
Submitted:
07 October 2025
Posted:
09 October 2025
You are already at the latest version
Abstract
Translation of mRNAs is a tightly regulated process in gene expression. In mRNA, the 5′ untranslated region (5′ UTR) controls ribosome recruitment and frequently contains structured elements that modulate translation efficacy. This study investigates stable structural motifs within 5' UTR of DDX23 mRNA, encoding a protein relevant for anticancer therapy, as potential regulators and targets for antisense oligonucleotides (ASOs). Despite bioinformatic predictions and transcriptomic validations suggesting RNA G-quadruplex (rG4) formation, comprehensive structural analysis using light-up assay and CD, UV, and NMR spectroscopy revealed that most putative rG4-forming sequences do not fold into stable rG4 structures, although one of them exists in an equilibrium between rG4 and alternative, likely hairpin, conformation. Reporter assays using a robust G4 stabilizer also argue against a significant regulatory role of rG4s in DDX23 mRNA translation. Instead, we identified and characterized a stable hairpin structure with potential regulatory function. Based on these findings, we designed fully locked nucleic acid (LNA)-modified ASOs to target this hairpin and regions flanking the upstream open reading frame (uORF) and start codon of the coding sequence. Reporter assay demonstrated that cap-proximal targeting achieved robust translation inhibition up to 80%. In contrast, targeting the efficiently translated uORF was ineffective, presumably due to steric hindrances from the ribosomal complex. The study yields crucial design principles for translation-regulating ASOs: avoid targeting regions shielded by efficient uORF translation and carefully tune ASO-RNA duplex stability to surpass endogenous structures without disrupting regulatory mechanisms. These findings provide insights into the regulation of DDX23 expression and establish a framework for developing ASO-based therapeutics with broad implications for mRNA targeting in anticancer applications.
Keywords:
DDX23
; hairpin
; LNA
; antisense oligonucleotides
; translation regulation
; uORF
1. Introduction
DDX23, a DEAD-box helicase family member, exhibits versatile regulatory functions in genome stability maintenance, RNA processing, and translational control [1,2]. The protein is an integral component of the U5 small nuclear ribonucleoprotein complex, facilitating spliceosomal conformational alterations through RNA duplex unwinding [2,3]. DDX23 maintains genomic stability via R-loop resolution [4] and participates in antiviral immunity through viral dsRNA detection and subsequent IRF3/NF-kB pathway activation [5,6,7].
DDX23 dysregulation is implicated in both cancer development and neurological disorders. Notable upregulation occurs in ovarian, pancreatic, and glial neoplasms, correlating with aggressive phenotypes and unfavorable clinical outcomes. In ovarian cancer, E2F1-driven DDX23 overexpression modulates FOXM1 splicing, promoting oncogenic FOXM1C isoform generation [8]. DDX23 activates Notch pathway in non-small cell lung cancer enhancing linc00630 expression [9]. In pancreatic ductal adenocarcinoma, METTL3-mediated m6A methylation stabilizes DDX23 mRNA, promoting gemcitabine resistance through PI3K/Akt pathway activation [10]. Glioma progression involves DDX23-mediated miR-21 biogenesis regulation [11]. Conversely, DDX23 deficiency or mutations compromises antiviral responses and neurological function, manifesting as developmental delay, autism spectrum disorder, seizures, and dysmorphism [7,12,13]. Current therapeutic approaches targeting DDX23 lack selectivity and include an antiparasitic drug and pan-helicase inhibitor ivermectin as well as estrogen receptor (ER) antagonists endoxifen and fulvestrant [11,14].
An approach to modulate gene expression with high specificity and effectiveness explores antisense oligonucleotides (ASOs) that are designed to bind to specific RNA sequences. A number of ASOs have been approved for commercial use [15], and they are increasingly recognized as powerful tools for uncovering the biological functions of genes. The main antisense mechanisms include RNase H1-dependent mRNA degradation and pre-mRNA splicing regulation for down- and up-regulation of gene expression, respectively [16]. However, ASOs can also utilize other mechanisms such as translation inhibition due to steric hindrance or no-go decay degradation [17,18,19]. To increase gene expression, ASOs are designed to disrupt mRNA 5’ UTR secondary structures or mask upstream open reading frames (uORF) that negatively affect translation of the coding sequence (CDS) [20,21].
Chemical modifications of ASOs have significantly enhanced their therapeutic potential through the improvement of target hybridization, stability, and cellular uptake. Striking examples include phosphorothioate (PS) backbones, 2′-O-methoxyethyl (2′-MOE) nucleotides, and locked nucleic acids (LNAs) that have proven effective in the treatment of genetic diseases [15]. LNAs represent ribonucleotide analogs featuring a methylene bridge between the 2’-O and 4’-C atoms of the ribose ring [22]. This structural constraint favors a 3’-endo conformation of the ribose moiety that results in optimization of hybridization characteristics and a context-dependent increase in melting temperature (Tm) up to 10 °C per modified nucleotide [23]. Additionally, LNA incorporation at ASO termini can enhance nuclease resistance, thus improving their therapeutic efficacy.
ASOs targeting 5’ UTR can interact with regulatory structural elements, such as hairpins and G-quadruplexes (rG4s) [21,24]. rG4 structures can fold within guanine (G)-rich sequences and consist of stacked G-quartets stabilized by a network of Hoogsteen hydrogen bonds and monovalent cations such as K+ [25]. In most cases, hairpins and rG4s function as translation inhibitors by acting as steric hindrances to the scanning pre-initiation complex (PIC) [25,26,27]. Previous transcriptomic analyses and bioinformatic predictions have indicated that DDX23 mRNA 5’ UTR contains putative rG4s [28,29]. Furthermore, RNA fold analysis suggests the presence of hairpin structures within the 5’ UTR [30,31]. Here, we employ spectroscopic analysis and LNA-based functional probing to characterize these structural elements and evaluate their regulatory capacity and targetability using dual luciferase assay.
2. Results and Discussion
2.1. Analysis of Potential Regulatory G-quadruplex Elements in 5’ UTR of DDX23 mRNA
The rG4 structure was previously identified in the ENST00000551468 transcript of DDX23 during investigation of its translation regulation by predominately rG4-resolving DEAH-box helicase DHX36 [28]. Although this finding is important, the later annotation (GENCODE release 48) implies that this specific isoform has a truncated 3’ end without the full coding sequence and 3’ UTR. Moreover, the sequence of the studied rG4 does not map to any other transcripts. Therefore, this rG4 has no influence on the isoforms coding full-length DDX23 proteoforms. Thus, we searched for rG4s in the MANE Select [32] isoform of DDX23 (ENST00000308025), which is more functionally relevant.
Computational analysis using QGRS software identified putative rG4-forming motifs within the DDX23 mRNA 5’ UTR exclusively (14-36 nt and 51-77 nt) and including proximal region of the coding sequence (14-36 nt and 76-96 nt) [33]. Transcriptomic techniques DMS-seq and G4RP-seq corroborated the formation of rG4 structures within 14-36 nt and 76-96 nt sequences (Figure S1) [29,34,35]. To study the putative rG4-folding in vitro, we synthesized RNA sequences that contain these rG4-forming motifs and 3-nt flanks, as well as control mutants with guanine-to-uridine substitutions (Figure 1A, Table S1): G1 (11-39 nt), G2 (48-80 nt) and G3 (73-99 nt).
We began the structural characterization of the sequences with thioflavin T (ThT)-based light-up assay [36] and used rTBA15 and BON1-2 with validated folding patterns as control rG4 and RNA duplex, respectively (Figure 2A) [37]. Only G3 sequence exhibited an enhancement in fluorescence intensity comparable to control rTBA15 that suggests rG4 formation.
Since the results of ThT light-up assay usually require additional validation due to possible false-positive outcomes [38], circular dichroism (CD) spectroscopy was used for additional structural analysis (Figure 2B). rG4s are typically parallel-stranded [26] and display characteristic CD spectra with positive and negative peaks at 263-267 nm and 240 nm, respectively [39,40,41]. However, the characteristic signals for RNA duplex also appear in the same spectral region, typically with maximum at 260-270 nm and with or without minimum at 230-240 nm [42,43,44,45]. This complicates the unambiguous assignment of secondary structure. The CD signal pattern of G3, with a maximum at 265 nm and a minimum at 237 nm, is most consistent with rG4. Furthermore, only the G-to-U mutant of G3 exhibited a decrease in ellipticity. This again indicates that G3 is the most probable rG4-forming sequence. For G1 and G2, the ellipticity of the mutants was higher, which reinforces doubts about their ability to form G4s, and we did not observe perfect alignment of signals with the characteristic peaks of rG4. Thus, among the three sequences, G3 has the highest probability of forming the rG4 structure; however, the slight increase in ellipticity compared to the mutant cast doubt on this conclusion and necessitate further verification.
Comparative CD analysis of native and mutated sequences was performed in potassium and lithium-containing buffers (Figure S2) [46,47]. K+ cations typically stabilize rG4 structures by coordinating the guanine O6 atoms within G-quartet, whereas Li+ cannot provide such stabilization due to its smaller ionic radius [48,49]. Only G3 exhibited modest spectral alterations upon K+-to-Li+ replacement, with peaks shifting towards characteristics of RNA A-form helix (positive and negative peaks at 268 nm and 235 nm, respectively). The minimal spectral changes observed for all other sequences, both wild type and mutant, further supported the inability of the G1 and G2 sequences to fold into rG4 structures.
The results obtained for G3 in vitro led us to investigate whether it influences the DDX23 mRNA translation when treated with pyridostatin (PDS), a potent G4 stabilizer [50]. Notably, rG4 structures positioned downstream of uORFs may enhance uORF translation, thus suppressing protein synthesis from CDS through synergistic inhibition [28]. To evaluate the impact of G3 stabilization, if formed in cellulo, a dual luciferase assay was conducted (Figure 3). The reporter plasmid encodes Renilla luciferase (Rluc) as a control as well as the native 5’ terminus of DDX23 mRNA, which is fused to firefly luciferase (Fluc) without AUG codon. Additionally, we utilized the same construct lacking the uORF start codon, in which ATG was mutated to AAG, and a construct with a different short 5’ UTR lacking rG4-forming motifs and uORFs (Figure 3A).
