
Submitted:
19 September 2025
Posted:
22 September 2025
You are already at the latest version
Abstract
The fields of RNA aptamers and epitranscriptomics have each emerged as powerful dimensions of RNA biology, yet their conceptual and mechanistic overlap remains largely unexplored. RNA aptamers as artificial single-stranded oligonucleotides selected through in vitro evolution (SELEX and related methods) are well known for their ability to fold into precise three-dimensional structures and bind specific targets with high affinity. These aptamers have been widely used in diagnostics, therapeutics, and molecular sensing. In parallel, the dynamic landscape of epitranscriptomic modifications, such as N⁶-methyladenosine (m⁶A), pseudouridine, and 5-methylcytosine, has been recognized as a critical regulator of RNA structure, stability, and function. In this perspective article, we chart unexplored territories at the interface of these two mechanisms, focusing on how RNA modifications may influence aptamer folding or binding, and how aptamers could, in turn, be harnessed to detect or modulate the epitranscriptomic state. We also highlight opportunities for cross-disciplinary integration in synthetic biology, post-transcriptional regulation, and RNA-based therapeutics. While the direct regulatory connections between these domains remain to be fully elucidated, we propose their potential convergence as a fertile ground for innovation. The primary aim of this article is to stimulate discussion within and between these research communities and to attract attention toward a promising frontier for future investigations.
Keywords:
RNA aptamers
; Epitranscriptomics
; RNA modifications
; RNA structure
; Post-transcriptional regulation
1. Introduction
In recent years, RNA biology has undergone a remarkable transformation, deriven by discoveries that have expanded our understanding of RNA beyond its traditional roles in coding, decoding, and catalysis. Two areas at the forefront of this shift, including RNA aptamer technology [1,2,3,4] and epitranscriptomics [5,6,7], have independently opened new avenues for molecular control, therapeutic development, and systems-level regulation of gene expression. However, despite their rapid advancement, these domains have largely evolved in parallel, with limited investigation into how they might conceptually or mechanistically intersect.
RNA aptamers, known as synthetic, short, and structured RNA oligonucleotide molecules selected through in vitro evolution techniques such as Systematic Evolution of Ligands by EXponential enrichment (SELEX), have been extensively utilized for their high affinity and specificity toward a broad range of molecular targets [3,8]. Their application in diagnostics [1], molecular imaging [9,10], targeted therapy [11,12], and biosensing [13,14] has been well documented. By adopting complex three-dimensional conformations, aptamers are capable of mimicking antibody-like behavior, yet retain the versatility and programmability inherent to nucleic acids [15].
Over the past decades, SELEX has been refined into several advanced variants, such as Cell-SELEX, Toggle-SELEX, High-Throughput SELEX (HT-SELEX), and Microfluidic SELEX, which is designed to improve specificity, speed, or physiological relevance of selection process [16]. These innovative approaches have enabled aptamer discovery for challenging and complex targets, including whole cells, small molecules, and membrane proteins, and facilitated integration with next-generation sequencing (NGS) and computational modeling [17,18,19]. Furthermore, novel non-SELEX selection methods, such as in vivo functional selection, which uses riboswitch-based reporters or survival assays, and machine learning-guided in silico aptamer prediction, are also emerging as promising tools for the rapid discovery and rational design of functional RNA binders [20,21]. These approaches offer alternatives to classical selection methods, particularly in contexts where functional output of the aptamers is prioritized. It is worth noting that RNA aptamer functions are critically dependent on RNA folding dynamics, secondary structure stability, and sequence-specific interactions, all of which are properties now known to be influenced by chemical modifications within the RNA itself [22]. In parallel, the field of epitranscriptomics has revealed that RNA molecules are subject to extensive and reversible chemical modifications that can impact their structure, localization, stability, and translational efficiency [7,23,24,25]. Numerous types of RNA modifications have been identified to date [26,27], and importantly, these modifications have been shown to influence RNA structure and interactions in both predictable and context-dependent ways [7,28,29].
Despite their structural impact, epitranscriptomic marks have received little attention regarding their effects on synthetic RNAs like aptamers. Conversely, the idea that aptamers could be designed to recognize specific RNA modifications or modulate modification-dependent functions remains underexplored. This gap presents an opportunity to integrate aptamer design with the chemical logic of the epitranscriptome to gain mechanistic insights and develop novel RNA tools (see Figure 1 and Figure 2). This perspective does not claim established links between aptamers and epitranscriptomics but highlights the theoretical basis and emerging evidence for their convergence. By examining how modifications influence aptamer function, or how aptamers might detect such states, new research directions may emerge. Framing this intersection aims to stimulate collaboration and uncover regulatory links and functional synergies that remain largely unexplored.
2. RNA Aptamers: Synthetic Precision in Targeting and Sensing
RNA aptamers have emerged as one of the most versatile synthetic tools in molecular biology and biotechnology [30,31]. They are typically generated through SELEX, the most widely used in vitro selection method. SELEX involves repeated rounds of binding, partitioning (which separates sequences bound to the target from unbound sequences), elution, and amplification to isolate high-affinity aptamers from large, randomized RNA libraries. During each SELEX cycle, sequences that bind to a specific molecular target are separated from non-binders, then reverse-transcribed, amplified, and transcribed back into RNA, progressively enriching the pool for functional aptamer candidates [32]. These short, single-stranded RNAs are capable of folding into well-defined three-dimensional structures that enable them to bind their targets with high specificity and affinity [33]. A wide array of molecules, including proteins, small metabolites, ions, and even other nucleic acids, can be targeted using aptamers, making them powerful agents for molecular recognition, sensing, and therapeutic modulation [34].
Central to their utility is the structural adaptability of RNA. Aptamers function by adopting complex secondary and tertiary structures, such as pseudoknots, stem-loops, and G-quadruplexes, which are essential for creating binding pockets or interaction surfaces that mimic those of natural receptors. The specificity of binding is not merely a function of nucleotide sequence, but also of precise spatial orientation enabled by the conformational plasticity of the RNA backbone [35,36]. Consequently, any factor that modulates RNA structure, either locally (e.g. nucleotide modifications) or globally (e.g. ionic conditions, folding motifs), can influence aptamer performance [34,37]. Recent advancements have enabled the design of aptamers with enhanced stability and function, including the use of chemical modifications (e.g., 2'-fluoro, 2'-O-methyl substitutions) to improve resistance to nucleases or to fine-tune binding characteristics [38,39]. While these modifications are typically introduced artificially during in vitro synthesis, there is growing interest in understanding how naturally occurring RNA modifications, such as those studied in epitranscriptomics, might influence aptamer behavior, particularly in endogenous or in vivo contexts.
Applications of aptamers have been diverse, ranging from diagnostic biosensors for infectious pathogens and cancer biomarkers to targeted therapeutics that inhibit specific protein-protein interactions or deliver drug payloads [33,40,41]. Aptamers have also been engineered into synthetic circuits for gene expression control and RNA imaging platforms, highlighting their modularity [42,43]. Despite this progress, aptamer design has largely proceeded independently of insights gained through epitranscriptomic research, a disconnect that may obscure subtle but impactful interactions between synthetic RNAs and the chemical environment of the transcriptome [44,45]. Given that RNA aptamers are, ultimately, RNA molecules functioning within complex cellular environments, a deeper understanding of how naturally occurring RNA modifications alter their folding and interaction dynamics may provide crucial insights. This could become particularly relevant as aptamers are increasingly used in cellulo and in vivo diagnostic, and therapeutic applications (theranostics), where they are exposed to endogenous RNA-modifying enzymes and RNA-binding proteins [8,46]. This potential intersection calls for closer investigation into the compatibility between aptamer function and the modified cellular transcriptome.
3. Epitranscriptomics: Decoding the Chemical Language of RNA Modifications
Epitranscriptomics refers to the study of biochemical modifications on RNA molecules that alter their fate and function without changing the underlying sequence [7,23,24,25]. To date, over 170 modifications have been identified, and they are now recognized as key regulators of post-transcriptional gene expression, affecting processes such as splicing, translation, localization, degradation, and structural dynamics [26,27]. Among the most studied modifications are N⁶-methyladenosine (m⁶A), 5-methylcytosine (m⁵C), pseudouridine (Ψ), and N¹-methyladenosine (m¹A), each of which can be deposited, interpreted, or removed by dedicated enzymatic machineries termed writers, readers, and erasers [6]. The dynamic and reversible nature of these modifications enables cell to fine-tune gene expression in response to developmental cues, environmental stressors, or disease states [47,48,49,50,51,52,53,54,55]. For example, m⁶A has been implicated in the regulation of mRNA stability and translation efficiency, often through interactions with reader proteins such as YTHDF1-3 [56]. Likewise, Ψ has been shown to enhance base-pairing stability and promote translational fidelity [57,58], while m⁵C can influence RNA export and transcript stability [59]. These modifications can reshape both local and global RNA structure, sometimes inducing conformational switches that affect binding by proteins or ribonucleoprotein complexes [60,61]. The structural consequences of RNA modifications are of particular relevance to both natural and synthetic RNAs. Modifications can either stabilize or destabilize secondary structure motifs depending on their chemical nature and position within the transcript [62]. For instance, m⁶A is known to weaken A–U base pairs and destabilize local duplexes, facilitating the formation of single-stranded regions that may be recognized by RNA-binding proteins or non-coding RNAs [60,63,64]. This structural flexibility raises compelling questions about how modifications might similarly affect the folding and function of synthetic RNA tools such as aptamers.
Technologies for mapping RNA modifications have rapidly evolved, ranging from antibody-based immunoprecipitation methods (e.g., m⁶A-seq) to direct detection using nanopore sequencing or chemical labeling approaches [65,66]. However, functional studies on how these modifications modulate interactions with structured RNAs, such as aptamers, remain relatively scarce. It remains unclear, for example, whether endogenous modification patterns are sufficiently conserved or dynamic to impact aptamer-based assays or therapeutics deployed in vivo [67,68]. Furthermore, the cellular context in which aptamers function often includes active RNA-modifying enzymes that could potentially install modifications onto synthetic RNAs, especially when they are delivered exogenously or transcribed intracellularly [69,70]. The interplay between such modifications and aptamer behavior, whether enhancing, impairing, or modulating their function, has yet to be systematically explored. Thus, the epitranscriptome is not merely a background feature of the cellular environment but may serve as a regulatory layer with implications for any RNA-based technology [44]. A more integrated understanding of how this chemical language intersects with engineered RNAs like aptamers could illuminate novel dimensions of RNA control and inspire innovative approaches in biotechnology and therapeutics.
4. Structural Plasticity and Modification Sensitivity: A Conceptual Bridge
The structural adaptability of RNA molecules is essential to their diverse functions [71]; RNA aptamers, in particular, rely on stable and predictable folding to exert their binding capabilities. These structures are determined not only by base-pairing rules but also by the conformational preferences encoded in the nucleotide sequence [39]. Increasingly, it has become evident that chemical modifications, such as those observed in the epitranscriptomic landscape, can influence RNA structure in ways that may be relevant to both endogenous and synthetic RNAs, including aptamers [70].
Post-transcriptional modifications often affect local folding by altering hydrogen-bonding potential, base stacking, or backbone flexibility [72,73,74]. For instance, N⁶-methyladenosine (m⁶A) tends to destabilize RNA duplexes and promote single-stranded configurations; this can lead to shifts in the secondary structure that modulate access to binding sites or regulatory motifs [60,63,75]. Pseudouridine, by contrast, increases the rigidity of local regions; its ability to form additional hydrogen bonds can stabilize helices and influence long-range interactions [57,76,77]. These structural effects can propagate through an RNA molecule, creating subtle but functionally significant changes in tertiary architecture [77,78].
