
Submitted:
03 June 2024
Posted:
05 June 2024
You are already at the latest version
Abstract
Fragile X syndrome (FXS) is an intellectual developmental disorder characterized, inter alia, by deficits in short-term processing of neural information such as sensory processing and working memory. The primary cause of FXS is the loss of fragile X messenger ribonucleoprotein (FMRP), which is profoundly involved in synaptic function and plasticity. Short-term synaptic plasticity (STSP) may play important roles in functions that are affected by FXS. Recent evidence points to a crucial involvement of presynaptic calcium sensor synaptotagmin-7 in STSP. However, how the loss of FMRP affects STSP and synaptotagmin-7 have been insufficiently studied. Further-more, males and females are affected differently by FXS, but the underlying mechanisms remain elusive. The aim of the present study is to investigate possible changes in STSP and the expres-sion of synaptotagmin-7 along the hippocampus of adult males and females in the Fmr1 knock-out (KO) rat model of FXS. We found that paired-pulse ratio and frequency facilita-tion/depression, two forms of STSP, as well as the expression of synaptotagmin-7, are normal in adult KO males, but paired-pulse ratio is increased in the ventral hippocampus of KO females. Furthermore, we found no gender-related but robust region-dependent difference in STSP. AM-PA receptors are similarly expressed in the two hippocampal segments of the two genotypes and in both genders. Also, basal excitatory synaptic transmission is higher in males compared with females. Interestingly, we found more than twofold higher levels of synaptotagmin-7, not syn-aptotagmin-1, in the dorsal compared with the ventral hippocampus in males of both genotypes and in wild type females. These results point to the susceptibility of the female ventral hippo-campus to FMRP loss. Importantly, the different levels of synaptotagmin-7 that parallel the higher score of synaptic facilitation in the dorsal vs ventral hippocampus suggest that synapto-tagmin-7 may play a pivotal role in defining the striking difference in STSP along the long axis of the hippocampus.
Keywords:
Synaptotagmin
; hippocampus
; Fragile X syndrome
; Fmr1
; short-term synaptic plasticity
; synaptic transmission
; septotemporal
; dorsoventral
; gender
; sex
1. Introduction
Fragile X syndrome (FXS) is a neurodevelopmental disorder and one of the most common inherited forms of intellectual disability [1,2,3,4], often associated with autistic behaviors [3,4,5]. Behaviorally, FXS is characterized by deficiencies in social communication, learning and memory, hypersensitivity, anxiety, and seizures [2,6]. FXS is primarily caused by a mutation associated with a CGG repeat expansion of the Fmr1 gene in the X chromosome, with full mutation consisting of more than 200 repeats, leading to a transcriptional silencing of Fmr1 gene [7,8,9]. The inactivation of Fmr1 gene leads to insufficient expression or total loss of fragile X messenger ribonucleoprotein (FMRP), which is broadly expressed throughout the body displaying high levels in the brain [10,11].
Due to the regulation of protein synthesis via binding with mRNAs, which is a main action of FMRP, the loss of FMRP in FXS leads to increased levels of several proteins in the brain [12,13,14,15]. Furthermore, FMRP can directly modulate other proteins such as ion channels [16,17]. FMRP is involved in many physiological processes including synaptogenesis, inter-neuronal signaling, synaptic plasticity, and neuronal excitability [15].
Main actions of FMRP are found at synapses [8,12,13,18,19,20,21,22] and numerous mRNA targets of FMRP are localized in the neuronal dendrites where it modulates local translation of several synaptic proteins including glutamate receptor subunits [23,24] that are involved in long-term synaptic plasticity and memory formation [25,26]. Expectedly, a great deal of the previous research concerning the consequences of FMRP loss on neuronal function has been focused on the postsynaptic compartment, and it is generally thought that most of the FMRP effects are related to synaptic plasticity [13,27]. Notably, the absence of FMRP in FXS is associated with dysregulation of several forms of synaptic plasticity that are expressed postsynaptically and require protein synthesis [28,29,30].
In addition to the impact that FXS has on learning-memory and long-term synaptic plasticity, the loss of FMRP is also associated with disrupted short-term processing of neural information including sensory processing and working memory [31,32]; these forms of neural information processing are proposed to engage short-term forms of synaptic plasticity [33,34]. Short-term forms of synaptic plasticity are thought to be critically supported by Ca2+-dependent mechanisms that engage components of the neurotransmitter release machinery and regulate the probability of neurotransmitter release during repetitive synaptic activity [35]. Interestingly, FMRP is expressed presynaptically [21,22,36,37], and recent evidence reveals significant actions of FMRP in modulating presynaptic function [18] suggesting its involvement in presynaptic forms of synaptic plasticity. For instance, FXS affects forms of long-term synaptic plasticity that reside on presynaptic mechanisms [20]. Furthermore, targets of FMRP include mRNAs that encode a plethora of presynaptic proteins involved in the process of neurotransmitter release [23,38]. For example, FMRP regulates neurotransmitter release by modulating calcium-depended potassium channels [17] and voltage-gated calcium channels [39]. Also, FMRP contributes to sustaining high-frequency synaptic activity by enhancing vesicle recycling through activity-dependent bulk endocytosis [40]. It is worth noting that some of these presynaptic actions are translation-independent [17].