We observed that uORF induces approximately 2.5-fold translation repression, as accessed by the change in the Fluc/Rluc ratio following uORF inactivation regardless the presence of PDS or DMSO (Figure 3B). This result aligns with efficient translation of uORF according to ribosome profiling data (Figure 1C). At the same time, PDS treatment induced a similar change in relative luciferase activities across all constructs tested (as evident after normalization to DMSO), calling into question the presence of rG4 or its influence on translation (Figure 3C).
Given the results of luciferase reporters, additional CD spectra were recorded in the presence of PDS. An increase in CD ellipticity was observed only for G1 (Figure 3D). However, considering previous experimental results, rG4 formation in the G1 sequence remains unlikely. The absence of the G3 response to the PDS treatment contradicted rG4 folding, despite the ThT-based light-up assay providing encouraging results and the CD spectral changes during K+-to-Li+ substitution (Figure 2A and Figure S2).
For further structural validation of G3 folding pattern, thermal difference spectra (TDS) and UV-melting analyses were used [51,52]. These methods revealed no characteristic hypochromic shift at 295 nm during melting, typically associated with rG4 structures, although TDS displayed minor negative peak around 295 nm (Figure 4A,B).
We hypothesized that G3 may fold into an alternative structure such as a hairpin. The potential hairpin formation is supported by the sequence’s capacity to form three stable canonical GC pairs and four GU wobble pairs [30,31,53]. CD melting of G3 was monophasic with Tm of 58 °C, indicating sufficient stability of the folded structure under physiological conditions (Figure 4C). Minimal hysteresis suggests an intramolecular structure.
The G3 structure was then investigated using 1H NMR spectroscopy (Figure 4D). A broad signal in the 10.8-11.8 ppm region may indicate rG4 formation, whereas other broad signals at 10.4, 11.9 and 13.4 ppm may correspond to the imino protons engaged in Watson-Crick base pairing. A poor signal-to-noise ratio, possibly due to aggregation or interconversion, prevented unambiguous structural assignment. The comparative analysis was conducted using two mutant sequences: mutG3 (a mutant sequence with no ability to form rG4) and h_mutG3 (designed to form exclusively rG4 structure through substitution of wobble pair-forming uridines with adenines) (Table S1). 1H NMR spectra of both mutants yielded clear interpretable signals. The sequence h_mutG3 demonstrated formation of a two-tetrad rG4, evidenced by eight signals from guanine imino atoms in the 10.8-11.8 ppm region, with minor fraction of an alternative rG4 conformation (Figure 4D). The mutG3 spectrum corresponded to the RNA hairpin-like conformation and contained four signals from imino protons of GU wobble pairs and four signals from canonical AU and GC pairs (Figure 4D). An additional signal may stem from a GU bifurcated pair [54,55] located in the region between the two double-stranded motifs. Comparative analysis of all three spectra suggests that G3 may form both rG4 and hairpin structures, as its 1H NMR spectrum contains imino proton signals in regions characteristic of both rG4 (as in h_mutG3) and Watson-Crick GC base pairs (as in mutG3).
CD spectra of mutG3 and h_mutG3 corroborated the structural assignments derived from the 1H NMR experiments. The CD spectrum of mutG3 had a positive peak at 260 nm, without minimum at 240 nm, which corresponds to a hairpin structure [42,56] (Figure S3). The presence of a positive peak at 263 nm and a negative peak at 240 nm indicated that h_mutG3 was folded into the rG4 structure. According to CD melting experiments, h_mutG3 was stabilized in the presence of 1 eq. PDS by 12 °C, while moderate (+7 °C) and little (+3 °C) changes in Tm for G3 and mutG3, respectively, were revealed (Figure 4E). The observed increase in G3 Tm does not contradict the absence of an increase in the CD maximum (Figure 3D), as has already been demonstrated for G4 ligands, including PDS [57,58]. In addition, UV melting proven to be ineffective for studying small RNAs that fold into alternative metastable structures [59]. These results confirmed that G3 could fold, at least partially, into rG4 in vitro.
To summarize this part, while the G3 sequence may adopt hairpin-like and rG4 conformations at equilibrium in vitro, as confirmed by spectroscopic methods, cell-based assays showed no response to the treatment with the G4 stabilizer PDS. Thus, none of the selected sequences forms a stable rG4 structure that may be regulatory. In contrast, a hairpin can be formed within the G3 sequence or the other part of the 5’ UTR. Such hairpin(s) could be involved in regulation of DDX23 mRNA translation.
2.2. Analysis of Regulatory Stem-Loop Elements
We identified the most probable hairpin within the 44-64 nt region by RNA fold [30,31]. Two oligoribonucleotides were synthesized to evaluate the influence of sequence context: hair_short (minimal hairpin-forming sequence) and hair_long (minimal hairpin-forming sequence with 6-nt flanks) (Table S1, Figure 1B). CD spectra showed maxima at 265 nm and minima at 233 nm (hair_short) and 236 nm (hair_long), consistent with A-form RNA helix formation (Figure 5A). CD melting analysis revealed Tm of 56 °C for hair_long and 66 °C for hair_short, indicating physiological stability despite the negative impact of additional 6-nt flanks on Tm (Figure 5B,C). Conditions with slow annealing produced alternative structures with uniform melting temperatures of 44 °C for both constructs (Figure 5B,C).
1H NMR spectra of both sequences contained three distinct signals that were attributed to guanine imino protons from Watson-Crick GC pairs (Figure 5D). Thus, CD and 1H NMR spectroscopies confirmed the hairpin structure predicted by RNA fold [30,31], which should be thermodynamically stable under cellular conditions.
The identified structured elements within 44-64 nt (hair_short) and 73-99 nt (G3) regions of DDX23 mRNA 5’ UTR may act as a translation-inhibiting element. While the thermal stability of hair_short appears insufficient for substantial translation inhibition (Tm around 60 °C, less than 13 kcal/mol based on ViennaRNAfold computational analysis [30,31]), its position downstream of the uORF start codon might facilitate synergistic inhibition of CDS translation, consistent with previous observations in other systems and partially explaining efficient uORF translation indicated by ribosome profiling data (Figure 1C) [60,61,62]. In turn, G3 also has Tm close to 60 °C and may promote the recognition of the uORF start codon and inhibit the CDS translation.
2.3 Evaluation of LNA-based ASOs Activity in DDX23 5’ UTR Translation Control
Since downregulation of DDX23 expression has shown therapeutic potential in cancer treatment, and current approaches are limited to non-selective helicase inhibitors and siRNAs with transient silencing effect [8,11], we designed a set of fully LNA-modified ASOs targeting start codons of uORF and CDS and/or the structured regions hair_short and G3 within DDX23 mRNA 5’ UTR and assessed their effects on the translation efficacy (Figure 6A, Table 1). LNA modifications were used to enhance nuclease resistance and hybridization properties, the latter being particularly important for unfolding of stem-loop structures. Previous study demonstrated that fully LNA-modified ASOs targeting 5’ UTR can equally or, in some cases, more effectively suppress mRNA translation compared to LNA-DNA chimeras with predicted ability to recruit RNase H [63]. Furthermore, fully modified blocker ASO targeting 5’ UTR can enhance the efficacy of RNase H1-activating ASOs that target mRNA coding regions [64].
Since the efficiency of ASO translation inhibition depends on the stability of ASO-RNA duplex, Tm was calculated using Tm predictor (Table 1) [65]. The predictor data show that all LNAs form duplexes with high stability under physiological conditions, exceeding the stability of the endogenous structures that have been detected in the 5’ UTR (with Tm < 70 °C). Therefore, LNAs are predicted to be highly effective in binding to mRNA and inhibiting translation. To evaluate LNA-modified ASO efficacy, we performed a dual luciferase assay. Fluc mRNA was synthesized using the wild-type DDX23 luciferase reporter construct and the construct with the inactivated uORF start codon (Figure 6B). For the control, we synthesized Rluc mRNA (Figure 6B) (see Materials and methods). HEK293T cells were co-transfected with Fluc reporter and Rluc control mRNAs as well as with ASOs LNA1-6 or the scramble LNA (SCR). The ASOs were used at 2, 10 and 100 nM concentrations, with SCR demonstrating no significant effects on the Fluc/Rluc ratio at any concentration compared to no ASO control (Figure 6C,D).
We observed translational repression with all ASOs targeting the DDX23 reporter mRNA of wild type, particularly at 100 nM concentration (Figure 6C). This result may be explained by the general inhibition of the translational machinery’s activity on the reporter mRNA. Nevertheless, the effects were not uniform across ASOs type and concentration. ASOs LNA1 and LNA2 target regions around the uORF start codon and should therefore mask it and repress uORF translation while enhancing CDS translation. However, they were the most effective inhibitors among ASOs under study at 10 and 100 nM concentrations (Figure 6C). This may be explained by their cap proximity, which hinders PIC loading. Interestingly, LNA ASOs exhibited a concentration dependency except for LNA3, which was less effective inhibitor. This difference was especially noticeable at 100 nM concentration (Figure 6C). Such effect may be explained by targeting of uORF or hair_short. To resolve this issue, we performed the same experiment with the DDX23 reporter mRNA lacking the uORF start codon. All LNAs showed inhibitory effects at 10 and 100 nM concentrations, which were concentration-dependent for all LNA ASOs (Figure 6D).
All LNA-modified ASOs except LNA2 and LNA3 demonstrated marginally diminished inhibitory effects for the no uORF reporter in comparison to the wild-type mRNA reporter (Figure 6E). Weaker effects observed for LNA2 in the mutated construct may be attributed to a nucleotide mismatch at its 3’ end because of the point substitution in the uORF start codon. Such mismatch should result in a reduction of the thermodynamic stability of the RNA–LNA duplex and also make it a more favorable substrate for RNA-helicases.