In the context of RNA aptamers, even minor structural disturbances can disrupt binding affinity or specificity. Typically, an aptamer’s function depends on the precise orientation of nucleotides within a folded domain. This folding creates complex three-dimensional structures formed by base-paired secondary elements (such as loops, stems, bulges, hairpins, and G-quadruplexes) [79], that enable highly specific molecular recognition through non-covalent interactions including hydrophobic and van der Waals forces, hydrogen bonding, electrostatic interactions, base stacking, and shape complementarity [80]. Thus, if a naturally occurring modification alters that geometry, either directly or allosterically, the aptamer’s ability to recognize its target could be compromised or, under certain conditions, enhanced [35]. Such a scenario suggests that epitranscriptomic marks could act as silent regulators of aptamer behavior, particularly in cellular environments where endogenous modifying enzymes are active. This idea is supported by previous studies demonstrating m6A-modified aptamers can exhibit changes in binding affinity or fluorescence upon enzymatic demethylation by RNA-modifying enzymes in cells [81,82].
The concept of a "modification-sensitive aptamer" has not yet been fully developed; however, several studies have indirectly hinted at the relevance of chemical context for RNA binding functionality [67,83,84]. Structural probing techniques such as SHAPE (Selective 2′-Hydroxyl Acylation analyzed by Primer Extension) have demonstrated that m⁶A and other modifications can shift RNA reactivity profiles; this implies a broader influence on folding pathways [85,86]. In the realm of aptamers, such effects could alter loop closure, pseudoknot formation, or other motifs necessary for high-affinity binding [87,88]. Experimental models for modification-sensitive aptamers could be developed by comparing aptamer performance in modified versus unmodified RNA backgrounds [22,89]. Synthetic systems could also be engineered to test how insertion of specific marks at key structural positions influences function. These models would allow for systematic evaluation of how epitranscriptomic environments modulate aptamer efficacy [90,91,92]. The significance of such interactions extends beyond structural interest; it suggests a potential bidirectional influence in which aptamers could be either regulated by, or responsive to, chemical signatures embedded in the transcriptome. Exploring these structural bridges can set the foundation for understanding compatibility, predictability, and robustness of aptamer-based applications in vivo [67].
5. RNA Modifications as Modulators or Targets of Aptamer-Based Tools
The recognition that RNA modifications influence not only the fate of transcripts but also their molecular interactions raises an important question: can these modifications be directly targeted by aptamer-based tools? While most aptamers are currently selected against proteins, small molecules, or ions, the concept of developing aptamers to recognize modified RNA bases remains largely unexplored, yet conceptually sound [67,93,94]. RNA modifications represent chemically distinct features that could, in principle, serve as epitopes for aptamer recognition [3,95,96]. The presence of a methyl group on adenosine (as in m⁶A) or an isomerized uridine (as in Ψ) introduces differences in hydrogen bonding patterns and steric properties that could be selectively bound by a structured RNA aptamer [82,97,98,99,100]. In theory, aptamers could be designed to distinguish between modified and unmodified bases within otherwise identical sequence contexts; such specificity would have profound implications for diagnostic and mechanistic applications. For instance, a selected RNA aptamer so called A-Pepper exhibits fluorescence changes depending on m⁶A status, and Broccoli-based assays similarly differentiate modified from unmodified RNA, demonstrating that aptamer-based tools can, resolve chemical differences arising from epitranscriptomic modifications [82,101].
Although few studies have directly attempted direct aptamer selection against RNA modifications, some conceptual precedents exist from DNA aptamer research, where modified nucleotides have been targeted successfully. For instance, DNA aptamers have been selected to bind specifically to methylated DNA sequences (e.g. methylated CpG dinucleotide sites) [102,103]. These examples suggest that, in principle, selective detection of modified versus unmodified bases is feasible. Expanding such efforts into RNA aptamers could provide valuable tools for detecting or isolating modified transcripts. These aptamers could be incorporated into biosensors or affinity capture platforms, enabling studies of RNA modification dynamics in live cells or complex mixtures [104].
Beyond sensing, there is also interest in whether RNA modifications could act as endogenous regulators of aptamer activity. Aptamers that are transcribed within cells, either from expression constructs or viral vectors, may acquire modifications from endogenous writer enzymes. If these modifications occur at functionally relevant positions, they could alter binding capacity or structural stability as shown in experimental studies and molecular dynamics (MD) simulations where specific chemical substitution in thrombin-binding aptamer significantly changed its structural stability and target affinity [105,106]. While this is often considered an undesirable side effect, it could also be repurposed as a control mechanism: aptamers could be designed to remain inactive until modified, thereby acting as on/off switches or conditional regulators. For instance, MD simulations of a thrombin-binding aptamer incorporating 5-furyl-2′-deoxyuridine revealed that the position of the modification within the sequence critically determines its impact on the aptamer–protein interaction sensitivity by modulating structural dynamics at the modification site, altering solvent exposure, influencing stacking with neighboring bases, and affecting interactions with thrombin, all without disrupting the global conformation of the aptamer [107].The design of such responsive systems would require a deeper understanding of the sequence preferences of modifying enzymes and the structural outcomes of various marks. It would also necessitate improved methods for site-specific installation of modifications in vitro, as well as the ability to map and verify modification status in cellular RNA populations at high resolution [108,109]. These tools are under active development in the epitranscriptomics field; their integration into aptamer design workflows could facilitate a new class of chemically-responsive RNA devices. In this conceptual framework, RNA modifications are not just obstacles to be accounted for, but potential control points or targets for aptamer-based strategies. Whether used to sense, respond to, or exploit the modified state of transcripts, aptamers could offer precise and programmable solutions for probing the epitranscriptome.
6. Potential for Synthetic Biology and Theranostics Integration
The growing interest in RNA technologies has positioned both RNA aptamers and epitranscriptomics as promising components of future synthetic biology systems. In this context, aptamers can be viewed as programmable elements capable of sensing and controlling molecular events [110,111], while epitranscriptomic modifications serve as regulatory inputs that shape RNA behavior [61,112]. When brought into conceptual proximity, these two systems suggest the potential for creating hybrid regulatory frameworks that are both chemically tunable and highly modular. RNA aptamers have already been incorporated into a variety of synthetic circuits; examples include small-molecule-responsive riboswitches [113,114], aptamer-controlled gene expression switches [115], and ligand-triggered RNA scaffolds for signal integration [116,117]. In many cases, the response of these systems relies on conformational changes that occur upon target binding [114,118]. This inherent sensitivity to structural transitions makes them potentially responsive to endogenous cues introduced by epitranscriptomic marks. For instance, modifications such as base methylation can work as internal signals by altering the secondary structure of RNA [60,119]. Typically, the addition of N6A-methyladenosine (m6A) on an RNA can change the base pairing and refold the RNA locally, a phenomenon known as the m6A-switch [63]. These modifications can make the previously hidden sequences accessible, or vice versa, in a way that resembles the conformational changes caused by ligand binding in aptamers [63,120]. Therefore, if an aptamer or its adjacent sequence is modified by a chemical mark through epitranscriptomic processes, the resulting conformational change could potentially modulate the aptamer’s binding or regulatory output [83,121]. In therapeutic contexts, aptamers have been explored as alternatives to antibodies for targeted delivery, inhibition of disease-associated proteins, and modulation of signaling pathways [122,123]. Several RNA aptamers have reached clinical evaluation stages, such as Pegaptanib a Food and Drug Administration (FDA) approved RNA aptamer medication to treat neovascular age-related macular degeneration by binding and inhibiting vascular endothelial growth factor (VEGF) [122,124]. However, in vivo functionality of aptamers often depends on both resistance to degradation and maintenance of structural fidelity, which are influenced by the cellular environment. In fact, aptamers selected in vitro sometimes fail to perform in vivo due to rapid nuclease digestion, suboptimal ionic conditions, or competition with other biomolecules [122].In this setting, RNA chemical modifications may either support or interfere with aptamer efficacy depending on the location and type of chemical marks introduced. For instance, incorporating 2′-fluoropyrimidines or 2′-O-methyl groups at the ribose 2′-position greatly increases nuclease resistance and serum stability without compromising target binding affinity [38,125]. However, addition of a locked nucleic acid (LNA) nucleotide into the loop of a thrombin-binding DNA aptamer was shown to stop its ability to bind thrombin [126]. Therefore, if the relationship between aptamer activity and RNA modification status can be better understood, opportunities may arise for engineering theranostic systems that are conditional upon the epitranscriptomic state of the target cell. For instance, aptamers could be engineered to function only in cells with elevated levels of m⁶A or other modifications, introducing a form of endogenous selectivity and improving therapeutic precision [81]. Conversely, aptamers could be used to block or enhance the activity of reader proteins, creating downstream effects on the stability or translation of modified transcripts. For example, a short RNA decoy corresponding to HIV’s TAR sequence was shown to bind and sequester the viral Tat protein, thereby inhibiting Tat’s downstream effects on gene expression [127].
Synthetic biology platforms could also benefit from aptamers that are sensitive to RNA modification states. In such designs, RNA circuits could include structural domains whose behavior depends on the presence or absence of specific marks, allowing cells to respond to physiological or pathological conditions in real time [81,82,128,129,130]. Regulatory switches could thus be activated or silenced based on the modification profile, introducing a novel axis of control that bridges molecular sensing and functional output [131,132]. The success of such applications will depend on the ability to map modification patterns accurately, identify modification hotspots within aptamer sequences, and predict structure-function outcomes [133,134,135]. While these requirements present technical challenges, advances in direct RNA sequencing [136,137], structural modeling [135], and in vivo RNA tracking [138] offer potential solutions. By incorporating RNA modifications as variables in aptamer design and deployment, synthetic biology and therapeutic systems may gain an additional layer of control that is both dynamic and biologically grounded.
7. Technological Convergence: Detecting, Mapping, and Manipulating Modifications
Technological advances in RNA research have brought aptamer-based platforms and epitranscriptomic mapping tools closer to potential integration [81,101,139]. This convergence offers practical opportunities for improving how RNA modifications are detected, quantified, and manipulated, especially in complex or living systems [94,134]. RNA aptamers, with their high specificity and tunable binding properties, can serve as customizable tools for addressing some of the limitations faced by current epitranscriptomic techniques. One of the main challenges in epitranscriptomics is the detection of modifications at single-nucleotide resolution [140]. Traditional antibody-based techniques, such as m⁶A-seq or MeRIP-seq, provide general information about modification enrichment but suffer from low resolution and cross-reactivity. Emerging methods such as direct RNA sequencing using nanopore technology offer site-specific information [141], yet they often require high RNA input and complex signal processing. In this landscape, RNA aptamers could fill a unique niche in epitranscriptomics. If aptamers are selected or engineered to bind specific modified nucleotides or the modified form of a transcript, they could be deployed as molecular probes for direct detection of epitranscriptomic states. This has been demonstrated a m6A-sensitive fluorescent RNA aptamer system, known as A-pepper, which reports m6A enzymes’ activity in vitro and in live cells based on fluorescent signal enhancement resulting from the methylation and demethylation of the aptamer [101]. Aptamers could also be used for affinity capture to isolate modified RNAs from a mixture, then enrichment could be followed by high-throughput sequencing to identify modification patterns. The feasibility of such systems has been shown by the selection of L-RNA aptamers that can selectively capture structured native RNA (D-RNA) [142,143]. Fluorescently labeled aptamers could also be used in imaging platforms to visualize modifications in fixed or live cells. For instance, fluorescent light-up aptamers (e.g., the Mango and Pepper families) have already demonstrated high-contrast RNA imaging in cells, indicating their potential for visualizing RNA modification dynamics [144,145,146]. In principle, an aptamer that binds a modified base, but not to the unmodified base, can be tagged with a fluorophore to serve as a fluorescent probe. For instance, DNA aptamers have already been selected to recognize specific post-translational modifications, such as particular histone acetylation with high selectivity and have been used in atomic force microscopy-based imaging of these modifications [147]. By extension, fluorescent RNA aptamer-based sensors could be developed to track RNA base modifications in cells, enabling researchers to track modification dynamics in response to stimuli or disease progression in real-time. Such applications would require aptamers that can discriminate between modified and unmodified bases, a level of specificity that remains technically challenging but conceptually feasible [81,101,148]. In this regard, aptamers have shown the ability to distinguish molecules differing by only a single methyl group. For example, researchers have developed an RNA aptamer that binds theophylline about 10,000-fold more tightly than caffeine, despite caffeine having just one extra methyl group, supporting their potential role in epitranscriptomic detection tools [149].