Synaptic facilitation, a form of STSP that manifested as an increase of synaptic response upon rapidly repetitive activation of presynaptic terminal, is attributed to a transient increase of transmitter release probability resulted from accumulating Ca2+ in the presynaptic terminal during repetitive activation [35,41]. Thus, STSP may be a result of modulation of the neurotransmitter release process which is fundamentally supported by several presynaptic proteins including synaptotagmins, synaptobrevin, syntaxin, and SNAP-25 [42]. Synaptotagmins constitute a protein family of 17 members that function as calcium sensors [43,44]. Notably, synaptotagmin-1 (Syt-1) displays low Ca2+ affinity and is needed for synchronous neurotransmitter release [45,46], while synaptotagmin-7 (Syt-7) displays high calcium affinity and functions as a Ca2+ sensor to trigger synaptic vesicle fusion with plasma membrane at conditions of low Ca2+ concentrations [47,48,49]. Experimental data suggests that Syt-7 plays a significant role in asynchronous neurotransmitter release that occurs during repetitive synaptic activation [45,50] and significantly contributes to synaptic facilitation [51,52,53,54,55] and normal brain function [56,57]. Notably, Syt-7 may facilitate neurotransmitter release during the period that follows an action potential in the presynaptic terminal, by effectively sensing the low levels of Ca2+ during this period [58]. Interestingly, a previous report suggested that Syt-1 and Syt-7 may be modulated by FMRP [23]. However, whether the suppressed expression of FMRP affects the levels of synaptotagmins in the brain is still not known. We hypothesized that the loss of FMRP in FXS may alter STSP by affecting the expression and/or the function of Syt-7.
It should be noted that changes in STSP have been found in the hippocampus and cortex of immature FXS animals [17,19,21,59,60]; however, the effects of FXS on STSP in adult animals have not yet been sufficiently clarified. Dorsal and ventral hippocampus express remarkably different patterns of STSP [61,62,63,64,65,66,67,68,69,70,71,72,73], and they are involved in different ways with brain functions such as cognition and emotion [74]. Interestingly, the dorsal and ventral hippocampus have been recently shown to respond differently to FXS with respect to network excitability, GABAergic inhibition, and rhythmogenesis [75,76]. It is therefore necessary to study possible effects of FXS on STSP separately in the two segments of the hippocampus.
Finally, no studies have previously examined the effects of FXS on STSP separately in males and females. It is essential to investigate the possible sex-related neurobiological differences in FXS to understand the distinct FXS-associated behavioral disturbances encountered in the two sexes [77,78], especially in view of the currently under-investigation of females with FXS/autism disorder [79,80].
Here, using a rat model of FXS, we show that two forms of STSP, namely paired-pulse ratio (PPR) and frequency facilitation/depression (FF/D), as well as the protein expression of Syt-1 and Syt-7 are not altered in the hippocampus of adult males with FXS. However, PPR is increased in the ventral hippocampus of adult female littermates. Furthermore, except for KO females, DH displays higher levels of Syt-7 compared with VH, that parallel the difference in STSP between the two segments of the hippocampus.
2. Results
2.1. Genotype Affects Synaptic Facilitation in VH of Female Rats
We examined two phenomena of short-term synaptic plasticity (STSP) using a frequency stimulation protocol. Specifically, we studied paired-pulse responses (paired-pulse ratio, PPR) and frequency facilitation or depression (FF/D) of synaptic responses in DH and VH of WT and KO male and female rats. Frequency stimulation was applied using similar conditioning responses (fEPSP, mV/ms) in WT (0.68±0.01) and KO (0.71±0.01), and in DH (0.69±0.01) and VH (0.69±0.011). We found that both DH and VH of male rats exhibit similar pattern of STSP in the two genotypes (Figure 1). Specifically, PPR, frequency facilitation and frequency depression did not significantly differ between WT and KO male rats either in DH (Figure 1A,B) or VH (Figure 1C,D). In contrast, we found a significant effect of genotype on STSP in VH but not DH of female rats (Figure 2). More specifically, we found that either PPR or FF/D did not significantly differ between WT and KO female DH (Figure 2A,B). However, we observed a significantly higher PPR at the inter-pulse interval (IPI) of 25 ms, but no other IPIs, in VH-KO compared with VH-WT (independent t-test, t28 = -2.14, p=0.041) (Figure 2C,D). These results suggest that Fmr1-KO is associated with frequency-dependent alterations in the properties of STSP specifically in the VH of female but not male rats.
2.2. STSP Differs Between DH and VH from Male and Female Rats of Both Genotypes
Then we compared PPR and FF/D between the two segments of the hippocampus (Figure 3), as well as between males and females (Figure 4). In WT male rats, we confirmed previous observations showing that PPR and FF/D remarkably differ between DH and VH [61,62,63,64,65,66,67,68,69,70,72,73,81], (Figure 3A). Furthermore, we demonstrate that the remarkable dorsoventral differences in STSP were also maintained in the hippocampus of KO male rats (Figure 3B). In addition, we report for the first time that similar robust dorsoventral differences also exist in the female hippocampus, both in WT (Figure 3C) and KO (Figure 3D) rats.
2.3. Expression of Synaptotagmin-1 in WT and KO Rat Hippocampus
Syt-1 is a presynaptic protein crucially involved in synchronous neurotransmitter release [45,46]. Figure 5A shows that in male rats, Syt-1 is similarly expressed in WT and KO DH (t11 = -0.821, p = 0.429, WT=7 and KO=6) and VH (t11 = -1.101, p = 0.295, WT=7 and KO=6). Furthermore, we found similar levels of Syt-1 between DH and VH in both WT (t12 = -0.648, p = 0.529) and KO male rats (t10 = -1.119, p = 0.289). In contrast to male rats, we detected significant differences in the levels of Syt-1 in the female hippocampus. Specifically, Syt-1 is expressed in higher levels in KO than WT DH (t10 = -3.32, p = 0.008, WT=5 and KO=7), but not VH (t9 = -1.941, p = 0.084, WT=5 and KO=7) (Figure 6A) of female rats. Furthermore, we found similar levels of Syt-1 between DH and VH in both WT (t8 = -1.52, p = 0.168) and KO female rats (t11 = -0.507, p = 0.622).
