
Submitted:
27 October 2025
Posted:
29 October 2025
You are already at the latest version
Abstract
This review synthesizes recent evidence on the use of the yellow mealworm (Tenebrio molitor) as a lever for circular bioeconomy strategies. We contrast industrial-scale production (standardized, capital-intensive, optimized for throughput) with arid-zone models (resource-constrained, climate-stressed, opportunity for decentralized valorization of local by-products). We examine protein and lipid yields, environmental and techno-economic metrics, supply-chain risks, regulatory bottlenecks, and quality aspects of feeds and foods. We highlight trade-offs between scale, resilience, and local integration, and identify practical priorities: stable substrates, harmonized standards, and risk-managed logistics. The review closes with a roadmap for scaling robust, region-adapted value chains.
Keywords:
mealworm
; Tenebrio molitor
; edible insects
; circular bioeconomy
; sustainable feed
; arid regions
; life-cycle thinking
; techno-economics
; supply chain
; by-products
; waste valorization
1. Introduction
The accelerating global population and growing environmental pressures are reinforcing the search for new food sources, especially sustainable proteins [1]. According to recent life-cycle assessments, mealworm farming has a comparatively low footprint. In Austria, Dreyer et al. estimated that producing 1 kg of edible protein results in 20.4 kg CO₂-eq, 213.7 MJ of non-renewable energy use, and 22.4 m² of land occupation. Compared with organic broiler, impacts were 18-72 % lower across most categories, except for freshwater eutrophication, which was slightly higher [2]. Within this transition, T. molitor stands out as a model species to explore (i) alternative protein production, (ii) integrated approaches linking nutrition and co-product valorisation, and (iii) circular bioeconomy strategies. The arid-zone model is here defined as a decentralized, low-input farming system. It operates under scarce water conditions, strong day/night temperature fluctuations, and relies on irregular local waste streams with minimal capital or energy infrastructure. Among edible insects, the yellow mealworm (Tenebrio molitor, T. molitor) is notable for its balanced nutritional profile, low environmental footprint, and potential to integrate into circular bioeconomy models [3]. Recent research has also highlighted the multi-sectoral value of its biomass. The exuviae are a promising source of high-quality chitosan with applications in food packaging and biomedical materials [4], while its frass and oils are increasingly incorporated into agricultural and cosmetic products [5,6]. Over the past five years, investigations have shifted from basic feasibility studies [7] to advanced applications. New findings include bioactive protein hydrolysates with antioxidant properties [8], functional peptides, and decentralized low-tech farming systems adapted to rural contexts [9]. Tachykinin-related peptides (TRPs) identified in T. molitor are of particular interest for their dual role: while they contribute to allergenic cross-reactivity, they also show potential in modulating immune responses and gut barrier integrity [10]. From a regulatory perspective, T. molitor has progressively gained Novel Food recognition in Europe, while pioneering policies outside the EU have supported its early commercialization. This evolving legal framework underscores the growing institutional interest in insect-based proteins. Technological innovation is also moving the sector forward. For example, machine learning-based monitoring systems are being developed to optimize insect farming and improve quality control at scale [11]. Research so far has revealed both opportunities and limits. Promising avenues include the economic use of co-products, the development of bioactive compounds, and socio-economic gains. However, these coexist with hurdles such as variable frass composition, allergenic risks, and uneven consumer acceptance [5,6] [12].
Ultimately, this review brings together key advances made between 2020 and 2025, with a focus on nutritional quality, functional innovation, regulatory progress, and scalable low-tech strategies. However, these findings also show how T. molitor can help build food systems that are resilient and efficient in their use of resources. This highlights a growing scientific consensus on its potential while also opening perspectives beyond nutrition, including possible applications in health and cosmetic sciences. In addition, this review explicitly considers the relevance of decentralized, low-input production models as potential pathways for resilience in arid regions. This perspective is contrasted with energy-intensive industrial scaling.
2. Regulatory Framework and Legitimacy
This section reviews the current regulatory landscape surrounding T. molitor, including authorization processes, safety assessments, and allergenicity evaluations critical for its integration into food and feed systems.
2.1. Regulatory Landscape and Approvals
Since 2021, the European Union has approved the use of processed insect proteins (PIPs) in poultry and pig feed, marking an important step in the integration of insects into the agri-food chain. It followed several years of safety assessments of insect-based products, particularly T. molitor, carried out by the EFSA Panel on Nutrition and Novel Foods (2021). Two years later, the same EFSA panel validated that both frozen and dried forms of T. molitor larvae are safe for human consumption as a Novel Food. This update reinforced their regulatory status (EFSA Panel on Nutrition and Novel Foods, 2025). Regulatory initiatives are also advancing outside Europe. In Asia, South Korea, through its Ministry of Agriculture (MAFRA), is promoting sustainable approaches that integrate insects into national agricultural policies. Regional consultations supported by FAO Korea Partnership (2022) have fostered South–South cooperation. These initiatives are now shaping harmonized regulations across Asia and Africa. They are also paving the way for the transnational recognition of T. molitor as a food and functional ingredient [13].
In January 2025, the European Commission extended this recognition by authorizing the commercialization of whole T. molitor powder treated with UV irradiation. This decision (Implementing Regulation (EU) 2025/89) facilitates its incorporation into processed foods such as breakfast cereals, dairy products, and snacks [14]. Meanwhile, in Switzerland, the sale of T. molitor for human consumption has been legal since 2017. This early regulation fostered the emergence of innovative start-ups in sustainable food and gave Switzerland a technological advantage over the rest of Europe, according to EFSA (2025).
From a microbiological perspective, several studies have shown that thermal treatment of larvae combined with fasting prior to processing eliminates pathogenic risks. These safety measures have been critical in securing approval for insect-based products in both human and animal nutrition, as reported by EFSA (2025). Future regulatory challenges include the harmonization of global standards, the standardization of processing methods, and the adaptation of legal frameworks to innovative products such as insect protein hydrolysates or functional lipid extracts. Several studies have also highlighted persistent regulatory ambiguities within the European framework. While Regulation (EU) 2015/2283 provides the foundation for approving insects as Novel Food, implementation across Member States remains heterogeneous, particularly regarding the classification of substrates and waste categories permitted for insect rearing. Meijer et al. (2025) emphasize the need for harmonized criteria concerning substrate safety, traceability, and cross-border market authorization. These measures are essential to facilitate industrial scaling of T. molitor production under consistent legal standards [15]. Żuk-Golaszewska et al. note that insect farming is gaining recognition under the Novel Food Regulation (EU) 2015/2283. However, national differences in labelling, substrate policies, and marketing standards still slow commercialization and add to consumer uncertainty [16].
2.2. Safety, Toxicology, and Allergenicity
Malematja et al. studied how heavy metals accumulate in feeder insects. Their work included cadmium (Cd), lead (Pb), mercury (Hg), arsenic (As), copper (Cu), and zinc (Zn). Tenebrio molitor, raised on substrates with controlled contamination, showed measurable uptake of Cd and Pb. The authors noted that “substrate composition directly shaped accumulation levels,” suggesting diet is a key driver [17]. These data serve as baseline values, but they also raise questions. How do larval and adult stages compare? And do current concentrations exceed EFSA or Codex Alimentarius thresholds? These gaps make clear that regulatory benchmarks are still needed. Machona et al. tested powdered T. molitor larvae in Sprague-Dawley rats for 70 days at 300, 1000, and 3000 mg/kg/day. They reported no deaths or clinical signs of toxicity. Body weight, food intake, and organ weight ratios stayed within normal ranges. A small change in liver weight appeared, though it was not statistically significant. “No adverse effects were observed under the tested conditions,” the authors concluded. Yet the study’s limited duration and the lack of genotoxicity or reproductive data underline the need for long-term trials before drawing final safety conclusions [18]. In its opinion on novel foods, the EFSA Panel on Nutrition, Novel Foods and Food Allergens (2021) [19] evaluated frozen and dried whole T. molitor larvae. Their conclusion was cautious: safe for use in products like cereal bars and biscuits, as long as allergenicity is controlled and clearly labelled. The assessment combined composition, processing, stability, and microbiological checks. It matched the toxicological trends reported in [18]. Nevertheless, EFSA acknowledged the absence of post-market epidemiological data, which is a common gap for novel foods. Allergenicity remains a central issue. Emilia et al. examined allergenicity in T. molitor using sera from patients allergic to crustaceans, mollusks, and dust mites. IgE binding to tropomyosin and arginine kinase was detected by ELISA and immunoblot. As they caution, “the presence of sIgE is not equal to allergy.” In plain terms, sensitization does not necessarily predict clinical reactions, and cross-reactivity with crustaceans and mites underpins the EU’s mandatory labelling, while clinical challenge tests remain scarce [20]. Reports of severe reactions, including anaphylaxis observed in some Asian cohorts, further support the need for careful monitoring as T. molitor enters human diets. As a practical perspective, indirect strategies to reduce allergenic exposure deserve attention. In a previous study, Debache (2021) [21] tested a novel feed combining boiled-dried T. molitor granules with date-fruit waste and observed improved broiler chicken performance without adverse effects. Using such intermediate steps, introducing insect proteins first via poultry diets before they enter human food chains, might help lower direct IgE-related risks while maintaining nutritional benefits. This remains a working hypothesis requiring validation. Nevertheless, it highlights a potential bridge between allergenicity research and innovative feed-to-food pathways that merit further exploration.
3. Cost and Environmental Impact
The economic viability of T. molitor production depends largely on two interlinked factors: (1) feedstock cost efficiency and (2) regional resource availability [22]. These parameters shape both the operational strategy and the long-term sustainability of production systems, regardless of scale.
3.1. Environmental Performance and Circularity
In recent years, several investigations have shown that T. molitor farming, when embedded within circular agri-food systems, can cut environmental impacts while still generating outputs of economic value. Among these outputs, frass stands out. Analyses report nitrogen levels between 2.8 % and 4.1 %, phosphorus (P₂O₅) from 1.5 % to 2.4 %, and potassium (K₂O) ranging from 1.4 % to 2.0 %, together with a range of beneficial microorganisms [23]. This composition makes frass a valuable organic fertilizer, able to promote plant growth and support soil microbial communities. To achieve consistent agronomic benefits, nutrient ratios must be adjusted to the needs of each crop, while also meeting EFSA and Codex Alimentarius safety limits for contaminants and heavy metals.
The choice of substrate plays a major role in environmental performance. Life-cycle assessments show that agro-industrial by-products, including malt residual pellets, wheat bran, or corn germ meal, help sustain high larval yields and close nutrient cycles [24,25]. In Lienhard et al., malt residues matched the growth performance of wheat bran while making productive use of a local waste stream [25]. Such substitutions can reduce carbon emissions by up to 50 % and cut water use by 40-60 % compared with conventional fishmeal production [6]. Similarly, an Austrian life-cycle assessment found that while heating drives most of the climate burden, T. molitor still outperformed organic broiler across nearly all impact categories [2]. Production scale is another determining factor. Small, low-tech farms, often relying on manual operation, can function with much lower energy requirements than fully automated industrial plants. These decentralized approaches can be well-suited to areas with limited resources, provided that strict biosecurity and quality standards are maintained to ensure consistent outputs [26,27]. Nevertheless, Table 1 reveals that small-scale’s labour costs (8-12 hrs/kg) negatively impact the Return on Investment (ROI) in high-wage areas, favouring automated systems [27]. Economic analyses suggest that integrating frass upcycling into the production chain can also improve environmental metrics. For example, in Dutch models, such integration was estimated to offset 20 % to 25 % of operational costs while simultaneously reducing the overall ecological footprint [28,29]. Policy incentives like EU tax breaks could help industrial systems overcome nutrient loop challenges while boosting circularity [28]. In this review, the “low-tech” column reflects an arid-zone production model: scarce water, wide day/night temperatures, and irregular substrates. Practical adaptations are simple: passive ventilation, only essential evaporative cooling, and strict substrate screening. These constraints help explain higher labour inputs and the variability seen for growth and frass.
As summarized in Table 1, small-scale systems excel in circularity but face scalability trade-offs, whereas industrial production prioritizes consistency at higher energy costs. This contrast is not only about scale but also geography. Small-scale, low-tech systems often match the realities of arid regions, where energy is costly and water is scarce. Local residues, like downgraded fruits or crop by-products, become the main resource. In these contexts, the “arid-zone model” is less about yield maximization and more about survival and resilience. Industrial farms, by contrast, depend on stable inputs and heavy infrastructure, which are rarely feasible in such environments.
3.2. Economic Feasibility and Production Models
The economic viability of T. molitor production depends largely on two factors: feedstock cost structures and the availability of local resources [22]. Controlled trials have shown that agricultural by-products such as malt pellets and corn germ meal can match the yields obtained with conventional wheat bran [25]. In a separate, unpublished trial conducted in Biskra, Algeria, larvae from the same batch were reared either on downgraded local dates, a readily available by-product in the region, or on assorted vegetable scraps. Both groups completed their development over the same period with no adverse effects observed, and growth rates were comparable, even without measuring protein or lipid contents. Notably, this dual advantage, lower costs and maintained performance, illustrates the potential of waste stream utilization as a driver of sustainable value chains.
In Europe, profitability depends above all on three factors. Feed conversion efficiency is the key driver. Labour productivity also matters but is often underestimated. Finally, the market value of insect proteins adds volatility that producers must manage [28,29,30]. Together, these factors explain why small-scale and industrial systems follow distinct economic paths. Small-scale operations often achieve financial stability sooner. Their reduced capital requirements and localized distribution networks shorten the path to breakeven. In contrast, industrial-scale production relies on substantial upfront investment but can deliver lower unit costs through economies of scale. Interestingly, such outcomes are more likely when policy support and adaptable business models are present [32].
