
Submitted:
22 August 2025
Posted:
26 August 2025
You are already at the latest version
Abstract
Background: Mesenchymal Stromal Cells (MSCs) exert their immunoregulatory properties after licensing by inflammatory signaling cues, e.g. interferon (IFN)-γ. However, MSCs licensing by IFN-γ may result in increased expression of Human Leukocyte Antigens (HLA) class II, which is related to rapid cell elimination, impairment of their immunosuppressive properties and patient sensitization. The aim of this study was to evaluate the impact of IFN-γ on mediated immunoregulation and HLA class II expression. Methods: In this study, Wharton’s Jelly (WJ)-MSCs were isolated from the human umbilical cords. Well-defined WJ-MSCs were submitted to IFN-γ exposure, and after 96 hours (hrs), evaluation of biomolecule secretion and HLA class II expression was performed. Typing of HLA alleles using the Next Generation Sequencing (NGS) platform was performed. Results: IFN-γ primed WJ-MSCs secreted a high amount of immunoregulatory biomolecules, while elevated expression of HLA-DRB1 was observed. NGS results analysis showed the possibility of WJ-MSCs cluster formation based on their frequency of detected HLA alleles and immunoregulatory potential. Conclusion: Taking into consideration that IFN-γ primed WJ-MSCs express HLA class II alleles, it is suggested that the HLA histocompatibility between allogeneic donor and recipient should be strongly considered to acquire the most beneficial outcome of the MSCs therapeutic strategy.
Keywords:
Mesenchymal Stromal Cells
; Human Leukocyte Antigens
; Interferon-γ
; autoimmune disorders
; advanced therapy medicinal products
1. Introduction
MSCs have attracted the attention of the scientific society as a therapeutic stem-cell-based approach for the proper administration of inflammatory, autoimmune and other immune-related diseases, which has remarkably risen over the last decade [1]. MSCs represent a multipotent cellular population of mesodermal origin, equipped with great regenerative and immunoregulatory abilities [2,3]. Notably, the application of either autologous or allogeneic MSCs in phase I and II clinical trials has provided significant evidence for their safety and tolerability in humans [4,5]. Recently, MSCs have gained FDA approval for the management of steroid-refractory acute graft-versus-host disease (SR-aGVHD) in pediatric patients aged 2 months and older [6].
Nowadays, MSCs can be efficiently isolated from a variety of different sources of human body, including the bone marrow (BM), adipose tissue (AT), stromal vascular fraction (SVF), Wharton’s Jelly (WJ) tissue of the umbilical cord (UC), amniotic fluid (AF), placenta (P), dental teeth (DT), etc [7,8]. Among them, BM represented the primary source for MSCs harvesting, since Alexander Fridenstein confirmed their presence and suggested their contribution in BM stroma formation and function[9]. Based on their origin, MSCs can be distinguished into embryonic stem cells, e.g. AF-MSCs, fetal stem cells, e.g. WJ-MSCs and adult stem cells, e.g. BM-MSCs, AT-MSCs, and SVF-MSCs. MSCs derived from different tissues exhibit distinct biological characteristics, including primarily the proliferation potential, stemness capacity, differentiation ability and immunomodulatory action [10]. In 2006, the International Society for Cell and Gene Therapy (ISCT) proposed specific criteria for the proper characterization of MSCs, which have also lately revised [11,12]. Among them, 1) the plastic adherence ability, 2) multi-directional differentiation to mesodermal committed cell lineages (“osteocytes”, “chondrocytes” and “adipocytes”) and 3) specific CD markers expression, including the positive expression of the CD73, CD90 and CD105 (> 90%) and the negative expression of CD34, CD45, CD11b, HLA-DR (< 3%), have been published [11,12]. The licensing of MSCs by inflammatory signaling cues is considered a fundamental step in presenting their immunoregulatory ability [13]. Interestingly, IFN-γ is considered the most potent stimulator, upon binding to IFNG receptor (IFNGR), which results in activation of Janus kinase (JAK)-Signal Transducer and Activator of Transcription (STAT) intracellular pathway. This in turn leads to Interferon regulatory factor 1 (IRF1) activation and expression of immunoregulatory membrane-bound and soluble-secreted biomolecules, including Programmed Death Ligand (PDL)-1, indoleamine-2,3-dioxygenase (IDO), nitric oxide (NO), prostaglandin E2 (PGE2), galectins, anti-inflammatory cytokines, e.g. interleukin (IL)-1β, IL10, IL13 and growth factors with known immunomodulatory action such as transforming growth factor- β1 (TGF-β1), fibroblast growth factor (FGF) hepatocyte growth factor (HGF), platelet derived growth factor (PDGF), and vascular endothelial growth factor (VEGF) [13]. In addition, IFN-γ is implicated in the signaling pathway of Mitogen Activated Protein 3 (MAP3) kinase, resulting in phosphorylation and nuclear translocation of p38 MAP kinase, which is further associated with gene expression of the aforementioned molecules, thus contributing to the adaptation of MSCs’ immunoregulatory role [13]. Once antigenic epitope presentation occurs by the professional antigen-presenting cells (APCs), then recognition by the cells of innate immunity, e.g. CD4+ or CD8+ T cells, is followed, resulting in the secretion of high amounts of pro-inflammatory cytokines such as IFN-γ, TNF-α and IL-1β, chemokines originating from the damaged cells and tissues, such as CCL2, CXCL10, CCL19 and pro-inflammatory cytokines, e.g. TNF-α, IL-1β, IL-3 and IL-6 [14]. In the context of immune response initiation, MSCs receive these signals, increase the expression of several surface molecules, including CXCR1, CXCR2, CXCR3, CXCR4, VCAM, ICAM, etc., for possible migration to the damaged tissue, and upon stimulation by the inflammatory cues, mainly by IFN-γ, increase the expression of HLA-class II, thus contributing to the antigen presentation process [15]. On the other hand, when the immune responses are persistent for a long time, higher levels of IFN-γ and other inflammatory biomolecules are produced. The aforementioned event could lead to the initiation of severe pathogenetic damage to host tissues, contribute to the accumulation in the inflamed area of different immune cells, including Th17 cells, thus inducing the occurrence of immune-related disorders [16,17]. To avoid this unfavored situation, and as part of the homeostatic action of MSCs, these stem cells can acquire an immunoregulatory phenotype to potentially tolerate the persistently overactivated immune responses [18]. For this purpose, currently MSCs are widely used as potent immunosuppressive agents in a great number of severe conditioned disorders, including the graft versus host disease (GvHD), occurred after hematopoietic stem cells (HSCs) or solid organ transplantation [19]. Interestingly, recipients after haploidentical HSCs transplantation are characterized by a high possibility of acute GvHD (aGvHD) occurrence (30-63%), within the first 100 days of transplantation [20]. AGvHD is also induced by treatment-related tissue damage, where the donor’s T cells recognize and attack against recipients' HLA-mismatched cells [21]. In addition, activated donor T cells migrate to other organs, causing extensive damage and production of high levels of inflammatory mediators, a process which resembles several autoimmune disorders. Similar aGvHD rates have also been reported in solid organ transplantation [22]. To reverse the adverse events of aGvHD, patients mostly receive steroids such as calcineurin inhibitors, however, a number of patients are less responsive to this therapeutic strategy, resulting in the development of steroid-resistant (SR) condition [21]. Given that MSCs exert strong immunoregulatory potential, this stem cell therapy may be proven life-saving for patients with SR-aGvHD. Interestingly, results from phase I/II clinical trials have shown the beneficial effects of third-party MSCs, which are associated with no-rejection episodes and overall long-term patient survival [23,24,25,26]. Overall, MSCs appear to play a beneficial role in suppressing overactive immune responses. However, enhanced HLA class II expression in IFN-γ primed MSCs can also be observed, which raises the following concern. Could the immunoregulatory properties of MSCs be compromised by the expression of HLA class II upon IFN-γ licensing? Elevated expression of HLA class II in IFN-γ primed MSCs can be further related to antigenicity acquisition, which could possibly elicit a host immune response against them, and rapid clearance of the infused stem cells. In addition, there is a possibility of donor-specific antibodies (DSA) production against the infused third-party MSCs, leading to patients’ sensitization to a great number of proteins [27].
Considering the clinical utility of MSCs in immune-related disorders as potent immunosuppressive agents, the aim of this study was to insightfully explore the induction HLA class II expression and the immunoregulatory properties of IFN-γ primed MSCs. The acquired data may offer additional insights into the role of MSCs as “sensors” and “switchers” implicated in both innate and adaptive immune responses. Interestingly, the acquired knowledge can be further utilized for the greater understanding of the MSCs’ immunobiology, which can be used as a basis for the development of “off-the-self” stem cell therapy, as an alternative therapeutic strategy for immune-related disorders.
2. Results
2.1. Characterization of Well-Defined Wj-Mscs
WJ-MSCs were successfully isolated from fresh hUC, characterized by spindle-shaped morphology under in vitro conditions. Moreover, the WJ-MSCs retained the fibroblastic-like morphology until P4 (Figure 1). To test if MSCs fulfilled the minimum criteria outlined by the ISCT, to be considered as well-defined stem cells, trilineage differentiation and immunophenotypic analysis were performed (Figure 1). WJ-MSCs successfully differentiated to “osteocytes”, “adipocytes” and “chondrocytes” as it was confirmed by alizarin Red S, Oil Red O and Toluidine blue stains, respectively (Figure 1). Immunophenotypic analysis showed positive expression for typical markers (CD73, CD90, CD105) and negative expression for CD11b, CD34 and CD45 (Figure 1 and Figure S2. Specifically, the expression of WJ-MSCs P4 for the classical CD markers, including the CD73, CD90 and CD105 was 96.7 ± 1.6 %, 96.3 ± 1.9 % and 96 ± 1.6%, respectively, while for CD45, CD34, HLA-DR, CD15, CD3, CD19, CD31 and CD11b was 1.1 ±0.5%, 0.6 ± 0.3%, 1.1 ± 0.5%, 1.2 ± 0.6 %, 0.6 ± 0.2%, 0.7 ± 0.3%, 0.5 ± 0.2% and 0.6 ± 0.3%, respectively (Table S1). WJ-MSCs P4 also presented intermediate expression for the CD49a (63.9 ± 9.8%) and CD44 (67.4 ± 8.3%). In addition, total cell number, CDT and CPD were determined from P1 to P4. Specifically, the total numbers of MSCs for P1, P2, P3 and P4 were 2.1 x 105 ± 3.9 x 105, 1.9 x 106 ± 3.3 x 106 , 5.8 x 106 ± 1.2 x 106 and 15.2 x 106 ± 2.4 x 106, respectively. The CDTs and CPDs of MSCs were 124.5 ± 14.4 h, 34.7 ± 5.2 h, 30.5 ± 8.3 h and 3.1 ± 0.3, 4.7 ± 0.4 and 6.1 ± 0.3 determined for P2, P3 and P4, respectively (Figure 1). The above results suggested that the MSCs were considered as well-defined stem cells based on the ISCT outlined criteria, and can be used for the next series of experimental procedures.
2.2. Impact of Ifn-Γ Priming On Wj-Mscs Characteristics
Next, INF-γ was used as a stimulatory cue for the priming of WJ-MSCs. Microscopically examination of IFN-γ primed WJ-MSCs showed the presence of an increased number of intracellular vesicles compared to non-primed WJ-MSCs (Figure 2). Moreover, total cell number and viability of IFN-γ primed and non-primed WJ-MSCs were examined. Interestingly, the number of non-primed and IFN-γ primed WJ-MSCs was 15.2 ± 1.8 x 106 and 15.4 ± 1.7 x 106 cells, respectively (Figure 2). The viability of non-primed WJ-MSCs was 92.2 ± 2.4% and 93.5 ± 3.1%, respectively (Figure 2). To further assess the impact of IFN-γ on MSCs’ metabolic activity, total ATP and the ratio of ADP/ATP were determined. The total ATP of non-primed and IFN-γ primed WJ-MSCs was 69.1 ± 11.8 μM and 70.4 ± 12.6 μM, respectively, whereas, of the positive control group was 1.8 ± 0.6 μM (Figure 2). The ADP/ ATP ratio of the non-primed and IFN-γ primed WJ-MSCs was 0.1 ± 0.05 for both groups, and 0.6 ± 0.1 for the positive control group (Figure 2). Between non-primed and IFN-γ primed WJ-MSCs, no statistically significant differences were observed. The only statistically significant differences were found between the non-primed and IFN-γ primed WJ-MSCs against the positive control group, both in total ATP determination (p < 0.01) and ADP/ ATP ratio (p < 0.01). CD markers expression showed elevation in HLA-DR and CD10 in IFN-γ primed WJ-MSCs (63.9 ± 9.8 % and 91.4 ± 4.2 %, respectively), whereas no change in expression levels of CD340 and CD49a were observed between IFN-γ primed and non-primed WJ-MSCs (Figure 2 and Table S2). No alteration in CD73, CD90, CD105, CD34, CD45, CD29, CD15, CD44, CD3, CD19, CD31 and CD11b was observed between non-primed and IFN-γ primed WJ-MSCs (Table S2). Statistically significant difference was observed between non-primed and IFN-γ primed WJ-MSCs regarding the expression of HLA-DR (p < 0.001) and CD10 (p < 0.001). Moreover, higher expression of p38 MAP kinase was observed in IFN-γ primed WJ-MSCs compared to non-primed WJ-MSCs, as indicated by the results of indirect immunofluorescence (Figure 2). In regards to MLR, in both direct and indirect approaches, IFN-γ primed and non-primed WJ-MSCs were able to decrease the proliferation of CB-T cells (Figure 2). Of particular interest, under direct contact conditions, non-primed WJ-MSCs resulted in a 19% reduction in T cell number, whereas IFN-γ primed WJ-MSCs achieved a 61% reduction (Figure 2 and Table S3). Under indirect contact, the reduction in T cell number was 15% and 50% for non-primed and IFN-γ primed WJ-MSCs (Figure 2 and Table S4). Statistically significant differences in MLR were observed between IFN-γ primed and non-primed WJ-MSCs in direct (p < 0.001) and indirect (p < 0.001) approaches. Gene expression analysis showed the positive expression of REX1, OCT4, NANOG, SOX9, KLF4 and GAPDH in non-primed WJ-MSCs (Figure 2 and Figure S3). On the other hand, IFN-γ primed WJ-MSCs characterized by lower expression of REX1, while no significant alteration in gene expression of OCT4, NANOG, SOX& and KLF4 was observed (Figure 2 and Figure S3). The above results were further confirmed by real-time PCR, where statistically significant difference in REX1 expression between IFN-γ primed and non-primed WJ-MSCs (p< 0.001) was indicated.