2.4. Higher Levels of Synaptotagmin-7 in DH vs VH
Then, motivated by the recently proposed positive relationship between synaptic facilitation and expression of Syt-7 [51] we measured the levels of Syt-7 in the hippocampus of male WT and KO rats. We found that Syt-7 is similarly expressed in WT and KO DH (t14 = -1.00, p = 0.333, WT=8 and KO=8) and VH (t8.6 = -1.523, p = 0.163, WT=8 and KO=7) of male rats. Remarkably, we found a significantly lower Syt-7 protein expression in VH compared with DH in both WT (t14 = 2.586, p = 0.022) and KO (t8.49 = 2.72, p = 0.025) male rats (Figure 5B).
Regarding female rats, we found no significant change between WT and KO DH (t13 = -0.401, p = 0.695, WT=7 and KO=8) and VH (t9.17 = -1.755, p = 0.112, WT=7 and KO=8). However, we found significantly lower levels of Syt-7 in VH compared with DH in WT (t7.64 = 2.7, p = 0.028, DH=7 and VH=7) but not KO (t14 = 1.0, p = 0.332, DH=8 and VH=8) female rats (Figure 6B). These results suggest that the lower levels of Syt-7 in VH vs DH conforms with the lower synaptic facilitation seen in VH vs DH in both sexes, at least in WT rats.
2.5. Similar Expression of AMPA Receptor Subunits GluA1 and GluA2 in WT and KO Rats
Next, considering that, in addition to presynaptic mechanisms, postsynaptic mechanisms including AMPA receptors can be involved in shaping the properties of STSP [82], we aimed to examine the levels of AMPA receptors by measuring the protein expression of AMPA receptor subunits GluA1 and GluA2 in the different conditions. As shown in Figure 7 and Figure 8, the protein expression of GluA1 and GluA2 subunits in the isolated CA1 region did not significantly differ between WT and KO dorsal or ventral hippocampus either in males (Figure 7) or females (Figure 8). Also, GluA1 and GluA2 subunit expression in the two segments of the hippocampus was similar in males and females of either genotype. Notably, the GluA1 subunit was similarly expressed in male and female dorsal (t8 = -0.032, p = 0.97, male=5 and female=5) and ventral hippocampus (t7 = 0.637, p = 0.544, male=5 and female=4) of WT rats, and in male and female dorsal (t4.32 = 0.528, p = 0.624, male=4 and female=5) and ventral hippocampus (t5 = 0.549, p = 0.607, male=3 and female=4) of KO rats. Likewise, the expression of GluA2 subunit did not significantly differ between male and female dorsal (t5.24 = 0.404, p = 0.702, male=5 and female=5) and ventral hippocampus (t7 = -0.240, p = 0.817, male=5 and female=4) of WT rats, and between male and female dorsal (t7 = -1.512, p = 0.174, male=4 and female=5) and ventral hippocampus (t5 = -0.888, p = 0.415, male=3 and female=4) of KO rats.
2.6. Similar Excitatory Synaptic Transmission in DH and VH of WT and KO Male Rats
We also examined the possible effect of genotype on basal excitatory synaptic transmission in DH and VH from male and female rats by constructing input-output (I-O) curves between stimulation current intensity and fEPSP (Figure 9). We found no significant effect of genotype on I-O relationship in male DH (F9,511 = 0.920, p = 0.507; WT=29, KO=23; Figure 9A) and VH (F9,465 = 1.122, p = 0.346; WT=26, KO=21; Figure 9B). Similarly, I-O curves were similar in WT and KO female DH (F9,326 = 0.034, p = 0.999; WT=15, KO=19; Figure 9C) and VH (F9,214 = 0.639, p = 0.763; WT=15, KO=8; Figure 9D). Regarding male rats, the present results conform with those of a previous study [76]. However, another study reported increased basal excitatory synaptic transmission in the DH of KO vs WT male rats [75]. This inconsistency apparently results from the different measurement approach used in the two studies to estimate excitatory synaptic transmission; in the first study the whole I-O curve as well as the average fEPSP calculated from the whole curve were compared between the two genotypes, while in the second study the average fEPSPs evoked with moderate stimulation current intensity was measured. The data of the present study suggests that the genotype does not significantly affect basal excitatory synaptic transmission in either segment of the hippocampus of male or female rats. Furthermore, the electrophysiological results conform with the similar levels of Syt-1 in the two genotypes, at least in males.
2.7. Increased Excitatory Synaptic Transmission in DH vs VH of WT Female Rats
We also examined the effect of hippocampal segment on basal synaptic transmission by comparing I-O relationships between DH and VH (see right panels in Figure 9). We found no significant dorsoventral difference in fEPSP/I curves in either WT male rats (F9,541 = 0.179, p = 0.996; DH=29, VH=26; Figure 9A) or KO male rats (F9,435 = 0.122, p = 0.999; DH=23, VH=21; Figure 9B). In contrast to male rats, we found increased excitatory synaptic transmission in the DH compared with VH of WT female rats (F9,289 = 2.837, p = 0.003; DH=15, VH=15; Figure 9C) but not KO female rats (F9,251 = 0.376, p = 0.946; DH=19, VH=8; Figure 9D). The results regarding male WT rats confirm previously reported observations [63,75,83,84,85]. Regarding female rats, to the best of our knowledge this is the first time that basal excitatory synaptic transmission is compared between DH and VH in female rats.
