
Submitted:
05 November 2025
Posted:
07 November 2025
You are already at the latest version
Abstract
In the continuous quest to enhance the efficiency and sustainability of flight, the natural world offers a plethora of strategies and adaptations that can be harnessed in aviation technology. This review paper explores the multifaceted approaches of energy harvesting and drag reduction observed in nature, emphasizing their potential applications in modern aircraft and drone design. It delves into the study of micro and macro structures in various species, such as the drag-reducing micro-structures of riblets on bird feathers. The paper further investigates the broader morphological adaptations in birds and insects, including topics such as beak shape, coloration, flight configurations, materials, molting, and airfoil design for their contributions to aerodynamic proficiency. In addition, this review highlights various energy harvesting techniques observed in nature, such as soaring and ground effect exploitation, and their potential integration into aircraft design for improved endurance. Through a comprehensive review of these natural phenomena, this work aims to provide valuable insights for the development of innovative, eco-friendly aviation technologies, contributing to the global effort to reduce the environmental impact of air travel while improving the viability of drones in the nano to micro range.
Keywords:
drag reduction
; energy harvesting
; flight efficiency
; bioinspiration
; biomimicry
; avian
; flight modes
1. Introduction
The quest for sustainable and efficient flight has led to a paradigm shift in the aviation industry, heavily influenced by the critical need to address environmental concerns and the ever-increasing fuel costs. This shift is not just a response to economic pressures but also a reflection of a broader societal commitment to reducing the carbon footprint of human activities. It is no secret the consumption of fossil fuels has significantly expanded in alignment with global economic growth, particularly in the realm of aviation. Year over year, there has been an increase in fuel and energy consumption in the aviation sector, due to increased air travel, shipping, and the "shrinkage" of the world, leading to an increase in greenhouse gas emissions [1]. In recent years, the impending energy crisis and energy-related pollution have quickly become increasingly social and environmental concerns.
Recent advances in the field have shown a promising direction toward integrating nature-inspired technologies for enhancing aerodynamic efficiency and energy management in aircraft. This approach, which harnesses the ingenuity of biological evolution, is not just an imitation of nature but an intelligent adaptation of its principles to modern engineering problems. Many studies have discussed the importance of drag reduction technologies, with numerous approaches presented in modern research. Several of these studies focus especially on skin friction drag and have illustrated its feasibility and importance for reducing energy consumption in many industries. Additionally, the combination of energy harvesting techniques observed in nature serves an equally important and interesting role. The combination of these bio-inspired concepts provides an ideal future for aviation, maximizing the aerodynamic efficiency while collecting energy from the surrounding environment [2,3,4,5,6,7,8].
This intersection of biology, aerodynamics, and material science opens a novel avenue for innovative solutions in aviation, as well as translation to other industries. The exploration of nature’s own mechanisms, evolved over millions of years, offers a treasure trove of insights. Bio-inspired design, a field at the crux of interdisciplinary research, harnesses these insights to drive technological advancements. By mimicking nature, engineers and scientists can develop more efficient, sustainable, and potentially transformative technologies for the aviation sector.
In the vast tapestry of nature, numerous creatures have evolved sophisticated micro and macro structures, alongside unique behavioral techniques, to enhance their survival in diverse and often harsh environments. At the microstructural level, a prominent example is the dermal denticles or micro-scales covering the skin of sharks. These structures have been extensively studied for their hydrodynamic efficiency, specifically their ability to significantly reduce drag, thereby enhancing the shark’s predatory efficiency [9,10,11,12,13].In parallel, avian riblets, tiny, streamlined protrusions on the surface of bird feathers, have been the subject of considerable study due to their similar drag-reducing properties. Works by Bechert et al., Chen et al., and Sharma et al. have elucidated how these structures can be mimicked to enhance aerodynamic performance in aviation [2,14,15,16,17,18,19].
On the macro scale, the morphological adaptations of birds, such as the aerodynamic efficiency of their beak shapes, body coloration, and wing geometries, offer a rich source of inspiration for aeronautical design. These adaptations, honed through millions of years of evolution, have been meticulously studied to understand their role in flight efficiency and energy conservation. The diversity of these natural designs presents a vast repository of ideas that can be translated into technological advancements in aircraft design [2,8,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125]. Moreover, nature’s repertoire includes a variety of energy harvesting techniques that are equally remarkable. Soaring birds, for instance, demonstrate an innate mastery of using thermal updrafts and the ground effect to conserve energy while covering vast distances. These natural strategies provide a blueprint for developing energy-efficient flight mechanisms, especially pertinent in the context of unmanned aerial vehicles (UAVs) and other aviation technologies. Detailed studies in these areas, encompassing both the micro and macro perspectives of nature’s designs, are not just of academic interest but hold substantial potential for practical applications in sustainable aviation technologies [46,98,102,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177].
In addition to the uptick in traditional aviation travel, the UAV or "drone" industry has also been rapidly expanding with growing needs for drones with diverse capabilities both in civilian and military applications ranging in size from fractions of a gram to thousands of kilograms [178]. With such a diverse range of sizes and configurations, there are hundreds if not thousands of application spaces for UAVs ranging from food delivery to space exploration and everything in between. With this increase in attention and varied set of application spaces, there has been a rising interest in avian flight as they operate in a similar flow regime. The ability to study and seek viable efficiency improvements from natural flyers that can be implemented in a range of applications is a significant opportunity. Although avian and other creatures in nature provide many opportunities to study potential improvement mechanisms, these aspects can often be difficult to study due to the complexity of their structures, control, agility, and rapid maneuverability. The challenge of unsteady fluid motion and unpredictability in conditions compounded with the different phases and styles of flapping motion used to adapt to these conditions is another layer of complexity in studying natural flyers [8].
Figure 1.
A flow chart showing the various considerations for energy saving mechanisms when designing an aircraft. Each topic is discussed in detail throughout this work.
Figure 1.
A flow chart showing the various considerations for energy saving mechanisms when designing an aircraft. Each topic is discussed in detail throughout this work.
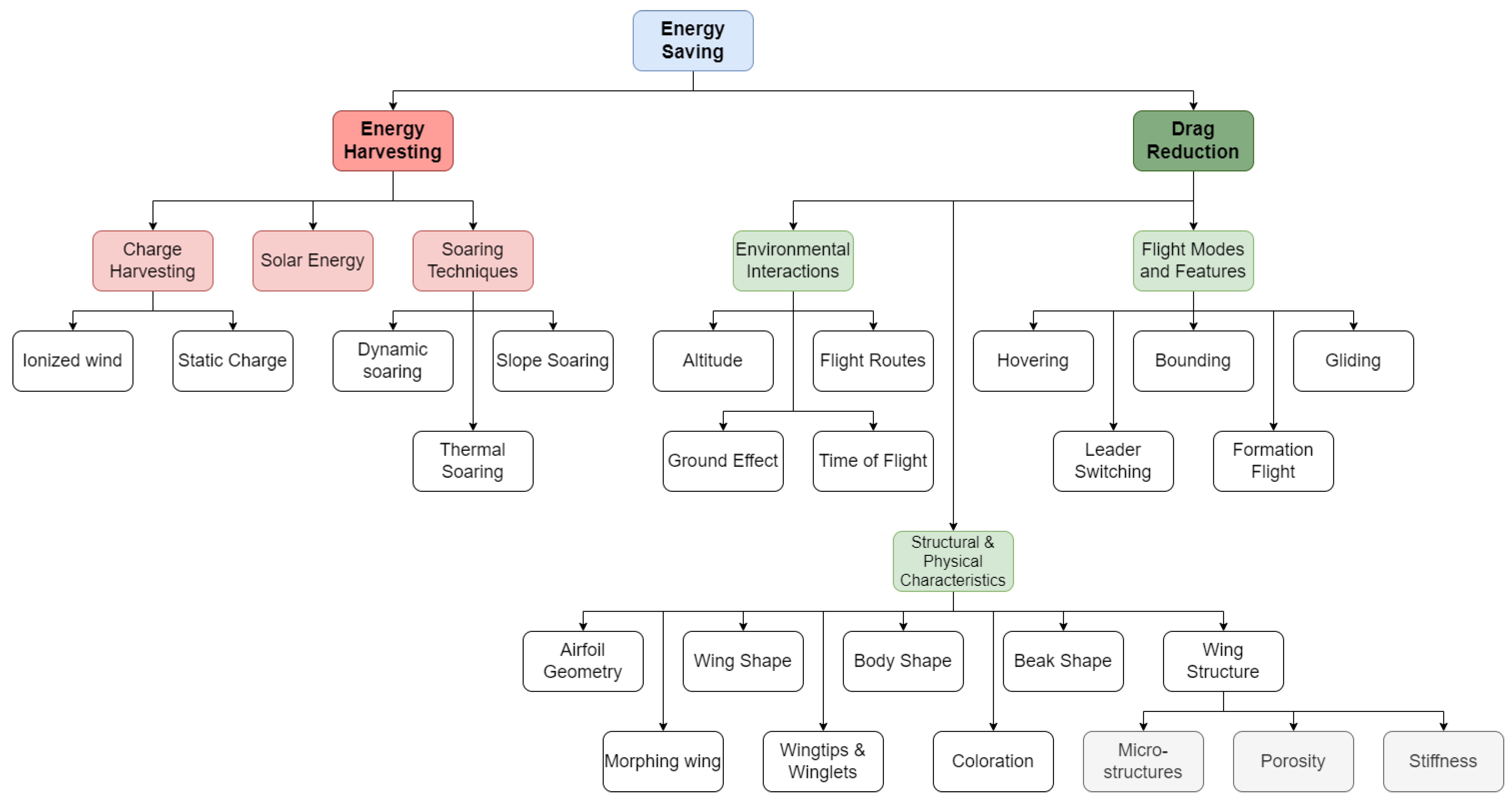
Many of the capabilities of interest are particularly of noteworthy and applicable to small UAVs in the mini, micro, and nano size ranges as they fly in similar flow regimes and compete with the same obstacles and challenges. This flight environment includes complex interactions with both natural and man-made obstacles, significant traffic, as well as significantly varying operational conditions, resulting in turbulence length scales on the order of meters [3]. The performance of vehicles on this scale is highly limited by the onboard energy or fuel and typically lacks endurance and range, greatly reducing the utility and usability. The addition of mechanisms for improving efficiency and harvesting locally available energy sources provides huge benefits to vehicles on this scale and has the potential to vastly improve their utility and expand usefulness. The use of techniques for improving the aerodynamic or flight efficiency can serve to improve the endurance of current and future UAV systems, while the development and implementation of energy harvesting systems can work in tandem to greatly extend the range and operational capacity. Many instances allow natural flyers to take advantage of naturally occurring updrafts such as thermals and spatial gradients, providing phenomena such as wind shears, or beneficial gusts.
In the context of this review, various energy-saving techniques for aviation are discussed, which, while primarily aimed at enhancing efficiency and reducing the energy expenditure of these systems, also inherently contribute to improvements in propulsion. These techniques, such as optimizing wingbeat frequency and adjusting wing shapes to minimize drag while maximizing lift, may directly or indirectly enhance propulsion by allowing for more effective use of energy and better control of air flows around the wings. However, the main focus of the review remains on energy efficiency rather than propulsion enhancement. Therefore, while the linkage between these areas is acknowledged and forms a subtle undercurrent throughout the discussions, it is not extensively elaborated upon, given that the direct implications for propulsion are not the central theme of the paper. This review focuses on the various drags encountered by birds and insects for the purpose of showing how to transfer this information to the design of more efficient low-speed air vehicles. Early pioneers like Leonardo da Vinci and the Wright brothers drew inspiration from birds and bats to design flying machines. However, as aviation technology advanced, the focus shifted towards engineering principles rather than biological observation. Modern aircraft have separated the key functions of lift generation, propulsion, and control. In contrast to conventional aircraft, birds and insects have evolved very efficient fully integrated lift, propulsion, and control systems through morphing bodies and flapping wings that realistically make it impossible to separate lift generation from thrust generation and therefore, to truly set aside the thrust generation discussion. With this in mind, the choice to set aside propulsion for this review ensures a concentrated exploration of energy savings without diluting the focus with the additional breadth of propulsion-focused technology which would require a lengthy addition. It should be noted that propulsion cannot be discounted when discussing many of the drag reduction techniques used during flapping flight, however, for the purposes of this review we will refrain from detailing their involvement.
1.1. Scaling Effects
Scaling principles are foundational in the study of aviation, and are of particular note when discussing flapping wing vehicles, where understanding the interaction between physical dimensions and aerodynamic forces is crucial. These principles illuminate how changes in size, from tiny insects to larger birds, drones, and commercial aircraft, affect flight capabilities and energy efficiency.
The development of flapping wing air vehicles, or ornithopters, has a long history that blends engineering with bio-inspired design. Early pioneers like Leonardo da Vinci conceptualized human-powered flapping wing machines, but practical implementations faced significant challenges. Modern advancements have revived interest in this field, led by researchers such as DeLaurier [179] and Bannasch[180]. DeLaurier’s work at the University of Toronto, as documented in [179] demonstrates the aerodynamic benefits of flapping wings through projects like the "Ornithopter No.1," which achieved manned flight. This work highlights the potential of flapping wings to improve lift-to-drag ratios and energy efficiency, essential for both micro and macro air vehicle applications. Rudolf Bannasch, in [180] further explored the translation of natural flight mechanisms into innovative designs. His research provides insights into how the flight adaptations of birds can inform more efficient aircraft technologies. These advancements suggest promising future applications for ornithopters, including enhanced maneuverability and energy efficiency in small unmanned aerial vehicles (UAVs).
Moreover, Kato et al. in [181] provided an in-depth exploration of the biological and physical principles underlying the locomotion of animals in water and air. This comprehensive work covers various aspects, from the hydrodynamics of fish swimming to the aerodynamics of insect and bird flight. It delves into the fluid dynamics, biomechanical properties, and evolutionary adaptations that enable efficient and effective movement in these mediums. The flapping flight mechanisms of insects and birds have informed the design of micro air vehicles (MAVs) with improved aerodynamic performance.
For flapping wing vehicles, scaling informs critical design decisions, influencing wing shape, flapping frequency, and overall vehicle architecture to optimize performance. By applying insights from biological flyers, engineers can design flapping wing vehicles that better navigate the complexities of aerodynamic forces at varying scales, improving maneuverability, stability, and energy use. This is particularly vital as advancements in micro air vehicles and biomimetic robots continue to push the boundaries of what is aerodynamically possible at smaller scales [182,183].
Shyy et al. delve into the topic of scaling and its pivotal role in understanding the aerodynamic properties of flyers operating at low Reynolds numbers, such as small birds, bats, and micro air vehicles in their book "Aerodynamics of Low Reynolds Number Flyers". This discussion is essential for determining how variables like size, shape, and flight dynamics influence aerodynamic forces and the resulting flight performance [182,184].
The authors emphasize the concept of geometric and kinematic scaling, which facilitates the comparison of diverse flyers through dimensionless parameters. The primary focus here is the Reynolds number, which serves as a fundamental measure relating viscous forces to inertial forces in fluid flow dynamics. By exploring the implications of different Reynolds numbers, the authors elucidate how these values impact aerodynamic performance, emphasizing the distinct challenges faced by small-scale flyers due to higher viscous forces.
The discussion extends to specific aerodynamic parameters, starting with wing area and wingspan. These are critical in determining the distribution and magnitude of lift and drag forces acting on a flyer. Shyy et al. illustrate how larger wingspans and optimized wing areas can enhance lift generation, thereby improving flight efficiency. The treatment of wing loading is particularly insightful. It is defined as the weight supported per unit wing area and is a crucial determinant of the flyer’s ability to maintain lift and maneuverability during flight. High wing loading indicates a greater weight relative to wing area, necessitating higher speeds to maintain lift, which the authors link to flight dynamics and energy efficiency. Considering this, the wing loading expressed in terms of a constant and the cube root of the weight results in two distinct recommended constants of 53 and 30.6 for aircraft and birds respectively [184,185]. Liu presents that both birds and aircraft follow the 1/3 power law when observing wingspans with different leading coefficients [185]. Another study discussed by Shyy et al. presents greater variation in the wing area when compared to the wingspan between groups of animals [184].
Aspect ratio, the ratio of wingspan to mean wing chord, is another focal point, highlighted for its impact on flight efficiency. Higher aspect ratios are shown to correlate with reduced drag and improved lift, characteristics beneficial for endurance and gliding efficiency. The authors use comparative analyses between different flyers to illustrate how aspect ratios adapt across species and vehicle designs to meet specific aerodynamic requirements.
Finally, the section addresses wing-beat frequency, a dynamic aspect of flapping flight. This parameter influences the aerodynamic power output necessary for maintaining flight. Shyy et al. detail how wing-beat frequency varies inversely with flyer size and directly impacts the mechanical and energetic aspects of flight. Smaller flyers, with their higher wing-beat frequencies, face different aerodynamic challenges compared to larger ones, influencing design considerations for MAVs and similar applications [184].
The article also discusses the size limitations of flapping wing MAVs based on the scaling laws. They point out that as the size of the flyer reduces, certain aerodynamic and structural limitations become pronounced. Smaller flyers, such as insects or small birds, need to flap their wings faster to remain airborne, which is a direct consequence of scaling laws affecting flapping frequency and power requirements. This fast flapping is limited by the physical capabilities of the flyer’s muscles and wing structures, which can only sustain a certain maximum frequency before failure risks occur. The natural frequency for these fliers is proportional to the square root of the ratio between gravitational acceleration and the flyer’s leg length [184]. Pennycuick performed an in-depth study on the wingbeat frequency and suggested an analytical expression to determine this frequency based on the mass, gravity, wingspan, wing area, and fluid density of the flyer and environment [186].
It was mentioned that the power required for flight and the structural strength of the muscles and bones determine the upper and lower limits of the flapping frequency, which in turn influences the overall feasible size of flapping flyers. Large flyers like some extinct pterosaurs might have reached the upper size limits for flapping flight due to these constraints, however, modern materials and mechanisms may allow us to surpass the limits of nature that have previously been thought rigid. Large flyers of the past likely could not flap fast enough to generate the necessary lift due to their large size, whereas smaller birds benefit from being able to flap faster but are also limited on the lower size end by muscle power and recovery rates.
This discussion highlights the intricate balance between body size, muscle power, and wing structure in determining the viability and performance of flapping wing MAVs, emphasizing the critical role of scaling laws in their design and functionality. These factors are important to consider when designing a flapping-wing UAV, but this review encompasses many different energy-saving techniques, and many do not require the use of flapping-wing systems. With this in mind, the combinatorial effect of many of the discussed mechanisms applies to both fixed and flapping-wing drones, indicating potential in a variety of application spaces.
2. Drag Reduction in Avian
2.1. Structural and Physical Characteristics of Avian
The natural world has long been a source of inspiration for technological innovation, particularly in the field of aerospace engineering. Among the most remarkable natural flyers are birds, whose unique adaptations for flight offer invaluable insights into the design of efficient, agile, and adaptive aerial vehicles [8,182,183]. This section aims to delve into the structural and physical characteristics of avian species to understand how their unique features have been developed to maximize flight efficiency. The discussion will explore a range of topics where both natural and engineered systems will be examined to understand how these traits and capabilities contribute to drag reduction and flight efficiency. This section will begin with the relationship between wing shape and aerodynamic performance, considering the benefits and advantages of morphing wing capabilities. Following this, the focus will shift to the specific airfoil shapes found in bird wings and how these geometries contribute to lift and drag characteristics. The role of wingtips and winglets in reducing induced drag, a critical factor in aerodynamic efficiency, micro-structures found in feathers, such as riblets, and their role in reducing skin friction drag will be examined, along with the moulting cycle and the porosity of bird wings. Other material properties of feathers, including their stiffness and flexibility, will also be investigated for their influence on flight. The overall body shape, as well as beak shapes of birds, will be discussed to review aerodynamic efficiency and maneuverability influences. These elements may provide indirect clues about flight characteristics, which often reflects feeding habits and lifestyle. Finally, the role of feather coloration in thermal boundary layer influence will be reviewed to determine the state of research on color-related efficiency gains.
A brief introduction to the differences in drag types should be considered prior to delving into this chapter. Flapping wings are capable of simultaneously generating lift and thrust, which inherently leads to the formation of vortices and the resultant induced drag, often referred to as vortex drag. This drag arises as an unavoidable consequence of achieving lift and thrust through vortex generation. In conventional fixed-wing aircraft, the minimization of induced drag is well understood and is typically achieved through an elliptic spanwise lift distribution. However, the seminal work of Ludwig Prandtl also highlights an alternate condition for minimizing drag that is not as widely recognized. Prandtl’s theory indicates that to achieve minimal drag, two conflicting aerodynamic considerations must be addressed: a narrow chord is desirable for reducing friction drag, while a long span is necessary for minimizing induced drag [187,188]. These aerodynamic goals, however, conflict with the structural requirements for strength and weight reduction. Prandtl demonstrated that to achieve the optimal balance between drag minimization and structural constraints, the downwash across the wingspan should follow a parabolic distribution [189]. This nuanced approach to drag minimization extends beyond the commonly discussed methods and underscores the complexity involved in optimizing aircraft performance.
For flapping-wing aircraft, the challenge of minimizing induced drag is even more intricate. The generation of induced drag in these systems stems from the shedding of vortices from the sharp trailing edges of the wings. The initial explanation of thrust generation via wing flapping, provided by Knoller and Betz, considered only quasi-steady wing motion and did not account for vortex shedding [190,191]. It was Birnbaum, a student of Prandtl, who first accurately described the physics of flapping-wing thrust generation in the early 1920s, demonstrating that flapping foils function analogously to two-dimensional propellers and turbines [192]. This understanding was further refined by Theodorsen’s analysis in the 1930s, which modeled inviscid two-dimensional flow [193].
Today, the fluid dynamics of two-dimensional thrust generation are well understood, particularly the role of vortex shedding in creating a time-averaged jet that aligns closely with experimental observations as documented by Jones et al. [194]. It is important to note that the propulsion mechanism in birds and insects, often referred to as "jet propulsion," is a critical component of their flight dynamics. Consequently, it is essential to present examples of the more complex three-dimensional jets produced in such systems and to compare computational models with experimental data in further studies to deepen our understanding of these phenomena such as several studies further discussed in this work [15,125,195,196,197,198,199,200].
2.1.1. Wing Shape, Airfoils, and Morphing Capabilities
The aerodynamic performance of both natural and engineered flying entities is significantly influenced by the shape and morphing capabilities of their wings [182,183]. This section aims to provide a comprehensive review of the current state of research in this area, focusing on the aerodynamic benefits of wing shape and morphing capabilities.
Aerodynamics and Ecomorphology of Avian Wings
Previous pioneering works in this field studied the ecomorphology of flexible feathers and morphing bird wings. The research delves into the complexities of morphing avian wings and feathers, particularly focusing on their aerodynamics during different flight modes such as take-off and gliding. The study found that bird wings are highly adapted to various aerodynamic conditions and are capable of changing shape almost instantaneously to meet aerodynamic demands [117]. The dissertation by Oorschot emphasized that wing shape has a significant impact on aerodynamic forces, particularly during flapping at low advance ratios like take-off. Extended wings outperform swept wings by about a third in flapping flight but not in gliding flight. Specifically, swept wings reduced horizontal forces (i.e., drag) by 69% during flapping compared to extended wings [117]. Another critical aspect of avian aerodynamics discussed in the dissertation is feather flexibility. The study indicates that feather flexibility may be beneficial in terms of stability and stall avoidance. Feather deflection likely delayed the onset of stall, thereby enhancing aerodynamic performance [117].
Figure 2.
a) Views of swept and extended wings from a sharp-shinned hawk (Accipiter striatus).– Birds are capable of morphing their wings into a swept and extended configuration, resulting in reduced area, increased leading edge angle, and reduction of wing-tip slots. b) Average ratios of the vertical to horizontal force coefficients (CV:CH) as a function of AOA of the wing for all 13 species observed with the shaded regions representing one standard deviation [117].
Figure 2.
a) Views of swept and extended wings from a sharp-shinned hawk (Accipiter striatus).– Birds are capable of morphing their wings into a swept and extended configuration, resulting in reduced area, increased leading edge angle, and reduction of wing-tip slots. b) Average ratios of the vertical to horizontal force coefficients (CV:CH) as a function of AOA of the wing for all 13 species observed with the shaded regions representing one standard deviation [117].
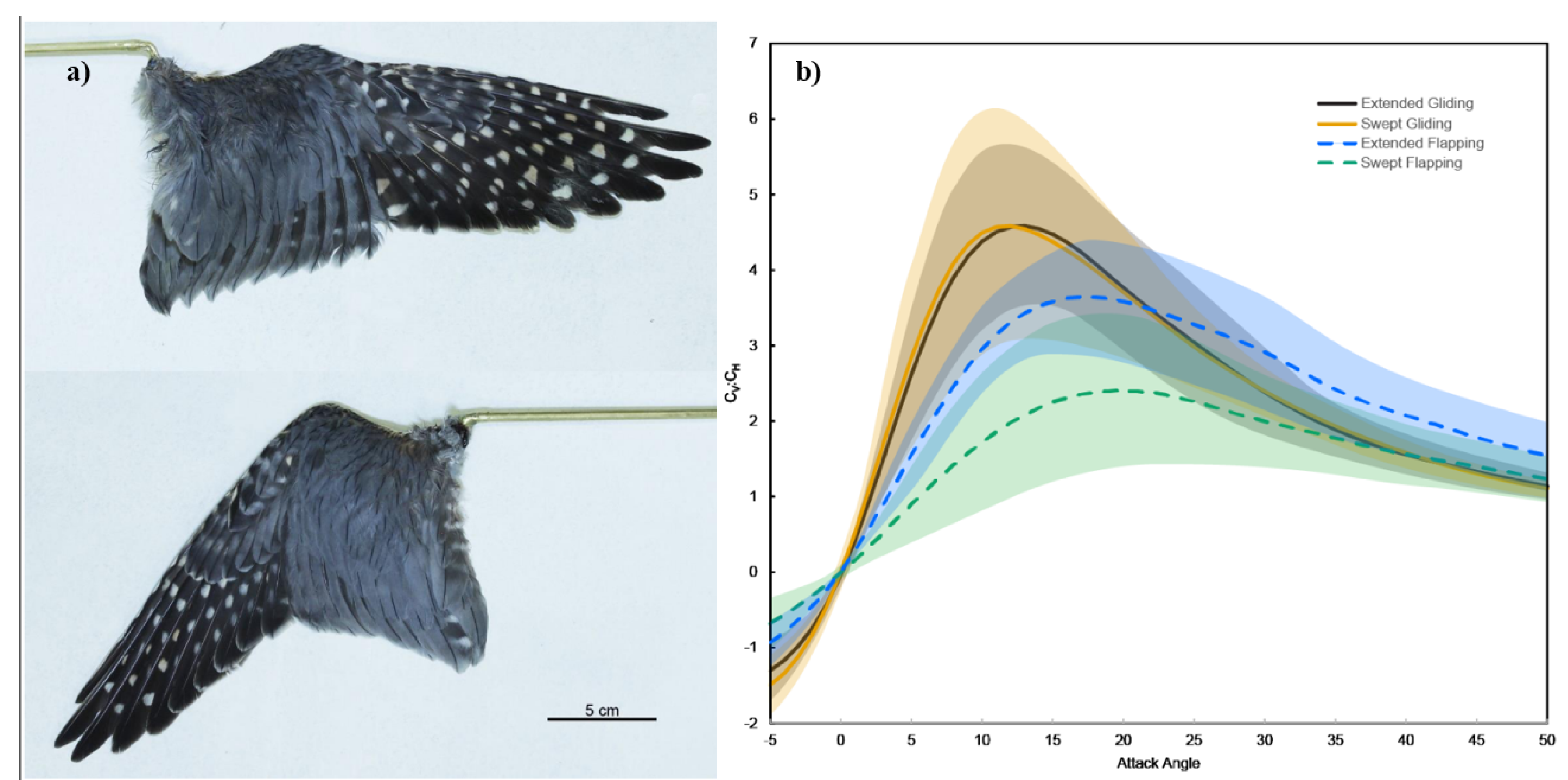
The study also explored the significance of wing tip slots and emarginate primary feathers in avian aerodynamics. These slots allow feathers to function as individual aerodynamic surfaces, providing separation for the feathers to bend, twist, and sweep independently. While it has been hypothesized that these features function like winglets on an airplane to break up the wingtip vortex and increase efficiency, the study suggests that they may serve more to increase lateral stability by redirecting aerodynamic forces [117].
Figure 3.
a) Schematic example depicting the skeleton anatomy and the feather arrangement on a typical bird wing [80]. b) A diagram showing the relevant parameters for determining emargination index [117]. c) Distal primary feathers of four species with varying degrees of emargination: RTHA: red-tailed hawk (Buteo jamaicensis), E=3.95; GHOW: great horned owl (Bubo virginianus), E=1.61; LAGU: laughing gull (Larus atricillia), E=0; LTDU: long-tailed duck (Clangula hyemalis), E=1.01. These are representative of many species in the study [117]. All figures are reprinted with permission; please see the Appendix A.
Figure 3.
a) Schematic example depicting the skeleton anatomy and the feather arrangement on a typical bird wing [80]. b) A diagram showing the relevant parameters for determining emargination index [117]. c) Distal primary feathers of four species with varying degrees of emargination: RTHA: red-tailed hawk (Buteo jamaicensis), E=3.95; GHOW: great horned owl (Bubo virginianus), E=1.61; LAGU: laughing gull (Larus atricillia), E=0; LTDU: long-tailed duck (Clangula hyemalis), E=1.01. These are representative of many species in the study [117]. All figures are reprinted with permission; please see the Appendix A.
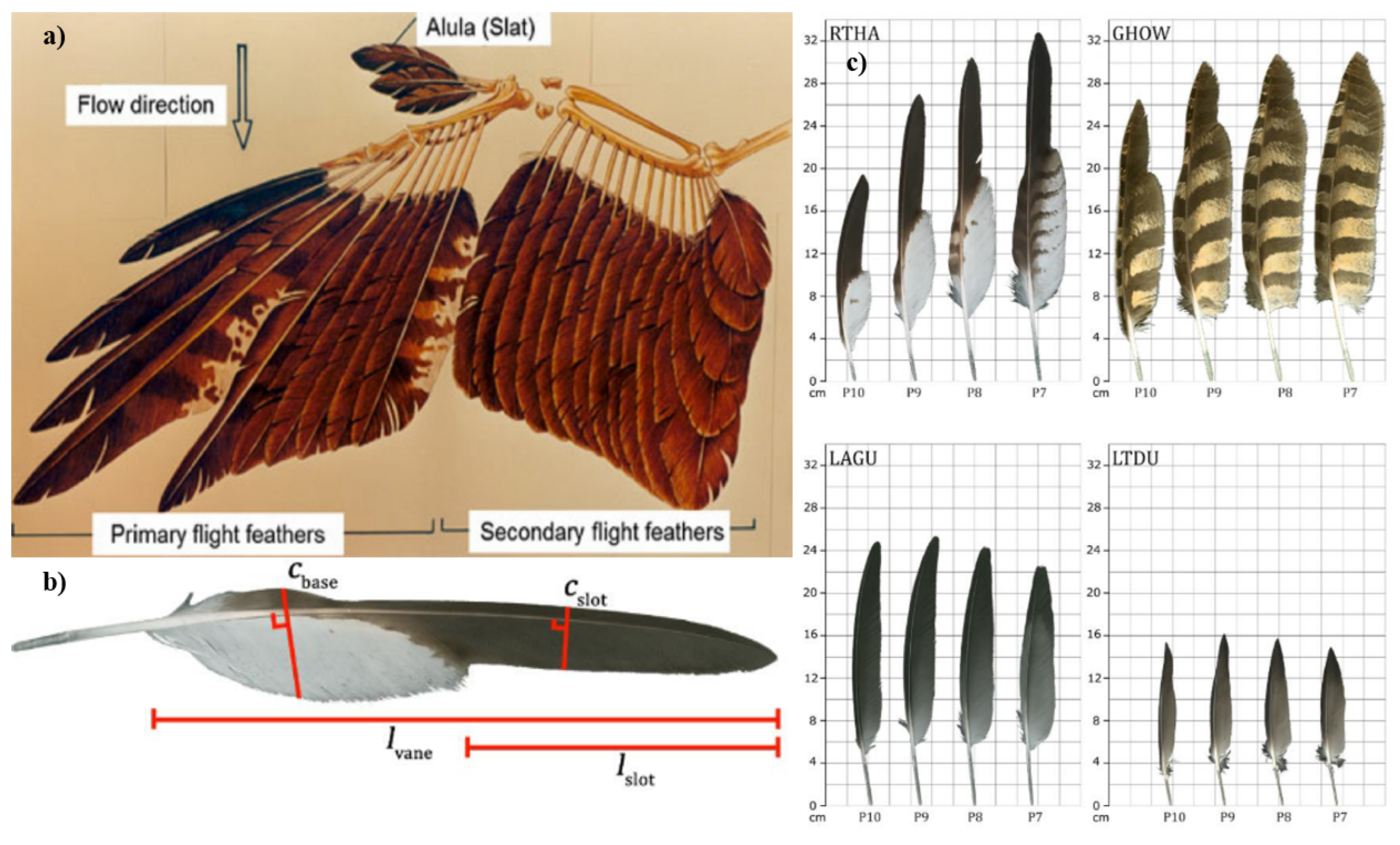
Wing folding and stroke tilting in flapping flight is another topic of interest with recent research, employing a biohybrid flapping robot modeled after the jackdaw bird to explore aerodynamic principles. One of the key findings is that folding the wing (increasing wing sweep) leads to a lower peak lift coefficient but also results in a lower drag coefficient compared to an extended wing, further corroborating the study by Oorschot [22]. This study also discussed the role of "stroke plane angle" in flight performance, indicating that varying this angle can either increase lift and reduce drag at low speeds or do the opposite at high speeds [22]. The developed biohybrid robot has advanced kinematic capabilities, allowing for a detailed study of aerodynamic forces. The paper emphasized the role of upstroke wing folding in enhancing aerodynamic efficiency, which could be beneficial for the design of flapping drones [22].
Aerodynamics of Aircraft Wings
There have been significant studies in the past focused on the aerodynamic characteristics of aircraft wings with and without bird feather-like winglets. A previous study used a rectangular wing model in a wind tunnel to compare aerodynamic characteristics. The primary focus was on how the use of bird feather-like winglets could reduce drag and increase lift. The study found a 25-30% reduction in drag coefficient and a 10-20% increase in lift coefficient at an angle of attack of 8 degrees as shown in the results tabulated in Table 1, Table 2 and Table 3 from Hossain et al. [63].
In addition to the primary findings, the study also compared its results with previous research from the Universiti Putra Malaysia, and the Georgia Institute of Technology. These tests showed that flat plate winglets set at zero degrees increased the lift curve slope by 10% for a maximum Reynolds number of 2.90×105. Another configuration led to a 15-22% increase in lift curve slope and a 15-20% decrease in drag [63,201].
The lift-to-drag ratio was found to increase with the angle of attack, reaching a maximum at 4 degrees. For a wing with a horizontal winglet at this angle, the maximum value of the lift-to-drag ratio ranged between 10.43 and 13.09, indicating better aerodynamic efficiency compared to wings without winglets [63]. Additional discussion regarding winglets and wingtips is conducted in Section 2.1.2
Application to Flapping Micro Air Vehicles (FMAVs)
Other studies have investigated the aerodynamics of eagle-inspired avian wing shapes, and how they can be applied to improve the performance of FMAVs. The study used a modified eagle wing design and compared its performance to a baseline wing in a wind tunnel. The modified eagle wing led to a 14% average improvement in lift force and up to a 21% increase in thrust force compared to the baseline wing [28]. The study attributes these improvements to the corrugated trailing edge and slotted wingtip of the modified eagle wing. These features contribute to the wing’s ability to generate higher lift and thrust forces, thereby enhancing the aerodynamic efficiency of FMAVs [28].
Investigations into avian ecomorphology have long established a correlation between migratory patterns and wing morphology, particularly the pointedness of wings. This attribute is traditionally linked to the necessity of reducing drag during sustained, powered flight, a critical factor for migratory species. Despite the prevalence of this understanding, the majority of past research has predominantly concentrated on bird species with relatively rounded wings, which exhibit distinct flight modes during foraging compared to migration. This focus has inadvertently overlooked a critical assessment of species where migratory and foraging flight modes are closely aligned.
Some recent work has challenged the conventional wisdom that migratory birds should have more pointed wings than sedentary species [8,65]. The study delves into the intricate relationship between wing shape and flight efficiency in avian species, particularly focusing on migratory and non-migratory birds. delved into the relationship between migration and wing shape within the Hirundinidae family, or swallows, known for their inherently pointed wing structures. This family presents a case in which the foraging flight closely resembles the migratory flight, providing a novel perspective in this field of study. Through a phylogenetic analysis, this research compared the wing shapes of various species pairs across eight genera of swallows, with a specific focus on their migratory behaviors [65].
Contrary to the established trend in other avian families, the findings of this study revealed a fascinating inversion of the usual correlation: migratory swallows exhibit less pointed wings compared to their sedentary counterparts. Furthermore, a linear decrease in wing pointedness was observed in Figure 4 with an increase in migratory distance. The study hypothesizes that this reversal in the migratory-swallow wing morphology could be attributed to a combination of factors. These include the adverse weather conditions prevalent in breeding grounds and the specific roosting habits of migratory swallows, which might impose conflicting selective pressures. The findings from this comparative analysis could have implications for the design of FMAVs, particularly those intended for long-duration flights. Understanding the aerodynamic trade-offs between different wing shapes could guide the design of more efficient FMAVs [65].
Advanced Wing Morphing Technologies
There have also been recent works focused on the aerodynamic advantages of flexible flaps inspired by avian feathers, particularly for fixed-wing drones. These studies employ force measurements and flow visualization in low-speed wind tunnels to explore the aerodynamic characteristics of these flaps. A key finding is that the stiffness of the flaps significantly impacts aerodynamic performance, particularly in reducing force fluctuations when the deformation synchronizes with strong vortex generation [89,202,203,204].
The study by Murayama found that the stiffness of these flaps significantly impacts aerodynamic performance, particularly in reducing force fluctuations when the deformation synchronizes with strong vortex generation. The paper suggests that attaching flexible flaps to the upper surface of a wing can be an effective biomimetic design strategy for enhancing aerodynamic robustness [89].
Figure 5.
a) Front and sides views of a peregrine falcon with popped-up feathers [203,205]. b) Schematic view of a flapped hinge on the suction side of the airfoil using a torsional spring [203]. c & d) Visualization of the induced effects of a flap at zero and high AOA respectively [202]. e) Mean streamwise velocity fields around a NACA0020 airfoil at an AOA of 17.5°. The top image shows a plain airfoil, and the bottom with a flap, where blue represents the shadow region, white indicates regions of negative streamwise velocity, and the color contour goes from 0 (blue) to 1.8* (red) [203]. f) Another schematic view showing the application of flexible flaps inspired by a bird wing [89]. g) View of a water tunnel flow visualization experiment, using dye to show water motion near the flap [204]. h) View of a seagull landing, showing the deflection of its wing flaps [204,206]. i) View of an experimental wing using an aluminum flap and j) shows a similar feather-inspired flap is also show allowing for “pop up” feathers [202]. All figures are reprinted with permission; please see the Appendix A.
Figure 5.
a) Front and sides views of a peregrine falcon with popped-up feathers [203,205]. b) Schematic view of a flapped hinge on the suction side of the airfoil using a torsional spring [203]. c & d) Visualization of the induced effects of a flap at zero and high AOA respectively [202]. e) Mean streamwise velocity fields around a NACA0020 airfoil at an AOA of 17.5°. The top image shows a plain airfoil, and the bottom with a flap, where blue represents the shadow region, white indicates regions of negative streamwise velocity, and the color contour goes from 0 (blue) to 1.8* (red) [203]. f) Another schematic view showing the application of flexible flaps inspired by a bird wing [89]. g) View of a water tunnel flow visualization experiment, using dye to show water motion near the flap [204]. h) View of a seagull landing, showing the deflection of its wing flaps [204,206]. i) View of an experimental wing using an aluminum flap and j) shows a similar feather-inspired flap is also show allowing for “pop up” feathers [202]. All figures are reprinted with permission; please see the Appendix A.
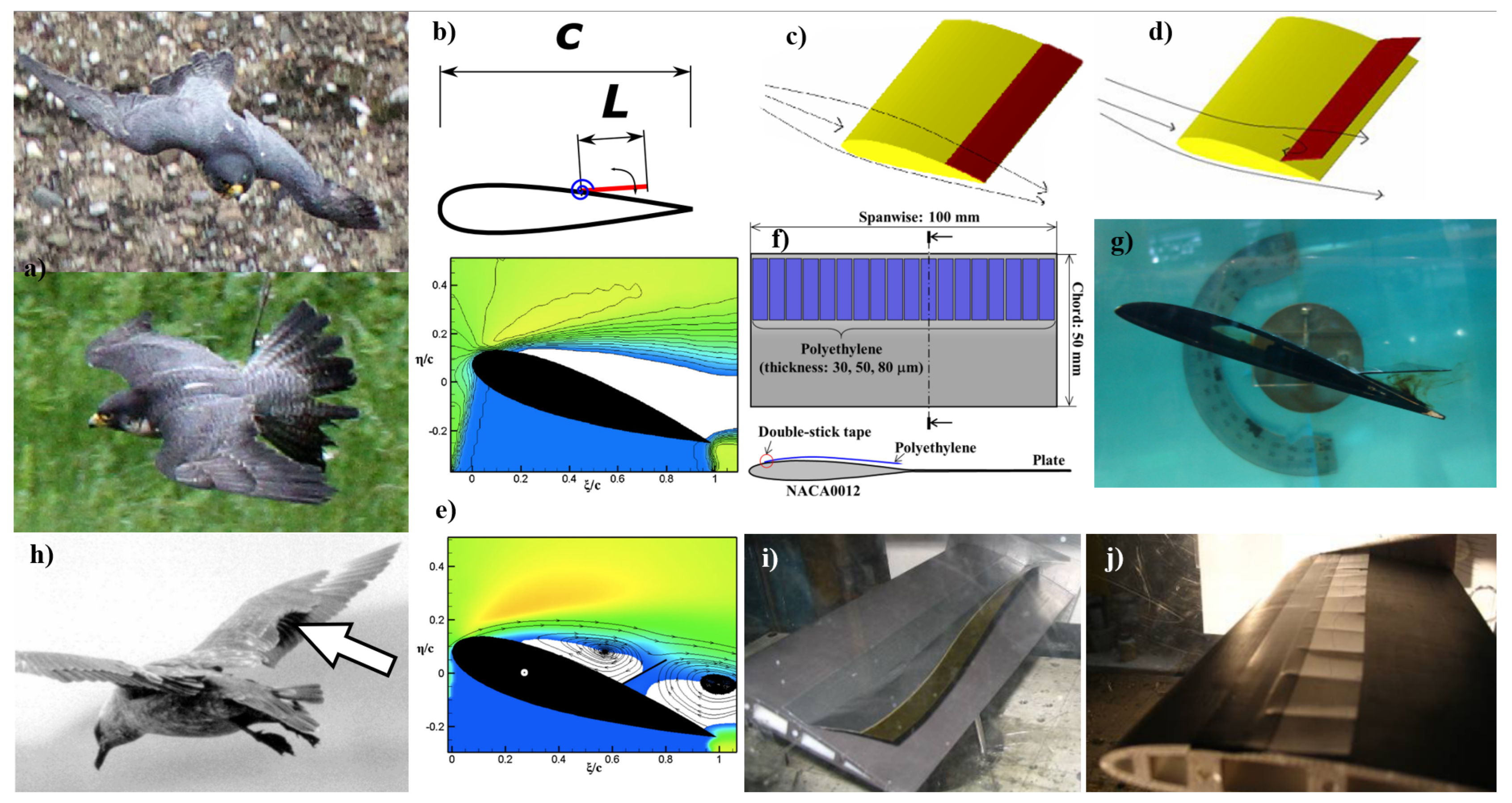
Traub and Coffman’s wind tunnel study provides additional experimental insights into the aerodynamic behavior of thin flat plate airfoils with leading and trailing edge flaps at low Reynolds numbers, ranging from 40,000 to 80,000. This regime is particularly relevant for MAVs, where the design challenges are intensified by viscous phenomena such as laminar separation bubbles. The experimental setup included techniques for streamlining visualization and surface flow analysis, ensuring high precision and repeatability. The airfoils tested included a circular arc profile and three conventional airfoils, with the performance measured against force balance and surface pressure metrics [114].
Results indicated that the flapped flat plate airfoils achieved superior lift-to-drag ratios compared to their conventional counterparts, with the highest lift-to-drag ratio corresponding to smooth on-flow onto the leading edge without separation, supporting the computation results of previous studies. This finding is pivotal for MAV design, suggesting that flaps can significantly enhance aerodynamic efficiency in these flow regimes [114].
An analytic prediction method was also developed to estimate the lift-dependent drag of these airfoils, providing a valuable tool for MAV designers. The study’s comprehensive experimental database offers a characterization of the effects of leading and trailing edge flaps, which can be directly applied to the optimization of MAV airfoils. The work also made comparisons with historical data and simulations using Xfoil, highlighting discrepancies at low Reynolds numbers and underscoring the importance of empirical data in MAV airfoil design. The increase in maximum lift-to-drag ratio with changes in Reynolds number is particularly noteworthy for MAVs operating across a range of conditions.
The use of passive mechanisms to improve the flight stability of drones is highly emphasized and hypothesizes that birds utilize flexible feathers to passively adapt to complex and changing wind environments [89,114,202,203,204].
Another point of interest is the aerodynamic advantages of wing tip slots in birds that soar over land. Tucker et al. employed a model wing in a wind tunnel, the study simulates the aerodynamic forces on bird wings. One of the key findings is that the slotted tips in bird wings serve to reduce induced drag. Specifically, the slotted tips function similarly to winglets (discussed further in Section 2.1.2) used in aircraft, spreading vorticity both horizontally and vertically, thereby reducing the kinetic energy left in the vortex sheets and consequently the induced drag [116]. Several studies have investigated this effect, comparing it to the "standard" wingtips we are generally familiar with. The conceptual aspect as well as examples of wingtip slots can be seen in Figure 6. Many studies have shown the advantages of these features, including increased agility and improved aerodynamic efficiency [28,46,80,115,116,207,208,209,210].
The review by Liu et al. further elaborates on these aerodynamic benefits, indicating that wingtip slots can also play a significant role in drag decomposition and variation in induced drag, offering more nuanced insights into their aerodynamic mechanism [80]. This study highlights the need for further exploration into the geometric and kinematic parameters of wingtip slot structure in unsteady states, emphasizing the importance of considering the flexibility of wingtips for an in-depth understanding of their aerodynamic impact.
Several studies discuss the concept of vorticity spreading as a mechanism by which the slotted tips or winglets spread the vortex sheets both horizontally and vertically. This is crucial for reducing the kinetic energy left in the vortex sheets, which in turn reduces induced drag [115,116,208]. Liu et al. (2021) also suggest placing emphasis on the study of the formation, development, and evolution of wingtip vortices on slotted wings to further understand this phenomenon [80].
Another focal point of the paper is the concept of "aspect ratio" and "tip feather configuration," which change between soaring and gliding flight. In gliding flight at higher velocities, the elbow and wrist are bent to reduce the span, thereby achieving a lower aspect ratio. This configuration also results in less exposure of the tip feather slots, affecting the induced drag [115,116]. Liu et al.’s review supports this perspective by highlighting the significance of geometric parameters like the shape and planar contour of winglets, which directly influence the wing’s aerodynamic properties [80].
Another study investigated the aerodynamic properties of avian wings as a function of unique wing shapes while considering morphing capabilities. One of the main goals of the study was to quantify these aerodynamic properties to understand shape-changing elastic structures better. Unlike typical airfoils, bird wings do not have a distinct stall point; they twist into the flow, which significantly reduces induced drag. This is facilitated by the slotted wingtips and primary feathers that are canted downward to generate lift in the thrust direction [84]. This study was inspired by birds like raptors, which change the shape of their wings to maximize their lift-to-drag ratio for different flight strategies such as soaring or diving [84].
The aerodynamic properties of bird wings from different flight style groups were investigated, finding that morphological differences manifest mainly as differences in aerodynamic drag rather than lift. The paper highlights that the aerodynamics of bird wings operate at lower, transitional Reynolds numbers, making them relevant for UAV design [84].
The role of wing shape in galliform birds like the White-tailed Ptarmigan, Sage Grouse, Wild Turkey, and California Quail has been thoroughly examined, particularly focusing on the trailing-edge notch in their wings. Findings indicate that this specialized feature enhances the maximum lift-to-drag ratio and stabilizes airflow around the wing. However, it also has the downside of reducing the maximum lift coefficient, which impacts efficiency during level flight [43].
The trailing-edge notch is particularly beneficial for birds that engage in vertical and slow flight, as it enhances performance in these flight modes at the cost of reduced efficiency in level flight. The study suggests that the size of the trailing-edge notch varies among species and is related to their ecology and behavior, and is particularly beneficial to birds that often fly when fleeing from predators and not much in between [43].
Figure 7.
Views of various galliform wings displaying the trailing edge notch characteristic: a) White-tailed Ptarmigan [43], b) California Quail [43], c) Sage Grouse [43], d) wild turkey [43], e) pheasant (Photo credit: Sadie-Michaela Harris), and f) peacock (Photo credit: Lisa Hubbard [214]). All figures are reprinted with permission; please see the Appendix A.
Figure 7.
Views of various galliform wings displaying the trailing edge notch characteristic: a) White-tailed Ptarmigan [43], b) California Quail [43], c) Sage Grouse [43], d) wild turkey [43], e) pheasant (Photo credit: Sadie-Michaela Harris), and f) peacock (Photo credit: Lisa Hubbard [214]). All figures are reprinted with permission; please see the Appendix A.

For instance, birds with short, rounded wings have deeper notches, which are more beneficial for vertical flight. The study also found an inverse relationship between the angle of attack and the coefficient of variation (CV) for lift and drag, suggesting that wings with trailing-edge notches should be more stable in flight and their performance more predictable [43].
The Influence Of Flight Style On The Aerodynamic Properties Of Avian Wings As Fixed Lifting Surfaces
A comprehensive examination of the aerodynamic properties of bird wings across various species and flight styles was conducted by Lees et al. to assess how wing morphology impacts aerodynamic performance, specifically in terms of lift and drag. One key observation is that the morphological differences between wings of birds with different flight styles are more pronounced in their effects on drag rather than lift. For instance, undulating flyers such as Corvids exhibit the lowest minimum drag coefficients. [76].
The study emphasizes that the aerodynamics of bird wings operate at lower, transitional Reynolds numbers, making them relevant for UAV design. It suggests that wing shape is principally linked to changes in aerodynamic drag, which is likely to be the primary driver during flapping flight rather than gliding [76].
The paper concludes that general wing-morphological measures based on planform cannot alone predict the gliding performance of avian wings. This conclusion comes from seeing that birds with different flight styles show different aerodynamic responses even when their wing shapes are taken into account. In other words, wing shape alone doesn’t give a full picture of a bird’s aerodynamic capabilities. [76].
Advanced computational models have been utilized to observe the properties of biomimetic wings inspired by white storks (ciconia ciconia). These models have been used to analyze the flow field around and through the wing tip cascade. Further validation of the aerodynamic performance of these bio-inspired wings comes from comparisons with empirical data on the soaring flight of free-flying birds [113].
The computational techniques implemented in this study include Reynolds-averaged Navier-Stokes (RANS) and hybrid RANS–LES (Large-Eddy Simulation) computations. These models have been applied to gain a comprehensive understanding of flow dynamics around and through the wing tip cascade. This approach offers an in-depth look at avian wing aerodynamics. The system of vortices linked to wing and tip design plays a significant role in the aerodynamic properties observed. This is especially pertinent for explaining how variations in wing shape and morphing capabilities can serve to minimize drag and optimize lift [113].
The intricate relation between avian wing shape, range of motion, and flight behavior was examined across 61 bird species by Baliga’s research group. The wing shape and range of motion were extracted and can been seen in Table 4. Employing a Bayesian framework for data analytics, traditional perspectives considering wing shape as the chief factor influencing flight behavior are challenged. Emphasis is instead placed on the wing’s range of motion as a significant determinant. The ability to dynamically alter wing shape during flight provides enhanced control over aerodynamic forces and moments. This dynamic capability is of particular importance in birds that display high-excursion flapping and bounding behaviors, as they show fewer restrictions in their range of motion. Emphasis is placed on the ability to dynamically alter wing shape as a key factor in achieving greater control over aerodynamic forces and moments. This aspect is especially crucial for species that display high-excursion flapping and bounding behaviors, as it allows for a less restricted range of motion [30]. Insights into the constraints imposed by natural selection on birds of varying sizes and flight styles offer valuable implications for engineering. Such understanding can inform the design of next-generation morphing wings, particularly in the realm of UAVs. [30].
State of Morphing Wing Research
The collective body of work on wing shape and morphing capabilities offers a comprehensive understanding of the aerodynamic properties and flight behaviors across both avian species and engineered systems. Earlier studies have established the fundamental principles of wing morphing for drag reduction and energy efficiency [8,22,28,63,65,117]. More recent research has expanded this understanding by exploring the role of specific wing features, such as wing tip slots and trailing-edge notches, in reducing induced drag and improving lift-to-drag ratios [43,84,89,116]. The importance of dynamic wing morphing and the range of motion have also been emphasized, particularly in the context of high-excursion flapping and bounding behaviors [8,30,84].
The collected findings have significant implications for the engineering and design of next-generation aircraft and unmanned aerial vehicles (UAVs). The concept of a morphing aircraft, inspired by various avian species, offers a promising avenue for future research [8,17,22,28,30,43,63,65,76,84,89,113,115,116,117,201,202,203,204,205,206,211]. The computational models employed in some studies provide a robust framework for analyzing aerodynamic properties, thereby informing the design of more efficient wing structures [113]. Furthermore, the insights into the aerodynamic benefits of specific wing features, such as flexible flaps and wing tip slots, offer practical design strategies for enhancing aerodynamic performance [89] [116]. The studies also suggest that understanding the natural limitations in birds of different sizes and flight styles can inform the design of more adaptive and efficient morphing wings for UAVs [30]. The intricate design of airfoils, particularly those inspired by avian species, has been the subject of extensive research due to its significant implications for the aerodynamics of UAVs and MAVs. This section aims to synthesize the collective findings from a variety of studies that delve into the aerodynamic efficiency of bird-like airfoils and their potential applications in aeronautical engineering.
Engineering Analysis of Avian Flight
The study of avian flight has yielded profound insights into the mechanics of wing morphology and its implications for engineering, particularly in the realm of UAVs and MAVs. The work by Liu et al. offers a detailed engineering perspective that bridges the gap between biological inspiration and aeronautical application [82].
Scaling Laws and Aerodynamic Efficiency At the heart of Liu’s study is an exploration of the scaling laws that govern bird flight. These laws are not mere curiosities but fundamental principles that dictate the feasibility of flight at various sizes and speeds. By understanding these scaling relationships, engineers can better predict the performance of UAVs and MAVs designed to mimic the flight of birds. The paper meticulously reviews the interplay between wing geometry, kinematics, and aerodynamics, providing a holistic view of the factors that contribute to efficient flight [82].
Detailed Wing Geometry Analysis Liu et al.’s research presented an in-depth analysis of the wing geometry of various birds like seagulls, mergansers, teals, and owls. Utilizing advanced laser scanning technology, the study comprehensively measured airfoil camber lines, thickness distributions, wing planforms, and twist distributions. This geometrical data is crucial in understanding the aerodynamic properties of avian wings and their potential application in UAV and MAV design [82].
Significantly, the study compared avian airfoils with high-lift, low-Reynolds-number airfoils like the S1223. This comparison highlights the aerodynamic efficiency of avian wings and offers insights into designing MAVs that can achieve similar performance. By analyzing the pressure coefficient distributions and structural similarities, the study bridged a key gap in applying biological insights to aeronautical engineering [82].
Kinematic Modeling for MAV Design Another major contribution of this work is the kinematic modeling of avian flight using a two-jointed arm model. This approach simplified the complex motion of bird wings, making it more feasible to replicate in MAV designs. The study’s kinematic data, derived from video analysis of birds in level flight, provides a foundational framework for designing MAVs that can mimic the efficient flapping motion of birds [82].
Morphing Wing Technology A focal point of the research is the aerodynamic efficiency of avian wing airfoils, examples of which can be seen in Figure 8 particularly as it pertains to the role of morphing wing technology in drag reduction. Birds, such as owls, have evolved wings that can change shape in real time, optimizing their aerodynamic profile for various flight conditions. This adaptability results in a higher lift-to-drag ratio, a coveted metric in aircraft design. The formation of a separation bubble, as observed on the owl airfoil, increases lift while simultaneously reducing drag, a feature that is highly desirable for UAVs and MAVs that require efficient lift mechanisms [73,82,101].
The owl airfoil, in particular, is highlighted for its superior aerodynamic performance. Several studies demonstrated that the owl airfoil maintains a higher lift-to-drag ratio across different kinematic modes, making it an ideal model for flapping wing aerial vehicles. The studies’ findings suggest that by incorporating the geometric and kinematic characteristics of the owl wing into UAV and MAV design, engineers could achieve significant advancements in aerodynamic efficiency [73,82,101].
Bio-Inspired Flapping Kinematics
The emulation of avian flight characteristics in aerial vehicle design has led to significant advancements in the aerodynamics of flapping wings, which have a characteristically stark contrast to gliding flight. The study by Lang et al. is particularly illuminating, focusing on the aerodynamic performance of the OWL05 airfoil in flapping flight(shown in Figure 8, modeled after the barn owl’s wing. This research is pivotal in understanding how the geometry of the airfoil contributes to its aerodynamic prowess, particularly in the context of flapping-wing MAV kinematics [73].
OWL05 Airfoil Geometry and Performance The OWL05 airfoil’s geometry is characterized by a round nose, high camber, thick leading edge, and sharp trailing edge. These features collectively facilitate a robust, near-constant drag coefficient, which is advantageous, especially at low Reynolds numbers typical of MAVs. The design of the airfoil allows for enhanced lift during the downstroke of flapping, a critical aspect of the bio-inspired kinematics derived from actual bird wing movements. This results in a higher peak lift and maintains positive lift over a wider proportion of the flapping cycle compared to other airfoils, such as the S1223 and Clark Y, [73].
The OWL05 airfoil not only demonstrates superior lift characteristics but also exhibits more energy-efficient flapping motion compared to sinusoidal motion. This efficiency is crucial for the prolonged operational capabilities of MAVs. The leading edge’s shape significantly affects the transition of the leading-edge vortex (LEV), with the sharper leading edge of other airfoils causing the LEV to roll up faster and with greater strength, which can be beneficial or detrimental depending on the specific flight requirements.
Wing Morphology’s Influence on Flapping Wing Model The study also highlights the influence of wing morphology on the flapping wing model. Different planforms manifest primarily in differences in drag rather than lift. For instance, barn owls with almost elliptical-shaped wings exhibit high flight efficiency during gliding flight due to their wing planform. This insight is crucial for the design of MAVs, where the optimization of the wing planform could lead to significant improvements in aerodynamic performance.
To summarize, the geometry of the owl-like airfoil contributes to its superior aerodynamic performance, with specific advantages in drag reduction and lift enhancement during bio-inspired flapping kinematics, as well as fixed-wing gliding flight. These findings are instrumental for the development of flapping MAVs, offering insights into the design of wings that mimic the highly efficient flight mechanisms of natural fliers.
Avian Airfoil Characterisitic
The intricate design of avian airfoils offers a wealth of knowledge for the development of efficient low Reynolds number airfoils where viscous phenomena dominate, which is particularly relevant for MAVs. The work by Rajesh Senthil Kumar et al. provided an analysis of the aerodynamic characteristics of various bird airfoils, including those of Seagulls, Mergansers, Teals, and Owls [101].
The study extends to the effects of feather roughness by superimposing the corrugation pattern of the Swift bird onto the Seagull airfoil. This modification demonstrates an improvement in the lift-to-drag ratio, suggesting that at the scale of MAVs, a corrugated surface can be beneficial. The cavities formed by the corrugation act as vortex-trapping cells, which effectively manage the flow separation and transition over the airfoil, thereby controlling the formation of laminar separation bubbles.
Geometric Characteristics and Performance Metrics The geometric characteristics of these avian airfoils—high camber, a thin aft region, and a thick leading edge—are crucial for generating aerodynamic forces in bird flight and serve as a template for MAV airfoil design. The designed airfoils feature a bulbous leading edge and a thin, feather-like aft region, aiming to promote a smooth transition from laminar to turbulent flow and manage the laminar separation bubble effectively.
The high camber of seagull and merganser airfoils parallels with high-lift, low-Reynolds-number airfoils used in modern MAVs, indicating an optimization for low Reynolds number flight conditions. The kinematics of avian wings, recovered from video recordings and expressed analytically, offer a framework for the design of flapping mechanisms in MAVs [101].
A notable point of interest discussed in these works is the unique structural adaptations of the owl wing, which are associated with silent flight, and can scarcely be detected. The studies describes the owl wing’s thin structure and small aspect ratio, which contribute to its stealthy flight capabilities. While the specific surface features that reduce noise may not be directly applicable to MAVs, the aerodynamic benefits derived from these features can inform the design of efficient and quiet MAVs [73,81,82,101].
Bird-Inspired Airfoil Design Approach
The quest for efficient UAVs and MAVs has led to the exploration of bird-like airfoil design processes for operations within a low Reynolds number regime ( typically - ). Ananda and Selig’s study provides a comprehensive approach to airfoil design, informed by the aerodynamic properties of bird wings, which are highly cambered with a thick leading edge and a thin aft region [26]. The designed airfoils feature a bulbous leading edge and a thin, feather-like aft region, aiming to promote a smooth transition from laminar to turbulent flow and manage the laminar separation bubble effectively. The performance of these airfoils is characterized by well-defined drag buckets, high lift-to-drag (Cl/Cd) performance, and predictable transition characteristics, essential for MAVs that require high wing loadings and efficient flight at high lift coefficients.
Design Methodology and Tools The design process employs multipoint inverse airfoil design methods and utilizes the PROFOIL tool, with the airfoils being analyzed using the XFOIL tool to predict their low Reynolds number flow characteristics. This methodology allows for the creation of airfoils that are optimized for MAVs operating at low Reynolds numbers, drawing heavily from avian flight characteristics. The ability to manage viscous effects such as the laminar separation bubble is a significant challenge in this flight regime. Bird wings have evolved to produce aerodynamic forces effectively at similar scales of Reynolds numbers, and the airfoils designed by Ananda and Selig aim to mimic this efficiency [26].
This work contributes a set of bird-like airfoil designs that are not only inspired by the efficiency of bird wings but are also tailored to address the unique challenges faced by MAVs at low Reynolds numbers. The insights gained from this study are useful for the advancement of MAV airfoil technology, ensuring that these vehicles can achieve high performance while navigating the complexities of aerodynamic forces at small scales.
Evolutionary Innovations in Avian Wings
The evolutionary trajectory of avian wings has led to a diverse array of adaptations, each serving to enhance the aerodynamic efficiency and flight capabilities of birds. Savile’s work sheds light on the adaptive evolution of the avian wing, emphasizing the development of wing slots and other features that contribute to drag reduction and lift enhancement [108].
Birds with elliptical wings, such as sparrows, thrushes, and other Passeriformes, exhibit a design optimized for quick take-offs and agile maneuvering. These wings are characterized by their rounded shape, which is broader and shorter compared to other wing types. The elliptical wing shape is highly effective in reducing air turbulence at the wingtips, thereby minimizing induced drag. This adaptation is particularly beneficial for birds that navigate through dense foliage or require sudden changes in flight direction. The airfoils are typically characterized by a uniform pressure distribution which aids in good maneuverability. The airfoil of these wings typically has a moderate camber and thickness, helping to maintain lift at various angles of attack. The smooth, rounded leading edge of the elliptical wing airfoil aids in reducing drag during flight maneuvers.
In contrast, High-speed wings are exemplified by species like swallows and falcons. These wings are slender and pointed, reducing the wing area to minimize drag, which is crucial for rapid flight. This wing type is most effective for birds that engage in high-speed chases or need to cover long distances quickly. The reduced drag not only enhances speed but also improves flight efficiency, making it easier for these birds to capture prey or migrate over long distances. They feature low-camber airfoils with a pronounced sweepback of the leading edge. This design minimizes drag during high-speed flight. The thin airfoil profile of high-speed wings reduces weight and drag, allowing for more efficient flight at higher speeds. The leading-edge sweepback also helps delay flow separation at high speeds, contributing to better lift-to-drag ratios.
Soaring wings are found in birds such as gulls and albatross. These wings are long and broad, maximizing the surface area for lift generation. Soaring wings are particularly suited for exploiting thermal currents to sustain flight with minimal energy expenditure. The large surface area enables these birds to glide effortlessly, conserving energy while covering vast areas during flight. The thin airfoil helps in reducing drag, which is crucial for maintaining lift over long periods without flapping. The large aspect ratio of the airfoil aids in creating a more efficient lift distribution along the wing span.
Birds that are adapted to carry heavy loads, such as pelicans and storks, possess high-lift wings. These wings are large and generate a significant amount of lift, supporting the bird’s weight and additional load. High-lift wings allow for efficient energy usage during slow, sustained flights, which is vital for birds that transport materials or engage in long-duration foraging. They often feature slotted wing tips and pronounced camber airfoils. These airfoils are designed to fly efficiently at low speeds, which is essential for birds that require agility and precision during hunting or landing. The slots in the wing tips help to manage the flow of air, reducing vortex formation and associated drag, while the cambered airfoil maximizes lift production at lower speeds.
Lastly, the hovering wing type is best represented by hummingbirds. These wings are small and highly aerodynamic, enabling these birds to perform extraordinary maneuvers such as hovering and even backward flight. The rapid wingbeat frequency and unique bone structure of hummingbirds facilitate this level of aerial agility [26,31,37,73,81,82,92,101,108,114].
