
Submitted:
05 September 2025
Posted:
05 September 2025
You are already at the latest version
Abstract
Rubisco activase (RCA) is an ATP-dependent enzyme that plays a crucial role in plant stress responses by regulating the catalytic activity of Rubisco. However, the alternative splicing and functional characteristics of the RCA gene exhibit notable species-specific diversity. The variable splice forms and functions of the RCA gene in mangrove plants remain poorly understood. We herein cloned the RCA genes in the leaves of mangrove plant Kandelia candel (L.) in response to combined flooding and salinity stress, and performed systematic expression analysis and functional validation. Our results demonstrated that the RCA gene undergoes alternative splicing to produce two isoforms, designated as KcRCAl (GenBank accession: MG492021) and KcRCAs (GenBank accession: MG492022), respectively. The KcRCAl encodes a 440-amino acid protein (42.49 kDa) belonging to the β-isoforms, while KcRCAs encodes a 474-amino acid protein (46.10 kDa) classified as the α-isoforms. Moreover, protein structure analysis revealed that both isoforms contain phosphorylation and lysine acetylation modification sites. Phylogenetic analysis indicated that KcRCA shares the closest evolutionary relationship with RCA from Cicer arietinum (chickpea) and Durio zibethinus (durian). Furthermore, RT-qPCR analysis revealed that the expression levels of KcRCAl and KcRCAs were significantly upregulated in K. Candel leaves under the combined stress condition. The following functional validation studies in transgenic Arabidopsis demonstrated that overexpression of the KcRCA genes enhances the plant's tolerance to resist flooding and salinity stress while improving antioxidant capacity, increased RCA and Rubisco activities, thereby maintaining photosynthetic efficiency under combined flooding and salinity stress. Our study not only provides new experimental evidence for understanding the molecular mechanisms of plant flooding and salinity stress, but also offers theoretical foundations for breeding flooding-and salinity-tolerant crops.
Keywords:
Kandelia candel
; flooding and salinity
; combined stress
; Rubisco activase
; gene cloning
; functional analysis
1. Introduction
Flooding and salinity stress are critical abiotic factors that severely impair plant growth and agricultural productivity [14]. In recent years, driven by intensified climate change and soil salinization, significant progress has been made in understanding plant responses to either flooding or salinity stress alone, including identifying key physiological adaptations and stress-related genes [2,3,4]. However, in natural environments (e.g., in coastal farmlands), these stresses often occur simultaneously, and their combined effects pose a more complex challenge [5]. How do plants integrate molecular and physiological responses to cope with the combined flooding and salinity stress, and what are the crucial antibiotic resistance genes (ARGs) involved in these adaptive mechanisms? Addressing this question is essential for understanding plant stress resilience and developing stress-resistant crop varieties.
As we know, two typical strategies for adaptation to flooding have been developed in plants over the course of long evolution including ‘escape’ and ‘quiescence’ [6]. The plants with the escape strategy respond to flooding stress by rapid elongation of stems and leaves, enabling plant organs to breach the water surface for access to oxygen and light. In contrast, the quiescence strategy allows plants to maintain essential life processes by reducing metabolic rates and optimizing carbon allocation [7]. Notably, the successful implementation of both strategies is heavily reliant on a continuous supply of energy [8].
Flooding stress often leads to multiple secondary stresses, especially reduced light intensity and impaired gas diffusion, which significantly diminish the efficiency of photosynthesis [9]. It has been reported that the certain aquatic plants have evolved special adaptive structures, such as the continuous gas film formed on the surface of rice leaves, which maintaining effective gas exchange under flooding stress [10].
Furthermore, as a terrestrial plant, maize exhibit a compensatory increase in photosynthetic rate during the initial stages of flooding (within 6 days) [11]. Our research on the mangrove species K. candel further reveals that this species can sustain high photosynthetic efficiency under effects of flooding stress through the synergistic action of protein phosphorylation modifications and transcriptional regulatory networks, continuously accumulating sucrose to provide sufficient carbon skeletons and energy for stress-resistant metabolism [12].
In response to salinity stress, plants also need to mobilize substantial energy for the regulation of ion homeostasis and the synthesis of osmoprotectants [13]. Based on the chloroplast proteome study of K. candel, it was found that salinity stress primarily induces the upregulation of photosystem-related proteins, while key enzymes of the Calvin cycle maintain stable expression. This differential regulation effectively mitigates the inhibition of net photosynthetic rate caused by salinity stress [14]. Therefore, maintaining an adequate energy supply is a crucial pathway for plants to withstand adverse stress conditions [15].
Ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO, EC 4.1.1.39), located in the chloroplast stroma, is the key rate-limiting enzyme in the photosynthetic carbon assimilation pathway, and its activity directly influences plant adaptability to environmental stress [16,17]. The catalytic function of this enzyme requires a specific activation process mediated by Rubisco activase (RCA) [18]. RCA provides energy through ATP hydrolysis, first dissociating ribulose-1,5-bisphosphate (RuBP) bound at the active site of RuBisco, and subsequently promoting the formation of a catalytically active ternary complex (ECM) between RuBisco, CO2, and Mg2+ [19]. As a member of the AAA+ superfamily, RCA exhibits typical ATPase activity [20]. In most higher plants, the RCA gene produces two main transcripts through alternative splicing: RCA1 (long form) and RCA2 (short form), which encode α (45-48 kDa) and β (41-43 kDa) isoforms of different molecular weights, respectively [21]. Phylogenetic analysis indicates that plants such as Arabidopsis [22], rice [23], and soybean [24] express both isoforms, while species like maize [25], tobacco [26], and cucumber [27] retain only the β isoform. It was recently reported that RCA functions as an important regulatory factor in plants in responses to temperature and drought stress [28]. Maintaining photosynthetic homeostasis is a key strategy for plant tolerance to flooding and salinity stress [29]. Our previous studies have confirmed that K. Candel can sustain normal photosynthesis to meet energy demands under individual flooding and salinity stress conditions [12,14]. Therefore, we hypothesize that RCA, which regulates photosynthesis, may play a significant role in the plant's response to the combined flooding and salinity stress.
In the context of global climate change, the genetic engineering of key photosynthetic enzymes has emerged as a significant strategy for enhancing crop yields [30]. As a direct regulatory factor for RuBisco activation, RCA is considered an ideal target for genetic improvement of photosynthesis [31]. Given the substantial differences in activity among the RCAs from various plant species, exploring efficient RCA gene resources from the plants in specialized habitats holds a considerable application potential [32]. However, the alternative splicing and functional characteristics of the RCA gene exhibit notable species-specific diversity [33,34]. The variable splice forms and functions of the RCA gene in mangrove plants that thrive in flooding and saline habitats within the intertidal zone remain poorly understood.
K. candel has evolved unique mechanisms for tolerance combined flooding and salinity stress during its long-term adaptation to intertidal environments, harboring a wealth of gene resources related to flooding and salinity stress [35,36]. Herein, we are the first to isolate the RCA gene from K. candel, identifying two variant splice variants of the RCA gene, designated KcRCAl and KcRCAs. Gene expression analysis and transgenic experiments showed that both KcRCAl and KcRCAs are the key genes involved in alleviating the adverse effects of the flooding and salinity stresses.
2. Materials and Methods
2.1. Materials and Treatments
Seedlings of Kandelia candel (L.) were collected from the Zhangjiangkou Mangrove National Nature Reserve in Zhangzhou, Fujian Province (23°55'N, 117°26'E). Healthy, mature, and similarly sized seedlings were selected and planted in nutrient pots (10 cm × 12 cm) filled with clean sand, with one seedling per pot. A total of 30 pots were arranged in a single plastic basin (42 cm × 30 cm × 14 cm) and irrigated with Hoagland nutrient solution. The seedlings were watered twice daily to replenish evaporated water, and the Hoagland nutrient solution was replaced every three days. After the seedlings developed two pair of true leaves, they were subjected to combined flooding and salinity stress treatments. Building on previous research regarding salinity stress in K. candel, we employed a treatment of 500 mmol•L-1 NaCl in this study [14]. Seedlings with relatively uniform heights (average height difference ± 3 cm) were selected and completely submerged in a Hoagland nutrient solution contenting 500 mmol•L-1 NaCl, with the solution level maintained 5 cm below the surface. Non-stressed seedlings serving as the control. All seedlings were grown in a greenhouse at an air temperature of 27–32 °C under natural sunlight. Leaf samples from both treantment and control groups were collected after 6, 12, 24, and 48 hours of treatment. The collected leaf samples were rapidly frozen in liquid nitrogen for 10 minutes and subsequently stored at -80 °C for RNA extraction. Each biological replicate consisted of 60 seedlings, with three biological replicates for each treatment.
