
Submitted:
05 September 2025
Posted:
09 September 2025
You are already at the latest version
Abstract
It was hypothesized that differences in production system and muscle type may influence the formation of lipid oxidation products (LOP) as well as protein oxidation (protein carbonyl compounds, PCC) during in vitro gastrointestinal digestion of chicken meat. To test our hypothesis, we investigated the formation of LOP and PCC after heating and in vitro gastrointestinal digestion of conventional and organic chicken breast and thigh meat and Wooden Breast meat. Prior to the in vitro digestion, thigh and breast meat was minced and heated. Digests of organic thigh meat had significantly higher levels of all LOP measured compared to conventional thigh meat (between +37% and +173%). Lower levels of LOP were found in digests of breast meat regardless of production system and Wooden Breast phenotype. LOP levels were associated with variations in heme-Fe, polyunsaturated fatty acids, carnosine, anserine and α-tocopherol. PCC levels were significantly higher in thigh meat than in breast meat after heating (+43%) and digestion (+25%), irrespective of the production system. The cut-dependent differences in composition and susceptibility to oxidation during in vitro digestion between organic and conventional chicken meat in the present study warrant further research to allow a comprehensive assessment of the health implications.
Keywords:
4-hydroxy-2-nonenal
; protein carbonyl compounds
; wooden breast
; antioxidants
; PUFA
1. Introduction
Worldwide, the majority of chicken meat (e.g., 90% in European Union countries) is produced in intensive production systems characterized by fast-growing chickens and indoor housing [1,2]. However, the success of the selective breeding programs in attaining fast- growth and large breast mass has coincided with an increased occurrence of muscle myopathies [3]. Among these, the Wooden Breast myopathy has gained attention as a muscular defect characterized by hardened ridges, a paler colour, and surface hemorrhagic lesions on the Pectoralis major muscle [4]. On the other hand, consumer interest in alternative production systems with slower-growing birds and outdoor access (e.g., organic production) has increased in recent years, based on the perception of improved welfare compared to conventional, intensive production systems [5,6].
These contrasting production systems differ in genetics, feeding, and housing practices, which can significantly influence meat quality and nutritional value [7]. For example, meat from slower-growing chickens whether raised in organic systems or not, generally contains higher proportions of polyunsaturated fatty acids (PUFA) and higher contents of heme-iron (heme-Fe), protein, α-tocopherol, and histidine-containing dipeptides (HCD) such as carnosine and anserine, compared to conventionally produced chicken meat [5,8,9,10]. Nutritional differences also exist between chicken meat, with thigh meat containing more fat and heme-Fe and a higher proportion of PUFA than breast meat [7]. Additionally, Wooden Breast meat has a lower protein content and higher lipid content than Normal Breast meat [11].
Such variations in meat composition may have direct implications for the oxidative stability of meat, not only during storage and heating, but also during gastrointestinal digestion. For instance, PUFA are essential nutrients for human health, however, when present in combination with heme-Fe in meat, their susceptibility to oxidative reactions leads to the formation of potentially harmful lipid- and protein oxidation products [12,13]. Oxidation of n-6 PUFA produces 4-hydroxy-2-nonenal (4-HNE) and hexanal (HEX), whereas oxidation of n-3 PUFA generates propanal (PROP). Both types of PUFA can generate malondialdehyde (MDA). MDA and 4-HNE are highly reactive compounds. The alkanals HEX and PROP are less reactive and have more limited effects in the body but are widely used to assess the extent of lipid oxidation. These lipid oxidation products (LOP) are more readily formed during the digestion of meat containing PUFA in the presence of heme-Fe. The pro-oxidant effects of heme-Fe were shown during in vitro digestion of chicken, pork and beef minced with lard to standardize the fatty acid (FA) profile [14]. Steppeler et al. [15] reported that higher levels of MDA and n-3 PUFA-derived oxidation products are formed during the digestion of fish and chicken meat compared to beef, due to their higher levels of PUFA. Furthermore, the interaction of amino acids with reactive oxygen species (ROS), LOP or Maillard reaction products results in the formation of protein carbonyl compounds (PCC), which are established markers of oxidative damage to proteins [16].
Excessive formation and intake of oxidation products may overwhelm the body’s antioxidant defense system, potentially causing oxidative stress and human health issues. For example, at lower concentrations in biological tissues or plasma (0.1–1 µmol/L), 4-HNE may exhibit hormetic effects, however moderate (1–10 µmol/L) and higher (up to 20 µmol/L) concentrations may lead to pathophysiological effects [17]. In addition, in vivo protein oxidation is often associated with aging and age-related diseases in humans [18]. This concern is particularly relevant given the growing consumer preference for organic and alternative production systems, which are often perceived as healthier options. Although previous studies have compared oxidative stability during storage across systems, there remains a knowledge gap on oxidation during gastrointestinal digestion of chicken meat produced in various production systems. The altered meat composition of the Wooden Breast myopathy may also affect its oxidative stability. Therefore, this study was designed to address this gap by assessing the oxidative stability during heating and in vitro gastrointestinal digestion of various chicken meats.
The current study comprised two experiments. We compared the oxidative stability of meat from two production systems (conventional vs. organic) and meat types (breast vs. thigh) in Experiment 1, and Wooden Breast and Normal Breast phenotypes within the conventional system in Experiment 2. In both experiments, we measured meat composition variables (heme-Fe, PUFA, α-tocopherol, carnosine, and anserine), LOP (4-HNE, HEX, PROP, TBARS), and PCC in heated meats and their corresponding in vitro gastrointestinal digests.
2. Materials and Methods
2.1. Chemicals
All digestive enzymes (α-amylase from hog pancreas [~50 U/mg; 10080], mucin from porcine stomach type II [M2378], pepsin from porcine gastric mucosa [>250 U/mg solid, P7000], lipase from porcine pancreas type II [10–400 U/mg protein; L3126], pancreatin from porcine pancreas [8 × United States Pharmacopeia specifications; P7545], and porcine bile extract [B8631]), the reagents (2-thiobarbituric acid [T5500], 1,3-cyclohexanedione [C101605], and 2,4-dinitrophenylhydrazine [D199303], and the analytical standards (HNE-DMA [purity >85%; H9538], 1,1,3,3-Tetramethoxypropane [purity >99%; 108383], alpha-tocopherol [purity >99%; PHR1031], hexanal [purity ≥98%; 115606] and propanal [purity ≥98%; 64409] were purchased from Merck in Diegem, Belgium. The analytical standards of L-carnosine and L-anserine were chemically synthesized by Flamma S.p.A. (Chignolo d’Isola, Bergamo, Italy). All other common chemicals were purchased from AnalytiChem Belgium NV (Zedelgem, Belgium).
2.2. Meat Sampling and Experimental Design
In Experiment 1 of the present study, meat from two distinct broiler production systems was used, i.e., conventional intensive (CON) and organic farming (ORG). The differences between these systems and the finisher phase diets are detailed in Table 1. Housing conditions in the CON system adhered to Council Directive 2007/43/EC [19], with temperature, humidity, and light conditions meticulously controlled, and a stocking density typically ranging from 33 kg/m2 up to 42 kg/m2. In contrast, chickens in the ORG system were housed and reared according to the EU Commission Regulation 889/2008 [20], which mandates access to fresh air, daylight, and outdoor space, with a minimum of 4 m2 of outdoor run per bird. Due to an avian influenza outbreak in Belgium during the sampling period, outdoor access was temporarily restricted on some ORG farms. This reduced the planned 6-week duration to 5, 4, 3, 2, and 1 weeks in the five farms. Chickens in both production systems were fed ad libitum. Five farms, distributed across Belgium, were selected from each production system. A total of 10 chickens per production system (two from each farm, randomly selected without regard to sex) were sampled after slaughtering in two slaughterhouses. The carcasses were transported to the laboratory in a cooling truck. On the second day post-slaughter, approximately 100g of meat from the breast (skinless Pectoralis major) and thigh cuts were sampled for analyses and in vitro gastrointestinal digestion. Carcasses with signs of Wooden Breast were excluded.
In Experiment 2, chickens displaying Wooden Breast were selected from the same five CON farms of Experiment 1 (n=10, two per farm). Wooden Breast was identified following the protocol of Tijare et al. [21] on a 4-point scale (0 = normal, 1 = mild, 2 = moderate, 3 = severe). To allow for more precise differentiation, a score of 2.5 was included to represent samples of which the severity condition was greater than moderate but not severe. Only carcasses with at least a score of 2 were selected, including 7 samples with a score of 2.5, 2 with a score of 3, and 1 with a score of 2. The breast was incised from the most affected part (tip of the breast). These Wooden Breast meat samples were compared to the similarly sampled tip of the Normal Breast samples (n=10) of the CON system from Experiment 1.
All meat samples were vacuum-packed and stored frozen at −80 °C until digestion and analysis. Samples were collectively defrosted and minced. Before the in vitro gastrointestinal digestion, 100 g of each sample was finely minced (Moulinex DP810, France) and heated at an oven temperature of 180 °C (Memmert UN110, Germany) until the core temperature reached 70 °C. Following the heating procedure, meats were minced again, vacuum packed, and stored at −80 °C until digestion or analysis. The characterization of meat was performed on heated meat samples, whereas the analyses of lipid oxidation (TBARS, 4-HNE, HEX, PROP) and protein oxidation products (PCC) were conducted on both the heated meats and their corresponding digests.
2.3. In Vitro Digestion
The in vitro gastrointestinal digestion was based on the protocol of Versantvoort et al. [22], as adapted for studying oxidative reactions in the gastrointestinal system by Van Hecke et al. [23]. The composition and concentration of digestive juices are shown in Supplementary Data (Table S1). This static in vitro model simulated digestion in the mouth, stomach, and small intestine aerobically at 37 °C. Briefly, 4.5 g of meat underwent simulated digestion, starting with a 5-minute exposure to 6 mL of saliva, followed by 2 hours in 12 mL of gastric fluid, and concluding with 2 hours incubation following the consecutive addition of 2 mL bicarbonate buffer, 12 mL duodenal fluid, and 6 mL bile. After digestion, all samples were homogenized at 9500 rpm for 5 s, and the resulting aliquots (termed digests) were stored at −80 °C for further analysis. All samples were digested on the same day with the same digestive juices to exclude possible day effects.
2.4. Meat Composition Analyses
All analyses of meat composition were performed on heated meat samples.
2.4.1. Fatty Acids
Lipids were extracted in duplicate from 3 g meat samples, using chloroform/methanol (2:1, v/v) following a modified method of Folch et al. [24], after which FA were analyzed as described by Raes et al. [25]. After methylation with NaOH/MeOH followed by HCl/MeOH, fatty acid methyl esters (FAME) were analyzed using a gas chromatograph (HP 7890A, Agilent Technologies, Diegem, Belgium) equipped with a fused silica SP-2560 capillary column (75 m × 0.18 mm i.d. × 0.14 µm thickness; Supelco Analytical, Bellefonte, PA, USA) and a flame ionization detector. Nonadecanoic acid (C19:0) was added as an internal standard to quantify the FA content.
