
Submitted:
09 September 2025
Posted:
11 September 2025
You are already at the latest version
Abstract
The aim of the present study is to evaluate the impact of chromium (Cr) supplementation on glucose and lipid metabolism in breast muscle in broilers under heat stress. A total of 220 one-day-old broiler chickens were reared in cages. At 29 days old, 180 birds were randomly assigned to three treatments (0, 400, and 800 µg Cr/kg, as chromium picolinate) and transferred to climate chambers (31 ± 1°C, 60 ± 7% humidity) for 14 days. The results showed that, compared with the heat stress group, the groups supplemented with 400 and 800 µg Cr/kg showed higher feed intake and average daily gain, breast muscle ratio, and lower feed conversion ratio and abdominal fat ratio; chickens supplemented with 400 and 800 µg Cr/kg showed significantly lower serum CORT, free fatty acids, and cholesterol levels compared with the heat stress (HS) group (P < 0.05). Fasting blood glucose and HOMA-IR were also significantly reduced, while fasting insulin was significantly increased in the Cr-supplemented groups (P < 0.05). Metabolomic analysis revealed that Cr supplementation regulated lipid and amino acid metabolism by altering key metabolites such as citric acid, L-glutamine, and L-proline, and modulating pathways including alanine, aspartate, and glutamate metabolism, and glycerophospholipid metabolism. Furthermore, Cr supplementation significantly upregulated the expression of PGC-1α, ABCA1, PPARα, and ABCG1 in both the hepatic and muscle tissue. This paper suggested that chromium supplementation may enhance energy metabolism and lipid transport by upregulating glucose and lipid metabolism genes, thereby mitigating heat stress-induced metabolic dysfunction and growth impairment.
Keywords:
chromium
; glucose metabolism
; lipid metabolism
; metabolomics
; skeletal muscle
1. Introduction
Heat stress is an increasingly harmful consequence caused by global warming, which has a wide range of detrimental effects on poultry production which includes reducing feed intake, growth rate, and feed conversion ratio, suboptimal carcass quality; excessive abdominal adiposity[1,2,3], while also exerting a detrimental effect on the immunity and organ function in broilers[4,5,6].
PPAR is a member of the nuclear receptor family involved in regulating lipid metabolism, lipoprotein metabolism, and glucose homeostasis[7]. One of the subtypes PPARα plays a crucial role in modulating gene expression associated with both peroxisomal and mitochondrial fatty acid oxidation. It also influences genes involved in fatty acid synthesis, transport, oxidation, and storage. By doing so, it contributes to the reduction of lipid accumulation and is intricately linked to the regulation of glucose and lipid metabolism as well as overall energy homeostasis[8]. The activation of PPARα can increase hepatic insulin activity, and reduce body fat deposition[9].
Chromium is an essential trace mineral commonly used as a dietary supplement for both humans and animals. Studies indicate that heat stress triggers the release of chromium from tissues, increases its excretion through urine, and decreases its retention, leading to chromium deficiency in the body[10]. As a result, the requirement for chromium becomes more critical. As is well known, heat stress can increase circulating concentrations of corticosterone in broilers. There is evidence showing that corticosterone can reduce insulin sensitivity in broilers, resulting in insulin resistance, which is associated with many diseases such as type II diabetes, obesity, and fatty liver disease[11]. In this case, chromium is known to enhance insulin sensitivity in broilers[12]. Supplementing the diet of broilers with chromium picolinate under heat stress has been shown to enhance growth performance [13], increase carcass yield[14], decrease abdominal fat deposition[15], improve the weights of immune organs[16], increase insulin, and decrease serum glucose[14].
As regards the skeletal muscle, the most relevant muscle in fast-growing broilers, accounting for 20–25% of their live body weight, which is considered a highly metabolically active tissue deeply involved in glucose and lipid metabolism[17]. Analyzing metabolomics in breast muscle could provide detailed information on glucose and lipid metabolism.
Our previous study showed that PPARα and ABCA1 were reduced in expression in heat-stressed broilers, which suggested that the PPARα gene may play a critical role in the mechanism of lipid metabolism under heat stress[18]. However, in light of previous studies, it can be found that researchers mainly focus on the lipid metabolism of the chromium effect. However, it is largely unknown how Cr supplementation impacts the glucose metabolism and lipid metabolism in the skeletal muscle and these related genes. Addtionally, it remains unclear whether the chromium supplementation can impact the subtype of PPAR, which in turn regulate glucose metabolism and lipid metabolism.
Therefore, the current study aimed to explore the effect of chromium on growth performance, blood parameters, breast muscle metabolism, and related gene expression under heat stress in broilers. We hypothesize that chromium dietary supplementation could affect the glucose and lipid metabolism, and PPARα may play a role in mitigating the impact of chromium under heat stress.
2. Materials and Methods
2.1. Experimental Design, Animals and Housing
A total of 220 one-day-old Arbor Acres male chickens were reared in 22 one-tier elevated cages (0.8 m × 0.8 m × 0.4 m) to 28 days old. The corn-soybean meal starter diet that meets NRC (1994) requirements for Arbor Acres chickens for this period was obtained from Huadu Commercial Company (Beijing, China) and served ad libitum with through feeders (0.82 m long × 0.07 m wide × 0.06 m deep). The water was available with nipple drinkers. The set of all environmental parameters followed the Arbor Acres management handbook. At the age of 29 d, 180 healthy male birds with similar body weight (1150 ± 70 g) were selected and randomly assigned to three treatments: 0 (heat stress group), 400, and 800 µg Cr/kg of supplemental diet (chromium picolinate) and transferred to three climate chambers. The environmental chambers were of identical size, each containing 6 cages with 10 chickens each. A cage was seen as a replicate. All treatments were conducted under a constant ambient temperature of 31 ± 1°C. The parameters of the chambers were set as follows: the humidity remained at 60 ± 7%, the airflow rate was under 0.5 m/s, and the ammonia concentration was controlled under 5 ppm. The lighting program was 24L:0D. The experiment lasted from 14 days to 42 days of age.
2.2. Diet
The basal diet was primarily composed of corn and soybean meal, and the composition is presented in Table 1. The heat stress group was subjected to the basal diet, while the diets of the other 2 groups were mixed with 400 and 800 µg Cr/kg chromium picolinate, respectively. Chickens were fed ad libitum and provided with free access to fresh drinking water throughout the experimental period. All diets were mixed thoroughly before feeding, and representative samples were analyzed for crude protein, metabolizable energy, and amino acid content to ensure diet consistency.
2.3. Measurements
Growth Performance: The body weight was determined at the start and the end of the experiment. The feed intake was recorded on a daily basis during the experiment. The average daily gain (ADG), average daily feed intake (ADFI), and feed conversion ratio (FCR) were calculated.
Body core temperature: 2 birds per replicate were randomly selected for the body core temperature determination by a thermometer once a week during the experiment.
Sample collection: At the end of the experiment, all chickens fasted for 12 h prior to the sampling. Thereafter, two chickens per replicate were selected based on their body weight closest to the cage average. The blood collection was performed using a blood collection needle at the wing vein. Subsequently, chickens were euthanized by CO₂ asphyxiation. The left breast muscle and abdominal fat were weighed, and their respective proportions were calculated as a percentage of body weight. The liver was collected and placed in a cryotube, immediately transferred to liquid nitrogen, and subsequently stored in a -80°C freezer.
LC-MS analysis: Breast muscle samples were analyzed using hydrophilic interaction liquid chromatography (HILIC) with an ACQUITY UPLC BEH Amide column (1.7 μm, Waters). Electrospray ionization (ESI) was applied in both positive and negative modes. The mobile phase system consisted of 25 mM ammonium acetate and 25 mM ammonium hydroxide in water (A), and acetonitrile (B). The gradient elution began with 85% B for 1 min, gradually decreased to 65% over 11 min, quickly dropped to 40% within 0.1 min and held for 4 min, followed by an increase to 85% within 0.1 min and a re-equilibration period of 5 min. For reversed-phase liquid chromatography (RPLC), an ACQUITY UPLC HSS T3 column (1.8 μm) was used. In positive mode, the mobile phase consisted of water and acetonitrile, both containing 0.1% formic acid, whereas in negative mode, 0.5 mM ammonium fluoride was added instead. The gradient profile was: 1% B for 1.5 min, increased to 99% over 11.5 min, maintained for 3.5 min, dropped to 1% within 0.1 min, and then equilibrated for 3.4 min. The column was operated at 25 °C with a flow rate of 0.3 mL/min, and 2 μL of each sample was injected for analysis.
