
Submitted:
01 August 2025
Posted:
05 August 2025
You are already at the latest version
Abstract
Short-term synaptic plasticity (STSP) and short-term neuronal dynamics (STND) are fundamental properties of neural circuits, essential for information processing and brain function. Emerging evidence suggests biological sex may influence these properties, yet sex-related differences in STSP and STND remain underexplored. This study investigates sex-specific differences in short-term synaptic plasticity (STSP) and neuronal dynamics (STND) along the dorsoventral axis of the rat hippocampus. Our findings reveal that both STSP and STND exhibit significant variation between female and male subjects. These differences are particularly pronounced in the ventral hippocampus, a region associated with affective and motivational processes. Given the role of short-term activity-dependent neuronal phenomena in modulating information processing and network function, these findings suggest potential functional implications for sex-specific cognitive and emotional regulation. The results highlight the importance of incorporating sex as a biological variable in studies of hippocampal physiology and its relation to behavior.
Keywords:
short-term synaptic plasticity
; short-term dynamics
; synaptic transmission
; excitability
; gender
; sex
; septotemporal
; dorsoventral
; hippocampus
; rat
1. Introduction
Short-term synaptic plasticity (STSP) refers to transient forms of activity-dependent changes in synaptic strength enabling several types of real-time information processing including frequency-dependent filtering, gain control, and adaptive responses to sensory inputs [1,2,3,4,5]. STSP is proposed to be crucial in brain functions such as working memory and decision making [6,7,8,9]. Despite its significance, the majority of existing studies on STSP have been conducted predominantly in male subjects, with limited consideration of female-specific mechanisms, and experimental results obtained from males could not be applicable to females [10,11,12].
However, emerging evidence reveals sex-specific differences in synaptic plasticity mechanisms at various levels of neural organization; for reviews see [11,12,13,14,15,16]. For instance, variations in the ability for long-term synaptic potentiation have been shown during the estrous cycle [17,18,19]. Furthermore, estradiol lowers the threshold for the induction of LTP [20,21] and enhances the NMDA receptor-dependent LTP at CA1 hippocampal synapses [22,23]. Also, the magnitude of hippocampal long-term potentiation has been found higher in male compared with female animals [24,25,26]. Even when a similar magnitude of LTP can be reached in females and males this is mediated by different molecular mechanisms. cAMP-regulated protein kinase is required for LTP in females but not males, and both L-type calcium channels and internal calcium stores appear to be required in females but not males [27]. Interestingly, region-specific sex-related differences have been reported in the hippocampus with the ventral hippocampus of female rats expressing higher levels of estrogen receptors than the dorsal hippocampus [28,29]. These findings could suggest that STSP dynamics may also diverge between males and females in ways that impact neural computations. Indeed, sex-related differences have described at the molecular, synaptic, and cellular levels that could impact on STSP and behavior. For instance, the nucleus accumbens of females displays larger readily releasable neurotransmitter pools compared to males [30], and medial prefrontal synapses display higher transmitter release probability in females than males [31]. These characteristics are expected to have a strong impact on STSP properties [1,2]. In addition, estrogens can influence spine density and dendritic arborization that normally fluctuate across the estrous cycle [32,33,34,35,36], thereby shaping STSP properties by altering electrotonic properties and synaptic integration [37,38].
In addition to STSP, which reflects activity-dependent changes at the synaptic level, the dynamics of neuronal firing, referred to as short-term neuronal dynamics (STND), play a critical role in shaping the output of neuronal circuits and thus the overall computational properties of brain networks [39,40,41,42,43]. Thus, STND represents rapid, transient changes in neuronal excitability and firing output that shape how local brain circuits respond to repeated or patterned input. It has been recently shown that STND in the hippocampal CA1 field while partially influenced by STSP, can be determined independently by network-level factors such as synaptic inhibition [44] and regional modulation by neuromodulators [44,45,46,47] supporting the idea that these dynamics are an emergent network property, not just a direct reflection of synaptic function.
Therefore, the dorsal and ventral hippocampus exhibit markedly distinct STND profiles [44], and age-related changes [48], suggesting functional specializations along the septotemporal axis and lifespan that contribute to localized information processing properties within the CA1 field. These dynamics are particularly relevant for short-term encoding of temporal information, and may serve to optimize gain control, signal amplification, and temporal filtering of input in hippocampal networks [41,42]. Furthermore, STND can modulate how inputs are transformed into output patterns over short timescales [43], especially during high-frequency oscillatory activity such as theta and gamma rhythms which are prominent in the hippocampus supporting learning and memory. Alterations in normal STND may have important implications for brain disorders. For instance, abnormal short-term dynamics have been implicated in neuropsychiatric diseases such as autism spectrum disorder and schizophrenia [49,50,51], where they may underlie symptoms like working memory dysfunction and impaired temporal filtering of information.
However, whether STND follows sex-related diversification is not known. Given the hippocampus’ key role in emotion and cognition, and the known sex-related differences in affective and cognitive behaviors, investigating STND in both sexes could reveal important physiological substrates for such differences. In addition, alterations in short-term dynamics have been linked to neuropsychiatric disorders, which often exhibit sex-biased prevalence and symptomatology. Therefore, knowing whether and how STND differs between females and males is crucial to understand sex-related specializations in information processing and may help explain sex-specific vulnerabilities to brain disorders.
In this study, we aimed to investigate sex-specific differences in STSP and STND across the dorsoventral axis of the rat hippocampus, recording field excitatory synaptic potentials (fEPSPs) and population spikes (PSs) from the CA1 field. We hypothesized that males and females would differ in both forms of short-term plasticity, and that these differences would vary with hippocampal region, reflecting distinct contributions to cognitive versus emotional processing which are associated with the dorsal and ventral hippocampus, respectively [52,53]. By examining frequency-dependent modulation of synaptic and neuronal responses, we sought to uncover sex- and region-dependent properties that may underlie differential hippocampal function and behavioral outcomes. Our results reveal significant sex-dependent and region-specific differences in short-term neuronal dynamics. These findings have important implications for understanding sex-specific mechanisms of information processing and broader brain functions.
2. Results
2.1. Similar Synaptic Transmission and Neuronal Excitability in Female amd Male Rats
We compared basal excitatory synaptic transmission and neuronal excitability between female and male rats by constructing input-output (I-O) curves relating stimulation current intensity to fEPSP or PS, as well as between fEPSP and PS (Figure 1). We found that the average fEPSP recorded from either dorsal or ventral hippocampus did not significantly differ between female and male rats (Figure 1A-B). Furthermore, we examined the excitation between the two sexes in both dorsal and ventral hippocampus. Similarly, our analysis revealed no significant effect of sex on PS either in the dorsal or the ventral hippocampus (Figure 1C-D). In addition, neuronal excitability, assessed by the PS/fEPSP ratio was found similar in female and male dorsal and ventral hippocampus (Figure 1E-F).
2.2. Short-Term Synaptic Plasticity (STSP) Differs Between Female and Male Hippocampus
To study STSP we delivered a ten-pulse stimulation train of varying frequency at Schaffer collaterals, and we recorded the fEPSP from the CA1 stratum radiatum using three stimulation current intensities (example recordings are shown in Figure 2). Specifically we adjusted the stimulation current intensity to alternatively produce: (a) a subthreshold fEPSP (0.4±0.01mV/ms); (b) a suprathreshold fEPSP (1.47±0.04 mV/ms) that evoked a PS of 1.0-2.0 mV (1.36±.01 mV); (c) a PS of 75% of its maximum amplitude (4.76±0.1 mV), and a corresponding fEPSP of 2.49±.05 mV/ms. Figure 3 (and supplementary Figure 1) shows the results of this stimulation protocol on the first conditioned response (fEPSP 2) in both dorsal and ventral hippocampus which is equivalent to paired-pulse facilitation. It is evident that the female hippocampus lags behind the male in terms of facilitation of synaptic responses, while at high frequencies of stimulation it shows an absence of facilitation or even depression.
Figure 4 (and supplementary Figure 2) shows the results of frequency stimulation on the steady state response, i.e., fEPSP 8-10. Here, female performance is expressed with a greater frequency depression of synaptic responses in females at relatively higher stimulation frequencies and increased facilitation of synaptic responses in males at intermediate stimulation frequencies.