2.8. Higher Excitatory Synaptic Transmission in Male vs Female Hippocampus
Comparing basal excitatory synaptic transmission between male and female rats we found that fEPSP was significantly higher in male than female WT rats, both in DH (F9,433 = 4.387, p < 0.001; male=29, female=15; Figure 10A) and VH (F9,397 = 8.136, p < 0.001; male=26, female=15; Figure 10B). Similarly, fEPSP was significantly higher in male than female KO rats in DH (F9,404 = 1.997, p = 0.039; male=23, female=19; Figure 10C) and VH (F9,511 = 1.86, p = 0.50; male=21, female=8; Figure 10D). Our observations concerning the gender-dependent effects on basal fEPSP in WT rats confirm the results of a previous study conducted in adult Long Evans rats which suggested that the increased basal excitatory synaptic transmission in males is most likely due to an excitatory effect of testosterone [86]. However, the effect of sex on basal synaptic transmission may depend on the strain of the animals used since another study reported greater fEPSPs in female than male Wistar rats [87]. In addition, we show that the effect of sex on basal transmission is maintained in KO rats, suggesting that the gender-related difference in basal excitatory synaptic transmission is unaffected by FXS.
3. Discussion
A key molecular signature of FXS is the loss of FMRP, a protein which plays profound roles in synaptic plasticity. It has been suggested that FMRP modulates presynaptic proteins that are involved in neurotransmitter release and short-term synaptic plasticity (STSP) including synaptotagmins. Recent evidence suggests that synaptotagmin-7 (Syt-7), a calcium sensor, plays a crucial role in synaptic facilitation, a ubiquitous form of STSP. Although some forms of STSP have been found altered in immature animals with FXS, whether the loss of FMRP affects STSP in adult animals remains elusive.
In the present study we investigated two forms of STSP, namely paired-pulse ratio (PPR) and frequency facilitation/depression (FF/D), and the expression of Syt-1, Syt-7, and AMPA receptors in the hippocampus of WT and Fmr1-KO rats. Notably, we prepared slices from the two segments of the hippocampus, the dorsal and the ventral, and from the two sexes, male and female. The main findings of this study are the following: a) STSP remains normal in both segments of the hippocampus of male rats and the DH of female KO rats but it is altered in the VH of female KO rats; b) STSP greatly differ between DH and VH, both in males and females; c) The large difference in STSP between DH and VH is paralleled by a corresponding difference in expression of Syt-7 both in males and females of either genotypes, except from the VH of female KO rats; d) basal excitatory synaptic transmission is higher in male than female hippocampus in both genotypes; e) AMPA receptors are similarly expressed in WT and KO, and no gender- or region-related difference was detected in receptor levels.
3.1. Effects of FXS on STSP and Synaptotagmins
In the present study we found that neither PPR nor FF/D are affected in the dorsal or ventral hippocampus of male KO rats. Our results on PPR are in keeping with previous studies reporting normal paired-pulse facilitation in the hippocampus of FXS rats [17,19,59,60]. However, regarding frequency facilitation, the present finding does not fit well with the results of previous reports which have shown increased responses at stimulation frequencies 20-100 Hz [17,19,60] or reduced responses at 10-20 Hz [21]. This inconsistency could be due to some important methodological differences. Notably, mouse models of FXS were used in previous studies while we used a rat model of FXS. However, most important for this discrepancy is the age difference of animals used in the present and previous studies. The previous studies were performed in immature animals aged between two and three weeks [17,19,21,59,60] and six weeks, while we used adult animals. FXS is a developmental disorder, and age matters in both neurobiological properties [88,89,90] and phenotypic traits [1]. For instance, seizures that are a common trait of FXS young individuals [2,91], often involving the anterior hippocampus (that corresponds to the ventral hippocampus in rodents), are absent in adults [2,91,92]. Moreover, the recently shown increase of GABAergic inhibition specifically in the ventral hippocampus of adult KO rats [75,76], the brain region most susceptible to epilepsy [93,94], may represent the result of homeostatic mechanisms activated during the development of individuals with the FXS, in an attempt to compensate for the increased excitability of the brain. Thus, it is tempting to speculate that the normal STSP we found in adult male KO rats may represent the result of adaptive mechanisms that attempt to restore the disrupted STSP in immature KO animals [17,19,21,60]. The data from a recent study using the valproic acid animal model of autism [95] are suggestive of the existence of such compensatory mechanisms during development.
Most of the previous studies that examined STSP in the hippocampus of FXS animals were performed in slices from the middle rather than the dorsal or the ventral hippocampus [17,19,21,60]; in one study no information was provided regarding the segment of the hippocampus used [59]. Furthermore, the previous studies did not clearly distinguish between males and females. Τhe present study is the first that examines the effects of FXS on hippocampal STSP separately in the two genders, and it reveals that FXS is associated with an increase in PPR specifically in the ventral hippocampus of adult female KO rats. To our best knowledge, there are no previously published data on STSP from female FXS animals. Given the multiple roles of FMRP in the synapse [8,12,13,18,19,20,21,22], these changes could be possibly related to the significant synaptic reorganization that takes place in the hippocampus of Fmr1-KO animals [21].
Curiously, frequency facilitation at stimulation frequencies around 40 Hz has been previously observed in studies where both males and females were used [19,60]. Interestingly, the increase in synaptic facilitation seen in the VH of KO female rats occurred at an IPI of 25 ms that corresponds to 40 Hz, the typical frequency of gamma oscillation that fundamentally supports processing of neural information in the brain [96], including processing of visual information, attention and episodic memory. Thus, the alteration in PPR observed here may be related to the previously reported impairment in sensory processing in females [97,98].
