
Submitted:
11 December 2024
Posted:
11 December 2024
You are already at the latest version
Abstract
Wetlands are among the most valuable ecosystems on the planet for biodiversity conservation, but also among the most threatened habitats in the world, strongly affected by human pressures and threats. The Mediterranean basin features numerous small lakes, ponds and pools, whose number and quality are decreasing at an alarming rate, and whose biodiversity is often little or not at all known. As a better knowledge of the biotic components of these minor water bodies is necessary, we carried out a phytosociological survey campaign in southern Tuscany (central Italy), an area with little information on the vegetation of aquatic and palustrine biotopes. Numerous previously unknown water bodies were localized and surveyed in this work, while the others already known were resurveyed. These investigations allowed us to identify 26 plant communities which can be classified into seven phytosociological classes. A new subassociation (Ranunculo ophioglossifolii-Callitrichetum stagnalis subass. ranunculetosum peltati) is newly described. The identification of the site-associated Natura2000 habitats led to the recognition of five habitats of conservation interest at national and European level. The results of these investigations allow to advance our knowledge of the flora and vegetation of these small but valuable natural sites, providing a basis for their preservation.
Keywords:
aquatic and palustrine habitats
; conservation
; freshwater ecosystem
; habitats
; phytosociology
; syntaxonomy
; vegetation
1. Introduction
Wetlands are among the most valuable ecosystems on the planet, as they fulfil multiple ecological services and are crucial for biodiversity conservation [1,2,3,4,5,6]. Nevertheless, wetlands are among the most threatened habitats in the world, strongly affected by human pressures and threats, such as transformation in agricultural and urban land, overexploitation, water pollution, flow modification, destruction or degradation of habitat, and alien species invasion [1,5,7,8,9]. This worrying global trend has been rapidly increasing in recent years, even for wetlands included in protected areas [10,11,12]. Consequently, it is not surprising that also in Europe and in Italy, freshwater ecosystems include many threatened habitats with an unfavorable conservation status [13,14,15,16,17,18].
In addition to large wetlands, for which a lot of information is often available [19,20,21], the Mediterranean basin also features numerous small lakes, ponds and pools, both in the mountains and in the lowlands, whose biodiversity is often little or not at all known [21,22]. Especially at relatively low elevations, many of these freshwater ecosystems can be of partially or completely artificial origin related to anthropogenic activities, but still highly relevant for plant diversity and conservation [7,23,24,25,26,27]. In the Mediterranean basin, small water bodies, permanent or more or less temporary, are generally widespread and provide important ecosystem benefits, but their quantity and quality are decreasing at an alarming rate [19,28]. A better knowledge of the biotic components of these minor water bodies is necessary for their conservation. Accordingly, we carried out a vegetation survey campaign in southern Tuscany (central Italy) and particularly in Maremma (Figure 1), an area where the vegetation information regarding aquatic and palustrine communities is scarce and, if present, mostly concerning large wetlands [29,30,31]. Numerous previously unknown water bodies were localized and surveyed in this work, while others were re-visited after several years since the first survey. The results of these investigations are the object of this contribute.
2. Materials and Methods
2.1. Study Area
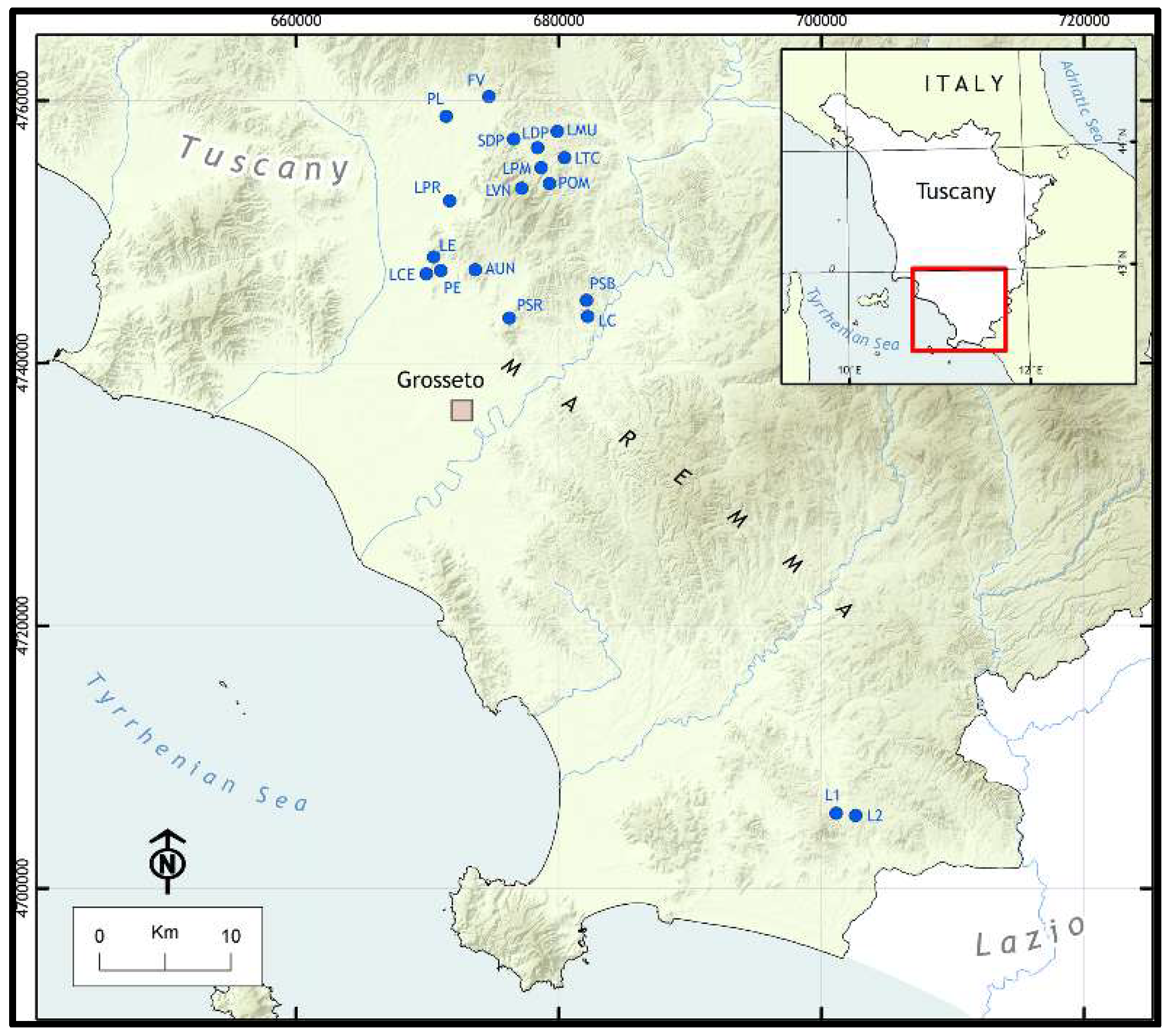
The 19 study sites are located in the province of Grosseto (42.7705N, 11.1121E), Figure 1, the southernmost and largest one in Tuscany. A general description of the main environmental characteristics and phytogeographical sectors of this territory is provided in Selvi [32] and Selvi in Giovacchini et al. [33]. Most of the sites are located in the northern hill-planitial sector, while only two belong to the southern hill-planitial sector. Their names, abbreviations, geographical details and information about inclusion in protected areas are given in Appendix S1.
2.2. Climate and Bioclimate
With 720–800 mm of mean annual rainfall, ca. 3 months of drought and 14–14.5 °C of mean annual temperature, the climate is typically meso-Mediterranean. According to the classification of Thornthwaite & Mather [34], the climate belongs to the type ‘‘second mesothermic, suboceanic (B’2b’4)”, and varies from “humid to subhumid (C2 s)” for the northern sites and from “subhumid to subarid (C1w2)” in the southern sites. As to the bioclimate, according to Pesaresi et al. [35], the study area has a Pluviseasonal oceanic Mediterranean bioclimate. Only the northern sites are near the border with the oceanic submediterranean Temperate bioclimate. As to the Italian ecoregion approach [36], all the sites are within the Thyrrenian Province of the Mediterranean Division, specifically in the Maremma Subsection (2B1b). Again, some of the northern sites are near the border with the Apennine Province of the Temperate Division, specifically near the Tuscan Basin Subsection (1C1b).
2.3. Geological Outline
The sites of the northern sector are mainly located around Monte Leoni (616 m a.s.l.), the highest elevation of a vast hill system mainly oriented along a NE-SW direction and covering an area of ca. 120 km2 between the Ombrone and the Bruna rivers, at the east and the west, respectively. The geological backbone of the area is formed by the so-called “Verrucano” formation, an assemblage of crystalline siliceous rocks of quartzitic-anagenetic type dating back to the upper Triassic [37,38,39,40]. The seven sites LDP, LMU, LTC, LPM, POM, SDP, and LVN lie in a vast low-elevation plateau (150-200 m a.s.l.) extending at the base of the northern side of Mt. Leoni. This plateau is formed by fluvio-lacustrine sands and loose conglomeratic material originated in a continental environment by the erosion of the siliceous rocks from the surrounding hill slopes and deposited in thick layers during the Pleistocene [41]. The area includes small springs, water veins and natural pools, part of which were transformed into small semi-permanent ponds (POM, LDP, LMU, LVN) or permanent lakes (LTC, LPM), to allow the watering of the livestock that was kept in the area by the local population since at least the 18th century. The LPR site is a shallow seasonal pond of natural origin found at the western foot of the Mt Leoni area, on sandy-silty soil. The FV site is similar, but lying on acid effusive rocks of rhyolitic type formed during local volcanic phases of the Pliocene. This site is included in a dense cork oak forest and was not used for livestock watering. The PL site is a seasonal artificial pond created in the plain of river Bruna at the NW of Mt. Leoni, with alluvial soil of sandy-silty texture, used since long time for cattle watering.
