
Submitted:
11 August 2025
Posted:
13 August 2025
You are already at the latest version
Abstract
Although wetlands are key habitats for biodiversity conservation, they are also among the most threatened ecosystems in the world. They are mainly affected by human pressures and threats, even when they are included in protected areas. The Padule di Fucecchio area is one of the largest and most significant inland marshes in Italy. It is also one of the wetlands of international importance listed in the Ramsar Convention. However, studies of the plant communities it hosts are surprisingly scarce and out of date. To address this, a survey of aquatic and marshy vegetation was conducted. This analysis provided an unprecedented census of the area's current aquatic and marsh vegetation, also reporting community types previously unknown in this territory. Some of these are completely new and are described here for the first time. However, a comparison with previous data revealed that significant changes to the vegetation structure have occurred in recent decades. The hydrophyte communities have almost completely disappeared and many of the most sensitive plants in the most sensitive marsh communities have become rarer or disappeared. They have mostly been replaced by more resilient native plants and invasive alien species.
Keywords:
aquatic and palustrine habitats
; biological invasions
; conservation
; freshwater ecosystems
; phytosociology
; syntaxonom
; vegetation
1. Introduction
Wetlands are ecosystems of significant ecological, cultural and economic value, playing a crucial role both locally and globally. They are vital for conserving biodiversity, as they are home to a wide variety of animal and plant species, some of which are endemic or endangered [1,2,3,4]. They also provide multiple ecological services, such as water purification, regulating surface water flow, filtering nutrients, protecting against flooding, acting as carbon sinks, and many others [3,5,6,7,8,9]. However, they are among the world's most threatened habitats, primarily due to human pressures and threats such as agricultural and urban land transformation, overexploitation, water pollution, flow modification, habitat destruction or degradation, and invasion by alien species [1,3,8,10,11,12], not to mention that they are also subject to negative global impacts, such as climate change [13,14]. This worrying general trend has increased rapidly in recent years, even among wetlands included in protected areas [15,16,17]. Consequently, it is not surprising that freshwater ecosystems in Europe and Italy also include many threatened habitats with an unfavourable conservation status [18,19,20,21,22,23]. Moreover, in the Mediterranean Basin, the general trends are exacerbated by the fact that many habitats that are not considered to be of conservation relevance in Central Europe become so further south. In addition, several of these habitats are not included in the European Habitats Directive [24] or any other protection lists, despite being extremely rare and threatened in southern European Mediterranean countries [25,26,27,28,29,30].
The Padule di Fucecchio area is one of the largest and most significant inland marshes in Italy. It is also one of the wetlands of international importance listed in the Ramsar Convention. It is home to several protected natural areas of European and local interest due to its rich natural environment and the unique features of its hydrogeology and landscape. This wetland is a crucial habitat for numerous bird species, including migratory waterfowl, and plays a vital role in maintaining regional biodiversity by providing essential breeding and feeding grounds for birds, as well as hosting a variety of invertebrates, amphibians and plants [31]. Bird populations are regularly monitored [32], and there are also numerous studies of the area's flora, despite these being rather dated. Several specimens collected in the area from the 19th century to the present day are deposited in Tuscan herbariums [33,34,35]. Conversely, studies on plant communities are surprisingly scarce and out of date. To address this, a vegetation survey focusing on aquatic and marshy vegetation was conducted in recent years. The results of these investigations are presented in this work, with the aim of comparing the current data with previous findings and outlining the changes that have occurred in the landscape.
2. Materials and Methods
2.1. Study Area
2.1.1. Topographic Location and Hydrological Features
The Padule di Fucecchio marsh, which covers an area of around 2,000 hectares, is located in northern-central Tuscany, spanning the provinces of Florence and Pistoia and extending minimally into the provinces of Lucca and Pisa. Its centre coordinates are 43.800086°N, 10.799955°E (Figure 1).
It encompasses the natural floodplain formed by streams and ditches originating from the Apennines to the north, the Montalbano Hills to the east, and the Cerbaie Hills to the west. Its only outlet is the Usciana canal, which flows into the Arno River. Located at an average altitude of around 13 m a.s.l., the marsh sits 5 m lower than the Arno floods, which can reach 18 m a.s.l. at the confluence with the Usciana canal. Therefore, the flow towards the Arno River is hindered by the large quantity of sediments deposited by the major watercourse along its banks since the beginning of the Quaternary [31,33,36]. This territory has experienced numerous geological and climatic events from the Upper Pliocene to the present day. These events have determined changes in its geomorphology and the distribution of plant communities [37,38]. Human exploitation of the area's natural resources (fishing, agriculture, harvesting marsh herbs, etc.) since ancient times has been no less relevant in shaping the landscape [31]. From this perspective, the most profound alterations are those resulting from historical reclamation. This process began in the Renaissance period with the Medici family and was resumed in the second half of the 18th century by the Lorraine family. It continued until quite recent times [39]. Currently, a dense network of canals crosses the marsh area, while the permanent lake area depicted in ancient maps (e.g. Figure 2), has long since disappeared. Only during the rainy seasons, temporary lakes and ponds are present, some of which are used for hunting waterfowl [31]. The management of the water level currently depends essentially on hydraulic manoeuvres regulated by a Land Reclamation Consortium. The effects of these manoeuvres on natural conservation, in terms of both flooding and drying times and the quality of inflowing water, have often been considered inadequate [40].
2.1.2. Vegetated Landscape and Presence of Protected Areas
The Padule di Fucecchio is extensively characterized by reed beds covering large surface areas, in mosaic with wet meadows and shrublands (Figure 2). Concentric bands or a mosaic of meadows, poplar groves and arable land develop around the marsh area [31]. This wetland hosts a Natura 2000 protected area, which is at the same time both Special Area of Conservation and Special Protection Area (code: IT5130007) and some other protected areas of provincial and local interest [24,31]. The Bosco di Chiusi (approximately 320 hectares), which is another biotope of high naturalistic value (Natura 2000 code IT5140010), is the last remaining lowland forest in Valdinievole. It is located on the eastern side of the Padule di Fucecchio [23].
2.1.3. Climate and Bioclimate, Geological Substrates
With an average annual rainfall of about 1,000 mm, approximately three months of moderate drought in summer and an average annual temperature of 14.8 °C, the climate is usually classified as meso/sub-Mediterranean [42,43]. According to the classification of Thornthwaite & Mather [44], the climate belongs to the type ‘‘second mesothermic, suboceanic”, with concentration of thermal efficiency in summer [43]. According to Pesaresi et al. [45], the study area has a temperate oceanic-sub-Mediterranean bioclimate.
In terms of geological substrates, the whole area is formed by alluvial sands, muds and silts derived from Pliocene-Quaternary deposits [46].
2.2. Data set and Data Analysis
The dataset comprises 155 original georeferenced relevés carried out in plant communities dominated by aquatic and palustrine species. The data were collected in in spring and summer of 2022 and 2023 using the classical phytosociological method by Braun–Blanquet [47,48] and its updates [49,50,51,52]. After transforming the original Braun–Blanquet cover-abundance scale into the Van der Maarel ordinal scale [53], aquatic coenoses were separated from palustrine ones, based on the growth forms [54] of the dominant species. A statistical analysis was not necessary for the few (16 relevés) and well-differentiated aquatic plant communities. However, the resulting matrix of palustrine plant communities (139 relevés and 131 species) was analysed using cluster analysis in the R environment [55]. This analysis used the Bray–Curtis dissimilarity function vegdist from the vegan package [56] and the Ward method from the hclust function in the stats package [55].
The names of plant species follow the Portal to the Flora of Italy [57]. The full names, including the authors, are provided in the tables. In the text, the names are given without the authors for brevity. The syntaxonomic nomenclature of classes, orders, and alliances mainly follows the Vegetation of Europe by Mucina et al. [58] and subsequent updates [59]. However, in certain instances, the interpretation of the Italian Vegetation Prodrome [60] was preferred, as detailed in the description paragraph. The syntaxonomical nomenclature is in accordance with the 4th edition of the ICPN [61]. All tables relating to vegetation types attributed to syntaxa that have already been described have been published as supplementary material, while the table relating to a new syntaxon that is described here has been published in the main body of the article. A comprehensive syntaxonomic scheme is provided in Appendix A, which includes the complete names of higher-rank syntaxa not cited in the text for brevity. All other syntaxa cited in the text but not included in the syntaxonomic scheme are also cited in full, including the authors and date.
3. Results and Discussion
The dendrogram resulting from the cluster analysis of the palustrine plant communities is shown in Figure 3, where the correspondence between relevés and plant communities is given.
The classification provided by the cluster analysis was followed not entirely but for 97.5%, as in accordance with the phytosociological procedure, greater importance was given to the floristic composition, both in terms of dominance and bioindication, to discriminate phytocoenoses. The case of the community dominated by Butomus umbellatus can be regarded as an example, since it is clustered together with Bolboschoenus glaucus coenoses due to the minor presence of B. glaucus in the relevé, but is nevertheless considered autonomous. In a small number of other cases, the cluster analysis can differentiate some relevés based on floristic heterogeneity. However, these should be classified in the same plant community type, as species dominance and ecology are more indicative of this classification. The analysis enabled the identification of 34 different plant community types, which can be syntaxonomically classified into eight classes: Lemnetea, Potamogetonetea, Isoëto-Nanojuncetea, Littorelletea uniflorae, Bidentetea, Phragmito-Magnocaricetea, Molinio-Arrhenatheretea, Franguletea. Each community type is described and commented on the following paragraphs.
3.1. Aquatic Plant Communities
Lemnetum minuto-gibbae Liberman Cruz, Pedrotti et Venanzoni 1988 (Table S1, rel. 1)
In the study area, Lemna minuta grows in small monophytic stands and seems to be currently rather rare. This community can be attributed to Lemnetum minuto-gibbae association, described for Lake Titicaca in Bolivia [62] and reported in Central Italy for the lakes Trasimeno [63], Porta [64] and Massaciuccoli [65]. Lemna minuta is a species with a high invasive potential, due to its plasticity in respect to chemical water features and its high vegetative propagation capacity [66]. According to the Portal to the Flora of Italy [57], this alien species is present in many Italian regions and shows an invasive behaviour. It can often replace native species such as L. minor.
Persicarietum amphibiae Pohjala 1933 nom. mut. Prunier et al. 2019 (Table S1, rels. 2-5)
In the Padule di Fucecchio, communities dominated by the aquatic ecophene of Persicaria amphibia are the only hydrophytic communities having currently appreciable surface areas. They are not uncommon in the marsh areas where ponds (clearings in the reed beds used for hunting) have relatively clear waters (Figure 4).
P. amphibia-dominated stands are more or less dense, poor in species, and generally colonizes ponds, canals and ditches with still or slowly flowing, shallow, meso-eutrophic to eutrophic waters, subject to regular drying out (in this case P. amphibia develops the terrestrial ecophene), on very clayey substrates [67]. In the past these coenoses were often attributed to Polygono-Potamogetonetum natantis Soó 1964 association, but ecological studies have demonstrated that habitat requirements of communities dominated by Potamogeton natans and by Persicaria amphibia are different and support to split them into separate associations [68]. This vegetation type was already reported for Tuscany [69], though with different syntaxonomic invalid names (e.g., Polygonetum natantis Soó 1927, nomen nudum). Other authors consider P. amphibia-dominated communities referable to Potamo natantis-Polygonetum natantis Knapp et Stoffers 1962 association [70], but according to Prunier et al. [67], the valid priority name is Polygonetum amphibium “associaatio” Pohjala 1933, which they propose to update and change in Persicarietum amphibiae.
Potamo pectinati-Myriophylletum spicati Rivas Goday 1964 corr. Conesa 1990 (Table S1, rels. 6-7)
Small, sporadic and species-poor stands dominated by Myriophyllum spicatum were found along the Pescia di Pescia stream. They can be attributed to the Potamo pectinati-Myriophylletum spicati association (in the past the invalid synonym Myriophylletum spicati Soó 1927 nom. nud. was often used), which has a broad ecology, but is more common in streams with eutrophic waters [71]. This vegetation type is considered an indicator of poor water quality [72], tolerating turbidity, drying and human disturbance [70,73]. It has been also found in other Tuscan marshes, lakes and ponds [65,74].
Potamogetonetum denso-nodosi O. de Bolòs 1957 (Table S1, rels. 8-9)
Small stands dominated by Potamogeton nodosus were found in one of the northern peripheral canals of the marsh. Although Potamogeton nodosus communities are sometimes considered to be typical of running waters [75], their presence also in standing waters is rather common [76,77]. These communities, once probably more frequent in the Padule di Fucecchio canals, can be attributed to an impoverished aspect of the Potamogetonetum denso-nodosi O. de Bolòs 1957 association, included in the Potamogetonion alliance, which has been reported both in standing and in flowing waters [63].
Callitriche platycarpa community (Table S1, rel. 10)
A small stand dominated by Callitriche platycarpa was found in a temporary pool within a mixed woodland area, on the border between the Padule di Fucecchio and the Bosco di Chiusi protected areas. This species is rather rare in Italy, and in Tuscany it is mainly known on the basis of old records [78]. Although a single community has recently been reported for the Tuscan Maremma [77], the phytosociological data available for Italy are very scarce. Felzines [79] reported the association Veronico beccabungae-Callitrichetum platycarpae Grube ex Felzines 2016 for France, indicated as Veronico beccabungae-Callitrichetum platycarpae Mériaux 1978 by Prunier et al. [67]. It belongs to the Ranunculion fluitantis alliance and indicates the reophile character of this community. However, this species can grow in different habitats [80]. As we found this coenosis in standing water, we provisionally attribute it to Ranunculion aquatilis alliance.
Callitriche stagnalis communities (Table S1, rels. 11-12)
In the same environment where we found the Callitriche platycarpa community, two small stands totally dominated by C. stagnalis occurred in shaded small ponds, subject to almost total drying up in the summer season and probably disturbed by trampling. C. stagnalis is typical of regions with hot summers and tolerates emergence and shading. It lives in shallow, meso-eutrophic, slightly acidic or neutral clear waters [67]. This species forms communities both in still and in running waters, showing a rather broad ecological behavior. Consequently, from a syntaxonomic point of view, the classification of this vegetation type is various and not univocal. In running waters, the association Callitrichetum stagnalis Kaiser 1926 (also indicated in the past with the synonym Callitrichetum stagnalis Segal 1964) was sometimes reported in Europe and in Italy [67,81], while for still waters other types of associations, such as Ranunculo ophioglossifolii-Callitrichetum stagnalis Brullo, Scelsi et Spampinato 2001 [77,82] have been described. Frequently, however, both because of their floristic poverty and the poor ecological characterization, communities dominated by this species are generically classified as phytocoenon or coenosis [74,83,84]. In our opinion, even for the monospecific stands surveyed in the study area, it is preferable to follow this type of interpretation, attributing them to the Ranunculion aquatilis alliance.