The choice of production model ultimately reflects regional assets and constraints. Regions rich in agro-industrial residues but with limited capital tend to favour decentralized, low-tech approaches. In arid-zone settings, the cost structure shifts from electricity to labour and logistics. Consequently, decentralized hubs can remain viable where wages are low, even if biomass is scarce. In contrast, high-wage economies inevitably favour automated, large-scale systems. This framing helps clarify the arid-zone model as a distinct pathway within the broader spectrum of mealworm production systems. Capital-intensive environments, by contrast, lend themselves to automated vertical integration, though careful energy management becomes essential to control ongoing costs [26]. Climate matters, and in arid contexts later-instar harvests can prioritize protein when cooling is limited. Gentle drying preserves lipids and vitamins. The rule is simple; less energy, tighter process windows, unchanged safety. To put it differently, these low-tech adaptations are not just cost-driven but also context-specific, directly linking production models to territorial resilience. Dutch models suggest that profitability can be reached within three to five years when co-products such as oil and frass are fully valorised [28]
4. Nutritional Quality and Food Innovation Potential
It is now well established, and has been for several years, that T. molitor larvae rank among the richest animal sources of protein, with values typically ranging from 44.2% to 60.2% of dry matter [33,34,35]. What makes their protein particularly valuable is its complete amino acid profile, which includes substantial amounts of lysine (K), leucine (L) and valine (V), similar to what we find in lean meats [33]. These proteins consist of both structural components like actin and myosin, and soluble fractions packed with essential amino acids. The lipid fraction is also of considerable interest, representing between 20.0% and 36.0% of dry matter [33]. In line with many other animal-derived sources, the fatty acid profile is largely shaped by polyunsaturated fats, with omega-6 linoleic acid making up 47-54% of total lipids. By contrast, omega-3 α-linolenic acid remains scarce (1.0-1.8%), which leads to an omega-6 to omega-3 ratio in the range of 25:1 to 35:1 [5,34]. The larvae further provide notable amounts of oleic acid (27-32%) and a saturated fraction (21-26%) dominated by palmitic acid at roughly 15% [33]. It should be noted that processing may influence these qualities. Lipid stability can decline over time, and drying steps may accelerate this effect. Heat during processing can also destroy certain vitamins. From a safety perspective, Hazard Analysis and Critical Control Points (HACCP) monitoring by Öllinger-Zierler et al. showed that properly processed batches maintain consistent microbiological quality. Key figures were total viable counts 3.5-4.2 log CFU/g, no detectable Salmonella or Listeria, Bacillus cereus spores <10² CFU/g, fungal loads around 2.3 log CFU/g (mainly Aspergillus and Penicillium), and no harmful levels of mycotoxins [35]. Recent innovations have successfully incorporated T. molitor into various food products, as highlighted in the comprehensive review by [12]. The applications range from protein-packed snacks to pasta and bakery items. What's particularly clever is how food technologists are overcoming potential sensory challenges, through fine milling, careful flavor masking, and strategic blending with familiar ingredients [12]. In bakery applications, T. molitor larval powder replacing 5-10% of wheat flour increases water absorption (+4-6%) and improves mixing stability. The resulting breads show greater volume and a softer crumb; above ~15% substitution, crumb darkening and roasted notes intensify. A practical mitigation is protease pre-treatment, which improves gas retention and helps maintain texture [36]. An observation repeatedly reported across studies tracking composition over larval development is that later instars can reach up to 56% protein (dry matter), with notably high levels of leucine, lysine, and valine, whereas early instars tend to carry more polyunsaturated fats and later instars accumulate a higher proportion of monounsaturated oleic acid [37]. For producers, this is more than a detail. It means harvest timing can be adapted in practice, either to secure the highest possible protein yield, or to target a specific fatty acid profile depending on the product’s intended use. Climate matters. In arid contexts, later-instar harvests can prioritize protein when cooling is limited. Gentle drying preserves lipids and vitamins. The rule is simple; less energy, tighter process windows, unchanged safety.
5. Applications in Animal Nutrition
The meal and oil derived from T. molitor are increasingly studied for their potential in animal feed, particularly for monogastric species (poultry, pigs, fish). This section explores recent scientific findings regarding the use of T. molitor in these species, highlighting zootechnical, digestive, and health-related benefits. Two subsections are dedicated to the main sectors concerned: poultry and other monogastrics.
5.1 Poultry (Broilers, Quails)
Several authors have reported that incorporating T. molitor into poultry diets delivers clear nutritional benefits [38,39,40]. Studies using inclusion levels as low as 5-10 % have still shown measurable gains in growth and feed efficiency [41,42,43]. In slow-growing chickens, full replacement of soybean meal with T. molitor can reduce feed intake and early growth. Performance, however, converged by ~95 days, with final outcomes comparable to conventional diets [44]. In trials with broilers, Biasato et al. reported an average final body weight of 2.51 kg and a feed conversion ratio of 1.58 when diets combined T. molitor meal and oil. Using both fractions together increased the energy density of the ration while maintaining protein digestibility above 85%. Under low-input production systems, these results were achieved without compromising carcass yield or meat quality, and egg parameters in quails also remained stable. Interestingly, the same feeding strategy supported robust zootechnical performance in resource-limited settings, making it compatible with decentralised micro-farming models. In the same trials, villus height increased by 7-10% while crypt depth stayed stable, a pattern likely linked to improved nutrient uptake. Diets also promoted a richer gut microbiota, with more lactic acid bacteria and fewer Clostridium spp [45]. Overall, these changes support better digestion and resilience, an advantage in low-input, decentralised poultry systems.
From a market perspective, poultry stands out as the most widely consumed and culturally accepted monogastric worldwide. It faces fewer religious restrictions than pork. It also requires less water and infrastructure than aquaculture. All these factors keep poultry in a leading position as the prime target for insect-based feed innovations.
5.2. Other Monogastrics (Pigs, Fish)
In pigs, diets incorporating T. molitor meal achieve protein digestibility rates of 91-99 %, with digestible indispensable amino acid scores (DIAAS, a measure of protein quality based on ideal digestibility of each essential amino acid) in the range of 89-92, values comparable to high-grade animal proteins such as poultry or fish meal [46]. These profiles include approximately 6.5 % lysine and 2.1 % methionine (of crude protein), both key amino acids for post-weaning piglets. Growth trials show that such inclusion maintains weight gain and supports a balanced gut microbiota [39,40,46]. In post-weaning conditions, daily gains around 400-420 g and feed conversion ratios equivalent to those obtained with soybean- or fishmeal-based diets have been reported, with no adverse effects on general health indicators [39]. In arid regions, ensuring that balance means relying on blends from residues that truly exist, not on idealized feeds. That practical choice often decides between nutritional adequacy and deficiency. In aquaculture, T. molitor has been widely tested as a sustainable replacement for fishmeal. In Nile tilapia, both partial and total substitution achieved growth rates and feed efficiency equal to, and sometimes exceeding, those from fishmeal diets, with average gains of 1.2-1.4 g/day and FCR values in the 1.3-1.4 range [6]. Fillet lipid profiles also shifted towards a higher proportion of polyunsaturated fatty acids [6]. In rainbow trout, even full replacement preserved growth performance, reaching FCR values of 1.0-1.1, and reduced liver fat deposition by 12-15 % [39]. As observed in poultry trials conducted by Jiang et al.; Effects of yellow mealworm larvae meal on the growth performance, serum biochemical parameters and caecal metabolome in broiler chickens [40],these insect-based diets may also favour beneficial gut microbiota, increasing lactic acid bacteria counts and reducing potential pathogens. Such effects, achieved in controlled feeding trials, could represent a practical advantage for disease resilience in aquaculture systems.
6. Bioactive Compounds and Human Health Applications
The nutritional quality of T. molitor has been increasingly linked to its bioactive peptide fraction. Recent work identified cryptides (2-20 amino acids) generated by controlled enzymatic hydrolysis of T. molitor proteins and further released during simulated human gastrointestinal digestion. Antioxidant capacity was confirmed via DPPH radical-scavenging (EC₅₀ ≈ 0.30-0.87 mg/mL). In LPS-stimulated human Caco-2 cells, digestion-derived hydrolysates down-regulated pro-inflammatory genes (TNF-α, IFN-γ, IL-6; modest effect on IL-1β) and up-regulated IL-4, with in silico docking suggesting interaction with the TLR4/MD2 complex [47]. On this basis, Hammer et al. further investigated the protein quality of T. molitor, using in vitro digestibility and DIAAS as indicators. The study also compared results with Acheta domesticus as a reference species. Interestingly, cricket proteins also showed digestibility above 90% [46]. For T. molitor, Khanal et al. confirmed similarly high digestibility and DIAAS values, consistent with the data already presented in Section 4 [3].
Recent syntheses reinforce this evidence base: comprehensive reviews describe multiple families of T. molitor-derived peptides (antioxidant, anti-inflammatory, antihypertensive, immunomodulatory) and consolidate analytical workflows (HPLC-MS-MS, targeted bioassays) supporting their health relevance [48,49]. In parallel to enzymatic routes, gentle non-thermal processing can also help. Perez et al. applied pulsed electric fields to defatted T. molitor and increased protein extraction yield while improving foaming capacity and foam stability. The treatment subtly altered protein structure, which likely explains the better techno-functionality for health-oriented formulations [50]. Evidence from process-standardized hydrolysates further supports these findings. An Alcalase-based protocol produced peptide fractions with both antioxidant activity and antimicrobial action, effective against Staphylococcus aureus and Escherichia coli [51]. Time-resolved hydrolysis experiments showed a progressive rise in free amino groups and a parallel increase in radical-scavenging capacity. These results support the antioxidant profile of T. molitor protein hydrolysates under optimized reaction windows [5]. Nevertheless, these findings emerge from a classic preclinical pipeline. In vitro assays show activity at milligram-per-millilitre concentrations, levels unlikely to be reached via diet. Small-animal models, often rodents, provide some in vivo support but are short-term and limited in physiological relevance. Validation then requires pharmacokinetic studies and, ultimately, human clinical trials. Without this translational ladder, claims on health applications remain premature.
Ferrazzano et al. examined the short tripeptide YAN (tyrosine-alanine-asparagine) in a rat model of hypertension [52]. The animals were treated and a fall in blood pressure was observed. Signs of endothelial improvement were also noted, and the effect was linked to inhibition of the angiotensin-converting enzyme (ACE) [52]. In parallel, Kotsou et al. examined peptide fractions obtained from T. molitor hydrolysates. In vitro, ACE-inhibitory activity was measured (IC₅₀ ≈ 0.25 mg/mL). Radical-scavenging activity was also detected, probably related to hydrophobic amino acids [12]. Extending this cardiometabolic perspective, Tan et al. (2022) identified T. molitor-derived heptapeptides, notably LPDQWDWR and APPDGGFWEWGD, with strong affinity for dipeptidyl peptidase-IV (DPP-IV), a key regulator of postprandial glucose homeostasis. Their inhibitory capacity (IC₅₀ ≈ 1.64 mg/mL) suggests potential for modulating glucose metabolism, an effect of particular interest for type 2 diabetes prevention [53]. These experimental findings are consistent with an earlier in silico pipeline that mined 384 T. molitor proteins and prioritized cuticular proteins as precursors of potent DPP-IV inhibitory peptides, thereby guiding targeted hydrolysis and validation strategies [54]. The discovery of such targeted peptides is also being revolutionized by new computational methods. The methodological landscape is evolving. Cross-species pipelines now combine enzymatic hydrolysis with computational tools such as deep-learning bioactivity prediction, molecular docking, and molecular dynamics. Gonzalez-de la Rosa et al. screened the T. molitor peptidome by LC-TIMS-MS/MS and, from an Alcalase hydrolysate (10 % DH), prioritized low-molecular-weight antimicrobial candidates using PeptideRanker filtering (> 0.8) and molecular docking. Top sequences included WLNSKGGF and GFIPYEPFLKKMMA; antifungal scores correlated moderately with hydrophobicity and pI. Experimental validation is still required [55].These approaches were first applied to non-insect proteins [56,57]. They may nevertheless be adapted to T. molitor and used to guide peptide discovery and early preclinical selection.
From methodology to experimental evidence, Brai et al. provide one example. Their study focused on T. molitor proteins and the inhibition of angiotensin-converting enzyme (ACE), a central regulator of blood pressure and an established target in hypertension therapy [58]. Among the detected sequences, the previously described tripeptide YAN had been identified earlier in this section by Ferrazzano et al. in a separate in vitro study, further supporting its ACE-inhibitory potential [52]. While known from other biological materials, it was possibly recovered here for the first time from mealworm proteins. The researchers also reported three further peptides, VVYPWTQ, AWYGANK, and LWDHKV, from the same insect source. Each showed ACE-inhibitory activity of a similar order. This combination of results may point to a broader spectrum of mealworm-derived bioactives relevant to cardiovascular health [58]. In line with these cardiovascular observations, Pessina et al. (2020) carried out an in vivo experiment on spontaneously hypertensive rats, a common model in human hypertension research. After oral intake of defatted T. molitor larval extracts, the animals’ systolic blood pressure dropped. Heart function also improved, with a noticeable rise in ejection fraction and fractional shortening. Interestingly, this was not the only change. The same treatment coincided with higher levels of brain-derived neurotrophic factor (BDNF) and a drop in lipid peroxidation. The authors also observed signs of reduced oxidative stress, along with lower amounts of pro-inflammatory cytokines such as TNF-α and IL-1β [59].
