Biol. 2025, 115, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-C.J.; Yeh, F.-L.J.; Yvon, R.; Simpson, K.; Jordan, S.; Chambers, J.; Wu, H.-M.; Cheung, A.Y. Extracellular pectin-RALF phase separation mediates FERONIA global signaling function. Cell 2023, 187, 312–330.e22. [Google Scholar] [CrossRef]
- Moussu, S.; Lee, H.K.; Haas, K.T.; Broyart, C.; Rathgeb, U.; De Bellis, D.; Levasseur, T.; Schoenaers, S.; Fernandez, G.S.; Grossniklaus, U.; et al. Plant cell wall patterning and expansion mediated by protein-peptide-polysaccharide interaction. Science 2023, 382, 719–725. [Google Scholar] [CrossRef]
- Schoenaers, S.; Lee, H.K.; Gonneau, M.; Faucher, E.; Levasseur, T.; Akary, E.; Claeijs, N.; Moussu, S.; Broyart, C.; Balcerowicz, D.; et al. Rapid alkalinization factor 22 has a structural and signalling role in root hair cell wall assembly. Nat. Plants 2024, 10, 494–511. [Google Scholar] [CrossRef]
- Lindner, H.; Müller, L.M.; Boisson-Dernier, A.; Grossniklaus, U. CrRLK1L receptor-like kinases: not just another brick in the wall. Current Opinion in Plant Biology 2012, 15, 659–669. [Google Scholar] [CrossRef]
- Gonneau, M.; Desprez, T.; Martin, M.; Doblas, V.G.; Bacete, L.; Miart, F.; Sormani, R.; Hématy, K.; Renou, J.; Landrein, B.; et al. Receptor Kinase THESEUS1 Is a Rapid Alkalinization Factor 34 Receptor in Arabidopsis. Curr. Biol. 2018, 28, 2452–2458.e4. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Shen, L.; Xiao, Y.; Vyshedsky, D.; Peng, C.; Sun, X.; Liu, Z.; Cheng, L.; Zhang, H.; Han, Z.; et al. Pollen PCP-B peptides unlock a stigma peptide–receptor kinase gating mechanism for pollination. Science 2021, 372, 171–175. [Google Scholar] [CrossRef]
- Feng, W.; Kita, D.; Peaucelle, A.; Cartwright, H.N.; Doan, V.; Duan, Q.; Liu, M.-C.; Maman, J.; Steinhorst, L.; Schmitz-Thom, I.; et al. The FERONIA Receptor Kinase Maintains Cell-Wall Integrity during Salt Stress through Ca2+ Signaling. Curr. Biol. 2018, 28, 666–675.e5. [Google Scholar] [CrossRef]
- Kong, Y.; Chen, J.; Jiang, L.; Chen, H.; Shen, Y.; Wang, L.; Yan, Y.; Zhou, H.; Zheng, H.; Yu, F.; et al. Structural and biochemical basis of Arabidopsis FERONIA receptor kinase-mediated early signaling initiation. Plant Commun. 2023, 4, 100559. [Google Scholar] [CrossRef]
- Feng, H.; Liu, C.; Fu, R.; Zhang, M.; Li, H.; Shen, L.; Wei, Q.; Sun, X.; Xu, L.; Ni, B.; et al. LORELEI-LIKE GPI-ANCHORED PROTEINS 2/3 Regulate Pollen Tube Growth as Chaperones and Coreceptors for ANXUR/BUPS Receptor Kinases in Arabidopsis. Mol. Plant 2019, 12, 1612–1623. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yeh, F.-L.; Cheung, A.Y.; Duan, Q.; Kita, D.; Liu, M.-C.; Maman, J.; Luu, E.J.; Wu, B.W.; Gates, L.; et al. Glycosylphosphatidylinositol-anchored proteins as chaperones and co-receptors for FERONIA receptor kinase signaling in Arabidopsis. eLife 2015, 4. [Google Scholar] [CrossRef]
- Zhou, K. Glycosylphosphatidylinositol-Anchored Proteins in Arabidopsis and One of Their Common Roles in Signaling Transduction. Front. Plant Sci. 2019, 10, 1022. [Google Scholar] [CrossRef] [PubMed]
- Mecchia, M.A.; Santos-Fernandez, G.; Duss, N.N.; Somoza, S.C.; Boisson-Dernier, A.; Gagliardini, V.; Martínez-Bernardini, A.; Fabrice, T.N.; Ringli, C.; Muschietti, J.P.; et al. RALF4/19 peptides interact with LRX proteins to control pollen tube growth in Arabidopsis. Science 2017, 358, 1600–1603. [Google Scholar] [CrossRef]
- Dünser, K.; Gupta, S.; Herger, A.; I Feraru, M.; Ringli, C.; Kleine-Vehn, J. Extracellular matrix sensing by FERONIA and Leucine-Rich Repeat Extensins controls vacuolar expansion during cellular elongation in Arabidopsis thaliana. EMBO J. 2019, 38. [Google Scholar] [CrossRef]
- Herger, A.; Gupta, S.; Kadler, G.; Franck, C.M.; Boisson-Dernier, A.; Ringli, C. Overlapping functions and protein-protein interactions of LRR-extensins in Arabidopsis. PLOS Genet. 2020, 16, e1008847. [Google Scholar] [CrossRef]
- Ge, Z.; Bergonci, T.; Zhao, Y.; Zou, Y.; Du, S.; Liu, M.-C.; Luo, X.; Ruan, H.; García-Valencia, L.E.; Zhong, S.; et al. Arabidopsis pollen tube integrity and sperm release are regulated by RALF-mediated signaling. Science 2017, 358, 1596–1600. [Google Scholar] [CrossRef]
- Campos, W.F.; Dressano, K.; Ceciliato, P.H.; Guerrero-Abad, J.C.; Silva, A.L.; Fiori, C.S.; Canto, A.M.D.; Bergonci, T.; Claus, L.A.; Silva-Filho, M.C.; et al. Arabidopsis thaliana rapid alkalinization factor 1–mediated root growth inhibition is dependent on calmodulin-like protein 38. J. Biol. Chem. 2018, 293, 2159–2171. [Google Scholar] [CrossRef]
- Li, L.; Chen, H.; Alotaibi, S.S.; Pěnčík, A.; Adamowski, M.; Novák, O.; Friml, J. RALF1 peptide triggers biphasic root growth inhibition upstream of auxin biosynthesis. Proc. Natl. Acad. Sci. 2022, 119. [Google Scholar] [CrossRef]
- Liu, Z.; Chu, X.; Ren, W.; Cheng, L.; Liu, C.; Wang, C.; Gao, S.; Dai, S.; Li, C. PCP-B peptides and CrRLK1L receptor kinases control pollination via pH gating of aquaporins in Arabidopsis. Dev. Cell 2025, 60, 1336–1347.e5. [Google Scholar] [CrossRef]
- Gjetting, S.K.; Mahmood, K.; Shabala, L.; Kristensen, A.; Shabala, S.; Palmgren, M.; et al. Evidence for multiple receptors mediating RALF-triggered Ca2+ signaling and proton pump inhibition. The Plant Journal 2020, 104, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Hager, A.; Menzel, H.; Krauss, A. Versuche und Hypothese zur Prim rwirkung des Auxins beim Streckungswachstum. Planta 1971, 100, 47–75. [Google Scholar] [CrossRef]
- Du, M.; Spalding, E.P.; Gray, W.M. Rapid Auxin-Mediated Cell Expansion. Annu. Rev. Plant Biol. 2020, 71, 379–402. [Google Scholar] [CrossRef] [PubMed]
- Fuglsang, A.T.; Kristensen, A.; Cuin, T.A.; Schulze, W.X.; Persson, J.; Thuesen, K.H.; Ytting, C.K.; Oehlenschlæger, C.B.; Mahmood, K.; Sondergaard, T.E.; et al. Receptor kinase-mediated control of primary active proton pumping at the plasma membrane. Plant J. 2014, 80, 951–964. [Google Scholar] [CrossRef]