Another potential direction involves using aptamers to manipulate the activity of the enzymes responsible for writing, reading, or erasing RNA modifications. Aptamers that bind to m⁶A writers like METTL3 [150], readers like YTHDF proteins [93], or erasers like ALKBH5 and FTO [81,82] could be used to inhibit or redirect their activity in a programmable manner. This type of aptamer function would parallel the use of inhibitory antibodies or small molecules but would offer improved tunability and intracellular delivery potential, especially if expressed as part of an RNA-based therapeutic system [37,110]. Furthermore, aptamer-controlled systems could be designed to act as responsive elements in RNA editing tools [131,151,152,153]. For instance, aptamers could gate the activity of an RNA-editing enzyme by binding a modification or its corresponding reader protein; such gating could be used to control editing efficiency or spatial targeting [152,154]. These systems would be useful in precision medicine, where therapeutic interventions must be tightly controlled and context-specific. The technological merging of aptamer platforms and epitranscriptomic analysis tools represents an underdeveloped but promising area. For this potential to be realized, more research is needed to develop high-affinity aptamers for modified bases, optimize delivery systems for aptamer-based tools, and integrate these elements into existing transcriptomic workflows. If successful, such integration could yield a powerful suite of molecular instruments for observing and controlling the RNA modification landscape.
8. Challenges and Considerations for Future Research
While the conceptual link between RNA aptamers and epitranscriptomics offers promising avenues for innovation, several challenges must be addressed before meaningful integration can occur. These span technical limitations and gaps in fundamental understanding, each requiring careful consideration to avoid premature conclusions or inefficient designs. One major limitation is the lack of systematic data on how endogenous RNA modifications affect synthetic aptamer structure and function. Most aptamer studies are performed in vitro under defined conditions, which do not reflect the chemical complexity of cellular RNA environments [155]. Aptamers that function reliably in buffer may behave unpredictably in cells, where post-transcriptional modifications and RNA-binding factors abound [122]. Without experimental systems that simulate modified RNA contexts, aptamer compatibility with the epitranscriptome remains unclear. Moreover, intracellularly expressed aptamers could become substrates for RNA-modifying enzymes. Whether such modifications occur randomly or follow enzyme-specific sequence preferences is poorly understood, as this has not been systematically studied. If key positions in an aptamer are chemically altered, e.g., by m6A methylation, binding affinity or structure may be disrupted. For instance, a Broccoli aptamer modified with m6A lost fluorescence until a demethylase removed the mark [82]. To improve stability and pharmacokinetics, chemical modifications like 2′-fluorinated pyrimidines, 2′-O-methyl ribose, or PEGylation have been employed. However, these do not always prevent enzymatic targeting and may not be compatible with in vivo expression [156].
Another challenge lies in selecting aptamers that recognize modified nucleotides with high specificity. Modified and unmodified bases often differ subtly, and aptamer binding depends on more than base identity; factors like structural context, solvent accessibility, and competition from cellular components also play roles [68,157]. Addressing this may require adapting SELEX protocols, for example by incorporating modified RNA libraries (e.g., 2′-fluoro, 2′-O-methyl, base analogs) to broaden interaction profiles and improve discrimination [158]. Structurally biased libraries may further enrich aptamers with desired conformations, enhancing their ability to distinguish modified from unmodified bases [159].
Integration into epitranscriptomic studies also raises concerns about off-target effects. Even highly specific aptamers may bind unintended targets in cellular contexts, especially if altered by endogenous modifications or protein interactions [160,161]. A methylated RNA aptamer, for example, failed to bind its ligand until demethylated [82]. Similarly, the DNA aptamer AS1411, targeting nucleolin, was found to bind fibronectin 1 in serum, negating its intended in vivo function [160]. These examples emphasize the need for thorough characterization under physiological conditions [161]. On the epitranscriptomic side, accurately mapping RNA modifications at single-base resolution remains a challenge [162,163]. Without spatial and temporal precision, predicting aptamer–modification interactions is difficult. Advances in direct RNA sequencing and structure-aware detection are helping, but reliable maps are still lacking.
Ethical and safety issues may also emerge in therapeutic settings [164,165,166]. If aptamer activity depends on modification status, misregulated epitranscriptomic profiles could cause unintended effects. Rigorous validation and control systems will be essential for clinical applications. Nonetheless, these challenges are not insurmountable. Addressing them systematically may enable the next generation of RNA tools that function within the chemical logic of the epitranscriptome.
9. Conclusions
RNA aptamers and epitranscriptomics are powerful yet largely separate areas of RNA biology. Aptamers offer programmable specificity and structural adaptability, while epitranscriptomic modifications serve as a chemical language regulating RNA behavior. Though each field has advanced independently, their intersection may hold untapped potential for research and innovation. This perspective explores how RNA modifications might affect aptamer folding or interactions, and how aptamers could detect or respond to modified RNA states, forming feedback between recognition and regulation. Applications in synthetic biology and therapeutics are discussed, along with conceptual and technical challenges. Rather than a conclusion, this article invites researchers across RNA structure, sensing, and synthetic biology to explore these overlaps and develop tools to probe or exploit the modified transcriptome. Bridging these domains may deepen our understanding of post-transcriptional regulation, enable smarter therapeutic systems, and blur the line between endogenous RNA control and synthetic design, opening new directions for RNA research.

Table 1.
Potential intersections between RNA aptamer function and epitranscriptomic mechanisms.
| Feature | RNA Aptamers | Epitranscriptomics | Possible Intersection |
|---|---|---|---|
| Functional dependency | Structure-dependent binding | Structure-dependent regulation | RNA modifications may alter aptamer folding or stability |
| Selection/design methods | SELEX-based in vitro evolution | Genetic, enzymatic, or chemical modification | Modified RNA libraries could be used for aptamer selection |
| Key chemical elements | Unmodified or chemically stabilized nucleotides | m⁶A, Ψ, m⁵C, etc. | Aptamers could be sensitive to or recognize these modifications |
| Role in gene regulation | Target inhibition; molecular recognition | mRNA splicing, stability, translation | Combined tools may control gene expression in a condition-specific manner |
| Therapeutic use | Targeted drug delivery; inhibition of proteins | Cancer, neurological diseases, immune regulation | Aptamer-based delivery of epitranscriptome-modifying agents |
| Response to cellular environment | May be degraded or misfolded | Responds to stress, development, signaling | Aptamer activity may vary depending on local modification state |
| Potential for biosensing | High specificity, real-time detection | Biomarker potential (modification levels) | Aptamers as sensors for detecting RNA modification patterns |
| Compatibility with in vivo systems | Requires stabilization strategies | Naturally occurring mechanisms | Modified-state-responsiveness could be integrated into aptamer designs |
| Regulatory feedback loops | Currently limited to design logic | Present in many cellular pathways | Aptamers could be designed to modulate reader/writer proteins |
| Application overlap | Synthetic biology, diagnostics, therapeutics | Functional genomics, disease mechanisms | Cross-disciplinary innovation in RNA-based systems |
Box 1. Key concepts and definitions
RNA Aptamers: Short, single-stranded RNA molecules
selected for their ability to bind specific targets with high affinity; their
function depends on precise 3D folding.
SELEX (Systematic Evolution of Ligands by EXponential
enrichment): An in vitro method for selecting aptamers from large RNA
libraries through iterative binding and amplification cycles.
Epitranscriptomics: The study of reversible chemical
modifications on RNA that influence structure, stability, localization, and
translation without altering sequence.
m⁶A (N⁶-methyladenosine): A common RNA modification
that modulates transcript fate by destabilizing local structures and
attracting specific reader proteins.
Pseudouridine (Ψ): An isomer of uridine that
enhances RNA backbone rigidity and base pairing; commonly found in non-coding
and coding RNAs.
RNA Modifications: Covalent chemical changes to RNA
bases; installed by writers, interpreted by readers, and removed by erasers
in a dynamic manner.
Writers, Readers, Erasers: Enzymes that install
(writers), recognize (readers), or remove (erasers) RNA modifications to
regulate RNA behavior.
RNA Secondary Structure: Local base pairing patterns
(e.g., stem-loops, bulges) that influence RNA stability and interaction
potential.
Aptamer Folding: The process by which aptamers adopt
specific 3D shapes essential for target binding; sensitive to sequence and
chemical context.
Structural Plasticity: The ability of RNA to adopt
multiple conformations depending on cellular or chemical cues; often altered
by modifications.
RNA Biosensors: Aptamer-based tools designed to
detect specific molecules, ions, or RNA modifications in vitro or in cells.
RNA-based Therapeutics: Strategies that use RNA
molecules (e.g., aptamers, siRNAs) to treat disease by modulating molecular
pathways.
Modification-sensitive Aptamers: Hypothetical or
designed aptamers whose structure or function is altered by the presence of
RNA modifications.
RNA Modification Mapping: Techniques used to
identify and locate chemical modifications across the transcriptome; often
requiring enrichment or sequencing.
Direct RNA Sequencing: A method that reads RNA
molecules without reverse transcription, allowing detection of certain base
modifications in situ.
Targeted RNA Delivery: The use of aptamers to guide
therapeutic cargos to specific cells or tissues based on target recognition.
Chemical Probing (e.g., SHAPE): Methods to study RNA
structure by modifying reactive groups; used to infer folding and
flexibility.
Aptamer-Protein Interactions: Non-covalent bindings
between aptamers and proteins; used for inhibition, sensing, or targeted
modulation.
Context-aware RNA Design: The development of RNA
molecules that function reliably within the biochemical environment of living
cells, including modification states.
References
- Kang KN, Lee YS. RNA Aptamers: A Review of Recent Trends and Applications. Adv Biochem Eng Biotechnol [Internet]. Springer, Berlin, Heidelberg; 2012 [cited 2025 Aug 19];131:153–69. Available from: https://link.springer.com/chapter/10.1007/10_2012_136.
- Lu X, Kong KYS, Unrau PJ. Harmonizing the growing fluorogenic RNA aptamer toolbox for RNA detection and imaging. Chem Soc Rev [Internet]. Chem Soc Rev; 2023 [cited 2025 Jun 6];52:4071–98. Available from: https://pubmed.ncbi.nlm.nih.gov/37278064/. [CrossRef]
- Razlansari M, Jafarinejad S, rahdar A, Shirvaliloo M, Arshad R, Fathi-Karkan S, et al. Development and classification of RNA aptamers for therapeutic purposes: an updated review with emphasis on cancer. Mol Cell Biochem 2022 4787 [Internet]. Springer; 2022 [cited 2025 Aug 19];478:1573–98. Available from: https://link.springer.com/article/10.1007/s11010-022-04614-x. [CrossRef]
- Lei X, Xia Y, Ma X, Wang L, Wu Y, Wu X, et al. Illuminating RNA through fluorescent light-up RNA aptamers. Biosens Bioelectron. Elsevier; 2025;271:116969. [CrossRef]
- Cerneckis J, Ming GL, Song H, He C, Shi Y. The rise of epitranscriptomics: recent developments and future directions. Trends Pharmacol Sci [Internet]. Elsevier Ltd; 2024 [cited 2025 Apr 20];45:24–38. Available from: https://www.cell.com/action/showFullText?pii=S0165614723002547. [CrossRef]
- Wiener D, Schwartz S. The epitranscriptome beyond m6A. Nat Rev Genet 2020 222 [Internet]. Nature Publishing Group; 2020 [cited 2025 Apr 5];22:119–31. Available from: https://www.nature.com/articles/s41576-020-00295-8. [CrossRef]
- Arzumanian VA, Dolgalev G V., Kurbatov IY, Kiseleva OI, Poverennaya E V. Epitranscriptome: Review of Top 25 Most-Studied RNA Modifications. Int J Mol Sci [Internet]. MDPI; 2022 [cited 2025 Apr 5];23:13851. Available from: https://www.mdpi.com/1422-0067/23/22/13851/htm. [CrossRef]
- Germer K, Leonard M, Zhang X. RNA aptamers and their therapeutic and diagnostic applications. Int J Biochem Mol Biol [Internet]. 2013 [cited 2025 Aug 15];4:27. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC3627066/.