2.3. Short-Term Neuronal Dynamics (STND) Differ Between Female and Male Hippocampus
We next examined neuronal excitation by assessing population spikes (PS) recorded from the stratum pyramidale under suprathreshold condition, producing a PS of 1.0–2.0 mV, and submaximal condition, producing a PS of 75% of its maximum amplitude (Figure 5 and supplementary Figure 3). A unified picture emerges from these measurements that differs from that concerning synaptic transmission. Specifically, in contrast to the lower facilitation of fEPSP 2 seen in the female compared with the male hippocampus, both dorsal and ventral, the onset response of the neuronal output (PS 2) is increased in the dorsal female hippocampus (Figure 5A-B) but in the ventral hippocampus is increased in males (Figure 5C). A similar boosting of the neuronal output in the was also seen for steady state response (PS 8-10) in the female vs male hippocampus (Figure 6A, and and supplementary Figure 4).
3. Discussion
The main findings of the present study were: 1) females display lower frequency facilitation and increased frequency depression of synaptic transmission (fEPSP) in both segments of the hippocampus; 2) in contrast, a more complicated pattern of changes has been seen in STND, where increased frequency facilitation of neuronal excitation (PS) has been seen in the female dorsal hippocampus, and the male ventral hippocampus with suprathreshold but not submaximal stimulation.
3.1. Possible Interpretations
Specific properties of female and male hippocampal circuit may offer explanations of these differences in short-term dynamics between the two biological sexes. For instance, a previous study using cell cultures has shown that the size of recycling pool is larger and more frequently used in male than female synapses [54]. A slow and/or small recycling pool should be associated with insufficiently readily releasable pool leading to limited ability for short-term synaptic facilitation and increased tendency to short-term synaptic depression since the recovery from depression depends on the replenishment kinetics, which are governed by the availability and mobilization of recycling vesicles [55,56,57]. Conversely, relatively fast and/or large recycling pool can sustain more effectively to high-frequency activity, such as used in the present study, since synapses depend heavily on recycling pools to maintain neurotransmitter output [55,56,57]. Therefore, a less effective recycling pool in the female hippocampus could reliably explain the lower scores of frequency facilitation and greater frequency depression of synaptic responses observed in females compared to males in this study.
The present results are in apparent discrepancy with those of a recent study which examined STSP in male and female rat hippocampus in a model of Fragile X syndrome [58]. That study did not find significant differences in paired-pulse ratio between male and female dorsal or ventral hippocampus. The observed discrepancies in STSP profiles may arise from several sources. Most notably, the current study did not control for estrous cycle, which is known to significantly affect measures of synaptic transmission, neuronal excitability, and structural plasticity [17,19,59]. Estrogen fluctuations during the estrous cycle can markedly alter GABAergic inhibition and glutamatergic transmission [57,58,10], potentially masking or inflating subtle sex differences in STSP. Another explanation for this discrepancy may lie in strain-related differences, as the previous study used Long Evans while Wistar rats were used in the present study. Differences in animal strain can influence plasticity-related properties, as strain-specific variations impact synaptic protein expression, dendritic architecture, and responses to hormonal status.
The present study is the first to systematically examine STND across both sexes and the dorsoventral axis of the hippocampus, revealing a novel, previously unappreciated functional asymmetry in how neuronal output is modulated during repetitive stimulation. The observed increased frequency facilitation of PS responses in the dorsal hippocampus of females and the ventral hippocampus of males suggests the existence of sex-related adaptations in network responsiveness, possibly associated with distinct cognitive and/or emotional processing properties in females and males. Mechanistically, these STND differences may arise from sex- and region-specific variations in synaptic inhibition and excitatory drive. It has been recently shown that inhibition plays a crucial role in shaping the properties of short-term changes in neuronal firing during high-frequency short bursts of activity [44]. Specifically, reduced inhibition leads to higher scores of frequency facilitation and lower frequency depression of PS [44].
Previous studies have documented sex-related differences in GABAergic inhibition with estrogens reducing GABAergic inhibition in the hippocampus of adult female rats, leading to increased excitability [57,58]. Furthermore, the expression of estrogens appears higher in the dorsal than ventral hippocampus, especially in GAD-positive interneurons in females [28], suggesting an estrogen-mediated disinhibition mechanism in females. This could explain the enhanced dorsal STND facilitation in females, and its ventral shift in males. Furthermore, substantial sex-related differences in dendritic structure, such as higher dendritic spine density and branching in the female hippocampus, especially during proestrus [32,36], and regional variance in dendritic complexity, could contribute to the observed regional differentiation in STND. These morphological features may shape the integration and propagation of repetitive synaptic inputs across the hippocampal circuit in a sex- and region-specific manner.
3.2. Implications
Short-term dynamics are critical for the encoding, amplification, gating, and filtering of incoming signals in a timescale of milliseconds to seconds [1,2,3,4,5,43,60], neural processes linked to various brain functions such as working memory, attentional gating, and emotional processing, which are known to exhibit sex-specific profiles. Therefore, the observed sex- and region-dependent differences in STSP and STND may reflect distinct modes of hippocampal information processing and have important implications for such brain functions between females and males, possibly conferring distinct computational advantages or vulnerabilities relevant to each sex.
In addition, considering the functional segregation along the dorsoventral hippocampal axis, with the dorsal segment primarily engaged in cognitive tasks such as spatial memory and attention, and the ventral segment more involved in affective processing and stress regulation [52,53,61], our findings suggest that sex differences in short-term dynamics may be specifically related to sex-specific cognitive and affective functions. For instance, enhanced short-term facilitation of neuronal output in the dorsal hippocampus of females may support sex-specific cognitive flexibility or working memory processing, e.g., optimizing short-term encoding in spatial memory tasks, while greater facilitation in the ventral hippocampus of males may alter emotional reactivity or stress responsiveness.
Furthermore, these physiological distinctions may underlie known sex biases in the prevalence and presentation of neuropsychiatric disorders. For instance, in autism spectrum disorder, which is typically more prevalent in males [62], altered short-term dynamics have been implicated in abnormal sensory filtering and social cognition [50,51,58], and males display higher susceptibility to alterations in synaptic plasticity than females [63]. Hence, sex-specific hippocampal dynamics could underlie differential vulnerability and phenotypic expression [11]. In contrast, depression, which is more prevalent in females and linked to dysfunction in the ventral hippocampus [64,65], may be exacerbated by reduced excitatory dynamics and enhanced synaptic and neuronal depression observed in female ventral CA1 circuits. Future research should aim to identify the molecular and circuit-level mechanisms underlying these sex-related differences in short-term dynamics. Particular emphasis should be placed on hormonal modulation, and neuronal subtype-specific contributions, as well as on possible roles of synaptic proteins and postsynaptic receptors.
5. Materials and Methods
5.1. Animals and Hippocampal Slice Preparation
Transverse hippocampal slices (550 μm thick) were obtained from 3–4-month-old female and male Wistar rats housed in the Laboratory of Experimental Animals, Department of Medicine, University of Patras (license: EL-13-BIOexp-04). Animals were maintained under stable temperature (20–22 °C) and a 12:12 h light-dark cycle with ad libitum access to food and water. All procedures complied with EU Directive 2010/63/EU and were approved by institutional and regional authorities (reg. no. 187531/626, 26/06/2018). Following deep anaesthesia with diethyl ether and decapitation, brains were quickly removed and placed in ice-cold oxygenated artificial cerebrospinal fluid (ACSF). Slices were prepared from the dorsal and ventral hippocampus (0.5–3.5 mm from each end) and transferred to an interface chamber perfused with ACSF (containing, in mM: 124 NaCl, 4 KCl, 2 CaCl2, 2 MgSO4, 26 NaHCO3, 1.25 NaH2PO4 and 10 glucose, equilibrated with 95% O2 and 5% CO2 gas mixture at pH=7.4) at 30 ± 0.5 °C. Slices were continuously humidified with a mixed gas consisting of 95% O2 and 5% CO2. After at least 90 minutes of recovery, electrophysiological recordings were initiated.
5.2. Electrophysiology and Data Acquisition
Field potential was recorded from the middle CA1 region following stimulation of the Schaffer collaterals. A bipolar platinum/iridium electrode (25 μm wire diameter, 100 μm inter-wire spacing; WPI, USA) delivered constant current pulses (100 μs duration, 20–260 μA). Field excitatory postsynaptic potentials (fEPSPs) and population spikes (PS) were recorded from stratum radiatum and stratum pyramidale, respectively, using 7 μm carbon fiber electrodes (Kation Scientific, USA) placed ~350 μm from the stimulation site. Recordings often captured both fEPSP and PS simultaneously. Signals were amplified ×500, filtered (0.5 Hz–2 kHz), digitized at 10 kHz, and stored for offline analysis (CED 1401-plus and Signal5.9 software, Cambridge Electronic Design, UK).