Still within the scope of human health, the incretin hormones GLP-1 and GIP regulate postprandial insulin release but are degraded by the enzyme DPP-4. This enzyme shortens incretin action, reducing their effect on glucose regulation. Berraquero-García et al. (2024) examined T. molitor peptides for their ability to inhibit DPP-4 (IC₅₀ ≈ 1.7 mg/mL). The material was processed through electrospraying and spray-drying to preserve this activity and enable nutraceutical formulation. Both encapsulation methods maintained the inhibitory effect, making them relevant for dietary strategies supporting metabolic function [60].Complementarily, fermenting T. molitor hydrolysates with Lactobacillus plantarum enhanced anti-inflammatory readouts in macrophages and improved gastrointestinal motility in a murine constipation model, suggesting microbiome-coupled routes to increase physiological relevance [61].
Turning to cognitive aspects of human health, recent research has explored potential brain-related benefits. Tran et al. tested a mixed extract from Zophobas atratus and T. molitor in mice model with scopolamine-induced memory impairment. Behavioural assessments, including the Morris water maze and Y-maze, showed marked gains in memory. In this model, the gains were accompanied by modulation of AMPK/SIRT1 and BDNF-Akt/mTOR pathways, which are key for synaptic plasticity, oxidative stress control, and neuronal repair [62]. LC-MS/MS analysis then pointed to two main groups of compounds: glycosides (31 %) and heterocycles (27 %). Both classes are known from other contexts to exert neuroprotective effects, particularly under inflammation or during aging [62]. Methanolic extracts of T. molitor containing phenolics also showed measurable antioxidant activity in DPPH and FRAP assays [63]. This highlights that, beyond peptides, insect-derived metabolites may also contribute to nutritional interest. Dermatological/dermocosmetic lines of evidence are also accumulating: a recent review synthesizes peptide-based anti-ageing, moisturizing and soothing mechanisms relevant to skin care [64]; in vivo, trypsin hydrolysates alleviated atopic dermatitis signs via TLR-MyD88-MAPK modulation [65]; and T. molitor oil showed moisturizing and cytoprotective effects on human keratinocytes consistent with repair-oriented topical uses [66]. Applications concerning animal health, antimicrobial stewardship, and agroecological systems are discussed separately in Section 8, which focuses on biotechnology and circularity.
Key bioactive compounds derived from T. molitor and their associated health effects are summarised in Table 2 for clarity and cross-reference.
7. Low-Tech Scalability & Biorefineries
7.1. Substrate Flexibility and Local Waste Streams
Low-tech T. molitor production offers a surprisingly adaptable solution for resource-limited regions. These resilient insects naturally tailor their growth to local conditions, as work from Shah et al. [22] demonstrated how larval development and body composition shift with dietary changes, essentially making the most of whatever food is available. This remarkable flexibility was also observed by Mahmoud et al. [26]. They reported distinct developmental patterns when switching between cereal byproducts and vegetable residues. In the same line, Greek researchers tested wheat bran, brewery spent grains, olive leaves, and mushroom waste, showing that larvae performed well on bran and spent grains, whereas other substrates gave inconsistent results [67]. To adapt rearing systems to local conditions, researchers have proposed simplified steps. Fondevila et al. demonstrated that tuning the starch-to-fiber ratio can significantly improve larval performance, making even low-quality residues more suitable for growth [68]. López-Gámez et al. observed that when dried vegetables or crop residues were added, the larvae found the diet more palatable. This adjustment also increased protein yield and improved nutrient availability [42]. In another trial, Yakti et al. tested bean and strawberry vegetative wastes. They applied simple pre-treatments such as autoclaving and solid-state fermentation with Trichoderma reesei. Results were mixed. Autoclaved bean residues maintained larval yields and enriched the biomass with minerals like calcium and iron. In contrast, strawberry residues, whether raw or pretreated, reduced larval performance. Still, they improved micronutrient content (e.g., manganese, zinc, iron). In short, beans worked, strawberries less so. The key message is that even modest processing can strongly change both growth outcomes and nutritional profiles [69]. Arid-zone case study, Biskra (Algeria). Larvae were reared on downgraded dates and compared with vegetable scraps. They completed development with similar growth. The trial was small and exploratory. Still, the message is simple. Where water and energy are scarce, local residues can keep T. molitor alive and productive, even without sophisticated equipment.
Finally, resource loops can close directly on-farm. In arid sandy soils, the application of T. molitor frass produced clear effects. Lettuce and tomato biomass increased by 20-28%, linked to higher foliar nitrogen and potassium levels that supported stronger plant growth [70]. Compared with a conventional mineral fertilizer, frass often achieved similar or even superior results. The benefit extended beyond yields, since soil microbial stability was also maintained, confirming the multifunctional role of frass within low-tech rearing systems.
7.2. Frass and Co-Product Valorisation
As previously outlined (see Section 3.1), frass nutrient values vary considerably with substrate and rearing conditions. Zunzunegui et al. reported ≈ 4.5 % N, 2.6 % P, and 2.5 % K, together with calcium (~2.4 %), magnesium (~0.8 %), and trace elements such as Zn, Cu, and Fe. They also noted that residual chitin fragments can “wake up” beneficial soil microbes. In greenhouse tomato trials, the same material provided not only a solid nutrient supply but also a clear biostimulant-like boost [23]. However, Lopes et al. caution that this composition is highly variable, shifting considerably with the original substrate and processing conditions, and point out that “frass” can include insect excreta, uneaten feed, larval cuticles and associated microbes. As a result, results differ across studies, and harmonised analytical protocols are needed [71]. Still, outcomes remain inconsistent. This is not surprising, given the mix of materials involved. Before strong biostimulant claims can be made, reproducibility and standardization must come first. Alternative routes have also been explored. Through solid-state fermentation, Aspergillus oryzae produced protease activities around 350-400 U/g when frass humidity was near ~ 65 %; activity remained stable for several days [72]. In parallel, anaerobic digestion studies reported biogas outputs of 44-668 m³ ton⁻¹ VS and methane yields of 26-502 m³ ton⁻¹ VS, placing them within the range of livestock manures [73]. In that study, performance depended on feedstock mix, operating conditions, and co-digestion strategies. A distinct environmental application comes from He et al. Carbonising T. molitor frass yielded an Fe-loaded biochar catalyst (Fe/FBC; surface area 90.65 m²/g; Fe(III) 6.0 %). In a photo-assisted Fenton-like process, the catalyst removed 67 % of total organic carbon from malachite-green dye in ~ 5 min and remained stable over reuse cycles. The main pathways involved N-demethylation, hydroxylation and chromophore disruption. Life-cycle analysis indicated lower impacts than standard composting, notably for climate change, aquatic toxicity and eutrophication [74].
In parallel, integrated biorefinery concepts underline the complementarity between insect rearing and co-product recovery. Oil fractions, protein hydrolysates and residual frass can be channelled into food, feed or energy loops [75]. Within this framework, protein hydrolysates surpassed 72% protein while demonstrating valuable emulsifying, foaming, and antioxidant capabilities. Importantly, smaller peptide fractions (< 3 kDa) exhibited potent antioxidant effects alongside moderate antimicrobial activity [47]. Similar high protein levels have been reported in other studies. Some hydrolysates even exceeded 70% protein, showing potential for food and nutraceutical applications due to their emulsifying and antioxidant properties [5,22,36]. For example, Chewaka et al. used nuruk extract to obtain very small peptides (< 1 kDa) reaching 71.6% protein, all while keeping their key functional properties [8].
7.3. Systemic Integration and Biorefinery Models
Several technical innovations aim to optimize resource recovery within circular systems. Kröncke et al. developed a zigzag air separator for larval isolation, combined with image-based neural networks for quality control. The same work noted that drying duration and temperature directly affect nutritional profiles. Oil extraction by screw pressing, followed by conversion of the press cake into insect meal using rolling mills, produced a feed comparable to fishmeal [37].
Moruzzo et al. emphasized in their review that T. molitor acts as a central agent in circular food and agriculture systems, turning diverse residues into proteins, lipids, chitosan, or biofertilizers [76]. Despite this potential, structural bottlenecks persist. Substrate logistics, lack of product standards, and consumer acceptability limit expansion. Vodenicharova (2023) added that in Europe, the absence of harmonized supply chain strategies, across pre-sale, on-sale, and post-sale stages, remains a major obstacle [77]. Recent work also points to hybrid biorefineries combining insect co-products with bioenergy production. Anaerobic digestion or biogas generation from frass and side-streams can provide both material and energetic returns, situating T. molitor within broader resource recovery frameworks [75].
8. Agroecological and Biotechnological Synergies (2020-2025): Towards a Circular Bioeconomy
T. molitor is now recognized as a flexible organism positioned between agroecology, biotechnology, and circular economy practices. Studies from the past five years show its use in waste recovery, the design of new biomaterials, and its integration into sustainable food and industrial systems. Unlike Section 6, which focuses on direct human health applications of bioactive compounds, this section emphasizes technological, agro-industrial, and cross-sectoral developments.
8.1. Cross-Sector Innovations and Integrated Applications
Recent work has expanded the upcycling of T. molitor beyond its nutritional potential toward biopolymers, oils, and system-wide applications. High-tech urban models such as vertical farming consolidate production in controlled environments using LEDs, sensors and automation, but raise energy-efficiency constraints [27]. Chitosan has been obtained from discarded larvae and exuviae, with yields close to those reported for marine sources. This insect-derived material shows reduced allergenic risk and a smaller environmental footprint [78,79,80]. One study reported antibacterial effects of insect chitosan against Pseudomonas aeruginosa, pointing to possible agro-industrial and biomedical uses [81]. New technologies are also streamlining protein processing. For example, Perez et al. used pulsed electric fields to boost protein recovery from mealworm flour. This method improved foaming properties without changing its nutritional value. Such non-thermal methods expand the functional applications of insect proteins in food and industrial formulations [50].
The oil fraction of T. molitor offers further technological opportunities. Martínez-Pineda et al. found that the oil contained about 43% oleic acid and 33% linoleic acid. γ-tocopherol and sterols were also identified. The extract showed high oxidative stability [82]. In food applications, South Korean studies [83,84] demonstrated that this oil can be blended with natural waxes to form stable oleogels. These oleogels successfully replaced shortening in bakery products, with no loss of quality. Larvae of T. molitor are able to consume many types of agro-industrial residues. For example, studies mention brewery by-products, cassava peels, date pits, and other side streams [21,42,68]. In addition, dairy by-products such as mozzarella whey and whey permeate have been successfully upcycled, with larvae showing increased protein content (≈ +7 %) and favourable fat composition [85]. On their own, such inputs have almost no commercial value. Yet the larvae manage to turn them into protein- and lipid-rich biomass. This process also significantly helps to lower rearing costs for producers. At present, most of these outputs remain at pilot scale. Oil, peptides or oleogels are produced in hundreds of kilos, rarely in tonnes. The near-term value lies in niche applications and full recovery of co-products, not in replacing bulk commodities. Frass, the coproduct of this activity, has been studied on its own. Zunzunegui et al. highlighted how leftover chitin fragments can stimulate soil microorganisms, which indirectly supports plant growth [23]. Interestingly, Muñoz-Seijas et al. placed frass within a biorefinery perspective. In that context, adding it to anaerobic digesters was linked with higher biogas yields [75].
In addition to organic waste, larvae can also process synthetic materials. This capacity for plastic biodegradation is mediated by their gut microbiota. Metagenomic studies by Mamtimin et al. revealed that T. molitor larvae fed with polystyrene harbor microbial consortia. These communities were dominated by Exiguobacterium and Pseudomonas, both known for polymer decomposition [86]. Nutritional co-substrates such as lysine and methionine further enhance degradation efficiency [87]. Enzymes such as feruloyl esterase-like PETases isolated from larvae confirm the direct enzymatic potential of this system [88]. Yet efficiency remains limited, and degradation rates are slow compared to industrial recycling. The potential for generating secondary microplastics also remains an open question. For now, these efficiency and safety constraints limit the large-scale relevance of this process. These caveats need to be kept in view when considering large-scale relevance. Of note, bioinformatics now plays a role here as well. Recent work on the T. molitor proteome has identified cuticular proteins as promising precursors of bioactive peptides, particularly DPP-IV inhibitors [54]. Although mainly discussed in health contexts, such in silico tools may also be adapted to predict functional traits of insect proteins and, by extension, support agro-industrial applications. In a separate but equally critical line of research, several teams have investigated how plant secondary metabolites interfere with insect physiology. This is a key consideration when using agricultural residues as feed. For instance, larvae of T. molitor exposed to high doses of the mycotoxin zearalenone showed weakened antioxidant defences and even a loss of locomotor activity [89]. A similar outcome was noted when phytochemicals forced T. molitor to reshape its digestive and detoxification enzymes, which required adjustments in its enzymatic toolkit [90]. More recently, Winkiel et al. showed that Solanaceae glycoalkaloids modulate energy metabolism in T. molitor, affecting glycolysis (phosphofructokinase, PFK), the TCA cycle (citrate synthase, CS), and β-oxidation (3-hydroxyacyl-CoA dehydrogenase, HADH) [91]. In a companion study, Winkiel et al. showed disruptions in lipid metabolism, with altered triacylglycerols and a drop in 3-hydroxyacyl-CoA dehydrogenase activity in the fat body [92]. In addition to glycoalkaloids, essential-oil volatiles such as (E)-2-decenal, furfural, 2-undecanone and (E,E)-2,4-decadienal impaired T. molitor reproduction in vivo. They reduced terminal oocyte size, vitellogenin expression and follicular patency, cutting oviposition by ~33–43% with lower hatchability; at 10⁻⁵ M, (E)-2-decenal also lowered female survival to ~39% [93]. In broilers, Biasato et al. found that including 5% Tenebrio molitor (TM) or a 1:1 blend of Hermetia illucens (HI) and T. molitor (TM) improved feed efficiency and carcass weight, whereas 10% HI or a 10% HI-TM blend yielded the weakest outcomes; meat-quality effects were variable [45]. In that study, feed conversion improved and intestinal morphology showed favourable changes [45].Additional work has explored effects on gut health. Interestingly, ingredients derived from T. molitor were shown to modulate the microbiota and to enhance immune responses in monogastric animals [40]. This places insect rearing within a wider perspective of livestock health and welfare, suggesting that it can contribute to One Health approaches.