- Chen W, Zhao X, Yang N, Li X. Single mRNA Imaging with Fluorogenic RNA Aptamers and Small-molecule Fluorophores. Angew Chemie [Internet]. John Wiley & Sons, Ltd; 2023 [cited 2025 Jun 6];135:e202209813. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/ange.202209813. [CrossRef]
- Lin B, Xiao F, Jiang J, Zhao Z, Zhou X. Engineered aptamers for molecular imaging. Chem Sci [Internet]. Royal Society of Chemistry; 2023 [cited 2025 Aug 19];14:14039–61. Available from: https://pubs.rsc.org/en/content/articlehtml/2023/sc/d3sc03989g. [CrossRef]
- Sundaram P, Kurniawan H, Byrne ME, Wower J. Therapeutic RNA aptamers in clinical trials. Eur J Pharm Sci. Elsevier; 2013;48:259–71. [CrossRef]
- Yang C, Jiang Y, Hao SH, Yan XY, Hong DF, Naranmandura H. Aptamers: an emerging navigation tool of therapeutic agents for targeted cancer therapy. J Mater Chem B [Internet]. Royal Society of Chemistry; 2021 [cited 2025 Aug 19];10:20–33. Available from: https://pubs.rsc.org/en/content/articlehtml/2022/tb/d1tb02098f. [CrossRef]
- Farjami E, Campos R, Nielsen JS, Gothelf K V., Kjems J, Ferapontova EE. RNA aptamer-based electrochemical biosensor for selective and label-free analysis of dopamine. Anal Chem [Internet]. American Chemical Society; 2013 [cited 2025 Aug 19];85:121–8. Available from: https://pubs.acs.org/doi/abs/10.1021/ac302134s. [CrossRef]
- Su Y, Hammond MC. RNA-based fluorescent biosensors for live cell imaging of small molecules and RNAs. Curr Opin Biotechnol. Elsevier Current Trends; 2020;63:157–66. [CrossRef]
- Kumar S, Mohan A, Sharma NR, Kumar A, Girdhar M, Malik T, et al. Computational Frontiers in Aptamer-Based Nanomedicine for Precision Therapeutics: A Comprehensive Review. ACS Omega [Internet]. American Chemical Society; 2024 [cited 2025 Aug 19];9:26838–62. Available from: https://pubs.acs.org/doi/full/10.1021/acsomega.4c02466. [CrossRef]
- Yadav L, Kumar S, Srivastava S, Golmei P. Aptamers: A Novel Class of Targeted Therapeutics. Biosens Aptamers [Internet]. Springer, Singapore; 2025 [cited 2025 Aug 20];49–85. Available from: https://link.springer.com/chapter/10.1007/978-981-96-8387-1_3.
- Liu Y, Wang N, Chan CW, Lu A, Yu Y, Zhang G, et al. The Application of Microfluidic Technologies in Aptamer Selection. Front Cell Dev Biol [Internet]. Frontiers Media S.A.; 2021 [cited 2025 Aug 20];9:730035. Available from: www.frontiersin.org. [CrossRef]
- Domsicova M, Korcekova J, Poturnayova A, Breier A. New Insights into Aptamers: An Alternative to Antibodies in the Detection of Molecular Biomarkers. Int J Mol Sci 2024, Vol 25, Page 6833 [Internet]. Multidisciplinary Digital Publishing Institute; 2024 [cited 2025 Aug 20];25:6833. Available from: https://www.mdpi.com/1422-0067/25/13/6833/htm. [CrossRef]
- El-Husseini DM, Sayour AE, Melzer F, Mohamed MF, Neubauer H, Tammam RH. Generation and Selection of Specific Aptamers Targeting Brucella Species through an Enhanced Cell-SELEX Methodology. Int J Mol Sci [Internet]. MDPI; 2022 [cited 2025 Aug 20];23:6131. Available from: https://www.mdpi.com/1422-0067/23/11/6131/htm. [CrossRef]
- Mohsen MG, Breaker RR. In vitro Selection and in vivo Testing of Riboswitch-inspired Aptamers. Bio-protocol [Internet]. Bio-protocol LLC; 2023 [cited 2025 Aug 20];13:e4775. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC10338711/. [CrossRef]
- Marton S, Reyes-Darias JA, Sánchez-Luque FJ, Romero-López C, Berzal-Herranz A. In Vitro and Ex Vivo Selection Procedures for Identifying Potentially Therapeutic DNA and RNA Molecules. Mol 2010, Vol 15, Pages 4610-4638 [Internet]. Molecular Diversity Preservation International; 2010 [cited 2025 Aug 20];15:4610–38. Available from: https://www.mdpi.com/1420-3049/15/7/4610/htm. [CrossRef]
- Chen Z, Luo H, Gubu A, Yu S, Zhang H, Dai H, et al. Chemically modified aptamers for improving binding affinity to the target proteins via enhanced non-covalent bonding. Front Cell Dev Biol. Frontiers Media S.A.; 2023;11:1091809. [CrossRef]
- Zhao BS, Roundtree IA, He C. Post-transcriptional gene regulation by mRNA modifications. Nat Rev Mol Cell Biol 2016 181 [Internet]. Nature Publishing Group; 2016 [cited 2025 Apr 12];18:31–42. Available from: https://www.nature.com/articles/nrm.2016.132. [CrossRef]
- Motorin Y, Helm M. RNA nucleotide methylation: 2021 update. Wiley Interdiscip Rev RNA [Internet]. John Wiley & Sons, Ltd; 2022 [cited 2025 Apr 9];13:e1691. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/wrna.1691. [CrossRef]
- Moshitch-Moshkovitz S, Dominissini D, Rechavi G. The epitranscriptome toolbox. Cell [Internet]. Elsevier B.V.; 2022 [cited 2025 Apr 12];185:764–76. Available from: https://www.cell.com/action/showFullText?pii=S0092867422001477. [CrossRef]
- Meyer KD, Jaffrey SR. The dynamic epitranscriptome: N6-methyladenosine and gene expression control. Nat Rev Mol Cell Biol 2014 155 [Internet]. Nature Publishing Group; 2014 [cited 2025 Apr 12];15:313–26. Available from: https://www.nature.com/articles/nrm3785. [CrossRef]
- Meyer KD. m6A-mediated translation regulation. Biochim Biophys Acta - Gene Regul Mech. Elsevier; 2019;1862:301–9.
- Deng L, Kumar J, Rose R, McIntyre W, Fabris D. Analyzing RNA posttranscriptional modifications to decipher the epitranscriptomic code. Mass Spectrom Rev [Internet]. John Wiley & Sons, Ltd; 2024 [cited 2025 Aug 26];43:5–38. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/mas.21798. [CrossRef]
- Mateos PA, Zhou Y, Zarnack K, Eyras E. Concepts and methods for transcriptome-wide prediction of chemical messenger RNA modifications with machine learning. Brief Bioinform [Internet]. Oxford Academic; 2023 [cited 2025 Aug 26];24:1–14. Available from:. [CrossRef]
- Jolly P, Estrela P, Ladomery M. Oligonucleotide-based systems: DNA, microRNAs, DNA/RNA aptamers. Essays Biochem [Internet]. Portland Press; 2016 [cited 2025 Aug 16];60:27–35. Available from: /essaysbiochem/article/60/1/27/78209/Oligonucleotide-based-systems-DNA-microRNAs-DNA. [CrossRef]
- Weigand JE, Suess B. Aptamers and riboswitches: Perspectives in biotechnology. Appl Microbiol Biotechnol [Internet]. Springer; 2009 [cited 2025 Aug 16];85:229–36. Available from: https://link.springer.com/article/10.1007/s00253-009-2194-2. [CrossRef]
- Stoltenburg R, Reinemann C, Strehlitz B. SELEX—A (r)evolutionary method to generate high-affinity nucleic acid ligands. Biomol Eng. Elsevier; 2007;24:381–403. [CrossRef]
- Sun H, Zu Y. Aptamers and Their Applications in Nanomedicine. Small [Internet]. John Wiley & Sons, Ltd; 2015 [cited 2025 Aug 16];11:2352–64. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/smll.201403073. [CrossRef]
- Ruscito A, DeRosa MC. Small-molecule binding aptamers: Selection strategies, characterization, and applications. Front Chem [Internet]. Frontiers Media S. A; 2016 [cited 2025 Aug 16];4:188509. Available from: www.frontiersin.org. [CrossRef]
- Zhou J, Rossi J. Aptamers as targeted therapeutics: current potential and challenges. Nat Rev Drug Discov 2016 163 [Internet]. Nature Publishing Group; 2016 [cited 2025 Aug 16];16:181–202. Available from: https://www.nature.com/articles/nrd.2016.199.
- Shraim AS, Abdel Majeed BA, Al-Binni MA, Hunaiti A. Therapeutic Potential of Aptamer-Protein Interactions. ACS Pharmacol Transl Sci [Internet]. American Chemical Society; 2022 [cited 2025 Aug 16];5:1211–27. Available from: https://pubs.acs.org/doi/abs/10.1021/acsptsci.2c00156. [CrossRef]
- Driscoll J, Gondaliya P, Zinn DA, Jain R, Yan IK, Dong H, et al. Using aptamers for targeted delivery of RNA therapies. Mol Ther [Internet]. Elsevier; 2025 [cited 2025 Aug 16];33:1344–67. Available from: https://www.cell.com/action/showFullText?pii=S1525001625001741. [CrossRef]
- Kratschmer C, Levy M. Effect of Chemical Modifications on Aptamer Stability in Serum. Nucleic Acid Ther [Internet]. Nucleic Acid Ther; 2017 [cited 2025 Aug 16];27:335–44. Available from: https://pubmed.ncbi.nlm.nih.gov/28945147/. [CrossRef]
- Nimjee SM, White RR, Becker RC, Sullenger BA. Aptamers as Therapeutics. Annu Rev Pharmacol Toxicol [Internet]. Annual Reviews Inc.; 2017 [cited 2025 Aug 16];57:61–79. Available from: https://www.annualreviews.org/content/journals/10.1146/annurev-pharmtox-010716-104558. [CrossRef]
- Sequeira-Antunes B, Ferreira HA. Nucleic Acid Aptamer-Based Biosensors: A Review. Biomed 2023, Vol 11, Page 3201 [Internet]. Multidisciplinary Digital Publishing Institute; 2023 [cited 2025 Aug 16];11:3201. Available from: https://www.mdpi.com/2227-9059/11/12/3201/htm. [CrossRef]
- Sheraz M, Sun XF, Wang Y, Chen J, Sun L. Recent Developments in Aptamer-Based Sensors for Diagnostics. Sensors 2024, Vol 24, Page 7432 [Internet]. Multidisciplinary Digital Publishing Institute; 2024 [cited 2025 Aug 16];24:7432. Available from: https://www.mdpi.com/1424-8220/24/23/7432/htm. [CrossRef]
- McKeague M, Wong RS, Smolke CD. Opportunities in the design and application of RNA for gene expression control. Nucleic Acids Res [Internet]. Oxford Academic; 2016 [cited 2025 Aug 16];44:2987–99. Available from:. [CrossRef]
- Paige JS, Wu KY, Jaffrey SR. RNA mimics of green fluorescent protein. Science (80- ) [Internet]. American Association for the Advancement of Science; 2011 [cited 2025 Aug 16];333:642–6. Available from: https://www.science.org/doi/10.1126/science.1207339. [CrossRef]
- Jonkhout N, Tran J, Smith MA, Schonrock N, Mattick JS, Novoa EM. The RNA modification landscape in human disease. RNA [Internet]. Cold Spring Harbor Laboratory Press; 2017 [cited 2025 Aug 16];23:1754–69. Available from: http://rnajournal.cshlp.org/content/23/12/1754.full. [CrossRef]
- Zhu Y, Zhu L, Wang X, Jin H. RNA-based therapeutics: an overview and prospectus. Cell Death Dis 2022 137 [Internet]. Nature Publishing Group; 2022 [cited 2025 Aug 16];13:1–15. Available from: https://www.nature.com/articles/s41419-022-05075-2.
