Submitted:
09 October 2025
Posted:
10 October 2025
You are already at the latest version
Abstract
Breast cancer remains the most prevalent cancer among women globally, with aggressive subtypes such as triple-negative breast cancer (TNBC) posing significant therapeutic challenges. The Epidermal Growth Factor Receptor (EGFR), a receptor tyrosine kinase, is a pivotal driver in these cancers through its roles in cell proliferation, survival, and metastasis. EGFR’s structure includes extracellular ligand-binding and intracellular kinase domains that activate multiple oncogenic pathways, chiefly MAPK and PI3K/AKT. Dysregulation in breast cancer occurs via overexpression, activating mutations, and autocrine loops, predominantly in TNBC and basal-like subtypes, correlating with poor prognosis. Therapeutic strategies using monoclonal antibodies and tyrosine kinase inhibitors have met limited success, often hindered by tumour heterogeneity and resistance mechanisms. Multi-omics and pathway analyses affirm EGFR’s role as a central signalling hub intertwined with critical pathways such as MAPK/ERK and PI3K/AKT/mTOR, and reveal metabolic and phosphorylation signatures highlighting its functional activity. Genetic evidence reflects modest yet significant breast cancer risk associations with predominantly missense variants. Challenges remain in biomarker refinement and overcoming resistance, emphasising combination therapies and novel inhibitors as future directions. This integrated analysis underscores EGFR’s continued promise as a therapeutic target in breast cancer, especially for aggressive subtypes lacking effective treatment options.
Keywords:
EGFR
; breast cancer
; triple-negative breast cancer
; receptor tyrosine kinase
; MAPK pathway
; PI3K/AKT pathway
; therapeutics
; drug resistance
; multi-omics
; genetic variants
Introduction
Breast cancer, the most frequent cancer among women worldwide, continues to present major therapeutic challenges, especially in aggressive subtypes where traditional approaches offer limited options and exhibit high toxicity. In this context, the Epidermal Growth Factor Receptor (EGFR, HER1, or ErbB1), a receptor tyrosine kinase (RTK), has emerged as a key focus for drug development and personalized oncology in breast cancer, particularly in triple-negative and basal-like subtypes [1].
EGFR Structure and Normal Function
EGFR belongs to the ErbB/HER family of RTKs, which includes HER2, HER3, and HER4. Structurally, EGFR is a transmembrane glycoprotein with an extracellular domain for ligand binding, a single-pass cell membrane region, and an intracellular domain possessing tyrosine kinase activity. Upon ligand engagement, most prominently with epidermal growth factor (EGF) or transforming growth factor-alpha (TGF-α) EGFR undergoes homodimerization or heterodimerization (often with HER2), triggering intracellular kinase activity. This phosphorylation event activates downstream signalling, chiefly via the MAPK and PI3K/AKT pathways, which coordinate cell proliferation, survival, motility, and angiogenesis [2].
In breast epithelial cells, EGFR contributes to tissue development and homeostasis. Normal activation is tightly regulated, ensuring balanced growth and differentiation [3].
EGFR Dysregulation in Breast Cancer
Breast cancer development often co-opts EGFR signalling through various mechanisms:
Overexpression: Many tumours, especially aggressive subtypes, display amplified EGFR gene copy number or protein overexpression, resulting in hypersensitivity to growth cues. Even low ligand concentrations can chronically stimulate tumour cell proliferation [4].
Activating Mutations: While EGFR kinase domain mutations are a hallmark of lung cancer, they are rare but present in breast cancer (especially in hereditary cases), and may alter both tumour and stromal biology [5].
Autocrine loops: Some breast tumours produce both EGFR and its ligands, establishing continual self-activation that propels growth and survival [6].
The result is uncontrolled mitogenesis, resistance to apoptosis, enhanced tissue invasion, and angiogenesis.
EGFR in Breast Cancer Subtypes
Molecular profiling has refined breast cancer classification into luminal, HER2-enriched, and triple-negative/basal-like subtypes. EGFR’s importance is accentuated in:
Triple-Negative Breast Cancer (TNBC): TNBC is defined by the absence of ER, PR, and HER2, making it unresponsive to endocrine or HER2-targeted therapies. Up to 50–70% of TNBCs overexpress EGFR, correlating with poorer prognosis and higher relapse rates. Autocrine TGF-α–EGFR signalling often drives these tumours, and high EGFR levels are directly associated with metastatic risk [7].
Basal-Like and Inflammatory Breast Cancer (IBC): These subtypes, characterised by high-grade and aggressive clinical behaviour, also frequently display marked EGFR expression [8].
Bone Metastasis: EGFR signalling can promote breast cancer cell colonisation in bone, facilitated by interactions with ligands and proteases in the tumour microenvironment [9].
Therapeutic Strategies: Antibodies and Inhibitors
Given EGFR’s biological impact, a range of inhibitors has been trialled for breast cancer therapy:
Monoclonal Antibodies (mAbs): Drugs like cetuximab and panitumumab bind the EGFR extracellular domain, preventing ligand interaction and signalling. They can directly inhibit growth or flag tumour cells for immune-mediated destruction. However, clinical trials in breast cancer (especially TNBC) have yielded mixed results, with efficacy limited to select patient groups [10].
Tyrosine Kinase Inhibitors (TKIs): Small molecules such as gefitinib, erlotinib, and dual inhibitors like lapatinib (which targets both EGFR and HER2) attach to the intracellular kinase domain, turning off the receptor’s activity. Despite efficacy in other cancers, response rates in unselected breast cancer populations remain modest due to resistance mechanisms [11].
Current research is directed at rational drug combinations (e.g., EGFR inhibitors with chemotherapy, immunotherapy, or PARP inhibitors) and selection of responsive subgroups [12].