2.3. Biomolecules Quantification
IFN-γ primed WJ-MSCs characterized by an increase in secreted biomolecules compared to non-primed WJ-MSCs (Figure 3). Specifically, the content of IL-1Ra, IL-6, IL-10, and IL-13 of IFN-γ primed WJ-MSCs was 811 ± 193 pg/ml, 86 ± 19 pg/ml, 238 ± 64 pg/ml and 150 ± 39 pg/ml, respectively. The content of the same cytokines for the non-primed WJ-MSCs was 86 ± 25 pg/ml, 32 ± 9 pg/ml, 43 ± 18 pg/ml and 41 ± 15 pg/ml, respectively (Figure 3A,B). Statistically significant differences were observed between the IFN-γ primed and non-primed WJ-MSCs regarding the aforementioned determined cytokines (p < 0.001). In a similar manner, the levels of TGF-β1, FGF, PDGF, VEGFA, HGF, NO and IDO of the IFN-γ primed WJ-MSCs was 873 ± 132 pg/ml, 887 ± 142 pg/ml, 949 ± 156 pg/ml, 1026 ± 182 pg/ml, 880 ± 184 pg/ml, 63 ± 16 μM and 1095 ± 258 pg/ml, respectively, while for the non-primed WJ-MSCs was 509 ± 111 pg/ml, 448 ± 73 pg/ml, 390 ± 105 pg/ml, 491 ± 119 pg/ml, 405 ± 158 pg/ml, 12 ± 5 μM and 328 ± 136 pg/ml (Figure 3A,B). Statistically significant differences were observed between IFN-γ primed and non-primed WJ-MSCs regarding the above-mentioned biomolecules (p < 0.001). In addition, uMAP analysis showed that 15 out of 20 MSCs samples (75%), characterized by increased immunoregulatory biomolecule secretion, however a 25% of the MSCs samples characterized by lower immunoregulatory biomolecule secretion, which was comparable to non-primed WJ-MSCs. Functional protein association network showed that IL-6 and TGF-β1 were the main orchestrators for anti-inflammatory responses, and actively interacted and possibly regulated the production of the immunoregulatory biomolecules secreted by the IFN-γ primed-WJ-MSCs.
2.4. Analysis of Hla Class I and Ii Alleles In Wj-Mscs
High-resolution HLA typing using the NGS approach was performed to determine the most frequent HLA class I and II alleles, presented by the WJ-MSCs. Regarding the HLA-A, the most frequent HLA alleles were the A*02:01:01 (23%), A*24:02:01 (18%), A*03:01:01 (15%), A*01:01:01 (13%), thus representing 69% of the total HLA-A alleles (Figure 4A). For the HLA-B, the most frequent HLA alleles were the B*51:01:01 (18%), *35:01:01 (10%), *35:03:01 (10%), *18:01:01 (8%), *44:02:01 (8%) and for the HLA-C, were the C*04:01:01 (28%), *12:03:01 (20%), *06:02:01 (8%) and *15:02:01 (8%), representing the 54% and 64% of the total HLA-B and C alleles, respectively (Figure 4A). In regards to HLA class II, the most frequent alleles for the HLA-DRB1, were DRB1*11:04:01 (15%), *11:01:01 (13%), *14:54:01 (10%), *13:01:01 (8%), *01:01:01 (8%) for the HLA-DQB1, were the DQB1*03:01:01 (40%), *05:03:01 (15%), *05:01:01 (13%), *05:02:01 (8%) and for the HLA-DPB1, were the DPB1*04:01:01 (50%), *04:02:01 (15%), *02:01:02 (10%) and *03:01:01 (8%), representing the 54%, 76% and 83% for the HLA-DRB1, -DQB1 and -DPB1 alleles, respectively (Figure 4A). The neighbor-joining trees for the HLA class I and II depicted the genetic distance between the HLA alleles. In addition, similar antigenic epitopes represented by different HLA alleles, found within the same phylogenetic branch (Figure 4B,C and Figure S4). Based on the HLA class I and II allele frequencies and biomolecules secretory profile, clustering of the different MSCs samples was performed as shown in figure 4 (4D). Indeed, 3 major MSCs clusters were formed, with the first cluster including primarily MSCs batches with the greatest immunoregulatory profile in terms of secretion and most frequent HLA class I and II alleles (Table S5). Moreover, nucleotide comparison of core exons in HLA class I and II molecules depicted significant alterations between the different HLA alleles (Figure 5). Specifically, for HLA class I, including HLA-A, B, and C alleles, multiple nucleotide alterations were observed in exon 2 position (pos) 74G-342C and exon 3 pos 343G-619G (Figures S5, S6 and S7). Regarding the HLA class II, also nucleotide alterations were observed in HLA-DRB1 pos 100C-369A, in HLA-DQB1 pos 109G-378A and HLA-DPB1 pos 100G-383A (Figures S8, S9 and S10). These nucleotide alterations result in different 3D structure formation of the different HLA alleles. However, when mismatches between donor and recipient exist, these nucleotide mismatches are responsible for the initiation of specific immune responses towards the foreign HLA alleles, which is further associated with the production of DSA.
3. Discussion
The clinical utility of MSCs has been demonstrated for over 20 years to treat severe steroid-resistant GvHD in patients undergone transplantation [28]. Since then, over 900 clinical trials (www.clinicaltrials.gov) have been conducted, involving both autologous and allogeneic “off-the-shelf” MSC applications. A great number of those clinical trials are focused on the proper administration of GvHD [29], autoimmune disorders [30], osteoarticular damaged tissue [31], while recently WJ-MSCs were applied in phase I/II trials for halting the cytokine release syndrome (CRS), mediated by SARS-CoV-2 pathogenic mechanism [32]. The Food and Drug Administration (FDA) approved in 2024, the MSCs application in pediatric patients suffering from steroid-resistant GvHD [33]. Several preclinical and clinical studies have prompted the idea to re-examine the immune-privileged status of MSCs and reconsider the concept of using them universally [34,35]. Whilst most studies have clearly shown the beneficial effect of MSCs in terms of immunosuppression, there is evidence in the literature indicating that HLA-mismatched MSCs can elicit host immune responses [34,35]. Previous studies have demonstrated that MSCs can modify their characteristics, notably exhibit increased expression of HLA class II molecules, when activated by inflammatory stimuli, in particular by interferon-gamma (IFN-γ) [13,36,37]. IFN-γ is considered a key mediator for the secretion of MSCs’ immunoregulatory biomolecules [38,39]. Accordingly, in this study, we investigated the impact of IFN-γ licensing on MSCs’ immunobiology, by exploring the acquired immunoregulatory phenotype and the altered HLA class II expression. The WJ-MSCs batches used in this study, characterized as well-defined stem cells, based on the criteria that have been outlined by the ISCT. In brief, WJ-MSCs presented fibroblastic spindle-shaped morphology, followed by successful trilineage differentiation (“osteocytes”, “adipocytes” and “chondrocytes”) and by typical CD markers expression (CD73, CD90 and CD105 > 95%, and CD15 CD34, CD45 < 3%). In addition to the markers above, WJ-MSCs showed moderate to high expression of CD29, CD44, and CD49α. Followed IFN-γ exposure, primed WJ-MSCs characterized by an increased number of intracellular vesicles when compared to non-primed WJ-MSCs, however, no other morphological alterations, in terms of size or shape, were observed. Similar observations in IFN-γ primed WJ-MSCs have been observed by other groups in the past, where primary senescent-like characteristics, including the accumulation of reactive oxygen species (ROS) after IFN-γ licensing, were observed [40]. However, in our study, no cytotoxicity of WJ-MSCs was evident after 96 hrs exposure to IFN-γ, as confirmed by the determination of cell number, viability, ATP and ADP/ATP ratio. No statistically significant differences regarding the characteristics mentioned above were found between IFN-γ-primed and non-primed WJ-MSCs. This suggested that the cytotoxicity reported by Yang ZX et. al. [42] may be attributed to variations in MSC origin (e.g. bone marrow-derived MSCs) and also to different applied experimental conditions compared to this study [41]. IFN-γ primed WJ-MSCs characterized by statistically significant increase in HLA-DR and CD10 expression compared to non-primed WJ-MSCs, while CD340 and CD49a didn’t present any significant alteration. CD340 can be considered as an advanced marker for MSCs characterization, mostly related to proliferation potential of MSCs, while CD49a represents the α1 integrin subunit, related to migration and homing abilities of MSCs [42]. No alteration in both characteristics was observed in MSCs after IFN-γ licensing. Regarding the increased expression of HLA-DR, it has already been shown that IFN-γ mediated phosphorylation of STAT1, results in strong activation of Class II Major Histocompatibility Complex Transactivator (CIITA) and eventually the transcription of all HLA class II genes (e.g. DR, DQ, DP) [43]. This is the primary reason why also in our study, production of membrane-bound HLA class II molecules was observed, upon IFN-γ licensing of MSCs. The increased expression of membrane-bound HLA-DR seems to be in accordance with the study of Le Blanc et. al. [36], van Megen et. al. [37] and Kuci et. al. [44]. Moreover, it has been shown that besides the elevated HLA class II expression in IFN-γ primed MSCs, no presence of co-stimulatory molecules including CD20L, CD80 (B7-1) and CD86 was evident in MSCs [45,46,47]. In regards to CD10 elevated expression, Kouroupis et al. [48], presented evidence for the potential link with the enhanced immunosuppressive properties exerted by the IFN-γ licensed MSCs. Taking into account the above, IFN-γ exposure may lead to increased expression of CD10, thus further contributing to the immunoregulatory phenotype shifting of WJ-MSCs. However, to properly clarify the association between IFN-γ and increased expression of CD10 by primed MSCs, further experimental procedures are required.
Knowing that, IFN-γ favors the immunoregulatory phenotype of MSCs through the activation of various intracellular signaling pathways, the next step of this study was the evaluation of altered p38 MAP kinase expression in primed MSCs. Indeed, IFN-γ primed WJ-MSCs characterized by high intranuclear localization of p38 MAP kinase, an event which is further associated with anti-inflammatory cytokine genes transcription. It is well known that upon exposure of MSCs to IFN-γ, activation of MAP kinase, including primarily the p38 MAP kinase, is performed, a process which is mediated by the activated JAK1/2 and STATs phosphorylation [49,50,51]. As part of a greater understanding of primed MSCs’ immunobiology, gene-level expression of REX1, OCT4, NANOG and SOX9 was evaluated. REX1 belongs to YY1 sub-family of transcription factors, which can act as a transcription regulator for a great number of genes, influencing primarily the stemness, cell growth and protein secretion [52,53]. In stem cells, REX1 is considered alongside OCT4 and NANOG a key-specific marker for stemness preservation. One of the primary functions of REX1 is the inhibition of the JAK/STAT pathway, thus acting in a competitive manner with the p38 MAP kinase [52,53]. In this study, non-primed WJ-MSCs were characterized by higher expression of REX1 and low detection of p38 MAP kinase (mainly cytoplasmic), while in IFN-γ primed WJ-MSCs, intranuclear translocation of p38 MAP kinase and significant decrease of REX1 expression were observed. No difference in gene expression levels of OCT4, KLF4, NANOG and SOX9, reflecting the preservation of stemness potential of MSCs after the IFN-γ stimulation, was observed. To further explore the immunoregulatory action of primed WJ-MSCs, MLR experiments were conducted. These experiments proved that primed WJ-MSCs, effectively induced the decrease of T cell number, thus confirming that IFN-γ plays a crucial role in immunoregulatory phenotype shifting upon exposure to IFN-γ, possibly through proper balance between REX1 expression and activation p38 MAP kinase signaling pathway.