2.2. Cloning of RCA Gene
Total RNA was extracted from the leaves of K. candel subjected to flooding stress using the TaKaRa Mini BEST Plant RNA Extraction Kit, following the manufacturer's instructions. The quality and integrity of the total RNA were assessed using 1% agarose gel electrophoresis and a NanoDrop ND-2000 spectrophotometer. cDNA synthesis and genomic DNA removal were performed using the TransScript® Uni One-Step gDNA Removal and cDNA Synthesis SuperMix, and the products were stored at -20°C. Based on the longest ORF sequence of RCA gene (Gene ID: CL9747Contig1, Supplementary Table S1) established in the preliminary transcriptome database of K. candel [37], cloning primers were designed using Primer Premier 5.0 software (Supplementary Table S2). PCR amplification of cDNA was conducted, followed by purification. The PCR products were ligated into the cloning vector pMD18-T and transformed into competent E. coli Trans 5α cells. Positive clones were screened and sequenced.
2.3. Bioinformatics Analysis of RCA Gene
The relative molecular weight, isoelectric point, and other physicochemical properties of the proteins were analyzed using the ExPASy online tool. The TargetP 1.1 Server was utilized to analyze the transit peptide segments of the proteins. The InterProScan online tool was employed to analyze the domains and functional sites of the amino acid sequences. The NetPhos 3.1 Server was used to predict phosphorylation sites in the proteins. Homologous amino acid sequences were identified using the BLAST tool in NCBI. A phylogenetic tree was constructed using MEGA 6.06 software. The software and websites used for the bioinformatics analysis of KcRCAl and KcRCAs protein structures are listed in Supplementary Table S3.
2.4. Measurement of H2O2 and O2•− Contents and Expression Analysis of KcRCAl and KcRCAs in K. Candel under Combined Stress Condition
The contents of H2O2 and O2•− were measured at defferent time points for both treatment and control groups of K. Candel under combined flooding and salinity stress conditions. The O2•− content was measured using the hydroxylamine oxidation method [38], while H2O2 content was quantified using the Micro Hydrogen Peroxide (H2O2) Assay Kit (Beijing Solarbio Science & Technology Co., Ltd., Beijing, China). Following the methodology described in Section 2.2. RNA was extracted from the leaves of K. candel for both treatment and control groups at various time points, and cDNA was subsequently synthesized. Primers were designed based on the sequenced KcRCAl and KcRCAs sequences, with the Actin gene of K. Candel serving as the internal control. The expression levels of KcRCAl and KcRCAs in the leaves under different treatments were analyzed using RT-qPCR with the SYBR® Premix ExTaq™ (Tli RNaseH Plus) kit, and relative gene expression levels were calculated using the 2−ΔΔCT method. The primer sequences are provided in Supplementary Table S4.
2.5. Arabidopsis Transformation Assay
The Gateway technology was employed to construct the expression vector for Arabidopsis. Primers with BP adapters were designed (Supplementary Table S5), and the target genes KcRCAl and KcRCAs were ligated to the entry vector pDONR207 via BP reaction, resulting in the plasmids pDONR207-KcRCAl and pDONR207-KcRCAs. The target genes were then ligated to the expression vector pEarleyGate104 via LR reaction, yielding the expression vectors pEarleyGate104-KcRCAl and pEarleyGate104-KcRCAs, which were transformed into Agrobacterium tumefaciens EHA105 competent cells to obtain positive strains. The floral dip method was utilized for Agrobacterium-mediated transformation of Arabidopsis, resulting in T0 generation seeds [39]. T0 seeds were screened for resistance to Basta (glufosinate) across multiple generations to obtain T2 generation transgenic plants, which were confirmed as positive by PCR. The transgenic plants were cultivated in a growth chamber maintained at 22 ℃and 16 h light/8 h dark photoperiodand. The expression levels of the RCA genes were assessed by RT-qPCR in T2 transgenic plants and wild-type(WT) plants, with the Actin gene of Arabidopsis serving as the internal control, following the methodology outlined in section 2.4. Primers were designed based on the mRNA sequences of Arabidopsis RCA-α and RCA-β isoform available in the GeneBank database (AEC09714.1, AEC09716.1). The primer sequences are provided in Supplementary Table S4.
2.6. Flooding and Salinity Tolerance Assay of Transgenic Arabidopsis
Thirty-day-old T2 transgenic plants and wild-type(WT) plants were subjected to combined flooding and salinity stress treatment, with a salt concentration of 200 mmol·L-1 NaCl. The treatment method was consistent with that described for K. Candel in Section 2.1. Samples were collected at 0 h, 6 h, 12 h, 24 h, and 48 h, and RT-qPCR was employed to detect the expression levels of the KcRCAl and KcRCAs genes in transgenic plants, following the methodology and primer described in section 2.4. The number of surviving plants was counted separately for transgenic and wild-type plants after 3 and 7 days of treatment, and the survival rates were calculated. The cultivation conditions for both the control and treatment groups were identical to those outlined in Section 2.5. Each biological replicate consisted of 45 seedlings, with three biological replicates for each treatment.
2.7. Determination of Physiological Indexes in Transgenic Arabidopsis under Combined Flooding and Salinity Tolerance
The activities of superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT) in the leaves, as well as the contents of superoxide anion (O2•−) , hydrogen peroxide (H2O2), and malondialdehyde (MDA), were measured. SOD activity was determined using the Hydroxylamine method SOD detection kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China); POD activity was measured using the Guaiacol microplate POD detection kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) ; CAT activity was assessed using the Ammonium molybdate microplate CAT detection kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China); The methods for measuring the contents of H2O2 and O2•− outlined in Section 2.4; and MDA content was determined using the thiobarbituric acid (TBA) method [40]. RCA activity was measured using a kit developed by GENMED, and Rubisco activity was assessed using a kit from Shanghai Enzyme-linked Biotechnology Co., Ltd.
2.8. Statistical Analysis of Data
Each measurement was performed with at least three biological replicates, and all data are presented as mean ± standard deviation. Statistical significance was determined using ANOVA or the Student’s t-test via GraphPad Prism 8 software, and difference at p < 0.05 and p < 0.01 was used to determine statistical significance.
3. Results
3.1. Isolation and Characterization of Two Distinct RCA Gene Isoforms in Kandelia Candel Leaves
The RCA gene was cloned from the K. candel leaves after being subjected to combined flooding and salinity stress for 6 hours, followed by bioinformatics analysis was performed. Using cDNA from the leaves of K. candel as a template, a 1500 bp gene fragment was successfully amplified via RT-PCR. The PCR product underwent an A-tailing reaction and was ligated into the cloning vector pMD18-T, followed by transformation into E. coli Trans 5α (Supplementary Figure S1). Positive clones were screened and sequenced, revealing the presence of two distinct RCA gene isoforms in K. candel: a long-chain isoform (KcRCAl, GenBank accession: MG492021) and a short-chain isoform (KcRCAs, GenBank accession: MG492022). The cDNA fragment length of the KcRCAl gene was 1461 bp, while that of the KcRCAs gene was 1425 bp, indicating a difference of 36 nucleotides between them. Furthermore, 30 positive clones from each treatment group (Control and Treatment) were randomly selected for sequencing, revealing that the occurrence probability of KcRCAl was approximately 1:3 compared to KcRCAs (Supplementary Table S6), suggesting a higher abundance of KcRCAs in K. candel.
The cDNA sequences of KcRCAl and KcRCAs were analyzed using ORF Finder software, which revealed that KcRCAl contains a premature stop codon, terminating translation at the 441 st amino acid. In contrast, the cDNA sequence of KcRCAs encodes 474 amino acid residues, with 34 additional residues at the C-terminus compared to KcRCAl. The KcRCAs protein sequence contains 37 differing amino acid residues (Figure 1): QAAQQVNLPVPPGCTDPAATNYDPTARSDDGSC.
Analysis of the physicochemical properties, including relative molecular weight and isoelectric point (Supplementary Table S7), indicated notable differences between KcRCAl and KcRCAs. The KcRCAl exhibited greater stability than KcRCAs, with KcRCAl classified as a stable protein and KcRCAs as an unstable protein.
Using the TargetP 1.1 Server, the amino acid sequences of KcRCAl and KcRCAs were analyzed, revealing that the first 58 amino acid residues (relative molecular weight of 5620.42 Da) are predicted to function as a chloroplast transit peptide. The mature KcRCAl protein is estimated to have a relative molecular weight of 42.49 kDa, suggesting it corresponds to the β isoforms of RCA (41-43 kDa), while the mature KcRCAs protein is estimated to have a relative molecular weight of 46.10 kDa, indicating it corresponds to the α isoforms of RCA (45-48 kDa). Thus, it can be inferred that the RCA gene in K. candel leaves encodes both α and β isoforms.
InterProScan analysis of the domains and functional sites of KcRCAl and KcRCAs revealed that the amino acid sequence from positions 138 to 332 contains a P-loop nucleoside triphosphate hydrolase functional domain, while the sequence from positions 163 to 304 corresponds to an ATPase, AAA-type, core functional domain, indicating ATP-binding biological function and belonging to the AAA+ superfamily of proteins (Supplementary Figure S2). The C-terminus of KcRCAl lacks conserved sequences; however, the C-terminus of KcRCAs contains a unique C-extension structure specific to the α isoforms: DPAATNYDPTARSDDGSC, which includes two cysteine residues (Cys) at positions 451 and 470 in the amino acid sequence, further confirming that KcRCAs is an α isoform and KcRCAl is a β isoform.