FMRP acts primarily to suppress translation of specific mRNAs, thereby affecting neuronal function and leading to the deficits observed in individuals with FXS [12,13]. However, not all mRNAs are FMRP targets [99]. Here, we show that neither Syt-1 nor Syt-7 are affected in adult FXS rats except Syt-1 which exhibits an increased expression in DH of adult female FXS rats. These results may indicate that synaptotagmins either escape from the consequences of FMRP loss, or their expression recovers during adulthood.
3.2. Dorsoventral Difference in STSP and a Possible Role of Synaptotagmin-7
In the present study we show that both forms of STSP, i.e., PPR and FF/D at CA3-CA1 synapses, greatly differ between the two segments of the hippocampus in male and female WT and KO rats. The difference in short-term forms of synaptic plasticity between DH and VH CA3-CA1 synapses has been repeatedly documented previously [61,62,63,64,65,66,67,68,69,70,72,73,81], representing one of the most prominent features of intrinsic diversification along the hippocampus. Therefore, the present results confirm previous ones and extend these findings to Fmr1-KO rats. However, even though this dorsoventral difference is well documented, the underlying mechanisms remain elusive. An early hypothesis, driven by the previously shown inverse relationship between basal probability of neurotransmitter release and synaptic facilitation [35,100], proposed that a high basal transmitter release probability may be responsible for the remarkably reduced synaptic facilitation of CA1 synapses in VH compared with DH [67,68,101]. However, this hypothesis has not been confirmed by more recent evidence [64,72,102].
Generally, several forms of STSP are thought to involve Ca2+-dependent mechanisms at the presynaptic terminal [35]. Notably, synaptic facilitation has been proposed to arise from an accumulation of Ca2+ in the cytosol of presynaptic terminal during repetitive activation, the so-called “residual Ca2+”, due to the slow action of mechanisms that restore Ca2+ concentration to resting levels, leading to an increased probability of neurotransmitter release [35,41,103]. The arrival of a subsequent action potential during the period of residual Ca2+ can lead to increased transmitter release (synaptic facilitation).
Presynaptic proteins that belong to the family of synaptotagmins, which function as Ca2+ sensors and trigger neurotransmitter release by interacting with the core membrane fusion proteins [44,45,58], appear to play fundamental roles in the process of neurotransmitter release and the Ca2+-dependent facilitation of release [43,44]. Among several members of this family, Syt-1 is a protein of synaptic vesicle membrane that functions as the primary sensor for Ca2+ in the presynaptic terminal, triggering the rapid fusion of synaptic vesicles with plasma membrane [45,46,104]. The low Ca2+ affinity of Syt-1 allows it to contribute decisively to the immediate transmitter release triggered by an action potential that leads to an abrupt and robust influx of Ca2+ in the vicinity of readily releasable synaptic vesicles [45,46,104].
Syt-7, on the other hand, exhibits high Ca2+ sensitivity and is activated at low Ca2+ concentrations, such as those occurring in the presynaptic terminal after an action potential invades the presynaptic terminal, when Ca2+ concentration falls but remains above the resting levels (residual Ca2+) [58]. Accordingly, recent data suggests that, in virtue of its property to be activated at low Ca2+ concentrations, Syt-7 can crucially support synaptic facilitation [51,52,53,54,55]. Thus, it might be expected that a relatively increased expression of Syt-7 in the presynaptic terminal would lead to a greater facilitation of synaptic transmission during rapidly repetitive activity. Indeed, the present study reveals a striking analogy between the dorsoventral difference in STSP and the levels of Syt-7 in the two segments of the hippocampus in both WT and KO male rats and WT female rats. It is therefore tempting to assume that the difference in synaptic facilitation observed between DH and VH could to some extent be attributed to the conspicuously different expression of Syt-7 between the two segments. The similar expression of Syt-7 in WT and KO rats suggests that Syt-7 remains unaffected by the loss of FMRP in adult rats. Thus, it will be interesting to see whether FXS affects the expression of Syt-7 in the hippocampus of immature WT and KO rats.
In addition to the presynaptic proteins that directly participate to the neurotransmitter release and facilitation of release process, other proteins might play important roles in synaptic facilitation. For instance, a recent study has revealed that activin A, a member of the transforming growth factor B family (TGF-β), in addition to its roles in a variety of functions such as cell proliferation and neuroprotection [105], it also contributes to shaping the frequency-dependent properties of synaptic facilitation in DH and VH [61]; however, activin A does not appear to be a crucial factor for the large dorsoventral difference in synaptic facilitation.
3.3. Conclusions
The present findings show that the loss of FMRP is associated with altered PPR in the VH of female KO rats. Furthermore, the increased synaptic facilitation seen in the dorsal vs ventral hippocampus of both genotypes is accompanied by a corresponding increased level of Syt-7, suggesting that Syt-7 plays a crucial role in differentiating the properties of STSP along the longitudinal axis of the hippocampus.
Here we could mention some limitations that are accompanied with the study. For example, we examined only two forms of STSP and found that they are not significantly altered in male KO animals. However, STSP includes a significant variety of phenomena that depend on both frequency and duration of activation (i.e., number of stimulation pulses), they may involve different mechanisms, potentially serve different functions, and may be affected differently in neurodevelopmental disorders. Therefore, we cannot rule out that there are changes in other STSP phenomena. Also, the increased PPR we found in the ventral hippocampus of KO females may possibly be underestimated, because WT females, due to their genetic diversity, can express FMRP to varying degrees. However, in the present study we did not quantitatively measure FMRP. Also, we did not measure estrogen levels, which could affect protein expression and synaptic plasticity.