- Trivedi J, Yasir M, Maurya RK, Tripathi AS. Aptamer-based Theranostics in Oncology: Design Strategies and Limitations. BIO Integr [Internet]. Compuscript; 2024 [cited 2025 Aug 15];5:993. Available from: https://www.scienceopen.com/hosted-document?doi=10.15212/bioi-2024-0002. [CrossRef]
- Haran V, Lenka N. Deciphering the Epitranscriptomic Signatures in Cell Fate Determination and Development. Stem Cell Rev Reports [Internet]. Humana Press Inc.; 2019 [cited 2025 Apr 12];15:474–96. Available from: https://link.springer.com/article/10.1007/s12015-019-09894-3. [CrossRef]
- Che YH, Lee H, Kim YJ. New insights into the epitranscriptomic control of pluripotent stem cell fate. Exp Mol Med 2022 5410 [Internet]. Nature Publishing Group; 2022 [cited 2025 Apr 12];54:1643–51. Available from: https://www.nature.com/articles/s12276-022-00824-x. [CrossRef]
- Livneh I, Moshitch-Moshkovitz S, Amariglio N, Rechavi G, Dominissini D. The m6A epitranscriptome: Transcriptome plasticity in brain development and function. Nat Rev Neurosci. Nature Research; 2020;21:36-51. [CrossRef]
- Zhang M, Zhai Y, Zhang S, Dai X, Li Z. Roles of N6-Methyladenosine (m6A) in Stem Cell Fate Decisions and Early Embryonic Development in Mammals. Front Cell Dev Biol [Internet]. Frontiers Media S.A.; 2020 [cited 2025 Apr 12];8:566543. Available from: www.frontiersin.org. [CrossRef]
- Yao Y, Liu P, Li Y, Wang W, Jia H, Bai Y, et al. Regulatory role of m6A epitranscriptomic modifications in normal development and congenital malformations during embryogenesis. Biomed Pharmacother [Internet]. Biomed Pharmacother; 2024 [cited 2024 Mar 21];173. Available from: https://pubmed.ncbi.nlm.nih.gov/38394844/. [CrossRef]
- Ahi EP. Regulation of Skeletogenic Pathways by m6A RNA Modification: A Comprehensive Review. Calcif Tissue Int 2025 1161 [Internet]. Springer; 2025 [cited 2025 Apr 16];116:1–23. Available from: https://link.springer.com/article/10.1007/s00223-025-01367-9. [CrossRef]
- Cayir A, Byun HM, Barrow TM. Environmental epitranscriptomics. Environ Res. Academic Press; 2020;189:109885.
- Ahi EP, Singh P. An emerging orchestrator of ecological adaptation: m6A regulation of post-transcriptional mechanisms. Mol Ecol. 2024;17545.
- Ahi EP, Schenekar T. The Promise of Environmental RNA Research Beyond mRNA. Mol Ecol [Internet]. John Wiley & Sons, Ltd; 2025 [cited 2025 Jun 19];34:e17787. Available from: https://onlinelibrary.wiley.com/doi/full/10.1111/mec.17787. [CrossRef]
- Sikorski V, Selberg S, Lalowski M, Karelson M, Kankuri E. The structure and function of YTHDF epitranscriptomic m6A readers. Trends Pharmacol Sci [Internet]. Elsevier Ltd; 2023 [cited 2025 Apr 16];44:335–53. Available from: https://www.cell.com/action/showFullText?pii=S0165614723000627. [CrossRef]
- Rodell R, Robalin N, Martinez NM. Why U matters: detection and functions of pseudouridine modifications in mRNAs. Trends Biochem Sci [Internet]. Elsevier Ltd; 2024 [cited 2024 Dec 23];49:12–27. Available from: http://www.cell.com/article/S0968000423002773/fulltext. [CrossRef]
- Monroe J, Eyler DE, Mitchell L, Deb I, Bojanowski A, Srinivas P, et al. N1-Methylpseudouridine and pseudouridine modifications modulate mRNA decoding during translation. Nat Commun 2024 151 [Internet]. Nature Publishing Group; 2024 [cited 2025 Aug 26];15:1–11. Available from: https://www.nature.com/articles/s41467-024-51301-0. [CrossRef]
- Chen YS, Yang WL, Zhao YL, Yang YG. Dynamic transcriptomic m5C and its regulatory role in RNA processing. Wiley Interdiscip Rev RNA [Internet]. John Wiley & Sons, Ltd; 2021 [cited 2025 Aug 26];12:e1639. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/wrna.1639. [CrossRef]
- Höfler S, Duss O. Interconnections between m6A RNA modification, RNA structure, and protein–RNA complex assembly. Life Sci Alliance [Internet]. Life Science Alliance; 2024 [cited 2025 Aug 26];7. Available from: https://www.life-science-alliance.org/content/7/1/e202302240.
- Fagre C, Gilbert W. Beyond reader proteins: RNA binding proteins and RNA modifications in conversation to regulate gene expression. Wiley Interdiscip Rev RNA [Internet]. John Wiley & Sons, Ltd; 2024 [cited 2025 Aug 26];15:e1834. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/wrna.1834. [CrossRef]
- Tanzer A, Hofacker IL, Lorenz R. RNA modifications in structure prediction – Status quo and future challenges. Methods. Academic Press; 2019;156:32–9. [CrossRef]
- Liu N, Dai Q, Zheng G, He C, Parisien M, Pan T. N6-methyladenosine-dependent RNA structural switches regulate RNA–protein interactions. Nat 2015 5187540 [Internet]. Nature Publishing Group; 2015 [cited 2025 Aug 20];518:560–4. Available from: https://www.nature.com/articles/nature14234. [CrossRef]
- Liu B, Shi H, Rangadurai A, Nussbaumer F, Chu CC, Erharter KA, et al. A quantitative model predicts how m6A reshapes the kinetic landscape of nucleic acid hybridization and conformational transitions. Nat Commun 2021 121 [Internet]. Nature Publishing Group; 2021 [cited 2025 Aug 26];12:1–17. Available from: https://www.nature.com/articles/s41467-021-25253-8. [CrossRef]
- Sağlam B, Akgül B. An Overview of Current Detection Methods for RNA Methylation. Int J Mol Sci 2024, Vol 25, Page 3098 [Internet]. Multidisciplinary Digital Publishing Institute; 2024 [cited 2025 Apr 16];25:3098. Available from: https://www.mdpi.com/1422-0067/25/6/3098/htm. [CrossRef]
- Zhong ZD, Xie YY, Chen HX, Lan YL, Liu XH, Ji JY, et al. Systematic comparison of tools used for m6A mapping from nanopore direct RNA sequencing. Nat Commun 2023 141 [Internet]. Nature Publishing Group; 2023 [cited 2025 Apr 16];14:1–14. Available from: https://www.nature.com/articles/s41467-023-37596-5. [CrossRef]
- Fallah A, Imani Fooladi AA, Havaei SA, Mahboobi M, Sedighian H. Recent advances in aptamer discovery, modification and improving performance. Biochem Biophys Reports. Elsevier; 2024;40:101852. [CrossRef]
- Troisi R, Sica F. Aptamers: Functional-Structural Studies and Biomedical Applications. Int J Mol Sci 2022, Vol 23, Page 4796 [Internet]. Multidisciplinary Digital Publishing Institute; 2022 [cited 2025 Aug 16];23:4796. Available from: https://www.mdpi.com/1422-0067/23/9/4796/htm. [CrossRef]
- Liu WW, Zheng SQ, Li T, Fei YF, Wang C, Zhang S, et al. RNA modifications in cellular metabolism: implications for metabolism-targeted therapy and immunotherapy. Signal Transduct Target Ther 2024 91 [Internet]. Nature Publishing Group; 2024 [cited 2025 Aug 16];9:1–30. Available from: https://www.nature.com/articles/s41392-024-01777-5. [CrossRef]
- Sasso JM, Ambrose BJB, Tenchov R, Datta RS, Basel MT, Delong RK, et al. The Progress and Promise of RNA Medicine-An Arsenal of Targeted Treatments. J Med Chem [Internet]. American Chemical Society; 2022 [cited 2025 Aug 16];65:6975–7015. Available from: https://pubs.acs.org/doi/full/10.1021/acs.jmedchem.2c00024. [CrossRef]
- Kornienko I V., Aramova OY, Tishchenko AA, Rudoy D V., Chikindas ML. RNA Stability: A Review of the Role of Structural Features and Environmental Conditions. Molecules [Internet]. Multidisciplinary Digital Publishing Institute (MDPI); 2024 [cited 2025 Jun 6];29:5978. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC11676819/. [CrossRef]
- Roundtree IA, Evans ME, Pan T, He C. Dynamic RNA Modifications in Gene Expression Regulation. Cell. Cell Press; 2017. p. 1187–200. [CrossRef]
- Stewart JM. RNA nanotechnology on the horizon: Self-assembly, chemical modifications, and functional applications. Curr Opin Chem Biol. Elsevier Current Trends; 2024;81:102479. [CrossRef]
- Hopfinger MC, Kirkpatrick CC, Znosko BM. Predictions and analyses of RNA nearest neighbor parameters for modified nucleotides. Nucleic Acids Res [Internet]. Oxford Academic; 2020 [cited 2025 Aug 27];48:8901–13. Available from:. [CrossRef]
- Roost C, Lynch SR, Batista PJ, Qu K, Chang HY, Kool ET. Structure and thermodynamics of N6-methyladenosine in RNA: A spring-loaded base modification. J Am Chem Soc [Internet]. American Chemical Society; 2015 [cited 2025 Apr 18];137:2107–15. Available from: https://pubs.acs.org/doi/abs/10.1021/ja513080v. [CrossRef]
- Kierzek E, Malgowska M, Lisowiec J, Turner DH, Gdaniec Z, Kierzek R. The contribution of pseudouridine to stabilities and structure of RNAs. Nucleic Acids Res [Internet]. Oxford Academic; 2014 [cited 2025 Apr 16];42:3492–501. Available from:. [CrossRef]
- Spenkuch F, Motorin Y, Helm M. Pseudouridine: Still mysterious, but never a fake (uridine)! RNA Biol [Internet]. Taylor & Francis; 2014 [cited 2025 Aug 27];11:1540–54. Available from: https://www.tandfonline.com/doi/abs/10.4161/15476286.2014.992278. [CrossRef]
- Jones AN, Tikhaia E, Mourão A, Sattler M. Structural effects of m6A modification of the Xist A-repeat AUCG tetraloop and its recognition by YTHDC1. Nucleic Acids Res [Internet]. Oxford Academic; 2022 [cited 2025 Aug 27];50:2350–62. Available from:. [CrossRef]
- Mayer G. The Chemical Biology of Aptamers. Angew Chemie Int Ed [Internet]. John Wiley & Sons, Ltd; 2009 [cited 2025 Aug 16];48:2672–89. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/anie.200804643. [CrossRef]
- Gelinas AD, Davies DR, Janjic N. Embracing proteins: structural themes in aptamer–protein complexes. Curr Opin Struct Biol. Elsevier Current Trends; 2016;36:122–32. [CrossRef]
- Shi Y, Lei Y, Chen M, Ma H, Shen T, Zhang Y, et al. A Demethylation-Switchable Aptamer Design Enables Lag-Free Monitoring of m6A Demethylase FTO with Energy Self-Sufficient and Structurally Integrated Features. J Am Chem Soc [Internet]. American Chemical Society; 2024 [cited 2025 Jul 20];146:34638–50. Available from: https://pubs.acs.org/doi/abs/10.1021/jacs.4c12884. [CrossRef]
- Svensen N, Jaffrey SR. Fluorescent RNA Aptamers as a Tool to Study RNA-Modifying Enzymes. Cell Chem Biol [Internet]. Elsevier Ltd; 2016 [cited 2025 Aug 16];23:415–25. Available from: https://www.cell.com/action/showFullText?pii=S2451945616300046. [CrossRef]