Online analysis using the NetPhos 3.1 Server predicted four phosphorylation sites in both KcRCAl and KcRCAs proteins, including one serine (Ser) phosphorylation site, one threonine (Thr) phosphorylation site, and two tyrosine (Tyr) phosphorylation sites. This finding is consistent with the phosphoproteomic data obtained from K. candel leaves [12].
NCBI BLAST sequence alignment analysis indicated that the sequences share over 80% similarity with 24 homologous amino acid sequences encoded by RCA genes. A phylogenetic tree constructed using MEGA 6.06 software, which included RCA genes from 24 plant species, revealed that KcRCAl (MG492021) and KcRCAs (MG492022) are most closely related to those of Cicer arietinum and Durio zibethinus, clustering with Populus euphratica, while showing more distant relationships with the RCA-encoding amino acid sequences of 20 other species, indicating they belong to two different subfamilies (Figure 2).
3.2. Analysis of H2O2 and O2•− Contents and KcRCA Gene Expression Levels in Kandelia candel Leaves Under Combined Stress of Flooding and Salinity
To assess the effects of combined flooding and salinity stress on the growth of K. candel, the contents of hydrogen peroxide (H2O2) and superoxide anions (O2•−) in the leaves were measured at 6, 12, 24, and 48 hours post-treatment, and morphological changes of the plants were observed and photographed after 48 hours. The results indicated that 6 hours after the stress treatment, the contents of H2O2 and O2•− in the leaves of K. candel were significantly higher than those in the control group, and remaining elevated thereafter (Figure 3, A, B). Notably, the seedlings of K. candel exhibited significant stress responses within 6 hours of combined flooding and salinity stress. After 48 hours, the leaves of the treatment group displayed slight wilting, although no obvious color changes were observed. Upon excavation, it was noted that the roots of the treatment group had turned black, with some fine roots exhibiting signs of rot (Figure 3, C, D). This indicates that the combined stress on K. candel for 48 hours resulted in significant phenotypic changes.
To verify the expression of the two splice variants of the RCA gene under stress conditions, this study analyzed the expression levels of KcRCAl and KcRCAs in the leaves of K. candel at various stress time points in both the treatment and control groups using RT-qPCR. Under flooding and salinity stress treatment, the expression levels of KcRCA in the leaves of K. candel demonstrated that combined stress significantly promoted the overall expression levels of both KcRCAl and KcRCAs (Figure 3, E, F). Notably, short-term flooding ( 6 hours) resulted in a significant enhancement of KcRCAl and KcRCAs expression levels in the leaves, which is advantageous for the plant's adaptation to flooded and saline environments.
3.3. Establishment and Molecular Confirmation of Arabidopsis Transgenic Plants
In this study, the RCA gene was transferred into Arabidopsis for functional verification, as no established genetic transformation system currently exists for K. candel. Transgenic Arabidopsis was successfully obtained through herbicide resistance and PCR identification. The results indicated that transgenic plants expressing KcRCAl exhibited a specific band of approximately 1500 bp (Supplementary Figure S3), while transgenic plants expressing KcRCAs also displayed a specific band of about 1500 bp (Supplementary Figure S4). No corresponding bands were amplified in wild-type Arabidopsis. Additionally, a specific band of approximately 1500 bp was cloned using the expression vector plasmid as a template. The PCR products from positive plants were sent for sequencing, and the results were consistent with the PCR detection, confirming that the exogenous genes KcRCAl and KcRCAs have been successfully integrated into the transgenic Arabidopsis. The results of vector construction (Supplementary Figure S5, 6) and Agrobacterium PCR detection (Supplementary Figure S7) are presented in the supplementary materials.
Through generation resistance screening, T2 generation transgenic Arabidopsis seedlings (Supplementary Figure S8,Supplementary Figure S9) were obtained and utilized for resistance verification of the transgenic lines. We analyzed the phenotypic characteristics and morphology of the transgenic plants. No significant differences in seedling were observed between the wild-type and transgenic plants (Figure 4, A-C). The RT-qPCR data indicated that the expression levels of the KcRCAl and KcRCAs genes in the transgenic plants were significantly higher than those in the wild-type plants (Figure 4, D, E). These results confirm the successful generation of transgenic Arabidopsis plants that overexpress the KcRCAl and KcRCAs genes.
3.4. Transgenic Arabidopsis Plants Exhibited Enhanced Tolerance to Flooding and Salinity Stress
The performance of wild-type and transgenic Arabidopsis seedlings were submerged in 200 mmol·L⁻¹ NaCl solution for flooding and salinity trentment. On Day 3 of stress, wild-type Arabidopsis leaves exhibited significant purpling and wilting, whereas the transgenic lines thrived(Figure 5, A). By monitoring the survival rates of the plants subjected to flooding and salinity stress treatments over several days, it was observed that, for the same duration of treatment, the survival rate of the transgenic plants was significantly higher than that of the wild-type plants. Furthermore, no significant difference was found between the two transgenic lines.(Figure 5,B, C). These phenotypic results indicate that the transgenic plants possess a higher tolerance to combined flooding and salinity stress compared to the wild-type plants.
In this study, RT-qPCR technology was employed to analyze the expression levels of the KcRCAl and KcRCAs genes in transgenic plants under combined flooding and salinity stress at various time points. The results indicated that the expression level of KcRCAl and KcRCAs genes were upregulate in the transgenic plants after 6 hours of treatment, remaining significantly higher than those in the control group. Although expression level decreased after 48 hours of treatment, they remained elevated compared to the control group.(Figure 6). The upregulation of RCA gene expression may be associated with the enhanced tolerance to combined flooding and salinity stress observed in the transgenic plants.
3.5. Physiological Analysis of Transgenic Arabidopsis Under the Combined Flooding and Salinity Stress
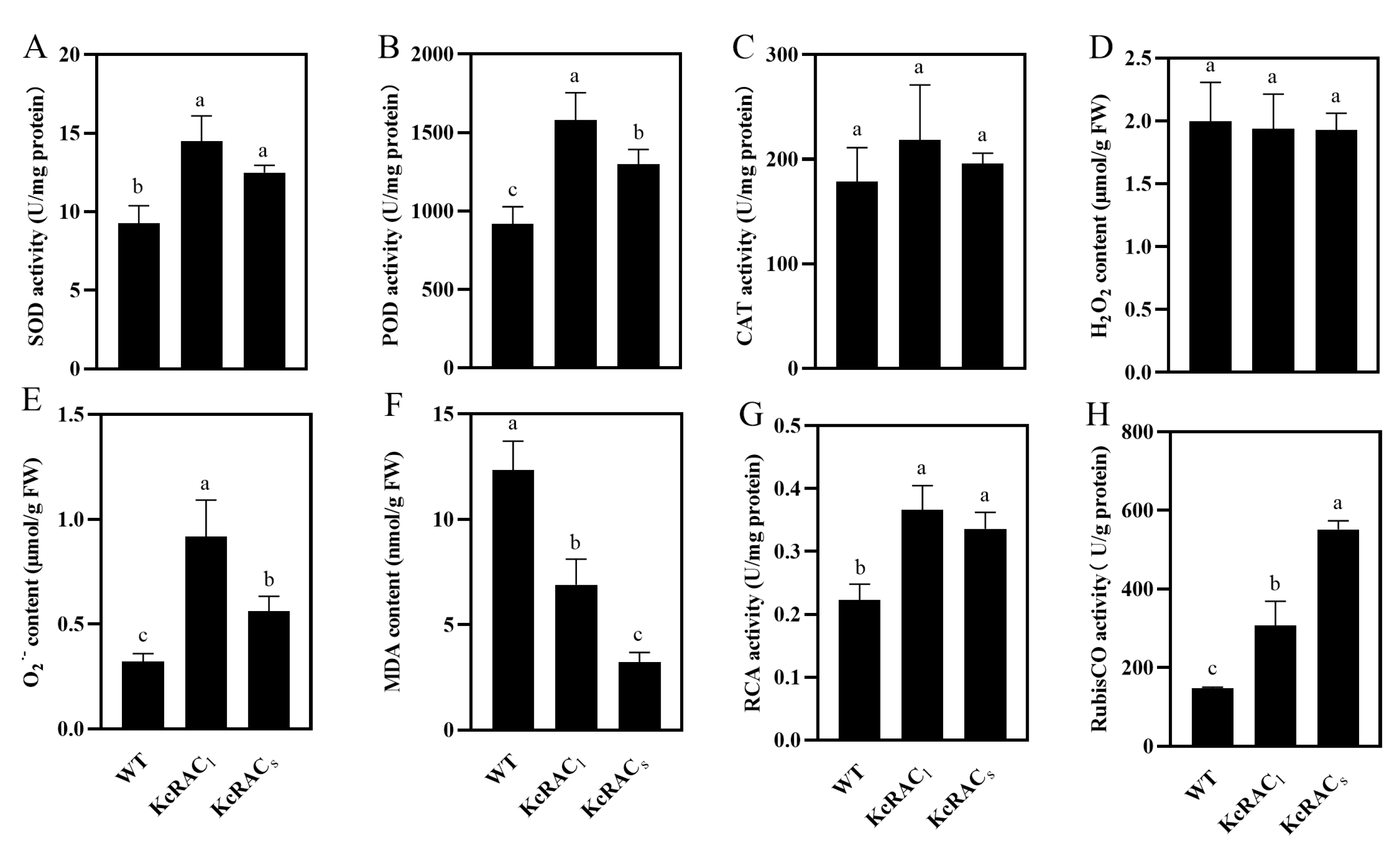
Physiological assessments of the transgenic plants' resistance for combined flooding and salinity tress and key photosynthetic enzyme activities revealed that, compared to wild-type plants, the activities of superoxide dismutase (SOD) and peroxidase (POD) in transgenic plants were significantly increased (Figure 7A,B), while catalase (CAT) activity did not exhibit significant changes (Figure 7 C). The content of hydrogen peroxide (H2O2) did not change significantly (Figure 7 D), whereas superoxide anion (O2•¯) content significantly increased (Figure 7 E) Meanwhile, malondialdehyde (MDA) content significantly decreased (Figure 7 F). These results indicate that the antioxidant capacity of plants expressing KcRCAl or KcRCAs has been enhanced to some extent. Furthermore, compared to wild-type plants, RCA activity in transgenic plants significantly increased, with Rubisco activity enhanced by 1.09-fold and 2.75-fold in KcRCAl and KcRCAs transgenic plants, respectively (Figure 7 G,H). This suggests that KcRCAl or KcRCAs contributes to the improvement of photosynthetic efficiency in transgenic plants.
