
Submitted:
29 October 2024
Posted:
31 October 2024
You are already at the latest version
Abstract
In this study, broilers were selected as the research object to investigate the effects and mechanism action of dietary gallic acid (GA) supplementation on growth performance, meat quality, antioxidant capacity, and muscle fiber-related gene expression. A total of 750 one-day-old healthy 817 male crossbred broiler chickens were divided into five treatment groups, with six replicates per group: control group (CON), basal diet, basal diet with lipopolysaccharide challenge (LPS), basal diet with 150, 300, 450 mg/kg GA supplementation and LPS challenge (GA150, GA300, GA450). On days 14, 17, and 20, chickens in the LPS, GA150, GA300, and GA450 groups received intramuscular injections of LPS, while chickens in the CON group received saline. Compared with the LPS group, adding GA (GA150 and GA300) significantly increased the average daily gain (ADG) of broiler chickens from 1 to 50 days old (p < 0.05), and had the tendency to increase average daily feed intake (ADFI) (p = 0.078). Dietary addition of 450 mg/kg GA significantly reduced the pH value of breast muscle at 45min after slaughter (p < 0.05), and as the level of GA addition increased, the pH value first decreased and then increased. Compared with the LPS group, adding 450mg/kg GA significantly increased the brightness of breast muscles at 45min and 24h after slaughter (p < 0.001). Dietary supplementation of 450 mg/kg GA significantly decreased the total volatile basic nitrogen (TVB-N) content of post-mortem breast muscle (p < 0.05). Adding 450 mg/kg GA could significantly increase C22:6n-3 concentration in breast muscle (p < 0.05). Different GA levels had no significant effect on the intramuscular fat (IMF) content of broiler breast muscle (p > 0.05). Adding GA (GA150 and GA300) significantly increased (p < 0.05) the total superoxide dismutase (T-SOD) activity in the breast muscle after LPS stimulation, significantly increased (p < 0.05) the catalase (CAT) activity, and significantly reduced (p < 0.05) malonic dialdehyde (MDA) production, and had the tendency to increase glutathione peroxidase (GSH-Px) activity (p = 0.066). In addition, dietary supplementation of GA significantly increased the expression levels of myosin heavy chain (MyHC) I and MyHC IIa in breast muscles (p < 0.05), and significantly decreased the expression level of MyHC IIx (p < 0.05). In summary, dietary addition of GA can alleviate the stress response on the growth performance of broiler chickens and improve antioxidant capacity and meat quality. The appropriate amount of dietary GA at each stage was 300 mg/kg.
Keywords:
gallic acid
; broiler
; lipopolysaccharide
; growth performance
; meat quality
; antioxidant capacity
1. Introduction
In poultry industry, antibiotics have been widely used as growth promoters with sub-therapeutic dosage due to their outstanding efficacy in feed conversion and animal growth [1]. However, as people's requirements for animal food safety and health are rapidly increasing, the use of antibiotics as preventive and therapeutic drugs in poultry production has been increasingly questioned. Blind and irregular use of antibiotics will not only lead to the emergence of bacterial resistance and resistant strains but also cause antibiotic residues in chicken meat and eggs, directly endangering the health of consumers [2]. Therefore, the search for feed additives with antimicrobial activity and other effects that improve the general health of animals and reduce the need for antibiotics contributes to the safeguarding of animal welfare and the improvement of livestock productivity, as well as to the improvement of consumer health [3]. Gallic acid (GA), also known as 3,4,5-trihydroxybenzoic acid, can be obtained by acid hydrolysis of tannins [4]. It has been reported that GA effectively inhibited inflammation and oxidation in vitro and in vivo and altered metabolic and bacterial profiles in the colitis model [5]. Their transfer to muscular mass has been hypothesized to enhance the antioxidant properties of meat, thereby improving its nutritional and technological quality [6]. Lipopolysaccharide (LPS), a component of the cell wall of Gram-negative bacteria, can cause severe oxidative stress, induce inflammation, change the distribution of nutrients, reduce growth potential, and induce various diseases, is widely used to build a stress model [7,8,9,10]. The aim of this study was to elucidate the effects of dietary GA-plant extract on growth performance, muscle quality and antioxidant capacity of broiler chickens after LPS stimulation, and to provide a basis for the application of its products in broiler production.
2. Materials and Methods
2.1. Animal and Experimental Design
The experimental protocol was approved by the Animal Care Committee of the Institute of Animal Science, Guangdong Academy of Agriculture Science, Guangzhou, P. R. China, with the approval number GAASISA-2022-003.
A total of 750 1-d male 817 crossbred broilers (39.50 g ± 0.20 g) were randomly assigned to 5 treatment groups (Table 1), each with 6 replicates of 25 broilers. Birds in the control group (CON) and LPS-challenged treatment (LPS) were fed a basal diet, and birds in the other 3 treatments received the basal diet with 150, 300, or 450 mg/kg added GA (GA150, GA300, GA450). On days 14, 17, and 20 of the trial, birds in the LPS, GA150, GA300, and GA450 treatments were injected intraperitoneally with 0.5 mL of LPS (0.5 mg/kg body weight), while birds in CON received an equal amount of normal saline.
GA (99.6% purity) was obtained from Wufeng Chicheng Biotechnology Co., Ltd (Yichang, Hubei, China). LPS from Escherichia coli serotype O55:B5 was purchased (L4005, Sigma-Aldrich Chemical Co. Ltd, St. Louis, MO).
2.2. Diets and Chicken Husbandry
The basic diet (Table 2) is formulated based on the nutritional requirements of fast-growing yellow-feathered broilers in China's "National Agricultural Industry Standard Nutrient Requirements for Yellow-feathered Broilers" (NY/T3645-2020) [11].
The 50-day experiment was carried out in the testing farm of the Institute of Animal Science, Guangdong Academy of Agricultural Sciences. Broilers were raised in floor pens and had ad libitum access to diets and water. Room temperature was maintained at 27 to 30 °C. For the first 3 d, the light was kept constant for 23 h per d, and then reduced by 2 h per d and maintained at 16 h per d.
2.3. Measurements of Growth Performance
The BW of the birds was measured at 1 and 50 d of age on a pen basis and feed intake was recorded daily. Mortality was checked daily, and dead birds were recorded and weighed to adjust estimates of gain, intake, and feed conversion ratio, as appropriate. The final body weight (FW), average daily feed intake (ADFI), average daily gain (ADG), and Feed-to-gain ratio (F: G) were calculated.
2.4. Sample Collection
On the 50th day of the experiment, two experimental chickens with a weight close to the average were selected from each treatment group for slaughter. The left breast muscle was taken to measure breast muscle color, pH value, cooking loss, drip loss, shear force, crude fat and other indicators. The right breast muscle specimen was taken and stored in a -80 °C ultra-low temperature refrigerator to detect the antioxidant index, shelf life index and related gene expression.
2.5. Determination of Meat Quality
The brightness (L*), redness (a*), and yellowness (b*) value of slaughtered pectoral muscles were measured by a colorimeter (CR-410, Minolta, Japan) after 45 min and 24 h at room temperature. Each sample was measured three times and the average value was taken. The pH of the slaughtered pectoral muscles was determined using a muscle pH meter (HI8242, HANNA, Italy) after 45 min and 24 h of standing at room temperature, with each sample measured three times and the average value taken. The drip loss was measured according to the procedure reported by Jin et al. [12]. Briefly, breast muscle chunks (size 2 × 3 × 4 cm) were weighed, and suspended in a cup at 4 °C for 24 h. Subsequently, the weight was recorded to calculate drip loss.
2.6. Determination of Intramuscular Fat and Total Volatile Base Nitrogen Contents
Approximately 20 g of pectoral muscle samples were weighed, the fascia was removed, minced, placed in a Petri dish, and freeze-dried at -80 °C for 48 h. Accurately take 2 g of freeze-dried sample seal it in a filter bag and extract it through a Soxhlet extraction device (XT15i, USA). After extraction, the filter paper tube was removed and the fat bottle was placed back into a constant temperature oven at 105 °C for 3.5 h, then removed, cooled and weighed to calculate the intramuscular fat (IMF) content. The total volatile base nitrogen (TVB-N) values were measured according to the method proposed by Chen et al. [13], with results expressed in milligrams of TVB-N per 100 g of sample.