Interestingly, in the case of mutation in the uORF start codon, LNA3 exhibits approximately the same inhibitory properties as the other ASOs (Figure 6D). Thus, the low efficiency of translation inhibition by LNA3 in the WT reporter is likely attributable to uORF translation. Hair_short might be suggested to enhance uORF translation, so LNA3, by unwinding it, would be expected to reduce uORF translation and correspondingly increase translation of the CDS, thereby yielding the observed low apparent efficacy of LNA3 due to mixed positive and negative effects. However, this explanation is unlikely. First, by unwinding the endogenous hairpin (Tm close to 60 °C), LNA3 forms an even more stable mRNA–LNA3 duplex (predicted Tm 82 °C, Table 1) closer to the uORF start codon, which could even enhance uORF translation and, relative to other ASOs, more effectively inhibit translation of the CDS. Second, although LNA4, like LNA5 and LNA6, masks the CDS start codon, its effects on translation should be different due to its partial overlap with hair_short. Because none of these outcomes is observed, we suppose that both the endogenous hairpin and the mRNA–LNA3 duplex are insufficiently stable to increase uORF translation efficiency at that distance (>15 nt) from the uORF start codon. Moreover, enhancement of translation by a downstream hairpin has been reported primarily for start codons in a weak context [61,62,66], whereas in DDX23 the context is strong: A at –3 and G at +4 positions (Figure 1) considering the optimal Kozak context [67].
A more plausible explanation is that the ribosome translating the uORF impedes LNA3 binding to the 5’ UTR and/or promotes the dissociation of already bound LNA3, which accounts for its low efficacy as a translation inhibitor. A similar effect is known for ASOs targeting CDS [64]; however, to our knowledge, it has not been described for uORFs. This model also explains the concentration-independent effects of LNA3 on translation of the wild-type construct (Figure 6C). Our explanation is indirectly supported by ribosome profiling data (Figure 1C) indicating high ribosome occupancy on the uORF of endogenous DDX23 mRNA, higher than on the CDS. This suggests that the LNA3 binding site is particularly well shielded by translating ribosomes.
Thus, our study of LNA-ASOs suggests a new hypothesis/design principle for ASOs inhibiting translation: an ASO should not target an efficiently translated uORF, because the high ribosome density in this region will preclude effective binding to the 5’ UTR. Furthermore, given that neither the unwinding of hair_short with LNA4 nor the formation of a more stable mRNA–LNA3 duplex closer to the uORF start codon has an effect on the efficiency of the CDS translation, we hypothesize that hair_short does not affect uORF translation efficiency, but rather may act as a weak inhibitor of the CDS translation by imposing steric hindrance to PIC scanning along the 5’ UTR or have a role unrelated to translation.
3. Materials and Methods
3.1 Oligonucleotide Synthesis and Sample Preparation
Unmodified RNAs were synthesized according to the optimized synthetic protocol on an automated ASM-800 DNA/RNA synthesizer (Biosset, Russia). For detritylation, capping and oxidation, 3% solution of dichloroacetic acid in CH2Cl2, propionic anhydride and N-methylimidazole, and iodine solution in tetrahydrofuran: water: pyridine mixture, respectively, were used. At the condensation stage, 0.1 M solutions of protected 2’-O-TBDMS-ribonucleotide 3’-phosphoramidites (ChemGen, USA) in absolute CH3CN and 0.25 M solution of activator 42 (Sigma-Aldrich, USA) in absolute CH3CN were used. Time of condensation was 7 min. The last 5’-O-dimetoxytrityl group was removed, and oligonucleotides were deprotected and cleaved from CPG support by the treatment with AMA (40% methylamine: 30% ammonium, 1:1) for 2 h at room temperature with subsequent treatment with NMP:TEA:TEA·3HF mixture for 1.5 h at 65 °C. After an addition of ethoxytrimethylsilane (Sigma-Aldrich, USA), precipitation with ethyl ether was carried out. All RNAs were isolated by preparative denaturating 15% PAGE. The RNA homogeneity was analyzed by analytical gel electrophoresis in denaturating 15% PAGE. Fully LNA-modified ASOs were purchased from Lumiprobe (Russia).
RNA solutions in 20 mM sodium phosphate buffer supplemented with 140 mM KCl (K+-buffer) or LiCl (Li+-buffer), pH 7.4, were annealed rapidly (heated to 90 °C and then snap-cooled on ice) to facilitate intramolecular folding. Thioflavin T (ThT) and pyridostatin (PDS) were obtained from Sigma-Aldrich (USA).
3.2 Thioflavin T (ThT) Light-Up Assays
Experiments were conducted using 384-well U-Bottom White Polystyrene microplates (CORNING, USA). Annealed RNA samples in K+-buffer and ThT were mixed to final concentrations of 2 µM and 1 µM, respectively, and incubated for 15 min. Measurements were performed at 25 °C. Fluorescence emission spectra were collected from 470 nm to 650 nm with excitation at 440 nm using a M200 PRO Tecan microplate reader (Tecan Group Ltd., Switzerland). Fluorescence intensities at 485 nm were used for data analysis.
3.3 Circular Dichroism (CD) and UV Spectroscopy and Melting
CD and UV measurements were performed using a Chirascan spectropolarimeter (Applied Photophysics, UK) with a 10 mm optical path quartz cuvette. CD spectra of annealed 2 µM RNA samples in K+- or Li+-buffer were recorded at 10 °C from 230 nm to 300 nm. TDS spectra were obtained by subtracting the absorption spectra in the range 230 nm to 320 nm recorded at 90 °C from those recorded at 10 °C. For the experiments with PDS, annealed 2 µM RNA samples in K+-buffer were supplemented with PDS (2.5 mM stock solution in DMSO) to 2 µM final concentration or equivalent DMSO volume. Each reported spectrum represents an average of 2 scans. Melting/annealing experiments were conducted with a heating/cooling rate of 1 °C/min from 20 °C to 90 °C, monitoring changes at 265 nm (CD melting) or 295 nm (UV melting).
3.4 NMR Experiments
Samples was prepared in DEPC-treated water (100 μM RNA oligonucleotide, 25 mM PBS, 100 mM KCl, 90:10 H2O/D2O, pH 7.0) and annealed rapidly by heating at 90 °C for 5 min and then snap-cooling on ice for 10 min. NMR spectra were acquired at 25 °C on Bruker Avance III 600 MHz spectrometer (Bruker, Germany) equipped with a triple resonance cryogenic probe and Bruker Topspin 3.2 software. 1H spectra were recorded using the standard zgesgp pulse sequence with water suppression at 4.7 ppm, 16 dummy scans and 1024 to 4096 scans. The pre-scan delay was set to 1 sec. FIDs were processed with an exponential window function and line broadening of 5.0 Hz.
3.5 Cell Culturing
The HEK293T cells (ATCC) were cultivated in DMEM/F12 medium (Servicebio), supplemented with L-alanyl-L-glutamine (Himedia, India), 10% fetal bovine serum (Himedia, India), 100 units/ml penicillin, and 100 μg/ml streptomycin (Himedia) at 37 °C in 5% CO2 atmosphere. Cells were routinely tested for the absence of mycoplasma using the MycoReport kit (Evrogen, Russia).
3.6 Plasmid Constructs
The plasmid for the dual luciferase assay, containing firefly (Fluc) and Renilla (Rluc) under different promoters, was made from the pGL3-Rluc vector (a kind gift from Radik R. Shafikov, unpublished). The vector backbone was amplified using Q5 High-Fidelity DNA Polymerase (NEB, USA) with primers 5’-AGACGAGTTGGCACGTCTCAGGAGAAGACGCCAAAAACATAAAG-3’ and 5’-TAAGAGCTCGGTACCTATCGATAGAGAAATGTTCTGGCAC-3’, and the EF-1α core promoter was amplified from the lentiCas9-Blast plasmid (Addgene #52962) using primers 5’-AACATTTCTCTATCGATAGGTACCGAGCTCTTAGGGCAGAGCGCACATCG-3’ and 5’-TTCTCCTGAGACGTGCCAACTCGTCTCGACCTAACCGGTCCTGTGTTCTG-3’. The products were assembled into the reporter vector using NEBuilder HiFi DNA Assembly Master Mix (NEB, USA), introducing two BsmBI sites after the EF-1α core promoter and immediately before the Fluc ORF without AUG codon. The 5’ end of the DDX23 transcript, including 20 codons of CDS, was amplified from cDNA, obtained by the reverse transcriptase Magnus and oligo(dT)15 primer (Evrogen, Russia), using primers 5’-GAACCTCGTCTCTAGGTTTCATCTCCGCGACCAG-3’ and 5’-TCAATGCGTCTCACTCCCCTTTCCTCCTTGGAAGGT-3’. The PCR-product was cloned into Esp3I sites of the reporter vector using Esp3I (Thermo Fisher, USA) and T4 ligase (Evrogen, Russia). The mutant with inactivated uORF start codon (no uORF) was obtained by overlap extension PCR of the wild-type sequence using primers 5’-GAAAGAAGGCGACGGCTCCGC-3’ and 5’-AGCCGTCGCCTTCTTTCC-3’.
3.7 Transfection
HEK293T cells were transfected with plasmids using the Lipofectamine 3000 Transfection Reagent Kit (Invitrogen, USA) in 96-well format according to the manufacturer’s protocol. In brief, 24,000 cells per well were seeded overnight in 200 µl of the medium for plasmid transfection, or 100 µl of the medium for mRNA transfection. 50 ng of plasmid DNA or a mixture containing 49 ng of Fluc reporter mRNA, 1 ng of Rluc control mRNA, and 10 pmol of LNA ASO, as well as 0.2 µl of Lipofectamine 3000 Transfection Reagent per well were used. If a treatment was required, the medium was replaced with a substance-supplemented medium prior to adding the transfection mixtures. The cells were harvested 4 hours after mRNA transfection and 24 hours after plasmid transfection for luciferase assays.
