
Submitted:
02 October 2025
Posted:
03 October 2025
You are already at the latest version
Abstract
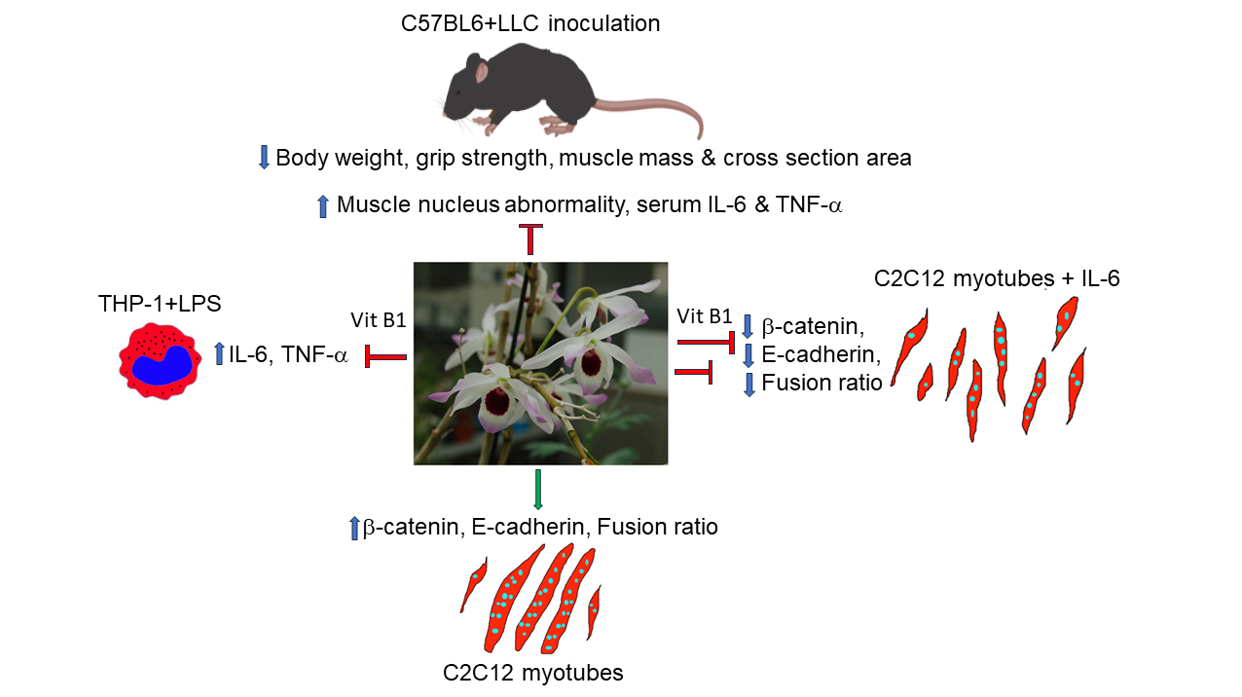
In this report, we showed that oral administration of Dendrobium Taiseed Tosnobile (DTT, also known as Taiwan Emperor No.1) allowed Lewis Lung Carcinoma (LLC) tumor-bearing mice to maintain body weight and grip strength in a dose-dependent manner. Histological analysis showed that treatment of DTT water extracts significantly reduced muscle fiber damage via inducing muscle regeneration and improved the cross-sectional area of the rectus femoris, soleus, and gastrocnemius of LLC tumor-bearing C57BL/6 female mice. Further studies revealed that DTT water extracts also reduced the expression of inflammatory cytokines such as IL-6 and TNF-α both in vitro and in vivo. Other analyses showed that DTT water extracts promoted the differentiation of C2C12 myoblast with or without IL-6 via maintaining myosin heavy chain (MyHC) level, suggesting DTT water extracts against muscle wasting via multiple manners. Interestingly, vitamin B1 was identified as an ingredient in DTT water extracts through an HPLC analysis. The vitamin B1 ameliorates IL-6 but not TNF- generation of active THP-1 cells and the protects C2C12 myotubes against IL-6. Further studies showed that DTT and vitamin B1 promoted the multi-nucleus fusion step of the C2C12 differentiation by inducing the expression of E-cadherin-catenin with or without IL-6 treatment. In summary, DTT water extracts protect muscle cells under cancer conditions through direct and indirect mechanisms, with vitamin B1 being a key functional ingredient that reduces IL-6 generation and aids muscle cell fusion against IL-6 treatment.
Keywords:
muscle wasting
; Dendrobium Taiseed Tosnobile
; IL-6
; E-cadherin
; MyHC
; C2C12
; multinucleation
1. Introduction
Skeletal muscle is the most abundant tissue of healthy people (about 45±5% of body mass) [1]. Individual skeletal muscle comprises bundles of muscle fibers enclosed through connective tissue sheathing [2]. The main component of each muscle fiber includes several striated bundles formed by the fusion of differentiated myocytes [3]. The main characteristics of myotubes are mature myofibrils containing multiple myofilaments, including actin, elastin, and MyHC [4]. Skeletal muscle fibers are generated by the repeated fusions of myocytes-myocytes and myocytes-myotubes during embryonic development. In adults, the fusion of myogenic satellite cells is essential to promote the growth and repair of myofibers after injury. The fusion of myocytes and myotube is a complicated process that includes the remodeling of both membrane and cytoskeleton, and dozens of proteins involved in this process, such as Interleukin-4 (IL-4), Vascular cell adhesion protein (v-CAM), myoferlin, MEK/ERK, and β-catenin [5,6,7,8,9]. The loss of function of these proteins affected the development or regeneration of muscle fibers. Skeletal muscles are essential in locomotion control, organ protection, protein reservoir, posture maintenance, and daily physiological activities [10]. The optimal size and function of muscles depend on the balance between biosynthesis and the destruction of muscle cells [11].
Muscle wasting (or muscle atrophy), a condition of gradual loss of muscle mass and function, is a vital characteristic found in older adults and patients with genetic defects, chronic diseases, or cancer cachexia [12,13]. Cachexia is the most critical comorbidity found in 40-80% of cancer patients, particularly at advanced stages of the disease, depending on the type and size of cancer [14], The incidence of cachexia makes the management of cancer patients more complicated, including poor response to treatments, poor appetite, and decreased body strength, which generally leads to a dramatic decrease in quality of life [15,16,17]. The consequence of cancer cachexia is medical futility and a poor survival rate, causing more than 25% of cancer deaths [18]. The pathogenetic characteristics of cancer cachexia are fat mass loss and skeletal muscle accompanied by anorexia; however, muscle wasting is considered the most critical trait since it is responsible for most of the symptoms associated with cancer cachexia and poor prognostic factor of survival [19,20,21]. To date, the underlying mechanisms of cancer cachexia are mainly involved in the up-regulation of inflammatory cytokines such as TNF-α and IL-6, which might correlate with the imbalance between protein synthesis and protein degradation in muscle cells [22,23,24,25]. Some medications, including megestrol acetate or Adlumiz® (a ghrelin activator), are appetite stimulants in which food intake and weight loss prevention have been reported in cancer cachexia patients; however, the improved effects were thoroughly validated and several adverse effects were reported [26,27,28,29]. In addition to treatment with appetite stimulators, muscle wasting attenuation was considered an alternative strategy for anticachectic treatments.
Herb medicine, an integral part of traditional Chinese medicine, has attracted considerable attention due to its distinct efficacy in treating various diseases [30]. Dendrobii (Shi-Hu), a medicinal orchid (family: Orchidaceae) classified as high-grade herbs in “shen nong ben cao jing,” have been widely used for improving health, such as antipyretic, ophthalmic, and tonic benefits [31,32,33]. Herba dendrobii exhibits multiple health benefits and is a valuable herb on the market ($ 3000/kg); however, due to the rare distribution in the natural environment and slow growth rate, Herba dendrobii yield was relatively low [34,35]. To overcome this obstacle, a new species, D. Taiseed Tosnobile (DTT), was developed by crossbreeding two medicinal species, D. tosaense and D. nobile, by the Taiwan Seed Improvement and Propagation Station [36]. DTT marks the high production and easy cultivation comparison with medicinal dendrobii [36]. The following research showed DTT’s biological safety and immunomodulation functions [36,37]. However, it remains to be elucidated whether DTT exerts other health benefits. Based on previous studies and pharmacopeia records, we hypothesize that Dendrobium’s immunomodulatory capability may facilitate cancer patients’ rehabilitation. This report investigates the effects of DTT water extracts on cancer-induced muscle wasting and potential underlying mechanisms. Our results showed that administering DTT water extracts effectively attenuated muscle wasting in LLC tumor-bearing mice. Further research revealed that DTT water extracts protected myotubes from injury directly and indirectly.
2. Materials and Methods
2.1. DTT Extraction and HPLC Procedure
Dendrobium Taiseed Tosnobile (DTT, Supp. Figure S1) was kindly provided by Dr. Chi-Luan Wen of the Taiwan Seed Improvement and Propagation Station (Taichung, Taiwan). The dried and powdered DTT stem was thoroughly mixed with pure water. The resulting mixture was shaken with an ultrasonic oscillator for approximately 60-90 minutes, filtered with 3M filter paper, and collected the filtrate. In addition, the residue obtained by filtration was used to repeat the above-mentioned pure water mixing-ultrasonic vibration-filtration steps three times. Then, all the collected filtrate is concentrated under reduced pressure to remove the pure water, thereby obtaining the dendrobium water extract. The HPLC fingerprint of DTT water extracts was provided by the Health Technology Center of Chung-Shan Medical University (https://rc.csmu.edu.tw/p/412-1018-23.php). Briefly, the dry matter weight of DTT (10 g) was dissolved in deionized water (1 L) and settled overnight in a refrigerator at 4 °C. The precipitate was filtered with 0.45 μm filter paper, and the filtrate was concentrated with a rotary evaporator. The net content of DTT (0.5 g) was mixed with 5 ml of sterile water in a tube. The chromatographic separation of DTT was carried out using a Shimadzu HPLC- Diode Array Detector system and equipped with an inertsil Octadecylsilyl-4 column chromatography (250 mm × 4.6 mm, five μm) column chromatography. The injection volume, flow rate, and detection wavelength were 10 μl, 1.0 ml/min, and 254 nm, respectively. Separation was performed using a gradient elution of water and acetonitrile based on the following procedure: The mobile phase used for the analysis was a gradient elution system consisting of solvent A (water, 0.1% acetic acid) and solvent B (1% acetic acid in acetonitrile). The gradient program was as follows: 0-2 min, 1% solvent B; 2-22 min, 30% solvent B; 22-32 min, 70% solvent B; 32-45 min, 1% solvent B.
2.2. Animal Model
Adult C57BL/6 (C57 black 6) mice (8 weeks of age, 20-22 g, female) were purchased from Lasco (http://www.biolasco.com.tw/index.php/tw/), followed by housing and acclimating to their cages and human handling for 7 days before the experiments. The mice were then housed in a room maintained at 22 ± 2 ◦C under a 12h light/dark cycle (5 mice per cage, lights on at 7:00 am). Laboratory Rodent Diet 5001 (Texas, USA), which was composed of 57.5% carbohydrates, 28.9% protein, and 13.6% fat, was applied as a daily routine diet. The animal experiments were conducted by the Guide for the Care and Use of Laboratory Animals prepared by the US National Institutes of Health and procedures approved by the Institutional Animal Care and Use Committee of Chung-Shan Medical University (CSMU IACUC No. 2422, 2021/03/02) and the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guideline. The LLC tumor-induced muscle-wasting model was conducted based on the previous study [38]. Briefly, 20 C57BL/6 mice were randomly assigned using a computer-based random order generator to 4 groups (5 mice per group with equal mean weights): regular, LLC, L+DWL (DTT water extracts low dose, 75 mg/kg), L+DWH (DTT water extracts high dose, 150 mg/kg). The mice were subcutaneously injected in the right inguinal region with 100 μl (1 × 105 cells) of LLC cells except for the standard group. Seven days after the injection of LLC cells, all mice had developed tumors, as confirmed by palpation of the injection site (approximately 0.5 cm in diameter). The tumor-bearing mice were then gavagely administered water, a low or a high dose of DTT water extracts daily for 18 consecutive days until the day of sacrifice.
2.3. Food Intake, Body Weight, and Grip Strength
Body weight and food intake were evaluated weekly by weighing the food and the animals before gavage administration of water or DTT water extracts. Both body weight and food intake were expressed in grams/week. The grip strength tests were performed using a BIO-GT3 tester (BIOSEB, FL, USA) according to the guidelines of the International Mouse Phenotyping Consortium [39]. Each mouse was placed on a metal grid connected to a horizontal force sensor. After the forelimbs of the mouse were stably grabbed on the grid, the tail was pulled backward at a 45◦ angle until the mouse was released from the grid, and the strength of the limb was recorded. The test for each mouse was repeated at least three times, and the results were averaged for analysis. On the day of the endpoint, all mice were sacrificed in a CO2 chamber. Organs (gastrocnemius and soleus, lung) and tumors were removed, weighed, and recorded.
2.4. Histological Analysis
The mice were sacrificed 28 days after the LLC implant for morphometric analysis, and the left hind leg was collected. The leg tissue samples were then dehydrated, embedded in paraffin, sectioned at 3 μm, mounted on positively charged microscope slides (Thomas Scientific, NJ, USA), and dried in a 50 °C oven for 30 min. For H/E staining, indicated muscle tissue sections were stained according to the previously described and manufacturer’s protocol (https://www.leicabiosystems.com). Fibers’ diameters, area, and nucleus numbers were measured in at least three randomly selected microscope fields and calculated using Fiji Image J software (https://imagej.net/software/fiji/downloads). For IHC staining, the proper hind muscle tissues were fixed in 10% formalin overnight at 25 °C. The Paraffin-embedded muscle was cut into 3 µm sections using a microtome and dried for two h at 37 °C or overnight at room temperature. The muscle sections were then deparaffinized and rehydrated using the following washing steps: 5 min xylene, three times; 5 min 100% ethanol, three times; 5 min 95% ethanol and 5 min 75% ethanol, followed by rinsing with distilled water. Next, sodium citrate buffer (10 mM; pH 6.0) was incubated with muscle slides using a microwave twice for 10 min for antigen retrieval. Muscle sections were incubated with 2% peroxidase at room temperature for 15 min, blocked with 5% BSA (w/v) at room temperature for 30 min, and incubated with anti-pan-MyHC antibody (Elabscience, Houston, Texas, USA) overnight at 4 °C. The tissue sections were then incubated with a biotinylated anti-rabbit secondary antibody for one hour at room temperature. Streptavidin-horseradish peroxidase and peroxidase substrate solution were added to muscle sections at room temperature for 10 min for signal development. IHC stains were analyzed under a light microscope.