2.7. Determination of Pectoral Muscle Fatty Acid Composition
Weigh about 1 g of pectoral muscle tissue sample and homogenize. Add 2 mL of n-hexane and shake at 50 °C for 30 min, add 3 mL of KOH methanol solution (0.4 mol/L) and shake at 50 °C for 30 min, then add 1 mL of water and 2 mL of n-hexane and mix well. The upper layer was separated, and the composition and proportion of fatty acids were determined by gas chromatography-mass spectrometry (GC-MS) using an Agilent 7890B-5977A gas chromatography-mass spectrometer (GC-MS).
2.8. Determination of Pectoral Muscle Antioxidant Index
Weighing approximately 1 g of broiler breast muscle samples, the tissue was then cut into pieces and homogenized to a concentration of 10% in normal saline at a ratio of 1:9 (sample: normal saline). Moreover, the commercially available reagent kits of the malonic dialdehyde (MDA, A003-1-2) content, total superoxide dismutase (T-SOD, A001-1-2), catalase (CAT, A007-1-1), glutathione peroxidase (GSH-Px, A005-1-2) and total anti-oxidative capacity(T-AOC, A015-1-2)were used to determine the concentration of MDA and the activities of antioxidant enzyme in muscle following the manufacturer’s instructions (Nanjing Jiancheng Biotech Limited., Nanjing, China).
2.9. Expression of Related Genes in Pectoral Muscle
Total RNA was extracted from jejunal mucosa using RNAiso plus (9109, TAKARA, Tokyo, JP) and reverse transcribed with the PrimeScript II 1st Strand cDNA Synthesis Kit (6210A, TAKARA). Real-time PCR was performed with SYBR PremixExTaq II (RR820A, TAKARA) and the real-time PCR system (ABI 7500, Applied Biosystems, Carlsbad, CA). The primers used are shown in Table 3. Results were normalized to the abundance of β-actin transcripts and relative quantification was calculated using the 2–△△CT method.
2.10. Statistical Analysis
A pen or individual bird was taken as the experimental unit. The effects of treatment were examined by multivariate analysis of variance (MANOVA) using SPSS 16.0 for Windows (SPSS Inc., Chicago, IL). The treatment means were compared by Duncan’s multiple range tests at P < 0.05 significance levels. Tabulated results were shown as means with standard error of mean (SEM).
3. Results
3.1. Growth Performance
As presented in Table 4, dietary supplementation with GA significantly increased the body weight of broiler chickens (p < 0.05), but had no significant effect on the F:G of broiler chickens (p > 0.05). Compared to the LPS-treated birds, supplementation with 150 and 300 mg/kg GA significantly increased the ADG (p < 0.05) and had a tendency to increase ADFI (p = 0.078).
3.2. Meat Quality
As presented in Table 5, dietary 450 mg/kg GA supplementation significantly decreased the pH of breast muscle at 45 min after slaughter compared to the CON group (p < 0.05), with a tendency of decreasing and then increasing with the increase of GA supplementation, and was the lowest at the addition of 300 mg/kg (p = 0.07). The addition of 450 mg/kg GA significantly increased the L* value of the breast muscle at 45 min after slaughter compared with the CON and LPS groups (p < 0.001). The addition of 450 mg/kg GA significantly (p < 0.05) decreased the volatile saline nitrogen content in the post-slaughter breast muscle compared to the CON group. Different GA levels did not significantly (p > 0.05) affect the 24h pH, a*, b*, and intramuscular fat content of breast muscle.
3.3. Intramuscular Fat and Total volatile Basic Nitrogen Contents
As presented in Table 6, different GA levels had no significant effect on the IMF content of breast muscles (p > 0.05). Compared with the CON group, adding 450 mg/kg GA significantly reduced the TVB-N content in postmortem breast muscle (p < 0.05).
3.4. Muscle Fatty Acid Composition
As presented in Table 7, the addition of GA had a trend effect (p = 0.063) on the content of C16:0 in the breast muscle of broilers after slaughter. 450 mg/kg GA could significantly increase C22:6n-3 (docosahexaenoic acid, DHA) concentration in breast meat(p < 0.05). Different GA levels did not (p > 0.05) affect other fatty acids, total saturated fatty acids (SFA), total polyunsaturated fatty acids (PUFA) and n-3/n-6 unsaturated fatty acid ratio in muscle.
3.5. Muscle Antioxidative Enzyme Activities and MDA Content
As presented in Table 8, the level of GA supplementation significantly (p < 0.05) affected T-SOD, CAT, GSH-Px activities and MDA content in breast muscle after slaughter. Compared with the LPS group, supplementation with 150 and 300 mg/kg GA significantly increased (p < 0.05) T-SOD activity, supplementation with 150 mg/kg GA significantly increased (p < 0.05) CAT activity, and supplementation with 300 and 450 mg/kg GA significantly decreased (p < 0.05) MDA content in breast muscle, and supplementation with 150, 300 and 450 mg/kg GA had a tendency to increase GSH-Px activity (p = 0.066). Neither LPS stimulation and supplementation with GA had no significant effect on T-AOC level (p > 0.05).
3.6. Muscle MyHC mRNA Expression
As presented in Figure 1, GA addition level significantly (p < 0.05) affected the gene expression of myosin heavy chain genes MyHC I, MyHC IIa and MyHC IIx in broiler breast muscle after slaughter. The addition of 300 mg/kg GA significantly (p < 0.05) increased the expression of MyHC I, and MyHC IIa in broiler breast muscle compared to the LPS group. The addition of 150, 300, and 450 mg/kg GA significantly decreased the expression of MyHC IIx compared to the LPS group (p < 0.05). Neither LPS stimulation nor the addition of GA had an effect on the expression of MyHC IIb (p > 0.05).
4. Discussion
Growth performance is one of the most important indicators to evaluate whether an additive is effective or not. While LPS stimulation, as a stress model, can change the content of water and electrolytes in the small intestine, causing an imbalance of homeostasis in the intestinal tract of broiler chickens, which can reduce the growth performance of poultry [14]. Zhang et al. injected LPS into the peritoneal cavity of AA chickens at the ages of 16, 18, and 20 days, which significantly reduced the body weight gain of broilers at the ages of 1 to 21 days [15]. This is similar to the results of the present experiment, therefore, alleviating the decline in growth performance caused by LPS stimulation could also reflect the effect of the additive. Samuel et al. increased feed conversion ratios from 21 to 42 days of age and 1 to 42 days of age by adding 75 to 100 mg/kg of GA to 1-day-old AA broiler diets [16]. This was similar to the results of the present experiment, where GA addition increased body weight and decreased FCR in starter stage, finshing stage, and whole stage compared to the LPS-stimulated group. Another study showed that the addition of grape pomace rich in GA by drinking water increased the body weight of broiler chickens by 5.8% and resulted in a cecal microbial composition that was more favorable for energy deposition in broiler chickens, thereby promoting weight gain in broiler chickens [17]. In addition, GA has been shown to have antimicrobial properties, induce changes in the microbiota toward a more favorable composition and activity, promote gastrointestinal epithelial cell proliferation and differentiation, and repair damaged mucosa to improve nutrient digestion and absorption [18,19,20]. Therefore, the positive effects on growth performance shown in this study can be explained by the fact that GA may counteract the impairment of LPS on the digestion and absorption capacity of broiler chickens by suppressing the inflammatory response and using the intestine as a target organ by maintaining its barrier function.