Each of these wing types demonstrates a distinct aerodynamic solution evolved by birds with distinctly represented features in the airfoils. The variety in wing shapes underscores the importance of structural adaptation in response to environmental demands and flight behaviors. By studying these diverse wing forms, significant insights can be gained into the aerodynamics of flight, which can also inform the design and development of aerial technologies.
Alula in Flight Dynamics The alula, a small feather or group of feathers found on the leading edge of the wings of most birds shown in Figure 9, and wing slots (discussed previously in Section 2.1.1) play a pivotal role during critical flight phases, such as takeoff and landing. The alula optimizes the wing contour for better lift, while the wing slots reduce drag and improve lift efficiency, underscoring the intricate relationship between wing morphology and aerodynamic demands [215,216,217,218,219,220,221,222,223,224,225,226,227,228,229,230,231,232]. The relationship between wing loading and lift is a focal point, with birds having short wings for maneuverability or frequent take-offs evolving slotting to supply added lift. This aspect of avian adaptation offers a parallel for MAV design, where managing wing loading is essential for performance [108]. Savile speculates on the early evolution of the wing, suggesting that while no existing birds may have a primitive wing in the strictest sense, the inefficient wing of the loons may offer a glimpse into the wing’s early evolutionary forms.
The alula plays a significant role in flight dynamics, particularly at high AOA. Research has shown that the deployment of the alula is influenced by the aerodynamic forces acting on the avian wings [215,216,217,218,219,220,221,222,224,225,227,228,231,232]. For instance, Austin and Anderson’s wind tunnel tests with static specimens of water duck wings revealed that the alula unfolds at specific AOA and velocity combinations, indicating a passive deflection mode, where the alula responds automatically to aerodynamic forces without active muscular control [228].
Contrastingly, other studies, such as those by Lee et al., suggest an active deflection mode controlled by bird muscles. These studies found that the alula deploys at very low flight speeds when aerodynamic forces are insufficient to support its deflection, implying active muscular control [229]. This dual-mode operation of the alula, both passive and active, plays a crucial role during various flight stages, blending the aerodynamic response with muscular control for optimal flight efficiency.
The alula’s small size misrepresents its significant impact on lift. Despite being only 17.5% of the wingspan length, it can enhance lift by up to 22%, making it an effective high-lift device [225]. Its strategic positioning at the wing’s leading edge allows it to manipulate airflow effectively, reducing the risk of stall at high AoAs and enhancing overall aerodynamic performance [232,233].
Another vital function of the alula is its role as a vortex generator. By inducing a stable LEV, the alula significantly increases lift, especially in small-scale birds, insects, and bats [221,223]. For some flying creatures, a stably attached LEV can boost lift by as much as 40%. In larger birds, while the LEV is still present during flight, it is less stable and can quickly transition to flow separation [218,224]. The alula’s position at the wing’s leading edge, particularly at the junction of the hand wing and arm wing, makes it pivotal in LEV development and management, contributing to enhanced aerodynamic efficiency [226,227].
In summary, the alula is a multifaceted aerodynamic feature that plays a critical role in the flight dynamics of birds. Its ability to act in both passive and active deflection modes, coupled with its impact on lift enhancement and LEV management, underlines its evolutionary significance in avian flight mechanics [215,217,219,230].
Implications for MAV Design The insights from research on the alula, have significant implications for the design and performance of MAVs and UAVs. The alula’s role in stall delay and lift enhancement at high angles of attack can be translated into MAV and UAV design to improve aerodynamic efficiency and maneuverability, particularly in challenging flight conditions. Additionally, the alula acts as a vortex generator, creating a stable LEV that significantly increases lift. This mechanism can be mimicked in MAV and UAV wing designs to improve lift capabilities and control in various flight scenarios, including low-speed flight or rapid directional changes. Bionic leading-edge devices, inspired by the alula, have been shown to increase the maximum lift coefficient and enhance stall angles, offering a way to achieve greater lift and aerodynamic efficiency, especially at high angles of attack. Such adaptations are particularly beneficial for MAVs and UAVs operating in urban environments or rugged terrains, where precise maneuvering and stability are crucial. Incorporating alula-inspired mechanisms into the design of these aerial vehicles can lead to improved performance, especially in complex flight conditions, underscoring the potential of bio-inspired design in aerospace engineering [215,216,217,218,219,220,221,222,223,224,225,226,227,228,229,230,231,232,233].
Future Research Directions For future research, Bao et al. recommend using higher-accuracy computational methods like Large Eddy Simulation (LES) or hybrid LES/RANS methods, combined with wind-tunnel experiments and Particle Image Velocimetry (PIV) technology, to further investigate the alula’s aerodynamic mechanisms [31].
Dynamic Airfoil Morphology of Birds
Carruthers et al. give an interesting analysis of the aerodynamic properties of avian wing aerofoils during free flight, particularly focusing on their adaptability and robustness in transitional Reynolds numbers. A distinctive feature of these aerofoils is their ability to deform in response to varying flight conditions, a capability that conventional aerial systems lack. The study offered a detailed reconstruction of the inner portion of an eagle’s wing during a rapid pitch-up maneuver at the end of a glide. This morphing ability of the airfoil is indicative of the bird’s capacity to maintain aerodynamic efficiency across different flight conditions. The feathers on the upper surface of the wing can deflect in response to airflow, signaling flow separation, and the wing’s adaptability [37].
Comparison with Technical Aerofoils When compared with technical aerofoils such as the Clark Y and the Selig S1223 shown in Figure 10 (also used in comparison by Lang et al. [73]), the bird-wing aerofoil sections outperformed the former but do not match the high-lift performance of the latter, which is engineered for maximum lift at low Reynolds numbers. However, the bird-wing aerofoil sections are also less sensitive to variations in shape, turbulence, and surface roughness, indicating a robustness that is beneficial for flight in natural and variable conditions [37].
Design Similarities with Model Aircraft The eagle aerofoil sections combine a high camber with a thick leading edge and a thin, flexible trailing edge that can acquire reflex camber under aerodynamic loading. This design is similar to the Jedelsky and Benedek profiles used in model aircraft, which also feature a thicker leading edge with a plate-like trailing portion. The lift and drag polars of the bird-wing sections suggest a lift curve slope close to the theoretical 2D lift curve slope predicted by thin aerofoil theory, with a stall angle at approximately 14 degrees. This performance offers valuable insights for UAV technologies, as their inherent flexibility and morphing capabilities enable efficient performance across a spectrum of flight conditions [37]. In addition, the introduction of flexible and morphing wing technologies is much more feasible on the scale of UAVs and MAVs.
Numerical Insights into Bird Airfoil Efficiency
The numerical study conducted by Omar et al. continued the analysis of the aerodynamic performance of bird airfoils, highlighting the stork and albatross as prime examples of aerodynamic excellence, something that is commonly discussed [3,31,33,35,39,46,52,56,57,58,59,60,62,68,80,92,97,98,99,100,102,104,105,106,107,108,113,116,117,119,121,127,129,132,137,139,142,143,145,146,147,148,154,157,158,159,160,161,163,166,167,169,172,175,177,208,212,216,234,235,236,237,238,239,240,241,242,243,3,208,212,216]. These birds exhibit airfoil shapes that are optimized for their specific flight behaviors, providing valuable models for MAV and UAV design. The stork’s airfoil is noted for its superior performance at high angles of attack, which is consistent with its migratory behavior involving long-range, high-altitude flights. This efficiency is attributed to its airfoil shape, which maintains lift and delays flow separation even at steep angles, as indicated by the absence of separation observed and a high lift coefficient of 1.77 at the stall angle. Conversely, the albatross airfoil excels at zero angle of attack, aligning with its gliding flight mode near the water surface over long distances. The albatross airfoil’s thickness and camber are tailored to its unique flight requirements, offering design cues for UAVs that need to glide efficiently, minimizing energy consumption, and perform soaring maneuvers discussed in more detail in Section 4.1 [92].
Base to Tip Aerodynamics and Porosity The study by Omar further emphasized the importance of considering the bird’s wing base and tip aerodynamics, as they generate different forces due to their varying camber and thickness. This differentiation could be applied to MAVs and UAVs to optimize lift and drag characteristics for specific flight phases. Furthermore, the investigation into the eagle’s airfoil permeability suggests that incorporating porous surfaces into MAV and UAV wing designs could lead to increased lift and aerodynamic efficiency at certain angles of attack. This bio-inspired feature, such as directional porosity, could be beneficial for controlling wake vortices and improving landing performance [92].
Biological Insights into MAV Design
Liu et al. provide a comprehensive geometric and kinematic description of avian wings, reviewing various bird wings, and elucidating the drag-reducing features of avian airfoils and their kinematics during flight. The study extracted the wing geometry of seagulls, mergansers, teals, and owls (see Figure 8), using a high-precision three-dimensional laser scanner. The authors present detailed analytical expressions for the camber line, thickness distribution, wing planform, chord distribution, and twist distribution. These empirical expressions are very beneficial for modeling and understanding the aerodynamic features that contribute to drag reduction in avian flight and serve as a significant contribution to the field of aerodynamics and bio-inspiration [81].
Kinematic Framework for MAVs The kinematic analysis underscores the importance of understanding the motion of the quarter-chord line and the relative rotation of local airfoil sections. This analysis is most beneficial for MAV design, where dynamic adjustments to wing shape and twist during flight can lead to significantly enhanced aerodynamic efficiency. The research highlights the high camber of seagull and merganser airfoils, drawing parallels with high-lift, low-Reynolds-number airfoils used in MAVs, indicating an optimization for low Reynolds number flight conditions often experienced by MAV-sized vehicles.
2.1.2. Wingtips and Winglets
While wingtips and winglets fall under the general category of wing shape, there has been so much work specifically focused on these aspects that we have provided them with their own section. The tip of the wing has proven to be an area of vast potential for aerodynamic improvement, with many works studying the morphological aspects of wingtips in birds.
Aerodynamic Performance of Wingtip Slots and Research Prospect
Liu et al. present a comprehensive examination of the aerodynamic performance of wingtip slots, drawing inspiration from avian species, which was discussed more in-depth in Section 2.1.1. The review systematically categorizes bird wing planforms and scrutinizes the drag reduction and lift enhancement capabilities of slotted high-lift wings. Their findings are substantiated by experiments on biological specimens and numerical simulations, which confirm the positive impact of slotted wingtips on aerodynamic performance [80].
Wingtip Sails and Ground-Effect Aircraft A significant aspect of this review is the exploration of wingtip sails, particularly in the context of ground-effect aircraft. Studies by Al-Atabi and Sun et al., as referenced in this paper, show that installing wingtip sails can reduce induced drag by about 35% and simultaneously improve lift. This is especially notable as the wing approaches the ground, a critical phase in aircraft operation [80,244,245].
The review also delves into the formation and impact of wingtip vortices on induced drag. Induced drag is a major contributor to total drag during various flight phases, and its reduction is crucial for efficient flight. The detailed analysis of wingtip vortices provides valuable insights into drag-reducing strategies.
Complexities in Wingtip Device Design An essential part of Liu’s review is the emphasis on the complexity involved in designing wingtip devices. The paper highlights that precise adjustment of each winglet is necessary to achieve the desired drag reduction effect, and is specific to the flow conditions they are designed for. This complexity necessitates a comprehensive study of the influence mechanisms of slotted wingtips’ parameters on aerodynamic performance.
The review identifies some inconsistencies in conclusions and gaps in the research scope. Liu et al. propose a research roadmap focusing on the study of geometric parameters of wingtip slots in both steady and unsteady states. They suggest the application of advanced technologies like PIV for flow field visualization and vortex analysis, pointing toward future directions in aerodynamic research.
Aerodynamics and Ecomorphology of Flexible Feathers and Morphing Wings
Van Oorschot’s comprehensive dissertation offers an in-depth exploration of avian aerodynamics, focussing on morphing capabilities in bird wings. This work, situated within the field of Organismal Biology, Ecology, and Evolution, investigates the morphing capabilities of bird wings and their aerodynamic implications, with a particular focus on the role of flexible feathers and wing morphing. A significant aspect of his dissertation is the exploration of wing tip slots (previously discussed) across various avian species. The study examines the aerodynamic role of emarginate primary feathers that form these wing tip slots, suggesting their near ubiquity among avians and potential adaptation for efficacy during vertical takeoff rather than efficiency during gliding flight [117].
Passive Aeroelastic Deflection of Avian Primary Feathers The dissertation examines how individual primary feathers deflect under aerodynamic load. This deflection reorients force, resulting in increased lateral stability and delayed stall characteristics compared to a rigid airfoil. The functional roles of wing tip shape and the emarginate primary feathers are explored, indicating that these features are influenced by ecological demands such as diet, flight style, and foraging behavior. This suggests that efficiency during forward translational flight is only one of the selective pressures acting on wing tips [117].
The flexibility of bird wings, particularly the passive morphing of feathers in response to airflow, is highlighted as a key factor enabling birds to navigate various aerodynamic conditions efficiently. This adaptive quality of bird wings is attributed to the deflection of feathers under aerodynamic load, leading to effective changes in wing shape and consequently, aerodynamic forces. Van Oorschot indicated that feather flexural stiffness varied with body mass, suggesting that feathers become more flexible as birds get heavier. This insight provides an understanding of the function of flexible feathers, suggesting that flexibility aids in passive roll stability and delays stalling during flight [117].
One of the pivotal findings is the relationship between aerodynamic loading and three-dimensional feather deflection. This relationship plays a crucial role in the force production of primary feathers from various raptor species. The dissertation details how these feathers, specifically the slotted primary feathers, function independently as aerodynamic surfaces due to their unique structure. The feathers’ ability to bend, twist, and sweep independently is crucial for their aerodynamic functionality, offering insights into passive roll stability and delayed stall in flight [117].
The study found that bending of the feathers resulted in the generation of lateral forces approximately 10% of the total lift. This was observed in comparison to the Clark-Y airfoil, which stalled at an attack angle () of 13.5°, whereas all feathers continued to increase lift production beyond the range of measurements (=27.5°). This result is attributed to the spanwise long-axis twist which reduced the local attack angle at the feather tips. This finding illustrates that feather flexibility enhances lift production and delays stall in flight [117].
Aerodynamic Analysis of Bionic Winglet-Slotted Wings
A recent study computationally explored the aerodynamics of bionic winglets inspired by bird wings, particularly focusing on slotted wings for use in UAVs. The study aimed to harness the aerodynamic advantages observed in bird wing tips for bionic UAVs. It highlights the design and aerodynamics of winglets that mimic the slotted wings of birds, known for their vortex reduction and enhanced roll stability. This research created an early examination of the functionality of winglets in diminishing lift-induced drag and enhancing aerodynamic performance through a morphing winglet mechanism. The challenges and benefits of replicating bird wing structures were discussed, with a focus on their role in reducing drag and enhancing stability and control. Experimental studies on various winglet configurations underscore their influence on aerodynamics, particularly in enhancing lift and reducing drag. The paper serves as an early foundation for future research in the development of bionic UAVs with superior endurance and control, exploring the potential of biomimetic approaches in aerospace engineering [46]. The work focussed on computational results that have not yet been validated, and only provides initial results to contemplate.
Varying Wingtip Devices
The study by Öztürk and Örs provided a comprehensive analysis of how different types of winglets and wingtip devices affect the aerodynamics of aircraft, some of which can be seen in Figure 11. The research emphasizes the use of CFD to analyze these effects, particularly focusing on the reduction of wingtip vortices [95]. The study explored the evolution of winglet and wingtip designs, underscoring their vital role in reducing induced drag and enhancing overall aircraft performance. This research produced a comparative analysis of different winglet designs and their impact on aerodynamic performance. By referencing specific studies and results, such as those conducted by Narayan and John using Ansys Fluent who found a max improvement in the lift-to-drag (L/D) of 22.59% with a multi-tip design, the paper demonstrates the superiority of certain designs in enhancing the lift-to-drag ratio [90]. This improvement in the L/D ratio translates into better fuel economy, extended range, and higher payload for aircraft. Among the various winglet designs, the blended winglet and maxi winglet, as studied by Gavrilović et al., improved the CL/CD ratio by 3.93% and 5.32% respectively, significantly enhancing the range and reducing fuel consumption [246]. Reddy et al. found that scimitar winglets increased the CL/CD ratio by 14.48% compared to plain wings, offering substantial aerodynamic advantages. Hossain et al. observed a 6% improvement in lifting and a 28% reduction in drag forces with elliptical winglets [63]. It was reported Ekinci found the best L/D value with a 45° winglet inclination angle, leading to a 4.78% increase in aircraft range [95]. In comparative studies, Wan et al. noted that modern spiroid winglets outperformed conventional winglets by 5.68% in terms of L/D value [247]. Furthermore, Falcão et al.’s research on adaptive and controllable winglets resulted in significant performance enhancements, including a 25.32% increase in CL value at minimum stall speed [248]. Overall, these studies underscore the critical role of winglets and wingtip devices in optimizing aerodynamic performance, with each design offering unique benefits in terms of lift, drag, and overall efficiency [95].
Study Methodology and Winglet Designs
Narayan and John’s comparative study on the effectiveness of different winglet designs in altering the tip vortex structure provides a framework to optimize enhancements in aircraft efficiency [90]. The paper presents a methodology involving computational simulations using Ansys Fluent and the Reynolds Averaged Navier–Stokes equations, focusing on the flow and vortex structure around the half wing. The winglets tested include various designs and configurations, providing a broad spectrum of data for analysis. A key finding of the study is the strong correlation between the size of the tip vortex and essential aerodynamic parameters like lift, drag, and pitching moment. This relationship is pivotal in understanding how modifications in winglet design can influence overall wing performance. The research highlights that multi-tipped winglets are particularly effective in dispersing vortex energy and reducing induced drag. The optimization of the number of tips is crucial for increasing the lift coefficient while minimizing frictional and vortex drag. The study concluded that among the various winglet designs tested, the multi-tip-3 winglets offer the best lift-to-drag ratio at 2.59%, making them an optimal choice under specific conditions for improving flight range and duration [90].
Experimental Optimization of a Wingtip Vortex Turbine
The thesis by Andrew Roberts shifted the focus from winglet designs to the design and experimental optimization of a Wingtip Vortex Turbine (WVT) for general aviation. This innovative concept aims at reducing induced drag by utilizing the vortex flow near the wingtips [103]. The thesis addressed the challenge of minimizing aircraft drag, with a specific focus on induced drag, which is particularly significant during cruising flight. Traditional drag reduction methods, such as winglets, have typically acted as stators to suppress vortex swirl. The WVT concept builds on this principle but adds a novel twist by operating in a rotating mode to extract energy from the vortex rather than mitigate it. Roberts details the design and testing of a 1/4-scale model of a thrust-producing WVT in a wind tunnel. The study experimented with various configurations, including 2 to 6-bladed designs, across different pitch angles. The 6-bladed configuration was found to be most efficient for drag reduction and power generation. The research indicates that winglets could reduce induced drag by about 13 percent, acting as stators to suppress vortex swirl. However, the thrust effects of a WVT alone are unlikely to significantly reduce overall drag for general aviation aircraft, with more pronounced benefits in heavier transport-type aircraft [103].
Power Generation and Drag Impact One of the intriguing aspects of the WVT is its potential for power generation. The model generated a power coefficient of 0.004 at cruise and a maximum of 0.0059 at a higher lift coefficient, equating to significant full-scale power outputs. In terms of drag impact, the WVT slightly increased overall drag at cruise but reduced overall drag at higher lift coefficients, depending on its stationary or rotating state. Roberts concludes that while the WVT can reduce drag, its efficiency as a thrust-producing device is lower than anticipated for general aviation aircraft. The thesis recommends concentrating more on vortex flow straightening and exploring airfoil and twist distribution modifications. It also suggests flight-testing of new WVT designs for further validation and optimization [103].
Gliding Harris’ Hawk Wingtip Slots
In his 1995 study, Tucker examined the aerodynamic efficiency of wingtip slots in soaring birds, using the Harris’ Hawk as a model. This research, pivotal in aerodynamics, demonstrates that slotted wingtips, similar to aircraft winglets, significantly reduce drag, marking an important contribution to biomimetic aviation design of the time [115].
Tucker’s experimental analysis revealed that the Harris’ Hawk, with its natural wingtip slots, exhibited substantially lower drag—only 70-90% of the drag when compared to its clipped counterpart. At a wing span of 0.8 m, the unclipped hawk exhibited a mean induced drag factor of 0.56, considerably lower than the 1.10 assumed for the clipped hawk, indicating a marked reduction in drag due to the wingtip slots. This drastic reduction is attributed to the slots spreading vorticity both horizontally and vertically, akin to winglets on aircraft. These findings emphasize the potential of integrating similar designs in aircraft for enhanced aerodynamic performance. The study underscores the significance of biomimicry in aircraft winglet design, suggesting that emulating the natural wingtip slots of birds like the Harris Hawk can lead to significant advancements in reducing drag and improving flight efficiency [115].
Wingtip Vortices in Flapping Wings
The work by Qin et al. explored the dynamics of wingtip vortices in a flapping wing model at a scale comparable to birds. The study is unique in providing an understanding of the aerodynamic behavior of wingtip vortices in flapping-wing systems. The research primarily focuses on the controlled evolution of wingtip vortices, examining how different wingtip designs affect the formation and behavior of these vortices. The methodology involves both computational and experimental PIV analyses, allowing for a comprehensive understanding of the aerodynamics involved [99].
The study discussed that wingtip vortices exhibit significant changes in vorticity concentration and vortex radius throughout the flapping cycle. Notably, with an increase in flapping frequency, the vortices become more dispersed, indicating a direct influence of flapping motion on vortex behavior. Qin’s research also highlighted that the trajectory of wingtip vortices in flapping flight is distinct from fixed-wing models. The study showed that the vortices in flapping wings follow a regular wave-like evolution trajectory, contrasting with the more stable helical structure observed in fixed wings. This indicates that flapping wing motion can give wingtip vortices a specific directionality, as opposed to random wandering. The findings suggest that flapping wing motion can effectively control the main shape and energy concentration of wingtip vortices. The proper orthogonal decomposition results indicate that flapping motion not only affects the shape of the vortices but also their energy distribution, leading to a more dynamic and controlled vortex evolution [99].
A Numerical Study of a Biomimetic Wing in Soaring Flight
Tangermann et al. present a numerical study on the aerodynamic behavior of a biomimetic wing in soaring flight, providing key insights into how the design principles of bird wings can be applied to enhance the aerodynamics of engineered flight systems. The research focused on replicating the aerodynamic characteristics of bird wings, particularly in soaring flight. Through RANS and hybrid RANS-LES CFD models, the study simulated the aerodynamic performance of biomimetic wings, offering a detailed analysis of lift generation, drag reduction, and overall flight efficiency. The performance results were compared and validated for plausibility with measurement data from live white storks. Central to the paper is the exploration of biomimetic wing designs, inspired by the structural and functional aspects of bird wings. The study examines how variations in wing geometry, such as aspect ratio and wingtip design, influence the aerodynamic performance of the wing in various flight conditions [113].
The research utilized RANS simulations to cover a large range of angles of attack and conducted scale-resolving simulation at a selected condition for detailed insight into the turbulent flow. The glide efficiency and sink rate results from measurements were reported to be slightly better than the values predicted from a series of RANS calculations. The deviations are attributed to uncertainties in the geometric design details. Nevertheless, the results closely match the experimental findings and provide a good quantitative prediction of efficient flying states [113].
The flow field analysis helped obtain a closer view of the flow field across the feather cascade. The simulation revealed how the flow from the lower side passes between the primary feathers, generating additional lift before forming a vortex. This is an important element in understanding the flow dynamics associated with multi-element wing tips and their contribution to aerodynamic performance. Tangermann indicated that the glide efficiency and sink rate from the measurements are slightly better than the predicted values, although the results are close to experimental findings and provide a good quantitative prediction of efficient flying states [113].
2.1.3. Feather’s Structure and Riblets
The fascinating structure of bird feathers, particularly herringbone riblets, has been the subject of extensive biomimetic research, focusing on drag reduction. This research realm not only provides insights into the natural world’s aerodynamic efficiencies but also opens up possibilities for their application in technological advancements. In avian flight, the alula and other feather structures act as natural boundary layer-tripping devices. These morphological adaptations are essential for maneuverability and stability, particularly at low Reynolds numbers where flow separation is more likely. For instance, the roughness of feathers has been shown to reduce flow separation during low-speed glides, enhancing the aerodynamic performance of birds such as swifts. Studies have demonstrated that these feather structures create small vortices that trip the boundary layer, promoting a turbulent flow state that delays separation and reduces drag [249,250].
Pioneering Research on Bird Feather Structures for Drag Reduction
The work of Chen et al. marked a significant milestone in understanding bird feather aerodynamics. Their meticulous analysis using Scanning Electron Microscopy (SEM) delved into the microstructure of pigeon feathers, focusing on the herringbone riblets. These studies were complemented by rigorous water tunnel testing of biomimetic surfaces, which demonstrated a remarkable drag reduction rate of up to 16%, significantly more effective than traditional U/V-microgroove riblets. Notably, their research revealed the crucial role of riblet orientation, finding that specific angles, particularly 30°, were optimal for drag reduction. This highlighted the importance of aligning riblet structures with flow direction, a principle observed in the natural configuration of bird feathers [2,16,17].
Bionic Surface Development Inspired by Feather Microstructure In parallel, Feng et al. explored the creation of bionic surfaces that replicated the microstructure of bird feathers. Their innovative use of hot-rolling technology on PVC substrates led to the formation of microriblet films, closely mimicking the natural design. Wind tunnel tests of these surfaces showed an average drag reduction rate of 10% across various Mach numbers, illustrating the potential of such designs in aerodynamic applications, particularly under subsonic airflow conditions. The research highlighted the role of micro-vortices within the riblets in reducing skin friction drag, offering a promising avenue for reducing aerodynamic drag in aviation and other high-speed travel forms [18].
Figure 12.
Examples of biological surfaces, avian riblets, and riblet inspired designs. a & b) Views of SEM shark skin denticles [14]. c) Example of sea leopard skin [14]. d & e) Views of SEM secondary flight feather riblets [16]. f) Example test surface with 3D riblets mimicking shark denticles [14]. g) Micrograph image of a toucan’s feather surface structure from a 3D topography interferometer [18]. h-j) Schematic views of riblet and herringbone riblet inspired surface microstructures [15]. k-m) Instantaneous vortical structures and wall shear from the schematic views of riblet structures [15]. n & o) Another example of iso-surfaces showing the Q-criterion with colors representing the streamwise velocity [125]. All figures are reprinted with permission; please see the Appendix A.
Figure 12.
Examples of biological surfaces, avian riblets, and riblet inspired designs. a & b) Views of SEM shark skin denticles [14]. c) Example of sea leopard skin [14]. d & e) Views of SEM secondary flight feather riblets [16]. f) Example test surface with 3D riblets mimicking shark denticles [14]. g) Micrograph image of a toucan’s feather surface structure from a 3D topography interferometer [18]. h-j) Schematic views of riblet and herringbone riblet inspired surface microstructures [15]. k-m) Instantaneous vortical structures and wall shear from the schematic views of riblet structures [15]. n & o) Another example of iso-surfaces showing the Q-criterion with colors representing the streamwise velocity [125]. All figures are reprinted with permission; please see the Appendix A.
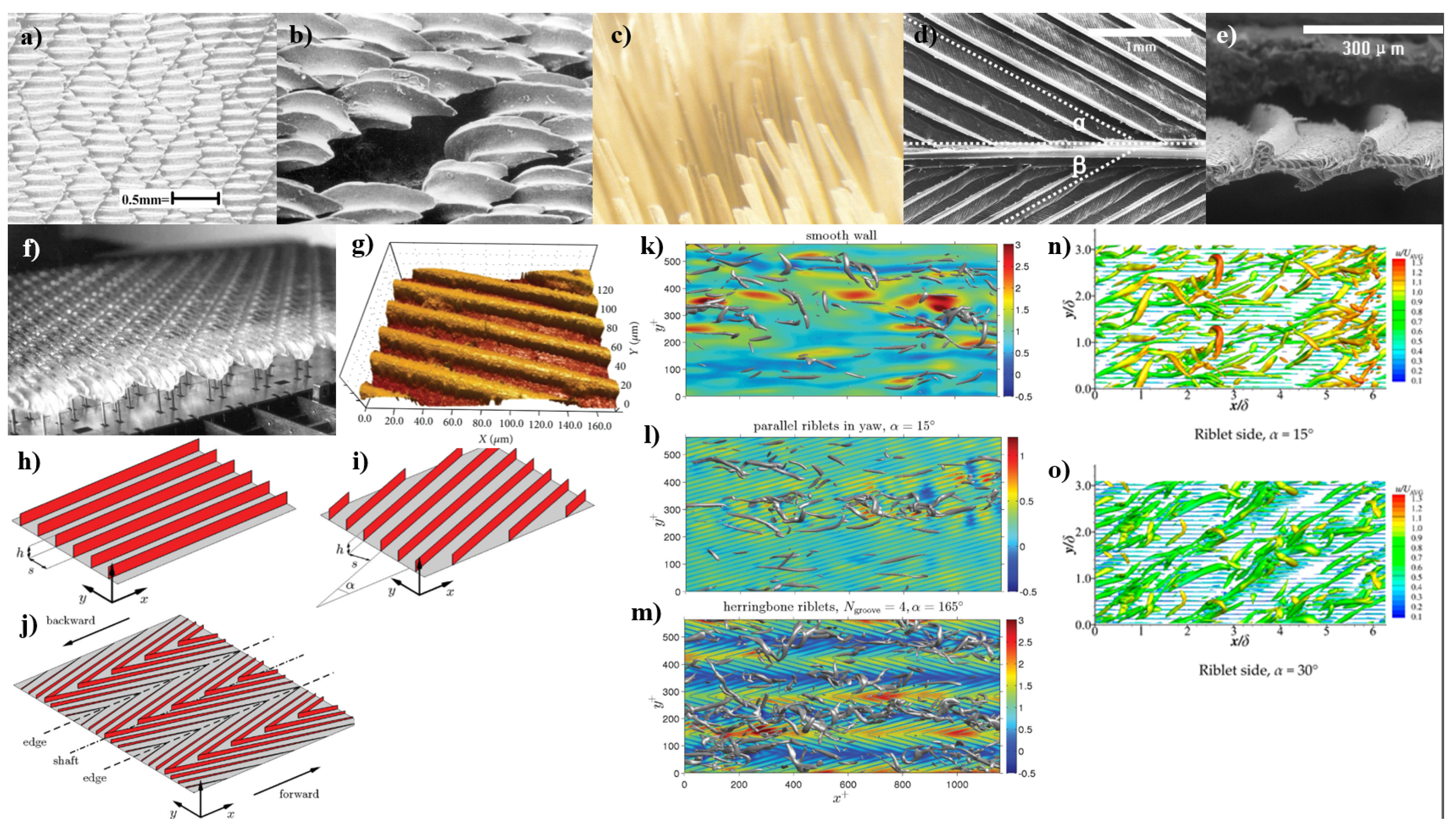
Insights from Various Biological Surfaces
Bechert et al. expanded the realm of biomimetic study by examining a diverse range of biological surfaces for their aerodynamic properties. This study included an exploration of shark skin replicas and the self-cleaning properties of lotus leaves, each offering unique insights into drag reduction and surface maintenance. The research emphasized the potential of adopting such nature-inspired designs in technological applications, especially in the aviation industry. The implementation of riblets, inspired by shark skin and bird feathers, in long-range commercial aircraft, was discussed as a strategy to reduce fuel consumption and maintenance costs, thereby enhancing the efficiency and environmental sustainability of aviation technology [14].
One of the primary focuses of the study is on reducing wall shear stress in turbulent flows. The research highlights how riblet surfaces, including shark skin replicas, can achieve this reduction. Specifically, the paper discusses the performance of various riblet surfaces, noting that the synthetic two-dimensional blade rib surfaces outperformed even the ambitious shark skin replica in terms of shear stress reduction. This finding is crucial as it suggests that while biological surfaces inspire these designs, technological enhancements can lead to even more efficient solutions for drag reduction. In addition to shear stress reduction, the paper also delves into separation control, a critical factor in the survival of certain species. The study emphasizes that biological surfaces sometimes combine shear stress reduction with separation control. This concept is exemplified by the analysis of bird feathers, where self-activated movable flaps (discussed more in-depth in Section 2.1.1) enhance the maximum lift of airfoils by about 20% without significant adverse effects under cruise conditions. The study’s findings in this area point towards potential applications in improving aircraft wing designs [14].
LES and DNS Analysis of Riblet Structures Furthering the understanding of riblet structures, Zhang and Yin focused on their aerodynamic effects, particularly on swept wings. Their study employed implicit LES to assess the impact of riblets in channel flow and on an infinite swept wing. They observed that riblets demonstrated a drag reduction effect when aligned at smaller incline angles with the flow direction. However, at higher angles, such as 30°, this effect reversed, leading to increased drag which contrasts the studies by Feng et al. This nuanced understanding of riblet efficiency under different simulation and flow conditions provided crucial insights for the design of aircraft wings and other aerodynamic structures [125].
The research extended to studying an infinite swept wing with a 30-degree angle, both with and without riblet films. The results showed that riblets demonstrated satisfactory drag reduction efficiency due to the mild cross-flow over most parts of the wing. The study underscored the significance of riblets in reducing both lift and friction drag, following the cosine law of a swept wing. The drag reduction was primarily induced by the friction drag, while the pressure drag was slightly increased [125].
The research by Benschop and Breugem took a similar route, delving into the complexities of herringbone riblet textures in turbulent channel flows. Employing direct numerical simulations, they analyzed the drag change mechanisms of these riblets, uncovering that their efficiency in drag reduction varied depending on the specific design parameters. For narrow feather widths, they noted an increase in drag due to the generation of fluctuating secondary flows by convergent/divergent riblets. In contrast, for larger feather widths, they observed a slight reduction in drag, suggesting the potential of these textures in certain aerodynamic applications. This study highlighted the intricate balance required in biomimetic designs, particularly when dealing with turbulent flow environments, and underscored the need for comprehensive testing and analysis to optimize these bio-inspired structures for practical use. The study extensively used the Fukagata-Iwamoto-Kasagi identity for drag decomposition to analyze the drag change mechanisms. The results indicated that convergent/divergent riblets in the texture are generally detrimental to turbulent drag reduction. This is due to the induced secondary flows that enhance mean and turbulent advection. The presence of these secondary flows was found to be a critical factor in determining whether the texture would increase or decrease drag [15].
The Role of Feather Morphology in Aerodynamic Performance Li et al. provided a unique perspective by examining the impact of morphological integrity on the drag reduction performance of pigeon feathers. One of the critical findings of the study is that the morphological integrity of feathers, where barbs are closely interconnected, plays an essential role in drag reduction performance. Intact feathers, characterized by their tightly connected barbs, were found to significantly reduce drag at low wind speeds compared to damaged and artificial feathers. This effect was attributed to the feather’s unique morphology, which enables strategies to maintain integrity and minimize wing damage, thereby optimizing drag reduction [77].
The research utilized wind tunnel experiments with force balance and smoke flow field visualization tests on different types of feathers. These experiments confirmed the hypothesis that intact feathers, due to their structural integrity, are better suited for drag reduction. Specifically, the study demonstrated that intact feathers experienced much lower drag force compared to damaged and artificial feathers at lower wind speeds. This implies that intact feathers have morphological adaptations for excellent drag reduction performance, particularly during critical flight phases like take-off or landing [77].
In a broader evolutionary context, the studies by Péter L. Pap et al. and Gergely Osváth et al. provided valuable insights into the structural properties and morphological variations of flight feathers across different bird species. These investigations revealed how feather morphology has adapted to various flight requirements and habitats, influenced by ecological and life-history traits [93,96]. For instance, Pap et al. used a phylogenetic comparative approach to analyze feather samples from 661 adult birds belonging to 137 species, finding significant relationships between feather morphology and flight type, habitat, wing morphology, and diet [96]. This comprehensive approach underscored the intricate relationship between ecological traits and aerodynamic efficiency, offering a window into the evolutionary dynamics of avian flight.
The research conducted by Rader et al. further enriched the understanding of bird wing evolution by examining the relationship between mechanical sensitivity and evolutionary dynamics. Their study, utilizing 3D scans of 1096 bird wings from 178 species, challenged the traditional view of morphological modularity in wing evolution. They demonstrated that evolutionary rate and morphological disparity in bird wings varied continuously along a mechanical sensitivity gradient, influenced by the biomechanics of flapping flight. This finding suggested that the evolutionary dynamics of bird wings are more closely aligned with a gradient of mechanical sensitivity than with strict modular divisions, offering a new perspective on the evolution of complex biological structures [100].
2.1.4. Moult and Porosity of the Wings
The study of avian aerodynamics, particularly focusing on wing porosity and the molting process, presents a comprehensive understanding of the natural mechanics of flight. This exploration into the intricacies of bird wings offers significant insights into both biological and engineered flight systems. Some examples of wing porosity experiments and molting can be seen in Figure 13.
Permeable Airfoils
The study by Omar et al. was previously discussed in Section 2.1.1, however, there was another aspect of this study that was not mentioned. A key aspect of the study was the investigation of permeability in the eagle’s airfoil. The researchers modeled one-third of the airfoil chord near the trailing edge as a flat plate with holes spaced every 0.25 cm. This permeable section allowed airflow from the lower surface to the upper surface, maintaining the same flow properties. The study found that permeability significantly increased the aerodynamic efficiency of the eagle’s airfoil at angles of attack less than 10°. Particularly at a zero angle of attack, the lift increased by 58%, indicating a substantial enhancement in the airfoil’s performance due to permeability [92]. This was also in good agreement with the study by Aldheeb et al. [8].
The permeable section of the eagle’s airfoil was shown to improve aerodynamic efficiency at low angles of attack. However, beyond a specific angle (stall angle), permeability adversely affected the flow, potentially due to the transition to turbulence ahead of the permeable section. The study concluded that the maximum lift coefficient of the eagle’s airfoil could be achieved at a lower angle of attack by applying permeability to one-third of the airfoil chord. The study also compared the performance of the permeable eagle’s airfoil with a nonpermeable version. The nonpermeable airfoil showed flow separation at its trailing edge at higher angles of attack, while the permeable airfoil did not exhibit such separation and maintained smoother streamlines. However, at very high angles of attack, the permeable airfoil experienced more severe separation than the nonpermeable airfoil, leading to a loss of lift [92].
Iosilevskii performed similar research into the aerodynamics of wings that incorporate a permeable aft segment, similar to the structure observed in bird wings. This study addressed how the permeability of the wing’s surface influences its aerodynamic performance, including the lift and the positioning of the aerodynamic center. One of the central findings is that the lift slope coefficient of a wing uniformly decreases with increasing permeability, shifting from the standard value of 2 to 2 times the relative width of the impermeable forward part. Concurrently, the aerodynamic center moves from its quarter-chord position to the quarter-chord point of the forward part, but this movement isn’t uniform. In cases where the width of the forward part is less than half of the chord, the aerodynamic center initially moves backward, then forward. [66].
The effectiveness of the permeable aft part of the wing varies with different parameters. The study notes that under certain conditions, particularly at a negative angle of attack, the aerodynamic center initially moves backward, then forward. This implies that accelerating a permeable wing at a constant angle of attack could lead to a decrease in its lift coefficient and a possible backward shift of its aerodynamic center. Such effects could render the permeable part of the wing less effective at high speeds, and combined effects might negatively impact the speed stability of UAVs or birds with such wings [66].
Porous Locations Hanna et al. investigate the effects of porosity on the aerodynamic performance of wings at moderate Reynolds numbers, using the NACA 0012 airfoil as a model. This study is especially relevant in the context of natural flyers like birds and bats, which operate in a similar Reynolds number regime and feature wings with varying degrees of porosity. The research highlights the complexities in aerodynamic behaviors at moderate Reynolds numbers, such as unpredictable performance characteristics including hysteresis loops in lift due to the formation and movement of laminar separation bubbles. These phenomena alter the effective shape of the airfoil and can lead to non-monotonic lift-drag profiles, posing significant challenges for small wings at this scale [55].
One of the significant findings of the study is that even small permeabilities in the wing can substantially affect its performance. The research demonstrated that introducing low porosity between the pressure and suction surfaces of the wing could remove counterintuitive behaviors such as negative lift production at small, positive angles of attack. This modification did not introduce a measurable drag or lift-to-drag penalty, indicating that subtle changes in wing structure can significantly impact aerodynamic efficiency [55].
The porous wing consistently performed better than the solid one, with the presence of holes in the porous wing almost completely straightening out the lift curve around zero degrees angle of attack. This was accompanied by little difference in drag coefficient, suggesting an overall improvement in aerodynamic efficiency for the porous wing. Dye trace visualization was used to analyze the flow patterns around both the solid and porous airfoils. For the porous airfoil, small-scale eddies formed in the boundary layer and wake, leading to small perturbations in the fluid flow. These perturbations contributed to the straightening of the lift curve near zero degrees angle of attack. The study found that the pressure difference between the suction and pressure surfaces may induce a fluctuating fluid flow through the wing, causing local disturbances to the boundary layer at the pores [55].
Aldheeb et al. provided comprehensive insights into how porosity affects aerodynamic performance, particularly in relation to lift and drag. The study uses a symmetric thin porous airfoil and a porous rectangular half-wing for its experiments conducted in a low-speed wind tunnel. The experimental results show a direct correlation between porosity and the aerodynamic coefficients of lift, drag, and pitching moment. As porosity increases, the lift coefficient consistently decreases, while the drag coefficient generally increases. This increase in drag is mainly due to the enhanced viscous effects caused by airflow through the porous regions [23].
Computation of Flow Past a Flat Plate with Porous Trailing Edge
Bae et al. investigate the flow past a flat plate with a porous trailing edge, employing a penalization method. This approach is motivated by prior works showing the benefits of porous treatments in significantly diminishing vortex shedding, reducing pressure fluctuations near the edge, and yielding more diffused side-edge vortex structures. The work discusses how porous treatment can significantly diminish vortex shedding, reduce the intensity of pressure fluctuation near the edge, and yield more diffused side-edge vortex structures. This is achieved by allowing flow transpiration through the pores and promoting local blowing and suction along the surface, as observed in the trailing edge of owl feathers. The study illustrates how a passive form of time-dependent blowing and suction along a porous surface can break the spatially coherent pressure field, thereby reducing drag and lift forces [29].
Bae et al. showed that with the porous treatment, the time-averaged drag coefficient decreases by 24%, and the amplitudes of drag and lift fluctuations are significantly reduced. This reduction in aerodynamic coefficient is generally accompanied by an increase in the vortex shedding frequency, which directly reflects on the dipole noise generated at the trailing edge. The research findings indicate that the fluid flows in porous media are accurately predicted by the present modeling, underscoring the effectiveness of porous surfaces in aerodynamic performance improvement. The results of this study have practical implications for the design of aerodynamic structures, particularly in reducing noise and improving performance. The study’s insights into the effects of passive porous surfaces on turbulent flow and noise are expected to inform future research and applications in areas such as aerospace and automotive engineering [29].
Experimental Study on the Pore Density
Kong et al.’s study delves into the intricate effects of porous media on airfoil aerodynamics. Utilizing time-resolved particle image velocimetry, the research compares the aerodynamic performance of airfoils with porous media of varying pore densities to a smooth airfoil model. The study’s main focus is on how different pore densities in porous wings impact aerodynamic forces, especially lift and drag. It was found that only porous media with the appropriate pore density can significantly improve the aerodynamic performance of the airfoil. The study determined that if the pore density of the porous media is too small, the aerodynamic performance is severely damaged across the entire range of angles of attack. Conversely, if the pore density is too large, the porous media may act like a spoiler, increasing the viscous effect and reducing the aerodynamic power of the airfoil, especially at small angles of attack [69].
laying porous media on the airfoil’s suction side led to a considerable reduction in negative pressure, particularly in the porous media area. This resulted in a decrease in the lift coefficient, which is consistent with previous force measurement results. The control effect of porous media was most considerable at a 6° angle of attack, while after the stall angle of attack, the porous media still had a large effect on the airfoil’s aerodynamic performance [69]. The flow visualization results indicated that the porous media with 20 pores per inch reduced the circulation of flow velocity around the suction side of the airfoil and destroyed the vortex structure. This broke the low-frequency large-scale vortex into a high-frequency granular vortex, inhibiting the amplitude of vortex fluctuation, effectively weakening the energy of different vortex modes, and thus improving the airfoil’s aerodynamic performance [69].
Yilmaz et al. similarly investigated the effects of porosity on the flight performance of flapping wings, focusing on how different numbers of pores and their angles of inclination influence lift and drag forces. Using 3D porous flapping wing flow analyses on Comsol Multiphysics, the study explored various configurations of pores on a wing. It was found that changing the number of pores in the wing, as well as the inclination angle of these pores, significantly affects the lift forces produced during flight. The study experimented with pore numbers of 36, 48, and 60 and pore inclination angles of 60, 70, and 80 degrees. It was observed that as the number of pores on the wing increases, the lift value moves away from zero, suggesting an improvement in lift generation with higher pore numbers [124].
Air Transmissivity of Feathers Müller and Patone’s study, "Air Transmissivity of Feathers," provides an in-depth analysis of the air transmissivity of bird feathers, focusing on the flight feathers and coverts of the European kestrel. The work involved testing the air transmissivity of feathers from a European kestrel (Falco tinnunculus) and 27 other bird species from 15 different families. The transmissivity was measured in both directions, from ventral to dorsal and vice versa. The mean difference between the two directions was found to be less than 10%, with the transmissivity from dorsal to ventral being slightly higher. This small but significant difference suggests that the feathers’ transmissivity varies by location and can impact the pressure distribution on a bird’s wing [88].
The research presented the transmissivity of the inner vanes of feathers from the 27 different species was similar to that of the kestrel feathers, ranging from 0.0001 to 0.0005 m3 s−1 N−1. This similarity across species indicates a common aerodynamic function of feather structure among various birds. The transmissivity of the outer vanes was found to be more than that of the inner vanes, which the researchers propose has significant aerodynamic implications. Morphological observations supported the transmissivity results, indicating that structural gaps in a perfectly intact feather, responsible for its transmissivity, must be very small. The study suggests that the enlargement of the ventral margins of the barbules and their apposition to the bases of adjacent barbules reduce these gaps, allowing the vanes to function as a relatively impervious surface while maintaining some level of transmissivity [88].
Biomimetic Self-Adaptive Porous Flaps
Mazellier et al. introduced a passive control system for drag reduction inspired by the molting and porosity of bird feathers, representing a novel approach in biomimetic design. The control device consists of flaps made from a combination of a rigid plastic skeleton and porous fabric, simulating the shaft and vane of a bird’s feather, respectively. These flaps are designed to mimic the main features of bird feathers and are fitted on the sides of a square cylinder, allowing each flap to freely rotate around its leading edge. This system is self-adaptive and activates based on flow separation at the corners of the cylinder, demonstrating the efficiency of passive control in aerodynamics [86].
They found that the implementation of this control system led to a significant drag reduction, approximately 22% on average, over a broad range of Reynolds numbers tested. This improvement in aerodynamic performance is attributed to the increase in base pressure in the controlled case. The control system modifies the pressure distribution on the lateral sides and base of the cylinder, leading to a noticeable change in the flow topology at a large scale in the vicinity of the controlled cylinder. The relative motion of the flaps and the flow dynamics were analyzed, showing that the dynamics of the flaps are sensitive to the Reynolds number. Furthermore, the study of the flow dynamics at a large scale suggests a lock-in coupling between the flap motion and vortex shedding, indicating a complex interaction between the control system and the surrounding flow [86].
Effects of Wing Damage and Molt Gaps Flight Performance
Hedenström’s research explores the impact of wing damage and molt gaps on the flight performance of vertebrates, particularly focusing on birds and bats [60,62]. His 1999 work used CFD to examine the effects of molt gap size, geometry, and position on the wing. It was found that the detrimental effect of molt gaps on flight performance increased with the wing’s aspect ratio. The impact was most significant when the molt gap was positioned well inside the wing, as the circulation declined close to the wing tip. In contrast, when the gap was at the wing tip, the performance was slightly increased because the lift distribution then became closer to the optimal elliptical distribution [62]. The 2023 work elaborated on the ecological consequences of wing damage in birds, indicating that such damage leads to reduced flight performance across various metrics. The study emphasized that during molt, birds can partially compensate for reduced flight performance through enlarged flight muscles. However, they tend to change their behavior, such as becoming more secretive, to minimize exposure during this vulnerable period [60]. Both studies explore the effects of natural molt gaps and experimentally created wing gaps. the works discussed how experimental manipulation, such as the symmetric removal or trimming of flight feathers, affects flight effort and performance. This approach offers valuable insights into life-history decisions during breeding, where the main effects monitored are the condition of the manipulated birds and the growth of their chicks [60].
Implications of Wing Feather Molt
Achache et al. investigated the aerodynamic implications of wing feather molt in Anna’s hummingbirds, focusing on changes in lift production during various molt stages. Using dynamically scaled hummingbird wings in fluids, they studied time-accurate aerodynamic loads and flow field measurements over different wing geometries that represent stages in the natural molt sequence of this species. The researchers found that depending on the molt stage, lift production can drop by more than 20% compared to the complete (non-moulting) wing. The largest effect on wing performance was observed when the medial flight feathers were missing, leading to significant flow fluctuations that reduced the effective disc area of the wing. The study underscores that the feathers in the medial section of the wing, which are sequentially shed only after the outermost feathers (P1-4) are fully regrown, play a critical role in maintaining lift during flight [21].
The study also provides power and efficiency analyses for hovering flight during molt under various plausible scenarios. It suggests that body mass reduction could be a compensatory mechanism that preserves the energetic costs of hover flight. Specifically, a 25% reduction in body mass during molt was measured for molting ruby-throated hummingbirds, indicating that body mass variation during the molting period is a significant factor in maintaining flight efficiency [21].
Passive Separation Control by Acoustic Resonance
Work by Yang and Spedding investigated the transformative effects of small holes in the suction surface of an Eppler 387 wing on its aerodynamics. This study is significant in understanding the sensitivity of wing aerodynamics to small geometric changes and the fluid environment, especially at transitional Reynolds numbers. found that the presence of small holes in the suction surface of the Eppler 387 wing significantly changes its aerodynamic performance. This effect is not due to the physical presence of the cavities alone, as they have no effect by themselves. Instead, the acoustic resonance within these cavities interacts with intrinsic flow instabilities, leading to notable changes in the flow pattern and aerodynamic performance of the wing [123].
The research showed that the small holes could promote a switch from a low-lift state to a high-lift state by forcing through acoustic resonance. This shift was observed to occur when the resonant modes in the cavities were close to the most unstable modes in the original flow. The large effect of these small holes underscores the sensitivity of wing aerodynamics to subtle features and their potential for passive flow control strategies [123].
The findings of this study have significant implications for wing design, particularly for MAVs operating in transitional Reynolds number regimes. The research also indicates the need for caution in interpreting and designing pressure tap measurements in such flow regimes, as small geometric cavities can substantially influence wing performance. The study suggests that the passive effects of the holes could be replaced with local, digital control of forces and moments on the wing through equivalent, local forcing [123].
2.1.5. Body Shape
Bird body shapes have evolved to meet the diverse aerodynamic demands of flight, both in the air and underwater. Despite the significant impact bird bodies can impart on aerodynamic performance, it seems there have been relatively few studies focused particularly on the aerodynamics of avian bodies and how they have adapted to maximize efficiency. According to Maybury, a considerable amount of controversy has surrounded the consideration of body drag, or parasite drag, in recent years, and his dissertation aimed to address this [85].
Evolutionary Advantage
Bird flight, encompassing diverse modes such as flapping, gliding, and diving which are discussed at length in Section 2.2, showcases evolutionary adaptations optimizing body shape for efficiency in the air and underwater. These adaptations are influenced by the mechanics and aerodynamics of flight, impacting bird behavior and evolution [85]. The evolution of bird body shape is not solely dictated by aerodynamic optimization, however, but is also constrained by their ancestral morphology. Birds, having evolved from small bipedal theropod dinosaurs, possess wings formed from forelegs. These structural constraints limit the extent to which their bodies can taper, affecting their aerodynamic profile. However, despite these constraints, certain anatomical features have evolved to reduce drag, such as dorso-ventral asymmetry and the role of the furled tail in controlling airflow over the body [85,251]. These features manipulate the boundary layer, controlling separation and transition from laminar to turbulent flow, which are essential for reducing drag.
Body Shape for Drag Reduction Maybury reports in his dissertation that body drag reduction in birds is achieved through various means. Birds typically exhibit a cavity on the dorsal surface behind the neck and a rapid narrowing of the body at the root of the tail, creating trapped vortices in these regions. This manipulation of airflow serves to reduce drag. Additionally, the furled tail functions akin to a splitter plate and wedge, reducing overall drag by controlling the size of the separation bubble formed beneath the tail. The ventral covert feathers also play a crucial role in drag reduction by aiding in the motion control of trapped vortices [85,251].
The research highlighted the importance of considering variations in bird morphology and behavior in drag modeling. Past models tended to oversimplify bird body drag, not accounting for factors like laminar boundary layer conditions and the unique body shape of each bird species.
Parasite Drag and Its Components Parasite drag in birds, comprising profile drag and skin friction, is a fundamental aspect of avian aerodynamics. Profile drag, influenced by body shape, results from airflow changes around the body. Skin friction, impacted by surface area and texture, arises from viscous forces against the body surface. The study challenged traditional models of estimating parasite drag and suggests that previous models may overestimate this drag factor by a significant margin. It highlights the importance of considering body shape, airspeed, and Reynolds number in accurately assessing drag. Additionally, the interaction between the body and wings significantly influences drag reduction. For instance, the alignment of feathers, treated with materials like wax or hairspray, can alter the drag coefficient, demonstrating the impact of surface texture and body shape on aerodynamic performance [85].
The study hypothesized that body shape scaling among birds affects body drag, with larger birds experiencing greater effects of boundary layer separation. Drag measurements correlated strongly with bird body mass, trunk length, minimum frontal projected area, and trunk fineness ratio. This indicated that multiple regression models would be highly effective in explaining the measured variation in drag and drag coefficient. Experimental treatments applied to European starlings revealed that drag coefficient based on frontal projected area decreased with airspeed. The lowest drag coefficients were observed in birds with rigid or smooth body surfaces. However, values ranging from 0.36 at lower speeds to 0.24 at higher speeds were considered representative of live birds in forward flight [85].
Feathers play a significant role in aerodynamics. Birds can control feather alignment to optimize boundary layer flow, potentially reducing drag. This active control mechanism is critical for maintaining aerodynamic efficiency during flight. Treatments like hairspray or wax were used to improve feather alignment, which significantly reduced the drag measured on frozen specimens. It was acknowledged that these procedures might overestimate drag by up to 32%, considering the challenges in achieving optimum posture and alignment [85].
Maybury contributed significantly to the understanding of bird body aerodynamics and enabled more accurate modeling of avian gliding and flapping flight. The work provided reliable estimates of body drag for small birds, essential for aerodynamic flight performance modeling in MAVs. The study used flow visualization techniques and controlled feather alignment to examine the aerodynamic properties of the boundary layer in various bird species [85].
Boundary Layer Dynamics The nature of the boundary layer, whether laminar or turbulent, significantly affects body drag. Turbulent boundary layers can reduce body drag by maintaining airflow attachment along the body’s rear, altering pressure distribution, and reducing total drag [85]. Maybury’s 2001 work aimed to understand how the tail, even when furled, significantly impacts body drag, a factor not previously considered in avian aerodynamics. It combined measurements of body drag and boundary layer dynamic pressure with flow visualization observations.
The experiment used mounted specimens of European starlings, focusing on the manipulation of their tails. Drag was measured using strain gauges, and the effect of tail size on drag coefficient was studied at various airspeeds. The study utilized two experimental protocols to systematically reduce tail size and examine its impact on body drag. The study found that the removal of the tail and covert feathers increased drag by 25% at a Reynolds number of 50,000, with a larger increase at lower Reynolds numbers. Each successive reduction in tail size led to an increased drag coefficient. This indicated that both the tail feathers and covert feathers significantly contribute to drag reduction by manipulating vortex shedding and body wake development [251].
Emperical Estimates of Body Drag In a similar study, Tucker aimed to accurately measure the body drag of a Harris’ hawk and a peregrine falcon. This was achieved by examining the drag in a wind tunnel on both a frozen, wingless body of a peregrine falcon and a smooth-surfaced model of that body. The study addressed technical challenges in measuring body drag and sought to provide accurate estimates for the body drag of birds similar in size and proportions to the falcon [252].
The study determined that the mean, minimum drag coefficients () for the frozen falcon body and the model were 0.24 and 0.14, respectively. These findings highlighted the significant effect of feathers on body drag, as the drag of the model was about 60% of that of the frozen body. Importantly, these values are lower than most other studies of avian body drag, which have reported estimates up to 0.41. Tucker underscored that the feathered surface contributed significantly to drag on the falcon body. The rougher surface of the feathered body, compared with the model, likely increased drag by thickening the boundary layer. The study also noted the importance of preening the feathers on the partially thawed body, adjusting body parts for minimum drag, and reducing and correcting for interference drag [252].
A critical aspect of the study was understanding and measuring interference drag, which arises from the air flow around a body mounted on a strut in a wind tunnel. This drag is not entirely corrected by subtracting the drag of the isolated strut, and the study provided techniques for measuring interference drag, indicating that corrections could reduce apparent body drag by more than 20% [252].
Pennycuick et al. also performed a similar study for large waterfowl and raptors to measure body frontal area and drag coefficients. It presented measurements on these two groups of birds, considering frontal area and drag coefficient as separate datasets. The study proposed relationships between frontal area and body mass, and between drag coefficient and Reynolds number [253].
The measurements indicated that the body frontal area of these birds varied with the two-thirds power of body mass. The wind tunnel measurements on frozen bodies provided drag coefficients ranging from 0.25 to 0.39 in the Reynolds number range of 145,000 to 462,000. These findings, combined with past studies, led to a practical rule for estimating the body drag coefficient in performance estimates. An intriguing observation, verified by Maybury, was that treating a snow goose with hair spray, to flatten the feathers, resulted in a 15% reduction in drag coefficient. This suggests that living birds might control their contour feathers to minimize drag during flight. The study highlighted the possible need to revise downwards the drag coefficient estimates for living birds based on observations of dead specimens. Pennycuick et al. also observed that all specimens exhibited a ’supercritical’ flow regime with turbulent boundary layers near the upstream end. The findings implied that the drag coefficients for large birds with well-streamlined bodies might be lower than previously thought, especially in the case of birds with features (e.g., long necks) that could increase drag [253].
Estimates of Body Drag During Dives
The study by Hedenström and Liechti aimed to measure the maximum dive speeds of passerine birds during their spring migration across the western Mediterranean. The focus was to estimate the body drag coefficient () during flight. Tracking radar was employed to measure the birds’ dives, with the assumption that when the speed reached a constant value, drag balanced the pull of gravity. The study calculated the total lift and drag for each bird track using established aerodynamic equations, and from these, the body drag coefficient was estimated. The work also discussed that a lower mean value from the determinations should not be ruled out as diving birds may control their speed below the theoretical maximum [61].
The mean estimated was found to be 0.37, with a range from 0.17 to 0.77. This study supports the notion that a default value of 0.4 is realistic for small passerines. The findings indicated that decreases with increasing Reynolds number within this range, suggesting that previous wind tunnel measurements using dead, frozen bird bodies or models might have overestimated . The aerodynamic performance of passerine birds, as indicated by this study, allows them to migrate long distances efficiently. Changes in values have significant implications for our understanding of flight performance, potentially increasing calculated flight ranges and characteristic flight speeds
Hydrodynamics in Diving Birds
, Although not necessarily critical for most aspects of flight, the consideration of hydrodynamic drag can still be compared to flight efficiency in air through similitude. In diving birds, hydrodynamic drag, influenced by body size, shape, and feather properties, becomes a primary factor. Lovvorn et al. aimed to investigate the effects of size, shape, and feathers on the hydrodynamic drag of avian divers. The study used a novel towing method without posts or strings that alter flow or angles of attack, allowing for a more accurate measurement of drag on birds. The researchers compared the hydrodynamic drag of models and frozen birds to examine the interaction between body shape and feather effects [83].
The research found that feathers can increase the drag of frozen birds by two to sixfold over that of cast models or other featherless bird shapes. This indicates that the roughness and vibration of feathers are significant factors in drag measurements. The magnitude of this ’feather factor’ varied with both speed and species, showing greater effects in foot-propelled divers that swim with their necks extended. The study demonstrated that differences in body shape among bird species had strong effects on patterns of change in drag with speed. For example, in auklets, minor variations in shape caused dramatic changes in drag, and these shape differences were not consistently related to size. This finding suggests that extrapolation of dimensionless CD/Re curves among ’similar’ species must be performed with caution. For larger birds, CD/Re curves varied consistently between wing-propelled divers with short or retracted necks and foot-propelled divers that swim with their neck extended. The study highlighted the need to consider species variations in drag at different speeds in the context of accelerational stroking. This is particularly relevant for birds that experience a range of instantaneous speeds during oscillatory strokes [83].
Experimental Challenges in Aerodynamics Research
The collective insights from this series of papers on avian body drag highlight the complexity and challenges inherent in both experimental and computational research in this field. One fundamental challenge is accurately capturing the influence of feathers on drag where feathers increased drag by two to sixfold compared to models. This underscores the difficulty in replicating the intricate properties of feathers and their interaction with fluid dynamics. Similarly, feather treatment has a substantial impact, such as smoothing with hair spray, on drag measurements, indicating the sensitivity of drag to minor variations in surface texture. Another challenge is the influence of body shape and size on drag, as different species exhibit varied drag patterns at different speeds. This complexity makes it challenging to develop accurate computational models, as slight variations in shape, which might not be apparent without detailed measurements, can lead to significant errors in drag estimation. These challenges are further compounded by the need to consider different body postures, as observed in various studies. For instance, Pennycuick and Lovvorn emphasize the varying effects of body shape and swimming mode on drag, necessitating careful consideration of these factors in both experimental setups and computational modeling. Collectively, these studies illustrate the multifaceted challenges in accurately modeling and measuring the hydrodynamic drag in avian species, necessitating a nuanced approach that accounts for the complex interplay of feathers, body shape, size, and behavior [61,83,85,251,252,253].
2.1.6. Beak Shape
The morphology of bird beaks has long been a subject of fascination in evolutionary biology, however, there have been very few studies investigating the aerodynamics of bird beaks and how the morphology can serve to improve efficiency. Traditionally, the diverse shapes of bird beaks have been attributed to dietary adaptations, feeding behaviors, and, by extension, ecological niches. However, recent research has begun to unravel a more complex picture, suggesting that non-dietary factors significantly influence beak morphology. This review aims to synthesize findings from key studies to understand better the drivers of beak shape evolution and its implications for aerodynamic performance.
Morphological and Ecological Correlations
Feeding Ecology and Beak Shape While beak shape has been commonly linked to feeding ecology, recent studies indicate that this relationship may not be as straightforward as previously thought. Navalon et al. observed a significant but limited correlation between beak morphology and feeding ecology across modern birds. Despite a discernible link, the variation in feeding ecology accounted for a relatively small portion of the observed variation in beak morphology, suggesting other influential factors [91]. Similarly, Van Wassenbergh and Baeckens’ study found that while there is a significant link between beak morphology and feeding ecology, much of the variation in beak morphology remains unexplained by feeding ecology alone. They suggested the lack of correlation with feeding ecology might be due to the multitude of nonfeeding functions of the beak, like singing, preening, fighting, and thermoregulation, which also influence its evolution. The study also highlights that while feeding ecology plays a significant role in beak morphology diversification, the functional and ecological trade-offs in beak evolution are also critical. They stress the need for further biomechanical analyses of the cranial musculoskeletal system of birds to understand better how beak shape affects the efficiency of performing certain tasks, giving rise to the question of aerodynamics [118].
Non-Dietary Factors in Beak Evolution A morphometric analysis conducted by Bright et al. on raptors to test the effects of genetic and developmental histories on avian craniofacial skeleton revealed that beak and skull morphology in birds is highly integrated and strongly influenced by size. They found that the beak is not an independently targeted module for selection, but instead, highly integrated with the skull. This integration accounts for almost 80% of the shape variation observed between species, overshadowing the role of morphological dietary adaptation. The study emphasized that changes in beak morphology invariably lead to predictable alterations in braincase morphology and vice versa, underscoring a strong developmental constraint on avian craniofacial morphology [32].
Drag Reduction at the Air-Water Interface While not focused on flight efficiency, Crandell et al. have performed research work focused on the drag reduction of the kingfishers during their dive from air into water. The work investigates the morphological adaptations these birds have developed specifically for diving, particularly the beak morphology. The study compares the beak morphology of terrestrial and mixed-foraging kingfisher species, using 3D beak models from 71 different species. In addition, functional testing was performed on 31 species representing various foraging strategies and body morphology. Physical simulations of dives with 3D printed beak models and CFD analyses were conducted in the study, and the findings revealed that diving kingfishers possess narrower bills in the dorsoventral and sagittal planes, and longer bills when size-corrected. These morphological traits contribute to lower peak decelerations during water entry, indicating a more efficient and quicker penetration into the water compared to terrestrial and mixed-foraging species. The CFD analyses confirm these trends and highlight that the morphology between the beak and head is key in reducing drag in aquatic species.
Figure 14.
a & b) Examples of a 3D scanned common kingfisher head showing the general variables of interest and model used for CFD analysis and printing [196]. c) water velocity calculated from CFD for beaks in the anterior-posterior direction showing how the beak morphology plays a significant role in hydrodynamic entry [196]. d) An overview of the feeding ecology considering, diet classes, food acquisition methods, shape of the beak, and leverage of the jaw muscles [118]. e) Typical water entry experiment sequence and force measurement for a 90° wedge used to test beak water-air interface interactions [254]. f) Breakdown of various avian beak types based on diet [255]. All figures are reprinted with permission; please see the Appendix A.