5.3. Stimulation Protocols and Quantification
Input–output curves were constructed to determine stimulation levels, and baseline pulses were applied every 30 s using current intensities that elicited near-threshold fEPSPs. For short-term synaptic plasticity (STP) analysis, we employed a frequency stimulation protocol comprising ten-pulse trains delivered at frequencies ranging from 0.1 to 100 Hz. This pattern mimics naturally occurring spike trains in hippocampal pyramidal neurons [66]. Trains were applied in random order, with 2-minute inter-train intervals to ensure synaptic recovery. To study activity-dependent changes, stimulation was applied at three defined intensities: Subthreshold: eliciting ~0.5 mV/ms fEPSP without PS; Suprathreshold: evoking a PS of 1.0-2.0 mV; Submaximal: evoking a PS at ~75% of maximal amplitude [44,67]. STP of fEPSP and PS was quantified by measuring the percent change of each of the nine conditioned responses (2nd –10th pulses) relative to the first (conditioning response). Steady-state responses were defined as the average of the 8th –10th pulses. The fEPSP slope was measured within 1 ms of the fiber volley onset, and PS amplitude was calculated as the vertical projection of the minimum peak relative to the line connecting the two flanking positive peaks.
5.5. Statistical Analysis
Comparisons between two groups were performed using the independent t-test. The assumption of equal variances and normal distribution of data was evaluated using Levene’s test and the Shapiro–Wilk test, respectively. Each hippocampal slice was considered an independent experimental unit. Data are expressed as mean ± S.E.M. In column graphs, values are presented as mean ± S.E.M., while box plots depict the median and interquartile range (25th to 75th percentiles; diamond-shaped box), mean (thick line), 5th and 95th percentiles (whiskers), and individual data points (superimposed). The number of slices and animals used (expressed as slices/animals) is reported throughout the text. Statistical analyses were performed based on the number of slices. All analyses were conducted using IBM SPSS Statistics version 27.
Author Contributions
A.M., G.T., M.T., and A.K. performed the experiments and data analysis. C.P. supervised the project and performed data analysis. A.M., and C.P. prepared and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Empeirikeion Foundation. A.M. is a recipient of a postgraduate fellowship from the Hellenic Foundation for Research and Innovation (HFRI). G.T. was financially supported by the “Polembros Shipping Limited”, as a recipient of Ph.D. fellowships.
Institutional Review Board Statement
The animal study protocol was approved by the Research Ethics Committee of the University of Patras and the Directorate of Veterinary Services of the Achaia Prefecture of Western Greece Region (reg. number: 5661/37, 18 January 2021). The treatment of animals and all experimental procedures used in this study were conducted in accordance with the European Communities Council Directive Guidelines for the care and use of Laboratory animals (2010/63/EU—European Commission).
Data Availability Statement
All data associated with this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
ACSF, artificial cerebrospinal fluid; fEPSP, field excitatory postsynaptic potential; PS, population spike, STSP, short-term synaptic plasticity; STND, short-term neuronal dynamics.
References
- Zucker, R.S.; Regehr, W.G. Short-Term Synaptic Plasticity. Annu. Rev. Physiol. 2002, 64, 355–405. [Google Scholar] [CrossRef]
- Jackman, S.L.; Regehr, W.G. The Mechanisms and Functions of Synaptic Facilitation. Neuron 2017, 94, 447–464. [Google Scholar] [CrossRef]
- Anwar, H.; Li, X.; Bucher, D.; Nadim, F. Functional roles of short-term synaptic plasticity with an emphasis on inhibition. Curr. Opin. Neurobiol. 2017, 43, 71–78. [Google Scholar] [CrossRef]
- Citri, A.; Malenka, R.C. Synaptic Plasticity: Multiple Forms, Functions, and Mechanisms. Neuropsychopharmacology 2008, 33, 18–41. [Google Scholar] [CrossRef] [PubMed]
- Hennig, M.H. Theoretical models of synaptic short term plasticity. Front. Comput. Neurosci. 2013, 7, 45. [Google Scholar] [CrossRef]
- Devaraju, P.; Yu, J.; Eddins, D.; Mellado-Lagarde, M.M.; Earls, L.R.; Westmoreland, J.J.; Quarato, G.; Green, D.R.; Zakharenko, S.S. Haploinsufficiency of the 22q11.2 microdeletion gene Mrpl40 disrupts short-term synaptic plasticity and working memory through dysregulation of mitochondrial calcium. Mol. Psychiatry 2016, 22, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Pals, M.; Stewart, T.C.; Akyürek, E.G.; Borst, J.P.; Morrison, A. A functional spiking-neuron model of activity-silent working memory in humans based on calcium-mediated short-term synaptic plasticity. PLOS Comput. Biol. 2020, 16, e1007936. [Google Scholar] [CrossRef]
- Mongillo, G.; Barak, O.; Tsodyks, M. Synaptic Theory of Working Memory. Science 2008, 319, 1543–1546. [Google Scholar] [CrossRef] [PubMed]
- Deco, G.; Rolls, E.T.; Romo, R. Synaptic dynamics and decision making. Proc. Natl. Acad. Sci. 2010, 107, 7545–7549. [Google Scholar] [CrossRef]
- Woolley, C.S. Acute Effects of Estrogen on Neuronal Physiology. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 657–680. [Google Scholar] [CrossRef]
- Gall, C.M.; Le, A.A.; Lynch, G. Sex differences in synaptic plasticity underlying learning. J. Neurosci. Res. 2021, 101, 764–782. [Google Scholar] [CrossRef]
- Kniffin, A.R.; Briand, L.A. Sex differences in glutamate transmission and plasticity in reward related regions. Front. Behav. Neurosci. 2024, 18, 1455478. [Google Scholar] [CrossRef]
- Fester, L.; Rune, G.M. Sexual neurosteroids and synaptic plasticity in the hippocampus. Brain Res. 2015, 1621, 162–169. [Google Scholar] [CrossRef]
- Foy, M.R. Ovarian hormones, aging and stress on hippocampal synaptic plasticity. Neurobiol. Learn. Mem. 2011, 95, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Baudry, M.; Bi, X.; Aguirre, C. Progesterone–estrogen interactions in synaptic plasticity and neuroprotection. Neuroscience 2013, 239, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Hyer, M.M.; Phillips, L.L.; Neigh, G.N. Sex Differences in Synaptic Plasticity: Hormones and Beyond. Front. Mol. Neurosci. 2018, 11, 266. [Google Scholar] [CrossRef] [PubMed]