Muscle pH value is an important index to evaluate muscle quality and the decomposition rate of muscle glycogen after poultry slaughter, which is mainly affected by phosphofructokinase activity [21]. The amount of lactic acid produced in muscle is related to the glycogen content in the muscle. The more glycogen reserves in muscle, the more lactic acid is produced during strenuous exercise, the more it accumulates, and the lower of the pH value, which leads to meat acidification. In addition, studies have shown that the pH value of breast muscle is negatively correlated with brightness, yellowness, and shear force [22], and differences in muscle fiber composition, density, and diameter can affect muscle pH [23]. In the present study, the addition of 150 mg/kg GA obviously reduced the pH value of breast muscle 45 min after slaughter. It has been reported that GA added to the diet can be distributed to tissues throughout the body [24], which could be responsible for the decrease in the pH value of muscle tissue. The meat color is one of the main quality attributes that directly influence consumer choice. Fresh chicken breasts are slightly pink, but may also appear white to yellow due to several factors [25]. It has been reported that the color of meat may be influenced by the concentration of myoglobin, its chemical and physical state, and the structure of the meat surface [25,26]. Drip loss is a major indicator of muscle water-holding capacity. Muscle with weak water-holding capacity have accelerated loss of total pigments and soluble nutrients, resulting in impaired meat color and flavor [27]. Muscle brightness was correlated with drip loss, with higher brightness being associated with poorer water-holding capacity and relatively higher drip loss. The results of the present study showed that the addition of 450 mg/kg GA had higher pectoral muscle brightness value compared to the LPS and CON groups. Increased brightness value and drip loss often imply accelerated muscle protein degradation [28], and lower pH reduces the hydrolytic potential of proteins, which reduces the muscle's water-locking capacity, and free water in the muscle seeps out of the surface of the muscle, which leads to an increase in brightness [29].
Oxidative reactions are still going on in the muscle after the animal is slaughtered. The activities of auto-antioxidant enzymes decreases continuously with the prolongation of post-slaughter muscle placement time, and if it decreases to a level that does not effectively reduce ROS, it can lead to muscle quality degradation and the production of toxic and hazardous substances, such as volatile saline nitrogen, thus affecting the shelf life of the product, which is not conducive to the health of the consumer ([30]Onk et al., 2019). Volatile saline nitrogen content, as an indicater of the proteolysis level, increased with storage time, thus accelerating muscle spoilage [31]. The addition of 450 mg/kg GA significantly reduced the volatile saline nitrogen content of pectoral muscle. In addition to its antioxidant function, this may be due to the fact that GA inhibited the growth of Pseudomonas spp. and Enterobacter spp. in muscle, which can use amino acids as a growth substrate to produce sulfur-containing compounds and amines with an off-flavor, resulting in a rapid increase in the volatile base nitrogen content [32,33]. The test results suggest that the addition of GA is beneficial to improve the storage stability of muscle and prolong the shelf life.
Eating foods rich in n-3 polyunsaturated fatty acids (n-3 PUFA) and essential fatty acids can play a natural preventive role in cardiovascular disease and other health problems [34,35]. The nutritional quality of chicken is an important factor affecting the health of consumers. Studies have found that PUFA can be effectively deposited in chicken and improve the quality of chicken [36]. The fatty acid composition of intramuscular fat, on the other hand, has implications for human health, and it is generally recognized that higher PUFA/SFA ratios are less likely to increase the incidence of cardiovascular disease and that PUFA/SFA > 0.45 barely increases the incidence of cardiovascular disease [37]. However, lipid oxidation increases linearly with increasing PUFA content, and the oxidative stability of unsaturated fatty acids (UFA) decreases with increasing levels of unsaturation. omega-3 long-chain unsaturated fatty acids, especially docosahexaenoic acid (DHA, C22:6n-3), are very sensitive to oxidation, and the body's antioxidant capacity has a significant effect on their content [38]. DHA can promote brain and retinal development, inhibit inflammation, and reduce the risk of cardiovascular disease and obesity [39]. Studies have shown that adding basil, thyme, and sage to the diet can significantly increase the healthy ALA (C18:3n-3), EPA (C20:5n-3), DHA, and total PUFA content and improve the fatty acid profile in broiler muscles [40]. In this experiment, the addition of GA not only improved the antioxidant properties of broiler thigh muscle but also enhanced its nutritional value by increasing health-promoting n-3 PUFA. Since there was no change in the unsaturated fatty acid content in the diets of each stage, the higher unsaturated fatty acid content in the thigh meat of broilers supplemented with natural feed additives may be due to the protective effect of antioxidants on the oxidative decomposition of unsaturated fatty acids. Therefore, GA may increase the relative content of DHA in breast muscle through antioxidant action, thus improving the nutritional quality of muscle.
Under physiological conditions, animals produce reactive oxygen species (ROS), which can cause damage to the body [41]. Multiple studies have shown that polyphenols can improve tissue oxidative status by scavenging free radicals and increasing the activity of antioxidant enzymes [42,43,44]. Dietary addition of plant polysaccharides, polyphenols and other bioactive substances can improve the antioxidant capacity of muscle and improve muscle quality [45,46]. GA is a good natural antioxidant that can eliminate ROS such as superoxide anions, hydrogen peroxide, and hydroxyl radicals [47,48]. Ramay et al. have shown that antioxidant enzymes such as CAT, SOD and GSH-Px can work together to reduce MDA content, remove excess ROS, reduce oxidative damage of breast muscle, and thus maintain better muscle mass [45]. In this experiment, the addition of GA to the diet could alleviate the effects of LPS-induced increase in muscle MDA content and decrease in T-SOD, GSH-PX and CAT activities. This is consistent with previous study that GA can improve the antioxidant capacity of broiler chicken muscle [16,49]. In summary, our results further confirmed that GA can effectively exert its antioxidant function to ensure animal health and growth.
Shear force is an indicator of muscle tenderness, which is inversely related to tenderness, and is related to myofiber diameter and fascicle membrane thickness. The myosin heavy chain method of classifying muscle fiber types is based on their ATPase activity and maximum contraction velocity, which classifies myosin heavy chains into four different major isoforms: slow oxidizing (I), fast oxidizing (IIa), fast fermenting (IIb), and intermediate (IIx) [50]. Aerobic endurance exercise in humans and animals effectively induces skeletal muscle type remodeling, i.e., the gradual transformation of skeletal muscle fibers from fast muscle (high proportion of MyHC type IIb fibers) to slow muscle (high proportion of MyHC type I fibers) [51]. This adaptive remodeling process of muscle fiber type is crucial for the body's energy homeostasis, fatigue alleviation, and improvement of livestock and poultry meat quality [52]. Type I fibers contain more mitochondria and aerobic metabolic enzymes such as cytochrome oxidase and succinate dehydrogenase, while type IIb fibers contain less mitochondria and more glycogen and glycolytic enzymes, and type IIa fibers are involved in glycolysis and aerobic oxidation. The heme content of type IIb fibers was lower than other types of fibers, the proportion of oxidized myosin heavy chain in muscle is negatively correlated with myofibril diameter, and the proportion of intermediate type is positively correlated with the proportion of myofibrils [53]. This experiment found that the addition of GA increased the gene expression of oxidized myosin heavy chain in the breast muscle of broiler chickens, reduced the gene expression of intermediate myosin heavy chain, promoted the differentiation of muscle fiber heavy chain type to the oxidized muscle fiber, reduced the diameter of breast muscle fibers and increased the density. This suggests that GA may reduce muscle shear force, increase muscle tenderness and improve meat quality by regulating the muscle fiber structure type. It is similar to the findings on polyphenol in fattening pigs [54].
Conclusions
In conclusion, dietary addition of GA can alleviate the negative impact of stress response on the growth performance and enhanced the antioxidant capacity of broiler chickens. It also promoted the differentiation of the heavy-chain type of glycolytic myofibrils to oxidized myofibrils, which effectively improved the quality of broiler breast muscle. The appropriate amount of dietary GA at each stage was 300 mg/kg.
Author Contributions
Conceptualization, T.X., Z.L., D.R.; Data curation, T.X., M.H., J.W., and S.T.; Formal analysis, T.X., Z.L., and D.R.; Investigation and Project administration, C.Z., J.Y., D.X., and Q.F.; Resources, F.D. and Z.C.; Supervision, S.J.; Writing—original draft, T.X., M.H., J.W.; Writing—review editing, Z.C., D.R., and S.J.; Funding acquisition, S.J.. All authors have read and agreed to the published version of the manuscript.
