
Submitted:
11 November 2025
Posted:
13 November 2025
You are already at the latest version
Abstract
Neural oscillations play a key role in modern neuroscience, linking perception and cognition through rhythmic coordination across distributed networks. Yet the conceptual roots of oscillatory theory trace back long ago. Between 1888 and 1890, Richard Avenarius depicted brain equilibrium as a rhythmic alternation between disturbance and restoration, anticipating the later discovery of EEG and several core concepts of modern neurodynamics. We reinterpret Avenarius’ concept of oscillatory equilibrium and his qualitative vocabulary through the framework of contemporary neural coding theories, encompassing rate, temporal, phase, population, predictive, correlation-based coding, etc. Avenarius’ cyclical sequences of excitation and compensation evoke the homeodynamic and error-corrective processes that govern energy minimization, while his account of oscillatory repetition, synchrony and contrast resonates with modern notions of synaptic adaptation, phase coherence, cross-frequency coupling, attentional modulation, predictive updating within hierarchical neural models. Avenarius’ framework provides also a basis for formulating testable hypotheses about yet unexplored principles of the neural code. From his conception of oscillatory equilibrium arise theoretical possibilities like metabolic–oscillatory coupling, where energy flux and neural rhythms jointly encode information; topological coding, where transient network geometries convey meaning; anti-phase coding, where contrast arises from oscillatory opposition; homeodynamic coding, where informational value lies in the trajectory toward equilibrium; habituation trajectory coding; affective coding; silent coding, etc. Unlike conventional historical analyses that regard philosophical physiology as outdated, we reinterpret it as a theoretical precursor to computational neuroscience, framing Avenarius’ model as a conceptual architecture that unites energy regulation, oscillatory synchronization and informational stability within a coherent dynamic framework.
Keywords:
predictive coding
; consciousness
; habituation
; neurodynamics
; topological representation
Introduction
Neural oscillations are now recognized as fundamental mechanisms underlying perception, cognition and consciousness. Contemporary neuroscience has identified multiple frequency bands, cross-frequency couplings and synchronization patterns that organize communication across distributed neural populations (Jazayeri and Afraz 2017; Huetz et al. 2019; Jeffery 2021; Yuste 2024; Geadah et al. 2024). Experimental findings converge on a model of the brain as a dynamic network of interacting oscillators, where information is transmitted and transformed through structured spatial and temporal coordination. Yet, while neural oscillations dominate present-day theory, their conceptual ancestry remains largely unacknowledged. The functional interpretation of brain oscillations was anticipated by late-nineteenth-century scientists (Ahmed and Cash, 2013). Richard Caton’s 1875 observation of rhythmic potentials in rabbits and monkeys first demonstrated that living tissue can generate spontaneous electrical rhythms (Caton 1875). In the 1890s, Adolf Beck confirmed and extended these results in frogs and dogs, arguing that oscillatory activity is a universal property of the nervous system (Coenen and Zayachkivska, 2013). Vladimir Pravdich-Neminsky’s 1912 photographic electrocerebrogram provided the first objective record of cortical rhythms, paving the way for Hans Berger’s human EEG experiments begun in 1924 and published in 1929 (Berger 1929). These studies collectively transformed speculative physiological ideas about “brain energy” into empirical evidence for structured electrical activity.
The philosophical and functional significance of brain oscillations was simultaneously being explored outside the laboratory. Among the first who theorized the oscillatory nature of thought and experience, Richard Avenarius stands out for proposing that every mental event corresponds to cyclical processes of excitation and stabilization able to restore equilibrium after disturbance. In the sequel, we revisit Avenarius’ account of the brain equilibrium as an alternation of oscillatory phases that integrate physiological variation with psychological experience. We systematically compare Avenarius’ oscillatory framework with current models of neural coding, situating his vital trains within the same taxonomy that encompasses rate, temporal, phase and population codes under a unified grammar of oscillatory equilibrium. By aligning his qualitative description of brain oscillations with modern coding principles, we reinterpret Avenarius’ system as an early dynamic model of informational representation, whose coherence and depth anticipate contemporary theories of neural computation. Also, we argue that a conceptual bridge between nineteenth-century empirio-criticism and twenty-first-century neuroscience may pave the way to novel, testable hypotheses on the structure and functioning of the neural code.
Kritik Der Reinen Erfahrung
In this chapter and the next, we present separately the core elements of Richard Avenarius’ Kritik der reinen Erfahrung and the principal mechanisms that define modern theories of neural coding. We aim to provide a concise, descriptive account of both frameworks before establishing their conceptual and functional correspondences in subsequent sections.
Avenarius’ Kritik der reinen Erfahrung (Avenarius 1888–1890) provides a systematic attempt to explain the relationship between physiological change and psychological experience within a unified naturalistic framework. In Part One ([numbered paragraphs 40–435]), Avenarius formulates a general model of energetic oscillations between disturbance and restoration, through which the brain sustains self-preservation by means of continuous compensatory fluctuations. The “vital train” ([181]) defines the temporal sequence of deviation from equilibrium, compensatory variation and return to stability. Equilibrium is never absolute but constantly renewed through oscillatory exchange between environment and organism.
Part Two ([436–963]) extends the general model to specific oscillatory correlates of various brain activities.
- In §§ [458–462], Avenarius analyzes how the form and magnitude of brain oscillations determine the qualities and intensities of sensory experience. The form of oscillation depends both on external conditions (stimuli) and on the specific preparation of the central system. By varying these conditions, different perceptual elements arise, such as light, sound, color, taste or odor. For instance, heating a filament produces “light,” vibrating a piano string generates “sound,” and applying cologne on the skin evokes a “burning” sensation. The magnitude of oscillation, in contrast, determines intensity: a stronger vibration produces a louder tone, brighter light or more vivid sensation. Avenarius concludes that perceptual quality depends on the form of oscillation, while intensity depends on its amplitude.
- In §§ [463–469], Avenarius examines how the relevance and direction of brain oscillations determine emotional life and bodily expression. An oscillation is considered more relevant when it is both large in magnitude and involves central subsystems with high physiological or experiential significance—shaped by individual predisposition and habitual practice. The resulting variations evoke affective responses, expressed as pleasure or displeasure. For example, a mother’s anxiety for a sick child, an artist’s frustration at a ruined color or a scientist’s excitement over a new discovery all reflect oscillations of different relevance and scope. Pleasure arises when oscillatory processes restore equilibrium between “work” and “nourishment,” while displeasure follows imbalance.
- In §§ [466–469], Avenarius introduces the notion of a value of indifference, the threshold where pleasure turns into displeasure, inspired by Wundt’s psychophysiology. He interprets emotional tone as a function of oscillatory direction: an increase in relevant oscillations produces displeasure, while a decrease generates pleasure. These rhythmic variations manifest physiologically through motor and visceral changes like muscle tension, breathing patterns, heart rate, sweating or warmth, each corresponding to specific oscillatory configurations. Thus, affective experience is not an added property of consciousness but an intrinsic modulation of oscillatory processes linking movement, sensation and emotion within a unified dynamic system.
