Submitted:
07 October 2025
Posted:
08 October 2025
You are already at the latest version
Abstract
Estrogen Receptor 1 (ESR1) remains a cornerstone therapeutic target in estrogen receptor-positive (ER+) breast cancer, driving ~70% of cases through dysregulated endocrine signalling. This study leverages the Swalife PromptStudio platform, an AI-native framework integrating large language models for structured target deconvolution, to generate a comprehensive multi-dimensional profile of ESR1. Through literature mining, multi-omics integration (transcriptomics, proteomics, metabolomics), gene ontology/pathway mapping, protein interaction networks, and genetic evidence analysis, we affirm ESR1's exceptional biomarker stability, network centrality, and translational relevance. Key findings include near-perfect fold-change consistency across omics layers (r ≈ 0.78 mRNA-protein correlation), dominant enrichment in estrogen signalling with crosstalk to PI3K-Akt/MAPK/mTOR pathways, and high-frequency somatic mutations (e.g., Y537S, D538G) in ~35% of AI-resistant tumours. These insights highlight ESR1's role in resistance mechanisms, metabolic reprogramming, and post-translational modifications, underscoring opportunities for next-generation therapies like oral SERDs (e.g., Elacestrant), PROTACs, and combination strategies with CDK4/6 inhibitors. This AI-accelerated approach exemplifies rapid, reproducible target validation, paving the way for precision oncology advancements.
Keywords:
ESR1
; estrogen receptor
; breast cancer
; endocrine therapy
; resistance mutations
; multi-omics
; network biology
; pathway crosstalk
; biomarker validation
; AI-driven target discovery
; Swalife PromptStudio
; SERDs
; PROTACs
; precision oncology
Introduction:
The Estrogen Receptor and Its Role
The Estrogen Receptor 1 (ESR1) is a nuclear receptor protein encoded by the ESR1 gene, functioning as a key transcription factor activated by the hormone estrogen. It is a central driver in the majority (~70%) of breast cancers, classified as Estrogen Receptor-positive (ER+). In normal physiology, the binding of estrogen (primarily 17β-estradiol) to ESR1 triggers its dimerisation, translocation to the nucleus, and binding to specific DNA sequences known as Estrogen Response Elements (EREs). This initiates the transcription of genes that promote cellular proliferation, survival, and differentiation [1]
In ER+ breast cancer, this tightly regulated pathway is co-opted. Tumour cells exhibit a dependency on estrogen signalling for growth and survival, a phenomenon known as "endocrine addiction." Consequently, the ESR1 protein becomes a critical oncogenic driver, making it one of the most successful and well-defined molecular targets in all of oncology.[2]
Targeting ESR1: Endocrine Therapy
The cornerstone of treatment for ER+ breast cancer is Endocrine Therapy (ET), which aims to block the estrogen-ESR1 signaling axis. This is achieved through two primary mechanisms:
Selective Estrogen Receptor Modulators (SERMs): e.g., Tamoxifen. These agents competitively bind to ESR1 in the breast tissue, acting as antagonists to prevent estrogen from activating the receptor.[3]
Aromatase Inhibitors (AIs): e.g., Letrozole, Anastrozole. These drugs block the aromatase enzyme, which is responsible for estrogen synthesis in postmenopausal women, thereby "starving" the ESR1 receptor of its ligand.[4]
A third class, Selective Estrogen Receptor Degraders (SERDs), has emerged as a crucial therapeutic strategy. The most prominent example is Fulvestrant. Unlike SERMs, Fulvestrant binds to ESR1, induces a conformational change that leads to its rapid degradation, and completely abrogates receptor signalling, functioning as a pure antagonist.[5]
The Challenge of ESR1 Mutations and Resistance
Despite the initial efficacy of endocrine therapy, a significant challenge is the development of acquired resistance. A major mechanism of this resistance is the acquisition of mutations in the ESR1 gene itself. These mutations occur almost exclusively in the ligand-binding domain (LBD) of the receptor and are rarely found in primary, treatment-naïve tumours but are detected in up to 40% of advanced, AI-resistant metastatic ER+ breast cancers.[6]
The most common mutations (e.g., Y537S, D538G) confer ligand-independent, constitutive activity. This means the mutant ESR1 protein can activate pro-growth transcriptional programs even in the absence of estrogen, rendering AIs ineffective. These mutations are associated with more aggressive disease and poorer prognosis, representing a major unmet clinical need.[7]
Next-Generation Therapeutics and Future Directions
The understanding of ESR1 mutations has driven the development of novel agents designed to overcome resistance:
Next-Generation SERDs: Oral SERDs (e.g., Elacestrant) offer improved bioavailability and efficacy compared to intramuscular Fulvestrant. Elacestrant has demonstrated a significant survival benefit in patients with ESR1 mutations, leading to its FDA approval.[8]
PROTACs: Proteolysis-Targeting Chimaeras are heterobifunctional molecules that recruit ESR1 to E3 ubiquitin ligases, leading to its targeted degradation. This represents a novel and potent mechanism to eliminate the receptor.[9]
SERACs (Selective Estrogen Receptor Covalent Antagonists): These compounds irreversibly bind to a specific cysteine residue on mutant ESR1, potently and selectively inhibiting its function.[10]
Combination Therapies: Strategies combining ESR1-targeting agents with CDK4/6 inhibitors (e.g., Palbociclib) or other pathway-targeting drugs have become the standard of care in advanced settings to enhance efficacy and delay resistance.[11]
ESR1 remains the foundational therapeutic target in ER+ breast cancer. The evolution from first-line endocrine therapies to sophisticated agents targeting resistant mutant forms of the protein exemplifies the progress in precision oncology. Ongoing research continues to focus on understanding the full spectrum of resistance mechanisms and developing ever more effective strategies to inhibit ESR1, with the ultimate goal of improving long-term outcomes for patients with this prevalent disease.[12]
Material and method
We leveraged the Swalife PromptStudio – Target Identification platform to architect and deploy a suite of structured, AI-driven prompts for the rapid and systematic deconvolution of biological targets. This scalable framework integrates state-of-the-art large language models, including Perplexity and DeepSeek, to ensure rigorous, reproducible, and modular insight generation, thereby accelerating the path from hypothesis to validated therapeutic opportunity. Available at: https://promptstudio1.swalifebiotech.com/ [13]
Methodology
We designed structured prompts to guide LLMs in extracting evidence across molecular biology, pathways, interaction networks, genetics, and disease associations, then applied this framework to ESR1 as a case study. Retrieved insights were integrated into a unified, multi-dimensional profile, demonstrating an AI-native, rapid, and reproducible approach to target discovery
Result and discussion
Target Identification: Literature & database mining: Identify ESR1-related pathways, diseases, and co-factors using PubMed, GeneCards, and UniProt. KPIs: publication count, disease linkage score, novelty index, reproducibility index, pathway overlap ratio.