- Warren, S.G.; Humphreys, A.G.; Juraska, J.M.; Greenough, W.T. LTP varies across the estrous cycle: enhanced synaptic plasticity in proestrus rats. Brain Res. 1995, 703, 26–30. [Google Scholar] [CrossRef]
- Good, M.; Day, M.; Muir, J.L. Cyclical changes in endogenous levels of oestrogen modulate the induction of LTD and LTP in the hippocampal CA1 region. Eur. J. Neurosci. 1999, 11, 4476–4480. [Google Scholar] [CrossRef] [PubMed]
- Bi, R.; Foy, M.R.; Vouimba, R.-M.; Thompson, R.F.; Baudry, M. Cyclic changes in estradiol regulate synaptic plasticity through the MAP kinase pathway. Proc. Natl. Acad. Sci. 2001, 98, 13391–13395. [Google Scholar] [CrossRef]
- Kramár, E.; Babayan, A.; Gall, C.; Lynch, G. Estrogen promotes learning-related plasticity by modifying the synaptic cytoskeleton. Neuroscience 2013, 239, 3–16. [Google Scholar] [CrossRef]
- Kramár, E.A.; Chen, L.Y.; Brandon, N.J.; Rex, C.S.; Liu, F.; Gall, C.M.; Lynch, G. Cytoskeletal Changes Underlie Estrogen's Acute Effects on Synaptic Transmission and Plasticity. J. Neurosci. 2009, 29, 12982–12993. [Google Scholar] [CrossRef]
- Montoya, D.C.; Carrer, H. Estrogen facilitates induction of long term potentiation in the hippocampus of awake rats. Brain Res. 1997, 778, 430–438. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Hojo, Y.; Kojima, H.; Ikeda, M.; Hotta, K.; Sato, R.; Ooishi, Y.; Yoshiya, M.; Chung, B.-C.; Yamazaki, T.; et al. Estradiol rapidly modulates synaptic plasticity of hippocampal neurons: Involvement of kinase networks. Brain Res. 2015, 1621, 147–161. [Google Scholar] [CrossRef]
- Maren, S.; Baudry, M. Properties and Mechanisms of Long-Term Synaptic Plasticity in the Mammalian Brain: Relationships to Learning and Memory. Neurobiol. Learn. Mem. 1995, 63, 1–18. [Google Scholar] [CrossRef]
- Monfort, P.; Gomez-Gimenez, B.; Llansola, M.; Felipo, V. Gender Differences in Spatial Learning, Synaptic Activity, and Long-Term Potentiation in the Hippocampus in Rats: Molecular Mechanisms. ACS Chem. Neurosci. 2015, 6, 1420–1427. [Google Scholar] [CrossRef]
- Safari, S.; Ahmadi, N.; Mohammadkhani, R.; Ghahremani, R.; Khajvand-Abedeni, M.; Shahidi, S.; Komaki, A.; Salehi, I.; Karimi, S.A. Sex differences in spatial learning and memory and hippocampal long-term potentiation at perforant pathway-dentate gyrus (PP-DG) synapses in Wistar rats. Behav. Brain Funct. 2021, 17, 1–11. [Google Scholar] [CrossRef]
- Jain, A.; Huang, G.Z.; Woolley, C.S. Latent sex differences in molecular signaling that underlies excitatory synaptic potentiation in the hippocampus. J. Neurosci. 2018, 39, 1897–18. [Google Scholar] [CrossRef] [PubMed]
- Shughrue, P.; Merchenthaler, I. Evidence for novel estrogen binding sites in the rat hippocampus. Neuroscience 2000, 99, 605–612. [Google Scholar] [CrossRef]
- Hart, S.A.; Patton, J.D.; Woolley, C.S. Quantitative analysis of ERα and GAD colocalization in the hippocampus of the adult female rat. J. Comp. Neurol. 2001, 440, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Deutschmann, A.U.; Kirkland, J.M.; Briand, L.A. Adolescent social isolation induced alterations in nucleus accumbens glutamate signalling. Addict. Biol. 2021, 27, e13077–e13077. [Google Scholar] [CrossRef] [PubMed]
- Knouse, M.C.; McGrath, A.G.; Deutschmann, A.U.; Rich, M.T.; Zallar, L.J.; Rajadhyaksha, A.M.; Briand, L.A. Sex differences in the medial prefrontal cortical glutamate system. Biol. Sex Differ. 2022, 13, 1–9. [Google Scholar] [CrossRef]
- Luine, V.; Frankfurt, M. Interactions between estradiol, BDNF and dendritic spines in promoting memory. Neuroscience 2013, 239, 34–45. [Google Scholar] [CrossRef]
- Li, C.; Brake, W.G.; Romeo, R.D.; Dunlop, J.C.; Gordon, M.; Buzescu, R.; Magarinos, A.M.; Allen, P.B.; Greengard, P.; Luine, V.; et al. Estrogen alters hippocampal dendritic spine shape and enhances synaptic protein immunoreactivity and spatial memory in female mice. Proc. Natl. Acad. Sci. 2004, 101, 2185–2190. [Google Scholar] [CrossRef]
- Tuscher, J.J.; Luine, V.; Frankfurt, M.; Frick, K.M. Estradiol-Mediated Spine Changes in the Dorsal Hippocampus and Medial Prefrontal Cortex of Ovariectomized Female Mice Depend on ERK and mTOR Activation in the Dorsal Hippocampus. J. Neurosci. 2016, 36, 1483–1489. [Google Scholar] [CrossRef]
- Castillo-Fernández, S.; Silva-Gómez, A.B. Changes in dendritic arborization related to the estrous cycle in pyramidal neurons of layer V of the motor cortex. J. Chem. Neuroanat. 2021, 119, 102042. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, P.A.S.; Choleris, E.; Galea, L.A.M. Structural plasticity of the hippocampus in response to estrogens in female rodents. Mol. Brain 2019, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Branco, T.; Häusser, M. Synaptic Integration Gradients in Single Cortical Pyramidal Cell Dendrites. Neuron 2011, 69, 885–892. [Google Scholar] [CrossRef]
- Poirazi, P.; Brannon, T.; Mel, B.W. Arithmetic of Subthreshold Synaptic Summation in a Model CA1 Pyramidal Cell. Neuron 2003, 37, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Bartley, A.F.; Dobrunz, L.E. Short-term plasticity regulates the excitation/inhibition ratio and the temporal window for spike integration in CA1 pyramidal cells. Eur. J. Neurosci. 2015, 41, 1402–1415. [Google Scholar] [CrossRef]
- Carvalho, T.P.; Buonomano, D.V. Differential Effects of Excitatory and Inhibitory Plasticity on Synaptically Driven Neuronal Input-Output Functions. Neuron 2009, 61, 774–785. [Google Scholar] [CrossRef]
- Galarreta, M.; Hestrin, S. Frequency-dependent synaptic depression and the balance of excitation and inhibition in the neocortex. Nat. Neurosci. 1998, 1, 587–594. [Google Scholar] [CrossRef]
- Abbott, L.F.; Varela, J.A.; Sen, K.; Nelson, S.B. Synaptic Depression and Cortical Gain Control. Science 1997, 275, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Silver, R.A. Neuronal arithmetic. Nat. Rev. Neurosci. 2010, 11, 474–489. [Google Scholar] [CrossRef] [PubMed]
- Koutsoumpa, A.; Papatheodoropoulos, C. Short-term dynamics of input and output of CA1 network greatly differ between the dorsal and ventral rat hippocampus. BMC Neurosci. 2019, 20, 1–30. [Google Scholar] [CrossRef]
- Tsotsokou, G.; Fassea, M.; Papatheodoropoulos, C. Muscarinic Modulation of Network Excitability and Short-Term Dynamics in the Dorsal and Ventral Hippocampus. . 2024, 2024. [Google Scholar] [CrossRef]