- In §§ [471–477], Avenarius explores the dependence of transexcitation, describing how oscillations in one sensory or motor subsystem can propagate through others, producing complex experiential effects. Sensations such as dizziness, shock or loss of balance arise from sudden oscillatory transfers within the central system. He cites Billroth’s anecdote of a soprano singing off-key, which caused a sharp toothache, an example of co-affective dependence where an auditory stimulus triggers a physiological reaction elsewhere. Avenarius distinguishes between proper feelings (directly related to one’s own oscillatory processes) and improper feelings (arising from cross-system propagation). Through repeated excitation, oscillations can deviate from their habitual form, producing a qualitative sense of “otherness”. This character manifests when the individual perceives difference, such as moving to a new country, learning a foreign language or encountering unfamiliar art. In contrast, when oscillations revert to their previous pattern, an individual sense of restored sameness and identity is achieved, captured in expressions such as “it’s just the same.”. Avenarius summarizes these relationships by describing the polarity of experiential differentiation that links novelty, familiarity and emotional tone to dynamic oscillatory processes of the nervous system.
- In §§ [478–498] Avenarius examines how exercise shapes oscillatory dependence and introduces a family of characters that grade experience between novelty and sameness. He defines Idential as the intermediate value that varies with transexcitation. Exercise of oscillations gives rise to a character of Fidentiality, the felt familiarity of well-practiced patterns exemplified by Heimat, whose deprivation elicits homesickness. When a less exercised oscillation is imposed on the nervous system, the opposed character emerges as Unfamiliarity, seen in reactions to cadavers by novice students, to stage tricks or to erratic social behavior. Fidentiality decomposes into three specific characters: reality, security and familiarity. Their unity often appears in everyday compounds such as the known road, the trusted physician or native currency. Avenarius traces how practice modulates these values: when exercise decreases, they drift toward their negative counterparts such as insecurity and attenuation, passing through a point of indifference. He details cultural and temporal modulations in which the present feels maximally real, the distant or past less so and copies, images and dreams hold reduced existential weight; yet repeated engagement can raise their existential value. He then outlines how social practice establishes what counts as normal, while unusual biological or social cases appear strange and how repeated exposure converts the strange into the familiar.
- In §§ [499–502] Avenarius analyzes the dependence of the articulation of fluctuations. The nervous system shifts from relative uniformity of internal links toward finer differentiation, where gradual change raises internal articulation while abrupt change can weaken cohesion. Formal separation picks out from a continuous whole, the specific “new work” and the part currently in focus: the workbench becomes distinct from the room, the corrected letter from the line, the striking advertisement from the page. What becomes formally separated gains prevalence; what recedes becomes a dead value, later recoverable by memory or by pedagogical maneuvers that vary familiar conditions (pointing, rearranging, naming). Excessive separation yields over-separation (confusion, disorder). Avenarius stresses that prevalence is not intensity: a loud letterform can be intense yet not prevail over content, while content may prevail with moderate intensity. Prevalence arises from changes against previously constant conditions and can decline if change is too slow or too slight. Habituation converts earlier separations into dead values (the miller ceases to hear the mill; silence becomes the separated value). Conversely, confused complexes can become separated through practice (a bustling city becomes comprehensible; Wagner’s overture shifts from chaos to lucid structure). Avenarius then tackles the principle of opposition of fluctuations. Material contrasts heighten mutual distinctness: complementary colors differentiate each other, bright stands out from dark, loud from quiet, heavy from light; analogous oppositions structure affect (joy against pain, love against hate) and life-world judgments (home against foreign). Yet contrasts have limits. If gaps are too large or too abrupt, prevalence collapses into confusion, stupor or blinding. The graded management of transitions in teaching and communication therefore regulates articulation and preserves comprehension. He concludes that contrast depends on the opposition of fluctuations, which enhances material distinctness within certain limits, but beyond those limits breaks articulation down into confusion and loss of structure.
The Kritik can thus be reconstructed as a proto-dynamical system in which oscillations mediate between physiological energy exchange and psychological meaning, hinting the empirical discovery of the brain electrical rhythms in animals.
2. The Neural Code
The neural code refers to the principles through which information is represented and transmitted within the nervous system. The following section outlines the main coding schemes that have been proposed to underlie these processes.
- Rate coding mechanisms encode information in the average firing rate of neurons over time. In firing rate coding, the number of spikes per unit time represents stimulus intensity, with higher firing rates typically corresponding to stronger stimuli (Gallistel 2017; Tomar 2019; Zhu et al., 2025). Time-averaged rate coding smooths neural activity over a time window to reduce variability in spike timing, while Poisson rate coding treats spike generation as a probabilistic process, where the rate parameter itself carries the information (Satuvuori and Kreuz, 2018; Liu et al., 2021).
- Temporal coding mechanisms rely on the precise timing of spikes. In spike timing coding, the exact moment a spike occurs relative to an internal or external reference carries meaning, as in first-spike latency (Li et al., 2018; Beckert et al.; Chen et al., 2024). Phase coding represents information through the timing of spikes within an oscillatory cycle, such as theta-phase alignment in the hippocampus (Seenivasan and Narayanan, 2020; Pacheco Estefan et al., 2021). Temporal pattern coding uses specific sequences of spikes to convey information (Madar et al., 2019), while interspike interval coding relies on the time between spikes to represent stimulus properties (Oswald et al., 2007; Koyama and Kostal, 2014). Synchrony coding occurs when groups of neurons fire simultaneously to signal the presence or relevance of a stimulus (Person and Raman, 2012; Baker et al., 2015; Rezaei et al., 2023)
- Population coding mechanisms involve distributed activity across multiple neurons (Georgopoulos et al., 1986; Runyan et al., 2017; Downer et al., 2017; LeMessurier and Feldman, 2018; Levitan et al., 2019; Downer et al., 2021; Stringer et al., 2021). In distributed coding, information is represented collectively by many neurons rather than by single units. Sparse coding makes use of small subsets of neurons, creating efficient representations. Vector coding models population responses as vectors in high-dimensional space, while basis function coding describes neural responses as components capable of representing arbitrary inputs. Redundancy reduction coding distributes information in a way that minimizes overlap and enhances efficiency.
- Correlation-based coding emphasizes interactions among neurons (Hong et al., 2012; Fox and Stryker, 2017; Montijn et al., 2014; Azeredo da Silveira and Rieke, 2021; Dora, et al. 2021; Tschantz et al., 2023; Zeng et al., 2023; Millidge et al., 2024). Population synchrony refers to correlations in firing across neurons that shape how information is encoded. Noise correlation coding focuses on correlated variability in responses, while Hebbian coding describes learning through the strengthening of correlated activity, exemplified by spike-timing-dependent plasticity.
- Predictive and Bayesian coding mechanisms treat neural computation as probabilistic inference (Aitchison and Lengyel, 2017; Bonetti et al., 2021; Pezzulo et al., 2022; Caucheteux et al., 2023; Lange et al., 2023; Chao et al., 2024; Taniguchi 2024). In Bayesian coding, neural populations represent probability distribution over sensory or cognitive variables. Predictive coding proposes that the brain continuously generates expectations about incoming inputs and adjusts them through error signals. The free energy principle unifies these perspectives by suggesting that the brain reduces uncertainty through internal modeling of expected sensory states.
- Specialized coding mechanisms apply to sensory or motor systems. Place coding in the hippocampus represents spatial position, while grid coding in the entorhinal cortex maps locations in a hexagonal grid (Mallory and Giocomo, 2018; Herzog et al., 2019; Zhou et al., 2024). Opponent coding describes systems that encode information through contrasting signals, as in color vision (Buchsbaum and Gottschalk, 1983; Derey et al., 2016; Rhodes et al., 2017; Hagihara and Lüthi, 2024). Rank order coding conveys information through the sequence in which neurons fire and time-to-first-spike coding uses the delay from stimulus onset to the first action potential as a signal (Bonilla et al., 2022; Liu et al., 2023; Sakemi et al., 2023; Kim et al., 2024; Li et al., 2024).