Considering that secreted immunoregulatory biomolecules have gained the interest of the scientific society, in terms of using them as “off-the-self” therapy, quantification of those proteins was performed in this study. IFN-γ primed WJ-MSCs secreted a great amount of immunoregulatory biomolecules, including cytokines such as IL-1RA, IL-6, IL-10, and IL-13, growth factors such as TGF-β1, FGF, PDGF, HGF and VEGF and other biomolecules including IDO and NO. Among them, the HGF had the greatest expression, followed by VEGFA, PDGF and IDO. The protein interactome revealed that TGF-β, IL-6, and IL-13 are key proteins actively engaging with other biomolecules. Indeed, it has been shown that IL-6 exert a pleotropic action, regulating possibly the balance between pro and anti-inflammatory cytokine production [54]. Moreover, supporting evidence is provided by the fact that IL-6-deficient mice failed to produce proper anti-inflammatory responses, an event which contributes to autoimmune disorders occurrence [54,55,56]. IL-6, produced by IFN-γ primed WJ-MSCs, can act also in autocrine manner, influencing the production of other anti-inflammatory cytokines and favoring the Th2 shifting. In the same line, IL-13 favors beyond the Th2 shifting, the M2 phenotype macrophage polarization. In addition, IL-13 can reduce the production levels of TNF-α, IL-1β and IL-3, through blockage of nuclear factor-kappa B (NF-kB) and c-Jun terminal kinase (JNKs), contributing in this way to the immunomodulation of the overactivated immune responses [57]. Recently, it has been shown that TGF-β1, independently of suppressor of mothers against decapentaplegic (SMADs) signaling pathway, is associated with the activation of p38 MAP kinase [58]. Indeed, activation of TGF-β1 type 1 receptor promotes the phosphorylation of SMAD2 and SMAD3, resulting in a plethora of biological events, e.g. cell growth arrest and apoptosis, as part of its canonical signaling pathway [58,59]. However, it has been shown that TGF-β1, can mediate the activation of other significant signaling proteins such as several MAP kinases, including extracellular signal-regulated kinases (ERKs), JNKs and p38 MAP kinase, by engaging a currently unknown SMAD-independent signaling pathway [58]. Knowing that TGF-β1, which is secreted by the IFN-γ primed WJ-MSCs, has an autocrine effect, this growth factor can influence diverse biological consequences, contributing to elevated levels of p38 MAP kinase, which finally is associated with the adaptation of immunoregulatory phenotype by the WJ-MSCs. It is noteworthy to be mentioned that a 25% of IFN-γ primed WJ-MSCs characterized by low secretion of immunoregulatory biomolecules. To further explain this phenomenon, IFNGR1 sequencing could be a future target of evaluation, to shed light on whether specific structural mutations impair the binding affinity of IFN-γ, resulting in poor MSCs’ response. However, this is beyond the scope of this study and could be a future direction to better understand the MSCs' immunobiology. Based on the current literature and from the results provided by this study, non-primed WJ-MSCs did not express membrane-bound HLA class II molecules [60]. On the other hand, IFN-γ mainly in combination with other inflammatory stimuli, can efficiently cause the priming of WJ-MSCs and the expression of membrane-bound HLA class II molecules [36]. Recent evidence has shown that MSCs are broadly applied as third-party allogeneic Advanced Therapeutic Medicinal Products (ATMPs) to a great number of immune-related diseases, including autoimmune disorders, GvHD and COVID-19 [1,29,30,32]. In the majority of cases, no HLA matching between donor and recipient is required, which may raise the question if allogeneic MSCs are capable of inducing DSAs production, resulting in rejection of cells, accompanied by the subsequent sensitization of the recipient. Trying to answer this question, high-resolution HLA typing showed that the WJ-MSCs used in this study, characterized by the most common HLA alleles in terms of frequency in the Greek population [61]. In addition, neighbor joining tree for HLA class I and II alleles showed the existence of several branches, with HLA belonging to the same branch to recognize foreign antigens in a similar manner. By combining the secretory profile of MSCs and their HLA class I and II molecules, clustering of the WJ-MSCs was achieved. Cluster analysis indicated that MSCs can be classified according to their secretory profiles and HLA allele frequencies into groups reflecting high, moderate, and low responses. As a future perspective, this could lead to the establishment of a cell biobank with well-defined MSC lines, to be readily accessible upon demand. Also, this may further explore the possibility of using precisely selected for a specific recipient MSCs (based on donor-recipient histocompatibility), which eventually elucidate the highest beneficial outcome for the patient. A notable example is the observation of specific HLA associations with disease susceptibility in various immune-related disorders, including multiple sclerosis (MS) [62,63,64,65]. For instance, high risk HLA alleles, including HLA-DRB1*15:01, HLA-DQB1*06:02 and HLA-DQB1*01:02 have been found in Caucasians patients suffering by MS [66]. In this way, myelin basic protein (MBP) and myelin oligondendrocyte glycoprotein (MOG) epitopes are strongly expressed by the aforementioned HLA alleles, triggering the CD4+ T cells and CD19+ B cells responses, and contributing to disease pathogenesis [67,68,69]. For this purpose, it is possible that the administration of precise selected third-party MSCs, bearing HLA alleles such as HLA-DRB1*01:01, HLA-DRB1*11:01, HLA-DRB1*07:01 and HLA-DQB1*06:02 with known protective action against MS, may exert better immunoregulatory action against the innate immunity cells in those patients [70,71].
Based on a meta-analytic study conducted by Bezstaroti et. al. [72] showed the possibility of DSA production against non-HLA-matched infused MSCs in kidney transplanted patients. It was indicated that to avoid repeated HLA mismatches high resolution HLA typing at 3 fields (all exons sequencing), should be performed. To address this, HLA typing with NGS was also performed in our study, revealing specific nucleotide alterations in exon 2 and 3 for HLA class I and exon 2 for HLA class II alleles. Interestingly, exon 2 and 3 in HLA class I and exon 2 in HLA class II contribute to the formation of the antigen binding groove and to 3D formation of the extracellular molecule. In this way, alterations in these positions can be further related to the induction of DSA production, when HLA mismatches exist between donor and recipient, resulting in the rapid clearance of the infused MSCs. Moreover, this event can also be associated with impaired immunoregulatory action by MSCs, due to their elimination by host defense. Besides the above, DSA production is actively related to patients’ sensitization, which represents a negative event especially for severely conditioned patients such as kidney transplanted or those who belong to organ waiting lists. Sensitization of the recipient represents a serious condition which hampers the possibility of finding a histocompatible donor, while also is responsible for aGvHD after transplantation. Moreover, recently it has been shown that nucleotide alterations in HLA between donor and recipient, beyond exon 3 for HLA class I and exon 2 of HLA class II, known as hidden patterns, may be subject to host immune recognition and production of DSAs [73]. Therefore, in the case of severely-conditioned patients undergone an immune-related disorder, where MSCs could be applied as a therapeutic strategy, the HLA histocompatibility of MSCs using the NGS approach should be performed and importantly taken into account by the physician for proper MSC line selection.
4. Materials and Methods
4.1. Ethical Statement
The human umbilical cords used for the isolation of MSCs were accompanied by informed consent regarding their research use, signed by the mothers a few days before the delivery. The informed consent was in accordance with the Declaration of Helsinki and conformed to the ethical standards of the Greek National Ethical Committee. The overall study has been approved by the Institution’s ethical board (Reference No. 1754, January 21, 2021).
4.2. Isolation and Expansion of Wj-Mscs
MSCs were isolated from the WJ tissue of the human umbilical cords (hUCs), which have been delivered to Hellenic Cord Blood Bank (HCBB) within 24 hrs after gestation. Totally, 50 full-term (with gestational weeks 38-40) hUCs derived either from normal or caesarian deliveries were used for the experiments performed in this study. After hUCs reception by the personnel of the HCBB, they were immediately processed or remained for a maximum of 48 hrs at 4o C. Before the MSCs isolation, the hUCs were extensively washed with Phosphate Buffer Saline 1x (PBS 1x, Sigma-Aldrich, Darmstadt, Germany), to remove any blood clots from the entire cord. Then, using sterile surgical tools, the umbilical arteries and vein were isolated and removed, leaving only the Wharton’s Jelly (WJ) tissue. The WJ tissue was dissected into small pieces (0.3 cm x 0.3 cm) and placed in a 6-well culture plate (Costar, Corning, Life, Canton, MA, United States) for 18 days (Figure S1). Complete culture medium consisted of a-Minimum Essentials Medium (MEM, Sigma-Aldrich, Darmstadt, Germany), supplemented with 20% Fetal Bovine Serum (FBS, Sigma-Aldrich) and 1% Penicillin-Streptomycin (Sigma-Aldrich, Darmstadt, Germany) and 1% L-glutamine (Sigma-Aldrich, Darmstadt, Germany) was added (1 ml/ well plate). The tissue cultures were incubated for a total period of 18 days at 37oC and 5% CO2. After 18 days of culturing, the plates were examined under a light microscope and upon confluency of at least 3 of 6 wells, passage was performed. To perform the MSCs passage, the culture medium was removed, followed by PBS 1x addition, which remained for 1-3 minutes (mins) maximum. The PBS 1x was completely removed, and 500 μl of trypsin solution (Trypsin-EDTA solution 0.05% w/v, Gibco, Thermo Fischer Scientific, Waltham, United States) / well, was added. The culture plates were placed in the incubator for 10 min at 37o °C. Detached cells were transferd in 75 cm2 culture flask. The whole procedure was repeated until passage 4 (P4).
4.3. Growth Kinetics of Wj-Mscs
To further validate the WJ-MSCs characteristics (from P1 to P4), total cell number, cell doubling time (CDT) and cumulative population doubling (CPD) were determined. Initially, a number of WJ-MSCs ranged from 1.5 – 2 x 105 was harvested from the 6 well-plates and transferred to 75 cm2 cell culture flasks (Costar, Corning Life, Canton, MA, United States), followed by passages at dedicated time points. Between each passage, MSCs number counting was performed using the Countess 3 Automated Cell Counter (ThermoFischer Scientific, Waltham, United States). For counting performance, also Trypan blue in dilution 1:1 was used (by applying 10 μl of MSCs yield to 10 μl of Trypan blue solution 1%), or if it was necessary, dilution of the harvested cells 1:5 or 1:10 was performed. The time required for P1 was 432 hrs, while for P2 to P4, the time required between each passage was 240 hrs.
The estimation of CDT was performed using the following equation:
CDT = log10(N/N0) ÷ log10(2) × T
The estimation of PD was determined using the following equation:
PD = log10(N/N0) ÷ log10(2)
The cumulative PD was obtained after the addition of the passage’s current PD to the calculated PD from the previous passage.
In the above equations, N was the number of cells at each passage, No was the number of initially plated WJ-MSCs, and T was the culture duration in hrs.
4.4. Immunophenotypic Analysis of Wj-Mscs
Immunophenotypic analysis of WJ-MSCs (n=30) at P4 was performed using a panel of 11 monoclonal antibodies (mAbs). Specifically, fluorescein (FITC) labelled antibodies against CD90, CD45, CD29, CD31, and HLA-ABC, phycoerythrin (PE) labelled mAbs against CD44, CD3, CD11b, and CD34, peridinin-chlorophyll-protein (PerCP) labelled antibodies against CD19, CD105 and HLA-DR, and allophycocyanin (APC) labelled antibodies against CD15 and CD73, were applied. The immunophenotypic analysis of WJ-MSCs was performed in FACSCanto II (BD, Biosciences, Franklin Lakes, NJ, United States), and all Abs used in the current protocol were purchased from BD Biosciences. WJ-MSCs P4 in a total number of 1.4 x 106 cells, were harvested, transferred to 7 polystyrene tubes (2 x 105 cells/tube) and incubated with the above mAbs for 20 min in the dark place. Then, all tubes were centrifuged at 500g for 6 min, the supernatant was discarded, followed by the addition of 1 ml of PBS 1x (Gibco, ThermoFischer Scientific, Waltham, USA). Finally, the tubes were loaded in the carousel, in a dedicated order, and immunophenotyping was performed. A total of 100.000 events were acquired for each parameter tested. Complete flow cytometric analysis was performed in FlowJo v10 (BD, Biosciences, Franklin Lakes, NJ, United States).
4.5. Trilineage Differentiation of Wj-Mscs
To confirm the multipotentiality of WJ-MSCs at P4 (n=5), trilineage differentiation to “osteocytes”, “adipocytes” and “chondrocytes” was performed. For this purpose, the StemPro Osteogenesis, Adipogenesis, and Chondrogenesis kits (Thermo Fischer Scientific, Waltham, USA) were used, following the manufacturer’s instructions. The successful differentiation of WJ-MSCs was confirmed with histological stains. Alizarin Red S, Oil Red O, and Toluidine blue (Sigma-Aldrich) were applied for the evaluation of calcium deposition formed by “osteocytes”, lipid droplets formed by “adipocytes”, and glycosaminoglycans (sGAGs) production by “chondrocytes”, respectively.
4.6. Ifn-Γ Priming of Wj-Mscs
WJ-MSCs P4 (n=20) were initially seeded in a 25 cm2 tissue culture flask with 10 ml of complete culture medium. The next day, the WJ-MSCs were microscopically examined for the successful seeding and washed 3 times with PBS 1x (Sigma-Aldrich) to completely remove residual complete culture medium. Then, a stimulation medium consisted of 10 ml α-MEM (Sigma-Aldrich), 1% Penicillin-Streptomycin (Sigma-Aldrich), 1% L-glutamine (Sigma-Aldrich) and 100 ng/μl IFN-γ (Invitrogen) was added to each sample. All WJ-MSCs samples were incubated at 37oC and 5% CO2 for 96 hrs, with only a change of the stimulated medium contained 100 ng/μl ΙFN-γ. After 96 hrs, the IFN- γ primed WJ-MSCs were used for the subsequent experiments.
4.7. Evaluation of Ifn-Γ Primed Wj-Mscs Properties
IFN-γ primed WJ-MSCs P4 were further analyzed for their morphological features and immunophenotypic analysis and compared with non-primed WJ-MSCs as a reference control group. For this purpose, WJ-MSCs were also seeded in a 25 cm2 tissue culture flask with 10 ml of complete culture medium. After 96 hrs, the cells were detached using trypsin-EDTA 0.05% solution (Gibco, Thermo Fishcer Scientific, Waltham, USA) and the cells were counted using trypan-blue in Countess 3 automated cell counter, as previously described. For immunophenotypic analysis of IFN-γ primed and non-primed WJ-MSCs, besides the CD markers described above, the use of CD340-Percp-Cy5-5A and CD10-APC was applied. The preparation of IFN-γ primed and non-primed WJ-MSCs and the whole immunophenotypic analysis using the flow cytometry BD FACSCanto II, was performed as described previously.