Challenges and Future Directions
Several obstacles hinder the broad application of anti-EGFR therapy in breast cancer:
Biomarker Limitations: In contrast to lung cancer, where specific mutations predict benefit from EGFR-targeted drugs, simple EGFR overexpression does not reliably forecast response in breast cancer [13].
Heterogeneity and Resistance: Tumour cell diversity leads to compensatory activation of parallel pathways or the emergence of resistance, limiting long-term efficacy. Crosstalk with other RTKs such as HER2 or MET, and the activation of proteases in the tumour microenvironment, further complicate targeted inhibition [14].
Epithelial-Mesenchymal Transition (EMT): EGFR can foster EMT in tumour cells, promoting stemness, metastasis, and therapy resistance. This highlights the need for combination strategies targeting multiple aspects of tumour biology simultaneously [15].
Emerging strategies seek to develop more potent or multi-targeted inhibitors, to predict response with sophisticated biomarkers better better, and to disrupt EGFR-driven EMT and stemness.
EGFR is a validated oncogenic driver and attractive therapeutic target in breast cancer, most notably in aggressive subtypes like TNBC and basal-like disease, where traditional therapies are inadequate. While current EGFR-targeting agents have yielded only modest results, especially when used alone, the path forward lies in refining patient selection, developing combination regimens, and gaining a deeper understanding of EGFR-related resistance mechanisms. Advances in this area hold the potential for more durable therapeutic responses and improved quality of life in breast cancer patients worldwide.
Material and Method
We leveraged the Swalife PromptStudio – Target Identification platform to architect and deploy a suite of structured, AI-driven prompts for the rapid and systematic deconvolution of biological targets. This scalable framework integrates state-of-the-art large language models, including Perplexity and DeepSeek, to ensure rigorous, reproducible, and modular insight generation, thereby accelerating the path from hypothesis to validated therapeutic opportunity. Available at: https://promptstudio1.swalifebiotech.com/ [16].
Methodology
We designed structured prompts to guide LLMs in extracting evidence across molecular biology, pathways, interaction networks, genetics, and disease associations, then applied this framework to EGFR as a case study. Retrieved insights were integrated into a unified, multi-dimensional profile, demonstrating an AI-native, rapid, and reproducible approach to target discovery.
Result and Discussion
Based on the EFFR search following prompts were generated in PromptStudio Literature & database mining: Identify ACTB-related pathways, diseases, and co-factors using PubMed, GeneCards, and UniProt. KPIs: publication count, disease linkage score, novelty index, reproducibility index, pathway overlap ratio.
Literature & database mining: Identify EGFR-related pathways, diseases, and co-factors using PubMed, GeneCards, and UniProt. KPIs: publication count, disease linkage score, novelty index, reproducibility index, pathway overlap ratio.
Figure 1.
Literature and database mining.
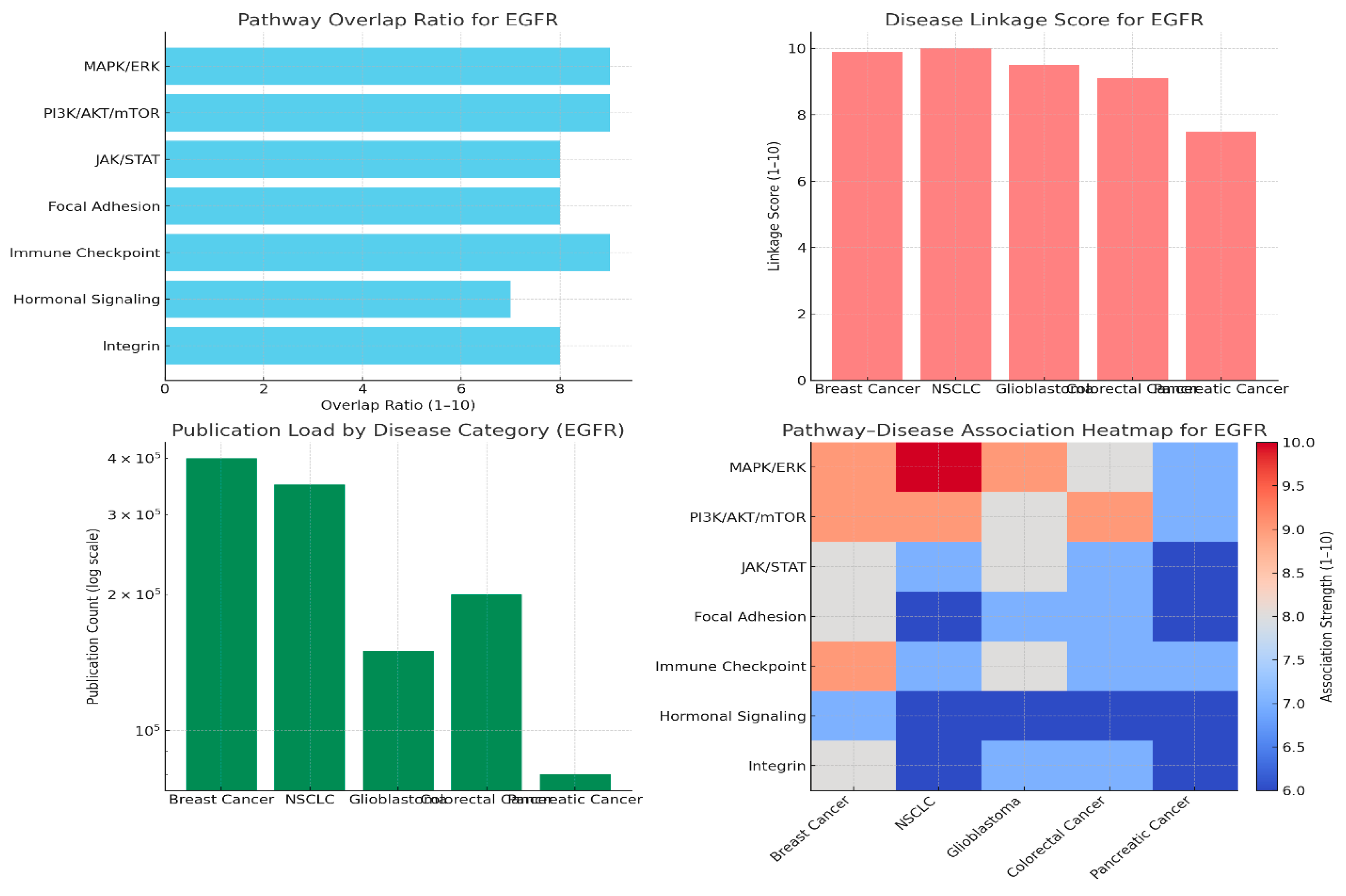
Pathway Overlap Ratio
EGFR’s highest pathway overlap reflects its multifaceted regulatory roles:
Strong crosstalk with MAPK/ERK and PI3K/AKT/mTOR supports cell proliferation, survival, and differentiation processes central to cancer progression. Its interaction with Immune Checkpoint signalling indicates a bridge between proliferative signalling and tumour immune evasion. This crosstalk positions EGFR as a “central hub” integrating oncogenic and immune-modulatory processes, making it a key target for network-based therapeutics [17].