- Kudla, J.; Becker, D.; Grill, E.; Hedrich, R.; Hippler, M.; Kummer, U.; Parniske, M.; Romeis, T.; Schumacher, K. Advances and current challenges in calcium signaling. New Phytol. 2018, 218, 414–431. [Google Scholar] [CrossRef] [PubMed]
- Day, I.S.; Reddy, V.S.; Ali, G.S.; Reddy, A. Analysis of EF-hand-containing proteins in Arabidopsis. Genome Biol. 2002, 3. [Google Scholar] [CrossRef]
- Gilroy, S.; Suzuki, N.; Miller, G.; Choi, W.-G.; Toyota, M.; Devireddy, A.R.; Mittler, R. A tidal wave of signals: calcium and ROS at the forefront of rapid systemic signaling. Trends Plant Sci. 2014, 19, 623–630. [Google Scholar] [CrossRef]
- Haruta, M.; Monshausen, G.; Gilroy, S.; Sussman, M.R. A Cytoplasmic Ca2+ Functional Assay for Identifying and Purifying Endogenous Cell Signaling Peptides in Arabidopsis Seedlings: Identification of AtRALF1 Peptide. Biochemistry 2008, 47, 6311–6321. [Google Scholar] [CrossRef] [PubMed]
- Frederick, R.O.; Haruta, M.; Tonelli, M.; Lee, W.; Cornilescu, G.; Cornilescu, C.C.; Sussman, M.R.; Markley, J.L. Function and solution structure of the Arabidopsis thaliana RALF8 peptide. Protein Sci. 2019, 28, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Wang, C.; Xi, Y.; Shao, Q.; Hou, C.; Li, L.; Luan, S. RALF signaling pathway activates MLO calcium channels to maintain pollen tube integrity. Cell Res. 2023, 33, 71–79. [Google Scholar] [CrossRef]
- Morato Do Canto, A.; Ceciliato, P.H.O.; Ribeiro, B.; Ortiz Morea, F.A.; Franco Garcia, A.A.; Silva-Filho, M.C.; et al. Biological activity of nine recombinant AtRALF peptides: Implications for their perception and function in Arabidopsis. Plant Physiology and Biochemistry 2014, 75, 45–54. [Google Scholar] [CrossRef]
- Yu, Y.; Assmann, S.M. Inter-relationships between the heterotrimeric Gβ subunit AGB1, the receptor-like kinase FERONIA, and RALF1 in salinity response. Plant Cell & Environment 2018, 41, 2475–2489. [Google Scholar]
- He, Y.; Chen, S.; Chen, X.; Xu, Y.; Liang, Y.; Cai, X. RALF22 promotes plant immunity and amplifies the Pep3 immune signal. J. Integr. Plant Biol. 2023, 65, 2519–2534. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhao, L.; Qi, F.; Htwe, N.M.P.S.; Li, Q.; Zhang, D.; Lin, F.; Shang-Guan, K.; Liang, Y. The receptor-like cytoplasmic kinase RIPK regulates broad-spectrum ROS signaling in multiple layers of plant immune system. Mol. Plant 2021, 14, 1652–1667. [Google Scholar] [CrossRef]
- Berken, A.; Thomas, C.; Wittinghofer, A. A new family of RhoGEFs activates the Rop molecular switch in plants. Nature 2005, 436, 1176–1180. [Google Scholar] [CrossRef]
- Smokvarska, M.; Bayle, V.; Maneta-Peyret, L.; Fouillen, L.; Poitout, A.; Dongois, A.; Fiche, J.-B.; Gronnier, J.; Garcia, J.; Höfte, H.; et al. The receptor kinase FERONIA regulates phosphatidylserine localization at the cell surface to modulate ROP signaling. Sci. Adv. 2023, 9. [Google Scholar] [CrossRef]
- Song, Y.; Wilson, A.J.; Zhang, X.-C.; Thoms, D.; Sohrabi, R.; Song, S.; Geissmann, Q.; Liu, Y.; Walgren, L.; He, S.Y.; et al. FERONIA restricts Pseudomonas in the rhizosphere microbiome via regulation of reactive oxygen species. Nat. Plants 2021, 7, 644–654. [Google Scholar] [CrossRef]
- Bergonci, T.; Ribeiro, B.; Ceciliato, P.H.; Guerrero-Abad, J.C.; Silva-Filho, M.C.; Moura, D.S. Arabidopsis thaliana RALF1 opposes brassinosteroid effects on root cell elongation and lateral root formation. J. Exp. Bot. 2014, 65, 2219–2230. [Google Scholar] [CrossRef]
- Friml, J. Auxin transport — shaping the plant. Current Opinion in Plant Biology 2003, 6, 7–12. [Google Scholar] [CrossRef]
- Friml, J.; Wiśniewska, J.; Benková, E.; Mendgen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 2002, 415, 806–809. [Google Scholar] [CrossRef]
- Petricka, J.J.; Winter, C.M.; Benfey, P.N. Control of Arabidopsis Root Development. Annu. Rev. Plant Biol. 2012, 63, 563–590. [Google Scholar] [CrossRef]
- Zhao, Y. Auxin Biosynthesis: A Simple Two-Step Pathway Converts Tryptophan to Indole-3-Acetic Acid in Plants. Mol. Plant 2012, 5, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Li, X.; Chen, J.; Chen, W.; Li, B.; Li, C.; Wang, L.; Li, J.; Zhao, X.; Lin, J.; et al. Receptor kinase complex transmits RALF peptide signal to inhibit root growth in Arabidopsis. Proc. Natl. Acad. Sci. 2016, 113, E8326–E8334. [Google Scholar] [CrossRef] [PubMed]
- Barbez, E.; Dünser, K.; Gaidora, A.; Lendl, T.; Busch, W. Auxin steers root cell expansion via apoplastic pH regulation in Arabidopsis thaliana. Proc. Natl. Acad. Sci. 2017, 114, E4884–E4893. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Zhang, Z.; Liu, Y.; Tao, L.; Liu, H. FERONIA regulates auxin-mediated lateral root development and primary root gravitropism. FEBS Lett. 2018, 593, 97–106. [Google Scholar] [CrossRef]
- Li, E.; Wang, G.; Zhang, Y.; Kong, Z.; Li, S. FERONIA mediates root nutating growth. Plant J. 2020, 104, 1105–1116. [Google Scholar] [CrossRef]
- Li, C.; Chen, J.; Li, X.; Zhang, X.; Liu, Y.; Zhu, S.; et al. FERONIA is involved in phototropin 1 -mediated blue light phototropic growth in Arabidopsis. JIPB 2022, 64, 1901–1915. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Li, R.; Cui, Y.; Chen, W.; Li, B.; Zhang, X.; Bu, Y.; Cao, Y.; Xing, J.; Jewaria, P.K.; et al. The RALF1-FERONIA interaction modulates endocytosis to mediate control of root growth in Arabidopsis. Development 2020, 147. [Google Scholar] [CrossRef] [PubMed]
- Planas-Riverola, A.; Gupta, A.; Betegón-Putze, I.; Bosch, N.; Ibañes, M.; Caño-Delgado, A.I. Brassinosteroid signaling in plant development and adaptation to stress. Development 2019, 146. [Google Scholar] [CrossRef]
- Kim, T.-W.; Wang, Z.-Y. Brassinosteroid Signal Transduction from Receptor Kinases to Transcription Factors. Annu. Rev. Plant Biol. 2010, 61, 681–704. [Google Scholar] [CrossRef]
- Chaudhary, A.; Hsiao, Y.-C.; Yeh, F.-L.J.; Župunski, M.; Zhang, H.; Aizezi, Y.; Malkovskiy, A.; Grossmann, G.; Wu, H.-M.; Cheung, A.Y.; et al. FERONIA signaling maintains cell wall integrity during brassinosteroid-induced cell expansion in Arabidopsis. Mol. Plant 2025, 18, 603–618. [Google Scholar] [CrossRef]