8.2. Biotechnologies and Innovative Materials
Applications of T. molitor-derived biomolecules extend into material sciences. Chitosan-based packaging films and coatings have demonstrated enhanced barrier properties, antimicrobial activity, and fruit preservation capacities [94,95,96]. These biodegradable solutions represent sustainable alternatives to petrochemical plastics. Microbiota-assisted biodegradation further broadens the portfolio of green biotechnologies. Mixed diets combining plastics and organic substrates modulate gut microbial communities toward polymer-degrading taxa [97], while co-feeding strategies improve larval survival under otherwise suboptimal diets [98]. Biomedical research has also leveraged T. molitor chitosan for advanced materials. Nanofibrils and bioengineered chitin–lignin complexes show compatibility with extracellular matrices, supporting applications in tissue scaffolding, wound healing, and controlled release systems [4,99,100]. These results place insect-derived chitosan as a promising renewable resource for next-generation biomaterials. Notably, insights from non-insect models reinforce this perspective. A recent study on tilapia skin successfully isolated cytoprotective peptides and validated their function in silico, showing protection of ovarian granulosa cells against oxidative stress [56]. Such workflows, combining peptide isolation with computational validation, highlight transferable strategies that could equally enhance the biotechnological exploitation of T. molitor. Agroecological studies also point to synergies between insect-derived polymers and plant systems. For example, insect chitosan has been shown to trigger defence pathways in crops, offering a sustainable alternative to synthetic fungicides [34]. This aligns the use of T. molitor with integrated pest management strategies.
8.3. Modular Biorefineries and Territorial Modeling
Integrated biorefineries represent one of the most promising avenues for scaling T. molitor. In practice, no fraction of T. molitor is wasted. Primary products include protein flour, oils, and chitosan. Secondary outputs cover frass and bioinformatic data. These flows lead to urban fertilization, animal feed, and sustainable agriculture, as shown in Figure 1.
Case studies in France and the Netherlands demonstrate that combining insect farming, peptide extraction, and urban fertilization can achieve profitability within three years [28,75]. Techno-economic models point out that substrate costs (~1 939 €/t DM), capital expenditure (~1 459 €/t DM), and energy use (~693 €/t DM) remain the primary economic bottlenecks [28,101]. These economic bottlenecks are summarized in Figure 2.
Life cycle assessments reveal lower greenhouse gas emissions and water use than conventional animal proteins, but also highlight non-renewable energy demand as a critical impact category [102,103]. This is consistent with an Austrian LCA of mealworm farming, which identified heating as a dominant hotspot while still outperforming organic broiler across most impact categories [2]. Comparative studies with other alternative proteins (e.g., Spirulina) show similar challenges of cost and scalability [104]. This has brought coproduct recycling and territorial integration into focus. Moreover, pilot projects indicate that the use of frass in agro-aquaculture systems supports nutrient recycling and reduces reliance on chemical fertilizers [5]. This demonstrates the capacity of T. molitor coproducts to contribute to climate-resilient territorial models. Beyond insects, predictive workflows developed for agro-industrial by products such as tomato residues have demonstrated the feasibility of identifying bioactive peptides entirely in silico [57]. Transferring such computational approaches to T. molitor could enhance integrated biorefineries, enabling not only the extraction of proteins and lipids but also the conversion of secondary flows in a circular economy framework.
8.4. Challenges and Prospects for Sustainable Industrialization
Scaling T. molitor production faces systemic barriers. Maintaining a steady supply of substrates remains a challenge. Energy efficiency is another unresolved issue, and seasonal variations still complicate production. Nevertheless, LCAs consistently flag non-renewable energy demand, largely from heating, as the dominant hotspot; the Austrian study confirms this even though mealworms still compare favourably to poultry systems [2]. Importantly, in slow-growing strains, full soybean replacement is not performance-neutral during early phases [44]. Plant secondary metabolites, especially Solanaceae glycoalkaloids, can blunt energy metabolism and disrupt lipid handling in T. molitor, likely contributing to the performance variability seen on plant-residue diets [91,92]. Environmental concerns also enter into the picture. One relates to competition with other industries for feedstock. Another is the ecological risk that could occur if insects escape and establish themselves in non-native ecosystems. Social acceptance continues to constrain expansion. Consumer surveys indicate greater acceptance of flours and processed forms than of whole insects [5,46]. These non-economic challenges are illustrated in Figure 3.
Camouflaged or blended ingredients, combined with transparent communication and traceability, appear critical for improving adoption. Regulatory frameworks are progressing unevenly. While EFSA approvals in Europe are expanding [19,105], other regions such as Africa lack harmonized standards despite traditional entomophagy practices [13]. Asia presents a mixed picture, with countries like South Korea moving faster than others [15]. Harmonized safety and labelling protocols will be essential for global trade.
8.5. Perspectives for Innovative Applications
In addition to the domain-specific applications detailed in Table 3, recent years have seen the emergence of transversal bioinformatic workflows and predictive in silico pipelines. These tools facilitate the discovery of novel bioactive peptides in T. molitor. Moreover, they provide transferable methodologies that can be applied to other organisms and by-products. This dual capability reinforces the systemic integration of insect biotechnology into a circular bioeconomy.
Table 3 provides a synthetic overview of these promising cross-sectoral opportunities identified between 2020 and 2025. Note that rows focused solely on human nutraceutical peptides (e.g., antihypertensive agents, DPP-IV inhibitors) have been excluded, as they are discussed in depth in Section 6. It is important to note that while Table 3 effectively highlights sectoral opportunities and functional benefits, it does not convey their environmental footprint. To provide this crucial perspective, Figure 4 contrasts the life cycle impacts of mealworm production with a conventional protein source, using data from recent LCA studies [102,103]. When considered together, Table 3 and Figure 4 offer a comprehensive view of both the industrial potential and the ecological positioning of T. molitor valorisation.
9. Conclusions
T. molitor has emerged as a versatile and promising resource, well suited to address contemporary challenges in sustainable food systems, circular economy, and territorial resilience. Its valorisation, into insect meal, oil, protein hydrolysates, or even as secondary by-products such as frass, opens a wide array of opportunities across agriculture, human and animal nutrition, and green biotechnologies. This circular logic also extends to biomedical and dermatological fields through the exploitation of secondary biomaterials. Chitin nanofibrils extracted from T. molitor, which mimic the structure of the extracellular matrix, have shown potential for tissue regeneration and advanced skin applications [4]. Recent studies confirm the effective integration of T. molitor into poultry diets, even at moderate inclusion rates (5-10%), with measurable improvements in zootechnical performance, digestibility, and meat quality [39,40,45]. Similar effects are observed in aquaculture, where T. molitor proves to be a promising substitute for fishmeal, yielding benefits in growth, liver health, and overall body composition [6,39]. Building on this, enzymatic hydrolysis of T. molitor proteins allows the production of high-value bioactive peptides with validated functional properties, including emulsifying, antioxidant, and sensory attributes [5,8,36,47]. These extracts have already been tested in fortified formulations such as bread, snacks, and dietary supplements [12,36]. Many of the reported bioactive effects remain confined to in vitro experiments or brief rodent trials, with limited physiological relevance. The inhibitory concentrations (IC₅₀) reported for certain bioactive peptides, such as DPP-IV inhibitors, suggest that their potency may be too low to be physiologically relevant in humans. Without pharmacokinetic and bioavailability data, translation into functional foods or nutraceuticals must therefore be considered uncertain until clinical data can be obtained. Regulatory approval in Europe has advanced rapidly (EFSA 2021; 2025), confirming safety and paving the way for commercialization. Yet, well-powered human trials are still missing. In vitro and animal data are promising, but they cannot, on their own, guarantee real benefits. Stability, bioavailability, and metabolic fate are still poorly understood. Addressing these issues is the necessary step if T. molitor is to move from experimental promise to credible nutrition and health applications.
One of the most overlooked issues is yield and feasibility. To give a sense of scale, global olive oil production reaches millions of tonnes each year. By contrast, outputs from T. molitor oil remain in the range of only a few hundred kilos to tonnes at pilot scale. This gap does not erase the promise of mealworm oils or peptides. It does, however, underline that positioning must be realistic, probably niche, not mainstream [1]. Recent pilot work confirms this limitation. Performance depends strongly on the type of agri-by-products used, with some supporting efficient growth and others leading to poor outcomes. At present, the near-term value lies more in niche applications and full valorisation of co-products than in replacing bulk commodities; this perspective helps to keep expectations realistic. In humid regions, abundant vegetation allows composting or manure recycling to sustain soils. By contrast, arid zones face chronic biomass scarcity, making it difficult to maintain fertility. In such contexts, insect farming, using downgraded fruits or wild inedible plants as feed, could deliver both protein and frass, offering one of the few circular options available. This remains largely unexplored and deserves targeted trials. The arid-zone pathway should therefore not be seen as anecdotal but as a systemic model: low-input, decentralized, and tied to local residues, where mealworms can provide resilience where conventional farming fails. Another point concerns contradictions across the literature. Some studies highlight strong frass fertilization effects or potent bioactive peptides, yet others report highly variable results or concentrations too weak to matter physiologically. These tensions are rarely acknowledged, but they are central if we want to move from enthusiasm to reproducibility. Frass composition is a moving target. Reproducibility through standardisation will be the key step before broad deployment. Without this, agronomic claims will remain case-specific. Finally, the experience in Biskra demonstrates that downgraded dates and insect-based granules can sustain poultry production even in harsh, arid conditions [21]. By contrast, frass trials in the literature mostly address plant tolerance to salinity and drought, without linking back to insect [70]. Collectively, this evidence suggests insect farming could serve as a resource in fragile ecosystems, but the evidence is still fragmentary. More direct trials in arid zones are needed before resilience claims can be generalized. Beyond efficiency, arid-zone systems could become a second model: decentralized, low-input, and based on local residues. This approach does not compete with industrial yield but offers territorial resilience where conventional farming fails. From an environmental perspective, T. molitor clearly outperforms beef or pork in greenhouse gas emissions, land use, and water consumption. Yet recent LCAs highlight non-renewable energy demand, driven largely by heating and ventilation in industrial facilities, as a persistent weak point [106]. This creates a core sustainability trade-off: energy-intensive industrial systems deliver volume and consistency, whereas lower-yield low-tech setups use less energy. Ecological risks from escapes remain moderate but warrant continued monitoring. Mirroring these technical issues are socio-economic hurdles. Consumer acceptance may be the most unpredictable bottleneck. A global review shows large regional contrasts: willingness to pay is higher in Asia, Africa, and Latin America, but in Europe it remains weak, shaped by neophobia and cultural barriers [109]. A recent synthesis by Biteau et al. indicates that uptake may remain modest even when products are relabelled and presented in familiar formats [110]. Transparent communication, culinary innovation, and harmonized labelling will likely be decisive for broader adoption. Taken together, the pathway looks promising. But it will require better communication and product design, and at the same time cleaner, more efficient process energy. Overall, these findings position T. molitor as a credible but not yet fully mature component of the circular bioeconomy. Its true value may lie less in isolated products than in systemic integration, where every stream (proteins, oils, chitin and frass) is utilised in interconnected loops. Future research must now shift from proof-of-concept to validation. Standardized methods and well-designed clinical trials are essential. Progress will also rely on cross-disciplinary collaboration, linking biotechnology, agroecology, and social science. This review has provided a transversal synthesis of T. molitor research (2020-2025), bringing together advances in nutrition, bioactive compounds, environmental sustainability, and novel applications. Perhaps more importantly, it has also highlighted the gaps that remain, stressing the urgent need for clinical validation. This review thus moves beyond summary by confronting key literature contradictions and proposing arid-zone integration as a novel, systemic pathway for circularity. It positions T. molitor as a promising model for sustainable food systems within a circular bioeconomy. Still, rigorous standardization will be necessary if this potential is to translate into industrial reality. The path forward now requires a concerted effort to address the identified challenges of bioefficacy, product standardization, energy efficiency, and consumer trust. Success will therefore depend on translating robust science into scalable, socially accepted, and economically viable applications within a truly circular framework. From these findings, a clear roadmap can now be drawn: (i) standardization of frass composition and safety protocols, (ii) well-powered human clinical trials on key bioactive peptides, (iii) pilot-scale LCAs of integrated production systems in arid regions, and (iv) cross-cultural studies on consumer acceptance drivers.
The key message is this. The yellow mealworm can turn even unexpected residues into valuable outputs, but translating promise into practice requires moving beyond enthusiasm toward reproducible science, regulatory clarity, and societal trust.
Author Contributions
The author was solely responsible for conceptualisation, investigation and writing (original draft and review/editing) of this manuscript.
Funding
This research was self-funded by the author and received no external funding from public, commercial, or non-profit sources.
Data Availability Statement
This review relies on published literature and the author’s synthesis of contextual field observations. All underlying data are available in the cited sources.