Callitriche brutia communities (Table S1, rels. 13-14)
Callitriche brutia is a Mediterranean-Atlantic species that can participate to different vegetation types. It can be found in aquatic communities of Ranunculion aquatilis alliance [77,85], or palustrine coenoses attributed to Oenanthion (today = Eleocharito palustris-Sagittarion sagittifoliae) alliance [86], or to Littorelletea class [77,86] or, more frequently, when it dominates in Mediterranean temporary wet habitats, to Isoëto-Nanojuncetea class [87,88,89,90,91]. In the study area, C. brutia-dominated communities have been sporadically found in a peripheral position with respect to Persicarietum amphibiae communities and in contact with marsh vegetation (Figure 5), in environments that dry out earlier and subject to greater fluctuations in water levels, so we refer them provisionally to the alliance Ranunculion aquatilis.
Potamo crispi-Ranunculetum trichophylli Imchenetzky 1926 (Table S1, rels. 15-16)
Ranunculus trichophyllus is one of the more common hydrophytes in Tuscany and can be found in several habitat types, both in flowing and in standing waters [26,74]. However, in the study area, R. trichophyllus-dominated communities are very rare. Small and paucispecific stands were only found in one of the northern peripheral canals of the marsh. These coenoses can be attributed to the association Potamo crispi-Ranunculetum trichophylli (in the past, the invalid synonym Ranunculetum trichophylli Soó 1927 was used), typical of oligo-mesotrophic to eutrophic waters and tolerating high levels of turbidity; this vegetation type tolerates strong water level variations and can live for a short time even in complete emersion [70].
3.2. Palustrine Plant Communities
Echinochloo cruris-galli-Sporoboletum schoenoidis Paradis & Lorenzoni 1994 nom. mut. nov. (Table S2)
The communities dominated by Sporobolus schoenoides grow in drying depressions at the edge of, or within, helophytic vegetation, such as Bolboschoenus laticarpus coenoses, or in the clearings resulting from the drying up of ponds and lakes (Figure 6).
S. schoenoides participates in various communities of ephemeral environments, gravitating mainly in coenoses of the Isoëto-Nanojuncetea class [92], but also in syntaxa indicating halophilous conditions, so much so that the association Heleochloetum schoenoidis Ţopa 1939 is often placed by some authors in the class Crypsietea aculeatae Vicherek 1973 [93,94]. Paradis [95] analysed different types of community, which he initially differentiated from Heleochloetum schoenoidis in several subassociations. These were subsequently formalised as new associations [96]. The comparison of these associations is used to evaluate the communities of the Padule di Fucecchio. Despite the presence of Cyperus michelianus in the Tuscan relevés, it is not possible to refer them to the association Crypsio schoenoidis-Cyperetum micheliani Martínez-Parras, Lorca, Esteban et Mesa 1988, which is always dominated by C. michelianus [97]. Furthermore, in the study area, the communities dominated by C. michelianus (see Table 1, sub Cyperetum micheliano-difformis) do not host S. schoenoides among the species and grow in rather different ecological situations, on more humid soils and in topologically distant sites from those dominated by S. schoenoides. The absence of Heliotropium supinum from Tuscany, where it was reported based on ancient finds not recently confirmed [57], does not even allow the attribution of our communities to Heliotropio supini-Heleochloetum schoenoidis Rivas Goday 1955, recently reported by Tomaselli et al. [88] for Puglia and otherwise synonymized with Crypsio schoenoidis-Cyperetum micheliani by Rivas-Martínez et al. [98]. The communities of the Padule di Fucecchio can instead be attributed to the association Echinochloo cruris-galli-Crypsidetum schoenoidis (whose name should be changed to Echinochloo cruris-galli-Sporoboletum schoenoidis), an association that grows in rather eutrophic and compact silty-clayey soils, in which the floristic composition is enriched with species linked to disturbed humid environments and constituting a sort of link towards the communities of the Bidentetea class [95]. As regards the classification at the higher levels of the association, originally placed by the authors in the class Isoëto-Nanojuncetea and in the alliance Heleochloion Br.-Bl. 1952, it should be pointed out that currently Mucina et al. [58] consider Heleochloion schoenoidis Br.-Bl. ex Rivas Goday 1956 a typically halophilous alliance belonging to the class Crypsietalia aculeatae. Since we believe that maintaining the original classification at the class level is appropriate, as was done by Tomaselli et al. [88], for example, we attribute the association to the Verbenion supinae Slavnić 1951 alliance. This alliance traditionally included many coenoses that gravitated around the Heleochloion Br.-Bl. 1952 alliance. In fact, some authors considered Verbenion supinae and Heleochloion to be synonyms [91].
Ranunculo flammulae-Juncetum bulbosi Oberdorfer 1957 (Table S3, rels. 1-2)
In a partially shaded site, in contact with aquatic Callitriche platycarpa vegetation, where the water level decreases, small communities dominated by Juncus bulbosus and Ranunculus flammula were detected. Juncus bulbosus has been indicated as a typical or frequent species in a large number of vegetation types [82,86,92,99], but when it is associated to Ranunculus flammula these communities can be attributed to Ranunculo flammulae-Juncetum bulbosi, an association which includes stands of low-growing perennial amphibious herbs occurring in the littoral zones of lakes, ponds, pools and ditches along forest roads, periodically flooded by oligotrophic to mesotrophic, rarely dystrophic water [100]. This vegetation type has been recorded in several western and central European countries [100,101,102,103] and can be attributed to Littorellion uniflorae alliance.
Juncus bulbosus - Gratiola officinalis communities (Table S3, rels. 3-4)
In sites ecologically similar and near to Ranunculo flammulae-Juncetum bulbosi communities, small stands dominated by Juncus bulbosus and with relevant cover values of Gratiola officinalis have been detected. G. officinalis typically grows in areas with wet soils for a fairly long period in spring, drying up in summer [77,86]. Some associations characterized by G. officinalis have been described (e.g., Alismo lanceolatae-Gratioletum officinalis Biondi & Bagella 2005) but our relevés are paucispecific and not dominated by G. officinalis. Therefore, this vegetation type could be interpreted as a situation in which the Ranunculo flammulae-Juncetum bulbosi association is replaced by these communities, in which G. officinalis, being less ecologically demanding than R. flammula, is advantaged by conditions of greater drying out and disturbance. These communities can be always attributed to the Littorellion uniflorae alliance.
Cyperetum micheliani-difformis ass. nova (Table 1, Holotypus: rel. 4)
In areas that flood during the winter and dry out during the summer, at the edges of reed beds and in clearings free of marsh vegetation, as well as along paths between reed beds, a vegetation type that is generally widespread throughout the study area grows, consisting of a mixture of native and alien species (Figure 7).
These communities typically have two layers. the lower one is often dominated by Cyperus michelianus, with the presence, sometimes with high cover, of Portulaca oleracea, as well as the alien plants Lindernia dubia, Amaranthus emarginatus and Eclipta prostrata. The taller layer is often dominated by another alien species, Cyperus difformis, and is frequently accompanied by Xanthium orientale (which sometimes has high cover or is dominant), Cyperus odoratus and Echinochloa crus-galli. In terms of their structure, physiognomy and, to some extent, their floristic composition, these communities have many similarities to the Cyperetum micheliano-glomerati association described by Pellizzari [104] for the eastern stretch of the Po River. However, this last vegetation type is characterised by two species of Cyperus absent in our relevés (C. glomeratus and C. squarrosus, the latter of which is yet to be found in Tuscany), as well as by Amaranthus tuberculatus. Compared to Cyperetum micheliano-glomerati, in the relevés of the study area higher cover values are provided by Cyperus difformis, a species that infests rice crops and is characteristic of the Oryzo-Cyperetum difformis association W.Koch 1954 [104,105], and partly by Xanthium orientale, which sometime becomes dominant. X. orientale participates in different communities linked to the disturbed environments of humid areas, gravitating towards Chenopodion rubri (e.g., Polygono lapathifolii-Xanthietum italici Pirola & Rossetti 1974) or Bidention (e.g., Xanthio-Polygonetum persicariae O.Bolós 1957) alliances. In our relevés, Amaranthus emarginatus is also present, while it is missing from the relevés by Pellizzari [104]. On average, the communities of the Padule di Fucecchio are much poorer in species, with an average of seven species per relevé compared to over 11 for the Po River. Given the differences highlighted, we propose establishing a new association named Cyperetum micheliano-difformis, which in the study area plays an ecological role similar to that of the Cyperetum micheliano-glomerati association in the Po River area. The characteristic species of the Cyperetum micheliano-difformis association are Cyperus michelianus, C. difformis, Xanthium orientale and Amaranthus emarginatus. Similarly to how Pellizzari [104] classified the association he described, the floristic composition allows us to place Cyperetum micheliano-difformis in the Bidention alliance. This is consistent with the ecology of the communities, which develop on muddy emergent soils in eutrophic environments subject to significant human disturbance. In our relevés, some stands dominated by Cyperus odoratus can also be noted, an aspect which is more closely linked to the Po River communities described by Pellizzari [104], together with some other relevés in which the frequency and/or cover values of alien species is lower and in which Cyperus michelianus forms dense carpets with high cover values. This species generally characterises the coenoses of the Isoëto-Nanojuncetea class. Such situations may suggest that, as hypothesised for other coenoses in which C. michelianus coexists with other alien species [106], the described association may constitute communities that have evolved into coenoses with a strong presence of alien species from an original Cyperetum michelianus Horvatic 1931 or similar syntaxa, favoured over time by increased disturbance and water eutrophication.
Bidens frondosa communities (Table S4, rels. 1-4)
This vegetation type is dominated by the alien and invasive American neophyte Bidens frondosa, with a constant and often huge presence of another invasive neophyte, Symphiotrychum squamatus. These communities are found near reed beds, ponds and canals, in slightly higher areas, where water retention is relatively less persistent, and the soil tends to dry more quickly. Similar coenoses have been referred by some authors [107] to Polygonetum hydropiperis Passarge 1965, the valid name of Polygono hydropiperis-Bidentetum (Koch 1926) Lohmeyer in Tüxen 1950, or to Leersio-Bidentetum tripartitae (Poli et J. Tüxen 1960) Zaliberova et al. 2000 bidentetosum frondosae (Felzines & Loiseau 2005), or to Cypero eragrostidi-Bidentetum frondosae [108]. Anyway, in all these cases, the paucispecific communities we detected lack some of the other characteristic or differential species of the above mentioned associations (i.e., Polygonum hydropiper, Leersia oryzoides, Cyperus eragrostis), and we therefore prefer not to attribute them at the level of association, but only to Bidention alliance.
Veronica peregrina communities (Table S4, rels. 5-7)
Veronica peregrina is an American neophyte that in the study area sometimes forms communities in which it is dominant, especially along the wet banks of ditches and canals. This species, already reported for the Fucecchio marshland by Roma-Marzio et al. [109], is considered as a casual alien species in Tuscany, even if this status should be changed in naturalized according to our relevées, showing rather dense and not ephemeral stands in the study area. This species forms communities belonging to several types of syntaxa, from Nanocyperion, e.g. Filaginello uliginosae-Veronicetum peregrinae Molero Brion. et Romo 1988 [110], to the Polygono-Poetea class [111]. On the other hand, according to Julve [112], the species is characteristic of the Bidention alliance. Our relevés, paucispecific and formed in addition to V. peregrina by a few other species from the neighbouring coenoses (Phragmites australis, Persicaria lapathifolia, Phalaris arundinacea) do not allow an in-deep syntaxonomic analysis on the community's attribution at higher levels. From an ecological point of view, however, the coenoses develop at the limit of the vegetation of tall helophytes, in wet and eutrophic soils, disturbed by human activities (mowing, trampling, etc.) and by water level fluctuations, suggesting a provisional placement in the Bidention alliance.
Portulaca oleracea communities (Table S4, rels. 8-9)
In the same desiccated areas where Echinochloo-Sporoboletum or Cyperetum micheliani-difformis are present, Portulaca oleracea sometimes becomes dominant for small stretches (Figure 8), and gives rise to paucispecific communities that can be considered an impoverished transition aspect between the most hygrophilous coenoses (Bidention alliance) and the driest and ruderal ones of the classes Chenopodietea Br.-Bl. in Br.-Bl. et al. 1952 or Stellarietea mediae Tüxen et al. ex von Rochow 1951, in which P. oleracea is often included [113,114]. This vegetation type, difficult to attribute to higher level syntaxa, can be provisionally referred to Bidention alliance.
Polygono lapathifolii-Xanthietum italici Pirola et Rossetti 1974 (Table S5, rels. 1-3)
In the study area, in the late summer-autumn period, on the desiccating and desiccated sandy-silty, eutrophic substrates emerging from the dried water bodies, large surfaces are covered by annual communities dominated by Xanthium orientale, an invasive alien species widespread in Italy, accompanied by Persicaria lapathifolia, Echinochloa crus-galli, Stachys palustris, and few other plants (Figure 9). This pioneer vegetation type is frequent along rivers, ponds and lake [69,115,116], and can be attributed to Polygono lapathifolii-Xanthietum italici association (in the past, X. italicum / X. orientale subsp. italicum was considered a distinct taxon but today is considered a synonym of X. orientale, see Tomasello [117]). Since these communities form later in the growing season than Cyperetum micheliani-difformis, depending on when the survey is carried out, it is sometimes possible to detect transitional features, where species from the earliest communities and those in progress coexist. (e.g., rels. 21-22 in Table 1).
Echinochloo-Polygonetum lapathifolii Soó et Csurös 1947 (Table S5, rels. 4-8)
In the areas bordering reed beds, canals and Bolboschoenus coenoses, on moist soils that have emerged for longer periods, communities dominated by Echinochloa crus-galli and/or Persicaria lapathifolia can be found. They can be probably attributed to Echinochloo-Polygonetum lapathifolii, an association growing on soils rich in nutrients and with high pH [118]. These communities are rather similar to Polygono-Xanthietum italici from an ecological view point, but can be distinguished from a floristic one, and have already been reported for other Tuscan and European wetlands [26,119,120]. Along the course of some large French rivers (Loire, Allier), for moist sandy areas that dry out progressively from mid-spring, Felzines and Loiseau [121] described an association named Persicario lapathifoliae-Echinochloetum cruris-gallii, whose relevés seem to be very similar to ours, though some floristic differences can be noted (e.g., the presence in French communities of Panicum capillare, Leersia oryzoides, Eragrostis pectinacea and others). The problem of a correct syntaxonomic attribution of Tuscan communities at the association level must be probably studied more in depth with a dedicated work, so they can be provisionally referred to Chenopodion rubri alliance.