- Sun, Y.; Han, Z.; Tang, J.; Hu, Z.; Chai, C.; Zhou, B.; Chai, J. Structure reveals that BAK1 as a co-receptor recognizes the BRI1-bound brassinolide. Cell Res. 2013, 23, 1326–1329. [Google Scholar] [CrossRef]
- Park, C.H.; Kim, T.-W.; Son, S.-H.; Hwang, J.-Y.; Lee, S.C.; Chang, S.C.; Kim, S.-H.; Kim, S.W.; Kim, S.-K. Brassinosteroids control AtEXPA5 gene expression in Arabidopsis thaliana. Phytochemistry 2010, 71, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Bergonci, T.; Silva-Filho, M.C.; Moura, D.S. Antagonistic relationship between AtRALF1and brassinosteroid regulates cellexpansion-related genes. Plant Signal. Behav. 2014, 9, e976146–e976146. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Structure and growth of plant cell walls. Nat. Rev. Mol. Cell Biol. 2023, 25, 340–358. [Google Scholar] [CrossRef]
- Lin, W.; Tang, W.; Pan, X.; Huang, A.; Gao, X.; Anderson, C.T.; Yang, Z. Arabidopsis pavement cell morphogenesis requires FERONIA binding to pectin for activation of ROP GTPase signaling. Curr. Biol. 2022, 32, 497–507.e4. [Google Scholar] [CrossRef] [PubMed]
- Biermann, D.; von Arx, M.; Munzert-Eberlein, K.S.; Xhelilaj, K.; Séré, D.; Stegmann, M.; Vert, G.; Wolf, S.; Engelsdorf, T.; Zipfel, C.; et al. A RALF-brassinosteroid signaling circuit regulates Arabidopsis hypocotyl cell shape. Curr. Biol. 2025, 35, 5002–5017.e5. [Google Scholar] [CrossRef] [PubMed]
- Colcombet, J.; Hirt, H. Arabidopsis MAPKs: a complex signalling network involved in multiple biological processes. Biochem. J. 2008, 413, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, X.; Qiang, X.; Li, X.; Li, X.; Zhu, S.; et al. EBP1 nuclear accumulation negatively feeds back on FERONIA-mediated RALF1 signaling. PLoS Biol. 2018, 16, e2006340. [Google Scholar] [CrossRef]
- Sonenberg, N.; Hinnebusch, A.G. New Modes of Translational Control in Development, Behavior, and Disease. Mol. Cell 2007, 28, 721–729. [Google Scholar] [CrossRef]
- Zhu, S.; Li, Y.; Wu, Y.; Shen, Y.; Wang, Y.; Yan, Y.; Chen, W.; Fu, Q.; Wang, Y.; Yu, X.; et al. The FERONIA-YUELAO module participates in translational control by modulating the abundance of tRNA fragments in Arabidopsis. Dev. Cell 2023, 58, 2930–2946.e9. [Google Scholar] [CrossRef]
- Lalande, S.; Merret, R.; Salinas-Giegé, T.; Drouard, L. Arabidopsis tRNA-derived fragments as potential modulators of translation. RNA Biol. 2020, 17, 1137–1148. [Google Scholar] [CrossRef]
- Stegmann, M. EBP1: A crucial growth regulator downstream of receptor kinases across kingdoms. PLOS Biol. 2018, 16, e3000056. [Google Scholar] [CrossRef]
- Staiger, D.; Zecca, L.; Kirk, D.A.W.; Apel, K.; Eckstein, L. The circadian clock regulated RNA-binding protein AtGRP7 autoregulates its expression by influencing alternative splicing of its own pre-mRNA. Plant J. 2003, 33, 361–371. [Google Scholar] [CrossRef]
- Steffen, A.; Elgner, M.; Staiger, D. Regulation of Flowering Time by the RNA-Binding Proteins AtGRP7 and AtGRP8. Plant Cell Physiol. 2019, 60, 2040–2050. [Google Scholar] [CrossRef]
- Clark, G.B.; Morgan, R.O.; Fernandez, M.-P.; Salmi, M.L.; Roux, S.J. Breakthroughs spotlighting roles for extracellular nucleotides and apyrases in stress responses and growth and development. Plant Sci. 2014, 225, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Guérin, A.; Herger, A.; Hou, X.; Schaufelberger, M.; Roulard, R.; Diet, A.; Roffler, S.; Lefebvre, V.; Wicker, T.; et al. Growth-inhibiting effects of the unconventional plant APYRASE 7 of Arabidopsis thaliana influences the LRX/RALF/FER growth regulatory module. PLOS Genet. 2024, 20, e1011087. [Google Scholar] [CrossRef] [PubMed]
- Rößling, A.K.; Dünser, K.; Liu, C.; Lauw, S.; Rodriguez-Franco, M.; Kalmbach, L.; et al. Pectin methylesterase activity is required for RALF1 peptide signalling output [Internet]. 2024 [cited 2024 Aug 11]. Available from: https://elifesciences.org/reviewed-preprints/96943v1.
- Manmohit Kalia, P.K. Pectin Methylesterases: A Review. J Bioprocess Biotech [Internet]. 2015 [cited 2024 Aug 11];05(05). Available from: https://www.omicsonline.org/open-access/pectin-methylesterases-a-review-2155-9821-1000227.php?aid=52733.
- Xu, F.; Chen, J.; Li, Y.; Ouyang, S.; Yu, M.; Wang, Y.; Fang, X.; He, K.; Yu, F. The soil emergence-related transcription factor PIF3 controls root penetration by interacting with the receptor kinase FER. Dev. Cell 2024, 59, 434–447.e8. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Shi, H.; Xue, C.; Wei, N.; Guo, H.; Deng, X.W. Ethylene-orchestrated circuitry coordinates a seedling’s response to soil cover and etiolated growth. Proc. Natl. Acad. Sci. 2014, 111, 3913–3920. [Google Scholar] [CrossRef]
- Fang, X.; Liu, B.; Shao, Q.; Huang, X.; Li, J.; Luan, S.; He, K. AtPiezo Plays an Important Role in Root Cap Mechanotransduction. Int. J. Mol. Sci. 2021, 22, 467. [Google Scholar] [CrossRef]
- Shen, Y.; Xie, Q.; Wang, T.; Wang, X.; Xu, F.; Yan, Z.; Li, X.; Ouyang, S.; Chen, J.; Wang, Y.; et al. RALF33–FERONIA signaling orchestrates postwounding root-tip regeneration via TPR4–ERF115 dynamics. Plant Cell 2025, 37. [Google Scholar] [CrossRef]
- Canher, B.; Lanssens, F.; Zhang, A.; Bisht, A.; Mazumdar, S.; Heyman, J.; Wolf, S.; Melnyk, C.W.; De Veylder, L. The regeneration factors ERF114 and ERF115 regulate auxin-mediated lateral root development in response to mechanical cues. Mol. Plant 2022, 15, 1543–1557. [Google Scholar] [CrossRef]
- Heyman, J.; Cools, T.; Canher, B.; Shavialenka, S.; Traas, J.; Vercauteren, I.; Van den Daele, H.; Persiau, G.; De Jaeger, G.; Sugimoto, K.; et al. The heterodimeric transcription factor complex ERF115–PAT1 grants regeneration competence. Nat. Plants 2016, 2, 16165. [Google Scholar] [CrossRef]