2.4.2. Heme-Fe
Heme-Fe was quantified by measuring the hematin concentration in duplicate after extraction with acetone, water, and HCl (40:2:1) as described by Hornsey [26]. The extract was left in a dark place at room temperature for 1 hour to allow all heme pigments to be converted to acid hematin. After this, the extract was filtered through filter paper (S&S 5892) and absorbance was measured spectrophotometrically at 640 nm (Genesys 10S UV–VIS, Thermo Scientific, Madison, USA). Hematin content was converted to heme-Fe using the formula: heme-Fe = (hematin concentration × atomic weight of Fe)/molecular weight of hematin.
2.4.3. α-Tocopherol
The analysis of α-tocopherol was conducted according to Claeys et al. [27]. In brief, 2 g of meat in duplicate was homogenized in ethanol at 78 °C for 30 minutes. The cooled extract was centrifuged with KCl, extracted twice with hexane, and evaporated under nitrogen. The resulting fraction and standards (0–20 μg/mL) were injected into an HPLC system (Agilent 1200 series, Diegem, Belgium) equipped with a Supelcosil LC18 column (25 cm × 4.6 mm, 5 μm) and a UV detector. The absorbance was measured at 292 nm, and α-tocopherol content was calculated by comparing peak areas with a standard curve.
2.4.4. Carnosine and Anserine
Carnosine and anserine concentrations were determined using a modified method of Kobe et al. [28]. Briefly, 1 g of minced meat in duplicate was homogenized in 45 mL of 0.01 mol/L phosphate buffer (pH 7.4) with an ultra-turrax (Type 18/10, Janke and Kunkel) at 9,500 rpm for 30 s. After adding 1 mL of acetonitrile to 2 mL of the homogenate, the mixture was stored overnight at 4 °C and centrifuged at 3,000 rpm for 10 minutes at 4 °C. The supernatant was filtered and 20 μL was injected into an HPLC column (EC250/4.6 Nucleosil 120-7 NH2, Machery-Nagel, Duren, Germany). Concentrations were determined by comparing peaks to standard solutions.
2.5. Oxidation Products in Meats and Digests
2.5.1. Lipid Oxidation Products
Concentrations of 4-HNE, HEX, and PROP in meat and digest samples were measured as fluorescent derivatives using 1,3-cyclohexanedione (CHD) through HPLC (Agilent 1200 series, Diegem, Belgium), following a method modified by Van Hecke et al. [29]. In pre-heated glass tubes, digests and meat samples were mixed with MeOH and CHD reagent, vortexed, and incubated in duplicate. After cooling, MeOH was added, and the mixture was filtered into dark HPLC vials. Separation was performed using a Supelcosil LC-18 column (25 cm × 4.6 mm, 5 μm, catalog no. 58295, Supelcosil), and aldehydes were detected with a fluorescence detector.
Thiobarbituric acid reactive substances (TBARS) in meats and digests were measured as MDA-equivalents according to the method of Grotto et al. [30]. Following alkaline hydrolysis, the absorbance of the MDA-TBA complex was measured spectrophotometrically at 532 nm in the n-butanol fraction (Genesys 10S UV–VIS, Thermo Scientific, Madison, USA).
2.5.2. Protein Oxidation Products
Protein oxidation in meat and digest samples was assessed by measuring PCC following the method of Ganhão et al. [31]. Three aliquots of digest samples (700 μL) or homogenized meat (200 μL from the 1:10 meat-to-phosphate buffer solution) were treated with trichloroacetic acid to precipitate proteins. One aliquot was used as a blank, while the others were treated with 2,4-dinitrophenylhydrazine (DNPH) to form stable 2,4-dinitrophenylhydrazone complexes. After incubation, excess DNPH and fat were removed by three washes with ethanol/ethyl acetate. Afterwards, guanidine HCL (6M) was added to dissolve proteins. Absorbance was measured spectrophotometrically at 280 nm and 370 nm (Genesys 10S UV–VIS, Thermo Scientific, Madison, USA). PCC concentrations were calculated by measuring the amount of 2,4-DNPH incorporated per mg of protein, using the following formula:
Chydrazone/Cprotein = A370/(εhydrazone370 × (A280 − A370 × 0.43)) × 1000
2.6. Statistical Analysis
All analyses were conducted using R (version 4.3.1) and RStudio (version 2023.09.0). Data visualization was performed using the ggplot2 package. In Experiment 1, data were analyzed using a two-way analysis of variance (ANOVA) model, which evaluated the main effects of production system (CON vs. ORG) and meat type (breast vs. thigh), as well as their interaction term. The ANOVA model was fitted using the ‘aov’ function in R. Residual normality was assessed through Q-Q plots, histograms, and the Shapiro-Wilk test, and data was log-transformed when needed. Estimated marginal means were calculated using the emmeans package, and Bonferroni-adjusted post-hoc tests were performed on all pairwise comparisons. A heatmap of Pearson correlation coefficients was generated using the pheatmap package. Principal Component Analysis (PCA) was conducted on the standardized numeric variables from the dataset using the ‘prcomp’ function. A biplot was generated to illustrate the PCA results, highlighting individual observations along with their respective groups. The significance level was set at P ≤ 0.05. A P-value greater than 0.05 but less than or equal to 0.1 was considered a trend.
In Experiment 2, a two-sample t-test was applied to assess mean differences between the two meat types (Normal Breast and Wooden Breast). The Shapiro-Wilk test for normality and Levene’s test for homogeneity of variances confirmed the assumptions for the t-test. The significance level was set at P ≤ 0.05.
3. Results
3.1. Experiment 1: Meat Composition
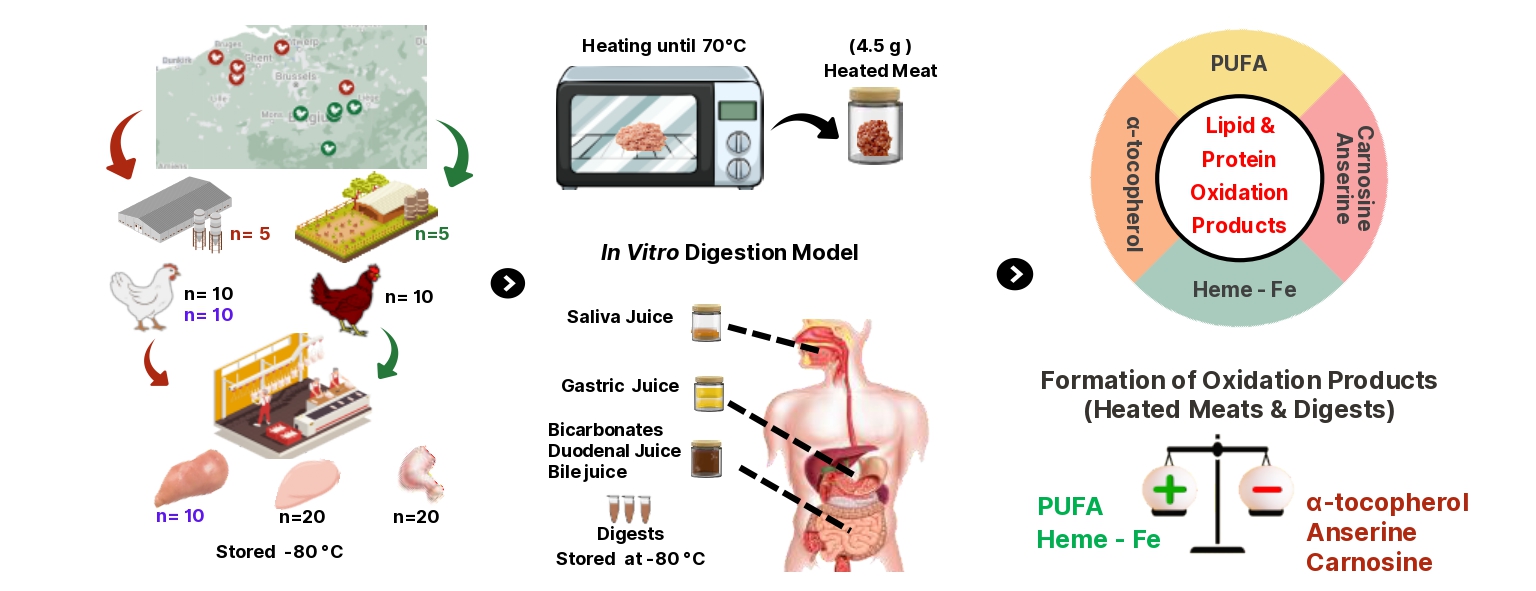
Table 2 summarizes the fatty acid profile, while Figure 1 shows concentrations of heme-Fe, α-tocopherol, carnosine, and anserine. The concentrations of total FA, SFA and MUFA were significantly influenced by the meat type only, being approximately three times higher in thigh meat compared to breast meat from both production systems. Levels of PUFA, encompassing both n-3 and n-6 PUFA, were mainly affected by the meat type, but significant effects of the production system, alongside its interaction with meat type, were observed as well. The lowest concentrations of total n-3 and total n-6 PUFA were observed in breast meat from both production systems followed by intermediate levels in CON thigh and the highest levels in ORG thigh. This pattern of n-3 PUFA and n-6 PUFA was reflected in the pattern of the levels of α-linolenic acid (ALA) and linoleic acid (LA) as the parent fatty acids, respectively. The long-chain n-3 derivative EPA was significantly higher in the CON thigh compared to the ORG breast with intermediate levels for the other groups. The level of DHA significantly increased in the order CON breast and CON thigh < ORG breast < ORG thigh. The n-6/n-3 PUFA ratio was not different between the groups.
The concentration of heme-Fe in the meats was significantly influenced by the production system, meat type and their interaction. The levels of heme-Fe were highest in ORG thighs, followed by CON thighs, and lowest in the breast meat irrespective of production system. Levels of α-tocopherol were significantly lower in breast compared to thigh meat (−26%), and higher in meats from the CON system compared to ORG meats (+48%). Anserine and carnosine concentrations were significantly affected by the production system, meat type, and their interaction. Specifically, breast meat had significantly higher levels of anserine (+157%) and carnosine (+161%) than thigh meat. ORG meats contained higher levels of anserine (+20%) and carnosine (+287%) than CON meats.
3.2. Experiment 1: Lipid and Protein Oxidation Products
Concentrations of 4-HNE, HEX, PROP, TBARS and PCC in meats (before digestion) and in digests (post-digestion), are presented in Figure 2. As anticipated, the levels of metabolites associated with the oxidation of n-6 PUFA (4-HNE and HEX), n-3 PUFA (PROP), and both n-6 and n-3 PUFA (TBARS) were multi-fold increased during the in vitro gastrointestinal digestion process. Levels of LOP in meats and digests were significantly affected by meat type and production system, with significant interaction effects observed primarily in the meats. ORG thigh contained significantly higher levels of all analyzed LOP, with increases ranging from 3.5 to 18 times in the heated meat samples and 1.6 to 4.4 times in the digests, compared to other meat types which did not differ among each other. Specifically, the digests of ORG thigh contained significantly higher concentrations of 4-HNE (4.4-fold), HEX (2.4-fold), PROP (1.6-fold) and TBARS (2-fold) compared to the mean of other digests. Chicken thigh meat from the ORG system had significantly higher PCC levels compared to CON breast, whereas intermediate levels were found in the ORG breast and CON thigh meat. After digestion, PCC increased significantly, demonstrating a meat type effect, with digests of thigh exhibiting a higher (+25%) concentration of PCC compared to digests of breast.