Blood analysis: Serum samples were obtained by centrifugation at 3000×g for 20 min at 4°C. The fasting blood glucose was measured by using the Roche glucometer (Accu-Chek Performa, Roche Diagnostics, USA). The fasting insulin was determined by a kit that was obtained from Nanjing Jiancheng Institute of Bioengineering (Nanjing Jiancheng, Nanjing, China). The Homeostasis model assessment of insulin resistance (HOMA-IR) was calculated by a formulation as follows: HOMA-IR = FBG (mmol/L) × FINS (mU/L) / 22.5. The levels of free fatty acids, cholesterol, and corticosterone (CORT) were measured using an automatic biochemical analyzer (Hitachi 7600, Hitachi Corporation, Japan). All the procedures were carried out according to the protocol of the manufacturer.
qPCR: Total RNA from breast muscle samples was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA) following the manufacturer’s protocol. cDNA synthesis and real-time quantitative PCR were performed on a LightCycler 96 instrument (Roche, Basel, Switzerland) using standard qPCR procedures. Transcript levels of ABCA1, ABCG1, PPARα, and PGC-1α were quantified in both liver and breast muscle tissues. Gene-specific primers were designed based on GenBank sequences (see Table 2), and β-actin was employed as the reference gene for normalization. Relative gene expression was calculated using the 2^-ΔΔCt method.
2.4. Statistical Analysis
All data were analyzed using a one-way ANOVA based on a single-factor experimental design, followed by Duncan’s multiple range test for post-hoc comparisons, implemented in SAS 9.2 software (SAS Institute Inc., Cary, NC, USA). Results were expressed as mean ± standard deviation (SD), with statistical significance defined at P < 0.05. For metabolomics, orthogonal partial least squares discriminant analysis (OPLS-DA) was employed to differentiate between groups, and metabolites with a variable importance in projection (VIP) score ≥ 1 and P < 0.05 (from independent sample t-tests) were considered significantly different. Identified differential metabolites between the HS and NT groups were annotated using KEGG IDs and analyzed through MetaboAnalyst. The pathways with P < 0.05 were deemed significantly affected.
3. Results
3.1. Growth Performance
All chickens remained free from diseases, and no mortality occurred during the experiment. As shown in Table 3, the ADFI and ADG in the 400 and 800 µg Cr/kg groups were significantly higher than those in the HS group (P < 0.05); the FCR was lower than the HS group. Additionally, the 400 and 800 µg Cr/kg groups both exhibited a significant increase in breast muscle rate and a significant decrease in abdominal fat rate compared to the HS group (P < 0.05). However, no significant differences in these parameters were observed between the 400 and 800 µg Cr/kg groups (P > 0.05).
3.2. LC-MS Analysis
The scores (OPLS-DA) plot in Figure 1 shows a clear separation between two groups, indicating distinct metabolic profiles. In the OPLS-DA model, the positive mode scores plot yielded R²X=0.470, R²Y=0.999, and Q²=0.779, while the negative mode scores plot resulted in R²X=0.359, R²Y=0.994, and Q²=0.854.
A total of 91 metabolites were identified as significantly different between the HS group and the group receiving 400 µg Cr/kg, as determined by OPLS-DA (VIP ≥ 1) combined with independent sample t-test (P < 0.05). Detailed information on these differential metabolites is provided in Table 5. Relative to the HS group, 35 differential metabolites were upregulated and 56 differential metabolites were downregulated in the 400 µg Cr/kg group. The key differential metabolites involved in metabolic pathways include citric acid, L-aspartic acid, L-glutamine, L-proline, and (R)-(+)-2-Pyrrolidone-5-carboxylic acid, phosphatidylcholine (14:0/20:4[8Z,11Z,14Z,17Z]), 9,10,13-trihydroxy-octadecadienoic acid, phosphatidylcholine (22:6[4Z,7Z,10Z,13Z,16Z,19Z]), phosphatidylserine (18:0/20:0), and a series of lysophosphatidylcholine (LysoPC) species, including LysoPC (16:0), LysoPC (20:4[5Z,8Z,11Z,14Z]), LysoPC (18:1[9Z]), LysoPC (17:0), LysoPC (18:0), LysoPC (P-18:1[9Z]), LysoPC (20:1[11Z]), and LysoPC (P-18:0). Compared with the HS group, the concentrations of citric acid, L-aspartic acid, L-aspartic acid, L-glutamine, L-proline, (R)-(+)-2-pyrrolidone-5-carboxylic acid, PC(14:0/20:4[8Z,11Z,14Z,17Z]), 9,10,13-Trihydroxy-octadecadienoic acid were higher, and others were lower in the 400 µg Cr/kg group (P < 0.05).
As shown in Figure 2, pathway analysis revealed that supplementation with 400 µg Cr/kg had a significant impact on 9 metabolic pathways compared with the HS group (P < 0.05). These pathways include alanine, aspartate, and glutamate metabolism; D-glutamine and D-glutamate metabolism; arginine biosynthesis; linoleic acid metabolism; glycine, serine, and threonine metabolism; glyoxylate and dicarboxylate metabolism; aminoacyl-tRNA biosynthesis; ABC transporters; and glycerophospholipid metabolism.

Table 4.
Differential metabolites involved in metabolic pathways in metabolomics in breast muscle of HS and EG broilers.
Table 4.
Differential metabolites involved in metabolic pathways in metabolomics in breast muscle of HS and EG broilers.
| Metabolites | VIP(HS&EG) | P-value | Change |
|---|---|---|---|
| Citric acid | 5.92 | 0.04 | ↑ |
| L-aspartic acid | 6.94 | 0.02 | ↑ |
| L-glutamine | 3.91 | 0.02 | ↑ |
| L-proline | 5.82 | 0.03 | ↑ |
| (R)-(+)-2-Pyrrolidone-5-carboxylic acid | 5.23 | 0.02 | ↑ |
| Phosphatidylcholine (14:0/20:4[8Z,11Z,14Z,17Z]) | 3.94 | 0.05 | ↑ |
| 9,10,13-trihydroxy-octadecadienoic acid | 4.69 | 0.002 | ↑ |
| Phosphatidylcholine (22:6[4Z,7Z,10Z,13Z,16Z,19Z]) | 5.35 | 0.05 | ↓ |
| Phosphatidylserine (18:0/20:0) | 3.59 | 0.02 | ↓ |
| LysoPC (16:0) | 10.43 | 0.005 | ↓ |
| LysoPC (20:4[5Z,8Z,11Z,14Z]) | 3.56 | <0.001 | ↓ |
| LysoPC (18:1[9Z]) | 5.97 | 0.03 | ↓ |
| LysoPC (17:0) | 8.95 | 0.002 | ↓ |
| LysoPC (18:0) | 6.93 | 0.02 | ↓ |
| LysoPC (P-18:1[9Z]) | 8.38 | <0.001 | ↓ |
| LysoPC (20:1[11Z]) | 3.56 | 0.02 | ↓ |
| LysoPC (P-18:0) | 5.80 | 0.03 | ↓ |
3.3. Blood Paraments
Table 5 shows the levels of CORT, free fatty acids, and cholesterol in serum in the 400 and 800 µg Cr/kg groups were significantly lower (P < 0.05), compared with the HS group (P < 0.05), whereas there was no significant difference in the 400 and 800 µg Cr/kg group (P > 0.05). As shown in Figure 3, the fasting blood glucose and HOMA-IR in the 400 and 800 µg Cr/kg groups were significantly lower than those in the HS group (P < 0.05), and the fasting insulin in the 400 and 800 µg Cr/kg groups was significantly higher than that in the HS group (P < 0.05), whereas there was no significant difference in the 400 and 800 µg Cr/kg groups (P > 0.05).
