
Submitted:
21 October 2025
Posted:
22 October 2025
You are already at the latest version
Abstract
Understanding how microbial communities from hyperarid environments respond to sporadic water availability is crucial for predicting ecosystem responses to unpredicta-ble precipitation related to climate change. Here, we experimentally simulated a short-term precipitation pulse (50 mm) in soils from the hyperarid Atacama Desert to evaluate shifts in microbial community composition, diversity, and functional potential, as well as changes in soil physicochemical properties. Using a microcosm experiment with time-structured sampling and high-throughput amplicon sequencing, we found that changes in soil properties after precipitation do not significantly influence micro-bial community composition. However, carbon dynamics exhibit significant variation. Actinobacteria and Proteobacteria dominated bacterial communities, exhibiting func-tional stability over time, which suggests high resilience. In contrast, fungal communi-ties, primarily composed of Ascomycota, displayed greater temporal variation in both taxonomic and functional profiles, particularly during intermediate hydration stages. Notably, fungal functional richness increased despite a limited taxonomic change, in-dicating lower redundancy and, thus, potentially higher community fragility. Our findings highlight distinct microbial strategies for coping with hydration pulses in ex-treme drylands, underscoring the role of fungi in nutrient cycling, decomposition, and soil structure, while also revealing their increased vulnerability to environmental fluc-tuations. Our results reinforce the importance of microbial dynamics in maintaining ecosystem functioning in pulse-reserve systems under ongoing climatic changes.
Keywords:
atacama desert
; microbial communities
; pulse precipitation
; functional resilience
1. Introduction
Drylands cover over 45% of Earth’s terrestrial surface [4], which harbors a third of the global human population [5]. Although previous studies have focused on the impact of current climate change trends, which are globally increasing the total dryland surface [6], the vulnerability of these habitats to unpredictable rainfall events remains understudied [7].
The pulse-reserve paradigm (PRP) explains how dryland ecosystems respond to infrequent rainfall, with episodic water pulses triggering bursts of biological activity [8,9,10]. Microbial growth in arid environments is driven by reserves—mainly carbon—stored during previous periods of moisture. During rainfall pulses, microorganisms use these reserves to grow, reproduce, and acquire new resources, replenishing what is needed to survive dry phases [10,11]. As moisture declines, microbes transition into dormancy, relying on accumulated reserves for survival [12]. The duration and intensity of these pulses strongly influence microbial dynamics: shorter pulses reduce growth efficiency and alter community composition, while longer pulses support net growth and effective reserve replenishment [11,12,13].
The short-term effects of a hydration pulse on desert soil microbial communities are significant and multifaceted, influencing microbial diversity, community composition, and metabolic activity. Following a hydration pulse, there is often a decrease in microbial diversity [8,11,12]. These patterns are attributed to the rapid growth of specific taxa that thrive in wet conditions, which can outcompete others during these brief periods of moisture availability. For instance, specialist phylotypes may become more prominent in response to specific pulse durations [12]. The composition of microbial communities undergoes significant shifts after hydration events [12,14]. Studies have shown that distinct microbial communities emerge under varying moisture conditions—one during high hydration, another during gradual desiccation, and a third in arid conditions [12]. These shifts reflect the community’s adaptation to fluctuating moisture levels. Hydration pulses trigger a surge in microbial metabolic activity, leading to increased respiration rates and CO2 release. This metabolic activation is crucial for nutrient cycling and can enhance overall soil fertility temporarily [14]. However, this activity can rapidly deplete resources if not replenished [12,13]. Microbial diversity often returns to pre-pulse levels as soils dry, a recovery essential for maintaining ecosystem resilience [11,15].
The Atacama Desert in northern Chile is the most arid desert on the planet and one of the oldest [16]. Here, the mean annual precipitation is only a few millimeters (< 2 mm), with rainfall events occurring approximately every 10-20 years [17,18]. In the core of the Atacama Desert, biological activity is triggered by rare precipitation events following prolonged drought, as exemplified by the “blooming desert” phenomenon [11,19]. During rainy periods, years that generally involve the El Niño phenomenon [20,21,22], increased water availability can trigger flowering desert events caused by a rapid biological response by microorganisms and dormant vegetation in the soil [20,21]. These increases in above and belowground productivity trigger new trophic cascades, altering the structure and functioning of this ecosystem [18,20,21].
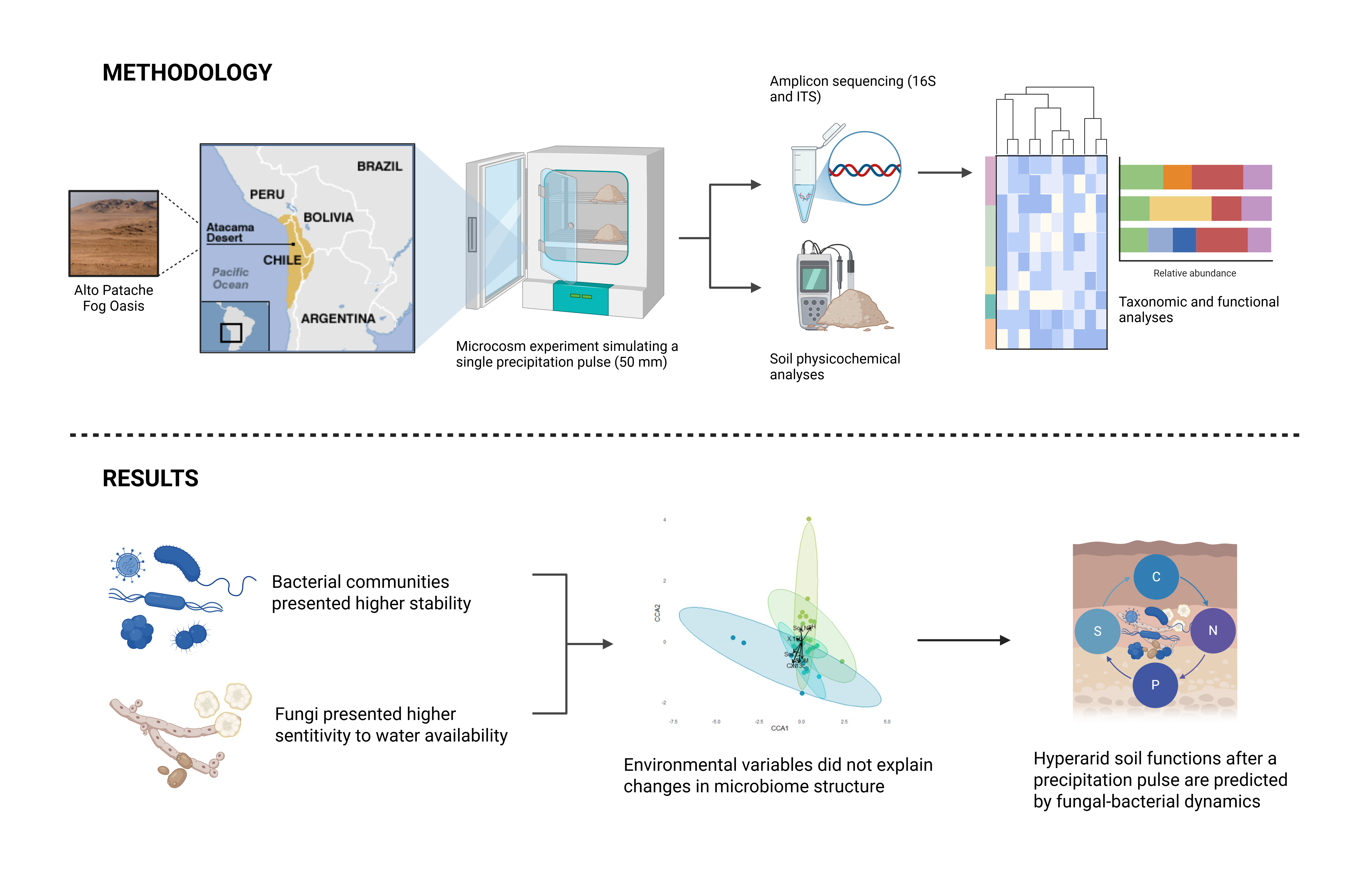
In this study, we experimentally evaluated the short-term effects of an intense precipitation pulse on the composition, taxonomic, and functional diversity of soil microbial communities in one of the world’s driest ecosystems. Specifically, we aimed to assess how a single hydration event - simulating the magnitude of the extraordinary 2015 rainfall in the Atacama Desert - alters microbial diversity, taxonomic structure, and the relative dominance of major microbial groups. Focusing on community responses across bacteria and fungi, we aimed to identify taxon-specific changes and patterns of resilience that reflect ecological strategies for coping with extreme moisture fluctuations in hyper-arid soils and infer how these changes in taxonomic diversity are related to functions.