The ponds in the south-western part of the study area (LE, LCE, PE, AUN) are mainly of artificial origin, and lie in the southern sector of the Mt Leoni hill complex, also consisting of the quartzitic Verrucano formation described above. The south-eastern ponds PSR, LC and PSB are also of artificial origin, but located in hilly area dominated by a clayey-calcareous formation of the mid Eocene-Paleocene [38].
The two sites in the southern sector are the Lagaccioli lakes (L1 and L2), which are placed in a heterogeneous geological area, with limestone, shales, clays, sandstones together with pebbles and muds of alluvial and lacustrine origin [29,40]. Their formation is thought to be of karstic origin [42].
2.4. Data Set and Data Analysis
Our dataset is composed of 88 original relevés carried out in plant communities dominated by aquatic and palustrine species. Data were collected in the spring 2024 (April-June) using the classical phytosociological method and its updates [43,44,45,46,47,48]. After transforming the original Braun–Blanquet cover-abundance scale into the ordinal scale in accordance with Van der Maarel [49], we separated aquatic coenoses from palustrine ones, based on the growth forms [50] of the dominant species. We then analysed the two resulting matrices through a cluster analysis in R environment [51], using the chord distance of the function vegdist of ´vegan´ package [52] and the median linkage of the function hclust of ´stats´ package [51].
Plant species names follow the Portal to the Flora of Italy [53] based on Bartolucci et al. [54], while the syntaxonomic nomenclature of classes, orders, and alliances follows mainly the Vegetation of Europe by Mucina et al. [55], and the Italian Vegetation Prodrome [56]. The syntaxonomical nomenclature is in accordance with the 4th edition of the ICPN [57].
3. Results
The dendrograms resulting from the cluster analysis (Figures S1 and S2) allowed us to identify 26 different plant communities. From a syntaxonomic viewpoint, they can be classified in seven classes: Charetea intermediae, Lemnetea, Potamogetonetea, Isoëto-Juncetea, Littorelletea uniflorae, Phragmito-Magnocaricetea and Molinio-Arrhenatheretea. Each community is described and commented in the following paragraphs. A comprehensive syntaxonomical scheme is reported in Appendix A.
Nitelletum hyalinae Corillion 1957 (Table S1, rel. 1)
Several algal species of Characeae were found in the sites, mostly in mosaic with aquatic vascular plant communities and often forming a more or less dense lower layer in the coenoses of the Potametea class. In some cases, the collected specimens lacked the diagnostic characters necessary for the identification at the species level, while in other cases it was possible to identify the following species: Chara vulgaris L., C. globularis Thuiller and Nitella hyalina (DC.) C.Agardh. The latter taxon formed a well distinct community in the small pond LVN. According to Bazzichelli & Abdelahad [58], in Italy this species is present in different habitats, such as ponds, lakes and running waters, and it is known from Lombardy, Veneto, Latium and Sicily. Nitella hyalina characterizes the association Nitelletum hyalinae Corillion 1957, gravitating in the Nitellion flexilis alliance which includes Atlantic to sub-Atlantic communities of moderately acidic to neutral waters, of low conductivity [59]. Although the association sometimes shows the co-presence of other Charophytes, in particular of Chara [59], it can also form coenoses rich in vascular plants such as hydrophytes and helophytes or also very species-poor and almost monospecific communities [60]. In the study area, the association was in contact, on one side, with the Potamogetonetum natantis, on the other side with the hygrophilous vegetation of the shallow water near the shores, as evidenced by the presence of Peplis portula L., Alisma lanceolatum With. and Agrostis stolonifera L.
Utricularietum australis Müller et Görs 1960 (Table S1, rel. 2)
This association is dominated by Utricularia australis R.Br., occurring in mesotrophic to naturally eutrophic water bodies, such as alluvial pools and oxbows but also in flooded sand pits and newly established wetlands [61]. It shows a pioneer behaviour, as recently pointed out by Viciani et al. [26] for some sites in the eastern Tuscan Apennines. In the study area, this community type was found at the lake of Piane di Materazzo (LPM). The association was developed in the shallow waters near to the shores, in a sheltered position at the edge of the reeds as shown by the several helophytic species in the relevé.
Potamogetonetum natantis Hild 1959 (Table S1, rel. 3-8)
The coenoses dominated by Potamogeton natans L. were widely represented and detected in four sites (LMU, LDP, LVN, POM). These communities can be attributed to the association Potamogetonetum natantis Hild 1959, occurring in oligo- to eutrophic water bodies with depths of 20–100 cm, with still or slowly flowing waters [62]. The association seems to be particularly widespread in small and low-depth water bodies, where it often forms very dense and paucispecific stands [26]. This can be confirmed also in our study area, where P. natans forms rather species-poor communities, mainly developing in shallow waters subjected to drastic variations in depth across the seasons. The occurrence of Phragmites australis (Cav.) Trin. ex Steud. indicates the contact of Potamogetonetum natantis with the tall helophytes belt (Phragmitetum australis) while, in very shallow waters, the presence of small helophytes such as Eleocharis multicaulis (Sm.) Desv., Juncus bulbosus L. or Peplis portula indicates a water dynamics favoring the affirmation of coenoses belonging to different classes during the summer desiccation (e.g. Littorelletea or Isoëto duriei-Juncetea bufonii).
Potamogetonetum pusilli von Soó 1927 (Table S1, rel. 9-12)
This association is typical of mesotrophic to eutrophic, clear waters of shallow parts of natural and artificial ponds, channels and rivers, especially along their lower courses, developing at various water depths, also in disturbed or in early succession stage habitats [62]. In the study area the association was found in three small ponds (AUN, LE, PE), all subjected to strong summer desiccation. In all the sites it was characterized by the presence of Chara species. In the ponds near Roselle (LE, PE) the association occupied the central and deeper waters compared to Ranunculion aquatilis coenoses which, instead, developed near the shores at few centimeters of depth.
Parvo-Potamogetono-Zannichellietum pedicellatae De Soó 1947 (Table S1, rel. 13-16)
According to Šumberová [62], stands dominated by Zannichellia palustris L. or mixed stands with Z. palustris and some narrow-leaved species of Potamogeton can be attributed to the association Parvo-Potamogetono-Zannichellietum pedicellatae, belonging to the Potamion alliance. In addition, this species can form communities also in brackish waters, gravitating in different syntaxa, such as the alliance Zannichellion pedicellatae.
The Parvo-Potamogetono-Zannichellietum pedicellatae grows in eutrophic to hypertrophic, often turbid, shallow freshwaters, mainly in ponds and pools and more rarely in slowly running streams [62]. In the study area, the association was found in two ponds (LC, PSB) where the ecological conditions and the floristic composition appear very similar. Zannichellia palustris grew on top of a dense layer of Chara species in the deepest part of the ponds, sometimes in contact with Potamo-Ranunculetum trichophylli association, present in more shallow waters. This community is also subject to strong water fluctuations in the ponds, already disappearing in early summer.
Potamogeton nodosus community (Table S1, rel. 17)
In a strongly disturbed pond (AUN) near Nomadelfia, in addition to the Potamogetonetum pusilli, a small stand dominated by Potamogeton nodosus Poiret was present. This vegetation type was characterized by an abundant presence of Chara vulgaris L. Although Potamogeton nodosus communities are sometimes considered communities typical of running waters, e.g. [63], their presence in standing waters is rather common [26,64]. The Nomadelfia community could be thus represent an impoverished aspect of the Potamogetonetum denso-nodosi O. de Bolòs 1957 association, included in the Potamion alliance, which was reported both in still and in flowing waters [65].
Potamogeton lucens community (Table S1, rel. 18)
A small and sparse stand dominated by Potamogeton lucens L. was found at the lake of Piane di Materazzo (LPM). Despite the low total cover value, this population is rather rich in hydrophytes, due to the presence of P. nodosus, P. pusillus and Utricularia australis. This community type represents an impoverished aspect of the association Potamogetonetum lucentis Hueck 1931, already reported for the southern Maremma (central Italy) by Lastrucci et al. [29].
Ranunculo ophioglossifolii-Callitrichetum stagnalis Brullo, Scelsi & Spampinato 2001 (Table S1, rel. 19-26)
subass. typicum (Table S1, rel. 19-21)
subass. ranunculetosum peltati subass. nova (Table S1, rel. 21-26) [holotypus rel. n. 26 Table S1 hoc loco]
This association was described for the Aspromonte massif (southern Calabria) by Brullo et al. [66], and is typical of long-time eutrophic flooded depressions subject to summer drying out. In the study area, this vegetation type showed the dominance of Callitriche stagnalis Scop., while Ranunculus ophioglossifolius Vill. was often present in the aquatic form with floating leaves. The subassociation typicum, corresponding to the original description by Brullo et al. [66] in accordance with the ICPN rules [57], was found at the edge of two very small ponds (LE, PSB), rapidly sloping and occupied in the deepest parts by communities dominated by Potamogeton pusillus or Zannichellia palustris. At the edge of two larger ponds (LCE, PE), the presence of Ranunculus peltatus, sometimes with high cover, supports the description of the new subassociation ranunculetosum peltati. This subassociation grows in shallow waters sloping less rapidly towards the deeper center, as indicated by the presence of palustrine species occurring in the wet soils around the ponds (Agrostis stolonifera, Glyceria fluitans (L.) R.Br., Ranunculus repens L.). The presence of Callitriche brutia Petagna highlights the potential for the development of the association Callitricho brutiae-Ranunculetum ophioglossifolii Gigante, Maneli & Venanzoni 2013. In the study area, this community finds its optimum at a later temporal phase, the late spring, when drying further reduces the depth of the water until the humid bottom of the pool is left uncovered.