Finally, the fact that data from different studies show that the effect of FXS on STSP varies during development is suggestive of the need to examine multiple developmental stages in a study. The elucidation of the developmental profile of STSP in both segments of the hippocampus would make it possible to identify the developmental stage in which the initial absence of FMRP is possibly followed by compensations that tend to maintain synaptic plasticity normal. Recent studies have suggested the possible existence of such compensatory mechanisms in the hippocampus of FXS rats. Development studies could constitute a proposed future perspective in investigating the effects of FXS on the neurophysiology of the hippocampus, contributing to the understanding of the causal relationship between loss of FMRP and STSP.
4. Materials and Methods
4.1. Animals
Fmr1 knockout and wildtype littermates Long Evans rats were obtained from the Medical College of Wisconsin (RRIDs: RGD_ 2308852 and RGD_ 11553873, respectively). Adult 3-4 months old of both sexes were used in this study. Specific pathogen-free rats were maintained at the Laboratory of Experimental Animals of the Department of Medicine of the University of Patras (license No: EL-13-BIOexp-04). Animals were maintained under a stable cycle of light-dark (12/12 h) and a temperature of 20-22 °C, and they had free access to food and water. The treatment of animals and all experimental procedures used in this study were conducted in accordance with the European Communities Council Directive Guidelines for the care and use of Laboratory animals (2010/63/EU – European Commission). Also, experimental procedures were approved by the Protocol Evaluation Committee of the Department of Medicine of the University of Patras and the Directorate of Veterinary Services of the Achaia Prefecture of Western Greece Region (reg. number: 5661/37, 18/01/2021). Furthermore, this animal study was re-viewed and approved by the Research Ethics Committee of the University of Patras. Rats were genotyped after each experiment using tail or brain tissue to test the expression of FMRP protein by means of Western blotting.
4.2. Slice Preparation
Slices were prepared from the dorsal and the ventral hippocampus as previously described [70]. Briefly, following decapitation of the animal under deep anesthesia with diethyl-ether, the brain was removed from the scull, and placed in chilled (2-4 oC) artificial cerebrospinal fluid (ACSF) containing (in mM) 124 NaCl, 4 KCl, 2 CaCl2, 2 MgSO4, 26 NaHCO3, 1.25 NaH2PO4 and 10 glucose. ACSF was equilibrated with 95% O2 and 5% CO2 gas mixture at a pH=7.4. After excising the two hippocampi from the hemispheres, we prepared 550 μm thick slices from the dorsal and ventral hippocampus cutting transversally to the long axis of the structures using a McIlwain tissue chopper. Slices were prepared from the segments of the hippocampus extending between 0.5 mm and 3.5 mm from each end. The slices were immediately transferred one by one, after preparation, in a home-made interface type (air-liquid) recording chamber where they maintained for the rest of the experiment continuously perfused with aCSF and humidified with gas mixture of the same composition as described above, at a temperature of 30±0.5 oC. Slices were perfused at a rate of ~1.5 ml/min and left to recover from the cut for at least one and a half hours after their placement in the recording chamber.
4.3. Electrophysiology
Recordings of evoked field excitatory postsynaptic potentials (fEPSPs) were performed from the stratum radiatum of the middle CA1 hippocampal region, using a 7 μm-thick carbon fiber electrode (Kation Scientific, Minneapolis, USA). fEPSPs were evoked using electrical stimulation of Schaffer collaterals by a home-made bipolar platinum/iridium wire electrode with a wire diameter of 25 μm (World Precision Instruments, USA), and an inter-wire distance of 100 μm. Stimulation current pulses of variable amplitude and stable duration of 100 μs were delivered using a DS3 constant current stimulator (Digitimer Ltd., UK). Stimulation and recording electrodes were placed in slices under visual guidance using three-axis mechanical micromanipulators (Narishige Group, Japan) and a stereo microscope (Olympus, Japan) under fibre optic lighting (Volpi AG, USA). We applied baseline stimulation at a frequency of 0.033 Hz using a current stimulation intensity evoking an fEPSP with a slope of about 1 mV/ms. We measured the maximum slope of the early rising phase of fEPSP. We assessed synaptic effectiveness by systematically constructing input-output (I-O) curves between increasing stimulation current intensity and fEPSP. Due to variation in stimulus intensity between experiments, current intensity was normalized in each experiment (i.e., slice) to the maximum current intensity value used. We studied STSP by using a frequency stimulation protocol as previously described [69,70]. Specifically, the frequency stimulation protocol consisted of short trains of ten pulses delivered at the frequencies of 5, 20, 40 and 100 Hz. Consecutive stimulation trains were separated by two-minute-long intervals. The effect of repeated stimulation on fEPSP was quantified by the percent change of either the second response or the 8-10th responses with respect to the first conditioning response. The former approach represents the paired-pulse ration (PPR), while the second approach provides a quantification of postsynaptic response change at steady-state. In addition, we calculated the average percentage of all conditioned responses (i.e., 2-10th).
The electrical signal was acquired and amplified X500 and then filtered at 0.5 Hz–2 kHz using Neurolog systems (Digitimer Ltd., UK). Analog signal was digitized at 10 kHz using a CED 1401-plus interface and the Signal software (Cambridge Electronic Design, Cambridge, UK). Signal was continuously monitored visually and auditory using an analog-to-digital oscilloscope (Hameg Instruments, Germany) and a Neurolog audio amplifier, respectively. Signal was stored on a computer disk for off-line analysis.