4. Discussion
4.1. Successfully Cloned Two Variable Splice Variants of RCA Gene from a Mangrove Plant
Combined stress presents a significant challenge to successful crop production, and the gene resources that enable plants to respond to such stresses are often found in species that thrive in specialized or extreme environments[41]. For instance, K. candel grows in coastal wetlands and has long been subjected to high levels of flooding and salinity stress. Consequently, it has developed a comprehensive suite of adaptive mechanisms to cope with the combined challenges of flooding and salinity, harboring a wealth of stress-resistant genes [42]. Photosynthesis, as the central process of plant energy metabolism, plays a crucial role in crop adaptation to environmental stress [43]. Previous studies have demonstrated that K. candel can sustain high photosynthetic efficiency by upregulating the expression of photosynthesis-related proteins while stabilizing the expression of key enzymes in the Calvin cycle, including Rubisco and RCA, in response to moderate salinity stress (100-200 mM NaCl) and flooding stress [10,31]. In this context, Rubisco serves as the key rate-limiting enzyme that catalyzes the carboxylation reaction of CO₂ with ribulose-1,5-bisphosphate (RuBP), and its activation is critically dependent on ATP hydrolysis mediated by Rubisco activase (RCA) [44,45]. This study successfully cloned two variable splice variants of RCA, KcRCAl and KcRCAs, from the leaves of K. candel for the first time. Bioinformatics analysis revealed that KcRCAl encodes 440 amino acids (42.49 kDa) and KcRCAs encodes 474 amino acids (46.10 kDa), both containing typical ATPase functional domains (P-loop NTP hydrolase domain and AAA+ ATPase core domain) [46]. Phylogenetic analysis indicated that the RCA of K. candel is most similar to that of Cicer arietinum and Durio zibethinus (>85%), clustering separately from Populus euphratica, while showing lower similarity (<70%) with the RCA genes of 20 other terrestrial plants. This confirms the conservation of the RCA gene and reveals the unique evolutionary characteristics of mangrove plants' RCA in intertidal habitats. As the first RCA gene cloned from a mangrove plant, this study provides important molecular targets for elucidating the mechanisms of salt and flood tolerance in plants.
4.2. Multifunctional Regulation of RCA in the Flooding and Salinity Tolerance Physiology of Kandelia Candel
Recent studies have shown that RCA not only maintains photosynthetic carbon assimilation efficiency by activating Rubisco but also plays a central regulatory role in plant responses to various environmental stresses [47,48]. For instance, in response to temperature stress, heat-tolerant cucumber (Cucumis sativus) significantly upregulated RCA gene expression and enhanced RCA enzyme activity, effectively maintaining Rubisco's catalytic efficiency and allowing net photosynthetic rates to remain above 80% of the control group under high-temperature conditions [27]. Under low-light stress, although the photosynthetic rate of cucumber significantly decreased, the transcription level of CsRCA exhibited a compensatory increase, significantly alleviating the reduction in Rubisco activity [49]. Additionally, in maize (Zea mays) under ozone stress, the mRNA levels of RCA in senescent leaves significantly increased in response to the stress [25]. While RCA genes have been shown to play roles in plant resistance to high temperatures, ozone, and low light, their functions in response to flooding and salinity stress have not been previously reported. Generally, environmental stress inhibits Rubisco activity, leading to a significant decrease in photosynthetic rates; however, RCA can enhance and maintain Rubisco's catalytic activity to regulate photosynthesis under stress [50,51]. This study found that the expression levels of KcRCA in the leaves of K. candel significantly increased under flooding and high salinity stress, suggesting that K. candel may compensate for the adverse effects of stress on Rubisco activity by increasing RCA expression to maintain normal photosynthesis under environmental stress. These results indicate that KcRCA in the leaves of K. candel plays a crucial role in resisting flooding and salinity stress, a finding that has been confirmed for the first time in salt-tolerant and flood-tolerant mangrove plants.
4.3. Molecular Characteristics and Functional Roles of Alternatively Spliced Isoforms of RCA in Kandelia Candel
The variable splicing of plant RCA genes exhibits significant species diversity. Phylogenetic analysis indicates that different plant groups have evolved unique regulatory mechanisms for RCA expression: in most higher plants (such as Arabidopsis, rice, spinach, and barley) [21,23,52,53], variable splicing of a single gene produces two isoforms of 41 kDa and 45 kDa; some species (such as tobacco and cucumber) retain only the expression of the smaller isoform [26,27]; while maize has evolved a unique 3' UTR variable splicing mechanism that generates different isoforms through post-translational modifications [54]. Additionally, plants like cotton and soybean have formed two independent RCA genes through gene duplication events [47,55]. This study reveals for the first time the variable splicing mechanism of RCA in the mangrove plant K. candel, identifying two transcripts, KcRCAl and KcRCAs, produced through alternative splicing. KcRCAl forms a smaller β isoform (42.49 kDa) via a premature stop codon (UAA), while KcRCAs encodes a complete α isoform (46.10 kDa). Notably, the α isoform contains two highly conserved cysteine residues (Cys247 and Cys392), suggesting its potential involvement in redox signaling. In response to environmental stress, K. candel can precisely maintain an α:β ratio of approximately 3:1(Supplymentary Table S6), a regulatory pattern that aligns with the reported functional division mechanism of RCA in rice [23], where the α isoform primarily activates Rubisco while the β isoform is responsible for maintaining its active state. These findings not only expand our understanding of the functional diversity of plant RCA but also provide new theoretical foundations for elucidating the unique adaptation strategies of intertidal plants. However, the specific molecular functions and regulatory networks of the two isoforms require further investigation.
4.4. Biotechnological Potential of RCA Gene Derived from Kandelia Candel for Enhancing Flooding and Salinity Tolerance in Crops
In recent years, RCA gene engineering has emerged as a significant strategy for enhancing crop photosynthetic efficiency and stress resistance. Multiple studies have demonstrated that exogenous overexpression of RCA genes can significantly improve the photosynthetic performance of transgenic plants [56]. For instance, overexpression of endogenous RCA genes in Arabidopsis and rice has been shown to enhance photosynthetic efficiency and improve adaptability to temperature stress and low-light environments [57,58,59]. This study is the first to clone the RCA gene from the highly salt- and flood-tolerant mangrove plant K. candel and validate its function through a heterologous expression system in Arabidopsis, successfully achieving RCA gene overexpression in Arabidopsis (Figure 4, D, E). The phenotypic analysis demonstrated that the transgenic plants exhibited significantly higher resistance to flooding and salinity stress compared to the wild type (Figure 5, A). Under the same stress duration, the survival rate of the transgenic lines was significantly higher than that of the wild type (Figure 5, B, C). These results indicate that overexpression of the KcRCA gene can enhance the tolerance of Arabidopsis to flooding and salinity stress. Meanwhile, this study also indicated that transgenic plants exhibited significantly enhanced antioxidant capacity, improved RCA enzyme activity, and better Rubisco activation status, thereby maintaining higher photosynthetic efficiency under stress conditions. These findings provide the first evidence that the RCA gene from mangrove plants possesses superior stress response characteristics, offering new high-quality gene resources for crop stress resistance breeding and pioneering a novel approach to crop improvement using genes from plants in unique habitats. Future research could validate the function of KcRCA in major crops, elucidate its molecular regulatory network, and develop molecular marker-assisted breeding technologies based on RCA. This study provides an important theoretical and technical foundation for utilizing specialized plant gene resources for crop improvement.