2.5. Cell Culture, Treatments, and Transfection
The C2C12 myoblasts (CRL-1772) were purchased from ATCC (VA, USA), C2C12 cells were maintained in DMEM (Dulbecco’s Modified Eagle’s Medium) containing 10% FBS (Fetal Bovine Serum, Gibco standard, Mexico) at 37 ◦C with 5% CO2. C2C12 myoblasts were seeded in 100 mm dishes (1×106 cells/dish) or 12-well plates (5×104 cells/well) in myogenic differentiation. The next day, the growth medium was replaced with a differentiation medium containing 2% horse serum with or without DTT water extracts. Six days later, the differentiated C2C12 myotubes were collected and subjected to the indicated assay. In the IL-6 treatment experiment, the IL-6 (100 ng/ml) was added to the differentiated C2C12 myotubes 6 days after replacing them with differentiated medium for another 48 h, followed by collecting and subjecting to the indicated assay. For transfection, the scramble or mouse E-cadherin siRNA (Small Interference RNA) purchased from Santa Cruz (Dallas, Texas, USA) were transfected into C2C12 cells with lipofectamine 2000 reagent twice within 48 h. The scramble RNA (scRNA) or E-cadherin siRNA transfectants were subjected to myogenic differentiation for another 6 days followed by indicated experiments.
2.6. Giemsa Staining and Measurement of Myotube Diameters
The differentiated C2C12 cells were washed with 1XPBS twice and fixed in absolute methanol for 5 min, after which the myotubes were stained with Giemsa dilution buffer (containing 1 mM KH2PO4) for 20 min and then observed by an optical microscope (SAGE, Taipei, Taiwan). For each condition, six pictures were randomly taken from each well of the six-well plates. The diameters of three different sites in each myotube were measured using Fiji Image J software (https://imagej.net/software/fiji/downloads). At least 200 myotubes were measured in one well, and the results were averaged for analysis.
2.7. Immunostaining
The differentiated C2C12 cells were fixed with 2% paraformaldehyde for 25 min before incubation with 0.05% Triton X-100 for 20 min and then blocked with 1% BSA for 60 min. Cells were probed with an anti-pan-MyHC antibody (E-AB-22021, Elabscience, Texas, USA) overnight at 4 °C, followed by incubation with a TRITC or FITC-conjugated goat anti-rabbit IgG antibody (Sigma, St. Louis, MO, USA, diluted 1:200) for one hour at 37 oC, washing with PBS three times and staining with DAPI for five minutes. The expression and location of target proteins were observed with a confocal laser scanning microscope.
2.8. Cytokines Analysis
Whole blood was drawn from the retroorbital sinus in experimental and control mice and collected in tubes. Blood was allowed to stay overnight at room temperature to clot, and serum (supernatant) was obtained by centrifugation at 3000 rpm at 4 °C for 5 min. The ebioscience ELISA kits (ThermoFisher, MA, USA) were grouped to determine the levels of TNF-α and IL-6 in collected serum samples using anti-TNF-α or IL-6 antibodies, respectively. All samples were analyzed in duplicate. For in vitro assay, the THP-1 cells (Cat. No. TIB-202, ATCC, VA, USA) were treated with 20 ng/ml phorbol myristate acetate (Sigma, MO, USA) for 48 h, followed by treatment with either LPS (LipoPolySaccharide) (100 ng/ml) alone or together with DTT water extracts for another eight hours. The medium was then collected and subjected to an ELISA assay kit using anti-mouse IL-6 or TNF-α antibody (ebioscience, Taipei, Taiwan), respectively.
2.9. Protein Extraction and Western Blot Analysis
The control or experimental cells were lysed in protein extraction solution followed by Bradford assay to determine protein concentrations. All samples were adjusted to an equal concentration of 50 μg/lane. The samples were then separated by 7-14% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane. After being blocked with 5% skim milk for three h, membranes stayed overnight at 4 °C with primary antibodies against pan-MyHC (E-AB-22021, Elabscience, Texas, USA) or actin (A2228, Merck, NJ, USA), GAPDH (sc-32233, Santa cruz, CA, USA), E-cadherin (GTX100443, GeneTex, CA, USA), β-catenin (ab6301, Abcam, MA, USA), respectively. The membranes were then incubated with horseradish peroxidase-conjugated goat anti-rabbit or mouse immunoglobulin G (1:4000 dilution; Santa cruz, CA, USA). The blots were developed using a Chemiluminescent Substrate kit (PerkinElmer, MA, USA).
2.10. Statistical Analysis
The statistical significance of differences was determined using Student’s t-test or ANOVA (Analysis of variance) with post hoc multiple comparison test (Tukey’s test) using Prism ver. 7.0 (GraphPad, CA, USA). A value of p< 0.05 was considered significant. For all analyzes, *p < 0.05, **p < 0.01 or ***p < 0.001 compared to the control group.
3. Results
3.1. DTT Water Extracts Attenuate the Loss of Tumor-Free Body Mass in LLC Tumor-Bearing Mice
The LLC tumor-implanted mice model, as described in the materials and methods section, was utilized in this study to investigate whether DTT water extracts mitigate cancer-associated muscle wasting. On day 7 after LLC implantation, tumor-bearing mice received DTT water extracts orally every day until day 28 (Figure 1A). At the experiment’s end, each mouse’s net body mass was determined after the tumor removal. The results showed no significant differences in total weight and food intake between the experimental and control groups (Figure 1B,C). However, the mean net body mass of mice in LLC was nearly 20% lower compared to that of the control (Figure 1D). Interestingly, the administration of either low (75 mg/kg) or high (150 mg/kg) dose of DTT water extracts effectively increased 10-15% of net body mass compared to LLC tumor-bearing mice (Figure 1E) with no dose effects observed. This result raises the possibility that a minimal dose of DTT water extracts in this experiment was beyond the working concentration to recover weight loss in a tumor-bearing mouse. Furthermore, the results also showed that the net body mass of DTT water extracts treated mice on day 28 was on average 40% higher compared to day 0 (Figure 1E). In comparison, control mice only increased by 25%; Since the food intake was similar between the different groups, this result suggests that the DTT water extracts exhibit a better conversion ratio of food intake to body mass.
3.2. DTT Water Extracts Recover Muscle Mass and Grip Strength in LLC Tumor-Bearing Mice
Our results showed that the DTT water extracts significantly attenuated the body mass loss in LLC tumor-bearing mice. Since LLC tumors were known to prime continuous decline of muscle mass and eventually impede its functionality [40], we investigated whether DTT water extracts ameliorated body mass loss via mass or functional muscle restoration in LLC tumor-bearing mice. First, we analyzed the effects of DTT water extracts on the grip strength of LLC tumor-bearing mice. The grip strengths of tumor-bearing mice decreased continuously as the experiment progressed and were reduced by 30-35% compared to the control on the day of sacrifice (Figure 2A). At the same time, the administration of DTT water extracts promoted LLC tumor-bearing mice maintained a higher grip strength throughout the experiment. On day of sacrifice, the LLC tumor-bearing mice coupled with DTT water extracts recovered about 20-25% grip strength compared to that of LLC tumor alone (Figure 2A). Furthermore, the measurement of F/W (force-over-body-weight) value on the day of sacrifice showed a reduction of F/W value in LLC tumor-bearing mice by 40% compared to the control (Figure 2B). Interestingly, administration of either low and high concentrations of DTT water extracts recovered F/W values in mice carrying LLC tumors almost similar to that of the control (increasing by 60-65% compared to that of the LLC tumor alone) without dosage effect (Figure 2B), resembled that of recovery of body mass (Figure 1E). The results implied that DTT water-extracts maintained muscle functionality by alleviating muscle atrophy in LLC tumor-bearing mice. To confirm this, mice’s muscle mass variance in various experimental conditions was investigated. As excepted, remarkable mass reductions of the gastrocnemius and soleus muscle were observed by 22-31% in the presence of the LLC tumor compared to those of the control mice (Figure 2C,D), while the administration of high (150 mg/kg) but not low (75 mg/kg) DTT water extracts significantly increased the mass of the gastrocnemius and soleus by 20-25% in LLC tumor-bearing mice, almost the same weight as that of the control (Figure 2C,D). The results suggested that DTT water extracts restored muscle functionality in LLC tumor-bearing mice by maintaining muscle mass.
3.3. DTT Water Extracts Recover the Cross-Section Area of the Hindlimb in LLC Tumor-Bearing Mice
To clarify how DTT water-extracts sustained muscle mass, we analyzed the effects of DTT water extracts on the CSA (cross-section area) of the hindlimb in tumor-bearing mice from LLC. The results of HE staining of the thigh and calf musculature were atrophic in LLC tumor mice, accompanied by reductions in perimysium, while the musculatures of LLC tumor-bearing mice treated with DTT water extracts resembled those of the control (Figure 3A,B). Further analysis revealed that the cross-section area of the rectus femoris (indicated by a yellow line) of tumor-bearing mice with LLC was smaller than that of the control (reduction of 32-36%) (Figure 3A,C). The addition of high (150 mg/kg) but not low (75 mg/kg) concentrations of DTT water extracts restored the loss of rectus femoris by 30-33% in LLC tumor-bearing mice and showed no significant difference compared to the control (Figure 3C). Similar results were also observed in gastrocnemius (indicated by the red line) and soleus (indicated by the green line), the CSA of these two musculatures was reduced by the LLC tumor by 24-30% while almost restored to the control level when engaged with a high concentration of DTT water extracts (Figure 3B,D,E). These results suggested a requirement for higher concentrations of DTT water extracts to combat muscle wasting induced by LLC tumors, and this might be due to the low active ingredients of DTT water extracts.
3.4. DTT Water Extracts Alleviate Myofiber Atrophy Induced by LLC Tumor
Next, we quantitatively measured the morphometric variations of the muscle fibers in mice under various experimental conditions. The results showed a significant reduction in the size of the perimysium. They were accompanied by an irregular shape of the myofibers of rectus femoris, gastrocnemius, and soleus in LLC tumor-bearing mice compared to the control (Supp. Figure S2A). The reduction ratio of the average minimum diameter (minferet) in these three muscle types of LLC tumor-bearing mice was 80%, 19%, and 28%, respectively, compared to that of the control (Figure 4A–C). Furthermore, the administration of low and high concentrations of DTT water extracts significantly increased the size of the myofibers of both the rectus femoris and soleus muscle by 65% and 26%, respectively, and almost fully recovered that of the gastrocnemius muscle in tumor-bearing mice with LLC (Figure 4A–C). The status of the nuclei of different musculatures was then investigated to prove myofiber atrophy, and the number and aggregation of the nuclei indeed increased in mice with LLC tumors compared to the control (Supp. Figure S2B). Quantitative results showed that the number of nuclei of the rectus femoris muscle in the LLC tumor-bearing increased by 85% compared to the control; similar patterns were also observed in the gastrocnemius and soleus muscle, which increased by 72% and 98%, respectively (Figure 4D–F). Administration of both low- and high-dose DTT water extracts in LLC tumor-bearing mice effectively reduced the abnormal nuclei number of rectus femoris to the level of control, while only high-dose DTT water extracts reduced the abnormal nuclei number of the gastrocnemius muscle (reduced by 15-21% vs. LLC tumor alone). In contrast, the low dose of DTT water extracts treatment only effectively reduced abnormal nuclei numbers in soleus muscle (reduced by 20-25%) in LLC tumor-bearing mice (Figure 4D–F). Our results indicated that DTT water extracts recover muscle wasting by alleviating myofiber atrophy in LLC tumor-bearing mice. Furthermore, the variations in recovery among different types of muscle might be due to the multiple ingredients in the DTT water extracts. The nucleus pattern was investigated to uncover the underlying mechanisms of recovery since it was reported that the centralized nucleus served as a hallmark of muscle regeneration [41]. The results showed the average ratio of the centralized nucleus of rectus femoris, gastrocnemius, and soleus were 2.8%, 2.3%, and 1.7%, respectively, in the control mice group. The ratio dropped to 1.0%, 1.7%, and 0.6% in the LLC-tumor mice group (Supp. Figures S2C and S4G–I), indicating a reduction of muscle regeneration in a tumorous condition. Interestingly, the administration of both low and high doses of DTT water extracts significantly increased the ratio of the centralized nucleus in rectus femoris, gastrocnemius, and soleus 2-3 folds in LLC-tumor mice group in a dose-dependent manner, nearly equal or higher than the control mice group (Figure 4G–I). These data strongly suggested that the induction of muscle regeneration might be one pathway of DTT water extracts against muscle wasting that is primed by tumorous conditions. Next, we investigated the architectures of muscle fibers in LLC tumor-bearing control mice with or without treatment with DTT water extracts. The results showed that the area of the soleus/plantaris region of muscle cells decreased significantly in LLC tumor-bearing mice. At the same time, the administration of DTT water extracts recovered the architecture of muscle fibers (Supp. Figure S3), which confirmed that the loss of muscle weight or area could be due to reductions of muscle cells.
3.5. DTT Water Extracts Attenuate the Levels of Inflammatory Cytokines IL-6 and TNF-α Both In Vitro and In Vivo
Both IL-6 and TNF-α were reported to be induced by tumors in both animal models and clinical investigations [42,43]. These two cytokines also showed the ability to induce muscle atrophy [24,25]. Therefore, we decided to examine the relationships between DTT water extracts and IL-6 and TNF-α. The serum of LLC tumor-bearing mice exhibited a higher amount of IL-6 (increased by 2.5 times) compared to that of the control, while administering a low dose of DTT water extracts reduced IL-6 by 1.8 times. The high dose of DTT water extracts nearly blocked IL-6 induction in LLC tumor-bearing mice (Figure 5A). However, the serum TNF-α increased 15-25 times by the LLC tumor compared to that of the control, the treatment of low or high doses of DTT water extracts reduced serum TNF-α by 27% and 39%, respectively (Figure 5A). These results indicated that the administration of DTT water extracts was inversely related to the levels of the inflammatory cytokines IL-6 and TNF-α. To further dissect the causality between DTT water extracts and these two cytokines, activated THP-1 cells (Human Acute Monocytic Leukemia Cells) were pretreated with different doses of DTT water extracts followed by LPS involvement. The results showed that 125 μg/ml of the DTT water extracts attenuated the expression of IL-6 induced by LPS by 18%. Furthermore, 500 μg/ml of DTT water extracts reduced the level of IL-6 by 35% (Figure 5B), showing a dose-dependent effect. On the contrary, the effects of DTT water extracts that interfered with TNF-α were milder; 500 μg/ml of DTT water extracts reduced the level of TNF-α only by 9% and by 28% at a higher concentration of 2000 μg/ml of DTT (Figure 5C). These results indicated that DTT water extracts inhibit the generation of both IL-6 and TNF-α and might contribute to the attenuation of tumor-induced muscle atrophy.
3.6. DTT Water Extracts Protect C2C12 Differentiation Against IL-6
We have shown that DTT water extracts ameliorate muscle wasting by reducing the generation of IL-6 and TNF-α. Next, we focus on the direct effects of DTT in protecting muscle cells against IL-6. C2C12 myoblasts were differentiated into myocyte (1 nucleus) or myotube (>= two nuclei) with or without DTT water extracts for six days, followed by IL-6 administration for another two days. The results showed an increasing ratio of differentiated C2C12 cells from 29% to 45% when engaged with DTT water extracts alone; Conversely, IL-6 treatment significantly reduced the differentiation ratio from 29% to 19% (Figure 6A). Pretreatment of DTT water extracts maintains the differentiation ratio of C2C12 at 41% even in the presence of IL-6 (Figure 6A,B), strongly implying that DTT water directly protects muscle cells against IL-6. Western blots showed the reduction of MyHC under IL-6 treatment (Figure 6C). Interestingly, pretreatment with DTT water extracts significantly reversed the expressions of murf-1 and MyHC of C2C12 cells in the presence of IL-6, consistent with the results of giemsa staining and supported our previous observation that DTT water extracts improve muscle atrophy by stabilizing muscle fibers in mice tumor model.