Phragmitetum australis Savič 1926 nom. corr. (Table S6, rels. 1-10)
Phragmites australis forms dense species-poor stands along submerged and emergent shores of lakes, swamps, pools, ponds, riverbanks, and canals [122,123]. These communities largely characterise the plant landscape of the Padule di Fucecchio, and can be attributed to the Phragmitetum australis association, which is quite common in Italy [122,123] and Tuscany [26,77,124,125]. In several large wetlands of Central Italy, these coenoses seem to be affected by the die-back syndrome and have shown a serious decline [27,126]. Despite that, no signs of die-back (e.g. the clumping habitus) were observed in the study area. This is probably due to the fact that in the Padule di Fucecchio strong seasonal water fluctuations, naturally and even artificially regulated, reduce the risk of permanent submergence, which seems to be an unfavorable ecological condition for Phragmitetum [27].
Typhetum latifoliae Nowiński 1930 (Table S6, rels. 11-13)
The communities dominated by Typha latifolia can be attributed to Typhetum latifoliae, an association very common in Italy, which grows in several different habitats, such as ponds, shores of lakes, banks of slow-running streams, deltas, swamps, canals and ditches [122], also tolerating the summer drying [127]. These communities are often very poor in species or even monospecific [128,129], a feature confirmed also in the study area, where this vegetation type is not very frequent and consists mainly of linear formations along peripheral ditches and canals.
Typhetum angustifoliae Pignatti 1953 (Table S6, rel. 14)
The communities dominated by Typha angustifolia can be attributed to Typhetum angustifoliae, an association typical of several wetland types which grows in mesotrophic to eutrophic waters, often monospecific or species-poor [122]. Though the association is rather common in Italy [122], and in Tuscany [65,125,130,131], in the Padule di Fucecchio it was found to be rather rare: we surveyed only one T. angustifolia-dominated community, in the widening of a ditch (probably of artificial origin), in a peripheral area of the marsh, between cultivated fields.
Schoenoplectetum lacustris Chouard 1924 nom. mut. nov. Lastrucci et al. 2023 (Table S6, rels. 15-19)
The communities dominated by Schoenoplectus lacustris can be attributed to the Schoenoplectetum lacustris association, which has a strong pioneer feature, and is reported for several habitat types, such as shores of mesotrophic to eutrophic lakes, ponds, or canals; it usually grows in deeper and longer-lasting water than other types of reed vegetation [122,125,132,133], and this is also confirmed in the study area.
Phalarido arundinaceae-Bolboschoenetum laticarpi Passarge 1999 corr. Krumbiegel 2006 (Table S6, rels. 20-29)
Bolboschoenus laticarpus, contrarily to B. glaucus, is also present in central Europe and lives only in freshwater habitats (Hroudová et al., 2009). In the study area, the communities in which it dominates are widespread, probably as much as Bolboschoenetum glauci or more. These dense and paucispecific coenoses can be probably attributed to Phalarido arundinaceae-Bolboschoenetum laticarpi, an association floristically rather heterogeneous living in floodplains of large rivers [134]. Phalaris arundinacea is not present in our relevé but is widespread in the area, while another differential species, Xanthium orientale, is frequent. To our knowledge, this vegetation type has not been previously reported for Italy, but it is likely not rare according to the updated distribution of the species [135].
Cyperetum longi Micevski 1957 (Table S7, rel. 1)
In the peripheral parts of the marsh, along the secondary ditches between meadows and arable land, small stands dominate by Cyperus longus are sporadically present. These communities grow in shallow water environments, which often dry out rather early, and can be attributed to Cyperetum longi, an association distributed in southern Europe and the Mediterranean [69,136,137], referable to Magnocaricion elatae alliance.
Caricetum ripariae Máthé et Kovács 1959 (Table S7, rels. 2-9)
This association lives in meso- and eutrophic wetlands emerged since late spring [122,123]. It was often found to be frequent in rather disturbed habitats [26,65]. In the study area the association is quite widespread along the edges of canals, around reed beds and in wet depressions. These communities are characterized by a large dominance of Carex riparia and by the presence of other species typical of upper syntaxonomic units, such as Phragmites australis, Bolboschoenus glaucus and B. laticarpus, Convolvulus sepium, Lythrum salicaria. They can be attributed to Magnocaricion gracilis alliance.
Caricetum vesicariae Chouard 1924 (Table S7, rels. 10-12)
Along the lower part of the ditch banks, in humid depressions and meadows, some communities dominated by Carex vesicaria were sporadically detected. They are referable to Caricetum vesicariae, a typical association of mesotrophic to eutrophic habitats, permanently flooded or saturated with water for most of the year [122,134]. Caricetum vesicariae was already known for lowland and montane wetlands in Tuscany [125,139] and can be attributed to Magnocaricion gracilis alliance.
Phalaridetum arundinaceae Libbert 1931 (Table S7, rels. 13-19)
Phalaris arundinacea-dominated communities occur in complexes of marsh vegetation in lowland river floodplains, in shallow depressions, in the littoral zones of water bodies and ditches with still or flowing water, generally eutrophic and base-rich, and in other similar environments [69,84,127]. In the study area, these communities have been detected along canals and ditches, where soils remain more or less constantly wet, and are not directly affected or disturbed by currents. For these reasons, they can be attributed to Phalaridetum arundinaceae, and not to Rorippo-Phalaridetum arundinaceae Kopecký 1961, an association typical of less humid and more disturbed sites, with longer exposure to conditions of complete emersion [70,127]. In any case, even if it has not been detected by us, the presence of Rorippo-Phalaridetum arundinaceae in the area is possible.
Leucojum aestivum communities (Table S7, rels. 20-21)
Leucojum aestivum can participate to different vegetation types, and can be found both in the understory of floodplain hygrophilous woods, dominated for example by Fraxinus angustifolia subsp. oxycarpa, Salix alba, etc. [23,140,141] and in herbaceous hygrophilous communities of Magnocaricetalia [119,142]. These communities, surveyed mostly for their rarity and their conservation interest, were present at the edge of the Padule, in a clearing of a partially shaded forest site, of which they can probably be considered a herbaceous sinusia. They can be provisionally attributed to Magnocaricion gracilis alliance.
Glycerio-Sparganietum neglecti Koch 1926 (Table S8, rels. 1-3)
Some communities dominated by Sparganium erectum L. sensu latu were found in the peripheral canals of the Padule di Fucecchio. According to Landucci et al. [122,123], they can be attributed to Glycerio-Sparganietum neglecti, which should be considered as a “macro-association” including other associations used in the past, such as Sparganietum erecti Roll 1938 and Sparganietum microcarpi (Weber 1976) Passarge 1978, due to the frequent uncertain taxonomic identification of the species in this group [143]. These communities usually grow in environments considered intermediate between Phragmition and Glycerio-Sparganion ecological conditions, so their attribution to the alliance level is not always univocal [122,123,134]. In the study area these canals generally have strong fluctuations of water levels, justifying the attribution of Glycerio-Sparganietum to the alliance Glycerio-Sparganion.
Eleochariteum palustris Savič 1926 (Table S9, rels. 1-3)
These communities are dominated by Eleocharis palustris and can be attributed to the Eleochariteum palustris association. They show a typical pioneer behavior, often developing in the wet soils emerging during the dry season [84]. In our relevés, we only found E. palustris subsp. palustris, but in the area, the presence of other subspecies is possible [144]. This association, typically poor in species, is rather common in Italy [77,122,125,145], but in the Padule di Fucecchio it is not so widespread. It was sometimes found along some canals or in the muddy areas surrounding the ponds (clearings for hunting) open in the reed beds.
Oenantho aquaticae-Rorippetum amphibiae Lohmeyer 1950 (Table S9, rels. 4-6)
Communities dominated by Rorippa amphibia generally occur in lowlands on lacustrine and fluvial sediment accumulations, and grow in marshes, ponds, pools, canals and lentic sections of rivers, where in summer water table often drops far enough to expose the bottom [127]. This vegetation type can be attributed to Oenantho aquaticae-Rorippetum amphibiae, an association sporadically present in the Padule di Fucecchio (Figure 10) and in other Tuscan wetlands [77].
Bolboschoenetum glauci Grechushkina, Sorokin et Golub 2011 (Table S9, rels. 7-13)
After taxonomic and distributional revisions of the genus Bolboschoenus by Hroudová et al. [146,147] for Europe and by Di Natale et al. [135] for Italy, the knowledge on the distribution of this complex of species (B. maritimus sensu lato) and on their dominated vegetation types have been deeply updated [134,148]. B. glaucus does not occur in inland central Europe, can grow both in freshwater and saline habitats and is mainly distributed in southern Europe, showing a thermophilic character [134,135]. In the study area, it forms dense, paucispecific and rather widespread communities in which it dominates, that can be attributed to the Bolboschoenetum glauci association, already indicated in other Tuscan wetlands [77,133].
Butometum umbellati Philippi 1973 (Table S9, rel. 14)
In the study area Butomus umbellatus is sporadically present in some vegetation types (e.g., in Schoenoplectetum lacustris coenoses), but rarely it forms communities in which it dominates. We could perform only one relevé of this type, that can be attributed to Butometum umbellati association. In Italy and in Tuscany this vegetation type is generally sporadic [69,138,149], and in the Padule di Fucecchio it is probably much less widespread than in the past.
Polypogono viridis-Paspaletum distichi Br.-Bl. in Br.-Bl., Gajewski, Wraber et Walas 1936 nom. mut. nov. (Table S10, rels. 1-10)
These communities are characterised by the dominance of the invasive alien species Paspalum distichum [150] and in the past have generally been attributed to the Paspalo distichi-Polypogonetum viridis association. This vegetation type has a mostly Mediterranean distribution and often results to be very poor in species [65,83,115,124]. Our relevés also confirm this feature, showing a strong dominance of P. distichum. In the study area, the species is found in the disturbed banks of the canals and around the ponds (clearings for hunting) in the reed beds. It forms dense mono- or paucispecific carpets in contact with hygrophilous herbaceous or helophytic communities. Only in few cases that can be interpreted as transition aspects (e.g., rel. 10 in Table S10), some stands can be co-dominated by other species, such as Cyperus sp.pl. From a nomenclatural viewpoint, the association name requires reversal, since P. distichum always dominates in both the original description [151] and the aforementioned works (including our relevés). We therefore propose changing it to Polypogono viridis-Paspaletum distichi (Polypogon viridis is the accepted name by several floras of Agrostis viridis and Agrostis semiverticillata). Furthermore, since the Paspalo distichi-Agrostion semiverticillatae alliance is based on the Paspalo distichi-Polypogonetum viridis association [152], the alliance name could also be changed to Polypogono viridis-Paspalion distichi Br.-Bl. in Br.-Bl. et al. 1952 nom. mut. nov. Regarding attribution to higher-level taxa, Mucina et al. [58] refer the Paspalo distichi-Agrostion semiverticillatae alliance to the Paspalo–Heleochloetalia Br.-Bl. ex Rivas Goday 1956 order, of the Bidentetea class. This class includes summer annual pioneer lacustrine bank vegetation. However, both Paspalum distichum and Polypogon viridis are perennial species. Therefore, we prefer to attribute the Polypogono viridis-Paspaletum distichi association to the Holoschoenetalia order of the Molinio-Arrhenatheretea class, in accordance with the Italian Prodrome [60].
Frangulo alni-Salicetum cinereae Graebner et Hueck 1931 (Table S11, rels. 1-4)
Salix cinerea-dominated communities are typical of marshes and grow in conditions of prolonged submersion. They are an infrequent example of the palustrine edapho-hygrophilous series in the temperate bioclimatic region [84]. These communities appear as species-poor, dense shrublands, generally in contact with plant communities of Phragmito-Magnocaricetea class, or directly bordering water bodies with or without aquatic vegetation. In Padule di Fucecchio, they are uncommon and can be considered a relic because their ecological niche has been occupied by Amorpha fruticosa, an alien shrub that is widespread in the marsh and has been favoured directly and indirectly by human disturbance. A. fruticosa is also present in our relevés and sometimes dominates. This vegetation type and the problems posed by A. fruticosa were already known for other Tuscan wetlands, both near and far from the study area [26,139]. As to the syntaxonomic attribution, in the past these communities have been attributed either to Salicetum cinereae Zólyomi 1931 or to Frangulo alni-Salicetum cinereae associations which, according to Douda [153], are synonyms of Salicetum pentandro-auritae Passarge 1957. However, the floristic description of Salicetum pentandro-auritae appears to differ significantly from that of S. cinerea communities described under previous names. Furthermore, Frangulo alni-Salicetum cinereae has also been used in recent times [141], prompting us to provisionally maintain this name.
3.3. Past and Present Vegetation
As mentioned in the introduction, literature studies on the vegetation of the Padule di Fucecchio are particularly scarce. Furthermore, the level of detail with which this work analyses the current vegetation has never been achieved before, making comparison with previous data difficult. Some phytosociological information, dated and at varying degrees of detail, on neighbouring areas [139,154,155], or on the very small provincial reserves within the Padule [156,157] is available, but most of the information originates from floristic studies, which provide general qualitative descriptions of the vegetation. Apart from some data relating to the Padule di Fucecchio contained in ancient Tuscan floras [158,159,160] or in specific works [161], the first comprehensive work on the flora in the study area is considered to be that by Nannizzi [162]. In this work, the plant landscape is also described in broad terms, although the direct observations date back to 1932-1933. Furthermore, Nannizzi's floristic list complicates matters as he also reports species found by previous authors without distinguishing between them. Consequently, if he does not mention them in the vegetation section, it is difficult to ascertain whether the taxon was discovered by him or only by others prior to his research.
Thirty years after Nannizzi's publication, Lorenzoni and Chiesura Lorenzoni [163] reported the presence and diffusion of some syntaxa in a preliminary note on ongoing research. However, the full study was never subsequently published. They mentioned: 1) Lemnetum minoris (Rubel 1930) Pignatti 1953, as very common; 2) Potameto-Utricularietum Br.-Bl. 1951, dominated by Utricularia vulgaris accompanied by various species of Potamogeton, as frequent in shallow waters; 3) Potameto-Vallisnerietum Br.-Bl. 1931, dominated by Myriophyllum spicatum, Potamogeton natans, Vallisneria spiralis, together with Potamogeton crispus, Spirodela polyrrhiza, as not very frequent; 4) Myriophylleto-Nupharetum Koch 1926, as the most common and widespread association, where Nymphoides peltata, Myriophyllum verticillatum, etc. predominate; 5) three associations of Phragmitetea, i.e. Typhetum angustifoliae (Allorge 1921) Pign. 1953, with both Typha angustifolia and T. latifolia, Phragmitetum communis (Allorge 1921) Pign. 1953 and Magnocaricetum elatae Koch 1926, without reporting their frequency. Regardless of the names they used for the associations, which are incorrect today, it is clear that the vegetation situation was very different from the current one. Lorenzoni & Chiesura Lorenzoni [163] state that, overall, the vegetation at the time of their study corresponds to that of 1932–1933. However, the reciprocal relationship between the communities has changed. Reclamation work involving the almost total elimination of marshy areas and the stabilisation of canal banks determined a clear separation between aquatic and terrestrial hygrophilous vegetation. This allowed anthropised vegetation containing alien species (e.g. Paspalum distichum, Amorpha fruticosa and Bidens frondosa), which were already present in 1932–1933, to become more widespread by 1968.