- Murphy, E.; Vu, L.D.; Broeck, L.V.D.; Lin, Z.; Ramakrishna, P.; van de Cotte, B.; Gaudinier, A.; Goh, T.; Slane, D.; Beeckman, T.; et al. RALFL34 regulates formative cell divisions in Arabidopsis pericycle during lateral root initiation. J. Exp. Bot. 2016, 67, 4863–4875. [Google Scholar] [CrossRef]
- Grierson, C.; Nielsen, E.; Ketelaarc, T.; Schiefelbein, J. Root Hairs. The Arabidopsis Book 2014, 12, e0172. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Lilley, C.J.; Urwin, P.E. Identification of Genes Involved in the Response of Arabidopsis to Simultaneous Biotic and Abiotic Stresses. Plant Physiol. 2013, 162, 2028–2041. [Google Scholar] [CrossRef]
- Hocq, L.; Pelloux, J.; Lefebvre, V. Connecting Homogalacturonan-Type Pectin Remodeling to Acid Growth. Trends Plant Sci. 2017, 22, 20–29. [Google Scholar] [CrossRef]
- Morcillo, R.J.L.; Leal-López, J.; Férez-Gómez, A.; López-Serrano, L.; Baroja-Fernández, E.; Gámez-Arcas, S.; Tortosa, G.; E López, L.; Estevez, J.M.; Doblas, V.G.; et al. RAPID ALKALINIZATION FACTOR 22 is a key modulator of the root hair growth responses to fungal ethylene emissions in Arabidopsis. Plant Physiol. 2024, 196, 2890–2904. [Google Scholar] [CrossRef]
- Molendijk, A.J.; Bischoff, F.; Rajendrakumar, C.S.V.; Friml, J.; Braun, M.; Gilroy, S.; Palme, K. Arabidopsis thaliana Rop GTPases are localized to tips of root hairs and control polar growth. EMBO J. 2001, 20, 2779–2788. [Google Scholar] [CrossRef]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.F.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.G.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef]
- Duan, Q.; Kita, D.; Li, C.; Cheung, A.Y.; Wu, H.-M. FERONIA receptor-like kinase regulates RHO GTPase signaling of root hair development. Proc. Natl. Acad. Sci. 2010, 107, 17821–17826. [Google Scholar] [CrossRef]
- Datta, S.; Prescott, H.; Dolan, L. Intensity of a pulse of RSL4 transcription factor synthesis determines Arabidopsis root hair cell size. Nat. Plants 2015, 1, 15138. [Google Scholar] [CrossRef]
- Yi, K.; Menand, B.; Bell, E.; Dolan, L. A basic helix-loop-helix transcription factor controls cell growth and size in root hairs. Nat. Genet. 2010, 42, 264–267. [Google Scholar] [CrossRef]
- Masucci, J.D.; Schiefelbein, J.W. Hormones act downstream of TTG and GL2 to promote root hair outgrowth during epidermis development in the Arabidopsis root. Plant Cell. 1996, 8, 1505–1517. [Google Scholar]
- Huang, G.; Li, E.; Ge, F.; Li, S.; Wang, Q.; Zhang, C.; Zhang, Y. Arabidopsis RopGEF4 and RopGEF10 are important for FERONIA-mediated developmental but not environmental regulation of root hair growth. New Phytol. 2013, 200, 1089–1101. [Google Scholar] [CrossRef]
- Franck, C.M.; Westermann, J.; Bürssner, S.; Lentz, R.; Lituiev, D.S.; Boisson-Dernier, A. The Protein Phosphatases ATUNIS1 and ATUNIS2 Regulate Cell Wall Integrity in Tip-Growing Cells. Plant Cell 2018, 30, 1906–1923. [Google Scholar] [CrossRef]
- Boisson-Dernier, A.; Franck, C.M.; Lituiev, D.S.; Grossniklaus, U. Receptor-like cytoplasmic kinase MARIS functions downstream of Cr RLK1L-dependent signaling during tip growth. Proc Natl Acad Sci USA 2015, 112, 12211–12216. [Google Scholar] [CrossRef]
- Donaldson, J.; Poratshliom, N.; Cohen, L. Clathrin-independent endocytosis: A unique platform for cell signaling and PM remodeling. Cell. Signal. 2009, 21, 1–6. [Google Scholar] [CrossRef]
- Borner, G.H.; Sherrier, D.J.; Weimar, T.; Michaelson, L.V.; Hawkins, N.D.; MacAskill, A.; Napier, J.A.; Beale, M.H.; Lilley, K.S.; Dupree, P. Analysis of Detergent-Resistant Membranes in Arabidopsis. Evidence for Plasma Membrane Lipid Rafts. Plant Physiol. 2005, 137, 104–116. [Google Scholar] [CrossRef]
- Chen, W.; Zhou, H.; Xu, F.; Yu, M.; Coego, A.; Rodriguez, L.; Lu, Y.; Xie, Q.; Fu, Q.; Chen, J.; et al. CAR modulates plasma membrane nano-organization and immune signaling downstream of RALF1-FERONIA signaling pathway. New Phytol. 2023, 237, 2148–2162. [Google Scholar] [CrossRef]
- Emenecker, R.J.; Holehouse, A.S.; Strader, L.C. Biological Phase Separation and Biomolecular Condensates in Plants. Annu. Rev. Plant Biol. 2021, 72, 17–46. [Google Scholar] [CrossRef]
- Diaz, M.; Sanchez-Barrena, M.J.; Gonzalez-Rubio, J.M.; Rodriguez, L.; Fernandez, D.; Antoni, R.; Yunta, C.; Belda-Palazon, B.; Gonzalez-Guzman, M.; Peirats-Llobet, M.; et al. Calcium-dependent oligomerization of CAR proteins at cell membrane modulates ABA signaling. Proc. Natl. Acad. Sci. 2015, 113, 201512779–405. [Google Scholar] [CrossRef]
- Kay, J.G.; Fairn, G.D. Distribution, dynamics and functional roles of phosphatidylserine within the cell. Cell Commun. Signal. 2019, 17, 1–8. [Google Scholar] [CrossRef]
- Platre, M.P.; Bayle, V.; Armengot, L.; Bareille, J.; Marquès-Bueno, M.d.M.; Creff, A.; Maneta-Peyret, L.; Fiche, J.-B.; Nollmann, M.; Miège, C.; et al. Developmental control of plant Rho GTPase nano-organization by the lipid phosphatidylserine. Science 2019, 364, 57–62. [Google Scholar] [CrossRef]
- Smokvarska, M.; Francis, C.; Platre, M.P.; Fiche, J.-B.; Alcon, C.; Dumont, X.; Nacry, P.; Bayle, V.; Nollmann, M.; Maurel, C.; et al. A Plasma Membrane Nanodomain Ensures Signal Specificity during Osmotic Signaling in Plants. Curr. Biol. 2020, 30, 4654–4664.e4. [Google Scholar] [CrossRef]
- Leung, J.; Giraudat, J. ABSCISIC ACID SIGNAL TRANSDUCTION. Annu. Rev. Plant Biol. 1998, 49, 199–222. [Google Scholar] [CrossRef]
- Raghavendra, A.S.; Gonugunta, V.K.; Christmann, A.; Grill, E. ABA perception and signalling. Trends Plant Sci. 2010, 15, 395–401. [Google Scholar] [CrossRef]
- Chen, J.; Yu, F.; Liu, Y.; Du, C.; Li, X.; Zhu, S.; Wang, X.; Lan, W.; Rodriguez, P.L.; Liu, X.; et al. FERONIA interacts with ABI2-type phosphatases to facilitate signaling cross-talk between abscisic acid and RALF peptide in Arabidopsis. Proc. Natl. Acad. Sci. 2016, 113, E5519–E5527. [Google Scholar] [CrossRef]
- Yu, F.; Qian, L.; Nibau, C.; Duan, Q.; Kita, D.; Levasseur, K.; Li, X.; Lu, C.; Li, H.; Hou, C.; et al. FERONIA receptor kinase pathway suppresses abscisic acid signaling in Arabidopsis by activating ABI2 phosphatase. Proc. Natl. Acad. Sci. 2012, 109, 14693–14698. [Google Scholar] [CrossRef]