Figure 14.
a & b) Examples of a 3D scanned common kingfisher head showing the general variables of interest and model used for CFD analysis and printing [196]. c) water velocity calculated from CFD for beaks in the anterior-posterior direction showing how the beak morphology plays a significant role in hydrodynamic entry [196]. d) An overview of the feeding ecology considering, diet classes, food acquisition methods, shape of the beak, and leverage of the jaw muscles [118]. e) Typical water entry experiment sequence and force measurement for a 90° wedge used to test beak water-air interface interactions [254]. f) Breakdown of various avian beak types based on diet [255]. All figures are reprinted with permission; please see the Appendix A.
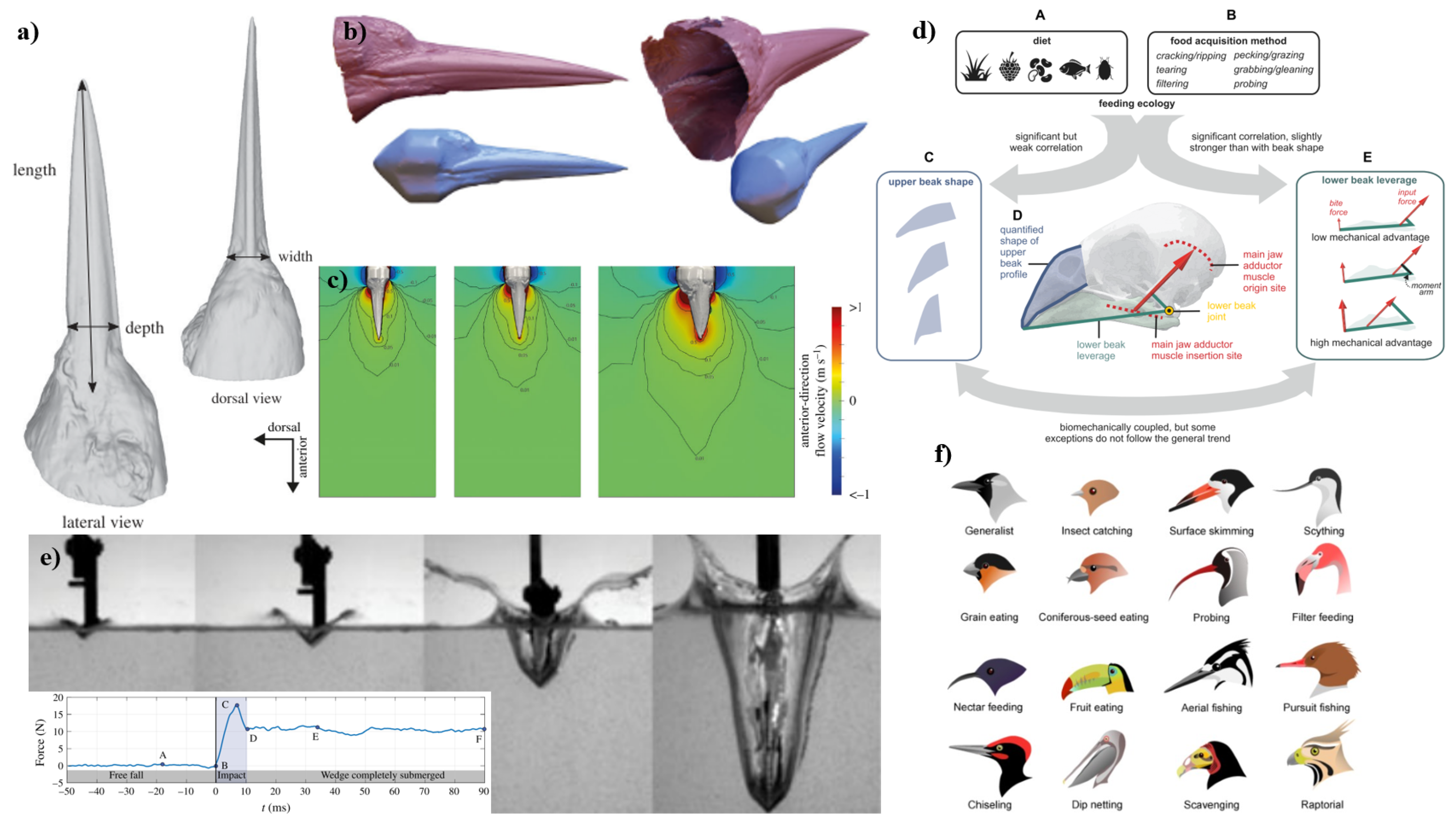
The study demonstrates that beak width in aquatically foraging species is less than in terrestrially foraging species, and although length and depth also differ between foraging groups, these differences are not significant once phylogeny is considered. Overall, the study provides evidence supporting adaptations for improved diving performance in aquatically foraging kingfishers relative to terrestrial and mixed-foraging species, suggesting a repeated evolution of functionally and morphologically more hydrodynamic beaks across the kingfisher phylogeny [196]. Similar studies have been conducted to observe the hydrodynamics of bird species that plunge dive without injury [254,256,257,258]. This is a promising result for the notion of morphological beak evolution for improved aerodynamics as well. Additionally, these bird beaks, along with several others were studied for drag optimization of the Southern Corridor High-Speed Rail, where they found a drag reduction in bio-based designs over the current design considerations [259].
Implications for Aerodynamic Performance The findings from these studies collectively suggest that beak shape and feeding ecology may not be as tightly woven together as previously thought, indicating additional factors may significantly contribute to the development of beak shape, including aerodynamic efficiency. While beak shapes are undoubtedly critical for various functions, including feeding, their evolution appears to be constrained by factors beyond ecological adaptation, such as size, integration with the skull, and genetic and developmental histories. This also implies that the aerodynamic performance of different beak shapes might be a by-product of these other factors rather than a direct evolutionary target, although there has been very little research into this topic as of now. Future studies should focus on the inclusion of aerodynamic analyses, integrated with biomechanical analysis and morphological data to further investigate the multifaceted drivers of beak shape evolution.
2.1.7. Color of the Feathers
The study of bird coloration has traditionally centered on its role in camouflage and mating, with plumage hues and patterns often interpreted as evolutionary adaptations for survival and reproductive success. Bright and conspicuous colors have been linked to mate attraction (Figure 15, while more subdued tones are associated with concealment from predators (Figure 16. This focus has led to a rich understanding of the ecological and behavioral implications of avian coloration. However, recent research has begun to unveil a more complex narrative, suggesting that these vibrant displays serve functions beyond the realms of mating and camouflage. Emerging studies are exploring the intriguing possibility that coloration, particularly in the context of wing patterns, might play a pivotal role in aerodynamic efficiency. This new line of inquiry represents a paradigm shift, extending the significance of avian coloration from a purely biological and ecological standpoint to one that intersects with the principles of physics and aerodynamics.
Avian Wing Coloration and Flight Efficiency
The intricate relationship between avian wing coloration and flight dynamics is a multifaceted subject that has garnered significant interest as of late in the field of aerodynamics and biomimetics. A series of studies have delved into various aspects of this relationship, revealing the complex interplay between coloration and flight mechanics.
Dark Wing Pigmentation and Flight Efficiency in the Larinae Goumas explored the influence of dark wing pigmentation on flight efficiency in the Larinae (gulls). His work hypothesized that the dark upperside of gulls’ wings, due to its heat-absorbing properties, reduces drag by decreasing air density around the wings. This theory was backed by empirical data showing a positive correlation between wing loading and both mantle darkness and the proportion of black on wingtips. Species with higher wing loading tend to have darker mantles and wingtips, suggesting an evolutionary adaptation for balancing maneuverability with long-distance flight efficiency. The study also considered factors beyond thermoregulation, such as species recognition and sexual selection, in the evolution of dark wing pigmentation [52].
Effects of Birds’ Wing Color on Flight Performance for Biomimetics
Hassanalian et al. investigated the thermal effects of birds’ wing color on their flight performance, particularly in large migrating birds like the albatross. The research showed that the dark coloration of the upper wing surfaces increases the temperature of the wing’s boundary layer, reducing drag forces and enhancing flight endurance, serving as additional evidence toward Goumas’ conclusions. This understanding is vital for biomimetic applications, especially in designing drones with enhanced endurance, mimicking the efficient flight mechanics of these birds [57].
In another study by Hassanalian, the research examined the thermal impacts of wing colorization of migrating birds on their skin friction drag and the choice of flight routes. This research focused on four species of birds, considering ocean flux and solar irradiance during migration. The findings indicated that the black on the top side of the wings and white on the bottom side (counter-shading) results in skin friction drag reduction, enhancing their flight endurance. This drag reduction is, of course, influenced by day and night conditions, suggesting that migratory birds may select flight routes that optimize conditions for reduced drag. The study also conducted a sensitivity analysis on the impact of sea surface temperature on wing performance, finding a strong correlation between optimal wing color arrangement [59].
Lastly, Hassanalian et al. focused on the thermal impact of migrating birds’ wing color on their flight performance. Observing large migrating birds like the albatross, the study once again found that the dark color on the top of the wing increases the boundary layer’s temperature over the wing, consequently reducing the skin drag force. This reduction in drag is an effective factor for the long endurance observed in migrating birds. The study aims to apply these findings to drone design, proposing efficient color configurations that could increase the endurance of drones by mimicking the color patterns of bird wings [237].
Thermal Impacts of Wing Coloration on Surface Temperature
Rogalla et al. conducted a similar study on the thermal impacts of wing coloration on surface temperature during bird flight. They found that darker wings heated up faster under solar radiation than lighter wings, potentially reducing skin friction drag and improving flight efficiency, corroborating Hassanalian et al.’s studies. This work used a combination of field and laboratory tests, including thermal imaging of live ospreys and wind tunnel experiments with differently colored wings. The findings revealed that temperature differentials on wings could produce convective currents towards the darker areas, increasing lift, and delaying flow separation above the wing, thereby increasing flight efficiency [105].
Rogalla et al. further explored the evolution of darker wings in seabirds concerning temperature-dependent flight efficiency. The study used phylogenetic analyses and wind tunnel experiments to investigate the relationship between darker wing pigmentation and flight efficiency. The analysis revealed that birds with higher glide performance have evolved darker wings, indicating an adaptation for improved flight under challenging oceanic conditions. The wind tunnel experiments showed that radiative heating of bird wings enhances flight efficiency, with both lift and drag decreasing under heated conditions, resulting in a higher lift-to-drag ratio [106].
Bioinspired Albatross Airfoil with Heated Top Wing Surface
Pellerito et al. [97] experimentally investigated the effects of heated wing surfaces on the aerodynamic performance of airfoils, inspired by the natural flight mechanisms of the albatross. The study showed that the surface temperature of black bird wings was significantly higher than white wings under solar radiation. An experimental setup with a bio-inspired albatross airfoil demonstrated that heating the top surface of the airfoil by 10°C decreased the drag coefficient by up to 60% and increased the lift coefficient by up to 70% at specific angles of attack. These findings suggest a novel approach to increasing flight efficiency in fixed-wing unmanned aerial vehicles by utilizing thermal effects.
Effects of Color Patterns Ahmed et al. present a new, but similar, theory stating that a bird’s dark color not only reduces drag but also improves overall flight performance. It posits that each color pattern leads to different types of flight performance improvement. Examples of these color patterns can be seen in Figure 17 and Figure 18. This is attributed to variations in hot and cold surfaces on the bird’s skin resulting from the interplay of light and dark feather colors. The research involved thermal imaging of real bird wings under infrared waves and testing a novel wind tunnel wing that could adjust the surface temperature in specific patterns. This setup aimed to evaluate the aerodynamic forces as a function of surface temperature and the hot-cold regions. Data from this experiment showed potential flight efficiency improvements of up to 20%, comparing the lift-to-drag ratio for specific heating cases [20].
The study revealed that certain heating cases could increase lift at low angles of attack, beneficial in cruise flight performance. The experimental results indicated that individual considerations of lift and drag in specific heating cases could improve these parameters by up to 20% and 7%, respectively. Additionally, significant reductions in drag at high angles of attack were noted when the middle regions of the wings were heated. The work also discussed how color patterns create unique flight characteristics due to the fluid-thermal interface influenced by temperature differences between black-and-white portions of the wing. The variations in these patterns were computationally modeled and experimentally verified in wind tunnel measurements. The findings indicate that the emissivity effects of different color patterns on a bird’s wings play a major role in flight efficiency. The observed improvements in aerodynamic performance due to these color patterns are seen as evolutionary adaptations that provide increased performance for bird species utilizing specific flight characteristics. This research holds biomimetic potential, suggesting that similar adaptations could be applied to engineered flying systems for enhanced efficiency [20].

Table 5.
Characteristics of different birds with black and white color for general comparison collected by Aboelezz et al., [20,24,25,27,33,34,35,36,38,39,40,41,42,44,45,47,48,49,50,51,53,54,56,64,67,68,70,71,72,74,75,78,79,87,94,98,102,104,107,109,111,112,119,120,121,122].
| Bird |
Weight (g) |
Wingspan (cm) |
Body length (cm) |
Aspect ratio |
Wing loading (N/m2) |
Flapping frequency (Hz) |
Max flight altitude (m) |
| Alpine Chough |
188–252 | 75–85 | 37–39 | 6.40 | 21.8 | - | 8,000 |
| Whooper Swan |
8500-10000 | 218-243 | 145-160 | 8.7 | 175 | 3.44 | 8,200 |
| Avocets | 260-290 | 77-80 | 42-45 | - | - | - | 3,000 |
| Yellow-billed Magpie |
150-170 | 61 | 43-54 | - | - | - | - |
| Pied Crow | 520 | 328-388 | 46-52 | - | - | - | - |
| Australian Magpie |
220-350 | 65-85 | 37-43 | 3.1 | - | - | - |
| Great Hornbill |
2000-4000 | 152 | 95-130 | - | - | - | - |
| Black-headed Ibis |
1100-1400 | 130 | 65-76 | - | - | - | - |
| Wandering Albatross |
5,900-12,700 | 250-350 | 107-135 | 15.6 | 150 | - | - |
| Manx Shearwater |
350-575 | 76-89 | 30-38 | - | - | - | - |
| White stork | 2300-4500 | 155-215 | 110-125 | 7.2 | 63 | - | 4,800 |
| Swallow-tailed Kite |
310-600 | 112-136 | 50-68 | - | - | - | - |
| Bearded Vulture |
4500-7800 | 231-283 | 94-125 | 8.9 | 79 | - | 7,300 |
| Bar-headed Goose |
2000-3000 | 140-160 | 68-78 | 8.5 | 92 | 3.75 | 8,800 |
| Common Crane |
4600-5400 | 180-240 | 100-130 | - | - | - | 10,000 |
| Andean Condor |
10100-12500 | 283-330 | 100-130 | - | - | - | 6,500 |
| Rüppell’s Vulture |
6400-9000 | 226-260 | 85-103 | - | - | 3 | 11,300 |
| Snow Goose |
2050-4500 | 135-165 | 64-79 | - | - | - | 3,050 |
| Whooping Crane |
4500-8500 | 200-230 | 132 | 4.1 | 50 | - | 950 |
| Indian Paradise Flycatcher |
20-22 | 86-92 | 19-22 | - | - | - | - |
| White-browed Wagtail |
30-36 | - | 21 | - | - | - | - |
| Hoopoe | 46-89 | 44-48 | 25-32 | - | - | - | - |
| Black and White Warbler |
8-15 | 18-22 | 11-13 | - | - | - | - |
| Great Shearwater |
670-995 | 105-122 | 43-51 | - | - | - | - |
| Magpie Lark | 64-118 | - | 25-30 | - | - | - | - |
2.1.8. Concluding Summary of Structural and Physical Characteristics of Avian
This review has extensively explored the structural and physical characteristics of avian species, revealing a multitude of adaptations that significantly contribute to their flight efficiency and maneuverability. Central to this discussion is the examination of wing shape and aerodynamics, highlighting the remarkable benefits of morphing wing capabilities. The dynamic nature of avian wing morphology underscores the potential for adaptable and efficient design principles in aerospace engineering.
Further, the airfoil geometries inherent to bird wings offer critical insights into lift and drag management, a fundamental aspect of aerodynamic performance. The intricate designs of wingtips and winglets, crucial in minimizing induced drag, present valuable lessons for enhancing the aerodynamic efficiency of engineered flight systems. Additionally, the detailed study of feather micro-structures, particularly riblets, emphasizes their role in reducing skin friction drag, an area ripe for application in aircraft design.
It should be noted that producing vortices by some of the above-mentioned structural and physical characteristics can generally increase drag due to added viscous effects, but their overall impact can be drag reducing by mitigating pressure drag through flow separation control. Vortex generators, for instance, introduce small vortices that mix higher-energy free-stream air with the slower boundary layer air, thereby delaying flow separation and reducing pressure drag. Similarly, riblets create streamwise vortices that help maintain attached flow over a surface, significantly reducing pressure drag. Furthermore, leading-edge vortices (LEVs) on delta wings stabilize airflow and reduce flow separation, enhancing aerodynamic performance. Thus, while vortices inherently increase drag initially, their strategic production and placement can lead to an overall reduction in drag by controlling and delaying flow separation.
The material properties of feathers, encompassing both stiffness and flexibility, are pivotal in the control and efficiency of avian flight. This understanding prompts a reevaluation of material selection and utilization in aerospace applications. Moreover, the overall body shape and the diversity in beak forms of birds are not merely physiological traits but also critical factors in their aerodynamic performance and maneuverability, reflecting their ecological niches and lifestyles. There are currently few research studies discussing the effects of beak and body shape on the aerodynamic performance of birds and leaves room for future works.
Lastly, the intriguing aspect of feather coloration and its influence on the thermal boundary layer offers a unique perspective on color-related efficiency gains in flight. This opens up a novel area of research, potentially leading to innovative approaches in thermal management in aerospace technology.
In sum, the structural and physical characteristics of avian species provide a plethora of design principles and adaptations that are invaluable to the field of aerospace engineering. By emulating these natural marvels, there lies the potential to revolutionize the design and efficiency of future aerial vehicles, making significant strides in drag reduction, lift enhancement, and maneuverability. The ongoing research and exploration in this domain promise to unveil further insights, potentially leading to groundbreaking advancements in aerospace technology.
2.2. Flight Modes and Features
2.2.1. Gliding Flight
In several different favorable conditions, birds have learned to use various modes of gliding flight to save energy without sacrificing flight speed or altitude in many cases. In Still air, gliding can still save energy, however, it comes at the cost of either speed or altitude. While encountering other unique wind phenomena, more complex and complicated possibilities arise to use gliding for prolonged durations and distances.
Hang-Gliding
Hang-gliding is likely one of the most common forms of gliding flight and can often be observed in our daily lives, both by birds and people. With the inspiration from avian efficiency and maneuverability, people developed the sport of hang-gliding by attempting to mimic the observed phenomena.
Exploitation of Up-Draughts in Avian Hang-Gliding Birds, particularly species such as kestrels and ospreys, have evolved remarkable skills in hang-gliding, utilizing up-draughts created by natural landscape features like hills, cliffs, and waves. These up-draughts redirect airflow upwards, allowing birds to maintain altitude with minimal wing-flapping. A study in the Netherlands on kestrels hang-gliding over a sea dike observed that these birds prefer positions about 6.5 meters above the windward slope in specific wind conditions. The upward wind angle in these preferred positions ranged between 6° and 7°, correlating with a glide ratio comfortably below the range found in controlled experiments, demonstrating the birds’ adeptness in optimizing flight parameters for energy efficiency [266]. These findings are supported by the work of Harvey and Inman, which further elaborates on the control challenges faced by birds and UAVs in turbulent atmospheric conditions [267]. This behavior is a manifestation of their ability to adapt to and exploit varying wind conditions, which also includes maintaining stable flight in turbulent atmospheric wind [150,267]. This adaptability and efficiency in energy use, evident in avian flight, are crucial considerations in the design of UAVs, particularly when aiming to mimic these natural flight strategies in varying and challenging atmospheric conditions.
Figure 19.
a) Andean Condor banking during gliding flight. Photo credit: Andy Reago & Chrissy McClarren. b) Great frigatebird gliding. Photo credit: Patrick Kavanagh. c) A golden eagle soaring on wind, Photo credit: Dona Hilkey. d) A small American white pelican flock soaring in V-formation. Photo credit: Claudio Contreras Koob. e) A turkey vulture gliding low to the ground to make use of its keen sense of smell. Photo credit: John Comisky. f) Gliding white stork. Photo credit: Luiz Lapa. g) Manx shearwater engaging in dynamic soaring over the ocean. Photo credit: Kirk Zufelt [268]. h) A white pelican in gliding flight above a body of water. Photo credit: Rui Ornelas [269]. i) A sooty tern in gliding flight. Photo credit: Stefan Hunt [270]. j) Bald eagle gliding low over the water. Photo credit: Andy Morffew. All figures are reprinted with permission; please see the Appendix A.
Figure 19.
a) Andean Condor banking during gliding flight. Photo credit: Andy Reago & Chrissy McClarren. b) Great frigatebird gliding. Photo credit: Patrick Kavanagh. c) A golden eagle soaring on wind, Photo credit: Dona Hilkey. d) A small American white pelican flock soaring in V-formation. Photo credit: Claudio Contreras Koob. e) A turkey vulture gliding low to the ground to make use of its keen sense of smell. Photo credit: John Comisky. f) Gliding white stork. Photo credit: Luiz Lapa. g) Manx shearwater engaging in dynamic soaring over the ocean. Photo credit: Kirk Zufelt [268]. h) A white pelican in gliding flight above a body of water. Photo credit: Rui Ornelas [269]. i) A sooty tern in gliding flight. Photo credit: Stefan Hunt [270]. j) Bald eagle gliding low over the water. Photo credit: Andy Morffew. All figures are reprinted with permission; please see the Appendix A.

Design Implications for Autonomous Aerial Vehicles The avian mastery of hang-gliding in varying wind conditions offers significant insights for the design of UAVs. Birds’ ability to maintain stable flight in turbulent conditions, as observed in species like kestrels and ospreys, suggests potential strategies for UAV stability in similar conditions [150]. The control mechanisms and sensing strategies employed by these birds could inform the development of UAVs capable of ultra-stable flight in turbulent atmospheric conditions. Moreover, the aerodynamic efficiency of birds in gliding flight, as compared to UAVs, indicates potential improvements in UAV design. Studies comparing the aerodynamic efficiency of gliding birds and UAVs reveal that birds’ wings are often cited as inspiration for efficient UAV wing designs [150]. The varying flight dynamics across different bird species, as observed in studies of Northern Bald Ibises during migratory flights, provide empirical evidence of the energy efficiency of intermittent gliding flight [239]. These insights could guide the development of UAVs that use similar intermittent gliding techniques to optimize energy use over long distances.
Dynamic Soaring in Avian Species
Dynamic soaring is a sophisticated flight technique often observed and studied in the albatross, enabling them to cover vast distances over the ocean with minimal wing flapping. Dynamic soaring will be discussed in additional detail in Section 4.1.1. This flight strategy is incredibly energy-efficient, allowing these birds to exploit natural wind gradients and updrafts over ocean waves [102]. Albatrosses utilize wind shear soaring and wave-slope soaring techniques, harnessing the vertical gradient of wind velocity and updrafts over waves. The energy gains from these methods are substantial, contributing significantly to the birds’ ability to maintain prolonged soaring [234]. A typical albatross feeding trip, for instance, may span over 10 days, covering approximately 1000 km per day [107]. These birds have adapted to exploit the energy available in the wind shear, particularly in regions known for strong and consistent winds like the ‘Roaring Forties’ and ‘Furious Fifties’ in the southern oceans [159].
The mathematical model developed by Denny simplifies the complex flight dynamics of albatrosses, illustrating the underlying physics of dynamic soaring while acknowledging the limitations of such a simplified approach. Despite these limitations, the model effectively captures the essence of dynamic soaring and is generally compatible with experimental observations. The large lift-to-drag ratios of albatrosses, their ability to maintain specific bank angles during turns, and the varying lengths of their dynamic soaring trajectories are key factors in their energy-efficient flight [234].
The flight dynamics of albatrosses involve complex maneuvers, including climbing upwind and then turning and descending downwind, effectively gaining energy from the wind’s velocity gradient. This is known as the Rayleigh cycle of wind-shear soaring, a concept first proposed by Lord Rayleigh in 1883 [102]. The Rayleigh cycle involves a series of semi-circular flight patterns, allowing the bird to gain energy by climbing into stronger winds at higher altitudes and then descending into lower wind speeds closer to the ocean surface. For additional information on dynamic soaring, please see the section in energy harvesting on dynamic soaring (Section 4.1.1).
Besides dynamic soaring, the Katzmayr effect is another critical factor in energy-saving flight that albatrosses might use [271]. This effect involves periodic changes in the angle of attack of an airfoil to enhance lift and reduce drag [272]. In wavy air conditions, the vertical component of wave-induced wind provides additional lift, which albatrosses can exploit by adjusting their wing angles while soaring over ocean waves. By harnessing these wind variations, they gain energy efficiently, enabling long-duration flights with minimal effort.
Implications for UAV Design The insights gleaned from the study of dynamic soaring in albatrosses have significant implications for the design and operation of UAVs, especially those intended for long-duration flights over vast areas, such as oceanic surveys. The ability of these birds to efficiently utilize wind energy for propulsion without the need for constant flapping can be translated into strategies for energy management in UAVs. Replicating the flight patterns of albatrosses, particularly their exploitation of wind gradients, could lead to the development of UAVs capable of prolonged autonomous flights with minimal energy expenditure [102,234,235,243]. In this context, Peter Lissaman’s contributions are essential, providing a quantitative analysis of energy extraction from wind variations [273]. In Lissaman’s work, the equations of motion are normalized using cruise speed and the acceleration of gravity, with a focus on the maximum lift-to-drag ratio as a critical parameter. His research includes various control schedules, illustrating how UAVs can perform energy-neutral cycles by appropriately adjusting the lift coefficient and bank angle. Furthermore, Lissaman’s models demonstrate how UAVs can mimic albatross flight patterns to achieve efficient, sustained travel by exploiting wind gradients. By incorporating these natural aerodynamic strategies, UAVs can achieve extended flight durations and operational efficiencies, enhancing their utility in environmental monitoring, maritime surveillance, and search-and-rescue operations.
2.2.2. Bounding Flight
Intermittent Flight Strategies in Birds
Intermittent flight, a remarkable aerodynamic strategy observed in birds, involves alternating between active flapping and passive phases. In gliding flight, birds optimize their trajectory to reduce energy expenditure, leveraging potential energy during ascent for subsequent gliding descent [236,274]. Bounding flight, predominantly seen in small birds with low aspect ratio wings, is characterized by cyclic variations in height, alternating between powered and passive phases as shown in the examples of Figure 20 and Figure 21 [241,275,276].
Bounding Flight: Kinematics and Aerodynamics Bounding flight in birds such as small passerines, parrots, and woodpeckers, showcases rhythmic variations in altitude and energy dynamics. During passive phases, birds fold their wings, leading to a unique undulating flight path. This pattern, involving energy loss to the air during passive phases and energy regain during active phases, serves as an energy-saving strategy [278,280].
Dynamic energy, combining kinetic and potential energies, fluctuates significantly in bounding flight. Theoretical models suggest that bounding flight can be more energy-efficient than steady-level flight at higher speeds, potentially offering up to 15% energy savings [241,279].
Muscle Activity During Intermittent Flight Studies of muscle activity during intermittent flight reveal that the pectoralis muscle is inactive during bounds and active during glides, indicating a reduction in metabolic cost during passive phases [277,280]. The variable activity of the supracoracoideus muscle across species during bounds further highlights the adaptability of muscle function in different flight styles. This muscle activity can be seen in Figure 20d.
In woodpeckers, variations in body mass and muscle composition, particularly the proportion of red and intermediate fibers in the pectoralis muscle, influence their ability to engage in flap-bounding flight. These findings challenge the notion that larger birds may be incapable of such flight patterns due to power constraints [280].
The adoption of intermittent flight patterns is influenced by flight speed and wing morphology as reported by Tobalske. For instance, zebra finches and other small birds with rounded wings primarily use bounding flight, while species with pointed wings exhibit a mix of bounding and gliding, particularly at higher speeds [279,280]. Bounding and undulating flights offer adaptive advantages beyond energy efficiency. These include optimization of flight performance and possibly enhanced evasive capabilities against predators as flight patterns become more difficult to predict [283].
Implications UAVs Keating’s review on bounding flight in birds discusses its potential application in UAV design. By emulating avian flight patterns, UAVs could achieve improvements in lift-to-drag ratios and handling qualities, particularly at higher speeds. This becomes especially relevant for flapping wing MAVs attempting to take advantage of various flight modes and the precise control that will be required to accurately and precisely enact them [284].
Research Directions and Future Applications The aerodynamic efficiency and energy-saving potential observed in bounding flight of birds present promising applications in UAV technology. Future UAV designs could incorporate adaptive wing mechanisms, such as stowable or variable-sweep wings, to optimize lift-to-drag ratios and maneuverability, similar to the wing folding observed in birds. These innovations could significantly enhance UAV performance, especially in challenging environmental conditions [284].
Developing autonomous control systems capable of replicating the intuitive adjustments seen in bounding flight is crucial. Such systems, coupled with advanced aerodynamic modeling and computational fluid dynamics simulations, would enable UAVs to efficiently navigate and operate in diverse atmospheric conditions. This research could extend well beyond the realm of MAVs and UAVs, influencing larger UAV designs and even robotic systems that utilize similar energy-efficient principles.
2.2.3. Hovering Flight
Hovering flight, characterized by an ability to maintain a fixed position in air, is particularly observed in bird species such as hummingbirds and certain sunbirds. This mode is employed by small passerines like the house sparrow for brief periods, mainly for tasks like nest approach or insect capture [236].
Kinematics and Morphology
Hovering flight kinematics, especially in hummingbirds, involves complex wing movement coordination. Birds generally orient their bodies vertically, with wingtips tracing a figure-eight motion. Wing beat frequencies vary across species, with notable differences in joint inversion during the backstroke [236,285]. The distinct morphology of hovering birds, such as elongated primary feathers and robust pectoral muscles, facilitates this complex flight mode [150].
Work by Vejdani et al. showed the substantial impact of wing kinematics and morphology on the performance of hovering flight. The study shows that long-axis wing rotation angle plays a crucial role in energy efficiency during hovering. For specific wing rotation amplitudes, there is an optimal flapping frequency for each stroke-plane orientation, closely corresponding to the wingbeat frequencies observed in diverse hummingbird species. This suggests that certain kinematic patterns are more energetically efficient and biologically plausible. It was determined that larger animals, like bats, require more power to maintain stable hovering due to their increased size and wing mass. This is particularly evident when comparing the hovering mechanisms of birds and bats, where the varying-wingspan mode becomes more feasible for achieving aerial station-keeping in larger animals. The study provides guidelines for relevant design limits for hovering flight vehicles, considering the trade-offs associated with different frequencies, flapping, and wing rotation angles. The key implication of this study is its relevance to the development of flapping robots capable of forward and hovering flight. The findings provide a foundation for designing aerial robots with low noise, high agility, and exceptional stability in unsteady environments. The research underscores the importance of considering the dynamics of body motion and aerodynamic force generation in designing effective flapping-wing robots [286].
Figure 22.
a) Schematic view of the wingtip path showing hummingbird hovering flight from both the top and side view [236]. b) A different schematic view showing the experimental setip to perform PIV analysis of birds hovering at a feeder. The left side of the image shows two different measurement planes used, while the right displays the expected wake structure to be observed [287]. c & d) PIV analysis images depicting the wake during (c) mid-downstroke and (d) upstroke [287]. e) Two more views of a hummingbird during upstroke and downstroke flapping during hovering flight. These images were illuminated using a laser sheet to perform PIV analysis [277]. f & g) Velocity vectors and vorticity fields displaying circulation on the wing of a hummingbird [277]. All figures are reprinted with permission; please see the Appendix A.
Figure 22.
a) Schematic view of the wingtip path showing hummingbird hovering flight from both the top and side view [236]. b) A different schematic view showing the experimental setip to perform PIV analysis of birds hovering at a feeder. The left side of the image shows two different measurement planes used, while the right displays the expected wake structure to be observed [287]. c & d) PIV analysis images depicting the wake during (c) mid-downstroke and (d) upstroke [287]. e) Two more views of a hummingbird during upstroke and downstroke flapping during hovering flight. These images were illuminated using a laser sheet to perform PIV analysis [277]. f & g) Velocity vectors and vorticity fields displaying circulation on the wing of a hummingbird [277]. All figures are reprinted with permission; please see the Appendix A.
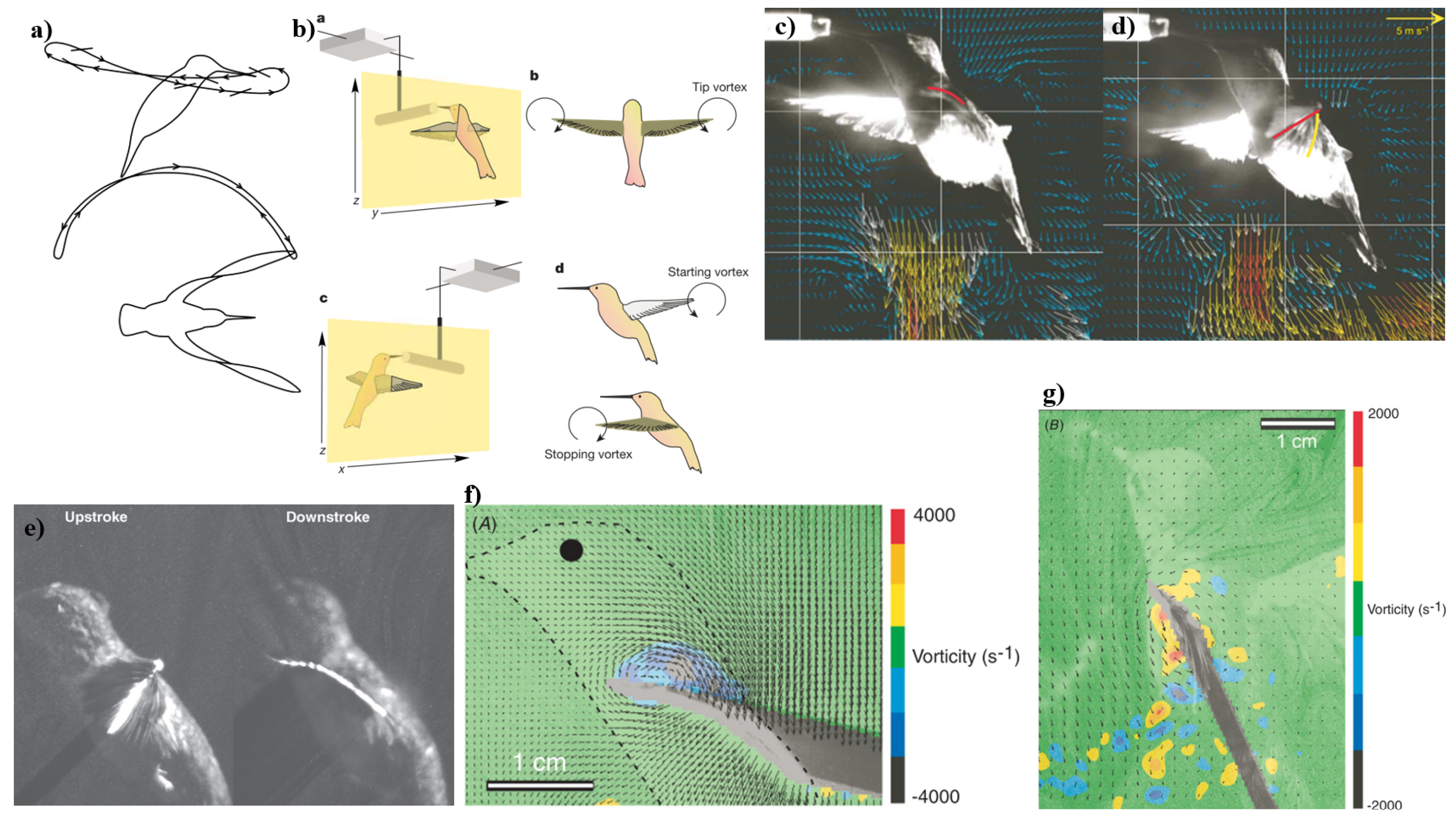
Energy and Efficiency Hovering is energy-intensive, requiring efficient wing joint rotation for maneuvering. Hummingbirds manage the high metabolic cost through nectar consumption, a high-energy food source, essential for sustaining this flight mode [236,288]. Studies have shown that the energy expenditure in hovering is significantly higher than in other flight modes, highlighting the evolutionary adaptations in these birds [289].
Maintaining a positive angle of attack during wing strokes is crucial in hovering. The interaction between the wings and air in hovering birds, especially regarding maximum wingtip velocities and aerodynamic control, is a subject of ongoing research, with quantitative flow visualization studies providing insights [287,290].
Hovering flight studies offer insights for engineering, especially in designing maneuverable and efficient aerial vehicles. Understanding the biomechanics and aerodynamics of hovering can inform small-scale drone development [290,291]. Further quantitative analysis of flow dynamics during hovering flight is necessary for understanding energy efficiency and aerodynamics, benefiting aerial robotics and biomimetic designs [239,290].
Hovering flight in birds demonstrates a complex interplay of biomechanics and aerodynamics. This flight mode, though energy-intensive, showcases evolutionary adaptations for efficiency and maneuverability, offering valuable insights for technology and engineering applications.
Wind-Hovering Flight
wind-hovering, distinct from hovering, is a technique where birds, such as various species of kestrels, maintain a stable position over the ground by flying against the wind at a speed that matches the wind’s velocity. This ability enables them to stay fixed in an earthbound frame of reference, which is particularly useful for hunting and observational purposes [285].
In wind-hovering, birds utilize the wind to counteract their forward motion, effectively achieving a stationary position relative to the ground. This technique requires precise adjustments in flight dynamics, where the bird must constantly adapt to the changing strength and direction of the wind to maintain its position. The stable position achieved in wind-hovering allows for detailed quantitative analysis of bird flight, as it offers a consistent frame of reference [285].
The aerodynamics of wind-hovering involve a strategic balance between the bird’s forward thrust and the opposing wind force. By aligning their body and adjusting their wing strokes against the wind, these birds can minimize the energy expenditure typically required for sustained flapping in hovering flight. This energy-efficient strategy is particularly advantageous for activities like hunting, where prolonged stationary observation is needed without expending excessive energy [150].
Implications for Avian Behavior and Flight Mechanics wind-hovering is a testament to the adaptability and precision of avian flight mechanics. It illustrates how birds have evolved to utilize natural elements like wind to their advantage, optimizing their energy expenditure while maintaining the ability to execute complex flight maneuvers. Studying wind-hovering can provide insights into the interplay between environmental factors and biomechanics in avian species [150,285].
Understanding the aerodynamics of wind-hovering in birds provides valuable insights for designing UAVs capable of stable flight in turbulent conditions. The glide angles and velocities observed in the wind-hovering kestrels, ranging from 7 to 12 m/s with glide angles of 6 to 7°, offer important data for replicating these conditions in UAV testing [150,266]. The methodology developed in this study opens avenues for further research into bird behavior, aerodynamic performance in gliding flight, and control responses to gusts and turbulence, which can significantly influence UAV design.
The study of wind-hovering behavior in birds offers valuable insights for the development of autonomous flying vehicles, especially those designed to operate in variable wind conditions. Understanding the mechanics and aerodynamics behind this flight mode could inform the design of drones and other aircraft capable of maintaining stability and efficiency in windy environments [150].
Further research is needed to explore the nuances of wind-hovering, particularly in how birds adjust their flight mechanics in response to varying wind speeds and directions. This could lead to a deeper understanding of flight stability and energy efficiency, potentially influencing the design of advanced aerial vehicles and systems [150,285].
Flight Adaptations and Efficiency
Migratory birds like the Brambling have evolved specific adaptations that enhance flight efficiency. These adaptations include features like prolonged and narrower wings, lower wing loading, and reduced wingbeat frequency and stroke amplitude, contributing to lower energy costs in flight compared to resident birds like the Eurasian Tree Sparrow. This efficiency is evident despite both species having comparable body mass, with studies showing that the Brambling has a lower maximum speed and power at any given speed, suggesting a strategy focused on energy minimization during flight [289].
Morphological Adaptations The morphological differences between migratory birds like the Brambling and resident species like the Eurasian Tree Sparrow highlight the evolutionary adaptations for energy-efficient, long-distance migration. The Brambling’s larger and longer wings, coupled with lower wing loading, enable reduced wingbeat frequency and stroke amplitude, essential for efficiency in long-journey flight. These adaptations allow for more energy-efficient flight at lower speeds, especially beneficial for migratory species [289].
The trade-off between flight speed and energy efficiency is critical, especially in migratory passerines like the Brambling. They exhibit higher flight energy efficiency, particularly at lower speed ranges, due to their adaptations in wing morphology and kinematics. In contrast, resident birds like the Eurasian Tree Sparrow, which do not undergo long migratory journeys, have adaptations favoring higher speed and maneuverability, useful for escaping predators or efficient foraging in diverse habitats [289].
Implications for Hovering and Windhovering Flight
Understanding the flight energetics and kinematics of different bird species, particularly the distinctions between migratory and resident birds, provides valuable insights into the biomechanics of hovering and wind-hovering flight. Migratory birds adapted for energy-efficient, long-distance travel might exhibit different hovering characteristics compared to resident birds, which may be more adept at quick, agile maneuvers. This differentiation is crucial in the context of developing and designing biomimetic flying robots or drones, where efficiency and maneuverability are key considerations [289].
2.2.4. Formation Flight
Formation flight, a phenomenon extensively observed in both avian species and aviation, showcases remarkable aerodynamic efficiency, leading to notable energy savings. Birds, especially larger species like geese and pelicans, demonstrate a strategic positioning in flight formations, often in a V-shaped or echelon arrangement. This behavior allows them to capitalize on the upwash effect from the bird in front, resulting in reduced drag and consequently lower energy expenditure [292,293,294]. Additionally, observations by Gould and Heppner on bird flock dynamics offer valuable perspectives on the formation behaviors in different species, contributing to a deeper understanding of the aerodynamic advantages these formations provide [295]. These insights have not only enhanced our understanding of avian flight mechanics but also inspired technological advancements in aviation, leading to novel approaches in aircraft design and operation to harness similar aerodynamic efficiencies [292,293,294,295].
Figure 23.
Various examples of birds flying in formation. a) Northern bald ibises flying in echelon formation flight [296]. b) Migrating white pelicans in formation flight. Photo credit: Don Henise [297]. c) Great white pelicans flying in an echelon formation over the surface of a river also employing the ground effect. Photo credit: R. Marzin, Galatee [298]. d) Geese migration flying in V-formation [299]. e) Eurasian cranes migrating in V-formation [300]. f) Snow geese flying in V-formation [301]. All figures are reprinted with permission; please see the Appendix A.
Figure 23.
Various examples of birds flying in formation. a) Northern bald ibises flying in echelon formation flight [296]. b) Migrating white pelicans in formation flight. Photo credit: Don Henise [297]. c) Great white pelicans flying in an echelon formation over the surface of a river also employing the ground effect. Photo credit: R. Marzin, Galatee [298]. d) Geese migration flying in V-formation [299]. e) Eurasian cranes migrating in V-formation [300]. f) Snow geese flying in V-formation [301]. All figures are reprinted with permission; please see the Appendix A.

Avian Based Evidence and Measurements
Studies on birds like pink-footed geese and great white pelicans provide indirect evidence of energy savings in formation flight. Research shows a reduction in heart rate and wing beat frequency during formation flight compared to solo flight, indicating significant energy savings [298,302].
Formation flight in birds illustrates a natural adaptation for energy efficiency, with evidence strongly supporting the energy-saving benefits of this behavior. This is particularly valuable for developing energy-efficient autonomous vehicles, where mimicking such natural formations could lead to significant advancements in power management.
Flight Formation in Large Birds Formation flight in large birds, such as pelicans, offers significant aerodynamic advantages. This behavior likely evolved to reduce flight power demands and energy expenditure when birds fly at optimal spacing. The study by Weimerskirch et al. focuses on the energy savings achieved by great white pelicans flying in a ’V’ formation [298].
Researchers observed pelicans in diagonal formation, which typically performed a few wing beats followed by a short glide, spending about 42.2% of their time flapping. The average wing-beat frequency decreased from the leader to the fourth position in the formation, indicating a reduction in individual energy expenditure [298].
Empirical evidence from the study indicates that formation flight confers a significant aerodynamic advantage, allowing birds to reduce their energy expenditure while maintaining similar speeds. In formation, each bird’s wing moves in an upwash field generated by the wings of other birds, leading to optimal spacing for maximal power reduction. The study’s findings suggest that pelicans flying in formation can achieve total energy savings of 11.4–14.0%, primarily by gliding for a greater proportion of their flight time [298].
The research by Weimerskirch et al. provides valuable insights into the aerodynamic benefits of formation flight in large birds. It highlights the importance of formation flight in reducing energy expenditure, underscoring its potential evolutionary advantage for group-migrating species like pelicans. The study’s findings contribute to our understanding of natural strategies for energy efficiency, which can be informative for the development of autonomous aerial vehicles and formation flying algorithms.
Energy Savings in Formation Flight of Pink-Footed Geese The study by Cutts and Speakman focused on pink-footed geese, analyzing wing-tip spacings in formation flight and the consequent induced power savings. The study highlights how optimal wing-tip spacing in formation flight reduces induced power requirements for birds, with geese achieving a 14% saving in induced power [302]. Geese save energy by flying in formation, although the mean observed spacing was below the theoretical maximum, suggesting difficulties in maintaining optimal positioning [302].
Wind conditions affect geese’s ability to maintain optimal formation positions. The study found a significant relationship between wind speed and depth spacing, indicating adjustments in varying wind conditions [302]. This work provides insights into the dynamics of formation flight in pink-footed geese, highlighting the aerodynamic benefits and challenges, particularly in varying wind conditions [302].
Induced Drag of a Bird Flock Higdon and Corrsin’s study focused on the induced drag in different bird flock formations, using mathematical models to analyze aerodynamic effects. The study explores how pressure differences across a bird’s wing generate lift and induce drag, particularly in flock configurations. Using a doublet-field strength model, the study approximates induced drag in different flock formations, indicating a decrease in induced drag in optimized formations. Different flock shapes significantly impact induced drag, with certain formations offering up to a 5% decrease in induced drag [303]. This work offers insights into optimizing formation shapes for reduced drag, applicable to natural bird flight and engineered flight systems.
Voelkl et al.’s study on Northern bald ibises examined the cooperative aspects of V-formation flight, exploring direct reciprocity in leading and following roles [304]. Similarly, Heppner et al. investigated the influence of visual angles on V-formation flight in Canadian geese, examining how visual perception impacts formation alignment [305].
The studies revealed a pattern of direct reciprocation in position-taking within formations, indicating a complex interplay of social and spatial factors [304,305]. Theoretical calculations by Voelkl et al. suggest that by flying in the upwash region at optimal wing-tip spacing, birds could save more than 50% of their energy costs relative to unaccompanied solo flight. The research revealed a strong correlation between the time a bird leads a formation and the time it benefits from flying in the wake of another bird. This finding indicates that birds actively seek positions in the wake of others and take turns leading. The majority of observed formations were pair formations consisting of only two individuals, suggesting that direct reciprocation in terms of mutual leading is key to solving the cooperation dilemma of formation flight. The size of formations varied, with pair formations consisting of only two individuals being the most common. This distribution closely matched the expected distribution for a model based on dyadic attraction, supporting the idea that direct reciprocity in formation flight is facilitated by frequent pairwise switches of the leading position. The study also found that the strength of dyadic ties during flight is not correlated with social proximity on the ground [304].
Formation Angles and Visual Alignment Heppner et al.’s analysis suggested that formation angles adopted by geese might be influenced by their visual tracking abilities, maintaining aerodynamic efficiency. An interesting focus point of the study is the detailed analysis of the visual field of Canadian geese. The researchers discovered that each eye of Canada Geese has a visual field of 135°, with a binocular overlap of 20° and a blind area at the back of the head of 29° on each side of the midline. These measurements suggest that when geese fly in a V-formation, each bird in the formation is not perfectly aligned along the optic axis of the bird following it. Instead, the formation allows for visual contact between birds without necessarily placing the leading bird within the sharpest focus of the following bird’s vision. Heppner et al. conducted geometric calculations to determine the angle of formation required for birds to be on the optic axis. The results indicated that for geese to fly so that each bird on a leg of the V-formation is on the visual axis of its following neighbor, the angle of the V should be around 128°. However, the observed angles of V-formation flying birds, such as Canada Geese, typically do not align with these calculated ideal angles. This suggests that other factors besides visual alignment, such as efficiency, might play a role in the formation structure [305].
Extended Formation Flight in Man-Made Aircraft
Ning’s foundational research delved into the realm of extended formation flight for man-made aircraft, presenting a shift from traditional aviation practices. This approach, inspired by the natural flight formations of birds, seeks to exploit the wake energy generated by leading aircraft to optimize drag reduction in trailing aircraft. Unlike conventional drafting techniques, this method emphasizes the strategic utilization of the upwash region, a critical element in achieving enhanced aerodynamic efficiency. The study suggests that, under realistic scenarios, formation flight can lead to up to a 13% reduction in fuel burn. Additionally, the formation types and arrangements play a critical role in optimizing drag reduction and ensuring safety. Extended formation flight, where aircraft are separated by larger distances, still retains much of the drag reduction benefits of close formation flight while enhancing safety [306].
The study explored the aerodynamic mechanisms underlying this drag-reduction strategy. Key to this is the precise positioning within the formation, which involves navigating the complex interplay between upwash benefits and the risks associated with downwash. An intriguing aspect of Ning’s research is the exploration of range scaling with formation size. The work presented diminishing returns in range improvement as more aircraft joined the formation. Ning’s research underscores the sensitivity of aircraft positioning, noting that optimal alignment is crucial for maximizing drag savings. Experimental work and computational studies have contributed to these insights. For example, wind-tunnel tests and flight tests have shown that the trailing aircraft in formation can reduce its power usage and achieve significant fuel flow reduction. The dissertation highlights various studies where drag reduction ranged from 8% to 18% in different scenarios. These findings are corroborated by computational models that predict similar levels of drag reduction in extended formations [306].
Figure 24.
a) Schematic diagram displaying the vertical velocity component induced by the wake of an aircraft [306]. b) View of a histogram-style plot showing the number of flaps occurring in the region between birds. Most of the flaps occurred at approximately 45° from the bird ahead. The dashed lines indicate the most favored positions [296]. c) A schematic showing long-track, cross-track, and vertical separation between two aircraft [307]. d) Schematic arrangement of wings in an arbitrarily shaped formation and approximation of the interference effects using single horseshoe vortices [292]. e) Formation flight example of two Dornier Do-28 aircraft [292]. f) Example of a novel flapping wing experiment to measure optimal overlap for flapping wing UAVs [308]. g) Another experimental setup for measuring optimal wing overlap in flapping formation flight using commercial systems [308]. h) Two F/A-18s performing a flight formation for NASA’s Autonomous Flight Project [306]. i) Relationship between wingtip spacing and depth from 393 paired pink-footed geese [302]. j) Wingtip spacing from 393 paired pink-footed geese [302]. k) Depth spacing from 393 paired pink-footed geese [302]. All figures are reprinted with permission; please see the Appendix A.
Figure 24.
a) Schematic diagram displaying the vertical velocity component induced by the wake of an aircraft [306]. b) View of a histogram-style plot showing the number of flaps occurring in the region between birds. Most of the flaps occurred at approximately 45° from the bird ahead. The dashed lines indicate the most favored positions [296]. c) A schematic showing long-track, cross-track, and vertical separation between two aircraft [307]. d) Schematic arrangement of wings in an arbitrarily shaped formation and approximation of the interference effects using single horseshoe vortices [292]. e) Formation flight example of two Dornier Do-28 aircraft [292]. f) Example of a novel flapping wing experiment to measure optimal overlap for flapping wing UAVs [308]. g) Another experimental setup for measuring optimal wing overlap in flapping formation flight using commercial systems [308]. h) Two F/A-18s performing a flight formation for NASA’s Autonomous Flight Project [306]. i) Relationship between wingtip spacing and depth from 393 paired pink-footed geese [302]. j) Wingtip spacing from 393 paired pink-footed geese [302]. k) Depth spacing from 393 paired pink-footed geese [302]. All figures are reprinted with permission; please see the Appendix A.
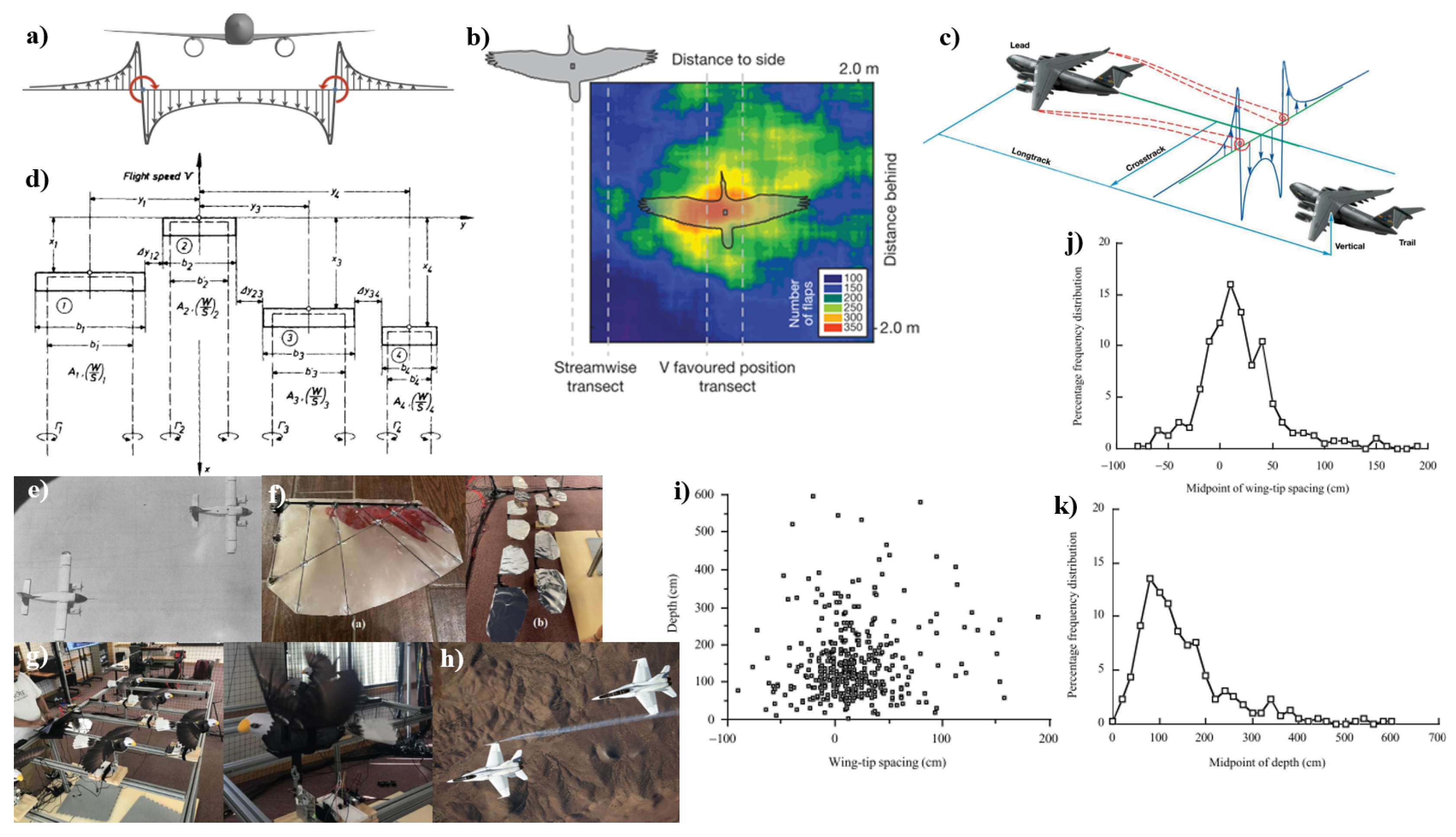
The study underscores that extended formation flight is a viable method for reducing drag in commercial, cargo, and military aircraft. This approach can be integrated into existing aircraft without the need for significant redesigns. The dissertation also discusses the potential for optimizing aircraft design specifically for formation flight, which could further enhance efficiency [306].
Formation Flight in Large Transport-Class Aircraft Formation flight in aircraft, inspired by avian species, offers similarly significant aerodynamic benefits. Pahle et al. discuss various studies that suggest potential drag reduction in the trailing aircraft by 15-28%, leading to considerable fuel savings, a concept particularly pertinent for large transport-class aircraft. Investigating these theoretical values, experiments with C-17 aircraft were performed and showed significant power and fuel flow reductions in formation flight. The experimental trials conducted by Pahle et al. demonstrated maximum average fuel flow reductions between 6.8% and 7.8%, and maximum average thrust reductions of 9.2%, compared to solo flight conditions. These reductions were most pronounced when the trail aircraft flew within the vortex area of influence generated by the lead aircraft. The flight tests were conducted at a single flight condition of 275 knots at 25,000 ft above mean sea level. The test points within the vortex influence of the lead aircraft were compared with tared test points outside the vortex influence. The pilots were able to fly the formation test points, but the workload was high, indicating that while formation flight can offer aerodynamic benefits, it requires careful consideration of operational feasibility and safety [307].
Despite potential benefits, formation flight poses considerable operational challenges, such as increased piloting difficulty and workload, especially in maintaining proximity to the vortex core, also discussed by Ning [306,307]. These experiments revealed increased turbulence and complexity in piloting as the aircraft drew closer to the vortex core, underscoring the need for advanced automated systems [307].
Flocking in UAVs Lambach’s thesis investigates the potential of UAV formation flying for drag reduction, emphasizing precision in position maintenance. The challenge lies in creating a UAV flock that efficiently reduces drag while avoiding collisions. The study focuses on the development of control methodologies that enable a flock of UAVs to fly in a V-formation, particularly during certain flight phases. MATLAB simulations tested various flocking strategies, demonstrating potential drag reduction and increased endurance in UAV formation flight. The simulation aims to navigate a flock through a set of waypoints while maintaining the appropriate formation position. The results from the simulation indicate that the formation geometric center control methodology is effective in reducing formation instabilities and enables the group to respond intelligently to positional disturbances. The research assumed effective two-dimensional management translates to three-dimensional operations, simplifying collision avoidance [309]. Lambach’s study provides a foundation for UAV flocking dynamics for drag reduction, highlighting both potential benefits and challenges.
2.2.5. Leader Switching in Flock
The dynamics of leader switching in formation flight, particularly in avian species, presents a fascinating intersection of aerodynamics, energy management, and evolutionary biology. This phenomenon, observed in various bird species during migratory flights, involves complex behavioral and aerodynamic interactions that lead to significant energy savings and flight efficiency. Recent studies have delved into the mechanics and implications of this behavior, exploring its benefits for both natural and engineered flight systems. This section reviews key findings from recent research, shedding light on the intricacies and advantages of leader-follower dynamics in V-shaped flight formations.
Figure 25.
Figures showing the concept of leader switching in formation flight. a) View of the major migration routes undertaken by Canadian geese each year [310]. These routes are often shared by other migratory birds such as the snow goose, sandhill crane, ducks, and surprisingly monarch butterflies. b) Schematic view of the upwash and downwash effects experienced during formation flight [310]. c) Example of leader/follower switching to conserve energy [310]. d & e) Schematic views of birds flying in formation flight with relevant measurement parameters for energy saving calculations [310,311]. f) View of the modeled drag reduction percentage with respect to 2s/b (the corresponding measurements are shown in d & e) for varying numbers of drones in formation [311]. g) A view of the total covered distance of a 14-bird Canadian goose flock with and without switching [310]. All figures are reprinted with permission; please see the Appendix A.
Figure 25.
Figures showing the concept of leader switching in formation flight. a) View of the major migration routes undertaken by Canadian geese each year [310]. These routes are often shared by other migratory birds such as the snow goose, sandhill crane, ducks, and surprisingly monarch butterflies. b) Schematic view of the upwash and downwash effects experienced during formation flight [310]. c) Example of leader/follower switching to conserve energy [310]. d & e) Schematic views of birds flying in formation flight with relevant measurement parameters for energy saving calculations [310,311]. f) View of the modeled drag reduction percentage with respect to 2s/b (the corresponding measurements are shown in d & e) for varying numbers of drones in formation [311]. g) A view of the total covered distance of a 14-bird Canadian goose flock with and without switching [310]. All figures are reprinted with permission; please see the Appendix A.
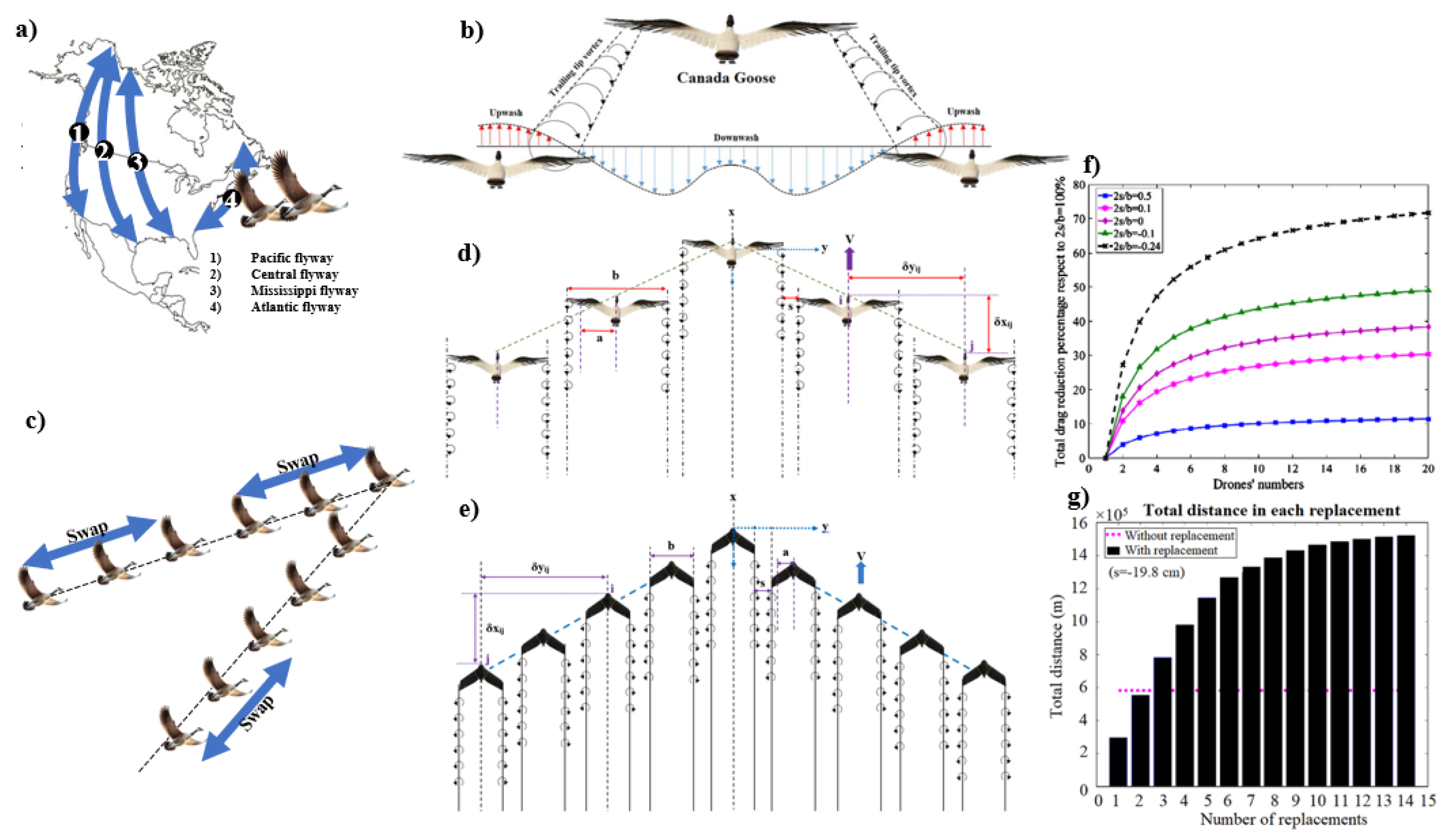
Leader-Follower Dynamics in V-Shaped Flight Formations
Mirzaeinia et al. presented a detailed analysis of energy efficiency in Canadian geese flocks, emphasizing the role of leader and follower switching in V-shaped formations. They found leader and follower switching in migrating birds can lead to considerable energy savings. It was found that with 14 geese in a flock, implementing position shifting (switching the leader and tail geese with those in the middle) could theoretically result in a minimum of 44.6% energy saving in their flight range and endurance compared to flocks without repositioning. The study provides an aerodynamic explanation for the observed repositioning behavior in V-shaped flocks. The lead and tail geese, which consume the most energy during flight, are replaced by those in the middle positions that have expended less energy. This repositioning helps maintain the flock’s overall energy efficiency and endurance. Extensive simulations showed that this leader-follower swapping mechanism is a viable strategy for managing energy expenditure during long migratory flights [310]. Kshatriya and Blake offered a similar theoretical model for determining the optimal flock size, which is a crucial factor in balancing the energy expenditure against the aerodynamic benefits of formation flight. The research highlighted that wing-tip spacing is a critical factor in formation flight. The benefit of formation flight increases with flock size, but there is a limiting value as the number of birds becomes indefinitely large. [312].