Callitricho brutiae-Ranunculetum peltati Pizarro & Rivas-Martinez 2002 subass. ranunculetosum trichophylli Lastrucci, Foggi, Selvi & Becattini 2007 (Table S1, rel. 27-31) and Potamo crispi-Ranunculetum trichophylli Imchenetzky 1926 (Table S1, rel. 32-35)
In two sites of the southern Maremma, Lastrucci et al. [29] detected the association Callitricho brutiae-Ranunculetum peltati Pizarro & Rivas-Martinez 2002, describing two subassociations, one (ranunculetosum trichophylli) for the Lagaccioli site and another one (callitrichetosum obtusangulae) for the Marruchetone lake. After almost 20 years, in the Lagaccioli ponds, the species of the association are still present. Although during the survey season the water level was rather high, some considerations concerning the different frequency of the various species in this community types can be done. Callitriche brutia was found only in a small patch at one of the two ponds, at the edge of the marsh vegetation, in very shallow water. It is possible that the depth of the water did not allow observation of this species in other parts of the ponds. For Ranunculus peltatus Schrank a situation similar to that reported by Lastrucci et al. [29] was observed, showing that only occasionally this was the dominant species. On the other hand, Ranunculus trichophyllus was almost always the dominant species in this community type, also forming populations in which the other two species were missing.
Stands dominated by R. trichophyllus were also detected in other sites than Lagaccioli, such as the Casalino pond (LC). These communities dominated by Ranunculus trichophyllus cannot be referred to Callitricho-Ranunculetum peltati subass. ranunculetosum trichophylli due to the total lack of the association guide species in the site. They can instead be referred to the association Potamo crispi-Ranunculetum trichophylli Imchenetzky 1926, in accordance with what was reported from other sites in Tuscany [26,30] or from the "Altipiani di Colfiorito" site in Umbria [67]. It is possible that at the Lagaccioli site both syntaxa are present and a sort of balance between them is regulated by the hydrological levels favoring or disfavoring the development of Callitriche brutia and Ranunculus peltatus. Another aspect that could limit the development of these two latter hydrophytes with emergent leaves is the marked increase, compared to the situation observed by Lastrucci et al. [29], of the aquatic ecophene of Persicaria amphibia (L.) Delarbre, present in almost all the Lagaccioli releveés, even with high cover values and a dominant role. Lastrucci et al. [29], in fact, indicated that this species, with its terrestrial ecophene, was particularly competitive during the drying period of the ponds but not during the aquatic phase of the habitat.
Lemno-Callitrichetum obtusangulae (Philippi 1978) Passarge 1992 (Table S1, rel. 36-37)
Callitriche obtusangula Le Gall communities can develop both in running and standing water habitats. In the first case, the coenoses can be referred mainly to the Batrachion fluitantis alliance, while in the second to the Ranunculion aquatilis alliance [68,69]. The association Lemno-Callitrichetum obtusangulae represents a kind of C. obtusangula-dominated vegetation reported for springs or small water bodies [68,69]. In the study area C. obtusangula-stands were only detected in one site (PSR), where the species covered almost the entire pond, developing above a layer of Chara sp. The association appears in an extremely impoverished form since only Ranunculus trichopyllus and Mentha aquatica were present.
Callitriche platycarpa community (Table S1, rel. 38)
In a small pound near Roselle, mainly occupied by communities belonging to Ranunculo ophioglossifolii-Callitrichetum stagnalis ranunculetosum peltati and to Potamogetonetum pusilli, a small patch dominated by Callitriche platycarpa Kütz. was found. Felzines [70] reported from France the association Veronico beccabungae-Callitrichetum platycarpae Grube ex Felzines 2016, belonging to the Ranunculion fluitantis alliance and indicating the reophile character of this community, though this species can grow in different kinds of habitats [71]. In Italy this species is rather rare and in Tuscany, in particular, known mainly from old records [72], so the lack of recent distribution and phytosociological data could affect a comprehensive understanding of the communities formed by this species.
At the edge of the natural pool close to Frantoio Verrocchio (FV), during the late spring drying, two vegetation types characterized by small helophytes were observed, behind a higher belt dominated by Eleocharis palustris (L.) Roem. & Schult. The first one develops on still moist soils due to the shading of the cork oak trees surrounding the pool. This community is rather dense and is dominated by Isolepis cernua associated with especially Alisma lanceolatum, both with rather high cover. Isolepis cernua (Vahl) Roem. & Schult. can form different communities belonging to syntaxa of Isoëto duriei-Juncetea bufonii class [73,74], but it is often found in species-poor communities that are difficult to place at the association level (see also Lastrucci et al. [64,75]) as in the case of the study area.
The second vegetation type develops on soils that dry up earlier, thus appearing as solidified mud at the time of our survey. This community differs from the previous one from a physiognomic point of view, being much sparser, and from a floristic point of view due to the presence of the rare Damasonium alisma Mill. The species was considered a characteristic of the Isoëtetalia order [73], although it can participate to associations attributed to other syntaxa, such as Damasonio alismae-Crypsietum aculeatae (Verbenion alliance, according to Brullo & Minissale [73]). This association was described by Rivas-Martinez et al. [76] from rather saline clay soils of the Guadalquivir brackish marshes in Spain and is characterized by the strong dominance of Sporobolus aculeatus (L.) P.M.Peterson, a species absent from our releveés.
The presence of small stands of Damasonium alisma in the southern Maremma and referred to the Isoëtetalia order had been already reported along a thin strip of the drying bottom at the eastern Lagaccioli pond [29]. In this study, however, it was not possible to observe again these stands, probably due to the high level of water at the time of the surveys.
Peplido portulae-Ludwigietum palustris Robbe ex J.-M. Royer, Felzines, Misset & Thévenin 2006 nom. mut. propos. (Table S2, rel. 5-6)
Ludwigia palustris (L.) Elliott dominated communities were observed in two small ponds of the study area (LVM, POM), developing on the muddy shores emerging during the drying period in late spring. In Maremma, L. palustris species-poor communities were recorded also from the Capalbio wetlands, at the Uccellina lake [29]. As pointed out by Lastrucci et al. [77] and Dítě et al. [78], L. palustris communities were placed by several authors in different syntaxonomic classes, such as Bidentetea, Isoëto duriei-Juncetea bufonii, Littorelletea or also Phragmito-Magnocaricetea. The presence of Peplis portula in one relevés allowed us to place the coenoses of the study area in the Peplido portulae-Ludwigietum palustris, a thermophilous association observed in different wetland types such as periodically inundated riverbanks, depressions and water reservoirs gravitating in the alliance Eleocharition soloniensis Philippi 1968 (in the past reported as Elatini-Eleocharition ovatae Pietsch 1973, today considered a synonym according to Mucina et al. [55]), of the class Isoëto duriei-Juncetea bufonii [78].
Callitricho brutiae-Juncetum bulbosi Gigante, Maneli & Venanzoni 2013 (Table S3, rel. 1-2)
In one site (LVN) of the Mt. Leoni area, small stands of Juncus bulbosus and Peplis portula were observed. The presence of Callitriche brutia allowed us to place these coenoses to the Callitricho brutiae-Juncetum bulbosi described by Gigante et al. [74] from some shallow ponds at the Ferretto site in Umbria, where the tufts of the dominant species Juncus bulbosus forms small bulges separated by shallow water depressions. Our releveés are more species-poor compared to those from Ferretto and the spaces between the tufts of J. bulbosus are occupied by Peplis portula and sometimes by Potamogeton natans, probably configuring a deeper-water variant of the typical association.
Junco bulbosi-Eleocharitetum multicaulis (Passarge 1955) Passarge 1999 (Table S3, rel. 3-5)
Eleocharis multicaulis was found to form dense stands along the partially flooded shores of some ponds and pools of the study area, on muddy soils gently sloping towards the submerged zones and also in more prolonged flooded sites, in contact with aquatic Potamogeton natans coenoses. On the external drier edge, this community is in contact with the humid meadows with Gratiola officinalis L. or with the taller vegetation dominated by Molinia arundinacea Schrank and Schoenus nigricans L. From a phytosociological point of view, the community of the study area differs from the Eleocharitetum multicaulis Allorge ex Tüxen 1937 especially from a floristic point of view, less for its synecology. This association represents a medium-low height amphibious turf developing on a more or less organic substrate [79] typical of oligotrophic and acidophilic habitats [80]. The association is characterized by species that are absent from our study area, such as Hypericum elodes L., Helosciadium inundatum (L.) W.D.J.Koch, Isolepis fluitans (L.) R.Br., Ranunculus baudotii Godr. (sub R. flaccidus subsp. confusus) or Potamogeton polygonifolius Pourr.; Eleocharitetum multicaulis can also show aspects rich in Sphagnum sp. pl. [81].
The more Mediterranean association Junco emmanuelis-Eleocharitetum multicaulis described by Rivas-Martinez et al. [76] appears very different from the floristic point of view, due the presence of species with restricted distribution such as Juncus emmanuelis A.Fern. & J.G.García and Avellara fistulosa (Brot.) Blanca & C.Díaz. Hence, the attribution of our communities to these associations does not seem appropriate. Passarge [82] described the association Junco bulbosi-Eleocharitetum multicaulis, which lacks the differential species listed above and is rich in Phragmition species such as Lycopus europaeus L., Lythrum salicaria L., Phragmites australis and other hygrophilous species like Juncus articulatus L. or Agrostis stolonifera, also present in our relevés. For these reasons, we provisionally place the E. multicaulis stands of the Maremma territory in the latter association. It is possible that further studies in Mediterranean Tuscany and possibly in other sites of the western Italian peninsula will lead to recognize such communities as a distinct syntaxon.
Phragmitetum australis Savič 1926 nom. corr. (Table S4, rel. 1-8)
This is a widespread association, rather common in Italy and Europe [83,84], developing in several types of wetlands, from meso- to eutrophic fresh or brackish waters, and on different types of soils. In the study area, however, the association was not very frequent, developing mainly in some wide and open ponds and small lakes (L1, LDP, LMU, LPM). where the shading of the surrounding forests was less intense. It was developed in more or less flooded soils, in contact with aquatic or other marsh vegetation types, often forming belts of variable width around the water bodies.