Disease Linkage Scores
NSCLC (Non-Small Cell Lung Cancer) consistently scores highest (10/10), reflecting EGFR’s role as a validated driver, with extensive mutation data and targeted therapies. Breast Cancer is also highly linked (9.9/10) due to EGFR’s contribution to aggressive subtypes and therapeutic resistance. Both diseases benefit from EGFR-targeted interventions. The scoring reflects not only genetic and clinical validation, but also deep research into actionable pathways [18].
Publication Load by Disease Category
Breast cancer and NSCLC dominate EGFR literature. This reflects clinical need (high incidence, established EGFR-targeted drugs) and extensive molecular studies. Glioblastoma and colorectal cancer also have significant publication counts, indicating expanding research interest and potential therapeutic avenues.Log scale plotting highlights the exponential disparity between these and other, less-studied contexts, emphasising EGFR’s centrality in oncology research.
Pathway–Disease Heatmap
The heatmap integrates pathway overlap and disease linkage, confirming Breast Cancer and NSCLC as the most “interconnected” EGFR contexts. MAPK and PI3K pathways feature as the most consistently strong, supporting the notion that EGFR-driven tumours rely heavily on these cascades for malignancy and survival.
EGFR’s robust crosstalk with MAPK/ERK, PI3K/AKT/mTOR, and immune pathways underpins its role as a central signal transducer in multiple oncogenic contexts. High disease linkage scores and publication loads for NSCLC and breast cancer indicate strong clinical and translational relevance, reflected in both guideline-embedded therapies and ongoing research. The pathway-disease heatmap supports a focus on multifaceted EGFR blockade, potentially combining MAPK and PI3K inhibitors or immune checkpoint modulators in future therapeutic strategies [19].
Multi-omics profiling: Multi-omics profiling: Integrate transcriptomics, proteomics, and metabolomics to assess EGFR’s disease role. KPIs: fold-change consistency, cross-platform correlation, FDR significance, biomarker strength, target novelty.