3.8 In vitro Transcription
Fluc reporter constructs were amplified (Q5 High-Fidelity DNA Polymerase, NEB) from plasmid reporters using primers 5’-GAGTACTTAATACGACTCACTATAGATCTCCGCGACCAGGAAACG-3’ and 5’-TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTCCGCCCCGACTCTAGAATTACACGG-3’, and the Rluc control was amplified from pGL4.70[hRluc] (Promega) using primers 5’-GAGTACTTAATACGACTCACTATAGTGGTTCTTTCCGCCTCAGGCCACCATGGCTTCCAAG-3’ and 5’-TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTCTCTAGAATTACTGCTCGTTCTTCAGCACG-3’ to add the T7 promoter and a poly(A) tail consisting of 50 adenosine bases. Capped mRNA was synthesized from these PCR products and was treated with TURBO DNase using the mMESSAGE mMACHINE T7 Transcription Kit (Invitrogen). The mRNA was subsequently purified with the CleanRNA Standard kit (Evrogen, Russia), and its integrity was assessed by denaturing urea polyacrylamide gel electrophoresis. RNA concentrations were measured using the Qubit RNA HS Assay Kit (Invitrogen).
3.9 Luciferase Assay
Luciferase activities were measured using the TransDetect Double-Luciferase Reporter Assay Kit (TransGen Biotech, China) according to manufacturer protocol in a 96-well format using Victor X5 plate reader (Perkin Elmer, USA).
3.10 Visualization of Ribosome Profiling
Aggregate ribosome profiling data were accessed using the RiboCrypt browser for ENST00000308025 in RFP modality for all merged Homo sapiens datasets (accessed on 22 September 2025). The link is available below.
3.11 Statistical Analysis
The samples were analyzed using the Shapiro-Wilk test, indicating normal distribution at p > 0.05. Statistical analysis was performed using GraphPad Prism 10.3.1 software (GraphPad Software).
4. Conclusions
Using fluorescence, CD, UV and 1H NMR spectroscopies, two structured elements were identified within DDX23 5’ UTR mRNA coding for the full-length protein and then, together with the sequences covering start codons, were targeted with LNA-modified ASOs. The study yields several significant outcomes. First, it suggests a new principle for designing translation-inhibiting ASOs: avoid targeting efficiently translated uORFs, as their high ribosome density blocks ASO binding. Second, there are several elements in DDX23 mRNA 5’ UTR relevant to its translation. The first is uORF, which represses the CDS translation, and the other are two structured elements within 44-64 nt and 76-96 nt. The hairpin hair_short within 44-64 nt likely inhibits translation of DDX23 mRNA blocking PIC scanning, without interference with uORF translation, 76-96 nt element may have a similar influence. ASO targeting 5’ proximal region of mRNA are effective translation inhibitors due to impaired PIC loading. To carefully modulate uORF translation, the thermal stability of ASO-RNA duplex formed should be carefully tuned to avoid creating a roadblock for translation machinery, more powerful than endogenous structures. These findings provide valuable insights for developing more effective ASOs targeting the main DDX23 transcript, and mRNA in general, and regulating its translation for potential anticancer applications.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1, Table S1: RNA oligonucleotides used in this study; Figure S1: rG4-forming sequences in accordance with G4Atlas database; Figure S2: CD spectra of putative rG4-forming sequences G1, G2 and G3 and their G-to-U mutants in K+- and Li+-containing buffers; Figure S3: CD spectra of G3, h_mutG3 and mutG3 ONs.
Author Contributions
Conceptualization, P.K., N.S. and A.A.; methodology, P.K., N.S., M.R. and A.A.; investigation, P.K., N.S., V.Z., V.B., A.C., D.R., E.K. and D.N.; writing—original draft preparation, P.K., N.S. and A.A.; writing—review and editing, M.R.; visualization, P.K., N.S. and A.A.; supervision, M.R.; project administration, A.A.; funding acquisition, M.R. and A.A. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by The Russian Scientific Foundation grant 24-15-00236 to A.A. (all experiments except for generating reporter plasmids) and by The Russian Scientific Foundation grant 21-64-00006-P to M.R. (construction of reporter plasmids).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ASO | Antisense oligonucleotide |
| CD | Circular dichroism |
| CDS | Coding sequence |
| CH2Cl2 | Methylene chloride |
| CH3CN | Acetonitrile |
| CPG | Controlled pore glass |
| DDX23 | DEAD-box helicase 23 |
| DEPC | Diethyl pyrocarbonate |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DMSO | Dimethyl sulfoxide |
| dsRNA | Double-stranded RNA |
| ER | Estrogen receptor |
| HF | Hydrofluoric acid |
| LNA | Locked nucleic acid |
| NMP | N-Methyl-2-pyrrolidone |
| NMR | Nuclear magnetic resonance |
| PBS | Phosphate-buffered saline |
| PDS | Pyridostatin |
| PIC | Pre-initiation complex |
| ppm | Parts per million |
| rG4 | RNA G-quadruplex |
| SCR | Scramble |
| rTBA15 | RNA 15-nt thrombin binding aptamer |
| TBDMS | tert-Butyldimethylsilyl |
| TDS | Thermal difference spectra |
| TEA | Triethylamine |
| ThT | Thioflavin T |
| Tm | Melting temperature |
| uORF | Upstream open reading frame |
| 5′ UTR | 5′ Untranslated region |
| UV | Ultraviolet |
References
- Cordin, O.; Beggs, J.D. RNA Helicases in Splicing. RNA Biol. 2013, 10, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Teigelkamp, S.; Mundt, C.; Achsel, T.; Will, C.L.; Lührmann, R. The Human U5 snRNP-Specific 100-kD Protein Is an RS Domain-Containing, Putative RNA Helicase with Significant Homology to the Yeast Splicing Factor Prp28p. RNA N. Y. N 1997, 3, 1313–1326. [Google Scholar]
- Segovia, D.; Adams, D.W.; Hoffman, N.; Safaric Tepes, P.; Wee, T.-L.; Cifani, P.; Joshua-Tor, L.; Krainer, A.R. SRSF1 Interactome Determined by Proximity Labeling Reveals Direct Interaction with Spliceosomal RNA Helicase DDX23. Proc. Natl. Acad. Sci. U. S. A. 2024, 121, e2322974121. [Google Scholar] [CrossRef] [PubMed]
- Sridhara, S.C.; Carvalho, S.; Grosso, A.R.; Gallego-Paez, L.M.; Carmo-Fonseca, M.; de Almeida, S.F. Transcription Dynamics Prevent RNA-Mediated Genomic Instability through SRPK2-Dependent DDX23 Phosphorylation. Cell Rep. 2017, 18, 334–343. [Google Scholar] [CrossRef]
- Ruan, J.; Cao, Y.; Ling, T.; Li, P.; Wu, S.; Peng, D.; Wang, Y.; Jia, X.; Chen, S.; Xu, A.; et al. DDX23, an Evolutionary Conserved dsRNA Sensor, Participates in Innate Antiviral Responses by Pairing With TRIF or MAVS. Front. Immunol. 2019, 10, 2202. [Google Scholar] [CrossRef]
- Abdullah, S.W.; Han, S.; Wu, J.; Zhang, Y.; Bai, M.; Jin, Y.; Zhi, X.; Guan, J.; Sun, S.; Guo, H. The DDX23 Negatively Regulates Translation and Replication of Foot-and-Mouth Disease Virus and Is Degraded by 3C Proteinase. Viruses 2020, 12, 1348. [Google Scholar] [CrossRef]
- Murillo, A.; Vera-Estrella, R.; Barkla, B.J.; Méndez, E.; Arias, C.F. Identification of Host Cell Factors Associated with Astrovirus Replication in Caco-2 Cells. J. Virol. 2015, 89, 10359–10370. [Google Scholar] [CrossRef]
- Zhao, C.; Li, Y.; Qiu, C.; Chen, J.; Wu, H.; Wang, Q.; Ma, X.; Song, K.; Kong, B. Splicing Factor DDX23, Transcriptionally Activated by E2F1, Promotes Ovarian Cancer Progression by Regulating FOXM1. Front. Oncol. 2021, 11, 749144. [Google Scholar] [CrossRef]
- Mao, G.; Jin, H.; Wu, L. DDX23-Linc00630-HDAC1 Axis Activates the Notch Pathway to Promote Metastasis. Oncotarget 2017, 8, 38937–38949. [Google Scholar] [CrossRef]
- Lin, C.; Li, T.; Wang, Y.; Lai, S.; Huang, Y.; Guo, Z.; Zhang, X.; Weng, S. METTL3 Enhances Pancreatic Ductal Adenocarcinoma Progression and Gemcitabine Resistance through Modifying DDX23 mRNA N6 Adenosine Methylation. Cell Death Dis. 2023, 14, 221. [Google Scholar] [CrossRef]
- Yin, J.; Park, G.; Lee, J.E.; Choi, E.Y.; Park, J.Y.; Kim, T.-H.; Park, N.; Jin, X.; Jung, J.-E.; Shin, D.; et al. DEAD-Box RNA Helicase DDX23 Modulates Glioma Malignancy via Elevating miR-21 Biogenesis. Brain 2015, 138, 2553–2570. [Google Scholar] [CrossRef]