Funding
The earmarked fund for China Agriculture Research System of MOF and MARA (CARS-41-G06); the National Key R&D Project (2021YFD1300404); the Science and Technology Planning Project of Guangdong Province (2023A0505050104), the Key Realm R&D Program of Guangdong Province (2020B0202090004); the Natural Science Foundation from Guangdong Province (2021A1515012412, 2021A1515010830, 2022A1515012069); the Science and Technology Plan Project of Guangzhou (202206010168), the Science and Technology Program of Guangdong Academy of Agricultural Sciences (202106TD, R2019PY-QF008). The Acknowledgement was extended to the Researchers Supporting Project (RSPD2024R731).
Institutional Review Board Statement
The experimental protocol was approved by the Animal Care Committee of the Institute of Animal Science, Guangdong Academy of Agriculture Science, Guangzhou, P. R. China, with the approval number GAASISA-2022-003.
Informed Consent Statement
Not applicable.
Data Availability Statement
All datasets collected and analyzed during the current study are available from the corresponding author by request, the availability of the data is restricted to investigators based at academic institutions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wierup, M. The control of microbial diseases in animals: alternatives to the use of antibiotics. Int. J. Antimicrob. Agents. 2000, 14, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Casewell, M.; Friis, C.; Marco, E.; McMullin, P.; Phillips, I. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef] [PubMed]
- De Cesare, A.; Sirri, F.; Manfreda, G.; Moniaci, P.; Giardini, A.; Zampiga, M.; Meluzzi, A. Effect of dietary supplementation with Lactobacillus acidophilus D2/CSL (CECT 4529) on caecum microbioma and productive performance in broiler chickens. PloS. One. 2017, 12, e0176309. [Google Scholar] [CrossRef] [PubMed]
- Arshad, R.; Mohyuddin, A.; Saeed, S.; Hassan, A.U. Optimized production of tannase and gallic acid from fruit seeds by solid state fermentation. Trop. J. Pharm. Res. 2021, 18, 911–918. [Google Scholar] [CrossRef]
- Li, Y.; Xie, Z.; Gao, T.; Li, L.; Chen, Y.; Xiao, D.; Liu, W.; Zou, B.; Lu, B.; Tian, X. A holistic view of gallic acid-induced attenuation in colitis based on microbiome-metabolomics analysis. Food. Funct. 2019, 10, 4046–4061. [Google Scholar] [CrossRef]
- Luciano, G.; Biondi, L.; Scerra, M.; Serra, A.; Mele, M.; Lanza, M.; Priolo, A. The effect of the change from a herbage-to a concentrate-based diet on the oxidative stability of raw and cooked lamb meat. Meat. Sci. 2013, 95, 212–218. [Google Scholar] [CrossRef]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef]
- Huang, Z.; Jin, S.; Lv, Z. Dietary genistein supplementation alters mRNA expression profile and alternative splicing signature in the thymus of chicks with lipopolysaccharide challenge. Poul. Sci. 2022, 101, 101561. [Google Scholar] [CrossRef]
- Zhong, W.J.; Yang, H.H.; Guan, X.X.; Xiong, J.B.; Sun, C.C.; Zhang, C.Y.; Luo, X.Q.; Zhang, Y.F.; Zhang, J.; Duan, J.X. Inhibition of glycolysis alleviates lipopolysaccharide-induced acute lung injury in a mouse model. J. Cell. Physiol. 2019, 234, 4641–4654. [Google Scholar] [CrossRef]
- Wang, Y.; Ye, J.; Zhang, S.; Chen, Z.; Fan, Q.; Jiang, S. Dietary supplementation with anthocyanin attenuates lipopolysaccharide-induced intestinal damage through antioxidant effects in yellow-feathered broiler chicks. Poul. Sci. 2023, 102. [Google Scholar] [CrossRef]
- NY/T3645-2020; Nutrient Requirements for Yellow-feathered Broilers. Ministry of Agriculture and Rural Affairs of the People’s Republic of China (PRC): Beijing, China, 2020.
- Jin, C.-l.; Wang, Q.; Zhang, Z.-m.; Xu, Y.-l.; Yan, H.-c.; Li, H.-c.; Gao, C.-q.; Wang, X.-q. Dietary supplementation with pioglitazone hydrochloride and chromium methionine improves growth performance, meat quality, and antioxidant ability in finishing pigs. J. Agric. Food. Chem. 2018, 66, 4345–4351. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, S.Z.; Chen, J.Y.; Chen, D.Z.; Deng, S.G.; Xu, B. Effect of cold plasma on maintaining the quality of chub mackerel (Scomber japonicus): Biochemical and sensory attributes. J. Sci. Food. Agric. 2019, 99, 39–46. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Li, Y.; Xiong, T.; Nie, X.; Zhang, H.; Zhu, C. Effect of dietary resveratrol supplementation on growth performance, antioxidant capacity, intestinal immunity and gut microbiota in yellow-feathered broilers challenged with lipopolysaccharide. Front. Microbiol. 2022, 13, 977087. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-Z.; Gong, J.-G.; Li, J.-H.; Hao, Y.-S.; Xu, H.-J.; Liu, Y.-C.; Feng, Z.-H. Dietary resveratrol supplementation on growth performance, immune function and intestinal barrier function in broilers challenged with lipopolysaccharide. Poult. Sci. 2023, 102, 102968. [Google Scholar] [CrossRef] [PubMed]
- Samuel, K.; Wang, J.; Yue, H.; Wu, S.; Zhang, H.; Duan, Z.; Qi, G. Effects of dietary gallic acid supplementation on performance, antioxidant status, and jejunum intestinal morphology in broiler chicks. Poult. Sci. 2017, 96, 2768–2775. [Google Scholar] [CrossRef]
- Salaheen, S.; Kim, S.-W.; Haley, B.J.; Van Kessel, J.A.S.; Biswas, D. Alternative growth promoters modulate broiler gut microbiome and enhance body weight gain. Front. Microbiol. 2017, 8, 2088. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug. Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Pandurangan, A.K.; Mohebali, N.; Esa, N.M.; Looi, C.Y.; Ismail, S.; Saadatdoust, Z. Gallic acid suppresses inflammation in dextran sodium sulfate-induced colitis in mice: Possible mechanisms. Int. Immunopharmacol. 2015, 28, 1034–1043. [Google Scholar] [CrossRef]
- Dhingra, M.S.; Dhingra, S.; Chadha, R.; Singh, T.; Karan, M. Design, synthesis, physicochemical, and pharmacological evaluation of gallic acid esters as non-ulcerogenic and gastroprotective anti-inflammatory agents. Med. Chem. Res. 2014, 23, 4771–4788. [Google Scholar] [CrossRef]
- Wang, C.; Matarneh, S.K.; Gerrard, D.; Tan, J. Modelling of energy metabolism and analysis of pH variations in postmortem muscle. Meat. Sci. 2021, 182, 108634. [Google Scholar] [CrossRef]
- Frerichs, C.; Beaulac, K.; Crowe, T.; Schwean-Lardner, K. The effects of simulated transport on the muscle characteristics of white-feathered end-of-cycle hens. Poult. Sci. 2021, 100, 101280. [Google Scholar] [CrossRef] [PubMed]
- Huo, W.; Weng, K.; Gu, T.; Zhang, Y.; Zhang, Y.; Chen, G.; Xu, Q. Effect of muscle fiber characteristics on meat quality in fast-and slow-growing ducks. Poult. Sci. 2021, 100, 101264. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Zhang, L.; Liao, P.; Xiao, Z.; Zhang, F.; Sindaye, D.; Xin, Z.; Tan, C.; Deng, J.; Yin, Y. Impact of gallic acid on gut health: focus on the gut microbiome, immune response, and mechanisms of action. Front. Immunol. 2020, 11, 580208. [Google Scholar] [CrossRef] [PubMed]
- Bennato, F.; Ianni, A.; Martino, C.; Grotta, L.; Martino, G. Evaluation of chemical composition and meat quality of breast muscle in broilers reared under light-emitting diode. Animals 2021, 11, 1505. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, G.; Lundström, K.; Tornberg, E. Contribution of pigment content, myoglobin forms and internal reflectance to the colour of pork loin and ham from pure breed pigs. Meat. Sci. 2001, 59, 141–151. [Google Scholar] [CrossRef]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef]