Table 5.
Effects of 400 and 800 µg Cr/kg of chromium picolinate supplementation on blood parameters in broilers under heat stress from 29 d to 42 d of age.
Table 5.
Effects of 400 and 800 µg Cr/kg of chromium picolinate supplementation on blood parameters in broilers under heat stress from 29 d to 42 d of age.
| Items2 | 0 | 400 | 800 | P-value |
|---|---|---|---|---|
| Cholesterol (mmol/L) | 4.80a ± 0.11 | 3.57b ± 0.52 | 3.21b ± 0.26 | <0.01 |
| Free fatty acids (μmol/L) | 106.68a ± 3.24 | 89.06b ± 1.46 | 106.68b ± 3.24 | <0.01 |
| Corticosterone (ng/mL) | 15.78a ± 0.51 | 12.14b ± 0.72 | 12.14 b ± 1.22 | 0.02 |
3.4. Genes Associated with Glycolipid Homeostasis
As shown in Figure 4, the expression of PGC-1α, PPARα, ABCA1 and ABCG1 in the liver in the 400 µg Cr/kg group was significantly higher than in the HS group (P < 0.05). As shown in Figure 5, the expression of PGC-1α, PPARα, ABCA1 and ABCG1 mRNA in breast muscle in the 400 µg Cr/kg group was significantly higher than in the HS group (P < 0.05), whereas there was no significant difference of ABCG1 in breast muscle between the two groups (P > 0.05).
4. Discussion
The current study was performed under heat stress at 31°C for 4 weeks. Our findings demonstrated that dietary Cr provision, regardless of the dosage, both increased the growth performance, mitigated the glucose and lipid metabolism disorders.
Provision of Cr in broiler has been proven to mitigate the adverse impact on performance caused by heat stress[19]. Evidences has indicated that the use of Cr supplementation as a nutritional strategy to alleviate growth performance[20] via improved nutrient metabolism [14,21], immune responses[22], antioxidant function and stress response[20,23] in broiler chickens exposed to heat stress. Our results demonstrated that dietary supplementation with 400 and 800 µg Cr/kg significantly improved the average daily feed intake (ADFI) and average daily gain (ADG) of broilers while reducing the feed conversion ratio (FCR). Moreover, Cr significantly increased the breast muscle yield and decreased the abdominal fat rate, suggesting a beneficial effect on carcass composition and meat quality. These findings were similar to studies conducted bySahin, et al. [24] andSahin, Hayirli, Orhan, Tuzcu, Akdemir, Komorowski and Sahin [19] , they found feed intake and body mass gain were enhanced after provision of 200, 400, 800, or 1200 μg Cr as Cr pic/kg diet.Sahin, Sahin, Onderci, Gursu and Cikim [24] found that abdominal fat was decreased in chicks supplemented with 4 × 105 μg Cr as Cr pic/kg diet.Zha, et al. [25] found FCR to be decreased in chicks supplemented with 500 μg Cr as Cr pic/kg diet or 500 μg Cr as Cr nano/kg diet and found that chicks supplemented with 500 μg Cr as Cr nano/kg diet had increased breast muscle mass. However, the results of the current study are not consistent with those ofAmatya, et al. [26] andMoeini, et al. [27]. The conflicting results among these studies could be attributed, in part, to the differing chromium sources and administration dosages.
In our study, Cr supplementation significantly lowered fasting blood glucose (FBG) and the HOMA-IR while markedly increasing fasting insulin levels. These results indicate that Cr enhances insulin sensitivity and glucose homeostasis under HS conditions. The reduction in HOMA-IR suggests that Cr mitigates insulin resistance, which is commonly observed during heat stress due to glucocorticoid overproduction and metabolic stress[28]. Chromium is generally accepted to be the active component in glucose tolerance factor, which increases the sensitivity of tissue receptors to insulin, resulting in increased glucose uptake by cells[29,30]. The observed improvements in growth performance and carcass traits may be attributed to the ability of Cr to enhance insulin sensitivity and promote efficient nutrient utilization, particularly glucose and lipid metabolism. Previous studies have reported that Cr, particularly in the form of chromium picolinate, chromium propionate, and chromium chloride, enhances insulin receptor activity and facilitates glucose transport, thereby improving feed efficiency and lean tissue deposition in poultry under thermal stress conditions[14,31]. Furthermore, Cr supplementation has been shown to reduce lipid peroxidation and oxidative damage caused by HS, thereby supporting better muscle development and reducing fat accumulation[32].
Heat stress can stimulate the hypothalamus-pituitary-adrenal axis, causing an increase in the blood corticosterone concentration[33]. Studies have shown that excessive secretion of glucocorticoids such as corticosterone can trigger oxidative damage leading to muscle protein hydrolysis and significantly affect glucose metabolism, lipid metabolism, and muscle development[34]. In the present study, broilers supplemented with 400 and 800 µg Cr/kg exhibited significantly lower serum levels of corticosterone (CORT), free fatty acids (FFA), and cholesterol compared to the control group. It is reported that increasing chromium supplementation in broilers linearly reduces serum corticosterone concentrations[10]. The decreased FFA and cholesterol levels might be attributed to decreased glucocorticoid secretion, as demonstrated in a study by Sahin, Sahin, Onderci, Gursu and Cikim [24].
In this study, supplementation with 400 µg Cr/kg under heat stress altered metabolic profiles in breast muscle metabolomics. Citric acid plays a critical role in the TCA cycle[35]. In addition, the L-aspartic acid and L-glutamine are involved in the malate-aspartate shuttle system, which promotes energy production[36]. On the other hand, L-glutamine and L-proline are vital components of the amino acid metabolism pathway and are involved in the antioxidant system[37], suggesting that Cr might reduce amino acid degradation by maintaining oxidative stability. PC is the important component of the cell membrane. One of the possible explanations is that the Cr might promote choletone and PC external to the cell by activating the ABC transport protein pathway. 9,10,13-Trihydroxy-octadecadienoic acid is the product of linoleic acid metabolism and has anti-inflammatory and antioxidant effects[38]. Therefore, the elevated levels of citric acid, L-aspartic acid, and L-glutamine suggest the TCA cycle, amino acid metabolism, and linoleic acid metabolism might be improved by Cr supplementation. In the KEGG results, the enrichment of pathways confirmed our supposition. Dietary supplementation with 400 µg Cr/kg significantly influenced nine metabolic pathways, including alanine, aspartate, and glutamate metabolism; D-glutamine and D-glutamate metabolism; arginine biosynthesis; linoleic acid metabolism; glycine, serine, and threonine metabolism; glyoxylate and dicarboxylate metabolism; aminoacyl-tRNA biosynthesis; ABC transporter pathways; and glycerophospholipid metabolism. We hypothesize that Cr might enhance amino acid metabolism by promoting the biosynthesis and utilization of key amino acids such as glutamine, arginine, and glycine, which are essential for protein synthesis, nitrogen balance, and antioxidant defense under HS conditions. Improved glycine, serine, and threonine metabolism may enhance the production of glutathione, a critical antioxidant, thereby reducing oxidative stress and cellular damage caused by HS. Similarly, the upregulation of linoleic acid metabolism indicates enhanced lipid mobilization and utilization, which may contribute to the reduction in abdominal fat rate observed in Cr-supplemented broilers.