- Fu, Q.; Li, H.; Wang, B.; Chen, W.; Wu, D.; Gao, C.; Yu, F.; Shen, W. The RALF1 peptide-FERONIA complex phosphorylates the endosomal sorting protein FREE1 to attenuate abscisic acid signaling. Plant Physiol. 2024, 197. [Google Scholar] [CrossRef]
- Gao, C.; Luo, M.; Zhao, Q.; Yang, R.; Cui, Y.; Zeng, Y.; Xia, J.; Jiang, L. A Unique Plant ESCRT Component, FREE1, Regulates Multivesicular Body Protein Sorting and Plant Growth. Curr. Biol. 2014, 24, 2556–2563. [Google Scholar] [CrossRef]
- Li, H.; Li, Y.; Zhao, Q.; Li, T.; Wei, J.; Li, B.; et al. The plant ESCRT component FREE1 shuttles to the nucleus to attenuate abscisic acid signalling. Nat Plants. 2019, 5, 512–524. [Google Scholar] [CrossRef]
- Mohanasundaram, B.; Pandey, S. Moving beyond the arabidopsis-centric view of G-protein signaling in plants. Trends Plant Sci. 2023, 28, 1406–1421. [Google Scholar] [CrossRef]
- Yu, Y.; Chakravorty, D.; Assmann, S.M. The G Protein β-Subunit, AGB1, Interacts with FERONIA in RALF1-Regulated Stomatal Movement. Plant Physiol. 2018, 176, 2426–2440. [Google Scholar] [CrossRef]
- Chakravorty, D.; Yu, Y.; Assmann, S.M. A kinase-dead version of FERONIA receptor-like kinase has dose-dependent impacts on rosette morphology and RALF 1-mediated stomatal movements. FEBS Letters 2018, 592, 3429–3437. [Google Scholar] [CrossRef]
- Zhao, C.; Jiang, W.; Zayed, O.; Liu, X.; Tang, K.; Nie, W.; Li, Y.; Xie, S.; Li, Y.; Long, T.; et al. The LRXs-RALFs-FER module controls plant growth and salt stress responses by modulating multiple plant hormones. Natl. Sci. Rev. 2020, 8, nwaa149. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of Reactive Oxygen Species and Antioxidant Defense in Plants under Salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef]
- Liu, X.; Wang, L.; Liu, L.; Li, Y.; Ogden, M.; Somssich, M.; Liu, Y.; Zhang, Y.; Ran, M.; Persson, S.; et al. FERONIA adjusts CC1 phosphorylation to control microtubule array behavior in response to salt stress. Sci. Adv. 2024, 10, eadq8717. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, W.; Li, Y.; Nie, H.; Cui, L.; Li, R.; Tan, L.; Peng, L.; Li, C.; Luo, J.; et al. FERONIA coordinates plant growth and salt tolerance via the phosphorylation of phyB. Nat. Plants 2023, 9, 645–660. [Google Scholar] [CrossRef]
- Gigli-Bisceglia, N.; van Zelm, E.; Huo, W.; Lamers, J.; Testerink, C. Arabidopsis root responses to salinity depend on pectin modification and cell wall sensing. Development 2022, 149. [Google Scholar] [CrossRef]
- Sato, T.; Maekawa, S.; Yasuda, S.; Yamaguchi, J. Carbon and nitrogen metabolism regulated by the ubiquitin-proteasome system. Plant Signal. Behav. 2011, 6, 1465–1468. [Google Scholar] [CrossRef]
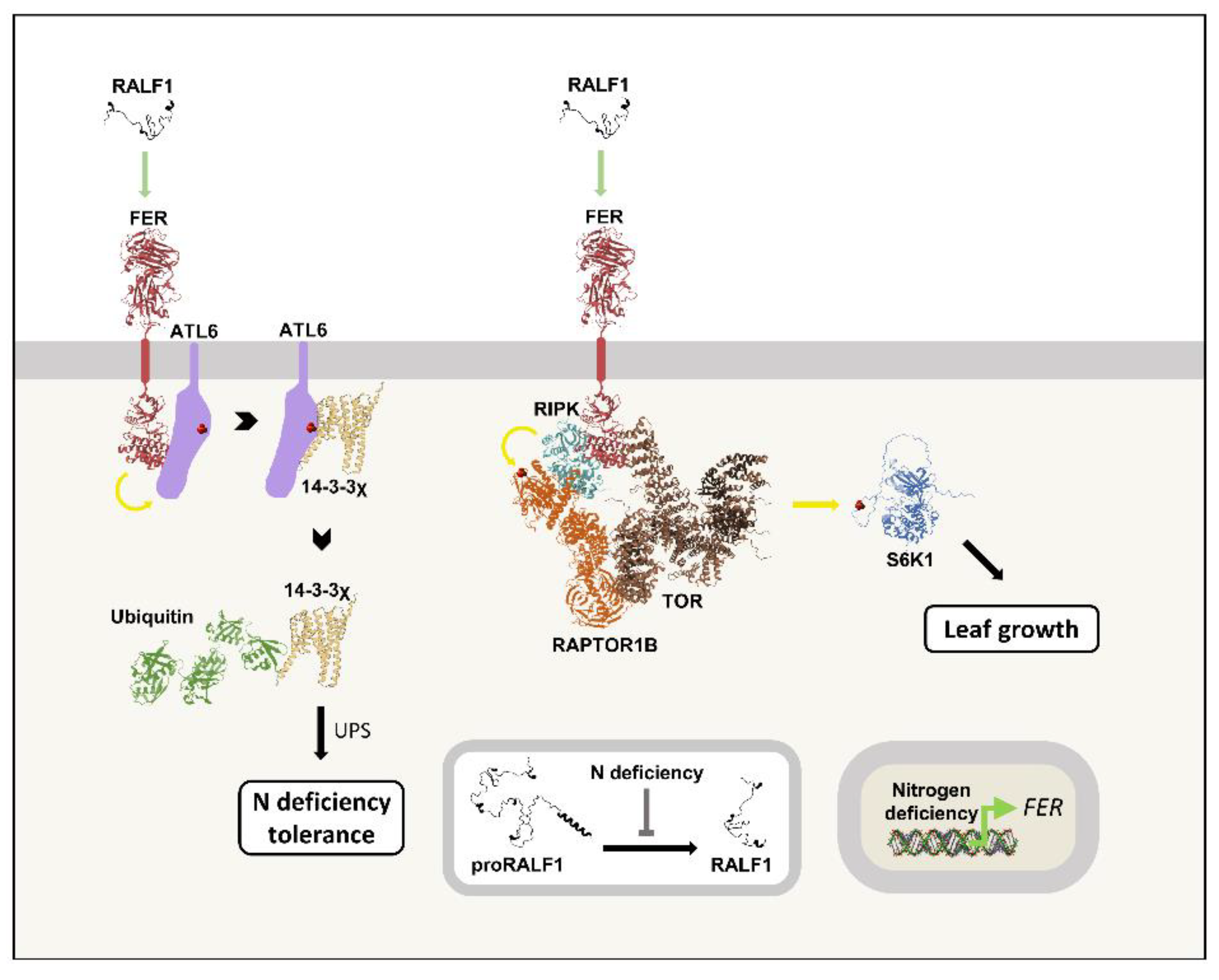
- Xu, G.; Chen, W.; Song, L.; Chen, Q.; Zhang, H.; Liao, H.; Zhao, G.; Lin, F.; Zhou, H.; Yu, F. FERONIA phosphorylates E3 ubiquitin ligase ATL6 to modulate the stability of 14-3-3 proteins in response to the carbon/nitrogen ratio. J. Exp. Bot. 2019, 70, 6375–6388. [Google Scholar] [CrossRef]
- Burkart, G.M.; Brandizzi, F. A Tour of TOR Complex Signaling in Plants. Trends Biochem. Sci. 2021, 46, 417–428. [Google Scholar] [CrossRef]
- Song, L.; Xu, G.; Li, T.; Zhou, H.; Lin, Q.; Chen, J.; Wang, L.; Wu, D.; Li, X.; Wang, L.; et al. The RALF1-FERONIA complex interacts with and activates TOR signaling in response to low nutrients. Mol. Plant 2022, 15, 1120–1136. [Google Scholar] [CrossRef]
- Wang, P.; Clark, N.M.; Nolan, T.M.; Song, G.; Whitham, O.G.; Liao, C.-Y.; Montes-Serey, C.; Bassham, D.C.; Walley, J.W.; Yin, Y.; et al. FERONIA functions through Target of Rapamycin (TOR) to negatively regulate autophagy. Front. Plant Sci. 2022, 13, 961096. [Google Scholar] [CrossRef]
- Hou, S.; Tsuda, K. Salicylic acid and jasmonic acid crosstalk in plant immunity. Kanyuka K, Hammond-Kosack K, editors. Essays in Biochemistry 2022, 66, 647–656. [Google Scholar]
- Guo, H.; Nolan, T.M.; Song, G.; Schnable, P.S.; Walley, J.W.; Yin, Y.; Guo, H.; Nolan, T.M.; Song, G.; Liu, S.; et al. FERONIA Receptor Kinase Contributes to Plant Immunity by Suppressing Jasmonic Acid Signaling in Arabidopsis thaliana. Curr. Biol. 2018, 28, 3316–3324.e6. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, J.-M. Plant Immunity Triggered by Microbial Molecular Signatures. Mol. Plant 2010, 3, 783–793. [Google Scholar] [CrossRef]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef]
- Kadota, Y.; Shirasu, K.; Zipfel, C. Regulation of the NADPH Oxidase RBOHD During Plant Immunity. Plant Cell Physiol. 2015, 56, 1472–1480. [Google Scholar] [CrossRef]
- Gronnier, J.; Franck, C.M.; Stegmann, M.; A DeFalco, T.; Abarca, A.; von Arx, M.; Dünser, K.; Lin, W.; Yang, Z.; Kleine-Vehn, J.; et al. Regulation of immune receptor kinase plasma membrane nanoscale organization by a plant peptide hormone and its receptors. eLife 2022, 11. [Google Scholar] [CrossRef]
- Boisson-Dernier, A.; Roy, S.; Kritsas, K.; Grobei, M.A.; Jaciubek, M.; Schroeder, J.I.; Grossniklaus, U. Disruption of the pollen-expressed FERONIA homologs ANXUR1 and ANXUR2 triggers pollen tube discharge. Development 2009, 136, 3279–3288. [Google Scholar] [CrossRef] [PubMed]
- Mang, H.; Feng, B.; Hu, Z.; Boisson-Dernier, A.; Franck, C.M.; Meng, X.; Huang, Y.; Zhou, J.; Xu, G.; Wang, T.; et al. Differential Regulation of Two-Tiered Plant Immunity and Sexual Reproduction by ANXUR Receptor-Like Kinases. Plant Cell 2017, 29, 3140–3156. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Bourdais, G.; Pan, H.; Robatzek, S.; Tang, D. Arabidopsis glycosylphosphatidylinositol-anchored protein LLG1 associates with and modulates FLS2 to regulate innate immunity. Proc. Natl. Acad. Sci. 2017, 114, 5749–5754. [Google Scholar] [CrossRef]