- Lan, R.; Zhao, Z.; Li, S.; An, L. Sodium butyrate as an effective feed additive to improve performance, liver function, and meat quality in broilers under hot climatic conditions. Poult. Sci. 2020, 99, 5491–5500. [Google Scholar] [CrossRef]
- Rey, A.; Menoyo, D.; Segura, J.; López-Bote, C.; Calvo, L. Combination of dietary glycaemic index and fasting time prior to slaughter as strategy to modify quality of pork. Meat. Sci. 2020, 161, 108013. [Google Scholar] [CrossRef]
- Onk, K.; Yalcintan, H.; Sari, M.; Isik, S.A.; Yakan, A.; Ekiz, B. Effects of genotype and sex on technological properties and fatty acid composition of duck meat. Poult. Sci. 2019, 98, 491–499. [Google Scholar] [CrossRef]
- Kim, H.-J.; Shin, D.-J.; Kim, H.-J.; Cho, J.; Kwon, J.-S.; Kim, D.; Jung, J.-H.; Jang, A. Assessment of chicken thigh meat quality of Ross 308 broiler of animal welfare certified farm. Anim. Biosci. 2022, 35, 1957. [Google Scholar] [CrossRef]
- Sujiwo, J.; Kim, D.; Jang, A. Relation among quality traits of chicken breast meat during cold storage: correlations between freshness traits and torrymeter values. Poult. Sci. 2018, 97, 2887–2894. [Google Scholar] [CrossRef] [PubMed]
- Jung SamOoel, J.S.; Bae YoungSik, B.Y.; Kim HyunJoo, K.H.; Jayasena, D.; Lee JunHeon, L.J.; Park HeeBok, P.H.; Heo KangNyung, H.K.; Jo CheoRun, J.C. Carnosine, anserine, creatine, and inosine 5′-Monophosphate contents in breast and thigh meats from 5 lines of Korean native chicken. Poult. Sci. 2013, 92, 3275–82. [Google Scholar] [CrossRef] [PubMed]
- Ruxton, C.; Reed, S.C.; Simpson, M.; Millington, K. The health benefits of omega-3 polyunsaturated fatty acids: a review of the evidence. J. Hum. Nutr. Diet. 2007, 20, 275–85. [Google Scholar] [CrossRef] [PubMed]
- Riediger, N.D.; Othman, R.A.; Suh, M.; Moghadasian, M.H. A systemic review of the roles of n-3 fatty acids in health and disease. J. Am. Diet. Assoc. 2009, 109, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Gou, Z.; Cui, X.; Li, L.; Fan, Q.; Lin, X.; Wang, Y.; Jiang, Z.; Jiang, S. Effects of dietary incorporation of linseed oil with soybean isoflavone on fatty acid profiles and lipid metabolism-related gene expression in breast muscle of chickens. Animal 2020, 14, 2414–2422. [Google Scholar] [CrossRef]
- Burghardt, P.R.; Kemmerer, E.S.; Buck, B.J.; Osetek, A.J.; Yan, C.; Koch, L.G.; Britton, S.L.; Evans, S.J. Dietary n-3: n-6 fatty acid ratios differentially influence hormonal signature in a rodent model of metabolic syndrome relative to healthy controls. Nutr. Metab. 2010, 7, 1–6. [Google Scholar] [CrossRef]
- Vlaicu, P.A.; Panaite, T.D.; Turcu, R.P. Enriching laying hens eggs by feeding diets with different fatty acid composition and antioxidants. Sci. Rep. 2021, 11, 20707. [Google Scholar] [CrossRef]
- Shen, Y.; Guo, C.; Lu, T.; Ding, X.-Y.; Zhao, M.-T.; Zhang, M.; Liu, H.-L.; Song, L.; Zhou, D.-Y. Effects of gallic acid alkyl esters and their combinations with other antioxidants on oxidative stability of DHA algae oil. Food. Res. Int. 2021, 143, 110280. [Google Scholar] [CrossRef]
- Mourão, J.L.; Pinheiro, V.; Prates, J.; Bessa, R.; Ferreira, L.; Fontes, C.; Ponte, P. Effect of dietary dehydrated pasture and citrus pulp on the performance and meat quality of broiler chickens. Poul. Sci. 2008, 87, 733–743. [Google Scholar] [CrossRef]
- Favier, A. Oxidative stress: value of its demonstration in medical biology and problems posed by the choice of a marker. Ann. Biol. Clin 1997, 9–16. [Google Scholar] [PubMed]
- Du, E.; Fan, Q.; Zhao, N.; Zhang, W.; Wei, J.; Chen, F.; Huang, S.; Guo, W. Supplemental magnolol improves the antioxidant capacity and intestinal health of broiler chickens. Anim. Sci. J. 2021, 92, e13665. [Google Scholar] [CrossRef]
- Lu, S.-H.; Chen, T.-H.; Chou, T.-C. Magnolol Inhibits RANKL-induced osteoclast differentiation of raw 264.7 macrophages through heme oxygenase-1-dependent inhibition of NFATc1 expression. J. Nat. Prod. 2015, 78, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, T.; Masuda, Y.; Izawa, T.; Ohnuma, T.; Ogura, K.; Hiratsuka, A. Magnolol protects PC12 cells from hydrogen peroxide or 6-hydroxydopamine induced cytotoxicity. J. Toxicol. Sci. 2019, 44, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Ramay, M.S.; Yalçın, S. Effects of supplemental pine needles powder (Pinus brutia) on growth performance, breast meat composition, and antioxidant status in broilers fed linseed oil-based diets. Poul. Sci. 2020, 99, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Long, L.; Zhang, H.; Wang, F.; Yin, Y.; Yang, L.; Chen, J. Research Note: Effects of polysaccharide-enriched Acanthopanax senticosus extract on growth performance, immune function, antioxidation, and ileal microbial populations in broiler chickens. Poult. Sci. 2021, 100, 101028. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.-C.; Duh, P.-D.; Tsai, H.-L. Antioxidant and pro-oxidant properties of ascorbic acid and gallic acid. Food. Chem. 2002, 79, 307–313. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, Y.; Tang, C.; Hou, Y.; Ai, X.; Chen, X.; Zhang, Y.; Wang, X.; Meng, X. Gallic acid: Pharmacological activities and molecular mechanisms involved in inflammation-related diseases. Biomed. Pharmacother. 2021, 133, 110985. [Google Scholar] [CrossRef]
- Starčević, K.; Krstulović, L.; Brozić, D.; Maurić, M.; Stojević, Z.; Mikulec, Ž.; Bajić, M.; Mašek, T. Production performance, meat composition and oxidative susceptibility in broiler chicken fed with different phenolic compounds. J. Sci. Food. Agric. 2015, 95, 1172–1178. [Google Scholar] [CrossRef]
- Zhang, L.; Yue, H.; Zhang, H.; Xu, L.; Wu, S.; Yan, H.; Gong, Y.; Qi, G. Transport stress in broilers: I. Blood metabolism, glycolytic potential, and meat quality. Poult. Sci. 2009, 88, 2033–2041. [Google Scholar] [CrossRef]
- Pette, D.; Staron, R.S. Mammalian skeletal muscle fiber type transitions. Int. Rev. Cytol. 1997, 170, 143–223. [Google Scholar] [CrossRef]
- Wang, T.; Xu, Y.Q.; Yuan, Y.X.; Xu, P.W.; Zhang, C.; Li, F.; Wang, L.N.; Yin, C.; Zhang, L.; Cai, X.C. Succinate induces skeletal muscle fiber remodeling via SUCNR1 signaling. EMBO. Rep. 2019, 20, e47892. [Google Scholar] [CrossRef]
- Li, R.; Li, B.; Jiang, A.; Cao, Y.; Hou, L.; Zhang, Z.; Zhang, X.; Liu, H.; Kim, K.-H.; Wu, W. Exploring the lncRNAs related to skeletal muscle fiber types and meat quality traits in pigs. Genes 2020, 11, 883. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Luo, J.; Yu, B.; Zheng, P.; Huang, Z.; Mao, X.; He, J.; Yu, J.; Chen, J.; Chen, D. Dietary resveratrol supplementation improves meat quality of finishing pigs through changing muscle fiber characteristics and antioxidative status. Meat. Sci. 2015, 102, 15–21. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of dietary GA on the mRNA expression of myosin heavy chain (MyHC) isoform genes in pectoral muscle 1. 1 Data are means ± SEM of six replicates per treatment (25 birds per replicate). 2 Values with various superscript letters in the same row are significantly different (p < 0.05). 3 CON: Basal diet and saline injection; LPS: Basal diet and LPS challenge; GA150: Basal diet supplemented with 150 mg/kg gallic acid and LPS challenge; GA300: Basal diet supplemented with 300 mg/kg gallic acid and LPS challenge; GA450: Basal diet supplemented with 450 mg/kg gallic acid and LPS challenge. 4 MyHC = Myosin heavy chain.