2. Materials and Methods
2.1. Study Site
The Atacama UC Oasis Niebla Alto Patache Station (20° 49’ S - 70° 09’ W) is located in the Atacama Desert, Iquique, Tarapacá Region (Figure 1). The topographical characteristics of this area allow limited entry of coastal fog or “camanchaca” inland, maintaining a relative humidity of 30% with a mean annual precipitation of <1 mm [23]. The mean annual temperature during the day is 25°C and 10°C at night. The vertical soil profile consists of alternating layers of sand and clay, which prevent water from diffusing into deeper layers, which in turn makes the soil virtually devoid of vegetation cover [24].
2.2. Soil Sampling
We collected composite soil samples (five subsamples) of topsoil (~5cm depth) using sterilized spatulas. Larger gravel and organic materials were removed from the samples by sieving them through a 2 mm mesh and then stored in plastic bags at room temperature. During the microcosm experiment, we divided the samples into two fractions. The first fraction was immediately frozen (−20 °C) for molecular analyses, and the second was air-dried for physicochemical analyses. We measured abiotic properties for each sample. Soil pH was determined using a pH sensor in a CaCl2 solution [25]. Soil organic matter (SOM) content was assessed by ignition [26]. Soil water holding capacity (WHC) was measured using gravimetric methods, employing 5 g of soil, a filter, and 10 ml of water, and calculating the percentage of water retained relative to the soil weight [27]. Gravimetric methods also determined moisture content in soil (SM) [25].
2.3. Microcosm Experiment
To evaluate the short-term effects of flooding pulses on soil properties and the structure of soil microbial communities, thirty pots containing 50 g of soil samples were flooded to simulate a 50mm-rainfall event, based on the last major flooding event in the Atacama Desert (2015), then incubated for 7 days in two culture chambers (111L CLIMACELL® Incubator, MMM Group) simulating the summer conditions of the Atacama Desert, with a mean temperature of 18°C, relative humidity of 30% and a photoperiod of 14 hours of light and 10 hours of darkness. Six microcosms were sacrificed just before the start of the experiment (time 0), and then at 12, 24, 96, and 168 hours after incubation.
2.4. DNA Extraction and Amplicon Sequencing
DNA extraction was performed using the DNeasy® PowerSoil® Pro Kit (Qiagen), following the manufacturer’s protocol, and DNA concentration was determined using a Qubit™ 4.0 fluorimeter (ThermoFisher Scientific). The 16S ribosomal RNA gene, targeting the V3-V4 region, and the ITS ribosomal RNA were sequenced at Zymo Research. Primers for sequencing were provided by the same company, and sequencing was performed using the Illumina® MiSeq™ platform. Zymo Research conducted the taxonomic assignment of fungi and bacteria using UCLUST in QIIME v.1.9.1, referencing their proprietary database. The absolute abundance of Amplicon Sequence Variants (ASVs) was determined by Zymo Research using qPCR, with a standard curve for both the 16S and ITS genes. We standardized sampling effort by applying rarefaction without replacement in both domains. Bacterial libraries ranged from 145 to 2545 reads per sample; all samples were rarefied to 145 reads (the minimum). Fungal libraries ranged from 159 to 60641 reads per sample; all samples were rarefied to 159 reads. Thresholds were selected to maximize sample retention across time points. After rarefaction, the bacterial dataset retained six replicates at all time points, except at 12 h, which retained five. In the fungal dataset, rarefaction reduced replication: the first three timepoints retained five replicates each, whereas the last two time points retained three replicates each.
2.5. Data Analysis
Microbial community analyses and biodiversity indices were assessed using the ‘microeco’ package [28] in R. To evaluate microbial community composition at the phylum level, we determined the relative abundance of major soil phyla at each time point and generated bar plots. For genus-level analyses, we used heatmaps, grouping genera by phyla. To assess the relationship between diversity indices and physicochemical factors over time, we performed a series of linear models (LM) where diversity metrics (i.e., Shannon, Chao1, Simpson) and soil physicochemical variables were treated as response variables and sampling time (0, 12, 24, 96, and 168 h) as a fixed factor. To determine the relationship between environmental factors and microbial communities, we performed a Canonical Correspondence Analysis (CCA) with a Hellinger transformation using the R package vegan [29]. Additionally, microbial functional composition was assessed by classifying genera using FUNGuild [30] for fungal functional groups and Tax4Fun2 [31] for bacterial functions. To compare microbial biodiversity and functional composition, we performed regression analyses between taxonomic richness and functional diversity, calculating the adjusted R2 and corresponding p-values. Statistical analyses were conducted using the R statistical software [32].
3. Results
3.1. Biogeochemical Changes During Short Pulse Precipitation
Soil physicochemical properties exhibited significant changes in response to increased water availability. Specifically, a significant increase was observed in the proportion of the δ13C isotope, soil carbon content (%C), and soil organic matter (SOM) throughout the experiment. Concurrently, soil pH progressively decreased, indicating a shift toward more neutral conditions compared to the initial measurements. In contrast, neither soil nitrogen content (%N) nor the proportion of the δ15N isotope showed significant changes over time (Figure 2).
We then evaluated whether these changes in soil properties influence soil bacterial and fungal community composition over time. In the case of bacteria, the first two dimensions (CCA1 and CCA2) account for 20% and 16.3% of the variance, respectively (Figure S1a). For fungi, the first and second axes of CCA account for 24.4% and 17.2% of the variance, respectively (Figure S1b). The analyses showed that, together, the environmental variables did not significantly explain the changes in microbial community composition for either group (global ANOVA CCA for bacteria: p = 0.495; for fungi: p = 0.300) (Table S1). Furthermore, none of the measured physicochemical variables individually showed a significant effect on bacterial or fungal composition, except for Soil C, which was significant for fungi (p = 0.015), but not for bacteria (p = 0.088) (Table S1). On the other hand, the PERMAVONA analysis based on Bray-Curtis’s dissimilarity index revealed that time had a clear and significant effect on both bacterial (R2 = 22.98%, p = 0.001) and fungal (R2 = 33.45%, p = 0.001) (Table S1) community structures.
3.2. The Impact of Pulse Precipitation on Soil Microbial Abundances and Diversity
The most dominant bacterial phyla were Actinobacteria (61.42%), Chloroflexi (10.77%), Proteobacteria (10.36%) and Firmicutes (6.82%) (Figure 3a), and their abundances varied over time after the water event. For example, Actinobacteria abundances decreased with time, while Proteobacteria showed an opposite trend (Figure 3a). At a more detailed taxonomic level, the most abundant bacterial genera across treatments were Rubrobacter, Gaiella, and Pseudonocardia, from the Actinobacteriota phylum. Additionally, some genera, such as Ammoniphilus (Firmicutes) and Gemmatimonas (Gemmatimonadota), increase in abundance during the later stages of this immediate response (Figure 3b). Bacterial alpha-diversity remains relatively high and stable, with no significant differences over time (Figure 3c).
The fungal communities were highly dominated by members of the Ascomycota (82.03%) phylum, followed by Basidiomycota (14.16%), Mortierellomycota (1.16%), Rozellomycota (1.13%), and Chytridiomycota (0.14%). However, no fungal phyla displayed any temporal trends (Figure 4a). At the genus level (Figure 4b), Alternaria (Ascomycota) stands out as one of the most abundant genera (representing 20.73% of the communities); particularly at 12 h (54%) and 24 h (44%) after the water pulse. In contrast, Fusarium and Oidiodendrum (Ascomycota) were more abundant at the beginning (31% and 33% at 0 h, respectively), and Fusarium and Colletotrichum were more abundant at the end of the experiment (33% and 21% at 168 h, respectively). Across the full experiment, Fusarium accounted for 13.0%, Colletotrichum for 4.5%, and Oidiodendrum for 6.1% of the total fungal community. Fungal diversity remains stable, though increasing toward the later stages, showing a significant positive trend in the Chao1 index (Figure 4c), suggesting this domain has a higher sensitivity to environmental changes.