- Burns, W.; Bird, L.M.; Heron, D.; Keren, B.; Ramachandra, D.; Thiffault, I.; Del Viso, F.; Amudhavalli, S.; Engleman, K.; Parenti, I.; et al. Syndromic Neurodevelopmental Disorder Associated with de Novo Variants in DDX23. Am. J. Med. Genet. A. 2021, 185, 2863–2872. [Google Scholar] [CrossRef]
- Zhang, R.; Chen, Y.; He, J.; Gou, H.-Y.; Zhu, Y.-L.; Zhu, Y.-M. WGCNA Combined with GSVA to Explore Biomarkers of Refractory Neocortical Epilepsy. IBRO Neurosci. Rep. 2022, 13, 314–321. [Google Scholar] [CrossRef]
- Asberger, J.; Erbes, T.; Jaeger, M.; Rücker, G.; Nöthling, C.; Ritter, A.; Berner, K.; Juhasz-Böss, I.; Hirschfeld, M. Endoxifen and Fulvestrant Regulate Estrogen-Receptor α and Related DEADbox Proteins. Endocr. Connect. 2020, 9, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Crooke, S.T.; Baker, B.F.; Crooke, R.M.; Liang, X.-H. Antisense Technology: An Overview and Prospectus. Nat. Rev. Drug Discov. 2021, 20, 427–453. [Google Scholar] [CrossRef] [PubMed]
- Dhuri, K.; Bechtold, C.; Quijano, E.; Pham, H.; Gupta, A.; Vikram, A.; Bahal, R. Antisense Oligonucleotides: An Emerging Area in Drug Discovery and Development. J. Clin. Med. 2020, 9, 2004. [Google Scholar] [CrossRef] [PubMed]
- Boiziau, C.; Kurfurst, R.; Cazenave, C.; Roig, V.; Thuong, N.T.; Toulmé, J.-J. Inhibition of Translation Initiation by Antisense Oligonucleotides via an RNase-H Independent Mechanism. Nucleic Acids Res. 1991, 19, 1113–1119. [Google Scholar] [CrossRef]
- Liang, X.; Nichols, J.G.; Hsu, C.-W.; Vickers, T.A.; Crooke, S.T. mRNA Levels Can Be Reduced by Antisense Oligonucleotides via No-Go Decay Pathway. Nucleic Acids Res. 2019, 47, 6900–6916. [Google Scholar] [CrossRef]
- Dias, N.; Dheur, S.; Nielsen, P.E.; Gryaznov, S.; Van Aerschot, A.; Herdewijn, P.; Hélène, C.; Saison-Behmoaras, T.E. Antisense PNA Tridecamers Targeted to the Coding Region of Ha-Ras mRNA Arrest Polypeptide Chain Elongation. J. Mol. Biol. 1999, 294, 403–416. [Google Scholar] [CrossRef]
- Liang, X.; Shen, W.; Sun, H.; Migawa, M.T.; Vickers, T.A.; Crooke, S.T. Translation Efficiency of mRNAs Is Increased by Antisense Oligonucleotides Targeting Upstream Open Reading Frames. Nat. Biotechnol. 2016, 34, 875–880. [Google Scholar] [CrossRef]
- Liang, X.; Sun, H.; Shen, W.; Wang, S.; Yao, J.; Migawa, M.T.; Bui, H.-H.; Damle, S.S.; Riney, S.; Graham, M.J.; et al. Antisense Oligonucleotides Targeting Translation Inhibitory Elements in 5′ UTRs Can Selectively Increase Protein Levels. Nucleic Acids Res. 2017, 45, 9528–9546. [Google Scholar] [CrossRef] [PubMed]
- Braasch, D.A.; Corey, D.R. Locked Nucleic Acid (LNA): Fine-Tuning the Recognition of DNA and RNA. Chem. Biol. 2001, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- McTigue, P.M.; Peterson, R.J.; Kahn, J.D. Sequence-Dependent Thermodynamic Parameters for Locked Nucleic Acid (LNA)−DNA Duplex Formation. Biochemistry 2004, 43, 5388–5405. [Google Scholar] [CrossRef] [PubMed]
- Cadoni, E.; De Paepe, L.; Manicardi, A.; Madder, A. Beyond Small Molecules: Targeting G-Quadruplex Structures with Oligonucleotides and Their Analogues. Nucleic Acids Res. 2021, 49, 6638–6659. [Google Scholar] [CrossRef]
- Varshney, D.; Spiegel, J.; Zyner, K.; Tannahill, D.; Balasubramanian, S. The Regulation and Functions of DNA and RNA G-Quadruplexes. Nat. Rev. Mol. Cell Biol. 2020, 21, 459–474. [Google Scholar] [CrossRef]
- Kamzeeva, P.N.; Alferova, V.A.; Korshun, V.A.; Varizhuk, A.M.; Aralov, A.V. 5’-UTR G-Quadruplex-Mediated Translation Regulation in Eukaryotes: Current Understanding and Methodological Challenges. Int. J. Mol. Sci. 2025, 26, 1187. [Google Scholar] [CrossRef]
- Svoboda, P.; Cara, A.D. Hairpin RNA: A Secondary Structure of Primary Importance. Cell. Mol. Life Sci. 2006, 63, 901–908. [Google Scholar] [CrossRef]
- Murat, P.; Marsico, G.; Herdy, B.; Ghanbarian, A.; Portella, G.; Balasubramanian, S. RNA G-Quadruplexes at Upstream Open Reading Frames Cause DHX36- and DHX9-Dependent Translation of Human mRNAs. Genome Biol. 2018, 19, 229. [Google Scholar] [CrossRef]
- Yu, H.; Qi, Y.; Yang, B.; Yang, X.; Ding, Y. G4Atlas: A Comprehensive Transcriptome-Wide G-Quadruplex Database. Nucleic Acids Res. 2023, 51, D126–D134. [Google Scholar] [CrossRef]
- Gruber, A.R.; Lorenz, R.; Bernhart, S.H.; Neuböck, R.; Hofacker, I.L. The Vienna RNA Websuite. Nucleic Acids Res. 2008, 36, W70–74. [Google Scholar] [CrossRef]
- Hofacker, I.L. Vienna RNA Secondary Structure Server. Nucleic Acids Res. 2003, 31, 3429–3431. [Google Scholar] [CrossRef] [PubMed]
- Morales, J.; Pujar, S.; Loveland, J.E.; Astashyn, A.; Bennett, R.; Berry, A.; Cox, E.; Davidson, C.; Ermolaeva, O.; Farrell, C.M.; et al. A Joint NCBI and EMBL-EBI Transcript Set for Clinical Genomics and Research. Nature 2022, 604, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Kikin, O.; D’Antonio, L.; Bagga, P.S. QGRS Mapper: A Web-Based Server for Predicting G-Quadruplexes in Nucleotide Sequences. Nucleic Acids Res. 2006, 34, W676–682. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Lejault, P.; Chevrier, S.; Boidot, R.; Robertson, A.G.; Wong, J.M.Y.; Monchaud, D. Transcriptome-Wide Identification of Transient RNA G-Quadruplexes in Human Cells. Nat. Commun. 2018, 9, 4730. [Google Scholar] [CrossRef]
- Guo, J.U.; Bartel, D.P. RNA G-Quadruplexes Are Globally Unfolded in Eukaryotic Cells and Depleted in Bacteria. Science 2016, 353, aaf5371. [Google Scholar] [CrossRef]
- Xu, S.; Li, Q.; Xiang, J.; Yang, Q.; Sun, H.; Guan, A.; Wang, L.; Liu, Y.; Yu, L.; Shi, Y.; et al. Thioflavin T as an Efficient Fluorescence Sensor for Selective Recognition of RNA G-Quadruplexes. Sci. Rep. 2016, 6, 24793. [Google Scholar] [CrossRef]
- Zaitseva, S.O.; Baleeva, N.S.; Zatsepin, T.S.; Myasnyanko, I.N.; Turaev, A.V.; Pozmogova, G.E.; Khrulev, A.A.; Varizhuk, A.M.; Baranov, M.S.; Aralov, A.V. Short Duplex Module Coupled to G-Quadruplexes Increases Fluorescence of Synthetic GFP Chromophore Analogues. Sensors 2020, 20, 915. [Google Scholar] [CrossRef]
- Liu, S.; Peng, P.; Wang, H.; Shi, L.; Li, T. Thioflavin T Binds Dimeric Parallel-Stranded GA-Containing Non-G-Quadruplex DNAs: A General Approach to Lighting up Double-Stranded Scaffolds. Nucleic Acids Res. 2017, 45, 12080–12089. [Google Scholar] [CrossRef]
- Kumari, S.; Bugaut, A.; Huppert, J.L.; Balasubramanian, S. An RNA G-Quadruplex in the 5′ UTR of the NRAS Proto-Oncogene Modulates Translation. Nat. Chem. Biol. 2007, 3, 218–221. [Google Scholar] [CrossRef]
- Hacht, A.V.; Seifert, O.; Menger, M.; Schütze, T.; Arora, A.; Konthur, Z.; Neubauer, P.; Wagner, A.; Weise, C.; Kurreck, J. Identification and Characterization of RNA Guanine-Quadruplex Binding Proteins. Nucleic Acids Res. 2014, 42, 6630–6644. [Google Scholar] [CrossRef]
- Pany, S.P.P.; Sapra, M.; Sharma, J.; Dhamodharan, V.; Patankar, S.; Pradeepkumar, P.I. Presence of Potential G-Quadruplex RNA-Forming Motifs at the 5′-UTR of PP2Acα mRNA Repress Translation. ChemBioChem 2019, 20, 2955–2960. [Google Scholar] [CrossRef] [PubMed]
- Harpster, C.; Boyle, E.; Musier-Forsyth, K.; Kankia, B. HIV-1 Genomic RNA U3 Region Forms a Stable Quadruplex-Hairpin Structure. Biophys. Chem. 2021, 272, 106567. [Google Scholar] [CrossRef] [PubMed]
- Bugaut, A.; Murat, P.; Balasubramanian, S. An RNA Hairpin to G-Quadruplex Conformational Transition. J. Am. Chem. Soc. 2012, 134, 19953–19956. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, M.; Sigurdsson, S.; Tomac, S.; Sen, S.; Rozners, E.; Sjöberg, B.-M.; Strömberg, R.; Gräslund, A. A Synthetic Model for Triple-Helical Domains in Self-Splicing Group I Introns Studied by Ultraviolet and Circular Dichroism Spectroscopy. Biochemistry 1996, 35, 4678–4688. [Google Scholar] [CrossRef]
- Chauca-Diaz, A.M.; Jung Choi, Y.; Resendiz, M.J.E. Biophysical Properties and Thermal Stability of Oligonucleotides of RNA Containing 7,8-dihydro-8-hydroxyadenosine. Biopolymers 2015, 103, 167–174. [Google Scholar] [CrossRef]
- Majumder, P.; Shukla, C.; Arya, A.; Sharma, S.; Datta, B. G-Quadruplexes in MTOR and Induction of Autophagy. Sci. Rep. 2024, 14, 2525. [Google Scholar] [CrossRef]
- G-Quadruplex Nucleic Acids: Methods and Protocols; Yang, D., Lin, C., Eds.; Methods in molecular biology; Humana Press: New York, NY, 2019; ISBN 978-1-4939-9666-7.