Subsequently Garbari [33] published a work on the flora and vegetation of Padule, presenting data collected between 1975-1977. This study also reported Nannizzi's species and observed qualitative and quantitative changes in the flora. Regarding the vegetation, Garbari [33] only conducted nine relevés, six of which were related to aquatic and marsh vegetation. These plots are large (80–400 m²), rather heterogeneous, and not in line with current trends of smaller dimensions and more homogeneous communities. However, they provide an overview of the vegetated landscape. His survey includes a reed bed with Phragmites australis; a community with Typha latifolia and several other marsh and aquatic species (Potamogeton sp. pl.); a community of hydrophytes with Lemna gibba, L. minor and Wolffia arrhiza; a community with Glyceria fluitans and Azolla filiculoides (sub A. caroliniana, see Lastrucci et al. [164]) with Nymphaea alba and Hydrocharis morsus-ranae; a sedge bed with Carex elata and others Carex; a community dominated by Oenanthe aquatica. with several other aquatic and marsh species. Garbari [33] concludes that, at the time of his study, the aquatic and marsh vegetation of the Padule di Fucecchio was dominated everywhere by two main aspects. One of these was attributable to Phragmitetea, with Pragmitetum communis having replaced Carex elata communities in most of the territory, although these were still present in residual strips. Another aspect was the vegetation in ditches and canals, which was dominated by Typha, Oenanthe, Sparganium, Alisma and Sagittaria, among others.
The observed plant landscape is fundamentally different, particularly with regard to the aquatic vegetation, but also with regard to the marsh communities. Almost all of the aquatic vegetation has disappeared from the Padule di Fucecchio, with only a few small patches remaining, mainly in ditches and canals with clearer water, or in unusual habitats such as small puddles along paths in the nearby forests. The most emblematic hydrophytes (Nymphaea alba, Hydrocharis morsus-ranae, Utricularia spp., various Potamogeton species, etc.) have apparently disappeared. Marsh vegetation is still dominated by Phragmites australis, but several marsh plants seem to have disappeared completely (e.g. Sagittaria sagittifolia) while others have decreased in number and spread (e.g. Oenanthe aquatica). Some species that play an important role today, such as Xanthium orientale and many other alien species (e.g. Eclipta prostrata, Lindernia dubia, etc.), were not even included in the floristic list by Garbari [33].
Why has the plant landscape changed so much? It is probably the result of a combination of factors. Following the Second World War, agricultural changes occurred, with a transition from an agricultural economy (the production of vegetable material for covering flasks and demijohns, as well as straw for chairs, was important in the area) and fishing, to a manufacturing-based economy. This led to highly polluted waters in the marsh. The accidental arrival of several alien plant and animal species, particularly the Louisiana crayfish (Procambarus clarkii) and nutria (Myocastor coypus) which spread to the area in the late 1990s [40]. From an avifaunistic point of view, the crayfish has been a valuable resource for many marsh birds of conservation interest. However, as has already been observed in other wetlands, the increasing presence of these invasive species has led to the decline and subsequent destruction of numerous aquatic and marshy plant communities [165,166]. The application to the whole area of conservation rules designed specifically for birdlife, which hindered the owners from continuing (at least partly) a traditional management of the territory, a practice which had in any case supported a certain biodiversity (both zoological and botanical) over time.
4. Conclusions
This study has enabled the current aquatic and marsh vegetation of the Padule di Fucecchio to be censused in unprecedented detail. It also reports the presence of community types that were previously unknown in the area, some of which are completely new and are described here for the first time. However, comparison with previous data has also enabled us to reasonably conclude that crucial changes to the vegetation structure have occurred in recent decades: hydrophyte communities have almost completely disappeared, and many sensitive marsh plant species have become rare or disappeared, being mostly replaced by more resilient native and invasive species. In addition to human disturbance and water pollution, the new vegetation arrangements were probably favoured by the arrival of alien fauna species that have had a profoundly negative impact on the state and dynamics of the vegetation. However, the avifaunal value of the area has remained substantially unchanged over time and, in some cases, has even improved [167]. For these reasons, aquatic and marsh vegetation types should be considered, like other types of plant communities of conservation interest, semi-natural coenoses and should be actively managed. However, it should be kept in mind that management that favours the protection of certain ecosystem components (e.g. birdlife) may not be adequate for the conservation of others (e.g. aquatic and marsh plants and vegetation). In a territory such as the Padule di Fucecchio, where it is possible to do so, areas with differentiated management should be created in agreement with local stakeholders, with the aim of conserving (and sometimes restoring) the various natural emergencies, both floristic and faunal.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Tables, divided in: Table S0: General Legend; Table S1: Aquatic plant communities; Table S2: Palustrine plant communities of Echinochloo cruris-galli-Sporoboletum schoenoidis association; Table S3: Palustrine plant communities of Littorellion uniflorae alliance; Table S4: Palustrine plant communities of Bidention tripartitae alliance; Table S5: Palustrine plant communities of Chenopodion rubri alliance; Table S6: Palustrine plant communities of Phragmition australis alliance; Table S7: Palustrine plant communities of Magnocaricion elatae and Magnocaricion gracilis alliances; Table S8: Palustrine plant communities of Glycerio-Sparganietum neglecti association; Table S9: Palustrine plant communities of Eleocharito palustris-Sagittarion sagittifoliae alliance; Table S10: Palustrine plant communities of Polypogono viridis-Paspaletum distichi association; Table S11: Palustrine plant communities of Frangulo alni-Salicetum cinereae association.
Author Contributions
Conceptualization, investigation, data curation, methodology, formal analysis, validation, writing—original draft preparation, writing—review and editing, L.L. and D.V.; All authors have read and agreed to the published version of the manuscript.
Funding
Daniele Viciani acknowledges the support of the National Biodiversity Future Center to University of Florence (Italy), funded by the Italian Ministry of University and Research, PNRR, Missione 4 Componente 2, Investimento 1.4, Project CN00000033.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Acknowledgments
We would like to thank Lorella Dell’Olmo for preparing Figures 1 and 2. We would also like to thank our friends Varaldo Macchi and Giovanni Riccioni, who have in-depth knowledge of the Fucecchio marshes. They accompanied us on several excursions and provided us with valuable information.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Syntaxonomic Scheme
| Lemnetea O. de Bolòs et Masclans 1955 | |
| Lemnetalia minoris O. de Bolòs et Masclans 1955 | |
| Lemnion minoris O. de Bolòs et Masclans 1955 | |
| Lemnetum minuto-gibbae Liberman Cruz, Pedrotti et Venanzoni 1988 | |
| Potamogetonetea Klika in Klika et Novák 1941 | |
| Potamogetonetalia Koch 1926 | |
| Potamogetonion Libbert 1931 | |
| Persicarietum amphibiae Pohjala 1933 nom. mut. Prunier et al. 2019 | |
| Potamo pectinati-Myriophylletum spicati Rivas Goday 1964 corr. Conesa 1990 | |
| Potamogetonetum denso-nodosi O. de Bolòs 1957 | |
| Callitricho hamulatae-Ranunculetalia aquatilis Passarge ex Theurillat in Theurillat et al. 2015 | |
| Ranunculion aquatilis Passarge ex Theurillat in Theurillat et al. 2015 | Callitriche platycarpa community |
| Callitrichetum stagnalis Kaiser 1926 | |
| Callitriche brutia communities | |
| Potamo crispi-Ranunculetum trichophylli Imchenetzky 1926 | |
| Isoëto-Nanojuncetea Br.-Bl. et Tx. in Br.-Bl. et al. 1952 | |
| Nanocyperetalia Klika 1935 | |
| Verbenion supinae Slavnić 1951 | |
| Echinochloo cruris-galli-Sporoboletum schoenoidis Paradis & Lorenzoni 1994 nom. mut. nov. | |
| Littorelletea uniflorae Br.-Bl. et Tx. ex Westhoff, Dijk & Passchier 1946 | |
| Littorelletalia uniflorae Koch ex Tx. 1937 | |
| Littorellion uniflorae Koch ex Klika 1935 | |
| Ranunculo flammulae-Juncetum bulbosi Oberdorfer 1957 | |
| Juncus bulbosus - Gratiola officinalis communities | |
| Bidentetea Tx., Lohmeyer et Preising ex von Rochow 1951 | |
| Bidentetalia Br.-Bl. et Tx. ex Klika et Hadač 1944 | |
| Bidention tripartitae Nordhagen ex Klika et Hadač 1944 | |
| Cyperetum micheliano-difformis ass. nova | |
| Bidens frondosa communities | |
| Veronica peregrina communities | |
| Portulaca oleracea communities | |
| Chenopodion rubri (Tx. in Poli et J. Tx. 1960) Hilbig et Jage 1972 | |
| Polygono lapathifolii-Xanthietum italici Pirola et Rossetti 1974 | |
| Echinochloo-Polygonetum lapathifolii Soó et Csurös 1947 | |
| Phragmito-Magnocaricetea Klika in Klika et Novák 1941 | |
| Phragmitetalia Koch 1926 | |
| Phragmition australis Koch 1926 nom. corr. | |
| Phragmitetum australis Savič 1926 nom. corr. | |
| Typhetum latifoliae Nowiński 1930 | |
| Typhetum angustifoliae Pignatti 1953 | |
| Schoenoplectetum lacustris Chouard 1924 nom. mut. Lastrucci et al. 2023 | |
| Phalarido arundinaceae-Bolboschoenetum laticarpi Passarge 1999 corr. Krumbiegel 2006 | |
| Magnocaricetalia Pignatti 1953 | |
| Magnocaricion elatae Koch 1926 | |
| Cyperetum longi Micevski 1957 | |
| Magnocaricion gracilis Géhu 1961 | |
| Caricetum ripariae Máthé et Kovács 1959 | |
| Caricetum vesicariae Chouard 1924 | |
| Phalaridetum arundinaceae Libbert 1931 | |
| Leucojum aestivum communities | |
| Nasturtio-Glycerietalia Pignatti 1953 | |
| Glycerio-Sparganion Br.-Bl. et Sissingh in Boer 1942 | |
| Glycerio-Sparganietum neglecti Koch 1926 | |
| Oenanthetalia aquaticae Hejný ex Balátová-Tuláčková et al. 1993 | |
| Eleocharito palustris-Sagittarion sagittifoliae Passarge 1964 | |
| Eleocharitetum palustris Savič 1926 | |
| Oenantho aquaticae-Rorippetum amphibiae Lohmeyer 1950 | |
| Bolboschoenetum glauci Grechushkina, Sorokin & Golub 2011 | |
| Butometum umbellati Philippi 1973 | |
| Molinio-Arrhenatheretea Tx. 1937 | |
| Holoschoenetalia Br.-Bl. ex Tchou 1948 | |
| Polypogono viridis-Paspalion distichi Br.-Bl. in Br.-Bl., Roussine & Nègre 1952 nom. mut. nov. | |
| Polypogono viridis-Paspaletum distichi Br.-Bl. in Br.-Bl., Gajewski, Wraber & Walas 1936 nom. mut. nov. | |
| Franguletea Doing ex Westhoff in Westhoff et Den Held 1969 | |
| Salicetalia auritae Doing 1962 | |
| Salicion cinereae T. Müller et Görs ex Passarge 1961 | |
| Frangulo alni-Salicetum cinereae Graebner et Hueck 1931 | |
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Acreman, M.; Hughes, K.A.; Arthington, A.H.; Tickner, D.; Dueñas, M.A. Protected areas and freshwater biodiversity: A novel systematic review distils eight lessons for effective conservation. Conserv. Lett. 2020, 13, e12684. [Google Scholar] [CrossRef]
- Albert, J.S.; Destouni, G.; Duke-Sylvester, S.M.; Magurran, A.E.; Oberdorff, T.; Reis, R.E.; Winemiller, K.O.; Ripple, W.J. Scientists’ warning to humanity on the freshwater biodiversity crisis. Ambio 2021, 50, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Fois, M.; Cuena-Lombraña, A.; Artufel, M.; Attard, V.; Cambria, S.; Farrugia, K.; Marignani, M.; Minissale, P.; Pace, A.; Sciandrello, S.; et al. Plant distribution and conservation in mediterranean islands’ lentic wetlands: there is more than aquatic species. Biodivers. Conserv. 2024, 2024 33, 1373–1392. [Google Scholar] [CrossRef]
- De Groot, R.; Brander, L.; Van Der Ploeg, S.; Costanza, R.; Bernard, F.; Braat, L.; Christie, M.; Crossman, N.; Ghermandi, A.; Hein, L.; et al. Global estimates of the value of ecosystems and their services in monetary units. Ecosyst Serv. 2012, 1, 50–61. [Google Scholar] [CrossRef]
- Zhang, L.L.; Yin, J.X.; Jiang, Y.Z.; Wang, H. Relationship between the hydrological conditions and the distribution of vegetation communities within the Poyang lake national nature reserve, China. Ecol. Inform. 2012, 11, 65–75. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Bernal, B.; Hernandez, M.E. Ecosystem services of wetlands. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2015, 11, 1–4. [Google Scholar] [CrossRef]