4.8. Estimation of Cell Proliferation Using The Atp Assay
The determination of the WJ-MSCs proliferation after IFN-γ priming was performed using the commercial ATP assay (MAK190, Sigma-Aldrich, Darmstadt, Germany). To estimate the cell proliferation, 1 x 105 non-primed WJ-MSCs P4 (n=10) or IFN-γ primed WJ-MSCs P4 (n=10), were seeded in 24-well plates (Costar, Corning Life, Canton, MA, United States). The following day, the cells from both groups were lysed with 100 μl ATP assay buffer and 20 μl of the cell lysates were transferred to 96-well plates, followed by the addition of the reaction buffer. The samples were incubated for 30 min at RT and then absorbance was measured in Tristar 5 (Berthold Technologies GmbH & Co.KG, Germany) photometer at 570 nm wavelength. Determination of the ATP concentration of each sample was performed by interpolation to a standard curve. The standard curve consisted of 0 (blank), 5, 10, 20, 50, 100, 150, and 200 nmol standards.
4.9. Adp/Atp Assay
To further validate the induced cell death by IFN-γ exposure to WJ-MSCs, the ADP/ATP assay was performed, using the commercially available kit (MAK135, Sigma-Aldrich, Darmstadt, Germany). The assay was performed in non-primed WJ-MSCs (n=10) and IFN-γ primed WJ-MSCs (n=10), following the manufacturer’s instructions. Briefly, the light intensity, which is specific to intracellular ATP concentration, is produced with the following reaction:
ATP + D-Luciferin + O2 → oxyluciferin + AMP + PPi + CO2 + light
In the next step, the ADP is converted to ATP, which further reacts with D-luciferin. The second light intensity determines the total ADP and ATP concentration. The light intensity was measured using a luminometer (Lucy 1, Anthos, Luminoskan, Labsystems, Germany) and expressed as the number of relative light units (RLUs). The determination of ADP/ATP ratio is performed using the following formula:
ADP/ATP ratio = (RLU C – RLU B) ÷ RLC A
Where RLU A is the initial luminescence measurement after the addition of the ATP reagent. RLU B is the luminescence measurement after 10 min of incubation, and RLU C is the measurement of light intensity after the addition of ADP reagent. For the performance of the assay, non-primed WJ-MSCs (n = 10) and IFN-γ primed WJ-MSCs (n=10) were seeded at a density of 2 × 105 cells in 24-well plates with 1 mL of complete culture medium. Finally, cell cultures were incubated for a total of 7 days in a humidified atmosphere and 5% CO2. WJ-MSCs with 10% v/v DMSO served as a positive control group. The determination of the ADP/ATP ratio was performed after 7 days.
4.10. Quantification of Biomolecules Produced from Primed WJ-MSCs
Biomolecules, including the anti-inflammatory cytokines and growth factors produced from IFN-γ primed WJ-MSCs were quantified using commercial ELISA assay kits (OriGene Technologies, Maryland, USA). Specifically, in the presented study, quantification of IL-1 receptor antagonist (RA), IL-6, Il-10, IL-13, TGF- β1, VEGFA, FGF-1, PDGF, HGF, IDO and NO, was performed following the manufacturer’s instructions. The quantification of biomolecules was performed on culture supernatants obtained from non-primed (n=20) and IFN-γ primed (n=20) WJ-MSCs. Non-primed and IFN-γ primed WJ-MSCs were washed 3 times with PBS 1x, and cultured with a medium containing only α-ΜΕΜ supplemented with 1% Penicillin-Streptomycin (Sigma-Aldrich, Darmstadt, Germany), 1% L-glutamine (Sigma-Aldrich, Darmstadt, Germany) for 48%. The supernatants were collected, and the performance of each enzyme-linked immunosorbent assay (ELISA) was evaluated. The final concentration of each biomolecule was estimated through interpolation to a standard curve.
4.11. Indirect Immunofluorescence for p38 MAP Kinase
Indirect immunofluorescence against p38 MAP kinase was performed in non-primed (n=5) and IFN-γ primed (n=5) WJ-MSCs. For this assay, 1 x 104 were seeded on culture slides (Sigma-Aldrich, Darmstadt, Germany). 1 ml of complete culture medium was used for the non-primed WJ-MSCs, and 1 ml of complete culture medium supplemented with 100 ng/ml IGN-γ was used for the primed WJ-MSCs. The slides were observed microscopically for confluency achievement by WJ-MSCs and then indirect immunofluorescence was initiated. WJ-MSCs of both groups were fixed with 10% v/v neutral formalin buffer (Sigma-Aldrich, Darmstadt, Germany). Then, antigen retrieval and blocking of the WJ-MSCs were applied, followed by the addition of the primary monoclonal antibody against human p38 MAP kinase (1:1000, Catalog No 506123, Sigma-Aldrich, Darmstadt, Germany). The primary antibody was incubated overnight at 4oC. The next day, the slides were rinsed in distilled H2O to remove non-specific antibody stain, followed by the addition of the secondary mouse-antihuman monoclonal antibody-FITC conjugated (1:100, Sigma-Aldrich, Darmstadt, Germany). The secondary antibody was incubated for 1 hour in the dark at RT. DAPI stain was used for the observation of cell nuclei. The slides were rinsed with distilled H2O and finally were glycerol mounted and checked under a fluorescent microscope (Leica SP5 II). The images were acquired using a camera mounted in a microscope coupled with the software LAS Sute v2 (Leica, Microsystems, Wetzlar, Germany).
4.12. Mixed Lymphocyte Reaction
Mixed Lymphocyte Reaction (MLR) was performed to assess the immunosuppressive properties of IFN-γ primed and non-primed WJ-MSCs, through direct and indirect contact. Mononuclear cells were isolated from 5 ml of two different CBUs, using the ficoll-paque method (Cytiva Ficoll-Paque™ PLUS Media, Sigma-Aldrich, Darmstadt, Germany), as has been previously described[74], followed by fluorescence-activated cell sorting (FACS) for CD3 T cell purification (FACS ARIA, BD, Biosciences, Franklin Lakes, NJ, United States). For the direct contact, 5 x 103 cord blood T cells (CB-T cells), referred to as stimulators, were seeded in a 24-well plate (Costar, Corning Life, Canton, MA, United States) and treated with mitomycin 25 μg/mL (Sigma-Aldrich, Darmstadt, Germany) for 30 min at 37 °C. An equal number of allogeneic CB-T cells (obtained from a different donor), referred to as responders, was added to the culture. Then, 5 x 103 of either IFN-γ primed or non-primed WJ-MSCs were applied to further assess their effectiveness in halting lymphocyte proliferation. In each well, 1 ml of cultured medium was used. For the indirect contact, the same number of stimulator and responder cells were seeded on the insert (with a pore size 0.4 μm). IFN-γ primed WJ-MSCs or non-primed WJ-MSCs, were seeded on the bottom of the 24-well plate. The culture medium used in the above tasks consisted of a-MEM with 15% FBS, 1% P-S and 1% L-glutamine. The total number of CB-T cells was counted with the Countess 3 automated cell counter (Thermo Fischer Scientific, Waltham, United States), as earlier described in this study. After 72 hrs, the CB-T cells were counted again, and the proliferation index was determined. In the above assays, as a positive control group, 5 x 103 CB-T cells treated with 5 μg/ ml phytohemagglutinin (PHA, Sigma-Aldrich, Darmstadt, Germany), and as a negative control, 5 x 103 unstimulated CB-T cells, were applied.
4.13. Gene Expression Analysis
To evaluate the gene expression, total mRNA was initially isolated from non-primed (n=3) and IFN-γ primed WJ-MSCs (n=3) using the Tri reagent (Sigma-Aldrich, Darmstadt, Germany) following the manufacturer’s instructions. Finally, the isolated mRNA was eluted in 30 μl of RNAse-free H2O. Quantification of mRNA concentration and purity from all samples was performed using the Nanodrop Lite spectrophotometer (Thermo Fischer Scientific) at A260 / A280 ratio. Then, reverse transcription (RT) PCR was performed to obtain the cDNA. For this purpose, the engineered M-MLV reverse transcriptase basic kit (EnzyQuest, Heraklion, Crete, Greece) was used. Briefly, 1 μg of total RNA was added to a reaction buffer contained 2 μl oligo (T)18 50 μM, 1 μl dNTPs mix 10 mM and 6.5 μl nuclease-free H2O, followed by an initial incubation at 65o °C for 5 min, and subsequently chilling on ice for 1 min. Then, a mastermix solution contained 3 μl nuclease-free H2O, 4 μl from 5x RT buffer, 2 μl DTT 100 mM and 1 μl of reverse transcriptase enzyme 200 U/ μl, was added to the reaction mix, reaching a final volume of 20 μl. RT-PCR was performed at 42o °C for 15 min. The obtained cDNA was checked for its validity both in 1% agarose gel electrophoresis and in a Nanodrop Lite spectrophotometer (Thermo Fischer Scientific) at A260 / A280 ratio, with an obtained OD > 1.7 in all samples. To further perform the gene expression analysis, PCR for REX1, OCT4, KLF4, NANOG, SOX9 and GAPDH (Table 1) was performed in non-primed (n=3) and IFN-γ primed (n=3) WJ-MSCs (in triplicate for each sample). PCR was performed in gradient PCR (T Gradient Thermocycler, Biometra, Analytik Jena, Germany), involved the following steps: 1) initial denaturation at 950 °C for 15s, denaturation at 94 °C for 30s, annealing at 59-61oC °C (dependent on primers used) for 90s, and final extension at 72o °C for 30s. The total number of cycles was 35, followed by electrophoresis on 2% agarose gel. For proper quantification of gene expression, Real Time PCR was also performed, using the Gentier 48E machine (Tianlong Science and Technology, China). The real time PCR was performed using the RN014S kit (Enzyquest, Heraklion, Crete, Greece), followed the manufacturer’s instructions and involved 40 cycles with the following steps 1) heat activation at 95o C for 15 min, denaturation at 95o C for 30s, annealing at 59-61oC for 30s, and final extension 72o C for 30s. The quantification of gene expression analysis was performed using the formula ΔCt = Ctg – Ctref and for the comparison between groups, the formula ΔΔCT = ΔCtnon-primed- ΔCTIFN-γ primed was used.
4.14. HLA Typing
Initially, genomic DNA (gDNA) from MSCs (n= 20) was isolated using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany), followed the manufacturer’s instructions. The concentration and purity of MSCs’ gDNA were estimated with the nanodrop spectrophotometer (260/ 280 nm). HLA typing for the HLA-A, -B, -C and -DR, -DQ, -DP was performed using the Omixon Holotype kit (Omixon, Budapest, Hungary), according to the manufacturer’s instructions. The analysis of the HLAs was performed using the HLA Twin Software (Omixon, Budapest, Hungary). Further analysis of HLA and neighbor joining- tree development for HLA class I and II was performed using the Molecular Evolutionary Genetics Analysis (MEGA) 12 software (National Institutes of Health). Nucleotide comparison of HLA class I and II alleles was performed using the JalView version 2 [75].
4.15. Multivariate Data Analysis using RStudio
The protein secretion profile of non-primed and IFN-γ primed WJ-MSCs was performed using the pheatmap package in RStudio. Clustering of non-primed and IFN-γ primed WJ-MSCs based on their secretory profile was performed using the unsupervised machine learning algorithms (factominer, factoextra, umap). Hierarchical clustering of WJ-MSCs based on their HLA identification was performed using the factoextra and hclust packages.
4.16. Statistical Analysis
Statistical analysis was performed using GraphPad Prism v6 (GraphPad Software, San Diego, CA, USA). Statistically significant differences observed between study groups were indicated using the nonparametric Kruskal-Wallis test. Moreover, the validity of the results was further confirmed using the unpaired nonparametric Mann-Whitney U test. Statistically significant differences were considered when the p-value was less than 0.05. Indicated values were presented as mean ± standard deviation.
5. Conclusions
In conclusion, WJ-MSCs represent a valuable source of third-party ATMPs, which are currently applied in clinical trials for immune-related disorders, to potentially reverse the pathological situation. WJ-MSCs can be non-invasively isolated and easily expanded in great numbers under standard in vitro culturing conditions. Moreover, stimulated WJ-MSCs by inflammatory signaling cues seem to retain their stemness and multipotency state for at least 96 hrs. In the context of a precise medicinal approach, high-resolution HLA typing of MSCs to select the most histocompatible cells for a specific donor should be performed to avoid any possibility for patient sensitization and rapid clearance of cells. Therefore, the establishment of a MSC biobank, where well-defined MSC lines will be stored, to be readily available for immediate use, presents great interest. This in turn, may favor the broader application of MSCs in significant conditions such as in corticosteroid-resistant GvHD and other immune-related disorders, making them ideal ATMPs for clinical translation and utility.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1. Overview of the experimental procedure. Figure S2. Representative histograms of flow cytometric analysis, Figure S3. Fold change of gene expression analysis , regarding the REX1, OCT4, NANOG, SOX9 and KLF4, for non-primed and IFN-γ primed WJ-MSCs, respectively, Figure S4. Neighbor joining tree of the most frequent HLA class I (A) and class II (B) alleles. Figure S5. Nucleotide sequence of HLA-A alleles. Figure S6. Nucleotide sequence of HLA-B alleles. Figure S7. Nucleotide sequence of HLA-C alleles. Figure S8. Nucleotide sequence of HLA-DRB1 alleles. Figure S9. Nucleotide sequence of HLA-DQB1 alleles. Figure S10. Nucleotide sequence of HLA-DPB1 alleles. Table S1. Percentage of CD markers expression in WJ-MSCs P4. Table S2. Percentage of CD markers expression in non-primed and IFN-γ primed WJ-MSCs, Table S3. Detailed information regarding the MLR-direct contact. Table S4. Detailed information regarding the MLR-indirect contact, Table S5. Characteristics of IFN-γ primed WJ-MSCs, including secretory profile, immunophenotype and HLA class I and II allele determination using the NGS approach.