Our previous results indicated that heat stress can reduce PPARα level in liver and suggested that PPAR might play an intermediate role in regulating lipid metabolism in broilers[18]. In the present study, the results indicated that the 400 µg Cr as pic/kg supplementation increased PPARα level to a large extent, which indicated that it helped to alleviate the adverse effects of heat stress on lipid metabolism. The beneficial effects of Cr on metabolic pathways are likely mediated through the activation of energy-regulating signaling cascades, such as the AMP-activated protein kinase (AMPK) and peroxisome proliferator-activated receptor gamma coactivator-1 alpha (PGC-1α) pathways. Previous studies have demonstrated that Cr can activate AMPK, a key cellular energy sensor, which in turn promotes glucose uptake, fatty acid oxidation, and mitochondrial biogenesis[39]. The increased expression of PGC-1α, PPARα, ABCA1, and ABCG1 observed in the current study further supports this mechanism, indicating that Cr enhances hepatic and muscular energy metabolism and lipid transport. Moreover, the significant upregulation of ABCA1 and ABCG1 in the liver suggests that Cr enhances cholesterol efflux and reverse cholesterol transport (RCT). ABCA1 and ABCG1 are membrane transporters responsible for exporting excess cholesterol to high-density lipoproteins (HDL), facilitating its transport to the liver for clearance[40]. The increased expression of these transporters indicates that Cr promotes cholesterol homeostasis and reduces the risk of hepatic lipid overload under HS conditions. Notably, the elevated expression of ABCA1 and ABCG1 in the liver suggests that Cr promotes cholesterol efflux and lipid clearance[41], contributing to improved lipid homeostasis under heat stress, which is consistent with our earlier statement. Interestingly, the expression of ABCG1 in the breast muscle was not significantly altered by Cr supplementation. This could be attributed to the low basal expression levels of ABCG1 in skeletal muscle tissue or its relatively weaker role in lipid transport compared to the liver.
5. Conclusions
In conclusion, chromium supplementation might improve energy metabolism and lipid transport in the liver and muscles by upregulating the expression of genes related to glucose and lipid metabolism, thereby alleviating the damage to the glucose and lipid metabolic function and growth performance under heat stress conditions.
Author Contributions
Conceptualization, G.W. and M.Z.; methodology, G.W.; software, J.F.; validation, Z.H. and M.Z.; formal analysis, X.L.; investigation, G.W.; resources, M.Y.; data curation, Z.H.; writing—original draft preparation, G.W.; writing—review and editing, X.L.; visualization, G.W.; supervision, M.Z.; project administration, M.Z.; funding acquisition, M.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (General Program 32272933). This work was also supported by the National Key R&D Program of China (2021YFD1300404) and the Science and Technology Innovation Project of the Chinese Academy of Agricultural Sciences (ASTIP-IAS09).
Institutional Review Board Statement
The experimental protocol had been approved by the Animal Experimental Welfare and Ethical Inspection Form of the Institute of Animal Science, Chinese Academy of Agricultural Sciences (permit number: IAS2020-112).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tang, L.-P.; Liu, Y.-L.; Zhang, J.-X.; Ding, K.-N.; Lu, M.-H.; He, Y.-M. Heat stress in broilers of liver injury effects of heat stress on oxidative stress and autophagy in liver of broilers. Poultry science 2022, 101, 102085. [Google Scholar] [CrossRef] [PubMed]
- Apalowo, O.O.; Ekunseitan, D.A.; Fasina, Y.O. Impact of Heat Stress on Broiler Chicken Production. Poultry 2024, 3, 107–128. [Google Scholar] [CrossRef]
- Kim, H.R.; Seong, P.; Seol, K.-H.; Park, J.-E.; Kim, H.; Park, W.; Cho, J.H.; Lee, S.D. Effects of heat stress on growth performance, physiological responses, and carcass traits in broilers. Journal of Thermal Biology 2025, 127, 103994. [Google Scholar] [CrossRef]
- Chaudhary, A.; Mishra, P.; Al Amaz, S.; Mahato, P.L.; Das, R.; Jha, R.; Mishra, B. Dietary supplementation of microalgae mitigates the negative effects of heat stress in broilers. Poultry science 2023, 102, 102958. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ouyang, J.; Deng, C.; Zhou, H.; You, J.; Li, G. Effects of dietary tryptophan supplementation on rectal temperature, humoral immunity, and cecal microflora composition of heat-stressed broilers. Frontiers in Veterinary Science 2023, 10, 1247260. [Google Scholar] [CrossRef]
- Sun, S.; Li, B.; Wu, M.; Deng, Y.; Li, J.; Xiong, Y.; He, S. Effect of dietary supplemental vitamin C and betaine on the growth performance, humoral immunity, immune organ index, and antioxidant status of broilers under heat stress. Tropical animal health and production 2023, 55, 96. [Google Scholar] [CrossRef]
- Kawasaki, M.; Kambe, A.; Yamamoto, Y.; Arulmozhiraja, S.; Ito, S.; Nakagawa, Y.; Tokiwa, H.; Nakano, S.; Shimano, H. Elucidation of molecular mechanism of a selective PPARα modulator, pemafibrate, through combinational approaches of X-ray crystallography, thermodynamic analysis, and first-principle calculations. International Journal of Molecular Sciences 2020, 21, 361. [Google Scholar] [CrossRef]
- Xu, J.; Xiao, G.; Trujillo, C.; Chang, V.; Blanco, L.; Joseph, S.B.; Bassilian, S.; Saad, M.F.; Tontonoz, P.; Lee, W.P. Peroxisome proliferator-activated receptor α (PPARα) influences substrate utilization for hepatic glucose production. Journal of Biological Chemistry 2002, 277, 50237–50244. [Google Scholar] [CrossRef]
- Biernacki, M.; Ambrożewicz, E.; Gęgotek, A.; Toczek, M.; Skrzydlewska, E. Long-term administration of fatty acid amide hydrolase inhibitor (URB597) to rats with spontaneous hypertension disturbs liver redox balance and phospholipid metabolism. Advances in Medical Sciences 2019, 64, 15–23. [Google Scholar] [CrossRef]
- Sahin, K.; Ozbey, O.; Onderci, M.; Cikim, G.; Aysondu, M.H. Chromium supplementation can alleviate negative effects of heat stress on egg production, egg quality and some serum metabolites of laying Japanese quail. Journal of Nutrition 2002, 132, 1265–1268. [Google Scholar] [CrossRef]
- Yuan, L.; Lin, H.; Jiang, K.; Jiao, H.; Song, Z. Corticosterone administration and high-energy feed results in enhanced fat accumulation and insulin resistance in broiler chickens. British poultry science 2008, 49, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.A.; Grimes, J.L.; Lloyd, K.E.; Krafka, K.; Lamptey, A.; Spears, J.W. Chromium propionate in broilers: effect on insulin sensitivity. Poultry Science 2016, 95, 1096–1104. [Google Scholar] [CrossRef]
- Toghyani, M.; Toghyani, M.; Shivazad, M.; Gheisari, A.; Bahadoran, R. Chromium Supplementation Can Alleviate the Negative Effects of Heat Stress on Growth Performance, Carcass Traits, and Meat Lipid Oxidation of Broiler Chicks without Any Adverse Impacts on Blood Constituents. Biological Trace Element Research 2012, 146, 171–180. [Google Scholar] [CrossRef]
- Sahin, K.; Sahin, N.; Kucuk, O. Effects of chromium, and ascorbic acid supplementation on growth, carcass traits, serum metabolites, and antioxidant status of broiler chickens reared at a high ambient temperature (32°C). Nutrition Research 2003, 23, 225–238. [Google Scholar] [CrossRef]