3.3. Experiment 1: Relationships Among Compositional Variables and Oxidation Products
Pearson correlation coefficients were used to assess relationships among the variables across all meats or digests, as shown in Supplementary Data (Figure S1). 4-HNE, HEX, PROP and TBARS in meats and digests were positively correlated with heme-Fe, total FA, n-3 PUFA, and n-6 PUFA. In digests, PCC were positively correlated with 4-HNE, HEX, PROP and TBARS. The antioxidants (α-tocopherol, carnosine and anserine) showed generally negative correlations with oxidation products (4-HNE, HEX, PROP, TBARS, and PCC) in meats and digests. To further investigate these relationships, a PCA biplot was performed as shown in Figure 3. A total of 73.7% of the variance in the data was explained, with PC1 accounting for 57.9% and PC2 for 15.8%. PC1 primarily separated the groups based on lipid oxidation products and FA profile. Markers of lipid oxidation loaded heavily along this dimension and were positively associated with n-3 PUFA, n-6 PUFA, and heme-Fe. In contrast, PC2 distinguished groups based on antioxidant content. The positioning of the groups along these components reflected differences in nutrient composition and oxidative profiles, with a more distinct separation between meat types than between production systems.
3.4. Experiment 2: Meat Composition
No significant differences were observed for total FA, the n-6/n-3 PUFA ratio, and the various FA fractions between Normal Breast and Wooden Breast, except for ALA, which was significantly higher in Wooden Breast (Table 3). This higher ALA contributed to a tendency toward a +45% increase in n-3 PUFA content in Wooden Breast meat. In addition, Wooden Breast meat contained significantly lower α-tocopherol levels (−28%) and tended to have lower anserine levels compared to Normal Breast meat, whereas no significant differences were observed for heme-Fe and carnosine (Figure 4).
3.5. Experiment 2: Lipid and Protein Oxidation Products
Concentrations of 4-HNE, HEX, PROP, TBARS and PCC in meats and digests are presented in Figure 5. In the meats, concentrations of 4-HNE, TBARS, PROP, and HEX were slightly higher in Wooden Breast compared to Normal Breast, with statistically significant differences for 4-HNE and trends observed for TBARS, PROP, and HEX. After digestion, the concentration of these oxidation products was generally increased. However, only HEX levels were significantly higher in Wooden Breast compared to Normal Breast.
4. Discussion
We investigated oxidative reactions in chicken meat during heating and in vitro gastrointestinal digestion, focusing on breast and thigh meat from CON and ORG production systems, as well as Wooden Breast and Normal Breast meat within a CON system. One of the major outcomes was the higher formation of LOP in thigh meat from ORG chickens during in vitro digestion. This appears to be largely driven by variations in meat composition which were influenced by both the production system and meat type. Although ORG meat may have a more favorable nutritional profile due to the rearing and feeding conditions, its higher susceptibility to oxidation may challenge the widespread belief that it is inherently superior for human health.
Higher proportions of PUFA (n-3 and n-6 PUFA) and heme-Fe content have been observed in ORG chicken meat [5,8,9], which aligns with the results of the present study. Furthermore, thigh meat, compared to breast meat, has been reported to contain higher levels of PUFA and heme-Fe, highlighting the influence of meat type on the nutritional composition of meat [7]. The higher heme-Fe content in thigh meat is associated with a greater proportion of oxidative muscle fibers compared to the predominant glycolytic fiber type in breast muscle [32]. In the present study, the combined effect of production system and meat type pronounced the differences in concentration of heme-Fe. Heme-Fe was reported to be higher in slow-growing chickens, particularly in thigh muscles compared to breast muscles [33]. Generally, it has been shown that in chickens, selection focused on growth and breast meat yield has led to a decrease in heme-Fe levels [34].
In contrast to studies that reported elevated α-tocopherol levels in meat from ORG chickens due to increased foraging behaviour [35], we observed lower α-tocopherol concentrations in ORG meat, consistent with the findings of Michiels et al. [36]. In the present study, the difference in α-tocopherol levels was not due to differences in feed levels, as similar α-tocopherol content was provided in the feed. Instead, the reduced α-tocopherol levels may be attributed to its increased utilization as an antioxidant in the body to mitigate higher oxidative stress from greater physical activity [37,38]. Moreover, it was found in a recent study in medium-growing chickens raised outdoors, that the most exploratory individuals exhibited lower plasma tocopherol concentrations and total antioxidant status compared to their more sedentary counterparts, confirming a link between these parameters and the animals’ physical activity [39]. In fact, oxidative homeostasis depends on the dietary intake of antioxidants, but also on the interaction of the tissue lipid profile with oxidative stress produced by kinetic activity [40]. The latter is related to the effect of locomotory activity on muscle metabolism and the production of free radicals. The adaptation to exercise is affected by genetic strain and could result in a different body antioxidant status [39]. Accordingly, the results obtained here could be affected by dietary inputs in the chicken body but also by differences in metabolism, which for certain molecules [41], and processes can be genetically driven [38,42]. Therefore, differences in the present study may be attributed to the different genotypes used for the CON and ORG production systems. In addition, the limited outdoor access of ORG chickens in this study could have restricted their foraging behaviour, thereby also reducing their intake of natural antioxidants. The concentration of anserine in the heated meats was notably higher than that of carnosine, confirming that anserine is the most abundant HCD in chicken skeletal muscle [10]. The higher concentration of HCD in breast meat compared to thigh, particularly in ORG breast, is likely related to the predominance of type IIB fast-twitch glycolytic fibers. These fibers rely on anaerobic metabolism, leading to increased lactic acid production, which necessitates more buffering compounds like anserine and carnosine to maintain the pH balance [43].
The higher oxidative susceptibility of ORG meat after heating was likely primarily due to its higher degree of unsaturation. In line with this Castellini et al. [37], reported higher TBARS levels in meat from ORG chickens compared to CON chickens, which was attributed to elevated levels of PUFA and heme-Fe. In addition to the influence of the production system, the meat type plays a crucial role in oxidative stability. Thigh meat, with its higher heme-Fe content and unsaturated FA profile, is naturally more prone to oxidation than breast meat, irrespective of the production system, and compared to other meats like beef and pork [12]. PCA analysis in the present study confirmed these findings, with oxidation products clustering strongly with ORG thigh meat. The antioxidant profile of the meat further influenced LOP formation. Lower levels of α-tocopherol and HCD in meat were associated with higher oxidation. This is consistent with findings from Michiels et al. [36], who reported elevated lipid oxidation and reduced α-tocopherol levels in chickens raised under conditions very similar to ORG systems (slow-growth, outdoor access) compared to CON systems (fast-growth, indoor confinement). α-tocopherol is a potent lipid-soluble antioxidant, reducing lipid oxidation by rapidly neutralizing radicals and protecting unsaturated fatty acids in a concentration-dependent manner [44]. Additionally, α-tocopherol delays the decomposition of hydroperoxides, scavenges ROS, and complexes metals, all of which help inhibiting the propagation phase of lipid oxidation [45]. HCD have been reported to contribute to the antioxidant status of chicken meat [46]. Specifically, carnosine inhibits lipid oxidation not only by scavenging free radicals and chelating pro-oxidant metals but also by inducing structural changes in the pro-oxidant metmyoglobin, thereby reducing its oxidative activity [47]. Also in the present study, lower levels of LOP were observed in meats with higher concentrations of HCD, however, the correlations only reached significance for anserine with LOP.
Notably, the formation of LOP increased during in vitro digestion compared to the levels observed in the heated meats. This is consistent with previous studies on the in vitro digestion of various muscle foods, e.g., Gorelik et al. [48], Steppeler et al. [15], and Van Hecke et al. [29,49] reporting similar increases in LOP levels during digestion. This increase was observed in all groups. A key factor contributing to this rise may be the conditions in the gastric phase of the in vitro model, which includes low pH and aeration to mimic the stomach’s aerobic environment. These conditions are ideal for promoting lipid peroxidation and co-oxidation of dietary constituents [50]. After digestion, 4-HNE, HEX, PROP, and TBARS levels remained highest in the ORG thigh digests compared to the other digests, consistent with the heated meat results. However, digestion revealed a unique redistribution of HEX and TBARS: levels were lowest in CON breast, intermediate in ORG breast and CON thigh, and highest in ORG thigh. The observed variation in LOP between heated and digested meats suggests that the in vitro digestion process itself may influence HEX and TBARS levels. This indicates that, beyond the role of meat composition in oxidation during heating, additional factors present during digestion may further contribute to LOP formation. As meat moves through the distinct stages of the digestion process, it encounters specific conditions: enzymatic action in the mouth phase, acidic gastric juices in the stomach phase, and bile and pancreatic enzymes in the intestinal phase. Each of these phases influences oxidative reactions, adding complexity to the formation of LOP as digestion progresses [13]. Post-digestion LOP levels were higher in thigh meat than in breast meat, consistent with findings from Van Hecke et al. [49]. Interestingly, the LOP in meats and digests exhibited similar correlations with n-3 PUFA and n-6 PUFA, whereas their precursors are either n-6 PUFA (precursor of 4-HNE and HEX) or n-3 PUFA (precursor of PROP). This may be attributed to the correlated pattern of n-3 and n-6 PUFA in the meats. In addition, the initial oxidative status of meat lipids also plays an important role. Primary oxidation products such as hydroperoxides, formed through reactions between ROS and PUFA during heating, can promote lipid peroxidation during digestion, leading to increased formation of secondary LOP [51]. Larsson et al. [52] demonstrated that higher initial oxidation levels in cod liver oil resulted in more TBARS during in vitro digestion, partly due to the depletion of endogenous antioxidants.
The observed lower levels of α-tocopherol in ORG meat may have reduced its capacity to inhibit LOP formation during in vitro digestion [51]. Supporting this, Kenmogne-Domguia et al. [53], reported significant losses of tocopherols during in vitro digestion of unsaturated oil emulsions, with α-tocopherol being rapidly consumed during the gastric phase. Moreover, the presence of pro-oxidant factors such as metmyoglobin further accelerates tocopherol depletion, thereby reducing the system’s ability to counteract lipid peroxidation. This indicates that while antioxidants are present, their capacity can be overwhelmed under highly oxidative conditions. The high PUFA and heme-Fe in ORG thigh meat likely created a strong oxidative environment that exceeded the protective potential of antioxidants. Similar findings have been reported for beef, suggesting that antioxidant defenses are effective under low oxidative conditions but insufficient when reactive substances are abundant [54]. Likewise, Van Hecke et al. [49] did not find the intrinsic levels of α-tocopherol, carnosine and anserine to be major determinants for the formation of oxidation products during in vitro digestion of meat from different species. On the other hand, the addition of extra antioxidants could partially mitigate oxidation in a study by Van Hecke et al. [55], who reported significantly lower levels of TBARS, HEX, and 4-HNE during in vitro digestion of high-fat beef mince supplemented with α-tocopherol.
Although there is limited information on the ability of antioxidant dipeptides like carnosine and anserine to reduce LOP during the digestion of muscle foods, both have shown the ability to inhibit low-density lipoproteins (LDL) oxidation, especially when LDL was oxidized in vitro with copper in their presence [56]. In that study, carnosine was found to be a more effective aldehyde quencher than anserine at equal concentrations. In the present study, the absolute amounts of anserine were approximately three times higher than those of carnosine in breast and thigh meat, which may overshadow carnosine’s antioxidant effects. Li et al. [57] reported a dual role of carnosine as both an antioxidant and pro-oxidant during in vitro digestion of a burger meal model. At higher concentration, carnosine exhibited antioxidant properties, whereas at moderate concentration, it acted as a pro-oxidant. In addition, the antioxidant efficacy of carnosine may be influenced by other meal factors, such as the fat content and cooking conditions. For example, the ability of carnosine to reduce lipid oxidation during in vitro digestion was more pronounced in moderately cooked, low-fat (1.3%) pork compared to high-fat (10%) pork [58]. In our study, these factors may have similarly contributed to the lower LOP formation in breast, possibly due to its higher carnosine and lower fat content compared to thigh.