3.7. Vitamin B1 Is an Ingredient of DTT Water Extracts Involved in Protecting Differentiated C2C12 Cells Against IL-6
To dissect the possible ingredients of DTT responsible for the protective effects of muscle cells against IL-6, we performed an HPLC assay to identify potential constituents of DTT water extracts as described in the Materials and Methods section. Various vitamin B derivatives, namely thiamin (vitamin B1), pyridoxine (vitamin B6), niacinamide (vitamin B3), and Riboflavin (vitamin B2), were detected at retention times of 3.685 min, 5.242 min, 9.665 min, and 20.337 min, respectively (Figure 7A). Among these vitamin B derivatives, thiamin accounted for the highest percentage, we then investigated the inhibitory effects of vitamin B in regulating pro-inflammatory cytokines such as IL-6 and TNF-α of active THP-1cells. The results showed that administration of vitamin B1 effectively reduced the generation of IL-6 but bot TNF-α in active THP-1 cells (supp. Figure S4), indicating that vitamin B1 involved in the partial reduction of pro-inflammatory cytokines. Next, we analyzed the effects of vitamin B1 on the protection of differentiated C2C12 cells against IL-6. To further clarify the effects of DTT water extracts or vitamin B1 in regulating C2C12 differentiation, the immunofluorescence of C2C12 differentiated cells under various treatments was performed using anti-pan-MyHC antibody. The results showed that both the fusion index (the numbers of nuclei of cells whose nucleus numbers are larger than one with MyHC positive were divided by total nuclei) and the differentiation index (the numbers of nuclei of cells with MyHC positive were divided by total nuclei) of C2C12 cells treated with DTT water extracts were higher than untreated (increased by 15-20%, Figure 7B,C), while vitamin B1 treatment did not show significant differences compared to untreated cells. IL-6 treatment reduced both fusion and differentiation index compared to untreated cells. Interestingly, the pre-addition of DTT water extracts showed a nearly 2-fold increase in both fusion and differentiation index of C2C12 cells treated with IL-6 alone. At the same time, vitamin B1 exhibited a similar but minor induction of the fusion and differentiation index with that of IL-6 treatment alone (increased by 90-95%, Figure 7B,C). Both the total numbers of nuclei and the numbers of MyHC-positive cells under IL-6 treatment were significantly lower than those of the untreated cells, explaining why neither fusion nor differentiation index showed a significant difference between the differentiated cells C2C12 treated or untreated with IL-6 (Figure 7B,D). Furthermore, the ratio of MyHC positive cells but not total nuclei numbers of C2C12 differentiated cells treated with DTT water extract was significantly higher than the untreated cells (increasing by 70-80%), while vitamin B1 showed similar trends compared to untreated cells (Figure 7D). Interestingly, pretreatment of DTT water extracts maintained a high ratio of MyHC positive cells in the presence of IL-6 (increased by 90-110% vs. IL-6 alone); on the other hand, pretreatment of vitamin B1 also exhibited a higher ratio of MyHC positive cells compared to IL-6 alone (increased by 55-65% vs. IL-6 alone). These results indicated that vitamin B1 acted as an ingredient in DTT water extracts to maintain differentiated C2C12 myotubes against IL-6. Further results showed that the number of myocytes (1 nucleus/cell plus MyHC positive) remained similar among untreated DTT water extracts and vitamin B1-treated C2C12 cells. Furthermore, the coexistence of DTT water extracts and IL-6 significantly reduced the ratio of myocytes compared to that of untreated, DTT water extracts, or IL-6 alone; while vitamin B1 pretreatment exhibited a similar but not significant reduction in myocyte numbers in the presence of IL-6 (Figure 7F). Unexpectedly, the numbers of myotubes of DTT water extracts or vitamin B1 treatment were lower than those of the untreated ones (reduced by 12-17% vs. untreated, Figure 7F) and even lower in the presence of IL-6 (reduced by 22-35% vs. untreated; reduced by 10-15% vs. IL-6 alone, Figure 7F). The following analysis of the nuclei of myotubes whose nucleus number/cell >1 with MyHC positive showed a significant increase in the nuclei numbers of DTT water extracts but not vitamin B1 treated C2C12 myotubes than untreated ones (increased by 75-92% vs. untreated, Figure 7F). Treatment with IL-6 significantly reduced nuclei numbers per myotube by 20-30% compared to those of untreated ones, surprisingly, pretreatment of DTT water extracts followed by administration of IL-6 dramatically increased nuclei numbers per myotube by 172-221% compared to IL-6 alone (Figure 7F). Similar trends were also observed when pretreatment with vitamin B1 followed by IL-6 (increased by 145-195%, Figure 7F, bottom left panel). Increased nuclei numbers while decreasing myotubes of C2C12 cells treated with DTT water extracts or vitamin B1 raise the possibility that DTT water extracts or vitamin B1 might enhance the multinucleation of the C2C12 myotube with or without the presence of IL-6. The numbers of nuclei per myotube were then calculated to validate the possible effects. As expected, the average number of nuclei in C2C12 myotubes treated with DTT water extracts increased by 55-110% compared to untreated (Figure 7F, bottom left panel). Furthermore, the co-presence of IL-6 and DTT water extracts induced higher average nuclei numbers of C2C12 myotubes up to 200-230% compared to untreated ones. A smaller induction was also observed as pretreatment of vitamin B1 followed by IL-6 (increased by 35-40% vs. IL-6 alone, Figure 7F, bottom left panel). In the analysis of the nuclei number distribution of the myotube, the highest distribution of nuclei numbers of untreated C2C12 myotubes was 2-5 per myotube (nearly 62%), and the nuclei number ratio greater than 10 per myotube was approximately 8-10%; however, the nuclei number ratio more than 10 per myotube increased significantly by up to 35-38% while the nuclei number ratio at 2-5 was reduced by 15% with the treatment of DTT water extracts (Figure 7F, bottom right panel). On the other hand, vitamin B1 or IL-6 treatment did not show any significant differences in the distributions of nuclei numbers per myotube compared to untreated ones (Figure 7F, bottom left panel). IL-6 treatment reduced the ratio of nuclei numbers more than 10 per myotube by 51-62% (vs. untreated), while the presence of DTT water extracts dramatically induced the ratio of high nucleus numbers (>10) per myotube from 4-7% to 22-27%. In contrast, the number of nuclei in 2-5 per myotube in the coexistence of IL-6 and DTT water extracts was reduced from 60% to 38% compared to IL-6 alone (Figure 7F, bottom right panel). Furthermore, pretreatment with vitamin B1 also showed a similar but smaller change in the high number of nuclei numbers (>10) per myotube in the presence of IL-6 compared to treatment with IL-6 alone (23-27% vs. 4-7%, Figure 7F, bottom right panel). By combining these results, DTT water extracts could promote the middle-late stage of myoblast differentiation, such as multinucleations of large myotubes (or myofilament) by fusing smaller myotubes or myocytes rather than promoting early differentiation of the C2C12 myoblast. In addition, the pathological effects of IL-6 on C2C12 differentiation would be reversed in the presence of DTT water extracts, which could explain the protective effects of DTT water extracts in cancer-induced muscle wasting. Regarding vitamin B1, despite treatment alone showing no significant effects as DTT water extracts in the presence of IL-6, vitamin B1 exhibited an induction capacity of the fusion index, differentiation index, and multinucleation of myotubes, indicating a possible role for vitamin B1 in promoting or maintaining multinucleations of the C2C12 myotube.
3.8. E-Cadherin Is Required for Multinucleation of C2C12 Myotubes Induced by DTT Water Extracts or Vitamin B1
Next, western blot analysis was performed to analyze the possible molecules affected by DTT water extracts with or without IL-6 during C2C12 differentiation. The results showed that MyHC was significantly induced by DTT water extracts (increased 1.5-2.4 times vs. untreated) while increasing by 45-55% when treated with vitamin B1 (vs. untreated, Figure 8A). Both E-cadherin and β-catenin were also significantly induced by the treatment of DTT water extracts, the expression of E-cadherin increased by 3-4.5 times compared to that of the untreated, while the treatment of vitamin B1 increased the level of E-cadherin by 0.7-1.5 times compared to untreated (Figure 8A). The addition of CHX (cycloheximide) reduced the levels of E-cadherin and β-catenin more significantly than MyHC in the presence of DTT water extracts (Figure 8A). These results indicated that DTT water extracts might induce these molecules via a way of de-novo protein synthesis. Furthermore, IL-6 treatment reduced MyHC, E-cadherin, and β-catenin levels (35%, 80%, and 60% vs. untreated); Pretreatment of DTT water extracts significantly restored the levels of all three proteins (Figure 8B). However, pretreatment with vitamin B1 restored MyHC, E-cadherin, and β-catenin levels at a minor trend (Figure 8B), indicating that some other ingredients of DTT water extracts could also participate in up-regulation of these molecules in the presence of both DTT water extracts and IL-6. It should be noted that the addition of MG132 only increased the level of β-catenin but not MyHC or E-cadherin (Figure 8B), suggesting that IL-6 reduced the levels of different proteins through different mechanisms. To uncover the role of E-cadherin in C2C12 myogenesis, myoblasts were transfected twice with scramble RNA or E-cadherin siRNA followed by myogenesis with or without DTT water extracts or Vitamin B1. The results showed that the knockdown of E-cadherin decreased the level of β-catenin and affected the myotube ratio with or without DTT water extracts or vitamin B1 (Figure 8C,D). Further quantifications showed a dramatic reduction in both the fusion and differentiation index with the knockdown of E-cadherin (down to 9-13%, Figure 8E). Furthermore, the administration of DTT water extracts induced the fusion index to 33-38% (scRNA plus DW vs. scRNA con, Figure 8E), and also decreased to 17-20% when the E-cadherin was knockdown (E-cadherin siRNA plus DW vs. scRNA plus DW, Figure 8E), suggesting that E-cadherin as an essential factor to increase the fusion index. On the other hand, vitamin B1 administration also exhibited a similar trend of fusion and differentiation index with that of DTT water extracts; as expected, the knockdown of E-cadherin also reduced the fusion and differentiation index of vitamin B1-treated C2C12 myotubes (Figure 8E). However, a detailed analysis showed that E-cadherin knockdown reduced both nuclei numbers per myotube and multinucleations of C2C12 cells, not statistically significant (Figure 8E). Consistent with previous results, DTT water extract administration increased the numbers of nuclei per myotube from 5-7 to 10-20 (scRNA plus DW vs. scRNA, Figure 8E, bottom left panel). The ratio of nuclei numbers > 15 of C2C12 myotubes treated with DTT water extracts increased from 7% to 26%, while the ratio of nuclei numbers between 2 and 5 of C2C12 myotubes treated with DTT water extracts decreased from 38 to 23% (scRNA plus DW vs. scRNA, Figure 8E, bottom right panel). Interestingly, the elimination of E-cadherin reduced the ratio of nuclei numbers >15 from 26% to 9% of C2C12 myotubes in the presence of DTT water extracts (scRNA plus DW vs. siRNA of E-cadherin plus DW, Figure 8E, bottom right panel). However, the presence of DTT water extracts still elevated the ratio of nuclei numbers >15 from 3% to 9% in the C2C12 myotubes drained with E-cadherin (E-cadherin siRNA vs. E-cadherin siRNA plus DW, Figure 8D,E), suggesting that other molecules involved in the DTT water tracts promoted multinucleations of C2C12 myotube. On the other hand, the elimination of E-cadherin also reduced the nuclei numbers >15 from 15% to 3% in vitamin B1 treated C2C12 myotubes (scRNA plus VitB vs siRNA with E-cadherin plus VitB, panel bottom right), suggesting that vitamin B1 could be responsible for some degrees of multinucleation of C2C12 myotubes induced by DTT water extracts by upregulating the level of E-cadherin. By combining earlier results, our investigations indicated that E-cadherin is a novel promoter in multinucleations of C2C12 myotubes induced by DTT water extracts in IL-6-mediated disturbance of muscle fibers in a cachexia condition.
4. Discussion
Skeletal muscles are essential for both metabolic health and daily activities. Loss of muscle mass or function usually affects the mobility of older people or patients with chronic diseases such as cancer. Preservation of muscle function is the key to maintaining the standard of daily living and the survival interval of patients or older adults. Therefore, the search for effective and low-toxicity compounds to ameliorate muscle wasting induced by diseases became a common goal in contemporary cachexia therapy or prevention. This report demonstrates that DTT water-extracts administration preserved grip strength and muscle mass in LLC tumor-bearing mice. Furthermore, we showed that shrinkage of the cross-section area, abnormal nuclei aggregations (which the lot nucleus of the myofiber aggregated caused by atrophy), and reduced centralized nucleus (a phenotype of muscle regeneration) and MyHC expressions were all ameliorated by treatment with DTT water extracts. Elevated serum IL-6 and TNF-α, which are both known to affect muscle mass and functions, were also reduced when paired with DTT water extracts. Furthermore, the in vitro assay showed that DTT water extracts promoted differentiated C2C12 myotubes in the presence of IL-6. These results implied a dual role for DTT water extracts in promoting myoblast differentiation and stabilizing myotubes. We then identified vitamin B derivatives as ingredients in DTT water extracts. Interestingly, our results showed that vitamin B1 was involved in stabilizing differentiated C2C12 in the presence of IL-6 rather than promoting C2C12 cell differentiation.
Dendrobii, one of the largest genera of Orchidaceae, reveals numerous medicinal effects including anti-diabetic, -angiogenic, -inflammatory, -fungal, -bacterial, anti-tumor, etc. [44,45,46,47,48]. Most of the effects are contributed by Dendrobium polysaccharides, alkaloids, or organic soluble compounds [35,49,50]. For example, gigantol from D. Draconis in attenuating the stemness of lung cancer cells [51]; denbinobin from D. nobile in blocking the growth or motility of lung cancer, ovary cancer, prostate cancer, and colon cells[31,52,53,54,55]; glucomannan from D. officinale in regulating the generation of IL-1β and TNF-α in THP-1 cells [56]; phenanthrenes from D. nobile suppressed LPS-induced NO generation in RAW 264.7 macrophages [57]. However, the effects of Dendrobium on improving muscle wasting induced by cachexia or aging have not yet been reported. The present study is the first report to show that DTT, a new and endemic Orchidaceae of Taiwan, exerts anti-muscle-wasting effects in tumor-bearing mice. Furthermore, we first reported vitamin B1 as one of the water-soluble ingredients of DTT in protecting differentiated C2C12 myocytes or myotubes against IL-6. DTT is a new hybrid orchid bred by Taiwan Seed Improvement and Propagation Station, and the related report on its characteristics includes immune modulating ability [36,37]. Our findings extend the potential applications of DTT to preserve muscle mass and functions under pathological conditions since DTT does not exert acute or subchronic toxicity and genotoxic effects [37] plus the characteristics of high yield and easy cultivation, suggesting that DTT could be a highly valuable medicinal herb in the market of healthy foods.