- Gardner, R.C.; Barchiesi, S.; Beltrame, C.; Finlayson, C.M.; Galewski, T.; Harrison, I.; Paganini, M.; Perennou, C.; Pritchard, D.E.; Rosenqvist, A.; et al. State of the World’s Wetlands and Their Services to People: A Compilation of Recent Analyses. Ramsar Briefing Note No. 7; Ramsar Convention Secretariat: Gland, Switzerland, 2015. [Google Scholar]
- Li, L.; Xu, H.; Zhang, Q.; Zhan, Z.; Liang, X.; Xing, J. Estimation methods of wetland carbon sink and factors influencing wetland carbon cycle: a review. Carbon Res. 2024 3, 50. [CrossRef]
- Hrivnák, R.; Kochjarová, J.; Oťaheľová, H.; Paľove-Balang, P.; Slezák, M.; Slezák, P. Environmental drivers of macrophyte species richness in artificial and natural aquatic water bodies–comparative approach from two central European regions. Ann. Limnol.-Int. J. Limnol. 2014, 50, 269–278. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Bacchetta, G. Knowledge gaps and challenges for conservation of Mediterranean wetlands: Evidence from a comprehensive inventory and literature analysis for Sardinia. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 2621–2631. [Google Scholar] [CrossRef]
- Ballut-Dajud, G.A.; Sandoval Herazo, L.C.; Fernández-Lambert, G.; Marín-Muñiz, J.L.; López Méndez, M.C.; Betanzo-Torres, E.A. Factors affecting wetland loss: A review. Land; 2022, 11, 434. [Google Scholar] [CrossRef]
- Xi, Y.; Peng, S.; Ciais, P.; Chen, Y. Future impacts of climate change on inland Ramsar wetlands. Nat. Clim. Chang. 2021, 2021 11, 45–51. [Google Scholar] [CrossRef]
- Leberger, R.; Geijzendorffer, I.R.; Gaget, E.; Gwelmami, A.; Galewski, T.; Pereira, H.M.; Guerra, C.A. Mediterranean wetland conservation in the context of climate and land cover change. Reg. Environ. Change 2020, 20, 67. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed]
- Fluet-Chouinard, E.; Stocker, B.D.; Zhang, Z.; Malhotra, A.; Melton, J.R.; Poulter, B.; Kaplan, J.O.; Goldewijk, K.K.; Siebert, S.; Minayeva, T.; et al. Extensive global wetland loss over the past three centuries. Nature 2023, 614, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Weng, B.; Yan, D.; Wang, K.; Li, X.; Bi, W.; Li, M.; Cheng, X.; Liu, Y. Wetlands of international importance: Status, threats and future protection. Int. J. Environ. Res. Public Health 2019, 16, 1818. [Google Scholar] [CrossRef]
- Janssen, J.A.M.; Rodwell, J.S.; Garcia Criado, M.; Gubbay, S.; Haynes, T.; Nieto, A.; Sanders, N.; Landucci, F.; Loidi, J.; Symank, A.; et al. European Red List of Habitats; Publications Office of European Union: Luxembourg, 2016; ISBN 978-92-79-61588-7. [Google Scholar]
- Zivkovic, L.; Biondi, E.; Pesaresi, S.; Lasen, C.; Spampinato, G.; Angelini, P. The third report on the conservation status of habitats (Directive 92/43/EEC) in Italy: Processes, methodologies, results and comments. Plant Sociol. 2017, 54, 51–64. [Google Scholar]
- Gigante, D.; Acosta, A.T.R.; Agrillo, E.; Armiraglio, S.; Assini, S.; Attorre, F.; Bagella, S.; Buffa, G.; Casella, L.; Giancola, C.; et al. Habitat conservation in Italy: The state of the art in the light of the first European Red List of Terrestrial and Freshwater Habitats. Rend. Lincei. Sci. Fis. e Nat. 2018, 29, 251–265. [Google Scholar] [CrossRef]
- Lazzaro, L.; Bolpagni, R.; Buffa, G.; Gentili, R.; Lonati, M.; Stinca, A.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; et al. Impact of invasive alien plants on native plant communities and Natura 2000 Habitats: State of the art, gap analysis and perspectives in Italy. J. Environ. Manag. 2020, 274, 111140. [Google Scholar] [CrossRef]
- Viciani, D.; Vidali, M.; Gigante, D.; Bolpagni, R.; Villani, M.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; Angiolini, C.; et al. A first checklist of the alien-dominated vegetation in Italy. Plant Sociol. 2020, 57, 29–54. [Google Scholar] [CrossRef]
- Gennai, M.; Gabellini, A.; Viciani, D.; Venanzoni, R.; Dell’Olmo, L.; Giunti, M.; Lucchesi, F.; Monacci, F.; Mugnai, M.; Foggi, B. The floodplain woods of Tuscany: Towards a phytosociological synthesis. Plant Sociol. 2021, 58, 1–28. [Google Scholar] [CrossRef]
- European Commission. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–50. [Google Scholar]
- Benavent-González, A.; Lumbreras, A.; Molina, J.A. Plant communities as a tool for setting priorities in biodiversity conservation: A novel approach to Iberian aquatic vegetation. Biod. Conserv. 2014, 23, 2135–2154. [Google Scholar] [CrossRef]
- Lastrucci, L.; Bonari, G.; Angiolini, C.; Casini, F.; Giallonardo, T.; Gigante, D.; Landi, M.; Landucci, F.; Venanzoni, R.; Viciani, D. Vegetation of Lakes Chiusi and Montepulciano (Siena, central Italy): Updated knowledge and new discoveries. Plant Sociol. 2014, 51, 29–55. [Google Scholar]
- Lastrucci, L.; Cerri, M.; Coppi, A.; Ferranti, F.; Ferri, V.; Foggi, B.; Lazzaro, L.; Reale, L.; Venanzoni, R.; Viciani, D.; et al. Understanding common reed die-back: A phytocoenotic approach to explore the decline of palustrine ecosystems. Plant Sociol. 2017, 54 (Suppl. S1), 15–28. [Google Scholar]
- Lastrucci, L.; Cerri, M.; Coppi, A.; Dell’Olmo, L.; Ferranti, F.; Ferri, V.; Filipponi, F.; Foggi, B.; Galardini, R.; Reale, L.; et al. Spatial landscape patterns and trends of declining reed-beds in peninsular Italy. Plant Biosyst. 2019, 2019 153, 427–435. [Google Scholar] [CrossRef]
- Angiolini, C.; Viciani, D.; Bonari, G.; Lastrucci, L. Habitat conservation prioritization: A floristic approach applied to a Mediterranean wetland network. Plant Biosyst. 2017, 151, 598–612. [Google Scholar] [CrossRef]
- Casavecchia, S.; Allegrezza, M.; Angiolini, C.; Biondi, E.; Bonini, F.; Del Vico, E.; Fanfarillo, E.; Foggi, B.; Gigante, D.; Gianguzzi, L.; et al. Proposals for improvement of Annex I of Directive 92/43/EEC: Central Italy. Plant Sociol. 2021, 58, 99–118. [Google Scholar] [CrossRef]
- Bartolini, A. La Riserva Naturale del Padule di Fucecchio. Dieci anni di gestione (1996-2006). Quad. del Padule di Fucecchio 2007, 4, 1–165. [Google Scholar]
- Puglisi, L.; Arcamone, E.; Franchini, M.; Giunchi, D.; Meschini, E.; Sacchetti, A.; Vanni, L.; Vezzani, A. Atlante degli uccelli nidificanti e svernanti in Toscana. 2. Distribuzione, abbondanza e conservazione. Centro Ornitologico Toscano, Regione Toscana, ETS Edizioni: Pisa, Italy, 2024.
- Garbari, F. Indagine floristica e vegetazionale sul padule di Fucecchio. In Progetto pilota per la salvaguardia e la valorizzazione del padule di Fucecchio; Min. Agr. For., Dir. Gen. Econ. Mont. For., Cons. Bon. Pad. Fucecchio: Pisa, Italy, 1980; pp. 217–263. [Google Scholar]
- Tomei, P.E.; Guazzi, E.; Kugler, P.C. Le Zone Umide della Toscana: Indagine Sulle Componenti Floristiche e Vegetazionali; Regione Toscana: Firenze, Italy, 2001; p. 167. [Google Scholar]
- Lastrucci, L.; Selvi, F.; Coppi, A.; Viciani, D. Ricerche botaniche del Museo di Storia Naturale dell'Università di Firenze sulle aree umide della Toscana. Notiz. Soc. Bot. Ital. 2024, 2024 6, 112–113. [Google Scholar]
- Padule di Fucecchio 2025. Available online: https://it.wikipedia.org/wiki/Padule_di_Fucecchio (accessed on 15 May 2025).
- Chetoni, R. Geologia; idrologia ed evoluzione paleogeografica del Padule di, Fucecchio, Dani, F.R., Eds.; Centro Ricerca, Documentazione e Promozione Padule di Fucecchio: Castelmartini (PT), Italy, 1999; pp. 41–51. [Google Scholar]
- Corsi, R. Flora e vegetazione nell’area del Padule di Fucecchio, In Il Padule di Fucecchio e il Lago di Sibolla. Natura e storia; Dani, F.R., Ed.; Centro Ricerca, Documentazione e Promozione Padule di Fucecchio: Castelmartini (PT), Italy, 1999; pp. 53–74. [Google Scholar]
- Malvolti, A. Il Padule nella storia. In Il Padule di Fucecchio e il Lago di Sibolla. Natura e storia. In Il Padule di Fucecchio e il Lago di Sibolla. Natura e storia; Dani, F.R., Ed.; Centro Ricerca, Documentazione e Promozione Padule di Fucecchio: Castelmartini (PT), Italy, 1999; pp. 21–40. [Google Scholar]
- Bartolini, A. CASTORE - Cartografia storica regionale. Carta topografica della Pianura del Valdarno di Sotto con i confini delle respettive Comunità dedotta dall’originale autentico eseguito nell’anno 1747(a cura di). Studio per la salvaguardia della biodiversità del S.I.R. Padule di Fucecchio; Centro Ricerca, Documentazione e Promozione Padule di Fucecchio: Castelmartini (PT), Italy, 2004. [Google Scholar]
- Regione Toscana. 2025. Available online: https://www502.regione.toscana.it/cartografia_storica_regionale_gi/cartografia_storica_regionale_smartviewer.html?img=11191 (accessed on 15 April 2025).
- Stazione meteorologica di Larciano Castelmartini. Available online: https://it.wikipedia.org/wiki/Stazione_meteorologica_di_Larciano_Castelmartini (accessed on 15 May 2025).
- Bigi, L.; Rustici, L. Regime idrico dei suoli e tipi climatici in Toscana. Regione Toscana; Dipart. Agric. For.: Firenze, Italy, 1984. [Google Scholar]
- Thornthwaite, C.W.; Mather, J.R. Instruction and tables for computing potential evapotraspiration and the water balance. Pubbl. Climatol. 1957, 10, 1–311. [Google Scholar]
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Carmignani, L.; Conti, P.; Cornamusini, G.; Pirro, A. Geological map of Tuscany (Italy). J. Maps 2013, 9, 487–497. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Plant Sociology: The Study of Plant Communities; McGraw-Hill: New York, NY, USA, 1932. [Google Scholar]
- 48 Braun-Blanquet, J. Pflanzensoziologie. Grundzuge der Vegetationskunde, 3rd ed.; Springer: Wien, Austria, 1964; pp. 1–330. [Google Scholar]
- Dengler, J.; Berg, C.; Jansen, F. New ideas for modern phytosociological monographs. Ann. Bot. 2005, 5, 193–210. [Google Scholar]
- Dengler, J.; Chytrý, M.; Ewald, J. Phytosociology. In Encyclopedia of Ecology; Jorgensen, S.E., Fath, B.D., Eds.; Elsevier: Oxford, UK, 2008; pp. 2767–2779. [Google Scholar]
- Biondi, E. Phytosociology today: Methodological and conceptual evolution. Plant Biosyst. 2011, 145 (Suppl. S1), 19–29. [Google Scholar] [CrossRef]
- Pott, R. Phytosociology: A modern geobotanical method. Plant Biosyst. 2011, 145 (Suppl. S1), 9–18. [Google Scholar] [CrossRef]
- Van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar]
- 54 Arrigoni, P.V. A classification of plant growth forms applicable to the floras and vegetation types of Italy. Webbia 1996, 50, 193–203. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org/ (accessed on 15 January 2025).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Ouellette, M.-H.; Cunha, E.R.; Smith, T.; Stier, A.; et al.; Vegan: Community Ecology Package R Package Version 2.6-6.1. 2024. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 January 2025).
- Portal to the Flora of Italy 2025. Available online: http://dryades.units.it/floritaly (accessed on 12 February 2025).
- Mucina, L.; Bultmann, H.; Diersen, K.; Theurillat, J.P.; Raus, T.; Carni, A.; Šumberova, K.; Willner, W.; Dengler, J.; Garcia, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19 (Suppl. S1), 3–264. [Google Scholar] [CrossRef]
- Chytrý, M.; Řezníčková, M.; Novotný, P.; Holubová, D.; Preislerová, Z.; Attorre, F.; Biurrun, I.; Blažek, P.; Bonari, G.; Borovyk, D.; et al. FloraVeg.EU – an online database of European vegetation, habitats and flora. Appl. Veg. Sci. 2024, 2024 27, e12798. [Google Scholar] [CrossRef]
- Biondi, E.; Blasi, C. Prodromo della Vegetazione Italiana. 2015. Available online: http://www.prodromo-vegetazione-italia.org/ (accessed on 12 February 2025).
- Theurillat, J.P.; Willner, W.; Fernandez-Gonzalez, F.; Bultmann, H.; Carni, A.; Gigante, D.; Mucina, L.; Weber, H. International Code of Phytosociological Nomenclature. 4th edition. Appl. Veg. Sci. 2021, 24, e12491. [Google Scholar] [CrossRef]
- Liberman Cruz, M.; Pedrotti, F.; Venanzoni, R. Le associazioni della classe Lemnetea del lago Titicaca (Bolivia). Riv. Idrobiol. 1988, 2(2-3), 377–388. [Google Scholar]
- Landucci, F.; Gigante, D.; Venanzoni, R. An application of the Cocktail method for the classification of the hydrophytic vegetation at Lake Trasimeno (Central Italy). Fitosociologia 2011, 48, 3–22. [Google Scholar]
- Lastrucci, L.; Valentini, E.; Dell’Olmo, L.; Vietina, B.; Foggi, B. Hygrophilous vegetation and habitats of conservation interest in the area of the Lake Porta (Tuscany, Central Italy). Atti Soc. Tosc. Sci. Nat. Mem. Serie B 2016, 2016 122, 131–146. [Google Scholar]
- Lastrucci, L.; Dell’Olmo, L.; Foggi, B.; Massi, L.; Nuccio, C.; Vicenti, C.; Viciani, D. Contribution to the knowledge of the vegetation of the Lake Massaciuccoli (northern Tuscany, Italy). Plant Sociol. 2017, 2017 54, 67–87. [Google Scholar] [CrossRef]
- Bramley, J.L.; Reeve, J.T.; Dussart, B.J. The distribution of Lemna minuta within the British Isles: identification; dispersal and niche constraints. In Plant Invasions. General Aspects and Special, Problems, Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; SPB Academic Publishing: Amsterdam, The Netherlands, 1995; pp. 181–185. [Google Scholar]
- Prunier, P.; Greulich, F.; Béguin, C.; Boissezon, A.; Delarze, R.; Hegg, O.; Klötlzi, F.; Pantke, R.; Steffen, J.; Steiger, P.; Vittoz, P.; Phytosuisse: un référentiel pour les associations végétales de Suisse. ISSN 2624-6171. Available online: www.infoflora.ch/it/ambienti/phytosuisse- (accessed on day month year).