Although the association and, more generally, P. australis stands, show serious decline in several large lakes in Italy [85], no signs of die-back were observed in the study area (e.g. the clumping habitus). This is probably because the strong water seasonal fluctuations in the examined sites reduce the risk of permanent submersion, which seems to be an unfavorable ecological condition for the Phragmitetum [86]. The vegetation resurvey on the Lagaccioli sites confirmed the current presence of the association, already reported by Lastrucci et al. [29].
Lysimachia vulgaris community (Table S4, rel. 9-10)
The presence of Lysimachia vulgaris L. community in the western Lagaccioli pond (L1) was reported already by Lastrucci et al. [29]. This association represented the more external hygrophilous community in contact with the woody vegetation around the pond. Our releveés confirmed its presence in the site, where it appeared more extended and abundant than in the past, probably at the expenses of other communities such as the Sparganietum, which is currently limited to the opposite bank of the ponds and no more in contact with the Lysimachia stands, as reported by Lastrucci et al. [29]. Although Lysimachia vulgaris is found in communities belonging to different syntax, see Lastrucci et al. [29], the presence of helophytes and also by some hydrophytes in the Lagaccioli stands, due to the prolonged submersion, confirms the position of these coenoses in the Magnocaricetalia order, Lastrucci et al. 2007 [29], and the Magnocaricion elatae alliance [87].
Glycerio-Sparganietum neglecti Koch 1926 (Table S5, rel. 1)
A stand with Sparganium erectum s.l. was detected in the western pond of Lagaccioli (L1), forming a narrow belt between aquatic coenoses with Ranunculus trichophyllus or Persicaria amphibia and Phragmitetum australis communities. According to Landucci et al. [83,84], Glycerio-Sparganietum neglecti Koch 1926 should be considered as a “macro-association” including other associations recognized in the past, such as Sparganietum erecti Roll 1938 and Sparganietum microcarpi (Weber 1976) Passarge 1978, due to the frequent uncertain taxonomic identification of the species of this group, see for Italy Lastrucci et al. [88]. Our research could not confirm the presence of the association in the eastern Lagaccioli pond (L2), where it had been previously indicated by Lastrucci et al. [29].
Glycerietum fluitantis Nowinski 1930 nom. inval. (Table S5, rel. 2-3)
The communities belonging to this association develop mainly on habitats in advanced stage of terrestrialization or periodically flooded, such as ponds, banks of channels and streams or inundated depressions [83]. Although G. fluitans is rather common the study area, coenoses dominated by this species that can be placed in the association were only found in two ponds (LCE, PL). These were developed on shallow waters, in contact with, respectively, aquatic coenoses and Eleocharitetum palustris, well-tolerating strong water fluctuation [89]. Concerning the nomenclatural problems of this association, see Lastrucci et al. [27]. Our research could not confirm these communities at the Lagaccioli sites (L1, L2), where, instead, it was previously reported by Lastrucci et al. [29].
Alismo lanceolatae-Gratioletum officinalis Biondi & Bagella 2005 (in mosaic with Junco tenageiae-Solenopsietum laurentiae Gigante, Maneli & Venanzoni, 2013) (Table S5, rel. 4-6)
Gratiola officinalis typically develops in areas with wet soils for a rather long period in spring, but drying up in the summer, see Gigante et al. [74]. From a syntaxonomical point of view, similarly to the coenoses at the Ferretto site (74 Gigante et al., 2013), it forms communities representing an impoverished aspect of Alismo lanceolatae-Gratioletum officinalis, described by Biondi & Bagella [90] from the Maddalena Archipelago (Sardinia). In the study area, the species formed dense stands along the wet shores of two small ponds (LPM, LTC). The presence in one relevé of Juncus conglomeratus L. suggests that also in the study area there is the potential for the presence of subassociation juncetosum conglomerati Gigante, Maneli & Venanzoni 2013.
It is important, however, to note that at least in two relevés the floristic composition is very peculiar and a well-defined pool of small hygrophytic species, such as Solenopsis laurentia (L.) C.Presl, Juncus tenageia L.f., J. pygmaeus Rich. ex Thuill., Cicendia filiformis (L.) Delarbre, Centaurium maritimum (L.) Fritsch, Juncus bulbosus and Eleocharis multicaulis was present. Many of these species were also found in the Ferretto site by Gigante et al. [74], where they formed acidophilic communities placed in the associations Junco tenageiae-Solenopsietum laurentiae Gigante, Maneli & Venanzoni, 2013 and Solenopsio laurentiae-Juncetum pygmaei V. Silva & Galán de Mera in V. Silva, Galán de Mera & Sérgio 2008, with the subassociation isolepidetum cernuae Gigante, Maneli & Venanzoni, 2013. At least one of our relevés (Table S5, rel. 6), with rather high cover of Solenopsis laurentia, could be considered as an aspect of the Junco tenageiae-Solenopsietum laurentiae in mosaic with the Alismo-Gratioletum association; also rel. 4 of Table S5 was rather rich in annual small helophytes.
It remains unclear whether the presence of Gratiola offinalis, with its higher biomass and shading effect, could be detrimental to the small species above. To this purpose, monitoring of this vegetation would help to understand whether the Alismo-Gratioletum, given the pioneer character of its dominant species, is a community that expands to the detriment of others, or the mosaic between species of different sizes remains stable over time.
Eleocharitetum palustris Savič 1926 (Table S5, rel. 7-13)
The communities belonging to this association have a strong pioneer character and are rather common in Italy [83,91]. Lastrucci et al. [27] reported the Eleocharitetum palustris association as particularly common in small ponds and pools, where it forms narrow strips or small stands often in shallow waters and disturbed habitats. In the study area, these patterns were confirmed and the association was found also in very small pools subject to rapid drying, often in contact with the Glycerietum fluitantis association. Due to the quick drying in late spring, in some very small areas such as the pool of Frantoio Verrocchio (FV), the Eleocharitetum palustris is present with dense stands only in the more humid central areas while it is unable to develop in the early drying marginal areas, where the empty spaces are occupied by a vegetation of small helophytes in mosaic with it. The vegetation resurvey at the Lagaccioli sites confirmed the presence in L1 of the association previously reported by Lastrucci et al. [29].
Oenantho aquaticae-Rorippetum amphibiae Lohmeyer 1950 (Table S5, rel. 14-19)
The association develops on periodically flooded habitats with strong water fluctuation and drying out in summer, such as depressions at the bottom of floodplains of rivers and streams, shorelines of dead branches and artificial water reservoirs, or slow-flowing lowland stream beds that may be overgrown with vegetation [92]. The habitats are often eutrophic, the substrate consists of a layer of mud and organic material of varying thickness. In the study area, the association was found only at the Lagaccioli sites (L1, L2), where it was formerly reported by Lastrucci et al. [29]. According to these authors, the association can be dominated by Rorippa amphibia in some stands and by Oenanthe aquatica in others. Our relevés confirmed the presence of a variant with Persicaria amphibia, sometimes becoming the dominant species, as previously already suggested by Lastrucci et al. [29].
Bolboschoenetum glauci Grechushkina, Sorokin & Golub 2011(Table S5, rel. 20)
At the Lagaccioli lakes, Lastrucci et al. [29] reported the presence of Bolboschoenus maritimus (L.) Palla vegetation (sub Scirpetum maritimi (Christiansen 1934) R.Tuxen 1937). During this study, we could verify that the species present at this site is Bolboschoenus glaucus (Lam.) S.G.Sm., based on the taxonomic revision by Di Natale et al. [93]. Consequently, the corresponding community can be attributed to the Bolboschoenetum glauci association, already reported from the close Acquato lake in Southern Maremma [31]. The association was detected in a restricted area of the western pond (L1), but it could be favored by the summer drying and thus develop over larger areas later in the season.
Callitricho brutiae-Ranunculetum ophioglossifolii Gigante, Maneli & Venanzoni 2013 (Table S6, rel. 1-5)
Ranunculus ophioglossifolius Vill. was rather common in the study area and, at least in four sites, it was found to form dominated or co-dominated communities. Dominated communities of this species in Italy were described by Biondi et al. [94] as Trifolio fragiferi-Ranunculetum ophioglossifolii for Marche Region, and by Gigante et al. [74] as Callitricho brutiae-Ranunculetum ophioglossifolii for some peculiar small acidophilic inland temporary ponds, near Trasimeno Lake (Ferretto site, Umbria). Both the associations were included in the alliance Oenanthion fistulosae de Foucault 2009 (for nomenclatural and syntaxonomic aspects see Mucina [55]) of the class Molinio-Arrhenatheretea. In addition, Brullo et al. [66] described the association Veronico beccabungae-Ranunculetum ophioglossifolii, included in Montio-Cardaminetea class, for some running-water habitats of the Aspromonte territory (Calabria, southern Italy).
As for the communities of the study area, several floristic and ecological affinities can be found with Callitricho-Ranunculetum, both for the presence of Callitriche brutia in some of our relevés and for the hydrological dynamism of the habitats, characterized by a strong seasonality. Also, the presence of Glyceria fluitans in some more humid areas (Table S6, rel. 3-5) supports the affinity with the association Callitricho-Ranunculetum by Gigante et al. [74], representing transitional aspects towards the subassociation glycerietosum fluitantis Gigante, Maneli & Venanzoni 2013. Gigante et al. [74] observed the large variability in the syntaxonomical interpretation of the communities with Ranunculus ophioglossifolius, ranging from aquatic syntaxa of the Ranunculion aquatilis alliance, to small helophyte syntaxa of the Isoëto duriei-Juncetea bufonii or Littorelletea classes, to Glycerio-Sparganion alliance and, as already mentioned, Montio-Cardaminetea and Molinio-Arrhenatheretea classes. Concerning in particular the Callitricho brutiae-Ranunculetum ophioglossifolii, neither the growth forms, nor the habitat, nor the phytogeographical characteristics of the examined sites seem particularly congruent with its inclusion in syntaxa of the order Molinetalia ceruleae, which includes wet meadows on mineral and peaty soils in temperate to subarctic zones of Europe [55]. One relevé (rel. 5) near “Poggio Romano” (LPR), with the presence of amphibious patches of Callitriche stagnalis, was considered here as a transitional aspect between an impoverished aspect of Callitricho-Ranunculetum and the aquatic association of Ranunculo ophioglossifolii-Callitrichetum stagnalis Brullo, Scelsi & Spampinato 2001.