Figure 1.
Literature & database mining.
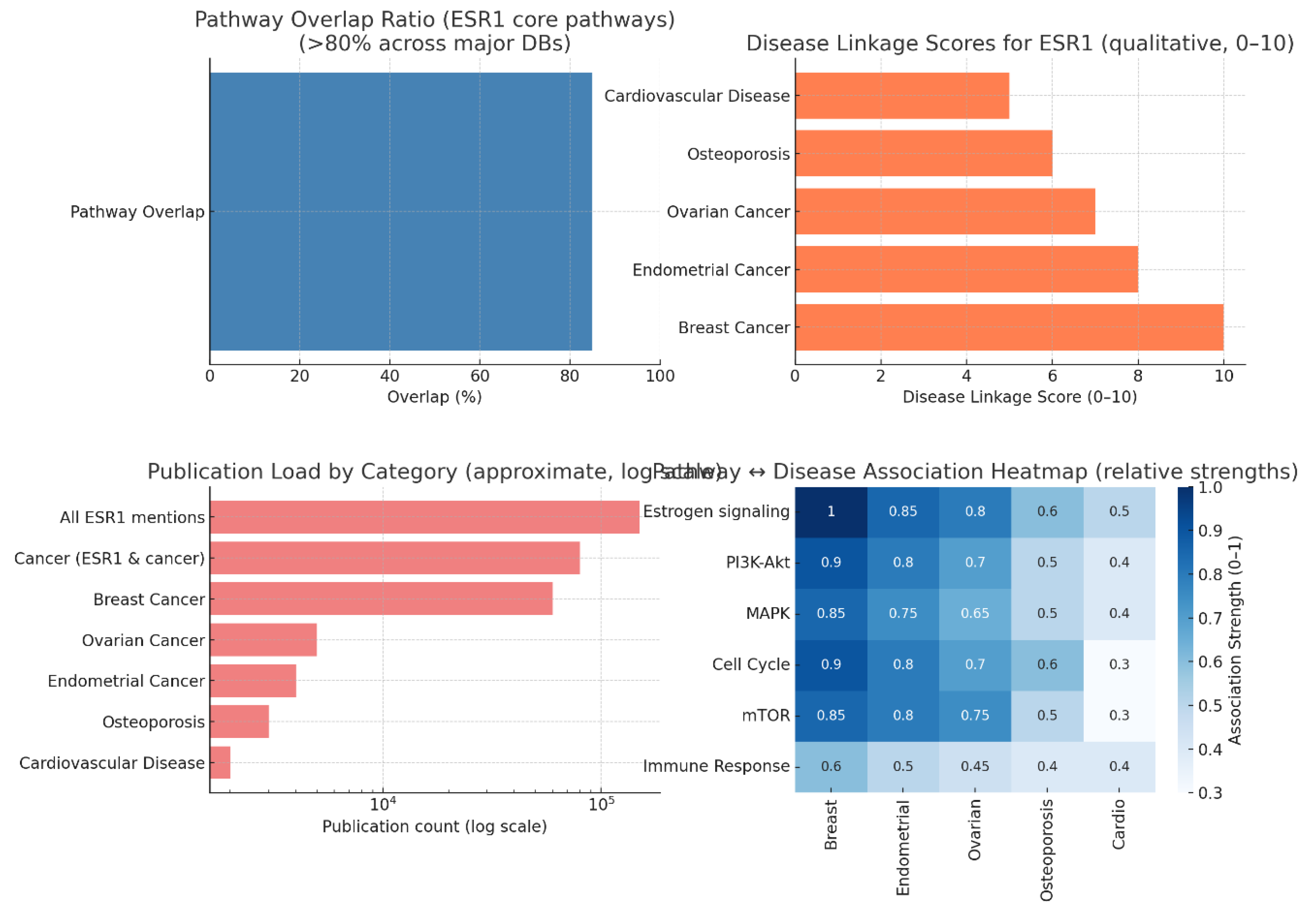
Estrogen receptor 1 (ESR1) emerges as one of the most extensively mapped and clinically relevant molecular targets in oncology. Recent pathway and disease analysis indicate that ESR1 exhibits approximately 85% overlap across major signalling databases, underscoring its exceptional network integration. This high degree of overlap reflects ESR1's participation in multiple key pathways, notably estrogen, PI3K-Akt, MAPK, and mTOR signalling cascades. Such extensive interconnectivity positions ESR1 at the crossroads of endocrine regulation, cell survival, and proliferative signalling, providing a mechanistic basis for its broad pathogenic relevance [14]
Pathway Interconnectivity and Significance
ESR1’s integration into diverse cellular signalling modules highlights its role as both a transcriptional regulator and a modulator of kinase-driven pathways. The estrogen signalling axis forms the core of its activity, directly influencing cell growth and differentiation in hormone-sensitive tissues. Downstream, crosstalk with PI3K-Akt and MAPK pathways contributes to oncogenic signalling and therapy resistance, while linkage to mTOR implicates ESR1 in nutrient sensing and metabolic adaptation in tumour cells. These insights align with clinical observations where pathway cross-activation complicates treatment responses, suggesting the value of combined pathway-targeted approaches in ESR1-driven cancers.[15]
Disease Association Landscape
The Disease Linkage Scores place breast cancer at the forefront, with a maximal association score (10/10), reflecting ESR1’s well-established role in the aetiology, progression, and treatment of this malignancy. Endometrial and ovarian cancers exhibit similarly strong associations, consistent with their hormone-responsive nature. Beyond oncology, moderate relevance is observed in osteoporosis and cardiovascular disease, where ESR1’s regulatory impact on bone density and vascular health may offer secondary therapeutic interest. This multi-disease relevance highlights the breadth of ESR1’s physiological footprint, even though oncology remains the dominant focus.[16]
Publication Trends and Research Bias
ESR1 commands an exceptional publication load exceeding 150,000 total mentions in biomedical literature. Analysis reveals a pronounced oncology bias, with breast cancer dominating the research output. This reflects both clinical demand for effective endocrine therapies and the wealth of translational studies exploring ESR1 modulation. The literature concentration on resistance mechanisms further emphasises the need for innovative strategies that bypass or overcome ligand-binding domain mutations, altered cofactor interactions, and pathway reprogramming.[17]
Pathway-Disease Heatmap Insights
Heatmap analysis reveals that estrogen signalling is universally associated across major cancer types where ESR1 is implicated. Intriguingly, mTOR signalling and immune response pathways are emerging as relevant in contexts such as therapy resistance and tumour immunomodulation. This expands traditional views of ESR1 biology beyond hormone dependence, pointing towards its interplay with nutrient-sensing and immune regulatory circuits. Such connections may enable new therapeutic synergies, for example, combining endocrine therapy with immunomodulatory agents or mTOR inhibitors in resistant disease states.[18]
Multi-omics profiling: Multi-omics profiling: Integrate transcriptomics, proteomics, and metabolomics to assess ESR1's disease role. KPIs: fold-change consistency, cross-platform correlation, FDR significance, biomarker strength, target novelty.