- Tsotsokou, G.; Sotiropoulou, I.-M.; Stampolitis, K.; Oikonomou, G.D.; Avdi, A.-P.; Papatheodoropoulos, C. Cannabinoid Modulation of Excitability and Short-Term Neuronal Dynamics in the Dorsal and Ventral Hippocampus. Biology 2025, 14, 642. [Google Scholar] [CrossRef]
- Miliou, A.; Papaleonidopoulos, V.; Trompoukis, G.; Papatheodoropoulos, C. Septotemporal variation in beta-adrenergic modulation of short-term dynamics in the hippocampus. IBRO Neurosci. Rep. 2021, 11, 64–72. [Google Scholar] [CrossRef]
- Trompoukis, G.; Tsotsokou, G.; Koutsoumpa, A.; Tsolaki, M.; Vryoni, G.; Papatheodoropoulos, C. Age-dependent modulation of short-term neuronal dynamics in the dorsal and ventral rat hippocampus. Int. J. Dev. Biol. 2022, 66, 285–296. [Google Scholar] [CrossRef]
- Kandilakis, C.L.; Papatheodoropoulos, C. Serotonin Modulation of Dorsoventral Hippocampus in Physiology and Schizophrenia. Int. J. Mol. Sci. 2025, 26, 7253. [Google Scholar] [CrossRef]
- Deng, P.-Y.; Sojka, D.; Klyachko, V.A. Abnormal Presynaptic Short-Term Plasticity and Information Processing in a Mouse Model of Fragile X Syndrome. J. Neurosci. 2011, 31, 10971–10982. [Google Scholar] [CrossRef] [PubMed]
- Leontiadis, L.J.; Felemegkas, P.; Trompoukis, G.; Tsotsokou, G.; Miliou, A.; Karagianni, E.; Rigas, P.; Papatheodoropoulos, C. Septotemporal Variation of Information Processing in the Hippocampus of Fmr1 KO Rat. Dev. Neurosci. 2024, 46, 353–364. [Google Scholar] [CrossRef]
- Bannerman, D.M.; Sprengel, R.; Sanderson, D.J.; McHugh, S.B.; Rawlins, J.N.; Monyer, H.; Seeburg, P.H. Hippocampal synaptic plasticity, spatial memory and anxiety. Nat Rev Neurosci 2014, 15, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Strange, B.A.; Witter, M.P.; Lein, E.S.; Moser, E.I. Functional organization of the hippocampal longitudinal axis. Nat. Rev. Neurosci. 2014, 15, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Sertel, S.M.; Blumenstein, W.; Mandad, S.; Shomroni, O.; Salinas, G.; Rizzoli, S.O. Differences in synaptic vesicle pool behavior between male and female hippocampal cultured neurons. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Stevens, C.F.; Wesseling, J.F. Activity-Dependent Modulation of the Rate at which Synaptic Vesicles Become Available to Undergo Exocytosis. Neuron 1998, 21, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Denker, A.; O Rizzoli, S. Synaptic vesicle pools: an update. Front. Synaptic Neurosci. 2010, 2, 135. [Google Scholar] [CrossRef]
- Neher, E.; Brose, N. Dynamically Primed Synaptic Vesicle States: Key to Understand Synaptic Short-Term Plasticity. Neuron 2018, 100, 1283–1291. [Google Scholar] [CrossRef]
- Tsotsokou, G.; Miliou, A.; Trompoukis, G.; Leontiadis, L.J.; Papatheodoropoulos, C. Region-Related Differences in Short-Term Synaptic Plasticity and Synaptotagmin-7 in the Male and Female Hippocampus of a Rat Model of Fragile X Syndrome. Int. J. Mol. Sci. 2024, 25, 6975. [Google Scholar] [CrossRef]
- Day, H.L.L.; Stevenson, C.W. The neurobiological basis of sex differences in learned fear and its inhibition. Eur. J. Neurosci. 2019, 52, 2466–2486. [Google Scholar] [CrossRef]
- Abbott, L.F.; Regehr, W.G. Synaptic computation. Nature 2004, 431, 796–803. [Google Scholar] [CrossRef]
- Fanselow, M.S.; Dong, H.-W. Are the Dorsal and Ventral Hippocampus Functionally Distinct Structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Werling, D.M.; Geschwind, D.H. Sex differences in autism spectrum disorders. Curr. Opin. Neurol. 2013, 26, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Mottron, L.; Duret, P.; Mueller, S.; Moore, R.D.; D’aRc, B.F.; Jacquemont, S.; Xiong, L. Sex differences in brain plasticity: a new hypothesis for sex ratio bias in autism. Mol. Autism 2015, 6, 1–19. [Google Scholar] [CrossRef]
- Bagot, R.C.; Parise, E.M.; Peña, C.J.; Zhang, H.-X.; Maze, I.; Chaudhury, D.; Persaud, B.; Cachope, R.; Bolaños-Guzmán, C.A.; Cheer, J.F.; et al. Ventral hippocampal afferents to the nucleus accumbens regulate susceptibility to depression. Nat. Commun. 2015, 6, 7062–7062. [Google Scholar] [CrossRef]
- Papp, M.; Gruca, P.; Lason, M.; Litwa, E.; Solecki, W.; Willner, P. Insufficiency of ventral hippocampus to medial prefrontal cortex transmission explains antidepressant non-response. J. Psychopharmacol. 2021, 35, 1253–1264. [Google Scholar] [CrossRef]
- Fenton, A.A.; Muller, R.U. Place cell discharge is extremely variable during individual passes of the rat through the firing field. Proc. Natl. Acad. Sci. 1998, 95, 3182–3187. [Google Scholar] [CrossRef]
- Koutsoumpa, A.; Papatheodoropoulos, C. Frequency-dependent layer-specific differences in short-term synaptic plasticity in the dorsal and ventral CA1 hippocampal field. Synapse 2021, 75, e22199. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Input-output curves between fEPSP and PS as a function of stimulation current intensity, and between PS and fEPSP, are shown for the dorsal hippocampus and the ventral hippocampus of female and male rats. The numbers of slices and rats used are indicated in parentheses. Average values derived from the input-output curves are presented in the corresponding inset graphs, where the results of independent t-tests are also shown.
Figure 1.
Input-output curves between fEPSP and PS as a function of stimulation current intensity, and between PS and fEPSP, are shown for the dorsal hippocampus and the ventral hippocampus of female and male rats. The numbers of slices and rats used are indicated in parentheses. Average values derived from the input-output curves are presented in the corresponding inset graphs, where the results of independent t-tests are also shown.
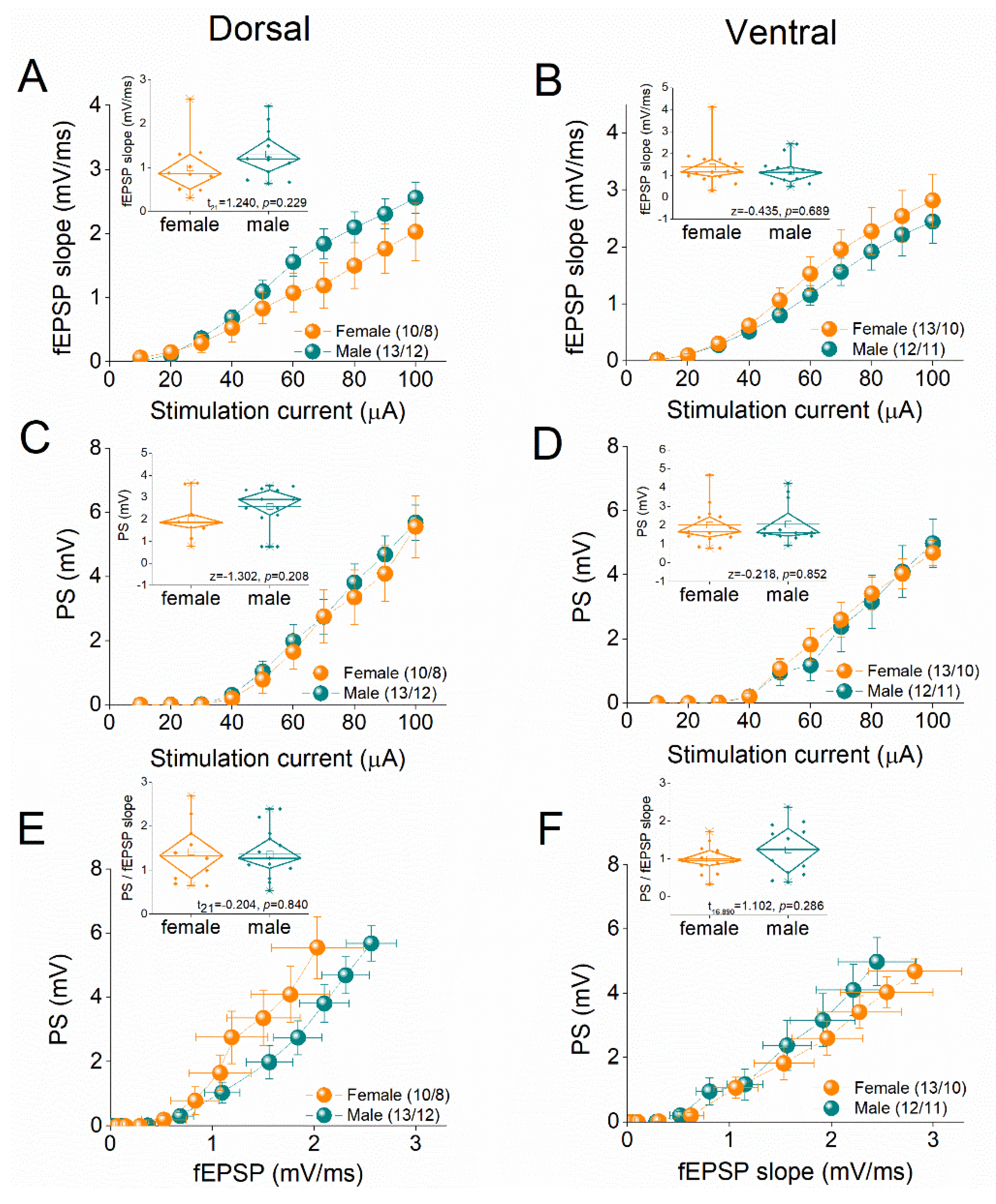
Figure 2.
Representative traces of evoked responses from the CA1 stratum radiatum (fEPSPs) and stratum pyramidale (PS) recorded during the repetitive stimulation of Schaffer collaterals at the dorsal (A) and ventral (B) hippocampal slices of female and male rats. Traces illustrate responses to subthreshold stimulation at 40 Hz, suprathreshold stimulation at 50 Hz and submaximal stimulation at 5 Hz. Calibration bars: 1 mV, 20 ms in traces of 40 Hz and 50 Hz; 3 mV, 200 ms in traces of 5 Hz. Stimulation artifacts are truncated for clarity.