Several studies reported the effects of tumors in eliciting muscle wasting in different animal models, for example, the lower mass of extensor digitorum longus, gastrocnemius, quadriceps, plantaris, tibialis, anterior triceps, and soleus muscles coupled with reduced grip strength were observed in mice engaging with tumor xenograft [58,59,60,61,62,63]. According to previous reports, our results showed a similar reduction ratio of the mass of the rectus femoris, gastrocnemius, and soleus muscles in a tumor-bearing mouse model (Figure 3). Additionally, a reduction in the cross-sectional area and the diameters of the rectus femoris, gastrocnemius and soleus muscles and multinucleations of myocytes were also observed (Figure 4), indicating extensive muscle wasting induced by LLC tumor xenograft. Administration of DTT water extracts recovered the grip strength, diameter, and number of nuclei of rectus femoris, gastrocnemius, and soleus muscles in a dose-dependent manner in tumor-bearing mice compared to control mice (Figure 3 and Figure 4). The masses and cross-section areas of these muscles, however, only recovered effects in the high dose of DTT water extracts (Figure 3 and Figure 4), raising the possibility that despite the physiological characteristics and outcomes of skeletal muscle being highly correlated in tumor-bearing mice, grip strength of skeletal muscles could depend more on the diameters and nucleus patterns of myofibers.
Zhang et al. reported that Baoyuan Jiedu Decoction ameliorated muscle wasting accompanied by tumor inhibition, implying an indirect pathway in recovering muscle functions in tumor-bearing mice [64]. Our results did not show significant inhibition of the tumor when treated with DTT water extracts (unpublished data); however, significant reductions in serum IL-6 and TNF-α, both highly correlated with muscle atrophy in a cancer cachexia model [65,66], were observed in the presence of DTT water extracts in tumor-bearing mice, and the in vitro results indicated that DTT water extracts reduced both IL-6 and TNF-α activated macrophage levels (Figure 5). Previous studies and our results showed that both inflammatory cytokines IL-6 and TNF-α, which are essential in the cachexia process, and the blockage of these cytokines by DTT water extracts could maintain muscle mass and functions even if the tumors remained. We also showed that DTT water extracts directly protected C2C12 myotubes under high-dose IL-6 treatment (Figure 6 and Figure 7). It should be noted that the role of IL-6 in the myogenesis of C2C12 is complicated; Steyn et al. showed that IL-6 promoted C2C12 cell differentiation, while Pelosi et al. showed that IL-6 impaired myogenesis of C2C12 [24,67]. The major difference between these controversial results is due to the concentration and time point of administration of IL-6. Generally, the administration of IL-6 of a lower dosage IL-6 (<100 ng/ml) at the start of differentiation will promote the differentiated index of C2C12 cells; however, the addition of IL-6 to differentiated C2C12 myotubes caused shrinkage of myotubes. Indeed, our unpublished results showed that the addition of both DTT water extracts and a low dose of IL-6 at the beginning of differentiation induced a higher differentiated index of C2C12 cells compared to treatment with water extracts alone, suggesting a dual role of IL-6 in promoting myoblast differentiation and altering differentiated myotubes. Regarding TNF-α, a similar pattern resembling IL-6 was observed when pretreating with DTT water extracts ameliorated such phenomena. Our results demonstrated that DTT water extracts protect muscle cells by both reducing the levels of inflammatory cytokines and stabilizing myotubes in the presence of both IL-6 and TNF-α. It should be noted that cytokine storms that involve surge of IL-6 and TNF-α, and after muscle catabolism, muscle anabolism impairment, and skeletal muscle atrophy occur in many other conditions such as burn [68] and sepsis [69,70], indicating that DTT could have importance in conditions beyond muscle wasting associated with cancer-induced skeletal muscle wasting. Therefore, further studies are warranted to investigate the effects of DTT on skeletal muscle wasting in various conditions.
DTT was crossbreeding by D. nobile and D. tosaense and exhibited the characteristics of high yield and easy planting. Yang et al. reported the immune stimulation of polysaccharide parts of DTT water extracts, which is contrary to our findings (Figure 5); this might be due to the amounts and purity of polysaccharides in different experiments. The physiological effects and corresponding active ingredients of the DTT water extracts have not been identified yet. In this report, we identified vitamin B derivatives, including B1, B2, B3, and B6 as novel components of DTT water extracts and further experiments showed that vitamin B1 exhibited stabilizing but not promoting effects in the C2C12 myogenesis process (Figure 7). Seon et al., mentioned the correlation between insufficient vitamin B12 intake and sarcopenia in elderly people [71]. Fuyuko et al. indicated that vitamin B1 or B12 intake was inversely correlated with muscle loss in older people with type 2 diabetes [72]. Takumi et al. demonstrated that vitamin B6-deficient mice harbored fewer quiescent satellite cells, which were required for de novo myogenesis [73]. The present study is the first report to indicate the novel role of vitamin B1 in protecting C2C12 myotubes against IL-6 treatment by attenuating the expression of murf-1, a muscle-specific F-box protein [74] responsible for proteolysis and muscle atrophy (Figure 7). However, the administration of vitamin B1 alone did not significantly promote C2C12 cell differentiation, which was not consistent with the findings that DTT water extracts induced C2C12 cell differentiation, raising the possibility that some other ingredients in DTT water extracts could be involved in the myogenesis-promoting process. Our findings provide additional molecular evidence to support previous reports on the correlations between vitamin B1 intake and muscle wasting.
Two members of the cadherin family, N and M cadherin, were known to be involved in promoting muscle regeneration; however, it was not clear which phase during muscle regeneration of these cadherins were involved [75,76]. On the other hand, β-catenin was involved in myogenesis through cooperation with smad7/MyoD to induce transcription of myogenic genes [77]. Furthermore, both M- and N- cadherin promoted myogenesis via regulating β-catenin functionality [78,79]. The precise role of E-cadherin in the regulation of muscle differentiation or regeneration has not been clarified. Our results showed that both E-cadherin and β-catenin were upregulated by administrating DTT water extracts while decreased by IL-6 treatment in the C2C12 myotube (Figure 8B). Furthermore, we also demonstrated that upregulation of E-cadherin was required for the multinucleation of C2C12 myotubes (Figure 8C–E). Interestingly, this result was consistent with the findings that both DTT water extracts and IL-6 affect the ratio of myotubes that contain more than 15 nuclei, implying a critical role of E-cadherin in the promotion of myotube fusion to generate myofibers. Since both DTT water extracts and vitamin B1 in gradient maintained the myotube fusion accompanied by up-regulation of E-cadherin under IL-6 treatment, it is reasonable to indicate that E-cadherin directly regulated myogenesis by promoting the myotube fusion process. The LLC allograft in the C57BL/6 mice model is widely used in the preclinical study of cachexia-related cancer symptoms [80,81]. Gradually reduced body weight, muscle weight, strength, and lipid weight are reminiscent of those of cancer cachexia patients [82]. Thus, our results from the animal study can potentially refer to the human pathological subject. However, there are some limitations to this study; the major limitation of this research was the lack of clinical evidence that DTT extracts against muscle wasting induced by cachexia or aging; future DTT research could focus on human subject research to evaluate the benefit of DTT extracts in attenuating muscle wasting induced by cachexia or aging. Second, the results of the animal study do not presumably predict human outcomes since there is a natural difference between the tumor environment in rodents and humans. Third, the sample sizes of mice in this animal study are relatively smaller, and this might impair the accuracy and validity of the results and translate them into clinical investigations.
5. Conclusions
Collectively, our and other previous findings demonstrated that DTT is a non-toxic, immunomodulated, and muscle-wasting prevention herb. It is worth further investigation into other physiological benefits of DTT, and this evidence might help to elevate the medical and economic values of DTT and encourage farmers to cultivate DTT to improve public health issues.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
All data were generated in-house, and no paper mill was used. All authors agree to be accountable for all aspects of work, ensuring integrity and accuracy. Chen-Chu Lin: Data Curation, Investigation, Methodology Resources, Writing—original draft. Wan-Ting Liao: Data Curation, Funding acquisition, Resources, Validation. Tsung-Ying Yang: Funding acquisition, Investigation. Jing-Hua Tsai: Conceptualization, Data Curation. Yi-Ju Lee: Methodology, Validation. Chi-Luan Wen: Methodology, Resources. Shih-Lan Hsu: Conceptualization, Resources. Chun-Chi Wu: Conceptualization, Data Curation, Investigation, Methodology, Project administration, supervision, Validation.
Institutional Review Board Statement
Ethical approval for the animals of subjects in this study was granted by the Animal Ethics Committee of Chung Shan Medical University (Approval No. 2422; Approval Date: 02-03-2021).
Data Availability Statement
The data generated in the present study may be requested from the corresponding author.
Acknowledgments
This work was supported by research grants from Dr. Chun-Chi Wu from Chung Shan Medical University (CSMU-INT-110-15), Dr. Tsung-Ying Yang from Taichung Veterans General Hospital (TCVGH-1113201D), and Dr Wan-Tin Liao from Show Chwan Memorial Hospital (SRD-110051).
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Karagounis, L. G.; Hawley, J. A., Skeletal muscle: increasing the size of the locomotor cell. The international journal of biochemistry & cell biology 2010, 42, (9), 1376-1379.
- Dave, H. D.; Shook, M.; Varacallo, M., Anatomy, skeletal muscle. In StatPearls [Internet], StatPearls Publishing: 2021.
- Hindi, S. M.; Tajrishi, M. M.; Kumar, A., Signaling mechanisms in mammalian myoblast fusion. Science signaling 2013, 6, (272), re2-re2.
- Bottinelli, R.; Reggiani, C., Human skeletal muscle fibres: molecular and functional diversity. Progress in biophysics and molecular biology 2000, 73, (2-4), 195-262.
- Rosen, G. D.; Sanes, J. R.; LaChance, R.; Cunningham, J. M.; Roman, J.; Dean, D. C. J. C., Roles for the integrin VLA-4 and its counter receptor VCAM-1 in myogenesis. 1992, 69, (7), 1107-1119.
- Chang, Y.-H.; Tsai, J.-N.; Chen, T.-L.; Ho, K.-T.; Cheng, H.-Y.; Hsiao, C.-W.; Shiau, M.-Y. J. M. o. I., Interleukin-4 promotes myogenesis and boosts myocyte insulin efficacy. 2019, 2019.
- Kurosaka, M.; Ogura, Y.; Sato, S.; Kohda, K.; Funabashi, T. J. S. M., Transcription factor signal transducer and activator of transcription 6 (STAT6) is an inhibitory factor for adult myogenesis. 2021, 11, (1), 14.
- Wu, Z.; Woodring, P. J.; Bhakta, K. S.; Tamura, K.; Wen, F.; Feramisco, J. R.; Karin, M.; Wang, J. Y.; Puri, P. L. J. M.; biology, c., p38 and extracellular signal-regulated kinases regulate the myogenic program at multiple steps. 2000, 20, (11), 3951-3964.
- Suzuki, A.; Pelikan, R. C.; Iwata, J. J. M.; biology, c., WNT/β-catenin signaling regulates multiple steps of myogenesis by regulating step-specific targets. 2015.
- Frontera, W. R.; Ochala, J., Skeletal muscle: a brief review of structure and function. Calcified tissue international 2015, 96, 183-195.
- Evans, W. J., Skeletal muscle loss: cachexia, sarcopenia, and inactivity. The American journal of clinical nutrition 2010, 91, (4), 1123S-1127S.
- Penna, F.; Ballarò, R.; Beltrá, M.; De Lucia, S.; Costelli, P., Modulating metabolism to improve cancer-induced muscle wasting. Oxidative Medicine and Cellular Longevity 2018, 2018.
- Holecek, M., Muscle wasting in animal models of severe illness. International journal of experimental pathology 2012, 93, (3), 157-171.
- Nishikawa, H.; Goto, M.; Fukunishi, S.; Asai, A.; Nishiguchi, S.; Higuchi, K., Cancer cachexia: its mechanism and clinical significance. International journal of molecular sciences 2021, 22, (16), 8491.
- Aoyagi, T.; Terracina, K. P.; Raza, A.; Matsubara, H.; Takabe, K., Cancer cachexia, mechanism and treatment. World journal of gastrointestinal oncology 2015, 7, (4), 17.
- Vaughan, V. C.; Martin, P.; Lewandowski, P. A., Cancer cachexia: impact, mechanisms and emerging treatments. Journal of cachexia, sarcopenia and muscle 2013, 4, 95-109.
- Castaneda, C., Muscle wasting and protein metabolism. Journal of Animal Science 2002, 80, (E-suppl_2), E98-E105.
- Argilés, J. M.; Busquets, S.; Stemmler, B.; López-Soriano, F. J., Cancer cachexia: understanding the molecular basis. Nature Reviews Cancer 2014, 14, (11), 754-762.
- Peixoto da Silva, S.; Santos, J. M.; Costa e Silva, M. P.; Gil da Costa, R. M.; Medeiros, R., Cancer cachexia and its pathophysiology: links with sarcopenia, anorexia and asthenia. Journal of cachexia, sarcopenia and muscle 2020, 11, (3), 619-635.
- Dodson, S.; Baracos, V. E.; Jatoi, A.; Evans, W. J.; Cella, D.; Dalton, J. T.; Steiner, M. S., Muscle wasting in cancer cachexia: clinical implications, diagnosis, and emerging treatment strategies. Annual review of medicine 2011, 62, 265-279.
- Baracos, V. E.; Mazurak, V. C.; Bhullar, A. S., Cancer cachexia is defined by an ongoing loss of skeletal muscle mass. Ann Palliat Med 2019, 8, (1), 3-12.
- Patel, H. J.; Patel, B. M., TNF-α and cancer cachexia: Molecular insights and clinical implications. Life sciences 2017, 170, 56-63.
- Bonetto, A.; Aydogdu, T.; Kunzevitzky, N.; Guttridge, D. C.; Khuri, S.; Koniaris, L. G.; Zimmers, T. A., STAT3 activation in skeletal muscle links muscle wasting and the acute phase response in cancer cachexia. PloS one 2011, 6, (7), e22538.
- Pelosi, M.; De Rossi, M.; Barberi, L.; Musarò, A., IL-6 impairs myogenic differentiation by downmodulation of p90RSK/eEF2 and mTOR/p70S6K axes, without affecting AKT activity. BioMed research international 2014, 2014.
- Adams, V.; Mangner, N.; Gasch, A.; Krohne, C.; Gielen, S.; Hirner, S.; Thierse, H.-J.; Witt, C. C.; Linke, A.; Schuler, G., Induction of MuRF1 is essential for TNF-α-induced loss of muscle function in mice. Journal of molecular biology 2008, 384, (1), 48-59.
- Garcia, V. R.; López-Briz, E.; Sanchis, R. C.; Perales, J. L. G.; Bort-Martí, S., Megestrol acetate for treatment of anorexia-cachexia syndrome. Cochrane Database of Systematic Reviews 2013, (3).
- Nakanishi, Y.; Higuchi, J.; Honda, N.; Komura, N., Pharmacological profile and clinical efficacy of anamorelin HCl (ADLUMIZ® Tablets), the first orally available drug for cancer cachexia with ghrelin-like action in Japan. Nihon Yakurigaku zasshi. Folia Pharmacologica Japonica 2021, 156, (6), 370-381.
- Bulchandani, D.; Nachnani, J.; Amin, A.; May, J., Megestrol acetate—associated adrenal insufficiency. The American Journal of Geriatric Pharmacotherapy 2008, 6, (3), 167-172.
- FARRAR, D. J., Megestrol acetate: promises and pitfalls. AIDS Patient Care and STDs 1999, 13, (3), 149-152.