- Hybrid coding schemes combine multiple strategies depending on function and context. Multiplexed coding integrates distinct mechanisms such as rate and phase coding to convey different aspects of information simultaneously (Baker et al., 2013; Hong et al., 2016; Ke et al., 2022; Hovhannisyan et al., 2023). Hierarchical coding organizes information processing in successive stages of increasing abstraction, as seen in the visual pathway from V1 to the inferotemporal cortex (Chen 2023; Gwilliams et al., 2025).
The neural code is therefore diverse and context-dependent, with different brain regions, sensory modalities and behavioral states relying on distinct yet often overlapping coding strategies. Understanding how these coding schemes interact remains a major challenge in neuroscience, as the brain likely employs a flexible combination of these principles to achieve adaptive and efficient information processing.
3. Parallels Between Avenarius’ Vital Trains and Modern Neural Codes
Our comparative analysis reveals a close functional correspondence between Avenarius’ description of oscillatory processes and the mechanisms that contemporary neuroscience designates as neural codes. In particular, the link between Avenarius’ Part Two [436–963] and current neuroscience of the neural code is remarkably rich, because both attempt to describe how patterns of physiological oscillations correspond to psychological states or meanings. A detailed comparison is presented below relating Avenarius’ main issues to their modern neurophysiological counterparts.
Brain oscillations as the basis of statements [455, 459–475]. Avenarius maintains that every statement depends upon oscillatory variations occurring in the brain. He identifies three primary dimensions of oscillatory behavior, each of which underlies distinct experiential qualities. Oscillation magnitude determines the intensity of perception, form differentiates qualitative aspects of sensations and direction introduces polarity and valence, thereby linking neural dynamics with emotional tone.
Modern neuroscience provides a striking parallel to this tripartite structure through the mechanisms of rate, temporal and phase coding.
- In rate coding, the number of spikes or the oscillatory power within a frequency band represents stimulus intensity; higher firing rates or stronger gamma-band amplitudes correspond to stronger stimuli, echoing Avenarius’[455,459–475 “magnitude” ([461]).
- The form of oscillation corresponds to waveform structure and spectral composition underlying temporal and resonance-based models of information processing.
- Finally, the direction of oscillation could match with, e.g., frontal alpha asymmetry correlating with emotional polarity and theta–gamma coupling distinguishing between positive and negative valence.
Together, Avenarius’ triadic structure seems to foreshadow the modern view of oscillatory coding where amplitude, frequency and phase jointly encode informational content and emotional meaning. His claim that every statement is grounded in oscillation thus may correspond to the contemporary notion that cognition and perception are emergent from structured rhythmic activity in neural ensembles.
Habituation, familiarity and identity [474–479]. Avenarius postulates that repeated identical oscillations generate sensations of familiarity, certainty and reality, whereas novel oscillations yield difference, variance and uncertainty. He thereby introduces the physiological roots of identity and recognition: repetition creates coherence, while variation produces novelty and surprise.
Modern neuroscience parallels these insights in several interconnected coding frameworks.
- Predictive coding interprets repeated stimuli as generating attenuated neural responses (a reduction in prediction error) much like Avenarius’ description of oscillatory convergence toward equilibrium.
- In rate and temporal coding, repetition reduces firing variability and sharpens spike-timing precision, increasing signal efficiency.
- The free-energy principle formalizes this process mathematically, describing how cortical circuits minimize the divergence between predicted and actual input (Kullback–Leibler divergence), a direct computational analogue of Avenarius’ “restoration” phase.
- Moreover, the emergence of “identity” through synchronized oscillations corresponds to phase locking and coherence, i.e., neural phenomena now regarded as signatures of perceptual binding and conscious unity.
Avenarius’ approach thus provides one of the earliest formulations of the neural mechanisms underlying predictive processing, linking oscillatory regularity with stability of experience and the emergence of coherent perceptual identity.
Contrast, attention and opposition [505]. In Avenarius’ system, attention arises from the opposition between oscillations: when two or more oscillatory fields interact, contrast emerges, allowing one component to prevail as the focus of perception.
- This model of contrast through opposition corresponds closely to contemporary theories of selective attention based on oscillatory competition, in which attention depends on the dynamic regulation of synchrony and desynchrony among neural populations.
- According to the communication-through-coherence hypothesis, synchronized assemblies amplify relevant information, whereas desynchronized activity is suppressed. This may stand for a functional analogue of Avenarius’ crucial concept of “extraction of a prevalent part from the background.”
- Furthermore, anti-phase coupling and cross-frequency desynchronization provide mechanisms for sensory discrimination and attentional gating, reflecting Avenarius’ observation that oscillatory opposition sharpens contrast and enhances awareness.
Overall, Avenarius’ oscillatory interplay foreshadows modern understandings of cortical coordination, in which cognitive focus emerges from rhythmic competition within large-scale networks, rather than from static hierarchies of processing.
Formation of concepts [512–552]). Avenarius attributes the formation of concepts, generalizations and truths to increasing oscillatory connectivity across the brain. When oscillations link repeatedly into longer and more stable chains, they produce enduring representations, what he calls things and ideas. This process culminates in the establishment of abstract laws and general categories ([552]).
- Modern neuroscience identifies comparable principles in population coding and vector coding, where distributed patterns of neuronal activity across large ensembles represent complex sensory or cognitive objects. Each concept may correspond to a stable attractor within the multidimensional state space of neural dynamics.
- The transition from repeated sensory oscillations to higher-level abstraction parallels the hierarchical organization of predictive models, in which recurrent associations generate more general predictive structures.
- Avenarius’ insight that generality results from the recurrence of identical oscillatory patterns seems to anticipate the Hebbian plasticity (“neurons that fire together wire together”) and Bayesian abstraction, in which probabilistic integration across repeated experiences yields conceptual knowledge.
Thus, Avenarius’ account of oscillatory chaining mechanisms foreshadows the construction of distributed cognitive maps and hierarchical representations in both neuroscience and artificial neural networks.
Structure of a specific vital train [775–857]. Avenarius defines the specific vital train as a tripartite oscillatory process composed of equilibrium/expectation, disturbance/variation and restoration/suppression. Each stage carries a distinct affective tone: expectation corresponds to stability, disturbance to uncertainty and discomfort and restoration to satisfaction or truth.
- This cyclical pattern maps directly onto the predictive-coding loop of modern neuroscience that comprises top-down predictions, bottom-up error signals and feedback corrections able to minimize free energy.
- The “pain of uncertainty” and “pleasure of certainty” may correspond to the dopaminergic reward and error signals that modulate learning and behavioral adaptation.
- Furthermore, Avenarius’ assertion that the final equilibrium is independent of the number of intermediate steps ([857]) parallels recurrent neural network convergence, where dynamic systems settle into stable attractors regardless of the complexity of their transient paths.
Avenarius’ vital train thus represents an early formal model of adaptive control and predictive homeostasis that bonds emotion, cognition and physiology within a single oscillatory cycle.
Reduction of variations and the search for laws [888–891]. In the later sections [§§888–891], Avenarius identifies three fundamental operations by which the nervous system restores equilibrium amid oscillatory disturbances.
The first is the reduction of variation, or the transformation of the unfamiliar into the familiar, an operation through which novel stimuli are integrated into existing patterns of experience.
The second is habituation, achieved through repetition, whereby recurrent oscillations lose their disruptive force and become stable components of the system’s dynamics.