Figure 2.
Representative traces of evoked responses from the CA1 stratum radiatum (fEPSPs) and stratum pyramidale (PS) recorded during the repetitive stimulation of Schaffer collaterals at the dorsal (A) and ventral (B) hippocampal slices of female and male rats. Traces illustrate responses to subthreshold stimulation at 40 Hz, suprathreshold stimulation at 50 Hz and submaximal stimulation at 5 Hz. Calibration bars: 1 mV, 20 ms in traces of 40 Hz and 50 Hz; 3 mV, 200 ms in traces of 5 Hz. Stimulation artifacts are truncated for clarity.
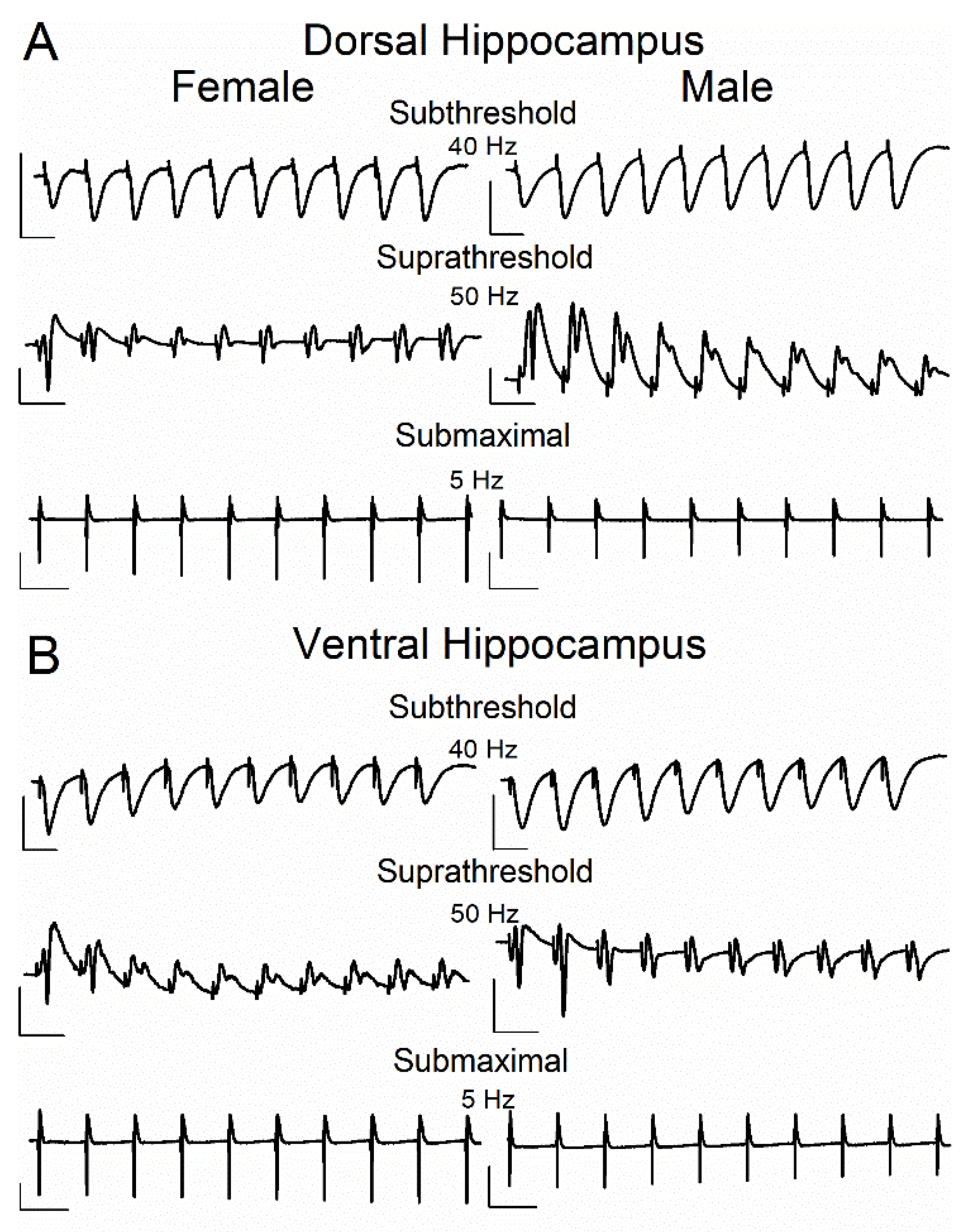
Figure 3.
Frequency-dependent modulation of fEPSP in the dorsal hippocampus (A-C) and ventral hippocampus (D-F) of female and male rats. Results for the first conditioned response (fEPSP 2) are presented under three stimulation intensities: subthreshold, suprathreshold, and submaximal. The average percent effects of frequency stimulation on fEPSP 2 are also illustrated using box plots positioned above the frequency-response graphs. Statistically significant differences are denoted by asterisks (independent t-test, p < 0.05). Numbers into parentheses in this and the following figures indicate the number of slices/rats used. Note that both the dorsal and ventral hippocampus of male rats exhibit greater synaptic facilitation than females under subthreshold and suprathreshold, but not submaximal stimulation conditions.
Figure 3.
Frequency-dependent modulation of fEPSP in the dorsal hippocampus (A-C) and ventral hippocampus (D-F) of female and male rats. Results for the first conditioned response (fEPSP 2) are presented under three stimulation intensities: subthreshold, suprathreshold, and submaximal. The average percent effects of frequency stimulation on fEPSP 2 are also illustrated using box plots positioned above the frequency-response graphs. Statistically significant differences are denoted by asterisks (independent t-test, p < 0.05). Numbers into parentheses in this and the following figures indicate the number of slices/rats used. Note that both the dorsal and ventral hippocampus of male rats exhibit greater synaptic facilitation than females under subthreshold and suprathreshold, but not submaximal stimulation conditions.
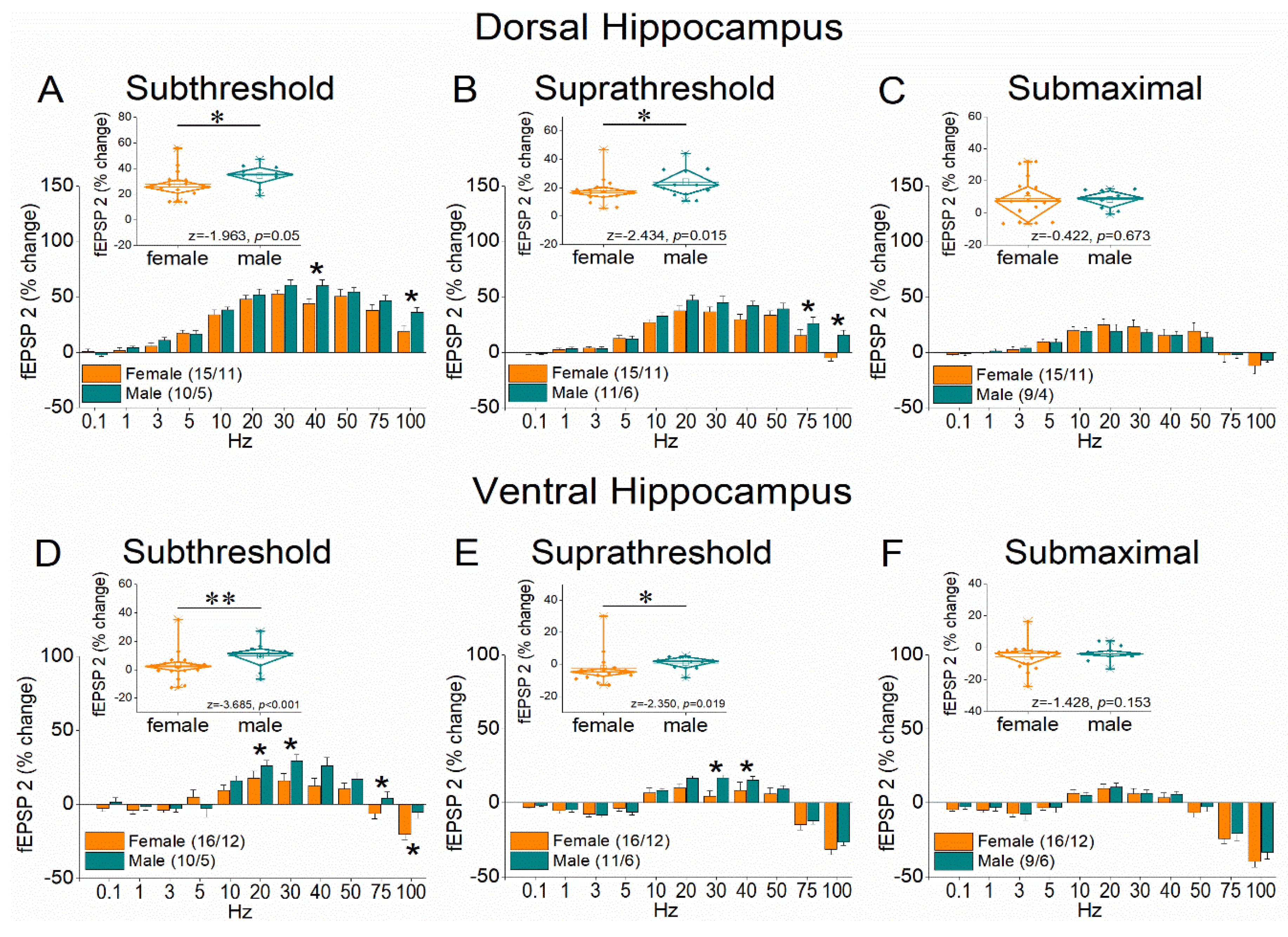
Figure 4.