The elevated PCC in thigh meat compared to breast meat reinforces the idea that the greater susceptibility of oxidative-type muscles to lipid oxidation, compared to glycolytic-type muscles, also extends to protein oxidation. Another factor that could have contributed to the higher PCC in thigh meat compared to breast is that primary as well as secondary LOP may serve as substrates for protein oxidation [59]. Estévez [16] suggested that reactive LOP, such as 4-HNE, can promote the formation of PCC in meat. Similarly, Van Hecke et al. [49] reported that 4-HNE may further enhance PCC formation during in vitro digestion of meat. This might explain the significant correlations and shared PCA clustering of PCC and LOP in digests, which was also reported by Van Hecke et al. [49]. Although Santé-L’houtellier et al. [60] reported that α-tocopherol exerts a protective effect against protein oxidation during the storage of lamb meat, we did not observe increased protein oxidation after heating and digestion of ORG meats, despite their lower α-tocopherol levels.
Minor differences in the FA profile between Wooden Breast and Normal Breast phenotypes were noticed in the present study, in line with Soglia et al. [61] and Thanatsang et al. [62], except for higher ALA concentrations in Wooden Breast, as well as a trend toward increased total n-3 PUFA levels. In partial agreement with Abasht et al. [63], we found no significant difference in carnosine, while anserine levels tended to be lower in Wooden Breast meat. The slightly higher levels of TBARS in Wooden Breast meat were consistent with the findings of Soglia et al. [61] and Xing et al. [64]. The former study indicated that although there were no differences in the FA profiles between Normal Breast and Wooden Breast, the Wooden Breast meat exhibited a greater susceptibility to lipid oxidation. In the present study, the modestly higher lipid oxidation observed in Wooden Breast may have been due to a combination of higher levels of n-3 PUFA and lower α-tocopherol levels. Other factors that could stimulate lipid oxidation in Wooden Breast phenotype include the continuous exposure of phospholipids to oxidative stress due to the hypoxic conditions followed by reoxygenation, which may enhance ROS formation [65]. Li et al. [66] underlined that high levels of lipid oxidation may be caused by increased metabolic waste, and poor blood and oxygen supply in the Wooden Breast phenotype. Although these processes occur in vivo, the oxidative damage encountered by the tissues could predispose the meat to increased lipid oxidation post-mortem or during digestion [67]. Moreover, Carvalho et al. [68] reported that Wooden Breast meat shows an altered redox balance, with increased abundance of proteins involved in oxidative damage repair and inflammation, as well as depletion of sulfur-containing protein components, which may compromise its antioxidant defense. Contrary to studies showing higher protein oxidation in Wooden Breast meat [61,62,64], our findings did not indicate significant differences in PCC between Normal and Wooden Breast meat, both before and after digestion. This may have been due to the similarities in muscle fiber composition, heme-Fe levels, and minor differences in secondary LOP between the two types of meat.
The higher susceptibility of ORG chicken meat to oxidation during digestion may compromise its nutritive value and reduce the bioavailability of nutrients (fatty acids and amino acids). This may seem counterintuitive as ORG foods are generally perceived as healthier and more nutritious due to their more ‘natural’ mode of production. However, there is a lack of comprehensive studies on the nutritional and health impact of consuming ORG versus CON meat [69]. In this context, the higher lipid oxidation observed in the present study does not necessarily translate to an increased health risk for humans. The health implications of oxidation products are complex and multifaceted, with their effects depending strongly on their concentrations. For instance, 4-HNE can modify LDL and contribute to foam cell formation, a factor in cardiovascular disease, however, at lower concentrations in plasma or vascular tissue, 4-HNE may induce adaptive cellular responses that enhance resistance to oxidative stress [17,70]. Our findings were derived from an in vitro model, which, although informative, does not fully replicate human physiological conditions. Therefore, caution is needed when extrapolating the present results to in vivo scenarios. In addition, a limitation of this study is that the two production systems compared differed not only in management practices, but also in breed, feeding regimen, and slaughter age. These confounding factors limit direct attribution of the observed oxidation patterns solely to the production system. Nevertheless, this reflects real market conditions, where consumers encounter meat derived from diverse commercial systems, and thus the findings remain relevant to the food matrix available to them.
5. Conclusions
This study showed that production system and meat type significantly and not independently impact oxidative reactions in chicken meat during in vitro gastrointestinal digestion. Thigh meat from an ORG system contained more heme-Fe and PUFA compared to the other meats, coinciding with increased formation of LOP during in vitro digestion, whereas PCC formation was not affected. One should be cautious in generalizing these findings to all ORG chicken meat given the multiple factors and practices determining differences between production systems and meat types. Future studies with more controlled comparisons of breed, feed, and age are warranted to isolate the specific role of production system in lipid oxidation. The concentrations of LOP found in the chicken meats and digests indicate potential biological significance. However, it is difficult to determine whether these levels are in a hormetic or pathophysiological range. This could be clarified through in vivo studies assessing postprandial plasma levels of oxidation products and their potential impact on human health in consumers of chicken meat differing in composition and antioxidant status. Further research could also aim at a comprehensive understanding of the effects and interactions of the numerous intrinsic and extrinsic factors in production systems on the oxidative status of meat. Furthermore, strategies to maintain the meat oxidative stability throughout the entire supply chain are essential. Specifically, food manufacturers should explore product specific antioxidant strategies, processing and packaging solutions to mitigate oxidation in chicken meat and help preserve its nutritional value. Such efforts are important to ensure that the perceived health benefits of ORG chicken meat are effectively delivered to consumers.
Supplementary Materials
The following supporting information is available online: Table S1: Composition of the simulated digestive juices (per liter) used in the in vitro digestion model. Figure S1: Visualization of Pearson correlations between variables of oxidation products [protein carbonyl compounds (PCC), thiobarbituric acid reactive substances (TBARS), propanal (PROP), hexanal (HEX), 4-hydroxy-2-nonenal (4-HNE)] in both meats (M) and digests (D) and meat characteristics [n-6 PUFA, n-3 PUFA, total FA, heme-Fe, Anserine, Carnosine, and α-tocopherol] from experiment 1, presented as a heatmap plot. Table S2: Selected oxidation products, their structures, and references.
Author Contributions
Conceptualization, T.V.H.; data curation, Z.A.; funding acquisition, S.D.S.; methodology, Z.A. and T.V.H.; supervision, E.K. and S.D.S.; writing—original draft, Z.A.; writing—review and editing, E.V., M.P. and C.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the European Union’s Horizon 2020 INTAQT project (grant agreement number 101000250) and the Higher Education Commission of Pakistan (HEC/HRD/OSS-III/Blg/2021/19305).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further information can be obtained from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 4-HNE | 4-hydroxy-2-nonenal |
| CON | conventional production system |
| DNPH | 2,4-dinitrophenylhydrazine |
| FA | fatty acid |
| HCD | histidine-containing dipeptides |
| HEX | hexanal |
| LDL | low-density lipoproteins |
| MDA | malondialdehyde |
| MUFA | monounsaturated fatty acids |
| ORG | organic production system |
| PCC | protein carbonyl compounds |
| PROP | propanal |
| PUFA | polyunsaturated fatty acids |
| ROS | reactive oxygen species |
| SFA | saturated fatty acids |
| TBA | thiobarbituric acid |
| TBARS | thiobarbituric acid reactive substances |
References
- Albrecht, A.; Hebel, M.; Mittler, M.; Hurck, C.; Kustwan, K.; Heitkönig, B.; Bitschinski, D.; Kreyenschmidt, J. Influence of Different Production Systems on the Quality and Shelf Life of Poultry Meat: A Case Study in the German Sector. Journal of Food Quality 2019, 1–11. [CrossRef]
- European Parliament. Directorate General for Parliamentary Research Services. The EU Poultry Meat and Egg Sector: Main Features, Challenges and Prospects: In Depth Analysis.; Publications Office: LU, 2019.
- Barbut, S.; Mitchell, R.; Hall, P.; Bacon, C.; Bailey, R.; Owens, C.M.; Petracci, M. Review: Myopathies in Broilers: Supply Chain Approach to Provide Solutions to Challenges Related to Raising Fast Growing Birds. Poultry Science 2024, 103, 103801. [Google Scholar] [CrossRef]
- Sihvo, H.-K.; Immonen, K.; Puolanne, E. Myodegeneration With Fibrosis and Regeneration in the Pectoralis Major Muscle of Broilers. Vet Pathol 2014, 51, 619–623. [Google Scholar] [CrossRef]
- Gálvez, F.; Domínguez, R.; Maggiolino, A.; Pateiro, M.; Carballo, J.; De Palo, P.; Barba, F.J.; Lorenzo, J.M. Meat Quality of Commercial Chickens Reared in Different Production Systems: Industrial, Range and Organic. Annals of Animal Science 2020, 20, 263–285. [Google Scholar] [CrossRef]
- Brown, S.N.; Nute, G.R.; Baker, A.; Hughes, S.I.; Warriss, P.D. Aspects of Meat and Eating Quality of Broiler Chickens Reared under Standard, Maize-Fed, Free-Range or Organic Systems. British Poultry Science 2008, 49, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Baéza, E.; Guillier, L.; Petracci, M. Review: Production Factors Affecting Poultry Carcass and Meat Quality Attributes. Animal 2022, 16, 100331. [Google Scholar] [CrossRef]
- Husak, R.L.; Sebranek, J.G.; Bregendahl, K. A Survey of Commercially Available Broilers Marketed as Organic, Free-Range, and Conventional Broilers for Cooked Meat Yields, Meat Composition, and Relative Value. Poultry Science 2008, 87, 2367–2376. [Google Scholar] [CrossRef]
- Michalczuk, M.; Zdanowska-Sąsiadek, Ż.; Damaziak, K.; Niemiec, J. Influence of Indoor and Outdoor Systems on Meat Quality of Slow-Growing Chickens. CyTA—Journal of Food 2016, 1–6. [CrossRef]
- Barbaresi, S.; Maertens, L.; Claeys, E.; Derave, W.; De Smet, S. Differences in Muscle Histidine-containing Dipeptides in Broilers. J Sci Food Agric 2019, 99, 5680–5686. [Google Scholar] [CrossRef]
- Petracci, M.; Soglia, F.; Madruga, M.; Carvalho, L.; Ida, E.; Estévez, M. Wooden-Breast, White Striping, and Spaghetti Meat: Causes, Consequences and Consumer Perception of Emerging Broiler Meat Abnormalities. Comp Rev Food Sci Food Safe 2019, 18, 565–583. [Google Scholar] [CrossRef]
- Min, B.; Nam, K.C.; Cordray, J.; Ahn, D.U. Endogenous Factors Affecting Oxidative Stability of Beef Loin, Pork Loin, and Chicken Breast and Thigh Meats. Journal of Food Science 2008, 73, C439–C446. [Google Scholar] [CrossRef] [PubMed]
- Van Hecke, T.; Goethals, S.; Vossen, E.; De Smet, S. Long-Chain n-3 PUFA Content and n-6/n-3 PUFA Ratio in Mammal, Poultry, and Fish Muscles Largely Explain Differential Protein and Lipid Oxidation Profiles Following In Vitro Gastrointestinal Digestion. Molecular Nutrition Food Res 2019, 63, 1900404. [Google Scholar] [CrossRef]
- Van Hecke, T.; Vanden Bussche, J.; Vanhaecke, L.; Vossen, E.; Van Camp, J.; De Smet, S. Nitrite Curing of Chicken, Pork, and Beef Inhibits Oxidation but Does Not Affect N -Nitroso Compound (NOC)-Specific DNA Adduct Formation during in Vitro Digestion. J. Agric. Food Chem. 2014, 62, 1980–1988. [Google Scholar] [CrossRef] [PubMed]
- Steppeler, C.; Haugen, J.-E.; Rødbotten, R.; Kirkhus, B. Formation of Malondialdehyde, 4-Hydroxynonenal, and 4-Hydroxyhexenal during in Vitro Digestion of Cooked Beef, Pork, Chicken, and Salmon. J. Agric. Food Chem. 2016, 64, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Estévez, M. Protein Carbonyls in Meat Systems: A Review. Meat Science 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Nègre-Salvayre, A.; Garoby-Salom, S.; Swiader, A.; Rouahi, M.; Pucelle, M.; Salvayre, R. Proatherogenic Effects of 4-Hydroxynonenal. Free Radical Biology and Medicine 2017, 111, 127–139. [Google Scholar] [CrossRef]
- Estévez, M.; Luna, C. Dietary Protein Oxidation: A Silent Threat to Human Health? Critical Reviews in Food Science and Nutrition 2017, 57, 3781–3793. [Google Scholar] [CrossRef]
- European Union. Council Directive 2007/43/EC of 28 June 2007 Laying down Minimum Rules for the Protection of Chickens Kept for Meat Production. Off. J. Eur. Union 2007, L 182, 19–28. Available online: http://data.europa.eu/eli/dir/2007/43/oj (accessed on 28 August 2025).