Molinia arundinacea-dominated communities were reported for several types of freshwater habitats, such as wet depressions along rivers [91,95,96] or lakes [77], as Molinietum arundinaceae Trinajstič 1965. For some wet areas in the ultramafic soils of the Upper Tiber Valley (eastern Tuscany), Lastrucci et al. [75]reported a variant of the association with high presence of Schoenus nigricans, showing some affinities with the association Molinio arundinaceae-Schoenetum nigricantis Rivas-Goday 1945 indicated for the Iberian Peninsula by Rivas-Martinez et al. [97]. According to Rivas-Goday [98], the variant dominated by Molinia arundinacea represents a transitional aspect between the association Molinio-Schoenetum nigricantis and the woody heath vegetation dominated by species of Calluna, Ulex and/or Erica. In the study area, stands with Molinia arundinacea and Schoenus nigricans were developed on gentle slopes surrounding the Eleocharis multicaulis vegetation near the shores of a pool (SDP) in the Mt. Leoni area. Among the other species characterizing M. arundinacea stands, some acidophilic or subacidophilic herbs such as Potentilla erecta (L.) Raeusch., Carex punctata Gaudin, Succisa pratensis Moench or Danthonia decumbens were present. In this vegetation type, the transition towards shrub acidophilic vegetation was indicated by Calluna vulgaris (L.) Hull.
Juncus effusus community (Table S6, rel. 8-9)
Juncus effusus L. was often observed in stands along the shores of lakes and ponds, forming a belt representing the border between the aquatic vegetation and the terrestrial communities surrounding the wetland area. This vegetation type was generally rich in more or less hygrophilous species, representing a transition between the marsh coenoses of the class Phragmito-Magnocaricetea and the wet meadows of the Molinio-Arrhenatheretea classes, see Lastrucci et al. [29,77,99,100]. In the study area, dense Juncus effusus-dominated coenoses were detected around the lake near the Neolithic village (LVN) of Mt. Leoni area.
According to Landucci et al. [84], when Juncus effusus communities are not particularly rich in elements of Phragmito-Magnocaricetea class, it is preferable to place them in the Molinio-Arrhenatheretea class. The same interpretation was applied for the Juncus effusus belt around the “Stagnone” lake of the Capraia Island in the Tuscan Archipelago [100].
4. Discussion and Conclusion
Our study pointed out that close field investigations, even in areas considered well known from a floristic and naturalistic point of view such as Tuscany, can bring to the light novel data or update existing information documenting the presence of rare species and plant communities of conservation interest. It is worth noting that 9 sites, out of 19, are not included in protected areas, and that, of these, only the Lagaccioli lakes were already known as of conservation relevance [29,42,101,102] The vegetation of all the other water bodies was previously unknown and examined here for the first time, though part of these are included in protected areas or Natura 2000 sites and floristic collections were made years ago by Selvi [32,103]. Considering that the habitats worthy of conservation at the European level and listed in the Habitats Directive [104,105] are mostly based on vegetation types [106,107,108,109,110,111,112,113], this study allows to recognize habitat types that occur in the study area. Recent scientific works pointed out that not all the habitats of real conservation importance in southern Europe and the Mediterranean are listed in the annexes of the Habitats Directive [84,114,115]. Among those recognized in the Directive, however, several are considered important at the continental scale, plus some others at the local one. As shown in Table S7, some of the detected communities can be attributed to the habitat “Oligotrophic to mesotrophic standing waters with vegetation of the Littorelletea uniflorae and/or Isoeto-Nanojuncetea”, some others to the habitat “Natural eutrophic lakes with Magnopotamion or Hydrocharition - type vegetation”. The Nitella-dominated communities belong to the habitat “Hard oligo-mesotrophic waters with benthic vegetation of Chara spp.), while the coenoses with Molinia arundinacea and Schoenus nigricans to the habitat “Mediterranean tall humid herb grasslands of the Molinio-Holoschoenion”. The communities that can be attributed to the habitat “Mediterranean temporary ponds” (see Table S7) are particularly relevant, as these environments, rich in rare species, are considered of priority importance for conservation at the European level [13,104].
In sum, the present study allows to advance our knowledge of the flora and vegetation of small but valuable wetland sites, providing a basis for their preservation and demonstrating that the acquisition of taxonomic and syntaxonomical knowledge based on herbarium and field surveys is still fundamental for the conservation of nature [116,117].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Appendix S1: names, abbreviations, geographical position and information about inclusion in protected areas of the investigated wetland sites; Figure S1: dendrogram resulting from the cluster analysis of aquatic plant communities; Figure S2: dendrogram resulting from the cluster analysis of palustrine plant communities; Table S1: Aquatic plant communities; Table S2: Plant communities of Isoeto-Nanojuncetea class; Table S3: Plant communities of Littorelletea class; Table S4: Plant communities of Phragmition and Magnocaricion alliances; Table S5: Plant communities of Sparganion and Oenanthion alliances; Table S6: Plant communities of Molinio-Arrhenatheretea class; Table S7: Correspondence between detected vegetation types and Natura 2000 habitats.
Author Contributions
Conceptualization, investigation, data curation, methodology, formal analysis, validation, writing—original draft preparation, writing—review and editing, L.L. and D.V.; investigation, data curation, validation, writing—review and editing, F.S., E.B. and A.S.
Funding
Federico Selvi: Daniele Viciani and Enrico Bajona acknowledge the support of the National Biodiversity Future Center to University of Florence (Italy), funded by the Italian Ministry of University and Research, PNRR, Missione 4 Componente 2, Investimento 1.4, Project CN00000033.
Data Availability Statement
Upon request to corresponding author.
Acknowledgments
We thank Lorella Dell’Olmo for preparing Figure 1.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Syntaxonomical scheme
Charetea intermediae F. Fukarek 1961
Nitelletalia W. Krause 1969
Nitellion flexilis W. Krause 1969
Nitelletum hyalinae Corillion 1957
Lemnetea O. de Bolòs et Masclans 1955
Lemnetalia minoris O. de Bolòs et Masclans 1955
Utricularion vulgaris Passarge 1964
Utricularietum australis Müller et Görs 1960
Potamogetonetea Klika in Klika et Novák 1941
Potamogetonetalia Koch 1926
Nymphaeion albae Oberd. 1957
Potamogetonetum natantis Hild 1959
Potamogetonion Libbert 1931
Potamogetonetum pusilli von Soó 1927
Parvo-Potamogetono-Zannichellietum pedicellatae De Soó 1947
Potamogeton nodosus community
Potamogeton lucens community
Callitricho hamulatae-Ranunculetalia aquatilis Passarge ex Theurillat in Theurillat et al. 2015
Ranunculion aquatilis Passarge ex Theurillat in Theurillat et al. 2015
Ranunculo ophioglossifolii-Callitrichetum stagnalis Brullo, Scelsi & Spampinato 2001
subass. typicum
subass. ranunculetosum peltati subass. nova
Callitricho brutiae-Ranunculetum peltati Pizarro & Rivas-Martinez 2002 subass. ranunculetosum trichophylli Lastrucci, Foggi, Selvi & Becattini 2007
Potamo crispi-Ranunculetum trichophylli Imchenetzky 1926
Lemno-Callitrichetum obtusangulae (Philippi 1978) Passarge 1992
Callitriche platycarpa community
Isoëto duriei-Juncetea bufonii Br.-Bl. & Tüxen ex Westhoff, Dijk & Paschier 1946
Isoëtetalia duriei Br.-Bl. 1936
Damasonium alisma community
Cicendio filiformis-Solenopsion laurentiae Brullo & Minissale 1998
Junco tenageiae-Solenopsietum laurentiae Gigante, Maneli & Venanzoni 2013
Nanocyperetalia Klika 1935
Nanocyperion Koch 1926
Isolepis cernua community
Eleocharition soloniensis Philippi 1968
Peplido portulae-Ludwigietum palustris Robbe ex J.-M. Royer, Felzines, Misset & Thévenin 2006
Littorelletea uniflorae Br.-Bl. et Tüxen ex Westhoff, Dijk & Passchier 1946
Littorelletalia uniflorae Koch ex Tx. 1937
Littorellion uniflorae Koch ex Klika 1935
Callitricho brutiae-Juncetum bulbosi Gigante, Maneli & Venanzoni 2013
Hyperico elodis-Sparganion Br.-Bl. et Tx. ex Oberd. 1957
Junco bulbosi-Eleocharitetum multicaulis (Passarge 1955) Passarge 1999
Phragmito-Magnocaricetea Klika in Klika et Novák 1941
Phragmitetalia Koch 1926
Phragmition australis Koch 1926 nom. corr.
Phragmitetum australis Savič 1926 nom. corr.
Magnocaricetalia Pignatti 1953
Magnocaricion elatae Koch 1926
Lysimachia vulgaris community
Nasturtio-Glycerietalia Pignatti 1953
Glycerio-Sparganion Br.-Bl. et Sissingh in Boer 1942
Glycerio-Sparganietum neglecti Koch 1926
Glycerietum fluitantis Nowinski 1930 nom. inval.