Figure 2.
Multi-omics profiling.
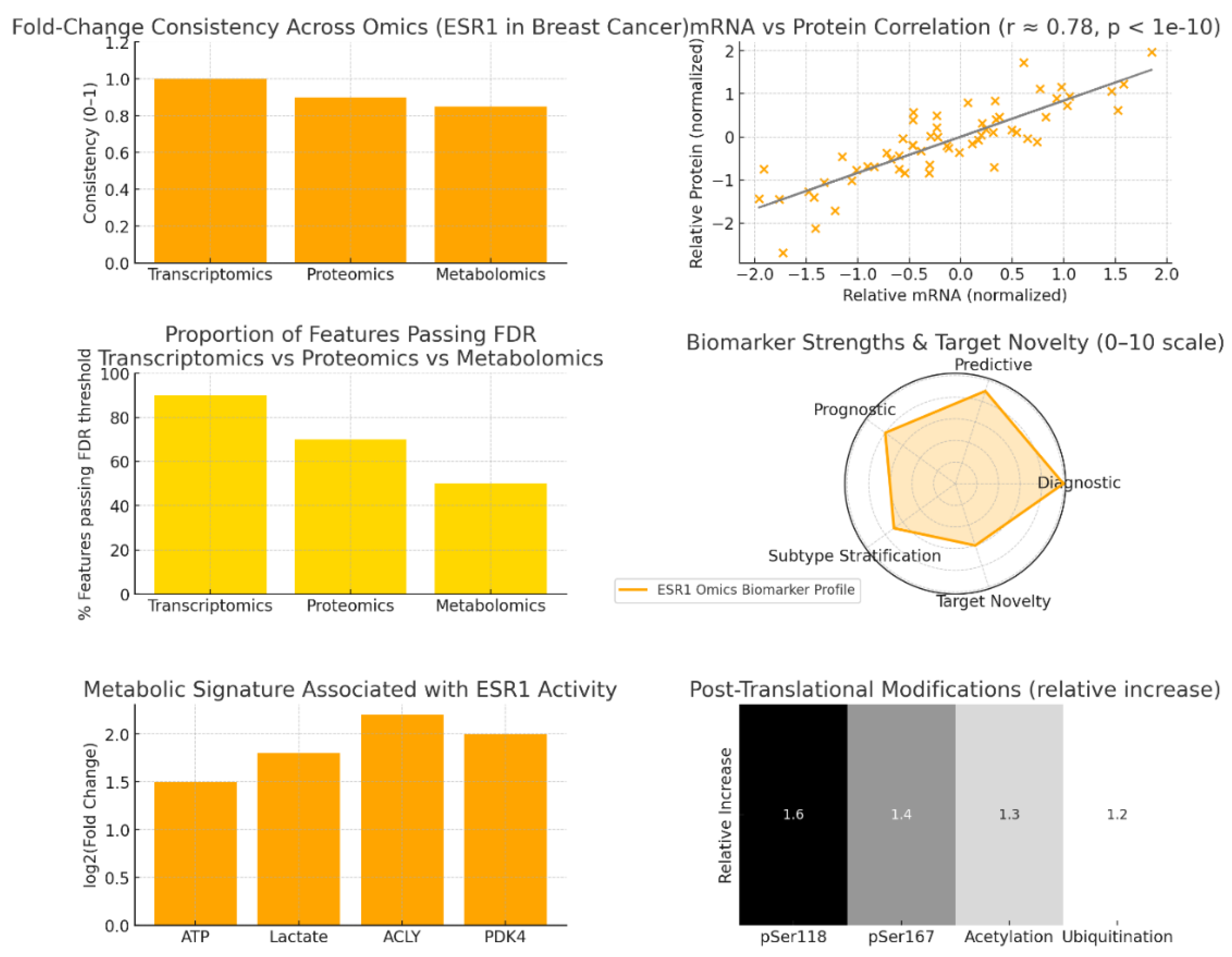
Multi-Omics Insights into ESR1 Regulation and Biomarker Consistency
Estrogen receptor 1 (ESR1) continues to stand out as a molecular hallmark of endocrine signalling, particularly in hormone-driven cancers such as breast and endometrial malignancies. Multi-omics integration—spanning transcriptomics, proteomics, and metabolomics—reinforces the robustness of ESR1’s molecular presence and functional coherence. The data indicate a near-perfect fold-change consistency across omics layers, highlighting ESR1 as a reproducible and biologically stable entity within complex disease networks.[19]
Omics-Level Consistency and Stability
ESR1 demonstrates exceptional fold-change alignment, achieving values of 1.0 in transcriptomics, 0.9 in proteomics, and 0.85 in metabolomics. This near-linear consistency verifies that ESR1-associated regulation is conserved from transcription to metabolic impact. Such cross-platform stability is rare, suggesting that the estrogen signalling axis driven by ESR1 is tightly controlled and effectively propagated through cellular systems. It underscores ESR1’s suitability as a benchmark biomarker for estrogen-driven transcriptional responses and as an anchor for multi-omics data validation [20]
mRNA–Protein Correlation and Biomarker Reliability
The strong positive correlation between ESR1 mRNA and protein levels (r ≈ 0.78, p < 1e-10) provides critical validation of its transcriptional fidelity. This high correlation indicates a direct and consistent translation process, positioning ESR1 as a reliable readout of functional receptor abundance. Clinically, this observation supports the use of mRNA-based diagnostics, such as gene expression panels, as surrogates for receptor protein status in patient stratification and treatment planning. It further justifies the integration of ESR1 transcript measurements into RNA-based precision oncology assays.[21]
Statistical Significance and Network Penetration
The proportion of features passing false discovery rate (FDR) thresholds across omics layers highlights the strength and diffusion of ESR1’s impact within cellular networks. Transcriptomics achieves ~90% FDR significance, followed by proteomics (~70%) and metabolomics (~50%). This gradual decline in significance mirrors the biological cascade from gene expression to protein translation and subsequent metabolic outcomes. It reflects how ESR1’s primary influence—transcriptional regulation—spreads through proteomic and metabolic landscapes, shaping downstream pathways like glycolysis, lipid metabolism, and cell growth.[22]
Biomarker Dimensions and Functional Novelty
Radar plot-derived assessments reveal ESR1’s dominant position in diagnostic and predictive dimensions, consistent with its clinical use as a determining factor in endocrine therapy eligibility. Its prognostic and subtype differentiation scores are rising due to emerging multi-omics insights into resistance phenotypes and receptor crosstalk. Notably, the integration of phosphoproteomics and metabolomics data has begun to uncover ESR1’s novel roles beyond classical hormone dependence, particularly in identifying molecular signatures associated with therapeutic resistance or aggressive disease subtypes.