5. Conclusions
In the present study, we successfully cloned the RCA gene from the mangrove plant K. candel and identified two variable splice variants, KcRCAl and KcRCAs, which encode a β isoform of 440 amino acids and an α isoform of 474 amino acids, respectively. Bioinformatics analysis revealed that both isoforms contain multiple phosphorylation sites and lysine acetylation sites, suggesting potential regulation by post-translational modifications. Phylogenetic analysis indicated that KcRCA is most closely related to the RCA genes of chickpeas and durians (similarity > 85%). Expression analysis demonstrated that combined flooding and salinity stress significantly induced the upregulation of KcRCAl and KcRCAs expression. Through a heterologous expression system in Arabidopsis, it was confirmed that plants expressing the KcRCA gene exhibited significantly enhanced antioxidant capacity, with RCA and Rubisco activities markedly increased, thereby maintaining photosynthetic efficiency under flooding and salinity stress. Our study firstly clarified the molecular mechanism at the molecular level by which mangrove plants adapt to the unique intertidal habitat through the regulation of RCA gene expression. It not only expanded the molecular theoretical basis for plant responses to cope with combined flooding and salinity stress, but also provided gene resources of significant application value for the genetic improvement of crops against stress.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: PCR identification of recombinant clones of pMD18-T-KcRCA; Figure S2: Conserved domains prediction for KcRCA; Figure S3: PCR assay of transgenic Arabidopsis thaliana with KcRCAl; Figure S4: PCR assay of transgenic Arabidopsis thaliana with KcRCAs; Figure S5: PCR identification of pDONR207-KcRCA; Figure S6: PCR identification of pEarleyGate104-KcRCA; Figure S7: PCR identification of transgenic Agrobacterium tumefaciens with pEarleyGate104-KcRCA; Figure S8: Transgenic Arabidopsis plants overexpressing KcRCAl and KcRCAs in a wild-type (WT) background; Figure S9: PCR analysis of T2 transgenic Arabidopsis thaliana containing the KcRCA gene. Table S1: Transcriptome and CDS sequence of CL9747Contig; Table S2: Primers used to amplify the complete KcRCA ORF; Table S3: The software and websites used for bioinformation analysis for the protein structure of KcRAC; Table S4: Primers used for RT-qPCR; Table S5: Primers used for Gateway Cloning (BP reaction); Table S6: The probability of KcRCAl and KcRCAs in Control and Flooding groups; Table S7: Physicochemical property of KcRCAl and KcRCAs proteins.
Author Contributions
Y.Z., J.Z. and J.X. conceived and designed the experiments; J.X. performed the gene cloning, transformation experiments, physiological assay, and wrote the manuscript; D.P. collected the plant materials, performed real-time PCR; C.L. analyzed the data; W.C. and S.Y. revised the manuscript critically. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Chinese National Natural Science Funds (Grant no. 331070542), the Natural Science Foundation of Fujian Province, China (Grant no. 2024J01892).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article or supplementary materials. The original contributions presented in this study are included in the article or supplementary material. Further inquiries can be directed toward the corresponding author.
Conflicts of Interest
The authors declare that have no conflicts of interest regarding the work in this study.
References
- Gui, G.; Zhang, Q.; Hu, W.; Liu, F. Application of multiomics analysis to plant flooding response. Front. Plant Sci. 2024, 15, 1389379. [Google Scholar] [CrossRef]
- Manghwar, H.; Hussain, A.; Alam, I.; Khoso, M.A.; Ali, Q.; Liu, F. Waterlogging stress in plants: Unraveling the mechanisms and impacts on growth, development, and productivity. Environ. Exp. Bot. 2024, 224, 105824. [Google Scholar] [CrossRef]
- Ngumbi, E.N. Could flooding undermine progress in building climate-resilient crops? Trends Plant Sci. 2025, 30, 85–94. [Google Scholar] [CrossRef]
- Komatsu, S.; Hiraga, S.; Yanagawa, Y. Proteomics techniques for the development of flood tolerant crops. J. Proteome Res. 2012, 11, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Jhan, L.H.; Yang, C.Y.; Huang, C.M.; Lai, M.C.; Huang, Y.H.; Baiya, S.; Kao, C.F. Integrative pathway and network analysis provide insights on flooding-tolerance genes in soybean. Sci. Rep. 2023, 13, 1980. [Google Scholar] [CrossRef]
- Li, G.J.; Wei, N.; Hou, H. Uncovering the secrets of how plants adapt to water stress. Plant Cell Environ. 2025, 10. [Google Scholar] [CrossRef]
- Chakraborty, K.; Ray, S.; Vijayan, J.; Molla, K.A.; Nagar, R.; Jena, P.; Mondal, S.; Panda, B.B.; Shaw, B.P.; Swain, P.; Chattopadhyay, K.; Sarkar, R.K. Preformed aerenchyma determines the differential tolerance response under partial submergence imposed by fresh and saline water flooding in rice. Physiol. Plant. 2021, 173, 1597–1615. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.S.; Li, Y.F.; Cai, Y.H.; Xie, Y.H.; Deng, Z.M.; Li, F.; Hou, Z.Y. Differential strategies to tolerate flooding in polygonum hydropiper plants originating from low- and high-elevation habitats. Front. Plant Sci. 2019, 9, 1970. [Google Scholar] [CrossRef]
- Silveira, J.A.G.; Carvalho, F.E.L. Proteomics, photosynthesis and salt resistance in crops: An integrative view. J. Proteomics. 2016, 143, 24–35. [Google Scholar] [CrossRef]
- Lin, C.; Zhang, Z.; Shen, X.; Liu, D.; Pedersen, O. Flooding-adaptive root and shoot traits in rice. Funct. Plant Biol. 2024, 51, FP23226. [Google Scholar] [CrossRef]
- Qi, M.; Liu, X.; Li, Y.; Song, H.; Yin, Z.; Zhang, F.; He, Q.; Xu, Z.; Zhou, G. Photosynthetic resistance and resilience under drought, flooding and rewatering in maize plants. Photosynth Res. 2021, 148, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Wang, L.; Tan, F.; Lu, S.; Lv, X.; Zaynab, M.; Cheng, C.L.; Abubakar, Y. S; Chen, S; Chen, W. Phosphoproteomics unveils stable energy supply as key to flooding tolerance in Kandelia candel. J. Proteomics. 2018, 176, 1–12. [Google Scholar] [CrossRef]
- Guo, Z.; Ma, D.; Li, J.; Wei, M.; Zhang, L.; Zhou, L.; Zhou, X.; He, S.; Wang, L.; Shen, Y.; Li, Q.Q.; Zheng, H.L. Genome-wide identification and characterization of aquaporins in mangrove plant Kandelia obovata and its role in response to the intertidal environment. Plant Cell Environ. 2022, 45, 1698–1718. [Google Scholar] [CrossRef]
- Wang, L.; Liang, W.; Xing, J.; Tan, F.; Chen, Y.; Huang, L.; Cheng, C.L.; Chen, W. Dynamics of chloroplast proteome in salt-stressed mangrove Kandelia candel (L.) Druce. J. Proteome Res. 2013, 12, 5124–36. [Google Scholar] [CrossRef]
- Nowicka, B.; Ciura, J.; Szymańska, R.; Kruk, J. Improving photosynthesis, plant productivity and abiotic stress tolerance-current trends and future perspectives. J. Plant Physiol. 2018, 231, 415–433. [Google Scholar] [CrossRef]