Mirzaeinia et al. extended these insights to artificial formations, demonstrating the potential for more than 21% improvement in flight time and distance of swarming drones after leader and tail repositioning. This approach, inspired by migrating birds, involves replacing the drones that have spent higher energy levels with those that have conserved more energy. The study showed that lead and tail drones in V-formation flight consume the highest level of energy. By reconfiguring the drones’ positions, energy efficiency can be significantly improved. The research presents theoretical modeling of aerodynamic drag forces in swarming flight and demonstrates that drones in a V-shaped formation can save up to 70% of their energy compared to solo flight. This reduction in drag force is attributed to the trailing aerodynamic effects of the front drones shown in Figure 25 and further discussed in Section 2.2.4, which leads to different energy consumption levels among the drones in the formation [311].
The paper outlines specific protocols for leader selection and load balancing in swarming fixed-wing drones. This approach is particularly useful when drones have different functionalities or are carrying different loads, as it allows for an optimal distribution of energy consumption across the swarm. The selection protocol is inspired by root node selection in spanning tree protocols used in computer networks [311].
Thien et al.’s study complemented this understanding by analyzing how the aerodynamic performance of a formation is affected by the leader’s wing configuration, including factors like incidence and dihedral angles. The study found that the incidence angle of the leader’s wing has a positive effect on increasing the lift and pitching moment, while reducing the drag of the rear wing. As the leader’s incidence angle increases, the rear wing experiences a higher drag reduction and increased lift. This suggests that the positioning and angle of the leader’s wing can significantly influence the aerodynamic efficiency of the entire formation. The dihedral angle of the leader’s wing was observed to have an adverse effect on the amount of drag reduction in the rear wing of the formation. A higher dihedral angle tends to linearly decrease the drag reduction benefits for the following wing. In terms of the taper ratio, Thien et al. showed that a low taper ratio in the leader’s wing leads to a higher lift increase in the trailing wings. Conversely, a high taper ratio results in a decrease in lift ratio for the trailing wing [313]. The research indicates that the optimal configuration for maximum drag reduction involves careful consideration of the leader’s wing shape and position. For instance, a rectangular platform of the leading wing can provide the highest drag reduction for the rear wing, emphasizing the importance of wing geometry in formation flight.
Evolutionary Aspects and Optimal Flock Dynamics Andersson and Wallander looked into the evolutionary biology aspect, suggesting that kin selection and reciprocity, especially in smaller groups, can influence the shape and dynamics of flight formations. The research suggested that kin selection plays a role in the formation of acute V formations, particularly in small flocks of geese, swans, cranes, and other large birds that migrate in family groups. The energetically more expensive leading position may be taken by adults as a form of parental care, with the gains in inclusive fitness making them willing to accept this less favorable position for the benefit of their relatives. This behavior indicates that in small flocks containing relatives, acute formations are often used, wherein the leader saves little or no energy. Andersson and Wallander also examined the concept of reciprocity, with individuals taking turns as leaders, sharing the energetic disadvantage of the front position. This phenomenon is expected mainly in small stable groups with individual recognition. The research suggests that in large flocks of unrelated individuals, acute formation shapes that save little energy for the leader may not be stable. Reciprocity in flight formations probably requires small groups in which individuals recognize each other [314]. Voelkl et al. provided empirical evidence of such behavior in Northern bald ibises, where juveniles demonstrate direct reciprocity in leading positions, balancing the energetic load across the group [304].
Collectively, these studies reveal the intricate balance of aerodynamics, energy management, and evolutionary biology in formation flight. They highlight the importance of strategic positioning, leader-follower dynamics, and cooperative behavior in achieving optimal energy efficiency. This understanding is crucial not only for comprehending natural avian flight patterns but also for informing the design and operation of energy-efficient aerial vehicles, such as drones.
Current State and Future Directions in Leader Switching Research
The current state of research on leader switching in formation flight indicates a deepening understanding of the aerodynamic and energy management benefits of this behavior. Studies have increasingly employed sophisticated models and empirical data to unravel the complexities of this phenomenon. While significant progress has been made, especially in understanding the mechanisms and advantages of leader switching, there remains ample scope for further research. Future studies could explore the application of these principles in advanced aerial vehicle design, as well as a deeper investigation into the evolutionary and ecological implications of such flight behaviors in various bird species.
2.3. Environmental Interactions
2.3.1. Time of Migration of Migratory Birds
Migration timing plays a crucial role in avian energy management and efficiency. Studies by Anderson et al., Brown et al., Somveille et al., Alerstam and Lindström, and Acácio et al. collectively discuss that the timing of migration can significantly influence the energy expenditure, route optimization, and overall migratory success in birds. These findings underscore the adaptive strategies birds employ to balance energy use, migration duration, and environmental conditions, offering insights into the design of energy-efficient aerial vehicles and biomimetic systems [238,315,316,317,318,319].
Energy-Minimizing Strategies in Herring Gulls and Lesser Black-Backed Gulls
The studies on Herring Gulls and Lesser Black-backed Gulls, by Anderson et al. and Brown et al. respectively, provide a deeper understanding of the energy-minimizing strategies employed by these birds during migration. Despite varying distances, both species prefer indirect routes, which are likely a strategic choice to manage energy consumption efficiently while navigating environmental challenges [315]. Anderson et al. show both short and long-distance migratory populations of Herring Gulls used indirect routes, contrary to the prediction that long-distance migrants would use more direct routes. Interestingly, the long-distance migrants were found to be the least direct in their routes. The research revealed that regardless of the distance traveled, Herring Gulls migrated at a slower overall migration speed than predicted by Optimal Migration Theory. This indicates that both short and long-distance migrants may prioritize minimizing energy expenditure over time. Contrary to the expectations, the study found that while long-distance migrants used more stopover days overall, relative to the distance traveled, all four populations used a similar number of stopover days. This suggests that the stopover strategy is not significantly influenced by the total migration distance and that stopovers are an important aspect of the migration strategy for both short and long-distance migrants [315].
For the lesser black-backed Gulls, the intriguing aspect lies in the consistency of their annual energy expenditure, regardless of their migration strategy. This consistency suggests a well-adapted energy management system that can efficiently distribute energy use throughout the year. However, the timing and distribution of energy expenditure vary, indicating a nuanced approach to optimizing energy use during different stages of their annual cycle [238]. Brown et al. found that the annual energy expenditure did not differ significantly among different migration strategies, which included birds wintering in locations ranging from France and the UK to West Africa. This indicates that birds, irrespective of their migration distance and wintering areas, manage to balance their energy budgets effectively throughout the year. The study’s findings suggest that the energetic costs of migration are offset by adjustments in behavior during other stages of the annual cycle. Despite varying migration distances and wintering areas, the time-activity budgets were remarkably similar across all migration strategies. This consistency in behavior suggests that the birds adjust their activities in a way that maintains a sustainable level of energy expenditure throughout the year. Notably, high inter-individual variation was observed among individuals using the same migration strategy, indicating that daily behavioral decisions may contribute more towards energy expenditure than the migration strategy itself [238].
Global Seasonal Distribution Driven by Energy Efficiency Energy efficiency is a driving factor in the global and seasonal distribution of birds, as seen in terrestrial bird species. The study by Somveille et al. proposes that bird migration is a behavior that allows highly mobile avian species to optimize their energy budget in response to fluctuating resources and interspecific competition. The model suggests that migration becomes more energetically efficient as interspecific competition reduces local energy supply, compelling species to migrate to areas with unexploited seasonal surpluses of energy. The results confirm the importance of energy supply competition in shaping the distribution of migrant and resident bird species across the world and throughout the year. The study’s mechanistic model demonstrates that birds’ distributions are not just a matter of geographical or climatic convenience but are intricately linked to optimizing energy expenditure. The model accurately predicted key global patterns of bird migration, such as the majority of breeding migrants occurring in the Northern Hemisphere around 50° N and redistributing to the southern part of the Northern Hemisphere for the non-breeding season. It also captured the peak in resident bird species in the tropics and the transition from avian communities that are net senders to those that are net receivers of breeding migrants. Comparisons with null models revealed that the best-fit model significantly outperformed alternatives, confirming that energy efficiency (the optimization of the balance between energy acquisition and energy expenditure) is a key mechanism underpinning bird migration patterns. This finding is supported by the strong correlation between simulated and empirical patterns of bird diversity and migration. By adjusting their migration patterns to align with resource availability and environmental conditions, birds exemplify a sophisticated strategy of energy management. This approach underscores the importance of energy efficiency as a key driver in the evolutionary adaptation and survival of bird species [316].
Optimization of Migration Birds are required to navigate trade-offs between rapid migration, energy-efficient travel, and safety, as underscored in the study by Alerstam and Lindström. This work looked into the decision-making process in bird migration, examining how various factors such as time, energy, and safety are balanced. The concept of optimal migration strategies is the main focus, which differs depending on whether the birds prioritize minimizing travel time, energy expenditure, or predation risk. The study suggested that birds maximizing migration speed are sensitive to variations in the rate of fat deposition along their migratory route. Conversely, energy-minimizing birds aim to reduce their total energy expenditure during migration, leading to different departure fat loads and stopover times compared to time-optimized migrants. The research discussed the role of fat deposition in determining a bird’s migratory behavior. Birds that accumulate fat at a certain rate will adjust their potential speed of migration with varying fat loads, impacting their choice of departure fat load and stopover sites. The study provides quantitative predictions about optimal fat loads and stopover times, illustrating how these are influenced by expected fat deposition rates and search/settling time costs at stopover sites. Alerstam and Lindström also examined the impact of predation risk on migratory strategies. They suggest that birds aiming to minimize predation risk during migration are likely to depart with smaller fat reserves compared to those prioritizing speed. This adaptation is expected to minimize the total mortality risk over the entire migratory journey [318].
Timing and Ecological Implications in White Storks The timing of migration in white storks has been shown to significantly affect their migratory performance, energy costs, and wintering destinations. The research conducted by Acácio et al. emphasized the significant impact of migration timing on the ecological success of white storks. Late departing storks, who experienced less supportive weather conditions, spent more energy during flight. They adopted shorter and straighter routes and spent fewer days on stopovers, potentially as a response to progressively deteriorating weather conditions in autumn. In contrast, early departing storks encountered more supportive weather conditions, spent less energy on migration and were exposed to westerly winds, leading them to likely end migration in eastern Sahel regions. The research showed that late departing storks faced stronger easterly winds and were more likely to end their migration in western areas of the Sahel. This pattern was observed in both adults and juveniles. The asynchronous migration timing of the population exposed individuals to different weather conditions, influencing their migratory routes and wintering areas. The study highlights that the choice of wintering grounds can influence individual energy expenditure and survival [319].
Historical Adaptability and Future Predictions
The research by Somveille et al. exploring the adaptability of migratory birds over the last 50,000 years discussed their remarkable evolutionary resilience. This study shows how migratory patterns have continuously evolved in response to changing climates, indicating a deep-rooted evolutionary mechanism for survival and efficient energy utilization. The ability of these birds to adapt their migration strategies over such an extended period is a testament to their extraordinary ecological flexibility. The presented model integrates annual energy budgets, considering both the energy assimilation and energetic costs associated with migration. It simulated bird species’ seasonal distributions, including breeding and non-breeding ranges, in a virtual world with similar geography and seasonal distribution of energy supply as the real world. The virtual bird species are distributed in a way that maximizes energetic fitness, optimizing the balance between energy assimilation and migration costs. Using this model, the study predicts the past global seasonal distribution of birds, indicating that the prevalence of avian migration has remained largely stable across the globe over the past 50,000 years. This suggests that bird migration has been continually important throughout glacial cycles. The results also show noticeable geographical variations in migration patterns due to changes in climate and energy availability [317].
The timing of migration has been presented in several past works as a pivotal element in the energy management and ecological success of migratory birds, as evidenced by the collective findings of the reviewed research. This body of work offers a detailed understanding of avian migration strategies, highlighting how birds adapt to environmental challenges and resource availability. The integration of these insights is critical for advancing bio-inspired engineering designs and conservation strategies. Understanding these natural migration patterns can guide the development of efficient, adaptable systems in both technology and ecological management.
2.3.2. Flight Routes of Migratory Birds
Bird migration, a marvel of nature, exemplifies an extraordinary journey, intricately optimized for energy efficiency. Migratory birds, in their quest for survival, have evolved to adeptly navigate a myriad of environmental variables and challenges. Several studies provide fascinating insights into these adaptations [320,321,322,323]. These studies collectively explore how wind patterns, weather conditions, and even human-induced factors like light pollution, profoundly influence migratory behavior. Birds’ responses to these factors reveal an intricate understanding of their environment, where each decision (from the choice of route to the timing of departure and the method of flight) is a testament to their remarkable capacity to optimize energy expenditure and ensure survival over long, arduous journeys. This section synthesizes these diverse studies to offer a comprehensive view of the strategies employed by migratory birds, underscoring their resilience and adaptability in the face of environmental challenges and changes.
Wind-Dependent Flight Routes
Nourani’s research on the Oriental honey-buzzard and Kranstauber et al.’s analysis of global migratory patterns both emphasize the critical role of wind in determining migratory efficiency [320,323]. Nourani illustrates how the Oriental honey-buzzard adjusts its flight routes in response to wind conditions, optimizing energy use during migration. The study presented that predicted changes in wind conditions due to climate change have the potential to significantly affect the suitability of traditional avian migratory routes. The Oriental honey-buzzards depend heavily on thermals and tailwinds to complete their sea crossing, and alterations in wind patterns can severely impact their migration. By mid-century, there is a likelihood of a slight shift and reduction in suitable areas for migration, followed by a complete loss of this section of the traditional route by the end of the century under both climatic scenarios (RCP4.5 and RCP8.5). The study also raised the possibility that suitable wind conditions over the migration region might occur later in the year due to climate change, leading to a temporal shift in migration timing. This shift could have far-reaching ecological consequences, including delayed arrival at wintering grounds and increased competition with other raptors [320].
Kranstauber et al. extend this understanding by demonstrating the potential evolutionary adaptation of migratory routes at a population level. They show that wind-assisted routes could significantly reduce travel time, indicating that birds might naturally select these routes for their energy-saving benefits. The research demonstrates that following an optimal route, calculated using empirical global wind data, can significantly reduce median travel time compared to the spatially shortest route. For instance, choosing a wind-optimized route reduced median travel time by 26.5%. This suggests that birds may adjust their migratory routes at a population level, resulting in the emergence of low-cost flyways representing a global network of aerial migratory pathways. This was determined by modeling optimal routes for migratory birds, taking into account wind speed and direction, which are interpolated to all nodes in a hexagonal grid. The model assumed a constant air speed for the bird, with tail and crosswinds influencing ground speed linearly [323]. These studies collectively highlight how wind patterns are not just navigational aids but vital factors influencing the evolution and optimization of migratory paths [320,323].
Meteorological Adaptations in Avian Migration Lanzone et al. and Rüppel et al. provided compelling insights into how birds adeptly navigate meteorological challenges during migration. Lanzone et al.’s study on golden eagles highlights their strategic use of different flight modes to adapt to wind variations, showcasing an advanced level of environmental sensitivity crucial for energy-efficient migration. Golden eagles responded to increased wind speeds by changing their flight mode from thermalling (using thermal updrafts for lift) to orographic flight (using wind deflected upwards by terrain). As wind speed increased, the eagles decreased their use of thermals and flew at lower altitudes. This behavior suggests a strategy to minimize energetic expenditures by adapting flight patterns to take advantage of the available lift, whether from thermals or orographic effects. The data showed that as the use of thermals decreased, there was a corresponding decrease in variation in flight speed and altitude. Eagles exhibited a more consistent flight pattern when relying on orographic lift compared to thermalling. This consistency indicates that the birds are optimizing their flight in relation to the changing wind conditions to maintain efficient migration [321].
Rüppel et al.’s research extends this understanding to songbirds, revealing how weather conditions intricately influence their migratory decisions. The migratory songbirds studied were more likely to depart for migration under favorable weather conditions, such as clear skies and no precipitation. The birds preferred light westward winds and southward wind directions, indicating a selection for tailwinds to assist their flight. This behavior aligns with previous studies suggesting that birds select for periods with no precipitation to depart, as rain can be hazardous during flight. Routing decisions, whether to fly offshore or follow the coastline, are predicted by wind direction. The study found that westward winds increase the probability of birds opting for offshore flights. Birds departed on average half an hour earlier for offshore routes than for coastal flights, suggesting that they maximize the available night time for flying across the sea. Additionally, the landing decisions of birds during migration were affected by changes in wind conditions and cloud cover. The probability of landing increased with cloud cover and with positive changes in the northward wind component. Birds that experienced a positive change in the northward wind component during flight were more likely to suspend migration [322].
Figure 26.
a) Current ensemble mode projection for the regions of suitable atmospheric conditions during migration of honey buzzards, with the dots representing en route locations of migrating birds, tracked using satellite transmitters [320]. b) Future projects for the suitable atmospheric conditions during the migration of honey buzzards under two different scenarios [320]. c) Optimal tracked flight routes over 21 years between Sweden and Ethiopia with the great circle route as a dashed line and color representing the average survival. Many of these routes outperform the great circle route [323]. d) Telemetry data collected from golden eagles migrating through the Appalachian Mountains, along with the above ground level (AGL) altitude in three 15 km regions of flight from three separate eagles. From top to bottom, the eagles are displaying orthographic lift, lift from thermals, and thermal streets [321]. e) Fall migratory paths of the eastern whip-poor-wills (yellow points are stopovers) with 20 km buffers surrounding the routes, overlayed on artificial light data collected by satellites [324]. f) Comparison of the a straight line path and the recorded flight path between two nightly stopovers, highlighting the avoidance of a major source of artificial light [324]. All figures are reprinted with permission; please see the Appendix A.
Figure 26.
a) Current ensemble mode projection for the regions of suitable atmospheric conditions during migration of honey buzzards, with the dots representing en route locations of migrating birds, tracked using satellite transmitters [320]. b) Future projects for the suitable atmospheric conditions during the migration of honey buzzards under two different scenarios [320]. c) Optimal tracked flight routes over 21 years between Sweden and Ethiopia with the great circle route as a dashed line and color representing the average survival. Many of these routes outperform the great circle route [323]. d) Telemetry data collected from golden eagles migrating through the Appalachian Mountains, along with the above ground level (AGL) altitude in three 15 km regions of flight from three separate eagles. From top to bottom, the eagles are displaying orthographic lift, lift from thermals, and thermal streets [321]. e) Fall migratory paths of the eastern whip-poor-wills (yellow points are stopovers) with 20 km buffers surrounding the routes, overlayed on artificial light data collected by satellites [324]. f) Comparison of the a straight line path and the recorded flight path between two nightly stopovers, highlighting the avoidance of a major source of artificial light [324]. All figures are reprinted with permission; please see the Appendix A.
These studies collectively illustrate the remarkable capacity of birds to respond to a range of meteorological conditions, adjusting their flight strategies to optimize travel efficiency. This behavioral flexibility, demonstrated across different bird species, underscores a fundamental aspect of avian migration: the ability to dynamically adapt to changing environmental conditions for optimal energy management and successful migration [321,322].
Analysis on the Impact of Artificial Light on Migratory Patterns
Thompson provided a critical examination of how artificial light affects the migratory behavior of Eastern Whip-poor-wills. One of the key findings is that migrating whip-poor-wills took more winding, less direct flight paths in response to more intense artificial light. This effect was observed on the scale of nightly flights from stopover to stopover along their fall migration routes. Interestingly, the effect of artificial light on the overall scale of migration routes was not found, suggesting that whip-poor-wills make migratory decisions based on environmental conditions or factors experienced while in flight. This study reveals that these birds significantly alter their migratory routes to circumvent areas with high artificial light intensity, indicating that whip-poor-wills may be sacrificing efficiency on their migratory flights to avoid exposure to bright artificial light. Such behavioral adjustments indicate a profound adaptation to human-induced environmental changes. The deviations caused by light pollution may result in longer and more complex migratory routes, underscoring the adaptability of these birds to urbanized environments but also highlighting the potential challenges they face, including increased energy expenditure and exposure to predators or other hazards. This research emphasizes the growing need to understand and mitigate the impacts of urbanization on wildlife, particularly migratory species like Eastern Whip-poor-wills, whose natural behaviors are being altered by human activities [324].
Integrating Avian Migration Strategies into Broader Contexts
The collective insights from these studies illuminate the extraordinary adaptability and navigational prowess of migratory birds. Strategies such as wind-assisted travel, weather-based route adjustments, and light pollution avoidance demonstrate an intricate understanding of environmental dynamics and resource optimization. These adaptive behaviors provide invaluable insights into the resilience of migratory species, offering lessons applicable beyond ornithology. The knowledge gained from these avian strategies can inform conservation efforts, guiding actions to mitigate impacts on migratory pathways. Additionally, these findings offer intriguing parallels for human technological advancements in the field of UAVs, where principles of energy efficiency and adaptive routing could be inspired by avian migration.
2.3.3. Changing Altitude
The optimization of flight altitude in migratory birds represents a fascinating intersection of aerodynamics, physiology, and environmental adaptation. This comprehensive review synthesizes findings from several pivotal studies to elucidate how birds achieve this optimization, particularly during migratory journeys. Understanding these dynamics is not merely an academic pursuit; it has practical implications in fields ranging from aviation safety to bio-inspired engineering. Migratory birds, traversing thousands of kilometers, face a myriad of challenges including varying atmospheric conditions, energy management, and physiological limitations. Their ability to modulate flight altitude in response to these challenges is a testament to a sophisticated interplay of innate behaviors and adaptive strategies. This review delves into the nuances of how weather conditions, wind patterns, and the birds’ unique physiological and bio-mechanical characteristics contribute to their remarkable efficiency in long-distance flights. The insights gleaned from these studies not only enhance our understanding of avian biology but also offer valuable lessons for human endeavors in flight and conservation efforts.
Influence of Weather and Wind Conditions
The studies by Shamoun-Baranes et al., Dokter et al., and Kemp et al. collectively underscore the profound impact of weather and wind conditions on avian flight altitudes [240,325,326]. They reveal a complex interaction where birds dynamically adjust their altitudes in response to changing weather patterns, predominantly to leverage tailwinds for migratory efficiency. Dokter et al. demonstrated using weather radar data how birds modulate their flight paths to optimize wind assistance. A key finding from the study is the observation of distinct bird density altitude profiles, which varied depending on weather conditions. The researchers noted that birds adjust their flight altitude to make optimal use of tailwinds along their migratory direction. For example, on certain nights, wind conditions at low altitudes were more favorable than at high altitudes, leading birds to migrate at lower altitudes. Conversely, on other occasions, favorable tailwinds at higher altitudes resulted in a significant portion of migration occurring at these higher elevations. Migration patterns observed at individual sites could strongly depend on weather conditions elsewhere, particularly in northern temperate climates where weather can vary significantly. This indicates that birds’ decisions to start migrating and their chosen altitudes are influenced not only by local weather conditions but also by conditions further along their migratory flyway [325].
Kemp et al. observed that nocturnal migrants, despite the availability of supportive winds at higher altitudes, often prefer lower flight paths unless lower altitudes offer adverse wind conditions. The study found that birds generally prefer to fly at lower altitudes unless wind conditions near the surface are unsupportive of migration. When surface winds are prohibitive, birds tend to increase their flight altitudes to seek out more supportive winds. However, even when flying higher than usual, birds tend to concentrate around the lowest altitude with acceptable, though not necessarily optimal, wind conditions. The generalized additive model analysis indicated an influence of temperature on flight altitudes, showing that birds specifically avoided colder layers [326]. These findings highlight an intricate decision-making process, balancing energy conservation, environmental conditions, and physiological demands, which birds navigate during migration.
Similarly, Mateos-Rodríguez and Liechti’s study provided insights into the decision-making processes of diurnal long-distance migratory birds concerning altitude selection in relation to wind conditions. Their work used radar tracking and revealed that these birds, despite having the capability to assess a wide range of altitudes, often select a limited range based on local maxima of tailwind support. This finding suggests that the birds’ altitude selection is not solely about finding the most favorable wind conditions. Instead, it reflects a complex balance of factors including energy conservation, predation avoidance, and physiological constraints. The study highlights that birds are making calculated decisions based on a combination of immediate environmental cues and inherent biological limitations, a process that is far more nuanced than previously understood [327].
Figure 27.
Altitude distributions of birds measured through radar studies. a) A map of the operational weather radars in western Europe. Those highlighted in red are sites used in the study by Dokter et al. [325]. b) Comparison of the bird density altitude profiles measured by (top) the bird radar and (middle) weather radars. The height integrated bird densities for each in the bottom plot (weather-red, bird-blue) [325]. c) Weighted average of the altitude distribution of proportional bird density (pBd) for spring and autumn [326]. d) Graphic depicting the typical flight altitude of various high altitude flyers [328]. e) Map showing the radar range used in Kemp et al.’s study [326]. All figures are reprinted with permission; please see the Appendix A.
Figure 27.
Altitude distributions of birds measured through radar studies. a) A map of the operational weather radars in western Europe. Those highlighted in red are sites used in the study by Dokter et al. [325]. b) Comparison of the bird density altitude profiles measured by (top) the bird radar and (middle) weather radars. The height integrated bird densities for each in the bottom plot (weather-red, bird-blue) [325]. c) Weighted average of the altitude distribution of proportional bird density (pBd) for spring and autumn [326]. d) Graphic depicting the typical flight altitude of various high altitude flyers [328]. e) Map showing the radar range used in Kemp et al.’s study [326]. All figures are reprinted with permission; please see the Appendix A.
Adaptations for High-Altitude Flight
Birds that elect to fly at high altitudes have developed unique adaptations at every step of the oxygen transport pathway to facilitate increased exercise requirements. These adaptations include an enhanced hypoxic ventilatory response, more effective breathing patterns, larger lungs, hemoglobin with higher oxygen affinity, changes in pulmonary and cardiovascular systems to enhance oxygen delivery, modifications in muscle structure to facilitate oxygen uptake, and multiple alterations in the metabolic properties of cardiac and skeletal muscle. Additionally, these birds often have larger wings compared to their lowland relatives, reducing the metabolic costs of staying aloft in low-density air [328,329]. Altshuler also explored how environmental and biotic factors, such as solar radiation, air temperature, humidity, and ecosystem composition, change systematically with elevation and influence avian physiology and flight performance. Birds residing at high altitudes or engaging in high-altitude migration need to adapt to these varying conditions, particularly in terms of thermoregulation and water balance [329].
Implications for Avian Flight Strategy and Future Research
While these studies collectively enhance our understanding of avian migration strategies, they also highlight a significant gap in research. These studies suggest that birds integrate a complex array of environmental, physiological, and bio-mechanical factors in their altitude selection, optimizing for energy efficiency and flight performance. However, there is a notable lack of comprehensive research beyond these foundational studies into the intricacies of flight altitude optimization during avian migration. This area remains ripe for further exploration, with potential implications for bio-inspired aerial vehicle design and predicting bird migration patterns concerning aviation safety. As such, this topic presents a rich opportunity for future studies to delve deeper into the complexities of avian aerodynamics and environmental interaction.
2.4. Ground Effects
When viewing the flight of large birds, particularly those with high aspect ratio wings, or wide narrow wings, it becomes increasingly apparent they often fly as close to the surface as possible. This presents the opportunity to ask why these birds repeatedly undertake what seems like such a difficult action in comparison to flying higher. Looking at just about any animal, the efficiency of their movement holds extreme value as it determines a great deal of the energy they expend, and thus how much food they must consume to keep up, which in turn also affects how much of their time they must expend to eat. Taking this into consideration, what benefit do these birds receive from flying low versus flying high? They do this to take advantage of the physics of the ground effect [126,128,129,133,135,137,138,142,155,156,159,161,165,167,169,176,177,330].
The ground effect comes into play when a bird flys above a surface no higher than its wingspan, and becomes more apparent the closer to the surface flight occurs [128,330,331]. As a bird comes closer to the surface they are flying over, the efficiency of flight increases due to the way the air behaves and is most commonly taken advantage of by birds with large aspect ratios, such as albatrosses, shearwaters, and black skimmers, and brown pelicans. This effect is primarily caused due to the ground interrupting the wingtip vortices and, thus the downwash from the wing. As an airfoil nears the ground, the wingtip vortices are cut off due to the surface resulting in smaller vortices, which in turn affects the downwash behind the wing. This downwash points the relative wind downward, and when interrupted by a surface, no longer does so as effectively, and because the lift is perpendicular to the relative wind, this results in a more vertical lift vector and thus reduced induced drag [128,331]. Flying close to a surface also has an impact on the air pressure. As an airfoil nears a surface, the air pressure on the underside of the wing increases, and this helps to improve the lift-to-drag ratio, and all of this combined indicates that an airfoil requires a lower angle of attack near a surface to provide the same amount of lift when not taking advantage of the ground effect(see Figure 28).
Ground effect in avian flight, a critical aerodynamic phenomenon, has been extensively explored through diverse research methodologies, including computational, experimental, observational, and theoretical studies. This synthesis aims to integrate these varied perspectives, presenting a comprehensive view of the current state of ground effect research in birds. Studies employing computational models reveal that birds achieve greater energy efficiency when flying close to the ground, optimizing wingtip vortices and exploiting ground plane effects [176]. Numerical investigations, like those by Tang et al. and Su et al., provide further understanding of the lift and drag characteristics in avian flapping flight, emphasizing the significance of ground effect [167,333].
Observational studies have significantly contributed to our understanding of how different bird species utilize ground effects. For instance, research has shown that ecological and behavioral factors heavily influence birds’ use of ground effect, with variations observed across species and environments. This is particularly evident in studies examining how birds adjust their flying strategies in relation to wind conditions and ground proximity, revealing the adaptive nature of their flight mechanics [133,135]. Experimental research complements these findings by demonstrating the aerodynamic benefits of specific wing designs when operating near the ground. Studies investigating seabird-like wings and NACA6409 wing sections have shown substantial improvements in lift coefficients in ground effect, highlighting the crucial role of wing morphology in energy-efficient flight [138,161]. Sakornsin and Atipan performed aerodynamic testing of several seabird-like wing models in a wind tunnel at speeds equivalent to a Reynolds Number of 240,000. These models were compared against a rectangular wing of the same aspect ratio to evaluate the effects of ground proximity on lift, drag coefficients, and lift-to-drag ratios at various angles of attack. The tests aimed to reveal how seabird wing shapes perform aerodynamically when in the ground effect compared to a conventional rectangular wing. The study found that seabird-like wings provide better aerodynamic performance when in the ground effect than the normal rectangular wing. Specifically, wings with delta wrist shapes demonstrated significant improvements in maximum lift-to-drag ratios, indicating that such designs could be particularly beneficial for WIG craft. The results showed an increase in lift and a reduction in drag coefficients as the wings approached the ground, highlighting the efficiency of seabird-like wing designs in ground effect conditions [161].
Theoretical research in the field of ground effect aerodynamics has been crucial in uncovering the fundamental principles that govern this phenomenon. The development of models for potential flow problems, as explored in the work of Baddoo et al., has been instrumental in advancing our understanding of ground effects in both natural and engineered flight systems [128]. Additionally, the studies by King and Rayner on the aerodynamics of wing formations and vortex wakes in the ground effect have provided significant insights. These studies focus on how wing configurations and vortex interactions can be optimized to reduce induced drag, offering valuable strategies for the design of more efficient aircraft [142,155]. The integration of these theoretical advancements is pivotal for enhancing our understanding of ground effect complexities and lays a foundation for future innovations in the design of aerial vehicles.
Blake’s study provides an in-depth analysis of the mechanics of gliding in birds, with a particular emphasis on the influence of the ground effect. This research offers key insights into how the ground effect can enhance the glide efficiency of birds, enabling them to conserve energy [129]. Similarly, Sachs focuses on the minimum wind strength required for dynamic soaring in albatrosses, highlighting the crucial role of environmental conditions in facilitating efficient flight through ground effect [159]. Withers also explores the aerodynamic cost of flight in the Black Skimmer, a species known for its low-altitude flight patterns, emphasizing the significance of ground effect in energy conservation during flight [177].
Other research presents evolutionary perspectives on ground effect utilization, particularly in bats which typically fly in alternate modes to gliding. This study reveals how bats modulate their flight mechanics to optimize ground effect usage, highlighting its importance in flight efficiency and energy savings [137]. In contrast, aerodynamic analyses of barn owls in gliding flight has also been studied, demonstrating how these birds adjust their flight dynamics to benefit from ground effect with minimal sound, thereby enhancing their aerodynamic performance [165].
The comprehensive exploration of ground effects in avian flight has revealed the complex interplay between aerodynamics, ecology, and evolutionary biology, greatly enhancing our understanding of avian flight mechanics [128,129,133,135,137,138,142,155,159,161,165,176,177]. Studies across various disciplines, from computational models and observational studies to experimental research and theoretical advancements, have collectively highlighted the significance of ground effect in energy efficiency and flight dynamics. This body of research not only provides deeper insights into natural avian flight but also offers valuable lessons for the design and optimization of aerial vehicles, demonstrating the importance of multidisciplinary approaches in advancing both biological understanding and technological innovation.
3. Drag Reduction in Flying Insects
3.1. Structural and Physical Characteristics of Insects
The study of the structural and physical characteristics of insects offers a fascinating glimpse into the world of aerodynamics and biomechanics. This subsection delves into various aspects, including wing structure, hairs, scales, color, and flight modes, all of which contribute uniquely to the flight capabilities and efficiency of insects. Each characteristic, from the microscopic structure of wing surfaces to the coloration patterns that may influence thermal regulation, plays a pivotal role in how these creatures interact with their environment. Understanding these features not only enlightens us about the intricate world of insect physiology but also paves the way for innovative applications in technology, particularly in the design of advanced aerial vehicles and energy-efficient systems. This comprehensive analysis aims to bridge the gap between biological understanding and technological innovation, drawing inspiration from nature’s design.
3.1.1. Wing Shape
The study of wing shape in flying insects once again stands at the intersection of biology and aerodynamics, offering additional insights into the mechanics of flight. This section delves into the nature of insect wing design, exploring how evolutionary pressures have shaped various species to achieve improved aerodynamic efficiencies. Research in this field not only unravels the complex interplay between wing morphology and flight performance but also illuminates potential applications in engineering, particularly in the design of bio-inspired aerial vehicles. By examining diverse insect species, ranging from the delicate flutter of butterfly wings to the robust flapping of locusts, this section brings forth a comprehensive understanding of how wing shape influences flight dynamics, energy utilization, and maneuverability in the natural world. The implications of these findings extend beyond biological insights, offering innovative cues for the development of more efficient and adaptable aerial technologies.
Morphological Influence on MAV Design
Insect wing morphology, encompassing aspect ratios, venation patterns, and flexibility, plays a pivotal role in the aerodynamics of flight. This aspect of wing design is not merely a biological curiosity but has significant implications for the field of aeronautics, particularly in the design and optimization of MAVs. The intricate structures of insect wings, with their unique configurations and properties, provide critical insights into efficient lift and thrust mechanisms. Understanding these natural design principles aids in enhancing MAV performance, leading to innovations in lift, maneuverability, and energy efficiency. The correlation between natural wing designs and artificial flight technology underscores a key area of biomimicry, where learning from nature informs human engineering, pushing the boundaries of what is achievable in MAV technology. Research by Whitney utilized experimental methods for testing the aerodynamic forces and three-dimensional kinematics generated by insect-scale flapping wings. The experiments involve mechanical drives to control wing movements, allowing precise manipulation of experimental parameters. The study also incorporates the use of Reynolds-number matching scaled-model experiments, where a scaled-up model of the wing is flapped in a high-viscosity fluid to match the aerodynamic forces and Reynolds numbers observed in actual insects. The research demonstrates significant variation in flapping kinematics across different insect species, with implications for aerodynamic efficiency. The study emphasizes the importance of understanding detailed wing deformations and real-time forces to optimize MAV design. The results suggest that passive wing dynamics could reduce the power required to flap the wings by either reducing inertial power consumption or allowing passive tailoring of wing shape to optimize aerodynamic efficiency [334].
Figure 29.
a) Adult wings of genetically modified Drosophila, displaying the change in wing morphology [335]. b) Changes in flight performance for the genetically modified Drosophila with 95% confidence intervals where the purple color denotes significant change relative to the control group [335]. c) Variation in forewing morphology of migrating monarch butterflies [336]. d) Validation experiment of CFD simulations using full fidelity wing kinematics of locusts [337]. e) View of the computed downwash distribution from the CFD simulation of full fidelity locust wing kinematics [337]. f) Tracings of successive frames from video of two Trichoptera in teathered flight, highlighting the deformation of the wings through the motion [338]. g) Example of a one-wing flapping mechanism for use in MAV systems [334]. h) Digitized wings of various small insects with a red line representing the longitudinal axis and b representing bristle spacing. Of particular note are the bristles on the wing and the vastly varying morphology [339]. All figures are reprinted with permission; please see the Appendix A.
Figure 29.
a) Adult wings of genetically modified Drosophila, displaying the change in wing morphology [335]. b) Changes in flight performance for the genetically modified Drosophila with 95% confidence intervals where the purple color denotes significant change relative to the control group [335]. c) Variation in forewing morphology of migrating monarch butterflies [336]. d) Validation experiment of CFD simulations using full fidelity wing kinematics of locusts [337]. e) View of the computed downwash distribution from the CFD simulation of full fidelity locust wing kinematics [337]. f) Tracings of successive frames from video of two Trichoptera in teathered flight, highlighting the deformation of the wings through the motion [338]. g) Example of a one-wing flapping mechanism for use in MAV systems [334]. h) Digitized wings of various small insects with a red line representing the longitudinal axis and b representing bristle spacing. Of particular note are the bristles on the wing and the vastly varying morphology [339]. All figures are reprinted with permission; please see the Appendix A.
The unique wing flexibility observed in butterflies represents a remarkable evolutionary adaptation, enhancing their flight efficiency significantly. Johansson and Henningsson’s research illustrated how during the upstroke, butterfly wings can form a cupped shape, effectively capturing air and generating increased thrust. This mechanism, often referred to as the ’clap and fling,’ is a sophisticated aerodynamic technique that reduces drag and maximizes lift, making it a crucial study point for bio-inspired design. They used high-speed video recordings to analyze the take-off and flight dynamics of butterflies. They specifically observed the clap technique, where the wings of a butterfly come together at the top of the upstroke, creating a pocket of air between them. By rapidly opening their wings, butterflies push this air pocket downwards, generating additional thrust that aids in flight. The analysis showed that the clap technique substantially increases the thrust compared to typical wing flapping. The unique wing shape of butterflies allows for this technique, enabling a more efficient take-off and flight. During the upstroke, the wings of a butterfly are peeled apart, creating a unified start vortex, which then forms a single vortex ring generated by all four wings. The implications of this finding are substantial in the realm of biomimetic engineering, particularly for the development of flapping drones and bio-inspired flying devices. By mimicking these natural motions, engineers can create more efficient, agile, and energy-saving aerial vehicles. [340].
The study of monarch butterflies, particularly regarding their migration behaviors and wing morphology, offers insight into evolutionary adaptations for energy-efficient flight. Flockhart’s and Satterfield’s research sheds light on how these butterflies, known for their extraordinary long-distance migrations, have developed wing sizes and shapes that optimize for endurance and energy efficiency. The larger and more angular wings observed in migratory monarchs, as compared to their non-migratory counterparts, indicate a natural selection process favoring traits that enhance aerodynamic performance over vast distances. This evolutionary adaptation is not only a testament to the resilience and adaptability of these creatures but also provides a valuable model for the design of aerial vehicles. The research by Flockhart et al. analyzed 613 monarch butterflies collected on Mexican wintering grounds from 1976 to 2014. Migratory distance from their natal areas in eastern North America was estimated through stable isotope analysis. The study measured wing morphology traits, such as length, width, area, roundness, and aspect ratio, to test their association with migratory distance. The findings showed that monarchs that migrated longer distances tended to have longer and larger wings. This relationship suggests positive selective pressure on wing length and area during migration. However, there was no significant relationship between migration distances and either wing roundness or aspect ratio. The results remained consistent when controlling for temperature, indicating that migratory distance plays a role in shaping wing morphology independently of temperature effects [341].
Similarly, Satterfield and Davis captured fall-migrating monarchs across an entire migration season and examined wing area, elongation (length/width), and redness. They used image analysis software for measuring these characteristics, with wing hue being an index of the degree of redness. Monarchs were divided into early, middle, and late groups based on capture date, and statistical analyses were conducted to assess the variation in wing characteristics. The work discussed that early migrants tended to have more elongated and redder forewings compared to late migrants. There was also a non-significant tendency for early migrants to have larger forewing areas. This pattern suggests that monarchs with more elongated and redder wings, which are associated with better flight performance, tend to migrate earlier. In contrast, those with less optimal wing characteristics likely make up the latest migrants [336]. By understanding and mimicking these natural adaptations, engineers can develop more efficient and resilient designs for long-distance aerial travel, as evidenced in the studies [336,341]. The implications of these findings extend beyond biology, offering practical applications in the enhancement of aerodynamic efficiency in human-engineered flying machines.
The mechanics of insect wing deformation, as explored in the work by Wootton, display insights into the aerodynamic performance of insect flight. These deformation patterns, characterized by their flexibility and adaptability, significantly enhance lift and thrust during flight. Wootton’s approach analyzed the detailed structure of insect wings and their deformation during flight, using simple physical models based on direct observations of insects in flight and manipulations of wings. The study examined aspects of wing geometry, such as corrugation, an M-shaped section, camber, and how these features influence bending, torsion, and profile change in wings. It was found that insect wings are smart structures whose three-dimensional shape during flight is largely determined by their elastic response to aerodynamic and inertial forces. The paper discusses the significance of relief structures, like corrugation, in supporting the wing and allowing it to bend usefully during the upstroke. The orientation of lines of transverse flexion is shown to determine the extent of torsional deformation in wings, which is essential for efficient flight [338]. This understanding is particularly relevant when juxtaposed with the flexibility and clap mechanism observed in butterfly wings. The clap and fling motion, as studied by Johansson, demonstrates a sophisticated strategy to maximize aerodynamic efficiency, where wing flexibility plays a central role [340].
The aerodynamic benefits of wing deformation in locusts, as highlighted By Young et al.’s research, present another fascinating example of nature’s engineering prowess. In locusts, the natural design and deformation of wings during flight play a critical role in enhancing aerodynamic efficiency. Unlike rigid or simplified wing models, the flexible and dynamic nature of locust wings allows for adaptations in flight that improve lift and thrust. These deformations are not random but are finely tuned adaptations evolved over millennia, and optimized for energy efficiency and effectiveness in various flight maneuvers. The study utilized high-speed digital video cameras to track over 100 natural features and marked points on the wings of locusts, reconstructing the deforming surface topography of the wings. The CFD simulations incorporated these detailed wing kinematics to provide a nuanced understanding of wing deformation during flight. The wings were modeled with a constant thickness based on cross-sections of the wing veins and membrane. The CFD simulations showed that the full-fidelity model, which included both camber and spanwise twist, achieved greater power economy than models without these features. Removing camber and twist dramatically changed the aerodynamics, leading to reduced lift power economy. The study highlights that wing deformation, particularly in locusts, is crucial for enhancing the efficiency of momentum transfer to the wake and directing the aerodynamic force vector appropriately for flight. This demonstrated the significant aerodynamic advantages of detailed wing kinematics in insects. The maintenance of attached flow aerodynamics and the ability to reduce flow separation, which is essential for high-lift aerodynamics, are achieved through tailored wing deformations. Understanding these natural mechanisms is not only vital for entomological studies but also offers valuable lessons for the design of more efficient and adaptable aerial vehicles, especially in applications where flexible, responsive wing structures could lead to significant improvements in performance [337].
The studies of wing deformation in locusts and the clap mechanism in butterflies represent complementary aspects of how insects utilize wing flexibility to optimize flight. These insights are invaluable in bio-inspired engineering, where mimicking such natural mechanisms can lead to the development of more efficient and innovative aerial vehicles. The deformation mechanics, combined with the clap and fling technique, provide a comprehensive framework for understanding how subtle variations in wing structure and motion can drastically improve aerodynamic performance. Integrating these concepts into the design of MAVs could lead to breakthroughs in flight efficiency, stability, and energy utilization, bridging the gap between natural evolution and technological innovation.
The study of small insect wing designs, particularly focusing on features like bristle structures, opens up a nuanced perspective on aerodynamics in the insect world. The presence of bristles, a seemingly minor feature, can significantly influence the aerodynamic efficiency of these tiny creatures. This aspect of wing morphology, unique to smaller insect species, is indicative of highly specialized evolutionary adaptations geared towards optimizing flight in varying environmental conditions [339].
Engels et al. employed three-dimensional computational fluid dynamics and parametrized wing models with bristle densities typical for small insects. The approach assesses flow fields around individual bristles and the entire wing, aerodynamic forces and moments, aerodynamic power, and Rankine–Froude efficiency of flight. The research focuses on how the efficiency of muscle power is translated to vertical force production by morphologically different wings. The results demonstrated that the decrease in aerodynamic efficiency with decreasing surface solidity is significantly smaller at low Reynolds numbers compared to higher Reynolds numbers. This finding suggests that small insects may fly with both bristled and solid wing surfaces at similar efficacy, while larger insects must use membranous wings for efficient production of flight forces. The study concludes that aerodynamic efficiency, rather than maximum force production, is key to the coexistence of bristled and solid wings in insects [339].
Fly wings, characterized by their unique shapes, vein patterns, and flexibility, demonstrate a complex interplay between physical structure and aerodynamic function. These features are not just incidental but are finely tuned for optimizing flight efficiency, maneuverability, and stability according to Krishna et al. The review highlights the complexities of insect wing structure, noting that wings are three-dimensional appendages with varying degrees of stiffness across their span. The tapered shape of fly wings improves span efficiency during root-flapping, but genetic modifications of wing shape have questioned whether the current shape results solely from an evolutionary selection process towards maximum aerodynamic performance. It is suggested that three-dimensional corrugation patterns of veins and membranes form valleys that channel axial flow components but do not trap vortices for lift enhancement as previously thought for more corrugated wings of dragonflies. The study of these wings offers a window into the sophisticated mechanisms insects use to control flight, providing valuable insights for aerodynamic modeling and design in engineering, especially in the creation of more efficient and responsive aerial vehicles [342]. This understanding underscores the potential of biomimicry in advancing aerial vehicle design, where mimicking these natural designs could lead to significant improvements in flight performance and energy efficiency.
Genetic Influence on Wing Shape
Ray et al. used RNA interference targeting the gene narrow to alter wing shape in developing Drosophila. This genetic manipulation resulted in wings with different shapes, specifically affecting the aspect ratio and overall wing planform. The study utilized a combination of high-speed videography and physical modeling to analyze the flight performance of these genetically altered flies. The altered wing shapes showed significant effects on key performance metrics such as maneuverability and agility. Flies with more tapered wings (higher aspect ratio) demonstrated enhanced agility, suggesting that a higher aspect ratio is beneficial for agile flight. However, the study also revealed that these performance improvements come at a cost, possibly requiring more power for flight and reducing efficiency. The aerodynamic model predicted that all the altered wing morphs displayed a decrease in aerodynamic efficiency compared to the control. This suggests that measurable differences in flight performance are a result of a balance between aerodynamic and mechanical modifications due to the shape change. The most extreme morphs, while showing improved agility, were more demanding to fly and less efficient [335].
This research extends into broader implications, suggesting that understanding and applying genetic influences on wing morphology could lead to innovative approaches in the design and optimization of aerial vehicles. By harnessing the principles observed in these genetic studies, engineers and designers could potentially create more efficient, adaptable, and robust MAVs. The ability to fine-tune wing characteristics at a genetic level in nature presents a novel avenue for biomimetic design, where small but precise modifications could yield significant improvements in performance and energy utilization [335,342].
Impact of Wing Planform on Aerodynamics
Additionally, other studies have researched the aerodynamic impact of various wing planform shapes in insects. This research provides a thorough understanding of how different wing shapes, such as elongated, broad, or tapered wings, influence the aerodynamics of insect flight. Li and Nabawy’s findings indicate that specific wing shapes can significantly affect lift, drag, and overall flight efficiency. The study analyzed how these variations in wing planform contribute to the agility, maneuverability, and energy efficiency of insect flight. They used numerical simulations, solving the incompressible Navier-Stokes equations, to evaluate the aerodynamic performance of different wing planforms. The study defined wing planforms using a beta-function distribution and employed kinematics representative of normal hovering flight. The simulations considered variations in aspect ratio, radial centroid location, and wing root offset, and their effects on aerodynamic force coefficients, flow structures, and aerodynamic efficiency. It was found that the wing with the lowest aspect ratio of 1.5 resulted in the highest peaks of lift and drag coefficients during stroke reversals. In contrast, wings with higher aspect ratios produced higher lift coefficient whilst leading to relatively small variations in aerodynamic efficiency for most aspect ratios. The influence of radial centroid location and wing root offset on aerodynamic performance was also systematically investigated, revealing distinct effects on the aerodynamic behavior of flapping wings [343].
In Liu et al.’s study, the focus is on optimizing wing shapes inspired by beetle hindwings for improving the aerodynamic performance of Flapping Wing MAVs (FWMAVs). This research delves into the unique aerodynamic properties of beetle hindwings, which are often overlooked in studies of insect flight. Geometric models based on the hindwings of these beetle species were created and fabricated with various parameters for wind tunnel testing. The experiments were conducted in a low-speed wind tunnel, focusing on optimizing wing shapes for high lift and efficiency. The tests assessed the impact of different parameters like aspect ratio, trailing edge length, root chord length, and wing area on the aerodynamic performance. This work highlights how these wings, with their specific shapes and structural features, contribute to efficient flight in beetles. By examining these characteristics, the study proposes innovative design principles for FWMAVs, aiming to replicate the efficient lift and thrust mechanisms observed in beetles. This approach to wing shape optimization opens new avenues in aerial vehicle design, showcasing the potential of integrating natural evolutionary designs into technological advancements [344].
The study by Engels et al. presented an analysis of the aerodynamic consequences of the three-dimensional wing structure in small dipteran insects. The researchers investigated wings from three fly species: Drosophila melanogaster, Musca domestica, and Calliphora vomitoria. They used CFD simulations to analyze the aerodynamic forces and power requirements during root flapping. Various wing models were generated to dissect the contribution of fine- and coarse-scale structures to aerial performance. These models included natural wings, wings without camber, and completely flat models. The study found that the three-dimensional design of fly wings, particularly the camber, has a pronounced effect on the temporal distribution of aerodynamic forces, power requirements, and Rankine–Froude efficiency. However, contrary to expectations, the coarse structure of fly wings (camber) attenuates aerodynamic efficiency by up to approximately 12% compared to a flat wing. The researchers did not find evidence for lift-enhancing trapped vortices in corrugation valleys at the tested Reynolds numbers. The results suggested that the primary function of the wing’s three-dimensional structure is to provide mechanical support against external forces rather than improving lift or saving energy costs associated with active wing flapping. The study supports the assumption that wing venation aligns closely with aerodynamic pressure distribution during flapping, giving interesting insights into the design of MAVs on the scale of insects [345].
In other research focusing on the impact of leading-edge vortices, Chen et al. discuss how variations in insect wing shape influence the size, strength, and dynamics of LEVs. These vortices are essential for lift generation in low-Reynolds-number flight, typical of small insects. The study used CFD simulations to compare different wing models, showing that forewing morphologies are crucial in LEV formation and significantly impact lift generation. The simulations incorporated the wing morphologies of a fruitfly (Drosophila melanogaster), a bumblebee (Bombus ignitus), and a hawkmoth (Manduca sexta), focusing on how these morphologies affect integral aerodynamic force coefficients, local pressure loading, and the LEV angle. The findings revealed that the forewing morphologies of these insects are well-matched to the formation of LEVs and are responsible for generating sufficient lift forces at the mean angles of attack and Reynolds numbers where these insects fly. Interestingly, the removal of the hindwing showed minimal change in LEV formation and pressure loading and even led to some improvement in power factor and aerodynamic efficiency. This suggests that the forewing morphology plays a dominant role in achieving low-Reynolds number aerodynamic performance in natural flyers as well as in revolving and/or flapping micro air vehicles [346].
3.1.2. Wing Structure and Hairs
Insects exhibit a remarkable variety of wing structures and hair arrangements, each contributing uniquely to their flight efficiency. This section explores how these adaptations not only facilitate flight but also offer insights into biomimetic applications in aerospace engineering. The intricate relationship between wing morphology and aerodynamics in insects like the Brown Lacewing (Micromus tasmaniae), butterflies, and members of the Mymaridae family will be discussed in detail.
Hair Structures
The Brown Lacewing’s wings demonstrate a dual-layer hair structure comprising macrotrichia and microtrichia, which enhance water repellency and potentially reduce flight drag. The study by Watson et al. utilized scanning electron microscopy and atomic force microscopy to examine the dual-layered hair structure on the lacewing’s wings, consisting of larger macrotrichia originating from the wing veins and smaller microtrichia scattered on the wing surface. This unique arrangement aids in repelling water and resisting contamination, which is crucial for maintaining the insect’s flight capabilities. The structure and dimensions of these hairs, including their length, spacing, and angle, were analyzed to understand their role in water repellency. The structured hair arrays on the brown lacewing’s wings effectively repel water at different length scales. The macrotrichia and microtrichia work together to minimize the contact area between the wing surface and water, thus enhancing the anti-wetting properties of the wings. This is particularly important for the brown lacewing, as its high wing surface area-to-body mass ratio makes it susceptible to being weighed down by water droplets [347]. The ability to repel water helps prevent the detrimental effects of water adhesion, such as increased weight and drag. Comparative analysis with other lacewing species highlights the evolutionary adaptations for water repellency, suggesting potential biomimetic applications for aerial vehicle design.
Studies on the aerodynamic characteristics of bristled wings in the smallest flying insects, like the Mymaridae family, reveal that these wings’ efficiency varies with Reynolds numbers and flapping strokes [348]. Shen et al. performed an analysis of the clap-and-fling mechanism, showing that bristle crossing and inter-wing gap reduction significantly influence aerodynamic forces and efficiency. The researchers collected wing morphological data from 38 specimens of the Mymaridae family and used these measurements to simulate bristled wing flight at Reynolds numbers ranging from 1 to 80. This led to the discovery of an unusual clap-and-fling pattern with bristle crossing, examining how this affects aerodynamic forces. The findings indicate that a lift-based stroke is superior to a drag-based stroke in terms of vertical force production and aerodynamic efficiency at 5 ≤ Re ≤ 20. Bristled wings using the clap-and-fling mechanism achieve both vertical force and efficiency augmentation, although bristle crossing incurs a substantial horizontal force and contributes little to vertical force augmentation. This suggests that bristle crossing in the clap-and-fling process may not be all that advantageous for flight efficiency. Understanding these aerodynamic characteristics is crucial for the miniaturization design of future bio-inspired MAVs.
Figure 30.
a) Image of a brown lacewing’s large hair arrangement on the wing veins [347]. b) Example of a water droplet resting on a brown lacewing’s wing [347]. c) SEM image revealing the dual layer of hairs on a brown lacewing [347]. d) An image of a bristled forewing from a Ptilomymar dianensis [348]. e-g) Examples of bristled wings from a featherwing beetle, chalid wasp, and the placentis beetle [349]. h)Experimental setup used to measure the flapping kinematics and perform PIV analysis in a tank filled with glycerine [350]. All figures are reprinted with permission; please see the Appendix A.
Figure 30.
a) Image of a brown lacewing’s large hair arrangement on the wing veins [347]. b) Example of a water droplet resting on a brown lacewing’s wing [347]. c) SEM image revealing the dual layer of hairs on a brown lacewing [347]. d) An image of a bristled forewing from a Ptilomymar dianensis [348]. e-g) Examples of bristled wings from a featherwing beetle, chalid wasp, and the placentis beetle [349]. h)Experimental setup used to measure the flapping kinematics and perform PIV analysis in a tank filled with glycerine [350]. All figures are reprinted with permission; please see the Appendix A.
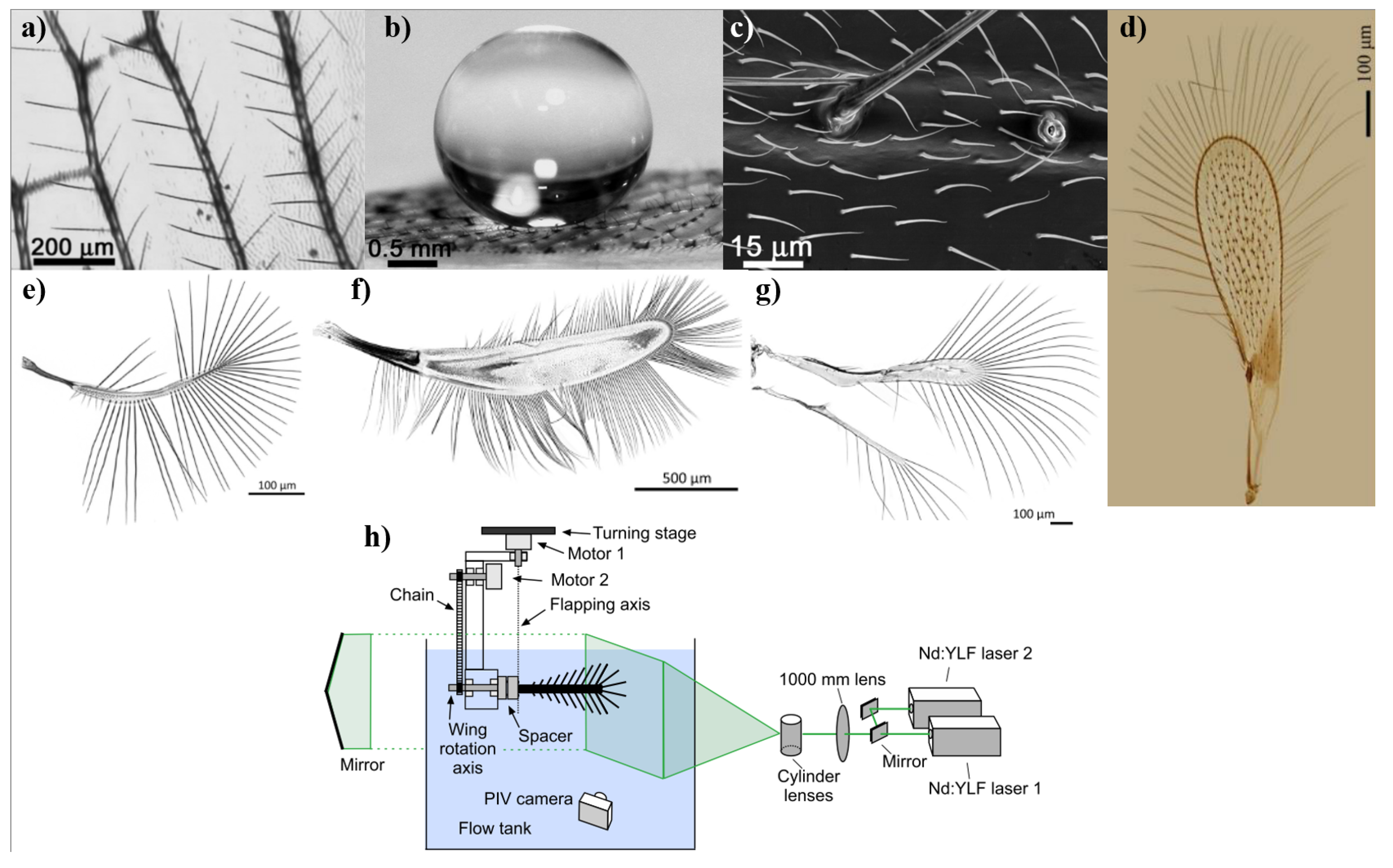
Kolomenskiy’s research presented an in-depth examination of the flight mechanics of miniature insects with bristled wings. The study highlights the functional advantages and aerodynamic properties of these wings, emphasizing their role in flight stability and maneuverability. Many of the smallest insects, such as those in the Mymaridae family, have evolved wings with elongated central blades and fringes of bristles, resembling bird feathers. These bristled wings are effective in producing the same aerodynamic forces as equally-sized membranous wings, yet are significantly lighter. The study emphasizes the importance of the wing’s three-dimensional structure, including bristle density and arrangement, in its aerodynamic performance. The work also discussed how these insects have adapted their wing beat kinematics to the low wing inertia and relatively high viscosity of their surrounding medium. This adaptation is manifested in U-shaped and figure-of-eight wing tip trajectories and pronounced wing clapping. The study provides insights into the flow blockage effect at low Reynolds numbers, showing that sparse arrays of slender bristles can function as virtually impermeable paddles. The aerodynamic stability of bristled wings in flapping flight was discussed as well, suggesting that the permeability effect of the bristles reduces force fluctuation caused by gusts. This feature, along with dynamic deformation of bristled wings, contributes to the flight performance of the smallest insects, enabling them to fly with competitive efficiency at the lowest possible cost [349].
O’Callaghan et al. presented a comprehensive analysis of the aerodynamic advantages and disadvantages of bristled wings in small flying insects, with a focus on the dependence on the Reynolds number. Their study utilized robotic wing models with various bristle spacings, from solid membranes to a few bristles, to mimic the wing structure of small insects. The wings were moved around their root using a generic lift-based kinematic pattern. The researchers assessed the lift coefficients, power coefficients, and Froude efficiency of these wings at different Reynolds numbers to understand the aerodynamic changes associated with bristled wing designs. The lift coefficients, power coefficients, and Froude efficiency decreased with increasing bristle spacing. Skin friction significantly attenuated lift production, which may result in negative coefficients at elevated bristle spacing and low Reynolds numbers. The research confirmed that for small insects, flying with bristled wings involves less change in energetic costs compared to larger insects. The study suggests that the co-occurrence of bristled and solid wings in miniature insects results from aerodynamic efficiency rather than maximizing force production [351].
O’Callaghan and Lehmann’s study shifted the focus to the aerodynamic performance of bristled wings due to LEVs, particularly in the tiny thrip Gynaikothrips ficorum. The research employed the same robotic model wings to analyze LEV circulation and its relation to bristle spacing. The wings were tested at a Reynolds number of 3.4, which is representative of small insect flight, in a flow tank filled with glycerine. 2D PIV analysis was applied to assess the flow development and LEV behavior during wing flapping. The findings indicate that the aerodynamic performance due to LEV circulation linearly decreases with increasing bristle spacing. The wings of Gynaikothrips ficorum, with their specific bristled structure, produce approximately 9% less aerodynamic force for flight compared to solid membranous wings. Additionally, at the stroke reversals, leading and trailing edge vortices dissipate quickly, allowing a quick build-up of counter-vorticity when the wing reverses direction. The results highlight that bristled wings in small insects are less effective for aerodynamic lift production. However, O’Callaghan and Lehmann point out that bristled wings co-exist with membranous wings in small insects, suggesting that bristled wings might be beneficial in certain conditions, like reducing drag during clap-and-fling kinematics at stroke reversal [350].
Flexural Stiffness in Insect Wings
Other studies examine the relationship between wing venation, material properties, and flexural stiffness in various insect species. The study by Combes et al. utilizes a combination of experimental measurements and finite element modeling to understand how venation patterns and material distribution contribute to wing stiffness. The flexural stiffness of wings from 16 insect species across six orders were measured. Flexural stiffness combines the material properties of the wing (Young’s modulus) and the geometric distribution of this material (second moment of area). The team performed static bending tests and digitized wing venation to quantify venation patterns. To account for phylogenetic effects, standardized independent contrasts of venation and stiffness measurements were calculated and analyzed for correlations. The study found that flexural stiffness is strongly correlated with wing size but is not significantly affected by the details of the venation pattern. There is a significant anisotropy between spanwise and chordwise flexural stiffness; spanwise stiffness is 1 to 2 orders of magnitude greater than chordwise stiffness in all species tested. To explore how wing structure contributes to stiffness anisotropy, a simplified finite element model of a hawkmoth wing was created. The model demonstrated that adding or strengthening veins in the leading edge of the wing significantly increases spanwise flexural stiffness, producing the observed anisotropy in real wings. This finding indicates that certain wing venation patterns are likely a common functional feature among insects [352].
Figure 31.
a) Finite element models based on Manduca sexta wings with the top row colors representing different regions of Young’s modulus. Model one has fixed values, while model two has a modulus that declines toward the trailing edge. Below this are the displacement results for a static point load applied at the wingtip, as well as a distributed load equivalent to the approximate pressure on a wing during steady flight [352]. b) SEM image of longitudinal veins (LV) and cross veins (CV), highlighting the morphological difference near the edge of the hind wing. The LV are relatively smooth and of circular cross-section, while the CV display an annulated pattern [353]. c) Example of a typical Schistocerca gregaria hind wing, displaying the distribution of the wing cell’s major axis length where CCL is the critical crack length [353]. d) Vibration modes of the wing model used by Reid et al. from the finite element model superimposed on an undeformed wing geometry. On the left is the first mode showing bending, and the right is the second mode showing torsion [354]. e) Schematic and physical realization of an experimental setup for testing the flapping motion of insect wings, simultaneously measuring reaction torques and annular position [354]. f) Schematic examples of circulation patterns and tracheae in insect wings, displaying the bi-directional flow in the veins, and variation in circulation patterns [355]. All figures are reprinted with permission; please see the Appendix A.
Figure 31.
a) Finite element models based on Manduca sexta wings with the top row colors representing different regions of Young’s modulus. Model one has fixed values, while model two has a modulus that declines toward the trailing edge. Below this are the displacement results for a static point load applied at the wingtip, as well as a distributed load equivalent to the approximate pressure on a wing during steady flight [352]. b) SEM image of longitudinal veins (LV) and cross veins (CV), highlighting the morphological difference near the edge of the hind wing. The LV are relatively smooth and of circular cross-section, while the CV display an annulated pattern [353]. c) Example of a typical Schistocerca gregaria hind wing, displaying the distribution of the wing cell’s major axis length where CCL is the critical crack length [353]. d) Vibration modes of the wing model used by Reid et al. from the finite element model superimposed on an undeformed wing geometry. On the left is the first mode showing bending, and the right is the second mode showing torsion [354]. e) Schematic and physical realization of an experimental setup for testing the flapping motion of insect wings, simultaneously measuring reaction torques and annular position [354]. f) Schematic examples of circulation patterns and tracheae in insect wings, displaying the bi-directional flow in the veins, and variation in circulation patterns [355]. All figures are reprinted with permission; please see the Appendix A.