The third is substitution, in which one pattern of change replaces another, allowing the system to preserve overall stability while adapting its configuration.
Together, these operations aim at the progressive elimination of variability, culminating in generalized stability, what he terms “laws.”
- In modern terms, this account describes the brain’s tendency toward homeostatic regulation and efficient coding, where synaptic scaling maintains stable firing statistics and redundancy reduction optimizes informational efficiency.
- Similarly, plasticity mechanisms allow substitutional learning, ensuring that changing inputs still produce coherent representations.
- Avenarius’ search for universal oscillatory laws prefigures computational neuroscience’s emphasis on compression and generalization, principles that also underpin deep learning networks.
The concept of cognition converging toward minimal predictive models is a remarkable anticipation of the brain’s optimization strategies as described by modern information theory.
Experience as character and content [931–963]. Avenarius distinguishes between experience as character, i.e., the variable oscillatory form associated with emotions and sensations and experience as content, the stable perceptual or conceptual element. He attributes both to specific oscillatory states and to each individual’s neural “education,” thereby introducing a form of experience-dependent individuality.
- This duality parallels the modern separation between dynamic neural activity and representational structure. In population and hybrid coding schemes, form may correspond to transient activity patterns, while content to stable network configurations. The dependence on individual history anticipates experience-dependent plasticity, whereby learning refines neural coding through long-term synaptic modification.
- Notably, Avenarius’ claim that imagined or hallucinatory experiences correspond to real oscillatory content ([961]) finds confirmation in modern neuroimaging studies showing that visual imagery activates cortical regions similar to those engaged during perception.
Avenarius’ treatment of character and content thus connects phenomenological variability and physiological stability, providing one of the earliest models of the subjective–objective continuum in brain function.
Oscillatory equilibrium and self-preservation [160–185,314,406,409]. In Part One, Avenarius formulates a general physiological law stating that the brain oscillates between disturbance and restoration to ensure self-preservation. In Part Two, this principle evolves into the continual re-stabilization of oscillatory patterns.
- This dynamic equilibrium corresponds to modern concepts of homeodynamic regulation, metastability and self-organized criticality in neural systems. Current models describe perception and cognition as trajectories through high-dimensional state spaces seeking temporary stability, i.e., energy minima, while maintaining flexibility for adaptation. This is effectively Avenarius’ “quiet, maximum-preservation state,” in which the organism sustains informational balance through rhythmic readjustment.
The alignment between Avenarius’ oscillatory equilibrium and modern theories of dynamic brain coordination underscores his concept of the brain as an open, adaptive system whose stability depends on perpetual motion rather than stasis.
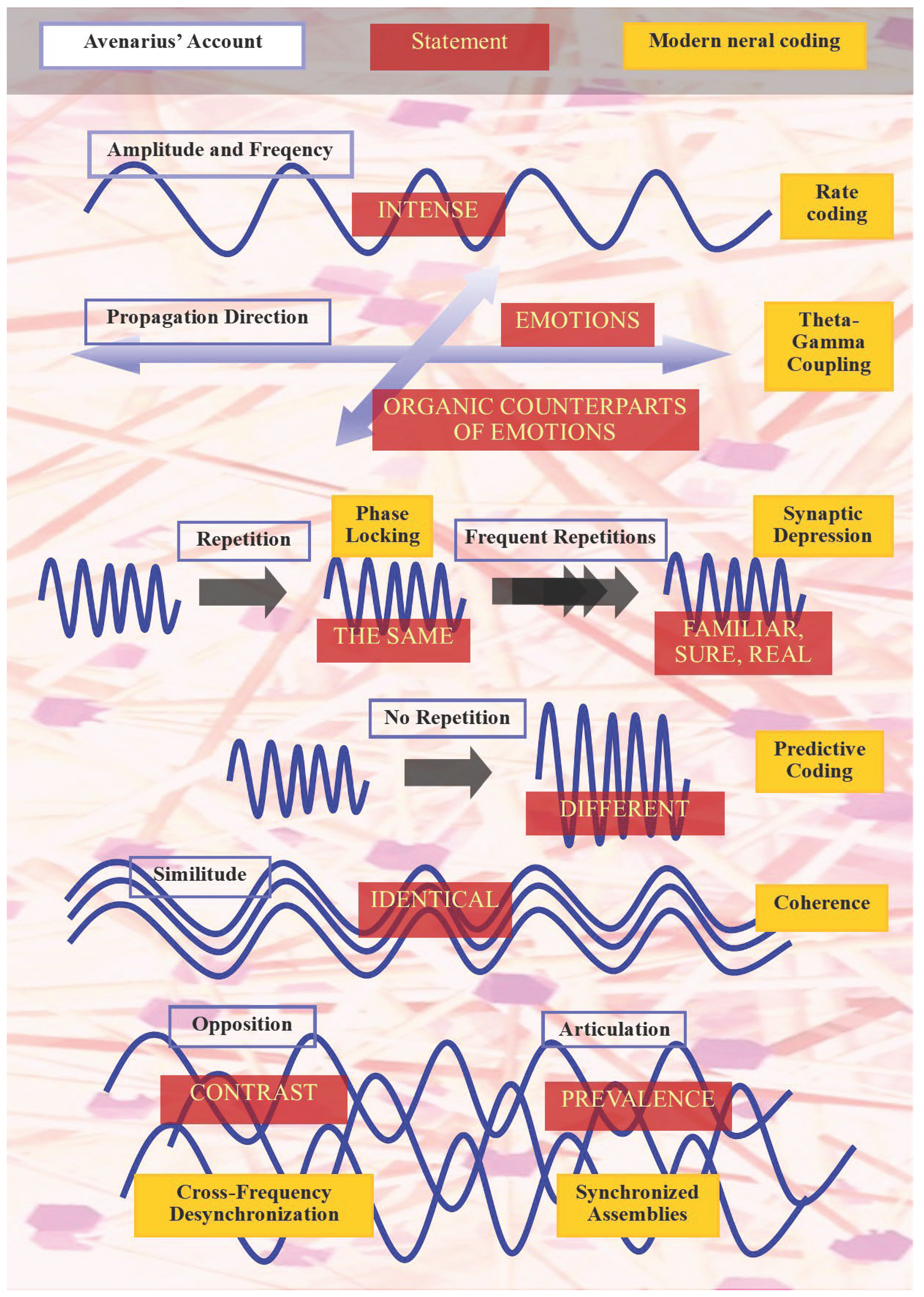
Taken together, Avenarius’ account (Figure) anticipates a unified neurodynamic theory in which cognition, emotion and perception emerge from continuous oscillatory equilibration between prediction and perturbation. His “statements” correspond to neural code patterns, i.e., configurations of spikes, phases, synchronies and correlations whose stability or disruption yields meaning and consciousness. Implicitly, he foresaw not only rate, temporal and phase coding but also population, correlation-based, predictive and Bayesian mechanisms, as well as hybrid schemes combining them hierarchically. The Kritik der reinen Erfahrung therefore represents one of the earliest articulations of a dynamic informational brain model, in which coherence, adaptation and self-stabilization form the organizing principles of mental life.
Figure 1.
Comparative mapping between Avenarius’ oscillatory features and statements and modern neural coding’s theories.
Figure 1.
Comparative mapping between Avenarius’ oscillatory features and statements and modern neural coding’s theories.