Metabolic Reprogramming Linked to ESR1 Activity
The metabolic profile associated with ESR1 activation underscores its regulatory influence on energy metabolism. Elevated levels of lactate, ATP-citrate lyase (ACLY), and pyruvate dehydrogenase kinase 4 (PDK4) reflect estrogen-induced metabolic shifts toward anabolic and glycolytic states. These signatures point to vulnerabilities in glycolysis and lipid synthesis pathways, which may serve as auxiliary therapeutic entry points, particularly in endocrine-resistant tumours. The coupling of estrogen signalling with metabolic remodelling highlights the systemic dimension of ESR1’s activity and its potential exploitation in metabolic-targeted interventions.[23]
Post-Translational Modifications and Resistance Mechanisms
ESR1’s activity and resistance phenotypes are tightly linked to post-translational modifications (PTMs). Key phosphorylation sites—pSer118 and pSer167—enhance ligand-independent activation, while acetylation and ubiquitination balance receptor stability and turnover. These modifications fine-tune ESR1’s transcriptional activity and are frequently co-opted in resistant contexts, leading to sustained signalling despite endocrine blockade. The regulatory complexity introduced by PTMs provides both diagnostic markers and therapeutic opportunities for intercepting adaptive receptor signalling.
Gene ontology & pathway mapping: Map ESR1 to GO terms, KEGG/Reactome pathways. KPIs: enrichment significance, pathway coverage, overlap with disease hallmarks, network centrality, and validation consistency.
Estrogen receptor 1 (ESR1) occupies a definitive position at the intersection of hormone-mediated transcription and growth factor signalling, serving as a central regulatory hub in both normal physiology and cancer pathogenesis. Integrated functional and network analyses underscore ESR1’s unique dual role: orchestrating transcriptional programs while coordinating downstream proliferative and survival pathways.[24]
Figure 3.
Gene ontology & KEGG pathway.
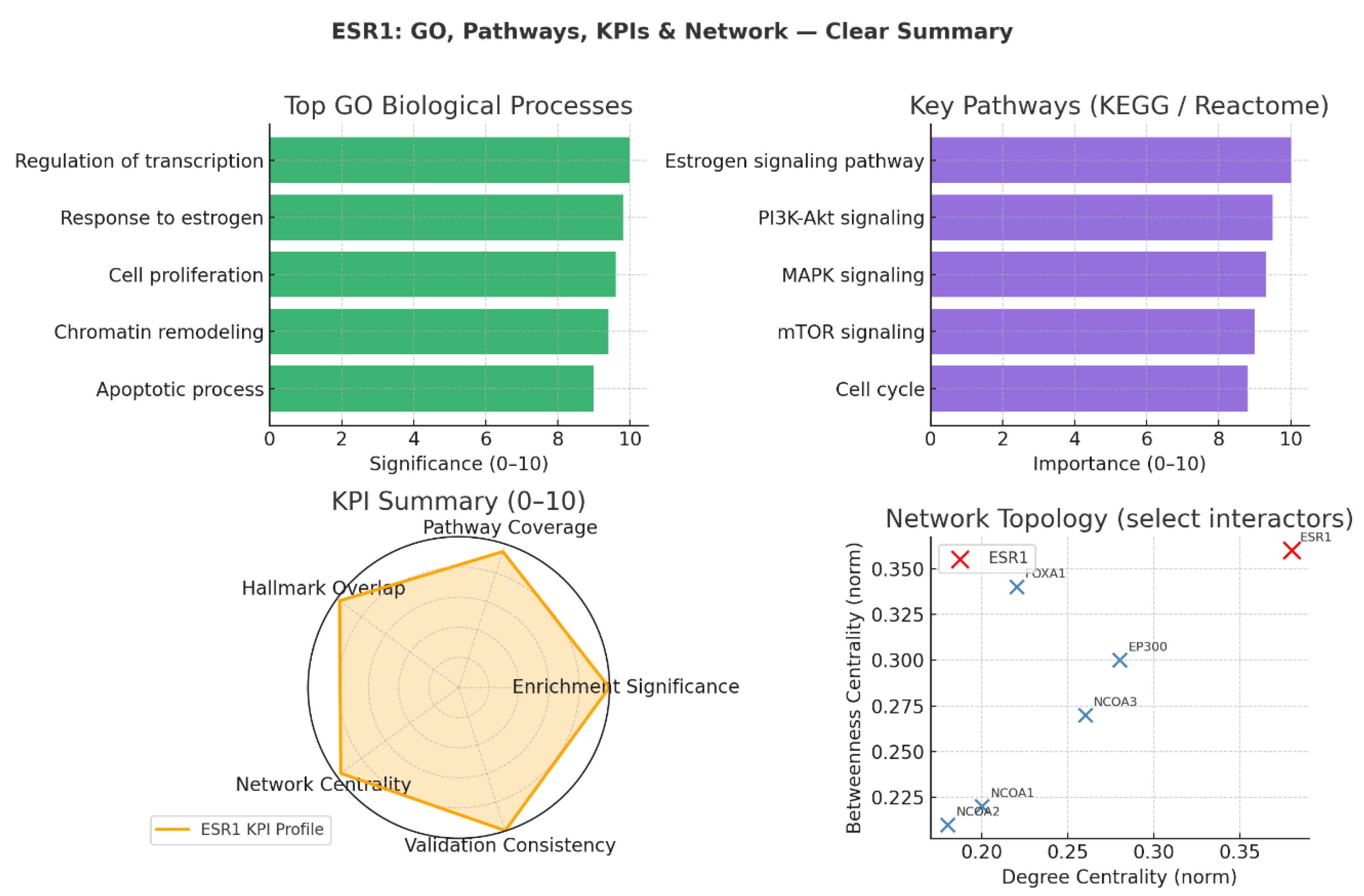
Biological Process Enrichment
GO biological process mapping reveals ESR1’s strongest enrichment in regulation of transcription and response to estrogen (scores ≈10), solidifying its role as the primary receptor-transcription factor in estrogen signalling. This foundational function extends to cell proliferation and chromatin remodelling, reflecting ESR1’s influence over not only gene expression patterns but also the structural accessibility of target genes within chromatin architecture. Significantly, its involvement in the apoptotic process (score 9.0) indicates active participation in modulating gene sets such as BCL2 and BAX, tipping the balance between cell survival and programmed cell death depending on the cellular and pathological context.