Author Contributions
Conceptualization, P.M.; Methodology, P.M.,T.C., E.G., V.H. and EF.-S.; Software, P.M. and T.C.; Validation, P.M., T.C. and E.M.; Formal Analysis, P.M.; Investigation, P.M. and E.M.; Resources, P.M. and E.M.; Data Curation, P.M. and T.C.; Writing – Original Draft Preparation, P.M.; Writing – Review & Editing, P.M., E.M., E.S. and C.S.-G.; Visualization, P.M.; Supervision, P.M., E.M., E.S. and C.S.-G; Project Administration, E.M., E.S. and C.S.-G.
Funding
This study was received no external funding
Institutional Review Board Statement
The overall study has been approved by the Institution’s ethical board with Reference No. 1754, January 21, 2021.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Conflicts of Interest
The authors declare no conflict of interest for this study.
References
- Jasim, S.A.; Yumashev, A.V.; Abdelbasset, W.K.; Margiana, R.; Markov, A.; Suksatan, W.; Pineda, B.; Thangavelu, L.; Ahmadi, S.H. Shining the Light on Clinical Application of Mesenchymal Stem Cell Therapy in Autoimmune Diseases. Stem Cell Research & Therapy 2022 13:1 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal Stem Cells. J Orthop Res 1991, 9, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Jiang, J.; Gu, Z.; Zhang, J.; Chen, Y.; Liu, X. Mesenchymal Stromal Cell Therapies: Immunomodulatory Properties and Clinical Progress. Stem Cell Research & Therapy 2020 11:1 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Kouchakian, M.R.; Baghban, N.; Moniri, S.F.; Baghban, M.; Bakhshalizadeh, S.; Najafzadeh, V.; Safaei, Z.; Izanlou, S.; Khoradmehr, A.; Nabipour, I.; et al. The Clinical Trials of Mesenchymal Stromal Cells Therapy. Stem Cells Int 2021, 2021, 1634782. [Google Scholar] [CrossRef]
- Galderisi, U.; Peluso, G.; Di Bernardo, G. Clinical Trials Based on Mesenchymal Stromal Cells Are Exponentially Increasing: Where Are We in Recent Years? Stem Cell Rev Rep 2021, 18, 23. [Google Scholar] [CrossRef]
- Blanc, K. Le; Dazzi, F.; English, K.; Farge, D.; Galipeau, J.; Horwitz, E.M.; Kadri, N.; Krampera, M.; Lalu, M.M.; Nolta, J.; et al. ISCT MSC Committee Statement on the US FDA Approval of Allogenic Bone-Marrow Mesenchymal Stromal Cells. Cytotherapy 2025, 27, 413–416. [Google Scholar] [CrossRef]
- Mushahary, D.; Spittler, A.; Kasper, C.; Weber, V.; Charwat, V. Isolation, Cultivation, and Characterization of Human Mesenchymal Stem Cells. Cytometry Part A 2018, 93, 19–31. [Google Scholar] [CrossRef]
- Costela-ruiz, V.J.; Melguizo-rodríguez, L.; Bellotti, C.; Illescas-montes, R.; Stanco, D.; Arciola, C.R.; Lucarelli, E. Different Sources of Mesenchymal Stem Cells for Tissue Regeneration: A Guide to Identifying the Most Favorable One in Orthopedics and Dentistry Applications. International Journal of Molecular Sciences 2022, 23, 6356. [Google Scholar] [CrossRef]
- Phinney, D.G. Alexander Friedenstein, Mesenchymal Stem Cells, Shifting Paradigms and Euphemisms. Bioengineering (Basel) 2024, 11. [Google Scholar] [CrossRef]
- Hass, R.; Kasper, C.; Böhm, S.; Jacobs, R. Different Populations and Sources of Human Mesenchymal Stem Cells (MSC): A Comparison of Adult and Neonatal Tissue-Derived MSC. Cell Commun Signal 2011, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D.G.; Sensebe, L. Mesenchymal Stem versus Stromal Cells: International Society for Cell & Gene Therapy (ISCT®) Mesenchymal Stromal Cell Committee Position Statement on Nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Polchert, D.; Sobinsky, J.; Douglas, G.W.; Kidd, M.; Moadsiri, A.; Reina, E.; Genrich, K.; Mehrotra, S.; Setty, S.; Smith, B.; et al. IFN-γ Activation of Mesenchymal Stem Cells for Treatment and Prevention of Graft versus Host Disease. Eur J Immunol 2008, 38, 1745. [Google Scholar] [CrossRef]
- Karp, J.M.; Leng Teo, G.S. Mesenchymal Stem Cell Homing: The Devil Is in the Details. Cell Stem Cell 2009, 4, 206–216. [Google Scholar] [CrossRef]
- Chan, J.L.; Tang, K.C.; Patel, A.P.; Bonilla, L.M.; Pierobon, N.; Ponzio, N.M.; Rameshwar, P. Antigen-Presenting Property of Mesenchymal Stem Cells Occurs during a Narrow Window at Low Levels of Interferon-γ. Blood 2006, 107, 4817–4824. [Google Scholar] [CrossRef]
- Waite, J.C.; Skokos, D. Th17 Response and Inflammatory Autoimmune Diseases. Int J Inflam 2011, 2012, 819467. [Google Scholar] [CrossRef]
- Langrish, C.L.; Chen, Y.; Blumenschein, W.M.; Mattson, J.; Basham, B.; Sedgwick, J.D.; McClanahan, T.; Kastelein, R.A.; Cua, D.J. IL-23 Drives a Pathogenic T Cell Population That Induces Autoimmune Inflammation. Journal of Experimental Medicine 2005, 201, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Jia, F.; Li, X.; Kong, Y.; Tian, Z.; Bi, L.; Li, L. Biophysical Cues to Improve the Immunomodulatory Capacity of Mesenchymal Stem Cells: The Progress and Mechanisms. Biomedicine & Pharmacotherapy 2023, 162, 114655. [Google Scholar] [CrossRef]
- Kelly, K.; Rasko, J.E.J. Mesenchymal Stromal Cells for the Treatment of Graft Versus Host Disease. Front Immunol 2021, 12. [Google Scholar] [CrossRef]
- Bonato, S.; Del Sordo, R.; Mancusi, A.; Terenzi, A.; Zei, T.; Iacucci Ostini, R.; Tricarico, S.; Piccinelli, S.; Griselli, M.; Marzuttini, F.; et al. Acute GvHD after HLA-Haploidentical Hematopoietic Cell Transplantation with Regulatory and Conventional T Cell Immunotherapy Does Not Adversely Affect Transplantation Outcomes. Blood 2021, 138, 2885. [Google Scholar] [CrossRef]
- Kadri, N.; Amu, S.; Iacobaeus, E.; Boberg, E.; Le Blanc, K. Current Perspectives on Mesenchymal Stromal Cell Therapy for Graft versus Host Disease. Cellular & Molecular Immunology 2023 20:6 2023, 20, 613–625. [Google Scholar] [CrossRef]
- Tenneti, P.; He, J.; Lalli, P.; Tenneti, P.; Grunwald, M.R.; Copelan, E.A.; Avalos, B.; Sanikommu, S.R.R. Incidence and Risk Factors Associated with Fatal Graft Vs Host Disease after Solid Organ Transplantation in United Network of Organ Transplant Database. Blood 2021, 138, 4067. [Google Scholar] [CrossRef]
- Pérez-Simon, J.A.; López-Villar, O.; Andreu, E.J.; Rifón, J.; Muntion, S.; Campelo, M.D.; Sánchez-Guijo, F.M.; Martinez, C.; Valcarcel, D.; Del Cañizo, C. Mesenchymal Stem Cells Expanded in Vitro with Human Serum for the Treatment of Acute and Chronic Graft-versus-Host Disease: Results of a Phase I/II Clinical Trial. Haematologica 2011, 96, 1072–1076. [Google Scholar] [CrossRef]
- Ringdén, O.; Uzunel, M.; Rasmusson, I.; Remberger, M.; Sundberg, B.; Lönnies, H.; Marschall, H.U.; Dlugosz, A.; Szakos, A.; Hassan, Z.; et al. Mesenchymal Stem Cells for Treatment of Therapy-Resistant Graft-versus-Host Disease. Transplantation 2006, 81, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Rasmusson, I.; Sundberg, B.; Götherström, C.; Hassan, M.; Uzunel, M.; Ringdén, O. Treatment of Severe Acute Graft-versus-Host Disease with Third Party Haploidentical Mesenchymal Stem Cells. Lancet 2004, 363, 1439–1441. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Frassoni, F.; Ball, L.; Locatelli, F.; Roelofs, H.; Lewis, I.; Lanino, E.; Sundberg, B.; Bernardo, M.E.; Remberger, M.; et al. Mesenchymal Stem Cells for Treatment of Steroid-Resistant, Severe, Acute Graft-versus-Host Disease: A Phase II Study. Lancet 2008, 371, 1579–1586. [Google Scholar] [CrossRef]
- Cherukuri, A.; Mehta, R.; Sharma, A.; Sood, P.; Zeevi, A.; Tevar, A.D.; Rothstein, D.M.; Hariharan, S. Post-Transplant Donor Specific Antibody Is Associated with Poor Kidney Transplant Outcomes Only When Combined with Both T-Cell-Mediated Rejection and Non-Adherence. Kidney Int 2019, 96, 202–213. [Google Scholar] [CrossRef]
- Le Blanc, K.; Rasmusson, I.; Sundberg, B.; Götherström, C.; Hassan, M.; Uzunel, M.; Ringdén, O. Treatment of Severe Acute Graft-versus-Host Disease with Third Party Haploidentical Mesenchymal Stem Cells. Lancet 2004, 363, 1439–1441. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hao, J.; Hu, Z.; Yang, Y.G.; Zhou, Q.; Sun, L.; Wu, J. Current Status of Clinical Trials Assessing Mesenchymal Stem Cell Therapy for Graft versus Host Disease: A Systematic Review. Stem Cell Res Ther 2022, 13, 1–22. [Google Scholar] [CrossRef]
- Shandil, R.K.; Dhup, S.; Narayanan, S. Evaluation of the Therapeutic Potential of Mesenchymal Stem Cells (MSCs) in Preclinical Models of Autoimmune Diseases. Stem Cells Int 2022, 2022, 6379161. [Google Scholar] [CrossRef]
- Carneiro, D. de C.; Araújo, L.T. de; Santos, G.C.; Damasceno, P.K.F.; Vieira, J.L.; Santos, R.R. dos; Barbosa, J.D.V.; Soares, M.B.P. Clinical Trials with Mesenchymal Stem Cell Therapies for Osteoarthritis: Challenges in the Regeneration of Articular Cartilage. Int J Mol Sci 2023, 24, 9939. [Google Scholar] [CrossRef]
- Grégoire, C.; Layios, N.; Lambermont, B.; Lechanteur, C.; Briquet, A.; Bettonville, V.; Baudoux, E.; Thys, M.; Dardenne, N.; Misset, B.; et al. Bone Marrow-Derived Mesenchymal Stromal Cell Therapy in Severe COVID-19: Preliminary Results of a Phase I/II Clinical Trial. Front Immunol 2022, 13, 932360. [Google Scholar] [CrossRef] [PubMed]
- Blanc, K. Le; Dazzi, F.; English, K.; Farge, D.; Galipeau, J.; Horwitz, E.M.; Kadri, N.; Krampera, M.; Lalu, M.M.; Nolta, J.; et al. ISCT MSC Committee Statement on the US FDA Approval of Allogenic Bone-Marrow Mesenchymal Stromal Cells. Cytotherapy 2025, 27, 413–416. [Google Scholar] [CrossRef]
- Hemeda, H.; Jakob, M.; Ludwig, A.K.; Giebel, B.; Lang, S.; Brandau, S. Interferon-Gamma and Tumor Necrosis Factor-Alpha Differentially Affect Cytokine Expression and Migration Properties of Mesenchymal Stem Cells. Stem Cells Dev 2010, 19, 693–706. [Google Scholar] [CrossRef]
- Ankrum, J.A.; Ong, J.F.; Karp, J.M. Mesenchymal Stem Cells: Immune Evasive, Not Immune Privileged. Nat Biotechnol 2014, 32, 252. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Tammik, C.; Rosendahl, K.; Zetterberg, E.; Ringdén, O. HLA Expression and Immunologic Propertiesof Differentiated and Undifferentiated Mesenchymal Stem Cells. Exp Hematol 2003, 31, 890–896. [Google Scholar] [CrossRef]
- Van Megen, K.M.; Van ’t Wout, E.J.T.; Motta, J.L.; Dekker, B.; Nikolic, T.; Roep, B.O. Activated Mesenchymal Stromal Cells Process and Present Antigens Regulating Adaptive Immunity. Front Immunol 2019, 10, 694. [Google Scholar] [CrossRef] [PubMed]
- Alvites, R.; Branquinho, M.; Sousa, A.C.; Lopes, B.; Sousa, P.; Maurício, A.C. Mesenchymal Stem/Stromal Cells and Their Paracrine Activity—Immunomodulation Mechanisms and How to Influence the Therapeutic Potential. Pharmaceutics 2022, 14, 381. [Google Scholar] [CrossRef]