- Odeh F, Nsairat H, Alshaer W, Ismail MA, Esawi E, Qaqish B, et al. Aptamers Chemistry: Chemical Modifications and Conjugation Strategies. Mol 2020, Vol 25, Page 3 [Internet]. Multidisciplinary Digital Publishing Institute; 2019 [cited 2025 Aug 16];25:3. Available from: https://www.mdpi.com/1420-3049/25/1/3/htm. [CrossRef]
- Eremeeva E, Fikatas A, Margamuljana L, Abramov M, Schols D, Groaz E, et al. Highly stable hexitol based XNA aptamers targeting the vascular endothelial growth factor. Nucleic Acids Res [Internet]. Oxford Academic; 2019 [cited 2025 Aug 16];47:4927–39. Available from:. [CrossRef]
- Low JT, Weeks KM. SHAPE-directed RNA secondary structure prediction. Methods. Academic Press; 2010;52:150–8. [CrossRef]
- Wilkinson KA, Merino EJ, Weeks KM. Selective 2′-hydroxyl acylation analyzed by primer extension (SHAPE): quantitative RNA structure analysis at single nucleotide resolution. Nat Protoc 2006 13 [Internet]. Nature Publishing Group; 2006 [cited 2025 Aug 16];1:1610–6. Available from: https://www.nature.com/articles/nprot.2006.249. [CrossRef]
- Oliveira R, Pinho E, Sousa AL, DeStefano JJ, Azevedo NF, Almeida C. Improving aptamer performance with nucleic acid mimics: de novo and post-SELEX approaches. Trends Biotechnol [Internet]. Elsevier Ltd; 2022 [cited 2025 Aug 16];40:549–63. Available from: https://www.cell.com/action/showFullText?pii=S0167779921002274. [CrossRef]
- Zhao L, Qi X, Yan X, Huang Y, Liang X, Zhang L, et al. Engineering Aptamer with Enhanced Affinity by Triple Helix-Based Terminal Fixation. J Am Chem Soc [Internet]. American Chemical Society; 2019 [cited 2025 Aug 16];141:17493–7. Available from: https://pubs.acs.org/doi/abs/10.1021/jacs.9b09292. [CrossRef]
- Soukup GA, Emilsson GAM, Breaker RR. Altering molecular recognition of RNA aptamers by allosteric selection. J Mol Biol. Academic Press; 2000;298:623–32. [CrossRef]
- Hoetzel J, Suess B. Structural Changes in Aptamers are Essential for Synthetic Riboswitch Engineering. J Mol Biol. Academic Press; 2022;434:167631. [CrossRef]
- Kelvin D, Suess B. Tapping the potential of synthetic riboswitches: reviewing the versatility of the tetracycline aptamer. RNA Biol [Internet]. Taylor & Francis; 2023 [cited 2025 Aug 16];20:457–68. Available from: https://www.tandfonline.com/doi/abs/10.1080/15476286.2023.2234732%4010.1080/tfocoll.2024.0.issue-Synthetic_RNA_Biology. [CrossRef]
- Hashmi MATS, Fatima H, Ahmad S, Rehman A, Safdar F. The interplay between epitranscriptomic RNA modifications and neurodegenerative disorders: Mechanistic insights and potential therapeutic strategies. Ibrain [Internet]. John Wiley & Sons, Ltd; 2024 [cited 2025 Aug 16];10:395–426. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/ibra.12183. [CrossRef]
- Arguello AE, Leach RW, Kleiner RE. In Vitro Selection with a Site-Specifically Modified RNA Library Reveals the Binding Preferences of N6-Methyladenosine Reader Proteins. Biochemistry [Internet]. American Chemical Society; 2019 [cited 2025 Aug 26];58:3386–95. Available from: https://pubs.acs.org/doi/abs/10.1021/acs.biochem.9b00485. [CrossRef]
- Brown A, Brill J, Amini R, Nurmi C, Li Y. Development of Better Aptamers: Structured Library Approaches, Selection Methods, and Chemical Modifications. Angew Chemie Int Ed [Internet]. John Wiley & Sons, Ltd; 2024 [cited 2025 Aug 26];63:e202318665. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/anie.202318665. [CrossRef]
- Felix AS, Quillin AL, Mousavi S, Heemstra JM. Harnessing Nature’s Molecular Recognition Capabilities to Map and Study RNA Modifications. Acc Chem Res [Internet]. American Chemical Society; 2022 [cited 2025 Aug 27]; Available from: https://pubs.acs.org/doi/abs/10.1021/acs.accounts.2c00287. [CrossRef]
- Guo W, Ma Y, Mou Q, Shao X, Lyu M, Garcia V, et al. Sialic acid aptamer and RNA in situ hybridization-mediated proximity ligation assay for spatial imaging of glycoRNAs in single cells. Nat Protoc 2025 207 [Internet]. Nature Publishing Group; 2025 [cited 2025 Aug 27];20:1930–50. Available from: https://www.nature.com/articles/s41596-024-01103-x. [CrossRef]
- Zhang Q, Zhao S, Su C, Han Q, Han Y, Tian X, et al. Construction of a Quantum-Dot-Based FRET Nanosensor through Direct Encoding of Streptavidin-Binding RNA Aptamers for N6-Methyladenosine Demethylase Detection. Anal Chem [Internet]. American Chemical Society; 2023 [cited 2025 Jan 3];95:13201–10. Available from: https://pubs.acs.org/doi/abs/10.1021/acs.analchem.3c02149. [CrossRef]
- Vögele J, Duchardt-Ferner E, Kruse H, Zhang Z, Sponer J, Krepl M, et al. Structural and dynamic effects of pseudouridine modifications on noncanonical interactions in RNA. RNA [Internet]. Cold Spring Harbor Laboratory Press; 2023 [cited 2025 Aug 27];29:790–807. Available from: http://rnajournal.cshlp.org/content/29/6/790.full. [CrossRef]
- Gu L, Zheng J, Zhang Y, Wang D, Liu J. Selection and Characterization of DNA Aptamers for Cytidine and Uridine. ChemBioChem [Internet]. John Wiley & Sons, Ltd; 2024 [cited 2025 Aug 27];25:e202300656. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/cbic.202300656. [CrossRef]
- Yoo H, Jo H, Oh SS. Detection and beyond: challenges and advances in aptamer-based biosensors. Mater Adv [Internet]. Royal Society of Chemistry; 2020 [cited 2025 Jun 6];1:2663–87. Available from: https://pubs.rsc.org/en/content/articlehtml/2020/ma/d0ma00639d. [CrossRef]
- Ying X, Huang C, Li T, Li T, Gao M, Wang F, et al. An RNA Methylation-Sensitive AIEgen-Aptamer Reporting System for Quantitatively Evaluating m6A Methylase and Demethylase Activities. ACS Chem Biol [Internet]. American Chemical Society; 2024 [cited 2025 Jul 20];19:162–72. Available from: https://pubs.acs.org/doi/abs/10.1021/acschembio.3c00613. [CrossRef]
- Catuogno S, Esposito CL, Ungaro P, De Franciscis V. Nucleic Acid Aptamers Targeting Epigenetic Regulators: An Innovative Therapeutic Option. Pharm 2018, Vol 11, Page 79 [Internet]. Multidisciplinary Digital Publishing Institute; 2018 [cited 2025 Aug 19];11:79. Available from: https://www.mdpi.com/1424-8247/11/3/79/htm. [CrossRef]
- Yin Y, Morgunova E, Jolma A, Kaasinen E, Sahu B, Khund-Sayeed S, et al. Impact of cytosine methylation on DNA binding specificities of human transcription factors. Science (80- ) [Internet]. American Association for the Advancement of Science; 2017 [cited 2025 Aug 19];356. Available from: https://www.science.org/doi/10.1126/science.aaj2239. [CrossRef]
- Ahi EP, Khorshid M. Potentials of RNA biosensors in developmental biology. Dev Biol [Internet]. Academic Press; 2025 [cited 2025 Jul 30];526:173–88. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0012160625002027. [CrossRef]
- Murray MT, Wetmore SD. Unlocking precision in aptamer engineering: a case study of the thrombin binding aptamer illustrates why modification size, quantity, and position matter. Nucleic Acids Res [Internet]. Oxford Academic; 2024 [cited 2025 Aug 19];52:10823–35. Available from:. [CrossRef]
- Valsangkar V, Vangaveti S, Lee GW, Fahssi WM, Awan WS, Huang Y, et al. Structural and Binding Effects of Chemical Modifications on Thrombin Binding Aptamer (TBA). Mol 2021, Vol 26, Page 4620 [Internet]. Multidisciplinary Digital Publishing Institute; 2021 [cited 2025 Aug 19];26:4620. Available from: https://www.mdpi.com/1420-3049/26/15/4620/htm. [CrossRef]
- Seelam Prabhakar P, A Manderville R, D Wetmore S. Impact of the Position of the Chemically Modified 5-Furyl-2′-Deoxyuridine Nucleoside on the Thrombin DNA Aptamer–Protein Complex: Structural Insights into Aptamer Response from MD Simulations. Mol 2019, Vol 24, Page 2908 [Internet]. Multidisciplinary Digital Publishing Institute; 2019 [cited 2025 Aug 19];24:2908. Available from: https://www.mdpi.com/1420-3049/24/16/2908/htm.
- Bowles IE, Orellana EA. Rethinking RNA Modifications: Therapeutic Strategies for Targeting Dysregulated RNA. J Mol Biol. Academic Press; 2025;437:169046. [CrossRef]
- Qi S, Duan N, Khan IM, Dong X, Zhang Y, Wu S, et al. Strategies to manipulate the performance of aptamers in SELEX, post-SELEX and microenvironment. Biotechnol Adv. Elsevier; 2022;55:107902. [CrossRef]
- Takahashi K, Galloway KE. RNA-based controllers for engineering gene and cell therapies. Curr Opin Biotechnol. Elsevier Current Trends; 2024;85:103026. [CrossRef]
- Hou Q, Jaffrey SR. Synthetic biology tools to promote the folding and function of RNA aptamers in mammalian cells. RNA Biol [Internet]. Taylor & Francis; 2023 [cited 2025 Aug 26];20:198–206. Available from: https://www.tandfonline.com/doi/abs/10.1080/15476286.2023.2206248%4010.1080/tfocoll.2024.0.issue-Synthetic_RNA_Biology. [CrossRef]
- Gilbert W V., Nachtergaele S. mRNA Regulation by RNA Modifications. Annu Rev Biochem [Internet]. Annu Rev Biochem; 2023 [cited 2025 Aug 26];92:175–98. Available from: https://pubmed.ncbi.nlm.nih.gov/37018844/.
- Vazquez-Anderson J, Contreras LM. Regulatory RNAs: charming gene management styles for synthetic biology applications. RNA Biol [Internet]. RNA Biol; 2013 [cited 2025 Aug 19];10:1778–97. Available from: https://pubmed.ncbi.nlm.nih.gov/24356572/.
- Shanidze N, Lenkeit F, Hartig JS, Funck D. A Theophylline-Responsive Riboswitch Regulates Expression of Nuclear-Encoded Genes. Plant Physiol [Internet]. Oxford Academic; 2020 [cited 2025 Aug 19];182:123–35. Available from:. [CrossRef]
- Khalil AS, Collins JJ. Synthetic biology: applications come of age. Nat Rev Genet 2010 115 [Internet]. Nature Publishing Group; 2010 [cited 2025 Aug 20];11:367–79. Available from: https://www.nature.com/articles/nrg2775. [CrossRef]
- Maung NW, Smolke CD. Higher-order cellular information processing with synthetic RNA devices. Science (80- ) [Internet]. American Association for the Advancement of Science; 2008 [cited 2025 Aug 20];322:456–60. Available from: https://www.science.org/doi/10.1126/science.1160311.