4. Towards Yet-Undiscovered Principles of Neural Coding
Avenarius’ model extends beyond the idea of oscillations as mere rhythmic events, presenting instead a general functional grammar that links disturbance and restoration, environmental variation and neural self-equilibration within a unified dynamic scheme. From this grammar, several unexplored possibilities can be logically derived, suggesting that scientists could extrapolate from Avenarius’ framework to formulate new hypotheses about yet-undiscovered mechanisms of neural coding.
Morphological or topological coding. Avenarius’ organization of the nervous system ([73,160–185,223]) describes functional subsystems linked by oscillatory compensation, emphasizing the form of coordination rather than its anatomical substrate.
From this arises the idea of morphological or topological coding, in which meaning is carried by the instantaneous topology of neural connectivity. Instead of relying solely on spike timing or firing rate, information could be inscribed in the transient geometric features of brain networks, e.g., loops, cavities or higher-dimensional simplices arising and dissolving during oscillatory reorganization.
Modern approaches such as topological data analysis and graph homology mapping could be employed to test whether shifts in network shape correlate with variations in perception or cognition, thereby assessing the feasibility of topological transformations as genuine carriers of information.
Metabolic–oscillatory coupling. Avenarius described oscillations also in terms of energetic equilibria between work and nourishment ([155,160]), suggesting that informational processes are inseparable from metabolic flux.
This notion supports the theoretical concept of metabolic–oscillatory coupling, where fluctuations in, e.g., glucose metabolism, astrocytic signaling or mitochondrial potential are integrated with electrical rhythms to form a thermodynamic code. Meaning would emerge from the dynamic equilibrium of energy exchange rather than from spikes alone.
Multimodal techniques combining EEG, fMRI and PET imaging could help evaluate whether variations in metabolic cycles systematically correspond to cognitive states, providing a possible empirical framework for exploring energetic dimensions of neural coding.
Counter-oscillatory or anti-phase coding. When describing attention and contrast, Avenarius ([505]) emphasized the importance of oscillatory opposition, where simultaneous rhythms interact through polarity rather than synchrony.
This leads to the hypothesis of counter-oscillatory or anti-phase coding, a mechanism in which information is stored not in aligned firing but in controlled phase inversion between oscillators. Such differential phase relations could allow the brain to encode contrast, context and selection without requiring precise coherence.
Tools such as phase-difference mapping and cross-frequency interaction analysis could be applied to determine whether specific anti-phase configurations consistently track attentional or perceptual distinctions.
Habituation-trajectory coding. Avenarius regarded habituation as the process by which oscillatory regularity transforms uncertainty into certainty ([474–479,775–857]).
This principle suggests the possibility of a habituation-trajectory coding where information resides in the rate and path of convergence toward equilibrium rather than in discrete signal events. The slope of stabilization, i.e., the temporal dynamics through which variance diminishes, would itself constitute a meaningful variable.
Learning-curve modeling and variance-reduction EEG paradigms could be designed to assess whether neural systems encode the pace of habituation as an informational quantity, potentially linking Avenarius’ description of familiarity formation to measurable adaptation rates.
Character-based or affective coding. In his discussion of characters ([469,931–953]), e.g., qualities such as pleasantness, familiarity and tension, Avenarius proposed that emotions and valuations correspond to modulations of oscillatory activity.
This anticipates the notion of character-based or affective coding, in which informational content is tagged by emotional modulation through amplitude and frequency envelopes. Instead of separating cognition and emotion, the oscillatory code would integrate them as a single representational structure.
Experimental paradigms using amplitude–frequency envelope analyses in affective EEG and fMRI could explore whether emotional states systematically alter the parameters of ongoing oscillations in a manner consistent with Avenarius’ predictions.
Equilibrium-seeking or homeodynamic coding. Every Avenarius’ vital train ends with a restoration of balance ([181–189,850]), implying that the informational significance of a process lies in its deviation from and return to equilibrium.
This idea supports a theoretical equilibrium-seeking or homeodynamic coding, where information corresponds to the trajectory leading the system back to stability. The value of a neural event would be determined not by its amplitude or rate, but by its position within the path of recovery.
Contemporary dynamical manifold modeling and free-energy minimization frameworks could be adapted to examine whether specific patterns of return-to-equilibrium behavior correlate with cognitive resolution or perceptual completion.
Plasticity-as-message coding. Because equilibrium in Avenarius’ system entails permanent organic transformation ([116,432]), it can be extended to plasticity-as-message coding, where structural modifications themselves carry information.
Processes such as dendritic branching, synaptic pruning and receptor redistribution would act as slow communication channels complementing fast oscillatory codes.
Techniques from structural connectomics, receptor mapping and long-term potentiation imaging could be used to test whether enduring morphological changes function as carriers of encoded experience, thus validating Avenarius’ claim that preservation and transformation coexist within the same oscillatory law.
Silent or potential-state coding. Avenarius insisted that quiescent or steady states of the brain ([314,409]) are not void of meaning but signify maximal preservation and readiness.
This concept suggests silent or potential-state coding, where the absence of visible oscillation encodes latent informational potential. Subthreshold activity, resting-state field organization and entropy minima could represent configurations of readiness awaiting activation.
High-sensitivity recordings of subthreshold dynamics and low-entropy field analyses might be applied to determine whether neural quietness corresponds to structured informational states, testing the possibility that stability itself functions as a code.
Overall, Avenarius’ functional grammar suggests that the neural code could represent a multilayered system of oscillatory correspondences. This unified speculative vision could redefine the neural code as a dynamic and living field of equilibrium processes rather than a fixed computational scheme with logical rules. Modern experimental techniques across electrophysiology, connectomics and computational modeling could be employed to evaluate these possibilities.
5. Conclusions: Lessons from the Past
Our approach establishes conceptual continuity between philosophical theories of mind and empirical models of neural computation, employing textual analysis and functional correspondence to reinterpret nineteenth-century terminology within the parameters of contemporary neuroscience. In this light, Avenarius emerges as a theoretical precursor of dynamical neuroscience, anticipating key principles that would later be validated through electrophysiological and computational research. Long before the advent of EEG, Avenarius described oscillatory processes in the brain as the physiological substrate of sensation, emotion and cognition. Still, Avenarius’ concept of oscillatory equilibrium emerges as an early formalization of the dynamic balance between variability and stability sustaining perception and cognition through rhythmic regulation of energy and information in the brain.
In comparison with approaches that treat separately energy metabolism, signal transmission and cognition, Avenarius’ account anticipates a unified informational principle in which these domains are mutually dependent (Sokoloski et al. 2021; Joiret et al. 2023; Shi et al. 2023). Predictive coding, dynamical systems theory and free-energy minimization share with the Kritik the principle that the brain continually restores equilibrium by minimizing error between internal and external states (Friston et al., 2006; Hemmatian et al., 2024; Lu 2024; Vallortigara and Vitiello, 2024). However, Avenarius’ formulation extends beyond signal optimization to include metabolic and experiential dimensions, prefiguring the energetic constraints central to modern neuroenergetics. His tripartite vital train parallels the structure of attractor dynamics but embeds it within a broader epistemic cycle linking perception and knowledge formation. Where information theory treats the neural code statistically, Avenarius conceives it functionally, as rhythmic coordination among physiological subsystems. Avenarius’ distinction between elements and characters anticipates the separation between representational content and affective modulation in current models of hierarchical processing. In contrast with later mechanistic theories, his framework preserves a descriptive fidelity to experience while remaining compatible with measurable physiological correlates. Therefore, the Kritik can be understood simultaneously as an early dynamic-systems model and a phenomenological analysis of oscillatory function.