- Portis, Jr.A.R. Rubisco activase-Rubisco's catalytic chaperone. Photosynth Res. 2003, 75, 11–27. [Google Scholar] [CrossRef]
- Sharkey, T.D. The discovery of rubisco. J. Exp. Bot. 2023, 74, 510–519. [Google Scholar] [CrossRef]
- Portis, Jr.A.R.; Li, C.; Wang, D.; Salvucci, M.E. Regulation of Rubisco activase and its interaction with Rubisco. J. Exp. Bot. 2008, 59, 1597–604. [Google Scholar] [CrossRef]
- Hayer-Hartl, M.; Hartl, F.U. Chaperone machineries of Rubisco-the most abundant enzyme. Trends Biochem. Sci. 2020, 45, 748–763. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.Y.; Thieulin-Pardo, G.; Hartl, F.U.; Hayer-Hartl, M. Rubisco activases: AAA+ Chaperones adapted to enzyme repair. Front. Mol. Biosci. 2017, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Werneke, J.M.; Chatfield, J.M.; Ogren, W.L. Alternative mRNA splicing generates the two ribulosebisphosphate carboxylase/oxygenase activase polypeptides in spinach and Arabidopsis. Plant Cell. 1989, 1, 815–25. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Stessman, D.J.; Wright, D.A.; Spalding, M.H.; Huber, S.C.; Ort, D.R. Arabidopsis plants expressing only the redox-regulated Rca-α isoform have constrained photosynthesis and plant growth. Plant J. 2020, 103, 2250–2262. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, X.; Zhou, Z.; Feng, X.; Yang, W.; Jiang, D. Two Rubisco activase isoforms may play different roles in photosynthetic heat acclimation in the rice plant. Physiol. plantarum. 2010, 139, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Harvey, C.M.; Cavanagh, A.P.; Kim, S.Y.; Wright, D.A.; Edquilang, R.G.; Shreeves, K.S.; Perdomo, J.A.; Spalding, M.H.; Ort, D.R.; Bernacchi, C.J.; Huber, S.C. Removal of redox-sensitive Rubisco activase does not alter Rubisco regulation in soybean. Photosynth. Res. 2022, 154, 169–182. [Google Scholar] [CrossRef]
- Leitao, L.; Maoret, J.J.; Biolley, J.P. Changes in PEP carboxylase, rubisco and rubisco activase mRNA levels from maize (Zea mays) exposed to a chronic ozone stress. Biol. Res. 2007, 40, 137–53. [Google Scholar] [CrossRef]
- Khairy, A.I.H.; Oh, M.J.; Lee, S.M.; Kim, D.S.; Roh, K.S. Nitric oxide overcomes Cd and Cu toxicity in in vitro-grown tobacco plants through increasing contents and activities of Rubisco and Rubisco activase. Biochim. Open. 2016, 2, 41–51. [Google Scholar] [CrossRef]
- Bi, H.; Liu, P.; Jiang, Z.; Ai, X. Overexpression of the rubisco activase gene improves growth and low temperature and weak light tolerance in Cucumis sativus. Physiol. Plant. 2017, 161, 224–234. [Google Scholar] [CrossRef]
- Li, X.; Yue, H.; Chu, Y.; Jia, Y. Comparative transcriptomes reveal molecular mechanisms of apple blossoms of different tolerance genotypes to chilling injury. Open Life Sci. 2023, 18, 20220613. [Google Scholar] [CrossRef]
- Carmo-Silva, A.E.; Salvucci, M.E. The regulatory properties of Rubisco activase differ among species and affect photosynthetic induction during light transitions. Plant Physiol. 2013, 161, 1645–55. [Google Scholar] [CrossRef]
- Nazari, M.; Kordrostami, M.; Ghasemi-Soloklui, A.A.; Eaton-Rye, J.J.; Pashkovskiy, P.; Kuznetsov, V.; Allakhverdiev, S.I. Enhancing photosynthesis and plant productivity through genetic modification. Cells. 2024, 13, 1319. [Google Scholar] [CrossRef]
- Wijewardene, I.; Shen, G.; Zhang, H. Enhancing crop yield by using Rubisco activase to improve photosynthesis under elevated temperatures. Stress Biol. 2021, 1, 2. [Google Scholar] [CrossRef]
- Sparrow-Muñoz, I.; Chen, T.C.; Burgess, S.J. Recent developments in the engineering of Rubisco activase for enhanced crop yield. Biochem. Soc. Trans. 2023, 51, 627–637. [Google Scholar] [CrossRef]
- Gunn, L.H.; Martin, Avila, E.; Birch, R.; Whitney, S.M. The dependency of red Rubisco on its cognate activase for enhancing plant photosynthesis and growth. Proc. Natl. Acad. Sci. USA. 2020, 117, 25890–25896. [CrossRef]
- Jurczyk, B.; Hura, K.; Trzemecka, A.; Rapacz, M. Evidence for alternative splicing mechanisms in meadow fescue (Festuca pratensis) and perennial ryegrass (Lolium perenne) Rubisco activase gene. J. Plant Physiol. 2015, 176, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Wang, L.; Chen, S.; Lv, X.; Lu, S.; Cheng, C.L.; Tan, F.; Chen, W. Protein acetylation as a mechanism for Kandelia candel's adaption to daily flooding. Tree Physiol. 2018, 38, 895–910. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, X.; Liang, M.; Tan, F.; Liang, W.; Chen, Y.; Lin, Y.; Huang, L.; Xing, J.; Chen, W. Proteomic analysis of salt-responsive proteins in the leaves of mangrove Kandelia candel during short-term stress. PLoS One. 2014, 9, e83141. [Google Scholar] [CrossRef]
- Wang, L.; Pan, D.; Lv, X.; Cheng, C.L.; Li, J.; Liang, W.; Xing, J.; Chen, W. A multilevel investigation to discover why Kandelia candel thrives in high salinity. Plant Cell Environ. 2016, 39, 2486–2497. [Google Scholar] [CrossRef]
- Jambunathan, N. Determination and detection of reactive oxygen species (ROS), lipid peroxidation, and electrolyte leakage in plants. Methods Mol. Biol. 2010, 639, 292–8. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–43. [Google Scholar] [CrossRef]
- Park, S.I.; Kim, J.J.; Shin, S.Y.; Kim, Y.S.; Yoon, H.S. ASR Enhances environmental stress tolerance and improves grain yield by modulating stomatal closure in rice. Front. Plant Sci. 2020, 10, 1752. [Google Scholar] [CrossRef]
- Renziehausen T, Frings S, Schmidt-Schippers R. 'Against all floods': plant adaptation to flooding stress and combined abiotic stresses. Plant J. 2024, 117, 1836–1855. [CrossRef]
- Liu S, Yang S, Liu H, Hu Q, Liu X, Wang J, Wang J, Xin W, Chen Q. Physiological and transcriptomic analysis of the mangrove species Kandelia obovata in response to flooding stress. Mar. Pollut. Bull. 2023, 196:115598. [CrossRef]
- Karthick, P.V.; Senthil, A.; Djanaguiraman, M.; Anitha, K.; Kuttimani, R.; Boominathan, P.; Karthikeyan, R.; Raveendran, M. Improving crop yield through increasing carbon gain and reducing carbon loss. Plants. 2024, 13, 1317. [Google Scholar] [CrossRef] [PubMed]
- Prywes, N.; Phillips, N. R.; Tuck, O. T.; Valentin-Alvarado, L.E.; Savage, D.F. Rubisco function, evolution, and engineering. Annu. Rev. Biochem. 2023, 92, 385–410. [Google Scholar] [CrossRef] [PubMed]
- Waheeda, K.; Kitchel, H.; Wang, Q.; Chiu, P.L. Molecular mechanism of Rubisco activase: dynamic assembly and Rubisco remodeling. Front. Mol. Biosci. 2023, 10, 1125922. [Google Scholar] [CrossRef]
- Nagarajan, R.; Gill, K.S. Evolution of Rubisco activase gene in plants. Plant Mol. Biol. 2018, 96, 69–87. [Google Scholar] [CrossRef]
- Yin, Z.; Meng, F.; Song, H.; Wang, X.; Xu, X.; Yu, D. Expression quantitative trait loci analysis of two genes encoding Rubisco activase in soybean. Plant Physiol. 2010, 152, 1625–1637. [Google Scholar] [CrossRef]
- Chen, Y; Wang, X.M.; Zhou, L.; He, Y.; Wang, D.; Qi, Y.H.; Jiang, D.A. Rubisco activase is also a multiple responder to abiotic stresses in rice. PLoS One. 2015, 10, e0140934.