Alismo lanceolatae-Gratioletum officinalis Biondi & Bagella 2005
Oenanthetalia aquaticae Hejný ex Balátová-Tuláčková et al. 1993
Eleocharito palustris-Sagittarion sagittifoliae Passarge 1964
Eleocharitetum palustris Savič 1926
Oenantho aquaticae-Rorippetum amphibiae Lohmeyer 1950
var. with Persicaria amphibia
Bolboschoenetum glauci Grechushkina, Sorokin & Golub 2011
Molinio-Arrhenatheretea Tx. 1937
Molinietalia caeruleae Koch 1926
Oenanthion fistulosae de Foucault 2009
Callitricho brutiae-Ranunculetum ophioglossifolii Gigante, Maneli & Venanzoni 2013
ranunculetosum ophioglossifolii Gigante, Maneli & Venanzoni 2013
glycerietosum fluitantis Gigante, Maneli & Venanzoni 2013
Holoschoenetalia Br.-Bl. ex Tchou 1948
Molinio-Holoschoenion Br.-Bl. ex Tchou 1948
Juncus effusus community
Molinietum arundinaceae Trinajstič 1965 var. with Schoenus nigricans
References
- Dudgeon, D.; Arthington A.H., Gessner M.O.; Kawabata, Z., Knowler, D.J.; Lévêque, C., et al. Freshwater biodiversity: importance, threats, status and conservation challenges. Biological Reviews 2006, 81, 163–182. [CrossRef]
- De Groot, R.; Brander, L.; Van Der Ploeg, S.; Costanza, R.; Bernard, F.; Braat, L.; Christie, M.; Crossman, N.; Ghermandi, A.; Hein, L.; et al. Global estimates of the value of ecosystems and their services in monetary units. Ecosyst Serv. 2012, 1, 50–61. [CrossRef]
- Zhang, L.L.; Yin, J.X.; Jiang, Y.Z.; Wang, H. Relationship between the hydrological conditions and the distribution of vegetation communities within the Poyang lake national nature reserve, China. Ecological Informatics 2012, 11, 65–75. [CrossRef]
- Mitsch, W.J.; Bernal, B.; & Hernandez, M.E. Ecosystem services of wetlands. International Journal of Biodiversity Science, Ecosystem Services & Management 2015, 11(1), 1-4. [CrossRef]
- Gardner, R.C.; Barchiesi, S.; Beltrame, C.; Finlayson, C.M.; Galewski, T.; Harrison, I.; Paganini, M.; Perennou, C.; Pritchard, D.E.; Rosenqvist, A.; Walpole, M. State of the World’s Wetlands and their Services to People: A compilation of recent analyses. Ramsar Briefing Note no. 7; Switzerland: Ramsar Convention Secretariat, Gland, Swiss, 2015.
- Acreman, M.; Hughes, K. A.; Arthington, A. H.; Tickner, D.; Dueñas, M. A. Protected areas and freshwater biodiversity: A novel systematic review distils eight lessons for effective conservation. Conservation Letters 2020, 13(1). [CrossRef]
- Hrivnák, R.; Kochjarová, J.; Oťaheľová, H.; Paľove-Balang, P.; Slezák, M.; Slezák, P. Environmental drivers of macrophyte species richness in artificial and natural aquatic water bodies–comparative approach from two central European regions. Annales de Limnologie-International Journal of Limnology 2014, 50(4): 269–278. [CrossRef]
- Taylor, N.G.; Grillas, P.; Al Hreisha, H.; Balkız, Ö.; Borie, M.; Boutron, O.; ... & Sutherland, W.J. The future for Mediterranean wetlands: 50 key issues and 50 important conservation research questions. Regional environmental change 2021, 21, 1-17. [CrossRef]
- Ballut-Dajud, G.A.; Sandoval Herazo, L.C.; Fernández-Lambert, G.; Marín-Muñiz, J.L.; López Méndez, M.C.; Betanzo-Torres, E. A. Factors affecting wetland loss: A review. Land 2022, 11(3), 434. [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biological Reviews 2019, 94(3), 849–873. [CrossRef]
- Fluet-Chouinard, E.; Stocker, B.D.; Zhang, Z.; Malhotra, A.; Melton, J.R.; Poulter, B.; ... & McIntyre, P.B. Extensive global wetland loss over the past three centuries. Nature 2023, 614(7947), 281-286. [CrossRef]
- Xu, T.; Weng, B.; Yan, D.; Wang, K.; Li, X.; Bi, W.; ... & Liu, Y. Wetlands of international importance: Status, threats and future protection. International journal of environmental research and public health 2019, 16(10), 1818. [CrossRef]
- Janssen, J.S., Rodwell, M.; Criado S.; Gubbay, T.; Haynes, A.; Nieto, N.; et al. European Red List of Habitats. Publications Office of European Union: Luxembourg, 2016. ISBN 978-92-79-61588-7. [CrossRef]
- Zivkovic, L.; Biondi, E.; Pesaresi, S.; Lasen, C.; Spampinato, G.; Angelini, P. The third report on the conservation status of habitats (Directive 92/43/EEC) in Italy: processes, methodologies, results and comments. Plant Sociology 2017 54(2), 51–64. [CrossRef]
- Gigante, D.; Acosta, A.T.R.; Agrillo, E.; Armiraglio, S.; Assini, S.; Attorre, F. et al. Habitat conservation in Italy: the state of the art in the light of the first European Red List of Terrestrial and Freshwater Habitats. Rendiconti Lincei. Scienze Fisiche e Naturali 2018, 29(2): 251–265. [CrossRef]
- Lazzaro, L.; Bolpagni, R.; Buffa, G.; Gentili, R.; Lonati, M.; Stinca, A. et al. Impact of invasive alien plants on native plant communities and Natura 2000 Habitats: state of the art, gap analysis and perspectives in Italy. Journal of Environmental Management 2020, 274, 111140 . [CrossRef]
- Viciani, D.; Vidali, M.; Gigante, D.; Bolpagni, R.; Villani, M.; Acosta, A.T.R. et al. A first checklist of the alien-dominated vegetation in Italy. Plant Sociology 2020, 57(1), 29–54. [CrossRef]
- Gennai, M.; Gabellini, A.; Viciani, D.; Venanzoni, R.; Dell’Olmo, L.; Giunti, M. et al. The floodplain woods of Tuscany: towards a phytosociological synthesis. Plant Sociology 2021, 58(1): 1–28. [CrossRef]
- Van den Broeck, M.; Waterkeyn, A.; Rhazi, L.; Grillas, P. & Brendonck, L. Assessing the ecological integrity of endorheic wetlands, with focus on Mediterranean temporary ponds. Ecological Indicators 2015, 54; 1-11. [CrossRef]
- Geijzendorffer, I.R.; Beltrame, C.; Chazee, L.; Gaget, E.; Galewski, T.; Guelmami, A.; ... & Grillas, P. A more effective Ramsar Convention for the conservation of Mediterranean wetlands. Frontiers in Ecology and Evolution 2019, 7, 21. [CrossRef]
- Fois, M.; Cuena-Lombraña, A. & Bacchetta, G. Knowledge gaps and challenges for conservation of Mediterranean wetlands: Evidence from a comprehensive inventory and literature analysis for Sardinia. Aquatic Conservation: Marine and Freshwater Ecosystems 2021, 31(9), 2621-2631. [CrossRef]
- Leberger, R.; Geijzendorffer, I.R.; Gaget, E.; Gwelmami, A.; Galewski, T.; Pereira, H.M.; Guerra, C.A. Mediterranean wetland conservation in the context of climate and land cover change. Regional Environmental Change 2020, 20, 1-11. [CrossRef]
- Gallego-Fernandez, J.B.; García-Mora, M.R.; Garcia-Novo, F. Small wetlands lost: a biological conservation hazard in Mediterranean landscapes. Environmental Conservation 1999, 26(3), 190-199.
- De Meester, L.; Declerck, S.; Stoks, R.; Louette, G.; Van De Meutter, F.; De Bie, T.; ... & Brendonck, L. Ponds and pools as model systems in conservation biology, ecology and evolutionary biology. Aquatic conservation: Marine and freshwater ecosystems 2005, 15(6), 715-725. [CrossRef]
- Angiolini, C.; Viciani, D.; Bonari, G.; Zoccola, A.; Bottacci, A.; Ciampelli, P. et al. Environmental drivers of plant assemblages: are there differences between palustrine and lacustrine wetlands? A case study from the northern Apennines (Italy). Knowledge and Management of Aquatic Ecosystems 2019, 420, 34. [CrossRef]
- Viciani, D.; Angiolini, C.; Bonari, G.; Bottacci, A.; Dell’Olmo, L.; Gonnelli, V.; Zoccola, A.; Lastrucci L. Contribution to the knowledge of aquatic vegetation of montane and submontane areas of Northern Apennines (Italy). Plant Sociology 2022, 59(1), 25–35. [CrossRef]
- Lastrucci, L.; Angiolini, C.; Bonari, G.; Bottacci, A.; Gonnelli, V.; Zoccola, A.; Mugnai, M.; Viciani, D.; Contribution to the knowledge of marsh vegetation of montane and submontane areas of Northern Apennines (Italy). Plant Sociology 2023, 60(1), 25–36. DOI 10.3897/pls2023601/03.
- Perennou, C.; Gaget, E.; Galewski, T.; Geijzendorffer, I.; Guelmami, A. Evolution of wetlands in Mediterranean region. Water resources in the Mediterranean Region 2020, 297-320. [CrossRef]
- Lastrucci, L.; Foggi, B.; Selvi, F.; Becattini, R. Contributo alla conoscenza della vegetazione e della flora delle aree umide nel comprensorio di Capalbio (Provincia di Grosseto, Italia centrale). Archivio Geobotanico 2007, 10(1-2), 1-30.
- Lastrucci, L; Bonari, G; Angiolini, C; Casini, F; Giallonardo, T; Gigante, D. et al. Vegetation of Lakes Chiusi and Montepulciano (Siena, central Italy): updated knowledge and new discoveries. Plant Sociology 2014, 51(2), 29–55. [CrossRef]
- Lastrucci, L.; Ferretti, G.; Mantarano, N.; Foggi, B. Vegetation and habitat of conservation interest of the lake Acquato (Grosseto – Central Italy). Plant Sociology 2019, 56(1), 19–30. [CrossRef]
- Selvi, F. A critical checklist of the vascular flora of Tuscan Maremma (Grosseto province, Italy). Flora Mediterranea 2010, 20, 47-139.