Frequency-dependent modulation of fEPSP in the dorsal hippocampus (A-C) and ventral hippocampus (D-F) of female and male rats. Results for the steady-state conditioned response (fEPSP 8-10) are presented across three stimulation intensities: subthreshold, suprathreshold, and submaximal. The average percent effects of frequency stimulation on fEPSP 8-10 are also illustrated using box plots positioned above the frequency-response graphs. Statistically significant differences are denoted by asterisks (independent t-test, p < 0.05). Note that the ventral hippocampus of male rats exhibits greater frequency facilitation, or lower frequency depression than females, under subthreshold stimulation intensity.
Figure 4.
Frequency-dependent modulation of fEPSP in the dorsal hippocampus (A-C) and ventral hippocampus (D-F) of female and male rats. Results for the steady-state conditioned response (fEPSP 8-10) are presented across three stimulation intensities: subthreshold, suprathreshold, and submaximal. The average percent effects of frequency stimulation on fEPSP 8-10 are also illustrated using box plots positioned above the frequency-response graphs. Statistically significant differences are denoted by asterisks (independent t-test, p < 0.05). Note that the ventral hippocampus of male rats exhibits greater frequency facilitation, or lower frequency depression than females, under subthreshold stimulation intensity.
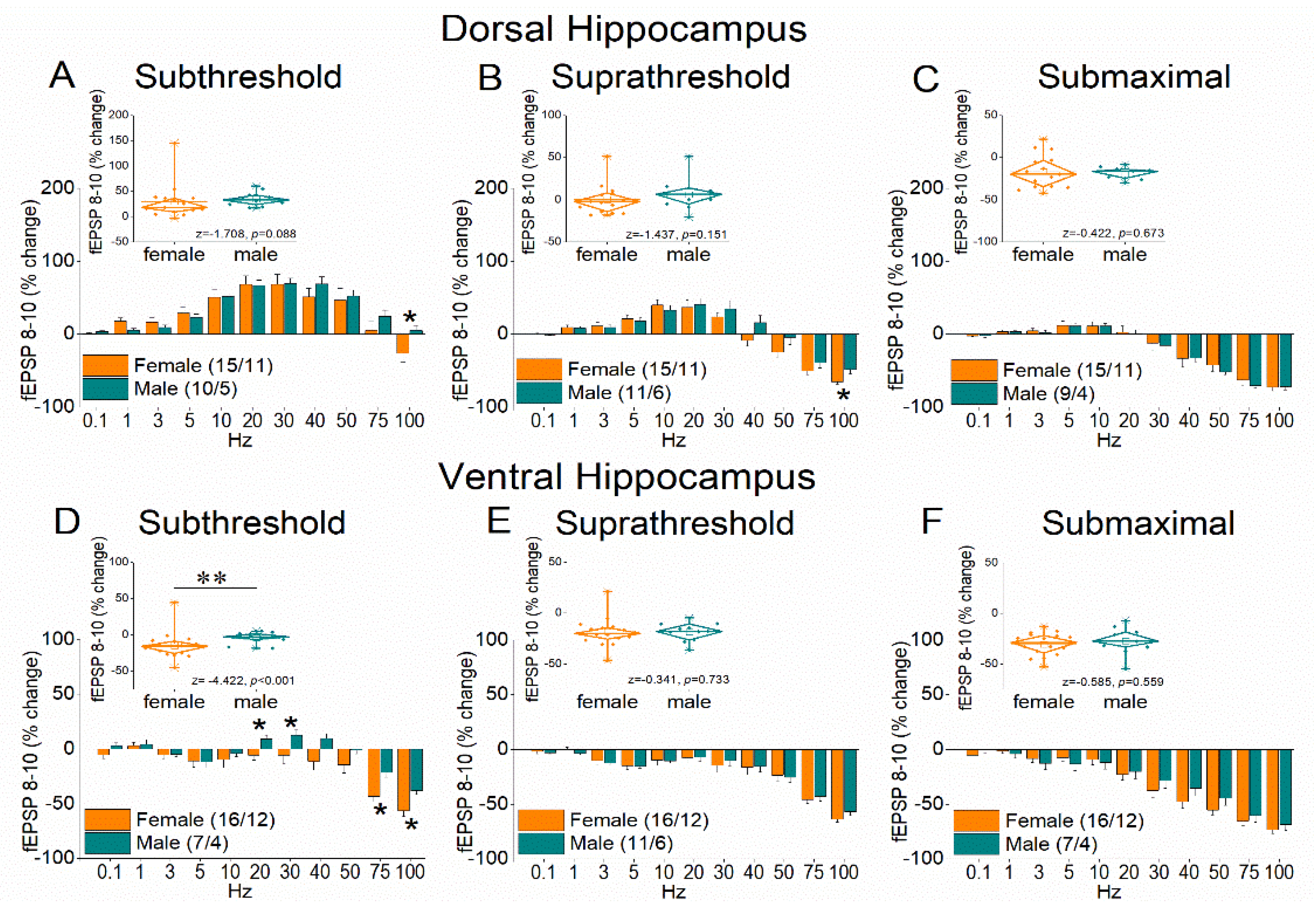
Figure 5.
Frequency-dependent modulation of PS in the dorsal and ventral hippocampus of female and male rats. Results for the first conditioned response (PS 2, onset response) are presented under two stimulation intensities: suprathreshold, and submaximal. The average percent effects of frequency stimulation on PS 2 are also illustrated using box plots positioned above the frequency-response graphs. Statistically significant differences are denoted by asterisks (independent t-test, p < 0.05). Note the region-specific response to frequency stimulation in the onset response between the two sexes under suprathreshold stimulation: frequency stimulation induces greater facilitation in the dorsal hippocampus of females and the ventral hippocampus of males.
Figure 5.
Frequency-dependent modulation of PS in the dorsal and ventral hippocampus of female and male rats. Results for the first conditioned response (PS 2, onset response) are presented under two stimulation intensities: suprathreshold, and submaximal. The average percent effects of frequency stimulation on PS 2 are also illustrated using box plots positioned above the frequency-response graphs. Statistically significant differences are denoted by asterisks (independent t-test, p < 0.05). Note the region-specific response to frequency stimulation in the onset response between the two sexes under suprathreshold stimulation: frequency stimulation induces greater facilitation in the dorsal hippocampus of females and the ventral hippocampus of males.