- Bhattacharyya, D.; Mirihana Arachchilage, G.; Basu, S. Metal Cations in G-Quadruplex Folding and Stability. Front. Chem. 2016, 4, 38. [Google Scholar] [CrossRef]
- Lyons, S.M.; Gudanis, D.; Coyne, S.M.; Gdaniec, Z.; Ivanov, P. Identification of Functional Tetramolecular RNA G-Quadruplexes Derived from Transfer RNAs. Nat. Commun. 2017, 8, 1127. [Google Scholar] [CrossRef]
- Rodriguez, R.; Müller, S.; Yeoman, J.A.; Trentesaux, C.; Riou, J.-F.; Balasubramanian, S. A Novel Small Molecule That Alters Shelterin Integrity and Triggers a DNA-Damage Response at Telomeres. J. Am. Chem. Soc. 2008, 130, 15758–15759. [Google Scholar] [CrossRef]
- Mergny, J.-L. Thermal Difference Spectra: A Specific Signature for Nucleic Acid Structures. Nucleic Acids Res. 2005, 33, e138. [Google Scholar] [CrossRef]
- Mergny, J.-L.; Lacroix, L. UV Melting of G-Quadruplexes. Curr. Protoc. Nucleic Acid Chem. 2009, Chapter 17, 17.1.1-17.1.15. [CrossRef]
- Varani, G.; McClain, W.H. The G·U Wobble Base Pair: A Fundamental Building Block of RNA Structure Crucial to RNA Function in Diverse Biological Systems. EMBO Rep. 2000, 1, 18–23. [Google Scholar] [CrossRef]
- Correll, C.C.; Freeborn, B.; Moore, P.B.; Steitz, T.A. Metals, Motifs, and Recognition in the Crystal Structure of a 5S rRNA Domain. Cell 1997, 91, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Ban, N.; Nissen, P.; Hansen, J.; Moore, P.B.; Steitz, T.A. The Complete Atomic Structure of the Large Ribosomal Subunit at 2.4 Å Resolution. Science 2000, 289, 905–920. [Google Scholar] [CrossRef] [PubMed]
- Baranowski, D.S.; Kotkowiak, W.; Kierzek, R.; Pasternak, A. Hybridization Properties of RNA Containing 8-Methoxyguanosine and 8-Benzyloxyguanosine. PLoS ONE 2015, 10, e0137674. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.; Shah, V.; Datta, B. Assessing G4-Binding Ligands In Vitro and in Cellulo Using Dimeric Carbocyanine Dye Displacement Assay. Molecules 2021, 26, 1400. [Google Scholar] [CrossRef]
- Pagano, A.; Iaccarino, N.; Abdelhamid, M.A.S.; Brancaccio, D.; Garzarella, E.U.; Di Porzio, A.; Novellino, E.; Waller, Z.A.E.; Pagano, B.; Amato, J.; et al. Common G-Quadruplex Binding Agents Found to Interact With i-Motif-Forming DNA: Unexpected Multi-Target-Directed Compounds. Front. Chem. 2018, 6, 281. [Google Scholar] [CrossRef]
- Micura, R.; Höbartner, C. On Secondary Structure Rearrangements and Equilibria of Small RNAs. ChemBioChem 2003, 4, 984–990. [Google Scholar] [CrossRef]
- Kiniry, S.J.; Michel, A.M.; Baranov, P.V. The GWIPS-viz Browser. Curr. Protoc. Bioinforma. 2018, 62, e50. [Google Scholar] [CrossRef]
- Wang, J.; Shin, B.-S.; Alvarado, C.; Kim, J.-R.; Bohlen, J.; Dever, T.E.; Puglisi, J.D. Rapid 40S Scanning and Its Regulation by mRNA Structure during Eukaryotic Translation Initiation. Cell 2022, 185, 4474–4487.e17. [Google Scholar] [CrossRef]
- Hedaya, O.M.; Venkata Subbaiah, K.C.; Jiang, F.; Xie, L.H.; Wu, J.; Khor, E.-S.; Zhu, M.; Mathews, D.H.; Proschel, C.; Yao, P. Secondary Structures That Regulate mRNA Translation Provide Insights for ASO-Mediated Modulation of Cardiac Hypertrophy. Nat. Commun. 2023, 14, 6166. [Google Scholar] [CrossRef]
- Braasch, D.A. Antisense Inhibition of Gene Expression in Cells by Oligonucleotides Incorporating Locked Nucleic Acids: Effect of mRNA Target Sequence and Chimera Design. Nucleic Acids Res. 2002, 30, 5160–5167. [Google Scholar] [CrossRef]
- Liang, X.-H.; Nichols, J.G.; Sun, H.; Crooke, S.T. Translation Can Affect the Antisense Activity of RNase H1-Dependent Oligonucleotides Targeting mRNAs. Nucleic Acids Res. 2018, 46, 293–313. [Google Scholar] [CrossRef]
- QIAGENE Tm Prediction GeneGlob Available online: https://geneglobe.qiagen.com/us/tools/tm-prediction.
- Xiang, Y.; Huang, W.; Tan, L.; Chen, T.; He, Y.; Irving, P.S.; Weeks, K.M.; Zhang, Q.C.; Dong, X. Pervasive Downstream RNA Hairpins Dynamically Dictate Start-Codon Selection. Nature 2023, 621, 423–430. [Google Scholar] [CrossRef]
- Kozak, M. The scanning model for translation: An update. J. Cell. Biol. 1989, 108, 229–241. [Google Scholar] [CrossRef]
Figure 1.
Putative rG4- and hairpin-prone sequences within 5’ UTR of DDX23 mRNA. RNA sequences that contain rG4-forming motifs predicted by QGRS mapper and 3-nt flanks (A) and hairpin-forming sequences predicted by RNA fold [30,31] (B). Aggregated ribosome profiling data shown using Ribocrypt (С). RFP—ribosome footprints, nt—nucleotide. 5’ UTR and a proximal region of the coding sequence of DDX23 mRNA are shown.
Figure 1.
Putative rG4- and hairpin-prone sequences within 5’ UTR of DDX23 mRNA. RNA sequences that contain rG4-forming motifs predicted by QGRS mapper and 3-nt flanks (A) and hairpin-forming sequences predicted by RNA fold [30,31] (B). Aggregated ribosome profiling data shown using Ribocrypt (С). RFP—ribosome footprints, nt—nucleotide. 5’ UTR and a proximal region of the coding sequence of DDX23 mRNA are shown.
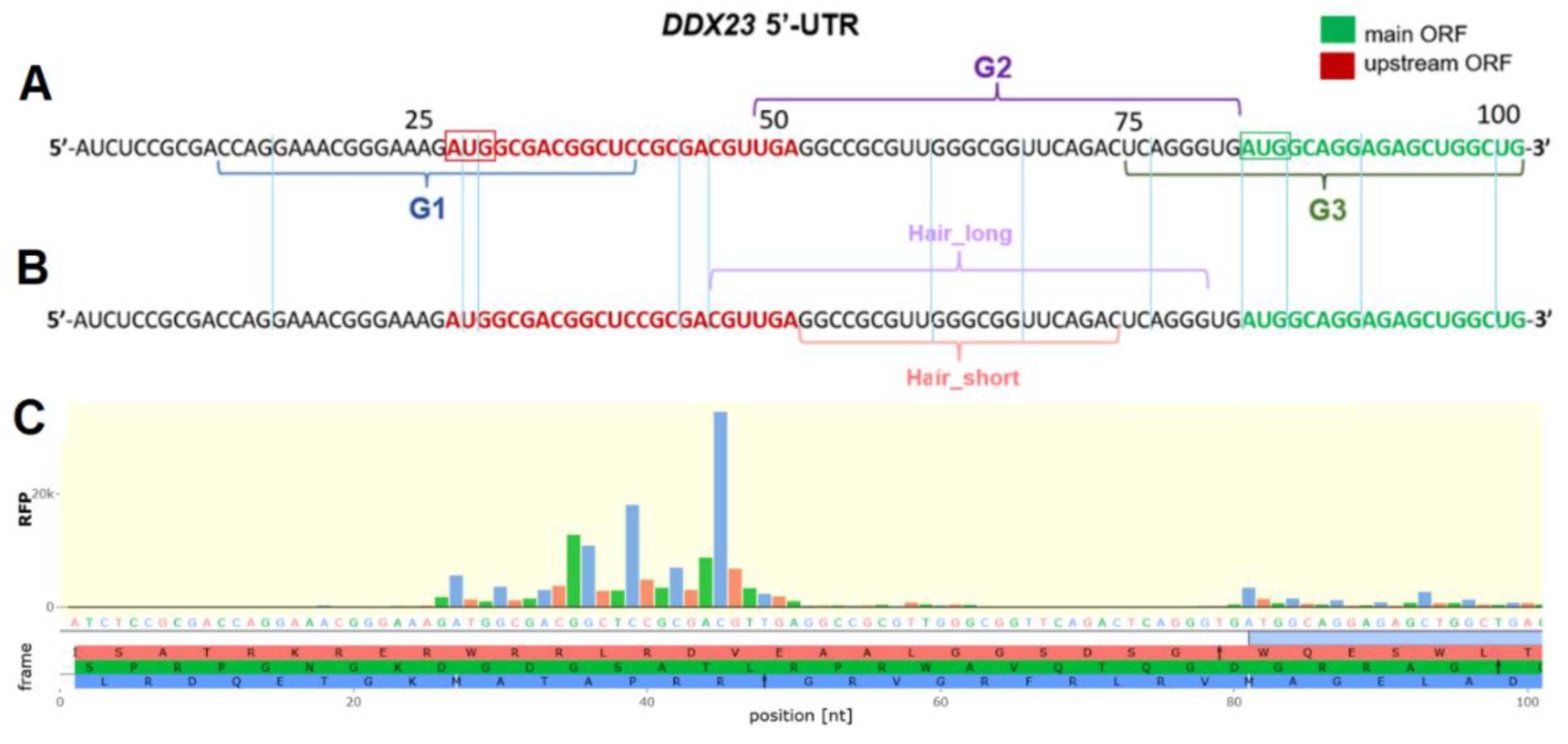
Figure 2.
Сharacterization of putative rG4-forming sequences using fluorescence and CD spectroscopy. ThT-based light-up assay (A). Conditions: K+-buffer, 2 µM RNA, 1 µM ThT, 25 °C, ex. 440 nm, em. 485 nm. CD spectra of putative rG4-forming sequences G1, G2 and G3 and their G-to-U mutants mutG1, mutG2 and mutG3 (B). Conditions: K+-buffer, 2 µM RNA, 15 °C.