- Kłosowski, S.; Tomaszewicz, H. Habitat requirements of Polygonetum natantis Soó 1927 and Potamogetonetum natantis Soó 1927 phytocenoses in north-eastern Poland. Acta Soc. Bot. Pol. 1986, 1986 55, 141–157. [Google Scholar] [CrossRef]
- Lastrucci, L.; Landi, M.; Angiolini, C. Vegetation analysis on wetlands in a Tuscan agricultural landscape (central Italy). Biologia 2010, 2010 65, 54–68. [Google Scholar] [CrossRef]
- Šumberová, K. Vegetace vodních rostlin zakorenenych ve dne (Potametea). Vegetation of aquatic plants rooted in the bottom. In Vegetace České republiky 3. Vodní a mokřadní vegetace [Vegetation of the Czech Republic 3. Aquatic and wetland vegetation]; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2011; pp. 100–247. [Google Scholar]
- Loidi, J.A.; Biurrun, I.G.; Herrera, M.G. La vegetación del centro-septentrional de España. Itinera Geobot. 1997, 1997 9, 161–618. [Google Scholar]
- Ceschin, S.; Zuccarello, V.; Caneva, G. Role of macrophyte communities as bioindicators of water quality Application on the Tiber River basin (Italy). Plant Biosyst. 2010, 144, 528–536. [Google Scholar] [CrossRef]
- Zervas, D.; Tsiripidis, I.; Bergmeier, E.; Tsiaoussi, V. A phytosociological survey of aquatic vegetation in the main freshwater lakes of Greece. Veg. Classific. Survey 2020, 2020 1, 53–75. [Google Scholar] [CrossRef]
- Viciani, D.; Angiolini, C.; Bonari, G.; Bottacci, A.; Dell’Olmo, L.; Gonnelli, V.; Zoccola, A.; Lastrucci, L. Contribution to the knowledge of aquatic vegetation of montane and submontane areas of Northern Apennines (Italy). Plant Sociol. 2022, 59, 25–35. [Google Scholar] [CrossRef]
- Hrivnák, R. Aquatic plant communities in the catchment area of the Ipel' river in Slovakia and Hungary. Part II. Class Potametea. Thaiszia 2002, 12, 137–160. [Google Scholar]
- Lastrucci, L.; Landucci, F.; Gonnelli, V.; Barocco, R.; Foggi, B.; Venanzoni, R. The vegetation of the upper and middle River Tiber (Central Italy). Plant Sociol. 2012, 49, 29–48. [Google Scholar]
- Lastrucci, L.; Selvi, F.; Bajona, E.; Sforzi, A.; Siccardi, E.; Viciani, D. Advancing Knowledge of Wetland Vegetation for Plant Diversity Conservation: The Case of Small Lakes, Ponds, and Pools in Maremma (Southern Tuscany, Central Italy). Land 2025, 2025 14, 218. [Google Scholar] [CrossRef]
- Lastrucci, L.; Saiani, D.; Mugnai, A.; Ferretti, G.; Viciani, D. Distribution novelties of the genus Callitriche (Plantaginaceae) in Italy from the study of the Herbarium Centrale Italicum collections. Mediterr. Bot. 2024, 45, e87474. [Google Scholar] [CrossRef]
- Felzines, J.C. Contribution au prodrome des vegetations de France: Les Potametea Klika in Klika & V. Novak 1941. Doc. Phytosoc. 2016, 3, 218–437. [Google Scholar]
- Passarge, H. Mitteleuropaische Potamogetonetea I. Phytocoenologia 1992, 20, 489–527. [Google Scholar] [CrossRef]
- Ceschin, S.; Salerno, G. La vegetazione del basso corso del Fiume Tevere e dei suoi affluenti (Lazio; Italia). Fitosociologia 2008, 45, 39–74. [Google Scholar]
- Brullo, S.; Scelsi, F.; Spampinato, F. La Vegetazione dell’Aspromonte; Laruffa Editore: Reggio Calabria, Italy, 2001. [Google Scholar]
- Baldoni, M.; Biondi, E. La vegetazione del medio e basso corso del fiume Esino (Marche-Italia centrale). Studia Botanica 1993, 11, 209–257. [Google Scholar]
- Venanzoni, R.; Gigante, D. Contributo alla conoscenza della vegetazione degli ambienti umidi dell’Umbria (Italia). Fitosociologia 2000, 37, 13–63. [Google Scholar]
- Lastrucci, L.; Foggi, B.; Selvi, F.; Becattini, R. Contributo alla conoscenza della vegetazione e della flora delle aree umide nel comprensorio di Capalbio (Provincia di Grosseto, Italia centrale). Arch. Geobot. 2007, 10, 1–30. [Google Scholar]
- Gigante, D.; Maneli, F.; Venanzoni, R. Mediterranean temporary wet systems in inland Central Italy: Ecological and phytosociological features. Plant Sociol. 2013, 50, 93–112. [Google Scholar]
- Bagella, S.; Caria, M.C.; Farris, E.; Filigheddu, R. Phytosociological analysis in Sardinian Mediterranean temporary wet habitats. Fitosociologia 2009, 46, 11–26. [Google Scholar]
- Tomaselli, V.; Beccarisi, L.; Brullo, S.; Cambria, S.; Forte, L.; Minissale, P.; Veronico, G. Phytosociological research on temporary ponds in Apulia (southern Italy), Medit. Bot. 2020, 2020 41, 15–41. [Google Scholar]
- Tomaselli, V.; Beccarisi, L.; Cambria, S.; Forte, L.; Minissale, P.; Sciandrello, S.; Veronico, G.; Brullo, S. Validation of associations for the temporary ponds of the class Isoeto-Nanojuncetea in Puglia (southern Italy), Medit. Bot. 2022, 2022 43, e80627. [Google Scholar]
- Brullo, S.; Brullo, C.; Sciandrello, S.; Tavilla, G.; Cambria, S.; Tomaselli, V.; Ilardi, V.; Giusso del Galdo, G.; Minissale, P. The plant communities of the class Isoëto-Nanojuncetea in Sicily. Plants 2022, 11, 1214. [Google Scholar] [CrossRef]
- Pedrotti, F.; Russo, G.; Murrja, E. Flora, plant communities and vegetation mapping of the lake d'Otri (Gargano, Apulia, Italy). Contribuţii Botanice 2022, LVII, 53–62. [Google Scholar] [CrossRef]
- Brullo, S.; Minissale, P. Considerazioni syntassonomiche sulla classe Isoeto-Nanojuncetea. Itinera Geobot. 1998, 11, 263–290. [Google Scholar]
- Eliáš, P.j.; Dítě, D.; Grulich, V.; Sádovský, M. Distribution and communities of Crypsis aculeata and Heleochloa schoenoides in Slovakia. Hacquetia 2008, 2008 7, 5–20. [Google Scholar] [CrossRef]
- Eliáš, P.j.; Dítě, D.; Dítě, Z. Halophytic Vegetation in the Pannonian Basin: Origin, Syntaxonomy, Threat, and Conservation. In Handbook of Halophytes, Grigore, M.N., Ed.; Springer: Cham, Swiss, 2020. [Google Scholar] [CrossRef]
- Paradis, G. Observations synécologiques sur des stations corses de trois thérophytes fini-estivales: Crypsis aculeata, Crypsis schoenoides et Chenopodium chenopodioides. Monde des Plantes 1992, 444, 1–21. [Google Scholar]
- Paradis, G.; Lorenzoni, C. Etude phytosociologique de communautés thérophytiques hygro-nitrophiles estivo-autumnales de la Corse (grpts. à Crypsis aculeata, C. schoenoides, Glinus lotoides et Chenopodium chenopodioides). Nouvelles propositions syntaxonomiques. 2e contribution. Monde des Plantes 1994 449, 19-26.
- Martínez-Parras, J.M.; Lorca, M.P.; Esteban, C.B.; Mesa, J.M. Algunas comunidades vegetales higrófilas e higronitrófilas estivo-autumnales de la provincia de Granada. Acta Bot. Barcinon. 1988, 1988 37, 271–279. [Google Scholar]
- Rivas-Martinez, S.; Fernandez–Gonzalez, F.; Loidi, J.; Lous. , M.; Penas, A. Syntaxonomical Checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobot. 2001, 14, 5–341. [Google Scholar]
- Dawson, F.H.; Szoszkiewicz, K. Relationships of some ecological factors with the associations of vegetation in British rivers. Hydrobiologia 1999, 1999 415, 117–122. [Google Scholar] [CrossRef]
- Šumberová, K.; Navrátilová, J.; Čtvrtlíková, M.; Hájek, M.; Bauer, P. Vegetace oligotrofních vod (Littorelletea uniflorae). Vegetation of oligotrophic water bodies. In Vegetace České republiky 3. Vodní a mokřadní vegetace [Vegetation of the Czech Republic 3. Aquatic and wetland vegetation]; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2011; pp. 268–308. [Google Scholar]
- Ninot, J.M.; Carreras, J.; Carrillo, E.; Vigo, J. Syntaxonomic conspectus of the vegetation of Catalonia and Andorra. I: Hygrophilous herbaceous communities. Acta Bot. Barc. 2000, 46, 191–237. [Google Scholar]
- Szankowski, M.; Kłosowski, S. Habitat variability of the Littorelletea uniflorae plant communities in Polish Lobelia lakes. Hydrobiologia 2006 570, 117–126. [CrossRef]
- Hrivnák, R.; Kochjarová, J.; Ot’ahel’ová, H.; Pal’ove-Balang, P.; Slezák, M.; Slezák, P. Environmental drivers of macrophyte species richness in artificial and natural aquatic water bodies–comparative approach from two central European regions. Ann. Limnol.-Int. J. Limnol. 2014, 50, 269–278. [Google Scholar] [CrossRef]
- Pellizzari, M. Cyperus-dominated vegetation in the eastern Po River. Plant Sociol. 2020, 2020 57, 1–16. [Google Scholar] [CrossRef]
- Piccoli, F.; Gerdol, R. Rice-field weed communities in Ferrara Province (northern Italy). Aquatic Bot. 1981, 1981 10, 317–328. [Google Scholar] [CrossRef]
- Melečková, Z.; Dítě, D.; Eliáš, P.j.; Schmidt, D. Cyperus glomeratus L. rediscovered in Slovakia. Hacquetia 2016, 2016 15, 93–100. [Google Scholar] [CrossRef]
- Šumberová, K.; Lososová, Z. Vegetace jednoletých nitrofilních vlhkomilných bylin (Bidentetea tripartitae). Vegetation of annual nitrophilous wetland herbs. In Vegetace České republiky 3. Vodní a mokřadní vegetace [Vegetation of the Czech Republic 3. Aquatic and wetland vegetation]; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2011; pp. 347–384. [Google Scholar]
- Amigo, J. Los herbazales terofíticos higronitrófilos en el noroeste de la Península Ibérica (Bidentetea tripartitae Tüxen; Lohmeyer & Preising ex von Rochow 1951). Lazaroa 2006, 27, 43–58. [Google Scholar]
- Roma-Marzio, F.; Lastrucci, L.; Guzzon, F.; Ardenghi, N.M.G.; Peruzzi, L.; Mossini, S. Nuove segnalazioni floristiche italiane 5. Flora vascolare (28-46). Not. Soc. Bot. Ital. 2018, 2, 205–210. [Google Scholar]
- Maiorca, G.; Crisafulli, A.; Puntillo, D.; Signorino, G.; Spampinato, G. Wetland vegetation of the Tarsia Lake Regional Nature Reserve (Calabria, southern Italy). Medit. Bot. 2020, 2020 41, 67–84. [Google Scholar] [CrossRef]
- Passarge, H. Pflanzengesellschaften Norostdeutschland. J. Cramer: Berlin, Stuttgart, Germany, 1996.
- Julve, P.; Les base de données botaniques. Baseveg. Baseflor. Programme Catminat. 2025. Available online: https://www.tela-botanica.org/ressources/donnees/telechargements/ (accessed on 15 Mars 2025).
- Chytrý, M.; Tichý, L. Diagnostic, constant and dominant species of vegetation classes and alliances of the Czech Republic: a statistical revision. Folia Fac. Sci. Nat. Univ. Masaryk. Brun. Biol. 2003, 108, 1–231. [Google Scholar]
- Lososová, Z. (2009): Portulacetum oleraceae Felföldy 1942. – In Vegetace České republiky. 2. Ruderální, plevelová, skalní a suťová vegetace [Vegetation of the Czech Republic 2. Ruderal, weed, rock and scree vegetation]; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2009; pp. 198–200. [Google Scholar]
- Biondi, E.; Baldoni, M. La vegetazione del fiume Marecchia (Italia Centrale). Biogeographia 1994, 17, 51–87. [Google Scholar] [CrossRef]
- Biondi, E.; Vagge, I.; Baldoni, M.; Taffetani, F. Biodiversità fitocenotica e paesaggistica dei fiumi dell’Italia centro-settentrionale: aspetti fitosociologici e sinfitosociologici. Studi Trent. Sci. Nat., Acta Biol. 2004, 80, 13–21. [Google Scholar]
- Tomasello, S. How many names for a beloved genus? – Coalescent-based species delimitation in Xanthium L. (Ambrosiinae, Asteraceae). Molec. Phylogenet. Evol. 2018, 127, 135–145. [Google Scholar] [CrossRef]
- Geisselbrecht-Taferner, L.; Mucina, L. 1993. Bidentetea tripartiti. In Pflanzengesellschaften Österreich. Teil 1, Mucina, L.; Grabherr, G., Ellmauer, T., Eds.; Eds.; Gustav Fischer Verlag: Jena-Stuttgart-New York, 1993; pp. 90–109. [Google Scholar]
- Sanda, V.; Öllerer, K.; Burescu, P. Fitocenozele din România. Ars Docendi: Bucureşti, Romania, 2008; 570 p.