- Kulkari, R.; Mandal, A.; Bhanja, S.; Goel, A.; Mehera, M. Dietary supplementation of chromium picolinate on productive performance and cost economics of coloured broiler chicken during hot-humid summer. International Journal of Livestock Research 2018, 8, 132–139. [Google Scholar] [CrossRef]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Increased fat synthesis and limited apolipoprotein B cause lipid accumulation in the liver of broiler chickens exposed to chronic heat stress. Poultry Science 2019, 98, 3695–3704. [Google Scholar] [CrossRef]
- Meloche, K.; Dozier III, W.; Brandebourg, T.; Starkey, J. Skeletal muscle growth characteristics and myogenic stem cell activity in broiler chickens affected by wooden breast. Poultry Science 2018, 97, 4401–4414. [Google Scholar] [CrossRef]
- Li, X.; Zhao, X.; Yu, M.; Zhang, M.; Feng, J. Effects of Heat Stress on Breast Muscle Metabolomics and Lipid Metabolism Related Genes in Growing Broilers. Animals 2024, 14, 430. [Google Scholar] [CrossRef]
- Sahin, N.; Hayirli, A.; Orhan, C.; Tuzcu, M.; Akdemir, F.; Komorowski, J.R.; Sahin, K. Effects of the supplemental chromium form on performance and oxidative stress in broilers exposed to heat stress. Poultry Science 2017, 96, 4317–4324. [Google Scholar] [CrossRef]
- Ghazi, S.; Habibian, M.; Moeini, M.M.; Abdolmohammadi, A.R. Effects of Different Levels of Organic and Inorganic Chromium on Growth Performance and Immunocompetence of Broilers under Heat Stress. Biological Trace Element Research 2012, 146, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Jahanian, R.; Rasouli, E. Dietary chromium methionine supplementation could alleviate immunosuppressive effects of heat stress in broiler chicks. Journal of Animal Science 2015, 93, 3355–3363. [Google Scholar] [CrossRef]
- Oba, A.; Lopes, P.C.F.; Boiago, M.M.; Silva, A.M.S.; Montassier, H.J.; de Souza, P.A. Productive and immunological traits of broiler chickens fed diets supplemented with chromium, reared under different environmental conditions. Revista Brasileira De Zootecnia-Brazilian Journal of Animal Science 2012, 41, 1186–1192. [Google Scholar] [CrossRef]
- Khan, R.U.; Naz, S.; Dhama, K. Chromium: Pharmacological Applications in Heat-Stressed Poultry. International Journal of Pharmacology 2014, 10, 213–217. [Google Scholar] [CrossRef]
- Sahin, K.; Sahin, N.; Onderci, M.; Gursu, F.; Cikim, G. Optimal dietary concentration of chromium for alleviating the effect of heat stress on growth, carcass qualities, and some serum metabolites of broiler chickens. Biological Trace Element Research 2002, 89, 53–64. [Google Scholar] [CrossRef]
- Zha, L.Y.; Zeng, J.W.; Chu, X.W.; Mao, L.M.; Luo, H.J. Efficacy of trivalent chromium on growth performance, carcass characteristics and tissue chromium in heat-stressed broiler chicks. Journal of the Science of Food and Agriculture 2009, 89, 1782–1786. [Google Scholar] [CrossRef]
- Amatya, J.L.; Haldar, S.; Ghosh, T.K. Effects of chromium supplementation from inorganic and organic sources on nutrient utilization, mineral metabolism and meat quality in broiler chickens exposed to natural heat stress. Animal Science 2004, 79, 241–253. [Google Scholar] [CrossRef]
- Moeini, M.M.; Bahrami, A.; Ghazi, S.; Targhibi, M.R. The Effect of Different Levels of Organic and Inorganic Chromium Supplementation on Production Performance, Carcass Traits and Some Blood Parameters of Broiler Chicken Under Heat Stress Condition. Biological Trace Element Research 2011, 144, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Beaupere, C.; Liboz, A.; Fève, B.; Blondeau, B.; Guillemain, G. Molecular mechanisms of glucocorticoid-induced insulin resistance. International journal of molecular sciences 2021, 22, 623. [Google Scholar] [CrossRef] [PubMed]
- Mertz, W.; Schwarz, K. Relation of glucose tolerance factor to impaired intravenous glucose tolerance of rats on stock diets. American Journal of Physiology-Legacy Content 1959, 196, 614–618. [Google Scholar] [CrossRef]
- Mertz, W. Chromium occurrence and function in biological systems. Physiological reviews 1969, 49, 163–239. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.J.; Li, X.M.; Zhou, Y.; Feng, J.H.; Zhang, M.H. Effects of Dietary Chromium Picolinate on Gut Microbiota, Gastrointestinal Peptides, Glucose Homeostasis, and Performance of Heat-Stressed Broilers. Animals 2022, 12. [Google Scholar] [CrossRef]
- Sahin, N.; Akdemir, F.; Tuzcu, M.; Hayirli, A.; Smith, M.; Sahin, K. Effects of supplemental chromium sources and levels on performance, lipid peroxidation and proinflammatory markers in heat-stressed quails. Animal feed science and technology 2010, 159, 143–149. [Google Scholar] [CrossRef]
- Del Barrio, A.S.; Mansilla, W.; Navarro-Villa, A.; Mica, J.; Smeets, J.; Den Hartog, L.; García-Ruiz, A. Effect of mineral and vitamin C mix on growth performance and blood corticosterone concentrations in heat-stressed broilers. Journal of Applied Poultry Research 2020, 29, 23–33. [Google Scholar] [CrossRef]
- Sato, H.; Takahashi, T.; Sumitani, K.; Takatsu, H.; Urano, S. Glucocorticoid generates ROS to induce oxidative injury in the hippocampus, leading to impairment of cognitive function of rats. Journal of clinical biochemistry and nutrition 2010, 47, 224–232. [Google Scholar] [CrossRef]
- Owen, O.E.; Kalhan, S.C.; Hanson, R.W. The key role of anaplerosis and cataplerosis for citric acid cycle function. Journal of Biological Chemistry 2002, 277, 30409–30412. [Google Scholar] [CrossRef]
- Broeks, M.H.; Meijer, N.W.; Westland, D.; Bosma, M.; Gerrits, J.; German, H.M.; Ciapaite, J.; van Karnebeek, C.D.; Wanders, R.J.; Zwartkruis, F.J. The malate-aspartate shuttle is important for de novo serine biosynthesis. Cell Reports 2023, 42. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Si, X.; Ji, Y.; Yang, Q.; Bai, J.; He, Y.; Jia, H.; Song, Z.; Chen, J.; Yang, L. l-Proline improves the cytoplasmic maturation of mouse oocyte by regulating glutathione-related redox homeostasis. Theriogenology 2023, 195, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Marzouk, M.M.; Hegazi, N.M.; El Shabrawy, M.O.; Farid, M.M.; Kawashty, S.A.; Hussein, S.R.; Saleh, N.A. Discriminative Metabolomics Analysis and Cytotoxic Evaluation of Flowers, Leaves, and Roots Extracts of Matthiola longipetala subsp. livida. Metabolites 2023, 13, 909. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.-C.; Liu, J.-M.; Lee, K.-I.; Tang, F.-C.; Fang, K.-M.; Yang, C.-Y.; Su, C.-C.; Chen, H.-H.; Hsu, R.-J.; Chen, Y.-W. Cr (VI) induces ROS-mediated mitochondrial-dependent apoptosis in neuronal cells via the activation of Akt/ERK/AMPK signaling pathway. Toxicology in Vitro 2020, 65, 104795. [Google Scholar] [CrossRef]
- Ji, A.; Wroblewski, J.M.; Cai, L.; de Beer, M.C.; Webb, N.R.; van der Westhuyzen, D.R. Nascent HDL formation in hepatocytes and role of ABCA1, ABCG1, and SR-BI. Journal of lipid research 2012, 53, 446–455. [Google Scholar] [CrossRef]
- Rosenson, R.S.; Brewer Jr, H.B.; Davidson, W.S.; Fayad, Z.A.; Fuster, V.; Goldstein, J.; Hellerstein, M.; Jiang, X.-C.; Phillips, M.C.; Rader, D.J. Cholesterol efflux and atheroprotection: advancing the concept of reverse cholesterol transport. Circulation 2012, 125, 1905–1919. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
OPLS-DA score plots of metabolite profiles in broilers from the heat stress (HT) and 400 µg Cr/kg (EG) groups. (A) Positive ionization mode; (B) Negative ionization mode. Yellow represents the EG group, and green represents the HT group. Distinct clustering indicates metabolic differences between treatments.
Figure 1.
OPLS-DA score plots of metabolite profiles in broilers from the heat stress (HT) and 400 µg Cr/kg (EG) groups. (A) Positive ionization mode; (B) Negative ionization mode. Yellow represents the EG group, and green represents the HT group. Distinct clustering indicates metabolic differences between treatments.