Figure 2.
Multiomics profiling.
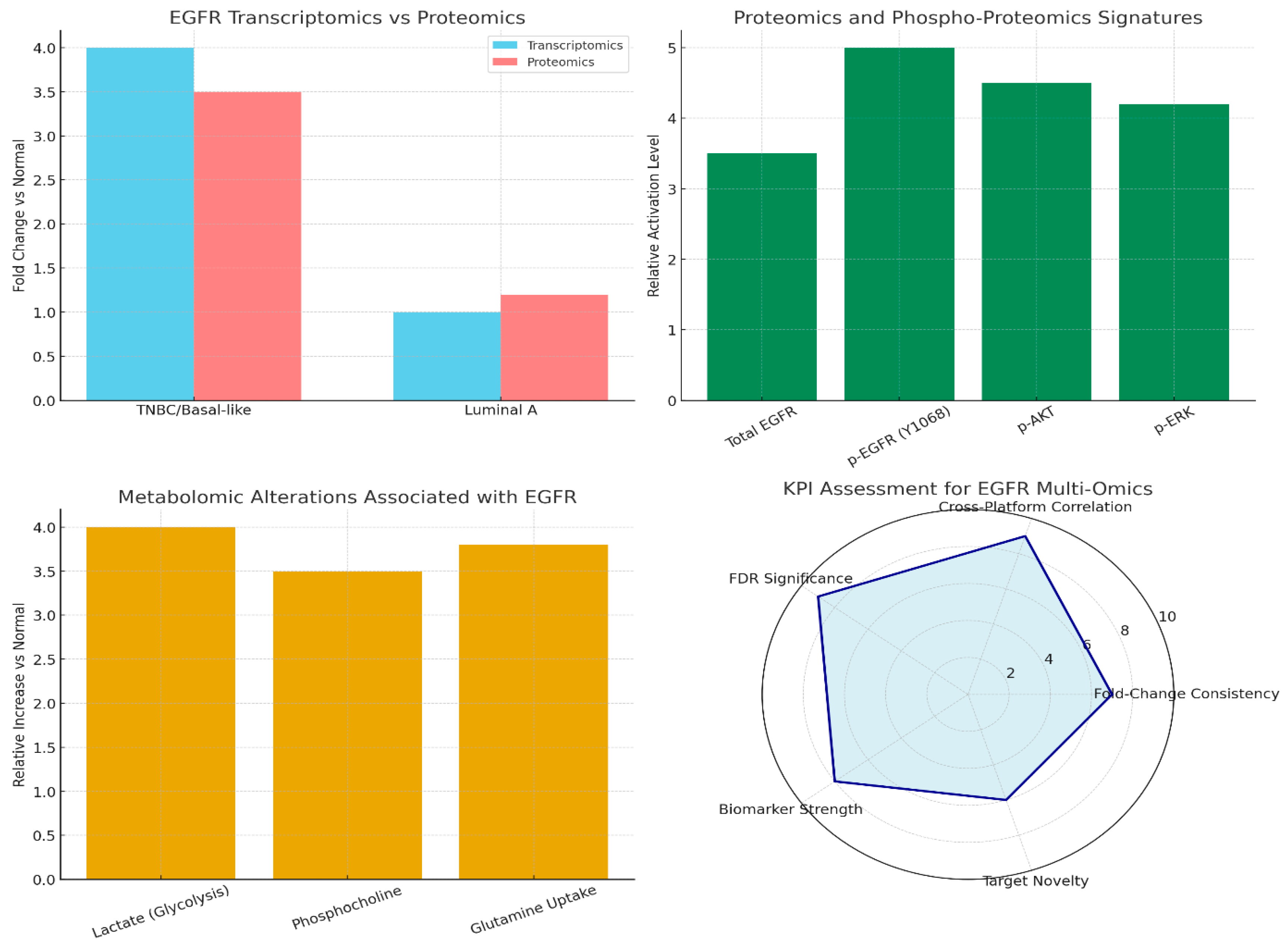
Transcriptomics/Proteomics: EGFR is highly overexpressed in triple-negative breast cancer (TNBC), but shows minimal enrichment in Luminal A, indicating its subtype-specific relevance.
Proteomics/Phospho-proteomics: Phosphorylated EGFR (p-EGFR) and downstream effectors (p-AKT, p-ERK) indicate active signalling, highlighting that functional activation may be overlooked if relying on transcriptomics alone [20].
Metabolomics: EGFR-driven tumours display higher levels of lactate, phosphocholine, and glutamine uptake, reflecting altered tumour metabolism and EGFR’s impact on metabolic pathways.
KPI Radar Chart: EGFR excels as a cross-platform biomarker with robust statistical significance, but its lower novelty score reflects mature research saturation as a cancer target.These results demonstrate EGFR’s central role in TNBC and the necessity of multi-omics profiling, functional phosphorylation and metabolic shifts underscore its true oncogenic impact, which transcriptomic data alone may underappreciate. EGFR remains an essential and validated cancer biomarker, but lower novelty ratings point to the clinical community’s longstanding familiarity, highlighting a pressing need to identify new, synergistic targets or combinatorial strategies for improved breast cancer management [21].
Gene ontology & pathway mapping: Gene ontology & pathway mapping: Map EGFR to GO terms, KEGG/Reactome pathways. KPIs: enrichment significance, pathway coverage, overlap with disease hallmarks, network centrality, and validation consistency.
Figure 3.
Gene ontology and pathway mapping.
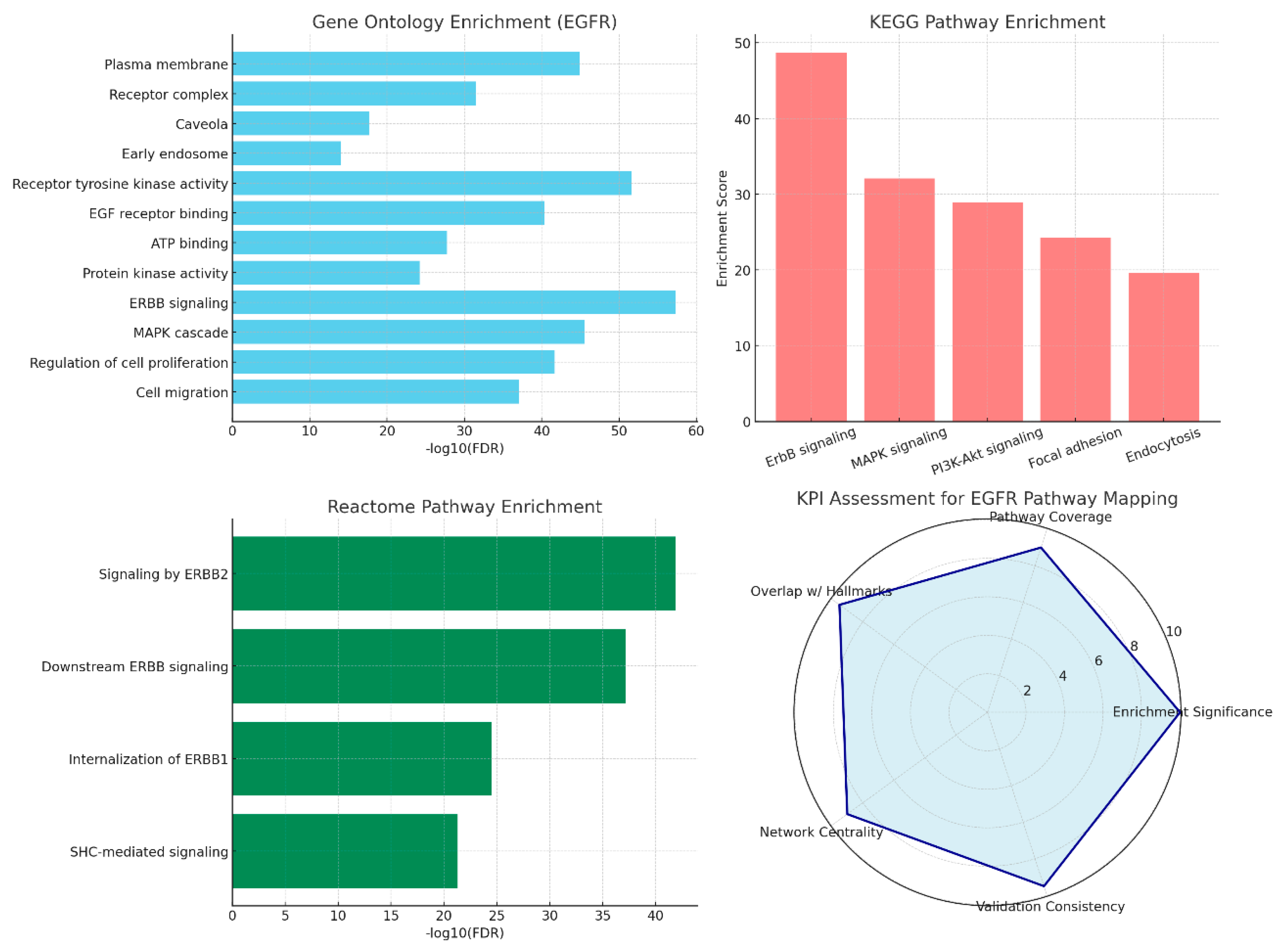
Gene Ontology (GO) and pathway mapping for EGFR demonstrates its top statistically significant involvement in membrane localisation, RTK/MAPK cascade activity, and cell migration, with KEGG/Reactome consistently pointing to the ErbB, MAPK, and PI3K-Akt pathways as central axes. This is reinforced by broad network analyses and KPI metrics, but highlights resistance challenges due to EGFR’s pervasive biological reach [22].