Our approach has intrinsic limitations since the mapping between “vital trains” and contemporary neural codes remains conceptual. In historical terms, translation from late nineteenth-century idioms introduces interpretive uncertainty, particularly regarding the energetic metaphors predating formal thermodynamics and information theory. While the parallel between oscillatory equilibrium and predictive coding is compelling, it may overstate the degree of functional identity between philosophical description and empirical mechanism. Our approach also relies on a selective alignment of modern coding theories, omitting stochastic and molecular forms of information processing that exceed the scope of Avenarius’ model.
Nevertheless, our synthesis paves the way to potential directions for empirical research. High-resolution EEG and MEG analyses could test whether specific oscillatory patterns predicted by Avenarius, such as synchronization correlating with identity or desynchronization with contrast, occur across sensory and cognitive tasks. Computational models could simulate vital trains as recurrent processes minimizing internal error through oscillatory feedback. In philosophical and methodological terms, our approach may enrich neurophenomenology by embedding subjective experience within quantifiable oscillatory dynamics. Furthermore, Avenarius’ emphasis on homeodynamic equilibrium could inform artificial intelligence architectures based on self-preservative regulation rather than static optimization. Still, we expanded this correspondence into predictive and speculative terrain. By extrapolating Avenarius’ functional grammar of disturbance and restoration, we outlined a coherent set of potential coding principles still unexplored in neuroscience.
In conclusion, we showed that Avenarius’ oscillatory model anticipated key mechanisms of contemporary coding strategies, predictive models and oscillatory neural computation. Recognizing this continuity situates present-day neurodynamics within a broader intellectual tradition and clarifies that the oscillatory nature of neural activity is a rediscovery of a deep conceptual structure. Intriguingly, Avenarius’ insights may also inspire future theoretical frameworks and guide the exploration of yet-unrecognized principles shaping the dynamic oscillatory patterns of the central nervous system.
Author Contributions
The Author performed: study concept and design, acquisition of data, analysis and interpretation of data, drafting of the manuscript, critical revision of the manuscript for important intellectual content, statistical analysis, obtained funding, administrative, technical and material support, study supervision.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial or not-for-profit sectors.
Institutional Review Board Statement
This research does not contain any studies with human participants or animals performed by the Author.
Data Availability Statement
All data and materials generated or analyzed during this study are included in the manuscript. The Author had full access to all the data in the study and took responsibility for the integrity of the data and the accuracy of the data analysis.
Acknowledgments
None.
Declaration of generative AI and AI-assisted technologies in the writing process
During the preparation of this work, the author used ChatGPT 4o to assist with data analysis and manuscript drafting and to improve spelling, grammar and general editing. After using this tool, the author reviewed and edited the content as needed, taking full responsibility for the content of the publication.
Conflicts of Interest
The Author does not have any known or potential conflict of interest including any financial, personal or other relationships with other people or organizations within three years of beginning the submitted work that could inappropriately influence or be perceived to influence their work. The Author transfers all copyright ownership, in the event the work is published. The undersigned author warrants that the article is original, does not infringe on any copyright or other proprietary right of any third part, is not under consideration by another journal and has not been previously published.
References
- Ahmed, Omar J. and Sydney S. Cash. “Finding Synchrony in the Desynchronized EEG: The History and Interpretation of Gamma Rhythms.” Frontiers in Integrative Neuroscience 7 (August 12, 2013): 58. [CrossRef]
- Aitchison, L., and M. Lengyel. “With or Without You: Predictive Coding and Bayesian Inference in the Brain.” Current Opinion in Neurobiology 46 (October 2017): 219–227. [CrossRef]
- Avenarius, Richard. Kritik der reinen Erfahrung. 2 vols. Leipzig: Verlag von Wilhelm Engelmann, 1888–1890.
- Azeredo da Silveira, R., and F. Rieke. “The Geometry of Information Coding in Correlated Neural Populations.” Annual Review of Neuroscience 44 (July 8, 2021): 403–424. [CrossRef]
- Baker, C. A., T. Kohashi, A. M. Lyons-Warren, X. Ma, and B. A. Carlson. “Multiplexed Temporal Coding of Electric Communication Signals in Mormyrid Fishes.” Journal of Experimental Biology 216, no. 13 (July 1, 2013): 2365–2379. [CrossRef]
- Baker, C. A., Huck, K. R., and Carlson, B. A. “Peripheral Sensory Coding through Oscillatory Synchrony in Weakly Electric Fish.” eLife 4 (August 4, 2015): e08163. [CrossRef]
- Beckert, M. V., Fischer, B. J., and Peña, J. L. “Effect of Stimulus-Dependent Spike Timing on Population Coding of Sound Location in the Owl’s Auditory Midbrain.” eNeuro 7, no. 2 (April 23, 2020): ENEURO.0244-19.2020. [CrossRef]
- Berger, Hans. “Über das Elektrenkephalogramm des Menschen.” Archiv für Psychiatrie und Nervenkrankheiten 87 (1929): 527–570. [CrossRef]
- Bonilla, L., J. Gautrais, S. Thorpe, and T. Masquelier. “Analyzing Time-to-First-Spike Coding Schemes: A Theoretical Approach.” Frontiers in Neuroscience 16 (September 26, 2022): 971937. [CrossRef]
- Bonetti, L., S. E. P. Bruzzone, N. A. Sedghi, N. T. Haumann, T. Paunio, K. Kantojärvi, M. Kliuchko, P. Vuust, and E. Brattico. “Brain Predictive Coding Processes Are Associated to COMT Gene Val158Met Polymorphism.” NeuroImage 233 (June 2021): 117954. [CrossRef]
- Buchsbaum, G., and A. Gottschalk. “Trichromacy, Opponent Colours Coding and Optimum Colour Information Transmission in the Retina.” Proceedings of the Royal Society B: Biological Sciences 220, no. 1218 (November 22, 1983): 89–113. [CrossRef]
- Caton, Richard. “The Electric Currents of the Brain.” British Medical Journal 2 (1875): 278. http://echo.mpiwg-berlin.mpg.de/MPIWG:4917F9YN.