Figure 2.
Сharacterization of putative rG4-forming sequences using fluorescence and CD spectroscopy. ThT-based light-up assay (A). Conditions: K+-buffer, 2 µM RNA, 1 µM ThT, 25 °C, ex. 440 nm, em. 485 nm. CD spectra of putative rG4-forming sequences G1, G2 and G3 and their G-to-U mutants mutG1, mutG2 and mutG3 (B). Conditions: K+-buffer, 2 µM RNA, 15 °C.
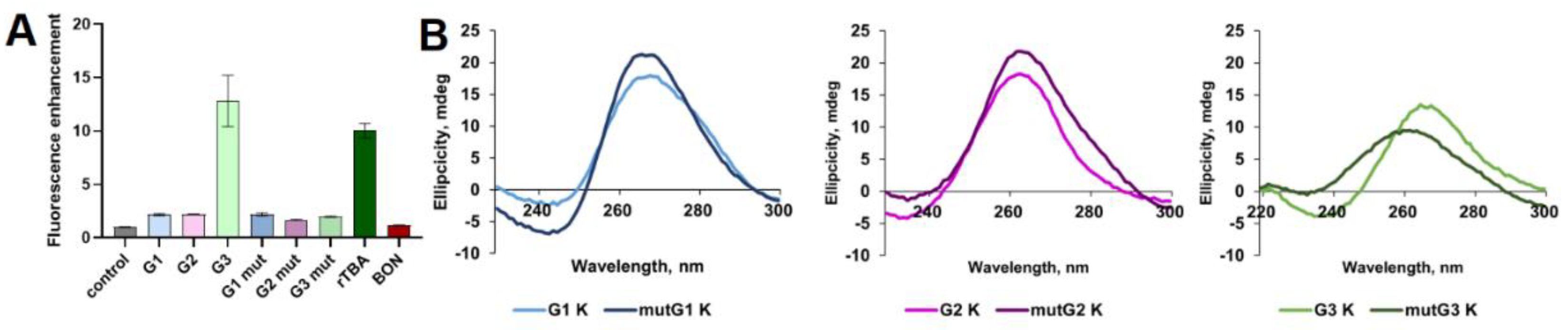
Figure 3.
Effect of PDS treatment on relative luciferase activities for reporter plasmids and CD spectra of putative rG4s. Plasmids encoding native DDX23 mRNA 5’ UTR (WT), DDX23 mRNA 5’ UTR with mutation in the uORF start codon (no uORF), and control short 5’ UTR lacking rG4s-prone motifs and uORF (short UTR) were used (A). Ratios of Fluc and Rluc activities for the reporter constructs tested (B). Normalized ratios of Fluc and Rluc activities to DMSO control (C). Means and standard deviations are shown. CD spectra of putative rG4-forming sequences G1, G2 and G3 in the presence of 1 eq. PDS or DMSO as control (D). Conditions: K+-buffer, 2 µM RNA, 15 °C.
Figure 3.
Effect of PDS treatment on relative luciferase activities for reporter plasmids and CD spectra of putative rG4s. Plasmids encoding native DDX23 mRNA 5’ UTR (WT), DDX23 mRNA 5’ UTR with mutation in the uORF start codon (no uORF), and control short 5’ UTR lacking rG4s-prone motifs and uORF (short UTR) were used (A). Ratios of Fluc and Rluc activities for the reporter constructs tested (B). Normalized ratios of Fluc and Rluc activities to DMSO control (C). Means and standard deviations are shown. CD spectra of putative rG4-forming sequences G1, G2 and G3 in the presence of 1 eq. PDS or DMSO as control (D). Conditions: K+-buffer, 2 µM RNA, 15 °C.
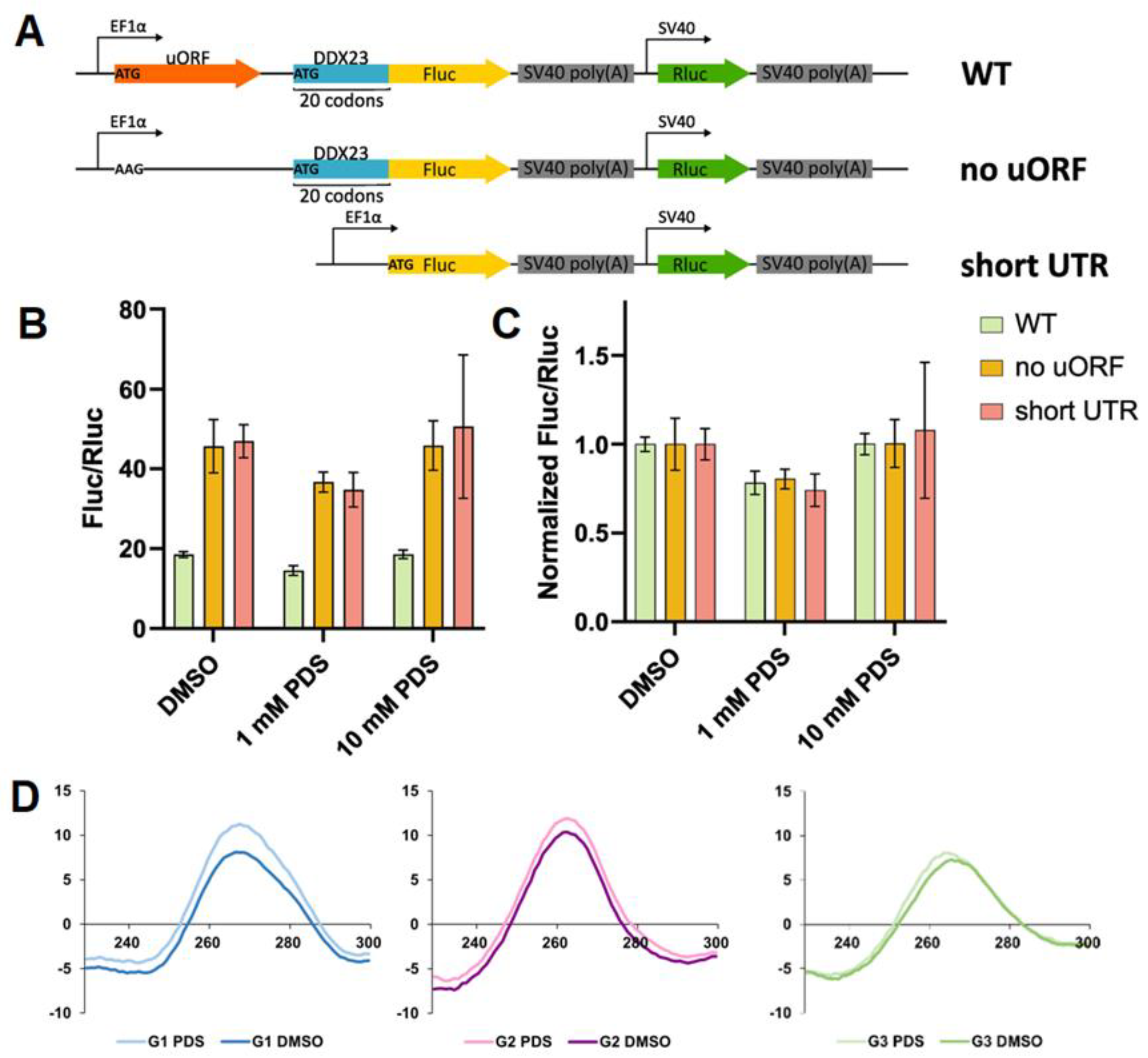
Figure 4.
Spectral characterization of G3 and its rG4- and hairpin-forming mutants h_mutG3 and mutG3, respectively. TDS spectra of G3 (A); UV melting curves registered at 295 nm for G3 (B); CD melting and annealing curves registered at 265 nm for G3 (C); Exchangeable protons region (9.0 to 14.0 ppm) of 1H NMR spectra for G3, h_mutG3 and mutG3 (D); CD melting curves registered at 265 nm for G3, h_mutG3 and mutG3 alone or in the presence of 1 eq. PDS (E). Conditions for A-C, E: K+-buffer, 2 µM RNA.
Figure 4.
Spectral characterization of G3 and its rG4- and hairpin-forming mutants h_mutG3 and mutG3, respectively. TDS spectra of G3 (A); UV melting curves registered at 295 nm for G3 (B); CD melting and annealing curves registered at 265 nm for G3 (C); Exchangeable protons region (9.0 to 14.0 ppm) of 1H NMR spectra for G3, h_mutG3 and mutG3 (D); CD melting curves registered at 265 nm for G3, h_mutG3 and mutG3 alone or in the presence of 1 eq. PDS (E). Conditions for A-C, E: K+-buffer, 2 µM RNA.
Figure 5.
Spectral characterization of hair_long and hair_short. CD spectra of hair_long and hair_short (A). Conditions: K+-buffer, 2 µM RNA; CD melting and annealing curves registered at 265 nm for hair_short (B) and hair_long (C). Conditions: K+-buffer, 2 µM RNA. Exchangeable protons region (9.0 to 14.0 ppm) of 1H NMR spectra for hair_long and hair_short (D).
Figure 5.
Spectral characterization of hair_long and hair_short. CD spectra of hair_long and hair_short (A). Conditions: K+-buffer, 2 µM RNA; CD melting and annealing curves registered at 265 nm for hair_short (B) and hair_long (C). Conditions: K+-buffer, 2 µM RNA. Exchangeable protons region (9.0 to 14.0 ppm) of 1H NMR spectra for hair_long and hair_short (D).