- Zimmermann, J.A.; Mcdevitt, T.C. Pre-Conditioning Mesenchymal Stromal Cell Spheroids for Immunomodulatory Paracrine Factor Secretion. Cytotherapy 2014, 16, 331–345. [Google Scholar] [CrossRef]
- Yang, Z.X.; Mao, G.X.; Zhang, J.; Wen, X.L.; Jia, B.B.; Bao, Y.Z.; Lv, X.L.; Wang, Y.Z.; Wang, G.F. IFN-γ Induces Senescence-like Characteristics in Mouse Bone Marrow Mesenchymal Stem Cells. Adv Clin Exp Med 2017, 26, 201–206. [Google Scholar] [CrossRef]
- Yang, Z.X.; Mao, G.X.; Zhang, J.; Wen, X.L.; Jia, B.B.; Bao, Y.Z.; Lv, X.L.; Wang, Y.Z.; Wang, G.F. IFN-γ Induces Senescence-like Characteristics in Mouse Bone Marrow Mesenchymal Stem Cells. Adv Clin Exp Med 2017, 26, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Fabre, H.; Ducret, M.; Degoul, O.; Rodriguez, J.; Perrier-Groult, E.; Aubert-Foucher, E.; Pasdeloup, M.; Auxenfans, C.; McGuckin, C.; Forraz, N.; et al. Characterization of Different Sources of Human MSCs Expanded in Serum-Free Conditions with Quantification of Chondrogenic Induction in 3D. Stem Cells Int 2019, 2019, 2186728. [Google Scholar] [CrossRef]
- Tur, J.; Farreras, C.; Sánchez-Tilló, E.; Vico, T.; Guerrero-Gonzalez, P.; Fernandez-Elorduy, A.; Lloberas, J.; Celada, A. Induction of CIITA by IFN-γ in Macrophages Involves STAT1 Activation by JAK and JNK. Immunobiology 2021, 226, 152114. [Google Scholar] [CrossRef]
- Kuçi, Z.; Piede, N.; Vogelsang, K.; Pfeffermann, L.M.; Wehner, S.; Salzmann-Manrique, E.; Stais, M.; Kreyenberg, H.; Bonig, H.; Bader, P.; et al. Expression of HLA-DR by Mesenchymal Stromal Cells in the Platelet Lysate Era: An Obsolete Release Criterion for MSCs? J Transl Med 2024, 22. [Google Scholar] [CrossRef]
- Herrero, C.; Pérez-Simón, J.A. Immunomodulatory Effect of Mesenchymal Stem Cells. Brazilian Journal of Medical and Biological Research 2010, 43, 425–430. [Google Scholar] [CrossRef]
- Ryan, J.M.; Barry, F.P.; Murphy, J.M.; Mahon, B.P. Mesenchymal Stem Cells Avoid Allogeneic Rejection. J Inflamm (Lond) 2005, 2, 8. [Google Scholar] [CrossRef]
- Le Blanc, K.; Davies, L.C. Mesenchymal Stromal Cells and the Innate Immune Response. Immunol Lett 2015, 168, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Kouroupis, D.; Kaplan, L.D.; Huard, J.; Best, T.M. CD10-Bound Human Mesenchymal Stem/Stromal Cell-Derived Small Extracellular Vesicles Possess Immunomodulatory Cargo and Maintain Cartilage Homeostasis under Inflammatory Conditions. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Valledor, A.F.; Sánchez-Tilló, E.; Arpa, L.; Park, J.M.; Caelles, C.; Lloberas, J.; Celada, A. Selective Roles of MAPKs during the Macrophage Response to IFN-γ. The Journal of Immunology 2008, 180, 4523–4529. [Google Scholar] [CrossRef]
- Pasquinelli, V.; Rovetta, A.I.; Alvarez, I.B.; Jurado, J.O.; Musella, R.M.; Palmero, D.J.; Malbrán, A.; Samten, B.; Barnes, P.F.; García, V.E. Phosphorylation of Mitogen-Activated Protein Kinases Contributes to Interferon γ Production in Response to Mycobacterium Tuberculosis. J Infect Dis 2012, 207, 340. [Google Scholar] [CrossRef]
- Zhang, S.; Kaplan, M.H. The P38 Mitogen-Activated Protein Kinase Is Required for IL-12-Induced IFN-γ Expression. The Journal of Immunology 2000, 165, 1374–1380. [Google Scholar] [CrossRef]
- Bhandari, D.R.; Seo, K.W.; Roh, K.H.; Jung, J.W.; Kang, S.K.; Kang, K.S. REX-1 Expression and P38 MAPK Activation Status Can Determine Proliferation/Differentiation Fates in Human Mesenchymal Stem Cells. PLoS One 2010, 5, e10493. [Google Scholar] [CrossRef]
- Mongan, N.P.; Martin, K.M.; Gudas, L.J. The Putative Human Stem Cell Marker, Rex-1 (Zfp42): Structural Classification and Expression in Normal Human Epithelial and Carcinoma Cell Cultures. Mol Carcinog 2006, 45, 887–900. [Google Scholar] [CrossRef]
- Covarrubias, A.J.; Horng, T. IL-6 Strikes a Balance in Metabolic Inflammation. Cell Metab 2014, 19, 898–899. [Google Scholar] [CrossRef]
- Atreya, R.; Mudter, J.; Finotto, S.; Müllberg, J.; Jostock, T.; Wirtz, S.; Schütz, M.; Bartsch, B.; Holtmann, M.; Becker, C.; et al. Blockade of Interleukin 6 Trans Signaling Suppresses T-Cell Resistance against Apoptosis in Chronic Intestinal Inflammation: Evidence in Crohn Disease and Experimental Colitis in Vivo. Nat Med 2000, 6, 583–588. [Google Scholar] [CrossRef]
- Xing, Z.; Gauldie, J.; Cox, G.; Baumann, H.; Jordana, M.; Lei, X.F.; Achong, M.K. IL-6 Is an Antiinflammatory Cytokine Required for Controlling Local or Systemic Acute Inflammatory Responses. Journal of Clinical Investigation 1998, 101, 311–320. [Google Scholar] [CrossRef]
- Zhu, C.; Zhang, A.; Huang, S.; Ding, G.; Pan, X.; Chen, R. Interleukin-13 Inhibits Cytokines Synthesis by Blocking Nuclear Factor-ΚB and c-Jun N-Terminal Kinase in Human Mesangial Cells. J Biomed Res 2010, 24, 308. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Hébert, M.C.; Zhang, Y.E. TGF-β Receptor-Activated P38 MAP Kinase Mediates Smad-Independent TGF-β Responses. EMBO Journal 2002, 21, 3749–3759. [Google Scholar] [CrossRef] [PubMed]
- Giarratana, A.O.; Prendergast, C.M.; Salvatore, M.M.; Capaccione, K.M. TGF-β Signaling: Critical Nexus of Fibrogenesis and Cancer. Journal of Translational Medicine 2024 22:1 2024, 22, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Locatetti, F.; Maccario, R.; Frassoni, F. Mesenchymal Stromal Cells, from Indifferent Spectators to Principal Actors. Are We Going to Witness a Revolution in the Scenario of Allograft and Immune-Mediated Disorders? Haematologica 2007, 92, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Mallis, P.; Siorenta, A.; Stamathioudaki, E.; Vrani, V.; Paterakis, G. Frequency Distribution of HLA Class I and II Alleles in Greek Population and Their Significance in Orchestrating the National Donor Registry Program. Int J Immunogenet 2024, 51, 164–172. [Google Scholar] [CrossRef]
- Haegert, D.G.; Muntoni, F.; Murru, M.R.; Costa, G.; Francis, G.S.; Marrosu, M.G. HLA-DQA1 and -DQB1 Associations with Multiple Sclerosis in Sardinia and French Canada: Evidence for Immunogenetically Distinct Patient Groups. Neurology 1993, 43, 548–552. [Google Scholar] [CrossRef]
- Marrosu, M.G.; Murru, M.R.; Costa, G.; Murru, R.; Muntoni, F.; Cucca, F. DRB1-DQA1-DQB1 Loci and Multiple Sclerosis Predisposition in the Sardinian Population. Hum Mol Genet 1998, 7, 1235–1237. [Google Scholar] [CrossRef]
- Olerup, O.; Hillert, J. HLA Class II-associated Genetic Susceptibility in Multiple Sclerosis: A Critical Evaluation. Tissue Antigens 1991, 38, 1–15. [Google Scholar] [CrossRef]
- Hollenbach, J.A.; Oksenberg, J.R. The Immunogenetics of Multiple Sclerosis: A Comprehensive Review. J Autoimmun 2015, 64, 13. [Google Scholar] [CrossRef]
- Kouri, I.; Papakonstantinou, S.; Bempes, V.; Vasiliadis, H.S.; Kyritsis, A.P.; Pelidou, S.H. HLA Associations with Multiple Sclerosis in Greece. J Neurol Sci 2011, 308, 28–31. [Google Scholar] [CrossRef]
- Wucherpfennig, K.W.; Catz, I.; Hausmann, S.; Strominger, J.L.; Steinman, L.; Warren, K.G. Recognition of the Immunodominant Myelin Basic Protein Peptide by Autoantibodies and HLA-DR2-Restricted T Cell Clones from Multiple Sclerosis Patients. Identity of Key Contact Residues in the B-Cell and T-Cell Epitopes. J Clin Invest 1997, 100, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Pette, M.; Fujita, K.; Kitze, B.; Whitaker, J.N.; Albert, E.; Kappos, L.; Wekerle, H. Myelin Basic Protein-Specific T Lymphocyte Lines from MS Patients and Healthy Individuals. Neurology 1990, 40, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- Pacini, G.; Ieronymaki, M.; Nuti, F.; Sabatino, G.; Larregola, M.; Aharoni, R.; Papini, A.M.; Rovero, P. Epitope Mapping of Anti-Myelin Oligodendrocyte Glycoprotein (MOG) Antibodies in a Mouse Model of Multiple Sclerosis: Microwave-Assisted Synthesis of the Peptide Antigens and ELISA Screening. J Pept Sci 2016, 22, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Barcellos, L.F.; Sawcer, S.; Ramsay, P.P.; Baranzini, S.E.; Thomson, G.; Briggs, F.; Cree, B.C.A.; Begovich, A.B.; Villoslada, P.; Montalban, X.; et al. Heterogeneity at the HLA-DRB1 Locus and Risk for Multiple Sclerosis. Hum Mol Genet 2006, 15, 2813–2824. [Google Scholar] [CrossRef]
- Dyment, D.A.; Herrera, B.M.; Cader, M.Z.; Willer, C.J.; Lincoln, M.R.; Sadovnick, A.D.; Risch, N.; Ebers, G.C. Complex Interactions among MHC Haplotypes in Multiple Sclerosis: Susceptibility and Resistance. Hum Mol Genet 2005, 14, 2019–2026. [Google Scholar] [CrossRef] [PubMed]
- Bezstarosti, S.; Erpicum, P.; Maggipinto, G.; Dreyer, G.J.; Reinders, M.E.J.; Meziyerh, S.; Roelen, D.L.; De Fijter, J.W.; Kers, J.; Weekers, L.; et al. Allogeneic Mesenchymal Stromal Cell Therapy in Kidney Transplantation: Should Repeated Human Leukocyte Antigen Mismatches Be Avoided? Front Genet 2024, 15, 1436194. [Google Scholar] [CrossRef] [PubMed]
- Vittoraki, A.G.; Fylaktou, A.; Tarassi, K.; Tsinaris, Z.; Siorenta, A.; Petasis, G.C.; Gerogiannis, D.; Lehmann, C.; Carmagnat, M.; Doxiadis, I.; et al. Hidden Patterns of Anti-HLA Class I Alloreactivity Revealed Through Machine Learning. Front Immunol 2021, 12. [Google Scholar] [CrossRef]
- Mallis, P.; Boulari, D.; Michalopoulos, E.; Dinou, A.; Spyropoulou-Vlachou, M.; Stavropoulos-Giokas, C. Evaluation of HLA-G Expression in Multipotent Mesenchymal Stromal Cells Derived from Vitrified Wharton’s Jelly Tissue. Bioengineering 2018, 5, 95. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—a Multiple Sequence Alignment Editor and Analysis Workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Evaluation of WJ-MSCs characteristics based on the minimum criteria outlined by ISCT. WJ-MSCs presented fibroblastic-like morphology, which was retained from P0 (A1), to P1(A2) and P3(A3). Immunophenotypic analysis of WJ-MSCs for CD73, CD90, CD105, CD45, HLA-DR, CD29, CD10, CD340, CD44 and CD15 (B). Successful differentiation of WJ-MSCs to “osteocytes”, “adipocytes” and “chondrocytes”, as it was confirmed by Alizarin Red O, Oil-Red O and Toluidine blue stains, respectively.
Figure 1.
Evaluation of WJ-MSCs characteristics based on the minimum criteria outlined by ISCT. WJ-MSCs presented fibroblastic-like morphology, which was retained from P0 (A1), to P1(A2) and P3(A3). Immunophenotypic analysis of WJ-MSCs for CD73, CD90, CD105, CD45, HLA-DR, CD29, CD10, CD340, CD44 and CD15 (B). Successful differentiation of WJ-MSCs to “osteocytes”, “adipocytes” and “chondrocytes”, as it was confirmed by Alizarin Red O, Oil-Red O and Toluidine blue stains, respectively.