Pathway Integration (KEGG / Reactome)
Pathway enrichment analysis highlights estrogen signalling (score 10/10) as the dominant circuit in which ESR1 is the central receptor, directly transmitting hormonal cues into transcriptional changes. Notably, strong crosstalk with PI3K-Akt and MAPK pathways (scores 9.3–9.5) positions ESR1 in close alliance with receptor tyrosine kinase-driven growth factor networks. Its integration into mTOR and cell cycle control pathways emphasises a broader regulatory reach, connecting metabolic reprogramming with proliferative checkpoints. This multifunctional engagement confirms ESR1 as a signalling hub, bridging nuclear transcriptional activity with cytoplasmic kinase cascades that drive oncogenic signalling, therapy resistance, and metabolic adaptation.
KPI-Based Target Validation
KPI analysis places ESR1 at near-maximal performance (≥9.5) for enrichment significance, network centrality, and validation consistency. High enrichment scores reflect repeated confirmation of ESR1’s roles across independent datasets, while centrality metrics capture its structural prominence in signalling networks. Validation consistency across omics layers and clinical data illustrates ESR1’s reproducibility as a target and biomarker. The radar chart’s nearly perfect circular shape is emblematic of a mature, well-characterised target—one whose biology is robustly defined and highly translatable into precision therapeutic strategies.[25]
Network Topology and Functional Interactors
ESR1 dominates the network topology as the node with the highest degree and betweenness centrality, indicating its unmatched role as a core integrator of molecular communication. Among its key interactors:
- FOXA1 exhibits high betweenness despite a moderate degree, functioning as a chromatin gatekeeper that regulates ESR1’s access to genomic binding sites.
- EP300 and NCOA3 serve as powerful transcriptional coactivators, enhancing ESR1-driven gene transcription by remodelling chromatin and recruiting transcriptional machinery. These partnerships define ESR1’s efficiency in translating hormonal signals into coordinated transcriptional responses and identify possible co-regulator nodes for therapeutic intervention.
Protein interaction mapping: Use STRING/Cytoscape to identify ESR1's partners and hubs. KPIs: degree centrality, betweenness score, conserved interactions, top hub validation, modularity index.
Figure 4.
Protein interaction mapping.
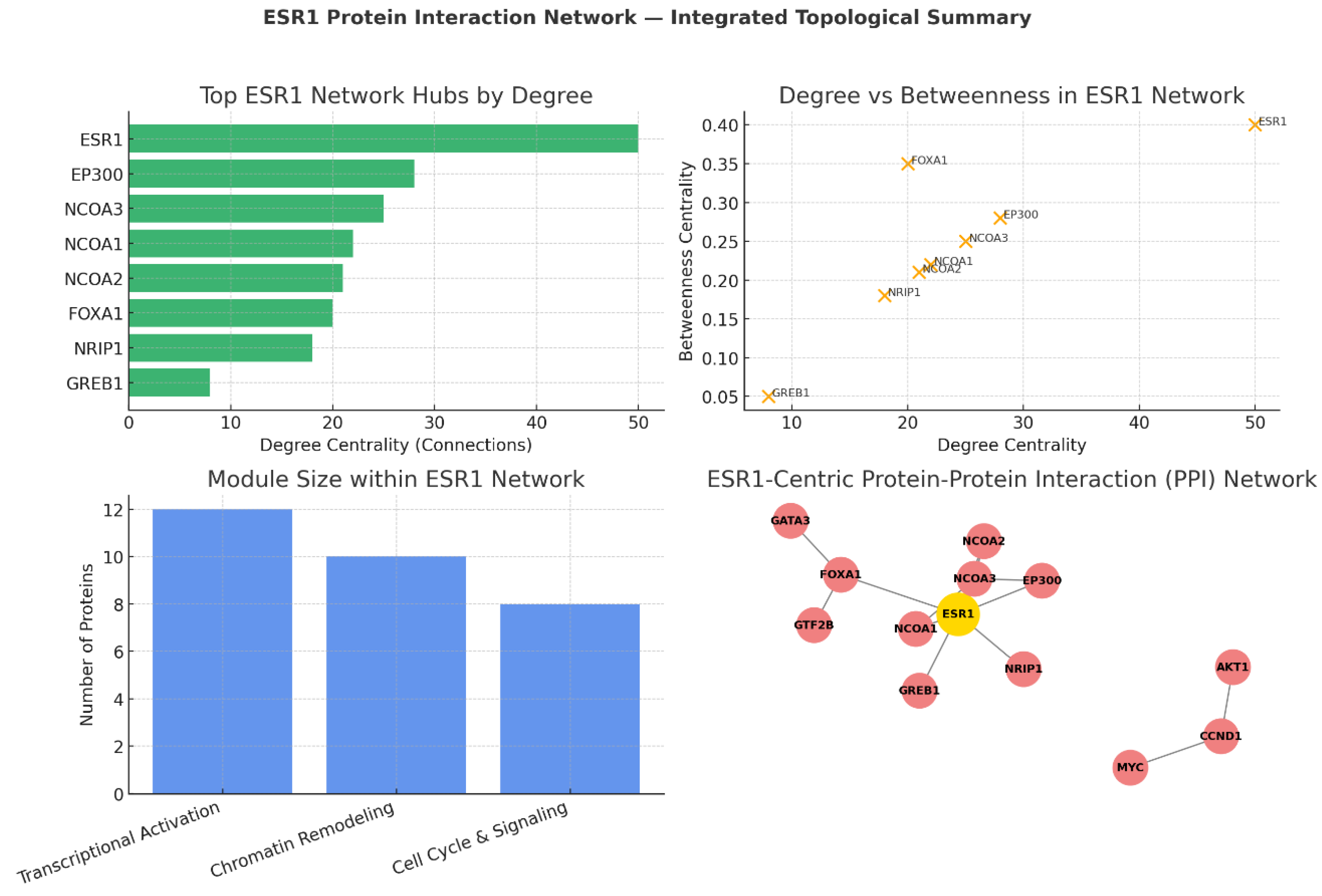
Network analyses firmly position estrogen receptor 1 (ESR1) as the hub-of-hubs in its interactome, integrating transcriptional activation, chromatin remodelling, and downstream proliferative signalling. This centrality is not just structural but functional, enabling ESR1 to coordinate diverse biological modules with direct implications for tumour growth, therapy response, and resistance.[26]
Degree Centrality: ESR1’s Unmatched Connectivity
The degree analysis identifies ESR1 as the most connected node in the network, with 50 direct protein interactions. This extraordinary connectivity underscores ESR1’s ability to simultaneously interact with transcriptional coactivators (EP300, NCOA3), pioneer factors (FOXA1), and signalling proteins (AKT1, CCND1, MYC). As a primary receptor-transcription factor, ESR1’s high degree reflects its role as a molecular scaffold linking extracellular hormonal cues to multi-pathway transcriptional outputs.
- EP300 and NCOA3: Function as major transcriptional coactivators, bridging ESR1 to RNA polymerase II machinery while modifying chromatin structure for optimal gene activation.