- Tang, J.; Wu, D.; Li, X.; Wang, L.; Xu, L.; Zhang, Y.; Xu, F.; Liu, H.; Xie, Q.; Dai, S.; et al. Plant immunity suppression via PHR1-RALF-FERONIA shapes the root microbiome to alleviate phosphate starvation. EMBO J. 2022, 41, e109102. [Google Scholar] [CrossRef]
- García-Gómez, P.; Bahaji, A.; Gámez-Arcas, S.; Muñoz, F.J.; Sánchez-López, Á.M.; Almagro, G.; Baroja-Fernández, E.; Ameztoy, K.; De Diego, N.; Ugena, L.; et al. Volatiles from the fungal phytopathogen Penicillium aurantiogriseum modulate root metabolism and architecture through proteome resetting. Plant, Cell Environ. 2020, 43, 2551–2570. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Yuan, B.; Chen, X.; Zhao, H.; Ali, Q.; Zheng, M.; Tan, Z.; Yao, H.; Zheng, S.; et al. Fusarium graminearum rapid alkalinization factor peptide negatively regulates plant immunity and cell growth via the FERONIA receptor kinase. Plant Biotechnol. J. 2024, 22, 1800–1811. [Google Scholar] [CrossRef]
- Masachis, S.; Segorbe, D.; Turrà, D.; Leon-Ruiz, M.; Fürst, U.; El Ghalid, M.; et al. A fungal pathogen secretes plant alkalinizing peptides to increase infection. Nat Microbiol. 2016, 1, 16043. [Google Scholar] [CrossRef]
- Liao, Y.; Wen, X.; Zheng, J.; Li, X.; Deng, X.; Tan, X.; Liang, X.; Yi, X.; Liao, H. RALF-like peptide improves the colonization of endophytic Colletotrichum tofieldiae through interacting with plant receptor-like kinase. Plant Pathol. 2023, 72, 1649–1661. [Google Scholar] [CrossRef]
- Pin, P.A.; Nilsson, O. The multifaceted roles of FLOWERING LOCUS T in plant development. Plant, Cell Environ. 2012, 35, 1742–1755. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Le, C.; Wang, Y.; Li, Z.; Jiang, D.; Wang, Y.; He, Y. Arabidopsis FLC clade members form flowering-repressor complexes coordinating responses to endogenous and environmental cues. Nat. Commun. 2013, 4, 1947. [Google Scholar] [CrossRef]
- Michaels, S.D.; Himelblau, E.; Kim, S.Y.; Schomburg, F.M.; Amasino, R.M. Integration of Flowering Signals in Winter-Annual Arabidopsis. Plant Physiol. 2005, 137, 149–156. [Google Scholar] [CrossRef]
- Wang, L.; Yang, T.; Lin, Q.; Wang, B.; Li, X.; Luan, S.; Yu, F. Receptor kinase FERONIA regulates flowering time in Arabidopsis. BMC Plant Biol. 2020, 20, 1–16. [Google Scholar] [CrossRef]
- Johnson, M.A.; Harper, J.F.; Palanivelu, R. A Fruitful Journey: Pollen Tube Navigation from Germination to Fertilization. Annu. Rev. Plant Biol. 2019, 70, 809–837. [Google Scholar] [CrossRef]
- Törnroth-Horsefield, S.; Wang, Y.; Hedfalk, K.; Johanson, U.; Karlsson, M.; Tajkhorshid, E.; et al. Structural mechanism of plant aquaporin gating. Nature 2006, 439, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Chebli, Y.; Kaneda, M.; Zerzour, R.; Geitmann, A. The Cell Wall of the Arabidopsis Pollen Tube—Spatial Distribution, Recycling, and Network Formation of Polysaccharides. Plant Physiol. 2012, 160, 1940–1955. [Google Scholar] [CrossRef]
- Wudick, M.M.; Feijó, J.A. At the Intersection: Merging Ca 2+ and ROS Signaling Pathways in Pollen. Mol. Plant 2014, 7, 1595–1597. [Google Scholar] [CrossRef] [PubMed]
- Seitz, P.L.; Qu, L.-J.; Dresselhaus, T.; Zhou, L.-Z. Spatial organization and trafficking dynamics of ANX/BUPS-RALF-LLG signaling complexes during pollen tube growth. Plant Reprod. 2025, 38, 1–10. [Google Scholar] [CrossRef]
- Boisson-Dernier, A.; Lituiev, D.S.; Nestorova, A.; Franck, C.M.; Thirugnanarajah, S.; Grossniklaus, U. ANXUR Receptor-Like Kinases Coordinate Cell Wall Integrity with Growth at the Pollen Tube Tip Via NADPH Oxidases. Nasrallah JB, editor. PLoS Biol. 2013, 11, e1001719. [Google Scholar] [CrossRef]
- Zhu, L.; Chu, L.; Liang, Y.; Zhang, X.; Chen, L.; Ye, D. The Arabidopsis CrRLK1L protein kinases BUPS1 and BUPS2 are required for normal growth of pollen tubes in the pistil. Plant J. 2018, 95, 474–486. [Google Scholar] [CrossRef]
- Sede, A.R.; Borassi, C.; Wengier, D.L.; Mecchia, M.A.; Estevez, J.M.; Muschietti, J.P. Arabidopsis pollen extensins LRX are required for cell wall integrity during pollen tube growth. FEBS Lett. 2018, 592, 233–243. [Google Scholar] [CrossRef]
- Wang, X.; Wang, K.; Yin, G.; Liu, X.; Liu, M.; Cao, N.; Duan, Y.; Gao, H.; Wang, W.; Ge, W.; et al. Pollen-Expressed Leucine-Rich Repeat Extensins Are Essential for Pollen Germination and Growth. Plant Physiol. 2017, 176, 1993–2006. [Google Scholar] [CrossRef]
- Fabrice, T.N.; Vogler, H.; Draeger, C.; Munglani, G.; Gupta, S.; Herger, A.G.; Knox, P.; Grossniklaus, U.; Ringli, C. LRX Proteins Play a Crucial Role in Pollen Grain and Pollen Tube Cell Wall Development. Plant Physiol. 2017, 176, 1981–1992. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Lu, J.; Zhang, Y.; Guo, J.; Lin, W.; Van Norman, J.M.; Qin, Y.; Zhu, X.; Yang, Z. Membrane receptor-mediated mechano-transduction maintains cell integrity during pollen tube growth within the pistil. Dev. Cell 2021, 56, 1030–1042.e6. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Lin, W.; Zhou, X.; Guo, J.; Dang, X.; Li, B.; Lin, D.; Yang, Z. Mechano-transduction via the pectin-FERONIA complex activates ROP6 GTPase signaling in Arabidopsis pavement cell morphogenesis. Curr. Biol. 2022, 32, 508–517.e3. [Google Scholar] [CrossRef]
- Ngo, Q.A.; Vogler, H.; Lituiev, D.S.; Nestorova, A.; Grossniklaus, U. A Calcium Dialog Mediated by the FERONIA Signal Transduction Pathway Controls Plant Sperm Delivery. Dev. Cell 2014, 29, 491–500. [Google Scholar] [CrossRef]
- Duan, Q.; Kita, D.; Johnson, E.A.; Aggarwal, M.; Gates, L.; Wu, H.-M.; Cheung, A.Y. Reactive oxygen species mediate pollen tube rupture to release sperm for fertilization in Arabidopsis. Nat. Commun. 2014, 5, 3129. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Trigo, S.; Blanco-Touriñán, N.; A DeFalco, T.; Wells, E.S.; E Gray, J.; Zipfel, C.; Smith, L.M. CrRLK1L receptor-like kinases HERK1 and ANJEA are female determinants of pollen tube reception. Embo Rep. 2019, 21, e48466. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Liu, M.-C.J.; Kita, D.; Jordan, S.S.; Yeh, F.-L.J.; Yvon, R.; Carpenter, H.; Federico, A.N.; Garcia-Valencia, L.E.; Eyles, S.J.; et al. FERONIA controls pectin- and nitric oxide-mediated male–female interaction. Nature 2020, 579, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kurten, E.L.; Monshausen, G.; Hummel, G.M.; Gilroy, S.; Baldwin, I.T. NaRALF, a peptide signal essential for the regulation of root hair tip apoplastic pH in Nicotiana attenuata, is required for root hair development and plant growth in native soils. Plant J. 2007, 52, 877–890. [Google Scholar] [CrossRef]