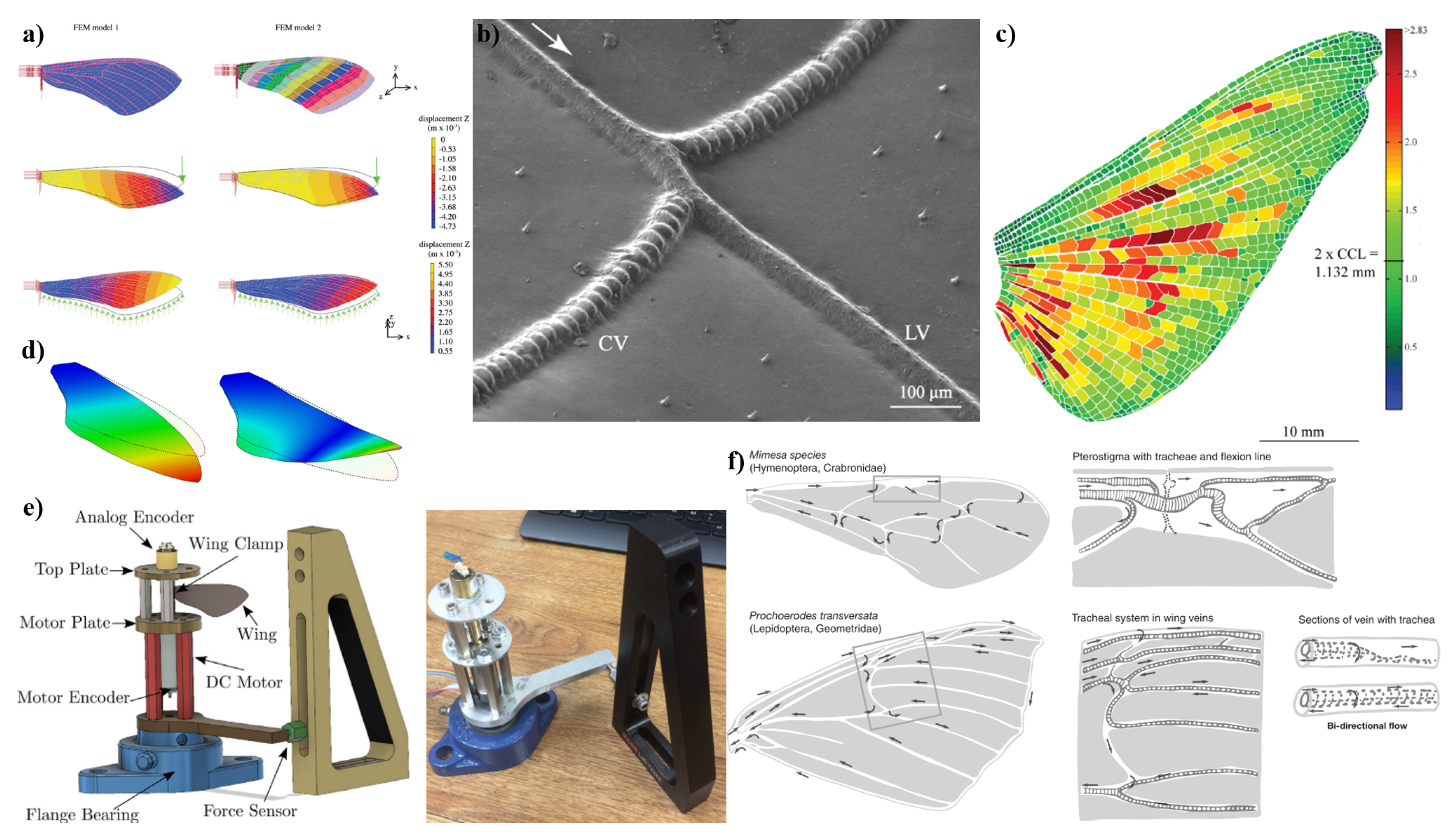
Reid et al. explored how wing flexibility impacts the energy efficiency of insect flight. The study combines field observations, laboratory experiments, and computational modeling to examine the biomechanical properties of insect wings. The team developed a novel methodology to determine the total torques and mechanical power of a flapping, flexible insect wing. They combined fluid-structure interaction modeling with a new method to estimate the inertial torques of a flexible wing. The approach involved both numerical simulation and experimental validation using a custom single-degree-of-freedom mechanical flapper to actuate a real hawkmoth wing. found that flexibility can reduce energy expenditures by almost 25% compared to a rigid wing if negative work is stored as potential energy and subsequently released to do positive work. The wing itself can store about 30% of the total energy required over a wingbeat. The model predicted experimental trends that rigid body models could not, suggesting that compliance should not be neglected in considering flight dynamics at this scale [354]. The findings highlight the importance of flexibility in flapping wing MAV design and suggest that tuned flexibility can greatly improve vehicle efficiency.
Veins Improve Fracture Toughness of Insect Wings Tangent to this, other studies investigate the role of veins in enhancing the fracture toughness of insect wings, particularly in desert locusts. Dirks and Taylor employed experimental techniques to measure the wings’ fracture toughness, stiffness, and strength. This involved mechanical testing to measure the fracture toughness of fresh and desiccated locust wings. The team manually induced small cracks at the edges of the wing samples, parallel to the longitudinal veins. The experiments were conducted using a standard tensile test machine, and the stress required to propagate a pre-existing crack was measured. The findings show that the wing veins act as effective barriers to crack propagation. The presence of cross veins significantly increases the wings’ structural fracture toughness by 50%, from 1.04 to 1.57 MPa·m. This indicates that the venation of locust wings not only qualitatively but also quantitatively follows a uniform pattern that enhances the wings’ durability. The study suggests that the vein pattern found in insect wings could inspire the design of more durable and lightweight artificial ’venous’ wings for MAVs [353].
Hemolymph circulation within insect wings, as detailed by Salcedo and Socha, is vital for maintaining wing functionality. The paper highlighted that hemolymph circulates through insect wings, playing a vital role in maintaining the functionality of sensory hairs, structural elements, and other living tissue within the wings. Circulation ensures that these components remain operational, contributing to the wings’ effectiveness as flight appendages and active sensors. The work noted that circulation in insect wings is critical for preserving wing flexibility, essential for aerodynamic performance. Without hemolymph circulation, wings can dry up and rapidly lose their flexibility, impacting flight mechanics. Salcedo and Socha also discussed the varying circulation patterns across different insect species and the methods used to study these patterns [355].
Inspiration from Insect Wing Scales
Zhang et al. provided a detailed classification of lepidopteran wing-scale structures based on their microstructural features. The study explores the optical properties of these structures and their physical mechanisms behind unique colorations. It also reviewed various biomimetic fabrication techniques inspired by these wing scales for applications in materials science. This comprehensive analysis offers valuable insights into designing new materials with unique optical properties. While the work touches upon the aerodynamic properties of butterfly wing scales, it primarily emphasizes the potential applications of these structures in the development of novel materials. These applications include optical gas sensors, structured color regulators, solar cells, anti-counterfeiting labels, and high-speed infrared imaging devices. [356].
Figure 32.
Example of butterfly dorsal scales using both SEM and optical microscopy. a) Example of the curvature of butterfly scales, showing the layers and the tip cupping upward [357]. b) image of scales that have been cut through to show a cross-section [357]. c) General view of the dorsal scales on a monarch butterfly wing [357]. d) Similar image of the monarch dorsal scales, but focused around a wing vein, displaying the perpendicular nature of the scales [357]. All figures are reprinted with permission; please see the Appendix A.
Figure 32.
Example of butterfly dorsal scales using both SEM and optical microscopy. a) Example of the curvature of butterfly scales, showing the layers and the tip cupping upward [357]. b) image of scales that have been cut through to show a cross-section [357]. c) General view of the dorsal scales on a monarch butterfly wing [357]. d) Similar image of the monarch dorsal scales, but focused around a wing vein, displaying the perpendicular nature of the scales [357]. All figures are reprinted with permission; please see the Appendix A.
More recent research investigated the aerodynamic functionality of microscopic scales on butterfly wings, particularly how they contribute to flight efficiency by reducing skin friction drag. Lang analyzed the effects of removing scales from monarch butterfly wings on their flight efficiency. The study used high-speed cameras to analyze the flapping motion and trajectory of monarch butterflies, first with their scales intact and then with the scales removed. The experiment revealed significant differences in flying efficiency between the two conditions. The removal of scales from the wings decreased the butterflies’ mean climbing efficiency by an average of 32%. This significant reduction in efficiency demonstrates the crucial role of scales in flight. The scales impart a unique, advantageous geometry to the wings, which includes microscopic cavities formed by the upward curve of the scales’ tips. These cavities improve the wing’s aerodynamics by decreasing skin friction. Lang further discusses the aerodynamics of butterfly flight, noting that butterflies, including monarchs, typically fly close to the ground but can reach higher altitudes during migration. The scales on their wings, besides providing coloration and water repellency, also play a significant role in reducing skin-friction drag by as much as 45%. This drag reduction is crucial for efficient flight, particularly given the butterflies’ large wings relative to their body size. These findings have substantial implications for biomimicry and aerospace engineering, suggesting that insights from butterfly wing scales can inspire advanced materials and surfaces in MAV design [358].
Prior to the research on wing scales reducing drag, Wilroy et al.’s research on butterfly-scale-inspired surface patterning investigated the impact on LEV growth and aerodynamic characteristics. The experiments were conducted at Reynolds numbers of 1416, 2833, and 5667, using a 2D flat plate with and without grooved patterning. The grooves, designed to mimic the scale patterns found on butterfly wings, were hypothesized to influence the LEV formation and growth. The study utilized experimental techniques to characterize the LEV, including the creation of secondary vortices and their vorticity, instability shedding from the shear layer, and saturation of the LEV. The findings showed that the presence of grooves significantly influenced the development of the LEV. A stronger secondary vortex was observed to lead to a lower maximum LEV circulation, thus enabling longer attachment and stability of the LEV on the wing. This mechanism was found to be particularly relevant to the aerodynamic benefits of butterfly wing scales, which have microgeometries that can stabilize the LEV by manipulating vortex interactions. Wilroy et al.’s results suggest that the optimized geometry of butterfly scales contributes to aerodynamic efficiency by managing the flow structures around the wings [359].
Sledgers et al. once again indicate that the microgeometry and surface patterning of wing scales play a significant role in enhancing aerodynamic efficiency. They compared the free-flight body kinematics and flapping of butterflies in two conditions: with wing scales and after the removal of wing scales. The team developed a method to measure butterfly flapping kinematics optically over long uninhibited flapping sequences. The experiment involved a total of 236 individual flights from 11 butterflies, providing a comprehensive dataset to analyze the impact of scales on flight efficiency. The results showed a significant decrease in mean climbing efficiency of 32.2% after the removal of the scales, with a confidence interval of 45.6% to 18.8%. Additionally, the flapping amplitude decreased by 7%, although the flapping frequency did not show a significant difference. These findings strongly suggest that the microscopic scale geometry and surface patterning on butterfly wings play a crucial role in improving their climbing efficiency, and are in good agreement with the work by Lang et al. [357]. Studies show a marked decrease in climbing efficiency and changes in flapping amplitude after the removal of these scales, underscoring their contribution to flight performance [357,358].
3.1.3. Wing Color
In the realm of aerodynamics and energy harvesting, the structural and physical characteristics of insects provide a fascinating blueprint for innovation. A potentially key aspect of these characteristics is wing coloration. Most insect wings are indeed transparent, a feature that serves various purposes including camouflage and reduced visibility to predators. However, there are notable exceptions where insects possess non-transparent, colored wings. These colored wings are not just for show; they can play significant roles in thermoregulation, communication, and mating rituals [360]. For instance, the vibrant colors found in butterfly wings are often used to attract mates or ward off predators. More intriguingly, the field of biomimicry and bio-inspiration have found other applications of these colored wings. Research suggests that the microstructures and pigments responsible for these colors can improve solar absorption, leading to potential efficiency gains in solar cell technology [361]. Similarly, these colors and solar absorption may benefit aerodynamic efficiency similar to that of bird wings. Additionally, the unique structural coloration of some insect wings has inspired advancements in non-reflective coatings and efficient, lightweight materials for use in aeronautics. The study of colored insect wings extends beyond mere aesthetics, offering innovative insights and applications in technology and energy efficiency. This section synthesizes research findings that explore the relationship between the color of insect wings and their flight efficiency, offering insights for potential applications in aerial vehicle design.
Energy-Harvesting Applications in Butterfly Wing Structures
The unique micro and nanostructures of butterfly wings not only contribute to their vivid coloration but also present significant opportunities in energy harvesting, potentially offering a blueprint for advanced materials in aerial vehicle design. These natural structures are adept at manipulating light, which could lead to significant advancements in drag reduction and energy efficiency in aerial vehicles. Butterfly wings inspired Osotsi et al. in the development of photocatalysts due to their unique microstructures that enhance light absorption and scattering. These photocatalysts have shown improved performance in environmental remediation and renewable energy harnessing, drawing from the efficiency of light manipulation by butterfly wings. These bio-inspired solar cells aim to mimic the efficient light absorption and trapping mechanisms of butterfly wings, potentially leading to more efficient and high-performance solar energy technologies [361].
Replicating these light-manipulating structures in aircraft design could result in innovative surface coatings or materials that enhance aerodynamic performance while reducing energy consumption. The study by Osotsi et al. emphasized the potential of integrating such natural efficiency mechanisms into aviation technologies, potentially revolutionizing aerodynamics and energy harvesting in the aerospace sector [361].
Furthermore, butterfly wing architectures could inform the development of bio-inspired sensors and renewable energy systems. This aligns with sustainable practices and opens new avenues for energy-efficient design in aerospace engineering. The exploration of butterfly wing structures thus serves as a compelling example of how nature-inspired designs can lead to more efficient and environmentally sustainable technology solutions in aviation [361].
The studies by Davis et al. provide compelling evidence on the impact of wing coloration in monarch butterflies on their flight performance [362,363]. The 2012 study meticulously measured the flight distance, duration, and speed of monarch butterflies, revealing a strong correlation between darker wing coloration, especially shades closer to red, and superior flight performance. This was evident in longer flight distances and indicates a potential link between the intensity of wing coloration and energy efficiency in flight. The results showed that monarchs with darker orange (approaching red) wings flew longer distances than those with lighter orange wings. This finding is consistent with previous observations that monarchs sampled during the fall migration are darker in hue (redder) than non-migratory monarchs. The study suggests that pigment deposition onto wing scales during metamorphosis could be linked with traits influencing flight, such as thorax muscle size, energy storage, or metabolism [362].
Using a tethered flight mill, Davis et al. measured various flight parameters of monarchs, quantifying the variation in wing coloration ranging from lighter to darker orange shades. The study adjusted for variables such as sex and morphological traits, ensuring the observed relationship between darker wing coloration and improved flight performance remained significant [362].
The research underscores the significance of physical traits like coloration in influencing energy efficiency. The darker wing coloration, closer to the red spectrum, is suggested to play a crucial role in energy-efficient flight, opening avenues for mimicking these natural coloration patterns in aircraft and UAV designs.
Extending their research, an additional study focused on the size of white spots in monarch butterflies and their impact on long-distance migration. Monarchs with larger white spots, particularly those collected at wintering sites, showed adaptations that possibly offer aerodynamic advantages for migration. This introduces a new perspective on how color patterns can enhance flight efficiency. The work used images of nearly 400 wild monarchs collected during the summer breeding period, fall migration, and at the Mexican overwintering sites. The analysis focused on determining if the relative coverage of the three primary wing colors changes along the migration journey. The study employed multivariate ANOVA (analysis of variance) and univariate ANOVA models to examine the variation in color percentages across different stages of the monarch’s life cycle (breeding, migration, wintering), as well as the effect of sex and year of collection. The analysis revealed that the relative surface area of two of the three primary colors varied across the summer, fall, and wintering stages. Specifically, the average percentage of the monarch forewing covered by black pigment declined by approximately 3 from the summer breeding stage to the overwintering stage. In contrast, the percentage of white on the wings increased by approximately 3% while orange did not vary significantly across migration stages. The study suggests there may be aerodynamic advantages induced by the coloration of monarch wings, indicating the white spot size on the wing margins may play a role in aerodynamic efficiency [363].
Figure 33.
Examples of butterfly wing coloration studies showing the feature changes between migratory and non-migratory species. a) Image of a monarch engaging in gliding flight, displaying the range of colors on the wing and their interaction with the sunlight shining on them. Photo credit: John Blair [363]. b) Schematic view showing the features of a standard butterfly wing scale, highlighting the complex geometry [361]. c) A comparison view of various closely related Danid butterflies examined for their wing coloration differences. Particularly, note the reduction in white spots of the non-migratory species. Photo credit: P. Barriga [363]. d) A display showing the expected solar exposure levels experienced by monarch along its migration south in the fall [363]. e) Comparison showing the relative white spot size of the Danid species shown in (d) [363]
Figure 33.
Examples of butterfly wing coloration studies showing the feature changes between migratory and non-migratory species. a) Image of a monarch engaging in gliding flight, displaying the range of colors on the wing and their interaction with the sunlight shining on them. Photo credit: John Blair [363]. b) Schematic view showing the features of a standard butterfly wing scale, highlighting the complex geometry [361]. c) A comparison view of various closely related Danid butterflies examined for their wing coloration differences. Particularly, note the reduction in white spots of the non-migratory species. Photo credit: P. Barriga [363]. d) A display showing the expected solar exposure levels experienced by monarch along its migration south in the fall [363]. e) Comparison showing the relative white spot size of the Danid species shown in (d) [363]
These insights could be influential for the field of aerospace engineering, suggesting that certain color patterns and spot sizes may enhance long-distance flight efficiency [362,363]. The integration of these principles in aerial vehicle design could lead to innovative approaches in energy harvesting and drag reduction in engineered flight systems.
These studies collectively highlight the potential of biomimicry, especially in harnessing the lessons from insect wing coloration for aerospace engineering. The integration of these natural efficiency mechanisms into aviation technology could revolutionize aerodynamics and energy harvesting, paving the way for more sustainable and efficient aerial vehicles.
3.1.4. Flight Modes
Hovering Flight
Hovering flight, a remarkable capability observed in various insect species, plays a crucial role in their survival and energy management strategies. This mode of flight is characterized by its stationary position in the air, demanding intricate wing kinematics and substantial energy expenditure. The following sections delve into various studies that explore the nuances of hovering flight, shedding light on the aerodynamic principles and energy efficiency tactics employed by these insects.
A study by Berman and Wang provided crucial insights into the optimization strategies of wing motion in hovering insects. Through detailed analysis of fruit flies, bumblebees, and hawkmoths, the research unveils a commonality in their wing kinematics aimed at balancing lift generation with minimal energy use. The research found that for each of the three insects, there exist specific wing kinematics that minimize power usage while still producing sufficient lift to maintain hovering flight. Their approach employed a hybrid optimization algorithm, integrating genetic algorithms with gradient-based optimization, to approximate natural insect flight patterns, allowing for the determination of the most energy-efficient wing movements during hovering flight. Berman and Wang compared the optimized kinematics to previously measured wing strokes of these insects. For instance, the optimal wing motion for the fruitfly is largely flat but slightly U-shaped, which is qualitatively similar to observed kinematics. The bumblebee and hawkmoth, on the other hand, show a figure-of-8 motion with larger stroke deviation and less constant forces and powers during the mid-stroke. These findings suggest that the observed wing strokes of these insects are close to the energy-minimizing kinematics. The work also examined the sensitivity of the optimal solutions to perturbations in various kinematic parameters. This analysis helped define why the optimal kinematics are at the values found and why most insects maintain the same leading edge throughout a stroke. It was noted that this type of stroke takes advantage of passive wing rotation, where aerodynamic forces help reverse the wing pitch, similar to the turning of a free-falling leaf [364]. This study not only enhances our understanding of insect biomechanics but also serves as a foundation for designing energy-efficient biomimetic UAVs.
Wang et al. presented a detailed examination of the effects of different flapping modes and wing kinematics on the hovering flight of insects. Utilizing numerical simulations based on two-dimensional unsteady Navier-Stokes equations, the study focused on the intricate dynamics of insect wing movements. The study considered four types of idealized flapping modes for insect wings: harmonic translation with harmonic rotation, harmonic translation with trapezoid rotation, trapezoid translation with harmonic rotation, and trapezoid translation with trapezoid rotation. These modes were analyzed to evaluate their influence on the aerodynamic performance of hovering insect wings. They found significant differences in instantaneous and time-averaged aerodynamic performances across varied flapping modes. The mode with harmonic translation and harmonic rotation is found to be the most energy-efficient, making it suitable for long-duration flight. In contrast, the harmonic translation-trapezoid rotation mode generates the largest instantaneous aerodynamic forces, which could be advantageous for maneuvering flight. Additionally, the effects of the Reynolds number on the time-averaged aerodynamic performance were systematically studied for each flapping mode. It was observed that the time-averaged lift force and lifting efficiency increase significantly with increasing Reynolds number, up to a certain point, after which the increase slows down. The study emphasized that the Reynolds number is a crucial factor affecting the aerodynamic performance of insect wings [365].
In past computational works on insect hovering, researchers emphasize the role of leading-edge vortices and wing tip vortices in generating lift during the upstroke, which constitutes a significant portion of the total lift in hovering flight. Their analysis demonstrates the impact of the Reynolds number on the formation and stability of these vortices, with notable differences observed between smaller and larger insects. Aono et al. found that during the early down- and upstroke of hovering, a horseshoe-shaped vortex is generated around the wing, which evolves into a doughnut-shaped vortex ring. This ring, with an intense jet stream in its core, forms the downwash. The doughnut-shaped vortex rings eventually break up into two circular vortex rings in the wake. A significant portion of the lift force (approximately 62% of the total lift generated over a full wingbeat cycle) is produced during the upstroke, primarily due to the presence of intensive and stable LEVs and wing tip vortices. The computed aerodynamic forces show reasonable agreement with experimental results, both in terms of mean force (vertical, horizontal, and sideslip forces) and the time course over one stroke cycle (lift and drag forces). This alignment with experimental data indicates the accuracy of the CFD model in capturing the complex aerodynamics of insect hovering flight. The study also estimates the hovering energetics based on the computed aerodynamic and inertial torques and powers. This approach provides an integrated view of the energetic costs associated with the complex wing motions in insect flight, contributing to a better understanding of the efficiency and adaptability of these animals [199].
Figure 34.
a) Schematic view of the 3D high-speed video setup used in measuring the hovering flight of drosophila [366]. b) Example images from the three cameras shown in the 3D video schematic with highlighted wings, displaying the ability to record kinematics [366]. c) Schematic of a scaled robotic wing to perform additional flapping experiments using an experimental setup to compare to the visual measurements [366]. d) Closer view of the experimental setup in (c) showing the forces measured in a plane orthogonal to the wingspan [366]. e) Example of thrust values from the aerodynamic force and torque measurements of drosophila during free hovering flight [366]. f) A multiblock grid system of a drosophila with a mesh showing the comparison to a real fruit fly for numerical simulation [199]. g) Example of a grid system for numerical simulation of insect wings with body-fitted curvilinear grids [198]. h) A selection of some possible wing kinematics displayed with the lines representing the wing chord [364]. i) Example of a comparison between optimized (solid line) and measured hawkmoon hovering kinematics [364]. All figures are reprinted with permission; please see the Appendix A.
Figure 34.
a) Schematic view of the 3D high-speed video setup used in measuring the hovering flight of drosophila [366]. b) Example images from the three cameras shown in the 3D video schematic with highlighted wings, displaying the ability to record kinematics [366]. c) Schematic of a scaled robotic wing to perform additional flapping experiments using an experimental setup to compare to the visual measurements [366]. d) Closer view of the experimental setup in (c) showing the forces measured in a plane orthogonal to the wingspan [366]. e) Example of thrust values from the aerodynamic force and torque measurements of drosophila during free hovering flight [366]. f) A multiblock grid system of a drosophila with a mesh showing the comparison to a real fruit fly for numerical simulation [199]. g) Example of a grid system for numerical simulation of insect wings with body-fitted curvilinear grids [198]. h) A selection of some possible wing kinematics displayed with the lines representing the wing chord [364]. i) Example of a comparison between optimized (solid line) and measured hawkmoon hovering kinematics [364]. All figures are reprinted with permission; please see the Appendix A.

The research by Lyu and Sun focused on the power requirements for hovering flight in insects of varying sizes. Their study showed a surprising uniformity in mass-specific power requirements across different insect sizes, suggesting an evolutionary optimization in insect muscle design for efficient flight. This uniformity sheds light on the consistent power needs relative to insect mass, regardless of size variations, offering critical insights into the biomechanics and energetics of insect flight which can be directly adapted to UAVs. The study discovered that despite a massive variation in size (spanning five orders of magnitude in mass), the required power per unit insect mass, or mass-specific power, remained approximately the same across all examined insects. This finding was unexpected as it suggests that both small and large insects expend similar amounts of power relative to their mass during flight. This consistency is attributed to the fact that as insect size decreases, wing speed decreases due to reduced wing length, but wing drag increases due to increased air-viscosity effect, resulting in an approximately unchanged mass-specific power [198].
A study by Fry et al. provided a comprehensive view of the aerodynamics, control, and energetics of hovering flight in Drosophila, or fruit flys. This study highlights the significance of generating substantial aerodynamic forces with a unique U-shaped wing trajectory. Continuous measurement of aerodynamic forces during free flight was performed, revealing insights into how fruit flies manage to generate sufficient lift while maintaining a balance of forces and moments for stable hovering. The research indicates that fruit flies utilize a complex wing motion driven by indirect flight muscles for power and direct control muscles for subtle adjustments. The study also assesses the effects of tethering on flight performance by comparing wing kinematics, forces, and moments under free and tethered conditions. It was found that tethering causes a significant distortion of the stroke pattern, leading to reduced translational forces and a prominent nose-down pitch moment. This distortion is likely due to a disruption of sensory feedback. The research provides a direct calculation of mean muscle power, offering an improved estimate of muscle efficiency and the effects of elastic storage [366].
Gliding Flight
Gliding flight in insects offers insights into evolutionary adaptations, aerodynamics, and energy efficiency that can be applied to small scale UAVs. This section examines various studies that have explored gliding flight in different insect species, highlighting their unique adaptations and implications for understanding flight efficiency.
Adaptive Evolution in Butterflies The investigation of flight behavior divergence in Morpho butterflies across different habitats has been an area of interest for past research. A study by Le Roy et al. revealed varying gliding phases among canopy and understory species, suggesting habitat-driven evolutionary flight adaptations. The kinematic analysis shows that canopy species, like M. cisseis and M. telemachus, exhibit longer gliding phases, indicative of efficient energy use during flight [367].
This study underscores the ecological influence on flight behavior and the adaptive evolution of flight in these butterflies. butterflies from species evolving in the cluttered understory habitat display powerful flapping phases, resulting in high flight speeds and advance ratios. In contrast, evolution in the open canopy resulted in more efficient gliding flight, characterized by reduced descent angles during gliding phases in canopy species. This divergence in wing shape and flight behavior between canopy and understory species has been documented and is indicative of the different selective pressures in these habitats. Canopy species were shown to have increased glide efficiency compared with understory species. This enhanced glide efficiency in canopy species is achieved through different means: some species, like M. rhetenor, have exceptionally high aspect ratio wings, while others enhance their span efficiency. Airflow visualizations confirmed that all gliding butterflies produce stable leading-edge vortices and streamwise wingtip vortices, crucial for efficient gliding. The study illustrates how adaptive evolution, driven by the contrasting microhabitats of canopy and understory, jointly alters wing shape and flight behavior, highlighting the variability in wing design required for different situations and environments [367].
Cespedes et al. explored the correlation between wing shape and flight behavior in Haeterini butterflies. The wing shapes of Haeterini butterflies are adapted for gliding flight in the ground effect along the rainforest floor. This adaptation is characterized by specific wing shape traits that enhance gliding efficiency. The research demonstrated a strong relationship between forewing shape and the relative amount of time spent using gliding versus flapping flight, suggesting that both wing shape and flight behavior have been shaped by selection. Cespedes et al. observed that male butterflies typically have forewing shapes that are aerodynamically more efficient for gliding flight than those of females. This suggests that selection acts differentially on male and female wing shapes, which might be linked to sex-specific flight behaviors and reproductive roles. For example, males often employ gliding flight during mate-searching and/or patrolling territories, while females use flapping flight for host plant searching and oviposition [368]. Both works by Le Roy et al. and Cespedes et al. discuss the development of wing shape in butterflies in response to habitat-specific flight behaviours.
Similar to the work by Le Roy and Cespedes, Mena et al. aimed to understand the morphological adaptations of butterflies in relation to their micro-environments in canopy and understory habitats. The study also found significant associations between vertical stratification and morphological traits, such as wing area and aspect ratio. This research supports the hypothesis that habitat preference influences flight morphology, contributing to the diversity in flying insects [369].
Aerodynamic Performance in Gliding Butterflies Hu et al. also investigated the gliding flight of butterflies, particularly focusing on species like Papilio ulysses and Danaus plexippus. The study reveals significant variations in lift and drag coefficients based on wing shape and angle of attack, highlighting the aerodynamic efficiency of these butterflies in gliding flight. This research has implications for the design of butterfly-like MAVs, suggesting that incorporating aspects of butterfly wing shape and gliding motion could lead to advancements in MAV technology. The research finds significant differences in aerodynamic characteristics between the two butterfly models. Danaus plexippus exhibits a higher lift coefficient and a lower drag coefficient before stalling compared to Papilio ulysses, leading to a lift-to-drag ratio that is twice as large at moderate angles of attack. This suggests that Danaus plexippus may have a flight superiority over Papilio ulysses, particularly in gliding flight. Hu et al. highlighted the importance of flow structures, particularly the LEV, in enhancing the lift force of butterflies. The leading-edge vortex of Danaus plexippus is stronger and more concentrated than that of Papilio ulysses, contributing to its higher lift and lower drag. Additionally, the reduction in lift-curve slope occurs at a lower AOA for Danaus plexippus, which is indicative of a stronger leading-edge vortex formation at these angles [370].
Prior to studies observing morphological adaptations in relation to environments, research was conducted to analyze flight performance in relation to wing shape in butterflies. This study correlates flight performance with wing shape, considering aspect ratio and non-dimensional moment parameters. The findings detail kinematic parameters in representative sequences of various flight modes, including fast climbs, hovering, and others. The butterflies displayed a wide range of complex flight patterns, each representing a sequence of modes and rapid maneuvers. The kinematic data from the film sequences analyzed show considerable inter- and intraspecific differences in flight parameters like wing velocity, amplitude, and stroke plane angle, suggesting a variety of aerodynamic mechanisms at play. The study emphasizes that wing design often represents a compromise between several functions, such as display, defense, thermoregulation, and flight. The research highlights that butterflies employ a range of aerodynamic tricks whose implications on wing design and flight performance are still being unraveled [371].
Dragonfly Gliding Mechanics and Thermoregulation Additional work provides an extensive analysis of the gliding flight mechanics in the dragonfly Sympetrum sanguineum. This study emphasizes the role of gliding in thermoregulation, showcasing how dragonflies utilize gliding for convective cooling while minimizing heat production. Wakeling and Ellington filmed the free gliding flight of Sympetrum sanguineum in a large flight enclosure and reconstructing the glide paths to analyze the aerodynamic forces. The reconstruction showed that the flights involved accelerations, and when the acceleration was constant, the lift and drag forces acting on the dragonfly could be accurately determined. The drag coefficient at zero angle of attack was found to range between 0.07 and 0.14, indicating that dragonfly wings exhibit exceptional steady-state aerodynamic properties compared to the wings of other insects. The maximum lift coefficient recorded from these glides was 0.93, though it’s noted that this may not be the maximum possible from the wings. The work extends to measuring lift and drag forces from isolated wings and bodies of Sympetrum sanguineum and the damselfly Calopteryx splendens in steady airflow. The maximum lift coefficients for these species were 1.07 and 1.15, respectively. These values are greater than those recorded for most other insects, except locusts, demonstrating the superior aerodynamic performance of dragonfly wings [372].
Summary of Flight Modes in Insects
The study of flight modes in insects, specifically hovering and gliding, reveals a fascinating intersection of biomechanics, aerodynamics, and ecological adaptation. Hovering flight, exemplified by its high energy demand and intricate wing kinematics, showcases an evolutionary mastery of precision and power management in insects like fruit flies and bumblebees. In contrast, gliding flight, observed in various butterfly and dragonfly species, exemplifies an energy-efficient locomotion strategy, utilizing passive aerodynamic forces and morphological adaptations for sustained flight. These flight modes not only reflect the diverse ecological niches inhabited by these insects but also offer valuable insights for biomimetic applications in aerial vehicle design, emphasizing efficiency and adaptability.
4. Energy Harvesting
4.1. Soaring
4.1.1. Dynamic Soaring
Dynamic soaring is a sophisticated flight technique, predominantly utilized by seabirds such as the albatross, enabling them to harness wind gradients for energy-efficient, long-distance oceanic travel. This method exemplifies the remarkable aerodynamic adaptations of avians, allowing them to exploit the atmospheric boundary layer over the sea where wind speeds vary significantly with altitude. These birds skillfully utilize the wind’s kinetic energy, transitioning between high and low wind zones, to sustain flight without the need for constant wing flapping [39,68,102,139,154,157,158,160,172,173].
This flight technique is not merely a product of evolutionary adaptation but also a testament to the complexity of avian flight mechanics. Dynamic soaring enables birds like the albatross to travel upwards of 15,000 thousand kilometers, a feat of endurance and efficiency [68]. The ability to utilize natural wind patterns for flight without substantial energy expenditure reflects a profound understanding of the physical principles of flight and energy utilization in the natural world [158].
Figure 35.
Schematic views of albatross dynamic soaring flight and its kinematic and kinetic breakdown [97].
Figure 35.
Schematic views of albatross dynamic soaring flight and its kinematic and kinetic breakdown [97].

Albatrosses are known for their remarkable ability to fly extensive distances, approximately 15,200 km, with minimal wing flapping, showcasing an extraordinary example of energy-efficient flight [68]. These birds, with a wingspan of about 3.5 m and weighing around 8.5 kg, maintain ground speeds exceeding 35 m/s [107], demonstrating their adeptness in dynamic soaring. The technique’s efficiency is so profound that it allows these birds to maintain high speeds for up to eight hours with little to no wing flapping [39].
Bio-Inspired Energy-Harvesting Mechanisms in Dynamic Soaring
Dynamic soaring is not just an avian flight technique but also a source of inspiration for small UAV design. This section delves into the energy-harvesting mechanisms inherent in dynamic soaring, emphasizing the energy transfer from the wind to the flyer. The classical four-phase Rayleigh cycle, comprising upwind climb, downwind turn, downwind dive, and upwind turn, serves as a framework to analyze energy gain. The study finds that in the air-relative frame, significant energy gain occurs during lower altitude climbs and dives due to large wind gradients, while in the inertial frame, it results from the lift vector inclined towards the wind speed direction during higher altitude maneuvers [143]. Exploring various dynamic soaring patterns, Liu et al. categorized them into closed (’O’ and ’8’ shapes) and traveling patterns (’’, ’’, ’C’, and ’S’ shapes). The relationship among these patterns was examined, validating the classification by reviewing past studies on dynamic soaring trajectories, which reveals a comprehensive understanding of optimal UAV flight paths [143]. The paper suggests that real-time wind field mapping on board a small-scale UAV is a promising study trend for autonomous dynamic soaring in real flight. Additionally, robust guidance and control for UAVs tracking optimized trajectories, especially in turbulent wind fields and near the surface, is a challenging yet vital area of research.
The albatross, distinguished by its large wingspan and specialized wing morphology, exemplifies the mastery of dynamic soaring. It employs strategic maneuvers, climbing into high wind speeds at an angle and then descending into lower wind speeds, to efficiently extract energy from the wind gradient. This method is a complex interplay of aerodynamics and bird physiology, allowing the albatross to sustain prolonged flights with minimal metabolic energy expenditure [154]. These types of birds also tend to live most of their lives in flight and only return to land to mate for short periods [173]. Pokhrel and Eisa hypothesized that dynamic soaring in albatrosses can be modeled as an autonomous, real-time extremum-seeking system. This concept suggests that albatrosses, and potentially UAVs, can dynamically optimize their flight path in real time without explicit knowledge of the wind profile or a predefined objective function. This is supported by the experimental observations of albatrosses, which demonstrated their ability to conduct dynamic soaring in real-time, autonomously adjusting to changing wind conditions. The work detailed the implementation of two extremum-seeking control structures incorporating the dynamic soaring problem. The control structures aimed to autonomously and in real-time find an energy-neutral or near-neutral dynamic soaring trajectory. This approach contrasts with the traditional optimal control problem configurations used in most dynamic soaring research, which often result in non-real-time algorithms and control solutions [154].
The efficiency of dynamic soaring significantly reduces the metabolic cost associated with long-distance travel in birds. The albatross’s proficiency in dynamic soaring is not just an aerodynamic achievement but also a result of physiological adaptations that are optimized for extended periods of flight. These adaptations are evident in their specialized wing structure and flight muscle composition which were described early on by Leonardo da Vinci [157].
Comparing dynamic soaring techniques across different seabird species reveals a variety of adaptations tailored to specific ecological niches and energy requirements. Differences in wing size, shape, and flight patterns across species highlight the diverse evolutionary strategies employed by seabirds to maximize energy efficiency in flight. Such comparative studies enhance our understanding of the biomechanics and ecological significance of dynamic soaring [139]. The research by Kempton et al. involved analyzing fine-scale flight trajectories reconstructed from bird-borne video data using a simplified flight dynamics model. The team developed a new metric, the horizontal wind effectiveness, to quantify how effectively the birds’ flight harvests energy from a shear layer. This metric was evaluated empirically for the trajectories and further assessed in relation to long-range, GPS-logged foraging trajectories. The results show that Manx shearwaters optimize their flight to increase the opportunity for dynamic soaring during favorable wind conditions. The birds’ undulations were found to be phased with their horizontal turning to optimize energy harvesting. This behavior was more pronounced during outbound flights when their destination was less constrained, suggesting a strategic modification of flight patterns to maximize energy gain [139].
The principles of avian dynamic soaring offer valuable insights for the design of energy-efficient aerial vehicles. Emulating the albatross’s soaring strategy could lead to significant advancements in drone technology, potentially enhancing flight duration and reducing energy consumption. This bio-inspired approach is particularly relevant for the development of fixed-wing amphibious drones, where optimizing energy efficiency is a key design consideration [139,143,154,172]. Wang et al. provided an in-depth analysis of the dynamic soaring technique, breaking it down into a four-phase cycle: windward climb, high altitude turn, leeward descent, and low altitude turn. The authors focused on the energy-extraction mechanisms during these phases, analyzing the variations in gliding performance based on different environmental and aerodynamic variables. The work utilized two types of reference frames, inertial and non-inertial, to deduce the energy harvesting equation. It examines the forces and angles involved in dynamic soaring and compares the energy harvesting mechanisms in these two frames. This approach helps to better understand the contribution of wind gradient and lift in the energy-extraction process. The results demonstrated that the lift force plays a significant role in energy harvesting in the inertial frame, while in the non-inertial frame, the energy comes primarily from the wind gradient, which generates a dynamic soaring thrust. Wang et al. suggested that the gradient wind is essential for dynamic soaring, as it can provide a continuous motion and energy source for the albatrosses [172].
Figure 36.
a) Schematic of dynamic soaring maneuvers conducted by an albatross in the presence of a wind shear [154,158]. b) Schematic to demonstrate potential soaring capabilities of a UAV system that can transition between urban environments and coastal regions [147]. c-e) Examples for potential dynamic soaring conditions, showing c)a large wind gradient over a sea surface, d) gradient soaring over thinner wind shears that ensue from waves, and e) a sharp ridge separating fast-moving and slow-moving air creating a gradient [147]. f) Leonardo da Vinci’s sketch illustrating the dynamic soaring flight from right to left [157]. g) Leonardo da Vinci’s sketch of winds interacting with vertical cliffs and deflecting upward [157]. All figures are reprinted with permission; please see the Appendix A.
Figure 36.
a) Schematic of dynamic soaring maneuvers conducted by an albatross in the presence of a wind shear [154,158]. b) Schematic to demonstrate potential soaring capabilities of a UAV system that can transition between urban environments and coastal regions [147]. c-e) Examples for potential dynamic soaring conditions, showing c)a large wind gradient over a sea surface, d) gradient soaring over thinner wind shears that ensue from waves, and e) a sharp ridge separating fast-moving and slow-moving air creating a gradient [147]. f) Leonardo da Vinci’s sketch illustrating the dynamic soaring flight from right to left [157]. g) Leonardo da Vinci’s sketch of winds interacting with vertical cliffs and deflecting upward [157]. All figures are reprinted with permission; please see the Appendix A.
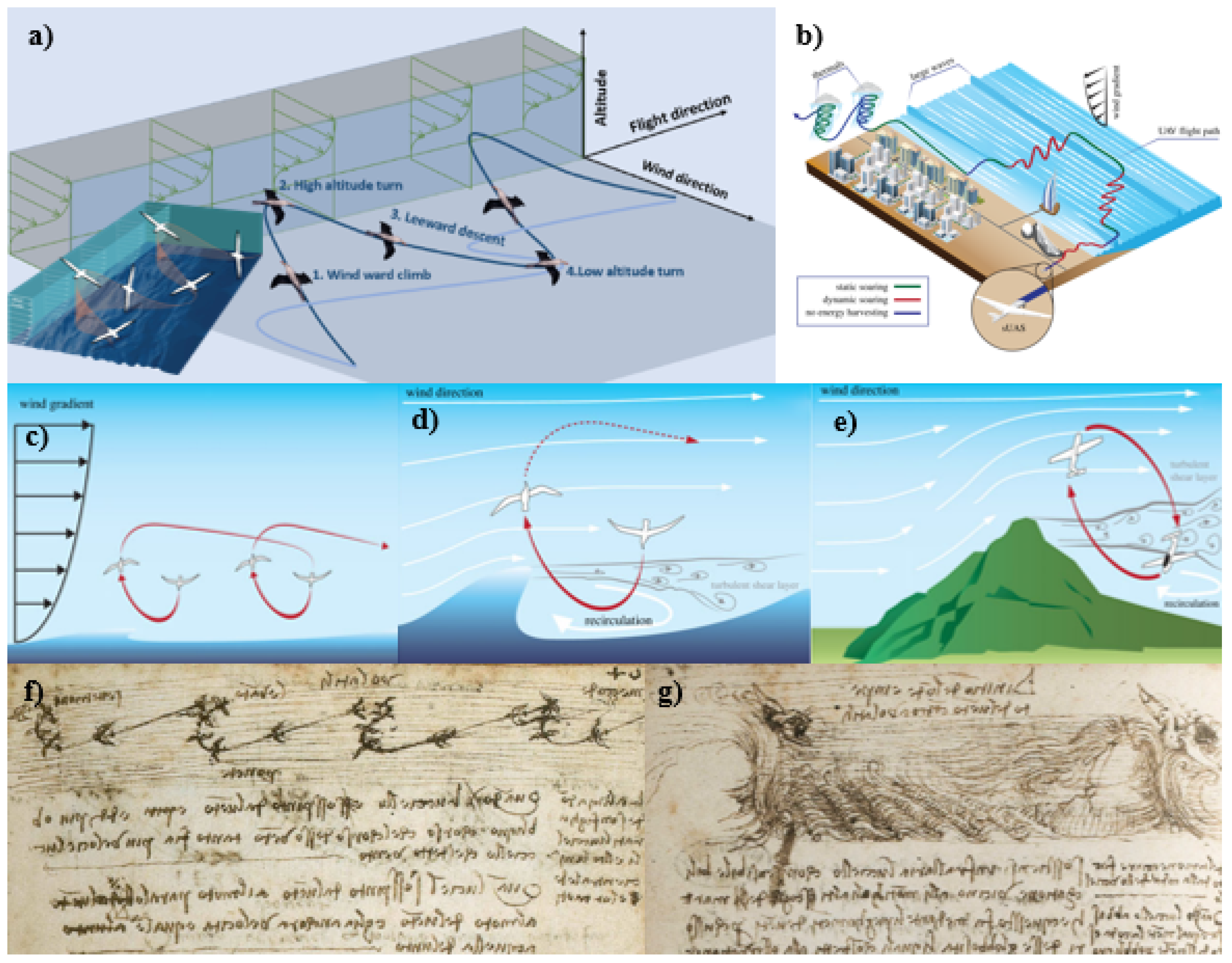
The principle of dynamic soaring hinges on utilizing wind behavior and wind shears. Albatrosses gain height above the ocean by exploiting wind speed and direction, a method further enhanced by interactions between wind and ocean waves, known as wave-slope soaring [102]. These birds use updrafts generated by waves to gain height and energy, effectively using the wind’s shear characteristics and vertical motions. Dynamic soaring in albatrosses involves specific movement categories: large-scale movements for long-distance flight at steady-state cruise speeds and small-scale, highly dynamic maneuvers necessary for dynamic soaring [102]. These movements exemplify the bird’s ability to adapt its flight mechanics to different environmental conditions, maximizing energy harvesting from natural wind patterns.
Multiple research studies have been carried out on albatrosses to investigate their ability to dynamically soar, their flight routes, and their overall flight characteristics. These studies have demonstrated that albatrosses can be classified as highly capable migratory birds. Due to their preference for flying over oceanic environments, various factors such as wind patterns, the effects of water waves, and ocean tides can significantly impact their flight performance. Additionally, the heat exchange within different boundary layers of the ocean can also influence the flight behavior of an albatross, irrespective of whether it is day or night. These aforementioned principles can be similarly applied to fixed-wing drone systems in order to harness energy from wind shears. By employing a comparable dynamic soaring pattern with fixed-wing drone systems, it is possible to achieve sustained level flight for extended periods of time. This idealized form of dynamic soaring can be most effectively described and governed by Mir’s equations of motion, which provide a framework for understanding the three-dimensional gliding flight of a fixed-wing drone.
Modeling and Application in UAV Design
The understanding of albatross flight mechanics has significant implications for UAV design, particularly in fixed-wing amphibious drone systems. The dynamic soaring patterns of albatrosses can be emulated to achieve sustained level flight in drones, enhancing their operational efficiency [147,243]. The application of these principles is governed by Zhao’s equations of motion, which describe the 3D gliding flight of fixed-wing drones. These equations take into account factors like wind shear, bank angle, and lift and drag forces, crucial for mimicking the flight characteristics of the albatross [243].
Of these equations, the mathematical modeling utilized three degrees of freedom to describe the motion. This model can be derived from (i,j,k) with the state x=(V, , , z, x, y). where V is the drone airspeed, is the angle between x and the projection of the airspeed V in the xy plane and is known as the air-relative heading angle, is the angle between V and the xy plane and is positive nose up and is known as the air-relative flight path angle [243]. The equations of motion shown by Mir et al. can be utilized to help better mimic the flight characteristics of an albatross, which in turn can be applied to fixed-wing systems. By employing these methods, a more efficient flight method can be achieved by utilizing an already naturally occurring dynamic soaring method to boost the effective range of this specific drone system. The energy harvesting technique in UAVs relies on extracting energy from wind shear near the Earth’s surface. Accurately modeling the wind shear gradient is essential for optimizing drone velocities and enhancing the effectiveness of dynamic soaring in UAV applications [243].
Wind Shear and Soaring Performance Wind shear, a crucial factor in dynamic soaring, is the difference in wind speed and sometimes direction over short atmospheric distances. Its characteristics, especially in relation to altitude, play a fundamental role in the performance of soaring. The modeling of wind shear, particularly over the sea, often follows an exponential model, as it is critical for the application of dynamic soaring techniques in UAVs [243]. Wind shear velocity over sea and ridges is often represented via an exponential model and can be seen in Mir et al.’s work, along with the wind shear gradient [243]. These equations for wind shear are used directly in the equations of motion for a drone and can have a great impact on the dynamic soaring performance of the model. Research into the dynamic soaring techniques of different seabird species has provided valuable insights into evolutionary adaptations. The comparative studies indicate that variations in wing structure, body mass, and flight behaviors are evolutionary responses to environmental demands. These adaptations enable species-specific optimization of flight efficiency and energy harvesting [139].
Theoretical frameworks have been developed to understand and model the dynamic soaring of seabirds. These models are critical for translating the principles of avian soaring into UAV design. They incorporate variables like airspeed, wind gradient, and turning radius, which are essential for replicating the soaring dynamics in mechanical systems. Mathematical modeling of dynamic soaring not only enhances our understanding of avian flight mechanics but also serves as a foundation for bio-inspired UAV design [172,243].
Environmental factors play a significant role in the optimization of dynamic soaring. The interaction between wind patterns, oceanic currents, and wave dynamics forms a complex system that seabirds exploit for energy-efficient flight. Understanding these interactions is crucial for developing UAVs that can utilize similar environmental advantages, thus optimizing their flight patterns for energy harvesting and efficiency [172].
The study of avian dynamic soaring opens up new possibilities in UAV technology. Future research is directed towards developing UAVs that can autonomously navigate and harvest energy from natural wind patterns, much like seabirds. This involves integrating advanced sensors, control algorithms, and energy-efficient designs to create UAVs capable of prolonged autonomous operations in various environmental conditions [147,243]. While the study of avian dynamic soaring has provided significant insights, it also presents unique challenges. The complexities of accurately simulating the wind conditions and the bird’s response to these conditions in a laboratory setting pose considerable difficulties. Additionally, translating these natural flight mechanisms into UAV design requires a deep understanding of both aerodynamics and avionics, a field that is still evolving [158].
Interdisciplinary Approaches and Collaborative Research The interdisciplinary nature of dynamic soaring research, involving aerodynamics, ecology, biology, and engineering, underscores the importance of collaborative efforts. Integrating knowledge from these diverse fields is essential for advancing our understanding of avian flight mechanisms and their application in UAV technology. Collaborative research efforts are paving the way for more sophisticated and energy-efficient UAV designs [139].
4.1.2. Thermal Soaring
Thermal soaring, a fundamental energy-harvesting strategy in avian flight, involves the utilization of rising thermal currents to gain altitude with minimal energy expenditure. This technique is not only pivotal in understanding bird flight mechanics but also plays a crucial role in the design and operation of UAVs. Birds exploit this natural phenomenon to conserve energy during long flights, effectively utilizing environmental resources. The study of thermal soaring extends from detailed aerodynamic analyses to evolutionary aspects, contributing to a broader understanding of flight mechanics and energy efficiency. It encompasses various disciplines, including meteorology, aerodynamics, and behavioral ecology, providing insights into the adaptive strategies of birds and their implications for UAV technology. The intricacies of thermal soaring, such as the detection and prediction of thermals, wind estimation, and the adaptability of birds to different soaring conditions, are essential for advancements in both biological understanding and technological innovation [127,136,140,149,150,151,163,164,168,170,171,174,373].
Historical Evolution of Thermal Soaring
The evolution of thermal soaring, a key energy-saving flight mechanism in birds, can be traced back to the early stages of avian evolution. This adaptive strategy has been a subject of interest, particularly in understanding how early birds like Sapeornis chaoyangensis developed soaring capabilities. Research indicates that these early birds displayed characteristics similar to modern soarers, suggesting an early adaptation to thermal soaring as an energy-efficient mode of flight. Studies like those by Serrano and Chiappe provide evidence of this evolutionary trajectory, revealing that even during the Cretaceous period, birds had already begun to exploit rising air currents for energy-efficient travel. This is indicated by morphofunctional analysis, including aspects like wing shape, lift-to-drag ratios, and gliding speeds compared to modern soaring birds. The study proposes that the ability for thermal soaring had evolved among large early birds by at least 125 million years ago. The development of thermal soaring in birds is closely tied to the evolutionary pressures of flight, where energy conservation is a critical factor. This evolutionary perspective sheds light on the origins and adaptations of thermal soaring, emphasizing its long-standing significance in avian flight dynamics [163].
Modeling Thermal Soaring
Many studies have developed analytical models of thermal soaring, which serve as a framework for UAV path planning and emphasize energy efficiency. Khaghani et al.’s model, validated through simulations, highlights the potential of using thermal soaring principles in UAVs for various applications. The researchers presented a simple hybrid control strategy designed to optimize the path for traveling between thermals without flapping, thereby maximizing energy efficiency. Khaghani et al. presented an analytical model for thermal soaring, aiming to replicate the flight behavior of birds using thermals for energy-efficient flying. This model is especially pertinent for UAV design in tasks like urban delivery and search and rescue and addressed previous limitations by considering horizontal wind and downdraft effects. The simulation phase, which included parameters set according to real conditions observed in Griffon Vulture and Magnificent Frigatebird, demonstrated the model’s accuracy and applicability. This validation emphasizes the potential of the model for guiding flying robots in a similar energy-efficient manner [140].
Other studies on crane migration and seabirds within wind farm areas showed the impact of meteorological factors on thermal soaring. These findings challenge previous assumptions about avian sea crossing and underscore the influence of temperature and wind conditions on soaring behavior [149,171]. Pekarsky et al. found that contrary to previous assumptions, cranes can and do utilize thermal updrafts over the sea to facilitate their migratory crossings, effectively debunking the paradigm that obligate soarers avoid sea crossings due to the absence of thermals. The study observed that cranes adapt their flight mode to the available atmospheric conditions, merging soaring and flapping flight to maximize energy efficiency and minimize migration time. This behavior indicates a highly flexible migration strategy that responds adaptively to the environmental conditions encountered en route [149]. Van Erp et al. found that thermal soaring occurs over the North Sea and is influenced by positive temperature differences between the sea surface and the air, with north and north-westerly winds enhancing these conditions. The research showed that birds flew at higher altitudes during thermal soaring than during non-soaring flight, increasing the proportion of flight time within the rotor-swept zone of wind turbines. The occurrence of thermal soaring was similar inside and outside of wind farm areas, suggesting that the presence of wind farms does not deter birds from utilizing thermal updrafts for soaring. Research on different bird species over central Israel and the osprey’s migration behavior also highlight the diversity in soaring strategies. These studies emphasize the role of wing loading, meteorological conditions, and adaptability in energy strategies among thermal soaring migrants [164,170]. The research by Shamoun-Barnes et al. found that each species exploited thermal convection differently, with maximum migration altitudes for each species being significantly explained by the model’s predictions of the upper boundary of thermal convection. This indicates a relationship between the birds’ migration altitudes and the conditions of thermal convection, which were affected by topography as well as changing atmospheric and environmental conditions. The study utilized thermal depth and intensity predictions from the ALPTHERM model to analyze the variance in migration altitudes across species [164]. Additionally, the findings by Thorup et al. suggest that ospreys, and potentially other thermal soaring migrants, are equally or even more prone than other migrants to stop and rest in rainy weather. This behavior may be attributed to the detrimental effect of rain on the development of thermals, forcing these soaring birds to switch to energetically costly flapping flight [170].
Harel’s research on Eurasian griffon vultures delved deeper into the cognitive processes involved in thermal soaring decision-making. The study used GPS data to analyze how these birds decide when to depart from thermal columns. It was found that short-term experiences, particularly from the last 20 minutes, played a significant role in their decision-making, rather than relying solely on long-term past experiences. This indicates a sophisticated integration of environmental cues and immediate feedback in their flight strategy and implies that vultures dynamically adjust their soaring behavior based on the most relevant environmental information available. The vultures were more likely to leave a thermal when the current climb rate was higher than recent experiences, suggesting an adaptive response to changing thermal conditions. The findings suggest that thermal soaring birds like Eurasian griffon vultures possess the cognitive ability to integrate multiple sources of environmental information over short time scales to make flight decisions. This capability allows them to effectively navigate and utilize the dynamic thermal landscape, enhancing their flight efficiency and reducing the energy costs associated with long-distance migration or foraging flights. This research not only contributes to understanding the decision-making processes in birds utilizing soaring flight but also provides insights for developing autonomous UAVs capable of similar dynamic decision-making [136]. Tallman et al.’s machine learning approach for thermal forecasting demonstrates the potential of using flight records, weather data, and satellite imagery for thermal prediction. This technology not only aids in free flight but also contributes to weather pattern prediction and UAV flight path optimization. The study developed a machine learning model based on convolutional neural networks, specifically utilizing the U-net architecture, to predict thermal locations. The model was trained on a dataset comprising satellite imagery, weather conditions, and recorded flight data from gliders that encountered thermals. The team aimed to provide a more accurate method of forecasting thermal locations to assist pilots in planning energy-efficient flights. The model demonstrated promising results, achieving a loss of .02% on validation samples, indicating high accuracy in thermal prediction. The research successfully created a custom dataset with thermal truth labels, which can be further expanded for future studies. One of the significant achievements of this research is the end-to-end approach for predicting thermals, combining satellite plus weather data, marking a considerable advancement in free flight navigation and safety [168].
The study of eagles and hummingbirds by Dutta and Chaugule provided insights into the aerodynamic efficiency and control surface dynamics in birds during thermal soaring. Their work discussed how eagles utilize thermal updrafts and orographic updrafts to subsidize their flights. Eagles are highlighted for their ability to switch between different modes of flight in response to atmospheric conditions. The distribution and availability of thermal and orographic updrafts, influenced by atmospheric perturbations and terrain topography, play a crucial role in the flight strategy of these raptors. The study emphasizes the eagle’s aerodynamic traits, such as slotted wings and control surfaces, which contribute to its high maneuverability and efficient energy use during flight [373]. An additional study explored glide polars and the performance of bird wings in thermal soaring. This study looked into the dynamics of glide polars, the performance of birds’ wings, and how different species adjust their flight strategies based on wing parameters. The research discussed various strategies employed by birds to maximize the benefits of thermal soaring, including the selection of optimal thermals and efficient circling techniques to gain maximum altitude. These strategies, informed by extensive observations and simulations, provide valuable insights into the development of autonomous soaring algorithms for UAVs, aiming to replicate the energy efficiency achieved by birds through millions of years of evolution [127].
Penn et al. introduced a novel wind tunnel configuration designed to study non-flapping wind hovering and soaring bird flight. This approach addresses the challenge of replicating the complex dynamic interactions between birds and natural wind patterns, including the turbulence and thermal updrafts they exploit for energy-efficient flight. The modifications to the wind tunnel enable the simulation of a range of outdoor conditions, such as those experienced by kestrels known for their windhovering behavior. This work also focuses on the stability of kestrels and other birds in hovering flight, despite turbulent atmospheric conditions. Investigating these control strategies, which are not yet fully understood, could lead to significant advancements in UAV design, particularly in windy conditions. The study details the required modifications to the RMIT Industrial Wind Tunnel for simulating natural bird wind hovering conditions. The initial results from flight trials with kestrels demonstrate the utility of this approach, highlighting its potential for uncovering new insights into bird behavior and flight mechanics. This controlled environment allows for an in-depth analysis of avian flight mechanics, critical for understanding the dynamics of thermal soaring. These trials marked a significant milestone in avian flight mechanics research, demonstrating the capability of the wind tunnel to support detailed studies of bird flight behaviors [150].
Pennycuick presented an interesting study on the field observations of thermals and thermal streets, challenging the classical theory of cross-country soaring in which an optimum gliding speed between thermals was suggested. The research, conducted over the Serengeti, involved measuring vertical air velocity to understand the dynamics of thermals and their effect on soaring flight and showed that glider pilots and birds often fly slower than this theoretical optimum speed, yet achieve faster cross-country speeds. This discrepancy was analyzed through a survey of real thermals over the Serengeti National Park, indicating that the ability to select stronger thermals and the presence of linear thermal streets can significantly enhance cross-country soaring efficiency [151].
Weinzierl introduced a method for wind estimation based on high-frequency GPS data from birds during thermal soaring. This approach, validated against ECMWF weather reanalysis data, demonstrated a systematic approach to evaluate wind conditions by observing the thermalling maneuvers of birds and provided accurate wind and airspeed estimates at flight locations. The methodology assumes a fixed horizontal mean wind speed and employs a maximum-likelihood optimization to minimize the error in wind estimate over short segments representing individual thermal loops. This study provides a novel perspective on understanding the interaction between wind patterns and bird flight behavior, with potential implications for ecological research and UAV design [174].
Interdisciplinary Insights and Future Directions
The research on thermal soaring, bridging the gap between biology and technology, has provided significant insights into both natural and engineered flight. Studies ranging from the evolutionary aspects of early birds to the latest in UAV design demonstrate how biological principles can enhance technological advancements. The future of thermal soaring research holds immense potential, with an emphasis on refining UAV design inspired by avian flight mechanics, and improving thermal prediction models for more efficient and adaptable flight paths. This interdisciplinary approach not only deepens our understanding of avian behavior but also paves the way for innovative solutions in aerial technology, leveraging the natural world’s efficiencies. As we continue to explore and integrate these insights, we can expect a new era of UAVs that are more energy-efficient, adaptable, and effective in varied environmental conditions [140,149,150,163,174].
4.1.3. Slope Soaring
Slope soaring, a critical flight mechanism in birds, harnesses environmental updrafts for energy-efficient travel, crucial for long-distance migration and conservation of energy. This strategy is extensively used by various bird species, including albatrosses and vultures, and is adapted to diverse environmental conditions. Albatrosses are particularly adept, utilizing wind-shear and wave-slope soaring for effective energy harvesting, even under challenging wind conditions [98,102]. Vultures exploit slope soaring for scavenging and long flights, minimizing energy expenditure while maximizing flight efficiency [153,164]. Petrels, alongside albatrosses, demonstrate remarkable use of slope soaring over sea waves, utilizing this technique for extensive foraging trips across the ocean [98,159]. The ecological significance of slope soaring is profound, influencing hunting, scavenging, and migratory behaviors. This flight strategy offers crucial insights into avian ecology and has significant implications for conservation efforts, highlighting the interplay between avian flight mechanics and environmental factors [132,144,164].
The efficiency of slope soaring, particularly in vultures, is highlighted by their minimal increase in heart rate during soaring and gliding flights, indicating exceptional energy efficiency [132]. Griffon vultures, for instance, exhibit distinct low-altitude slope soaring patterns along cliffs, often in a figure-eight shape, differentiating it from other flight types like thermal soaring [141]. The ecological significance of this flight mechanism is evident in their hunting and migratory behaviors, where efficient use of wind currents against slopes conserves energy for long flights [153]. Turkey Vultures show adaptability in their reliance on thermals and updrafts for migration, altering flight modes with atmospheric conditions and demonstrating a mix of thermal and slope soaring strategies [145]. Furthermore, research on Andean Condors reveals their reliance on different uplift types, including slope soaring, with significant time spent in thermals and orographic uplift, indicating their adaptability to seasonal changes [175].
Additionally, slope soaring is observed in avian species like brown pelicans, which utilize a special case known as wave-slope soaring. This method involves flying over ocean waves to take advantage of updrafts, significantly reducing the cost of transport. The study by Stoked and Lucas, employing potential flow theory and an energetics-based approach, found that wave-slope soaring can lead to a 60-70% reduction in transport costs under typical ocean conditions, with potential enhancements based on flight height and wave amplitude. They developed a theoretical model to quantify the energy savings associated with wave-slope soaring. This model considers the airflow induced by near-shoaling ocean waves and the consequent impact on the aerodynamics of a soaring bird. The study analyzes the brown pelican’s flight over near-shoaling coastal waves, emphasizing the cost of transport as the primary metric for energy efficiency. This energy-efficient flight mode is crucial for brown pelicans, aiding their survival and foraging over large home ranges. The insights gained from this study contribute to our understanding of avian flight behaviors and have applications in designing energy-efficient unmanned aerial vehicles [166].
Slope soaring also extends to non-avian species, notably in Pteropus bats, also known as flying foxes. These bats demonstrate morphological adaptations for soaring, such as lower wing loadings and broader wingtips, which result in larger wing areas. Such adaptations are crucial for exploiting thermals, even when weak, thereby minimizing the energy costs of flight. The research by Norberg et al. involved comparing two species pairs from American Samoa and the Comoro Islands, with one species in each pair using soaring flight and the other relying mostly on continuous flapping flight. The team analyzed the morphological data of these species to calculate theoretical glide polars and circling envelopes for both soaring and non-soaring species. The study also compared circling performance, crucial for soaring in thermals and maneuverability. The findings suggest that the soaring species have longer wings and lower wing loadings than the non-soaring species. This observation aligns with the hypothesis that morphological adaptations for improved soaring ability help minimize energetic flight costs. The soaring bats demonstrated lower sinking speeds during gliding, higher glide ratios, and smaller radii of turn, all favored by low wing loading. The morphological differences, although small, indicate adaptations for better gliding/soaring performance. These findings underscore the ecological implications of flight ecology in Pteropus bats, showing how diurnality and morphological adaptations play a pivotal role in their energy-efficient soaring behavior [148].
Weather conditions significantly impact slope soaring. Golden Eagles show a preference for migrating under conditions conducive to thermal development. Age and seasonal variations play a crucial role in influencing their response to orographic lift, with younger eagles showing different patterns compared to older ones [131]. Duerr at al. collected GPS locations from 47 golden eagles along 83 migration tracks and paired each location with meteorological data. The research aimed to determine the weather correlates of soaring flight modes and understand how eagles respond to environmental conditions that favor the development of thermals and orographic lift. The findings show that the migratory flight of golden eagles was positively associated with weather conditions promoting thermal lift, both during spring and fall migration across all ecoregions and age classes. Interestingly, the eagles showed different migratory responses to weather conditions that encourage orographic lift, varying by age and season. The study demonstrates that eagles preferentially migrate when conditions are suitable for thermal development, suggesting they seek to maximize flight efficiency and minimize energy expenditure [131].