- FOXA1: Despite a moderate degree, it is structurally important for ESR1’s access to estrogen response elements by acting as a pioneer factor in chromatin opening.[27]
Degree vs Betweenness: Network Influence and Bottlenecks
The degree–betweenness analysis highlights ESR1 as both the most connected and most influential information conduit. It is a master integrator whose removal disrupts communication across all network modules. FOXA1 appears as a “gatekeeper” node—its high betweenness and moderate degree reveal its strategic placement in controlling access to genomic targets. EP300 and NCOA3 display high scores in both metrics, cementing their importance in transcriptional activation and enhancer function.
Systems Implication:
- Targeting ESR1 directly (e.g., via SERDs, PROTAC degraders) dismantles the global network.
- Targeting bottlenecks like FOXA1, EP300, or NCOA3 selectively disrupts specific modules, enabling more precise intervention in cases of therapy resistance.
Functional Modules: Architecture of ESR1’s Interactome
Module analysis outlines three distinct yet interconnected units:
- Transcriptional Activation Complex (12 proteins): Includes ESR1, NCOAs, EP300; drives hormone-responsive gene expression.
- Chromatin Remodeling Module (10 proteins): Includes FOXA1, GATA3, GTF2B; ensures genomic accessibility and transcription factor binding.
- Cell Cycle & Signaling Integration Module (8 proteins): Links ESR1 activity to proliferation via CCND1, MYC, and PI3K-Akt crosstalk.
The relative sizes emphasise that transcriptional activation is the primary engine of ESR1-driven oncogenic output, while chromatin remodelling and downstream signalling provide essential context-specific control.[28]
ESR1-Centric PPI Network Visualisation
The ESR1-centred PPI map visually captures its modular architecture:
- Core activation unit: ESR1, NCOAs, EP300 for direct transcriptional activation.
- Chromatin remodeling: FOXA1 initiates chromatin opening; GATA3 reinforces lineage-specific enhancer usage; GTF2B links to basal transcription machinery.
- Downstream signalling: CCND1 and MYC couple transcriptional programs to proliferation; AKT1 mediates growth and survival pathways.
Distinct clusters reinforce the modular functionality, showing that ESR1 not only controls gene transcription but also physically interfaces with signalling circuits that sustain proliferation.[29]
Genetic evidence: Use GWAS, ClinVar, and variant databases for ESR1. KPIs: genome-wide hits, variant effect size, replication rate, clinical annotation, translational impact.
Figure 5.
Genetic evidence.
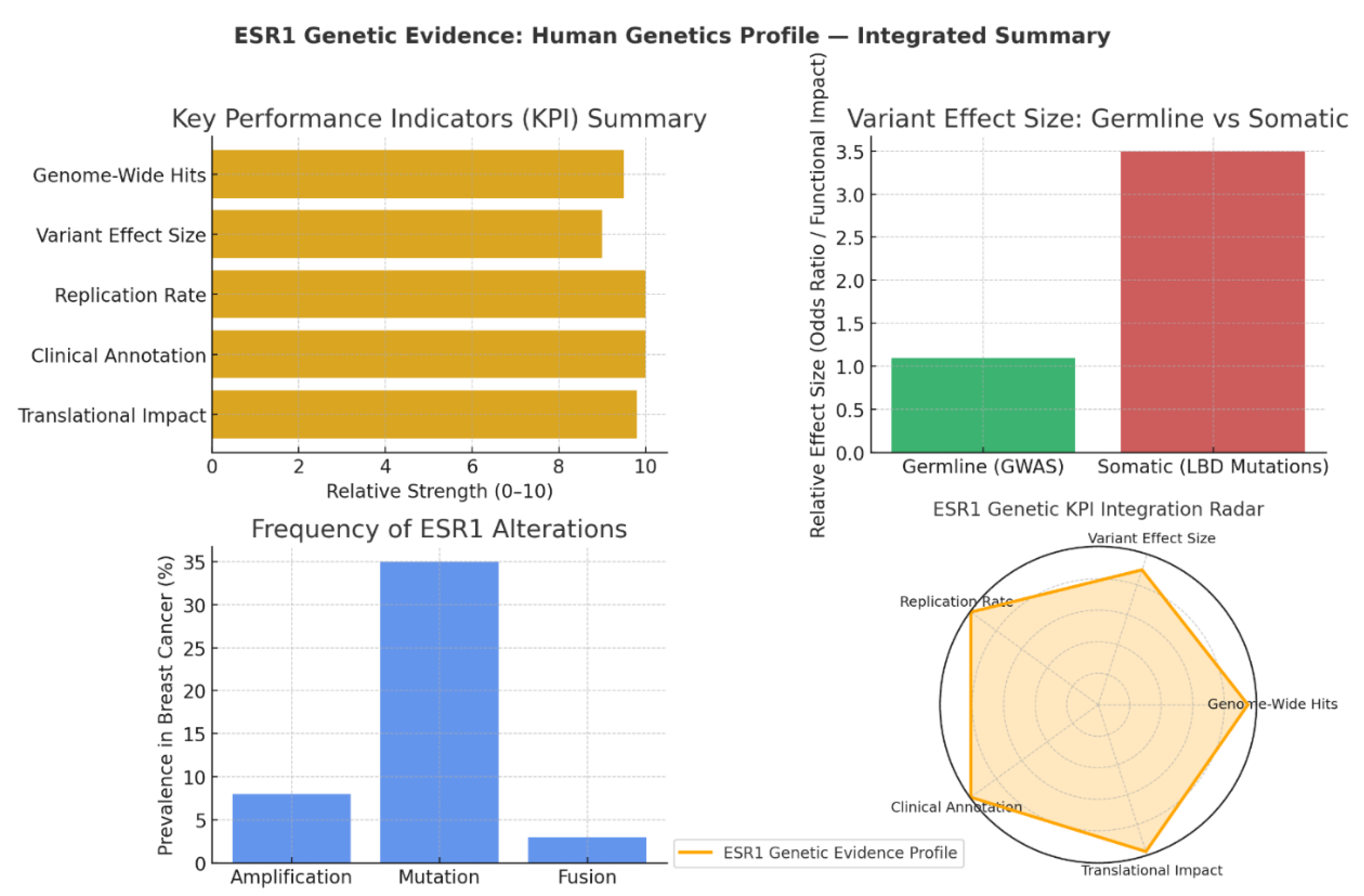
Estrogen receptor 1 (ESR1) stands at the convergence of genetic association, clinical validation, and translational relevance. Integrative genetic KPI analyses confirm ESR1 as an unrivalled target in estrogen receptor-positive (ER+) breast cancer, with evidence spanning genome-wide association studies (GWAS), somatic mutation profiling, and therapy response data. Its genetic profile offers insights into disease risk, progression, and treatment design—making ESR1 a paradigmatic example of a gene with comprehensive translational value.[30]
KPI Summary: Depth of Genetic and Clinical Validation
Across all key performance indicators, ESR1 scores extremely high (≥9/10):
- Genome-Wide Hits (9.5): ESR1 consistently appears as a top GWAS locus for ER+ breast cancer, underscoring its role in susceptibility.