- European Commission. Commission Regulation (EC) No 889/2008 of 5 September 2008 Laying down Detailed Rules for the Implementation of Council Regulation (EC) No 834/2007 on Organic Production and Labelling of Organic Products with Regard to Organic Production, Labelling and Control. Off. J. Eur. Union 2008, L 250, 1–84. Available online: https://eur-lex.europa.eu/eli/reg/2008/889/oj (accessed on 28 August 2025).
- Tijare, V.V.; Yang, F.L.; Kuttappan, V.A.; Alvarado, C.Z.; Coon, C.N.; Owens, C.M. Meat Quality of Broiler Breast Fillets with White Striping and Woody Breast Muscle Myopathies. Poultry Science 2016, 95, 2167–2173. [Google Scholar] [CrossRef] [PubMed]
- Versantvoort, C.H.M.; Oomen, A.G.; Van De Kamp, E.; Rompelberg, C.J.M.; Sips, A.J.A.M. Applicability of an in Vitro Digestion Model in Assessing the Bioaccessibility of Mycotoxins from Food. Food and Chemical Toxicology 2005, 43, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Van Hecke, T.; Basso, V.; De Smet, S. Lipid and Protein Oxidation during in Vitro Gastrointestinal Digestion of Pork under Helicobacter Pylori Gastritis Conditions. J. Agric. Food Chem. 2018, 66, 13000–13010. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A SIMPLE METHOD FOR THE ISOLATION AND PURIFICATION OF TOTAL LIPIDES FROM ANIMAL TISSUES. Journal of Biological Chemistry 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Raes, K.; Smet, S.D.; Demeyer, D. Effect of Double-Muscling in Belgian Blue Young Bulls on the Intramuscular Fatty Acid Composition with Emphasis on Conjugated Linoleic Acid and Polyunsaturated Fatty Acids. Anim. Sci. 2001, 73, 253–260. [Google Scholar] [CrossRef]
- Hornsey, H.C. The Colour of Cooked Cured Pork. I.—Estimation of the Nitric oxide-Haem Pigments. J Sci Food Agric 1956, 7, 534–540. [Google Scholar] [CrossRef]
- Claeys, E.; Vossen, E.; De Smet, S. Determination of A-tocopherol by Reversed-phase HPLC in Feed and Animal-derived Foods without Saponification. J Sci Food Agric 2016, 96, 522–529. [Google Scholar] [CrossRef]
- Kobe, R.; Ishihara, Y.; Takano, J.; Kitami, H. Simultaneous Determination of Anserine and Carnosine in Chicken Meat by Hydrophilic Interaction Chromatography on an Aminopropyl Bonded Silica Gel Column. BUNSEKI KAGAKU 2011, 60, 859–863. [Google Scholar] [CrossRef]
- Van Hecke, T.; Ho, P.L.; Goethals, S.; De Smet, S. The Potential of Herbs and Spices to Reduce Lipid Oxidation during Heating and Gastrointestinal Digestion of a Beef Product. Food Research International 2017, 102, 785–792. [Google Scholar] [CrossRef]
- Grotto, D.; Santa Maria, L.D.; Boeira, S.; Valentini, J.; Charão, M.F.; Moro, A.M.; Nascimento, P.C.; Pomblum, V.J.; Garcia, S.C. Rapid Quantification of Malondialdehyde in Plasma by High Performance Liquid Chromatography–Visible Detection. Journal of Pharmaceutical and Biomedical Analysis 2007, 43, 619–624. [Google Scholar] [CrossRef]
- Ganhão, R.; Morcuende, D.; Estévez, M. Protein Oxidation in Emulsified Cooked Burger Patties with Added Fruit Extracts: Influence on Colour and Texture Deterioration during Chill Storage. Meat Science 2010, 85, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Soyer, A.; Özalp, B.; Dalmış, Ü.; Bilgin, V. Effects of Freezing Temperature and Duration of Frozen Storage on Lipid and Protein Oxidation in Chicken Meat. Food Chemistry 2010, 120, 1025–1030. [Google Scholar] [CrossRef]
- Pellattiero, E.; Tasoniero, G.; Cullere, M.; Gleeson, E.; Baldan, G.; Contiero, B.; Dalle Zotte, A. Are Meat Quality Traits and Sensory Attributes in Favor of Slow-Growing Chickens? Animals 2020, 10, 960. [Google Scholar] [CrossRef] [PubMed]
- Berri, C.; Wacrenier, N.; Millet, N.; Le Bihan-Duval, E. Effect of Selection for Improved Body Composition on Muscle and Meat Characteristics of Broilers from Experimental and Commercial Lines. Poultry Science 2001, 80, 833–838. [Google Scholar] [CrossRef]
- Dal Bosco, A.; Mugnai, C.; Mattioli, S.; Rosati, A.; Ruggeri, S.; Ranucci, D.; Castellini, C. Transfer of Bioactive Compounds from Pasture to Meat in Organic Free-Range Chickens. Poultry Science 2016, 95, 2464–2471. [Google Scholar] [CrossRef]
- Michiels, J.; Tagliabue, M.M.; Akbarian, A.; Ovyn, A.; De Smet, S. Oxidative Status, Meat Quality and Fatty Acid Profile of Broiler Chickens Reared under Free-Range and Severely Feed-Restricted Conditions Compared with Conventional Indoor Rearing. Avian Biology Research 2014, 7, 74–82. [Google Scholar] [CrossRef]
- Castellini, C.; Mugnai, C.; Dal Bosco, A. Effect of Organic Production System on Broiler Carcass and Meat Quality. Meat Science 2002, 60, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, S.; Mancinelli, A.C.; Dal Bosco, A.; Ciarelli, C.; Amato, M.G.; Angelucci, E.; Chiattelli, D.; Castellini, C. Intake of Nutrients (Polyunsaturated Fatty Acids, Tocols, and Carotenes) and Storage Efficiency in Different Slow-Growing Chickens Genotypes Reared in Extensive Systems. PLoS ONE 2022, 17, e0275527. [Google Scholar] [CrossRef] [PubMed]
- Bonnefous, C.; Collin, A.; Guilloteau, L.A.; Germain, K.; Ravon, L.; Bordeau, T.; Chartrin, P.; Godet, E.; Cailleau-Audouin, E.; Couroussé, N.; et al. Performance, Meat Quality and Blood Parameters in Four Strains of Organic Broilers Differ According to Range Use. Sci Rep 2024, 14, 30854. [Google Scholar] [CrossRef]
- Mattioli, S.; Angelucci, E.; Castellini, C.; Cartoni Mancinelli, A.; Chenggang, W.; Di Federico, F.; Chiattelli, D.; Dal Bosco, A. Effect of Genotype and Outdoor Enrichment on Productive Performance and Meat Quality of Slow Growing Chickens. Poultry Science 2024, 103, 104131. [Google Scholar] [CrossRef]
- Perini, F.; Wu, Z.; Cartoni Mancinelli, A.; Soglia, D.; Schiavone, A.; Mattioli, S.; Mugnai, C.; Castellini, C.; Smith, J.; Lasagna, E. RNAseq Reveals Modulation of Genes Involved in Fatty Acid Biosynthesis in Chicken Liver According to Genetic Background, Sex, and Diet. Animal Genetics 2023, 54, 338–354. [Google Scholar] [CrossRef]
- Coudert, E.; Baéza, E.; Chartrin, P.; Jimenez, J.; Cailleau-Audouin, E.; Bordeau, T.; Berri, C. Slow and Fast-Growing Chickens Use Different Antioxidant Pathways to Maintain Their Redox Balance during Postnatal Growth. Animals 2023, 13, 1160. [Google Scholar] [CrossRef]
- Jung, S.; Bae, Y.S.; Kim, H.J.; Jayasena, D.D.; Lee, J.H.; Park, H.B.; Heo, K.N.; Jo, C. Carnosine, Anserine, Creatine, and Inosine 5′-Monophosphate Contents in Breast and Thigh Meats from 5 Lines of Korean Native Chicken. Poultry Science 2013, 92, 3275–3282. [Google Scholar] [CrossRef]
- Banerjee, R.; Verma, A.K.; Siddiqui, M.W. Applications in Foods of Animal Origin; Apple Academic Press: New York, NY, USA, 2017; ISBN 978-1-315-36591-6. [Google Scholar]
- Domínguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.; Lorenzo, J.M. A Comprehensive Review on Lipid Oxidation in Meat and Meat Products. Antioxidants 2019, 8, 429. [Google Scholar] [CrossRef] [PubMed]
- Charoensin, S.; Laopaiboon, B.; Boonkum, W.; Phetcharaburanin, J.; Villareal, M.O.; Isoda, H.; Duangjinda, M. Thai Native Chicken as a Potential Functional Meat Source Rich in Anserine, Anserine/Carnosine, and Antioxidant Substances. Animals 2021, 11, 902. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Zhuang, H.; Zhou, G.; Zhang, J. Investigation of Inhibition of Lipid Oxidation by L-Carnosine Using an Oxidized-Myoglobin-Mediated Washed Fish Muscle System. LWT 2018, 97, 703–710. [Google Scholar] [CrossRef]