Turkey Vultures, known for their adaptability, utilize a combination of thermal and slope soaring, adjusting their flight strategies based on environmental variations [145]. Mandel’s dissertation presented various findings, including how birds adjust their migratory paths based on the availability of rugged terrain for slope soaring. It also discusses how these adjustments differ depending on the weather and terrain phenomena. For instance, during spring migration, birds engage less in slope-soaring and increase their activity period. On breeding grounds, birds do not alter their course for any weather or terrain phenomena, suggesting a focus on providing food to young without regard to route optimization [145]. Duriez et al. talk about how the presence of strong thermals on sunny days facilitates easier soaring flight for these birds, whereas cloudy conditions might lead to less efficient thermal uplift, requiring more flapping and increased energy expenditure [132]. Their study focused on two species of vultures, equipped with bio-loggers that recorded their GPS positions, 3-axis accelerations, and electrocardiogram data. The experiments were conducted at a site that offers natural soaring conditions, allowing the birds to take off and land freely during the recording sessions. The study aimed to understand how heart rate varies with flight mode and how weather conditions, particularly solar radiation, influence soaring activity. The results indicate that heart rate increased threefold during take-off and landing, characterized by prolonged flapping flight, compared to baseline levels when the birds were perched or walking. However, within 10 minutes after the initial flapping phase, heart rate in soaring and gliding flight dropped to levels slightly lower than baseline, suggesting that soaring and gliding are energetically cheap modes of flight. The study also found that weather conditions significantly affected flight performance, with heart rate being noticeably higher during cloudy conditions when thermal ascending air currents were less available [132]. Furthermore, studies suggest that the height of the planetary boundary layer correlates with navigation strategies in these birds, indicating slope soaring along mountain ridges under specific atmospheric conditions [145].
Golden eagles and turkey vultures exhibit distinct preferences for uplift types during migration, a difference that is closely linked to their morphology and migration timing. Golden eagles prefer stronger orographic uplifts due to their larger size and later migration timing when thermal uplifts are less prevalent. Turkey vultures, in contrast, show a higher likelihood of using strong thermal uplifts, leveraging their lighter build and earlier migration timing to take advantage of the abundant thermals [130]. Bohrer et al. showed that golden eagles display a higher degree of flexibility in their flight strategies compared to other large raptors. They often modify their flight behavior in response to varying environmental conditions, demonstrating an exceptional ability to adjust to different types of uplift. This adaptability is particularly evident during their long migratory flights, where they encounter a wide range of atmospheric conditions. In contrast, turkey vultures have been observed to rely more heavily on thermal soaring, often using a mix of thermal and slope soaring strategies. This reliance on thermals is attributed to their lighter body structure and the need for energy-efficient travel over long distances [130]. The variations in flight strategies between these species underscore the complex interplay between physical morphology, environmental conditions, and energy expenditure, offering valuable insights into the adaptive mechanisms of these birds for conservation efforts.
In Eastern North America, the migratory performance of soaring birds such as Golden Eagles is heavily influenced by weather conditions, particularly atmospheric thermals and wind patterns. The flight path straightness and overall migration efficiency of these birds are significantly affected by these conditions, emphasizing the importance of favorable weather for efficient migration [146]. The study by Miller et al. analyzed GPS tracking data from 18 adult, 13 sub-adult, and 15 juvenile golden eagles migrating across eastern North America. By correlating these tracks with atmospheric data, the study aimed to understand how conditions supportive of thermal convection and orographic lift affected the birds’ flight efficiency and path straightness. Golden eagles’ migratory performance, measured in terms of flight path directness and efficiency, was significantly influenced by weather conditions conducive to thermal updrafts. Eagles adjusted their migratory behaviors based on the availability of thermals and orographic lift, showcasing flexibility in flight strategies to optimize energy expenditure. The study noted that atmospheric stability and the presence of lateral winds, particularly during spring migration, had a notable impact on the birds’ ability to maintain direct migratory paths. Golden Eagles in North America adjust their migratory routes and behaviors in response to seasonal variations. They tend to use more thermal soaring in seasons with strong thermal activity and switch to more slope soaring or gliding in less favorable thermal conditions [146].
Andean Condors similarly demonstrate how seasonal changes impact the costs of moving between thermals. During seasons with less favorable thermal conditions, these birds show an increased reliance on slope soaring and other energy-saving flight strategies. This adaptability highlights the birds’ capability to optimize their energy expenditure across different environmental conditions. The research by Williams et al. also indicated a correlation between seasonal changes and the flying behavior of Andean Condors. During winter, when thermal uplift is less available, these birds tend to spend more time in orographic uplifts, adjusting their flight patterns to the changing availability of natural lift sources [175]. This seasonal variation in flight strategies underscores the importance of understanding the ecological dynamics of these birds for effective conservation efforts.
Pennycuick’s 1971 study offered a comprehensive look at the soaring behavior of various East African bird species, with a particular focus on Rüppell’s Griffon Vultures (Gyps rueppellii). These vultures prominently utilize slope lift along nesting cliffs, an adaptation that is crucial for their scavenging lifestyle in cliff-rich habitats [152]. The importance of slope and wave lift is emphasized for these birds, especially given their need to cover vast areas during scavenging and migration activities. Further insights from the study indicated that these vultures, along with other bird species in the region, have developed specialized soaring techniques to cope with the diverse and often challenging topographical and meteorological conditions prevalent in East Africa. The study notes that the effective use of slope lift not only conserves energy but also enables these birds to maintain a constant lookout for food sources while soaring [152]. In addition to Rüppell’s Griffon Vultures, other species such as the African White-backed Vulture and the Lappet-faced Vulture also demonstrate effective use of slope soaring in these regions. These species have shown remarkable adaptability in their flight strategies, optimizing their energy expenditure and enhancing their foraging efficiency in response to the varied ecological niches they inhabit [152].
The research further underscores the ecological significance of slope soaring in these birds’ lifestyles, highlighting how this flight technique is intricately linked to their survival and reproductive strategies. The adaptability and efficiency of slope soaring in these East African birds provide key insights into the broader ecological dynamics of avian species in diverse habitats.
Research by Santo et al. on species like black kites demonstrated their adaptability to varying environmental conditions for energy-efficient travel. Understanding these diverse soaring strategies provides valuable insights into avian migration, flight ecology, and conservation. Santos et al. showed that black kites effectively use orographic uplifts along slopes for faster progression during migration, showcasing their ability to exploit different atmospheric conditions. Additionally, the research highlighted how black kites adjust their flight strategies in response to changing weather patterns and landscape features. They exhibit flexibility in using both thermal and slope soaring to optimize their flight efficiency. This adaptability is crucial for their survival, especially during long migratory journeys where energy conservation is key. It was also discussed that black kites, along with other raptors, have developed a keen sense of exploiting wind and thermal currents, which is indicative of their complex cognitive abilities and deep understanding of their environment. This ability not only aids in their migratory and foraging activities but also has implications for their ecological impact and the ecosystems they inhabit [162].
Stokes and Lucas discussed how slope soaring facilitates energy-efficient travel and migration. Its adaptability to different environmental conditions and its impact on migration and ecological strategies are critical in the natural world [166]. The ability of birds and other animals to leverage natural wind patterns and atmospheric conditions for slope soaring illustrates a remarkable adaptation for energy conservation and efficient movement. Their study presented a theoretical framework to estimate the energy savings associated with wave-slope soaring compared to steady, level flight both in and out of ground effect. The analysis demonstrated that brown pelicans could reduce their cost of transport by approximately 60-70% typical ocean conditions. This significant reduction in energy demands highlights the efficiency of wave-slope soaring as a flight strategy in calm wind conditions. Further discussion regarding slope soaring also underscored its significance in the context of climate change and habitat alterations, suggesting that incorporating mechanisms to exploit similar environmental energy sources in UAVs could enhance flight endurance. The theoretical model and empirical evidence provided offer insights into the mechanics of slope soaring and its advantages for energy-efficient travel [166].
4.2. Solar Energy Harvesting
Drone Hubs for Solar Charging
In recent years the drone industry has greatly expanded both in production and application, leading to the implementation of drone design for over 200 applications [178]. Drones have become a widely used modern technology and the ability to collect energy from available sources is critical. Considering the nature of the work conducted by many of these drones, it becomes apparent a system to house drones in a variety of locations is necessary due to the energy consumption experienced as they travel to various operational destinations. By providing drone hubs, the operational duration of UAVs could be greatly increased as they will be stationed much closer to the intended area of work, minimizing unnecessary travel distance [332].
Increasing the system’s overall operational endurance requires overcoming constant energy consumption during missions, as well as maximizing the capability to easily and efficiently recharge and refuel drone systems. Not only would this hub provide a mass transportation and housing method for drones not in use, but it would also provide the capabilities of a mobile recharging station. As drones return from their directives, the energy systems used will be depleted. Therefore, this platform could be utilized as a recharging and refueling base employing several systems to do so, such as solar collection, harvesting wave energy, or via the collection of hydrogen through electrolysis. This would allow for the use of drones powered by both electricity and hydrogen, providing the basis for multiple drone types designed for several applications [332].
Historically, marine observations have predominantly relied on stationary sensors mounted on fixed platforms and buoys, employing a range of devices and concepts to host various common sensors for thorough marine observations [374,375,376,377]. The primary considerations for these long-term observations have been cost efficiency, durability, and reliability. Over the years, innovative methods for marine observation and data collection have emerged, including water, aerial, and space-based systems. Techniques such as using buoys and drifters for surface-level oceanic data collection have been notable, yet these methods are somewhat limited in their operational scope [378]. The constrained functionality of these observation platforms makes them less than ideal for comprehensive, sustained marine environment monitoring.
Figure 37.
Views of a floating drone hub concept to collect solar power and serve as a safe haven for drones during adverse weather. a & b) Views of the top of a floating drone hub, providing a landing location for aerial vehicles, along with charging [332]. c) A view of the underside of a drone hub capable of housing aquatic drones [332]. d) concept of area coverage of floating drone hubs for ecosystem monitoring [332]. e) View of bio-inspiration for a floating drone hub based on Amazonian water lilies, which provide platforms for birds, frogs, and other amphibious creatures [332].
Figure 37.
Views of a floating drone hub concept to collect solar power and serve as a safe haven for drones during adverse weather. a & b) Views of the top of a floating drone hub, providing a landing location for aerial vehicles, along with charging [332]. c) A view of the underside of a drone hub capable of housing aquatic drones [332]. d) concept of area coverage of floating drone hubs for ecosystem monitoring [332]. e) View of bio-inspiration for a floating drone hub based on Amazonian water lilies, which provide platforms for birds, frogs, and other amphibious creatures [332].
The proposition of integrating drone docking systems introduces a revolutionary platform for sensor deployment, facilitating the collection of widespread oceanic data over extended periods and across various locations. These advanced docking systems, envisioned to either replace traditional buoys or complement existing data collection setups using moorings, are set to substantially enhance the capacity for data collection. This would be achieved through the utilization of autonomous drones for continuous, minimal human intervention, and environmentally friendlier data gathering [332]. Such systems aim to simplify and improve the consistency of monitoring marine life, leveraging wire-guided profiling vehicles for the acquisition of physical, chemical, and biological data beneath the water surface with significant endurance [379,380].
Solar Powered UAVs
Shashati et al. discussed the development of a UAV enhanced with solar power to increase its flight range and endurance. The focus of the study was to integrate ultra-lightweight solar panels as a secondary power source to extend the UAV’s operating time and capabilities across various applications. The research demonstrated that incorporating solar panels on a fixed-wing UAV can significantly improve its performance by enabling longer flight durations than those possible with battery power alone. The design process involved using 3D modeling software for conceptual design and structural testing, the selection of suitable airfoils through aerodynamic analysis using XFLR5, and the implementation of a solar energy harvesting system optimized with a Maximum Power Point Tracker. SunPower flexible solar cells were selected for their high efficiency and flexibility, which are crucial for minimizing drag and fitting the airfoil shape. The solar panels were arranged in series and parallel to achieve the required voltage and current, ensuring the motor had a sufficient power supply while minimizing the weight of the necessary battery. The NACA 4412 airfoil was chosen for the main wing due to its favorable lift-to-drag ratio and ease of fabrication. The UAV’s design aimed at balancing the trade-off between performance enhancement and weight increase from the solar panels. Preliminary tests compared the endurance of the system with and without solar panels, demonstrating the potential for significant improvements in flight time due to the additional solar power [381].
A similar project delved into the development and evaluation of a quadcopter powered by both solar energy and a lithium-polymer battery. This hybrid power system aims to maximize flight duration by utilizing solar panels for direct energy capture during daylight and storing excess energy for use when solar power is unavailable. Nileshkumar et al. integrated solar panels with a quadcopter, focusing on energy management and flight time optimization. The solar panels were used to collect energy during the day for immediate use and to charge the onboard battery for night-time operation. The research included practical analysis through real flight data collection, showcasing the impact of solar assistance on flight modes and energy consumption. The data illustrates the quadcopter’s ability to operate efficiently with solar energy, resulting in increased flight time and reduced battery discharge rates. The project addresses several challenges associated with long-duration drone flights, such as maintaining sufficient current flow for lift-off and minimizing energy consumption to match the power provided by solar panels. A significant achievement of the study is the design of a solar power management system circuit, which achieves an 80% charging efficiency ratio. This innovation enables the drone to gain 10-15 minutes of extra flight endurance during peak sunlight hours by effectively utilizing solar-generated power. By achieving more than 10 minutes of increased flight time with solar cell use, the study provides a promising outlook for the application of renewable energy sources in unmanned aerial vehicles. The findings suggest that with appropriate solar technology integration and energy management, drones can achieve longer flight times, expanding their usability across various applications while adhering to sustainability principles [382].
Buzdugan et al. provided an extensive exploration into optimizing solar-powered UAVs through numerical modeling. The study centers on a UAV with a 151 cm wingspan, powered by a combination of photovoltaic cells, a battery pack, and a fuel cell. Through numerical modeling, the researchers aimed to identify the optimal energy source configuration for different flight stages, ensuring a steady and efficient power supply throughout the mission. The model incorporated simulations of UAV performance under various conditions using MATLAB/SIMULINK, taking into account the technical specifications developed in Buzdugan’s doctoral thesis. The findings reveal that an integrated approach, combining photovoltaic cells with a battery and a fuel cell, enables the UAV to maintain power for extended periods and across different times of the day. The research proposes an energy management system equipped with a maximum power point tracking function to dynamically select the optimal energy source based on flight conditions and propulsion system requirements. This system allows for mixed or individual mode power supply from the three sources, ensuring uninterrupted and efficient power delivery. The study concludes that for UAVs to achieve high-altitude and long-distance missions, it is necessary to utilize multiple energy sources and an effective energy management system. This approach not only maximizes the efficiency of solar power but also enhances the overall performance of UAVs in various operational scenarios [383].
Figure 38.
a) First prototype of solar glider by Shashati et al., highlighting the solar cells conformed to the wing surface [381]. b) [383]. c) View of NASA’s long endurance flying wing design history [384]. d) View of the Helios prototype (HP01) in flight [384]. e) Isometric view of the Newsolar concept designed to maximize solar area [385]. All figures are reprinted with permission; please see the Appendix A.
Figure 38.
a) First prototype of solar glider by Shashati et al., highlighting the solar cells conformed to the wing surface [381]. b) [383]. c) View of NASA’s long endurance flying wing design history [384]. d) View of the Helios prototype (HP01) in flight [384]. e) Isometric view of the Newsolar concept designed to maximize solar area [385]. All figures are reprinted with permission; please see the Appendix A.
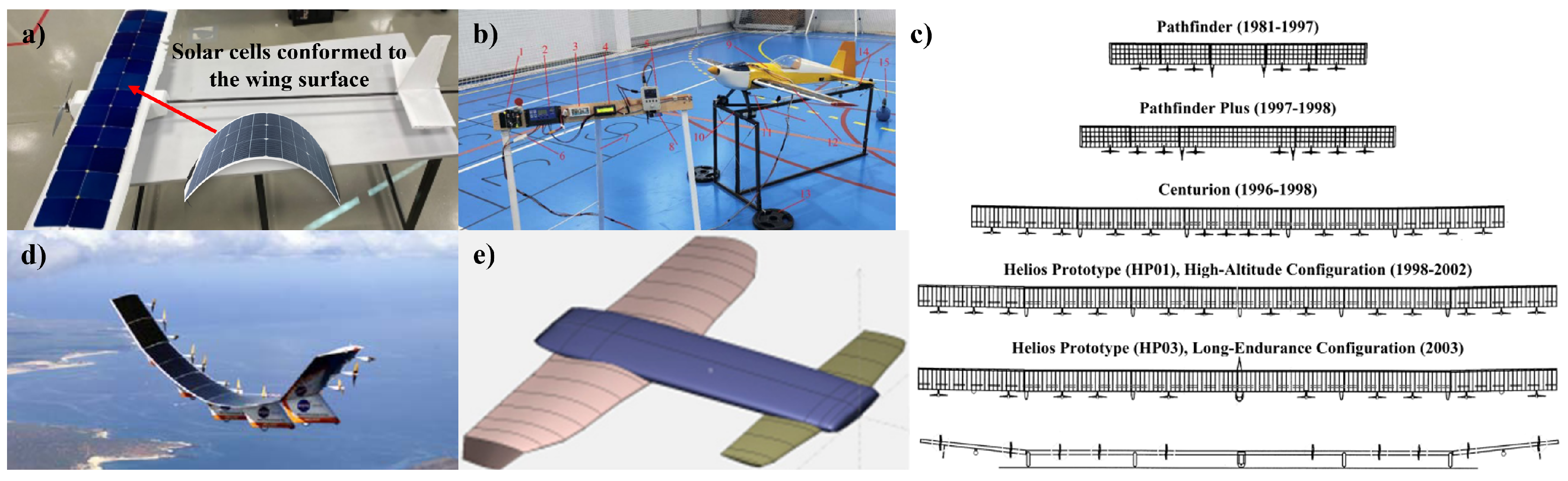
Chen classified drones based on operational altitude, which includes low-altitude drones, high-altitude drones, and near-space drones. The study emphasizes the pivotal role of solar drones in a variety of applications, from mapping and surveillance to natural disaster early warning systems, highlighting the advantages of solar energy systems in extending the range and endurance of UAVs. His analysis covered the evolution of solar UAVs, including significant milestones like the development of the Mozi II aircraft, which boasts a wingspan of 15 m and can operate for 12 hours under ideal conditions. Chen also discussed the challenges related to solar cell conversion efficiency, battery performance, and aircraft skin adaptation for solar integration. Highlighting advancements like GaAs thin-film solar cells, the study underscores the technological strides in enhancing UAV performance through solar power. For low-altitude UAVs, the work identified thin-film monocrystalline silicon solar cells as a preferred choice due to their balance of efficiency, flexibility, and resistance to environmental factors. For high-altitude long-endurance UAVs, Chen talked about the importance of optimizing solar photovoltaic cells’ conversion efficiency and the need for batteries that offer high energy density and stability under varying conditions. The study suggests that advancements in solar cell technology and energy storage are crucial for maximizing the operational capabilities of solar UAVs [386].
Dinca et al. explored the trade-offs between aerodynamic efficiency and electrical power in the design of solar-powered UAVs. The authors proposed a new solar UAV configuration that sacrificed high aerodynamic efficiency for a higher surface area available for solar cells, thereby increasing the amount of energy available on board. The research focused on the importance of optimizing solar UAV designs for specific mission requirements. The conventional approach has favored high-aspect-ratio wings to maximize aerodynamic efficiency. However, this design choice limits the surface area available for mounting solar cells, thereby constraining the UAV’s energy harvesting capability. The proposed "Newsolar" configuration aims to balance these considerations by offering a design that may have lower aerodynamic efficiency but significantly increases the potential for solar energy harvesting. The Newsolar configuration, with its enhanced solar power capacity, opens up new possibilities for UAV missions that do not require continuous flight during nighttime. This configuration is particularly suited for missions with heavy payloads and high energy consumption, where the extended daylight flight time would be highly beneficial. The payload capacity of this configuration was determined to be 2.5 times greater than a traditional glider configuration. The authors suggest that further detailed studies and a design phase are necessary to refine this UAV configuration before it is ready for practical deployment [385].
Another example of solar harvesting for UAVs was produced by NASA during the Pathfinder, Centurion, and Helios Prototype projects as shown in Figure 38. Noll et al. provided a comprehensive analysis of the Helios Prototype vehicle’s operational challenges and mishap. The Helios Prototype, a significant project under NASA’s Environmental Research Aircraft and Sensor Technology program, was developed to explore the potential of solar-powered flight for high-altitude, long-endurance missions. The vehicle featured two configurations: the HP01 aimed at high-altitude operations using solar cells and batteries, and the HP03 designed for long-duration flights leveraging both solar power and a hydrogen-air fuel cell system for night-time operation. The work detailed the events leading to the HP03 configuration’s failure, where the vehicle encountered unexpected aerodynamic instability, leading to structural failure and the subsequent loss of the aircraft. This incident underscored the critical importance of addressing the complex interplay between aerodynamic design, environmental conditions, and vehicle control systems for such innovative UAVs. The analysis revealed that a combination of factors, including the vehicle’s response to atmospheric turbulence and its structural flexibility, contributed to the mishap. Specifically, the vehicle entered a persistent high dihedral state that it could not recover from, ultimately leading to its destruction. The authors discussed the necessity for advanced, multidisciplinary analysis methods appropriate for vehicles with significant morphing capabilities and high flexibility, and the importance of thorough ground-test procedures to validate these new analysis methods. The document stresses the need for real-time monitoring systems capable of detecting and responding to critical flight parameters, suggesting enhancements in the design and operational protocols to improve the safety and reliability of future high-altitude, long-endurance solar UAV missions [384].
4.3. Ionized Winds and Energy Harvesting
The field of energy harvesting has surged in popularity, driven by the abundant energy sources in the environment that can be transformed into usable forms in parallel with the need to find alternative green energy sources. While many harvesting methods capitalize on mechanical energy, the potential of chemical and electrical energy present in nature cannot be overlooked, with numerous strategies available for their extraction, conversion, and utilization [333,387,388]. The energy thus obtained finds applications across a broad spectrum, from powering miniature devices like pacemakers and wearable technology to energizing larger machinery including drones. A variety of natural phenomena, including water flows, wind movements, mechanical vibrations, temperature differentials, and more, have been harnessed to convert diverse forms of energy into electrical power that is practical for use [389].
The exploration of capturing energy from storm phenomena, particularly lightning, has been a subject of scientific inquiry for over forty years, with ongoing research efforts today seeking to advance our understanding and capabilities in this area [390,391,392]. To grasp the concept of lightning energy harvesting, it’s necessary to delve into the fundamental mechanics of a lightning strike, which originates from electrostatic charges accumulating within cloud formations shown in Figure 39c & d [393]. This process begins when water vapor rises from large bodies of water and is transported by atmospheric wind patterns. The friction generated as water molecules and air particles interact results in the accumulation of electrostatic charges within the clouds [394].
Cloud dynamics play a crucial role in this process, where turbulent winds carry water droplets upwards to altitudes between 35,000 and 70,000 feet, well beyond the freezing point, thus forming ice particles [395]. Concurrently, descending currents carry these frozen particles downwards, facilitating collisions between updrafts and downdrafts. This interaction dislodges electrons from ascending particles, depositing them onto descending particles, creating a cloud with a negatively charged base and a positively charged top, separated by the atmosphere’s insulating properties. When the electrostatic field’s strength surpasses the atmosphere’s insulation capacity, it breaks down, leading to a rapid discharge of electricity aimed at neutralizing the charge difference across the cloud layers.
Enhancing the detail on the mechanism of charge separation within clouds could provide further insight into potential energy harvesting techniques. For instance, understanding the specific conditions that lead to the optimal charge buildup and the precise nature of the interactions between particles within the cloud could inform more efficient designs for lightning energy harvesting systems. Moreover, integrating recent advancements in materials science and electrical engineering could offer innovative solutions for capturing and storing the immense energy released during a lightning strike, potentially transforming how we think about renewable energy sources. This approach not only deepens our comprehension of a natural phenomenon but also paves the way for groundbreaking applications in energy technology.
The accumulation of charges within clouds is the primary driver of lightning, which manifests in three distinct forms: intra-cloud, inter-cloud, and cloud-to-ground lightning, each characterized by unique interactions between charged regions [399,400]. Inter-cloud and intra-cloud lightning share similar mechanisms, involving the exchange of charges between different parts of the same cloud or between two separate clouds, respectively. Cloud-to-ground lightning, however, holds more significance for public safety due to its potential for striking the ground and posing a risk to life and property. This type of lightning occurs when the negatively charged bottom of a cloud attracts positive charges from the ground. These ground charges ascend towards the cloud, moving up along elevated objects in their vicinity, such as trees and buildings. As the atmospheric insulation breaks down, negative charges from the cloud extend toward the ground in the form of branching tendrils, seeking a pathway to discharge. When these tendrils approach the ground closely, a "positive streamer" from the ground rises to meet them, completing the circuit and resulting in a lightning strike. The resistances within the cloud () and between the cloud and the ground () are crucial factors in this process. Typically, is lower than , given the denser medium within the cloud and the relatively greater distance between the cloud and the ground. This difference in resistance plays a pivotal role in the development and eventual discharge of lightning [398].
Enhancing this description with additional insights, such as the statistical likelihood of each type of lightning or the specific conditions that favor the formation of one type over the others, could offer a more comprehensive understanding of these phenomena. Moreover, exploring the implications of these electrical interactions for energy harvesting technologies, especially in the context of cloud-to-ground lightning, could reveal potential avenues for harnessing this natural power source more effectively. Further research into the material properties that influence and , as well as advancements in atmospheric and electrical engineering, could improve our ability to predict, and perhaps even control, the occurrence of lightning for energy production and safety purposes.
Capturing the immense energy released by a typical lightning stroke, which is around 1 Gigajoule—equivalent to about 30% of the average monthly energy consumption of a US household—presents a significant challenge with the current state of technology, leading to the absence of a matured method for its effective harvesting. Research in this domain explores innovative approaches, such as transforming water into hydrogen for use as a chemical fuel source [401], or harnessing the electrical energy from lightning to trigger the evaporation of metals. Another avenue of investigation aims to leverage the substantial electrical current generated by a lightning stroke to electrolyze water molecules, producing hydrogen and oxygen which, when recombined, yield an exothermic reaction [402].
Three principal strategies have emerged for the extraction of electrostatic energy from the atmosphere, each characterized by distinct mechanisms and potential applications. These include capturing the charge prior to a lightning strike, post-lightning strike energy capture, and the generation of ions through electro-aerodynamics for propulsion purposes. Pre-lightning strategies focus on converting static charge into direct current, taking care to avoid inadvertently triggering a lightning strike. Post-lightning approaches explore the use of capacitors designed to capture corona discharge, the residual electrical activity following a lightning event. The electro-aerodynamic method, on the other hand, employs an electric field to accelerate ions, which could potentially facilitate aircraft propulsion, a concept further elaborated in discussions on electro-aerodynamic propulsion [403].
Enhancing this overview with a deeper dive into the feasibility, efficiency, and safety concerns associated with each method could provide a clearer picture of the current challenges and future possibilities in the field of atmospheric energy harvesting. For example, examining the materials and technologies needed to implement these strategies, as well as their environmental impact and practical applications, could offer valuable insights. This could pave the way for breakthroughs in our ability to harness the vast power of lightning and other atmospheric energy sources, contributing to the development of sustainable and renewable energy solutions.
Electro-Aerodynamic Propulsion
Typically, the propulsion of drones relies on the process of combustion, batteries, electric motors, and propellers, to generate required thrust. The new electro-aerodynamic approach that recently has been proposed by MIT researchers uses ionic wind to generate thrust as seen in Figure 39 b [403]. Xu et al. introduced a novel solid-state propulsion system based on electro-aerodynamics, a method that propels planes using electrical forces to accelerate ions in a fluid, thereby generating thrust without moving parts, nearly silently, and without combustion emissions. This study marked a significant milestone by demonstrating the first successful flight of a heavier-than-air aircraft powered by such a system. The research team designed and constructed a fixed-wing aircraft with a five-meter wingspan, incorporating an ultralight high-voltage power converter to generate the necessary electric fields for ion acceleration. The propulsion system, operating at a high voltage of 40 kilovolts, uses a corona discharge method to produce ions. These ions, accelerated by an electric field, collide with neutral air molecules, transferring momentum and creating an ’ionic wind’ that results in forward thrust. The aircraft’s performance was assessed through ten flights, highlighting the feasibility of steady-level flight and providing a proof of concept for electro-aerodynamic propulsion in aviation [403].
The study successfully demonstrated that the solid-state electroaerodynamic propulsion system could sustain powered flight, overcoming previously believed limitations regarding thrust-to-power ratio and thrust density. Experimental results showed that the aircraft achieved steady-level flight over distances of 40-45 meters, with an average height gain, proving that electro-aerodynamic propulsion can generate sufficient thrust for flight without the drawbacks of conventional propulsion systems. The achieved thrust-to-power ratio was comparable to that of conventional propulsion methods, such as jet engines, but at a significantly lower overall efficiency, which was expected given the experimental nature and the prototype’s design constraints. This research opens up new avenues for developing aircraft and aerodynamic devices that are quieter, mechanically simpler, and emission-free. The solid-state nature of electro-aerodynamic propulsion offers potential for miniaturization and integration into a variety of aircraft designs, including urban drones, high-altitude environmental monitoring planes, and potentially commercial aviation, although significant advancements are needed to increase efficiency and thrust density to competitive levels [403].
The innovative design of an ionized plane by MIT researchers represents a groundbreaking advancement in aviation technology, marking the introduction of the first aircraft capable of flight without any moving parts [396,403,404]. This aircraft, a light, drone-like vehicle, harnesses the power of ionized wind for propulsion by installing high-voltage electrodes beneath the wings, generating a strong electric field. The leading-edge electrodes, charged to 40 kV, attract a high positive charge, while a thicker wire along the wing’s trailing edge acts as the negative counterpart. When activated, these electrodes create a magnetic-like effect that strips electrons from nearby air molecules, leaving behind positively charged ions. These ions are then drawn towards the negatively charged electrodes at the wing’s rear. As they move, they push neutral air molecules in the same direction, creating a flow of air or ’ion wind.’ This flow generates relative air velocity across the wing, producing the necessary thrust for propulsion. This mechanism allows the drone to move forward, demonstrating a novel method of flight that eliminates the need for conventional moving parts, such as propellers or turbines.
Over the last decade, the advancement of drone-swarming technology has significantly accelerated, driven by a quest to improve mission success rates and speed up mission performance. The reliance on a single drone for missions is fraught with risks due to the unpredictable nature of challenges that drones often encounter. By deploying swarms of drones, these risks are substantially reduced, enhancing efficiency and ensuring that the failure of one drone doesn’t affect the mission’s outcome. Swarm intelligence, which is the collective, coordinated, flexible, self-organized, and robust behavior of a group following certain rules, plays a pivotal role in this context.
Additionally, formation flight, particularly in the form of V-shaped swarm reformation, is explored as a means to address various challenges in drone design and application, especially in terms of flight efficiency. For instance, Mirzaeinia et al. [405] discuss the concept of V-shape swarm reformation to minimize flight drag and conserve energy for drones positioned strategically within the formation. This rearrangement helps reduce overall energy consumption during flight, thereby enhancing energy efficiency and extending the operational range.
Herkenhoff et al. expanded on this idea by proposing swarm reformation to improve the phase of cloud charge harvesting during flight. As depicted in Figure 39e, a drone swarm can adjust its formation based on proximity to cloud charges, with Figure 39e (left) showing a swarm maintaining the greatest distance from both top and bottom cloud charges. In contrast, Figure 39e (middle) and Figure 39e (right) illustrate how a swarm can reorganize itself into a chain formation to get closer to the top and bottom charges. This strategy is designed to enhance the ionic propulsion method by positioning drones with lower energy levels closer to negatively charged clouds and those with more energy further away. Cycling drones through different positions within the swarm, much like the energy-saving benefits seen in V-formation flight, is hypothesized to improve overall swarm efficiency and endurance [406].
Inspired by the hydrophobic and self-cleaning properties observed in plant leaves and insect wings, Herkenhoff et al. investigated the application of hair-like structures on drone wings to harvest static charge from the atmosphere. These microscopic and nano-structured surfaces exhibit capabilities such as superhydrophobicity and reducing wettability, enabling a "self-cleaning" mechanism. Insects, through their wing topographical structuring, manage not only to repel water but also to potentially harvest electrostatic energy from the atmosphere due to these unique surface properties. The experimental approach involved using a high-voltage power supply and a parallel plate capacitor to simulate the effects of electrostatic fields on various test objects, mimicking the hair-like structures on insect wings. Different geometrical shapes and materials were placed between the parallel plates to observe the effects of electric field strengths on these structures. The research aimed to understand how these hair-like structures could harvest static charge and improve aerodynamic efficiency by potentially reducing drag and increasing lift, drawing parallels to the way shark scales operate to minimize friction drag. The study found that implementing hair-like structures on drone wings could enhance aerodynamic performance passively while also aiding in the reception of charged particles from the atmosphere. It was hypothesized that when an electrostatic charge is applied, these hairs would stand normal to the wing surface, thereby increasing the tendency to harvest static charges. The experiments demonstrated the movement of electroreceptive hairs might produce a beneficial aerodynamic effect, suggesting a potential for static charge harvesting in improving the flight performance of micro-UAVs [406].
Gavrilović et al. conducted a series of flight simulations and outdoor flight tests, focusing on the exploitation of atmospheric turbulence and wind gradients. The study examines the effect of thrust in vertical gusts, the potential benefits of MAVs in atmospheric energy harvesting, and the development of a bioinspired sensory system for wind field estimation. The experimental setup included a 1.2 m flying wing equipped with an integrated inertial measurement unit, differential pressure sensors, a magnetic encoder, and a GPS for precise navigation and control. The study found significant improvements in MAV performance through active control of command surfaces in energy-harvesting modes compared to auto-stabilization or fixed-stick flight. The analysis highlighted the importance of wind profiles with higher turbulence intensity for energy transfer to the aircraft. Additionally, the paper introduces a biologically inspired sensory system that estimates the wind field, enabling the exploitation of atmospheric energy through tailored equipment and control algorithms. Initial flight tests aimed to determine the average power consumption of the motor for altitude hold tasks and to evaluate the performance of sensors in various atmospheric conditions. Subsequent flights focused on the autonomous exploitation of atmospheric phenomena, such as thermals and wind gradients, demonstrating the potential for substantial reductions in motor power consumption and significant altitude gains through energy harvesting [407].
The work conducted by Kiziroglou et al. presented a comprehensive investigation into the potential of harvesting static charge on aircraft for powering monitoring sensors. This exploration is motivated by the ongoing pursuit of cost-effective and maintenance-efficient solutions in aircraft operation, where wireless sensing technologies offer substantial advantages but are often hindered by the practical challenges of battery replacement and cabling complexities and weight. The researchers evaluated several innovative methods for energy harvesting, including the utilization of materials with varying air-flow charging rates, the strategic discharge of the fuselage’s accumulated static charge to neutral bodies, and the induction of charge movement by the fuselage’s electrostatic field. Additionally, they explored the inductive capture of energy from corona discharges at static discharge wicks, a phenomenon associated with the fuselage’s interaction with the air. Kiziroglou et al. demonstrated direct charging of a storage capacitor from static electricity, achieving a significant voltage of 25V on a 25nF capacitor. This accomplishment underscores the feasibility of integrating static charge harvesting mechanisms into aircraft systems to supply power to sensors, potentially leading to the realization of energy-autonomous sensor networks onboard. The paper also discussed the installation considerations and potential advantages of each harvesting method, contributing valuable insights into the practical application of these technologies in aviation contexts. Furthermore, the study delved into the mechanics of static charge generation and collection, offering detailed analyses of various charge transfer mechanisms between insulating surfaces under relative motion. It proposes innovative approaches for maximizing the efficiency of static charge collection, such as embedding electrodes within insulating materials and employing moving electrodes to scan charged areas. The exploration of fuselage static charge utilization, particularly through field-effect induction and the exploitation of corona discharges, presents intriguing possibilities for enhancing the energy independence of aircraft sensor systems [408].
Xie et al. assessed the practicality of harvesting electrostatic energy generated on carbon fiber composite (CFC) aircraft during flight. This research is pivotal as it shifts the paradigm from merely dissipating static charges, which are viewed as a potential hazard, to utilizing these charges as a viable energy source. Through a series of laboratory experiments, the research team was able to demonstrate that it is feasible to collect static electricity generated in flight using capacitive collectors. These experiments were designed to mimic the generation and collection of static charges under various configurations, providing a comprehensive analysis of the potential for electrostatic energy harvesting on aircraft. The results of the study indicated that specific configurations of capacitive collectors could efficiently harvest static charges. The experiments explored both grounded and ungrounded collector setups, offering valuable insights into the mechanisms and efficiency of static electricity collection on CFC aircraft. This exploration is significant because it lays the groundwork for developing new energy harvesting systems that could potentially enhance the energy efficiency of aircraft operations.
In the study by Mohanty et al. they also developed a novel concept for harvesting electrostatic energy on flying composite aircraft. The research focuses on the potential of collecting static charges through capacitive collectors, reviewing existing energy harvesting systems, and the electrification of CFC aircraft. The team designed detailed model experiments to assess different configurations for electrostatic energy harvesting on airplanes, utilizing a corona discharging device to generate static charge and testing the collection of these charges under various setups. The research introduced a method for treating local electrostatic energy on aircraft by utilizing capacitive collectors to gather static charges. They experimented with collecting these charges using either a capacitor or a metal sheet positioned within the electric field created by the corona discharging device. Four distinct configurations were tested to evaluate the effectiveness of static charge collection and demonstrated the feasibility of collecting local static electricity generated during flight. The study found that modifying certain portions of an aircraft’s body to act as capacitors can successfully store the static charge generated from air friction during onboard navigation. These findings suggest that the static charge, typically seen as a nuisance due to its potential to interfere with aircraft communication equipment, can instead be harnessed as a valuable energy source.
5. Concluding Remarks
In the realm of bio-inspired aviation technology, the integration of natural mechanisms into aircraft and UAV design has shown significant promise. However, the body of research in this field often yields disparate results, reflecting the complexity and multifaceted nature of biological systems and their adaptation to engineered applications. As an example, the study of drag reduction techniques inspired by avian and insect physiology has led to varying conclusions. For instance, the riblet structures found on shark skin and bird feathers have been extensively researched for their potential to reduce skin friction drag. While Bechert et al. [14] reported significant drag reduction when applying these riblet structures to aircraft surfaces, subsequent studies by Feng et al. [18] found a different impact on drag reduction under certain flight conditions. These disparities can be attributed to differences in experimental conditions such as turbulence intensity and flow uniformity, as well as scale and material differences. The microstructural precision required to replicate biological riblets might not be achieved uniformly in all studies, leading to inconsistent results.
The disparities in findings across studies on bio-inspired aviation technologies highlight the complexities of translating natural mechanisms into engineered systems. These differences underscore the importance of considering environmental context, scale, design specifics, and flight regimes when evaluating the applicability of biological adaptations. Further research, with a focus on standardizing experimental conditions and exploring scalable solutions, is essential to bridge the gap between biological inspiration and practical engineering applications.
Induced drag, or vortex drag, is a critical factor in the aerodynamic efficiency of aircraft. It arises from the generation of lift and manifests as trailing vortices that form as air flows from the high-pressure region beneath the wing to the low-pressure region above it, creating spiraling air patterns. The minimization of induced drag, particularly in three-dimensional flows, is essential for enhancing the performance, range, and endurance of both fixed-wing and flapping-wing drones, as well as other types of aircraft. As previously discussed, there is a significant understanding of the minimization of vortex drag in fixed-wing systems, however, there are many aspects we can still glean from nature for both fixed and flapping-wing systems.
In fixed-wing aircraft, induced drag is one of the primary components of total drag at lower speeds and higher angles of attack. Traditional methods of minimizing induced drag involve optimizing wing design, particularly through elliptic spanwise lift distribution, which has been well-documented in conventional aviation [187,188,194]. The work of Ludwig Prandtl laid the foundation for understanding how to achieve minimal induced drag by balancing aerodynamic forces with structural constraints. His theories emphasize the need for longer wingspans, which reduce induced drag by spreading out the vortices generated at the wing tips over a larger area, thereby reducing their intensity.
For drones, especially those designed for long-range or endurance missions, minimizing induced drag is crucial. Fixed-wing drones often operate at lower Reynolds numbers compared to larger manned aircraft, making them more susceptible to the negative effects of induced drag. By minimizing this drag, these drones can achieve greater flight efficiency, allowing for extended missions, reduced energy consumption, and improved overall performance [182,184]. In military and surveillance applications, where operational efficiency and endurance are critical, minimizing induced drag directly contributes to the drone’s ability to perform extended missions without the need for frequent refueling or battery recharging.
Flapping-wing aircraft, inspired by the flight mechanisms of birds and insects, present a unique challenge in the minimization of induced drag due to their unsteady and complex flow dynamics. Unlike fixed-wing aircraft, where induced drag can be addressed significantly through wing shape optimization, flapping-wing vehicles must manage the vortices generated by the periodic motion of the wings and be optimized for multiple modes of flight. These vortices are essential for thrust and lift generation, particularly in hovering or slow-speed flight, but they also contribute heavily to induced drag. Minimizing this induced drag in flapping-wing drones involves a delicate balance between maintaining the necessary vortex structures for lift and thrust and reducing the energy lost to these vortices. Advances in 3D CFD have enabled more sophisticated modeling of these 3-D unsteady flows, providing insights into how wing flexibility, wing kinematics, and other design factors can be optimized to reduce induced drag while maintaining or enhancing aerodynamic performance [15,125,195,196,197,198,199,200]. It is our belief that several of the techniques observed by natural flyers may translate to both fixed and flapping wing systems, providing better efficiency and adaptability for future systems.
The compilation of bio-inspired techniques for enhancing flight efficiency in aviation, as outlined in the summary Table 6, underscores the ingenuity and resourcefulness nature offers. These strategies, distilled into applicable technologies, span across various aspects of aircraft design and operation, including wing morphology, surface textures, flight patterns, and energy management.
Birds and insects, through millions of years of evolution, have honed mechanisms that reduce drag, optimize lift, and harvest energy from their environment. The summarized techniques, from morphing wing technologies to ionized wind energy harvesting, present an intricate tapestry of potential applications in modern aviation. The ingenuity encapsulated in features such as the alula-inspired leading-edge devices for increased lift, or the corrugation patterns of dragonfly wings that inform robust airfoil designs, illustrates a wealth of design cues that can be adapted for improved aerodynamic efficiency.
A pivotal benefit of bio-inspiration in aviation is the holistic enhancement it offers. These strategies do not exist in isolation in their natural contexts and, likewise, present the opportunity for combined application in engineering designs. For instance, the integration of flexible flaps and feather riblet structures can concurrently reduce drag and modulate airflow over the wings for diverse flight conditions. Similarly, the combination of wing porosity concepts and charge harvesting techniques could lead to multifunctional structures that not only manage airflow more effectively but also contribute to the aircraft’s power needs.
Moreover, the deployment of formation flight principles, alongside real-time wing shape optimization, can revolutionize energy consumption patterns for fleets of UAVs. By synchronizing these strategies, swarms of UAVs, MAVs or other aircraft can achieve reduced drag through aerodynamic positioning while actively adapting to variable flight conditions, optimizing energy use across the formation.
In embracing these bio-inspired methodologies, it is crucial to adopt a multidisciplinary approach, bridging aerodynamics, material science, and control systems to explore the synergistic effects of combined techniques. The potential intersection of these methods promises to unlock new levels of efficiency, where the sum of integrated bio-inspired strategies is greater than its parts. The future of aviation lies not only in the adoption of individual bio-inspired innovations but in the architectural integration of these methods into cohesive, intelligent systems. This integration aims to synergize shape, surface, and flight path adaptations to create aircraft that are not only efficient but are responsive to the dynamic nature of their operational environment. The horizon of bio-inspired aviation is vast and promising, poised to usher in an era of sustainable, efficient, and intelligent aircraft design.
Minimizing induced drag is a complex but essential aspect of modern aircraft design, particularly for fixed-wing and flapping-wing drones. While the principles of induced drag are well-understood in the context of conventional fixed-wing aircraft, the unique challenges posed by flapping-wing systems require future research and innovation. As drones and other aircraft continue to evolve, driven by the need for greater efficiency, range, and performance, the importance of optimizing aerodynamic design will only grow. Through advances in computational modeling, material science, and bio-inspired design, the aerospace industry can continue to push the boundaries of what is possible, leading to more efficient and capable aircraft across all domains of flight.
Author Contributions
Conceptualization, M.H. and B.H.; methodology, B.H.; investigation, B.H.; resources, B.H.; writing—original draft preparation, B.H.; writing—review and editing, B.H. and M.H.; visualization, B.H.; supervision, M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
DURC Statement
Current research is limited to the biomimicry, which is beneficial for flight efficiency and does not pose a threat to public health or national security. Authors acknowledge the dual-use potential of the research involving biomimicry and confirm that all necessary precautions have been taken to prevent potential misuse. As an ethical responsibility, authors strictly adhere to relevant national and international laws about DURC. Authors advocate for responsible deployment, ethical considerations, regulatory compliance, and transparent reporting to mitigate misuse risks and foster beneficial outcomes.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A. Permissions
The following is the statement of copyright from the American Institute of Aeronautics and Astronautics (AIAA) that I signed during the submission of the above papers: "Copyright Form A: I assign copyright to my work to AIAA, giving Institute all rights to it except that I and the organization by which I was employed at the time I wrote the manuscript have the right of further reproductions, in part or in full, provided they are not for sale. AIAA does not claim any patent, trademark, or proprietary rights other than copyright."
, is reprinted with permission from Springer Nature: Journal of Bionic Engineering, License: 5771421339822.
, is reprinted with permission from Dr. Andrew Davis.
Figure 9(a), is reprinted with permission: https://creativecommons.org/licenses/by/4.0/Creative Commons.
Figures 9(b,d,e), are reprinted with permission from IOP Publishing for electronic distribution: Smart Materials and Structures, License: 1457966-2.
Figures 9(c,f) , are reprinted with permission from Springer Nature for electronic distribution: Journal of Bionic Engineering, License: 5772081488023.
Figures 9(g-i) reproduced from Elsevier Masson SAS for electronic distribution. All rights reserved.
Figures 11(b,c) reproduced from Elsevier Masson SAS. All rights reserved.
Figure 9(j) , is reprinted with permission from Annual Reviews: Annual Review of FluidMechanics, License: 1458398-8.
Figure 5(a) , is reprinted with permission: Creative Commons.
Figure 5(b,e) , are reprinted with permission: Creative Commons.
Figure 5(c,d,j) , are reprinted with permission from the American Institute of Aeronautics and Astronautics (AIAA): 46th AIAA Aerospace Sciences Meeting and Exhibit, License: 1458902-1.
Figure 5(f) , is reprinted with permission: Creative Commons.
Figure 5(g-h) , are reprinted with permission from the American Institute of Aeronautics and Astronautics (AIAA): 39th AIAA Fluid Dynamics Conference, License: 1458902-2.
Figure 5(h) , is reprinted with permission from the American Institute of Aeronautics and Astronautics (AIAA): 42nd AIAA Aerospace Sciences Meeting and Exhibit, License: 1458902-3.
Figure 6(a) , is reprinted with permission from The Company of Biologists Ltd.: The journal of experimental biology.
Figure 6(c) , is reprinted with permission: Creative Commons.
Figure 6(g) , is reprinted with permission: Creative Commons.
Figure 6(h) , is reprinted with permission from Elsevier Science and Technology Journals: Materials Today : Proceedings, License: 1458298-1.
Figure 6(i) , is reprinted with permission from The Royal Society (U.K.): Journal of the Royal Society interface, License: 1458961-2.
Figure 6(j) , is reprinted with permission from AIP Publishing: Physics of Fluids, License: 5743160979679.
Figure 6(k) , is reprinted with permission from The American Society of Mechanical Engineers (ASME): 2005 ASME International Mechanical Engineering Congress and Exposition (IMECE).
Figure 6(l) , is reprinted with permission from IOP Publishing, Ltd: Bioinspiration and Biomimetics, License: 1457965-1.
Figure 7(a-d) , are reprinted with permission from Oxford University Press: Ornithology, License: 5743230089954.
Figure 15(a) , is reprinted with permission from Tim Laman.
Figure 15(b) , is reprinted with permission from Jill Duncan and Ken Bissett.
Figure 15(c) , is reprinted with permission from Wolfe R.
Figure 15(d) , is reprinted with permission from Chris Venetz.
Figure 15(e) , is reprinted with permission from Yeray Seminario.
Figure 15(f) , is reprinted with permission from Pascal De Munck.
Figure 14(a-c) , is reprinted with permission from The Royal Society (U.K.): Journal of the Royal Society interface, License: 1458961-4.
Figure 14(d) , is reprinted with permission from Dr. Simon Baeckens.
Figure 14(e) , is reprinted with permission from Cambridge University Press.
Figure 11(a) , is reprinted with permission: Creative Commons.
Figure 11(b-c) reproduced from Elsevier Masson SAS for electronic distribution. All rights reserved.
Figure 11(d) , is reprinted with permission from Dr. Tung Wan.
Figure 11(e) , is reprinted with permission from Dr. Afzal Suleman.
Figure 11(f-h) , are reprinted with permission from the American Institute of Aeronautics and Astronautics (AIAA): Journal of aircraft, License: 1458902-4.
Figure 12(a-c,f) , are reprinted with permission from Springer Nature: The Science of Nature, License: 5744361043300.
Figure 12(d-e) , are reprinted with permission from Dr. Anne-Kristin Wohlbier.
Figure 12(g) , is reprinted with permission: Creative Commons.
Figure 12(h-m) , are reprinted with permission: Creative Commons.
Figure 12(n-o) , are reprinted with permission: Creative Common.
Figure 13(a-b) , are reprinted with permission from Springer Nature BV: Acta mechanica, License: 1457259-1.
Figure 13(c) , is reprinted with permission from Elsevier Science and Technology Journals: Computers and fluids, License: 1458298-2.
Figure 13(d-f) , are reprinted with permission from the American Institute of Aeronautics and Astronautics (AIAA): AIAA Aviation 2019 Forum, License: 1458902-5.
Figure 13(g-h) , are reprinted with permission: Creative Commons.
Figure 13(i) , is reprinted with permission: Creative Commons.
Figure 13(j) , is reprinted with permission from Helen von Holten.
Figure 20(a,e) and Figure 22(e-g) , are reprinted with permission from IOP Publishing, Ltd: Bioinspiration and Biomimetics, License: 1457966-3.
Figure 20(b-c,g) , are reprinted with permission from the Company of Biologists Ltd.: The journal of experimental biology.
Figure 20(d,f) , are reprinted with permission from Oxford University Press: Integrative and Comparative Biology, License: 5743250768643.
Figure 20(h) , is reprinted with permission from Elsevier Science and Technology Journals: Journal of theoretical biology, License: 1458298-3.
Figure 21(a-e,h) , are reprinted with permission. (https://moorhen.me.uk/)
Figure 21(f-g,i-k) , are reprinted with permission from Sue Chaplin.
Figure 24(a,h) , are reprinted with permission: Creative Common.
Figure 24(b) and Figure 23(a) , are reprinted with permission from Springer Nature: Nature, License: 5743871467428.
Figure 24(c) , is reprinted with permission from the American Institute of Aeronautics and Astronautics (AIAA): Atmospheric Flight Mechanocs Conference, License: 1476830-1.
Figure 24(d-e) , are reprinted with permission from Brill Publishers: Israel Journal of Ecology and Evolution.
Figure 24(f-g) , are reprinted with permission from the American Institute of Aeronautics and Astronautics (AIAA): AIAA SciTech, License: 1458902-6.
Figure 24(i-k) , are reprinted with permission from The Company of Biologists Ltd.: The journal of experimental biology.
Figure 19(d), is reprinted with permission from Claudio Contreras Koob. emailed
Figure 19(e), is reprinted with permission from John Comisky. emailed
Figure 19(g) , is reprinted with permission from Kirk Zufelt.
Figure 22(a) , is reprinted with permission from Oxford Publishing Press: Oxford Publishing Limited, License: 91621.
Figure 22(b-d) , are reprinted with permission from Springer Nature BV: Nature, License: 1457966-13.
Figure 25(a-d,g) , are reprinted with permission from Springer Nature BV: Swarm intelligence, License: 1457959-1.
Figure 25(e,f) reproduced from Elsevier Masson SAS for electronic distribution. All rights reserved.
Figure 27(a-b) , are reprinted with permission: Creative Commons.
Figure 27(c,e) , are reprinted with permission from John Wiley and Sons - Books: IBIS, License: 1458968-1.
Figure 27(d) , is reprinted with permission from The Company of Biologists Ltd.: The journal of experimental biology.
Figure 26(a-b) , are reprinted with permission from The Royal Society (U.K.): Proceedings of the Royal Society of London. Series B: Biological Sciences. Biological sciences, License: 1458961-6.
Figure 26(c) , is reprinted with permission from John Wiley and Sons - Books: Ecology letters, License: 1458969-1.
Figure 26(d) , is reprinted with permission from The Royal Society (U.K.): Biology letters, License: 1458961-8.
Figure 26(e,f) , are reprinted with permission from Stephanie Thompson.
Figure 34(a-e) , are reprinted with permission from The Company of Biologists Ltd.: The journal of experimental biology.
Figure 34(f) , is reprinted with permission from The Company of Biologists Ltd.: The journal of experimental biology.
Figure 34(g) , is reprinted with permission from Elsevier Science and Technology Journals: Journal of insect physiology, License: 1458298-4.
Figure 34(h-i) , are reprinted with permission from Cambridge University Press: Journal of Fluid Mechanics.
Figure 33(a,c-e) , are reprinted with permission from Dr. Andrew Davis.
Figure 33(b) , is reprinted with permission: Creative Commons.
Figure 29(a-b) , are reprinted with permission: Creative Commons.
Figure 29(c) , is reprinted with permission from Dr. Andrew Davis.
Figure 29(f) , is reprinted with permission: Creative Commons.
Figure 29(g) , is reprinted with permission from Dr. John Peter Whitney.
Figure 29(h) , is reprinted with permission: Creative Commons.
Figure 31(a) , is reprinted with permission from Oxford University Press: American Entomologist, License: 5743271043687.
Figure 31(b-c) , are reprinted with permission: Creative Commons.
Figure 31(d-e) , are reprinted with permission from IOP Publishing, Ltd: Bioinspiration and Biomimetics, License: 1457966-5.
Figure 31(f) , is reprinted with permission from Oxford University Press: Integrative and Comparative Biology, License: 5743271277116.
Figure 30(a-c) , are reprinted with permission from Elsevier: Biophysical Journal, License: 5743210634543.
Figure 30(d) , is reprinted with permission: Creative Commons.
Figure 30(e-g) , are reprinted with permission: Creative Commons.
Figure 30(h) , is reprinted with permission: Creative Commons.
Figure 32(a-d) , are reprinted with permission from IOP Publishing, Ltd: Bioinspiration and Biomimetics, License: 1457966-7.
Figure 36(a) , is reprinted with permission from IOP Publishing, Ltd: Bioinspiration and Biomimetics, License: 1457966-9.
Figure 36(b-e) , are reprinted with permission: Creative Commons.
Figure 38(a) , is reprinted with permission from The Institute of Electrical and Electronics Engineers (IEEE): IEEE.
Figure 38(e) , is reprinted with permission: Creative Commons.
References
- Sources of Greenhouse Gas Emissions, 2023.
- Chen, H.; Rao, F.; Shang, X.; Zhang, D.; Hagiwara, I. Biomimetic drag reduction study on herringbone riblets of bird feather. Journal of Bionic Engineering 2013, 10, 341–349. [Google Scholar] [CrossRef]
- Gavrilovic, N.; Mohamed, A.; Marino, M.; Watkins, S.; Moschetta, J.M.; Benard, E. Avian-inspired energy-harvesting from atmospheric phenomena for small UAVs. Bioinspiration and Biomimetics 2018, 14. [Google Scholar] [CrossRef] [PubMed]
- Aboelezz, A.; Herkenhoff, B.; Hassanalian, M. Effects of avian wings color patterns on their flight performance: experimental and computational studies. Journal of Bionic Engineering 2022. [Google Scholar] [CrossRef]
- Gammill, M.; Sherman, M.; Raissi, A.; Hassanalian, M. Energy Harvesting Mechanisms for a Solar Photovoltaic Plant Monitoring Drone: Thermal Soaring and Bioinspiration. In Proceedings of the AIAA Scitech 2021 Forum. American Institute of Aeronautics and Astronautics, 2021, p. 1053. [CrossRef]
- Hassanalian, M.; Mohammadi, S.; Acosta, G.; Guido, N.; Bakhtiyarov, S. Surface temperature effects of solar panels of fixed-wing drones on drag reduction and energy consumption. Meccanica 2021, 56, 3–22. [Google Scholar] [CrossRef]
- Omer, A.M. Energy, environment and sustainable development. Renewable and Sustainable Energy Reviews 2008, 12, 2265–2300. [Google Scholar] [CrossRef]
- Aldheeb, M.A.; Asrar, W.; Sulaeman, E.; Omar, A.A. A review on aerodynamics of nonflapping bird wings. Journal of Aerospace Technology and Management 2016, 8, 7–17. [Google Scholar] [CrossRef]
- Walsh, M.J. Riblets as a Viscous Drag Reduction Technique. AIAA Journal 1983, 21, 485–486. [Google Scholar] [CrossRef]
- Lang, A.; Habegger, M.L.; Motta, P. Shark Skin Drag Reduction. In Encyclopedia of Nanotechnology; Bhushan, B., Ed.; Springer Netherlands: Dordrecht, 2012; pp. 2394–2400. [Google Scholar] [CrossRef]
- Dean, B.; Bhushan, B. Shark-skin surfaces for fluid-drag reduction in turbulent flow: A review. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 2010, 368, 4775–4806. [Google Scholar] [CrossRef]
- Walsh, M.; Lindemann, A. Optimization and application of riblets for turbulent drag reduction. In 22nd Aerospace Sciences Meeting; Aerospace Sciences Meetings, American Institute of Aeronautics and Astronautics, 1984. [CrossRef]
- Domel, A.G.; Saadat, M.; Weaver, J.C.; Haj-Hariri, H.; Bertoldi, K.; Lauder, G.V. Shark skin-inspired designs that improve aerodynamic performance. Journal of the Royal Society Interface 2018, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bechert, D.W.; Bruse, M.; Hage, W.; Meyer, R. Fluid mechanics of biological surfaces and their technological application. Naturwissenschaften 2000, 87, 157–171. [Google Scholar] [CrossRef]
- Benschop, H.O.; Breugem, W.P. Drag reduction by herringbone riblet texture in direct numerical simulations of turbulent channel flow. Journal of Turbulence 2017, 18, 717–759. [Google Scholar] [CrossRef]
- Chen, H.W.; Rao, F.G.; Zhang, D.Y.; Shang, X.P. Drag reduction study about bird feather herringbone riblets. Applied Mechanics and Materials 2014, 461, 201–205. [Google Scholar] [CrossRef]
- Chen, H.; Rao, F.; Shang, X.; Zhang, D.; Hagiwara, I. Flow over bio-inspired 3D herringbone wall riblets. Experiments in Fluids 2014, 55. [Google Scholar] [CrossRef]
- Feng, B.; Chen, D.; Wang, J.; Yang, X. Bionic research on bird feather for drag reduction. Advances in Mechanical Engineering 2015, 7. [Google Scholar] [CrossRef]
- Sharma, V.; Dutta, S. Experimental and Numerical Investigation of Bio-Inspired Riblet for Drag Reduction. Journal of Fluids Engineering 2022, 145. [Google Scholar] [CrossRef]
- Aboelezz, A.; Rubin, Z.; Hassanalian, M. Heated Boundary Layer and Aerodynamic Efficiency of Airfoils: Birds Coloration and Bioinspiration. In Proceedings of the AIAA Scitech 2021 Forum. American Institute of Aeronautics and Astronautics, jan 2021, AIAA SciTech Forum, p. 848. [CrossRef]
- Achache, Y.; Sapir, N.; Elimelech, Y. Hovering hummingbird wing aerodynamics during the annual cycle. Ii. implications of wing feather moult. Royal Society Open Science 2018, 5. [Google Scholar] [CrossRef]
- Ajanic, E.; Paolini, A.; Coster, C.; Floreano, D.; Johansson, C. Robotic Avian Wing Explains Aerodynamic Advantages of Wing Folding and Stroke Tilting in Flapping Flight. Advanced Intelligent Systems 2023, 5, 1–12. [Google Scholar] [CrossRef]
- Aldheeb, M.; Asrar, W.; Sulaeman, E.; Omar, A.A. Aerodynamics of porous airfoils and wings. Acta Mechanica 2018, 229, 3915–3933. [Google Scholar] [CrossRef]
- Allen, R. The Whooping Crane; National Audobon Society, 1952.
- Alonso, J.C.; Bautista, L.M.; Alonso, J.A. Sexual size dimorphism in the Common Crane, a monogamous, plumage-monomorphic bird. Ornis Fennica 2019, 96, 194–204. [Google Scholar] [CrossRef]
- Ananda, G.K.; Selig, M.S. Design of bird-like airfoils. AIAA Aerospace Sciences Meeting, 2018 2018, pp. 1–21. [CrossRef]
- Anderton, J.; Rasmussen, P. Birds of South Asia The Ripley Guide, 2 ed.; Smithsonian Institution & Lynx Edicions, 2005; p. 315.
- Azargoon, Y.; Djavareshkian, M.H. Unsteady characteristic study on the flapping wing with the corrugated trailing edge and slotted wingtip. Aerospace Science and Technology 2023, 139. [Google Scholar] [CrossRef]
- Bae, Y.; Jeong, Y.E.; Moon, Y.J. Computation of flow past a flat plate with porous trailing edge using a penalization method. Computers and Fluids 2012, 66, 39–51. [Google Scholar] [CrossRef]
- Baliga, B.; Szabo, I.; Altshuler, D.L. Range of motion in the avian wing is strongly associated with flight behavior and body mass. Science Advances 2019, 5. [Google Scholar] [CrossRef]
- Bao, H.; Song, B.; Yang, W.; Xuan, J.; Xue, D. The progress of aerodynamic mechanisms based on avian leading-edge alula and future study recommendations. Aerospace 2021, 8. [Google Scholar] [CrossRef]
- Bright, J.A.; Marugán-Lobón, J.; Cobb, S.N.; Rayfield, E.J. The shapes of bird beaks are highly controlled by nondietary factors. Proceedings of the National Academy of Sciences of the United States of America 2016, 113, 5352–5357. [Google Scholar] [CrossRef]
- Brooke, M. Albatrosses and petrels across the world; Vol. 11, Oxford University Press, 2004.
- Brown, C. Plumages And Measurements Of The Bearded Vulture In Southern Africa. Ostrich 1989, 60, 165–171. [Google Scholar] [CrossRef]
- Bushnell, D.M.; Moore, K.J. Drag reduction in nature. Annual Review of Fluid Mechanics 1991, 23, 65–79. [Google Scholar] [CrossRef]
- Carboneras, C.; Jutglar, F.; Kirwan, G. Great Shearwater Ardenna gravis, 2020. [CrossRef]
- Carruthers, A.C.; Walker, S.M.; Thomas, A.L.; Taylor, G.K. Aerodynamics of aerofoil sections measured on a free-flying bird. Proceedings of the Institution of Mechanical Engineers, Part G: Journal of Aerospace Engineering 2010, 224, 855–864. [Google Scholar] [CrossRef]
- Carwardine, M.; Natural History Museum (London, E. Animal Records; Sterling, 2008.
- Catry, P.; Phillips, R.A.; Croxall, J.P. Sustained fast travel by a gray-headed albatross (Thalassarche chrysostoma) riding an antarctic storm. Auk 2004, 121, 1208–1213. [Google Scholar] [CrossRef]
- Czikeli, H. Whooping Crane, 1982.
- del Hoyo, J.; Elliott, A.; Sargatal, J. Ostrich to Ducks, 1992.
- del Hoyo, J.; Elliott, A.; Sargatal, J.; Christie, D.; de Juana, E. Manx Shearwater (Puffinus puffinus); Lynx Edicions, 2013.
- Drovetski, S.V. Influence Of The Trailing-Edge Notch On Flight Performance Of Galliforms. Auk 1996, 113, 802–810. [Google Scholar] [CrossRef]
- Dunn, J. Accidentals, Extinct Species. National Geographic Field Guide to the Birds of North America (fifth ed.) 2006, p. 467.
- Dunning, J.B. CRC Handbook of Avian Body Masses; CRC Press, 2007.
- Dutta, P.; Nagar, O.P.; Sahu, S.K.; Savale, R.R.; Gokul Raj, R. Aerodynamic analysis of bionic winglet- slotted wings. Materials Today: Proceedings 2022, 62, 6701–6707. [Google Scholar] [CrossRef]
- Eder, H.; Fiedler, W.; Neuhäuser, M. Evaluation of aerodynamic parameters from infrared laser tracking of free-gliding white storks. Journal of Ornithology 2015, 156, 667–677. [Google Scholar] [CrossRef]
- Elphick, J. The Atlas of Bird Migrations: Tracing the Great Journeys of the World’s Birds; Struik Publishers, 2007; pp. 88–89.
- Elphick, J. The Atlas of Bird Migration: Tracing the Great Journeys of the World’s Birds, illustrate ed.; Firefly Books, 2011.
- Ferguson-Lees, J. Raptors of the World; Houghton Mifflin Harcourt, 2001.
- Gargiulo, C. Distribución y situación actual del cóndor andino (Vultur gryphus) en las sierras centrales de Argentina. PhD thesis, Universidad de Buenos Aires Facultad, 2012.
- Goumas, M. Dark wing pigmentation as a mechanism for improved flight efficiency in the Larinae. Communications Biology 2022, 5, 1–6. [Google Scholar] [CrossRef]
- Halsey, L.G.; Portugal, S.J.; Smith, J.A.; Murn, C.P.; Wilson, R.P. Recording raptor behavior on the wing via accelerometry. Journal of Field Ornithology 2009, 80, 171–177. [Google Scholar] [CrossRef]
- Hancock, J.; Kushlan, J.; Kahl, M.P. Storks, Ibises, and Spoonbills of the World, 1 ed.; Academic Press, 1992.