- Jiang, Z.; Sun, X.; Ai, X.; Wang, M.; Bi, H.; Wang, H. Responses of Rubisco and Rubisco activase in cucumber seedlings to low temperature and weak light. Ying Yong Sheng Tai Xue Bao. 2010, 21, 2045–2050. (in Chinese). [Google Scholar]
- Somkuwar, R.G.; Dhole, A.M. Somkuwar, R.G.; Dhole, A.M. Understanding the photosynthesis in relation to climate change in grapevines. Theor. Biosci. 2025, 10.1007/s12064-025-00435-w. Advance online publication. [CrossRef] [PubMed]
- Amaral, J.; Lobo, A.K.M.; Carmo-Silva, E. Regulation of Rubisco activity in crops. New Phytol, 2024, 241, 35–51. [Google Scholar] [CrossRef]
- Ma, L.; Liu, C.; Qu, C.; Yin, S.; Liu, J.; Gao, F.; Hong, F. Rubisco activase mRNA expression in spinach: modulation by nanoanatase treatment. Biol. Trace Elem. Res. 2008, 122, 168–78. [Google Scholar] [CrossRef]
- Aliakbari, M.; Cohen, S.P.; Lindlöf, A.; Shamloo-Dashtpagerdi, R. Rubisco activase A (RcaA) is a central node in overlapping gene network of drought and salinity in Barley (Hordeum vulgare L.) and may contribute to combined stress tolerance. Plant Physiol. Bioch. 2021, 161, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Suárez, M.; Ayala-Ochoa, A.; Lozano-Franco, J.; García-Torres, I.; Díaz-Quiñonez, A.; Ortíz-Navarrete, V.F.; Sánchez-de-Jiménez, E. Rubisco activase chaperone activity is regulated by a post-translational mechanism in maize leaves. J. Exp. Bot. 2004, 55, 2533–2539. [Google Scholar] [CrossRef]
- Salvucci, M. E.; Van de Loo F., J.; Stecher, D. Two isoforms of Rubisco activase in cotton, the products of separate genes not alternative splicing. Planta. 2003, 216, 736–744. [Google Scholar] [CrossRef]
- Carmo-Silva, A. E.; Salvucci, M.E. The activity of Rubisco's molecular chaperone, Rubisco activase, in leaf extracts. Photosynth Res. 2011, 108, 143–155. [Google Scholar] [CrossRef]
- Feng, Y.; Wu, H.; Liu, H.; He, Y.; Yin, Z. Effects of OsRCA overexpression on Rubisco activation state and photosynthesis in maize. Plants. 2023, 12, 1614. [Google Scholar] [CrossRef] [PubMed]
- Wijewardene, I.; Mishra, N.; Sun, L.; Smith, J.; Zhu, X.; Payton, P.; Shen, G.; Zhang, H. Improving drought-, salinity-, and heat-tolerance in transgenic plants by co-overexpressing Arabidopsis vacuolar pyrophosphatase gene AVP1 and Larrea Rubisco activase gene RCA. Plant Sci. 2020, 296, 110499. [Google Scholar] [CrossRef]
- Suganami, M.; Suzuki, Y.; Tazoe, Y.; Yamori, W.; Makino, A. Co-overproducing Rubisco and Rubisco activase enhances photosynthesis in the optimal temperature range in rice. Plant physiol. 2021, 185, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Masumoto, C.; Fukayama, H.; Makino, A. Rubisco activase is a key regulator of non-steady-state photosynthesis at any leaf temperature and, to a lesser extent, of steady-state photosynthesis at high temperature. Plant J. 2012, 71, 871–880. [Google Scholar] [CrossRef]
Figure 1.
KcRCA’s cDNA sequence and its deduced amino acid sequence.

Figure 2.
Phylogenetic analysis of RCA from Kandelia candel and other plant species.
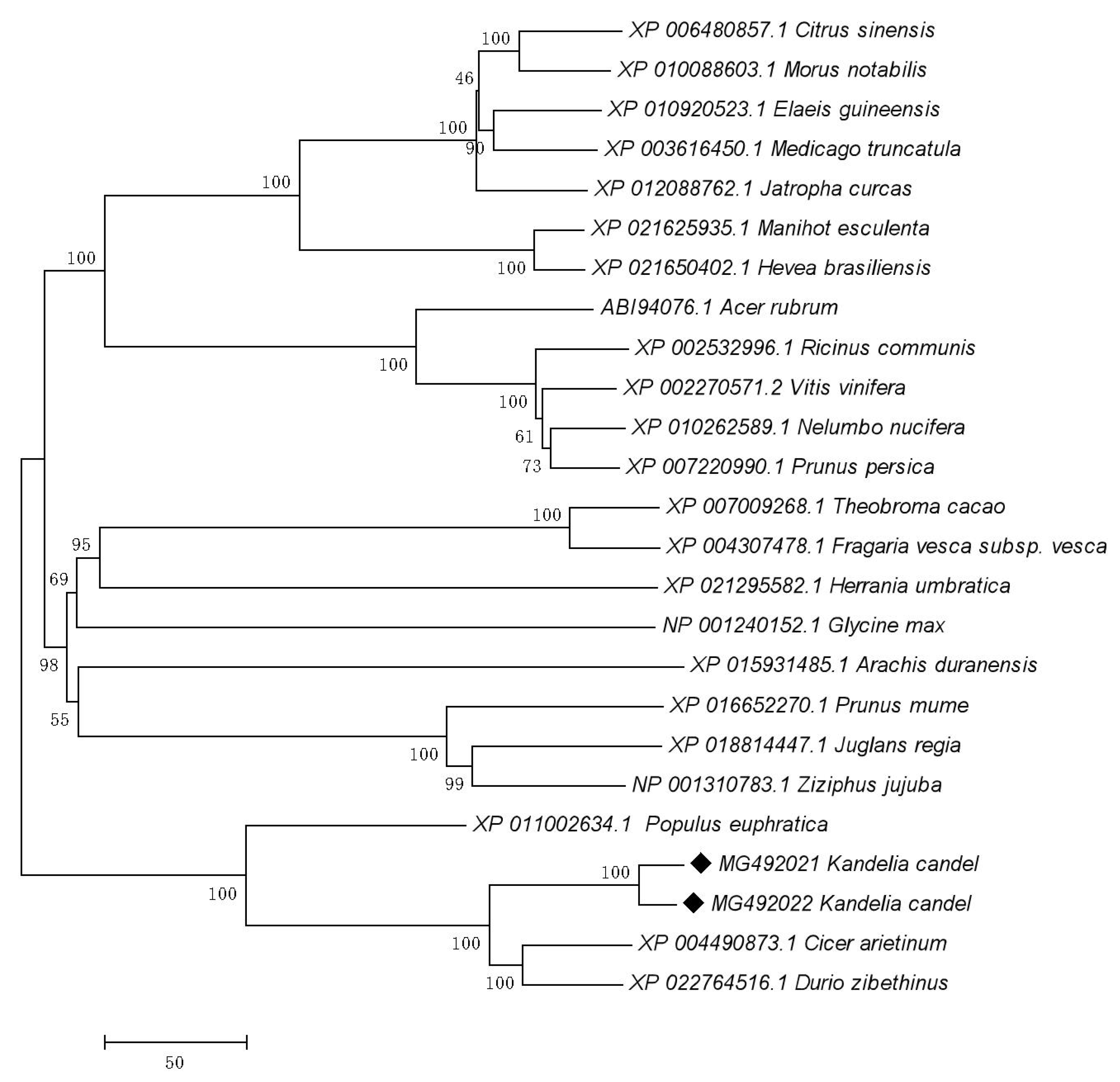
Figure 3.
Relative expression levels of KcRCA from K. candel leave under combined flooding and salinity stress. (A) and (B) H2O2 and O2•− contents in K. candel leave at various time points under combined stress condition. (C) and (D) Images of K. Candel seedling from the control and treatment groups after 48 h of combined stress condition. (E) and (F) Relative expression levels of KcRCAl and KcRCAs from K. candel leave under combined stress condition. .
Figure 3.
Relative expression levels of KcRCA from K. candel leave under combined flooding and salinity stress. (A) and (B) H2O2 and O2•− contents in K. candel leave at various time points under combined stress condition. (C) and (D) Images of K. Candel seedling from the control and treatment groups after 48 h of combined stress condition. (E) and (F) Relative expression levels of KcRCAl and KcRCAs from K. candel leave under combined stress condition. .
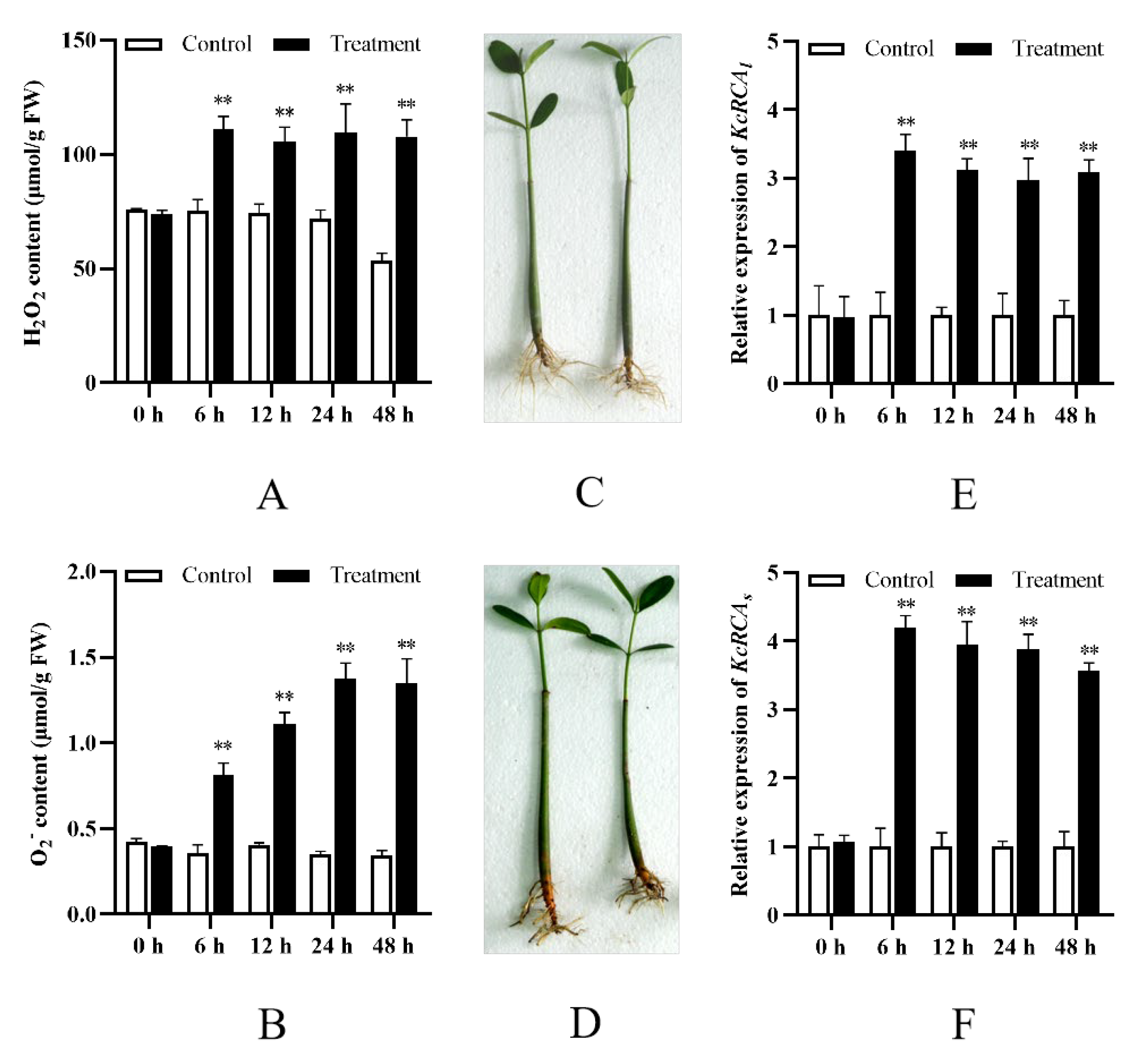
Figure 4.