- Neacșu, A.; Arsene, G.G. Pioneer vegetation on water shores (Bidenti-Polygonetum hydropiperi Lohmeyer in R. Tüxen 1950 and Echinochloo-Polygonetum lapathifolii Soó et Csűrös 1974) in the Romanian Banat. Res. J. Agricult. Sci. 2017, 49, 122–135. [Google Scholar]
- Felzines, J.C.; Loiseau, J.E. Les groupements fluviatiles des Bidentetea de la Loire moyenne, du bas Allier et de la Dordogne moyenne. Modifications apportées à la synsystématique de la classe des Bidentetea. Bull. Soc. Bot. Centre-Ouest 2005, 36, 159–204. [Google Scholar]
- Landucci, F.; Gigante, D.; Venanzoni, R.; Chytr, M. Wetland vegetation of the class Phragmito-Magno-Caricetea in central Italy. Phytocoenologia 2013, 43, 67–100. [Google Scholar] [CrossRef]
- Landucci, F.; Šumberova, K.; Tichý, L.; Hennekens, S.; Aunina, L.; Biță-Nicolae, C.; Borsukevych, L.; Bobrov, A.; Čarni, A.; De Bie, E.; et al. Classification of the European marsh vegetation (Phragmito-Magnocaricetea) to the association level. Appl. Veg. Sci. 2020, 23, 297–316. [Google Scholar] [CrossRef]
- Lastrucci, L.; Paci, F.; Raffaelli, M. The wetland vegetation of the Natural Reserves and neighbouring stretches of the Arno river in the Arezzo province (Tuscany; Central Italy). Fitosociologia 2010, 47, 29–59. [Google Scholar]
- Lastrucci, L.; Angiolini, C.; Bonari, G.; Bottacci, A.; Gonnelli, V.; Zoccola, A.; Mugnai, M.; Viciani, D. Contribution to the knowledge of marsh vegetation of montane and submontane areas of Northern Apennines (Italy). Plant Sociol. 2023, 60, 25–36. [Google Scholar] [CrossRef]
- Lastrucci, L.; Lazzaro, L.; Coppi, A.; Foggi, B.; Ferranti, F.; Venanzoni, R.; Cerri, M.; Ferri, V.; Gigante, D.; Reale, L. Demographic and macro-morphological evidence for common reed dieback in central Italy. Plant Ecol. Divers. 2017, 10, 241–251. [Google Scholar] [CrossRef]
- Šumberová, K.; Hájková, P.; Chytrý, M.; Hroudová, Z.; Sádlo, J.; Hájek, M.; Hrivnák, R.; Navrátilová, J.; Hanáková, P.; Ekrt, L.; Ekrtová, E. Vegetace rákosin a vysokých ostřic (Phragmito-Magno-Caricetea). Marsh vegetation. In Vegetace České republiky 3. Vodní a mokřadní vegetace [Vegetation of the Czech Republic 3. Aquatic and wetland vegetation. In Vegetace České republiky 3. Vodní a mokřadní vegetace [Vegetation of the Czech Republic 3. Aquatic and wetland vegetation]; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2011; pp. 385–580. [Google Scholar]
- Hrivnák, R. The plant communities of Phragmitetalia in the catchment area of the Ipel’river (Slovakia and Hungary), 2, Tall-sedge dominated wetlands (Magnocaricion elatae). Biologia 2004, 2004 59, 457–476. [Google Scholar]
- Fernez, T.; Causse, G. Synopsis phytosociologique des groupements végétaux d’Île-de-France. Version 1 - avril 2015. Conservatoire Botanique National du Bassin Parisien - Muséum national d’Histoire naturelle, délégation Île-de-France, Direction régionale et interdépartementale de l’environnement et de l’énergie Île-de-France, 2015; 89 p.
- Lastrucci, L.; Foggi, B.; Mantarano, N.; Ferretti, G.; Calamassi, R.; Grigioni, A. La vegetazione del laghetto “Lo Stagnone” (Isola di Capraia; Toscana). Atti Soc. Tosc. Sci. Nat. Mem. Ser. B 2010, 116, 17–25. [Google Scholar]
- Mereu, L.; Lastrucci, L.; Viciani, D. Contributo alla conoscenza della vegetazione del fiume Pesa (Toscana, Italia Centrale). Studia Botanica 2012, 2012 29, 105–143. [Google Scholar]
- Poldini, L. La vegetazione del Carso isontino e triestino. Lint: Trieste, Italy, 1989.
- Lastrucci, L.; Ferretti, G.; Mantarano, N.; Foggi, B. Vegetation and habitat of conservation interest of the lake Acquato (Grosseto—Italy). Plant Sociol. 2019, 56, 19–30. [Google Scholar]
- Hroudová, Z.; Hrivnák, R.; Chytrý, M. Classification of inland Bolboschoenus-dominated vegetation in Central Europe. Phytocoenologia 2009, 2009 39, 205–215. [Google Scholar] [CrossRef]
- Di Natale, S.; Lastrucci, L.; Hroudova, Z.; Viciani, D. A review of Bolboschoenus species (Cyperaceae) in Italy based on herbarium data. Plant Biosyst. 2022, 156, 261–270. [Google Scholar] [CrossRef]
- Horvat, I.; Glavač, V.; Ellenberg, H. Vegetation Südosteuropas. G. Fischer Verlag: Stuttgart, Germany, 1974.
- Devillers, P.; Devillers-Terschuren, J. Application and development of the Palearctic habitat classification in the course of the setting up of the Emerald Project, Malta. Council of Europe, Strasbourg, France, 2001.
- Buchwald, R. Vegetazione e odonatofauna negli ambienti acquatici dell’Italia Centrale. Braun-Blanquetia 1994, 11, 1–77. [Google Scholar]
- Lastrucci, L.; Viciani, D.; Nuccio, C.; Melillo, C. Indagine vegetazionale su alcuni laghi di origine artificiale limitrofi al Padule di Fucecchio (Toscana, Italia Centrale). Ann. Mus. Civ. Rovereto 2008, 23, 169–203. [Google Scholar]
- Hahn, I.; Gergely, A.; Barabás, S. Changes in the active floodplain vegetation of the Szigetköz. Ann. Bot. Roma 2011, 1, 1–8. [Google Scholar]
- Poldini, L.; Vidali, M.; Castello, M.; Sburlino, G. A novel insight into the remnants of hygrophilous forests and scrubs of the Po Plain biogeographical transition area (Northern Italy). Plant Sociol. 2020, 2020 57, 17–69. [Google Scholar] [CrossRef]
- Postolache, G. Diversitatea cenotaxonomică a vegetației Republicii Moldova. Revista Botanică 2022, 25, 24–43. [Google Scholar]
- Lastrucci, L.; Gambirasio, V.; Prosser, F.; Viciani, D. First record of Sparganium oocarpum in Italy and new regional distribution data for S. erectum species complex. Plant Biosyst. 2024, 158, 595–600. [Google Scholar] [CrossRef]
- Lastrucci, L.; Lazzaro, L.; Lunardi, L.; Fiorini, G.; Viciani, D. Morphometric surveys on Eleocharis palustris (L.) Roem. et Schult. (Cyperaceae). The contribution of herbarium investigations to the delimitation of its subspecies and their distribution in Italy. Ann. Bot. Roma 2020, 10, 97–105. [Google Scholar] [CrossRef]
- Angiolini, C.; Viciani, D.; Bonari, G.; Zoccola, A.; Bottacci, A.; Ciampelli, P.; Gonnelli, V.; Lastrucci, L. Environmental drivers of plant assemblages: Are there differences between palustrine and lacustrine wetlands? A case study from the northern Apennines (Italy). Knowl. Manag. Aquat. Ecosyst. 2019, 420, 34. [Google Scholar] [CrossRef]
- Hroudová, Z.; Zákravský, P.; Frantík, T. Ecological differentiation of Central European Bolboschoenus taxa and their relationship to plant communities. Folia Geobot. 1999, 1999 34, 77–96. [Google Scholar] [CrossRef]
- Hroudová, Z.; Zákravský, P.; Ducháček, M.; Marhold, K. Taxonomy, distribution and ecology of Bolboschoenus in Europe. Ann. Bot. Fenn. 2007, 2007 44, 81–102. [Google Scholar]
- Grechushkina, N.A.; Sorokin, A.N.; Golub, V.B. The plant communities with domination of Phragmites australis and Bolboschoenus glaucus in the territory of Russian coast of the Azov Sea. Nauchnyje Soobschchestva 2011, 2011 20, 105–115. [Google Scholar]
- Prosser, F.; Sarzo, A. Flora e vegetazione di fossi nel settore trentino del fondovalle dell’Adige (Trentino-Italia settentrionale). Ann. Mus. civ. Rovereto. Sez.: Arch., St., Sci. Nat. 2003, 18, 89–144. [Google Scholar]
- Celesti-Grapow, L.; Alessandrini, A.; Arrigoni, P.V.; Banfi, E.; Bernardo, L.; Bovio, M.; et al. Inventory of the non-native flora of Italy. Plant Biosyst. 2009, 2009 143, 386–430. [Google Scholar] [CrossRef]
- Braun-Blanquet, J.; Gajewski, W.; Wraber, M.; Walas, J. Classe des Rudereto-Secalinetales. Groupements messicoles; culturaux et nitrophiles-rudérales du cercle de végétation méditerranéen. Prodr. Group. Vég. Montpellier 1936, 3, 1–37. [Google Scholar]
- Braun-Blanquet, J.; Roussine, N.; Nègre, R. Les Groupements Végétaux de la France Méditerranéenne. Centre National de la Recherche Scientifique: Montpellier, 1952.
- Douda, J. 1957. In Vegetace České republiky. 4. Lesní a křovinná vegetace [Vegetation of the Czech Republic 4. Forest and shrub vegetation]; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2013; pp. 180–182. [Google Scholar]
- Tomei, P.E.; Cenni, M. Il Bosco di Chiusi e la Paduletta di Ramone (PT): note floristiche e vegetazionali. Quad. Mus. Stor. Nat. Livorno 1986, 1986 7, 55–79. [Google Scholar]
- Arrigoni, P.V. Documenti per la carta della vegetazione delle Cerbaie (Toscana settentrionale). Parlatorea 1997, 2, 39–71. [Google Scholar]
- Corsi, R. vegetazione. In Lungo le rotte migratorie. Progetti di ricerca sulla vegetazione, l'avifauna e le specie aliene; Venturato, E., Petrini, R., Eds.; Centro Ricerca, Documentazione e Promozione Padule di Fucecchio: Castelmartini (PT), Italy, 2001; pp. 23–120. [Google Scholar]
- Franzese, M. Studio delle comunità vegetazionali ed aggiornamento cartografico della vegetazione 2004. Centro di Ricerca, Documentazione e Promozione del Padule di Fucecchio, 2004; 43 pp.
- Caruel, T. Prodromo della Flora Toscana. Le Monnier: Firenze, 1860–1864, 767 pp.
- Sandri, G.; Fantozzi, P. Contribuzione alla Flora della Val di Nievole. Nuovo Giorn. Bot. Ital., n.s. 1895, 2(1-4, 129–333. [Google Scholar]
- Baroni, E. Supplemento Generale al “Prodromo della Flora Toscana di T. Caruel”. Società Botanica Italiana: Firenze, 1897–1908, 633 pp.
- Pampanini, R. Gli esponenti più rimarchevoli e più rari della Flora toscana nel censimento dei Monumenti Naturali d'Italia. Nuovo Giorn. Bot. Ital., n.s. 925, 32, 5–35. [Google Scholar]
- Nannizzi, A. Ricerche idrobiologiche sul padule di Fucecchio. II. Fanerogame. Boll. Pesca, Piscicolt. Idrobiol. 1938, 14, 161–179. [Google Scholar]
- Lorenzoni, G.G.; Chiesura Lorenzoni, F. Osservazioni sull'evoluzione della vegetazione del Padule di Fucecchio (Toscana). Nuovo Giorn. Bot. Ital. 1968, 1968 101, 423–424. [Google Scholar]
- Lastrucci, L.; Fiorini, G.; Lunardi, L.; Viciani, D. Herbarium survey on the genus Azolla (Salviniaceae) in Italy: distributive and taxonomic implications. Plant Biosyst. 2019, 2019 153, 710–719. [Google Scholar] [CrossRef]
- Rodríguez, C.F.; Bécares, E.; Fernández-Aláez, M.; Fernández-Aláez, C. Loss of diversity and degradation of wetlands as a result of introducing exotic crayfish. Biol. Invasions 2005, 2005 7, 75–85. [Google Scholar] [CrossRef]
- Prigioni, C.; Balestrieri, A.; Remonti, L. Food habits of the coypu, Myocastor coypus, and its impact on aquatic vegetation in a freshwater habitat of NW Italy. Folia Zool. 2005, 2005 54, 269–277. [Google Scholar]
- Zarri, E.; Natura in Toscana. C.U.A. (Censimento Uccelli Acquatici e svernanti) Padule di Fucecchio 2025. Available online: https://www.naturaintoscana.it/c-u-a-padule-di-fucecchio-2025/ (accessed on 15 May 2025).
- Löbl, I.; Klausnitzer, B.; Hartmann, M.; Krell, F.T. The silent extinction of species and taxonomists—An appeal to science policymakers and legislators. Diversity 2023, 2023 15, 1053. [Google Scholar] [CrossRef]
- Eckert, I.; Bruneau, A.; Metsger, D.A.; Joly, S.; Dickinson, T.A.; Pollock, L.J. Herbarium collections remain essential in the age of community science. Nat. Commun. 2024, 15, 7586. [Google Scholar] [CrossRef]
- Liu, D. World view. Nature 2024, 633, 741. Available online: https://www.nature.com/articles/d41586-024-03072-3 (accessed on 7 November 2024). [CrossRef]
Figure 1.
The study area's location and limits.
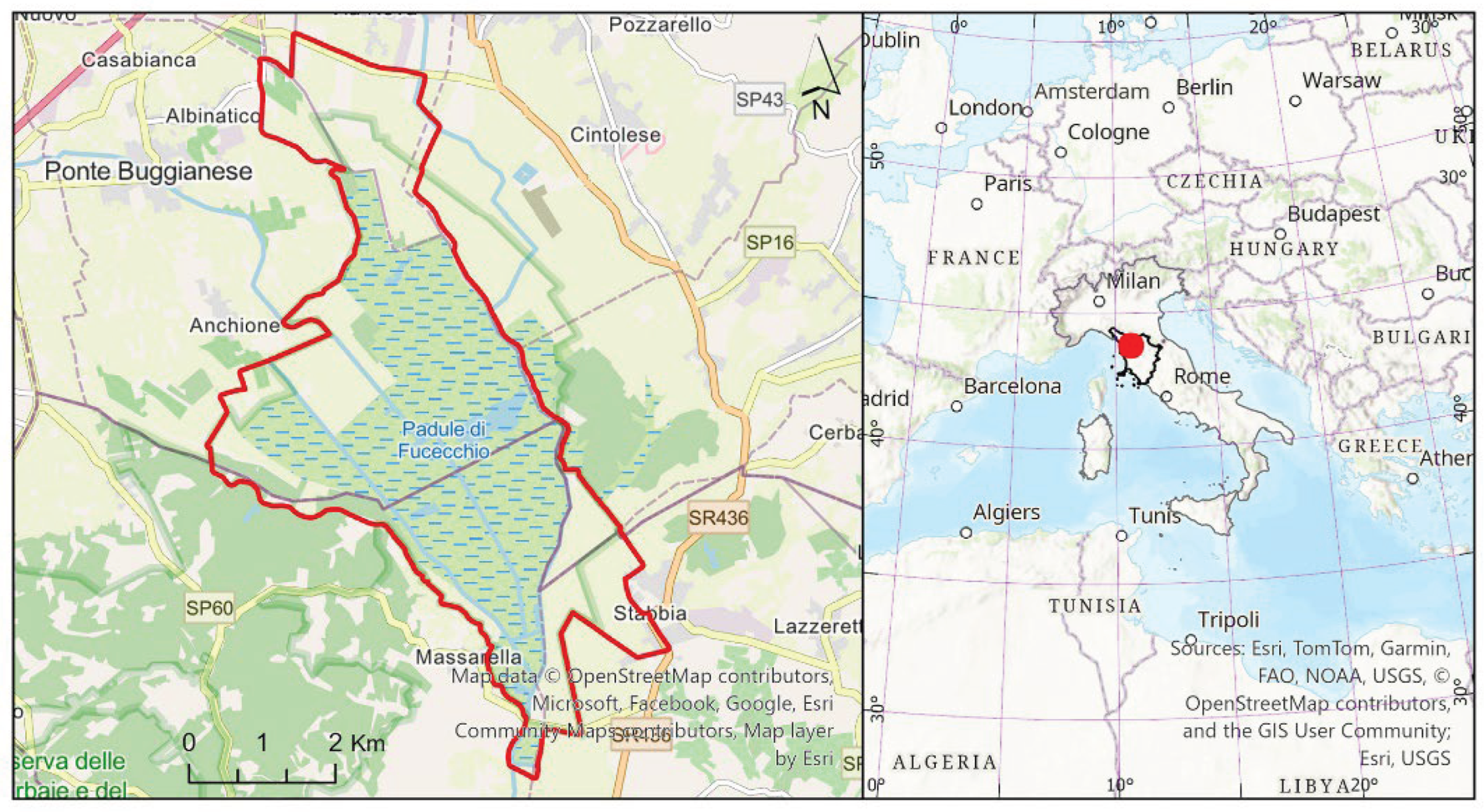
Figure 2.