- Hanna, Y.G.; Spedding, G.R. Aerodynamic performance improvements due to porosity in wings at moderate re. In Proceedings of the AIAA Aviation 2019 Forum, 2019, number June, pp. 1–11. [CrossRef]
- Harrison, P. Seabirds , an Identification Guide. The Wilson Journal of Ornithology 1984, 96, 333–335. [Google Scholar]
- Hassanalian, M.; Abdelmoula, H.; Ben Ayed, S.; Abdelk, A. Effects of birds’ wing color on their flight performance for biomimetic purposes. 25th AIAA/AHS Adaptive Structures Conference, 2017 2017, pp. 1–8. [CrossRef]
- Hassanalian, M.; Abdelmoula, H.; Ben Ayed, S.; Abdelkefi, A. Thermal impact of migrating birds’ wing color on their flight performance: Possibility of new generation of biologically inspired drones. Journal of Thermal Biology 2017, 66, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Hassanalian, M.; Ayed, S.B.; Ali, M.; Houde, P.; Hocut, C.; Abdelkefi, A. Insights on the thermal impacts of wing colorization of migrating birds on their skin friction drag and the choice of their flight route. Journal of Thermal Biology 2018, 72, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Hedenström, A. Effects of wing damage and moult gaps on vertebrate flight performance. Journal of Experimental Biology 2023, 226. [Google Scholar] [CrossRef]
- Hedenström, A.; Liechti, F. Field estimates of body drag coefficient on the basis of dives in passerine birds. Journal of Experimental Biology 2001, 204, 1167–1175. [Google Scholar] [CrossRef]
- Hedenström, A.; Sunada, S. On the aerodynamics of moult gaps in birds. Journal of Experimental Biology 1999, 202, 67–76. [Google Scholar] [CrossRef]
- Hossain, A.; Rahman, A.; Hossen, J.; Iqbal, P.; Shaari, N.; Sivaraj, G.K. Drag reduction in a wing model using a bird feather like winglet. Jordan Journal of Mechanical and Industrial Engineering 2011, 5, 267–272. [Google Scholar]
- Hua, J.; Kong, F.; Liu, H.H. Unsteady thermodynamic computational fluid dynamics simulations of aircraft wing anti-icing operation. Journal of Aircraft 2007, 44, 1113–1117. [Google Scholar] [CrossRef]
- Huber, G.H.; Turbek, S.P.; Bostwick, K.S.; Safran, R.J. Comparative analysis reveals migratory swallows (Hirundinidae) have less pointed wings than residents. Biological Journal of the Linnean Society 2017, 120, 228–235. [Google Scholar] [CrossRef]
- Iosilevskii, G. Aerodynamics of permeable membrane wings. European Journal of Mechanics, B/Fluids 2011, 30, 534–542. [Google Scholar] [CrossRef]
- Johnsgard, P.A. Cranes of the World; Indiana University Press, 1983.
- Jouventin, P.; Weimerskirch, H. Satellite tracking of Wandering albatrosses. Nature 1990, 343, 746–748. [Google Scholar] [CrossRef]
- Kong, W.; Dong, H.; Wu, J.; Zhao, Y.; Jin, Z. Experimental Study on the Effect of Porous Media on the Aerodynamic Performance of Airfoils. Aerospace 2023, 10, 1–16. [Google Scholar] [CrossRef]
- Kricher, J. Black-and-white Warbler Mniotilta varia, 2020. [CrossRef]
- Kuyt, E. WHOOPING CRANE. Hinterland Who’s Who 1993.
- Kuyt, E. Aerial radio-tracking of whooping cranes migrating between Wood Buffalo National Park and Aransas National Wildlife Refuge, 1981-84; Vol. 74, Authority of the Minister of Environment Canadian Wildlife Service, 1992.
- Lang, X.; Song, B.; Yang, W.; Song, W. Aerodynamic performance of owl-like airfoil undergoing bio-inspired flapping kinematics. Chinese Journal of Aeronautics 2021, 34, 239–252. [Google Scholar] [CrossRef]
- Laybourne, R.C. Collision between a Vulture and an Aircraft at an Altitude of 37,000 Feet. The Wilson Bulletin 1974, 86, 461–462. [Google Scholar]
- Lee, S.Y.; Scott, G.R.; Milsom, W.K. Have wing morphology or flight kinematics evolved for extreme high altitude migration in the bar-headed goose? Comparative Biochemistry and Physiology - C Toxicology and Pharmacology 2008, 148, 324–331. [Google Scholar] [CrossRef]
- Lees, J.J.; Dimitriadis, G.; Nudds, R.L. The influence of flight style on the aerodynamic properties of avian wings as fixed lifting surfaces. PeerJ 2016, 2016. [Google Scholar] [CrossRef]
- Li, Q.; Shen, H.; Han, Q.; Ji, A.; Dai, Z.; Gorb, S.N. Effects of Morphological Integrity of Secondary Feather on Their Drag Reduction in Pigeons. Journal of Bionic Engineering 2022, 19, 1422–1438. [Google Scholar] [CrossRef]
- Likoff, L. Adelie Penguin to Budgerigar; Number v. 6 in The Encyclopedia of Birds, Facts on File, 2007.
- Lincoln, F. Migration of Birds, illustrate ed.; U.S. Government Printing Office, 1999.
- Liu, D.; Song, B.; Yang, W.; Yang, X.; Xue, D.; Lang, X. A Brief Review on Aerodynamic Performance of Wingtip Slots and Research Prospect. Journal of Bionic Engineering 2021, 18, 1255–1279. [Google Scholar] [CrossRef]
- Liu, T.; Kuykendoll, K.; Rhew, R.; Jones, S. Avian wing geometry and kinematics. AIAA Journal 2006, 44, 954–963. [Google Scholar] [CrossRef]
- Liu, T.; Wang, S.; Liu, H.; He, G. Engineering perspective on bird flight: Scaling, geometry, kinematics and aerodynamics. Progress in Aerospace Sciences, 2023; 100933. [Google Scholar] [CrossRef]
- Lovvorn, J.R.; Liggins, G.A.; Borstad, M.H.; Calisal, S.M.; Mikkelsen, J. Hydrodynamic drag of diving birds: Effects of body size, body shape and feathers at steady speeds. Journal of Experimental Biology 2001, 204, 1547–1557. [Google Scholar] [CrossRef]
- March, A.I.; Bradley, C.W.; Garcia, E. Aerodynamic Properties Of Avian Flight As A Function Of Wing Shape. In Proceedings of the International Mechanical Engineering Congress and Exposition (IMECE), Orlando, Florida, 2005; pp. 1–9. [CrossRef]
- Maybury, W.J. The aerodynamics of bird bodies. PhD thesis, University of Bristol, 2000.
- Mazellier, N.; Feuvrier, A.; Kourta, A. Biomimetic bluff body drag reduction by self-adaptive porous flaps. Comptes Rendus - Mecanique 2012, 340, 81–94. [Google Scholar] [CrossRef]
- Mowbray, T.; Fred, C.; Barbara, G. Snow Goose (Chen caerulescens), 2000. [CrossRef]
- Müller, W.; Patone, G. Air transmissivity of feathers. Journal of Experimental Biology 1998, 201, 2591–2599. [Google Scholar] [CrossRef] [PubMed]
- Murayama, Y.; Nakata, T.; Liu, H. Flexible Flaps Inspired by Avian Feathers Can Enhance Aerodynamic Robustness in low Reynolds Number Airfoils. Frontiers in Bioengineering and Biotechnology 2021, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Narayan, G.; John, B. Effect of winglets induced tip vortex structure on the performance of subsonic wings. Aerospace Science and Technology 2016, 58, 328–340. [Google Scholar] [CrossRef]
- Navalón, G.; Bright, J.A.; Marugán-Lobón, J.; Rayfield, E.J. The evolutionary relationship among beak shape, mechanical advantage, and feeding ecology in modern birds*. Evolution 2019, 73, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Omar, A.; Rahuma, R.; Emhemmed, A. Numerical investigation on aerodynamic performance of bird’s airfoils. Journal of Aerospace Technology and Management 2020, 12, 1–17. [Google Scholar] [CrossRef]
- Osváth, G.; Vincze, O.; David, D.C.; Nagy, L.J.; Lendvai, Á.Z.; Nudds, R.L.; Pap, P.L. Morphological characterization of flight feather shafts in four bird species with different flight styles. Biological Journal of the Linnean Society 2020, 131, 192–202. [Google Scholar] [CrossRef]
- Owen, D. The rufous and white forms of an Asiatic paradise flycatcher, Terpsiphone paradisi. Museum of Zoology, University of Michigan 1959.
- Öztürk, Ş.; Örs, İ. An overview for effects on aerodynamic performance of using winglets and wingtip devices on aircraft. International Journal of Aeronautics and Astronautics 2020, 1, 1–11. [Google Scholar]
- Pap, P.L.; Osváth, G.; Sándor, K.; Vincze, O.; Bărbos, L.; Marton, A.; Nudds, R.L.; Vágási, C.I. Interspecific variation in the structural properties of flight feathers in birds indicates adaptation to flight requirements and habitat. Functional Ecology 2015, 29, 746–757. [Google Scholar] [CrossRef]
- Pellerito, V.; Hassanalian, M.; Sedaghat, A.; Sabri, F.; Borvayeh, L.; Sadeghi, S. Performance analysis of a bioinspired albatross airfoil with heated top wing surface: experimental study. AIAA Propulsion and Energy Forum and Exposition, 2019. [CrossRef]
- Pennycuick, J. The flight of petrels and albatrosses (procellariiformes), observed in South Georgia and its vicinity. Philosophical Transactions of the Royal Society of London. B, Biological Sciences 1982, 300, 75–106. [Google Scholar] [CrossRef]
- Qin, S.; Weng, Z.; Li, Z.; Xiang, Y.; Liu, H. On the controlled evolution for wingtip vortices of a flapping wing model at bird scale. Aerospace Science and Technology 2021, 110, 106460. [Google Scholar] [CrossRef]
- Rader, J.; Hedrick, T.L. Morphological evolution of bird wings follows a mechanical sensitivity gradient determined by the aerodynamics of flapping flight Mechanical sensitivity drives wing shape evolution. bioRxiv 2022, pp. 1–32.
- Rajesh Senthil Kumar, T.; Shriram, S.V.; Chowdary, G.P.; Sagar, J.; Ramakrishnananda, B. Aerodynamic characteristics of avian airfoils. AIP Conference Proceedings 2019, 2134, 1–5. [Google Scholar] [CrossRef]
- Richardson, P.L. How do albatrosses fly around the world without flapping their wings? Progress in Oceanography 2011, 88, 46–58. [Google Scholar] [CrossRef]
- Roberts, A. The Design and Experimental Optimization of a Wingtip Vortex Turbine for General Aviation Use. PhD thesis, Embry Riddle Aeronautical University, 1997.
- Robertson, R. Albatrosses (Diomedeidae), 2 ed.; Cengage Gale, 2002.
- Rogalla, S.; D’Alba, L.; Verdoodt, A.; Shawkey, M.D. Hot wings: Thermal impacts of wing coloration on surface temperature during bird flight. Journal of the Royal Society Interface 2019, 16. [Google Scholar] [CrossRef]
- Rogalla, S.; Nicolaï, M.P.; Porchetta, S.; Glabeke, G.; Battistella, C.; D’Alba, L.; Gianneschi, N.C.; Van Beeck, J.; Shawkey, M.D. The evolution of darker wings in seabirds in relation to temperature-dependent flight efficiency. Journal of the Royal Society Interface 2021, 18. [Google Scholar] [CrossRef]
- Sachs, G.; Traugott, J.; Nesterova, A.P.; Dell’Omo, G.; Kümmeth, F.; Heidrich, W.; Vyssotski, A.L.; Bonadonna, F. Flying at No Mechanical Energy Cost: Disclosing the Secret of Wandering Albatrosses. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Savile, D.B.O. Adaptive Evolution in the Avian Wing. Society for the Study of Evolution 1957, 11, 212–224. [Google Scholar]
- Sinclair, I.; Hockey, P. SASOL Larger Illustrated Guide to Birds of Southern Africa; New Holland Publishers, 2005.
- Stetson, K.F.; Kimmel, R.L. Surface temperature effects on boundary-layer transition. AIAA Journal 1992, 30, 2782–2783. [Google Scholar] [CrossRef]
- Stevenson, T.; Fanshawe, J. Birds of East Africa: Kenya, Tanzania, Uganda, Rwanda, Burundi Second Edition, 2 ed.; Princeton University Press, 2020.
- Swift, K.M. An Experimental Analysis of the Laminar Separation Bubble At Low Reynolds Numbers. PhD thesis, The University of Tennessee, 2009.
- Tangermann, E.; Ercolani, G.; Klein, M. Aerodynamic Behavior of a Biomimetic Wing in Soaring Flight – A Numerical Study. Flow, Turbulence and Combustion 2022, 109, 1155–1173. [Google Scholar] [CrossRef]
- Traub, L.W.; Coffman, C. Efficient low-Reynolds-number airfoils. Journal of Aircraft 2019, 56, 1987–2003. [Google Scholar] [CrossRef]
- Tucker, V.A. Drag Reduction by Wing Tip Slots in a Gliding Harris’ Hawk, Parabuteo Unicinctus. Journal of Experimental Biology 1995, 198, 775–781. [Google Scholar] [CrossRef]
- Tucker, V.A. Gliding Birds: Reduction of Induced Drag by Wing Tip Slots Between the Primary Feathers. Journal of Experimental Biology 1993, 180, 285–310. [Google Scholar] [CrossRef]
- Van Oorschot, B.K. Aerodynamics and Ecomorphology of Flexible Feathers and Morphing Bird Wings. PhD thesis, University of Montana, 2017.
- Van Wassenbergh, S.; Baeckens, S. Digest: Evolution of shape and leverage of bird beaks reflects feeding ecology, but not as strongly as expected. Evolution 2019, 73, 621–622. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.; Bishop, C.M.; Woakes, A.J.; Butler, P.J. Heart rate and the rate of oxygen consumption of flying and walking barnacle geese (Branta leucopsis) and bar-headed geese (Anser indicus). Journal of Experimental Biology 2002, 205, 3347–3356. [Google Scholar] [CrossRef]
- Warham, J., Ed. The Behaviour, Population Biology and Physiology of the Petrels; Vol. 21, Harcourt Brace & Company, 1998; p. 114. [CrossRef]
- Warham, J. Wing loadings, wing shapes, and flight capabilities of procellariiformes. New Zealand Journal of Zoology 1977, 4, 73–83. [Google Scholar] [CrossRef]
- Whiteman, L. The High Life. Audubon 2000, 102, 104–108. [Google Scholar]
- Yang, S.L.; Spedding, G.R. Passive separation control by acoustic resonance. Experiments in Fluids 2013, 54. [Google Scholar] [CrossRef]
- Yilmaz, A.; Tekeci, A.; Ozyetkin, M.; Demircali, A.; Unsal, K.; Uvet, H. The effect of pore structure in flapping wings on flight performance. arXiv 2021, arXiv:2106.04390. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, Y. Study on riblet drag reduction considering the effect of sweep angle. Energies 2019, 12. [Google Scholar] [CrossRef]
- Adhikari, D.R.R.; Bhattacharya, S. Ground Effect-Enhanced Unsteady Aerodynamic of Rapid-Pitch-Up Motions for Diverse Flying Objectives in Birds. In AIAA SCITECH 2024 Forum; AIAA SciTech Forum, American Institute of Aeronautics and Astronautics, 2024. [CrossRef]
- Ákos, Z.; Nagy, M.; Leven, S.; Vicsek, T. Thermal soaring flight of birds and unmanned aerial vehicles. Bioinspiration and Biomimetics 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Baddoo, P.J.; Kurt, M.; Ayton, L.J.; Moored, K.W. Exact solutions for ground effect. Journal of Fluid Mechanics 2020, 891, 2, [1912.02713]. [CrossRef]
- Blake, R.W. Mechanics of gliding in birds with special reference to the influence of the ground effect. Journal of Biomechanics 1983, 16, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Bohrer, G.; Brandes, D.; Mandel, J.T.; Bildstein, K.L.; Miller, T.A.; Lanzone, M.; Katzner, T.; Maisonneuve, C.; Tremblay, J.A. Estimating updraft velocity components over large spatial scales: Contrasting migration strategies of golden eagles and turkey vultures. Ecology Letters 2012, 15, 96–103. [Google Scholar] [CrossRef]
- Duerr, A.E.; Miller, T.A.; Lanzone, M.; Brandes, D.; Cooper, J.; O’Malley, K.; Maisonneuve, C.; Tremblay, J.A.; Katzner, T. Flight response of slope-soaring birds to seasonal variation in thermal generation. Functional Ecology 2015, 29, 779–790. [Google Scholar] [CrossRef]
- Duriez, O.; Kato, A.; Tromp, C.; Dell’Omo, G.; Vyssotski, A.L.; Sarrazin, F.; Ropert-Coudert, Y. How cheap is soaring flight in raptors? A preliminary investigation in freely-flying vultures. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Finn, J.; Carlsson, J.; Kelly, T.; Davenport, J. Avoidance of headwinds or exploitation of ground effect-why do birds fly low. Journal of Field Ornithology 2012, 83, 192–202. [Google Scholar] [CrossRef]
- Guo, D.; Zhang, X.; Zhang, J.; Li, H. An Interface Management Approach for Civil Aircraft Design; Vol. 680 LNEE, 2021; pp. 435–446. [CrossRef]
- Hainsworth, F.R. Induced Drag Savings From Ground Effect and Formation Flight in Brown Pelicans. Journal of Experimental Biology 1988, 135, 431–444. [Google Scholar] [CrossRef]
- Harel, R.; Nathan, R. The characteristic time-scale of perceived information for decision-making: Departure from thermal columns in soaring birds. Functional Ecology 2018, 32, 2065–2072. [Google Scholar] [CrossRef]
- Johansson, L.C.; Jakobsen, L.; Hedenström, A. Flight in Ground Effect Dramatically Reduces Aerodynamic Costs in Bats. Current Biology 2018, 28, 3502–3507.e4. [Google Scholar] [CrossRef]
- Jung, K.H.; Chun, H.H.; Kim, H.J. Experimental investigation of wing-in-ground effect with a NACA6409 section. Journal of Marine Science and Technology 2008, 13, 317–327. [Google Scholar] [CrossRef]
- Kempton, J.A.; Wynn, J.; Bond, S.; Evry, J.; Fayet, A.L.; Gillies, N.; Guilford, T.; Kavelaars, M.; Juarez-Martinez, I.; Padget, O.; et al. Optimization of dynamic soaring in a flap-gliding seabird affects its large-scale distribution at sea. Science Advances 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Khaghani, J.; Nekoui, M.; Nasiri, R.; Ahmadabadi, M.N. Analytical Model of Thermal Soaring: Towards Energy Efficient Path Planning for Flying Robots. IEEE International Conference on Intelligent Robots and Systems 2018, pp. 7589–7594. [CrossRef]
- Khosravifard, S.; Venus, V.; Skidmore, A.K.; Bouten, W.; Muñoz, A.R.; Toxopeus, A.G. Identification of Griffon Vulture’s Flight Types Using High-Resolution Tracking Data. International Journal of Environmental Research 2018, 12, 313–325. [Google Scholar] [CrossRef]
- King, R.M.; Gopalarathnam, A. Ideal aerodynamics of ground-effect and formation flight. AIAA Paper 2004, 42, 11500–11511. [Google Scholar] [CrossRef]
- Liu, D.N.; Hou, Z.X.; Guo, Z.; Yang, X.X.; Gao, X.Z. Bio-inspired energy-harvesting mechanisms and patterns of dynamic soaring. Bioinspiration and Biomimetics 2017, 12. [Google Scholar] [CrossRef]
- Mallon, J.M.; Bildstein, K.L.; Katzner, T.E. In-flight turbulence benefits soaring birds. Auk 2016, 133, 79–85. [Google Scholar] [CrossRef]
- Mandel, J.T. Using Movement Ecology To Understand Flight Behavior In Soaring Birds 2009.
- Miller, T.A.; Brooks, R.P.; Lanzone, M.J.; Brandes, D.; Cooper, J.; Tremblay, J.A.; Wilhelm, J.; Duerr, A.; Katzner, T.E. Limitations and mechanisms influencing the migratory performance of soaring birds. Ibis 2016, 158, 116–134. [Google Scholar] [CrossRef]
- Mohamed, A.; Taylor, G.K.; Watkins, S.; Windsor, S.P. Opportunistic soaring by birds suggests new opportunities for atmospheric energy harvesting by flying robots. Journal of the Royal Society Interface 2022, 19. [Google Scholar] [CrossRef]
- Norberg, U.M.L.; Brooke, A.P.; Trewhella, W.J. Soaring And Non-Soaring Bats Of The Family Pteropodidae 2000. 664, 651–664.
- Pekarsky, S.; Shohami, D.; Horvitz, N.; Bowie, R.C.K.; Kamath, P.L.; Markin, Y.; Getz, W.M.; Nathan, R. Cranes soar on thermal updrafts behind cold fronts as they migrate across the sea. In Proceedings of the Proceedings of Royal Society B, 2024, number 291. [CrossRef]
- Penn, M.; Yi, G.; Watkins, S.; Martinez Groves-Raines, M.; Windsor, S.P.; Mohamed, A. A method for continuous study of soaring and windhovering birds. Scientific Reports 2022, 12, 1–9. [Google Scholar] [CrossRef]
- Pennycuick, C.J. Field Observations of Thermals and Thermal Streets , and the Theory of Cross-Country Soaring Flight. Journal of Avian Biology 1998, 29, 33–43. [Google Scholar] [CrossRef]
- Pennycuick, C.J. Soaring Behaviour and Performance of Some East African Birds, Observed From a Motor-Glider. Ibis 1972, 114, 178–218. [Google Scholar] [CrossRef]
- Pennycuick, C.J. The Soaring Flight of Vultures. Scientific American 1973, 229, 102–109. [Google Scholar] [CrossRef]
- Pokhrel, S.; Eisa, S.A. A novel hypothesis for how albatrosses optimize their flight physics in real-time: an extremum seeking model and control for dynamic soaring. Bioinspiration and Biomimetics 2023, 18. [Google Scholar] [CrossRef] [PubMed]
- Rayner, J.M. On the aerodynamics of animal flight in ground effect. Royal Society, 1991; 119–128. [Google Scholar]
- Rayner, J.M.; Thomas, A.L. On the vortex wake of an animal flying in a confined volume. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 1991, 334, 107–117. [Google Scholar] [CrossRef]
- Richardson, P.L. Leonardo da Vinci’s Discovery of the Dynamic Soaring by Birds in Wind Shear. Royal Society, 2018; 285–301. [Google Scholar] [CrossRef]
- Richardson, P.L.; Wakefield, E.D.; Phillips, R.A. Flight speed and performance of the wandering albatross with respect to wind. Movement Ecology 2018, 6, 1–15. [Google Scholar] [CrossRef]
- Sachs, G. Minimum shear wind strength required for dynamic soaring of albatrosses. Ibis 2005, 147, 1–10. [Google Scholar] [CrossRef]
- Sachs, G. In-flight measurement of upwind dynamic soaring in albatrosses. Progress in Oceanography 2016, 142, 47–57. [Google Scholar] [CrossRef]
- Sakornsin, R.; Atipan, S. Experimental Investigation of Seabird-Like Wings in Ground Effect. Journal of Aeronautics, Astronautics and Aviation 2019, 51, 213–224. [Google Scholar] [CrossRef]
- Santos, C.D.; Hanssen, F.; Muñoz, A.R.; Onrubia, A.; Wikelski, M.; May, R.; Silva, J.P. Match between soaring modes of black kites and the fine-scale distribution of updrafts. Scientific Reports 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Serrano, F.J.; Chiappe, L.M. Aerodynamic modelling of a Cretaceous bird reveals thermal soaring capabilities during early avian evolution. Journal of the Royal Society Interface 2017, 14. [Google Scholar] [CrossRef]
- Shamoun-Baranes, J.; Leshem, Y.; Yom-Tov, Y.; Liechti, O. Differential Use of Thermal Convection by Soaring Birds over Central Israel. The Condor 2003, 105, 208–218. [Google Scholar] [CrossRef]
- Song, J. Fly low: The ground effect of a barn owl (Tyto alba) in gliding flight. Proceedings of the Institution of Mechanical Engineers, Part C: Journal of Mechanical Engineering Science 2021, 235, 308–318. [Google Scholar] [CrossRef]
- Stokes, I.A.; Lucas, A.J. Wave-slope soaring of the brown pelican. Movement Ecology 2021, 9, 1–13. [Google Scholar] [CrossRef]
- Su, J.Y.; Tang, J.H.; Wang, C.H.; Yang, J.T. A numerical investigation on the ground effect of a flapping-flying bird. Physics of Fluids 2013, 25. [Google Scholar] [CrossRef]
- Tallman, J.; Yu, D.; Pantoja, M. Thermal detection for free flight. Journal of Physics: Conference Series 2021, 1828, 1–3. [Google Scholar] [CrossRef]
- Tang, J.H.; Su, J.Y.; Wang, C.H.; Yang, J.T. Numerical investigation of the ground effect for a small bird. Journal of Mechanics 2013, 29, 433–441. [Google Scholar] [CrossRef]
- Thorup, K.; Alerstam, T.; Hake, M.; Kjellén, N. Traveling or stopping of migrating birds in relation to wind: An illustration for the osprey. Behavioral Ecology 2006, 17, 497–502. [Google Scholar] [CrossRef]
- van Erp, J.; Sage, E.; Bouten, W.; van Loon, E.; Camphuysen, K.C.; Shamoun-Baranes, J. Thermal soaring over the North Sea and implications for wind farm interactions. Marine Ecology Progress Series 2023, 723, 185–200. [Google Scholar] [CrossRef]
- Wang, W.; An, W.; Song, B. The Mechanisms of Albatrosses’ Energy-Extraction During the Dynamic Soaring; Vol. 680 LNEE, Springer Singapore, 2021; pp. 507–516. [CrossRef]
- Warham, J. The behaviour, population biology and physiology of the petrels; Academic Press, 1996. [CrossRef]
- Weinzierl, R.; Bohrer, G.; Kranstauber, B.; Fiedler, W.; Wikelski, M.; Flack, A. Wind estimation based on thermal soaring of birds. Ecology and Evolution 2016, 6, 8706–8718. [Google Scholar] [CrossRef]
- Williams, H.J.; Shepard, E.L.; Holton, M.D.; Alarcón, P.A.; Wilson, R.P.; Lambertucci, S.A. Physical limits of flight performance in the heaviest soaring bird. Proceedings of the National Academy of Sciences of the United States of America 2020, 117, 17884–17890. [Google Scholar] [CrossRef]
- Willis, D.J.; Peraire, J.; Breuer, K.S. A computational investigation of bio-inspired formation flight and ground effect. Collection of Technical Papers - AIAA Applied Aerodynamics Conference 2007, 2, 1140–1174. [Google Scholar] [CrossRef]
- Withers, P.C.; Timko, P.L. The Significance of Ground Effect to the Aerodynamic Cost of Flight and Energetics of the Black Skimmer ( Rhyncops Nigra ). Journal of Experimental Biology 1977, 70, 13–26. [Google Scholar] [CrossRef]
- Hassanalian, M.; Abdelkefi, A. Classifications, applications, and design challenges of drones: A review. Progress in Aerospace Sciences 2017, 91, 99–131. [Google Scholar] [CrossRef]
- Mueller, T.J. Fixed and Flapping Wing Aerodynamics for Micro Air Vehicle Applications; Vol. 195, Progress in Astronautics and Aeronautics, American Institute of Aeronautics and Astronautics: Reston, VA, 2001.
- Bannasch, R., From Soaring and Flapping Bird Flight to Innovative Wing and Propeller Constructions. In Fixed and Flapping Wing Aerodynamics for Micro Air Vehicle Applications; Mueller, T.J., Ed.; American Institute of Aeronautics and Astronautics: Reston, VA, 2001; Vol. 195, Progress in Astronautics and Aeronautics, pp. 449–477.
- Kato, N.; Kamimura, S. Biomechanisms of Swimming and Flying; Springer: Tokyo, New York, 2004. [Google Scholar]
- Shyy, W.; Aono, H.; Kang, C.k.; Liu, H. An introduction to flapping wing aerodynamics, 2013. https://doi.org/LK - https://nmt.on.worldcat.org/oclc/847664062.
- Harvey, C.; de Croon, G.; Taylor, G.K.; Bomphrey, R.J. Lessons from natural flight for aviation: then, now and tomorrow. Journal of Experimental Biology 2023, 226. [Google Scholar] [CrossRef]
- Shyy, W.; Lian, Y.; Tang, J.; Viieru, D.; Liu, H. Aerodynamics ofLow Reynolds Number Flyers; Cambridge University Press: Cambridge, 2008. [Google Scholar]
- Liu, T. Comparative scaling of flapping- and fixed-wing flyers. AIAA Journal 2006, 44, 24–33. [Google Scholar] [CrossRef]
- Pennycuick, C.J. Wingbeat frequency of birds in steady cruising flight: New data and improved predictions. Journal of Experimental Biology 1996, 199, 1613–1618. [Google Scholar] [CrossRef]
- Prandtl, L. Induced Drag of Multiplanes. Technical report, NASA, 1924.
- Prandtl, L. The Mechanics of Viscous Fluids. 1935.
- Jones, R.T. Wing Theory; Princeton University Press, 1990.
- Knoller, R.; Verein, O.F. Die Gesetze des Luftwiderstandes; Verlag des Österreichischer Flugtechnischen Vereines: Wien SE - 14 pages : illustrations ; 24 cm, 1909. https://doi.org/LK-https://worldcat.org/title/841616578.
- Betz, A. Ein Beitrag zur Erklärung des Segelfluges. Zeitschrift für Flugtechnik und Motorluftschiffahrt 1912, 3, 269–272. [Google Scholar]
- Birnbaum, W. Flugmechanik der Flügelschläge. Zeitschrift für Flugtechnik und Motorluftschiffahrt 1912, 15, 1–8. [Google Scholar]
- Theodorsen, T.; Mutchler, W.H.; for Aeronautics, U.S.N.A.C. General theory of aerodynamic instability and the mechanism of flutter; National Advisory Committee for Aeronautics: Washington, D.C. SE - 23 pages : illustrations ; 26 cm, 1935. https://doi.org/LK - https://worldcat.org/title/46448356.
- Jones, K.D.; Dohring, C.M.; Platzer, M.F. Experimental and Computational Investigation of the Knoller-Betz Effect. AIAA Journal 1998, 36, 1240–1246. [Google Scholar] [CrossRef]
- Yang, L.J.; Feng, A.L.; Lee, H.C.; Esakki, B.; He, W. The three-dimensional flow simulation of a flapping wing. Journal of Marine Science and Technology (Taiwan) 2018, 26, 297–308. [Google Scholar] [CrossRef]
- Crandell, K.E.; Howe, R.O.; Falkingham, P.L. Repeated evolution of drag reduction at the air–water interface in diving kingfishers. Journal of the Royal Society Interface 2019, 16. [Google Scholar] [CrossRef]
- Reddy, S.R.; Sobieczky, H.; Dulikravic, G.S.; Abdoli, A. Multi-element winglets: Multi-objective optimization of aerodynamic shapes. Journal of Aircraft 2016, 53, 992–1000. [Google Scholar] [CrossRef]
- Lyu, Y.Z.; Sun, M. Power requirements for the hovering flight of insects with different sizes. Journal of Insect Physiology 2021, 134, 104293. [Google Scholar] [CrossRef] [PubMed]
- Aono, H.; Liang, F.; Liu, H. Near- and far-field aerodynamics in insect hovering flight: An integrated computational study. Journal of Experimental Biology 2008, 211, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Roll, J.; Liu, Y.; Troolin, D.R.; Deng, X. Three-dimensional vortex wake structure of flapping wings in hovering flight. Journal of the Royal Society Interface 2014, 11. [Google Scholar] [CrossRef] [PubMed]
- Arora, P.R.; Hossain, A.; Edi, P.; Jaafar, A.A.; Younis, T.S.; Saleem, M. Drag Reduction in Aircraft Model Using Elliptical Winglet. The Institution of Engineer Malaysia 2005, 66, 6–13. [Google Scholar]
- Kernstine, K.H.; Moore, C.J.; Cutler, A.; Mittal, R. Initial Characterization of Self-Activated Movable Flaps, “Pop-Up Feathers”. In Proceedings of the 46th AIAA Aerospace Sciences Meeting and Exhibit, Reno, Nevada; 2008. [Google Scholar] [CrossRef]
- Rosti, M.E.; Kamps, L.; Bruecker, C.; Omidyeganeh, M.; Pinelli, A. The PELskin project-part V: towards the control of the flow around aerofoils at high angle of attack using a self-activated deployable flap. Meccanica 2017, 52, 1811–1824. [Google Scholar] [CrossRef]
- Schlüter, J.U. Lift enhancement at low Reynolds numbers using pop-up feathers. In Proceedings of the 39th AIAA Fluid Dynamics Conference, 2009, number June. [CrossRef]
- Ponitz, B.; Schmitz, A.; Fischer, D.; Bleckmann, H.; Brücker, C. Diving-flight aerodynamics of a peregrine falcon (Falco peregrinus). PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Schatz, M.; Knacke, T.; Thiele, F.; Meyer, R.; Hage, W.; Bechert, D.W. Separation Control by Self-Activated Movable Flaps. In Proceedings of the 42nd AIAA Aerospace Sciences Meeting & Exhibit, Reno, NV, 2004. [CrossRef]
- Guerrero, J.E.; Maestro, D.; Bottaro, A. Biomimetic spiroid winglets for lift and drag control. Comptes Rendus - Mecanique 2012, 340, 67–80. [Google Scholar] [CrossRef]
- KleinHeerenbrink, M.; Christoffer Johansson, L.; Hedenström, A. Multi-cored vortices support function of slotted wing tips of birds in gliding and flapping flight. Journal of the Royal Society Interface 2017, 14. [Google Scholar] [CrossRef] [PubMed]
- Hui, Z.; Cheng, G.; Chen, G. Experimental investigation on tip-vortex flow characteristics of novel bionic multi-tip winglet configurations. Physics of Fluids 2021, 33. [Google Scholar] [CrossRef]
- La Roche, U.; Palffy, S. WING-GRID, a Novel Device for Reduction of Induced Drag on Wings. In Proceedings of the International Civil Aviation Convention, Switzerland; 1996; pp. 2303–2309. [Google Scholar]
- Rong, J.; Liu, H. Effects of owl-inspired leading-edge serrations on tandem wing aeroacoustics. AIP Advances 2022, 12, 1–21. [Google Scholar] [CrossRef]
- Lynch, M.; Mandadzhiev, B.; Wissa, A. Bioinspired wingtip devices: A pathway to improve aerodynamic performance during low Reynolds number flight. Bioinspiration and Biomimetics 2018, 13. [Google Scholar] [CrossRef]
- Brothers, E. Airbus unveils hybrid-electric regional airliner concept, 2019.
- Hubbard, L. Peacock at the Cincinnati Zoo, 2016.
- A leading-edge alula-inspired device (LEAD) for stall mitigation and lift enhancement for low Reynolds number finite wings. In Proceedings of the ASME Conference on Smart Materials, Adaptive Structures and Intelligent Systems, San Antonio, TX, 2018; Vol. 2, pp. 1–9. [CrossRef]
- Chamorro, L.P.; Lynch, M.K. Alula-inspired Leading Edge Device for Low Reynolds Number Flight. In Proceedings of the ASME Conference on Smart Materials, Adaptive Structures and Intelligent Systems, Stowe, VT, 2017; pp. 1–12.
- Mandadzhiev, B.A.; Lynch, M.K.; Chamorro, L.P.; Wissa, A.A. An experimental study of an airfoil with a bio-inspired leading edge device at high angles of attack. Smart Materials and Structures 2017, 26. [Google Scholar] [CrossRef]
- Shyy, W.; Liu, H. Flapping wings and aerodynamic lift: The role of leading-edge vortices. AIAA Journal 2007, 45, 2817–2819. [Google Scholar] [CrossRef]
- Linehan, T.; Mohseni, K. Investigation of a sliding alula for control augmentation of lifting surfaces at high angles of attack. Aerospace Science and Technology 2019, 87, 73–88. [Google Scholar] [CrossRef]
- Muijres, F.T.; Johansson, L.C.; Hedenström, A. Leading edge vortex in a slow-flying passerine. Biology Letters 2012, 8, 554–557. [Google Scholar] [CrossRef]
- Muijres, F.T.; Johansson, L.C.; Barfield, R.; Wolf, M.; Spedding, G.R.; Hedenström, A. Leading-Edge Vortex Improves Lift in. Science 2008, 319, 1250–1253. [Google Scholar] [CrossRef]
- Videler, J.J.; Stamhuis, E.J.; Povel, G.D. Leading-edge vortex lifts swifts. Science 2004, 306, 1957–1960. [Google Scholar] [CrossRef]
- Lentink, D.; Dickson, W.B.; Van Leeuwen, J.L.; Dickinson, M.H. Leading-edge vortices elevate lift of autorotating plant seeds. Science 2009, 324, 1438–1440. [Google Scholar] [CrossRef]
- Eldredge, J.D.; Jones, A.R. Leading-edge vortices: Mechanics and modeling. Annual Review of Fluid Mechanics 2019, 51, 75–104. [Google Scholar] [CrossRef]
- Meseguer, J.; Franchini, S.; Pérez-Grande, I.; Sanz, J.L. On the aerodynamics of leading-edge high-lift devices of avian wings. In Proceedings of the Proceedings of the Institution of Mechanical Engineers, Part G: Journal of Aerospace Engineering, 2005, Vol. 219, pp. 63–68. [CrossRef]
- Linehan, T.; Mohseni, K. On the maintenance of an attached leading-edge vortex via model bird alula. Journal of Fluid Mechanics 2020, 897, 2001.03964. [Google Scholar] [CrossRef]
- Linehan, T.; Mohseni, K. Scaling trends of bird’s alular feathers in connection to leading-edge vortex flow over hand-wing. Scientific Reports 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Austin, B.; Anderson, A.M. The alula and its aerodynamic effect on avian flight. ASME International Mechanical Engineering Congress and Exposition, Proceedings (IMECE) 2007, 7, 797–806. [Google Scholar] [CrossRef]
- Lee, S.I.; Kim, J.; Park, H.; Jabłoński, P.G.; Choi, H. The function of the alula in avian flight. Scientific Reports 2015, 5, 1–5. [Google Scholar] [CrossRef]
- Ito, M.R.; Duan, C.; Wissa, A.A. The function of the alula on engineered wings: A detailed experimental investigation of a bioinspired leading-edge device. Bioinspiration and Biomimetics 2019, 14. [Google Scholar] [CrossRef]
- Hubel, T.Y.; Tropea, C. The importance of leading edge vortices under simplified flapping flight conditions at the size scale of birds. Journal of Experimental Biology 2010, 213, 1930–1939. [Google Scholar] [CrossRef] [PubMed]
- Sander, A. The Role of the Alula in Avian Flight and Its application to Small Aircraft: A Numerical Study. PhD thesis, University of Grogingen, 2018.
- Ge, C.; Ren, L.; Liang, P.; Zhang, C.; Zhang, Z. High-Lift effect of bionic slat based on owl wing. Journal of Bionic Engineering 2013, 10, 456–463. [Google Scholar] [CrossRef]
- Denny, M. Dynamic soaring : aerodynamics for albatrosses Dynamic soaring : aerodynamics for 2009. [CrossRef]
- Pennycuick, C.J. Gust soaring as a basis for the flight of petrels and albatrosses (Procellariiformes). Avian Science 2002, 2, 1–12. [Google Scholar]
- Videler, J.J. Bird flight modes. In Avian Flight; Oxford Academic, 2006; chapter 6, pp. 118–155. [CrossRef]
- Hassanalian, M.; Abdelmoula, H.; Ben Ayed, S.; Abdelkefi, A. Thermal impact of migrating birds’ wing color on their flight performance: Possibility of new generation of biologically inspired drones. Journal of Thermal Biology 2017, 66, 27–32. [Google Scholar] [CrossRef]
- Brown, J.M.; Bouten, W.; Camphuysen, K.C.; Nolet, B.A.; Shamoun-Baranes, J. Energetic and behavioral consequences of migration: an empirical evaluation in the context of the full annual cycle. Scientific Reports 2023, 13, 1–15. [Google Scholar] [CrossRef]
- Mizrahy-Rewald, O.; Perinot, E.; Fritz, J.; Vyssotski, A.L.; Fusani, L.; Voelkl, B.; Ruf, T. Empirical Evidence for Energy Efficiency Using Intermittent Gliding Flight in Northern Bald Ibises. Frontiers in Ecology and Evolution 2022, 10. [Google Scholar] [CrossRef]
- Shamoun-Baranes, J.; van Loon, E.; van Gasteren, H.; van Belle, J.; Bouten, W.; Buurma, L. A Comparitive Analysis Of The Influence Of Weather On The Flight Altitudes Of Birds. American Meteorological Society 2006. [Google Scholar] [CrossRef]
- Rayner, J.M. Bounding and undulating flight in birds. Journal of Theoretical Biology 1985, 117, 47–77. [Google Scholar] [CrossRef]
- Rayner, J.M.V. A New Approach to Animal Flight Mechanics. Journal of Experimental Biology 1979, 80, 17–54. [Google Scholar] [CrossRef]
- Mir, I.; Eisa, S.A.; Maqsood, A. Review of dynamic soaring: technical aspects, nonlinear modeling perspectives and future directions. Nonlinear Dynamics 2018, 94, 3117–3144. [Google Scholar] [CrossRef]
- Al-Atabi, M. Aerodynamics of wing tip sails mushtak al-atabi. 2006; 1, 89–98. [Google Scholar]
- Sun, C.; Dai, C. Experimental Study on Ground Effect of a Wing with Tip Sails; Vol. 126, Elsevier B.V., 2015; pp. 559–563. [CrossRef]
- Gavrilović, N.N.; Rašuo, B.P.; Dulikravich, G.S.; Parezanović, V.B. Commercial aircraft performance improvement using winglets. FME Transactions 2015, 43, 1–8. [Google Scholar] [CrossRef]
- Wan, T.; Chou, H.C.; Lien, K.W. Aerodynamic efficiency study of modern spiroid winglets. ICAS-Secretariat - 25th Congress of the International Council of the Aeronautical Sciences 2006 2006, 2, 707–713. [Google Scholar]
- Falcão, L.; Gomes, A.A.; Suleman, A. Design and analysis of an adaptive wingtip. In Proceedings of the 52nd AIAA/ASME/ASCE/AHS/ASC Structures, Structural Dynamics and Materials Conference, 2011, number April. [CrossRef]
- Van Bokhorst, E.; De Kat, R.; Elsinga, G.E.; Lentink, D. Feather roughness reduces flow separation during low Reynolds number glides of swifts. Journal of Experimental Biology 2015, 218, 3179–3191. [Google Scholar] [CrossRef]
- Servini, P. Roughing up wings: boundary layer separation over static and dynamic roughness elements. Doctoral dissertation, UCL (University College London), 2018.
- Maybury, W.J.; Rayner, J.M. The avian tail reduces body parasite drag by controlling flow separation and vortex shedding. Proceedings of the Royal Society B: Biological Sciences 2001, 268, 1405–1410. [Google Scholar] [CrossRef]
- Tucker, V.A. Body drag, feather drag and interference drag of the mounting strut in a peregrine falcon, Falco peregrinus. Journal of Experimental Biology 1990, 149, 449–468. [Google Scholar] [CrossRef]
- Pennycuick, C.J.; Obrecht, H.H.; Fuller, M.R. Empirical Estimates of Body Drag of Large Waterfowl and Raptors. Journal of Experimental Biology 1988, 135, 253–264. [Google Scholar] [CrossRef]
- Vincent, L.; Xiao, T.; Yohann, D.; Jung, S.; Kanso, E. Dynamics of water entry. Journal of Fluid Mechanics 2018, 846, 508–535. [Google Scholar] [CrossRef]
- Sterling, J.; Jaramillo, A.; Hayes, F. Bird Guide Training Curriculum: Advanced Level; National Audubon Society, 2015; p. 5.
- Ropert-Coudert, Y.; Grémillet, D.; Ryan, P.; Kato, A.; Naito, Y.; Le Maho, Y. Between air and water: The plunge dive of the Cape Gannet Morus capensis. Ibis 2004, 146, 281–290. [Google Scholar] [CrossRef]
- Stephenson, R. Diving energetics in lesser scaup (Aythyta affinis, Eyton). Journal of Experimental Biology 1994, 190, 155–178. [Google Scholar] [CrossRef]
- Chang, B.; Croson, M.; Straker, L.; Gart, S.; Dove, C.; Gerwin, J.; Jung, S. How seabirds plunge-dive without injuries. Proceedings of the National Academy of Sciences of the United States of America 2016, 113, 12006–12011. [Google Scholar] [CrossRef]
- Foo, C.T.; Omar, B.; Taib, I. Shape Optimization of High-Speed Rail by Biomimetic. MATEC Web of Conferences 2017, 135, 1–13. [Google Scholar] [CrossRef]
- Laman, T. Vogelkop Lophorina ML62128001, 2017.
- Bissett, K.; Duncan, J. Victoria’s Riflebird (Ptiloris victoriae) ML384020521, 2021. [CrossRef]
- R, W. Greater Sage-Grouse (Centrocercus urophasianus) ML432171241, 2022. [CrossRef]
- Venetz, C. Wilson’s Bird-of-Paradise (Diphyllodes respublica) ML588187181, 2023. [CrossRef]
- Seminario, Y. Crimson-backed Tanager (Ramphocelus dimidiatus) ML314404811, 2013. [CrossRef]
- De Munck, P. Resplendent Quetzal (Pharomachrus mocinno) ML251030911, 2019. [CrossRef]
- Videler, J.; Groenewold, A. Field measurements of hanging flight aerodynamics in the kestrel Falco tinnunculus. Journal of Experimental Biology 1991, 155, 519–530. [Google Scholar] [CrossRef]
- Harvey, C.; Inman, D.J. Aerodynamic efficiency of gliding birds vs comparable UAVs: A review. Bioinspiration and Biomimetics 2021, 16. [Google Scholar] [CrossRef]
- Zufelt, K. Manx Shearwater ML216800971, 2020.
- Ornelas, R. White Pelican Pelecanus onocrotalus, 2006.
- Hunt, S. High Soarer, 2015.
- Patel, C.; Lee, H.T.; Kroo, I. Extracting energy from atmospheric turbulence with flight tests. Technical Soaring 2009, 33, 100–108. [Google Scholar]
- Katzmayr, R. Effect of periodic changes in angle of attack on behavior of airfoils. Technical Report NACA-TM-147, NACA, 1922.
- Lissaman, P. Wind energy extraction by birds and flight vehicles. In Proceedings of the 43rd AIAA aerospace sciences meeting and exhibit. AIAA, January 2005, p. 241.
- Rayner, J. The intermittent flight of birds. In Scale effects in animal locomotion; Academic Press: London, 1977; chapter The interm, pp. 437–444.
- Rayner, J.M. A vortex theory of animal flight. Part 1. The vortex wake of a hovering animal. Journal of Fluid Mechanics 1979, 91, 697–730. [Google Scholar] [CrossRef]
- Rayner, J.M. A vortex theory of animal flight. Part 1. The vortex wake of a hovering animal. Journal of Fluid Mechanics 1979, 91, 697–730. [Google Scholar] [CrossRef]
- Tobalske, B.W. Hovering and intermittent flight in birds. Bioinspiration and Biomimetics 2010, 5. [Google Scholar] [CrossRef]
- Tobalske, B.W.; Peacock, W.L.; Dial, K.P. Kinematics of flap-bounding flight in the zebra finch over a wide range of speeds. Journal of Experimental Biology 1999, 202, 1725–1739. [Google Scholar] [CrossRef]
- Tobalske, B.W. Morphology, velocity, and intermittent flight in birds’. American Zoologist 2001, 41, 177–187. [Google Scholar] [CrossRef]
- Tobalske, B.W. Scaling of muscle composition, wing morphology, and intermittent flight behavior in woodpeckers. Auk 1996, 113, 151–177. [Google Scholar] [CrossRef]
- Battell, R.; Battell, M. Image of the Day.
- Chaplin, S. Bounding Flight, 2019.
- Rayner, J.M.; Viscardi, P.W.; Ward, S.; Speakmanf, J.R. Aerodynamics and energetics of intermittent flight in birds1. American Zoologist 2001, 41, 188–204. [Google Scholar] [CrossRef]
- Keating, H.a. A Literature Review on Bounding Flight in Birds With Applications to Micro Uninhabited Air Vehicles. Semantic Scholar.
- Videler, J.J.; Weihs, D.; Daan, S. Intermittent Gliding in the Hunting Flight of the Kestrel, Falco Tinnunculus L. Journal of Experimental Biology 1983, 102, 1–12. [CrossRef]
- Vejdani, H.R.; Boerma, D.B.; Swartz, S.M.; Breuer, K.S. The dynamics of hovering flight in hummingbirds, insects and bats with implications for aerial robotics. Bioinspiration and Biomimetics 2019, 14, 0–30. [Google Scholar] [CrossRef] [PubMed]
- Warrick, D.R.; Tobalske, B.W.; Powers, D.R. Aerodynamics of the hovering hummingbird. Nature 2005, 435, 1094–1097. [Google Scholar] [CrossRef]
- Rayner, J.M. Avian flight energetics. Annual review of physiology 1982, 44, 109–119. [Google Scholar] [CrossRef]
- Wang, Y.; Yin, Y.; Ren, Z.; Jiang, C.; Sun, Y.; Li, J.; Nabi, G.; Wu, Y.; Li, D. A comparison of flight energetics and kinematics of migratory Brambling and residential Eurasian Tree Sparrow. Avian Research 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Chai, P.; Chen, J.S.; Dudley, R. Transient hovering performance of hummingbirds under conditions of maximal loading. Journal of Experimental Biology 1997, 200, 921–929. [Google Scholar] [CrossRef]
- Muijres, F.T.; Johansson, L.C.; Bowlin, M.S.; Winter, Y.; Hedenström, A. Comparing aerodynamic efficiency in birds and bats suggests better flight performance in birds. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Hummel, D. Formation flight as an energy -saving mechanism. Israel Journal of Zoology 1995, 41, 261–278. [Google Scholar] [CrossRef]
- Hummel, D. Formation Flight as an Energy-Saving Mechanism. Progress in Aerospace Sciences 2000, 35, 77–88. [Google Scholar]
- Martinez-Ponce, J.; Urban, C.; Armanini, S.F.; Agarwal, R.K.; Hassanalian, M. Aerodynamic Analysis of V-Shaped Flight Formation of Flapping-Wing Drones: Analytical and Experimental Studies. In Proceedings of the AIAA SCITECH 2022 Forum. American Institute of Aeronautics and Astronautics, dec 2021, AIAA SciTech Forum, p. 1979. [CrossRef]
- Gould, L.; Heppner, F. The Vee Formation of Canada Geese. The Auk 1974, 91, 494–506. [Google Scholar] [CrossRef]
- Portugal, S.J.; Hubel, T.Y.; Fritz, J.; Heese, S.; Trobe, D.; Voelkl, B.; Hailes, S.; Wilson, A.M.; Usherwood, J.R. Upwash exploitation and downwash avoidance by flap phasing in ibis formation flight. Nature 2014, 505, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Henise, D. American White Pelican, 2016.
- Weimerskirch, H.; Martin, J.; Clerquin, Y.; Alexandre, P.; Jiraskova, S. Energy saving in flight formation. Nature 2001, 413, 697–698. [Google Scholar] [CrossRef]
- Benson, J. ’V’ is for Vamoose, 2007.
- Hajihusseini, H. Eurasian Cranes migrating to Meyghan Salt Lake, 2010.
- Scott, J. V-Formation Flight of Birds, 2005.
- Cutts, C.J.; Speakman, J.R. Energy savings in formation flight of pink-footed geese. Journal of Experimental Biology 1994, 189, 251–261. [Google Scholar] [CrossRef]
- Higdon, J.J.L.; Corrsin, S. Induced Drag of a Bird Flock. The American Naturalist 1978, 112, 727–744. [Google Scholar] [CrossRef]
- Voelkl, B.; Portugal, S.J.; Unsölde, M.; Usherwood, J.R.; Wilsond, A.M.; Fritz, J. Matching times of leading and following suggest cooperation through direct reciprocity during V-formation flight in ibis. Proceedings of the National Academy of Sciences of the United States of America 2015, 112, 2115–2120. [Google Scholar] [CrossRef]
- Heppner, F.H.; Convissar, J.L.; Moonan, D.E.; John, G.; Anderson, T.; Auk, S.T.; Jan, N. Visual Angle and Formation Flight in Canada Geese (Branta canadensi. The Auk: Ornithological Advances 1985, 102, 195–198. [Google Scholar] [CrossRef]
- Ning, S.A. Aircraft Drag Reduction Through Extended Formation Flight. PhD thesis, STANFORD UNIVERSITY, 2011.
- Pahle, J.; Berger, D.; Venti, M.; Duggan, C.; Faber, J.; Cardinal, K. An initial flight investigation of formation flight for drag reduction on the C-17 aircraft. AIAA Atmospheric Flight Mechanics Conference 2012 2012, pp. 1–13. [CrossRef]
- Martinez-ponce, J.; Herkenhoff, B.; Aboelezz, A.; Hassanalian, M. Load Distribution on “ V ” and Echelon Formation Flight of. In Proceedings of the AIAA SciTech Forum, Orlando, FL, 2024; Number January. [CrossRef]
- Lambach, J.L. Integrating UAS Flocking Operations with Formation Drag, Thesis. Air Force Institue of Technology 2014, p. 99.
- Mirzaeinia, A.; Heppner, F.; Hassanalian, M. An analytical study on leader and follower switching in V-shaped Canada Goose flocks for energy management purposes. Swarm Intelligence 2020, 14, 117–141. [Google Scholar] [CrossRef]
- Mirzaeinia, A.; Hassanalian, M.; Lee, K.; Mirzaeinia, M. Energy conservation of V-shaped swarming fixed-wing drones through position reconfiguration. Aerospace Science and Technology 2019, 94, 105398. [Google Scholar] [CrossRef]
- Kshatriya, M.; Blake, R.W. Theoretical model of the optimum flock size of birds flying in formation. Journal of Theoretical Biology 1992, 157, 135–174. [Google Scholar] [CrossRef]
- Thien, H.P.; Moelyadi, M.A.; Muhammad, H. Effects of Leaders Position and Shape on Aerodynamic Performances of V Flight Formation. In Proceedings of the International Conference on Intelligent Unmanned System; 2008; pp. 43–49. [Google Scholar]
- Andersson, M.; Wallander, J. Kin selection and reciprocity in flight formation? Behavioral Ecology 2004, 15, 158–162. [Google Scholar] [CrossRef]
- Anderson, C.M.; Gilchrist, H.G.; Ronconi, R.A.; Shlepr, K.R.; Clark, D.E.; Fifield, D.A.; Robertson, G.J.; Mallory, M.L. Both short and long distance migrants use energy-minimizing migration strategies in North American herring gulls. Movement Ecology 2020, 8, 1–12. [Google Scholar] [CrossRef]
- Somveille, M.; Rodrigues, A.S.; Manica, A. Energy efficiency drives the global seasonal distribution of birds. Nature Ecology and Evolution 2018, 2, 962–969. [Google Scholar] [CrossRef]
- Somveille, M.; Wikelski, M.; Beyer, R.M.; Rodrigues, A.S.; Manica, A.; Jetz, W. Simulation-based reconstruction of global bird migration over the past 50,000 years. Nature Communications 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Alerstam, T.; Lindström, Å. Optimal bird migration: the relative importance of time, energy and safety. – In: Gwinner, E. (ed) Bird Migration: The Physiology and ecophysiology. Springer-Verlag, Berlin. pp. 331-351. Springer 1990, pp. 331–351.
- Acácio, M.; Catry, I.; Soriano-Redondo, A.; Silva, J.P.; Atkinson, P.W.; Franco, A.M. Timing is critical: consequences of asynchronous migration for the performance and destination of a long-distance migrant. Movement Ecology 2022, 10, 1–16. [Google Scholar] [CrossRef]
- Nourani, E.; Yamaguchi, N.M.; Higuchi, H. Climate change alters the optimal wind-dependent flight routes of an avian migrant. Proceedings of the Royal Society B: Biological Sciences 2017, 284, 2–7. [Google Scholar] [CrossRef]
- Lanzone, M.J.; Miller, T.A.; Turk, P.; Brandes, D.; Halverson, C.; Maisonneuve, C.; Tremblay, J.; Cooper, J.; O’Malley, K.; Brooks, R.P.; et al. Flight responses by a migratory soaring raptor to changing meteorological conditions. Biology Letters 2012, 8, 710–713. [Google Scholar] [CrossRef]
- Rüppel, G.; Hüppop, O.; Lagerveld, S.; Schmaljohann, H.; Brust, V. Departure, routing and landing decisions of long-distance migratory songbirds in relation to weather. Royal Society Open Science 2023, 10. [Google Scholar] [CrossRef]
- Kranstauber, B.; Weinzierl, R.; Wikelski, M.; Safi, K. Global aerial flyways allow efficient travelling. Ecology Letters 2015, 18, 1338–1345. [Google Scholar] [CrossRef]
- Thompson, S. Migratory behaviour of Eastern Whip-poor- wills ( Antrostomus vociferus ): quantifying return rates and the effects of artificial light on flight paths. PhD thesis, University of Manitoba, 2023.
- Dokter, A.M.; Liechti, F.; Stark, H.; Delobbe, L.; Tabary, P.; Holleman, I. Bird migration flight altitudes studied by a network of operational weather radars. Journal of the Royal Society Interface 2010, 8, 30–43. [Google Scholar] [CrossRef]
- Kemp, M.U.; Shamoun-Baranes, J.; Dokter, A.M.; van Loon, E.; Bouten, W. The influence of weather on the flight altitude of nocturnal migrants in mid-latitudes. International Journal of Avian Science 2013, 155, 734–749. [Google Scholar] [CrossRef]
- Mateos-Rodríguez, M.; Liechti, F. How do diurnal long-distance migrants select flight altitude in relation to wind? Behavioral Ecology 2012, 23, 403–409. [Google Scholar] [CrossRef]
- Scott, G.R. Elevated performance: The unique physiology of birds that fly at high altitudes. Journal of Experimental Biology 2011, 214, 2455–2462. [Google Scholar] [CrossRef]
- Altshuler, D.L. The physiology and biomechanics of avian flight at high altitude. Integrative and Comparative Biology 2006, 46, 62–71. [Google Scholar] [CrossRef]
- Thomas, B. Bird Flight Over Water | Loyola University Center for Environmental Communication, 2011.
- Cutler, C. Ground effect: Why You Float During Landing, 2020.
- Herkenhoff, B.; Hassanalian, M. Potential Applications and Integration of Ground Effects on Amphibious Drones: Shearwaters and Bioinspiration. AIAA AVIATION 2022 Forum 2022, pp. 1–9. [CrossRef]
- Tang, L.; Yang, Y.; Kiong Soh, C.; Halvorsen, E.; Nguyen, S.D.; Mann, B.P.; Pozzi, M.; Zhu, M.; Benasciutti, D.; Moro, L.; et al. Advances in Energy Harvesting Methods; Springer, 2013. [CrossRef]
- Whitney, J. Design and Performance of Insect-Scale Flapping-Wing Vehicles. PhD thesis, Harvard University, 2012.
- Ray, R.P.; Nakata, T.; Henningsson, P.; Bomphrey, R.J. Enhanced flight performance by genetic manipulation of wing shape in Drosophila. Nature Communications 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Satterfield, D.A.; Davis, A.K. Variation in wing characteristics of monarch butterflies during migration: Earlier migrants have redder and more elongated wings. Animal Migration 2014, 2, 1–7. [Google Scholar] [CrossRef]
- Young, J.; Walker, S.M.; Bomphrey, R.J.; Taylor, G.K.; Thomas, A.L. Details of insect wing design and deformation enhance aerodynamic function and flight efficiency. Science 2009, 325, 1549–1552. [Google Scholar] [CrossRef]
- Wootton, R. The geometry and mechanics of insect wing deformations in flight: A modelling approach. Insects 2020, 11, 1–19. [Google Scholar] [CrossRef]
- Engels, T.; Kolomenskiy, D.; Lehmann, F.O. Flight efficiency is a key to diverse wing morphologies in small insects. Journal of the Royal Society Interface 2021, 18. [Google Scholar] [CrossRef]
- Johansson, L.C.; Henningsson, P. Butterflies fly using efficient propulsive clap mechanism owing to flexible wings: Butterflies fly using efficient propulsive clap mechanism owing to flexible wings. Journal of the Royal Society Interface 2021, 18. [Google Scholar] [CrossRef]
- Flockhart, D.T.; Fitz-gerald, B.; Brower, L.P.; Derbyshire, R.; Altizer, S.; Hobson, K.A.; Wassenaar, L.I.; Norris, D.R. Migration distance as a selective episode for wing morphology in a migratory insect. Movement Ecology 2017, 5, 1–9. [Google Scholar] [CrossRef]
- Krishna, S.; Cho, M.; Wehmann, H.N.; Engels, T.; Lehmann, F.O. Wing design in flies: Properties and aerodynamic function. Insects 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Li, H.; Nabawy, M.R. Wing Planform Effect on the Aerodynamics of Insect Wings. Insects 2022, 13. [Google Scholar] [CrossRef]
- Liu, C.; Li, P.; Song, F.; Sun, J. Wing shape optimization design inspired by beetle hindwings in wind tunnel experiments. Computers in Biology and Medicine 2021, 135, 104642. [Google Scholar] [CrossRef]
- Engels, T.; Wehmann, H.N.; Lehmann, F.O. Three-dimensional wing structure attenuates aerodynamic efficiency in flapping fly wings. Journal of the Royal Society Interface 2020, 17. [Google Scholar] [CrossRef]
- Chen, D.; Kolomenskiy, D.; Liu, H. The impact of insect wing shape on the formation of leading edge vortex. The Proceedings of the Bioengineering Conference Annual Meeting of BED/JSME 2017, 2017.29, 2F22. [CrossRef]
- Watson, J.A.; Cribb, B.W.; Hu, H.M.; Watson, G.S. A dual layer hair array of the brown lacewing: Repelling water at different length scales. Biophysical Journal 2011, 100, 1149–1155. [Google Scholar] [CrossRef]
- Shen, T.; Tu, Z.; Li, D.; Kan, Z.; Xiang, J. Aerodynamic Characteristics of Bristled Wings in Flapping Flight. Aerospace 2022, 9. [Google Scholar] [CrossRef]
- Kolomenskiy, D. Recent Progress in the Biomechanics of Flight of Miniature Insects with Bristled Wings. Journal of the Physical Society of Japan 2023, 92, 1–5. [Google Scholar] [CrossRef]
- O’Callaghan, F.; Lehmann, F.O. Flow development and leading edge vorticity in bristled insect wings. Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology 2023, 209, 219–229. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, F.; Sarig, A.; Ribak, G.; Lehmann, F.O. Efficiency and Aerodynamic Performance of Bristled Insect Wings Depending on Reynolds Number in Flapping Flight. Fluids 2022, 7, 1–15. [Google Scholar] [CrossRef]
- Combes, S.A.; Daniel, T.L. Flexural stiffness in insect wings: Effects of wing venation and stiffness distribution on passive bending. American Entomologist 2005, 51, 42–44. [Google Scholar] [CrossRef]
- Dirks, J.H.; Taylor, D. Veins improve fracture toughness of insect wings. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Reid, H.E.; Schwab, R.K.; Maxcer, M.; Peterson, R.K.; Johnson, E.L.; Jankauski, M. Wing flexibility reduces the energetic requirements of insect flight. Bioinspiration and Biomimetics 2019, 14. [Google Scholar] [CrossRef]
- Salcedo, M.K.; Socha, J.J. Circulation in insect wings. Integrative and Comparative Biology 2020, 60, 1208–1220. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, W.; Gu, J.; Fan, T.; Liu, Q.; Su, H.; Zhu, S. Inspiration from butterfly and moth wing scales: Characterization, modeling, and fabrication. Progress in Materials Science 2015, 68, 67–96. [Google Scholar] [CrossRef]
- Slegers, N.; Heilman, M.; Cranford, J.; Lang, A.; Yoder, J.; Habegger, M.L. Beneficial aerodynamic effect of wing scales on the climbing flight of butterflies. Bioinspiration and Biomimetics 2017, 12. [Google Scholar] [CrossRef]
- Lang, A.W. Microscopic scales enhance a butterfly’s flying efficiency, 2023. [CrossRef]
- Wilroy, J.; Wahidi, R.A.; Lang, A. Effect of butterfly-scale-inspired surface patterning on the leading edge vortex growth. Fluid Dynamics Research 2018, 50. [Google Scholar] [CrossRef]
- Badejo, O.; Skaldina, O.; Gilev, A.; Sorvari, J. Benefits of insect colours : a review from social insect studies. Oecologia 2020, 194, 27–40. [Google Scholar] [CrossRef]
- Osotsi, M.I.; Zhang, W.; Zada, I.; Gu, J.; Liu, Q.; Zhang, D. Butterfly wing architectures inspire sensor and energy applications. National Science Review 2021, 8. [Google Scholar] [CrossRef]
- Davis, A.K.; Chi, J.; Bradley, C.; Altizer, S. The redder the better: Wing color predicts flight performance in monarch butterflies. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Davis, A.K.; Herkenhoff, B.; Vu, C.; Barriga, P.A.; Hassanalian, M. How the monarch got its spots: Long-distance migration selects for larger white spots on monarch butterfly wings. PloS one 2023, 18, e0286921. [Google Scholar] [CrossRef]
- Berman, G.J.; Wang, Z.J. Energy-minimizing kinematics in hovering insect flight. Journal of Fluid Mechanics 2007, 582, 153–168. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, C.; Zhu, X. Influences of flapping modes and wing kinematics on aerodynamic performance of insect hovering flight. Journal of Mechanical Science and Technology 2020, 34, 1603–1612. [Google Scholar] [CrossRef]
- Fry, S.N.; Sayaman, R.; Dickinson, M.H. The aerodynamics of hovering flight in Drosophila. Journal of Experimental Biology 2005, 208, 2303–2318. [Google Scholar] [CrossRef]
- Le Roy, C.; Amadori, D.; Charberet, S.; Windt, J.; Muijres, F.T.; Llaurens, V.; Debat, V. Adaptive evolution of flight in Morpho butterflies. Science 2021, 374, 1158–1162. [Google Scholar] [CrossRef]
- Cespedes, A.; Penz, C.M.; Devries, P.J. Cruising the rain forest floor: Butterfly wing shape evolution and gliding in ground effect. Journal of Animal Ecology 2015, 84, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Mena, S.; Kozak, K.M.; Cárdenas, R.E.; Checa, M.F. Forest stratification shapes allometry and flight morphology of tropical butterflies: Stratification and butterfly morphology. Proceedings of the Royal Society B: Biological Sciences 2020, 287, 1–10. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, J. Experimental investigation on aerodynamic performance of gliding butterflies. AIAA Journal 2010, 48, 2454–2457. [Google Scholar] [CrossRef]
- Betts, C.R.; Wootton, R.J. Wing Shape and Flight Behaviour in Butterflies (Lepidoptera: Papilionoidea and Hesperioidea): A Preliminary Analysis. Journal of Experimental Biology 1988, 138, 271–288. [Google Scholar] [CrossRef]
- Wakeling, J.M.; Ellington, C.P. Dragonfly flight: I gliding flight and steady-state aerodynamic forces. Journal of Experimental Biology 1997, 200, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Chaugule, S. Thermal Soaring and Control Surface Dynamics of an Eagle and the Hummingbird’s Flapping Flight Aerodynamics. International Journal for Research in Applied Science and Engineering Technology 2022, 10, 1161–1171. [Google Scholar] [CrossRef]
- Hawkins, S.; Vale, M.; Firth, L.; Burrows, M.; Mieszkowska, N.; Frost, M. Sustained Observation of Marine Biodiversity and Ecosystems. Oceanography: Open Access 2013, 01, 1–4. [Google Scholar] [CrossRef]
- Palumbi, S.R.; Sandifer, P.A.; Allan, J.D.; Beck, M.W.; Fautin, D.G.; Fogarty, M.J.; Halpera, B.S.; Incze, L.S.; Leong, J.A.; Norse, E.; et al. Managing for ocean biodiversity to sustain marine ecosystem services. Frontiers in Ecology and the Environment 2009, 7, 204–211. [Google Scholar] [CrossRef]
- Duffy, J.E.; Amaral-Zettler, L.A.; Fautin, D.G.; Paulay, G.; Rynearson, T.A.; Sosik, H.M.; Stachowicz, J.J. Envisioning a marine biodiversity observation network. BioScience 2013, 63, 350–361. [Google Scholar] [CrossRef]
- Zolich, A.; Palma, D.; Kansanen, K.; Fjørtoft, K.; Sousa, J.; Johansson, K.H.; Jiang, Y.; Dong, H.; Johansen, T.A. Survey on Communication and Networks for Autonomous Marine Systems. Journal of Intelligent and Robotic Systems: Theory and Applications 2019, 95, 789–813. [Google Scholar] [CrossRef]
- Rynne, P.F.; Von Ellenrieder, K.D. Unmanned autonomous sailing: Current status and future role in sustained ocean observations. Marine Technology Society Journal 2009, 43, 21–30. [Google Scholar] [CrossRef]
- Allen, B.; Austin, T.; Forrester, N.; Goldsborough, R.; Kukulya, A.; Packard, G.; Purcell, M.; Stokey, R. Autonomous docking demonstrations with enhanced REMUS technology. In Proceedings of the OCEANS; 2006; pp. 1–6. [Google Scholar] [CrossRef]
- Meindl, A. Guide to moored buoys and other ocean data acquisition systems. Data Buoy Cooperation Panel 1996, 8, 40. [Google Scholar]
- Shashati, M.; Ain, A.; Zeyoudi, S.A.; Ain, A.; Ain, A.; Khatib, O.A.; Ain, A.; Ain, A.; Hakim, A. Design and Fabrication of a Solar Powered Unmanned Aerial Vehicle ( UAV ). 2023 Advances in Science and Engineering Technology International Conferences (ASET) 2023, pp. 1–10. [CrossRef]
- Nileshkumar, S.K.; Prince, P.; Mukulbhai, K.J. Design and Development of Solar Drone. International Journal for Research in Applied Science & Engineering Technology 2023.