Figure 1.
Effects of dietary GA on the mRNA expression of myosin heavy chain (MyHC) isoform genes in pectoral muscle 1. 1 Data are means ± SEM of six replicates per treatment (25 birds per replicate). 2 Values with various superscript letters in the same row are significantly different (p < 0.05). 3 CON: Basal diet and saline injection; LPS: Basal diet and LPS challenge; GA150: Basal diet supplemented with 150 mg/kg gallic acid and LPS challenge; GA300: Basal diet supplemented with 300 mg/kg gallic acid and LPS challenge; GA450: Basal diet supplemented with 450 mg/kg gallic acid and LPS challenge. 4 MyHC = Myosin heavy chain.
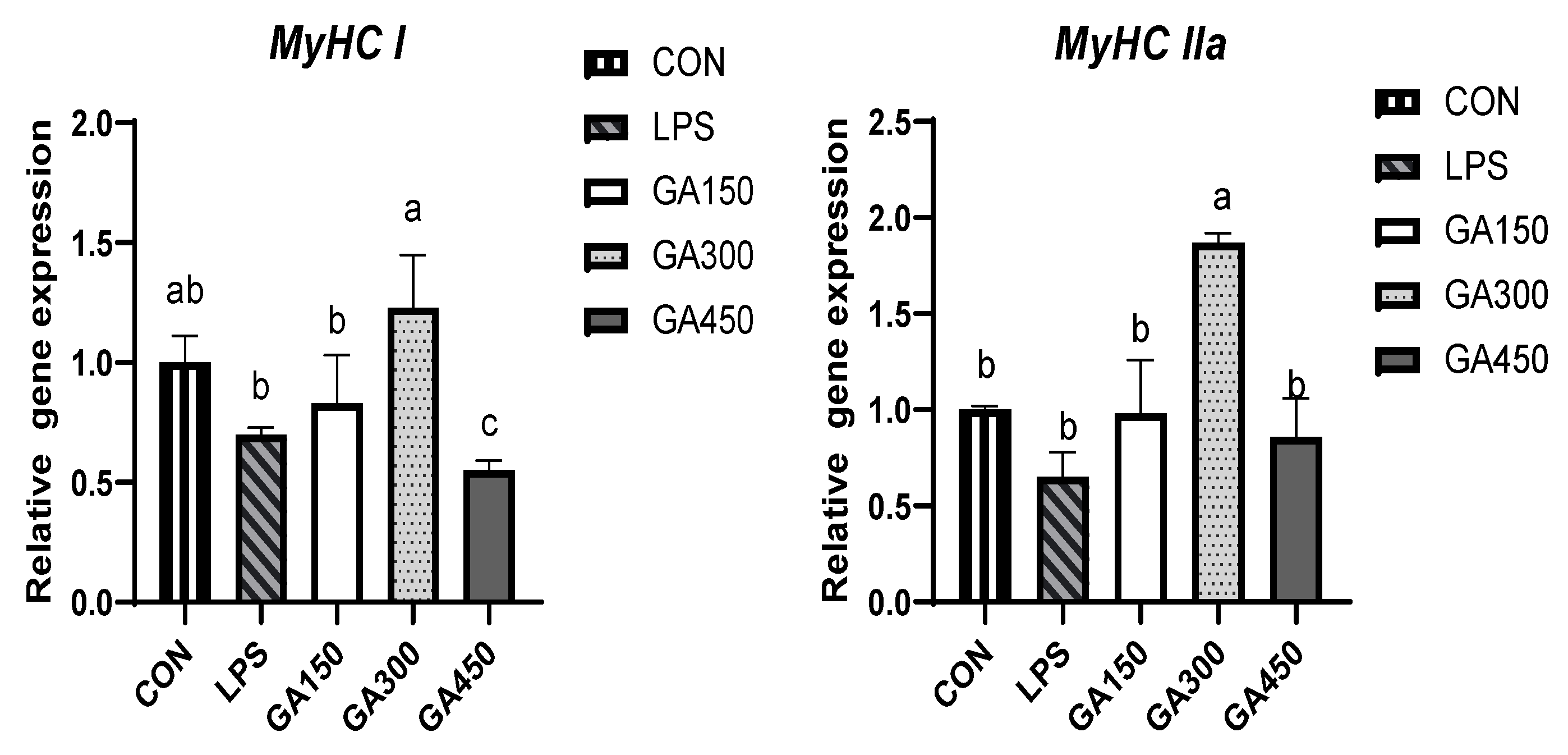
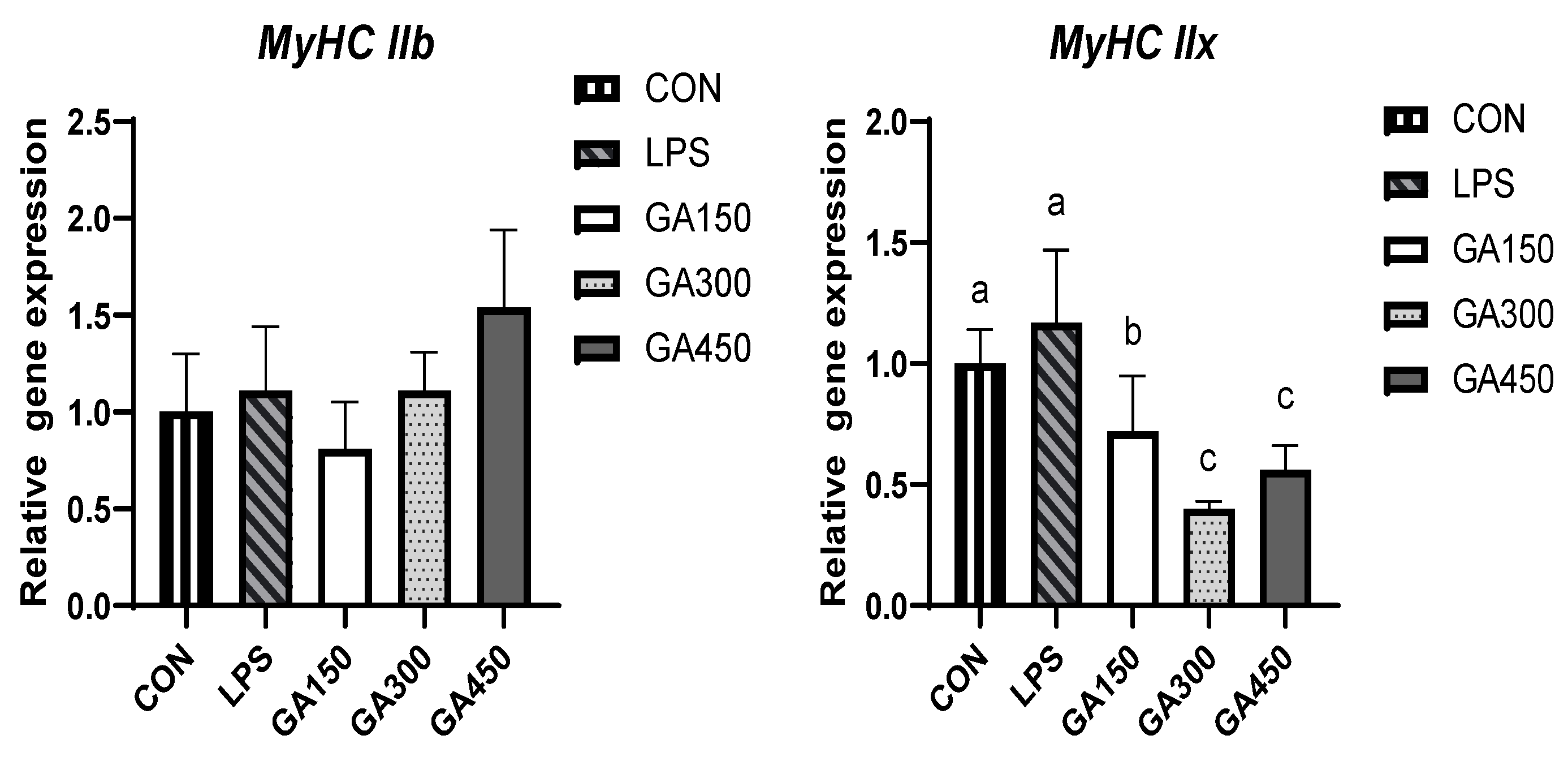

Table 1.