On the left, a historical topographic map of the Valdarno di Sotto plain, showing the Padule di Fucecchio marshes. The map was drawn up between 1770 and 1780, based on the original from 1747 [41]. On the right, the current aerial photograph of the same area.
Figure 2.
On the left, a historical topographic map of the Valdarno di Sotto plain, showing the Padule di Fucecchio marshes. The map was drawn up between 1770 and 1780, based on the original from 1747 [41]. On the right, the current aerial photograph of the same area.
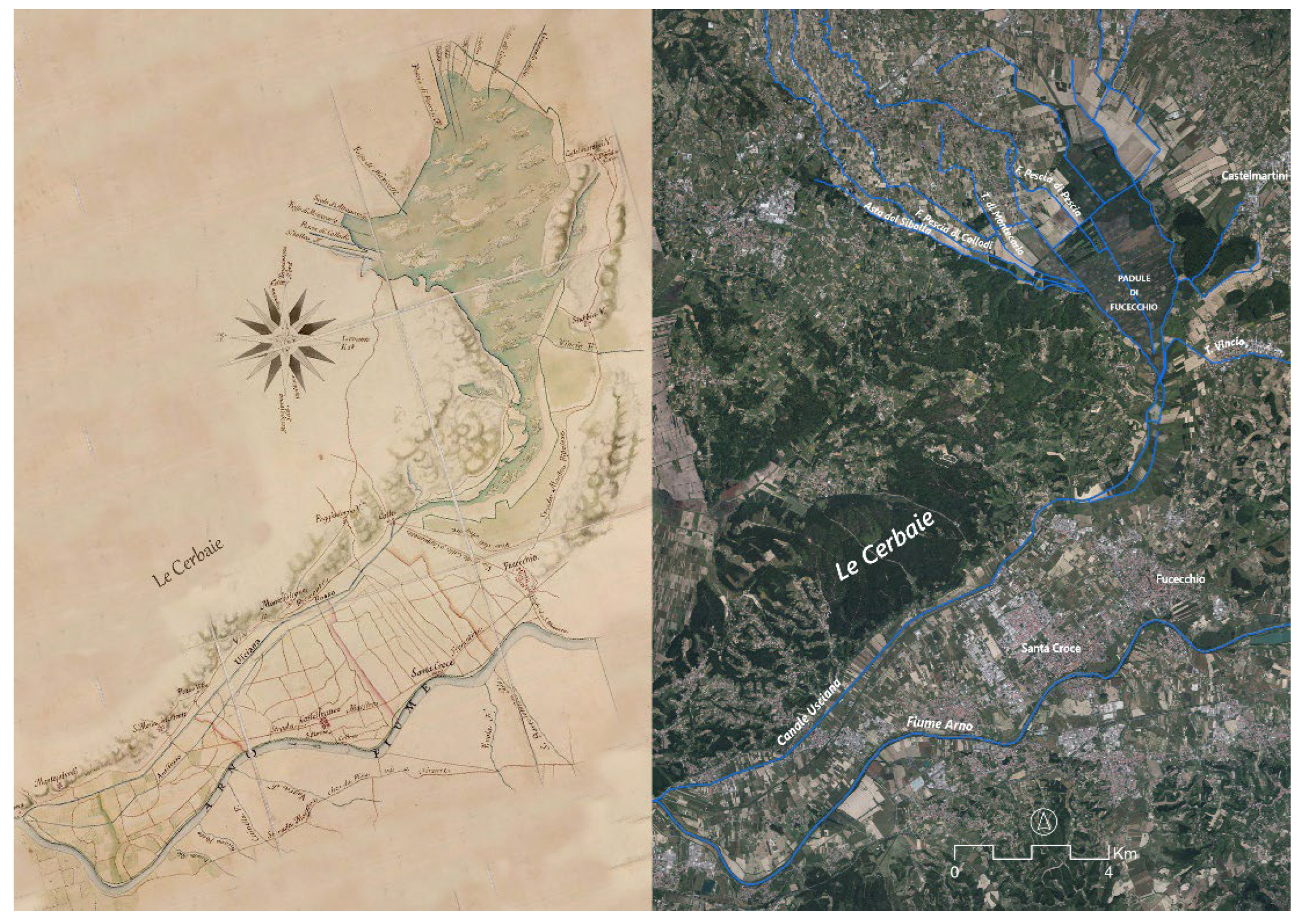
Figure 3.
Dendrogram resulting from the cluster analysis of palustrine plant communities. The numbers corresponding to the community type names are placed below the corresponding relevé numbers (see in the headings of Table 1 and Tables S1-S11 the line “Relevé number in cluster dendrogram”). 1: Polypogono viridis-Paspaletum distichi; 2: Caricetum ripariae; 3: Schoenoplectetum lacustris; 4: Bolboschoenetum glauci; 5: Frangulo alni-Salicetum cinereae; 6: Leucojum aestivum communities; 7: Juncus bulbosus - Gratiola officinalis communities; 8: Ranunculo flammulae-Juncetum bulbosi; 9: Eleocharitetum palustris; 10: Oenantho aquaticae-Rorippetum amphibiae; 11: Veronica peregrina communities; 12: Phragmitetum australis; 13: Bidens frondosa communities; 14: Phalaridetum arundinaceae; 15: Caricetum vesicariae; 16: Typhetum latifoliae; 17: Typhetum angustifoliae; 18: Glycerio-Sparganietum neglecti; 19: Cyperetum micheliano-difformis; 20: Echinochloo cruris-galli-Sporoboletum schoenoidis; 21: Phalarido arundinaceae-Bolboschoenetum laticarpi; 22: Portulaca oleracea communities; 23: Polygono lapathifolii-Xanthietum italici; 24: Echinochloo-Polygonetum lapathifolii.
Figure 3.
Dendrogram resulting from the cluster analysis of palustrine plant communities. The numbers corresponding to the community type names are placed below the corresponding relevé numbers (see in the headings of Table 1 and Tables S1-S11 the line “Relevé number in cluster dendrogram”). 1: Polypogono viridis-Paspaletum distichi; 2: Caricetum ripariae; 3: Schoenoplectetum lacustris; 4: Bolboschoenetum glauci; 5: Frangulo alni-Salicetum cinereae; 6: Leucojum aestivum communities; 7: Juncus bulbosus - Gratiola officinalis communities; 8: Ranunculo flammulae-Juncetum bulbosi; 9: Eleocharitetum palustris; 10: Oenantho aquaticae-Rorippetum amphibiae; 11: Veronica peregrina communities; 12: Phragmitetum australis; 13: Bidens frondosa communities; 14: Phalaridetum arundinaceae; 15: Caricetum vesicariae; 16: Typhetum latifoliae; 17: Typhetum angustifoliae; 18: Glycerio-Sparganietum neglecti; 19: Cyperetum micheliano-difformis; 20: Echinochloo cruris-galli-Sporoboletum schoenoidis; 21: Phalarido arundinaceae-Bolboschoenetum laticarpi; 22: Portulaca oleracea communities; 23: Polygono lapathifolii-Xanthietum italici; 24: Echinochloo-Polygonetum lapathifolii.
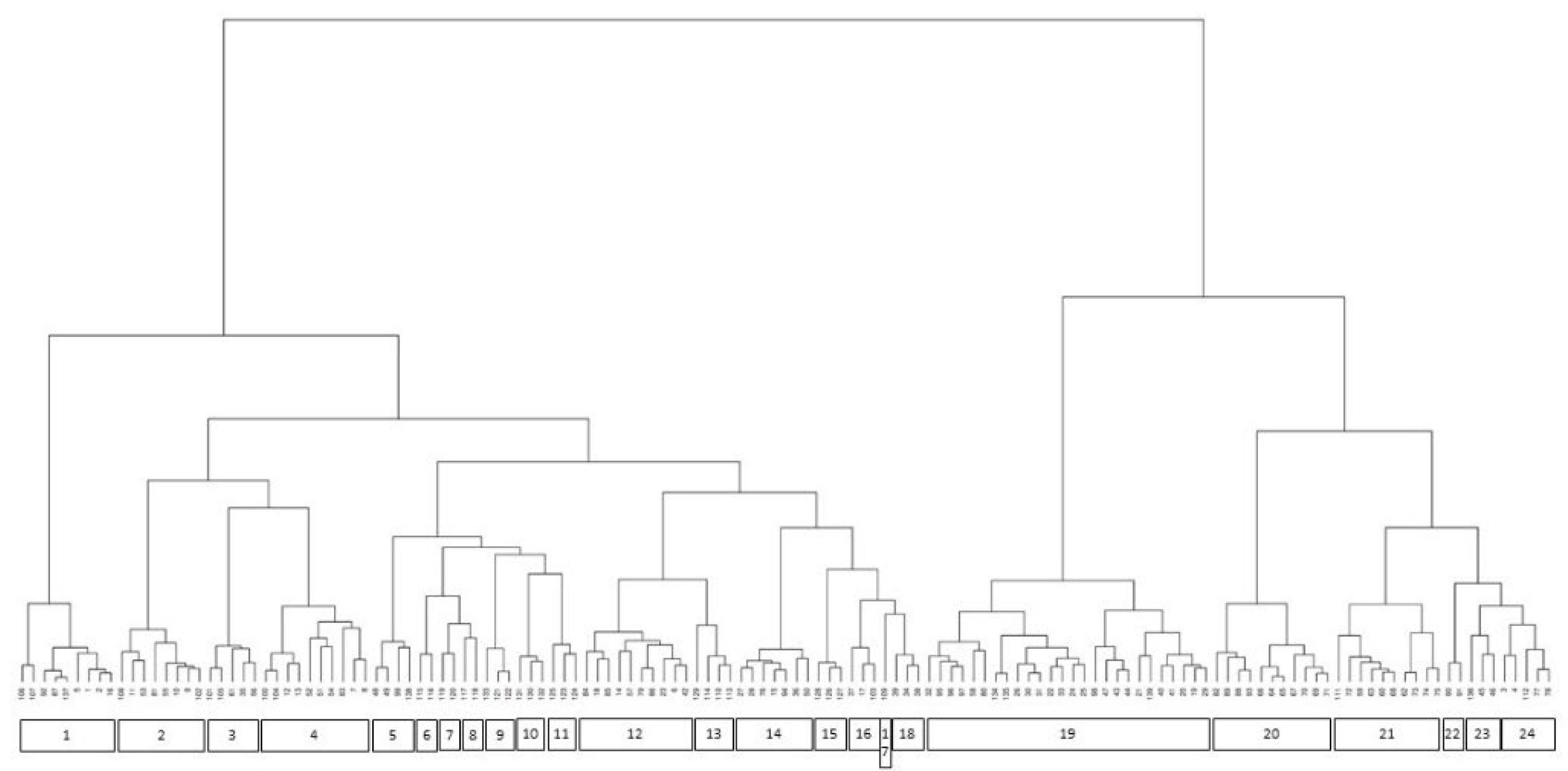
Figure 4.
A flooded pond in April, partly covered by Persicarietum amphibiae vegetation.

Figure 5.
A small area of a flooded pond in April, mostly covered by Callitriche brutia communities.
Figure 5.
A small area of a flooded pond in April, mostly covered by Callitriche brutia communities.

Figure 6.
A: a dried-up pond in August, whose bottom is home to Echinochloo cruris-galli-Sporoboletum schoenoidis communities. B: a Sporobolus schoenoides plant is shown in detail.
Figure 6.
A: a dried-up pond in August, whose bottom is home to Echinochloo cruris-galli-Sporoboletum schoenoidis communities. B: a Sporobolus schoenoides plant is shown in detail.

Figure 7.
An aspect of Cyperus-dominated communities (Cyperetum micheliani-difformis).

Figure 8.
A dried-up pond in August, with Portulaca oleracea-dominated communities growing on the bottom.
Figure 8.
A dried-up pond in August, with Portulaca oleracea-dominated communities growing on the bottom.

Figure 9.
A: A flooded canal in early May. B: The same canal in August, now dry, with Polygono lapathifolii-Xanthietum italici communities growing on the riverbed. C: A drying pond in early July, with a belt dominated by Bolboschoenus laticarpus in the foreground. D: Another pond in August, with Polygono lapathifolii-Xanthietum italici communities growing on the bottom.
Figure 9.
A: A flooded canal in early May. B: The same canal in August, now dry, with Polygono lapathifolii-Xanthietum italici communities growing on the riverbed. C: A drying pond in early July, with a belt dominated by Bolboschoenus laticarpus in the foreground. D: Another pond in August, with Polygono lapathifolii-Xanthietum italici communities growing on the bottom.

Figure 10.
An aspect of Oenantho aquaticae-Rorippetum amphibiae communities.


Table 1.
Palustrine plant communities of Cyperetum micheliani-difformis ass. nova (see the Supplementary Table S0 General Legend for the meaning of the cover-abundance value numbers).
Table 1.
Palustrine plant communities of Cyperetum micheliani-difformis ass. nova (see the Supplementary Table S0 General Legend for the meaning of the cover-abundance value numbers).
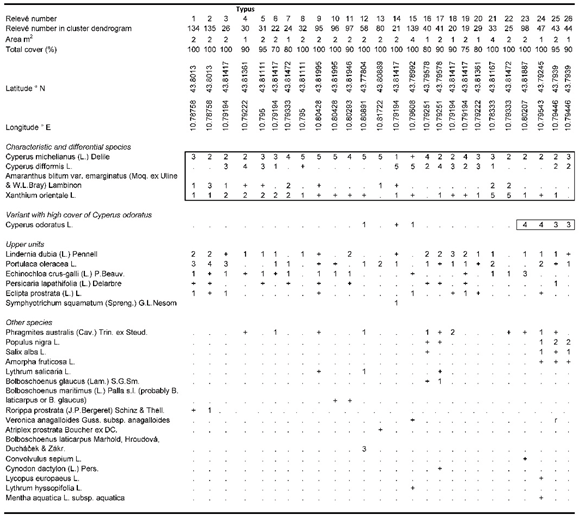 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