- Buzdugan, A.; Bălteanu, A. Numerical Modeling of an Energy Management System for a UAV Design Powered by Photovoltaic Cells. Review of the Air Force Academy 2022, 2, 63–70. [Google Scholar] [CrossRef]
- Noll, T.E.; Ishamel, S.D.; Perez-Davis, M.E.; Tiffany, G.C.; Madura, J.; Gaier, M.; Brown, J.M.; Wierzbanowski, T. Technical Findings , Lessons Learned , and Recommendations Resulting from the Helios Prototype Vehicle Mishap. In Proceedings of the NATO/RTO AVT-145 Workshop on Design Concepts, Florence, Italy, 2007.
- Dinca, L.; Corcau, J.I.; Voinea, D.G. Solar UAVs—More Aerodynamic Efficiency or More Electrical Power? Energies 2023, 16. [Google Scholar] [CrossRef]
- Chen, Y. Overview of Solar UAV Power System. Academic Journal of Science and Technology 2022, 4, 4–6. [Google Scholar] [CrossRef]
- Sudevalayam, S.; Kulkarni, P. Energy harvesting sensor nodes: Survey and implications. IEEE Communications Surveys and Tutorials 2011, 13, 443–461. [Google Scholar] [CrossRef]
- Kazmierski, T.J.; Beeby, S., Eds. Energy Harvesting Systems; 2011.
- Karami, M.A.; Inman, D.J. Powering pacemakers from heartbeat vibrations using linear and nonlinear energy harvesters. Applied Physics Letters 2012, 100. [Google Scholar] [CrossRef]
- Loughan, M.; Shoup, D.; Heise, M. Methods and Devices for Harvesting Ionic Energy to Produce Electricity, 2020.
- Lai, J.C. Airborne energy harvester for storing atmospheric static electrical energy.pdf, 2014.
- Huang, T.Y. ENERGY STORAGE DEVICE, 2015.
- Saunders, C. Charge separation mechanisms in clouds. Space Science Reviews 2008, 137, 335–353. [Google Scholar] [CrossRef]
- MacGorman, D.R.; Rust, W.D.; Williams, E.R. The electrical nature of storms. Oxford University Press, 1998.
- Bloemink, H. Static electricity measurements for lightning warnings - an exploration. Royal Netherlands Meteorological Institute 2013.
- Chu, J. MIT engineers fly first-ever plane with no moving parts, 2018.
- Cowen, L. Cloud Physics.
- Harrison, R.G.; Nicoll, K.A.; Ambaum, M.H. On the microphysical effects of observed cloud edge charging. Quarterly Journal of the Royal Meteorological Society 2015, 141, 2690–2699. [Google Scholar] [CrossRef]
- Marshall, T.C.; Stolzenburg, M. Electrical energy constraints on lightning. Journal of Geophysical Research: Atmospheres 2002, 107. [Google Scholar] [CrossRef]
- Jantunen, J.; Mäkelä, J.; Kaikuranta, T.; Reunamäki, J.; Aloha, T.; Uusitalo, A.; Kalliojarvi, K. Detection of Lightning, 2009.
- Helman, D.S. Catching lightning for alternative energy. Renewable Energy 2011, 36, 1311–1314. [Google Scholar] [CrossRef]
- Srinivasan, K.; Jason, G. Lightning as atmospheric electricity. Canadian Conference on Electrical and Computer Engineering 2006, pp. 2258–2261. [CrossRef]
- Xu, H.; He, Y.; Strobel, K.L.; Gilmore, C.K.; Kelley, S.P.; Hennick, C.C.; Sebastian, T.; Woolston, M.R.; Perreault, D.J.; Barrett, S.R. Flight of an aeroplane with solid-state propulsion. Nature 2018, 563, 532–535. [Google Scholar] [CrossRef] [PubMed]
- Varnava, C. A charged flight. Nature Electronics 2018, 1, 614. [Google Scholar] [CrossRef]
- Mirzaeinia, A.; Hassanalian, M.; Lee, K.; Mirzaeinia, M. Performance enhancement and load balancing of swarming drones through position reconfiguration. In Proceedings of the AIAA Aviation 2019 Forum, 2019, number June, pp. 1–17. [CrossRef]
- Herkenhoff, B.K.; Gee, P.; Cantu, D.; Mirzaeinia, A.; Silva, C.L.; Hassanalian, M. Harnessing Electrostatic Energy to Power Micro Drones. In Proceedings of the AIAA Scitech 2023 Forum, National Harbor, MD, 2023; Number January.
- Gavrilović, N.; Bronz, M.; Moschetta, J.m.; Gavrilović, N.; Bronz, M.; Bioinspired, J.m.M.; Harvesting, E.; Gavrilovi, N.; Bronz, M.; Moschetta, J.m. Bioinspired Energy Harvesting from Atmospheric Phenomena for Small Unmanned Aerial Vehicles. Journal of Guidance, Control, and Dynamics 2020. [CrossRef]
- Kiziroglou, M.E.; Becker, T.; Yeatman, E.M.; Schmid, U.; Evans, J.W.; Wright, P.K. Comparison of methods for static charge energy harvesting on aircraft. Smart Sensors, Actuators, and MEMS VIII 2017, 10246, 1–6. [Google Scholar] [CrossRef]
- Herkenhoff, B.; Fisher, J.; Dinelli, C.; Hassanalian, M. Energy harvesting techniques for hybrid amphibious drones in the marine environment. In Proceedings of the AIAA Propulsion and Energy 2020 Forum. American Institute of Aeronautics and Astronautics, 2020, p. 3959. [CrossRef]
- Herkenhoff, B.; Morris, I.; Ebrahimkhanlou, A.; Hassanalian, M. Application and Potential Design of an Amphibious Drone Hub: Water Lily and Bioinspiration Concepts. AIAA Aviation and Aeronautics Forum and Exposition, AIAA AVIATION Forum 2021 2021, pp. 1–13. [CrossRef]
- Rubin, Z.; Aboelezz, A.; Herkenhoff, B.; Hassanalian, M. Drones for venus exploration: Energy harvesting mechanisms and thermal-based flight control. AIAA Scitech 2021 Forum 2021, pp. 1–8. [CrossRef]
- Herkenhoff, B.K.; Fisher, J.M.; Hassanalian, M. Performance analysis of solar fixed-wing drones on venus. In Proceedings of the 18th VEXAG Meeting. NASA, 2020, [arXiv:1011.1669v3].
- Meyer, A. Migratory birds, Cranes, Nature image, 2023.
Figure 4.
Plot of wingtip roundedness against migration distance for swallows adapted from [65]. The linear regression shows a significant downward trend: r2 = 0.6057, P = 0.00023.
Figure 4.
Plot of wingtip roundedness against migration distance for swallows adapted from [65]. The linear regression shows a significant downward trend: r2 = 0.6057, P = 0.00023.
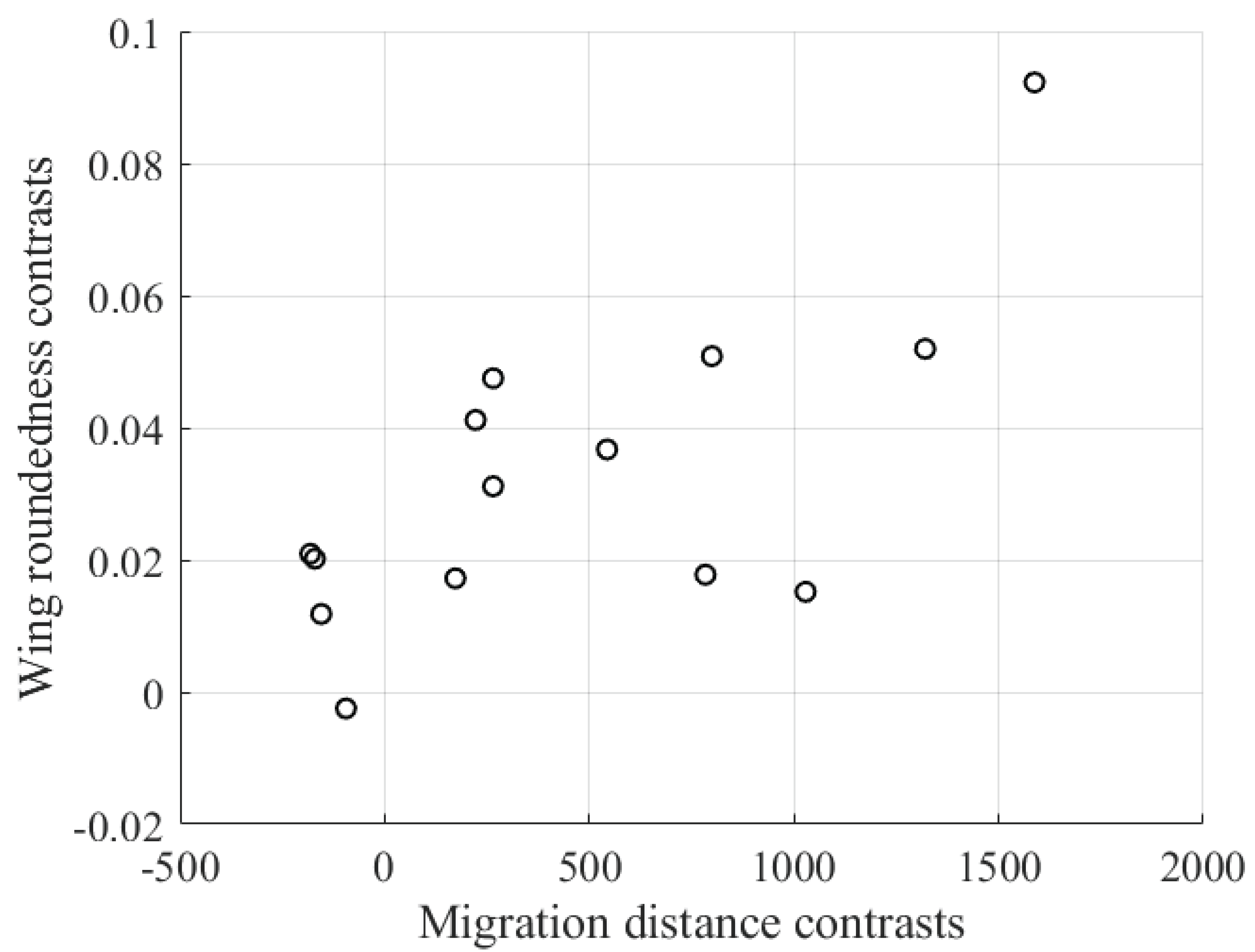
Figure 6.
a) Figures featuring the wingtip slots of a Harris hawk showing the horizontal and vertical separation [116]. b) Illustrations emphasizing the wingtip slots horizontal and vertical spacing [115]. c) Schematic showing another example of primary wingtip slots of an owl in gliding flight [80,211]. Views of wing tip slots in d) Kea, e) Pacific Brown Pelican, f) Red Tailed Hawk, g) and a Bald Eagle [80,207]. h) Wingtip slots introduced into a CAD model for testing and simulation [46]. i) Conceptual flow visualization showing the measurement planes for the slotted wing of a jackdaw [80,208]. j) Example bionic wing with varying numbers of wingtip slots [80,209]. k) Example of flow streamlines over a Great Horned Owl wingtip [80,84]. l) Another example of a bionic wing for testing wingtip slots installed in a wind tunnel for testing [80,212]. m) Conceptual aircraft applying wingtip slots designed by Airbus called the “bird of prey” [80,213]. n) Flight test of a grid like wingtip system similar to wingtip slots [80,210]. All figures are reprinted with permission; please see the Appendix A.
Figure 6.
a) Figures featuring the wingtip slots of a Harris hawk showing the horizontal and vertical separation [116]. b) Illustrations emphasizing the wingtip slots horizontal and vertical spacing [115]. c) Schematic showing another example of primary wingtip slots of an owl in gliding flight [80,211]. Views of wing tip slots in d) Kea, e) Pacific Brown Pelican, f) Red Tailed Hawk, g) and a Bald Eagle [80,207]. h) Wingtip slots introduced into a CAD model for testing and simulation [46]. i) Conceptual flow visualization showing the measurement planes for the slotted wing of a jackdaw [80,208]. j) Example bionic wing with varying numbers of wingtip slots [80,209]. k) Example of flow streamlines over a Great Horned Owl wingtip [80,84]. l) Another example of a bionic wing for testing wingtip slots installed in a wind tunnel for testing [80,212]. m) Conceptual aircraft applying wingtip slots designed by Airbus called the “bird of prey” [80,213]. n) Flight test of a grid like wingtip system similar to wingtip slots [80,210]. All figures are reprinted with permission; please see the Appendix A.
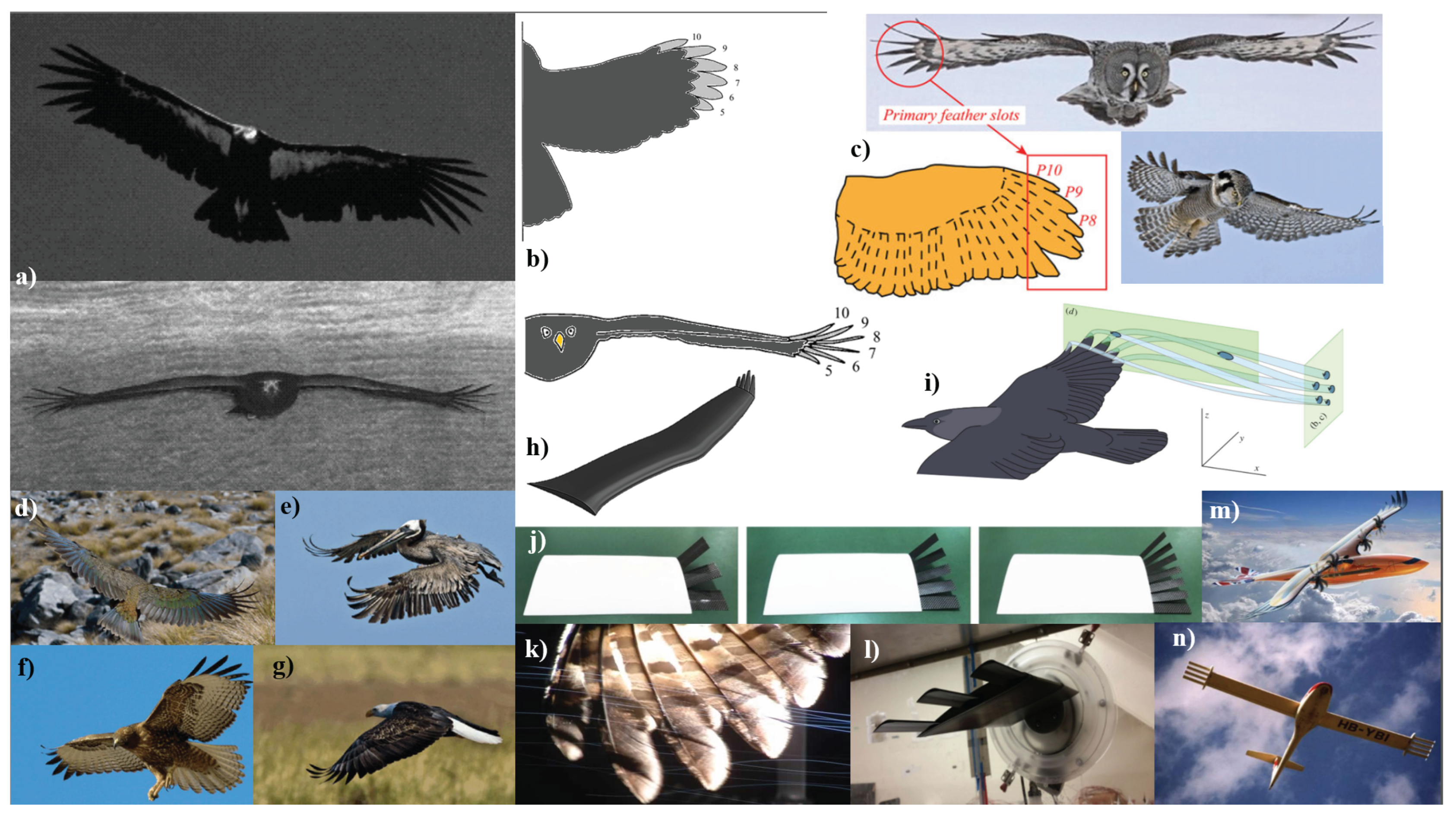
Figure 8.
Examples of several different avian airfoils that highlight the unique features of each type of flyer [73,82,101].
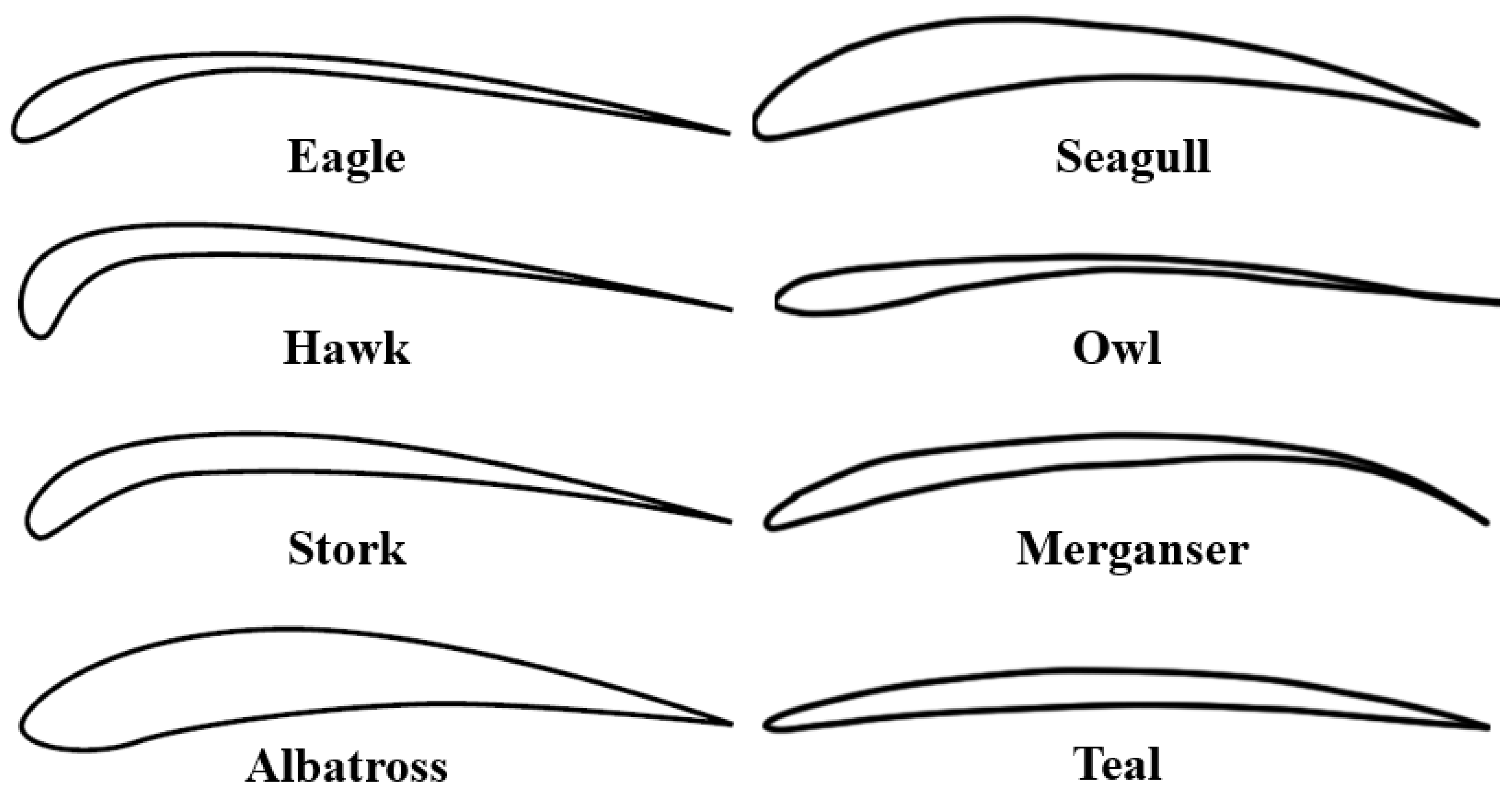
Figure 9.
a) Deflection of a pigeon’s alula [31]. b) Examples of hawk alula [217]. c) Example of an extended alula on and owl’s wing flying near the surface [233]. d) Schematic views of alula tip vortices interacting with a wing [217]. e) Application of an alula device is attached through an adjustable connector [217]. f) Schematic view of a wing alula [233]. g-i) Additional examples of avian alula on a bald eagle and hawk [219]. j) Schematic of fluxes induced by an alula over a fat plate [224]. All figures are reprinted with permission; please see the Appendix A.
Figure 9.
a) Deflection of a pigeon’s alula [31]. b) Examples of hawk alula [217]. c) Example of an extended alula on and owl’s wing flying near the surface [233]. d) Schematic views of alula tip vortices interacting with a wing [217]. e) Application of an alula device is attached through an adjustable connector [217]. f) Schematic view of a wing alula [233]. g-i) Additional examples of avian alula on a bald eagle and hawk [219]. j) Schematic of fluxes induced by an alula over a fat plate [224]. All figures are reprinted with permission; please see the Appendix A.
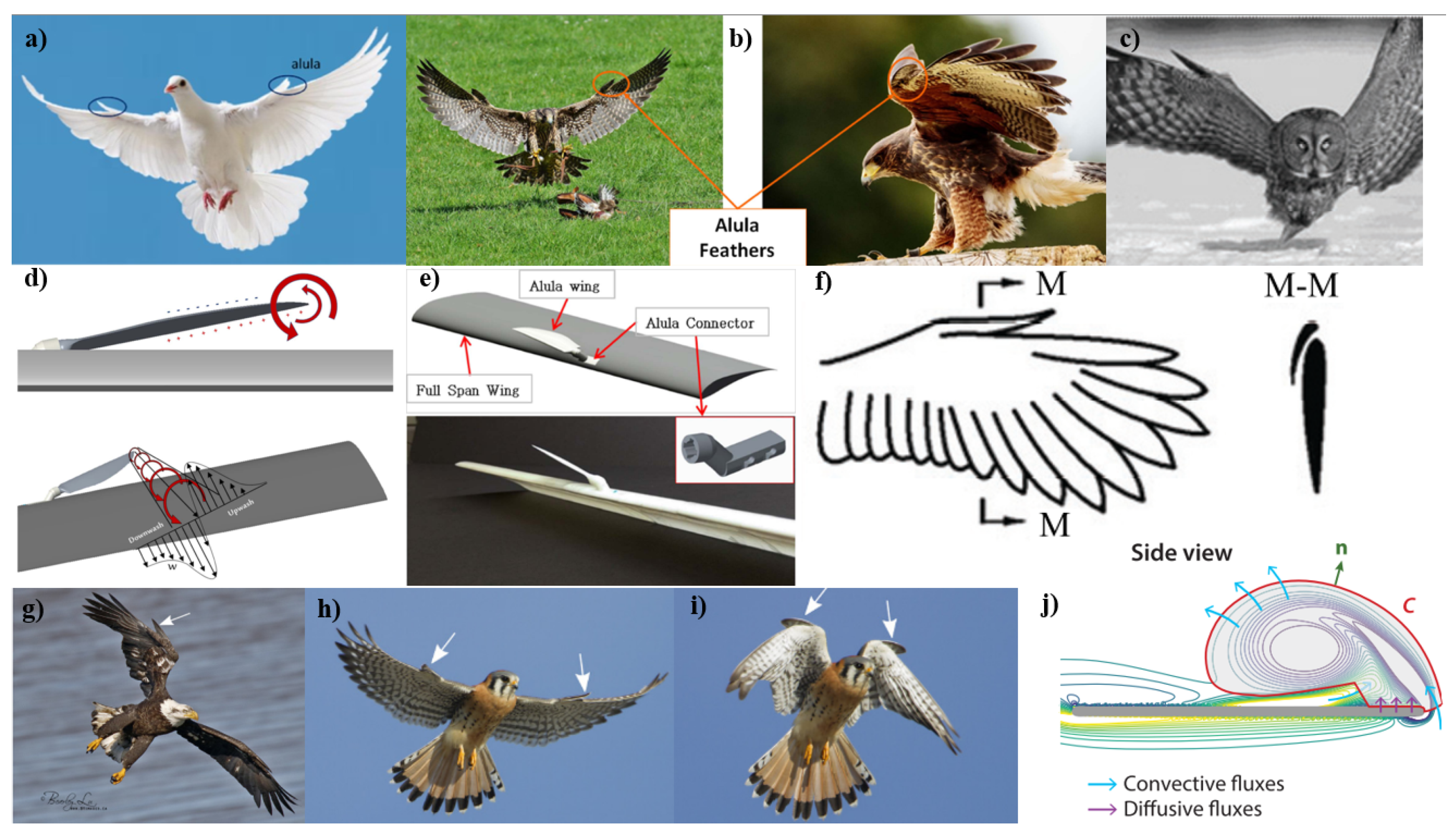
Figure 10.
ClarkY and s1223 airfoils used as reference technical airfoils in studies by Carruthers and Lang [37,73].
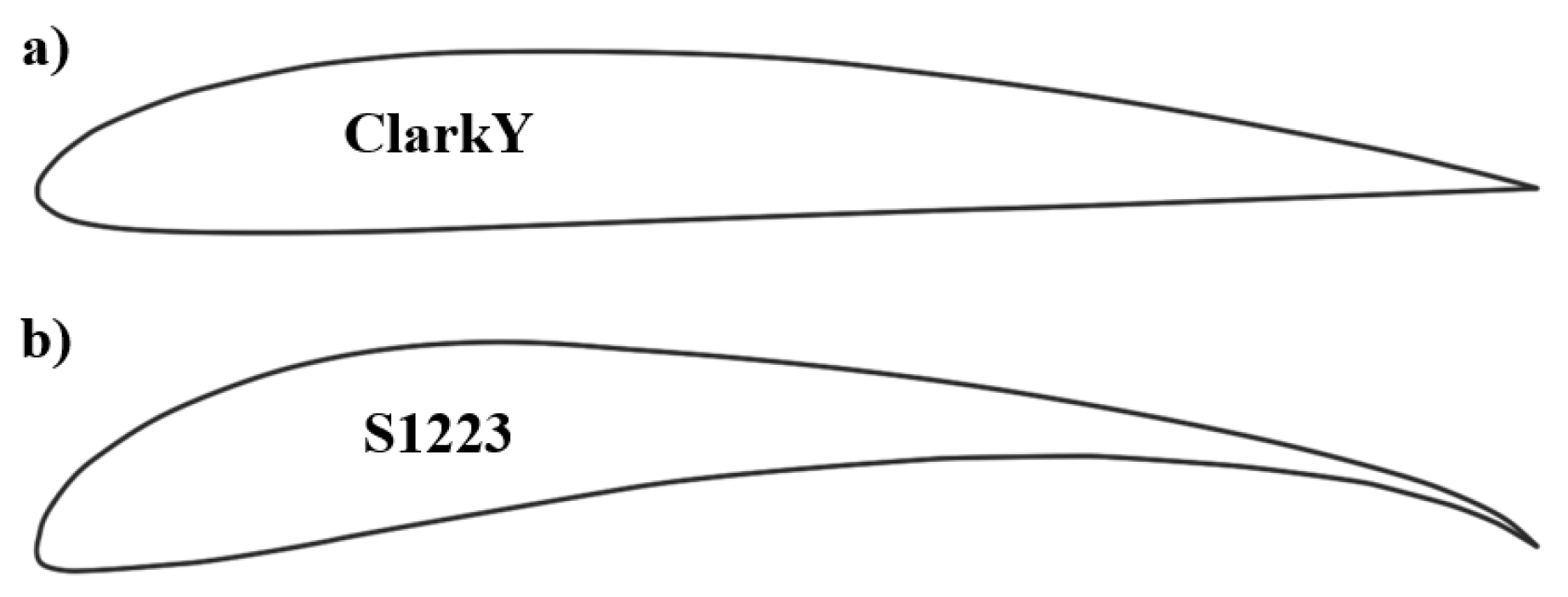
Figure 11.
Examples of various wingtip devices and designs: a) vortex turbine [103], b & c) multi-tip wingtips [90], d) spiroid winglet [247], e) controllable wingtip [248], f) three element winglet [197], g-h) streamlines around a two and three part winglet [197]. All figures are reprinted with permission; please see the Appendix A.
Figure 11.
Examples of various wingtip devices and designs: a) vortex turbine [103], b & c) multi-tip wingtips [90], d) spiroid winglet [247], e) controllable wingtip [248], f) three element winglet [197], g-h) streamlines around a two and three part winglet [197]. All figures are reprinted with permission; please see the Appendix A.
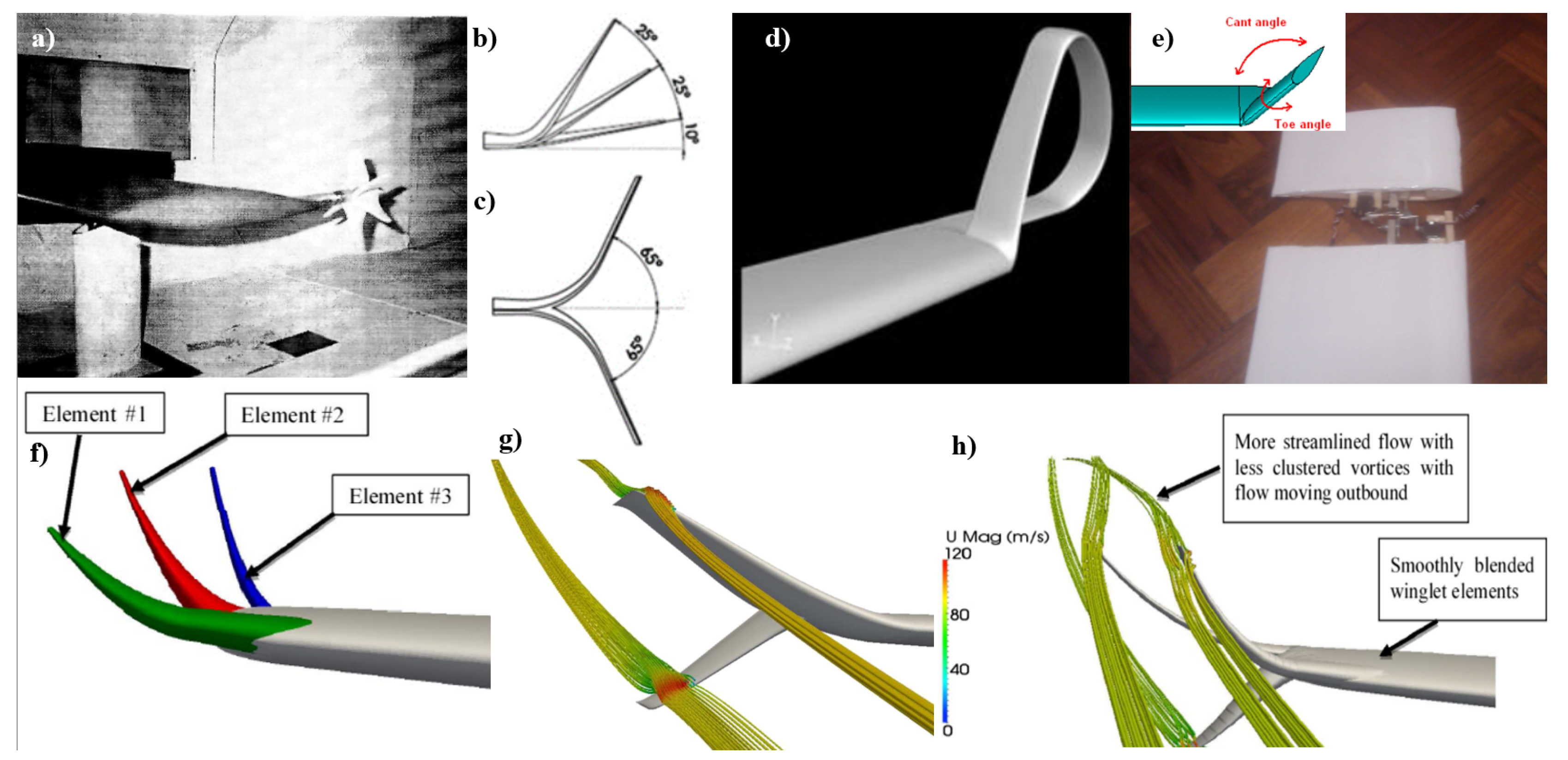
Figure 13.
Various examples of wing porosity experiments and molting. a) Schematic view of an airfoil surface with impermeable and permeable sections [23]. b) Sketch of the wind tunnel setup used to measure the effects of permeable wings and airfoils [23]. c) Example instantaneous pressure field around a flat plate with a permeable section [29]. d) CAD schematic of an experimental porous wing [55]. e) L/D curve comparing a solid vs. porous wing [55]. f) Instantaneous dye stream images for varying AOA (left solid, right porous) highlighting the changes in the flow field [55]. g) Schematic of an airfoil with porous media inserted along the span [69]. h) Time averaged velocity field comparing varying media density with a solid airfoil [69]. i) Top view of 3D scanned Anna’s hummingbird wings used in Achache’s molting experiments [21]. j) Example of a red kite wing in with deformed feathers in the molting process. Image Credit: Helen von Holten [60]. All figures are reprinted with permission; please see the Appendix A.
Figure 13.
Various examples of wing porosity experiments and molting. a) Schematic view of an airfoil surface with impermeable and permeable sections [23]. b) Sketch of the wind tunnel setup used to measure the effects of permeable wings and airfoils [23]. c) Example instantaneous pressure field around a flat plate with a permeable section [29]. d) CAD schematic of an experimental porous wing [55]. e) L/D curve comparing a solid vs. porous wing [55]. f) Instantaneous dye stream images for varying AOA (left solid, right porous) highlighting the changes in the flow field [55]. g) Schematic of an airfoil with porous media inserted along the span [69]. h) Time averaged velocity field comparing varying media density with a solid airfoil [69]. i) Top view of 3D scanned Anna’s hummingbird wings used in Achache’s molting experiments [21]. j) Example of a red kite wing in with deformed feathers in the molting process. Image Credit: Helen von Holten [60]. All figures are reprinted with permission; please see the Appendix A.
Figure 15.
Examples of avian color patterns used for mating purposes. a) Vogelkop Bird-of-Paradise. Photo credit: Tim Laman; Cornell Lab of Ornithology | Macaulay Library [260]. b) Victoria’s Riflebird. Photo credit: Jill Duncan and Ken Bissett; Cornell Lab of Ornithology | Macaulay Library [261]. c) Grater Sage-Grouse. Photo credit: Wolfe R; Cornell Lab of Ornithology | Macaulay Library [262] d) Wilson’s Bird-of-Paradise. Photo credit: Chris Venetz; Cornell Lab of Ornithology | Macaulay Library [263]. e) Crimson-backed Tanager. Photo credit: Yeray Seminario; Cornell Lab of Ornithology | Macaulay Library [264]. f) Resplendent Quetzal. Photo credit: Pascal De Munck; Cornell Lab of Ornithology | Macaulay Library [265]. All figures are reprinted with permission; please see the Appendix A.
Figure 15.
Examples of avian color patterns used for mating purposes. a) Vogelkop Bird-of-Paradise. Photo credit: Tim Laman; Cornell Lab of Ornithology | Macaulay Library [260]. b) Victoria’s Riflebird. Photo credit: Jill Duncan and Ken Bissett; Cornell Lab of Ornithology | Macaulay Library [261]. c) Grater Sage-Grouse. Photo credit: Wolfe R; Cornell Lab of Ornithology | Macaulay Library [262] d) Wilson’s Bird-of-Paradise. Photo credit: Chris Venetz; Cornell Lab of Ornithology | Macaulay Library [263]. e) Crimson-backed Tanager. Photo credit: Yeray Seminario; Cornell Lab of Ornithology | Macaulay Library [264]. f) Resplendent Quetzal. Photo credit: Pascal De Munck; Cornell Lab of Ornithology | Macaulay Library [265]. All figures are reprinted with permission; please see the Appendix A.

Figure 16.
Example of birds using unique color patterns for camouflage purposes. a) Rose-ringed Parakeet. Photo credit: Donald Hobern. b) American Bittern. Photo credit: Dan. e) Western Screech-Owl. Photo credit: Baker County Tourism. d) Double-banded Sandgrouse (Pterocles bicinctus) female. Photo credit: Bernard DuPont. e) Great Horned Owl. Photo credit: Mark Gunn. All figures are reprinted with permission; please see the Appendix A.
Figure 16.
Example of birds using unique color patterns for camouflage purposes. a) Rose-ringed Parakeet. Photo credit: Donald Hobern. b) American Bittern. Photo credit: Dan. e) Western Screech-Owl. Photo credit: Baker County Tourism. d) Double-banded Sandgrouse (Pterocles bicinctus) female. Photo credit: Bernard DuPont. e) Great Horned Owl. Photo credit: Mark Gunn. All figures are reprinted with permission; please see the Appendix A.

Figure 17.
Several examples of black and white bird species: (a) alpine chough, (b) whooper swan, (c) avocets, (d) yellow-billed magpie, (e) pied crow, (f) magpie-lark, (g) black-and-white-casqued hornbill, (h) black-headed ibis, (i) albatross, (j) manx shearwater, (k) black skimmer, (l) white stork, (m) swallow-tailed kite, (n) bearded vulture, (o) bar-headed goose, (p) common crane, (q) Andean condor, (r) rüppell’s vulture, (s) snow goose, (t) whooping crane, (u) Indian paradise flycatcher, (v) white-browed wagtail, (w) hoopoe, and (x) black and white warbler [20].
Figure 17.
Several examples of black and white bird species: (a) alpine chough, (b) whooper swan, (c) avocets, (d) yellow-billed magpie, (e) pied crow, (f) magpie-lark, (g) black-and-white-casqued hornbill, (h) black-headed ibis, (i) albatross, (j) manx shearwater, (k) black skimmer, (l) white stork, (m) swallow-tailed kite, (n) bearded vulture, (o) bar-headed goose, (p) common crane, (q) Andean condor, (r) rüppell’s vulture, (s) snow goose, (t) whooping crane, (u) Indian paradise flycatcher, (v) white-browed wagtail, (w) hoopoe, and (x) black and white warbler [20].

Figure 18.
Observed black and white coloration pattern examples for birds’ wings [20].
Figure 18.
Observed black and white coloration pattern examples for birds’ wings [20].
Figure 20.
a) Diagram providing an example of intermittent flap-bounding and flap-gliding flight. During intermittent bounds, the wings are held tight against the body, while during intermittent glides they are extended. [277]. b & c) Example schematic of wing and body kinematics measured for a zebra finish, showing the wingtip elevation at the start and end of the downstroke, and the dorsal view of the wings and body posture during flap-bounding [278]. d) Kinematics and muscle activity during bounding flight of a zebra finch in a wind tunnel. The bounds are highlighted in gray, while the rest is flapping flight [277,279]. e) Schematic view and PIV analysis displaying the trailing vortices formed during the fully flexed wing phase of bounding flight [277]. f) A chart displaying the wing posture during non-flapping flight as a function of flight speed [277,279]. g) Position of a finch’s eye measured during a bounding cycle used to calculate the body lift and drag [278]. h) Estimated mean power consumption during bounding flight for various values of the flapping ratio [241]. All figures are reprinted with permission; please see the Appendix A.
Figure 20.
a) Diagram providing an example of intermittent flap-bounding and flap-gliding flight. During intermittent bounds, the wings are held tight against the body, while during intermittent glides they are extended. [277]. b & c) Example schematic of wing and body kinematics measured for a zebra finish, showing the wingtip elevation at the start and end of the downstroke, and the dorsal view of the wings and body posture during flap-bounding [278]. d) Kinematics and muscle activity during bounding flight of a zebra finch in a wind tunnel. The bounds are highlighted in gray, while the rest is flapping flight [277,279]. e) Schematic view and PIV analysis displaying the trailing vortices formed during the fully flexed wing phase of bounding flight [277]. f) A chart displaying the wing posture during non-flapping flight as a function of flight speed [277,279]. g) Position of a finch’s eye measured during a bounding cycle used to calculate the body lift and drag [278]. h) Estimated mean power consumption during bounding flight for various values of the flapping ratio [241]. All figures are reprinted with permission; please see the Appendix A.
Figure 21.
This figure shows several example photos of various bird species performing bounding flight during “standard” flight and prior to landing. a) View of a great spotted woodpecker engaging in bounding flight [281]. b & c) Views of a fieldfare in its version of bounding flight [281]. d) Another view of a great spotted woodpecker engaging in bounding flight [281]. e) View of a green woodpecker in bounding flight [281]. f) An orange-crowned warbler exiting bounding flight to land [282]. g) A hooded oriole in bounding flight [282]. h) View of a small fieldfare flock with 2 members performing bounding flight [281]. i) View of a green jay exiting bounding flight prior to landing [282]. j) A cardinal exiting bounding flight to land [282]. k) View of a golden-fronted woodpecker performing bounding flight to land [282]. All figures are reprinted with permission; please see the Appendix A.
Figure 21.
This figure shows several example photos of various bird species performing bounding flight during “standard” flight and prior to landing. a) View of a great spotted woodpecker engaging in bounding flight [281]. b & c) Views of a fieldfare in its version of bounding flight [281]. d) Another view of a great spotted woodpecker engaging in bounding flight [281]. e) View of a green woodpecker in bounding flight [281]. f) An orange-crowned warbler exiting bounding flight to land [282]. g) A hooded oriole in bounding flight [282]. h) View of a small fieldfare flock with 2 members performing bounding flight [281]. i) View of a green jay exiting bounding flight prior to landing [282]. j) A cardinal exiting bounding flight to land [282]. k) View of a golden-fronted woodpecker performing bounding flight to land [282]. All figures are reprinted with permission; please see the Appendix A.

Figure 28.
View of ground effect employed by (a) a shearwater and (b) an amphibious drone [332].
Figure 28.
View of ground effect employed by (a) a shearwater and (b) an amphibious drone [332].
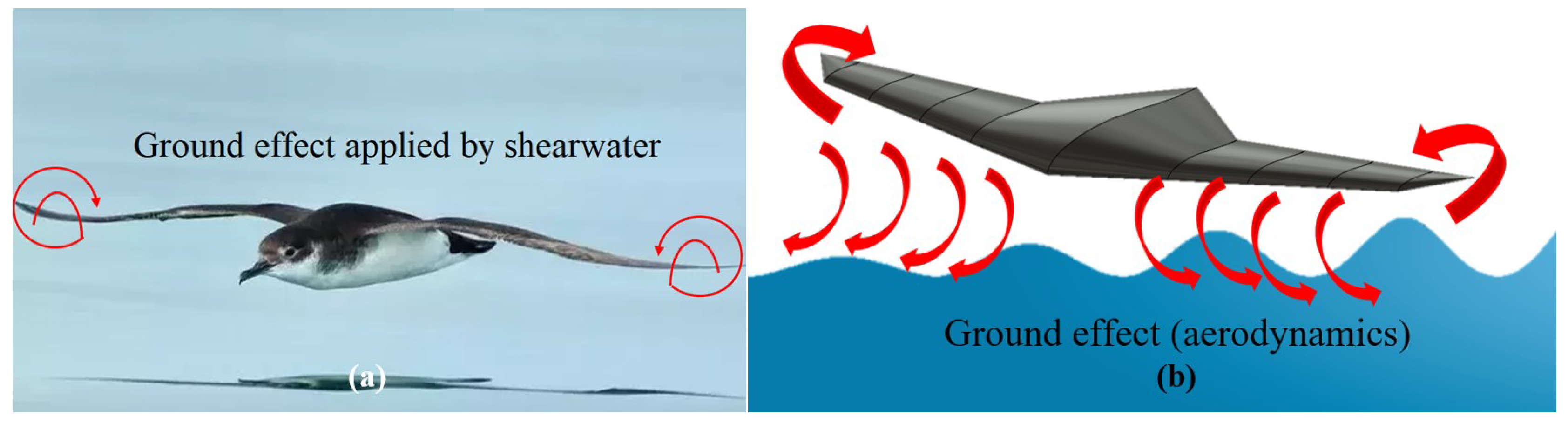
Figure 39.
a)Example of a cloud-to-ground lightning bolt and the different lines or energy formed by the stepped leader of the lightning. The red circles indicate two lower-energy lines of the lightning bolt. b) An ionized plane designed by MIT researchers [396]. c) Example of the distinct types of lightning [397]. d) Electrical circuit model of the cloud charges [398]. e) Example of a potential static charge harvesting system through formation flight of UAVs.
Figure 39.
a)Example of a cloud-to-ground lightning bolt and the different lines or energy formed by the stepped leader of the lightning. The red circles indicate two lower-energy lines of the lightning bolt. b) An ionized plane designed by MIT researchers [396]. c) Example of the distinct types of lightning [397]. d) Electrical circuit model of the cloud charges [398]. e) Example of a potential static charge harvesting system through formation flight of UAVs.
Table 1.
A comparison table summarizing the aerodynamic lift performance of wings with and without bird feather-like winglets [63].
Table 1.
A comparison table summarizing the aerodynamic lift performance of wings with and without bird feather-like winglets [63].
| Lift Coefficient | Angle of attack | |||||
|---|---|---|---|---|---|---|
| Winglet Configuration | Reynolds Number (10⌃5) | 0 | 4 | 8 | 12 | 14 |
| 1.66 | 0.228 | 0.594 | 0.804 | 0.725 | 0.666 | |
| Without winglet | 2.08 | 0.256 | 0.605 | 0.787 | 0.721 | 0.589 |
| 2.5 | 0.308 | 0.677 | 0.88 | 0.797 | 0.735 | |
| 1.66 | 0.405 | 0.678 | 0.85 | 0.68 | 0.572 | |
| Winglet in horizontal (0 degree) | 2.08 | 0.433 | 0.668 | 0.915 | 0.81 | 0.722 |
| 2.5 | 0.414 | 0.725 | 0.972 | 0.83 | 0.759 | |
| 1.66 | 0.442 | 0.787 | 0.993 | 0.812 | 0.78 | |
| Winglet 60 degree inclined | 2.08 | 0.456 | 0.817 | 0.956 | 0.876 | 0.75 |
| 2.5 | 0.481 | 0.803 | 0.99 | 0.854 | 0.828 | |
Table 2.
A comparison table summarizing the aerodynamic drag performance of wings with and without bird feather-like winglets [63].
Table 2.
A comparison table summarizing the aerodynamic drag performance of wings with and without bird feather-like winglets [63].
| Drag Coefficient | Angle of attack | |||||
|---|---|---|---|---|---|---|
| Winglet Configuration | Reynolds Number (10) | 0 | 4 | 8 | 12 | 14 |
| 1.66 | 0.088 | 0.105 | 0.156 | 0.216 | 0.258 | |
| Without winglet | 2.08 | 0.0849 | 0.1029 | 0.152 | 0.236 | 0.289 |
| 2.5 | 0.0667 | 0.0878 | 0.136 | 0.191 | 0.218 | |
| 1.66 | 0.0616 | 0.065 | 0.103 | 0.175 | 0.193 | |
| Winglet in horizontal (0 degree) | 2.08 | 0.0546 | 0.0576 | 0.0936 | 0.145 | 0.164 |
| 2.5 | 0.0534 | 0.0554 | 0.085 | 0.117 | 0.131 | |
| 1.66 | 0.076 | 0.0935 | 0.118 | 0.16 | 0.193 | |
| Winglet 60 degree inclined | 2.08 | 0.0642 | 0.0907 | 0.104 | 0.152 | 0.171 |
| 2.5 | 0.052 | 0.071 | 0.091 | 0.139 | 0.159 | |
Table 3.
A comparison table summarizing the aerodynamic drag performance of wings with and without bird feather-like winglets [63].
Table 3.
A comparison table summarizing the aerodynamic drag performance of wings with and without bird feather-like winglets [63].
| Pitching moment coefficient | Angle of attack | |||||
|---|---|---|---|---|---|---|
| Winglet Configuration | Reynolds Number (10) | 0 | 4 | 8 | 12 | 14 |
| 1.66 | -0.086 | -0.098 | -0.12 | -0.32 | -0.373 | 0.258 |
| Without winglet | 2.08 | -0.091 | -0.125 | -0.197 | -0.425 | -0.502 |
| 2.5 | -0.113 | -0.143 | -0.27 | -0.55 | -0.643 | 0.218 |
| 1.66 | -0.294 | -0.373 | -0.524 | -1.075 | -1.12 | 0.193 |
| Winglet in horizontal (0 degree) | 2.08 | -0.319 | -0.404 | -0.577 | -0.983 | -1.1 |
| 2.5 | -0.353 | -0.424 | -0.613 | -1.045 | -1.207 | 0.131 |
| 1.66 | -0.385 | -0.486 | -0.686 | 1.207 | -1.47 | 0.193 |
| Winglet 60 degree inclined | 2.08 | -0.406 | -0.531 | -0.709 | -1.103 | -1.28 |
| 2.5 | -0.481 | -0.549 | -0.704 | -1.09 | -1.24 | 0.159 |
Table 4.
Extracted wing shapes and maximum range of motion for 61 species of bird, adapted from [30].
Table 4.
Extracted wing shapes and maximum range of motion for 61 species of bird, adapted from [30].
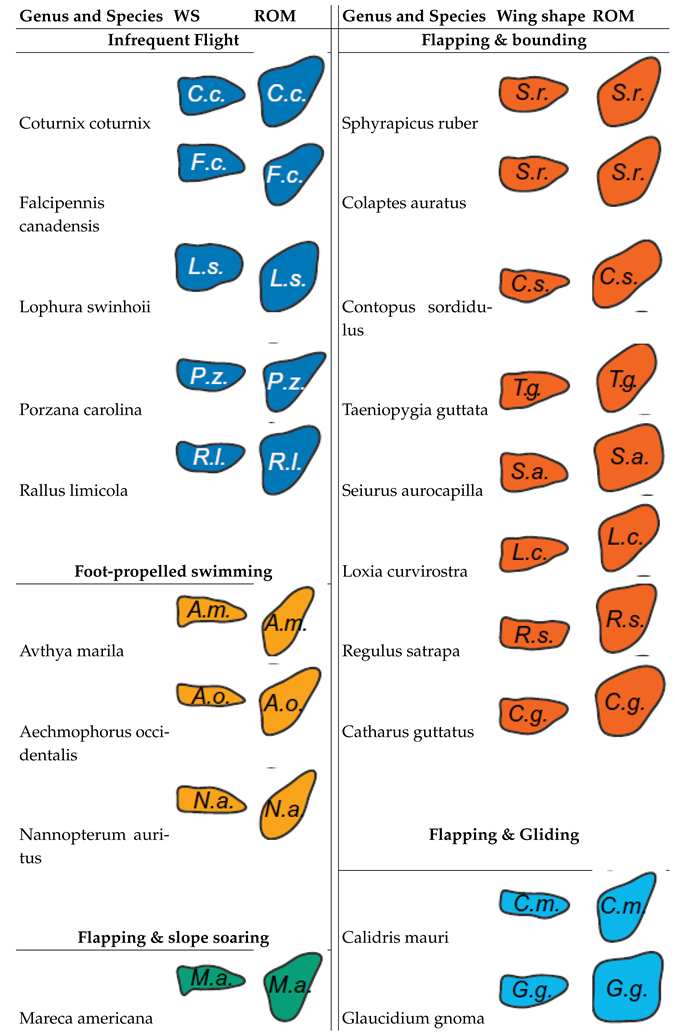 |
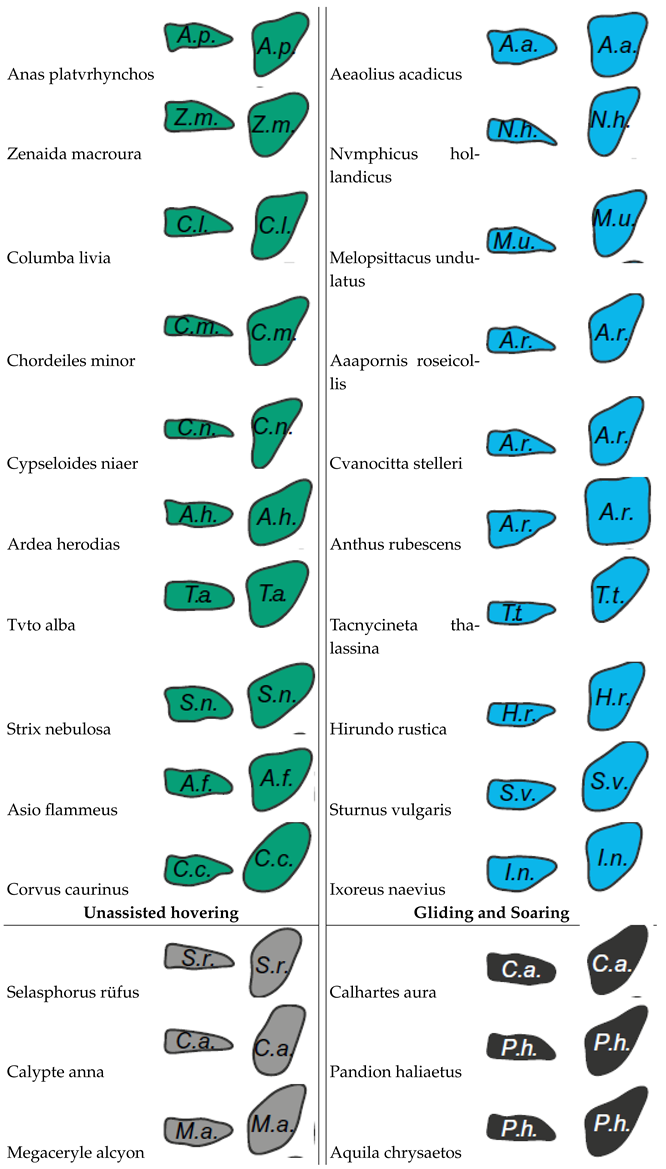 |
 |
Table 6.
A summary table collected various techniques for energy savings, drag reduction, and energy harvesting. The table highlights the key application spaces, as well as what area of an aircraft is of concern, and a brief description of the impacts.
Table 6.
A summary table collected various techniques for energy savings, drag reduction, and energy harvesting. The table highlights the key application spaces, as well as what area of an aircraft is of concern, and a brief description of the impacts.
| Method | Aircraft Type | Component | Impact on Efficiency/Flight Paths/Decisions | Drag Component Addressed | Adapted from Bionics Studies |
| Morphing Wing Technologies | UAVs, MAVs, Commercial, Flapping-wing, Fixed-wing | Wings, Airfoil Design | Dynamic adjustments to optimize aerodynamic efficiency and lead to a higher lift-to-drag ratio | Drag due to lift, Pressure drag | Partially |
| Wingtips and Winglets | UAVs, MAVs, Commercial, Flapping-wing, Fixed-wing | Wings, Tails | Significantly reduce induced drag, thereby improving the lift-to-drag ratio | Drag due to lift | Yes |
| Feather Riblet Structure | UAVs, MAVs, Commercial, Flapping-wing, Fixed-wing | External Surfaces | Can significantly reduce viscous drag, thereby improving the lift-to-drag ratio | Friction drag (laminar, turbulent) | Yes |
| Wing Porosity | UAVs, MAVs, Flapping-wing, Fixed-wing | Wings, Tails | Can increase lift and aerodynamic efficiency, particularly at certain angles of attack, improving landing performance | Pressure drag, Drag due to lift | Yes |
| Flexible Flaps | UAVs, MAVs, Fixed-wing, Flapping-wing | Wings | Enhances aerodynamic robustness, potentially beneficial for the design of flapping drones | Drag due to lift, Pressure drag | No |
| Wing Tip Slots and Emargination | UAVs, MAVs, Commercial, Fixed-wing, Flapping-wing | Wingtips | Reduces induced drag, increases agility and aerodynamic efficiency | Drag due to lift | Yes |
| Alula/ Leading-edge Devices | UAVs, MAVs, Commercial, Fixed-wing, Flapping-wing | Leading Edge of Wings | Increases maximum lift coefficient, enhances stall angles | Drag due to lift | Yes |
| Airfoil Optimization | UAVs, MAVs, Commercial, Fixed-wing, Flapping-wing | Airfoils, Wings | Tailored for different flight behaviors and environmental conditions, optimizing aerodynamics | Friction drag (laminar, turbulent), Pressure drag | No |
| Bio-inspired Flapping Kinematics | Flapping-wing | Wings | Superior lift characteristics, more energy-efficient flapping motion | Drag due to lift | Yes |
| Aerodynamic Advantages of Flexible Flaps | UAVs, Fixed-wing drones | Flaps on Wings | Reduces force fluctuations, enhances lift-to-drag ratios | Drag due to lift, Pressure drag | No |
| Corrugation Patterns | UAVs, MAVs, Commercial, Fixed-wing drones, Flapping-wing | Wings, Tails | Improves lift-to-drag ratio, manages flow separation | Friction drag (laminar, turbulent), Pressure drag | Yes |
| Body Shapes | UAVs, MAVs, Commercial, Flapping-wing, Fixed-wing | Body/ Fuselage | Optimizing body shape for drag reduction can lead to significant improvements in flight efficiency | Pressure drag | Partially |
| Beak Shape | UAVs, MAVs, Commercial, Flapping-wing, Fixed-wing | Nose | Optimizing nose shape for drag reduction can lead to significant improvements in flight efficiency | Pressure drag | No |
| Color | UAVs, MAVs, Commercial, Flapping-wing, Fixed-wing | External Surfaces | Can reduce drag and improve lift by changing the thermal gradient based on color pattern | Friction drag | No |
| Bounding Flight | MAVs, Flapping-wing | Flight Mode Control | Can increase energy efficiency and maneuverability at higher speeds | Drag due to lift | No |
| Hovering Flight | UAVs, MAVs, Flapping-wing | Flight Mode Control | Allows aircraft to remain in a fixed position for observation or interaction without landing, but generally does not increase efficiency | Drag due to lift | No |
| Formation Flight | Multi-agent Systems, Swarming Drones, Flapping-wing, Fixed-wing | Flight Mode Control, Flight Formation Communication | Reduces drag, leads to fuel savings, but can cause instabilities in flight | Drag due to lift | Yes |
| Leader Switching | Multi-agent Systems, Swarming Drones, Flapping-wing, Fixed-wing | Flight Mode Control, Flight Formation Communication | Balances energy expenditure, leading to extended flight range and endurance | Drag due to lift | No |
| Time of Flight | UAVs, MAVs, Flapping-wing, Fixed-wing | Environmental Monitoring, Autonomous Navigation Systems | Aligns migration with favorable conditions; optimizes energy use and survival | Drag due to lift | No |
| Flight Route Optimization | UAVs, MAVs, Commercial, Flapping-wing, Fixed-wing | Environmental Monitoring, Autonomous Flight Path Design | Utilizes wind patterns for energy efficiency; adapts to meteorological changes | Drag due to lift | No |
| Altitude Adjustment | UAVs, MAVs, Commercial, Flapping-wing, Fixed-wing | Flight Path Optimization | Exploits environmental conditions for energy savings; adapts to meteorological variations | Drag due to lift | No |
| Wing Scales/Microstructures for Drag Reduction | UAVs, MAVs | External surfaces, Particularly Wing Surfaces | Reduces drag through surface texture modifications | Friction drag | Yes |
| Light manipulating Microstructures | UAVs, MAVs, Commercial, Flapping-wing, Fixed-wing | Wings, Fuselage | Enhances light absorption, potentially improving solar energy harvesting and aerodynamic efficiency | Friction drag | No |
| Electro Aerodynamic Propulsion | UAVs, MAVs, Fixed-wing | Propulsion/Wings, External Surfaces | Potential to provide an additional propulsion source without mechanical motion | Pressure drag | No |
| Solar Panel Integration | UAVs, MAVs, Commercial, Flapping-wing, Fixed-wing | Wings, Fuselage | Extends operational time and range, harnessing renewable energy for enhanced endurance | Friction drag, Pressure drag | No |
| Static Charge Harvesting | UAVs, MAVs, Flapping-wing, Fixed-wing | External Surfaces | Harvests electrostatic energy, potentially improving aerodynamic performance and energy harvesting from the environment | Friction drag | No |
| Bio-inspired Sensory Systems | UAVs, MAVs, Flapping-wing, Fixed-wing | Navigation and Control Systems | Utilizes wind turbulence for energy, enhancing MAV flight performance | Friction drag | No |
| Ground Effect Exploitation | UAVs, MAVs, Commercial Aircraft | Close to Ground Operations | Reduces drag, enhances lift near the ground | Drag due to lift | Yes |
| Thermal Soaring | UAVs, MAVs, Flapping-wing, Fixed-wing | Utilizing Thermal Updrafts | Extends flight duration without power | Drag due to lift | No |
| Slope Soaring | UAVs, MAVs, Flapping-wing, Fixed-wing | Exploiting Wind against Terrain | Permits sustained flight by utilizing orographic lift | Drag due to lift | No |
| Dynamic Soaring | UAVs, MAVs, Flapping-wing, Fixed-wing | Open Air Operations | Utilizes wind gradient for energy gain | Drag due to lift | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