- Giovacchini, P.; Falchi, V.; Vignali, S.; Radi, G.; Passalacqua, L.; Corsi, F.; Porciani, M.; Farsi, F. Atlante degli anfibi della provincia di Grosseto (2003-2013) Quaderni Volume n. 6 della Collana “Quaderni delle Aree Protette”; Provincia di Grosseto, 2015.
- Thornthwaite, C.W.; Mather, J. R. Instruction and tables for computing potential evapotraspiration and the water balance. Pubbl. Climatol. 1957, 10 (3): 1-311.
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. Journal of Maps 2017, 13, 955–960. [CrossRef]
- Blasi, C.; Capotorti, G.; Copiz, R.; Guida, D.; Mollo, B.; Smiraglia, D.; Zavattero, L. Classification and mapping of the ecoregions of Italy. Plant Biosyst. 2014, 148(6), 1255–1345. [CrossRef]
- Gelmini, R. Ricerche geologiche nel Gruppo di M. Leoni (Grosseto; Toscana). I) La geologia di M. Leoni tra Montepescali e il fiume Ombrone. Mem. Soc. Geol. It. 1969, 8, 755-796.
- Lazzarotto, A. Elementi di Geologia. In: Giusti, F. (ed) La Storia Naturale della Toscana Meridionale; Pizzi editore: Siena, Italy, 1993; pp. 19-87.
- Aldinucci, M.; Brogi, A.; Sandrelli, F. The metamorphic units of the eastern side of Monte Leoni (Northern Apennines, Italy). Boll. Soc. Geol. It. 2005, 124, 313-332.
- Carmignani, L.; Conti, P.; Cornamusini, G.; Pirro, A. Geological map of Tuscany (Italy). Journal of Maps 2013, 9, 487–497. [CrossRef]
- Motta, S. Note Illustrative della carta geologica d’Italia; foglio 128 Grosseto; Servizio Geologico d’Italia, 1969.
- Selvi, F.; Stefanini, P.; Biotopi naturali e aree protette nella Provincia di Grosseto. Componenti floristiche e ambienti vegetazionali. Città di Castello; TipoLitografia Petruzzi Provincia di Grosseto: Città di Castello, Italy, 2006.
- Braun-Blanquet, J. Plant sociology: The study of plant communities; McGraw-Hill: New York, United States, 1932.
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde; Springer, 3rd ed.: Wien, Austria, 1964; pp. 1–330.
- Dengler, J.; Berg, C.; Jansen, F. New ideas for modern phytosociological monographs. Ann. Bot. (Roma) 2005, 5, 193–210.
- Dengler, J.; Chytrý, M.; Ewald, J. Phytosociology in Jørgensen, S.E.; Fath, B.D.; Encyclopedia of Ecology, Elsevier: Oxford, UK, 2008; pp. 2767–2779.
- Biondi, E. Phytosociology today: Methodological and conceptual evolution. Plant Biosyst. 2011, 145 (Suppl. S1), 19–29. [CrossRef]
- Pott, R. Phytosociology: A modern geobotanical method. Plant Biosyst. 2011, 145 (Suppl. S1), 9–18. [CrossRef]
- Van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114.
- Arrigoni, P.V. A classification of plant growth forms applicable to the floras and vegetation types of Italy. Webbia 1996, 50(2), 193-203.
- R Core Team -2024 R: A language and environment for statistical computing. R Foundation for Statistical Computing: Vienna; Austria. URL https://www.R-project.org/.
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre P.; McGlinn D. et al. 2024 Vegan: Community Ecology Package. R package version 2.6-6.1. https://CRAN.R-project.org/package=vegan.
- Portal to the Flora of Italy 2024 [accessed 2024 Sep] http://dryades.units.it/floritaly.
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Alessandrini, A.; Ardenghi, N.M.G.; Bacchetta, G.; Banfi, E.; Barberis, G.; Bernardo, L.; Bouvet, D.; Bovio, M.; Calvia, G.; Castello, M.; Cecchi, L.; Del Guacchio, E.; Domina, G.; Fascetti, S.; Gallo, L.; Gottschlich, G.; Guarino, R.; Gubellini, L.; Hofmann, N.; Iberite, M.; Jiménez-Mejías, P.; Longo, D.; Marchetti, D.; Martini, F.; Masin, R.R.; Medagli, P.; Peccenini, S. et al. A second update to the checklist of the vascular flora native to Italy. Plant Biosyst. 2024, 158(2), 219–296. [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A. et al. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Applied Vegetation Science 2016, 19(suppl. 1), 3–264. [CrossRef]
- Biondi, E.; Blasi, C. 2015 Prodromo della Vegetazione Italiana. [accessed 2024 Sept] http://www.prodromo-vegetazione-italia.org/.
- Theurillat, J.P.; Willner, W.; Fernández-González, F.; Bültmann, H.; Čarni, A.; Gigante, D. et al. International Code of Phytosociological Nomenclature. 4th edition. Applied Vegetation Science 2021, 24, e12491. [CrossRef]
- Bazzichelli, G.; Abdelahad, N.; Alghe d’acqua dolce d’Italia. Flora analitica delle Caroficee - Ministero dell’Ambiente, Università degli Studi di Roma La Sapienza; Centro Stampa Università: Rome, Italy, 2009.
- Mangeat, M.; Contribution à la connaissance de la Characée Nitella hyalina (De Candolle) C. Agardh, 1824, dans le nord-est de la France. Les Nouvelles Archives de la Flore jurassienne et du nord-est de la France 2014, 12, 48-61.
- Csiky, J.; Purger, D.; Blaženčić, J. New occurrence and distribution of Nitella hyalina (DC .) Agardh (Characeae) and the first report on Nitelletum hyalinae Corillion 1957, in Croatia. Arch. Biol. Sci. 2014, 66(1), 203-208. [CrossRef]
- Šumberová, K. Vegetace volně plovoucích vodních rostlin (Lemnetea). Vegetation of free floating aquatic plants. In: Chytrý M. Ed.); Vegetace České republiky 3 Vodní a mokřadní vegetace. [Vegetation of the Czech Republic 3 Aquatic and wetland vegetation]; Academia: Praha, Czech Republic, 2011; pp. 43–99.
- Šumberová K. Vegetace vodních rostlin zakořeněných ve dně (Potametea). Vegetation of aquatic plants rooted in the bottom. In Chytrý M. (Ed.). Vegetace České republiky 3 Vodní a mokřadní vegetace. [Vegetation of the Czech Republic 3 Aquatic and wetland vegetation]. Academia: Praha, Czech Republic, 2011; 100-247.
- Hrivnák, R.; Aquatic plant communities in the catchment area of the Ipel' river in Slovakia and Hungary. Part II. Class Potametea. Thaiszia 2002, 12, 137-160.
- Lastrucci, L.; Landucci, F.; Gonnelli, V.; Barocco, R.; Foggi, B.; Venanzoni, R. The vegetation of the upper and middle River Tiber (Central Italy). Plant Sociology 2012, 49(2), 29-48. DOI: 10.7338/pls2012492/02.
- Landucci, F.; Gigante, D.; Venanzoni, R. An application of the Cocktail method for the classification of the hydrophytic vegetation at Lake Trasimeno (Central Italy). Fitosociologia 2011, 48(2), 3-22.
- Brullo, S.; Scelsi, F.; Spampinato, F. La vegetazione dell’Aspromonte. Laruffa Editore: Reggio Calabria, Italy, 2001.
- Tardella, F.M.; Di Agostino, V.M. Vegetation of the "Altipiani di Colfiorito" wetlands (central Apennines, Italy). Plant Sociology 2020, 57(2), 113-132. [CrossRef]
- Sburlino, G.; Tomasella, M.; Oriolo, G.; Poldini, L.; Bracco, F. La vegetazione acquatica e palustre dell'Italia nord-orientale. 2 – La classe Potametea Klika in Klika et V. Novák 1941. Fitosociologia 2008, 45(2), 3-40.
- Caldarella, O.; Lastrucci, L.; Bolpagni, R.; Gianguzzi, L. Contribution to the knowledge of Mediterranean wetland vegetation: Lemnetea and Potamogetonetea classes in Western Sicily. Plant Sociology 2021, 58(1), 107–131. [CrossRef]
- Felzines, J.C.; Contribution au prodrome des végétations de France: les Potametea Klika in Klika & V. Novák 1941. Doc. Phytosoc 2016, série 3, 3, 218-437.
- Passarge, H.; Mitteleuropäische Potamogetonetea I. Phytocoenologia 1992, 20(4), 489-527.
- Lastrucci, L.; Saiani, D.; Mugnai, A.; Ferretti, G., Viciani, D. Distribution novelties of the genus Callitriche (Plantaginaceae) in Italy from the study of the Herbarium Centrale Italicum collections. Mediterr. Bot. 2024, 45(2); e87474. [CrossRef]
- Brullo, S., Minissale, P. Considerazioni syntassonomiche sulla classe Isoëto-Nanojuncetea. Itinera Geobotanica 1998, 11, 263–290.
- Gigante, D.; Maneli, F.; Venanzoni, R. Mediterranean temporary wet systems in inland Central Italy: ecological and phytosociological features. Plant Sociology 2013, 50(2), 93-112. [CrossRef]
- Lastrucci, L.; Foggi, B.; Gonnelli, V.; Gusmeroli, E.; La vegetazione delle aree umide dei substrati ultramafici dell’Alta Valtiberina (Arezzo; Italia centrale). Studia Botanica 2006, 24 (2005), 9-44.
- Rivas-Martínez, S.; Costa, M.; Castroviejo, S.; Valdés, E. Vegetación de Doñana (Huelva, España). Lazaroa 1980, 2, 5-189.