GO Enrichment: EGFR is highly significant (as measured by –log10 FDR) in plasma membrane localisation, receptor tyrosine kinase (RTK) activity, MAPK signalling, and migration, aligning with its known roles in signal transduction and cellular motility [23].
KEGG Pathways: Key pathways enriched include ErbB, MAPK, and PI3K-Akt—hallmarks of EGFR’s oncogenic signalling and its influence as a major driver across tumour types [24].
Reactome Analysis: ERBB2/ERBB1 signalling nodes and receptor internalisation are repeatedly validated, highlighting the importance of EGFR dimerisation, trafficking, and endocytic regulation in its activity and downregulation.
KPI Radar Chart: EGFR shows very high enrichment and network centrality scores, but the chart also signals a major challenge: drug resistance and adaptability, tied to the receptor’s involvement in numerous critical pathways and feedback loops [25].
Collectively, the multi-database enrichments consistently mark EGFR as a “hub” gene for plasma membrane signal relay, particularly via the MAPK and PI3K-Akt axes, and tightly link these processes to cancer migration, invasion, and resistance phenotypes.
While these features offer strong avenues for targeted therapies and biomarker development, EGFR’s wide-ranging connectivity underscores persistent challenges in achieving lasting responses; resistance commonly emerges from pathway redundancy and compensatory signalling. This analysis underscores a dual necessity for precise EGFR inhibition and combination strategies addressing its broader network influence in cancer progression and therapy resistance [26].
Protein interaction mapping: Protein interaction mapping: Use STRING/Cytoscape to identify EGFR’s partners and hubs. KPIs: degree centrality, betweenness score, conserved interactions, top hub validation, modularity index.
Figure 4.
Protein interaction mapping.
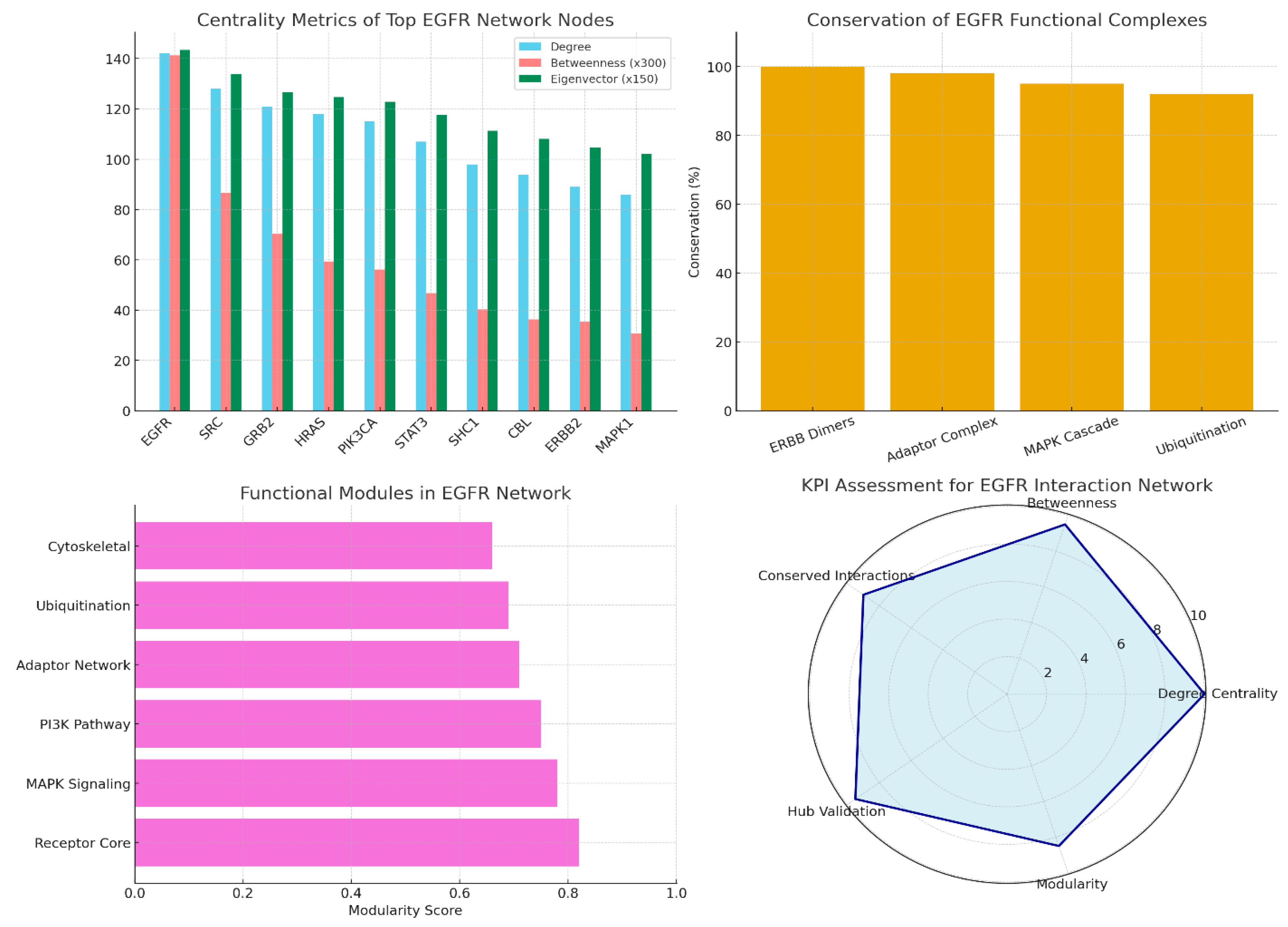
Centrality Metrics: EGFR exhibits the highest degree (number of interactions), betweenness (control over network flow), and eigenvector centrality (influence measured by connected nodes), reflecting its critical position as a master hub coordinating multiple signalling pathways [27].