- Caucheteux, C., A. Gramfort, and J. R. King. “Evidence of a Predictive Coding Hierarchy in the Human Brain Listening to Speech.” Nature Human Behaviour 7, no. 3 (March 2023): 430–441. [CrossRef]
- Chao, Z. C., M. Komatsu, M. Matsumoto, K. Iijima, K. Nakagaki, and N. Ichinohe. “Erroneous Predictive Coding across Brain Hierarchies in a Non-Human Primate Model of Autism Spectrum Disorder.” Communications Biology 7, no. 1 (July 12, 2024): 851. [CrossRef]
- Chen, Z. S. “Hierarchical Predictive Coding in Distributed Pain Circuits.” Frontiers in Neural Circuits 17 (March 3, 2023): 1073537. [CrossRef]
- Chen, L., Karilanova, S., Chaki, S., Wen, C., Wang, L., Winblad, B., Zhang, S. L., Özçelikkale, A., and Zhang, Z. B. “Spike Timing-Based Coding in Neuromimetic Tactile System Enables Dynamic Object Classification.” Science 384, no. 6696 (May 10, 2024): 660–665. [CrossRef]
- Coenen, Anton, Zayachkivska, Oksana. “Adolf Beck: A Pioneer in Electroencephalography in Between Richard Caton and Hans Berger.” Advances in Cognitive Psychology 9, no. 4 (December 31, 2013): 216–221. [CrossRef]
- Derey, K., G. Valente, B. de Gelder, and E. Formisano. “Opponent Coding of Sound Location (Azimuth) in Planum Temporale Is Robust to Sound-Level Variations.” Cerebral Cortex 26, no. 1 (January 2016): 450–464. [CrossRef]
- Dora, S., S. M. Bohte, and C. M. A. Pennartz. “Deep Gated Hebbian Predictive Coding Accounts for Emergence of Complex Neural Response Properties Along the Visual Cortical Hierarchy.” Frontiers in Computational Neuroscience 15 (July 28, 2021): 666131. [CrossRef]
- Downer, J. D., Niwa, M., and Sutter, M. L. “Hierarchical Differences in Population Coding within Auditory Cortex.” Journal of Neurophysiology 118, no. 2 (August 1, 2017): 717–731. [CrossRef]
- Downer, J. D., Bigelow, J., Runfeldt, M. J., and Malone, B. J. “Temporally Precise Population Coding of Dynamic Sounds by Auditory Cortex.” Journal of Neurophysiology 126, no. 1 (July 1, 2021): 148–169. [CrossRef]
- Falcone, R., Weintraub, D. B., Setogawa, T., Wittig, J. H. Jr, Chen, G., and Richmond, B. J. “Temporal Coding of Reward Value in Monkey Ventral Striatal Tonically Active Neurons.” Journal of Neuroscience 39, no. 38 (September 18, 2019): 7539–7550. [CrossRef]
- Fox, K., and M. Stryker. “Integrating Hebbian and Homeostatic Plasticity: Introduction.” Philosophical Transactions of the Royal Society B 372, no. 1715 (March 5, 2017): 20160413. [CrossRef]
- Friston, K., J. Kilner, and L. Harrison. “A Free Energy Principle for the Brain.” Journal of Physiology Paris 100, nos. 1–3 (July–September 2006): 70–87. [CrossRef]
- Geadah, V., Barello, G., Greenidge, D., Charles, A. S. and Pillow, J. W. “Sparse-Coding Variational Autoencoders.” Neural Computation 36, no. 12 (November 19, 2024): 2571–2601. [CrossRef]
- Georgopoulos, A. P., Schwartz, A. B., and Kettner, R. E. “Neuronal Population Coding of Movement Direction.” Science 233, no. 4771 (September 26, 1986): 1416–1419. [CrossRef]
- Gwilliams, L., A. Marantz, D. Poeppel, and J. R. King. “Hierarchical Dynamic Coding Coordinates Speech Comprehension in the Human Brain.” Proceedings of the National Academy of Sciences of the United States of America 122, no. 42 (October 21, 2025): e2422097122. [CrossRef]
- Hagihara, K. M., and A. Lüthi. “Bidirectional Valence Coding in Amygdala Intercalated Clusters: A Neural Substrate for the Opponent-Process Theory of Motivation.” Neuroscience Research 209 (December 2024): 28–33. [CrossRef]
- Hemmatian, B., L. R. Varshney, F. Pi, and A. K. Barbey. “The Utilitarian Brain: Moving beyond the Free Energy Principle.” Cortex 170 (January 2024): 69–79. [CrossRef]
- Herzog, L. E., L. M. Pascual, S. J. Scott, E. R. Mathieson, D. B. Katz, and S. P. Jadhav. “Interaction of Taste and Place Coding in the Hippocampus.” Journal of Neuroscience 39, no. 16 (April 17, 2019): 3057–3069. [CrossRef]
- Hong, S., M. Negrello, M. Junker, A. Smilgin, P. Thier, and E. De Schutter. “Multiplexed Coding by Cerebellar Purkinje Neurons.” eLife 5 (July 26, 2016): e13810. [CrossRef]
- Hong, S., S. Ratté, S. A. Prescott, and E. De Schutter. “Single Neuron Firing Properties Impact Correlation-Based Population Coding.” Journal of Neuroscience 32, no. 4 (January 25, 2012): 1413–1428. [CrossRef]
- Hovhannisyan, H., A. Rodríguez, E. Saus, M. Vaneechoutte, and T. Gabaldón. “Multiplexed Target Enrichment of Coding and Non-Coding Transcriptomes Enables Studying Candida spp. Infections from Human Derived Samples.” Frontiers in Cellular and Infection Microbiology 13 (January 24, 2023): 1093178. [CrossRef]
- Huetz, C., Souffi, S., Adenis, V. and Edeline, J. M. “Neural Code: Another Breach in the Wall?” Behavioral and Brain Sciences 42 (November 28, 2019): e232. [CrossRef]
- Jazayeri, M. and Afraz, A. “Navigating the Neural Space in Search of the Neural Code.” Neuron 93, no. 5 (March 8, 2017): 1003–1014. [CrossRef]
- Jeffery, K. J. “How Environmental Movement Constraints Shape the Neural Code for Space.” Cognitive Processing 22, suppl. 1 (September 2021): 97–104. [CrossRef]
- Joiret, M., Leclercq, M., Lambrechts, G., Rapino, F., Close, P., Louppe, G. and Geris, L. “Cracking the Genetic Code with Neural Networks.” Frontiers in Artificial Intelligence 6 (April 6, 2023): 1128153. [CrossRef]
- Ke, J. C., X. Chen, W. Tang, M. Z. Chen, L. Zhang, L. Wang, J. Y. Dai, J. Yang, J. W. Zhang, L. Wu, Q. Cheng, S. Jin, and T. J. Cui. “Space-Frequency-Polarization-Division Multiplexed Wireless Communication System Using Anisotropic Space-Time-Coding Digital Metasurface.” National Science Review 9, no. 11 (October 18, 2022): nwac225. [CrossRef]
- Kim, Y., A. Kahana, R. Yin, Y. Li, P. Stinis, G. E. Karniadakis, and P. Panda. “Rethinking Skip Connections in Spiking Neural Networks with Time-To-First-Spike Coding.” Frontiers in Neuroscience 18 (February 14, 2024): 1346805. [CrossRef]
- Koyama, S., and Kostal, L. “The Effect of Interspike Interval Statistics on the Information Gain under the Rate Coding Hypothesis.” Mathematical Biosciences and Engineering 11, no. 1 (February 2014): 63–80. [CrossRef]
- Lange, R. D., S. Shivkumar, A. Chattoraj, and R. M. Haefner. “Bayesian Encoding and Decoding as Distinct Perspectives on Neural Coding.” Nature Neuroscience 26, no. 12 (November 23, 2023): 2063–2072. [CrossRef]
- LeMessurier, A. M., and Feldman, D. E. “Plasticity of Population Coding in Primary Sensory Cortex.” Current Opinion in Neurobiology 53 (December 2018): 50–56. [CrossRef]