- Kundert K, Lucas JE, Watters KE, Fellmann C, Ng AH, Heineike BM, et al. Controlling CRISPR-Cas9 with ligand-activated and ligand-deactivated sgRNAs. Nat Commun 2019 101 [Internet]. Nature Publishing Group; 2019 [cited 2025 Aug 20];10:1–11. Available from: https://www.nature.com/articles/s41467-019-09985-2. [CrossRef]
- Winkler W, Nahvi A, Breaker RR. Thiamine derivatives bind messenger RNAs directly to regulate bacterial gene expression. Nat 2002 4196910 [Internet]. Nature Publishing Group; 2002 [cited 2025 Aug 20];419:952–6. Available from: https://www.nature.com/articles/nature01145. [CrossRef]
- Spitale RC, Flynn RA, Zhang QC, Crisalli P, Lee B, Jung JW, et al. Structural imprints in vivo decode RNA regulatory mechanisms. Nat 2015 5197544 [Internet]. Nature Publishing Group; 2015 [cited 2025 Aug 26];519:486–90. Available from: https://www.nature.com/articles/nature14263. [CrossRef]
- Feagin TA, Maganzini N, Soh HT. Strategies for Creating Structure-Switching Aptamers. ACS Sensors [Internet]. American Chemical Society; 2018 [cited 2025 Aug 26];3:1611–5. Available from: https://pubs.acs.org/doi/full/10.1021/acssensors.8b00516. [CrossRef]
- Akhter S. Mechanism of ligand binding to target RNA aptamer. Biophys J [Internet]. Elsevier BV; 2024 [cited 2025 Aug 20];123:544a-545a. Available from: https://www.cell.com/action/showFullText?pii=S0006349523040006. [CrossRef]
- Lyu M, Chan CH, Chen Z, Liu Y, Yu Y. Advantages, applications, and future directions of in vivo aptamer SELEX: A review. Mol Ther Nucleic Acids [Internet]. Cell Press; 2025 [cited 2025 Aug 20];36. Available from: https://www.cell.com/action/showFullText?pii=S2162253125001295. [CrossRef]
- Bayani A, Abosaoda MK, Rizaev J, Nazari M, Jafari Z, Soleimani Samarkhazan H. Aptamer-based approaches in leukemia: a paradigm shift in targeted therapy. Clin Exp Med 2025 251 [Internet]. Springer; 2025 [cited 2025 Aug 20];25:1–14. Available from: https://link.springer.com/article/10.1007/s10238-025-01724-w. [CrossRef]
- Ng EWM, Shima DT, Calias P, Cunningham ET, Guyer DR, Adamis AP. Pegaptanib, a targeted anti-VEGF aptamer for ocular vascular disease. Nat Rev Drug Discov 2006 52 [Internet]. Nature Publishing Group; 2006 [cited 2025 Aug 20];5:123–32. Available from: https://www.nature.com/articles/nrd1955. [CrossRef]
- Ni S, Zhuo Z, Pan Y, Yu Y, Li F, Liu J, et al. Recent Progress in Aptamer Discoveries and Modifications for Therapeutic Applications. ACS Appl Mater Interfaces [Internet]. American Chemical Society; 2021 [cited 2025 Aug 20];13:9500–19. Available from: https://pubs.acs.org/doi/full/10.1021/acsami.0c05750. [CrossRef]
- Doessing H, Vester B. Locked and Unlocked Nucleosides in Functional Nucleic Acids. Mol 2011, Vol 16, Pages 4511-4526 [Internet]. Molecular Diversity Preservation International; 2011 [cited 2025 Aug 20];16:4511–26. Available from: https://www.mdpi.com/1420-3049/16/6/4511/htm. [CrossRef]
- Bunka DHJ, Platonova O, Stockley PG. Development of aptamer therapeutics. Curr Opin Pharmacol. Elsevier; 2010;10:557–62.
- Li Q, Liu J, Guo L, Zhang Y, Chen Y, Liu H, et al. Decoding the interplay between m6A modification and stress granule stability by live-cell imaging. Sci Adv [Internet]. American Association for the Advancement of Science; 2024 [cited 2025 Aug 26];10:5689. Available from: https://www.science.org/doi/10.1126/sciadv.adp5689. [CrossRef]
- Marayati BF, Thompson MG, Holley CL, Horner SM, Meyer KD. Programmable protein expression using a genetically encoded m6A sensor. Nat Biotechnol 2024 429 [Internet]. Nature Publishing Group; 2024 [cited 2025 Aug 26];42:1417–28. Available from: https://www.nature.com/articles/s41587-023-01978-3. [CrossRef]
- Tegowski M, Meyer KD. Studying m6A in the brain: a perspective on current methods, challenges, and future directions. Front Mol Neurosci. Frontiers Media SA; 2024;17:1393973. [CrossRef]
- Tang W, Hu JH, Liu DR. Aptazyme-embedded guide RNAs enable ligand-responsive genome editing and transcriptional activation. Nat Commun 2017 81 [Internet]. Nature Publishing Group; 2017 [cited 2025 Aug 26];8:1–8. Available from: https://www.nature.com/articles/ncomms15939. [CrossRef]
- Horner SM, Thompson MG. Challenges to mapping and defining m6A function in viral RNA. RNA [Internet]. Cold Spring Harbor Laboratory Press; 2024 [cited 2025 Aug 26];30:482–90. Available from: http://rnajournal.cshlp.org/content/30/5/482.full. [CrossRef]
- Shachar R, Dierks D, Garcia-Campos MA, Uzonyi A, Toth U, Rossmanith W, et al. Dissecting the sequence and structural determinants guiding m6A deposition and evolution via inter- and intra-species hybrids. Genome Biol [Internet]. BioMed Central Ltd; 2024 [cited 2025 Aug 26];25:1–29. Available from: https://link.springer.com/articles/10.1186/s13059-024-03182-1. [CrossRef]
- Yang Y, Lu Y, Wang Y, Wen X, Qi C, Piao W, et al. Current progress in strategies to profile transcriptomic m6A modifications. Front Cell Dev Biol. Frontiers Media SA; 2024;12:1392159. [CrossRef]
- Varenyk Y, Spicher T, Hofacker IL, Lorenz R. Modified RNAs and predictions with the ViennaRNA Package. Bioinformatics [Internet]. Bioinformatics; 2023 [cited 2025 Aug 26];39. Available from: https://pubmed.ncbi.nlm.nih.gov/37971965/. [CrossRef]
- Esfahani NG, Stein AJ, Akeson S, Tzadikario T, Jain M. Evaluation of Nanopore direct RNA sequencing updates for modification detection. bioRxiv [Internet]. Cold Spring Harbor Laboratory; 2025 [cited 2025 Aug 26];2025.05.01.651717. Available from: https://www.biorxiv.org/content/10.1101/2025.05.01.651717v1.
- Wu Y, Shao W, Yan M, Wang Y, Xu P, Huang G, et al. Transfer learning enables identification of multiple types of RNA modifications using nanopore direct RNA sequencing. Nat Commun 2024 151 [Internet]. Nature Publishing Group; 2024 [cited 2025 Aug 26];15:1–19. Available from: https://www.nature.com/articles/s41467-024-48437-4. [CrossRef]
- Sheehan CJ, Marayati BF, Bhatia J, Meyer KD. In situ visualization of m6A sites in cellular mRNAs. Nucleic Acids Res [Internet]. Oxford Academic; 2023 [cited 2025 Aug 26];51:e101–e101. Available from:. [CrossRef]
- He B, Chen Y, Yi C. Quantitative mapping of the mammalian epitranscriptome. Curr Opin Genet Dev. Elsevier Current Trends; 2024;87:102212. [CrossRef]
- Motorin Y, Helm M. General Principles and Limitations for Detection of RNA Modifications by Sequencing. Acc Chem Res [Internet]. American Chemical Society; 2024 [cited 2025 Aug 26];57:275–88. Available from: https://pubs.acs.org/doi/full/10.1021/acs.accounts.3c00529. [CrossRef]
- Hendra C, Pratanwanich PN, Wan YK, Goh WSS, Thiery A, Göke J. Detection of m6A from direct RNA sequencing using a multiple instance learning framework. Nat Methods [Internet]. Nat Methods; 2022 [cited 2025 Jul 23];19:1590–8. Available from: https://pubmed.ncbi.nlm.nih.gov/36357692/. [CrossRef]
- Sczepanski JT, Joyce GF. Binding of a structured d-RNA molecule by an l-RNA aptamer. J Am Chem Soc [Internet]. American Chemical Society; 2013 [cited 2025 Aug 20];135:13290–3. Available from: https://pubs.acs.org/doi/abs/10.1021/ja406634g. [CrossRef]
- Dey S, Sczepanski JT. In vitro selection of l-DNA aptamers that bind a structured d-RNA molecule. Nucleic Acids Res [Internet]. Oxford Academic; 2020 [cited 2025 Aug 20];48:1669–80. Available from:. [CrossRef]
- Cawte AD, Unrau PJ, Rueda DS. Live cell imaging of single RNA molecules with fluorogenic Mango II arrays. Nat Commun 2020 111 [Internet]. Nature Publishing Group; 2020 [cited 2024 Dec 27];11:1–11. Available from: https://www.nature.com/articles/s41467-020-14932-7. [CrossRef]
- Chen Z, Chen W, Reheman Z, Jiang H, Wu J, Li X. Genetically encoded RNA-based sensors with Pepper fluorogenic aptamer. Nucleic Acids Res [Internet]. Oxford Academic; 2023 [cited 2025 Aug 20];51:8322–36. Available from:. [CrossRef]
- Wang Q, Xiao F, Su H, Liu H, Xu J, Tang H, et al. Inert Pepper aptamer-mediated endogenous mRNA recognition and imaging in living cells. Nucleic Acids Res [Internet]. Oxford Academic; 2022 [cited 2025 Aug 20];50:e84–e84. Available from:. [CrossRef]
- Lin L, Fu Q, Williams BAR, Azzaz AM, Shogren-Knaak MA, Chaput JC, et al. Recognition imaging of acetylated chromatin using a DNA aptamer. Biophys J [Internet]. Biophysical Society; 2009 [cited 2025 Aug 20];97:1804–7. Available from: https://www.cell.com/action/showFullText?pii=S0006349509012296. [CrossRef]
- Gao W, Xu L, Jing J, Zhang X. A far red emissive RNA aptamer–fluorophore system for demethylase FTO detection: design and optimization. New J Chem [Internet]. The Royal Society of Chemistry; 2023 [cited 2025 Aug 20];47:5238–43. Available from: https://pubs.rsc.org/en/content/articlehtml/2023/nj/d3nj00043e. [CrossRef]
- Jenison RD, Gill SC, Pardi A, Polisky B. High-Resolution Molecular Discrimination by RNA. Science (80- ) [Internet]. American Association for the Advancement of Science; 1994 [cited 2025 Aug 20];263:1425–9. Available from: https://www.science.org/doi/10.1126/science.7510417. [CrossRef]
- Pham H, Kumar M, Martinez AR, Ali M, Lowery RG. Development and validation of a generic methyltransferase enzymatic assay based on an SAH riboswitch. SLAS Discov. Elsevier; 2024;29:100161. [CrossRef]
- Berens C, Groher F, Suess B. RNA aptamers as genetic control devices: The potential of riboswitches as synthetic elements for regulating gene expression. Biotechnol J [Internet]. John Wiley & Sons, Ltd; 2015 [cited 2025 Aug 26];10:246–57. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/biot.201300498. [CrossRef]
- Khosravi HM, Jantsch MF. Site-directed RNA editing: recent advances and open challenges. RNA Biol [Internet]. Taylor & Francis; 2021 [cited 2025 Aug 26];18:41–50. Available from: https://www.tandfonline.com/doi/abs/10.1080/15476286.2021.1983288. [CrossRef]
- Ai X, Ding S, Zhou S, Du F, Liu S, Cui X, et al. Enhancing RNA editing efficiency and specificity with engineered ADAR2 guide RNAs. Mol Ther Nucleic Acids [Internet]. Cell Press; 2025 [cited 2025 Aug 26];36:102447. Available from: https://www.cell.com/action/showFullText?pii=S2162253125000010. [CrossRef]
- Flamand MN, Ke K, Tamming R, Meyer KD. Single-molecule identification of the target RNAs of different RNA binding proteins simultaneously in cells. Genes Dev [Internet]. Cold Spring Harbor Laboratory Press; 2022 [cited 2025 Aug 26];36:1002–15. Available from: http://genesdev.cshlp.org/content/36/17-18/1002.full. [CrossRef]
- Okuda M, Fourmy D, Yoshizawa S. Use of Baby Spinach and Broccoli for imaging of structured cellular RNAs. Nucleic Acids Res [Internet]. Oxford Academic; 2017 [cited 2025 Aug 20];45:1404–15. Available from:. [CrossRef]
- Al Shanaa O, Rumyantsev A, Sambuk E, Padkina M. In Vivo Production of RNA Aptamers and Nanoparticles: Problems and Prospects. Mol 2021, Vol 26, Page 1422 [Internet]. Multidisciplinary Digital Publishing Institute; 2021 [cited 2025 Aug 20];26:1422. Available from: https://www.mdpi.com/1420-3049/26/5/1422/htm. [CrossRef]
- Zhang HL, Lv C, Li ZH, Jiang S, Cai D, Liu SS, et al. Analysis of aptamer-target binding and molecular mechanisms by thermofluorimetric analysis and molecular dynamics simulation. Front Chem [Internet]. Frontiers Media S.A.; 2023 [cited 2025 Aug 20];11:1144347. Available from: https://zdock.umassmed.edu/. [CrossRef]
- Yang X, Li N, Gorenstein DG. Strategies for the discovery of therapeutic aptamers. Expert Opin Drug Discov [Internet]. Taylor & Francis; 2011 [cited 2025 Aug 20];6:75–87. Available from: https://www.tandfonline.com/doi/abs/10.1517/17460441.2011.537321. [CrossRef]
- Vorobyeva MA, Davydova AS, Vorobjev PE, Pyshnyi D V., Venyaminova AG. Key Aspects of Nucleic Acid Library Design for in Vitro Selection. Int J Mol Sci 2018, Vol 19, Page 470 [Internet]. Multidisciplinary Digital Publishing Institute; 2018 [cited 2025 Aug 20];19:470. Available from: https://www.mdpi.com/1422-0067/19/2/470/htm. [CrossRef]
- Liu J, Ren Z, Sun Y, Xu L, Wei D, Tan W, et al. Investigation of the Relationship between Aptamers’ Targeting Functions and Human Plasma Proteins. ACS Nano [Internet]. American Chemical Society; 2023 [cited 2025 Aug 20];17:24329–42. Available from: https://pubs.acs.org/doi/abs/10.1021/acsnano.3c10238. [CrossRef]
- Kelly L, Maier KE, Yan A, Levy M. A comparative analysis of cell surface targeting aptamers. Nat Commun 2021 121 [Internet]. Nature Publishing Group; 2021 [cited 2025 Aug 20];12:1–14. Available from: https://www.nature.com/articles/s41467-021-26463-w. [CrossRef]
- Grozhik A V., Jaffrey SR. Distinguishing RNA modifications from noise in epitranscriptome maps. Nat Chem Biol 2018 143 [Internet]. Nature Publishing Group; 2018 [cited 2025 Aug 26];14:215–25. Available from: https://www.nature.com/articles/nchembio.2546.