Experimental design.
| Treatment | CON | LPS | GA150 | GA300 | GA450 |
|---|---|---|---|---|---|
| Dietary supplementation with gallic acid, mg/kg | 0 | 0 | 150 | 300 | 450 |
1 CON: Basal diet and saline injection; LPS: Basal diet and LPS challenge; GA150: Basal diet supplemented with 150 mg/kg gallic acid and LPS challenge; GA300: Basal diet supplemented with 300 mg/kg gallic acid and LPS challenge; GA450: Basal diet supplemented with 450 mg/kg gallic acid and LPS challenge.
Table 2.
Composition and nutrient levels of the basal diet (air-dry basis, %).
| Items | 1 to 21 d | 22 to 42 d | 43 to 50 d |
|---|---|---|---|
| Ingredients, % | |||
| Corn | 52.07 | 51.80 | 51.00 |
| Wheat | 5.00 | 10.00 | 15.00 |
| Soybean meal | 33.00 | 25.80 | 20.60 |
| Corn gluten meal | 3.00 | 4.50 | 5.00 |
| Soybean oil | 2.28 | 3.50 | 4.40 |
| L-Lysine HCl | 0.25 | 0.24 | 0.20 |
| DL-Methionine | 0.17 | 0.14 | 0.08 |
| Limestone | 1.25 | 1.16 | 1.10 |
| CaHPO4 | 1.68 | 1.56 | 1.32 |
| NaCl | 0.30 | 0.30 | 0.30 |
| Premix 1 | 1.00 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 | 100.00 |
| Nutritional levels 2 | |||
| ME/(Kcal/kg) | 2960 | 3060 | 3155 |
| CP % | 23.11 | 21.40 | 19.38 |
| Ca % | 1.33 | 1.21 | 1.19 |
| Total phosphorus % | 0.76 | 0.7 | 0.71 |
| Nonphytate phosphorus P % | 0.47 | 0.41 | 0.36 |
| Lysine % | 1.29 | 1.15 | 0.96 |
| Methionine % | 0.52 | 0.48 | 0.40 |
| Threonine % | 0.86 | 0.81 | 0.67 |
1 Premix provided the following per kilogram of the diet during 1 to 21 d of age: VA 12 000 IU, VD3 600 IU, VE 45 IU, VK3 2.5 mg, VB1 2.4 mg, VB2 5 mg, VB6 2.8 mg, VB12 16 μg, pantothenic acid 12 mg, niacin 42 mg, biotin 0.12 mg, choline chloride 1 300 mg, Fe 80 mg, Cu 7 mg, Mn 80 mg, Zn 85 mg, Se 0.15 mg, I 0.7 mg; Premix provided the following per kilogram of the diet during 22 to 42 d of age: VA 9 000 IU, VD3 600 IU, VE 45 IU, VK3 2.5 mg, VB1 2.4 mg, VB2 5.0 mg, VB6 2.8 mg, VB12 16 μg, pantothenic acid 12 mg, niacin 42 mg, biotin 0.12 mg, choline chloride 1 000 mg, Fe 80 mg, Cu 7 mg, Mn 80 mg, Zn 80 mg, Se 0.15 mg, I 0.7 mg; Premix provided the following per kilogram of the diet during 43 to 50 d of age: VA 6 000 IU, VD3 500 IU, VE 25 IU, VK3 1.7 mg, VB1 1.0 mg, VB2 4.0 mg, VB6 0.6 mg, VB12 8.0 μg, pantothenic acid 8.0 mg, niacin 20 mg, biotin 0.02 mg, choline chloride 750 mg, Fe 80 mg, Cu 7 mg, Mn 80 mg, Zn 75 mg, Se 0.15 mg, I 0.7 mg. 2 CP, Ca and Total phosphorus were measured values, while the others were calculated.
Table 3.
Primers for real-time PCR of pectoral muscle.
| Gene | GenBank ID | Primers sequence (5’-3’) | Tm, ℃ |
|---|---|---|---|
| β-actin | NM_001001611.2 | F: GAGAAATTGTGCGTGACATCA | 56 |
| R: CCTGAACCTCTCATTGCCA | |||
| MyHC I | NM_204587.2 | F: ACGGGCCTGATCAACCAAAA | 56 |
| R: GGCCTTCTTGGCTTTCTCCT | |||
| MyHC IIa | NM_204228.3 | F: GGTCAACAAGCTCCGAGTGA | 56 |
| R: CAGGCCACTTTACTGCCTCA | |||
| MyHC IIb | NM_001013396.1 | F: GGGAGACCTGAATGAAATGGAG | 56 |
| R: CTTCCTGTGACCTGAGAGCATC | |||
| MyHC IIx | NC_052549.1 | F: AGACACATTTGCTGCACTGG | 56 |
| R: GGTCCGTGCCTTTCAGTTTT |
1 MyHC = Myosin heavy chain.
Table 4.
Effect of dietary GA supplementation on growth performance in broiler chickens 1.
| Items | CON | LPS | GA150 | GA300 | GA450 | SEM 4 | p-value |
|---|---|---|---|---|---|---|---|
| IBW (g) | 39.41 | 39.55 | 39.53 | 39.44 | 39.52 | 0.05 | 0.526 |
| FBW (kg) | 1.69 b | 1.71 b | 1.78 a | 1.75 ab | 1.75 ab | 0.02 | 0.031 |
| ADFI (g/d) | 65.17 | 65.98 | 66.78 | 65.41 | 66.40 | 0.35 | 0.078 |
| ADG (g/d) | 33.17 b | 33.98 b | 34.77 a | 33.84 b | 34.54 a | 0.20 | 0.039 |
| F: G | 1.96 | 1.94 | 1.92 | 1.93 | 1.92 | 0.01 | 0.278 |
| Mortality (%) | 4.00 | 9.33 | 7.33 | 5.33 | 9.34 | 2.57 | 0.466 |
1 Data are means of six replicates per treatment (25 birds per replicate). 2 Values with various superscript letters in the same row are significantly different (p < 0.05). 3 CON: Basal diet and saline injection; LPS: Basal diet and LPS challenge; GA150: Basal diet supplemented with 150 mg/kg gallic acid and LPS challenge; GA300: Basal diet supplemented with 300 mg/kg gallic acid and LPS challenge; GA450: Basal diet supplemented with 450 mg/kg gallic acid and LPS challenge. 4 SEM: Standard error of the mean. 5 IBW = Initial body weight; FBW = Final body weight; ADFI = Average daily feed intake; ADG = average daily gain; F:G = Feed-to-gain ratio.
Table 5.