- Wachtel-Galor, S.; Benzie, I. F., Herbal medicine. Lester Packer, Ph. D. 2011, 1.
- SONG, G.-Q., Research progress on pharmacological activities of medical plants from Dendrobium Sw. Chinese Traditional and Herbal Drugs 2014, 2576-2580.
- Teoh, E. S.; Teoh, E. S., Traditional Chinese Medicine, Korean Traditional Herbal Medicine, and Japanese Kanpo Medicine. Medicinal Orchids of Asia 2016, 19-31.
- Ramesh, T.; Koperuncholan, M.; Praveena, R.; Ganeshkumari, K.; Vanithamani, J.; Muruganantham, P.; Renganathan, P., Medicinal properties of some Dendrobium orchids–A review. Journal of Applied and Advanced Research 2019, 4, (4), 119-128.
- Xing, X.; Cui, S. W.; Nie, S.; Phillips, G. O.; Goff, H. D.; Wang, Q., A review of isolation process, structural characteristics, and bioactivities of water-soluble polysaccharides from Dendrobium plants. Bioactive Carbohydrates and Dietary Fibre 2013, 1, (2), 131-147.
- Teixeira da Silva, J. A.; Ng, T. B., The medicinal and pharmaceutical importance of Dendrobium species. Applied Microbiology and Biotechnology 2017, 101, 2227-2239.
- Yang, L.-C.; Hsieh, C.-C.; Wen, C.-L.; Chiu, C.-H.; Lin, W.-C., Structural characterization of an immunostimulating polysaccharide from the stems of a new medicinal Dendrobium species: Dendrobium Taiseed Tosnobile. International journal of biological macromolecules 2017, 103, 1185-1193.
- Yang, L.-C.; Liao, J.-W.; Wen, C.-L.; Lin, W.-C., Subchronic and Genetic Safety Assessment of a New Medicinal Dendrobium Species: Dendrobium Taiseed Tosnobile in Rats. Evidence-Based Complementary and Alternative Medicine 2018, 2018.
- McLean, J. B.; Moylan, J. S.; Andrade, F. H., Mitochondria dysfunction in lung cancer-induced muscle wasting in C2C12 myotubes. Frontiers in physiology 2014, 5, 503.
- Castro, B.; Kuang, S. J. B.-p., Evaluation of muscle performance in mice by treadmill exhaustion test and whole-limb grip strength assay. 2017, 7, (8), e2237-e2237.
- Ballarò, R.; Costelli, P.; Penna, F., Animal models for cancer cachexia. Current opinion in supportive and palliative care 2016, 10, (4), 281-287.
- Cadot, B.; Gache, V.; Gomes, E. R. J. N., Moving and positioning the nucleus in skeletal muscle–one step at a time. 2015, 6, (5), 373-381.
- Oliff, A.; Defeo-Jones, D.; Boyer, M.; Martinez, D.; Kiefer, D.; Vuocolo, G.; Wolfe, A.; Socher, S. H. J. C., Tumors secreting human TNF/cachectin induce cachexia in mice. 1987, 50, (4), 555-563.
- Rupert, J. E.; Narasimhan, A.; Jengelley, D. H.; Jiang, Y.; Liu, J.; Au, E.; Silverman, L. M.; Sandusky, G.; Bonetto, A.; Cao, S. J. J. o. E. M., Tumor-derived IL-6 and trans-signaling among tumor, fat, and muscle mediate pancreatic cancer cachexia. 2021, 218, (6), e20190450.
- Wang, K.; Wang, H.; Liu, Y.; Shui, W.; Wang, J.; Cao, P.; Wang, H.; You, R.; Zhang, Y., Dendrobium officinale polysaccharide attenuates type 2 diabetes mellitus via the regulation of PI3K/Akt-mediated glycogen synthesis and glucose metabolism. Journal of Functional Foods 2018, 40, 261-271.
- Yue, H.; Liu, Y.; Qu, H.; Ding, K., Structure analysis of a novel heteroxylan from the stem of Dendrobium officinale and anti-angiogenesis activities of its sulfated derivative. International journal of biological macromolecules 2017, 103, 533-542.
- Xing, Y.-M.; Chen, J.; Cui, J.-L.; Chen, X.-M.; Guo, S.-X., Antimicrobial activity and biodiversity of endophytic fungi in Dendrobium devonianum and Dendrobium thyrsiflorum from Vietman. Current microbiology 2011, 62, 1218-1224.
- Qiang, Z.; Ko, C.-H.; Siu, W.-S.; Kai-Kai, L.; Wong, C.-W.; Xiao-Qiang, H.; Liu, Y.; Bik-San Lau, C.; Jiang-Miao, H.; Leung, P.-C., Inhibitory effect of different Dendrobium species on LPS-induced inflammation in macrophages via suppression of MAPK pathways. Chinese journal of natural medicines 2018, 16, (7), 481-489.
- Liang, J.; Li, H.; Chen, J.; He, L.; Du, X.; Zhou, L.; Xiong, Q.; Lai, X.; Yang, Y.; Huang, S., Dendrobium officinale polysaccharides alleviate colon tumorigenesis via restoring intestinal barrier function and enhancing anti-tumor immune response. Pharmacological research 2019, 148, 104417.
- Huang, S.; Chen, F.; Cheng, H.; Huang, G., Modification and application of polysaccharide from traditional Chinese medicine such as Dendrobium officinale. International Journal of Biological Macromolecules 2020, 157, 385-393.
- Cui, S.; Li, L.; Yu, R. T.; Downes, M.; Evans, R. M.; Hulin, J.-A.; Makarenkova, H. P.; Meech, R., β-Catenin is essential for differentiation of primary myoblasts via cooperation with MyoD and α-catenin. Development 2019, 146, (6), dev167080.
- Charoenrungruang, S.; Chanvorachote, P.; Sritularak, B.; Pongrakhananon, V., Gigantol, a bibenzyl from Dendrobium draconis, inhibits the migratory behavior of non-small cell lung cancer cells. Journal of Natural Products 2014, 77, (6), 1359-1366.
- Kuo, C.-T.; Hsu, M.-J.; Chen, B.-C.; Chen, C.-C.; Teng, C.-M.; Pan, S.-L.; Lin, C.-H., Denbinobin induces apoptosis in human lung adenocarcinoma cells via Akt inactivation, Bad activation, and mitochondrial dysfunction. Toxicology Letters 2008, 177, (1), 48-58.
- Lee, Y. H.; Park, J. D.; Beak, N. I.; Kim, S. I.; Ahn, B. Z., In vitro and in vivo antitumoral phenanthrenes from the aerial parts of Dendrobium nobile. Planta Medica 1995, 61, (02), 178-180.
- Lu, T.-L.; Han, C.-K.; Chang, Y.-S.; Lu, T.-J.; Huang, H.-C.; Bao, B.-Y.; Wu, H.-Y.; Huang, C.-H.; Li, C.-Y.; Wu, T.-S., Denbinobin, a phenanthrene from Dendrobium nobile, impairs prostate cancer migration by inhibiting Rac1 activity. The American journal of Chinese medicine 2014, 42, (06), 1539-1554.
- Yang, K.-C.; Uen, Y.-H.; Suk, F.-M.; Liang, Y.-C.; Wang, Y.-J.; Ho, Y.-S.; Li, I.-H.; Lin, S.-Y., Molecular mechanisms of denbinobin-induced anti-tumorigenesis effect in colon cancer cells. World Journal of Gastroenterology: WJG 2005, 11, (20), 3040.
- Huang, Y.-P.; He, T.-B.; Cuan, X.-D.; Wang, X.-J.; Hu, J.-M.; Sheng, J., 1, 4-β-d-Glucomannan from Dendrobium officinale Activates NF-к B via TLR4 to Regulate the Immune Response. Molecules 2018, 23, (10), 2658.
- Hwang, J. S.; Lee, S. A.; Hong, S. S.; Han, X. H.; Lee, C.; Kang, S. J.; Lee, D.; Kim, Y.; Hong, J. T.; Lee, M. K., Phenanthrenes from Dendrobium nobile and their inhibition of the LPS-induced production of nitric oxide in macrophage RAW 264.7 cells. Bioorganic & Medicinal Chemistry Letters 2010, 20, (12), 3785-3787.
- Brown, J. L.; Lawrence, M. M.; Ahn, B.; Kneis, P.; Piekarz, K. M.; Qaisar, R.; Ranjit, R.; Bian, J.; Pharaoh, G.; Brown, C. J. J. o. C., Sarcopenia; Muscle, Cancer cachexia in a mouse model of oxidative stress. 2020, 11, (6), 1688-1704.
- Khal, J.; Wyke, S.; Russell, S. T.; Hine, A. V.; Tisdale, M. J. J. B. j. o. c., Expression of the ubiquitin-proteasome pathway and muscle loss in experimental cancer cachexia. 2005, 93, (7), 774-780.
- Roberts, B.; Frye, G.; Ahn, B.; Ferreira, L.; Judge, A. J. B.; communications, b. r., Cancer cachexia decreases specific force and accelerates fatigue in limb muscle. 2013, 435, (3), 488-492.
- Otis, J. S.; Lees, S. J.; Williams, J. H. J. B. c., Functional overload attenuates plantaris atrophy in tumor-bearing rats. 2007, 7, (1), 1-9.
- Levolger, S.; van den Engel, S.; Ambagtsheer, G.; IJzermans, J. N.; de Bruin, R. W. J. N.; Aging, H., Quercetin supplementation attenuates muscle wasting in cancer-associated cachexia in mice. 2021, 6, (1), 35-47.
- Al-Majid, S.; McCarthy, D. O. J. B. r. f. n., Cancer-induced fatigue and skeletal muscle wasting: the role of exercise. 2001, 2, (3), 186-197.
- Zhang, Y.; Han, X.; Ouyang, B.; Wu, Z.; Yu, H.; Wang, Y.; Liu, G.; Ji, X. J. E.-B. C.; Medicine, A., Chinese herbal medicine Baoyuan Jiedu Decoction inhibited muscle atrophy of cancer cachexia through Atrogin-l and MuRF-1. 2017, 2017.
- Bonetto, A.; Aydogdu, T.; Jin, X.; Zhang, Z.; Zhan, R.; Puzis, L.; Koniaris, L. G.; Zimmers, T. A. J. A. J. o. P.-E.; Metabolism, JAK/STAT3 pathway inhibition blocks skeletal muscle wasting downstream of IL-6 and in experimental cancer cachexia. 2012, 303, (3), E410-E421.
- Webster, J. M.; Kempen, L. J.; Hardy, R. S.; Langen, R. C. J. F. i. p., Inflammation and skeletal muscle wasting during cachexia. 2020, 11, 597675.
- Steyn, P. J.; Dzobo, K.; Smith, R. I.; Myburgh, K. H. J. I. j. o. m. s., Interleukin-6 induces myogenic differentiation via JAK2-STAT3 signaling in mouse C2C12 myoblast cell line and primary human myoblasts. 2019, 20, (21), 5273.
- Ono, Y.; Saito, M.; Sakamoto, K.; Maejima, Y.; Misaka, S.; Shimomura, K.; Nakanishi, N.; Inoue, S.; Kotani, J. J. F. i. P., C188-9, a specific inhibitor of STAT3 signaling, prevents thermal burn-induced skeletal muscle wasting in mice. 2022, 13, 1031906.
- Ono, Y.; Maejima, Y.; Saito, M.; Sakamoto, K.; Horita, S.; Shimomura, K.; Inoue, S.; Kotani, J. J. S. r., TAK-242, a specific inhibitor of Toll-like receptor 4 signalling, prevents endotoxemia-induced skeletal muscle wasting in mice. 2020, 10, (1), 694.
- Ono, Y.; Sakamoto, K. J. P. o., Lipopolysaccharide inhibits myogenic differentiation of C2C12 myoblasts through the Toll-like receptor 4-nuclear factor-κB signaling pathway and myoblast-derived tumor necrosis factor-α. 2017, 12, (7), e0182040.
- Chae, S. A.; Kim, H.-S.; Lee, J. H.; Yun, D. H.; Chon, J.; Yoo, M. C.; Yun, Y.; Yoo, S. D.; Kim, D. H.; Lee, S. A. J. I. J. o. E. R.; Health, P., Impact of vitamin B12 insufficiency on sarcopenia in community-dwelling older Korean adults. 2021, 18, (23), 12433.
- Takahashi, F.; Hashimoto, Y.; Kaji, A.; Sakai, R.; Kawate, Y.; Okamura, T.; Kondo, Y.; Fukuda, T.; Kitagawa, N.; Okada, H. J. N., Vitamin intake and loss of muscle mass in older people with type 2 diabetes: a prospective study of the KAMOGAWA-DM cohort. 2021, 13, (7), 2335.
- Komaru, T.; Yanaka, N.; Kumrungsee, T. J. N., Satellite Cells Exhibit Decreased Numbers and Impaired Functions on Single Myofibers Isolated from Vitamin B6-Deficient Mice. 2021, 13, (12), 4531.
- Gomes, M. D.; Lecker, S. H.; Jagoe, R. T.; Navon, A.; Goldberg, A. L. J. P. o. t. N. A. o. S., Atrogin-1, a muscle-specific F-box protein highly expressed during muscle atrophy. 2001, 98, (25), 14440-14445.
- Donalies, M.; Cramer, M.; Ringwald, M.; Starzinski-Powitz, A., Expression of M-cadherin, a member of the cadherin multigene family, correlates with differentiation of skeletal muscle cells. Proceedings of the National Academy of Sciences 1991, 88, (18), 8024-8028.
- George-Weinstein, M.; Gerhart, J.; Blitz, J.; Simak, E.; Knudsen, K. A., N-cadherin promotes the commitment and differentiation of skeletal muscle precursor cells. Developmental biology 1997, 185, (1), 14-24.
- Kang, J.-S.; Feinleib, J. L.; Knox, S.; Ketteringham, M. A.; Krauss, R. S., Promyogenic members of the Ig and cadherin families associate to positively regulate differentiation. Proceedings of the National Academy of Sciences 2003, 100, (7), 3989-3994.
- Charrasse, S.; Meriane, M.; Comunale, F.; Blangy, A.; Gauthier-Rouvière, C., N-cadherin–dependent cell–cell contact regulates Rho GTPases and β-catenin localization in mouse C2C12 myoblasts. The Journal of cell biology 2002, 158, (5), 953-965.
- Wróbel, E.; Brzóska, E.; Moraczewski, J., M-cadherin and β-catenin participate in differentiation of rat satellite cells. European journal of cell biology 2007, 86, (2), 99-109.
- Li, L.; Wazir, J.; Huang, Z.; Wang, Y.; Wang, H. J. G.; Diseases, A comprehensive review of animal models for cancer cachexia: Implications for translational research. 2024, 11, (6), 101080.
- Pototschnig, I.; Feiler, U.; Diwoky, C.; Vesely, P. W.; Rauchenwald, T.; Paar, M.; Bakiri, L.; Pajed, L.; Hofer, P.; Kashofer, K. J. J. o. c., sarcopenia; muscle, Interleukin-6 initiates muscle-and adipose tissue wasting in a novel C57BL/6 model of cancer-associated cachexia. 2023, 14, (1), 93-107.
- Mueller, T. C.; Bachmann, J.; Prokopchuk, O.; Friess, H.; Martignoni, M. E. J. B. c., Molecular pathways leading to loss of skeletal muscle mass in cancer cachexia–can findings from animal models be translated to humans? 2016, 16, 1-14.