3.3. The Impact of Pulse Precipitation on Soil Microbial Composition and Function
The ecological functions associated with bacteria remained stable over time, with aerobic chemoheterotrophy being the most frequently observed function (Figure 5a). Additionally, a less frequent but also stable function is nitrate reduction. In the case of fungi, a greater number of ecological functions were observed, the most frequent including roles as plant pathogens and saprotrophs (Figure 5b), particularly related to the presence of Alternaria in the intermediate stages (12 and 24 hours) and Fusarium, though less dominant, in the early and late stages (0 and 168 hours) (Figure S2). The main fungal lifestyles are as soil and aquatic fungi, which abundances change with water availability (Figure 5b).
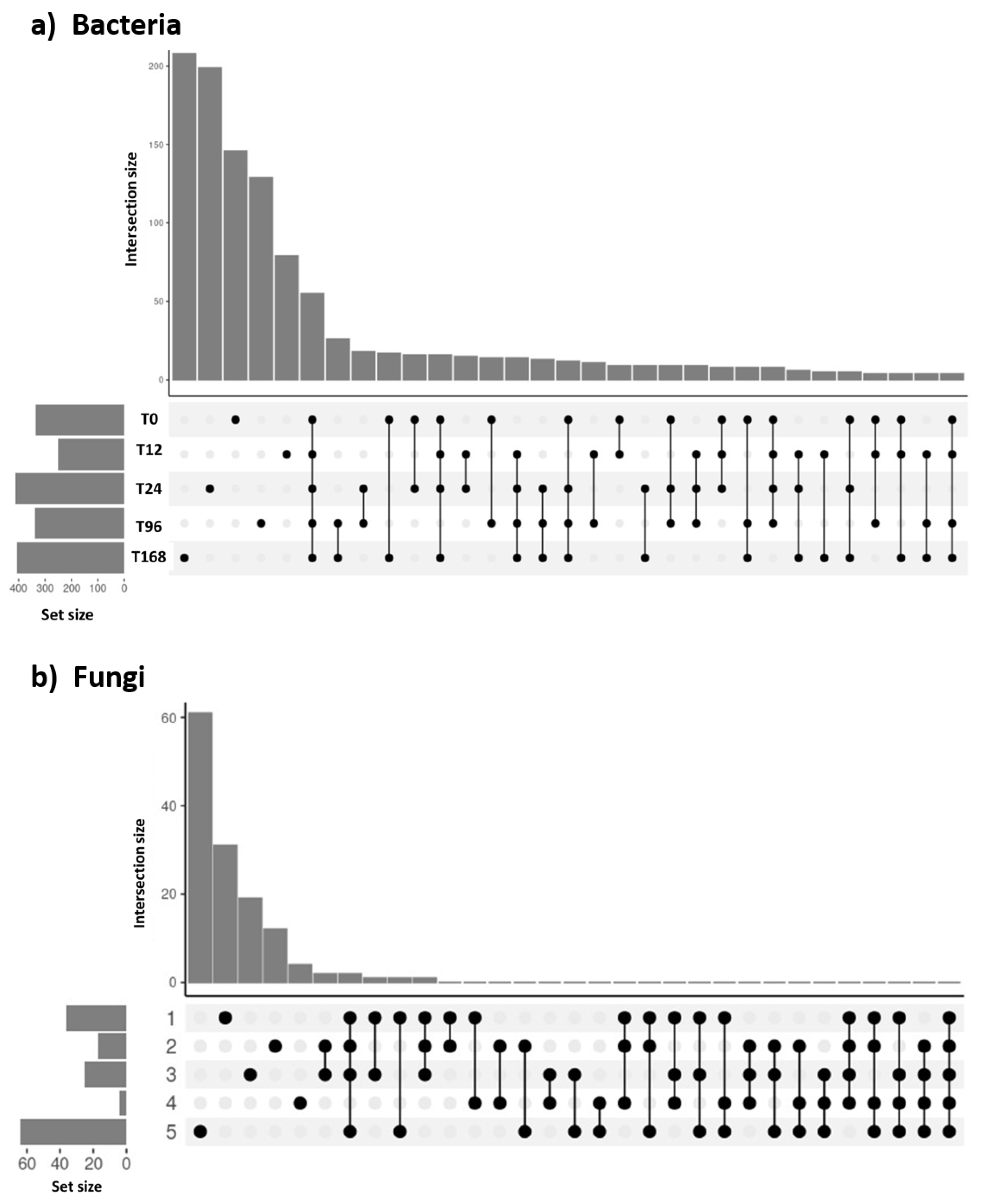
The bacterial community composition over time revealed that early-stage communi-ties harbored fewer unique species, while species tend to become more specific in the later stages (Figure 6a). In contrast, fungal community composition exhibited the opposite trend, with more specific fungal species found in the initial stages and less specificity observed as time progressed (Figure 6b), except at 168 hours, which shows the most unique composition among timepoints.
To assess a relative functional stability in these microbial groups, we evaluated the relationship between functional and taxonomic richness. Functional richness in bacterial communities does not correlate with taxonomic richness, as both metrics remained stable over time (Figure S3a). However, we observed a sharp increase in fungal community functional richness while taxonomic richness remained much lower in comparison (Figure S3b).
4. Discussion
Short-term precipitation pulses in hyperarid soils can generate rapid and multidimensional responses in both abiotic and biotic components, despite the extreme water limitations that typically constrain microbial activity [33]. In this study, we demonstrate that even a brief hydration event can trigger shifts in soil chemical properties, particularly in carbon dynamics, alongside changes in microbial composition and functional potential. These changes were not immediate or linear but rather reflected a mosaic of transient responses driven by microbial physiological traits and ecological interactions. Our results support the notion that microbial communities in pulse-reserve systems exhibit differential strategies for coping with water pulses, with bacteria maintaining stable functions through metabolic versatility and a possible functional asynchrony, while fungi respond with greater compositional specificity and functional shifts.
The fungi-bacteria ratio in desert soils can shift markedly during hydration pulses, driven by microbial community interactions and resource availability [15]. Initially, bacterial activity increases due to sudden water and nutrient inputs, often outpacing fungal growth in the immediate response [12,13]. Moreover, mycelium-forming fungi can facilitate bacterial growth by redistributing water and nutrients within the soil, creating microhabitats that enhance bacterial survival and proliferation [34], therefore favoring bacterial dominance over fungi immediately after a pulse. Subsequently, as moisture levels stabilize or decrease, fungi may regain prominence due to their ability to form resilient structures, such as spores and hyphae, that can withstand dry conditions [15]. Both groups, bacteria and fungi, display physiological flexibility to alternate between active and passive phases, a key survival pattern in extreme arid environments. These findings suggest that short water pulses can reset soil microbial trajectories and influence key ecosystem functions in desert environments.
An increase in the δ13C isotope, soil carbon content (%C), and soil organic matter (SOM) (Figure 2) could be due to the Birch effect, where the decomposition of previously protected or newly available labile organic carbon is accelerated with the rapid flush of microbial activity after a water pulse [35,36]. Microbes often preferentially metabolize organic compounds containing the lighter 12C isotope due to lower energy requirements for breaking those bonds. This leaves behind a residual SOM pool that is relatively enriched in the heavier 13C isotope, thus increasing the overall δ13C value [37]. The decomposition processes activated by the precipitation pulse cause the release of various organic acids. Microbial respiration from increased metabolic activity during a precipitation pulse also produces carbon dioxide (CO2). When this CO2 dissolves in soil water, it forms carbonic acid (H2CO3), a weak acid that directly contributes hydrogen ions (H+) to the soil solution, thus lowering the pH [38]. However, the increased water flow can lead to the leaching of basic cations, especially calcium in the calcium carbonate-rich (CaCO3) Atacama Desert soils, from the upper soil layers [39]. The rate of pH decrease might be slowed due to the CaCO3 buffering capacity (Figure 2). Together with the production of CO2, it can influence the carbonate equilibrium, affecting soil pH in nuanced and complex ways, and possibly temporarily and/or locally lowering the pH [40,41].
In this context, bacteria tend to have greater adaptability than fungi to changes in environmental conditions [36,42]. They are therefore able to form specialized communities as soil conditions change during the pulse [12,33]. Fungi form highly specialized ecological networks but with less immediate plasticity and probably struggle to adapt to the abrupt input of water [43,44,45]. In this sense, the interactions between fungal genera and between fungi and bacteria become essential to produce balanced ecosystems [43]. For instance, fungi like Alternaria [46] and Oidiodendrum [47] can influence bacterial populations through their different decomposition activities. This could also help modulate carbon dynamics in the soil, as seen in Figure 2.