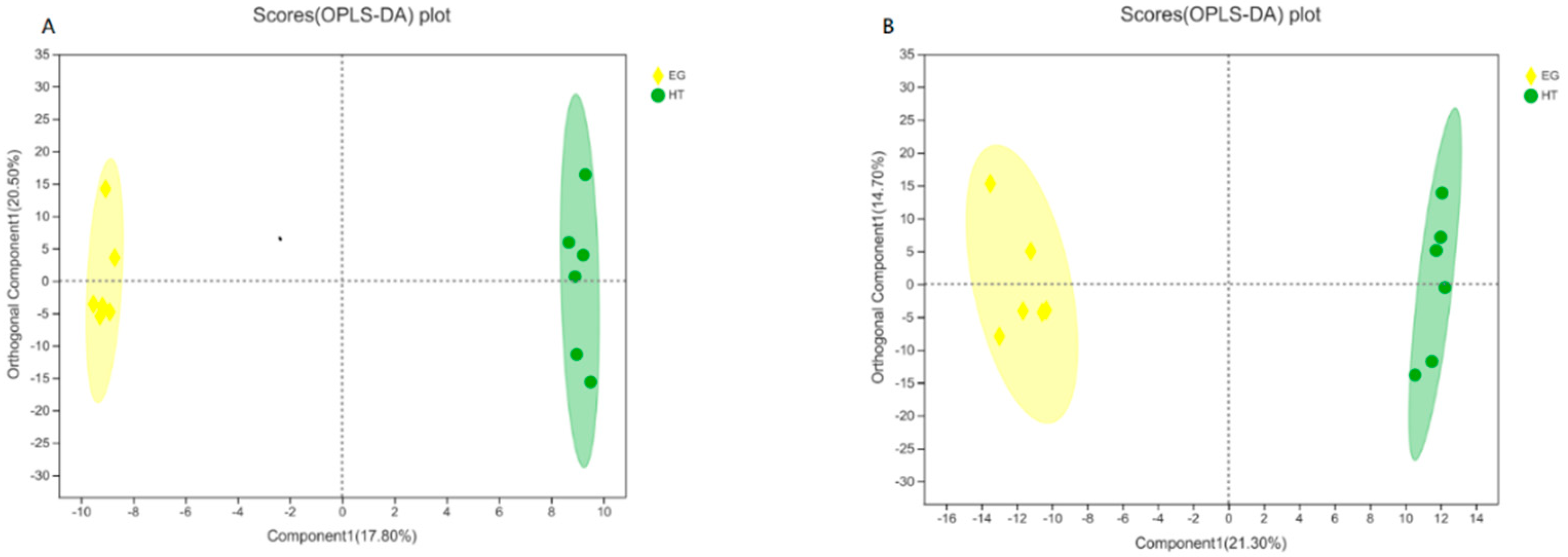
Figure 2.
Pathway enrichment of the identified metabolites was conducted using the KEGG database in broilers from the heat stress (HS) group and the 400 µg Cr/kg supplementation (EG) group. The x-axis means the pathway, while the y-axis represents the enrichment ratio, defined as the proportion of pathway-enriched metabolites to the total metabolites associated with that pathway. A higher enrichment rate signifies a greater degree of enrichment. The color gradient of the bars represents the significance of enrichment, and the dark color bars indicating more significant enrichment of the KEGG term. Statistical significance is denoted as follows: P-value or FDR < 0.001 is marked as ***, P-value or FDR < 0.01 is marked as **, and P-value or FDR < 0.05 is marked as *.
Figure 2.
Pathway enrichment of the identified metabolites was conducted using the KEGG database in broilers from the heat stress (HS) group and the 400 µg Cr/kg supplementation (EG) group. The x-axis means the pathway, while the y-axis represents the enrichment ratio, defined as the proportion of pathway-enriched metabolites to the total metabolites associated with that pathway. A higher enrichment rate signifies a greater degree of enrichment. The color gradient of the bars represents the significance of enrichment, and the dark color bars indicating more significant enrichment of the KEGG term. Statistical significance is denoted as follows: P-value or FDR < 0.001 is marked as ***, P-value or FDR < 0.01 is marked as **, and P-value or FDR < 0.05 is marked as *.
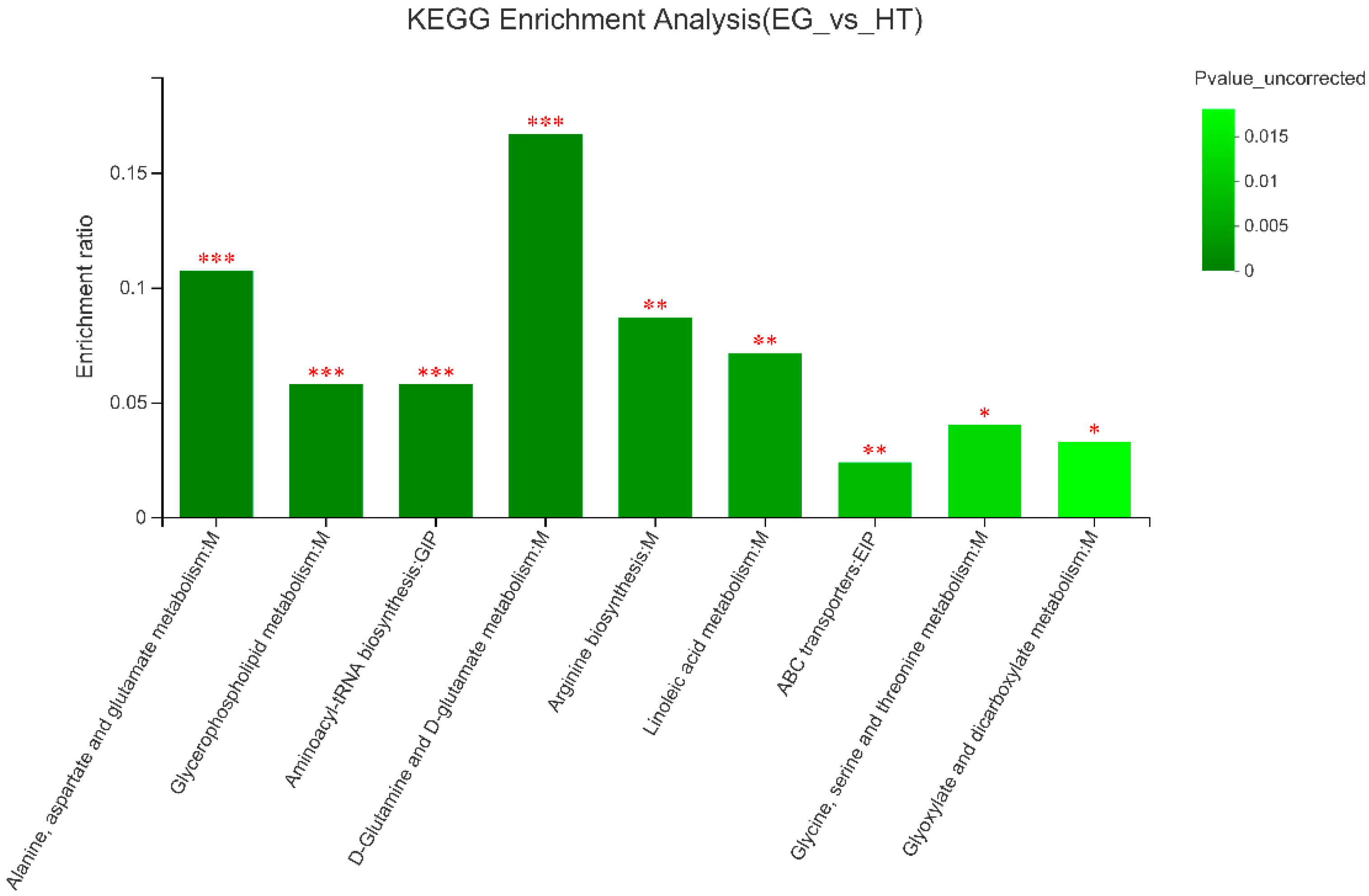
Figure 3.
Effects of 400 and 800 µg Cr/kg of chromium picolinate supplementation on blood parameters in broilers under heat stress from 29 d to 42 d of age.
Figure 3.
Effects of 400 and 800 µg Cr/kg of chromium picolinate supplementation on blood parameters in broilers under heat stress from 29 d to 42 d of age.