Figure 1.
DTT water extracts reduce body weight loss in tumor-bearing mice with LLC. (A) Treatment schedule for experimental animal study, Lewis lung cancer cells were implanted subcutaneously in the right flank of mice (1 × 10^5/mouse) on day 0. DTT water extracts (75 or 150 mg/kg/day) were started orally on day seven after tumor inoculation (tumor was apparent), while the tumor control group received an equal volume of water. From day seven, all groups of mice were subjected to a measurement of body weight week and grip strength once a week until day 28. The mice were then sacrificed and related muscle tissues such as the left hind limb, gastrocnemius, and soleus of the right hind limb were collected for the indicated experiments. (B) Each group’s mean consecutive body weights were recorded every week until sacrifice. (C) The mean food intake of each group was recorded every day until sacrifice. (D) On the day of sacrifice, each group’s mean body weight (w/o tumor) was measured and compared with that of the control group as a related loss ratio, n=5/group, ***p<0.001, *p<0.05. (E) The mean body weight without tumors on the day of the sacrifice of each group (n=5/group) was presented, ***p<0.001, *p<0.05. Multiple group comparisons were achieved by one-way ANOVA with Tukey’s post hoc.
Figure 1.
DTT water extracts reduce body weight loss in tumor-bearing mice with LLC. (A) Treatment schedule for experimental animal study, Lewis lung cancer cells were implanted subcutaneously in the right flank of mice (1 × 10^5/mouse) on day 0. DTT water extracts (75 or 150 mg/kg/day) were started orally on day seven after tumor inoculation (tumor was apparent), while the tumor control group received an equal volume of water. From day seven, all groups of mice were subjected to a measurement of body weight week and grip strength once a week until day 28. The mice were then sacrificed and related muscle tissues such as the left hind limb, gastrocnemius, and soleus of the right hind limb were collected for the indicated experiments. (B) Each group’s mean consecutive body weights were recorded every week until sacrifice. (C) The mean food intake of each group was recorded every day until sacrifice. (D) On the day of sacrifice, each group’s mean body weight (w/o tumor) was measured and compared with that of the control group as a related loss ratio, n=5/group, ***p<0.001, *p<0.05. (E) The mean body weight without tumors on the day of the sacrifice of each group (n=5/group) was presented, ***p<0.001, *p<0.05. Multiple group comparisons were achieved by one-way ANOVA with Tukey’s post hoc.
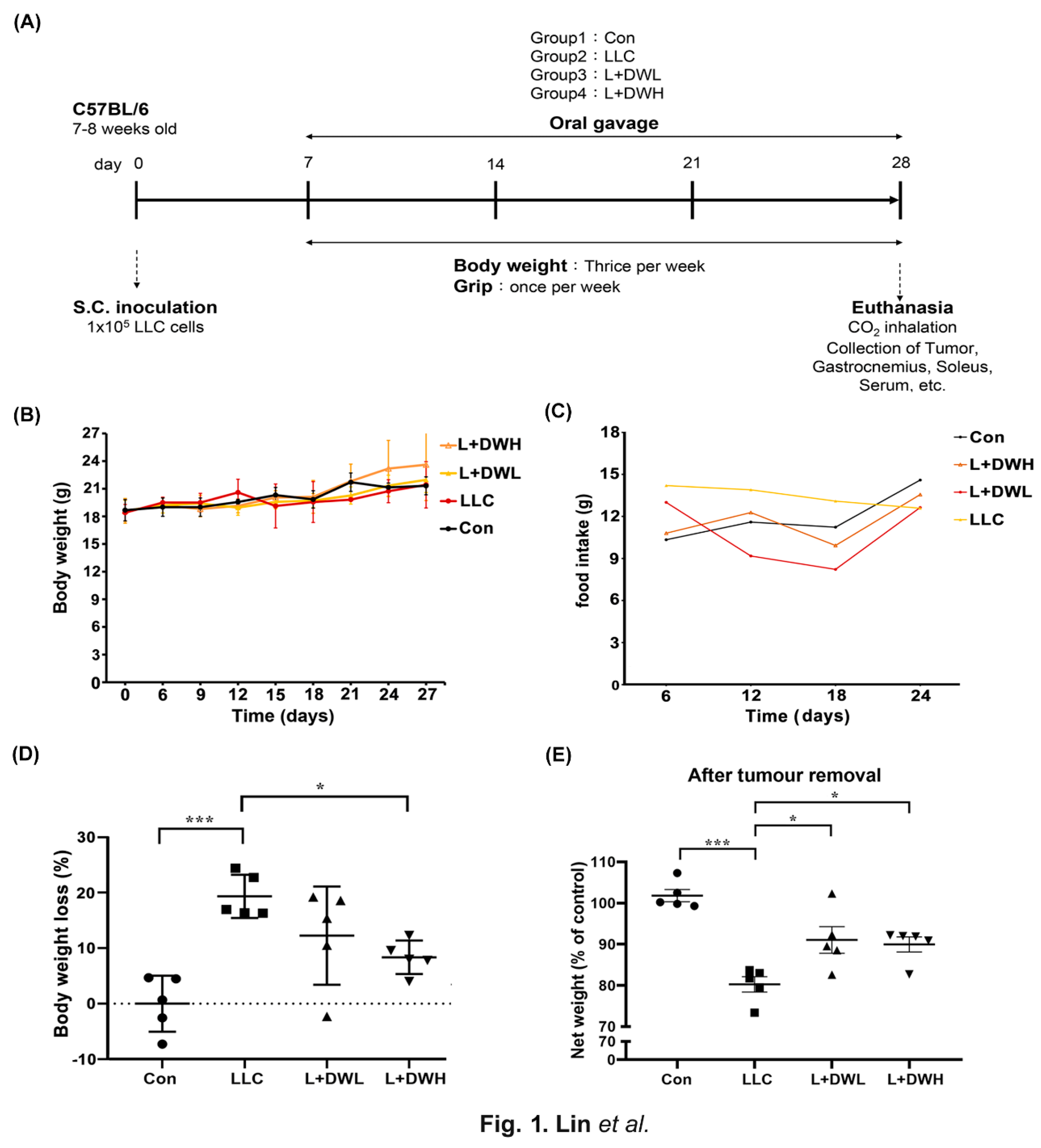
Figure 2.
DTT water extracts retrieve the muscle functionality of LLC tumor-bearing mice. (A)The grip strength of each group was measured once a week until day 28 and was shown. (B) On day 28, each group’s grip strength relative to body weights (without tumor) was recorded, and the average values were then calculated. Data are expressed as means ± SD, ***p < 0.001 compared to the control group, **p <0.01 compared to the tumor group. After the euthanasia of mice on day 28, the mass of (C) gastrocnemius and (D) soleus of each mouse from a different group was measured. n=5/group, ***p<0.001, *p<0.05, multiple group comparisons were achieved by one-way ANOVA with a Tukey’s post hoc.
Figure 2.
DTT water extracts retrieve the muscle functionality of LLC tumor-bearing mice. (A)The grip strength of each group was measured once a week until day 28 and was shown. (B) On day 28, each group’s grip strength relative to body weights (without tumor) was recorded, and the average values were then calculated. Data are expressed as means ± SD, ***p < 0.001 compared to the control group, **p <0.01 compared to the tumor group. After the euthanasia of mice on day 28, the mass of (C) gastrocnemius and (D) soleus of each mouse from a different group was measured. n=5/group, ***p<0.001, *p<0.05, multiple group comparisons were achieved by one-way ANOVA with a Tukey’s post hoc.
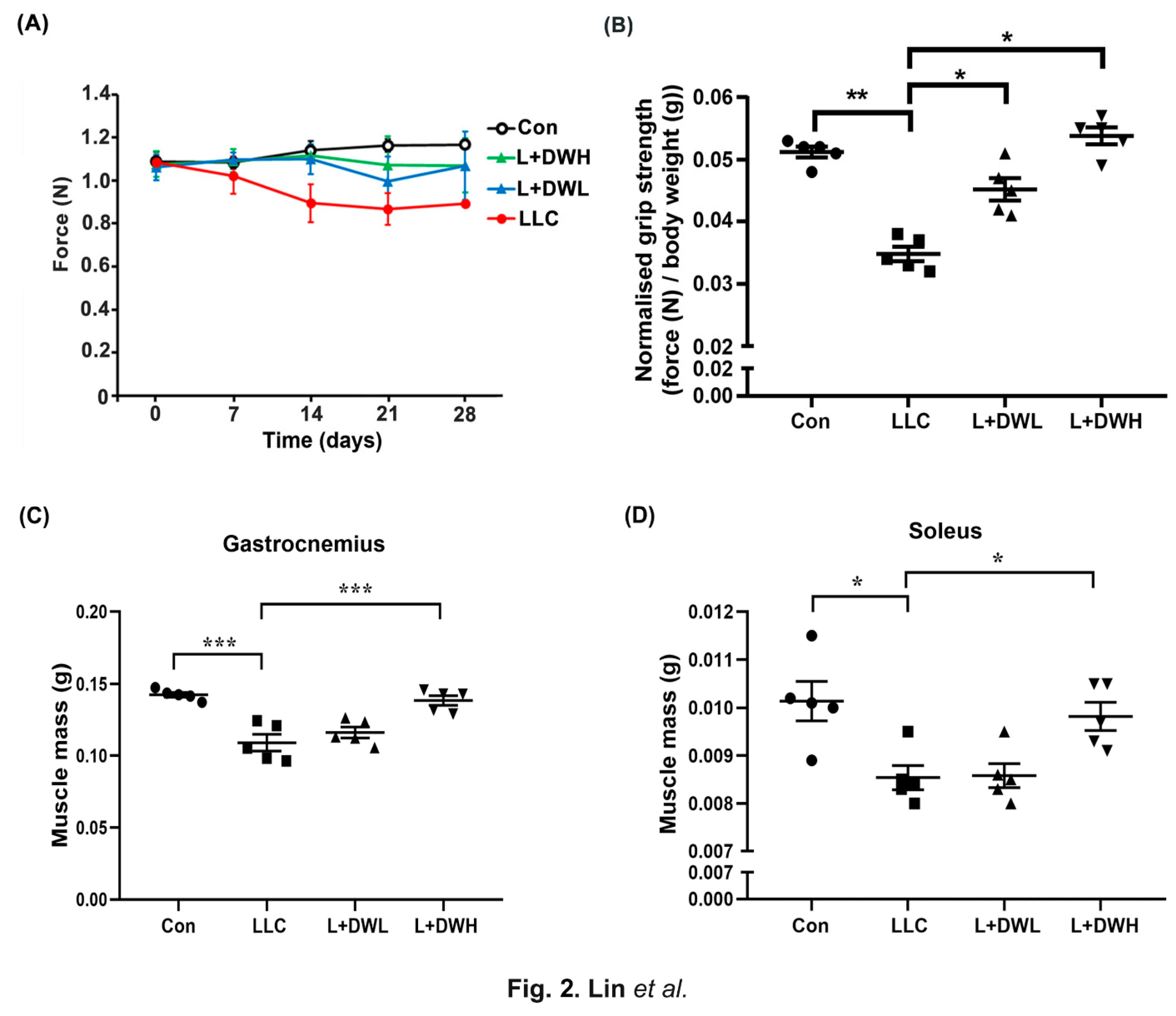
Figure 3.
DTT water extracts ameliorate muscle CSA reduction in LLC tumor-bearing mice. After euthanasia of the mice on day 28, the histopathological analysis of (A) rectus femoris, (B) soleus and gastrocnemius of mice from control, LLC, LLC plus DTT water extracts low dosage and LLC plus DTT water extracts high dosage were performed using H/E stain, scale bar = 1mm. The cross-section area (CSA) of (C) (yellow circle line), (D) soleus (green circle line), and (E) gastrocnemius (red circle line) was calculated and quantified. n=5/group, **p<0.005, *p<0.05, multiple group comparisons were achieved by one-way ANOVA with Tukey’s post hoc.
Figure 3.
DTT water extracts ameliorate muscle CSA reduction in LLC tumor-bearing mice. After euthanasia of the mice on day 28, the histopathological analysis of (A) rectus femoris, (B) soleus and gastrocnemius of mice from control, LLC, LLC plus DTT water extracts low dosage and LLC plus DTT water extracts high dosage were performed using H/E stain, scale bar = 1mm. The cross-section area (CSA) of (C) (yellow circle line), (D) soleus (green circle line), and (E) gastrocnemius (red circle line) was calculated and quantified. n=5/group, **p<0.005, *p<0.05, multiple group comparisons were achieved by one-way ANOVA with Tukey’s post hoc.
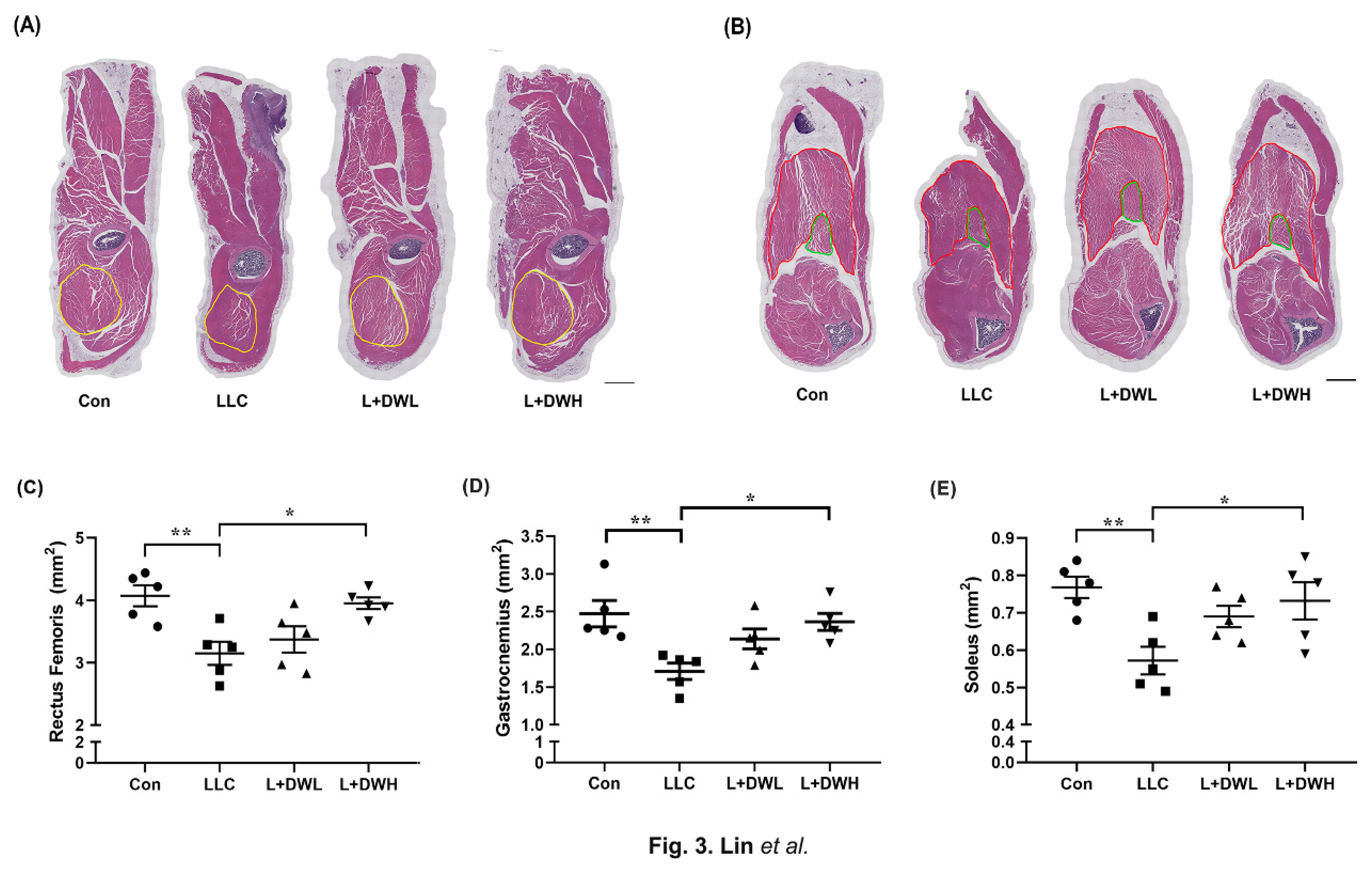
Figure 4.