Ascomycetes are versatile fungi capable of decomposing a variety of organic materials, including dead microbial biomass and organic matter derived from other sources [47]. Their ability to break down complex compounds and their saprophytic activity are crucial for nutrient cycling in nutrient-poor soils [48,49]. At the genus level, Alternaria stands out as one of the most abundant genera, particularly in the intermediate stages (12 and 24 hours), while Fusarium, Colletotrichum and Oidiodendrum are more abundant at both earlier and later stages (Figure 4b). Alternaria species likely thrive on decomposing organic matter from microbial sources or other detritus, helping to recycle nutrients back into the soil [46]. In fact, Alternaria contributed the most to saprotrophic functions (Figure S2). Genera like Fusarium and Oidiodendrum can also have these ecological functions by decomposing recalcitrant materials, enhancing soil fertility over time [47,50]. Fusarium and Alternaria dominate as endophytes, saprotrophs and plant pathogens. However, they seem to outcompete each other at different stages of the pulse, with the first strongly present in drier soil (0 and 168 hours), and the latter present in wetter stages (12 and 24 hours), hinting at a possible functional asynchrony (Figure S1).
In the case of bacteria, the Actinobacteria phylum is recognized for its ability to break down complex organic matter. The decrease in abundance could be due to increased competition from other groups and changes in nutrient availability [51] (Figure 3a). On the other hand, Proteobacteria is a highly diverse phylum that includes many metabolically versatile bacteria. Its increase in abundance could be due to their ability to rapidly colonize new niches, utilize a wide range of substrates, or form beneficial associations with other organisms [51,52] (Figure 2A). These patterns were also observed in other studies of desert soil microbiome stimulation by hydration events [19,53,54]. The most abundant bacterial genera, such as Rubrobacter, Gaiella and Pseudonocardia (Figure 3b), are often associated with dry environments, and are known for their role in carbon cycling and nutrient mobilization in harsh conditions [55]. Additionally, some genera, such as Ammoniphilus and Gemmatimonas, increase in abundance during the later stages of this immediate response, suggesting a shift towards bacteria that can utilize nitrogen sources or thrive under specific nutrient conditions that arise after initial hydration events [56]. This shift could reflect changes in soil chemistry or microbial interactions as communities mature.
Aerobic chemoheterotrophy and nitrate reduction are the most prominent bacterial functions, with the former being much more abundant than the latter, but both being stable over time (Figure 5a). The stability in bacterial functions suggests that these communities are well-adapted to their environment and consistently rely on organic carbon sources for energy and growth, primarily through the decomposition of available organic matter, which is critical in ecosystems with low to no vegetation. Versatility in nutrient uptake is crucial in extreme environments, especially in pulse-reserve systems, where conditions can abruptly change in an instant [10].
Although no plants are present in the studied soils, the presence of fungal functions as plant pathogens (Figure 5b) suggests that these fungi may still play a role in breaking down organic matter [57]. The presence of filamentous mycelium suggests that fungi are likely forming extensive networks, which enhance nutrient absorption and decomposition processes within the soil. Fungal mycelium can create microhabitats that support bacterial growth by providing moisture and nutrients, while bacteria can help decompose organic matter more efficiently when associated with fungal partners [34,43]. In this sense, the interactions between bacterial and fungal communities can enhance overall ecosystem functioning [58].
Certain functional groups in fungi exhibited changes over time. For example, the increase in the variety of decay substrates and the presence of aquatic fungi during the intermediate stages highlight a shift in fungal community dynamics as conditions evolve (Figure 4b). This indicates a response to changing moisture levels or organic matter availability. This, along with the simultaneous rise of fungi with filamentous mycelium, suggests that as microbial communities mature, they may begin to explore and exploit new niches or resources that become available due to changes in environmental conditions or interactions with other microorganisms [15].
The presence of these functions in this analysis, however, only indicates their potential, not actual functional activity, as these data are annotations associated with a specific taxonomic group. Nonetheless, the relationship between functional potential and microbial taxonomic composition can inform us about functional stability, which is particularly important in light of current climatic trends [59,60]. Altogether, the results suggest that fungal communities at each timepoint may be more specialized, less functionally stable, and possibly more fragile and sensitive to environmental conditions than bacterial communities (Figure 6b and Figure S3). This pattern has been observed in recent studies [61,62,63], where fungi resulted to have less taxonomic richness than bacterial communities, allowing them to form unique communities and highly specialized ecological networks for each niche, but consequently causing them to have less adaptability to changing environments.
The fact that we are learning how fragile fungal communities in arid and hyperarid soils are to these infrequent pulse precipitation events is of the highest importance in the face of climate change [64]. Fungi can produce a wide variety of extracellular enzymes that enable them to decompose various types of organic matter, regulating carbon and nutrient balance in the soil, an especially vital process in environments where organic matter is scarce [65]. Fungi also contribute to soil structure formation by creating networks of mycelium that bind soil particles together. This process enhances soil stability and improves water retention capabilities, which is critical in hyperarid regions where moisture is limited. Additionally, it creates habitats for other microorganisms, promoting biodiversity within the soil ecosystem [34,43,65]. Moreover, deserts are among the largest carbon sinks on the planet [66,67], making carbon management a crucial aspect of soil conservation. These results reinforce the crucial role of soil microbial communities in arid and hyperarid ecosystems, particularly in carbon and nitrogen cycling, as well as overall soil health.
In conclusion, soil microbial communities of the Atacama Desert exhibit dynamic responses to precipitation pulses. Bacterial communities, dominated by Actinobacteria and Proteobacteria, show temporal shifts in relative abundance patterns, with Proteobacteria increasing over time. Fungal communities, primarily composed of Ascomycota, exhibit greater variation, with shifts in many functional groups occurring in response to changes in water and nutrient availability during the pulse. Both bacteria and fungi play crucial roles in nutrient cycling, decomposition, and the formation of soil structure. However, fungal communities may be more sensitive to environmental changes. Understanding the intricate interactions between these microbial groups is crucial to comprehend the resilience and functioning of arid ecosystems, particularly in the face of climate change.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: CCA to assess the effect of environmental factors on community composition; Figure S2: Relative contribution of fungal genera to the main functional guilds; Figure S3: Relationship between functional richness and taxonomic richness; Table S1: Summary of Canonical Correspondence Analysis (CCA) and PERMANOVA results.
Author Contributions
Conceptualization, Marlene Manzano and Fernando Alfaro; Data curation, María del Pilar Fernandez-Murillo; Formal analysis, Antonia Beggs and María del Pilar Fernandez-Murillo; Funding acquisition, Fernando Alfaro; Investigation, Antonia Beggs, María del Pilar Fernandez-Murillo and Marlene Manzano; Methodology, Marlene Manzano and Fernando Alfaro; Project administration, Fernando Alfaro; Resources, Fernando Alfaro; Software, María del Pilar Fernandez-Murillo; Supervision, Fernando Alfaro; Validation, Antonia Beggs and María del Pilar Fernandez-Murillo; Visualization, Antonia Beggs and María del Pilar Fernandez-Murillo; Writing – original draft, Antonia Beggs; Writing – review & editing, Cameron Wagg and Jean-Baptiste Ramond. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by ANID (Chile), Fondecyt 1220358.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original data presented in the study are openly available in FigShare at 10.6084/m9.figshare.30290302.