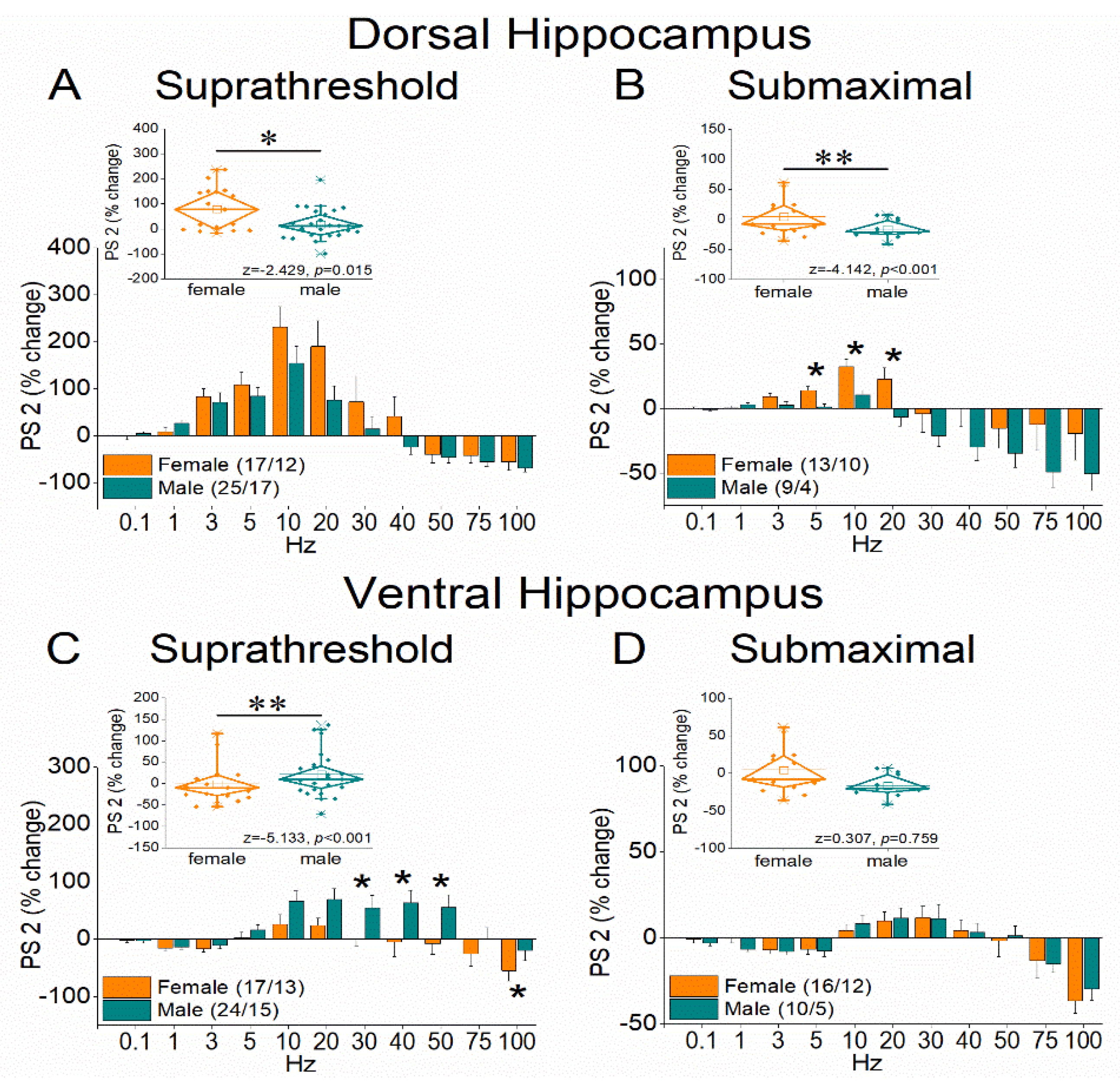
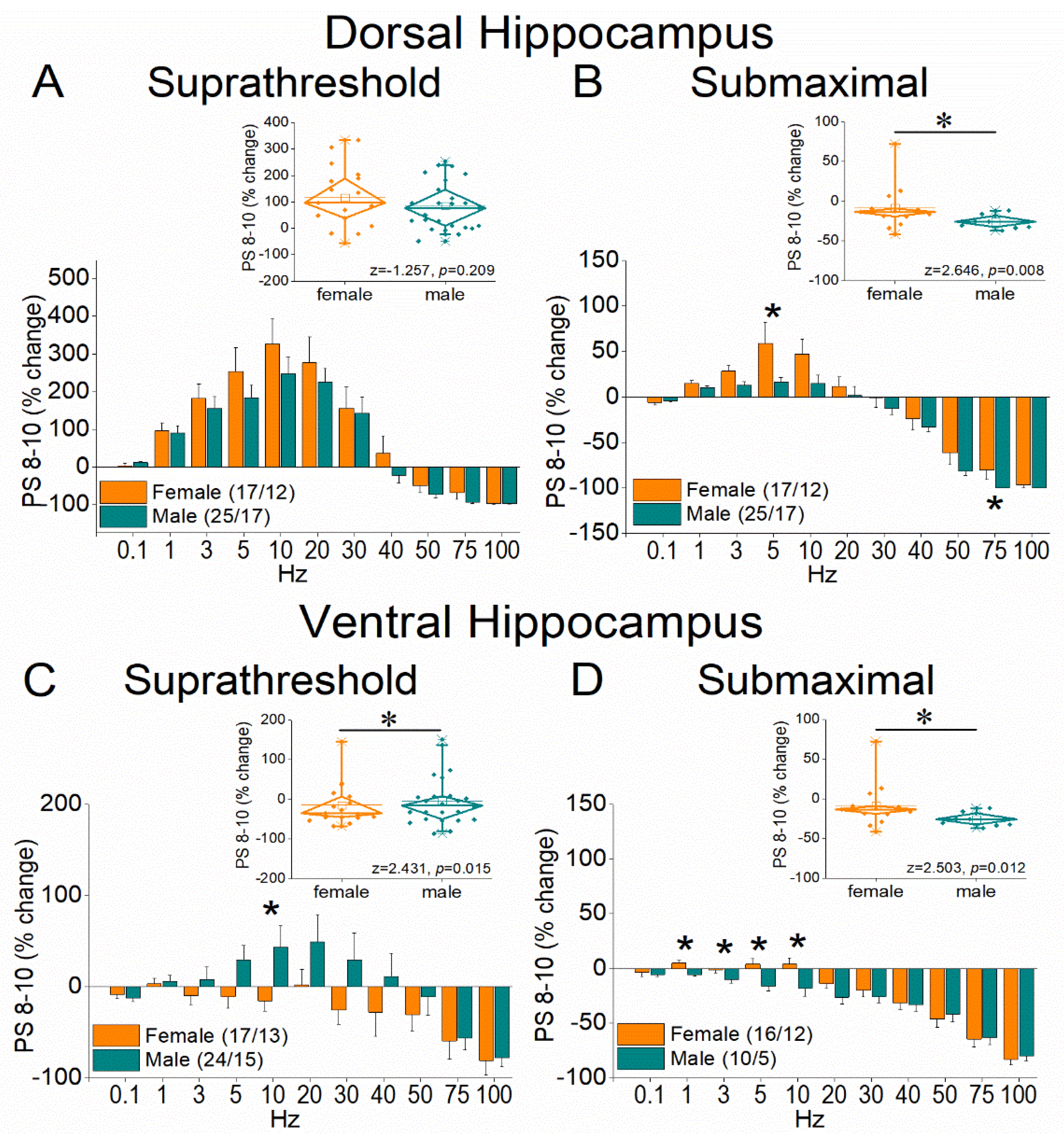
Figure 6.
Frequency-dependent modulation of PS in the dorsal and ventral hippocampus of female and male rats. Results for the steady-state conditioned response (PS 8-10) are presented across two stimulation intensities: suprathreshold, and submaximal. The average percent effects of frequency stimulation on fEPSP 8-10 are also illustrated using box plots positioned above the frequency-response graphs. Statistically significant differences are denoted by asterisks (independent t-test, p < 0.05). Note that the ventral hippocampus of male rats exhibits greater frequency facilitation under suprathreshold stimulation and greater frequency depression under submaximal stimulation, suggesting a broader dynamic range of responses compared to the ventral hippocampus of female rats.
Figure 6.
Frequency-dependent modulation of PS in the dorsal and ventral hippocampus of female and male rats. Results for the steady-state conditioned response (PS 8-10) are presented across two stimulation intensities: suprathreshold, and submaximal. The average percent effects of frequency stimulation on fEPSP 8-10 are also illustrated using box plots positioned above the frequency-response graphs. Statistically significant differences are denoted by asterisks (independent t-test, p < 0.05). Note that the ventral hippocampus of male rats exhibits greater frequency facilitation under suprathreshold stimulation and greater frequency depression under submaximal stimulation, suggesting a broader dynamic range of responses compared to the ventral hippocampus of female rats.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