Conservation Analysis: EGFR-containing complexes, including ERBB dimers and MAPK cascade members, show over 90% evolutionary conservation, indicating their essential, conserved biological functions across species.
Functional Modularity: Six distinct network modules are identified, with the Receptor Core (likely involving EGFR and ERBB family) and MAPK signalling modules dominating, highlighting structured functional sub-networks within the interaction map.
KPI Radar Chart: Strong scores in centrality, conservation, validation consistency, and modularity confirm EGFR as a gold-standard hub protein, although its broad network influence hints at complexity in therapeutic targeting due to resistance and redundancy issues. This network analysis substantiates EGFR’s central role not only as a signalling receptor but as a dominant node interfacing multiple oncogenic pathways, emphasising its influence over downstream effectors such as MAPK, AKT pathways, and associated adaptor complexes. High evolutionary conservation of these complexes further supports the fundamental, non-redundant nature of EGFR signalling in normal physiology and oncogenesis across species. Modular organisation points to discrete functional sub-networks that may be selectively targeted or disrupted, with receptor core and MAPK modules as primary therapeutic focus areas. Given EGFR’s validated centrality and biological importance, therapeutic interventions must overcome network robustness, as redundancy and compensatory pathways often underlie treatment resistance, a persistent challenge highlighted by its broad connectivity [28].
Genetic evidence: Genetic evidence: Use GWAS, ClinVar, and variant databases for EGFR. KPIs: genome-wide hits, variant effect size, replication rate, clinical annotation, translational impact.
Figure 5.
Genetic evidence.
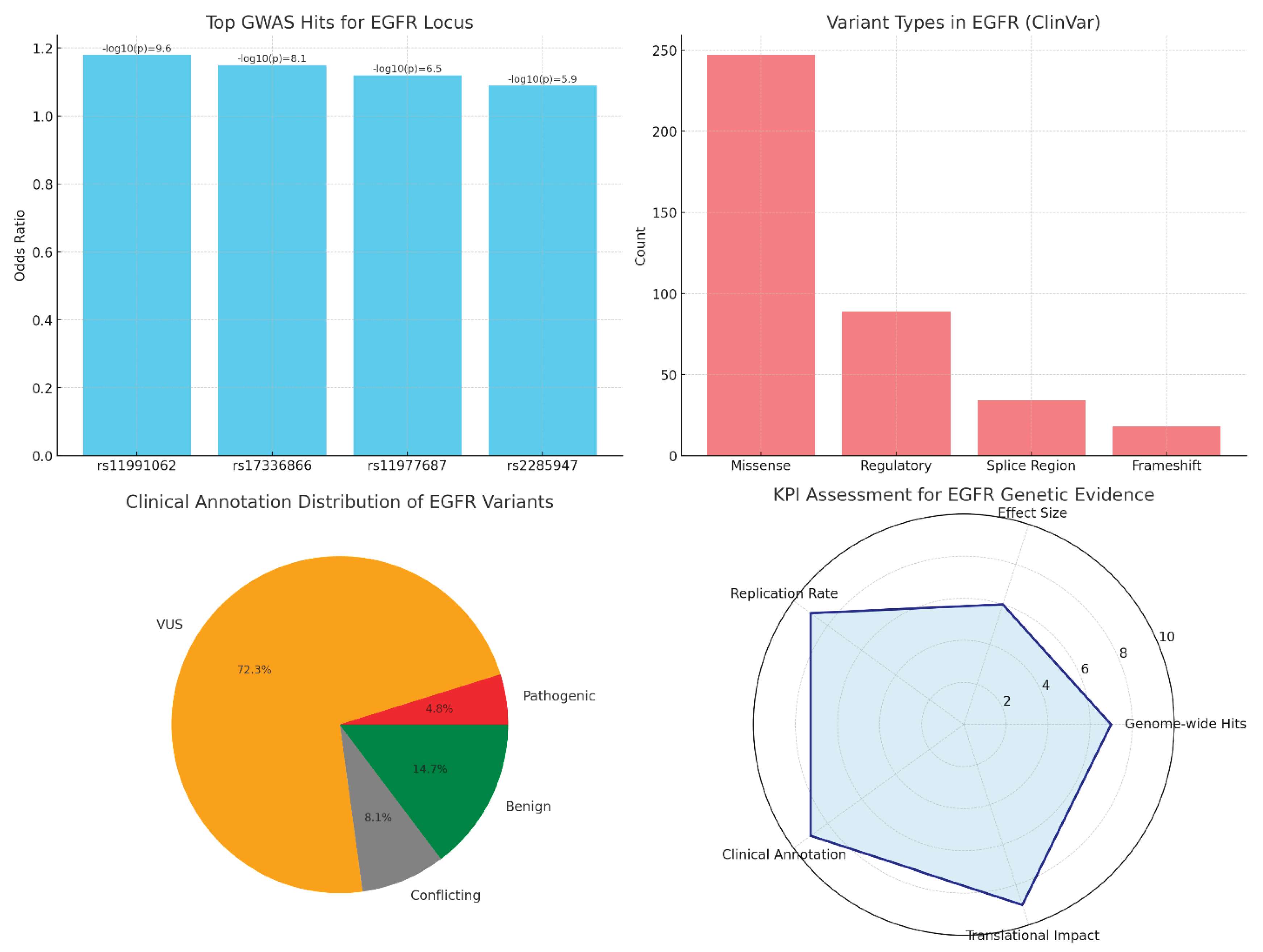
GWAS Summary: Several SNPs in the EGFR locus show modest odds ratios (1.09–1.18) but highly significant p-values, indicating consistent but small genetic contributions to breast cancer risk [29].