Acknowledgments
Many thanks to Miguel Ángel Castellani for providing the helpful map of the study site. During the preparation of this manuscript, the authors used Gemini 2.5 for the purposes of rephrasing cited publications. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Milena Holmgren, Paul Stapp, Chris R. Dickman, Carlos Gracia, Sonia Graham, Julio R. Gutiérrez, Christine Hice, Fabián Jaksic, Douglas A. Kelt, Mike Letnic, Mauricio Lima, Bernat C. López, Peter L. Meserve, W Bryan Milstead, Gary A. Polis, M Andrea Previtali, Michael Richter, Santi Sabaté, Francisco A. Squeo. Extreme Climatic Events Shape Arid and Semiarid Ecosystems. Frontiers in Ecology and the. 2006;4(2):87–95. [CrossRef]
- Orlando J, Alfaro M, Bravo L, Guevara R, Carú M. Bacterial diversity and occurrence of ammonia-oxidizing bacteria in the Atacama Desert soil during a “desert bloom” event. Soil Biol Biochem. 2010;42(7):1183–8. [CrossRef]
- Wang X-B, Azarbad H, Leclerc L, Dozois J, Mukula E, Yergeau É. A drying-rewetting cycle imposes more important shifts on soil microbial communities than does reduced precipitation. mSystems. 2022;7(4):e0024722. [CrossRef]
- Prăvălie R. Drylands extent and environmental issues. A global approach. Earth Sci Rev. 2016;161:259–78. [CrossRef]
- Plaza C, Zaccone C, Sawicka K, Méndez AM, Tarquis A, Gascó G, et al. Soil resources and element stocks in drylands to face global issues. Sci Rep. 2018;8(1):13788. [CrossRef]
- Huang J, Yu H, Guan X, Wang G, Guo R. Accelerated dryland expansion under climate change. Nat Clim Chang. 2016;6(2):166–71. [CrossRef]
- Mirzabaev A, Annagylyjova J, Amirova I. Environmental degradation. In: The Aral Sea Ba-sin. Routledge; 2019. p. 67–85.
- Noy-Meir I. Desert ecosystems: Environment and producers. Annu Rev Ecol Syst. 1973;4(1):25–51. [CrossRef]
- Reynolds JF, Kemp PR, Ogle K, Fernández RJ. Modifying the ‘pulse–reserve’ paradigm for deserts of North America: precipitation pulses, soil water, and plant responses. Oecologia. 2004;141(2):194–210. [CrossRef]
- Collins SL, Sinsabaugh RL, Crenshaw C, Green L, Porras-Alfaro A, Stursova M, et al. Pulse dynamics and microbial processes in aridland ecosystems: Pulse dynamics in aridland soils. J Ecol. 2008;96(3):413–20. [CrossRef]
- Garcia-Pichel F, Sala O. Expanding the pulse–reserve paradigm to microorganisms on the basis of differential reserve management strategies. Bioscience. 2022;72(7):638–50. [CrossRef]
- Kut P, Garcia-Pichel F. Nimble vs. torpid responders to hydration pulse duration among soil microbes. Commun Biol. 2024;7(1):455. [CrossRef]
- Collins SL, Belnap J, Grimm NB, Rudgers JA, Dahm CN, D’Odorico P, et al. A multiscale, hierarchical model of pulse dynamics in arid-land ecosystems. Annu Rev Ecol Evol Syst. 2014;45(1):397–419. [CrossRef]
- Št’ovíček A, Azatyan A, Soares MIM, Gillor O. The impact of hydration and temperature on bacterial diversity in arid soil mesocosms. Front Microbiol. 2017;8:1078. [CrossRef]
- Šťovíček A, Kim M, Or D, Gillor O. Microbial community response to hydration-desiccation cycles in desert soil. Sci Rep. 2017;7:45735. [CrossRef]
- Dion P, Nautiyal CS, editors. Microbiology of extreme soils. 2008th ed. Berlin, Germany: Springer; 2014.
- Houston J, Hartley AJ. The central Andean west-slope rainshadow and its potential contribution to the origin of hyper-aridity in the Atacama Desert. Int J Climatol. 2003;23(12):1453–64. [CrossRef]
- Miller FP, Vandome AF, McBrewster J, editors. Climate of Chile. Alphascript Publishing; 2010.
- Leung PM, Bay SK, Meier DV, Chiri E, Cowan DA, Gillor O, et al. Energetic basis of microbial growth and persistence in desert ecosystems. mSystems. 2020;5(2). [CrossRef]
- Jaksic FM. Ecological effects of El Niño in terrestrial ecosystems of western South America. Ecography (Cop). 2001;24(3):241–50. [CrossRef]
- Araya JP, González M, Cardinale M, Schnell S, Stoll A. Microbiome dynamics associated with the Atacama flowering desert. Front Microbiol. 2020;10(3160). [CrossRef]
- Martínez-Harms J, Guerrero PC, Martínez-Harms MJ, Poblete N, González K, Stavenga DG, et al. Mechanisms of flower coloring and eco-evolutionary implications of massive blooming events in the Atacama Desert. Front Ecol Evol. 2022;10. [CrossRef]
- McKay CP, Friedmann EI, Gómez-Silva B, Cáceres-Villanueva L, Andersen DT, Landheim R. Temperature and moisture conditions for life in the extreme arid region of the Atacama desert: four years of observations including the El Niño of 1997-1998. Astrobiology. 2003 Summer;3(2):393–406. [CrossRef]
- García J-L, Lobos-Roco F, Schween JH, del Río C, Osses P, Vives R, et al. Climate and coastal low-cloud dynamic in the hyperarid Atacama fog Desert and the geographic distribution of Tillandsia landbeckii (Bromeliaceae) dune ecosystems. Osterr Bot Z. 2021;307(5). [CrossRef]
- Webster R. Soil sampling and methods of analysis - edited by M.r. carter & E.g. gregorich. Eur J Soil Sci. 2008;59(5):1010–1. [CrossRef]
- Eyherabide M, Rozas HS, Barbieri P, Echeverría H. Comparación de métodos para determinar carbono orgánico en suelo. 2014;32:13–9. https://www.scielo.org.ar/pdf/cds/v32n1/v32n1a02.pdf.
- Nelson JT, Adjuik TA, Moore EB, VanLoocke AD, Ramirez Reyes A, McDaniel MD. A simple, affordable, do-it-yourself method for measuring soil maximum water holding capacity. Commun Soil Sci Plant Anal. 2024;55(8):1190–204. [CrossRef]
- Liu C, Cui Y, Li X, Yao M. microeco: an R package for data mining in microbial community ecology. FEMS Microbiol Ecol. 2021;97(2). [CrossRef]
- Dixon P. VEGAN, a package of R functions for community ecology. J Veg Sci. 2003;14(6):927–30 [software]. [CrossRef]
- Nguyen NH, Song Z, Bates ST, Branco S, Tedersoo L, Menke J, et al. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016;20:241–8 [software]. [CrossRef]
- Wemheuer F, Taylor JA, Daniel R, Johnston E, Meinicke P, Thomas T, et al. Tax4Fun2: prediction of habitat-specific functional profiles and functional redundancy based on 16S rRNA gene sequences. Environ Microbiome. 2020;15(1):11 [software]. [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. 2025. https://www.R-project.org/.
- León-Sobrino C, Ramond J-B, Coclet C, Kapitango R-M, Maggs-Kölling G, Cowan DA. Temporal dynamics of microbial transcription in wetted hyperarid desert soils. FEMS Microbiol Ecol. 2024;100(3). [CrossRef]
- Worrich A, Stryhanyuk H, Musat N, König S, Banitz T, Centler F, et al. Mycelium-mediated transfer of water and nutrients stimulates bacterial activity in dry and oligotrophic environments. Nat Commun. 2017;8(1):15472. [CrossRef]
- Iovieno P, Bååth E. Effect of drying and rewetting on bacterial growth rates in soil: Rewetting and bacterial growth in soil. FEMS Microbiol Ecol. 2008;65(3):400–7. [CrossRef]
- Wang X-B, Azarbad H, Leclerc L, Dozois J, Mukula E, Yergeau É. A drying-rewetting cycle imposes more important shifts on soil microbial communities than does reduced precipitation. mSystems. 2022;7(4):e0024722. [CrossRef]
- Krüger N, Finn DR, Don A. Soil depth gradients of organic carbon-13 – A review on drivers and processes. Plant Soil. 2024;495(1–2):113–36. [CrossRef]
- Brady NC, Weil RR. Elements of the nature and properties of soils. 4th ed. Upper Saddle River, NJ: Pearson; 2018.