- Sarkar A, Gasperi W, Begley U, Nevins S, Huber SM, Dedon PC, et al. Detecting the epitranscriptome. Wiley Interdiscip Rev RNA [Internet]. John Wiley & Sons, Ltd; 2021 [cited 2025 Aug 26];12:e1663. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/wrna.1663. [CrossRef]
- Kovacevic KD, Gilbert JC, Jilma B. Pharmacokinetics, pharmacodynamics and safety of aptamers. Adv Drug Deliv Rev. Elsevier; 2018;134:36–50. [CrossRef]
- Hostiuc M, Scafa A, Iancu B, Iancu D, Isailă OM, Ion OM, et al. Ethical implications of developing RNA-based therapies for cardiovascular disorders. Front Bioeng Biotechnol. Frontiers Media SA; 2024;12:1370403. [CrossRef]
- Mubarak G, Zahir FR. Recent Major Transcriptomics and Epitranscriptomics Contributions toward Personalized and Precision Medicine. J Pers Med 2022, Vol 12, Page 199 [Internet]. Multidisciplinary Digital Publishing Institute; 2022 [cited 2025 Aug 26];12:199. Available from: https://www.mdpi.com/2075-4426/12/2/199/htm. [CrossRef]
Figure 1.
Potential examples of RNA modification mediated improvement in RNA aptamers synthesis processes.
Figure 1.
Potential examples of RNA modification mediated improvement in RNA aptamers synthesis processes.
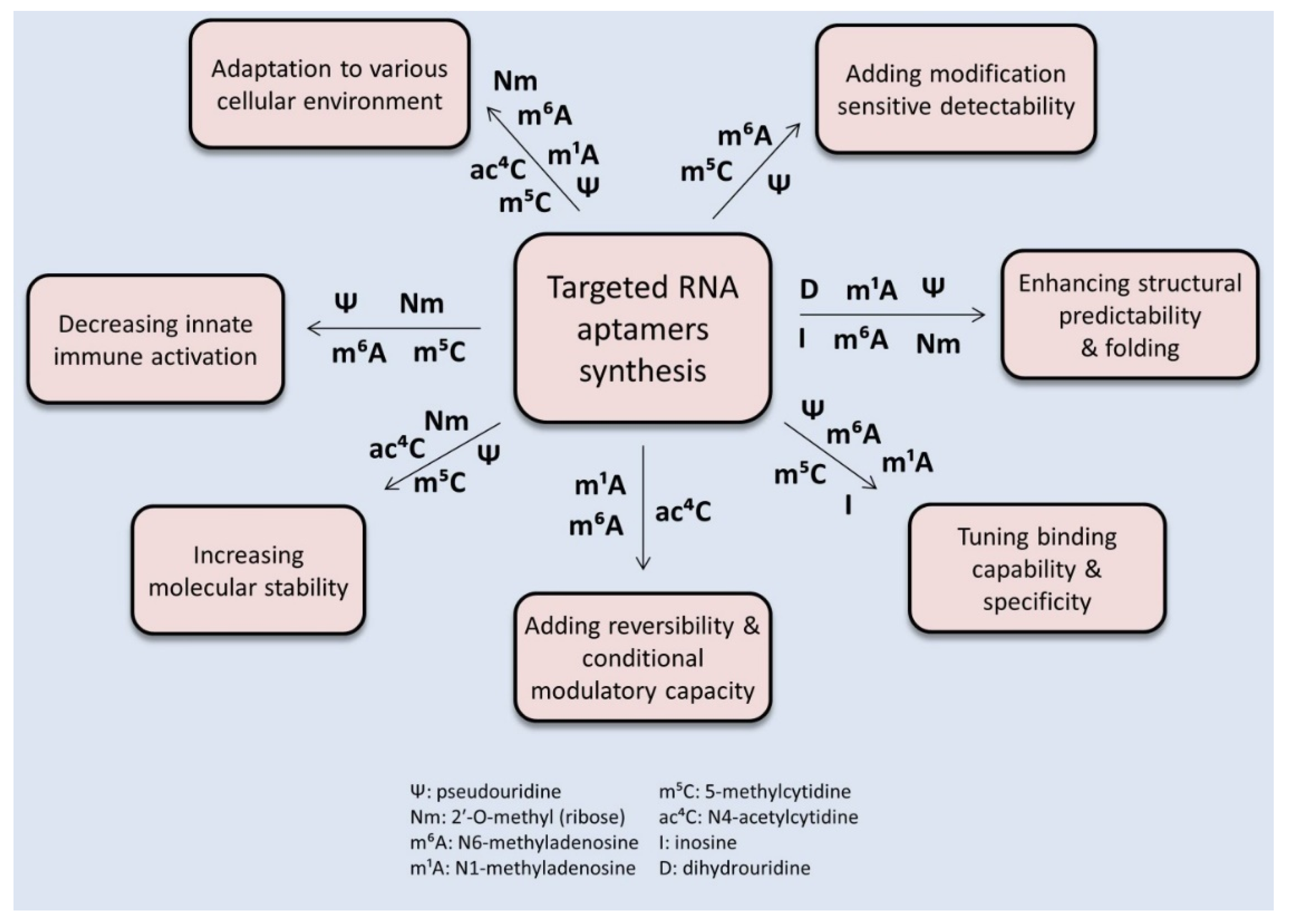
Figure 2.
Potential examples of aptamer-based improvement in epitranscriptomics research.
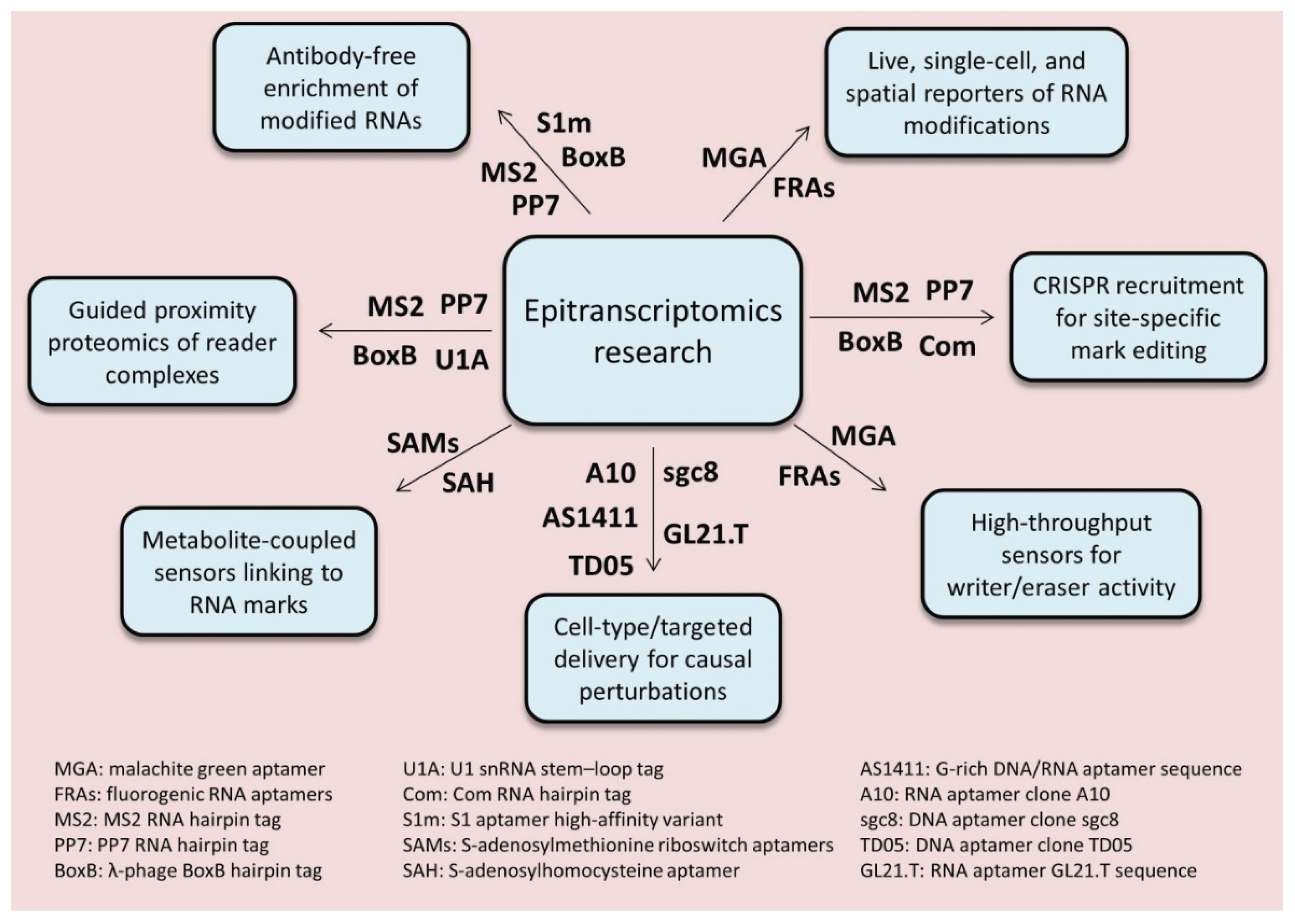
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