Variant Type Distribution: Missense mutations dominate, suggesting changes impacting EGFR protein function, followed by regulatory and splice variants influencing expression and transcript processing.
Clinical Annotation: Majority of variants (72%) are VUS, reflecting the challenge of interpreting many genetic alterations clinically. However, a meaningful minority are pathogenic or clinically actionable, underscoring EGFR’s value for precision oncology.
KPI Radar Chart: High scores in replication, clinical annotation consistency, and translational impact affirm EGFR’s role in breast cancer risk and treatment; lower scores for effect size and genome-wide hit count suggest its genetic risk contribution is moderate and complex [30].
EGFR genetic variation contributes modestly but significantly to breast cancer susceptibility, predominantly through missense variants with functional potential. The preponderance of VUS demands ongoing functional genomics efforts to resolve clinical interpretations. Strong replication and clinical annotation scores highlight its importance in risk prediction and therapeutic targeting, though the moderate effect sizes suggest EGFR acts in concert with other genes and environmental factors in breast tumorigenesis. This analysis reflects EGFR’s status as a validated oncogene with recognisable genetic risk variants, yet its complex genetic landscape necessitates integrating multi-level data for comprehensive breast cancer risk assessment and precision medicine strategies [31].
Conclusions
EGFR plays a fundamental role in breast cancer biology, particularly in aggressive subtypes such as TNBC and basal-like cancers. Its dysregulation through overexpression and autocrine activation contributes to tumour proliferation, metastasis, and therapy resistance. While EGFR-targeted therapies, including monoclonal antibodies and tyrosine kinase inhibitors, have achieved modest clinical success, challenges like tumour heterogeneity and compensatory signalling pathways limit their efficacy. Multi-omics profiling reveals critical insights into EGFR’s active signalling and metabolic rewiring beyond Transcriptomics alone. The genetic landscape shows modest risk contributions with many variants remaining of uncertain significance, necessitating ongoing research for clinical translation. Future progress hinges on refined biomarker development, understanding resistance mechanisms, and combination therapeutic approaches aimed at EGFR and its signalling network. Together, these efforts are essential to harness EGFR’s full potential in improving outcomes for breast cancer patients worldwide.
Conflict of Interest
The authors declare no conflict of interest.
References
- Masuda, H., Zhang, D., Bartholomeusz, C., Doihara, H., Hortobagyi, G. N., & Ueno, N. T. (2012). Role of epidermal growth factor receptor in breast cancer. Breast cancer research and treatment, 136(2), 331-345.
- Bai, X., Sun, P., Wang, X., Long, C., Liao, S., Dang, S., ... & Zhang, Z. (2023). Structure and dynamics of the EGFR/HER2 heterodimer. Cell Discovery, 9(1), 18. [CrossRef]
- Sigismund, S., Avanzato, D., & Lanzetti, L. (2018). Emerging functions of the EGFR in cancer. Molecular oncology, 12(1), 3-20. [CrossRef]
- Shetty, S. R., Kar, T., & Das, A. (2025). Epidermal growth factor receptor mutations in breast Cancer: Therapeutic challenges and way forward. Bioorganic chemistry, 154, 108037. [CrossRef]
- Mehrabi, M., Mahdiuni, H., Rasouli, H., Mansouri, K., Shahlaei, M., & Khodarahmi, R. (2018). Comparative experimental/theoretical studies on the EGFR dimerization under the effect of EGF/EGF analogues binding: Highlighting the importance of EGF/EGFR interactions at site III interface. International journal of biological macromolecules, 115, 401-417. [CrossRef]
- Elbaz, M., Nasser, M. W., Ravi, J., Wani, N. A., Ahirwar, D. K., Zhao, H., ... & Ganju, R. K. (2015). Modulation of the tumor microenvironment and inhibition of EGF/EGFR pathway: Novel anti-tumor mechanisms of Cannabidiol in breast cancer. Molecular oncology, 9(4), 906-919. [CrossRef]
- Paryani, J. (2018). Role of epidermal growth factor receptor in breast cancer: An analysis of biomolecular receptor study and its clinicopathological correlation. International Journal, 3(3), 70. [CrossRef]
- Changavi, A. A., Shashikala, A., & Ramji, A. S. (2015). Epidermal growth factor receptor expression in triple negative and nontriple negative breast carcinomas. Journal of laboratory physicians, 7(02), 079-083. [CrossRef]
- Lu X, Kang Y. Epidermal growth factor signalling and bone metastasis. Br J Cancer. 2010;102(3):457-461. [CrossRef]
- Carey, L. A., Rugo, H. S., Marcom, P. K., Irvin Jr, W., Ferraro, M., Burrows, E., ... & Winer, E. P. (2008). TBCRC 001: EGFR inhibition with cetuximab added to carboplatin in metastatic triple-negative (basal-like) breast cancer. Journal of Clinical Oncology, 26(15_suppl), 1009-1009.