- Kwon, O.; Moon, H.; Jeong, A.; Yeom, G.; Park, C. Rice small secreted peptide, OsRALF26, recognized by FERONIA-like receptor 1 induces immunity in rice and Arabidopsis. Plant J. 2024, 118, 1528–1549. [Google Scholar] [CrossRef]
- Jiang, W.; Li, C.; Li, L.; Li, Y.; Wang, Z.; Yu, F.; Yi, F.; Zhang, J.; Zhu, J.-K.; Zhang, H.; et al. Genome-Wide Analysis of CqCrRLK1L and CqRALF Gene Families in Chenopodium quinoa and Their Roles in Salt Stress Response. Front. Plant Sci. 2022, 13, 918594. [Google Scholar] [CrossRef]
- Wieghaus, A.; Prüfer, D.; Gronover, C.S. Loss of function mutation of the Rapid Alkalinization Factor (RALF1)-like peptide in the dandelion Taraxacum koksaghyz entails a high-biomass taproot phenotype. PLOS ONE 2019, 14, e0217454. [Google Scholar] [CrossRef]
- Mingossi, F.B.; Matos, J.L.; Rizzato, A.P.; Medeiros, A.H.; Falco, M.C.; Silva-Filho, M.C.; Moura, D.S. SacRALF1, a peptide signal from the grass sugarcane (Saccharum spp.), is potentially involved in the regulation of tissue expansion. Plant Mol. Biol. 2010, 73, 271–281. [Google Scholar] [CrossRef]
- Fan, Y.; Bai, J.; Wu, S.; Zhang, M.; Li, J.; Lin, R.; et al. The RALF2-FERONIA-MYB63 module orchestrates growth and defense in tomato roots. New Phytologist 2024, 243, 1123–1136. [Google Scholar] [CrossRef]
- Ginanjar, E.F.; Teh, O.; Fujita, T. Characterisation of rapid alkalinisation factors in Physcomitrium patens reveals functional conservation in tip growth. New Phytol. 2021, 233, 2442–2457. [Google Scholar] [CrossRef] [PubMed]
- Haruta, M.; Constabel, C.P. Rapid Alkalinization Factors in Poplar Cell Cultures. Peptide Isolation, cDNA Cloning, and Differential Expression in Leaves and Methyl Jasmonate-Treated Cells. Plant Physiol. 2003, 131, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Gidrol, X.; Chrestin, H.; Mounoury, G.; D'Auzac, J. Early Activation by Ethylene of the Tonoplast H+-Pumping ATPase in the Latex from Hevea brasiliensis. Plant Physiol. 1988, 86, 899–903. [Google Scholar] [CrossRef]
- Sui, J.; Xiao, X.; Yang, J.; Fan, Y.; Zhu, S.; Zhu, J.; Zhou, B.; Yu, F.; Tang, C. The rubber tree RALF peptide hormone and its receptor protein kinase FER implicates in rubber production. Plant Sci. 2022, 326, 111510. [Google Scholar] [CrossRef]
- Covey, P.A.; Subbaiah, C.C.; Parsons, R.L.; Pearce, G.; Lay, F.T.; Anderson, M.A.; Ryan, C.A.; Bedinger, P.A. A Pollen-Specific RALF from Tomato That Regulates Pollen Tube Elongation. Plant Physiol. 2010, 153, 703–715. [Google Scholar] [CrossRef]
- Kou, X.; Sun, J.; Wang, P.; Wang, D.; Cao, P.; Lin, J.; Chang, Y.; Zhang, S.; Wu, J. PbrRALF2-elicited reactive oxygen species signaling is mediated by the PbrCrRLK1L13-PbrMPK18 module in pear pollen tubes. Hortic. Res. 2021, 8, 222. [Google Scholar] [CrossRef]
- Zhou, L.-Z.; Wang, L.; Chen, X.; Ge, Z.; Mergner, J.; Li, X.; Küster, B.; Längst, G.; Qu, L.-J.; Dresselhaus, T. The RALF signaling pathway regulates cell wall integrity during pollen tube growth in maize. Plant Cell 2023, 36, 1673–1696. [Google Scholar] [CrossRef]
- Kim, E.; Kim, J.; Hong, W.; Kim, E.Y.; Kim, M.; Lee, S.K.; Min, C.W.; Kim, S.T.; Park, S.K.; Jung, K.; et al. Rice pollen-specific OsRALF17 and OsRALF19 are essential for pollen tube growth. J. Integr. Plant Biol. 2023, 65, 2218–2236. [Google Scholar] [CrossRef]
- Kim, J.-H.; Son, Y.-J.; Kim, E.-J.; Jung, K.-H.; Kim, Y.-J. Pollen tube-expressed RUPO forms a complex with OsMTD2 and OsRALF17 and OsRALF19 peptides in rice (Oryza sativa). J. Plant Physiol. 2025, 305, 154421. [Google Scholar] [CrossRef]
- Liu, L.; Zheng, C.; Kuang, B.; Wei, L.; Yan, L.; Wang, T. Receptor-Like Kinase RUPO Interacts with Potassium Transporters to Regulate Pollen Tube Growth and Integrity in Rice. PLOS Genet. 2016, 12, e1006085. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Zhou, J.; Gu, Y.; Zeng, Y.; Lu, K.; Peng, S.; Tan, X. Genome-wide identification of the RALF gene family in Camellia oleifera Abel. and the potentiality of CoRALF50 in pollen tube growth. Plant Sci. 2025, 359, 112603. [Google Scholar] [CrossRef]
- Chevalier, E.; Loubert-Hudon, A.; Matton, D.P. ScRALF3, a secreted RALF-like peptide involved in cell–cell communication between the sporophyte and the female gametophyte in a solanaceous species. Plant J. 2012, 73, 1019–1033. [Google Scholar] [CrossRef]
- Germain, H.; Matton, D.P.; Chevalier, É.; Caron, S. Characterization of five RALF-like genes from Solanum chacoense provides support for a developmental role in plants. Planta 2004, 220, 447–454. [Google Scholar] [CrossRef]
- Loubert-Hudon, A.; Mazin, B.D.; Chevalier, É.; Matton, D.P. The ScRALF3 secreted peptide is involved in sporophyte to gametophyte signalling and affects pollen mitosis I. Plant Biol. 2020, 22, 13–20. [Google Scholar] [CrossRef]
- Wang, P.; Yao, S.; Kosami, K.; Guo, T.; Li, J.; Zhang, Y.; Fukao, Y.; Kaneko-Kawano, T.; Zhang, H.; She, Y.; et al. Identification of endogenous small peptides involved in rice immunity through transcriptomics- and proteomics-based screening. Plant Biotechnol. J. 2019, 18, 415–428. [Google Scholar] [CrossRef]
- Jia, Y.; Li, Y. Genome-Wide Identification and Comparative Analysis of RALF Gene Family in Legume and Non-Legume Species. Int. J. Mol. Sci. 2023, 24, 8842. [Google Scholar] [CrossRef]
- Wang, W.; He, Z.; Wang, D.; Xu, G.; Yang, J.; Feng, X.; Liang, X.; Dou, D.; Chen, R. Functional analysis of RALF peptides in soybean immunity and disease resistance. Phytopathol. Res. 2025, 7, 1–14. [Google Scholar] [CrossRef]