- Nawaz R, Parkpian P, Garivait H, Anurakpongsatorn P, DeLaune RD, Jugsujinda A. Impacts of acid rain on base cations, aluminum, and acidity development in highly weathered soils of Thailand. Commun Soil Sci Plant Anal. 2012;43(10):1382–400. [CrossRef]
- Ramnarine R, Wagner-Riddle C, Dunfield KE, Voroney RP. Contributions of carbonates to soil CO2 emissions. Can J Soil Sci. 2012;92(4):599–607. [CrossRef]
- Knapp WJ, Tipper ET. The efficacy of enhancing carbonate weathering for carbon dioxide sequestration. Front Clim. 2022;4(928215). [CrossRef]
- Wang S, Zuo X, Awada T, Medima-Roldán E, Feng K, Yue P, et al. Changes of soil bacterial and fungal community structure along a natural aridity gradient in desert grassland ecosystems, Inner Mongolia. Catena. 2021;205(105470):105470. [CrossRef]
- Boer W de, Folman LB, Summerbell RC, Boddy L. Living in a fungal world: impact of fungi on soil bacterial niche development. FEMS Microbiol Rev. 2005;29(4):795–811. [CrossRef]
- Fierer N, Jackson RB. The diversity and biogeography of soil bacterial communities. Proc Natl Acad Sci U S A. 2006;103(3):626–31. [CrossRef]
- Zhang S, Cao Z, Liu S, Hao Z, Zhang X, Sun G, et al. Soil fungal diversity, community structure, and network stability in the southwestern Tibetan Plateau. J Fungi (Basel). 2025;11(5). [CrossRef]
- Fernandes C, Casadevall A, Gonçalves T. Mechanisms of Alternaria pathogenesis in animals and plants. FEMS Microbiol Rev. 2023;47(6). [CrossRef]
- Kravchenko IK, Tikhonova EN, Baslerov RV, Kolganova TV. Cultivable fungal diversity in The Forest soil during litter degradation: Microcosm study at different temperature regimes. Journal of Agriculture and Environment. 2022;2(22). [CrossRef]
- Abdullah M. Al-Sadi, Badriya Al-Khatri, Abbas Nasehi, Muneera Al-Shihi, Issa H. Al-Mahmooli, Sajeewa S.N. Maharachchikumbura. High Fungal Diversity and Dominance by Ascomycota in Dam Reservoir Soils of Arid Climates. International Journal of Agriculture and Biology. 2025;19(4):682–8.
- Egidi E, Delgado-Baquerizo M, Plett JM, Wang J, Eldridge DJ, Bardgett RD, et al. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat Commun. 2019;10(1):2369. [CrossRef]
- Legeay J, Basiru S, Ziami A, Errafii K, Hijri M. Response of Alternaria and Fusarium species to low precipitation in a drought-tolerant plant in Morocco. Microb Ecol. 2024;87(1):127. [CrossRef]
- Crits-Christoph A, Robinson CK, Barnum T, Fricke WF, Davila AF, Jedynak B, et al. Colonization patterns of soil microbial communities in the Atacama Desert. Microbiome. 2013;1(1):28. [CrossRef]
- Nessner Kavamura V, Taketani RG, Lançoni MD, Andreote FD, Mendes R, Soares de Melo I. Water regime influences bulk soil and Rhizosphere of Cereus jamacaru bacterial communities in the Brazilian Caatinga biome. PLoS One. 2013;8(9):e73606. [CrossRef]
- Angel R, Conrad R. Elucidating the microbial resuscitation cascade in biological soil crusts following a simulated rain event: Microbial resuscitation in biological soil crusts. Environ Microbiol. 2013;15(10):2799–815. [CrossRef]
- Jordaan K, Lappan R, Dong X, Aitkenhead IJ, Bay SK, Chiri E, et al. Hydrogen-oxidizing bacteria are abundant in desert soils and strongly stimulated by hydration. mSystems. 2020;5(6). [CrossRef]
- Demergasso C, Neilson JW, Tebes-Cayo C, Véliz R, Ayma D, Laubitz D, et al. Hyperarid soil microbial community response to simulated rainfall. Front Microbiol. 2023;14:1202266. [CrossRef]
- Ortega R, Miralles I, Domene MA, Meca D, Del Moral F. Ecological practices increase soil fertility and microbial diversity under intensive farming. Sci Total Environ. 2024;954(176777):176777. [CrossRef]
- 57Li Z, Liu X, Zhang M, Xing F. Plant diversity and fungal richness regulate the changes in soil multifunctionality in a semi-arid grassland. Biology (Basel). 2022;11(6):870. [CrossRef]
- Jiao S, Peng Z, Qi J, Gao J, Wei G. Linking bacterial-fungal relationships to microbial diversity and soil nutrient cycling. mSystems. 2021;6(2). [CrossRef]
- Wagg C, Schlaeppi K, Banerjee S, Kuramae EE, van der Heijden MGA. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. Nat Commun. 2019;10(1):4841. [CrossRef]
- Biggs CR, Yeager LA, Bolser DG, Bonsell C, Dichiera AM, Hou Z, et al. Does functional redundancy affect ecological stability and resilience? A review and meta-analysis. Ecosphere. 2020;11(7). [CrossRef]
- Chen Y, Xi J, Xiao M, Wang S, Chen W, Liu F, et al. Soil fungal communities show more specificity than bacteria for plant species composition in a temperate forest in China. BMC Microbiol. 2022;22(1):208. [CrossRef]
- Fu F, Li Y, Zhang B, Zhu S, Guo L, Li J, et al. Differences in soil microbial community structure and assembly processes under warming and cooling conditions in an alpine forest ecosystem. Sci Total Environ. 2024;907(167809):167809. [CrossRef]
- Li J, Yang H, Duan YY, Dan Sun X, Pang XP, Guo ZG. Fungi contribute more than bacteria to the ecological uniqueness of soil microbial communities in alpine meadows. Glob Ecol Conserv. 2024;55(e03246):e03246. [CrossRef]
- Cuartero J, Querejeta JI, Prieto I, Frey B, Alguacil MM. Warming and rainfall reduction alter soil microbial diversity and co-occurrence networks and enhance pathogenic fungi in dryland soils. Sci Total Environ. 2024;949(175006):175006. [CrossRef]
- Frąc M, Hannula SE, Bełka M, Jędryczka M. Fungal biodiversity and their role in soil health. Front Microbiol. 2018;9:707. [CrossRef]
- Millennium Ecosystem Assessment (MEA). Ecosystems and human well-being: desertification synthesis. World Resources Institute, Washington, DC. 2005. https://www.millenniumassessment.org/documents/document.355.aspx.pdf.
- Liu Z, Wang G. Concept and methodology for Scope 4 carbon emission accounting. Carbonsphere. 2025;1(9510002). [CrossRef]
Figure 1.
Map showing the location of the study site and sampling area, in the Atacama UC Oasis Niebla Alto Patache Station. Coordinates are in UTM.
Figure 1.
Map showing the location of the study site and sampling area, in the Atacama UC Oasis Niebla Alto Patache Station. Coordinates are in UTM.
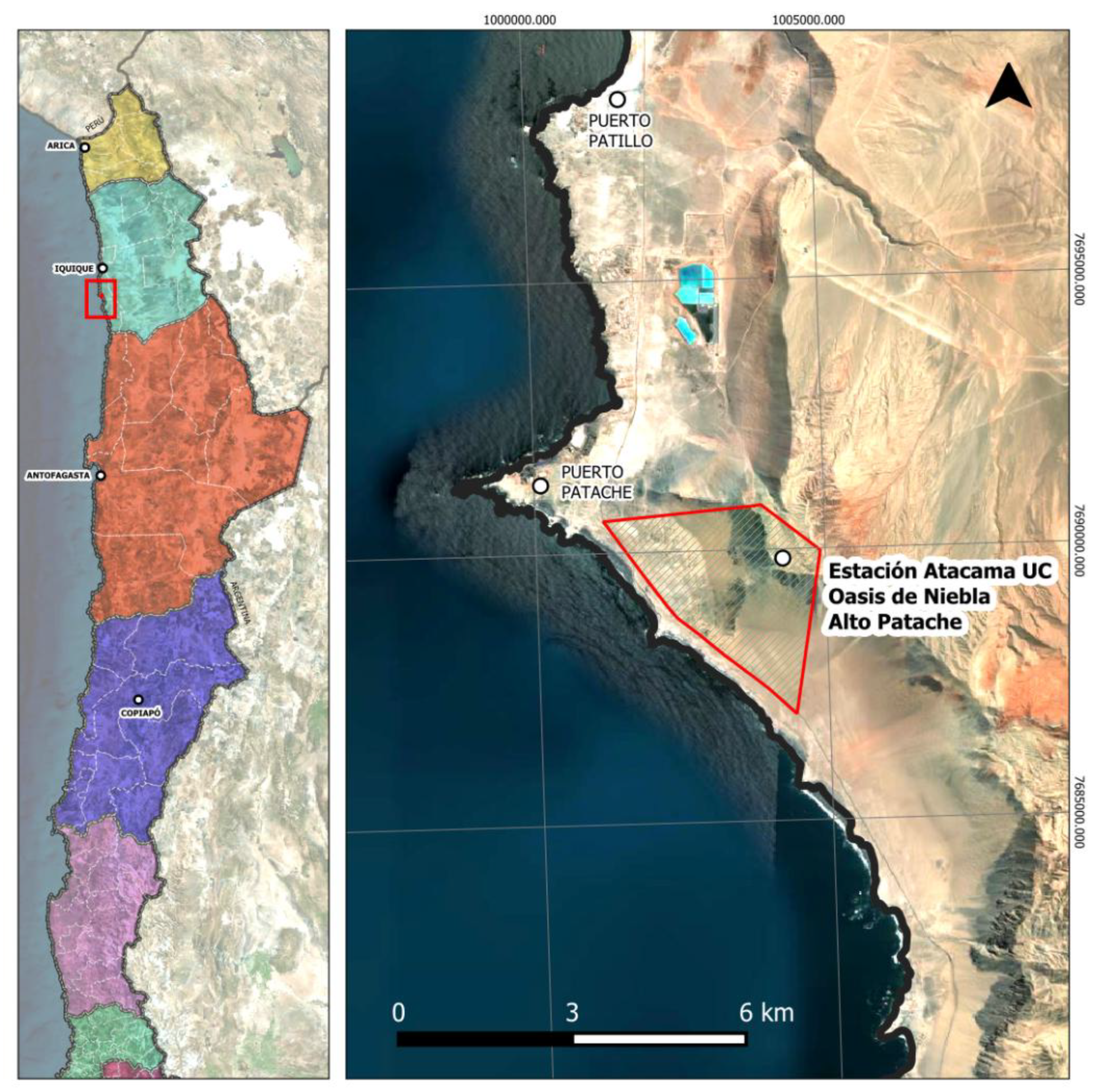
Figure 2.