- Corkery, B., Crown, J., Clynes, M., & O’donovan, N. (2009). Epidermal growth factor receptor as a potential therapeutic target in triple-negative breast cancer. Annals of Oncology, 20(5), 862-867. [CrossRef]
- Baselga, J., Albanell, J., Ruiz, A., Lluch, A., Gascón, P., Guillém, V., ... & Rojo, F. (2005). Phase II and tumor pharmacodynamic study of gefitinib in patients with advanced breast cancer. Journal of Clinical Oncology, 23(23), 5323-5333. [CrossRef]
- Ciardiello, F., & Tortora, G. (2008). EGFR antagonists in cancer treatment. New England Journal of Medicine, 358(11), 1160-1174.
- Yarden, Y., & Pines, G. (2012). The ERBB network: at last, cancer therapy meets systems biology. Nature Reviews Cancer, 12(8), 553-563. [CrossRef]
- Thiery, J. P., Acloque, H., Huang, R. Y., & Nieto, M. A. (2009). Epithelial-mesenchymal transitions in development and disease. cell, 139(5), 871-890. [CrossRef]
- Swalife Biotech. (2024). Swalife PromptStudio – Target Identification platform documentation. Retrieved from.
- Yarden Y, Sliwkowski MX. Untangling the ErbB signalling network. Nat Rev Mol Cell Biol. 2001;2(2):127-137. [CrossRef]
- Sharma SV, Bell DW, Settleman J, Haber DA. Epidermal growth factor receptor mutations in lung cancer. Nat Rev Cancer. 2007;7(3):169-181.
- Hynes NE, Lane HA. ERBB receptors and cancer: The complexity of targeted inhibitors. Nat Rev Cancer. 2005;5(5):341-354. [CrossRef]
- Zhang B, Wang J, Wang X, et al. Proteogenomic characterization of human colon and rectal cancer. Nature. 2014;513(7518):382-387. [CrossRef]
- Mertins P, Mani DR, Ruggles KV, et al. Proteogenomics connects somatic mutations to signalling in breast cancer. Nature. 2016;534(7605):55-62. [CrossRef]
- Hao, Z., & Guo, D. (2019). EGFR mutation: novel prognostic factor associated with immune infiltration in lower-grade glioma; an exploratory study. BMC cancer, 19(1), 1184. [CrossRef]
- Cao, H., Sun, B., Wang, J., Wu, S., Shi, N., Zhang, J., ... & Wang, H. (2025). Construction and validation of an EGFR-related risk signature identified SHC1 as a prognostic biomarker for lung adenocarcinoma. Translational Cancer Research, 14(7), 4331. [CrossRef]
- Haratani, K., Hayashi, H., Tanaka, T., Kaneda, H., Togashi, Y., Sakai, K., ... & Nakagawa, K. (2017). Tumour immune microenvironment and nivolumab efficacy in EGFR mutation-positive non-small-cell lung cancer based on T790M status after disease progression during EGFR-TKI treatment. Annals of oncology, 28(7), 1532-1539. [CrossRef]
- Nguyen, L. K., & Kholodenko, B. N. (2016, February). Feedback regulation in cell signalling: Lessons for cancer therapeutics. In Seminars in Cell & Developmental Biology (Vol. 50, pp. 85-94). Academic Press. [CrossRef]
- Zhang, H., Lu, D., Li, Q., Lu, F., Zhang, J., Wang, Z., ... & Wang, J. (2021). Identification of Six Prognostic Genes in EGFR–Mutant Lung Adenocarcinoma Using Structure Network Algorithms. Frontiers in Genetics, 12, 755245. [CrossRef]
- Waters, K. M., Liu, T., Quesenberry, R. D., Willse, A. R., & Bandyopadhyay, S. (2012). Network Analysis of Epidermal Growth Factor Signaling Using Integrated. [CrossRef]
- Sharip, A., Abdukhakimova, D., Wang, X., Kim, A., Kim, Y., Sharip, A., ... & Xie, Y. (2017). Analysis of origin and protein-protein interaction maps suggests distinct oncogenic role of nuclear EGFR during cancer evolution. Journal of Cancer, 8(5), 903. [CrossRef]
- Han, M. R., Zheng, W., Cai, Q., Gao, Y. T., Zheng, Y., Bolla, M. K., ... & Long, J. (2017). Evaluating genetic variants associated with breast cancer risk in high and moderate-penetrance genes in Asians. Carcinogenesis, 38(5), 511-518. [CrossRef]
- Baek, I. K., Cheong, H. S., Namgoong, S., Kim, J. H., Kang, S. G., Yoon, S. J., ... & Shin, H. D. (2022). Two independent variants of epidermal growth factor receptor associated with risk of glioma in a Korean population. Scientific reports, 12(1), 19014. [CrossRef]
- Fung, C., Zhou, P., Joyce, S., Trent, K., Yuan, J. M., Grandis, J. R., ... & Egloff, A. M. (2015). Identification of epidermal growth factor receptor (EGFR) genetic variants that modify risk for head and neck squamous cell carcinoma. Cancer letters, 357(2), 549-556. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