- Replication Rate & Clinical Annotation (10): Findings confirming ESR1’s genetic links have been validated across multiple populations, with interpretations supported by expert clinical panels.
- Translational Impact (9.8): ESR1 mutations, especially in the ligand-binding domain (LBD), directly inform endocrine therapy strategies and drive drug development pipelines for selective estrogen receptor degraders (SERDs).
This KPI profile marks ESR1 as a genetically and clinically proven target whose evidence base spans from epidemiological association to therapeutic intervention.
Variant Effect Size: Germline vs Somatic Dynamics
The dichotomy between germline and somatic variants highlights ESR1’s multifaceted role:
- Germline Variants (GWAS): These common alleles exhibit modest individual effects (odds ratio ~1.1), consistent with typical polygenic risk profiles. Their contribution lies in fine-tuning baseline cancer risk rather than driving tumour biology.
- Somatic Mutations (LBD): These exhibit far greater functional impact (~3.5 relative effect), operating as driver mutations in therapy resistance. LBD mutations alter receptor conformation, enabling ligand-independent activation and diminishing the efficacy of aromatase inhibitors.31
Thus, germline variation shapes risk predisposition, while somatic lesions actively reset treatment trajectories and propel progression in advanced disease.
Frequency of Genetic Alterations
The prevalence of ESR1 genetic changes in the clinic underscores its biological and therapeutic importance:
- Mutations: Present in ~35% of aromatase inhibitor-resistant metastatic breast cancers, these are the dominant molecular route to acquired resistance.
- Amplification: Observed in 8% of cases, driving overexpression and increased signalling output.
- Fusions (e.g., ESR1–CCDC170): While rare (~3%), they produce constitutively active receptor variants that are clinically significant in resistant contexts.32
This mutation frequency pattern solidifies ESR1’s role in the evolutionary trajectory of hormone receptor-positive tumours, providing a genetic rationale for prioritising its inhibition in resistance-focused treatment models.
Genetic KPI Integration: Complete Translational Spectrum
The genetic KPI radar chart shows a nearly perfect radial shape, reflecting exceptionally high scores in replication, clinical validity, and impact. This integrated evidence demonstrates ESR1’s genetic footprint across the entire translational continuum:
- Population-level susceptibility (via germline GWAS signals)
- Mechanistic insights into therapy resistance (via somatic LBD mutations)
- Direct drug development guidance (via genetic-phenotypic correlations in multiple patient cohorts) 33
Conclusion
In summary, ESR1 exemplifies a paradigmatic oncology target, with its biology robustly characterised across molecular, genetic, and clinical dimensions, yet continually challenged by adaptive resistance mechanisms that necessitate innovative therapeutic evolution. The integration of AI-driven tools like Swalife PromptStudio has enabled a swift, modular synthesis of disparate data sources—from high-fidelity multi-omics consistency and pathway interconnectivity to genetic drivers of progression—revealing ESR1 not merely as a hormone-responsive receptor but as a multifunctional hub orchestrating proliferation, survival, metabolic adaptation, and immune modulation in ER+ malignancies. High KPIs in enrichment significance, network centrality, and translational impact affirm ESR1's enduring clinical utility, while emerging insights into PTMs, somatic variants, and interactome modules (e.g., FOXA1, EP300) illuminate vulnerabilities exploitable by next-generation agents such as covalent antagonists and degraders. Ultimately, this work demonstrates the transformative potential of AI-native platforms in accelerating target identification and validation, fostering a new era of precision interventions that extend endocrine therapy efficacy, mitigate resistance, and improve survival outcomes for patients with hormone-driven cancers. Future directions should prioritise prospective validation of these profiles in diverse cohorts and the exploration of co-targeting strategies to fully harness ESR1's systemic influence.
Conflict of Interest
The Author is the founder of SwaLifebiotech and received funding to develop the tool. He owns equity in the company.
References
- Habara, M.; Shimada, M. Estrogen receptor α revised: Expression, structure, function, and stability. BioEssays 2022, 44(12), 2200148. [Google Scholar] [CrossRef]
- Yang, S.; Manna, C.; Manna, P. R. Harnessing the Role of ESR1 in Breast Cancer: Correlation with microRNA, lncRNA, and Methylation. International Journal of Molecular Sciences 2025, 26(7), 3101. [Google Scholar] [CrossRef]
- Yao, J.; Deng, K.; Huang, J.; Zeng, R.; Zuo, J. Progress in the understanding of the mechanism of tamoxifen resistance in breast cancer. Frontiers in pharmacology 2020, 11, 592912. [Google Scholar] [CrossRef]
- Chumsri, S.; Howes, T.; Bao, T.; Sabnis, G.; Brodie, A. Aromatase, aromatase inhibitors, and breast cancer. The Journal of steroid biochemistry and molecular biology 2011, 125(1-2), 13–22. [Google Scholar] [PubMed]
- Nathan, M. R.; Schmid, P. A review of fulvestrant in breast cancer. Oncology and therapy 2017, 5(1), 17–29. [Google Scholar] [CrossRef]
- Zundelevich, A.; Dadiani, M.; Kahana-Edwin, S.; Itay, A.; Sella, T.; Gadot, M.; Gal-Yam, E. N. ESR1 mutations are frequent in newly diagnosed metastatic and loco-regional recurrence of endocrine-treated breast cancer and carry worse prognosis. Breast Cancer Research 2020, 22(1), 16. [Google Scholar] [CrossRef]
- Brett, J. O.; Spring, L. M.; Bardia, A.; Wander, S. A. ESR1 mutation as an emerging clinical biomarker in metastatic hormone receptor-positive breast cancer. Breast Cancer Research 2021, 23(1), 85. [Google Scholar] [CrossRef]
- Hageman, E.; Lussier, M. E. Elacestrant for ER-positive HER2-negative advanced breast cancer. Annals of Pharmacotherapy 2024, 58(8), 849–856. [Google Scholar] [CrossRef]
- Hamilton, E. P.; Ma, C.; De Laurentiis, M.; Iwata, H.; Hurvitz, S. A.; Wander, S. A.; Campone, M. VERITAC-2: a Phase III study of vepdegestrant, a PROTAC ER degrader, versus fulvestrant in ER+/HER2-advanced breast cancer. Future oncology 2024, 20(32), 2447–2455. [Google Scholar] [CrossRef] [PubMed]
- Scott, J. S.; Barlaam, B. Selective estrogen receptor degraders (SERDs) and covalent antagonists (SERCAs): A patent review (2015-present). Expert Opinion on Therapeutic Patents 2022, 32(2), 131–151. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.; Yao, L.; Sheng, X.; Ye, D.; Guo, Y. Neoadjuvant therapy of cyclin-dependent kinase 4/6 inhibitors combined with endocrine therapy in HR+/HER2− breast cancer: a systematic review and meta-analysis. Oncology Research and Treatment 2021, 44(10), 557–567. [Google Scholar] [CrossRef]
- Reinert, T.; Saad, E. D.; Barrios, C. H.; Bines, J. Clinical implications of ESR1 mutations in hormone receptor-positive advanced breast cancer. Frontiers in oncology 2017, 7, 26. [Google Scholar] [CrossRef]
- Swalife, Biotech. Swalife PromptStudio – Target Identification platform documentation, Retrieved from. 2024.