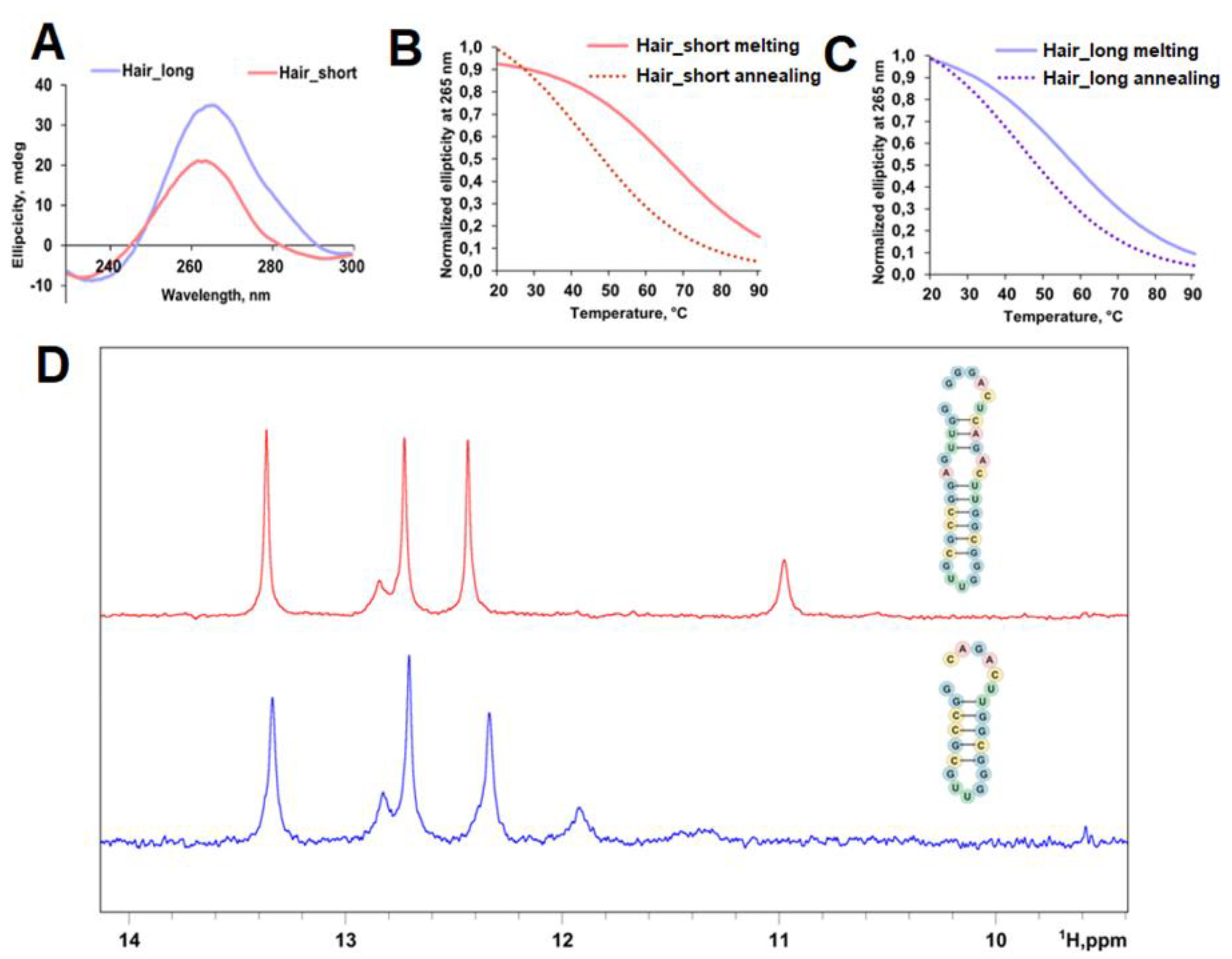
Figure 6.
Design and effects of LNA-ASOs at different concentrations used on translation efficiency of mRNA containing the DDX23 5’ UTR. Designed ASOs targeting structured and/or putative regulatory elements (A). 5’ UTR and a proximal region of the coding sequence of DDX23 mRNA are shown. Schemes of the reporter mRNA used (B). Results for the wild-type 5’ UTR (C), 5’ UTR with point substitution with no uORF (D). Comparison of results for wild-type and no uORF constructs at 100 nM LNA concentrations (E). Means and standard deviations are shown. For accurate normalization, Fluc/Rluc ratios were divided by the median of only mRNA samples for each condition. Ordinary two-way ANOVA with Šidák correction for multiple comparisons were performed. *—adjusted p-value < 0.05, **—adjusted p-value < 0.01, ***—adjusted p-value < 0.001, ****—adjusted p-value < 0.0001.
Figure 6.
Design and effects of LNA-ASOs at different concentrations used on translation efficiency of mRNA containing the DDX23 5’ UTR. Designed ASOs targeting structured and/or putative regulatory elements (A). 5’ UTR and a proximal region of the coding sequence of DDX23 mRNA are shown. Schemes of the reporter mRNA used (B). Results for the wild-type 5’ UTR (C), 5’ UTR with point substitution with no uORF (D). Comparison of results for wild-type and no uORF constructs at 100 nM LNA concentrations (E). Means and standard deviations are shown. For accurate normalization, Fluc/Rluc ratios were divided by the median of only mRNA samples for each condition. Ordinary two-way ANOVA with Šidák correction for multiple comparisons were performed. *—adjusted p-value < 0.05, **—adjusted p-value < 0.01, ***—adjusted p-value < 0.001, ****—adjusted p-value < 0.0001.
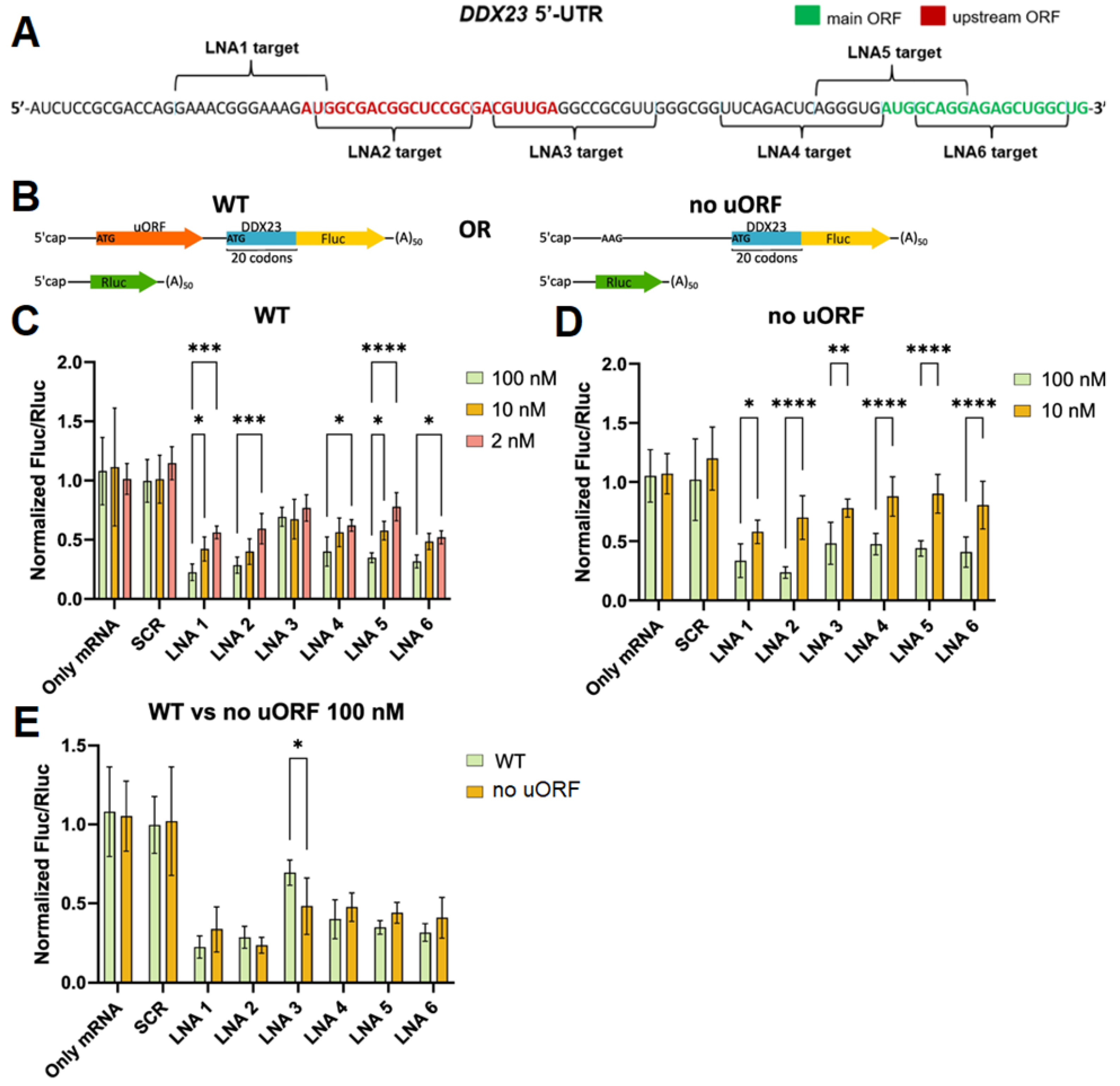

Table 1.
LNA-modified ASOs targeting the uORF and CDS start codons as well as the putative regulatory elements in DDX23 5’ UTR.
Table 1.
LNA-modified ASOs targeting the uORF and CDS start codons as well as the putative regulatory elements in DDX23 5’ UTR.
| Code | Sequence 5’→3’ | Target region |
LNA/RNA duplex Tm, °C |
GC, % |
| LNA1 | CATCTTTCCCGTTC | Upstream region to the uORF start codon | 134 | 50 |
| LNA2 | GCGGAGCCGTCGCCA | Downstream region to the uORF start codon | 98 | 80 |
| LNA3 | AACGCGGCCTCAACG | 5’ segment of hair_short and the uORF stop codon | 82 | 67 |
| LNA4 | CACCCTGAGTCTGAA | 3’ segment of hair_short and 5’ segment of G3 | 120 | 53 |
| LNA5 | TCCTGCCATCACCCT | Central region of G3 and the CDS start codon | 123 | 60 |
| LNA6 | AGCCAGCTCTCCTGC | 3’ segment of G3 | 104 | 67 |
| SCR | CATACGTCTATACGCT | Negative control | - | 44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