- Wang, W.; Chen, R.; Wang, D.; Zhang, W.; Zhang, Q.; Zhang, Y.; Xu, G.; Yang, J.; Cheng, Z.; Yang, S.; et al. A pair of GPI-anchored proteins regulate soybean immunity and disease resistance in coordination with GmLMM1 and GmRALFs. Plant Biotechnol. J. 2025, 23, 4353–4366. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liang, X.; Bao, Y.; Yang, S.; Zhang, X.; Yu, H.; Zhang, Q.; Xu, G.; Feng, X.; Dou, D. A malectin-like receptor kinase regulates cell death and pattern-triggered immunity in soybean. Embo Rep. 2020, 21, e50442. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Fang, X.; Yu, T.; Chen, J.; Yan, F. Turnip mosaic virus pathogenesis and host resistance mechanisms in Brassica. Hortic. Plant J. 2024, 10, 947–960. [Google Scholar] [CrossRef]
- Rui, P.; Jia, Z.; Fang, X.; Yu, T.; Mao, W.; Lin, J.; et al. A plant viral effector subverts FER - RALF1 module-mediated intracellular immunity. Plant Biotechnology Journal 2025, 23, 2734–2751. [Google Scholar] [CrossRef]
- Shen, W.; Yuan, M.; Chen, L.; Zhang, X. Comparative analysis of rapid alkalinization factor peptide-triggered plant immunity in citrus and closely related species. Plant Physiol. Biochem. 2025, 224, 109941. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, D.; Chen, J.; Wu, D.; Feng, X.; Yu, F. Nematode RALF-Like 1 Targets Soybean Malectin-Like Receptor Kinase to Facilitate Parasitism. Front. Plant Sci. 2021, 12, 775508. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Y.; Jiang, H.; Shui, Z.; Zhong, Y.; Shang, J.; Yang, H.; Sun, X.; Du, J. Genome-wide characterization of soybean RALF genes and their expression responses to Fusarium oxysporum. Front. Plant Sci. 2022, 13, 1006028. [Google Scholar] [CrossRef]
- Prusky, D.; Yakoby, N. Pathogenic fungi: leading or led by ambient pH? Molecular Plant Pathology 2003, 4, 509–516. [Google Scholar] [CrossRef]
- Malinovsky, F.G.; Fangel, J.U.; Willats, W.G.T. The role of the cell wall in plant immunity. Front. Plant Sci. 2014, 5, 178. [Google Scholar] [CrossRef]
- Wei, H.; Yang, R.; Xue, Z.; Zhu, J.; Zhang, Q.; Luan, Y. Molecular Traits of Rapid Alkalinization Factor Family and Functional Analysis of SlRALF2 in Tomato Resistance to Phytophthora infestans. J. Agric. Food Chem. 2025, 73, 3622–3634. [Google Scholar] [CrossRef] [PubMed]
- He, Y.-H.; Zhang, Z.-R.; Xu, Y.-P.; Chen, S.-Y.; Cai, X.-Z. Genome-Wide Identification of Rapid Alkalinization Factor Family in Brassica napus and Functional Analysis of BnRALF10 in Immunity to Sclerotinia sclerotiorum. Front. Plant Sci. 2022, 13, 877404. [Google Scholar] [CrossRef]
- Merino, M.C.; Guidarelli, M.; Negrini, F.; De Biase, D.; Pession, A.; Baraldi, E. Induced expression of the Fragaria × ananassa Rapid alkalinization factor-33-like gene decreases anthracnose ontogenic resistance of unripe strawberry fruit stages. Mol. Plant Pathol. 2019, 20, 1252–1263. [Google Scholar] [CrossRef]
- van Rhijn, P.; Vanderleyden, J. The Rhizobium-plant symbiosis. Microbiol. Rev. 1995, 59, 124–142. [Google Scholar] [CrossRef]
- Solís-Miranda, J.; Juárez-Verdayes, M.A.; Nava, N.; Rosas, P.; Leija-Salas, A.; Cárdenas, L.; Quinto, C. The Phaseolus vulgaris Receptor-Like Kinase PvFER1 and the Small Peptides PvRALF1 and PvRALF6 Regulate Nodule Number as a Function of Nitrate Availability. Int. J. Mol. Sci. 2023, 24, 5230. [Google Scholar] [CrossRef]
- Combier, J.-P.; Küster, H.; Journet, E.-P.; Hohnjec, N.; Gamas, P.; Niebel, A. Evidence for the Involvement in Nodulation of the Two Small Putative Regulatory Peptide-Encoding Genes MtRALFL1 and MtDVL1. Mol. Plant-Microbe Interactions 2008, 21, 1118–1127. [Google Scholar] [CrossRef]
- Li, F.; Mai, C.; Liu, Y.; Deng, Y.; Wu, L.; Zheng, X.; He, H.; Huang, Y.; Luo, Z.; Wang, J. Soybean PHR1-regulated low phosphorus-responsive GmRALF22 promotes phosphate uptake by stimulating the expression of GmPTs. Plant Sci. 2024, 348, 112211. [Google Scholar] [CrossRef]
- Mamaeva, A.; Lyapina, I.; Knyazev, A.; Golub, N.; Mollaev, T.; Chudinova, E.; Elansky, S.; Babenko, V.V.; Veselovsky, V.A.; Klimina, K.M.; et al. RALF peptides modulate immune response in the moss Physcomitrium patens. Front. Plant Sci. 2023, 14, 1077301. [Google Scholar] [CrossRef]
- Lin, H.; Han, X.; Feng, X.; Chen, X.; Lu, X.; Yuan, Z.; Li, Y.; Ye, W.; Yin, Z. Molecular traits and functional analysis of Rapid Alkalinization Factors (RALFs) in four Gossypium species. Int. J. Biol. Macromol. 2022, 194, 84–99. [Google Scholar] [CrossRef]
- Zhang, R.; Li, Y.; Li, Y.; Ma, H.; Zuo, C.; Yang, J.; Ma, Y.; Kong, J.; Wei, X.; Zhu, L.; et al. Polyploidization-driven formation of the GhRALF30L gene cluster confers basal thermotolerance in cotton male reproductive organs via GhFERA1/GhCAPA1. Sci. Adv. 2025, 11, eady1386. [Google Scholar] [CrossRef]
- Jing, X.-Q.; Shi, P.-T.; Zhang, R.; Zhou, M.-R.; Shalmani, A.; Wang, G.-F.; Liu, W.-T.; Li, W.-Q.; Chen, K.-M. Rice kinase OsMRLK63 contributes to drought tolerance by regulating reactive oxygen species production. Plant Physiol. 2023, 194, 2679–2696. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.; Wang, Y.; Gao, Y.; Li, R.; Wang, G.; Li, W.; Liu, W.; Chen, K. The plasma membrane NADPH oxidase OsRbohA plays a crucial role in developmental regulation and drought-stress response in rice. Physiol. Plant. 2015, 156, 421–443. [Google Scholar] [CrossRef]
- Sun, W.; Qin, Y.; Cao, Y.; Tong, J.; Shi, L.; Liu, H.; Wang, G.; Zheng, X.; Zou, L.; Jiang, L.; et al. Genome-wide identification and functional characterization of rapid alkalinization factor 6 as a key peptide regulator of abiotic stress tolerance in Tartary buckwheat. Plant Sci. 2025, 362, 112747. [Google Scholar] [CrossRef]
- Qiao, Q.; Ren, Z.; Fu, X.; Qiao, W.; Sheng, F.; Li, S.; Xiao, D.; He, L. Genome-wide exploration and characterization of the RALFs and analysis of its role in peanut (Arachis hypogaea L.). BMC Plant Biol. 2025, 25, 1–21. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