Effect of dietary GA supplementation on meat quality of broilers 1.
| Items | CON | LPS | GA150 | GA300 | GA450 | SEM 4 | p-value |
|---|---|---|---|---|---|---|---|
| pH45min | 5.98 a | 5.91 ab | 5.97 a | 5.86 ab | 5.85 b | 0.02 | 0.035 |
| pH24h | 5.61 | 5.74 | 5.68 | 5.58 | 5.67 | 0.01 | 0.234 |
| L*45min | 57.74 bc | 56.86 c | 58.47 ab | 58.27 ab | 59.56 a | 0.47 | 0.008 |
| a*45min | 13.28 | 13.50 | 13.25 | 13.17 | 11.90 | 0.16 | 0.163 |
| b*45min | 18.00 | 17.03 | 17.28 | 17.42 | 17.93 | 0.23 | 0.130 |
| L*24h | 62.84 | 61.62 | 61.84 | 61.40 | 62.65 | 0.89 | 0.726 |
| a*24h | 11.67 | 12.20 | 12.98 | 13.00 | 11.80 | 0.16 | 0.269 |
| b*24h | 19.18 | 18.11 | 18.31 | 18.17 | 21.18 | 0.26 | 0.072 |
| Shear force, N | 28.77 | 31.19 | 26.42 | 26.01 | 17.21 | 0.90 | 0.109 |
| Drip loss, % | 2.34 | 2.82 | 2.41 | 2.42 | 2.31 | 0.06 | 0.669 |
1 Data are means of six replicates per treatment (25 birds per replicate). 2 Values with various superscript letters in the same row are significantly different (p < 0.05). 3 CON: Basal diet and saline injection; LPS: Basal diet and LPS challenge; GA150: Basal diet supplemented with 150 mg/kg gallic acid and LPS challenge; GA300: Basal diet supplemented with 300 mg/kg gallic acid and LPS challenge; GA450: Basal diet supplemented with 450 mg/kg gallic acid and LPS challenge. 4 SEM: Standard error of the mean.
Table 6.
Effect of dietary EA supplementation on IMF and TVB-N contents of breast muscle in broilers 1.
Table 6.
Effect of dietary EA supplementation on IMF and TVB-N contents of breast muscle in broilers 1.
| Items | CON | LPS | GA150 | GA300 | GA450 | SEM 4 | p-value |
|---|---|---|---|---|---|---|---|
| IMF, % | 1.54 | 1.26 | 1.30 | 1.44 | 1.60 | 0.01 | 0.509 |
| TVB-N, mg/100g | 12.72 a | 12.09 ab | 12.43 ab | 12.08 ab | 11.64 b | 0.14 | 0.007 |
1 Data are means of six replicates per treatment (25 birds per replicate). 2 Values with various superscript letters in the same row are significantly different (p < 0.05). 3 CON: Basal diet and saline injection; LPS: Basal diet and LPS challenge; GA150: Basal diet supplemented with 150 mg/kg gallic acid and LPS challenge; GA300: Basal diet supplemented with 300 mg/kg gallic acid and LPS challenge; GA450: Basal diet supplemented with 450 mg/kg gallic acid and LPS challenge. 4 SEM: Standard error of the mean.5 IMF = Intramuscular fat; TVB-N = Total volatile basic nitrogen.
Table 7.
Effect of dietary GA supplementation on fatty acid composition of breast muscle in broilers (%) 1.
Table 7.
Effect of dietary GA supplementation on fatty acid composition of breast muscle in broilers (%) 1.
| Items | CON | LPS | GA150 | GA300 | GA450 | SEM 4 | p-value |
|---|---|---|---|---|---|---|---|
| C14:0 | 0.48 | 0.50 | 0.48 | 0.48 | 0.45 | 0.02 | 0.274 |
| C16:0 | 21.57 | 22.57 | 21.82 | 21.42 | 20.85 | 0.45 | 0.063 |
| C18:0 | 8.51 | 8.92 | 8.39 | 8.39 | 8.33 | 0.39 | 0.821 |
| C20:0 | 0.22 | 0.23 | 0.22 | 0.22 | 0.21 | 0.01 | 0.844 |
| C22:0 | 0.72 | 0.73 | 0.69 | 0.80 | 0.65 | 0.08 | 0.741 |
| Total SFA | 32.26 | 32.78 | 32.01 | 31.93 | 31.01 | 0.80 | 0.138 |
| C16:1 | 2.86 | 3.25 | 3.20 | 2.62 | 2.52 | 0.28 | 0.248 |
| C18:1 | 26.72 | 26.9 | 27.13 | 25.95 | 25.17 | 0.84 | 0.462 |
| Total MUFA | 29.58 | 30.15 | 30.33 | 28.57 | 27.68 | 1.00 | 0.319 |
| C18:2n-6 | 28.32 | 27.87 | 29.35 | 29.65 | 29.35 | 0.97 | 0.663 |
| C18:3n-3 | 0.29 | 0.29 | 0.30 | 0.30 | 0.31 | 0.01 | 0.920 |
| C20:2n-6 | 0.59 | 0.54 | 0.60 | 0.66 | 0.59 | 0.05 | 0.578 |
| C20:3n-6 | 4.02 | 3.88 | 4.03 | 4.34 | 5.00 | 0.47 | 0.578 |
| C20:5n-3 | 0.93 | 0.92 | 0.93 | 0.98 | 1.13 | 0.12 | 0.724 |
| C22:6n-3 | 0.48 b | 0.49 b | 0.51 ab | 0.54 ab | 0.68 a | 0.07 | 0.019 |
| Total PUFA | 34.64 | 34.09 | 36.01 | 36.04 | 36.97 | 0.86 | 0.155 |
| n-6 | 32.88 | 32.33 | 34.28 | 34.28 | 34.86 | 0.84 | 0.201 |
| n-3 | 1.73 | 1.77 | 1.77 | 1.86 | 2.11 | 0.18 | 0.511 |
| n-3/n-6 | 0.05 | 0.05 | 0.05 | 0.06 | 0.06 | 0.00 | 0.667 |
1 Data are means of six replicates per treatment (25 birds per replicate). 2 Values with various superscript letters in the same row are significantly different (p < 0.05). 3 CON: Basal diet and saline injection; LPS: Basal diet and LPS challenge; GA150: Basal diet supplemented with 150 mg/kg gallic acid and LPS challenge; GA300: Basal diet supplemented with 300 mg/kg gallic acid and LPS challenge; GA450: Basal diet supplemented with 450 mg/kg gallic acid and LPS challenge. 4 SEM: Standard error of the mean. 5 SFA = Saturated fatty acids; MUFA = Monounsaturated fatty acids; PUFA = Polyunsaturated fatty acids. .
Table 8.
Effect of dietary GA supplementation on antioxidant capacity of breast muscle in broilers 1.
Table 8.
Effect of dietary GA supplementation on antioxidant capacity of breast muscle in broilers 1.
| Items | CON | LPS | GA150 | GA300 | GA450 | SEM 4 | p-value |
|---|---|---|---|---|---|---|---|
| T-SOD, U/mg prot | 301.41 c | 318.84 bc | 362.90 a | 357.21 a | 339.63 ab | 2.05 | 0.001 |
| GSH-Px, U/mg prot | 31.18 | 37.75 | 43.99 | 44.28 | 43.77 | 3.37 | 0.066 |
| CAT, U/mg prot | 32.55 bc | 30.30 c | 39.63 a | 37.62 ab | 32.89 bc | 9.79 | 0.013 |
| T-AOC, U/mg prot | 1.31 | 1.27 | 1.31 | 1.25 | 1.27 | 0.01 | 0.242 |
| MDA, nmol/mg prot | 2.41 a | 2.45 a | 1.76 ab | 1.59 b | 1.54 b | 0.01 | 0.038 |
1 Data are means of six replicates per treatment (25 birds per replicate). 2 Values with various superscript letters in the same row are significantly different (p < 0.05). 3 CON: Basal diet and saline injection; LPS: Basal diet and LPS challenge; GA150: Basal diet supplemented with 150 mg/kg gallic acid and LPS challenge; GA300: Basal diet supplemented with 300 mg/kg gallic acid and LPS challenge; GA450: Basal diet supplemented with 450 mg/kg gallic acid and LPS challenge. 4 SEM: Standard error of the mean. 5 T-SOD = Total superoxide dismutase; GSH-Px = Glutathione peroxidase; CAT = Catalase; T-AOC = Total anti-oxidative capacity; MDA = Malonic dialdehyde.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