DTT water extracts restore the architecture of the muscle fibers of LLC tumor-bearing mice. The minferet diameter of the (A) rectus femoris, (B) gastrocnemius and (C) soleus of each mouse was calculated and quantified. Four distinct fields of the nuclei of (D) rectus femoris, (E) gastrocnemius and (F) soleus were calculated and quantified. The ratio of the centralized nucleus of (G) rectus femoris, (H) gastrocnemius and (I) soleus was calculated and quantified. n=5/group, ***p<0.001, **p<0.005, *p<0.05, multiple group comparisons were achieved by one-way ANOVA with a Tukey’s post hoc.
Figure 4.
DTT water extracts restore the architecture of the muscle fibers of LLC tumor-bearing mice. The minferet diameter of the (A) rectus femoris, (B) gastrocnemius and (C) soleus of each mouse was calculated and quantified. Four distinct fields of the nuclei of (D) rectus femoris, (E) gastrocnemius and (F) soleus were calculated and quantified. The ratio of the centralized nucleus of (G) rectus femoris, (H) gastrocnemius and (I) soleus was calculated and quantified. n=5/group, ***p<0.001, **p<0.005, *p<0.05, multiple group comparisons were achieved by one-way ANOVA with a Tukey’s post hoc.
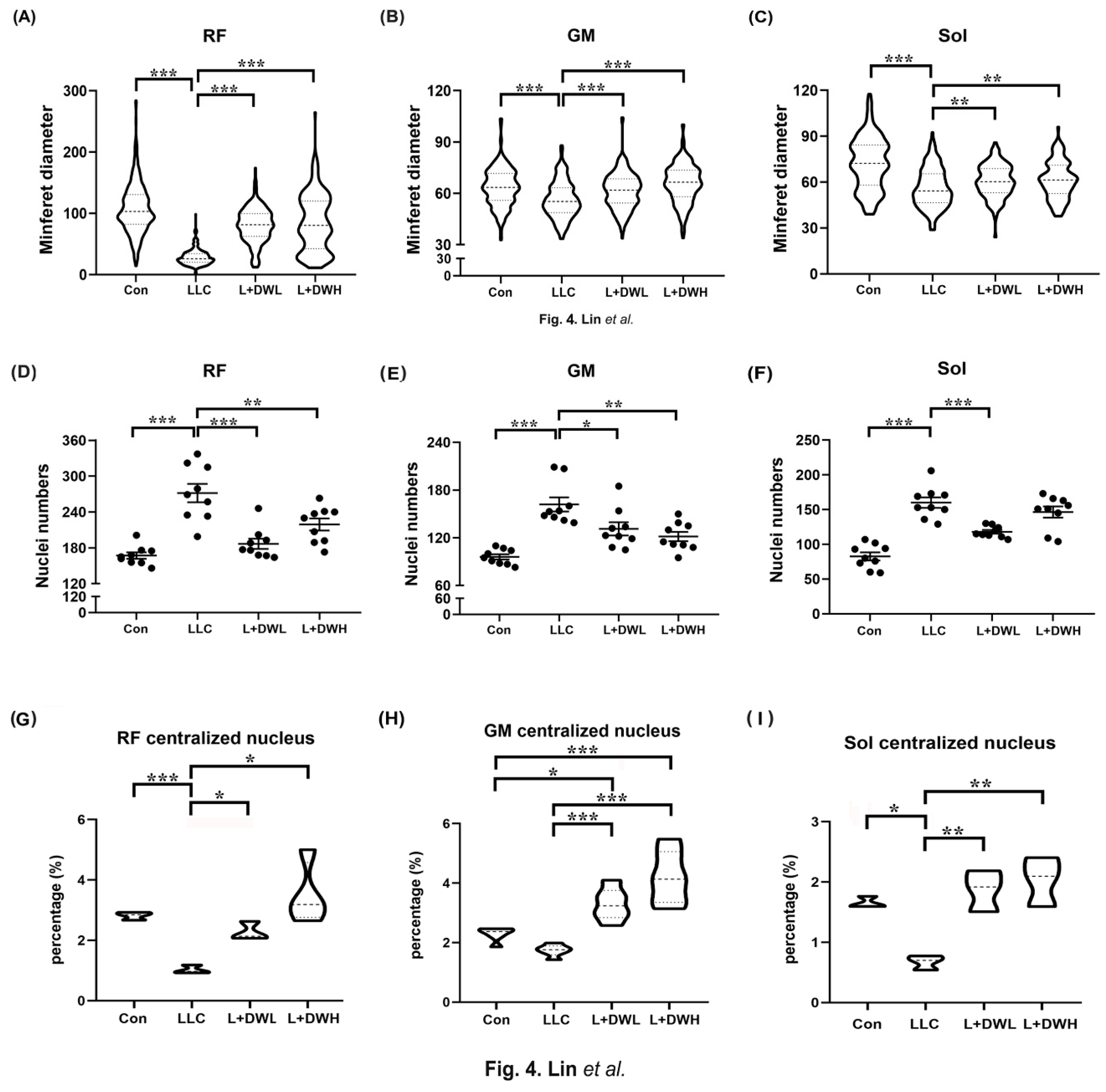
Figure 5.
Upregulation of serum IL-6 and TNF-alpha was inhibited by DTT water extracts in tumor-bearing mice. (A) After the euthanasia of mice on day 28, the serum of mice was collected, and the concentrations of the cytokines IL-6 and TNF-α were determined. (B) and (C) THP-1 cells were co-treated with LPS (100 ng/ml) with different doses of DTT water extracts for eight hours, the medium was then collected and subjected to ELISA to reveal the levels of IL-6 or TNF-α. ***p<0.001, **p<0.005, *p<0.05, multiple group comparisons were achieved by one-way ANOVA with a Tukey’s post hoc.
Figure 5.
Upregulation of serum IL-6 and TNF-alpha was inhibited by DTT water extracts in tumor-bearing mice. (A) After the euthanasia of mice on day 28, the serum of mice was collected, and the concentrations of the cytokines IL-6 and TNF-α were determined. (B) and (C) THP-1 cells were co-treated with LPS (100 ng/ml) with different doses of DTT water extracts for eight hours, the medium was then collected and subjected to ELISA to reveal the levels of IL-6 or TNF-α. ***p<0.001, **p<0.005, *p<0.05, multiple group comparisons were achieved by one-way ANOVA with a Tukey’s post hoc.
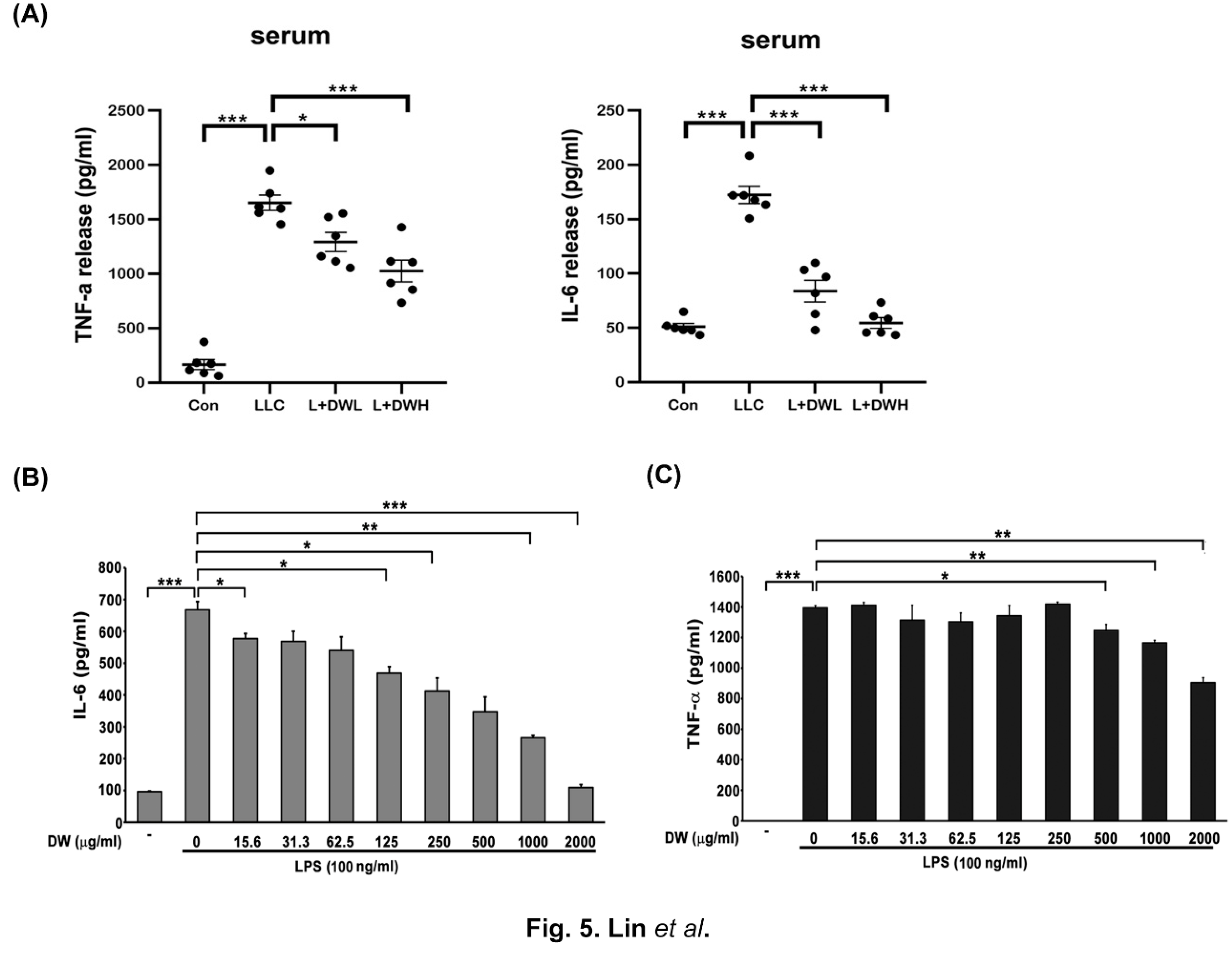
Figure 6.
The reductions in C2C12 myotubes by IL-6 were reversed in the presence of DTT water extracts. The C2C12 myoblasts were subjected to differentiation with 2% horse serum for 6 days with or without the presence of DTT water extracts (800 μg/ml); the differentiated cells were then treated with 100 ng/ml of IL-6 for another 2 days. (A) Representative images were revealed using Giemsa stain. (B) GEMSA positive signals were counted and calculated, ***p<0.001, **p<0.005. (C) The expressions of MyHC of different experimental C2C12 cells were revealed by Western blot using an anti-pan-MyHC or murf-1 antibody, respectively. The quantification results are shown in the bottom chart, ***p<0.001, *p<0.05.
Figure 6.
The reductions in C2C12 myotubes by IL-6 were reversed in the presence of DTT water extracts. The C2C12 myoblasts were subjected to differentiation with 2% horse serum for 6 days with or without the presence of DTT water extracts (800 μg/ml); the differentiated cells were then treated with 100 ng/ml of IL-6 for another 2 days. (A) Representative images were revealed using Giemsa stain. (B) GEMSA positive signals were counted and calculated, ***p<0.001, **p<0.005. (C) The expressions of MyHC of different experimental C2C12 cells were revealed by Western blot using an anti-pan-MyHC or murf-1 antibody, respectively. The quantification results are shown in the bottom chart, ***p<0.001, *p<0.05.
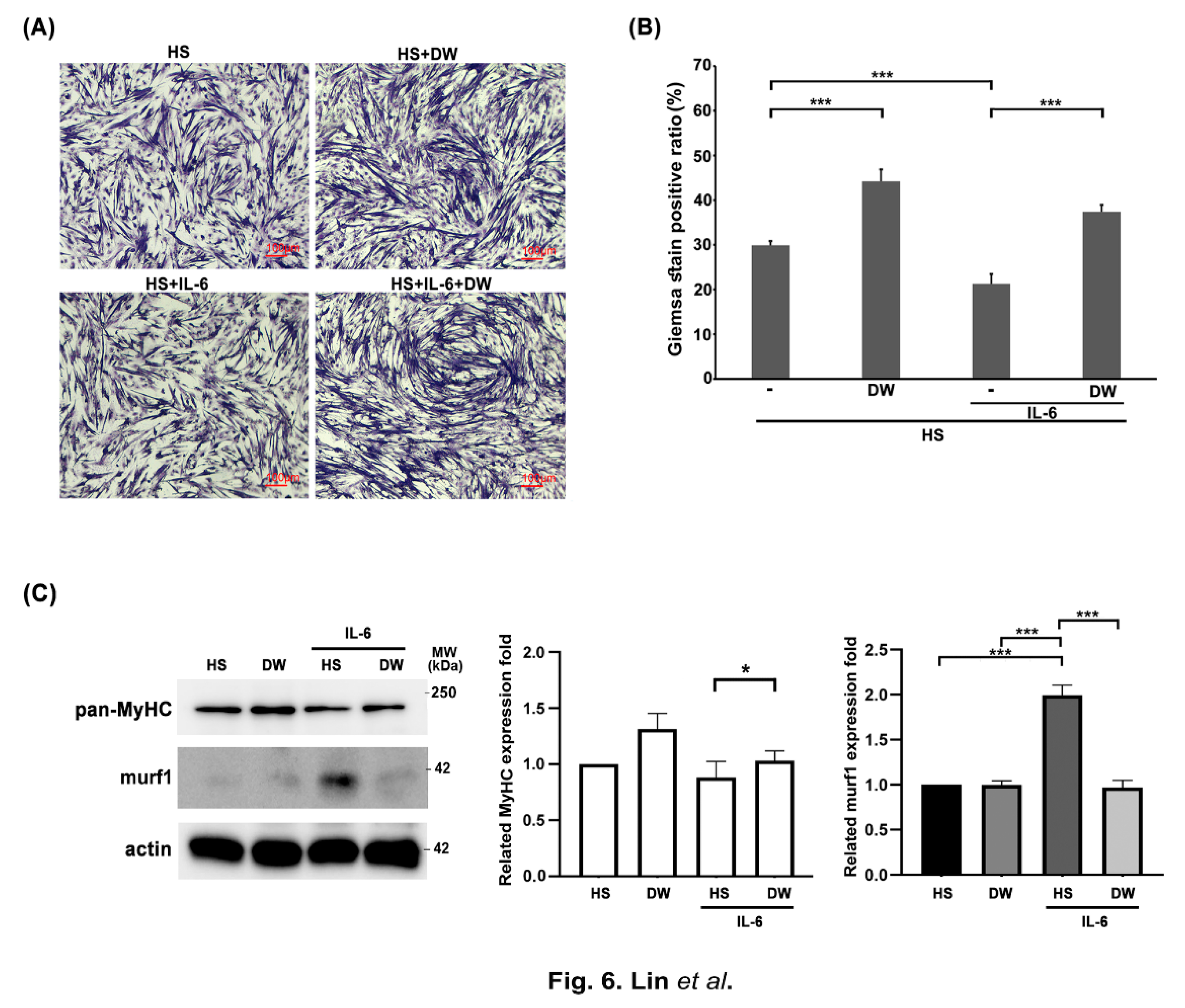
Figure 7.