Acknowledgements
The author used AI-based tools to assist with reference management and style editing. All scientific content, data interpretation and conclusions are the sole responsibility of the author.
Conflicts of Interest
The author declares no competing interests.
References
- D. Despommier, ‘Vertical farming: a holistic approach towards food security’, Front. Sci., vol. 2, Sept. 2024. [CrossRef]
- M. Dreyer et al., ‘Environmental life cycle assessment of yellow mealworm (Tenebrio molitor) production for human consumption in Austria – a comparison of mealworm and broiler as protein source’, Int. J. Life Cycle Assess., vol. 26, no. 11, pp. 2232–2247, Nov. 2021. [CrossRef]
- P. Khanal et al., ‘Yellow mealworms (Tenebrio molitor) as an alternative animal feed source: A comprehensive characterization of nutritional values and the larval gut microbiome’, J. Clean. Prod., vol. 389, p. 136104, Feb. 2023. [CrossRef]
- M. S. Marín-Morales, C. C. Ibarra-Herrera, and M. J. Rivas-Arreola, ‘Obtention and Characterization of Chitosan from Exuviae of Tenebrio molitor and Sphenarium purpurascens’, ACS Omega, vol. 10, no. 16, pp. 17015–17023, Apr. 2025. [CrossRef]
- M. Maciejewska, A. Dąbrowska, and M. Cano-Lamadrid, ‘Sustainable Protein Sources: Functional Analysis of Tenebrio molitor Hydrolysates and Attitudes of Consumers in Poland and Spain Toward Insect-Based Foods’, Foods, vol. 14, no. 2, p. 333, Jan. 2025. [CrossRef]
- M. S. Khan et al., ‘Mealworms (Tenebrio molitor L.) as a Substituent of Protein Source for Fisheries and Aquaculture: A Mini Review: Mealworms as a Substituent of Protein Source’, MARKHOR J. Zool., pp. 19–25, Mar. 2025. [CrossRef]
- Tlak Gajger and S. A. Dar, ‘Plant Allelochemicals as Sources of Insecticides’, Insects, vol. 12, no. 3, p. 189, Feb. 2021. [CrossRef]
- L. S. Chewaka, C. S. Park, Y.-S. Cha, K. T. Desta, and B.-R. Park, ‘Enzymatic Hydrolysis of Tenebrio molitor (Mealworm) Using Nuruk Extract Concentrate and an Evaluation of Its Nutritional, Functional, and Sensory Properties’, Foods, vol. 12, no. 11, p. 2188, May 2023. [CrossRef]
- Montalbán, S. Martínez-Miró, A. Schiavone, J. Madrid, and F. Hernández, ‘Growth Performance, Diet Digestibility, and Chemical Composition of Mealworm (Tenebrio molitor L.) Fed Agricultural By-Products’, Insects, vol. 14, no. 10, p. 824, Oct. 2023. [CrossRef]
- Urbański et al., ‘Tachykinin-related peptides modulate immune-gene expression in the mealworm beetle Tenebrio molitor L.’, Sci. Rep., vol. 12, no. 1, p. 17277, Oct. 2022. [CrossRef]
- P. Majewski, P. Zapotoczny, P. Lampa, R. Burduk, and J. Reiner, ‘Multipurpose monitoring system for edible insect breeding based on machine learning’, Sci. Rep., vol. 12, no. 1, p. 7892, May 2022. [CrossRef]
- K. Kotsou, T. Chatzimitakos, V. Athanasiadis, E. Bozinou, C. G. Athanassiou, and S. I. Lalas, ‘Innovative Applications of Tenebrio molitor Larvae in Food Product Development: A Comprehensive Review’, Foods, vol. 12, no. 23, p. 4223, Nov. 2023. [CrossRef]
- Food and Agriculture Organization Of United Nations, ‘FAO Korea Partnership Newsletter – 1st Quarter 2022, Issue #2’. Food and Agriculture Organization of the United Nations (FAO), 2022. [Online]. Available: https://openknowledge.fao.org/items/66121641-98fb-4877-9e68-2b53b65ffa33.
- AGRINFO, ‘Latest novel food authorisations – January 2025’. COLEAD, Feb. 05, 2025. [Online]. Available: https://agrinfo.eu/book-of-reports/latest-novel-food-authorisations-january-2025/.
- N. Meijer, R. A. Safitri, W. Tao, and E. F. Hoek-Van Den Hil, ‘Review: European Union legislation and regulatory framework for edible insect production – Safety issues’, animal, p. 101468, Feb. 2025. [CrossRef]
- K. Żuk-Gołaszewska, R. Gałęcki, K. Obremski, S. Smetana, S. Figiel, and J. Gołaszewski, ‘Edible Insect Farming in the Context of the EU Regulations and Marketing—An Overview’, Insects, vol. 13, no. 5, p. 446, May 2022. [CrossRef]
- E. Malematja, T. G. Manyelo, N. A. Sebola, S. D. Kolobe, and M. Mabelebele, ‘The accumulation of heavy metals in feeder insects and their impact on animal production’, Sci. Total Environ., vol. 885, p. 163716, Aug. 2023. [CrossRef]
- Machona, M. Mutanga, F. Chidzwondo, and R. Mangoyi, ‘Sub-chronic toxicity determination of powdered Tenebrio molitor larvae as a novel food source’, Toxicol. Rep., vol. 12, pp. 111–116, June 2024. [CrossRef]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA) et al., ‘Safety of frozen and dried formulations from whole yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283 (EFSA 2021)’, EFSA J., vol. 19, no. 8, Aug. 2021. [CrossRef]
- M. Emilia et al., ‘IgE-based analysis of sensitization and cross-reactivity to yellow mealworm and edible insect allergens before their widespread dietary introduction’, Sci. Rep., vol. 15, no. 1, Jan. 2025. [CrossRef]
- K. Debache, ‘Growth performance of novel food based on mixture of boiled-dried granulated Tenebrio molitor larvae and date-fruit waste in broiler chicken farming’, Asian J. Agric., vol. 5, no. 1, Feb. 2021. [CrossRef]
- A. Shah et al., ‘Mealworm (Tenebrio molitor) rearing and growth optimization as a sustainable food source using various larval diets under laboratory conditions’, Entomol. Exp. Appl., vol. 172, no. 9, pp. 827–836, Sept. 2024. [CrossRef]
- Zunzunegui, J. Martín-García, Ó. Santamaría, and J. Poveda, ‘Analysis of yellow mealworm (Tenebrio molitor) frass as a resource for a sustainable agriculture in the current context of insect farming industry growth’, J. Clean. Prod., vol. 460, p. 142608, July 2024. [CrossRef]
- S. Oh and C. Lu, ‘Vertical farming - smart urban agriculture for enhancing resilience and sustainability in food security’, J. Hortic. Sci. Biotechnol., vol. 98, no. 2, pp. 133–140, Mar. 2023. [CrossRef]
- Lienhard, R. Rehorska, B. Pöllinger-Zierler, C. Mayer, M. Grasser, and S. Berner, ‘Future Proteins: Sustainable Diets for Tenebrio molitor Rearing Composed of Food By-Products’, Foods, vol. 12, no. 22, p. 4092, Nov. 2023. [CrossRef]
- M. A. Mahmoud, A. O. Abotaleb, and R. A. Zinhoum, ‘Evaluation of various diets for improved growth, reproductive and nutritional value of the yellow mealworm, Tenebrio molitor L.’, Sci. Rep., vol. 15, no. 1, May 2025. [CrossRef]
- Sowmya, M. Anand, C. Indu Rani, G. Amuthaselvi, and P. Janaki, ‘Recent developments and inventive approaches in vertical farming’, Front. Sustain. Food Syst., vol. 8, Sept. 2024. [CrossRef]
- H. H. Niyonsaba, I. L. Groeneveld, I. Vermeij, J. Höhler, H. J. Van Der Fels-Klerx, and M. P. M. Meuwissen, ‘Profitability of insect production for T. molitor farms in The Netherlands’, J. Insects Food Feed, vol. 10, no. 6, pp. 895–902, Dec. 2023. [CrossRef]
- H. H. Niyonsaba, J. Höhler, J. Kooistra, H. J. Van Der Fels-Klerx, and M. P. M. Meuwissen, ‘Profitability of insect farms’, J. Insects Food Feed, vol. 7, no. 5, pp. 923–934, Aug. 2021. [CrossRef]
- Truzzi et al., ‘Influence of Feeding Substrates on the Presence of Toxic Metals (Cd, Pb, Ni, As, Hg) in Larvae of Tenebrio molitor: Risk Assessment for Human Consumption’, Int. J. Environ. Res. Public. Health, vol. 16, no. 23, p. 4815, Nov. 2019. [CrossRef]
- Noyens, F. Schoeters, M. Van Peer, S. Berrens, S. Goossens, and S. Van Miert, ‘The nutritional profile, mineral content and heavy metal uptake of yellow mealworm reared with supplementation of agricultural sidestreams’, Sci. Rep., vol. 13, no. 1, p. 11604, July 2023. [CrossRef]
- E. Aguilar-Toalá, A. M. Vidal-Limón, and A. M. Liceaga, ‘Advancing Food Security with Farmed Edible Insects: Economic, Social, and Environmental Aspects’, Insects, vol. 16, no. 1, p. 67, Jan. 2025. [CrossRef]
- Kotsou, T. Chatzimitakos, V. Athanasiadis, E. Bozinou, and S. I. Lalas, ‘Exploiting Agri-Food Waste as Feed for Tenebrio molitor Larvae Rearing: A Review’, Foods, vol. 13, no. 7, p. 1027, Mar. 2024. [CrossRef]
- Y. K. Dalmoro, C. H. Franceschi, and C. Stefanello, ‘A Systematic Review and Metanalysis on the Use of Hermetia illucens and Tenebrio molitor in Diets for Poultry’, Vet. Sci., vol. 10, no. 12, p. 702, Dec. 2023. [CrossRef]
- B. Pöllinger-Zierler et al., ‘Tenebrio molitor (Linnaeus, 1758): Microbiological Screening of Feed for a Safe Food Choice’, Foods, vol. 12, no. 11, p. 2139, May 2023. [CrossRef]
- Verni et al., ‘Optimizing Tenebrio molitor powder as ingredient in breadmaking: Impact of enzymatic hydrolysis on dough techno-functional properties and bread quality’, Future Foods, vol. 11, p. 100665, June 2025. [CrossRef]
- Kröncke, S. Wittke, N. Steinmann, and R. Benning, ‘Analysis of the Composition of Different Instars of Tenebrio molitor Larvae using Near-Infrared Reflectance Spectroscopy for Prediction of Amino and Fatty Acid Content’, Insects, vol. 14, no. 4, p. 310, Mar. 2023. [CrossRef]
- J. H. Yoon and C. Kong, ‘Determination of ileal digestibility of tryptophan in tryptophan biomass for broilers using the direct and regression methods’, Anim. Feed Sci. Technol., vol. 304, p. 115732, Oct. 2023. [CrossRef]