- Gorelik, S.; Kanner, J.; Schurr, D.; Kohen, R. A Rational Approach to Prevent Postprandial Modification of LDL by Dietary Polyphenols. Journal of Functional Foods 2013, 5, 163–169. [Google Scholar] [CrossRef]
- Van Hecke, T.; Goethals, S.; Vossen, E.; De Smet, S. Long-Chain n-3 PUFA Content and n-6/n-3 PUFA Ratio in Mammal, Poultry, and Fish Muscles Largely Explain Differential Protein and Lipid Oxidation Profiles Following In Vitro Gastrointestinal Digestion. Molecular Nutrition Food Res 2019, 63, 1900404. [Google Scholar] [CrossRef]
- Kanner, J.; Gorelik, S.; Roman, S.; Kohen, R. Protection by Polyphenols of Postprandial Human Plasma and Low-Density Lipoprotein Modification: The Stomach as a Bioreactor. J. Agric. Food Chem. 2012, 60, 8790–8796. [Google Scholar] [CrossRef]
- Kenmogne-Domguia, H.B.; Meynier, A.; Boulanger, C.; Genot, C. Lipid Oxidation in Food Emulsions Under Gastrointestinal-Simulated Conditions: The Key Role of Endogenous Tocopherols and Initiator. Food Dig. 2012, 3, 46–52. [Google Scholar] [CrossRef]
- Larsson, K.; Cavonius, L.; Alminger, M.; Undeland, I. Oxidation of Cod Liver Oil during Gastrointestinal in Vitro Digestion. J. Agric. Food Chem. 2012, 60, 7556–7564. [Google Scholar] [CrossRef]
- Kenmogne-Domguia, H.B.; Moisan, S.; Viau, M.; Genot, C.; Meynier, A. The Initial Characteristics of Marine Oil Emulsions and the Composition of the Media Inflect Lipid Oxidation during in Vitro Gastrointestinal Digestion. Food Chemistry 2014, 152, 146–154. [Google Scholar] [CrossRef]
- Mahecha, L.; Dannenberger, D.; Nuernberg, K.; Nuernberg, G.; Hagemann, E.; Martin, J. Relationship between Lipid Peroxidation and Antioxidant Status in the Muscle of German Holstein Bulls Fed n -3 and n -6 PUFA-Enriched Diets. J. Agric. Food Chem. 2010, 58, 8407–8413. [Google Scholar] [CrossRef]
- Van Hecke, T.; Wouters, A.; Rombouts, C.; Izzati, T.; Berardo, A.; Vossen, E.; Claeys, E.; Van Camp, J.; Raes, K.; Vanhaecke, L.; et al. Reducing Compounds Equivocally Influence Oxidation during Digestion of a High-Fat Beef Product, Which Promotes Cytotoxicity in Colorectal Carcinoma Cell Lines. J. Agric. Food Chem. 2016, 64, 1600–1609. [Google Scholar] [CrossRef]
- Barski, O.A.; Xie, Z.; Baba, S.P.; Sithu, S.D.; Agarwal, A.; Cai, J.; Bhatnagar, A.; Srivastava, S. Dietary Carnosine Prevents Early Atherosclerotic Lesion Formation in Apolipoprotein E–Null Mice. ATVB 2013, 33, 1162–1170. [Google Scholar] [CrossRef]
- Li, Y.Y.; Yaylayan, V.; Palin, M.; Ngapo, T.M.; Cliche, S.; Sabik, H.; Gariépy, C. Dual Effects of Dietary Carnosine during in Vitro Digestion of a Western Meal Model with Added Ascorbic Acid. Journal of Food Science 2024, 89, 710–726. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Yaylayan, V.; Palin, M.; Sullivan, B.; Fortin, F.; Cliche, S.; Sabik, H.; Gariépy, C. Protective Effects of Dietary Carnosine during In-vitro Digestion of Pork Differing in Fat Content and Cooking Conditions. J. Food Biochem. 2021, 45, e13624. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, S.; Ahn, D.U. Protein Oxidation: Basic Principles and Implications for Meat Quality. Critical Reviews in Food Science and Nutrition 2013, 53, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Santé-Lhoutellier, V.; Engel, E.; Aubry, L.; Gatellier, P. Effect of Animal (Lamb) Diet and Meat Storage on Myofibrillar Protein Oxidation and in Vitro Digestibility. Meat Science 2008, 79, 777–783. [Google Scholar] [CrossRef]
- Soglia, F.; Laghi, L.; Canonico, L.; Cavani, C.; Petracci, M. Functional Property Issues in Broiler Breast Meat Related to Emerging Muscle Abnormalities. Food Research International 2016, 89, 1071–1076. [Google Scholar] [CrossRef]
- Thanatsang, K.V.; Malila, Y.; Arayamethakorn, S.; Srimarut, Y.; Tatiyaborworntham, N.; Uengwetwanit, T.; Panya, A.; Rungrassamee, W.; Visessanguan, W. Nutritional Properties and Oxidative Indices of Broiler Breast Meat Affected by Wooden Breast Abnormality. Animals 2020, 10, 2272. [Google Scholar] [CrossRef] [PubMed]
- Abasht, B.; Mutryn, M.F.; Michalek, R.D.; Lee, W.R. Oxidative Stress and Metabolic Perturbations in Wooden Breast Disorder in Chickens. PLoS ONE 2016, 11, e0153750. [Google Scholar] [CrossRef]
- Xing, T.; Zhao, X.; Xu, X.; Li, J.; Zhang, L.; Gao, F. Physiochemical Properties, Protein and Metabolite Profiles of Muscle Exudate of Chicken Meat Affected by Wooden Breast Myopathy. Food Chemistry 2020, 316, 126271. [Google Scholar] [CrossRef]
- Sihvo, H.-K.; Airas, N.; Lindén, J.; Puolanne, E. Pectoral Vessel Density and Early Ultrastructural Changes in Broiler Chicken Wooden Breast Myopathy. Journal of Comparative Pathology 2018, 161, 1–10. [Google Scholar] [CrossRef]
- Li, B.; Dong, X.; Puolanne, E.; Ertbjerg, P. Effect of Wooden Breast Degree on Lipid and Protein Oxidation and Citrate Synthase Activity of Chicken Pectoralis Major Muscle. LWT 2022, 154, 112884. [Google Scholar] [CrossRef]
- Estévez, M. Oxidative Damage to Poultry: From Farm to Fork. Poultry Science 2015, 94, 1368–1378. [Google Scholar] [CrossRef]
- Carvalho, L.M.; Rocha, T.C.; Delgado, J.; Díaz-Velasco, S.; Madruga, M.S.; Estévez, M. Deciphering the Underlying Mechanisms of the Oxidative Perturbations and Impaired Meat Quality in Wooden Breast Myopathy by Label-Free Quantitative MS-Based Proteomics. Food Chemistry 2023, 423, 136314. [Google Scholar] [CrossRef]
- Prache, S.; Lebret, B.; Baéza, E.; Martin, B.; Gautron, J.; Feidt, C.; Médale, F.; Corraze, G.; Raulet, M.; Lefèvre, F.; et al. Review: Quality and Authentication of Organic Animal Products in Europe. Animal 2022, 16, 100405. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxidative Medicine and Cellular Longevity 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of production system (s), meat type (m), and their interaction term (s×m) on heme-Fe, α-tocopherol, carnosine and anserine in conventional breast (CON BR), organic breast (ORG BR), conventional thigh (CON TH) and organic thigh (ORG TH) in Experiment 1 (n=10 each). Means with different superscripts are significantly different. P-values for the main effects of production system (Ps), meat type (Pm), and their interaction (Ps×m) were derived from an ANOVA model.
Figure 1.
Effect of production system (s), meat type (m), and their interaction term (s×m) on heme-Fe, α-tocopherol, carnosine and anserine in conventional breast (CON BR), organic breast (ORG BR), conventional thigh (CON TH) and organic thigh (ORG TH) in Experiment 1 (n=10 each). Means with different superscripts are significantly different. P-values for the main effects of production system (Ps), meat type (Pm), and their interaction (Ps×m) were derived from an ANOVA model.
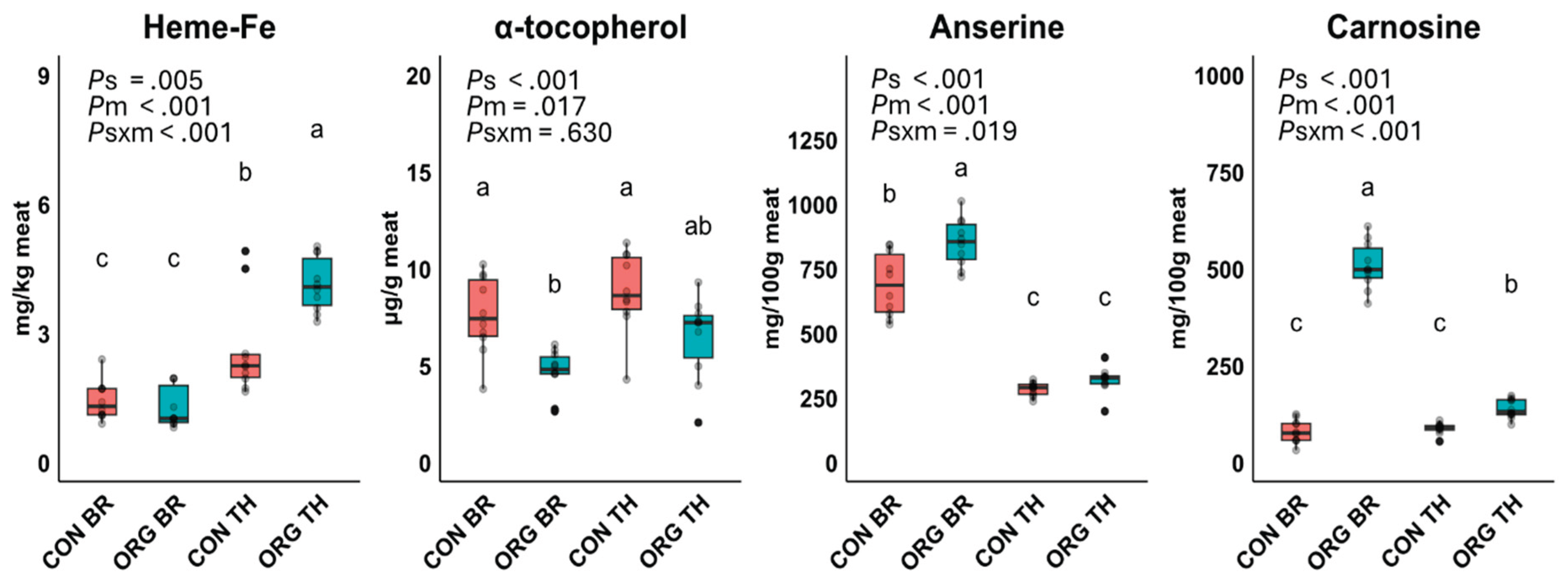
Figure 2.
Effect of production system (s), meat type (m), and their interaction term (s×m) on products of lipid oxidation (4-hydroxy-2-nonenal, 4-HNE; hexanal, HEX; propanal, PROP; thiobarbituric acid reactive substances, TBARS) and protein oxidation (protein carbonyl compounds, PCC) in meats (M) and digests (D) of conventional breast (CON BR), organic breast (ORG BR), conventional thigh (CON TH) and organic thigh (ORG TH) in Experiment 1 (n=10 each. Means with different superscripts are significantly different. P-values for the main effects of production system (Ps), meat type (Pm), and their interaction (Ps×m) were derived from an ANOVA model.