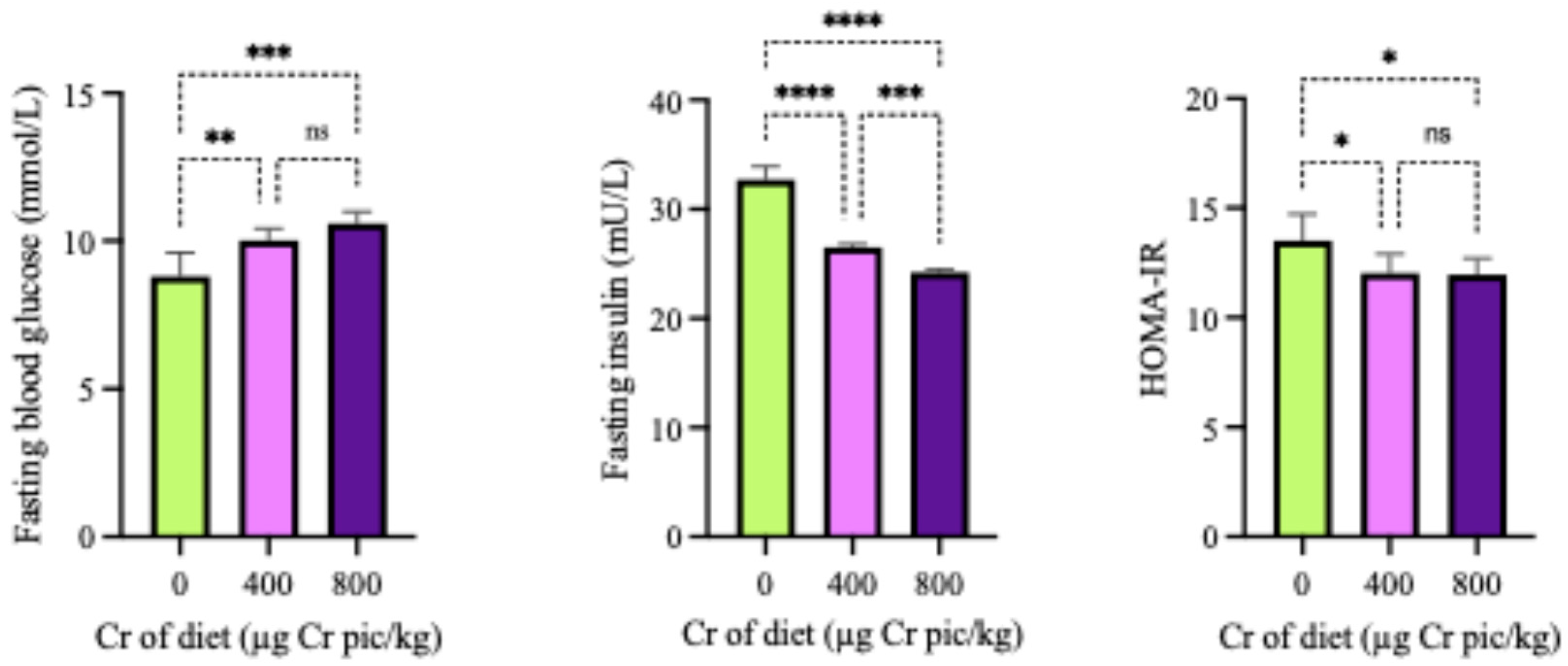
Figure 4.
Effects of 400 µg Cr/kg supplementation on the mRNA expression of genes involved in glycolipid metabolism in the liver of heat-stressed broilers. The relative expression levels of PGC-1α (A), PPARα (B), ABCA1 (C), and ABCG1 (D) are shown. Each bar represents the mean ± SD (n = 6). Bars labeled with different superscripts (a, b) indicate significant differences at P < 0.05.
Figure 4.
Effects of 400 µg Cr/kg supplementation on the mRNA expression of genes involved in glycolipid metabolism in the liver of heat-stressed broilers. The relative expression levels of PGC-1α (A), PPARα (B), ABCA1 (C), and ABCG1 (D) are shown. Each bar represents the mean ± SD (n = 6). Bars labeled with different superscripts (a, b) indicate significant differences at P < 0.05.
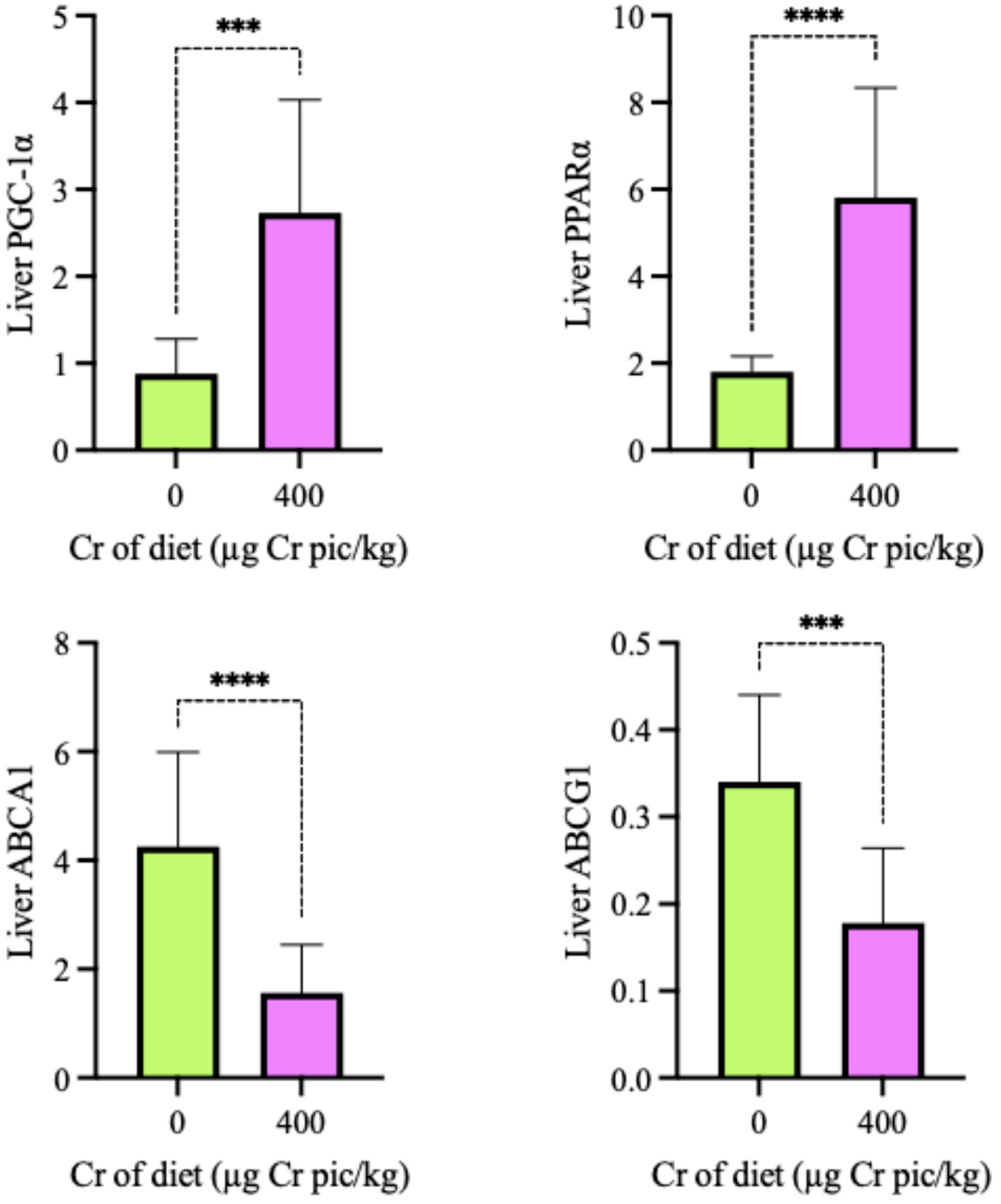
Figure 5.
Impact of 400 µg Cr/kg supplementation on the mRNA expression of glycolipid metabolism genes in the breast muscle of heat-stressed broilers. The expression levels of PGC-1α (A), PPARα (B), ABCA1 (C), and ABCG1 (D) are shown. Each bar represents the mean ± SD (n = 6). Bars labeled with different superscripts (a, b) indicate significant differences at P < 0.05.