Establishment of Arabidopsis transgenic plants overexpressing KcRCAl and KcRCAs in a wild-type (WT) background. (A) Wild type(WT). (B) T2 transgenic seedlings with the KcRCAl gene. (C) T2 transgenic seedlings with the KcRCAs gene. (D) Relative expression of KcRCAl gene in the transgenic plants. (E) Relative expression of KcRCAs gene in the transgenic plants. Bar = 0.5 cm.
Figure 4.
Establishment of Arabidopsis transgenic plants overexpressing KcRCAl and KcRCAs in a wild-type (WT) background. (A) Wild type(WT). (B) T2 transgenic seedlings with the KcRCAl gene. (C) T2 transgenic seedlings with the KcRCAs gene. (D) Relative expression of KcRCAl gene in the transgenic plants. (E) Relative expression of KcRCAs gene in the transgenic plants. Bar = 0.5 cm.

Figure 5.
Flooding and salinity stress tests and phenotypic analysis of transgenic plants and wild-type (WT) plants. (A) Phenotypic images of plants undergoing combined stress condition for 3 and 7 days, respectively. (B) Statistical graph depicting the survival rates of WT and transgenic plants. Bar = 1 cm.
Figure 5.
Flooding and salinity stress tests and phenotypic analysis of transgenic plants and wild-type (WT) plants. (A) Phenotypic images of plants undergoing combined stress condition for 3 and 7 days, respectively. (B) Statistical graph depicting the survival rates of WT and transgenic plants. Bar = 1 cm.
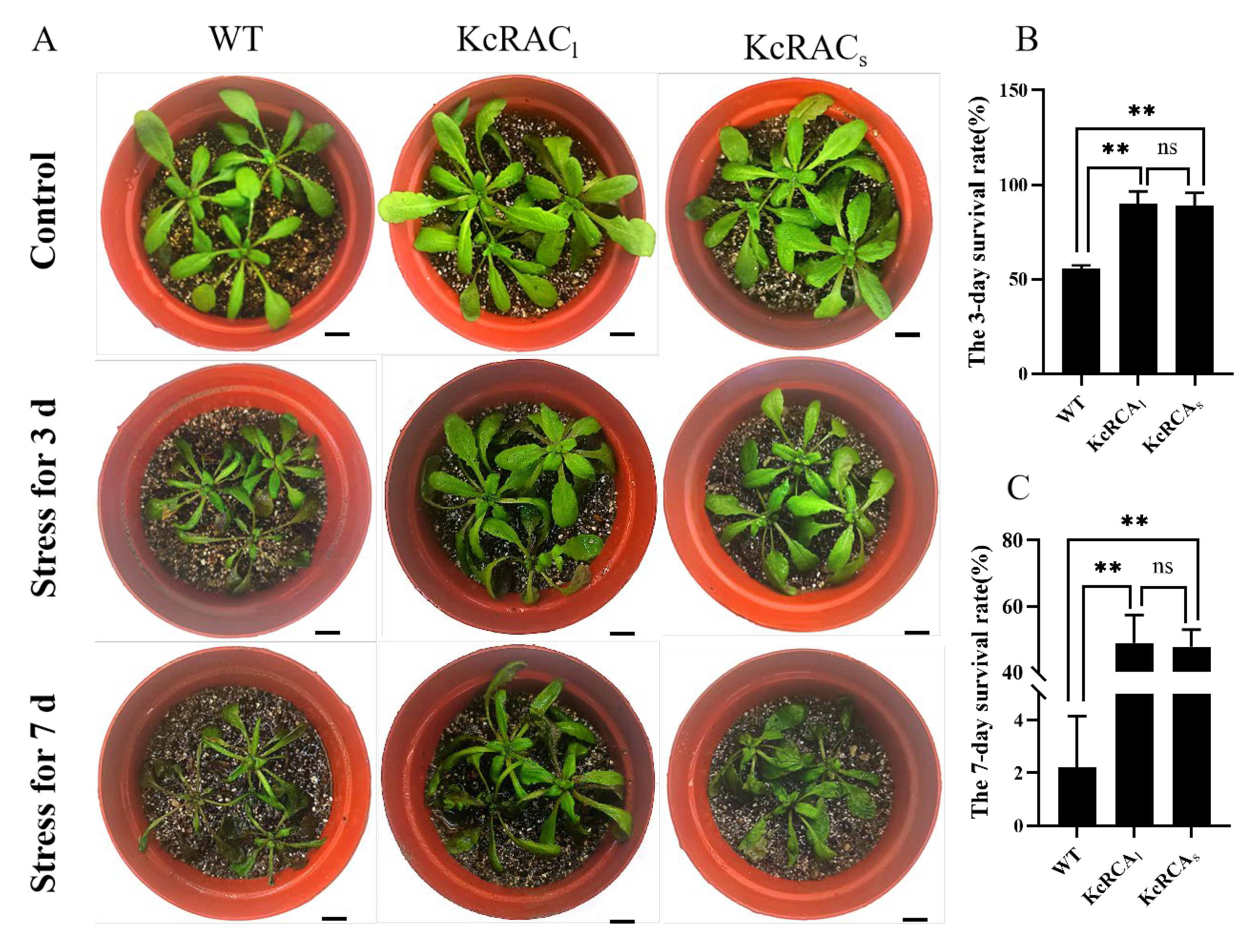
Figure 6.
Relative expression levels of the KcRCAl and KcRCAs genes in transgenic plants under combined flooding and salinity stress treatments at various time points. (A) KcRCAl Transgenetic plant. (B) KcRCAs Transgenetic plant.
Figure 6.
Relative expression levels of the KcRCAl and KcRCAs genes in transgenic plants under combined flooding and salinity stress treatments at various time points. (A) KcRCAl Transgenetic plant. (B) KcRCAs Transgenetic plant.
Figure 7.
Physiological analysis of transgenic plants under combined flooding and salinity stress for 48 h. WT: Wild-type Arabidopsis; KcRCAl: Arabidopsis transformed with the KcRCAl gene; KcRCAs: A. thaliana transformed with the KcRCAs gene. (A) Superoxide dismutase (SOD) activity. (B) Peroxidase (POD) activity. (C) Catalase (CAT) activity. (D) Superoxide radical (O2•−) content. (E) Hydrogen peroxide (H2O2) content. (F) Malondialdehyde (MDA) content. (G) Ribulose-1,5-bisphosphate carboxylase/oxygenase activase (RCA) activity. (H) Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) activity. Different lowercase letters indicate significant differences among the sample time or plant tissue (P < 0.05).
Figure 7.
Physiological analysis of transgenic plants under combined flooding and salinity stress for 48 h. WT: Wild-type Arabidopsis; KcRCAl: Arabidopsis transformed with the KcRCAl gene; KcRCAs: A. thaliana transformed with the KcRCAs gene. (A) Superoxide dismutase (SOD) activity. (B) Peroxidase (POD) activity. (C) Catalase (CAT) activity. (D) Superoxide radical (O2•−) content. (E) Hydrogen peroxide (H2O2) content. (F) Malondialdehyde (MDA) content. (G) Ribulose-1,5-bisphosphate carboxylase/oxygenase activase (RCA) activity. (H) Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) activity. Different lowercase letters indicate significant differences among the sample time or plant tissue (P < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