- S. Moutinho, A. Oliva-Teles, S. Martínez-Llorens, Ó. Monroig, and H. Peres, ‘Total fishmeal replacement by defattedHermetia illucens larvae meal in diets for gilthead seabream (Sparus aurata) juveniles’, J. Insects Food Feed, vol. 8, no. 12, pp. 1455–1468, Dec. 2022. [CrossRef]
- T. Jiang et al., ‘Effects of yellow mealworm (Tenebrio molitor) larvae meal on the growth performance, serum biochemical parameters and caecal metabolome in broiler chickens’, Ital. J. Anim. Sci., vol. 23, no. 1, pp. 813–823, Dec. 2024. [CrossRef]
- S. Vasilopoulos et al., ‘Growth performance, welfare traits and meat characteristics of broilers fed diets partly replaced with whole Tenebrio molitor larvae’, Anim. Nutr., vol. 13, pp. 90–100, June 2023. [CrossRef]
- G. López-Gámez, R. Del Pino-García, M. A. López-Bascón, and V. Verardo, ‘Improving Tenebrio molitor Growth and Nutritional Value through Vegetable Waste Supplementation’, Foods, vol. 13, no. 4, p. 594, Feb. 2024. [CrossRef]
- Petkov et al., ‘Low-Fat Tenebrio molitor Meal as a Component in the Broiler Diet: Growth Performance and Carcass Composition’, Insects, vol. 15, no. 12, p. 979, Dec. 2024. [CrossRef]
- J. Nieto, J. Plaza, J. Lara, J.-A. Abecia, I. Revilla, and C. Palacios, ‘Performance of Slow-Growing Chickens Fed with Tenebrio molitor Larval Meal as a Full Replacement for Soybean Meal’, Vet. Sci., vol. 9, no. 3, p. 131, Mar. 2022. [CrossRef]
- Biasato et al., ‘Can a mixture of Hermetia illucens and Tenebrio molitor meals be feasible to feed broiler chickens? A focus on bird productive performance, nutrient digestibility, and meat quality’, Poult. Sci., vol. 104, no. 7, p. 105150, July 2025. [CrossRef]
- L. Hammer et al., ‘Mealworm larvae (Tenebrio molitor) and crickets (Acheta domesticus) show high total protein in vitro digestibility and can provide good-to-excellent protein quality as determined by in vitro DIAAS’, Front. Nutr., vol. 10, p. 1150581, July 2023. [CrossRef]
- T. Gonzalez-de La Rosa, S. Montserrat-de La Paz, and F. Rivero-Pino, ‘Production, characterisation, and biological properties of Tenebrio molitor-derived oligopeptides’, Food Chem., vol. 450, p. 139400, Aug. 2024. [CrossRef]
- W. Ma, J. Yang, H. Suo, and J. Song, ‘Tenebrio molitor proteins and peptides: Cutting-edge insights into bioactivity and expanded food applications’, Food Biosci., vol. 68, p. 106369, June 2025. [CrossRef]
- P. Stephan, J. E. D. S. Sarkis, J. S. D. Rosa, and M. L. Cocato, ‘Tenebrio Molitor: Investigating the Scientific Foundations and Proteomic and Peptidomic Potential’, Food Nutr. Sci., vol. 16, no. 04, pp. 427–435, 2025. [CrossRef]
- T. Perez, F. Casanova, L. S. Queiroz, H. O. Petersen, P. J. García-Moreno, and A. H. Feyissa, ‘Protein extraction from yellow mealworm (Tenebrio molitor) assisted by pulsed electric fields: Effect on foaming properties’, LWT, vol. 213, p. 117041, Dec. 2024. [CrossRef]
- C. V. Villanova et al., ‘Bioactive peptides from Tenebrio molitor: physicochemical and antioxidant properties and antimicrobial capacity’, An. Acad. Bras. Cienc., vol. 96, no. suppl 1, p. e20231375, 2024. [CrossRef]
- G. F. Ferrazzano, F. D’Ambrosio, S. Caruso, R. Gatto, and S. Caruso, ‘Bioactive Peptides Derived from Edible Insects: Effects on Human Health and Possible Applications in Dentistry’, Nutrients, vol. 15, no. 21, p. 4611, Oct. 2023. [CrossRef]
- J. Tan et al., ‘Tenebrio molitor Proteins-Derived DPP-4 Inhibitory Peptides: Preparation, Identification, and Molecular Binding Mechanism’, Foods, vol. 11, no. 22, p. 3626, Nov. 2022. [CrossRef]
- Dávalos Terán, K. Imai, I. M. E. Lacroix, V. Fogliano, and C. C. Udenigwe, ‘Bioinformatics of edible yellow mealworm ( Tenebrio molitor ) proteome reveal the cuticular proteins as promising precursors of dipeptidyl peptidase-IV inhibitors’, J. Food Biochem., vol. 44, no. 2, Feb. 2020. [CrossRef]
- T. Gonzalez-de La Rosa, E. Marquez-Paradas, M. J. Leon, S. Montserrat-de La Paz, and F. Rivero-Pino, ‘Exploring Tenebrio molitor as a source of low-molecular-weight antimicrobial peptides using a n in silico approach: correlation of molecular features and molecular docking’, J. Sci. Food Agric., vol. 105, no. 3, pp. 1711–1736, Feb. 2025. [CrossRef]
- H. Yin et al., ‘Isolation, identification and in silico analysis of two novel cytoprotective peptides from tilapia skin against oxidative stress-induced ovarian granulosa cell damage’, J. Funct. Foods, vol. 107, p. 105629, Aug. 2023. [CrossRef]
- Morena, C. Cencini, E. Calzoni, S. Martino, and C. Emiliani, ‘A Novel Workflow for In Silico Prediction of Bioactive Peptides: An Exploration of Solanum lycopersicum By-Products’, Biomolecules, vol. 14, no. 8, p. 930, July 2024. [CrossRef]
- Brai et al., ‘Proteins from Tenebrio molitor: An interesting functional ingredient and a source of ACE inhibitory peptides’, Food Chem., vol. 393, p. 133409, Nov. 2022. [CrossRef]
- Pessina et al., ‘Antihypertensive, cardio- and neuro-protective effects of Tenebrio molitor (Coleoptera: Tenebrionidae) defatted larvae in spontaneously hypertensive rats’, PLOS ONE, vol. 15, no. 5, p. e0233788, May 2020. [CrossRef]
- Berraquero-García, L. Martínez-Sánchez, E. M. Guadix, and P. J. García-Moreno, ‘Encapsulation of Tenebrio molitor Hydrolysate with DPP-IV Inhibitory Activity by Electrospraying and Spray-Drying’, Nanomaterials, vol. 14, no. 10, p. 840, May 2024. [CrossRef]
- H.-S. Jo, D.-B. Song, S.-H. Lee, K.-S. Lee, J. Yang, and S.-M. Hong, ‘Concurrent Hydrolysis–Fermentation of Tenebrio molitor Protein by Lactobacillus plantarum KCCM13068P Attenuates Inflammation in RAW 264.7 Macrophages and Constipation in Loperamide-Induced Mice’, Foods, vol. 14, no. 11, p. 1886, May 2025. [CrossRef]
- B. Tran, H. Lee, M.-G. Ji, L. Ngo Hoang, and S.-J. Lee, ‘The synergistic extract of Zophobas atratus and Tenebrio molitor regulates neuroplasticity and oxidative stress in a scopolamine-induced cognitive impairment model’, Front. Aging Neurosci., vol. 17, Apr. 2025. [CrossRef]
- R. I. Torres-Acosta et al., ‘Insects with Phenolics and Antioxidant Activities to Supplement Mezcal: Tenebrio molitor L.1 and Schistocerca piceifrons Walker2’, Southwest. Entomol., vol. 46, no. 3, Sept. 2021. [CrossRef]
- T. N. Ngoc, J.-Y. Moon, and Y.-C. Lee, ‘Insights into Bioactive Peptides in Cosmetics’, Cosmetics, vol. 10, no. 4, p. 111, Aug. 2023. [CrossRef]
- Fan, N. E. Wedamulla, Y.-J. Choi, Q. Zhang, S. M. Bae, and E.-K. Kim, ‘Tenebrio molitor Larva Trypsin Hydrolysate Ameliorates Atopic Dermatitis in C57BL/6 Mice by Targeting the TLR-Mediated MyD88-Dependent MAPK Signaling Pathway’, Nutrients, vol. 15, no. 1, p. 93, Dec. 2022. [CrossRef]
- R. Verheyen, F. Meersman, I. Noyens, S. Goossens, and S. Van Miert, ‘The Application of Mealworm (Tenebrio molitor) Oil in Cosmetic Formulations’, Eur. J. Lipid Sci. Technol., vol. 125, no. 3, Mar. 2023. [CrossRef]
- Vrontaki, C. Adamaki-Sotiraki, C. I. Rumbos, A. Anastasiadis, and C. G. Athanassiou, ‘Valorization of local agricultural by-products as nutritional substrates for Tenebrio molitor larvae: A sustainable approach to alternative protein production’, Environ. Sci. Pollut. Res., vol. 31, no. 24, pp. 35760–35768, May 2024. [CrossRef]
- G. Fondevila, A. Remiro, and M. Fondevila, ‘Growth performance and chemical composition of tenebrio molitor larvae grown on substrates with different starch to fibre ratios’, Ital. J. Anim. Sci., vol. 23, no. 1, pp. 887–894, Dec. 2024. [CrossRef]
- W. Yakti et al., ‘Utilising common bean and strawberry vegetative wastes in yellow mealworm (Tenebrio molitor) substrates: effects of pre-treatment on growth and composition’, Sci. Rep., vol. 15, no. 1, Mar. 2025. [CrossRef]
- Verardi et al., ‘Tenebrio molitor Frass: A Cutting-Edge Biofertilizer for Sustainable Agriculture and Advanced Adsorbent Precursor for Environmental Remediation’, Agronomy, vol. 15, no. 3, p. 758, Mar. 2025. [CrossRef]
- G. Lopes et al., ‘BugBook: Critical considerations for evaluating and applying insect frass’, J. Insects Food Feed, pp. 1–28, June 2025. [CrossRef]
- Muñoz-Seijas, H. Fernandes, D. Outeiriño, M. G. Morán-Aguilar, J. M. Domínguez, and J. M. Salgado, ‘Potential use of frass from edible insect Tenebrio molitor for proteases production by solid-state fermentation’, Food Bioprod. Process., vol. 144, pp. 146–155, Mar. 2024. [CrossRef]
- D. Magro, D. Lovarelli, J. Bacenetti, and M. Guarino, ‘The potential of insect frass for sustainable biogas and biomethane production: A review’, Bioresour. Technol., vol. 412, p. 131384, Nov. 2024. [CrossRef]
- L. He et al., ‘Fabrication and environmental assessment of photo-assisted Fenton-like Fe/FBC catalyst utilizing mealworm frass waste’, J. Clean. Prod., vol. 256, p. 120259, May 2020. [CrossRef]
- N. Muñoz-Seijas, H. Fernandes, J. M. Domínguez, and J. M. Salgado, ‘Recent Advances in Biorefinery of Tenebrio molitor Adopting Green Technologies’, Food Bioprocess Technol., vol. 18, no. 2, pp. 1061–1078, Feb. 2025. [CrossRef]