Changes in soil physicochemical properties in response to increased water availability. Ordinary Least Squares regressions illustrates the distribution of values and linear trends for δ15N, δ13C, nitrogen content, carbon content, pH, and soil organic matter across five time points (0, 12, 24, 96, and 168 hours). The diversity panel includes the adjusted R2 and P values from linear model tests for temporal differences. Statistical significance was considered at P < 0.05, indicated with a continuous line.
Figure 2.
Changes in soil physicochemical properties in response to increased water availability. Ordinary Least Squares regressions illustrates the distribution of values and linear trends for δ15N, δ13C, nitrogen content, carbon content, pH, and soil organic matter across five time points (0, 12, 24, 96, and 168 hours). The diversity panel includes the adjusted R2 and P values from linear model tests for temporal differences. Statistical significance was considered at P < 0.05, indicated with a continuous line.
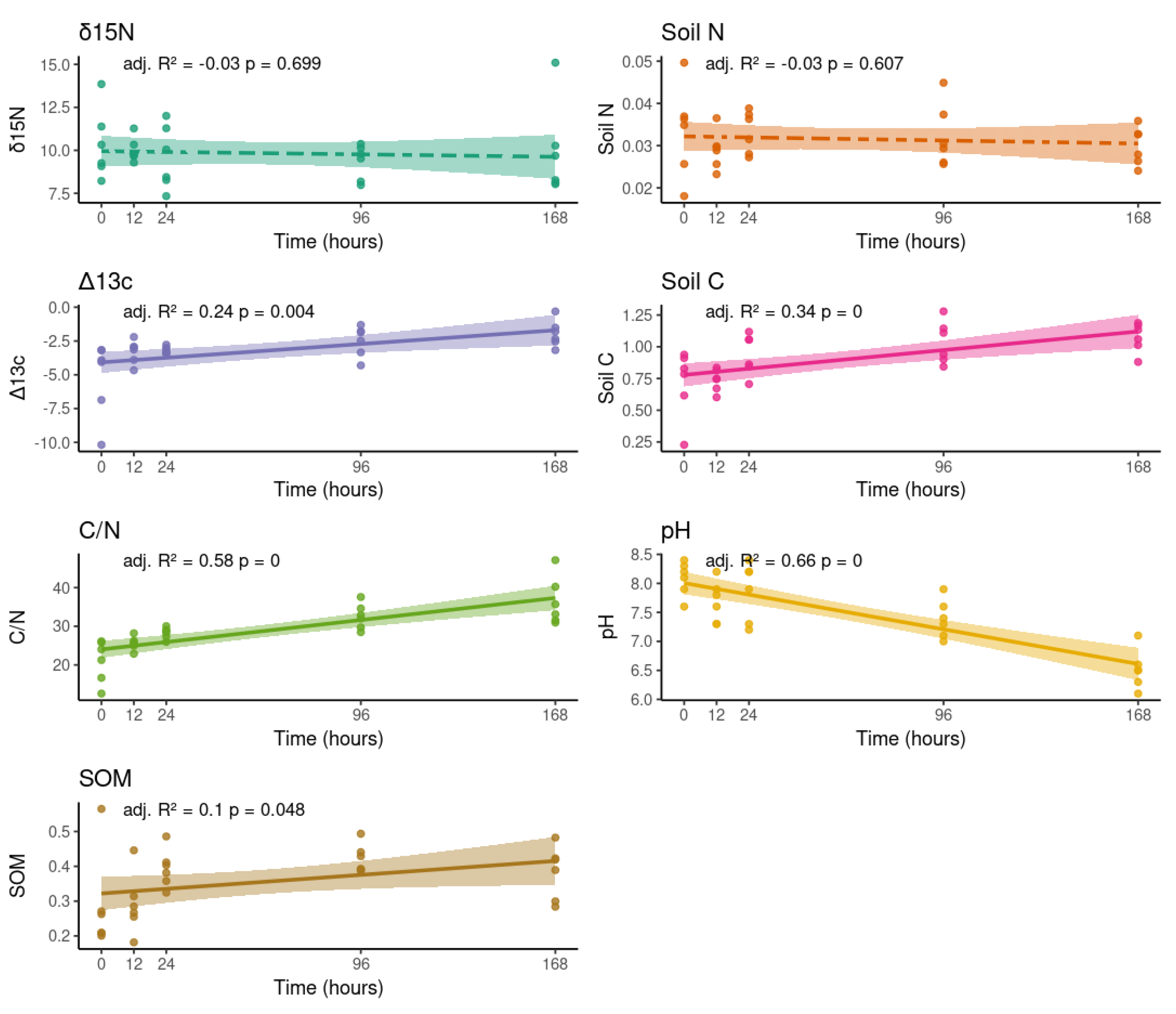
Figure 3.
(a) Bar plot showing the relative abundance of the five most dominant bacterial phyla at each sampling time. Phyla with significant differences (P < 0.05) are marked with an asterisk. (b) Heatmap of the relative abundance of bacterial genera, grouped by phylum on the y-axis and by sampling time on the x-axis, with each sample detailed. (c) Diversity indices (Shannon, Simpson, and Chao1) linear trends for bacterial communities across sampling times (p-values < 0.05 are considered significant, indicated with a continuous line).
Figure 3.
(a) Bar plot showing the relative abundance of the five most dominant bacterial phyla at each sampling time. Phyla with significant differences (P < 0.05) are marked with an asterisk. (b) Heatmap of the relative abundance of bacterial genera, grouped by phylum on the y-axis and by sampling time on the x-axis, with each sample detailed. (c) Diversity indices (Shannon, Simpson, and Chao1) linear trends for bacterial communities across sampling times (p-values < 0.05 are considered significant, indicated with a continuous line).