Figure 2.
Effect of production system (s), meat type (m), and their interaction term (s×m) on products of lipid oxidation (4-hydroxy-2-nonenal, 4-HNE; hexanal, HEX; propanal, PROP; thiobarbituric acid reactive substances, TBARS) and protein oxidation (protein carbonyl compounds, PCC) in meats (M) and digests (D) of conventional breast (CON BR), organic breast (ORG BR), conventional thigh (CON TH) and organic thigh (ORG TH) in Experiment 1 (n=10 each. Means with different superscripts are significantly different. P-values for the main effects of production system (Ps), meat type (Pm), and their interaction (Ps×m) were derived from an ANOVA model.
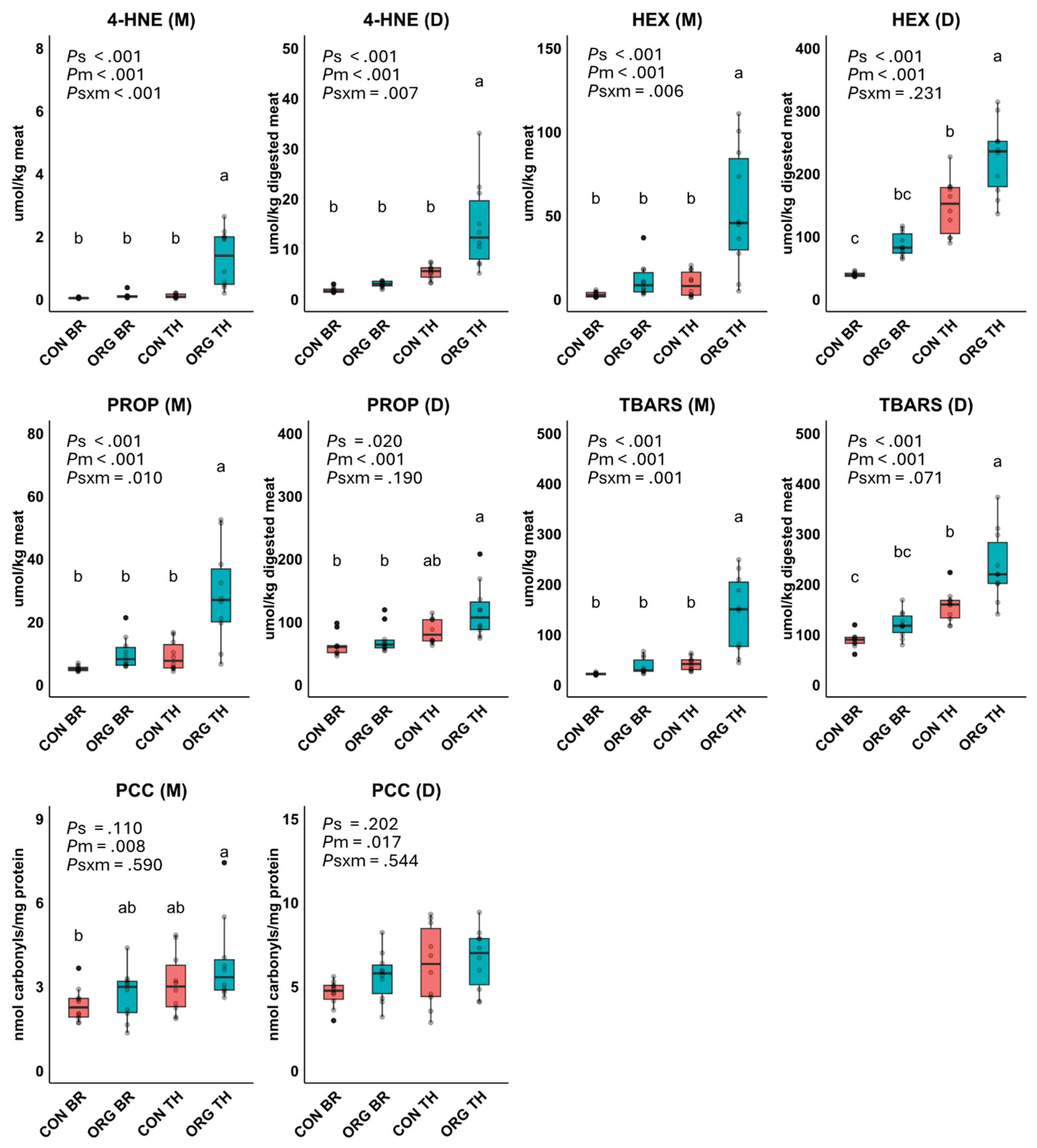
Figure 3.
Principal component analysis (PCA) biplot from Experiment 1, showing the distribution of oxidation products (4-hydroxy-2-nonenal, 4-HNE; hexanal, HEX; propanal, PROP; thiobarbituric acid reactive substances, TBARS; protein carbonyl compounds, PCC) in meats (M) and digests (D), as well as meat composition variables (n-6 PUFA, n-3 PUFA, total FA, heme-Fe, α-tocopherol, anserine, carnosine) across experimental groups: conventional breast (CON BR), conventional thigh (CON TH), organic breast (ORG BR) and organic thigh (ORG TH). The ellipse around each group represents the 95% confidence interval, highlighting the separation between groups. The percentage of variance explained by each principal component (PC1 and PC2) is indicated in parentheses.
Figure 3.
Principal component analysis (PCA) biplot from Experiment 1, showing the distribution of oxidation products (4-hydroxy-2-nonenal, 4-HNE; hexanal, HEX; propanal, PROP; thiobarbituric acid reactive substances, TBARS; protein carbonyl compounds, PCC) in meats (M) and digests (D), as well as meat composition variables (n-6 PUFA, n-3 PUFA, total FA, heme-Fe, α-tocopherol, anserine, carnosine) across experimental groups: conventional breast (CON BR), conventional thigh (CON TH), organic breast (ORG BR) and organic thigh (ORG TH). The ellipse around each group represents the 95% confidence interval, highlighting the separation between groups. The percentage of variance explained by each principal component (PC1 and PC2) is indicated in parentheses.
Figure 4.
Effect of Normal Breast (NB; n=10) and Wooden Breast (WB; n=10) phenotype on heme-Fe, α-tocopherol, carnosine and anserine in meats in Experiment 2. P-values were based on t-test.
Figure 4.
Effect of Normal Breast (NB; n=10) and Wooden Breast (WB; n=10) phenotype on heme-Fe, α-tocopherol, carnosine and anserine in meats in Experiment 2. P-values were based on t-test.
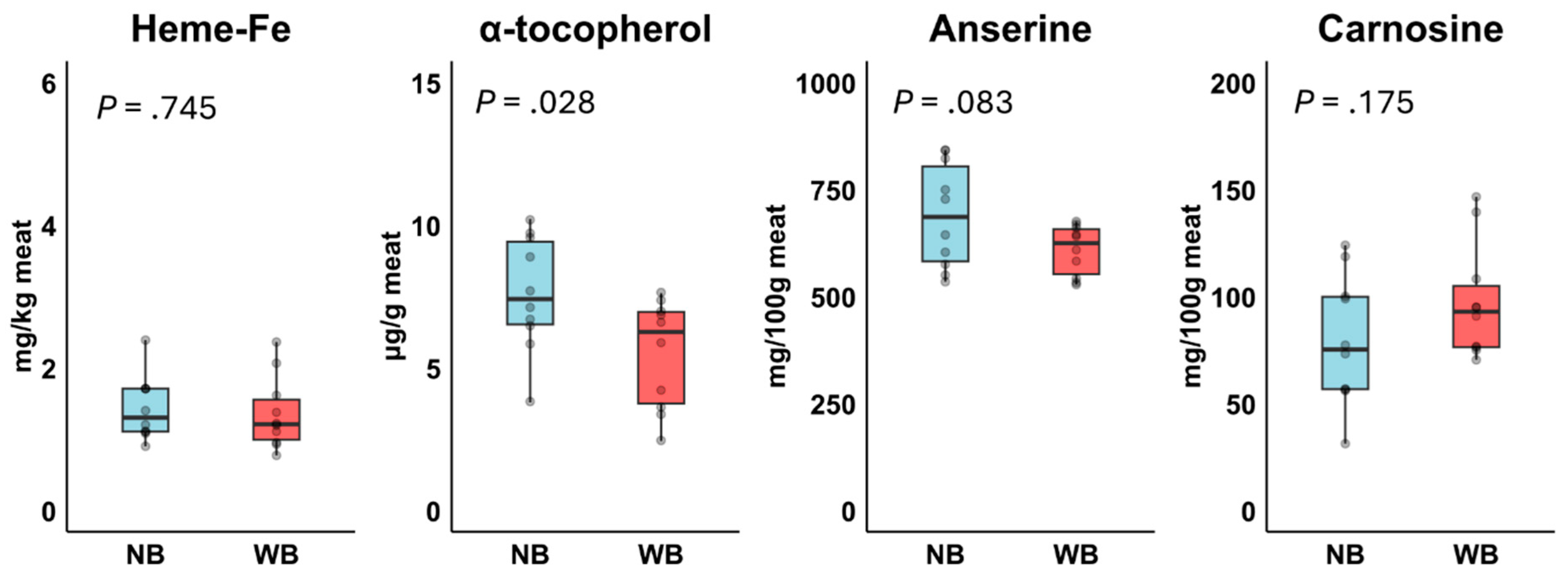
Figure 5.
Effect of Normal Breast (NB; n=10) and Wooden Breast (WB; n=10) phenotype on products of lipid oxidation (4-hydroxy-2-nonenal, 4-HNE; hexanal, HEX; propanal, PROP, thiobarbituric acid reactive substances, TBARS) and protein oxidation (protein carbonyl compounds, PCC) in meats (M) and digests (D) in Experiment 2. P-values were based on t-test.
Figure 5.
Effect of Normal Breast (NB; n=10) and Wooden Breast (WB; n=10) phenotype on products of lipid oxidation (4-hydroxy-2-nonenal, 4-HNE; hexanal, HEX; propanal, PROP, thiobarbituric acid reactive substances, TBARS) and protein oxidation (protein carbonyl compounds, PCC) in meats (M) and digests (D) in Experiment 2. P-values were based on t-test.