- R. Moruzzo, F. Riccioli, S. Espinosa Diaz, C. Secci, G. Poli, and S. Mancini, ‘Mealworm (Tenebrio molitor): Potential and Challenges to Promote Circular Economy’, Animals, vol. 11, no. 9, p. 2568, Aug. 2021. [CrossRef]
- Maria Vodenicharova, ‘Supply Chain Challenges of Insect Protein’, Nanotechnol. Percept., vol. 20, no. S11 (Special Issue), pp. 1570–1582, 2024.
- M. Azzi, S. Elkadaoui, J. Zim, J. Desbrieres, Y. El Hachimi, and A. Tolaimate, ‘Tenebrio Molitor breeding rejects as a high source of pure chitin and chitosan: Role of the processes, influence of the life cycle stages and comparison with Hermetia illucens’, Int. J. Biol. Macromol., vol. 277, p. 134475, Oct. 2024. [CrossRef]
- Terkula Iber, N. Azman Kasan, D. Torsabo, and J. Wese Omuwa, ‘A Review of Various Sources of Chitin and Chitosan in Nature’, J. Renew. Mater., vol. 10, no. 4, pp. 1097–1123, 2022. [CrossRef]
- Izadi, H. Asadi, and M. Bemani, ‘Chitin: a comparison between its main sources’, Front. Mater., vol. 12, Mar. 2025. [CrossRef]
- Nafary, S. A. Mousavi Nezhad, and S. Jalili, ‘Extraction and Characterization of Chitin and Chitosan from Tenebrio Molitor Beetles and Investigation of Its Antibacterial Effect Against Pseudomonas aeruginosa’, Adv. Biomed. Res., vol. 12, no. 1, Apr. 2023. [CrossRef]
- M. Martínez-Pineda et al., ‘Exploring the Potential of Yellow Mealworm (Tenebrio molitor) Oil as a Nutraceutical Ingredient’, Foods, vol. 13, no. 23, p. 3867, Nov. 2024. [CrossRef]
- Kim and I. Oh, ‘The Characteristic of Insect Oil for a Potential Component of Oleogel and Its Application as a Solid Fat Replacer in Cookies’, Gels, vol. 8, no. 6, p. 355, June 2022. [CrossRef]
- S. Jeong and I. Oh, ‘Characterization of mixed-component oleogels: Beeswax and glycerol monostearate interactions towards Tenebrio Molitor larvae oil’, Curr. Res. Food Sci., vol. 8, p. 100689, 2024. [CrossRef]
- Brai et al., ‘Upcycling Milk Industry Byproducts into Tenebrio molitor Larvae: Investigation on Fat, Protein, and Sugar Composition’, Foods, vol. 13, no. 21, p. 3450, Oct. 2024. [CrossRef]
- T. Mamtimin et al., ‘Gut microbiome of mealworms (Tenebrio molitor Larvae) show similar responses to polystyrene and corn straw diets’, Microbiome, vol. 11, no. 1, May 2023. [CrossRef]
- X. Wang et al., ‘Enhancing Plastic Decomposition in Mealworms (Tenebrio molitor): The Role of Nutritional Amino Acids and Water’, Adv. Energy Sustain. Res., vol. 6, no. 6, June 2025. [CrossRef]
- T. Mamtimin et al., ‘Novel Feruloyl Esterase for the Degradation of Polyethylene Terephthalate (PET) Screened from the Gut Microbiome of Plastic-Degrading Mealworms (Tenebrio Molitor Larvae)’, Environ. Sci. Technol., vol. 58, no. 40, pp. 17717–17731, Oct. 2024. [CrossRef]
- M. Janković-Tomanić, B. Petković, J. S. Vranković, and V. Perić-Mataruga, ‘Effects of high doses of zearalenone on some antioxidant enzymes and locomotion of Tenebrio molitor larvae (Coleoptera: Tenebrionidae)’, J. Insect Sci., vol. 24, no. 3, May 2024. [CrossRef]
- M. Chamani et al., ‘From Digestion to Detoxification: Exploring Plant Metabolite Impacts on Insect Enzyme Systems for Enhanced Pest Control’, Insects, vol. 16, no. 4, p. 392, Apr. 2025. [CrossRef]
- M. J. Winkiel, S. Chowański, K. Walkowiak-Nowicka, M. Gołębiowski, and M. Słocińska, ‘A tomato a day keeps the beetle away – the impact of Solanaceae glycoalkaloids on energy management in the mealworm Tenebrio molitor’, Environ. Sci. Pollut. Res., vol. 31, no. 48, pp. 58581–58598, Sept. 2024. [CrossRef]
- M. J. Winkiel, S. Chowański, M. Gołębiowski, S. A. Bufo, and M. Słocińska, ‘Solanaceae Glycoalkaloids Disturb Lipid Metabolism in the Tenebrio molitor Beetle’, Metabolites, vol. 13, no. 12, p. 1179, Nov. 2023. [CrossRef]
- Walkowiak-Nowicka, J. Mirek, S. Chowański, R. Sobkowiak, and M. Słocińska, ‘Plant secondary metabolites as potential bioinsecticides? Study of the effects of plant-derived volatile organic compounds on the reproduction and behaviour of the pest beetle Tenebrio molitor’, Ecotoxicol. Environ. Saf., vol. 257, p. 114951, June 2023. [CrossRef]
- S. Mwita et al., ‘Chitosan Extracted from the Biomass of Tenebrio molitor Larvae as a Sustainable Packaging Film’, Materials, vol. 17, no. 15, p. 3670, July 2024. [CrossRef]
- M. Liu et al., ‘Preparation of chitosan/Tenebrio molitor larvae protein/curcumin active packaging film and its application in blueberry preservation’, Int. J. Biol. Macromol., vol. 275, p. 133675, Aug. 2024. [CrossRef]
- M. Benashvili et al., ‘Effect of mealworm (Tenebrio molitor L.) chitosan coating on the postharvest qualities of strawberries’, Postharvest Biol. Technol., vol. 228, p. 113657, Oct. 2025. [CrossRef]
- Ilijin et al., ‘The impact of co-fed plastic diet on Tenebrio molitor gut bacterial community structure’, Sci. Rep., vol. 15, no. 1, July 2025. [CrossRef]
- X. Wang and T. Tang, ‘Effects of Polystyrene Diet on the Growth and Development of Tenebrio molitor’, Toxics, vol. 10, no. 10, p. 608, Oct. 2022. [CrossRef]
- Morganti, G. Morganti, and M.-B. Coltelli, ‘Natural Polymers and Cosmeceuticals for a Healthy and Circular Life: The Examples of Chitin, Chitosan, and Lignin’, Cosmetics, vol. 10, no. 2, p. 42, Feb. 2023. [CrossRef]
- Mersmann, V. G. L. Souza, and A. L. Fernando, ‘Green Processes for Chitin and Chitosan Production from Insects: Current State, Challenges, and Opportunities’, Polymers, vol. 17, no. 9, p. 1185, Apr. 2025. [CrossRef]
- Leipertz, H. Hogeveen, and H. W. Saatkamp, ‘Economic supply chain modelling of industrial insect production in the Netherlands’, J. Insects Food Feed, vol. 10, no. 8, pp. 1361–1385, Mar. 2024. [CrossRef]
- G. Vinci, S. A. Prencipe, L. Masiello, and M. G. Zaki, ‘The Application of Life Cycle Assessment to Evaluate the Environmental Impacts of Edible Insects as a Protein Source’, Earth, vol. 3, no. 3, pp. 925–938, Aug. 2022. [CrossRef]
- S. Modahl and A. Brekke, ‘Environmental performance of insect protein: a case of LCA results for fish feed produced in Norway’, SN Appl. Sci., vol. 4, no. 6, June 2022. [CrossRef]
- S. Villaró-Cos, J. L. Guzmán Sánchez, G. Acién, and T. Lafarga, ‘Research trends and current requirements and challenges in the industrial production of spirulina as a food source’, Trends Food Sci. Technol., vol. 143, p. 104280, Jan. 2024. [CrossRef]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA) et al., ‘Safety of frozen and dried forms of whole yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283 (EFSA 2025)’, EFSA J., vol. 23, no. 1, Jan. 2025. [CrossRef]
- Y. Malila et al., ‘Current challenges of alternative proteins as future foods’, Npj Sci. Food, vol. 8, no. 1, p. 53, Aug. 2024. [CrossRef]
- Foughar, M. Arrobas, and M. Â. Rodrigues, ‘Mealworm Larvae Frass Exhibits a Plant Biostimulant Effect on Lettuce, Boosting Productivity beyond Just Nutrient Release or Improved Soil Properties’, Horticulturae, vol. 10, no. 7, p. 711, July 2024. [CrossRef]
- V. Ninkuu et al., ‘Phenylpropanoids metabolism: recent insight into stress tolerance and plant development cues’, Front. Plant Sci., vol. 16, June 2025. [CrossRef]
- Z. Abro et al., ‘Global review of consumer preferences and willingness to pay for edible insects and derived products’, Glob. Food Secur., vol. 44, p. 100834, Mar. 2025. [CrossRef]
- C. Biteau, T. Bry-Chevalier, D. Crummett, R. Ryba, and M. St. Jules, ‘Beyond the buzz: insect-based foods are unlikely to significantly reduce meat consumption’, Npj Sustain. Agric., vol. 3, no. 1, p. 35, June 2025. [CrossRef]
Figure 1.
Integrated modular biorefinery for T. molitor. Center: rearing and life cycle. Ring 1: primary outputs (protein flour, oils, exuviae). Ring 2: intermediate valorisation (fertiliser, biogas, oleogels, chitosan, in-silico data). Outer nodes: territorial uses (urban fertilisation, animal feed, field crops, packaging/materials and biomedical). Arrows indicate transformation or use; no reverse flows are implied.
Figure 1.
Integrated modular biorefinery for T. molitor. Center: rearing and life cycle. Ring 1: primary outputs (protein flour, oils, exuviae). Ring 2: intermediate valorisation (fertiliser, biogas, oleogels, chitosan, in-silico data). Outer nodes: territorial uses (urban fertilisation, animal feed, field crops, packaging/materials and biomedical). Arrows indicate transformation or use; no reverse flows are implied.
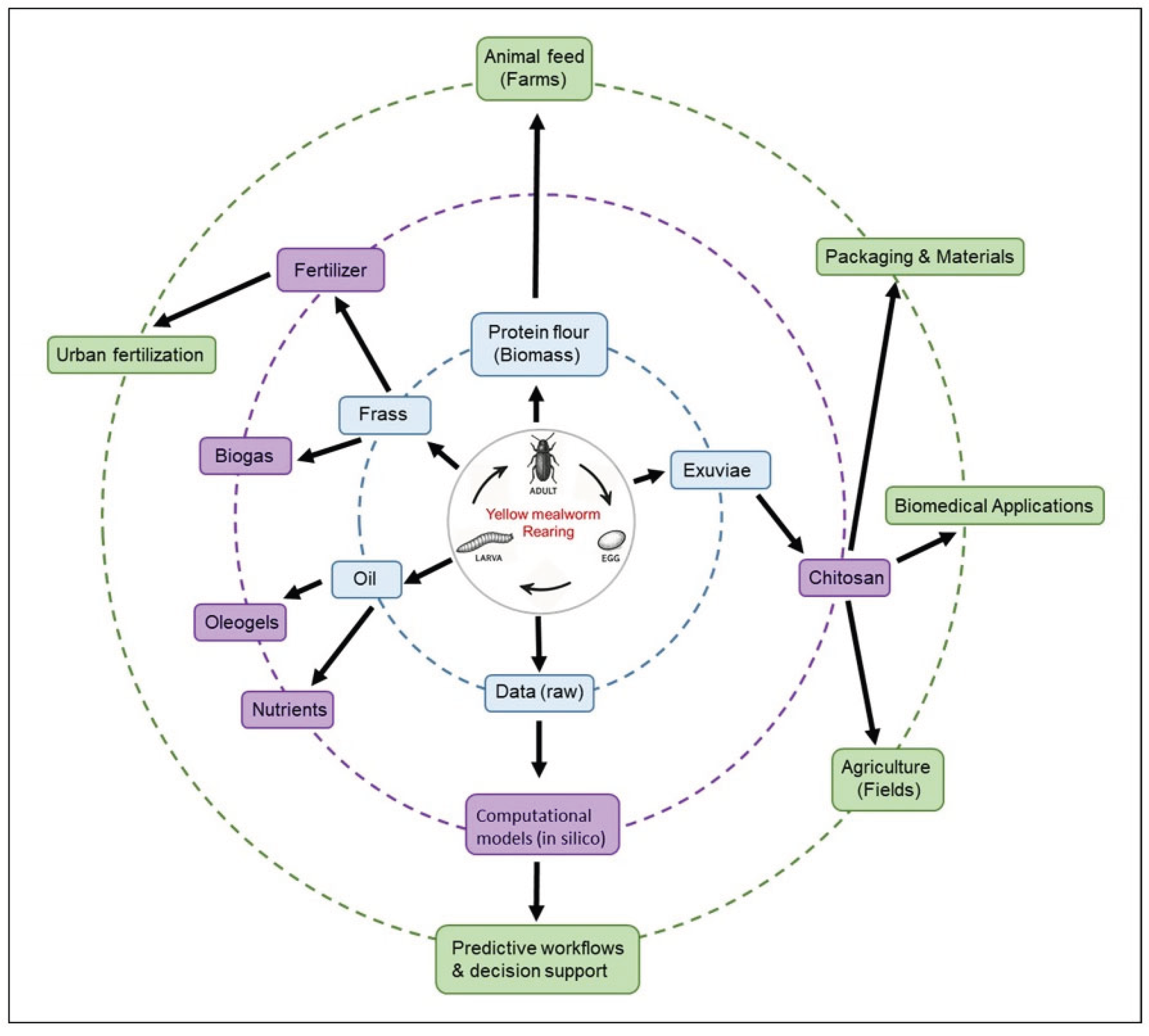
Figure 2.
Bottlenecks for scaling T. molitor biorefineries (€/t DM). Substrate cost dominates, followed by capital expenditure and energy use.
Figure 2.
Bottlenecks for scaling T. molitor biorefineries (€/t DM). Substrate cost dominates, followed by capital expenditure and energy use.
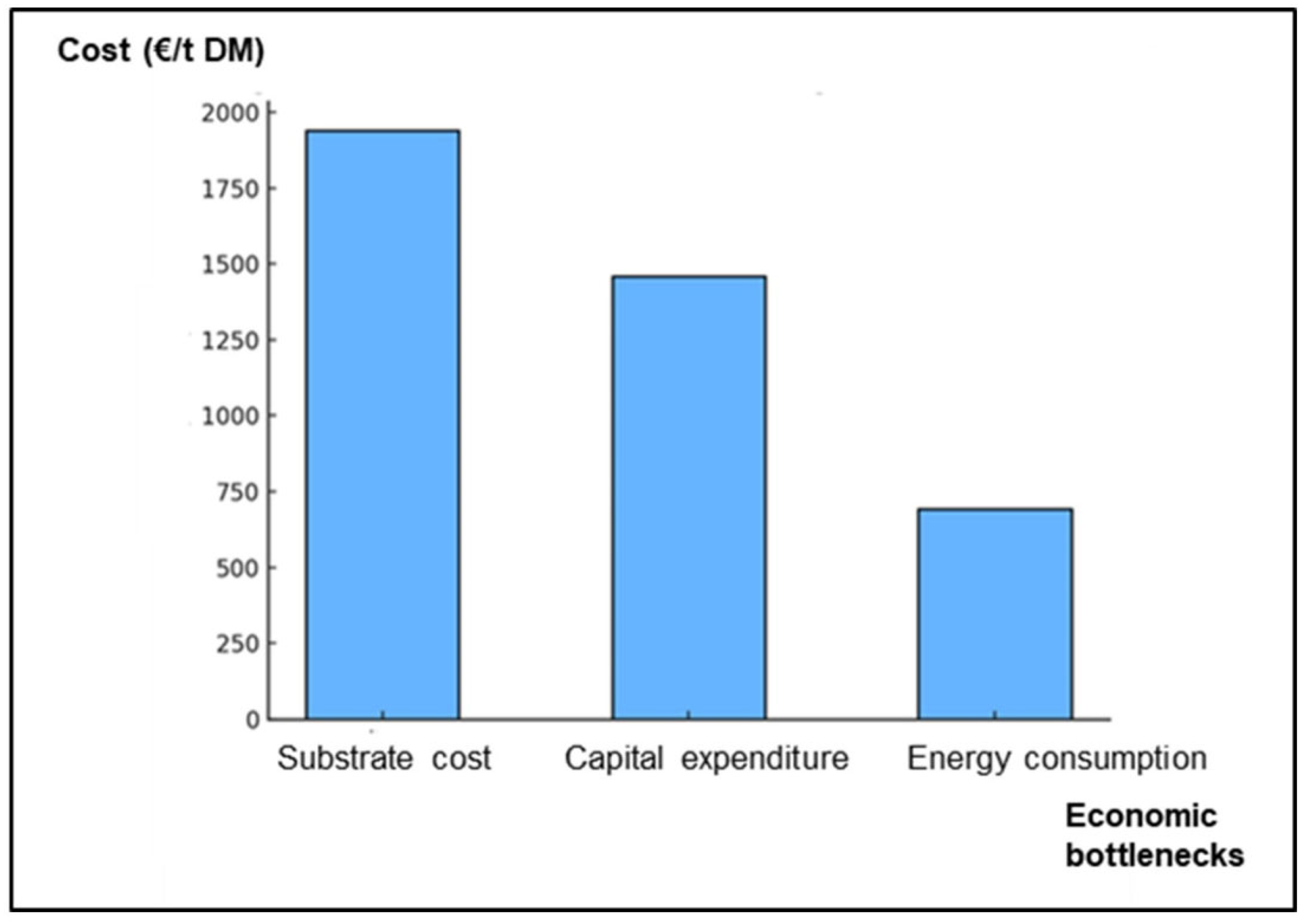
Figure 3.
Non-economic bottlenecks expressed as qualitative scores (1-5). Social acceptance is a strong barrier, while ecological risk is of moderate concern. These scores are indicative, based on consumer surveys [5,46] and ecological risk assessments, not quantitative.