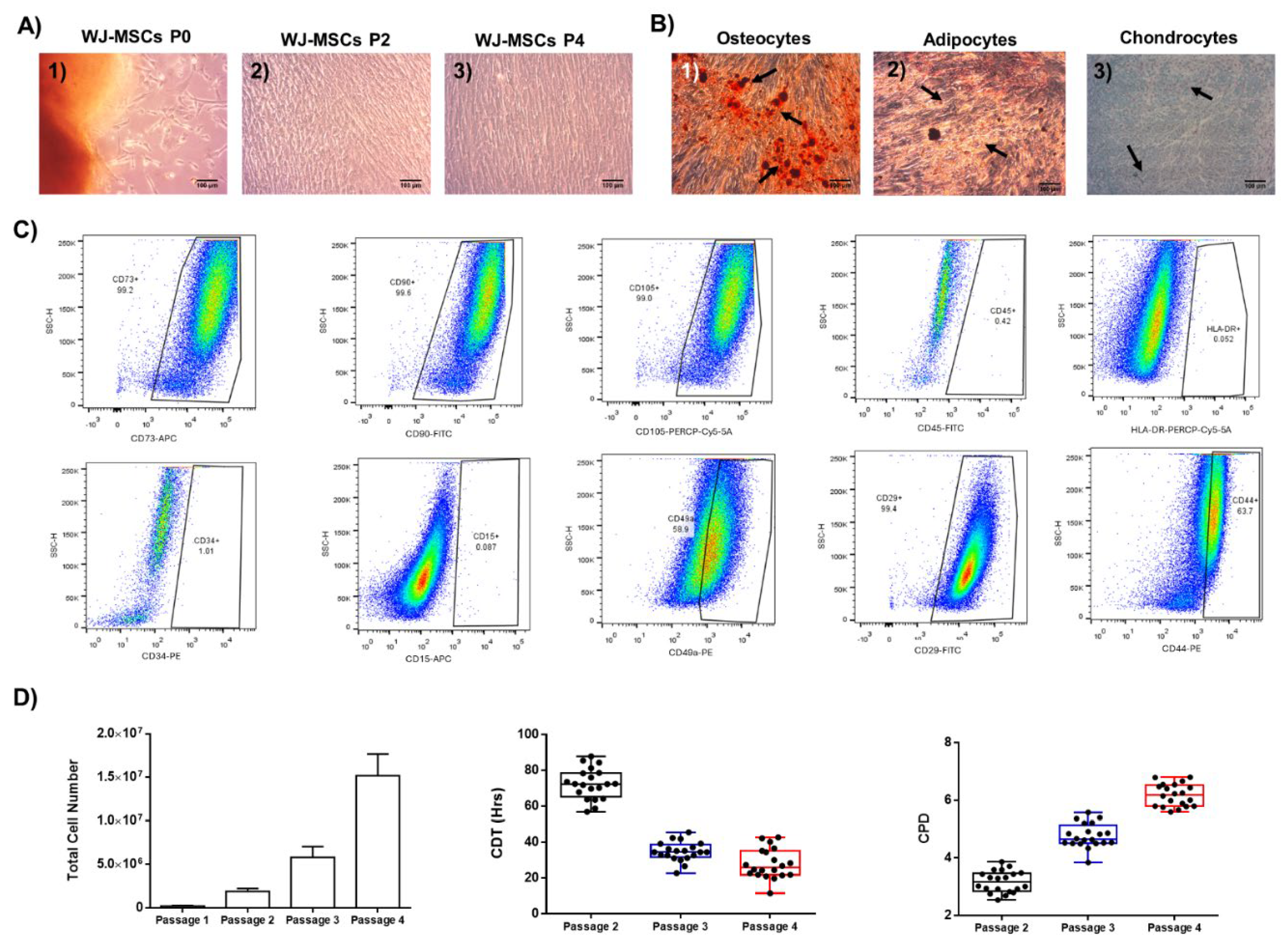
Figure 2.
Impact of IFN-γ on MSCs' phenotypic characteristics. Microscopic evaluation of non-primed WJ-MSCs (A1) and IFN-γ primed WJ-MSCs (A2). Black arrows in image A2 indicated the presence of intracellular vesicles in IFN-γ primed MSCs. Microscopic images were presented with original magnification 20x, and scale bars 50 μm. Total cell number, viability, total ATP and ADP/ATP ratio measured in non-primed and IFN-γ primed WJ-MSCs (B). No statistically significant differences were observed regarding the total cell number (p = 0.969), viability (p = 0.232), total ATP (p = 0.832) and ADP/ATP ratio (p = 0.999) between the non-primed and IFN-γ primed MSCs. The only statistically significant difference was observed regarding the total ATP (p < 0.01) and ADP/ATP ratio (p < 0.01) between the positive control group and the other study groups. Surface marker expression of HLA-DR, CD10, CD340, and CD49a of non-primed and IFN-γ primed WJ-MSCs (C). Indirect immunofluorescence against p38 MAP kinase counterstained with DAPI, in non-primed and IFN-γ primed WJ-MSCs (D). In IFN-γ primed WJ-MSCs, p38 MAP kinase stain (green) was observed in combination with DAPI stain (blue). Microscopic images were presented with original magnification of 20x and 40x, and scale bars of 50 and 10 μm, respectively. MLR was performed either in direct or indirect contact with ΙFN-γ primed and non-primed WJ-MSCs. Statistically significant reduction in CB-T cell number was observed with the co-culture of WJ-MSCs (IFN-γ primed and non-primed WJ-MSCs) in both MLR (p < 0.001). Statistically significant differences were observed between the IFN-γ primed WJ-MSCs and non-primed WJ-MSCs either in direct (p < 0.01) or in direct contact (p < 0.01) MLR. Gene expression analysis, regarding the REX1, OCT4, NANOG, SOX9, KLF4 and GAPDH in non-primed and IFN-γ primed WJ-MSCS (F). Performance of real-time PCR for the evaluation of gene expression, regarding the REX1, OCT4, NANOG, SOX9, KLF4 and GAPDH. GAPDH served as a house-keeping gene. Statistically significant difference was observed between non-primed and IFN-γ primed WJ-MSCs regarding the REX1 (p < 0.001).
Figure 2.
Impact of IFN-γ on MSCs' phenotypic characteristics. Microscopic evaluation of non-primed WJ-MSCs (A1) and IFN-γ primed WJ-MSCs (A2). Black arrows in image A2 indicated the presence of intracellular vesicles in IFN-γ primed MSCs. Microscopic images were presented with original magnification 20x, and scale bars 50 μm. Total cell number, viability, total ATP and ADP/ATP ratio measured in non-primed and IFN-γ primed WJ-MSCs (B). No statistically significant differences were observed regarding the total cell number (p = 0.969), viability (p = 0.232), total ATP (p = 0.832) and ADP/ATP ratio (p = 0.999) between the non-primed and IFN-γ primed MSCs. The only statistically significant difference was observed regarding the total ATP (p < 0.01) and ADP/ATP ratio (p < 0.01) between the positive control group and the other study groups. Surface marker expression of HLA-DR, CD10, CD340, and CD49a of non-primed and IFN-γ primed WJ-MSCs (C). Indirect immunofluorescence against p38 MAP kinase counterstained with DAPI, in non-primed and IFN-γ primed WJ-MSCs (D). In IFN-γ primed WJ-MSCs, p38 MAP kinase stain (green) was observed in combination with DAPI stain (blue). Microscopic images were presented with original magnification of 20x and 40x, and scale bars of 50 and 10 μm, respectively. MLR was performed either in direct or indirect contact with ΙFN-γ primed and non-primed WJ-MSCs. Statistically significant reduction in CB-T cell number was observed with the co-culture of WJ-MSCs (IFN-γ primed and non-primed WJ-MSCs) in both MLR (p < 0.001). Statistically significant differences were observed between the IFN-γ primed WJ-MSCs and non-primed WJ-MSCs either in direct (p < 0.01) or in direct contact (p < 0.01) MLR. Gene expression analysis, regarding the REX1, OCT4, NANOG, SOX9, KLF4 and GAPDH in non-primed and IFN-γ primed WJ-MSCS (F). Performance of real-time PCR for the evaluation of gene expression, regarding the REX1, OCT4, NANOG, SOX9, KLF4 and GAPDH. GAPDH served as a house-keeping gene. Statistically significant difference was observed between non-primed and IFN-γ primed WJ-MSCs regarding the REX1 (p < 0.001).
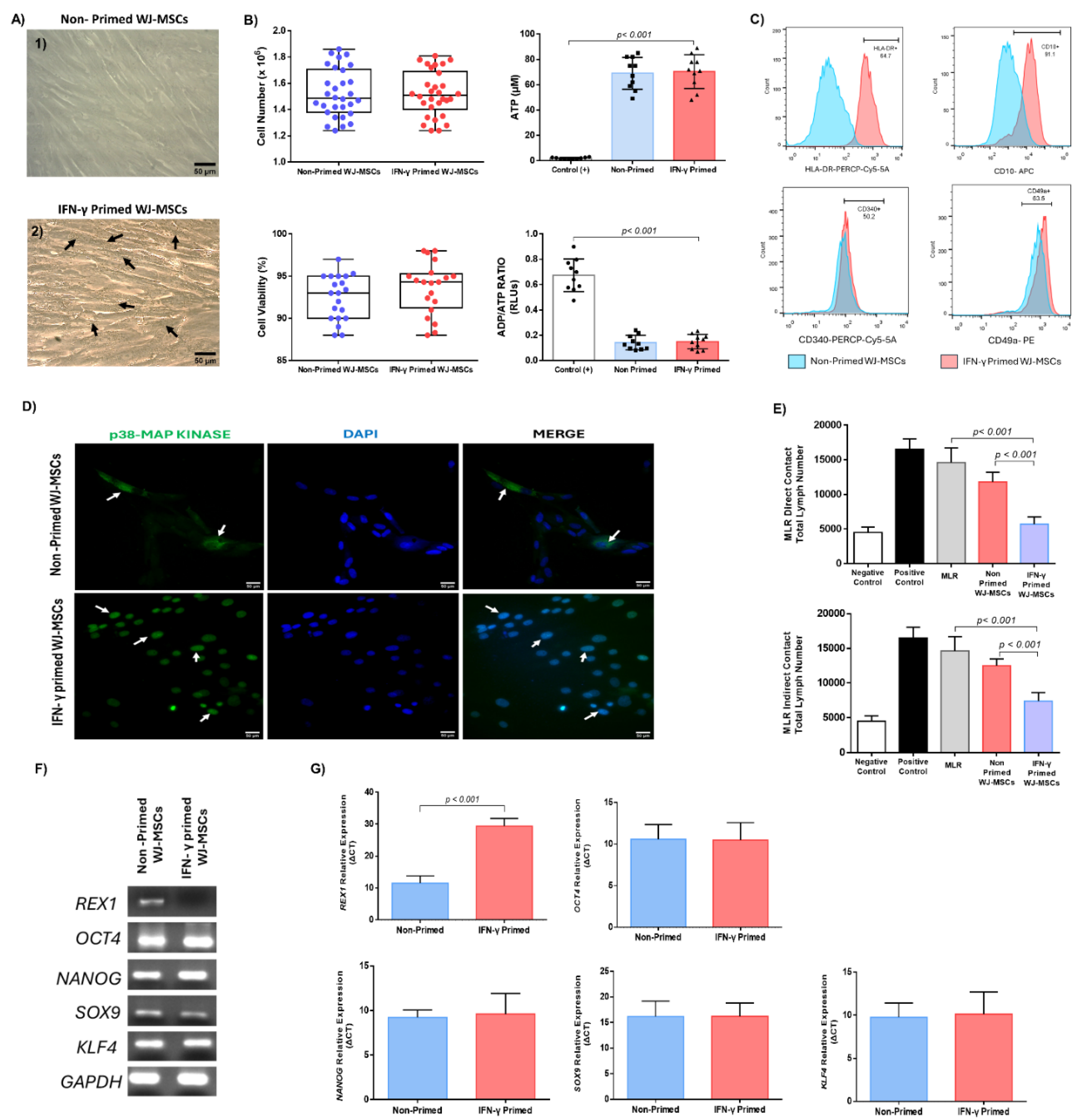
Figure 3.
Biomolecules quantification of IFN-γ primed and non-primed WJ-MSCs. Quantification of anti-inflammatory cytokines, including the IL-1Ra (A), IL-6 (B), IL-10 (C) and IL-13 (D), exerted by non-primed and IFN-γ primed WJ-MSCs. Quantification of growth factors involved in immunoregulation, including the TGF-β1 (Ε), FGF (F), PDGF (D), VEGFA (H), HGF (I), produced by non-primed and IFN-γ primed WJ-MSCs. Quantification of other immunomodulatory biomolecules, such as IDO and NO, exerted from non-primed and IFN-γ primed WJ-MSCs. Statistically significant difference was observed in all quantified biomolecules between non-primed and IFN-γ primed WJ-MSCs (p < 0.001).
Figure 3.
Biomolecules quantification of IFN-γ primed and non-primed WJ-MSCs. Quantification of anti-inflammatory cytokines, including the IL-1Ra (A), IL-6 (B), IL-10 (C) and IL-13 (D), exerted by non-primed and IFN-γ primed WJ-MSCs. Quantification of growth factors involved in immunoregulation, including the TGF-β1 (Ε), FGF (F), PDGF (D), VEGFA (H), HGF (I), produced by non-primed and IFN-γ primed WJ-MSCs. Quantification of other immunomodulatory biomolecules, such as IDO and NO, exerted from non-primed and IFN-γ primed WJ-MSCs. Statistically significant difference was observed in all quantified biomolecules between non-primed and IFN-γ primed WJ-MSCs (p < 0.001).