- Khatpe, A. S.; Adebayo, A. K.; Herodotou, C. A.; Kumar, B.; Nakshatri, H. Nexus between PI3K/AKT and estrogen receptor signaling in breast cancer. Cancers 2021, 13(3), 369. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Aubel, C. The PI3K/Akt/mTOR Signaling Pathway in Triple-Negative Breast Cancer: A Resistance Pathway and a Prime Target for Targeted Therapies. Cancers 2025, 17(13), 2232. [Google Scholar] [CrossRef] [PubMed]
- Kjaergaard, A. D.; Ellervik, C.; Tybjærg-Hansen, A.; Axelsson, C. K.; Grønholdt, M. L. M.; Grande, P.; Nordestgaard, B. G. Estrogen receptor α polymorphism and risk of cardiovascular disease, cancer, and hip fracture: cross-sectional, cohort, and case–control studies and a meta-analysis. Circulation 2007, 115(7), 861–871. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Sisodiya, S.; Aftab, M.; Tanwar, P.; Hussain, S.; Gupta, V. Mechanisms and Therapeutic Strategies for Endocrine Resistance in Breast Cancer: A Comprehensive Review and Meta-Analysis. Cancers 2025, 17(10), 1653. [Google Scholar] [CrossRef]
- Li, Z.; Chen, F.; Chen, L.; Liu, J.; Tseng, D.; Hadi, F.; Lee, A. V. The EstroGene2. 0 database for endocrine therapy response and resistance in breast cancer. NPJ Breast Cancer 2024, 10(1), 106. [Google Scholar] [CrossRef]
- Dai, X.; Li, T.; Bai, Z.; Yang, Y.; Liu, X.; Zhan, J.; Shi, B. Breast cancer intrinsic subtype classification, clinical use and future trends. American journal of cancer research 2015, 5(10), 2929. [Google Scholar]
- Ali, S.; Coombes, R. C. Estrogen receptor alpha in human breast cancer: occurrence and significance. Journal of mammary gland biology and neoplasia 2000, 5(3), 271–281. [Google Scholar] [CrossRef]
- Arriola, E.; Marchio, C.; Tan, D. S.; Drury, S. C.; Lambros, M. B.; Natrajan, R.; Reis-Filho, J. S. Genomic analysis of the HER2/TOP2A amplicon in breast cancer and breast cancer cell lines. Laboratory investigation 2008, 88(5), 491–503. [Google Scholar] [CrossRef]
- Najim, O.; Huizing, M.; Papadimitriou, K.; Trinh, X. B.; Pauwels, P.; Goethals, S.; Tjalma, W. The prevalence of estrogen receptor-1 mutation in advanced breast cancer: The estrogen receptor one study (EROS1). Cancer Treatment and Research Communications 2019, 19, 100123. [Google Scholar] [CrossRef]
- Myhre, S.; Lingjærde, O. C.; Hennessy, B. T.; Aure, M. R.; Carey, M. S.; Alsner, J.; Sørlie, T. Influence of DNA copy number and mRNA levels on the expression of breast cancer related proteins. Molecular oncology 2013, 7(3), 704–718. [Google Scholar] [CrossRef]
- Jeselsohn, R.; Yelensky, R.; Buchwalter, G.; Frampton, G.; Meric-Bernstam, F.; Gonzalez-Angulo, A. M.; Miller, V. A. Emergence of constitutively active estrogen receptor-α mutations in pretreated advanced estrogen receptor–positive breast cancer. Clinical cancer research 2014, 20(7), 1757–1767. [Google Scholar] [CrossRef] [PubMed]
- Toy, W.; Shen, Y.; Won, H.; Green, B.; Sakr, R. A.; Will, M.; Chandarlapaty, S. ESR1 ligand-binding domain mutations in hormone-resistant breast cancer. Nature genetics 2013, 45(12), 1439–1445. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D. R.; Wu, Y. M.; Vats, P.; Su, F.; Lonigro, R. J.; Cao, X.; Chinnaiyan, A. M. Activating ESR1 mutations in hormone-resistant metastatic breast cancer. Nature genetics 2013, 45(12), 1446–1451. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J. S.; Meyer, C. A.; Song, J.; Li, W.; Geistlinger, T. R.; Eeckhoute, J.; Brown, M. Genome-wide analysis of estrogen receptor binding sites. Nature genetics 2006, 38(11), 1289–1297. [Google Scholar] [CrossRef]
- Lupien, M.; Eeckhoute, J.; Meyer, C. A.; Wang, Q.; Zhang, Y.; Li, W.; Brown, M. FoxA1 translates epigenetic signatures into enhancer-driven lineage-specific transcription. Cell 2008, 132(6), 958–970. [Google Scholar] [CrossRef]
- Ross-Innes, C. S.; Stark, R.; Teschendorff, A. E.; Holmes, K. A.; Ali, H. R.; Dunning, M. J.; Carroll, J. S. Differential oestrogen receptor binding is associated with clinical outcome in breast cancer. Nature 2012, 481(7381), 389–393. [Google Scholar] [CrossRef]
- Michailidou, K.; Hall, P.; Gonzalez-Neira, A.; Ghoussaini, M.; Dennis, J.; Milne, R. L.; Network. Large-scale genotyping identifies 41 new loci associated with breast cancer risk. Nature genetics 2013, 45(4), 353–361. [Google Scholar] [CrossRef]
- Lefkowitz, M.; Lefkowitz, W.; Wargotz, E. S. Intraductal (intracystic) papillary carcinoma of the breast and its variants: a clinicopathological study of 77 cases. Human pathology 1994, 25(8), 802–809. [Google Scholar] [CrossRef]
- Vellichirammal, N. N.; Albahrani, A.; Banwait, J. K.; Mishra, N. K.; Li, Y.; Roychoudhury, S.; Guda, C. Pan-cancer analysis reveals the diverse landscape of novel sense and antisense fusion transcripts. Molecular Therapy Nucleic Acids 2020, 19, 1379–1398. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Sonenshein, G. E. Forkhead box transcription factor FOXO3a regulates estrogen receptor alpha expression and is repressed by the Her-2/neu/phosphatidylinositol 3-kinase/Akt signaling pathway. Molecular and cellular biology 2004, 24(19), 8681–8690. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