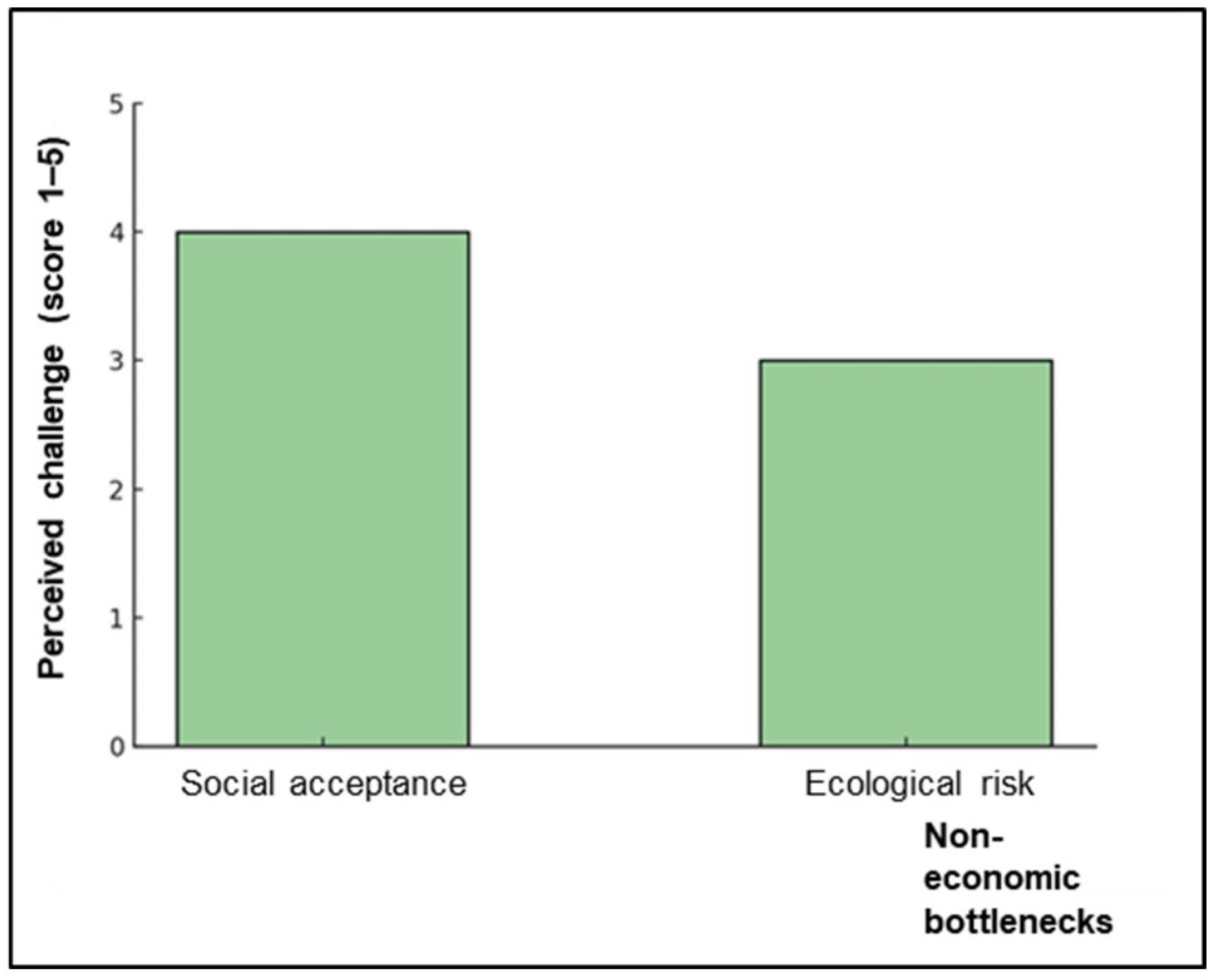
Figure 4.
Comparative environmental performance of T. molitor and beef (illustrative). Radar diagram based on life cycle assessment (LCA) results. Axes cover greenhouse gas emissions, water use, energy demand, and land use. T. molitor shows lower impacts in GHG emissions, water, and land use, while energy demand remains a relative weakness. Scores are normalized (1 = highest impact, 5 = lowest impact), so higher values indicate better environmental performance. Values are indicative, derived from published LCA studies. Note: Data adapted from recent LCA studies, including Malila et al. (2024) [106] and Modahl & Brekke (2022) [103].
Figure 4.
Comparative environmental performance of T. molitor and beef (illustrative). Radar diagram based on life cycle assessment (LCA) results. Axes cover greenhouse gas emissions, water use, energy demand, and land use. T. molitor shows lower impacts in GHG emissions, water, and land use, while energy demand remains a relative weakness. Scores are normalized (1 = highest impact, 5 = lowest impact), so higher values indicate better environmental performance. Values are indicative, derived from published LCA studies. Note: Data adapted from recent LCA studies, including Malila et al. (2024) [106] and Modahl & Brekke (2022) [103].
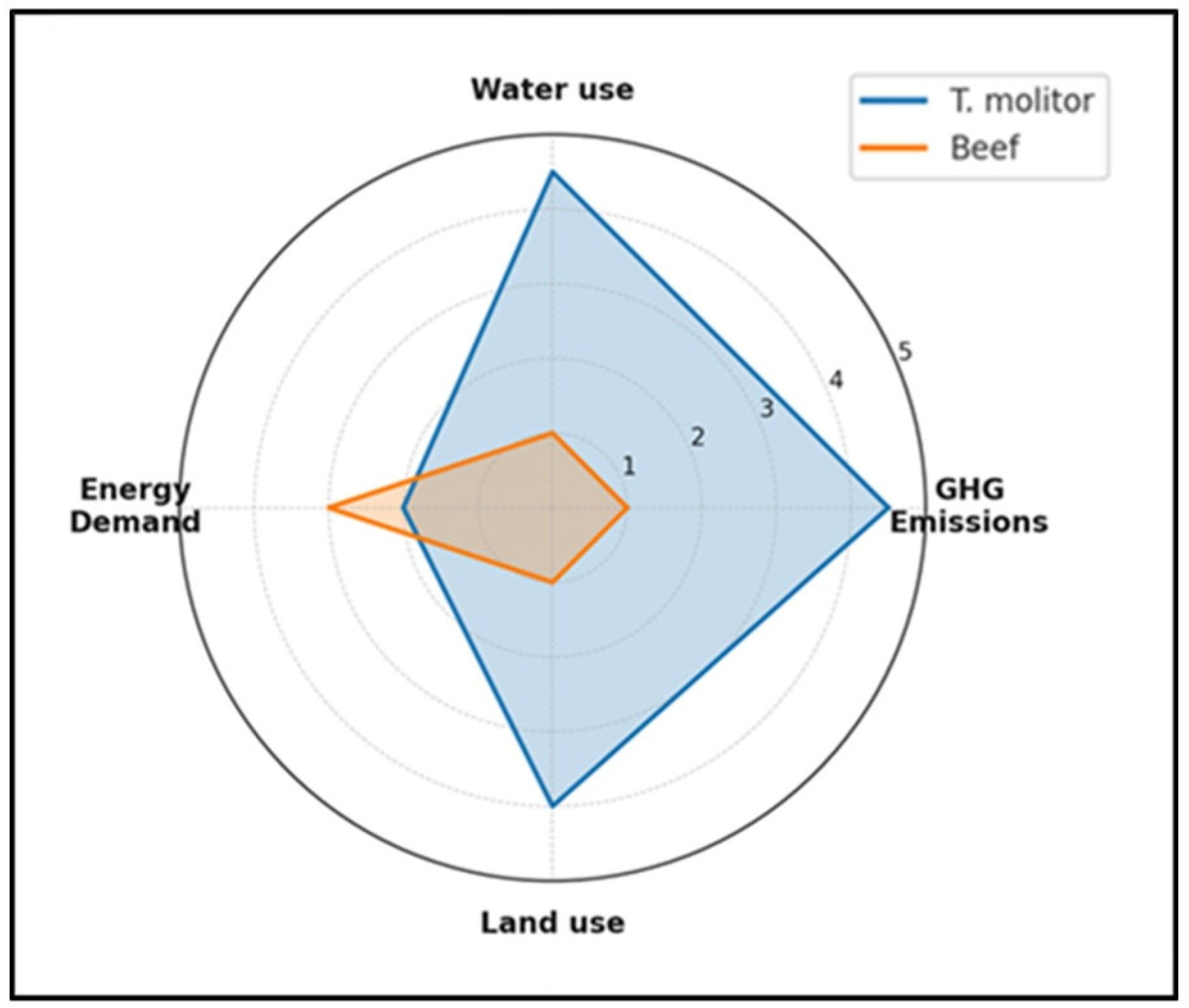

Table 1.
Comparative sustainability of T. molitor production systems, contrasting small-scale/low-tech with industrial/large-scale approaches.
Table 1.
Comparative sustainability of T. molitor production systems, contrasting small-scale/low-tech with industrial/large-scale approaches.
| Parameter | Small-Scale/Low-Tech | Industrial/Large-Scale | Key Implications |
| Substrate | Local by-products (high variability) [30,31] | Standardized diets (consistent nutrients) [31] | Small farms adapt to waste but face contamination risks. |
| Energy Use | 15–20 kWh/kg (passive systems)[25] | 25–30 kWh/kg (HVAC + automation)[26] | Industrial cuts labor costs but increases energy demand. |
| Labor | 8–12 hrs/kg (manual processes)[26] | 1–2 hrs/kg (automated)[26] | Critical for ROI in high-wage regions [26]. |
| Frass Quality | Variable NPK; occasional As exceedance [30] | Uniform NPK; EFSA-compliant [30] | Small-scale requires blending (e.g., rice hulls [31]). |
| Circularity | High (local waste recycling)[25] | Moderate (logistics constraints) [28] | Policy incentives (e.g., EU tax breaks) could boost industrial circularity. |
Note: Data compiled from multiple sources, including Lienhard et al. [25], Mahmoud et al. [26], Khan et al. [6], Niyonsaba et al. [28]. Energy estimates exclude transport emissions for decentralized systems. The low-tech column is particularly relevant for arid-zone conditions, where energy is scarce and residues are the main resource.
Table 2.
Bioactive compounds from T. molitor and their reported health effects. AA = amino acids; YAN = Tyrosine-Alanine-Asparagine; DPP-IV = dipeptidyl peptidase IV; GI = gastrointestinal; TLR = Toll-like receptor; MAPK = mitogen-activated protein kinase.
Table 2.
Bioactive compounds from T. molitor and their reported health effects. AA = amino acids; YAN = Tyrosine-Alanine-Asparagine; DPP-IV = dipeptidyl peptidase IV; GI = gastrointestinal; TLR = Toll-like receptor; MAPK = mitogen-activated protein kinase.
| Bioactive Compound | Health Effect | Reference |
| Cryptides (2–20 AA) | Antioxidant, anti-inflammatory | [47] |
| YAN | Antihypertensive | [52,58] |
| Hydrophobic fractions | Cardiovascular protection | [12] |
| LPDQWDWR, APPDGGFWEWGD | Type 2 diabetes | [53] |
| VVYPWTQ, AWYGANK, LWDHKV | Antihypertensive | (32) |
| Defatted extract | Cardioprotective, anti-inflammatory | [59] |
| DPP-IV inhibitors | Type 2 diabetes | [60] |
| Glycosides & heterocycles | Neuroprotective | [62] |
| Alcalase hydrolysates (standardized process) | Antioxidant; antimicrobial vs. S. aureus, E. coli | [51] |
| Protein hydrolysates (time-resolved) | Increasing antioxidant capacity during hydrolysis | [5] |
| Fermented hydrolysates (L. plantarum) | Anti-inflammatory; improved GI motility (mouse) | [61] |
| Phenolic compounds (methanolic extracts) | Antioxidant (DPPH, FRAP) | [63] |
| Dermocosmetic peptides (review) | Anti-ageing, moisturizing, soothing (skin) | [64] |
| Trypsin hydrolysates (in vivo, dermatitis) | Atopic dermatitis amelioration (TLR-MyD88-MAPK) | [65] |
| T. molitor oil (keratinocytes) | Moisturizing; cytoprotective for skin repair | [66] |
Note: References compiled from multiple studies between 2020 and 2025; values indicate reported experimental outcomes under controlled conditions. These examples collectively highlight the potential of T. molitor proteins and extracts as functional ingredients in health-oriented applications.
Table 3.
Cross-sectoral opportunities for T. molitor.
| Application Domain | Valorized Form of T. molitor | Function / Benefit | References |
| Medical / Biomaterials | Chitin nanofibrils | Skin regeneration, tissue engineering | [4,99,100,102] |
| Agriculture | Frass, chitosan | Organic fertilizer, soil enhancer, crop protection | [5,12,23,24,25,34,42,67,107,108] |
| Animal Nutrition | Meal, peptides | Alternative protein source, gut health, immune modulation | [9,19,39,40,42,43,45] |
| Green Biotechnologies | Chitosan | Biodegradable antimicrobial films | [34,94,95,96] |
| Food Technology | Oil, oleogels | Fat replacement, nutrient-rich oil | [37,82,83,84] |
| Industrial Processes | Whole larvae, oils, meal | Sensor-based monitoring, scalability, optimization | [28,37,75,101,103,104] |
| Bioinformatics & Predictive Workflows (Transversal) | Proteome mining, in silico modeling | Bioactive-peptide prediction; by-product upcycling; computational workflows | [54,56,57] |
Scope note: Table 3 highlights cross-sectoral, non-nutraceutical applications. Rows focused solely on human nutraceutical peptides (covered in Section 6) are excluded. Biomaterials and dermocosmetic uses remain in scope when their function comes from material properties (e.g., chitin nanofibrils), not from ingestible peptides.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