- Levitan, D., Lin, J. Y., Wachutka, J., Mukherjee, N., Nelson, S. B., and Katz, D. B. “Single and Population Coding of Taste in the Gustatory Cortex of Awake Mice.” Journal of Neurophysiology 122, no. 4 (October 1, 2019): 1342–1356. [CrossRef]
- Li, M., Xie, K., Kuang, H., Liu, J., Wang, D., Fox, G. E., Shi, Z., Chen, L., Zhao, F., Mao, Y., and Tsien, J. Z. “Neural Coding of Cell Assemblies via Spike-Timing Self-Information.” Cerebral Cortex 28, no. 7 (July 1, 2018): 2563–2576. [CrossRef]
- Li, F., D. Li, C. Wang, G. Liu, R. Wang, H. Ren, Y. Tang, Y. Wang, Y. Chen, K. Liang, Q. Huang, M. Sawan, M. Qiu, H. Wang, and B. Zhu. “An Artificial Visual Neuron with Multiplexed Rate and Time-to-First-Spike Coding.” Nature Communications 15, no. 1 (May 1, 2024): 3689. [CrossRef]
- Liu, S., V. C. H. Leung, and P. L. Dragotti. “First-Spike Coding Promotes Accurate and Efficient Spiking Neural Networks for Discrete Events with Rich Temporal Structures.” Frontiers in Neuroscience 17 (October 2, 2023): 1266003. [CrossRef]
- Liu, J., Wang, Y., and Zhao, H. “Calculating Orthologous Protein-Coding Sequence Set Probability Using the Poisson Process.” Journal of Computational Biology 28, no. 10 (October 2021): 961–974. [CrossRef]
- Lu, W. “Bayesian Brain Computing and the Free-Energy Principle: An Interview with Karl Friston.” National Science Review 11, no. 5 (January 17, 2024): nwae025. [CrossRef]
- Madar, A. D., Ewell, L. A., and Jones, M. V. “Temporal Pattern Separation in Hippocampal Neurons Through Multiplexed Neural Codes.” PLoS Computational Biology 15, no. 4 (April 22, 2019): e1006932. [CrossRef]
- Mallory, C. S., and L. M. Giocomo. “Heterogeneity in Hippocampal Place Coding.” Current Opinion in Neurobiology 49 (April 2018): 158–167. [CrossRef]
- Millidge, B., M. Tang, M. Osanlouy, N. S. Harper, and R. Bogacz. “Predictive Coding Networks for Temporal Prediction.” PLoS Computational Biology 20, no. 4 (April 1, 2024): e1011183. [CrossRef]
- Montijn, J. S., M. Vinck, and C. M. Pennartz. “Population Coding in Mouse Visual Cortex: Response Reliability and Dissociability of Stimulus Tuning and Noise Correlation.” Frontiers in Computational Neuroscience 8 (June 2, 2014): 58. [CrossRef]
- Oswald, A. M., Doiron, B., and Maler, L. “Interval Coding. I. Burst Interspike Intervals as Indicators of Stimulus Intensity.” Journal of Neurophysiology 97, no. 4 (April 2007): 2731–2743. [CrossRef]
- Pacheco Estefan, D., Zucca, R., Arsiwalla, X., Principe, A., Zhang, H., Rocamora, R., Axmacher, N., and Verschure, P. F. M. J. “Volitional Learning Promotes Theta Phase Coding in the Human Hippocampus.” Proceedings of the National Academy of Sciences of the United States of America 118, no. 10 (March 9, 2021): e2021238118. [CrossRef]
- Person, A. L., and Raman, I. M. “Synchrony and Neural Coding in Cerebellar Circuits.” Frontiers in Neural Circuits 6 (December 11, 2012): 97. [CrossRef]
- Pezzulo, G., T. Parr, and K. Friston. “The Evolution of Brain Architectures for Predictive Coding and Active Inference.” Philosophical Transactions of the Royal Society B: Biological Sciences 377, no. 1844 (February 14, 2022): 20200531. [CrossRef]
- Rezaei, M. R., Saadati Fard, R., Popovic, M. R., Prescott, S. A., and Lankarany, M. “Synchrony-Division Neural Multiplexing: An Encoding Model.” Entropy 25, no. 4 (March 30, 2023): 589. [CrossRef]
- Rhodes, G., S. Pond, L. Jeffery, C. P. Benton, A. L. Skinner, and N. Burton. “Aftereffects Support Opponent Coding of Expression.” Journal of Experimental Psychology: Human Perception and Performance 43, no. 3 (March 2017): 619–628. [CrossRef]
- Runyan, C. A., Piasini, E., Panzeri, S., and Harvey, C. D. “Distinct Timescales of Population Coding across Cortex.” Nature 548, no. 7665 (August 3, 2017): 92–96. [CrossRef]
- Sakemi, Y., K. Yamamoto, T. Hosomi, and K. Aihara. “Sparse-Firing Regularization Methods for Spiking Neural Networks with Time-to-First-Spike Coding.” Scientific Reports 13, no. 1 (December 21, 2023): 22897. [CrossRef]
- Satuvuori, E., and Kreuz, T. “Which Spike Train Distance Is Most Suitable for Distinguishing Rate and Temporal Coding?” Journal of Neuroscience Methods 299 (April 1, 2018): 22–33. [CrossRef]
- Seenivasan, P., and Narayanan, R. “Efficient Phase Coding in Hippocampal Place Cells.” Physical Review Research 2, no. 3 (September 11, 2020): 033393. [CrossRef]
- Shi, Y., Bi, D., Hesse, J. K., Lanfranchi, F. F., Chen, S. and Tsao, D. Y. “Rapid, Concerted Switching of the Neural Code in Inferotemporal Cortex.” bioRxiv [Preprint] (December 13, 2023): 2023.12.06.570341. [CrossRef]
- Sokoloski, S., Aschner, A. and Coen-Cagli, R. “Modelling the Neural Code in Large Populations of Correlated Neurons.” eLife 10 (October 5, 2021): e64615. [CrossRef]
- Stringer, C., Michaelos, M., Tsyboulski, D., Lindo, S. E., and Pachitariu, M. “High-Precision Coding in Visual Cortex.” Cell 184, no. 10 (May 13, 2021): 2767–2778.e15. [CrossRef]
- Taniguchi, T. “Collective Predictive Coding Hypothesis: Symbol Emergence as Decentralized Bayesian Inference.” Frontiers in Robotics and AI 11 (July 23, 2024): 1353870. [CrossRef]
- Tomar, R. “Review: Methods of Firing Rate Estimation.” Biosystems 183 (September 2019): 103980. [CrossRef]
- Tschantz, A., B. Millidge, A. K. Seth, and C. L. Buckley. “Hybrid Predictive Coding: Inferring, Fast and Slow.” PLoS Computational Biology 19, no. 8 (August 2, 2023): e1011280. [CrossRef]
- Vallortigara, G., and G. Vitiello. “Brain Asymmetry as Minimization of Free Energy: A Theoretical Model.” Royal Society Open Science 11, no. 7 (July 31, 2024): 240465. [CrossRef]
- Yuste, R. “Breaking the Neural Code of a Cnidarian: Learning Principles of Neuroscience from the ‘Vulgar’ Hydra.” Current Opinion in Neurobiology 86 (June 2024): 102869. [CrossRef]
- Zeng, C., S. Zhao, B. Chen, A. Zeng, and S. Li. “Feature-Correlation-Aware History-Preserving-Sparse-Coding Framework for Automatic Vertebra Recognition.” Computers in Biology and Medicine 160 (June 2023): 106977. [CrossRef]
- Zhou, Y. Q., V. Puliyadi, X. Chen, J. L. Lee, L. Y. Zhang, and J. J. Knierim. “Vector Coding and Place Coding in Hippocampus Share a Common Directional Signal.” Nature Communications 15, no. 1 (December 5, 2024): 10630. [CrossRef]
- Zhu, H., He, F., Zolotavin, P., Patel, S., Tolias, A. S., Luan, L., and Xie, C. “Temporal Coding Carries More Stable Cortical Visual Representations Than Firing Rate Over Time.” Nature Communications 16, no. 1 (August 4, 2025): 7162. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