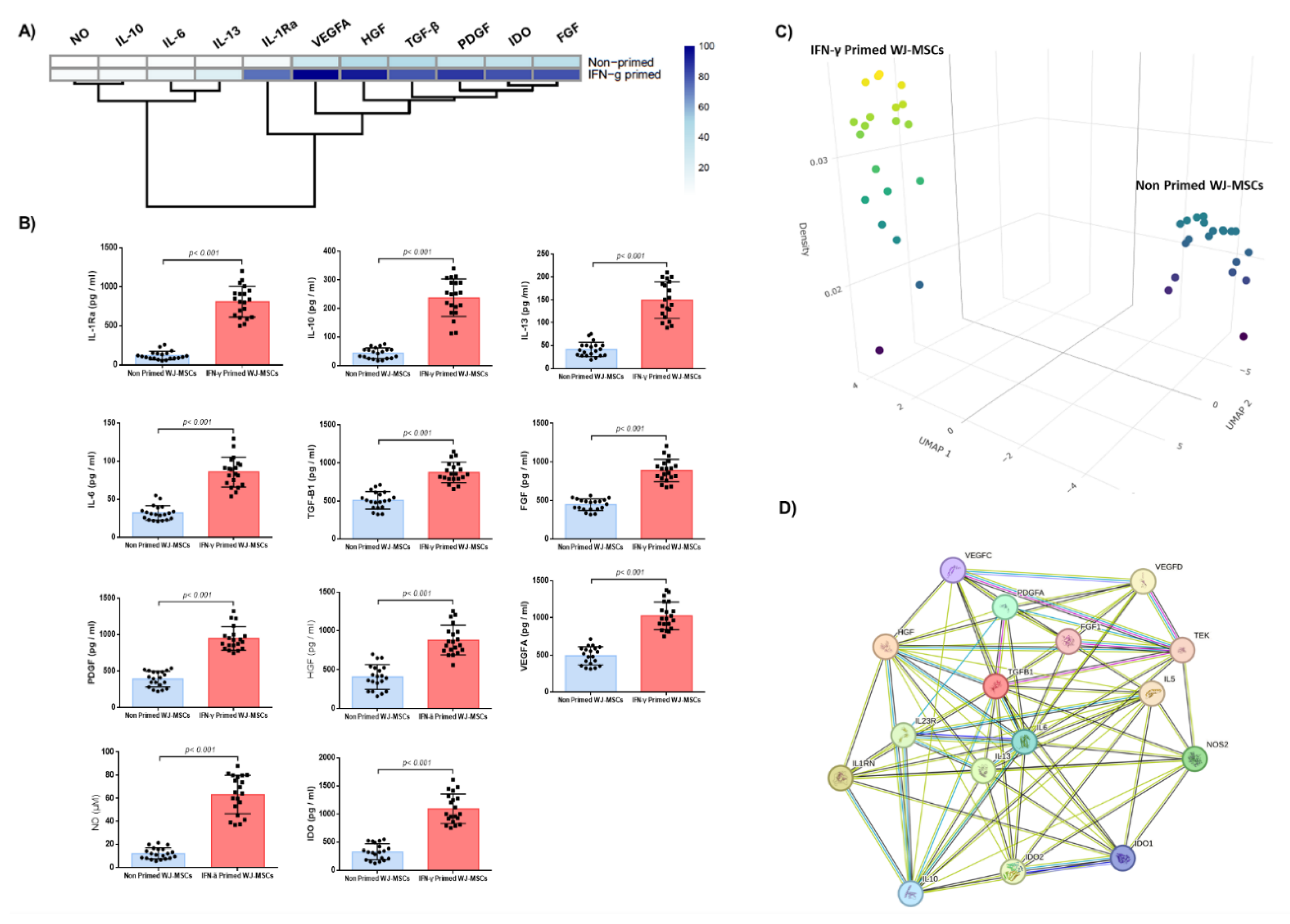
Figure 4.
Determination of HLA class I and II allele frequencies of WJ-MSCs. HLA class I and II frequencies in WJ-MSCs (A). Neighbor joining tree of the most frequent HLA class I, including the HLA-A, HLA-B and HLA-C (B) and class II, including the HLA-DRB1, HLA-DQB1 and HLA-DPB1 (C) alleles. Hierarchical clustering of WJ-MSCs based on the HLA class I and II frequencies and biomolecule secretory profile (D).
Figure 4.
Determination of HLA class I and II allele frequencies of WJ-MSCs. HLA class I and II frequencies in WJ-MSCs (A). Neighbor joining tree of the most frequent HLA class I, including the HLA-A, HLA-B and HLA-C (B) and class II, including the HLA-DRB1, HLA-DQB1 and HLA-DPB1 (C) alleles. Hierarchical clustering of WJ-MSCs based on the HLA class I and II frequencies and biomolecule secretory profile (D).
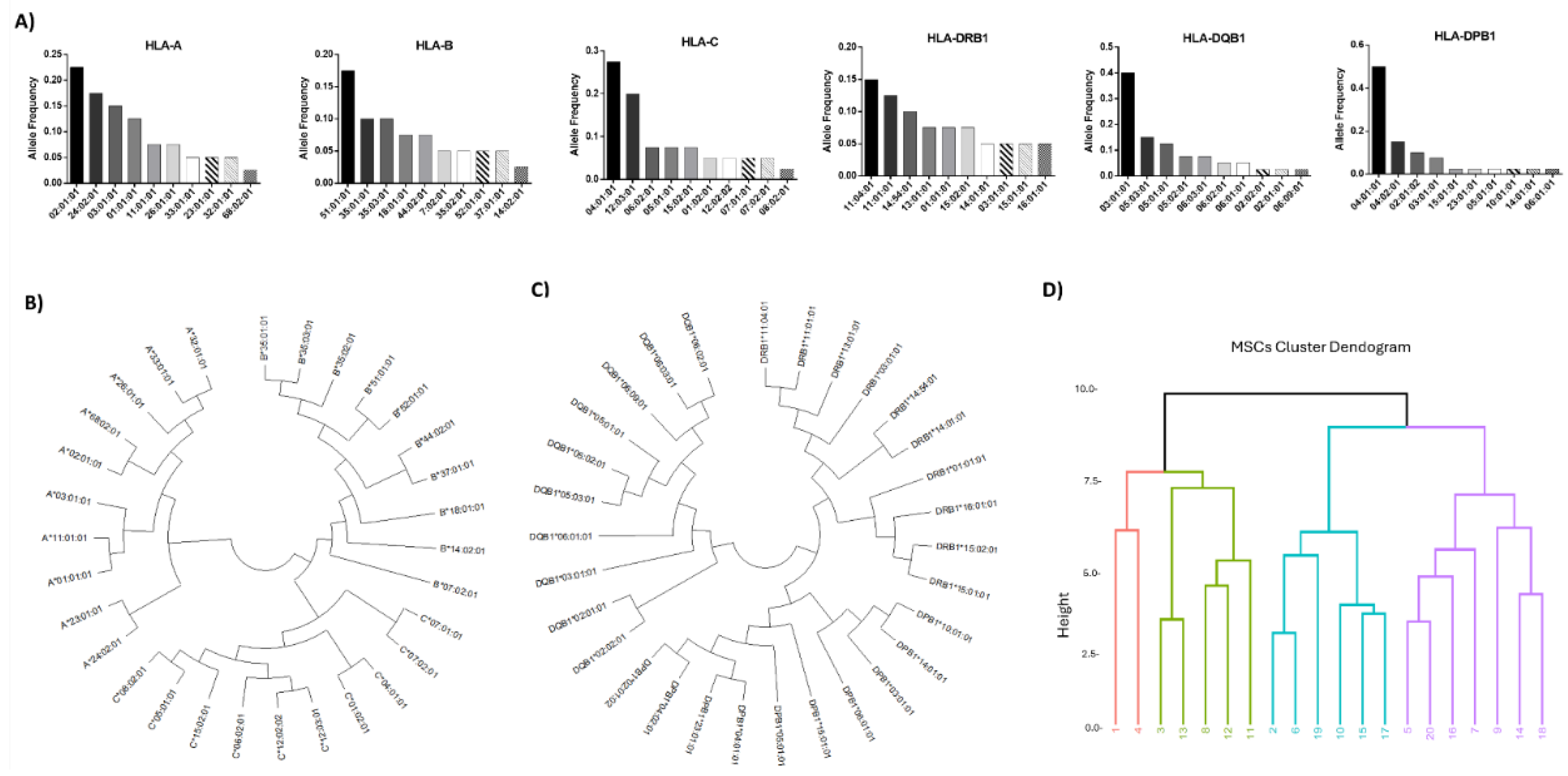
Figure 5.
Representative images of HLA class I (HLA-A alleles) and class II (HLA-DRB1 alleles) nucleotide comparison regarding exons 2 and 3. Multiple nucleotide changes were observed in exons 2 and 3 within the different alleles of HLA-A (A) and in exon 2 within the different alleles of HLA-DRB1 (B).
Figure 5.
Representative images of HLA class I (HLA-A alleles) and class II (HLA-DRB1 alleles) nucleotide comparison regarding exons 2 and 3. Multiple nucleotide changes were observed in exons 2 and 3 within the different alleles of HLA-A (A) and in exon 2 within the different alleles of HLA-DRB1 (B).
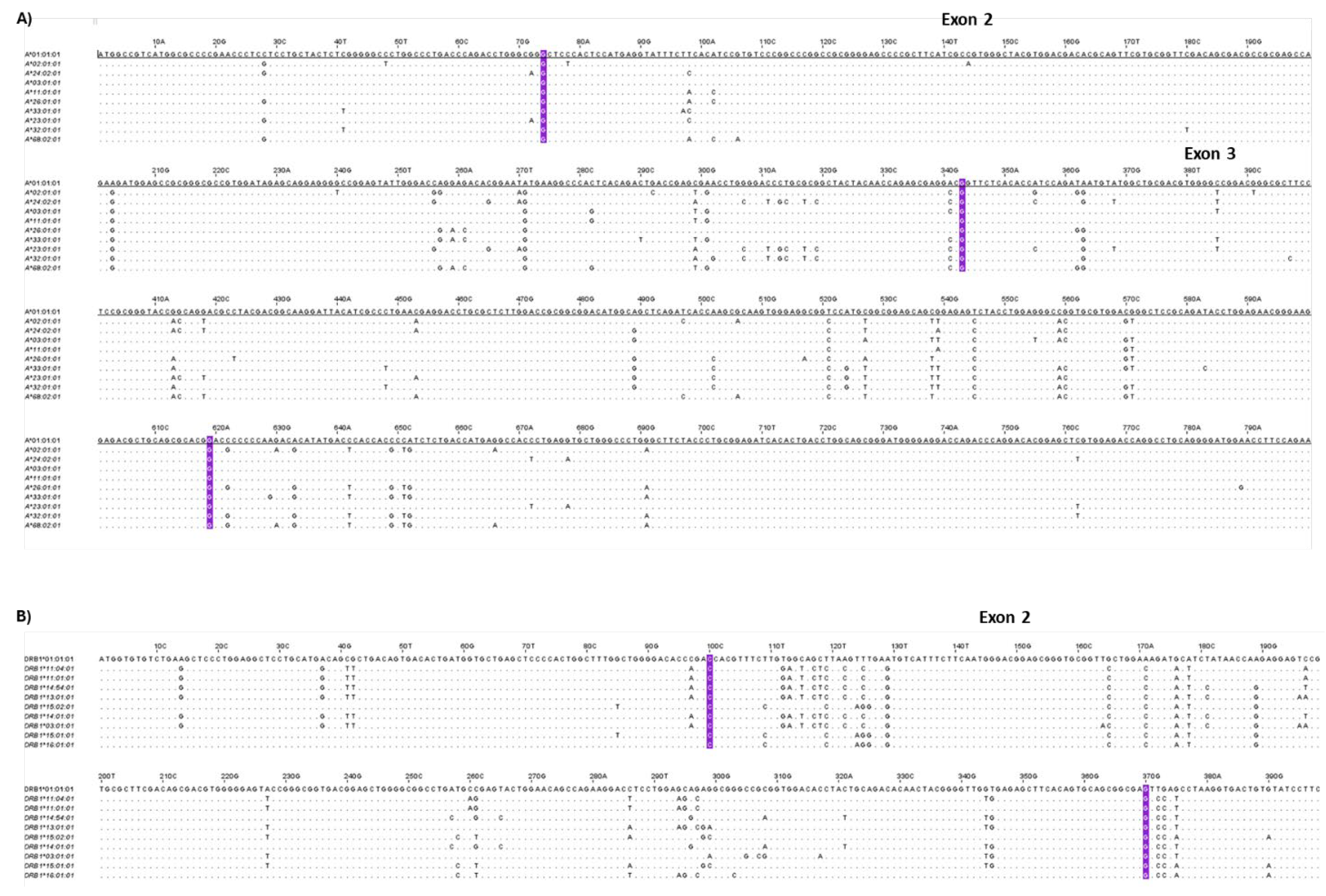

Table 1.
Primer pairs used for gene expression analysis.
| Gene | Forward Sequence | Location | Reverse Sequence | Location | Amplicon Size |
| REX1 | CCCTGGAATACGTCCCCAAG | 718-737 | CATCCTGTGAGGACTGGACC | 1227-1208 | 510 |
| OCT4 | GTGTTCAGCCAAAAGACCATCT | 532-553 | GGCCTGCATGAGGGTTTCT | 687-669 | 156 |
| KLF4 | CGGACATCAACGACGTGAG | 563-581 | GACGCCTTCAGCACGAACT | 701-683 | 139 |
| NANOG | TTTGTGGGCCTGAAGAAAACT | 83-103 | AGGGCTGTCCTGAATAAGCAG | 198-178 | 116 |
| GAPDH | GGAGCGAGATCCCTCCAAAAT | 108-128 | GGCTGTTGTCATACTTCTCATGG | 304-282 | 197 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