- Lastrucci, L.; Viciani, D.; Nuccio, C.; Melillo, C. Indagine vegetazionale su alcuni laghi di origine artificiale limitrofi al Padule di Fucecchio (Toscana, Italia Centrale). Ann. Mus. Civ. Rovereto 2008, 23(2007), 169–203.
- Dítě, D.; Eliáš, P. Jr.; Dítě, Z.; Šimková, A. Recent distribution and phytosociological affiliation of Ludwigia palustris in Slovakia. Acta Soc Bot Pol. 2017, 86(1), 3544. [CrossRef]
- De Foucault, B.; Contribution au prodrome des végétations de France: les Littorelletea uniflorae Braun-Blanq. & Tüxen ex Westhoff, Dijk, Passchier & Sissingh 1946. J. Bot. Soc. Bot. France 2010, 52, 43-78.
- Wattez, J.R.; Gehu, J.M. Groupements amphibies acidoclines relictuels ou disparus du nordde la France. Doc. Phytosociol. 1982, N.S. 6, 263-278.
- Dierssen, K.; Die Vegetation des Gildehauser Venns (Kreis Grafschaft Bentheim). Beih. Ber. Naturhist. Ges. 1973, 8, 1-120.
- Passarge, H. Pflanzengesellschaften Norostdeutschlands 2 II. Helocyperosa uns Caespitosa. J. Cramer in der Gebrüder Borntraeger Verlagsbuchhandlung: Berlin, Stuttgart, Germany, 1999; 451 pp.
- Landucci, F.; Gigante, D.; Venanzoni, R.; Chytrý, M. Wetland vegetation of the class Phragmito-Magno-Caricetea in central Italy. Phytocoenologia, 2013, 43, 67–100. [CrossRef]
- Landucci, F.; Šumberová, K.; Tichý, L.; Hennekens, S.; Aunina, L.; Biță-Nicolae, C. et al. Classification of the European marsh vegetation (Phragmito-Magnocaricetea) to the association level. Applied Vegetation Science 2020, 23, 297–316. [CrossRef]
- Lastrucci, L.; Lazzaro, L.; Coppi, A.; Foggi, B.; Ferranti, F.;Venanzoni, R.; Cerri, M.; Ferri, V.; Gigante, D.; Reale, L. Demographic and macro-morphological evidence for common reed dieback in central Italy. Plant Ecology & Diversity 2017, 10, 2-3, 241-251. [CrossRef]
- Lastrucci, L.; Cerri, M.; Coppi, A.; Ferranti, F.; Ferri, V.; Foggi, B. et al. Understanding common reed die-back: a phytocoenotic approach to explore the decline of palustrine ecosystems. PlantnSociology 2017, 54(2) Suppl.1, 15–28. [CrossRef]
- Venanzoni, R.; Properzi, A.; Bricchi, E.; Landucci, F.; Gigante, D. The Magnocaricetalia Pignatti 1953 (Phragmito-Magnocaricetea Klika in Klika et Novák 1941) Plant Communities of Italy. In: Pedrotti, F. (ed.) Climate Gradients and Biodiversity in Mountains of Italy, Geobotany Studies. Springer International Publishing, 2018, 135-173.
- Lastrucci, L.; Gambirasio, V.; Prosser, F.; Viciani, D. First record of Sparganium oocarpum in Italy and new regional distribution data for S. erectum species complex. Plant Biosyst. 2024, 158 (4). [CrossRef]
- Stančić, Z. Marshland vegetation of the class Phragmito-Magnocaricetea in Croatia. Biologia 2007, 62, 297-314. [CrossRef]
- Biondi, E.; Bagella, S. Vegetazione e paesaggio vegetale dell’arcipelago di La Maddalena (Sardegna nord-orientale). Fitosociologia 2005, 42(2) suppl.1, 3-99.
- Venanzoni, R., Gigante, D. Contributo alla conoscenza della vegetazione degli ambienti umidi dell’Umbria (Italia). Fitosociologia 2000, 37 (2), 13–63.
- Hrivnák, R.; Spoločenstvá zväzu Oenanthion aquaticae v povodí rieky Ipeľ [The plant communities of Oenanthion aquaticae in the catchment area of the river Ipeľ]. Bull. Slov. Bot. Spoločn 2003, 25, 169 – 183.
- Di Natale, S.; Lastrucci, L.; Hroudova, Z.; Viciani D. A review of Bolboschoenus species (Cyperaceae) in Italy based on herbarium data. Plant Biosyst. 2022, 156(1), 261–270. [CrossRef]
- Biondi, E.; Casavecchia, S.; Raketic, Z. The guazzi vegetation and the plant landscape of the alluvial plane of the last stretch of the Musone River (Central Italy). Fitosociologia 2002, 39(1), 45-70.
- Biondi, E.; Baldoni, M.; La vegetazione del fiume Marecchia (Italia Centrale). Biogeographia 1994, 17, 51-87.
- Biondi, E.; Vagge, I.; Baldoni, M.; Taffetani, F. La vegetazione del Parco Fluviale Regionale del Taro (Emilia Romagna). Fitosociologia 1997, 34, 69-110.
- Rivas-Martínez, S.; Fernández–González; F., Loidi, J.; Lousã, M.; Penas, Á. Syntaxonomical Checklist of vascular plant communities of Spain and Portugal to association level. Itinera Geobotanica 2001, 14, 5-341.
- Rivas-Goday, S. Contribución al conocimiento del Schoenetum nigricantis de Vasconia. Bol. Real Soc. Esp. Hist. Nat. 1945, 43, 261-273.
- Lastrucci, L.; Paci, F.; Raffaelli, M. The wetland vegetation of the Natural Reserves and neighbouring stretches of the Arno river in the Arezzo province (Tuscany; Central Italy). Fitosociologia 2010, 47(1), 29–59.
- Lastrucci, L.; Foggi, B.; Mantarano, N.; Ferretti, G.; Calamassi, R.; Grigioni, A. La vegetazione del laghetto “Lo Stagnone” (Isola di Capraia; Toscana). Atti Soc. Tosc. Sci. Nat.; Mem. Ser. B 2010, 116 (2009), 17–25.
- Tomei, P.E.; Guazzi, E. Le zone umide della Toscana; lista generale delle entità vegetali. Atti Mus. Civ. St. Nat. Grosseto 1993, 15, 107-152.
- Tomei, P.E.; Guazzi, E.; Kugler, P.C. Le zone umide della Toscana: indagine sulle componenti floristiche e vegetazionali. Ed. Regione Toscana: Firenze, Italy, 2001; pp. 167.
- Selvi, F. Flora vascolare del Monte Leoni (Toscana Meridionale). Webbia 1998, 52, 265-306.
- European Commission -1992 Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off J Eur Union 206, 7–50.
- European Commission -2013 Interpretation Manual of European Union Habitats - Version EUR 28; April 2013 European Commission DG-ENV; Brussels; 146 pp.
- Evans, D. The habitats of the European Union Habitats Directive. Biology and Environment. Proceedings of the Royal Irish Academy 2006, 106, 167–173.
- Evans, D. Interpreting the habitats of Annex I. Past, present and future. Acta Botanica Gallica 2010, 157, 677–686.
- Biondi, E.; Blasi, C.; Burrascano, S.; Casavecchia, S.; Copiz, R.; Del Vico, E. et al. Manuale Italiano di interpretazione degli habitat della Direttiva 92/43/CEE. Società Botanica Italiana. Ministero dell’Ambiente e della Tutela del Territorio e del Mare, 2009; http://vnr.unipg.it/habitat.
- Biondi, E.; Burrascano, S.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Galdenzi, D. et al. Diagnosis and syntaxonomic interpretation of Annex I Habitats (Dir. 92/43/EEC) in Italy at the alliance level. Plant Sociology 2012, 49(1), 5-37. [CrossRef]
- Bunce, R.G.H.; Bogers, M.M.B.; Evans, D.; Halada, L.; Jongman, R.H.G.; Mücher, C.A.;…Olsvig-Whittaker, L. The significance of habitats as indicators of biodiversity and their links to species. Ecological Indicators 2013, 33, 19–25. [CrossRef]
- Gigante, D.; Foggi, B.; Venanzoni, R.; Viciani, D.; Buffa, G. Habitats on the grid: The spatial dimension does matter for red-listing. Journal for Nature Conservation 2016, 32, 1–9. [CrossRef]
- Viciani, D.; Dell’Olmo, L.; Ferretti, G.; Lazzaro, L.; Lastrucci, L.; Foggi, B. Detailed Natura 2000 and Corine Biotopes habitat maps of the island of Elba (Tuscan Archipelago; Italy). Journal of Maps 2016, 12(3), 492–502. [CrossRef]
- Viciani, D.; Dell’Olmo, L.; Foggi, B.; Ferretti, G.; Lastrucci, L. & Gennai, M. Natura 2000 habitat of Mt. Argentario promontory (southern Tuscany; Italy). Journal of Maps 2018, 14(2), 447-454. [CrossRef]
- Angiolini, C.; Viciani, D.; Bonari, G.; Lastrucci, L. Habitat conservation prioritization: A floristic approach applied to a Mediterranean wetland network. Plant Biosyst. 2017, 151(4), 598–612. [CrossRef]
- Casavecchia, S.; Allegrezza. M.; Angiolini, C.; Biondi, E.; Bonini, F.; Del Vico, E. et al. Proposals for improvement of Annex I of Directive 92/43/EEC: Central Italy. Plant Sociology 2021, 58(2), 99–118. [CrossRef]
- Eckert, I.; Bruneau, A.; Metsger, D.A. et al. Herbarium collections remain essential in the age of community science. Nat. Commun. 2024, 15, 7586. [CrossRef]
- Liu, D. World view. Nature, 2024, 633, 741 https://www.nature.com/articles/d41586-024-03072-3.
Figure 1.
The location of study area and the distribution of the 19 study sites are showed.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