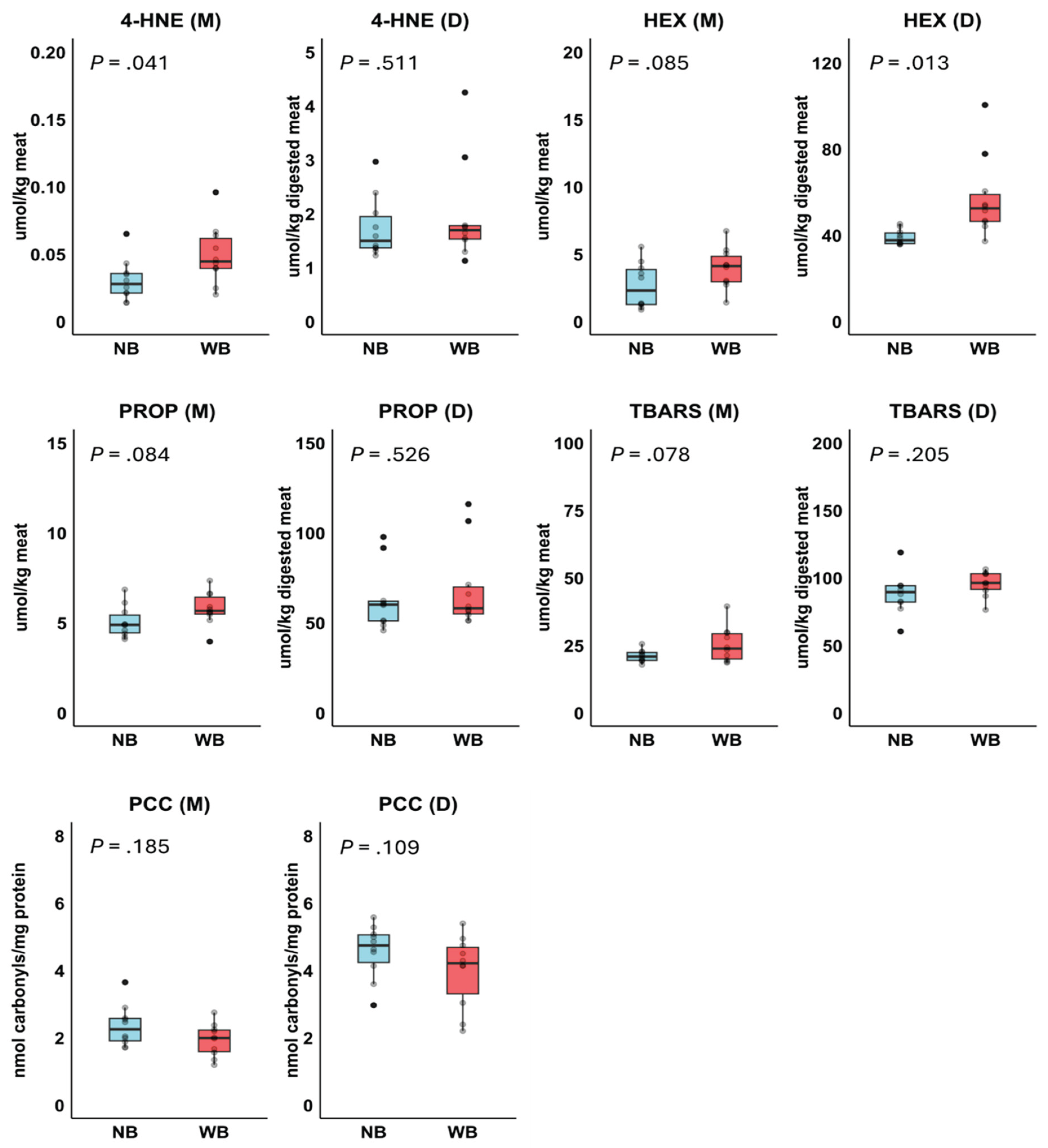

Table 1.
Characteristics and chemical composition of the finisher phase diets in the conventional and organic production system.
Table 1.
Characteristics and chemical composition of the finisher phase diets in the conventional and organic production system.
| Production system | ||
|---|---|---|
| Characteristics | Conventional | Organic |
| Breed | Ross 308 | Sasso (Ruby XL) |
| Age at slaughter (days) | 39.00–42.00 | 73.00–76.00 |
| Finishing phase (days) | 14.00 | 21.00 |
| Live weight at slaughter (kg) | 2.63–2.85 | 2.30–2.50 |
| Stocking density (kg/m2) | 42.00 | 21.00 |
| Outdoor access | No | Yes |
| Indoor enrichments | No | Yes |
| Diet composition (finisher phase) | ||
| Protein % | 21.33 | 20.56 |
| Fat % | 7.40 | 4.00 |
| Ash % | 5.60 | 5.40 |
| Fiber % | 2.60 | 2.90 |
| Methionine % | 0.75 | 0.30 |
| Lysine % | 1.25 | 0.94 |
| Calcium % | 0.73 | 0.8 |
| Phosphorus % | 0.43 | 0.60 |
| Na % | 0.16 | 0.20 |
| Vitamin A (IU/kg) | 13200.00 | 10000.00 |
| Vitamin D3 (IU/kg) | 3630.00 | 3000.00 |
| Vitamin E (mg/kg) | 50.00 | 55.00 |
| Se (mg/kg) | 0.52 | 0.30 |
| Iron (mg/kg) | 19.80 | 70.00 |
| Zn (mg/kg) | 66.00 | 70.00 |
| Cu (mg/kg) | 19.84 | 15.40 |
Table 2.
Fatty acid profile of heated breast and thigh meats from conventional (CON) and organic (ORG) production system used in the in vitro gastrointestinal digestion of Experiment 1.
Table 2.
Fatty acid profile of heated breast and thigh meats from conventional (CON) and organic (ORG) production system used in the in vitro gastrointestinal digestion of Experiment 1.
| Trait | Unit | Breast | Thigh | P-value | |||||
|---|---|---|---|---|---|---|---|---|---|
| CON | ORG | CON | ORG | RMSE | S | M | S×M | ||
| Total FA | g/100g | 1.06 ± 0.23b | 0.77 ± 0.07b | 2.79 ± 0.80a | 3.05 ± 0.62a | 0.49 | 0.921 | <0.001 | 0.102 |
| SFA | mg/100g | 343.10 ± 80.17b | 243.55 ± 23.79b | 896.28 ± 261.54a | 875.34 ± 187.78a | 158.54 | 0.263 | <0.001 | 0.458 |
| MUFA | mg/100g | 316.06 ± 91.00b | 183.01± 35.06b | 906.07 ± 286.64a | 878.45 ± 234.79a | 182.34 | 0.191 | <0.001 | 0.398 |
| PUFA | mg/100g | 267.45 ± 51.32c | 234.65 ± 24.43c | 684.95 ± 193.32b | 980.76 ± 164.29a | 123.09 | 0.002 | <0.001 | <0.001 |
| n-3 PUFA | mg/100g | 22.74 ± 5.63c | 21.80 ± 2.55c | 56.94 ± 16.37b | 85.54 ± 17.07a | 11.60 | <0.001 | <0.001 | <0.001 |
| ALA | mg/100g | 10.67 ± 4.02c | 7.68 ± 1.19c | 41.90 ± 13.65b | 63.42 ± 16.74a | 10.44 | 0.011 | <0.001 | 0.001 |
| EPA | mg/100g | 2.21 ± 0.87ab | 1.39 ± 0.45b | 2.68 ± 1.27a | 2.28 ± 0.64ab | 0.81 | 0.031 | 0.017 | 0.447 |
| DHA | mg/100g | 2.84 ± 0.88c | 4.94 ± 1.15b | 3.42 ± 1.18c | 7.52 ± 1.39a | 1.10 | <0.001 | <0.001 | 0.010 |
| n-6 PUFA | mg/100g | 244.68 ± 49.77c | 213.75 ± 22.26c | 627.23 ±184.65b | 894.48 ± 148.52a | 115.65 | 0.004 | <0.001 | <0.001 |
| LA | mg/100g | 197.10 ± 46.70c | 157.42 ± 18.9c | 565.42 ± 176.54b | 803.28 ± 145.32a | 111.42 | 0.011 | <0.001 | <0.001 |
| AA | mg/100g | 33.17 ± 3.70c | 45.30 ± 4.20b | 44.93 ± 9.38b | 71.62 ± 10.13a | 7.08 | <0.001 | <0.001 | 0.004 |
| n-6/n-3 PUFA | ratio | 11.35 ± 3.79 | 9.82 ± 0.69 | 11.53 ± 3.20 | 10.54 ± 0.71 | 2.40 | 0.126 | 0.596 | 0.741 |
FA = fatty acid; SFA = saturated fatty acids; MUFA = monounsaturated fatty acids; PUFA = polyunsaturated fatty acids (n-3 PUFA + n-6 PUFA); n-3 PUFA = omega-3 PUFA (C18:3n-3; C20:3n-3; C20:4n-3; C20:5n-3; C22:5n-3; C22:6n-3); ALA = α-linolenic acid (C18:3n-3); EPA = eicosapentaenoic acid (C20:5n-3); DHA = docosahexaenoic acid (C22:6n-3); n-6 PUFA = omega-6 PUFA (C18:2n-6; C18:3n-6; C20:2n-6; C20:3n-6; C20:4n-6; C22:4n-6; C22:5n-6); LA = linoleic acid (C18:2n-6); AA = arachidonic acid (C20:4n-6); n-6/n-3 PUFA = ratio of n-6 PUFA to n-3 PUFA. Values are means ± standard deviation (n=10). Means with different superscripts are significantly different. P-values of the main effects of production system (S), meat type (M), and their interaction (S×M) were derived from an ANOVA model. RMSE = root mean square error.
Table 3.
Fatty acid profile of heated Normal Breast and Wooden Breast meats used in the in vitro gastrointestinal digestion of Experiment 2.
Table 3.
Fatty acid profile of heated Normal Breast and Wooden Breast meats used in the in vitro gastrointestinal digestion of Experiment 2.
| Trait | Unit | Normal Breast | Wooden Breast | RMSE | P-value |
|---|---|---|---|---|---|
| Total FA | g/100g | 1.06 ± 0.23 | 1.42 ± 0.70 | 0.49 | 0.156 |
| SFA | mg/100g | 343.10 ± 80.17 | 476.43 ± 244.78 | 173.34 | 0.131 |
| MUFA | mg/100g | 316.06 ± 91.00 | 430.10 ± 233.32 | 167.43 | 0.173 |
| PUFA | mg/100g | 267.45 ± 51.32 | 353.12 ± 162.57 | 114.78 | 0.140 |
| n-3 PUFA | mg/100g | 22.74 ± 5.63 | 32.91 ± 17.06 | 12.27 | 0.090 |
| ALA | mg/100g | 10.67 ± 4.02 | 21.70 ± 14.83 | 10.23 | 0.043 |
| EPA | mg/100g | 2.21 ± 0.87 | 2.15 ± 0.81 | 0.79 | 0.878 |
| DHA | mg/100g | 2.84 ± 0.88 | 2.63 ± 0.78 | 0.78 | 0.592 |
| n-6 PUFA | mg/100g | 244.68 ± 49.77 | 320.34 ± 146.45 | 103.46 | 0.149 |
| LA | mg/100g | 197.10 ± 46.70 | 277.17 ± 137.60 | 96.94 | 0.110 |
FA = fatty acid; SFA = saturated fatty acids; MUFA = monounsaturated fatty acids; PUFA = polyunsaturated fatty acids (n-3 PUFA + n-6 PUFA); n-3 PUFA = omega-3 PUFA (C18:3n-3; C20:3n-3; C20:4n-3; C20:5n-3; C22:5n-3; C22:6n-3); ALA = α-linolenic acid (C18:3n-3); EPA = eicosapentaenoic acid (C20:5n-3); DHA = docosahexaenoic acid (C22:6n-3); n-6 PUFA = omega-6 PUFA (C18:2n-6; C18:3n-6; C20:2n-6; C20:3n-6; C20:4n-6; C22:4n-6; C22:5n-6); LA = linoleic acid (C18:2n-6); AA = arachidonic acid (C20:4n-6); n-6/n-3 PUFA = ratio of n-6 PUFA to n-3 PUFA. Values are means ± standard deviation (n=10). P-values were determined using t-test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