Figure 5.
Impact of 400 µg Cr/kg supplementation on the mRNA expression of glycolipid metabolism genes in the breast muscle of heat-stressed broilers. The expression levels of PGC-1α (A), PPARα (B), ABCA1 (C), and ABCG1 (D) are shown. Each bar represents the mean ± SD (n = 6). Bars labeled with different superscripts (a, b) indicate significant differences at P < 0.05.
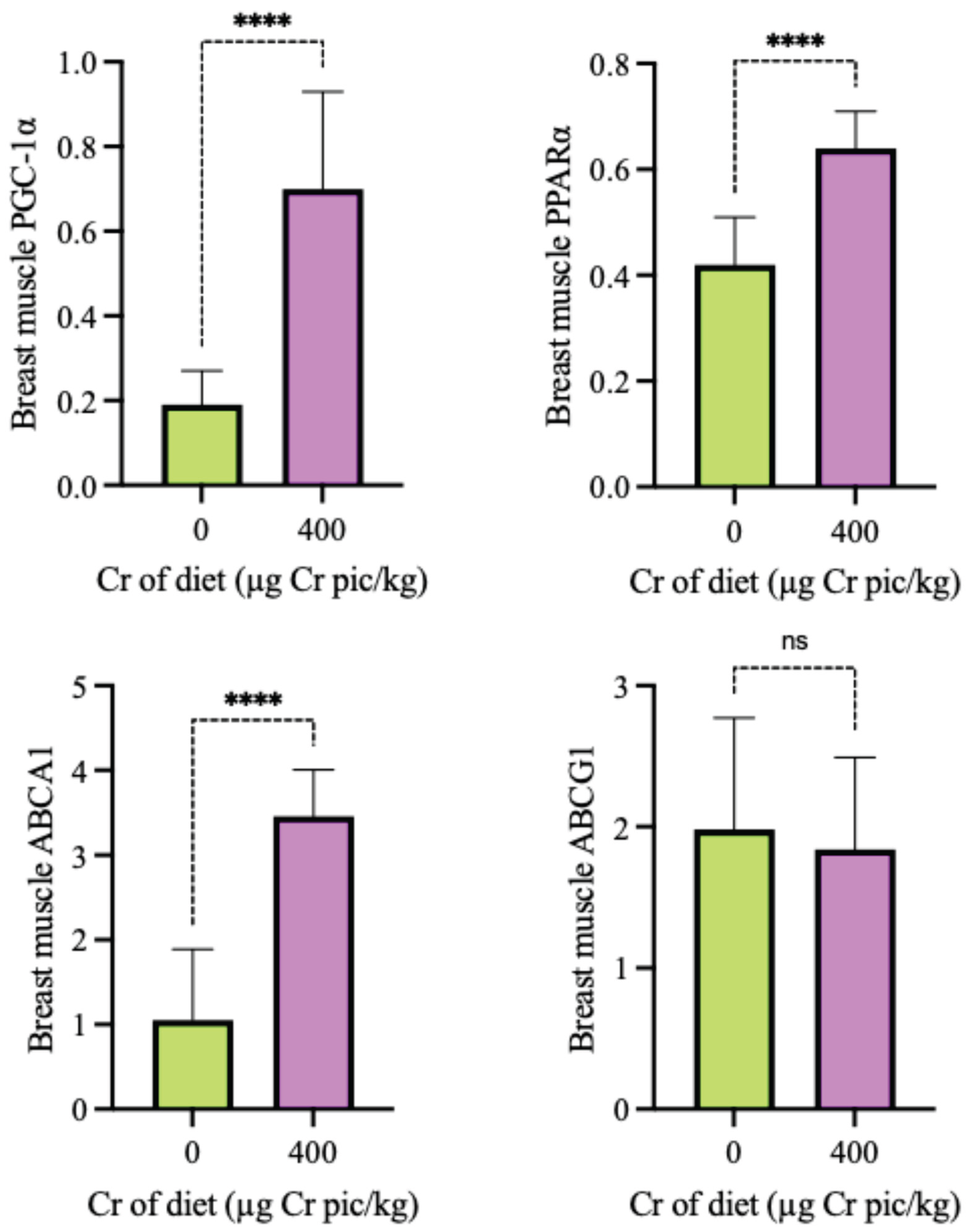
Table 1.
Composition and nutrient levels of the basal diet (as-fed basis) %.
| Items | Content |
|---|---|
| Ingredients | |
| Corn | 56.51 |
| Soybean meal | 35.52 |
| Limestone | 1.00 |
| Soybean oil | 4.50 |
| NaCl | 0.30 |
| dL-Met | 0.11 |
| CaHPO4 | 1.78 |
| Premix1 | 0.28 |
| Total | 100.00 |
| Nutrient levels | |
| ME/(MJ/Kg) | 12.73 |
| CP | 20.07 |
| AP | 0.40 |
| Ca | 0.90 |
| Met | 0.42 |
| Lys | 1.00 |
| Met+Cys | 0.78 |
1 Premix provided the following per kg of the diet: VA 10 000 IU, VD3 3 400 IU, VE 16 IU, VB1 2.0 mg, VK3 2.0 mg, VB2 6.4 mg, VB12 0.012 mg, VB6 2.0 mg, choline 500 mg, pantothenic acid calcium 10 mg, folic acid 1 mg, biotin 0.1 mg, nicotinic acid 26 mg, Zn (ZnSO₄·7H₂O) 40 mg, Mn (MnSO₄·H₂O) 80 mg, Fe (FeSO₄·7H₂O) 80 mg, I (KI) 0.35 mg, Cu (CuSO₄·5H₂O) 8 mg, Se (Na₂SeO₃) 0.15 mg.
Table 2.
Primers used for quantitative RT-PCR.
| Primer name1 | Primer sequence 5’-3’2 | Product size (bp) | GenBank accession number |
|---|---|---|---|
| β-actin | F: CTGTGTTCCCATCTATCGT | 270 | NM_205518.2 |
| R: TCTTCTCTCTGTTGGCTTTG | |||
| ABCA1 | F: TCATCCACCGCCGCCACATT | 223 | NM204145 |
| R: GGCTGAGGAAGGCACTGAAGTC | |||
| ABCG1 | F: AACCAGTGGCTTGGATAGTGC | 298 | XM_025145525.1 |
| R: CCTTACCAGTCGGCTGTTCTG | |||
| PPARα | F: CAAACCAACCATCCTGACGAT | 22 | NM_001001464.1 |
| R: GGAGGTCAGCCATTTTTTGGA | |||
| PGC-1α | F: TGCAGCGCGATCTGAATG | 110 | NM_001006457.1 |
| R: GTTCTTGTCCTTGAGCCACTGAT | |||
| R: CAAGACTGACTGTGAAGGCATCCA |
1β-actin, beta-actin. 2F, forward; R, reverse.
Table 3.
Effects of chromium picolinate supplementation on growth performance of broilers under heat stress from 29 d to 42 d of age. 1.
Table 3.
Effects of chromium picolinate supplementation on growth performance of broilers under heat stress from 29 d to 42 d of age. 1.
| Items2 | 0 | 400 | 800 | P-value |
| ADFI/g | 97.21b ± 4.22 | 105.02a ± 2.61 | 108.55a ± 5.56 | 0.02 |
| ADG/g | 56.37b ± 3.15 | 64.37a ± 1.58 | 68.51a ± 4.11 | <0.01 |
| FCRg/g | 1.73a ± 0.04 | 1.63b ± 0.04 | 1.59b ± 0.06 | <0.01 |
| Breast muscle rate (%) | 17.85b ± 1.27 | 19.18a ± 1.18 | 18.90a ± 0.7 | 0.01 |
| abdominal fat rate (%) | 0.77a ± 0.20 | 0.59b ± 0.22 | 0.56b ± 0.10 | 0.02 |
a-b Means within a column with different superscripts are different at P < 0.05. 1 All means are reported as means ± SD. 2ADFI = average daily gain; ADG = average feed intake; FCR = ADFI/ADG.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