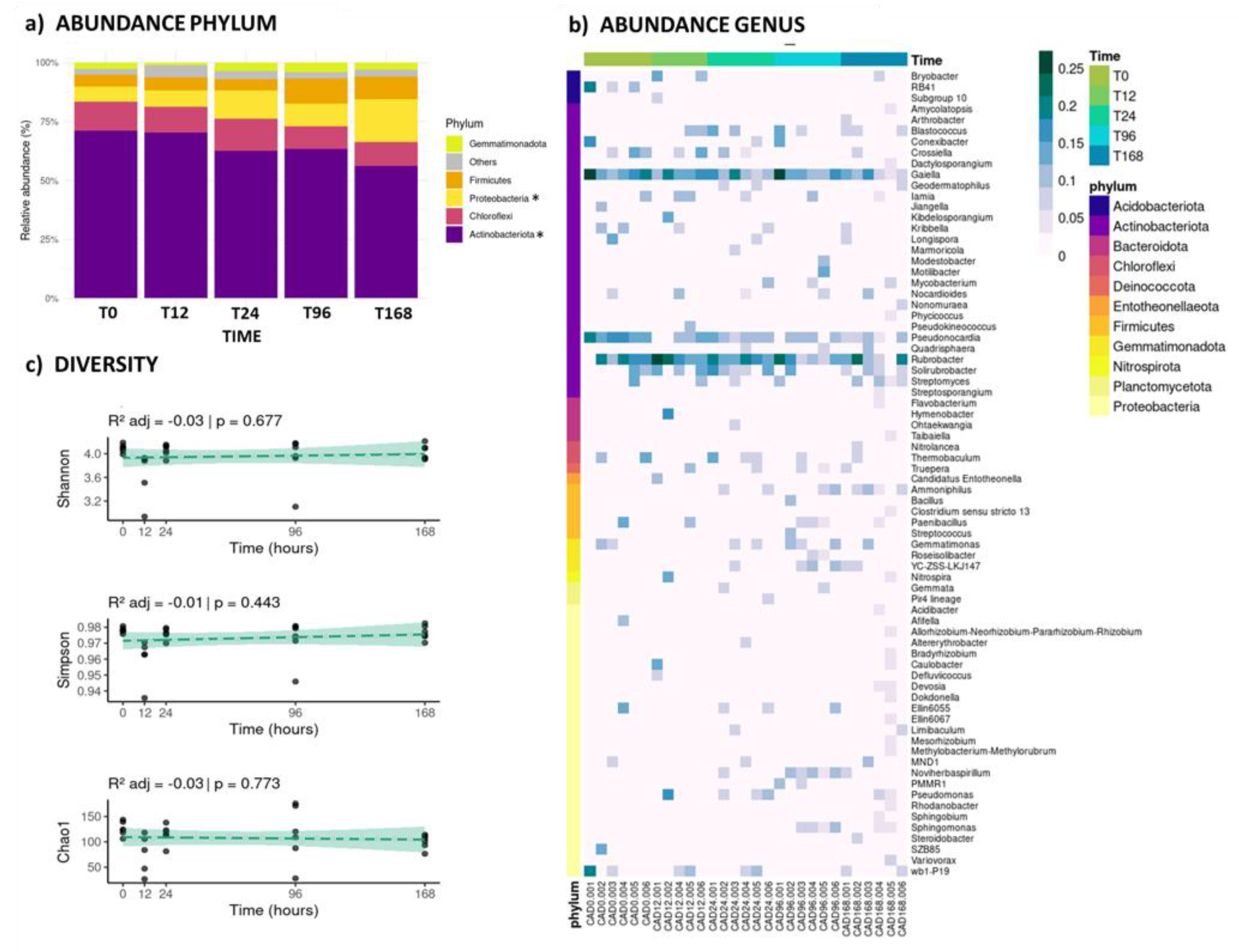
Figure 4.
(a) Bar plot showing the relative abundance of the five most dominant fungal phyla at each sampling time. (b) Heatmap of the relative abundance of fungal genera, grouped by phylum on the y-axis and by sampling time on the x-axis, with each sample detailed. (c) Diversity indices (Shannon, Simpson, and Chao1) linear trends for fungal communities across sampling times (p-values < 0.05 are considered significant, indicated with a continuous line).
Figure 4.
(a) Bar plot showing the relative abundance of the five most dominant fungal phyla at each sampling time. (b) Heatmap of the relative abundance of fungal genera, grouped by phylum on the y-axis and by sampling time on the x-axis, with each sample detailed. (c) Diversity indices (Shannon, Simpson, and Chao1) linear trends for fungal communities across sampling times (p-values < 0.05 are considered significant, indicated with a continuous line).
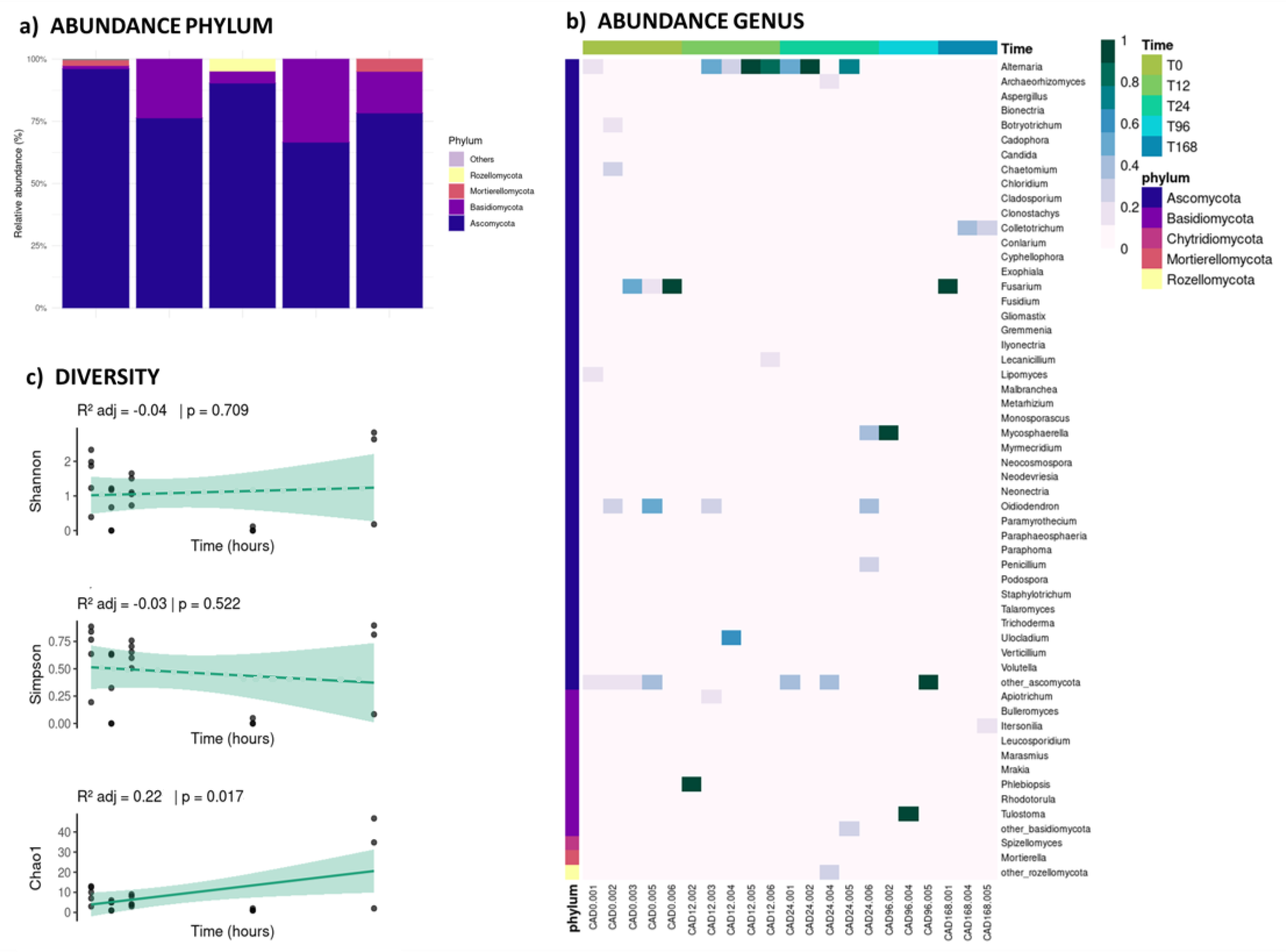
Figure 5.
Heatmaps of functions associated with taxonomy, grouped by functional categories and sampling time for bacteria (a) and fungi (b).
Figure 5.
Heatmaps of functions associated with taxonomy, grouped by functional categories and sampling time for bacteria (a) and fungi (b).

Figure 6.
Upset plots illustrating the number of ASVs shared among bacterial (a) and fungal (b) communities at different sampling times. The matrix layout displays the intersections among the five sampling times, sorted by size. Dark circles in the matrix indicate the sets included in each intersection.
Figure 6.
Upset plots illustrating the number of ASVs shared among bacterial (a) and fungal (b) communities at different sampling times. The matrix layout displays the intersections among the five sampling times, sorted by size. Dark circles in the matrix indicate the sets included in each intersection.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