Vitamin B1 acts as a potential functional ingredient of DTT water extracts to promote the multinucleation of C2C12 myotubes reduced by IL-6 (A) HPLC chromatogram revealed the different vitamin B derivatives of DTT water extracts. Peaks with an R time of 3.685, 5.242, 9.665, and 20.337 min corresponding to thiamine (vitamin B1), Pyrimidine HCl, niacinamide (vitamin B3), and Riboflavin (vitamin B2), respectively. (B) The C2C12 myoblast was subjected to differentiation with 2% horse serum for 6 days with or without the presence of DTT water extracts (800 μg/ml) or vitamin B1 (5 μM); the differentiated myotube was then treated with 100 ng/ml of IL-6 for another 2 days. Representative images were revealed by immunofluorescence staining with anti-pan-MyHC antibody. The red color indicates MyHC staining and blue color stands for nucleus. (C)The fusion and differentiation index were calculated and quantified, respectively. ***p<0.001, **p<0.005, multiple group comparisons were achieved by one-way ANOVA with Tukey’s post hoc. (D) The total and MyHC-positive nucleus numbers were calculated and quantified, ***p<0.001, **p<0.005, *p<0.05, and multiple group comparisons were achieved by one-way ANOVA with Tukey’s post hoc. (E) The numbers of myocytes and myotubes, the total and mean nuclei numbers of myotubes, and the distributions of the nuclei numbers of C2C12 myotubes were calculated and quantified, ***p<0.001, **p<0.005, *p<0.05, multiple group comparisons were achieved by two-way ANOVA with Tukey’s post hoc.
Figure 7.
Vitamin B1 acts as a potential functional ingredient of DTT water extracts to promote the multinucleation of C2C12 myotubes reduced by IL-6 (A) HPLC chromatogram revealed the different vitamin B derivatives of DTT water extracts. Peaks with an R time of 3.685, 5.242, 9.665, and 20.337 min corresponding to thiamine (vitamin B1), Pyrimidine HCl, niacinamide (vitamin B3), and Riboflavin (vitamin B2), respectively. (B) The C2C12 myoblast was subjected to differentiation with 2% horse serum for 6 days with or without the presence of DTT water extracts (800 μg/ml) or vitamin B1 (5 μM); the differentiated myotube was then treated with 100 ng/ml of IL-6 for another 2 days. Representative images were revealed by immunofluorescence staining with anti-pan-MyHC antibody. The red color indicates MyHC staining and blue color stands for nucleus. (C)The fusion and differentiation index were calculated and quantified, respectively. ***p<0.001, **p<0.005, multiple group comparisons were achieved by one-way ANOVA with Tukey’s post hoc. (D) The total and MyHC-positive nucleus numbers were calculated and quantified, ***p<0.001, **p<0.005, *p<0.05, and multiple group comparisons were achieved by one-way ANOVA with Tukey’s post hoc. (E) The numbers of myocytes and myotubes, the total and mean nuclei numbers of myotubes, and the distributions of the nuclei numbers of C2C12 myotubes were calculated and quantified, ***p<0.001, **p<0.005, *p<0.05, multiple group comparisons were achieved by two-way ANOVA with Tukey’s post hoc.
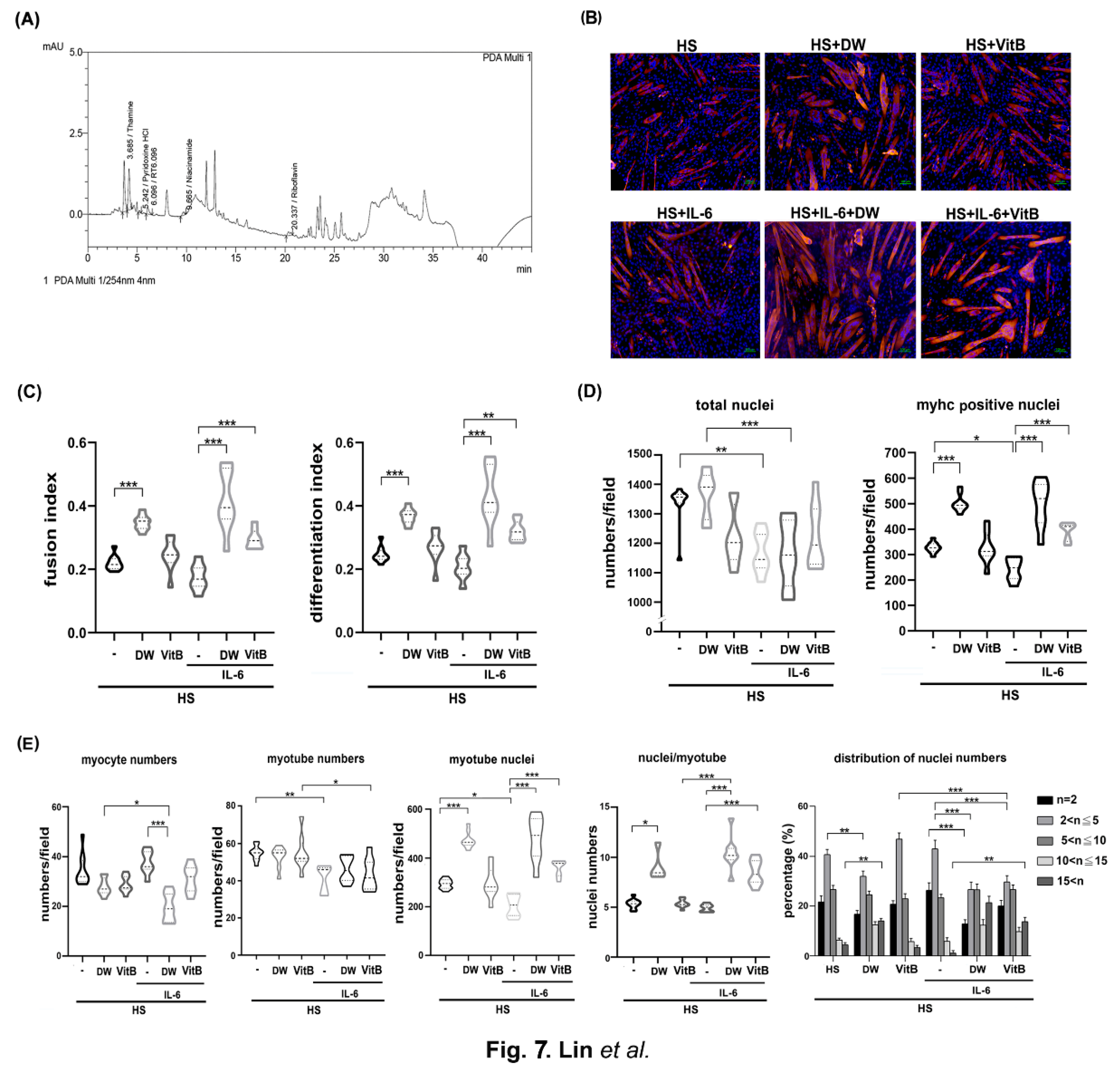
Figure 8.
E-cadherin/β-catenin is accompanied by multinucleations of C2C12 myotubes induced by DTT water extracts or vitamin B1. (A) The C2C12 myoblasts were subjected to differentiation for 6 days with or without the presence of DTT water extracts (800 μg/ml) or vitamin B1 (5 μM). Two hours before harvesting, cells were treated with cycloheximide followed by collection, extraction, and subjected to western blotting using anti-pan-MyHC, E-cadherin, β-catenin, or β-actin antibodies, respectively. The expression of β-actin served as an internal control. Quantification results of western blots are shown in the bottom chart, ***p<0.001, **p<0.005, *p<0.05 (B) The C2C12 myoblast was subjected to differentiation for 6 days with or without the presence of DTT water extracts (800 μg/ml) or vitamin B1 (5 μM); the differentiated myotube was then added with 100 ng/ml of IL-6 for another 2 days. Two hours before harvesting, cells were treated with MG132, and western blots were performed. Quantification results of western blots are shown in the bottom chart, ***p<0.001, **p<0.005, *p<0.05. The scRNA or E-cadherin siRNA-transfected C2C12 cells were subjected to differentiation for 6 days with or without DTT water extracts (800 μg/ml) or vitamin B1 (5 μM). The cells were then collected and subjected to (C) a western assay using an anti-E-cadherin antibody, the quantification results of western blot are shown as the bottom chart, **p<0.005, *p<0.05, or (D) immunofluorescence using anti-pan-MyHC antibody. (E) The fusion index, number of differentiation index, nuclei/myotube, and nuclei distribution ratio from C2C12 cells of different groups were counted and calculated. ***p<0.001, **p<0.005, *p<0.05, multiple group comparisons were achieved by two-way ANOVA with Tukey’s post hoc.
Figure 8.
E-cadherin/β-catenin is accompanied by multinucleations of C2C12 myotubes induced by DTT water extracts or vitamin B1. (A) The C2C12 myoblasts were subjected to differentiation for 6 days with or without the presence of DTT water extracts (800 μg/ml) or vitamin B1 (5 μM). Two hours before harvesting, cells were treated with cycloheximide followed by collection, extraction, and subjected to western blotting using anti-pan-MyHC, E-cadherin, β-catenin, or β-actin antibodies, respectively. The expression of β-actin served as an internal control. Quantification results of western blots are shown in the bottom chart, ***p<0.001, **p<0.005, *p<0.05 (B) The C2C12 myoblast was subjected to differentiation for 6 days with or without the presence of DTT water extracts (800 μg/ml) or vitamin B1 (5 μM); the differentiated myotube was then added with 100 ng/ml of IL-6 for another 2 days. Two hours before harvesting, cells were treated with MG132, and western blots were performed. Quantification results of western blots are shown in the bottom chart, ***p<0.001, **p<0.005, *p<0.05. The scRNA or E-cadherin siRNA-transfected C2C12 cells were subjected to differentiation for 6 days with or without DTT water extracts (800 μg/ml) or vitamin B1 (5 μM). The cells were then collected and subjected to (C) a western assay using an anti-E-cadherin antibody, the quantification results of western blot are shown as the bottom chart, **p<0.005, *p<0.05, or (D) immunofluorescence using anti-pan-MyHC antibody. (E) The fusion index, number of differentiation index, nuclei/myotube, and nuclei distribution ratio from C2C12 cells of different groups were counted and calculated. ***p<0.001, **p<0.005, *p<0.05, multiple group comparisons were achieved by two-way ANOVA with Tukey’s post hoc.
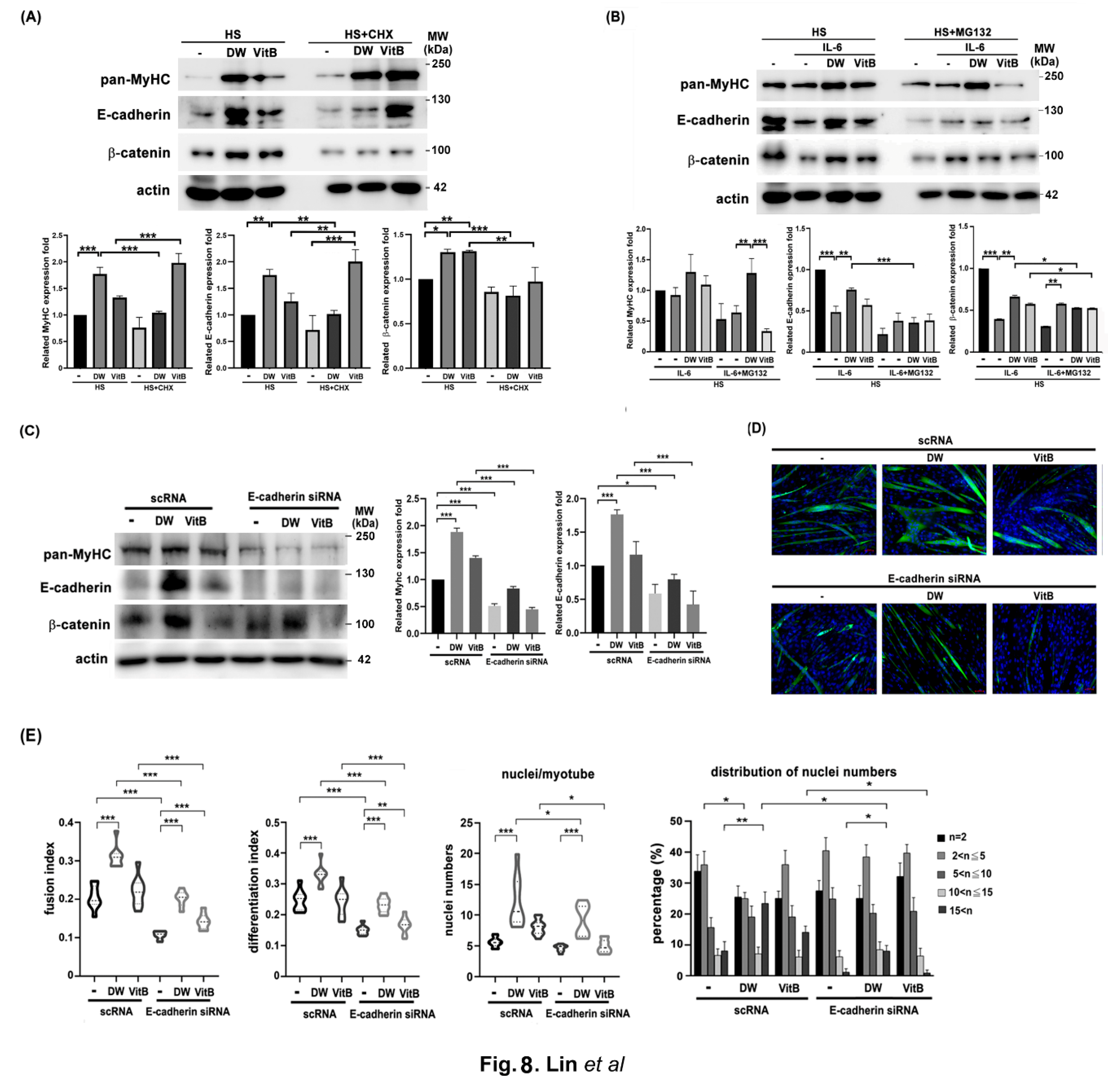
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