4.4. Immunoblotting
The CA1 region of the dorsal and ventral hippocampus of males and females, WT and KO rats and the remaining brain tissue or tail tissue were stored at −80 °C for protein expression analysis. Following SDS-PAGE electrophoresis on 10% polyacrylamide gels, proteins were transferred to polyvinylidene difluoride (PVDF) membrane at 400 mA for 90 min and membranes were blocked with PBS containing 0.1% Tween-20 (PBST) and 5% nonfat dried milk, at room temperature. Membranes were next incubated overnight at 4 °C with the following primary antibodies diluted in PBST, 3% dried milk: rabbit anti-FMRP polyclonal (1:1500, #17722, Abcam, Cambridge, UK), mouse anti-synaptotagmin 1 monoclonal (1:1000, #MAB5200, Millipore Sigma, Burlington, MA, USA), mouse anti-synaptotagmin 7 monoclonal (1:350, #MA5-27654, Thermo Fisher Scientific, Waltham, MA, USA), rabbit anti-GluA1 polyclonal (1:2000, #D4N9V, Cell Signaling Technology, Danvers, MA, USA), rabbit anti-GluA2 polyclonal (1:2000, #E1L8U, Cell Signaling Technology, Danvers, MA, USA), and rabbit anti-β-actin polyclonal (1:15,000, #E-AB-20058, Elabscience, Houston, TX, USA) antibodies. The blots were rinsed with PBST and then incubated with either goat anti-rabbit or anti-mouse secondary horseradish peroxidase-conjugated IgG antibodies for 60 min at RT. Immunodetection was carried out using an Enhanced Chemiluminescence detection system. The bands were visualized on ChemiDoc MP (BioRad, Hercules, CA, USA) with 1 to 10 min exposures. Optical density measurements from each band were defined as ROD units with ImageLab 6.1. The ROD of each band was quantified relative to the ROD of β-actin, which serves as a gel-loading control. Then, the ratio (ROD of protein of interest)/(ROD β-actin) was normalized with the same ratio of an internal sample, which was loaded in all gels.
4.5. Statistical Analysis
Values throughout the text represent mean±S.E.M. For comparisons between two different populations of data we used the independent t-test that accounted for unequal variances whenever needed. For comparisons of data with repeated measures we used the two-way ANOVA (UNIANOVA). The experimental unit was the slice in electrophysiology and the rat in western blotting. The IBM SPSS Statistics 27 software package was used for all statistical analyses.
Author Contributions
G.T., A.M., L.J.L., and G.Tr. performed electrophysiological experiments, and data analysis. G.T. and A.M. performed western blot experiments and analyzed the data. C.P. designed and supervised the project, performed data analysis, and prepared and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was co-financed by the European Union and Greek national funds through the Operational Program Competitiveness, Entrepreneurship and Innovation, under the call RESEARCH – CREATE – INNOVATE (project code: T2EDK - 02075).
Institutional Review Board Statement
The animal study protocol was approved by the Research Ethics Committee of the University of Patras and the Directorate of Veterinary Services of the Achaia Prefecture of Western Greece Region (reg. number: 5661/37, 18 January 2021). The treatment of animals and all experimental procedures used in this study were conducted in accordance with the European Communities Council Directive Guidelines for the care and use of Laboratory animals (2010/63/EU—European Commission).
Data Availability Statement
All data associated with this study are available from the corresponding author upon reasonable request.
Acknowledgments
G. Tsotsokou was financially supported by Andreas Mentzelopoulos Foundation and the “Polembros Shipping Limited”, as a recipient of Ph.D. fellowships.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
ACSF, artificial cerebrospinal fluid; fEPSP, field excitatory postsynaptic potential; DH, dorsal hippocampus; FF/D, frequency facilitation/depression; FXS, fragile X syndrome; KO, knockout; PPR, paired-pulse ratio; STSP, short-term synaptic plasticity; Syt-1, synaptotagmin-1; Syt-7, synaptotagmin-7; VH, ventral hippocampus; WT, wild-type.
References
- Hagerman, R.J.; Berry-Kravis, E.; Hazlett, H.C.; Bailey, D.B., Jr.; Moine, H.; Kooy, R.F.; Tassone, F.; Gantois, I.; Sonenberg, N.; Mandel, J.L.; et al. Fragile X syndrome. Nature reviews. Disease primers 2017, 3, 17065. [Google Scholar] [CrossRef] [PubMed]
- Kidd, S.A.; Lachiewicz, A.; Barbouth, D.; Blitz, R.K.; Delahunty, C.; McBrien, D.; Visootsak, J.; Berry-Kravis, E. Fragile X syndrome: a review of associated medical problems. Pediatrics 2014, 134, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.B., Jr.; Mesibov, G.B.; Hatton, D.D.; Clark, R.D.; Roberts, J.E.; Mayhew, L. Autistic behavior in young boys with fragile X syndrome. Journal of autism and developmental disorders 1998, 28, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, R.J.; Jackson, A.W., 3rd; Levitas, A.; Rimland, B.; Braden, M. An analysis of autism in fifty males with the fragile X syndrome. American journal of medical genetics 1986, 23, 359–374. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, W.E.; Kidd, S.A.; Andrews, H.F.; Budimirovic, D.B.; Esler, A.; Haas-Givler, B.; Stackhouse, T.; Riley, C.; Peacock, G.; Sherman, S.L.; et al. Autism Spectrum Disorder in Fragile X Syndrome: Cooccurring Conditions and Current Treatment. Pediatrics 2017, 139, S194–s206. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, R.J. Lessons from fragile X regarding neurobiology, autism, and neurodegeneration. Journal of developmental and behavioral pediatrics : JDBP 2006, 27, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, A.J.; Pieretti, M.; Sutcliffe, J.S.; Fu, Y.H.; Kuhl, D.P.; Pizzuti, A.; Reiner, O.; Richards, S.; Victoria, M.F.; Zhang, F.P.; et al. Identification of a gene (FMR-1) containing a CGG repeat coincident with a breakpoint cluster region exhibiting length variation in fragile X syndrome. Cell 1991, 65, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Bassell, G.J.; Warren, S.T. Fragile X syndrome: loss of local mRNA regulation alters synaptic development and function. Neuron 2008, 60, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Rylaarsdam, L.; Guemez-Gamboa, A. Genetic Causes and Modifiers of Autism Spectrum Disorder. Frontiers in cellular neuroscience 2019, 13, 385. [Google Scholar] [CrossRef]
- Gholizadeh, S.; Halder, S.K.; Hampson, D.R.J.B.r. Expression of fragile X mental retardation protein in neurons and glia of the developing and adult mouse brain. 2015, 1596, 22–30. [Google Scholar]
- Zorio, D.A.; Jackson, C.M.; Liu, Y.; Rubel, E.W.; Wang, Y.J.J.o.C.N. Cellular distribution of the fragile X mental retardation protein in the mouse brain. 2017, 525, 818–849. [Google Scholar]
- Beckel-Mitchener, A.; Greenough, W.T. Correlates across the structural, functional, and molecular phenotypes of fragile X syndrome. Mental retardation and developmental disabilities research reviews 2004, 10, 53–59. [Google Scholar] [CrossRef]
- Bear, M.F.; Dölen, G.; Osterweil, E.; Nagarajan, N. Fragile X: translation in action. Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology 2008, 33, 84–87. [Google Scholar] [CrossRef]
- Fyke, W.; Velinov, M. FMR1 and Autism, an Intriguing Connection Revisited. Genes 2021, 12. [Google Scholar] [CrossRef]
- Richter, J.D.; Zhao, X. The molecular biology of FMRP: new insights into fragile X syndrome. Nat Rev Neurosci 2021, 22, 209–222. [Google Scholar] [CrossRef]
- Brown, M.R.; Kronengold, J.; Gazula, V.R.; Chen, Y.; Strumbos, J.G.; Sigworth, F.J.; Navaratnam, D.; Kaczmarek, L.K. Fragile X mental retardation protein controls gating of the sodium-activated potassium channel Slack. Nature neuroscience 2010, 13, 819–821. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.Y.; Rotman, Z.; Blundon, J.A.; Cho, Y.; Cui, J.; Cavalli, V.; Zakharenko, S.S.; Klyachko, V.A. FMRP regulates neurotransmitter release and synaptic information transmission by modulating action potential duration via BK channels. Neuron 2013, 77, 696–711. [Google Scholar] [CrossRef]
- Wang, X.; Sela-Donenfeld, D.; Wang, Y. Axonal and presynaptic FMRP: Localization, signal, and functional implications. Hearing research 2023, 430, 108720. [Google Scholar] [CrossRef]
- Deng, P.Y.; Sojka, D.; Klyachko, V.A. Abnormal presynaptic short-term plasticity and information processing in a mouse model of fragile X syndrome. The Journal of neuroscience : the official journal of the Society for Neuroscience 2011, 31, 10971–10982. [Google Scholar] [CrossRef] [PubMed]
- Koga, K.; Liu, M.G.; Qiu, S.; Song, Q.; O’Den, G.; Chen, T.; Zhuo, M. Impaired presynaptic long-term potentiation in the anterior cingulate cortex of Fmr1 knock-out mice. The Journal of neuroscience : the official journal of the Society for Neuroscience 2015, 35, 2033–2043. [Google Scholar] [CrossRef] [PubMed]
- Klemmer, P.; Meredith, R.M.; Holmgren, C.D.; Klychnikov, O.I.; Stahl-Zeng, J.; Loos, M.; van der Schors, R.C.; Wortel, J.; de Wit, H.; Spijker, S.; et al. Proteomics, ultrastructure, and physiology of hippocampal synapses in a fragile X syndrome mouse model reveal presynaptic phenotype. The Journal of biological chemistry 2011, 286, 25495–25504. [Google Scholar] [CrossRef]
- Monday, H.R.; Kharod, S.C.; Yoon, Y.J.; Singer, R.H.; Castillo, P.E. Presynaptic FMRP and local protein synthesis support structural and functional plasticity of glutamatergic axon terminals. Neuron 2022, 110, 2588–2606 e2586. [Google Scholar] [CrossRef]
- Darnell, J.C.; Van Driesche, S.J.; Zhang, C.; Hung, K.Y.; Mele, A.; Fraser, C.E.; Stone, E.F.; Chen, C.; Fak, J.J.; Chi, S.W.; et al. FMRP stalls ribosomal translocation on mRNAs linked to synaptic function and autism. Cell 2011, 146, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Bhakar, A.L.; Dölen, G.; Bear, M.F. The pathophysiology of fragile X (and what it teaches us about synapses). Annual review of neuroscience 2012, 35, 417–443. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.G. NMDA receptors and memory encoding. Neuropharmacology 2013, 74, 32–40. [Google Scholar] [CrossRef]
- Jia, Z.; Lu, Y.M.; Agopyan, N.; Roder, J. Gene targeting reveals a role for the glutamate receptors mGluR5 and GluR2 in learning and memory. Physiology & behavior 2001, 73, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.R.; Bray, S.M.; Warren, S.T. Molecular mechanisms of fragile X syndrome: a twenty-year perspective. Annu Rev Pathol 2012, 7, 219–245. [Google Scholar] [CrossRef] [PubMed]
- Nosyreva, E.D.; Huber, K.M. Metabotropic receptor-dependent long-term depression persists in the absence of protein synthesis in the mouse model of fragile X syndrome. Journal of neurophysiology 2006, 95, 3291–3295. [Google Scholar] [CrossRef] [PubMed]
- Meredith, R.M.; Holmgren, C.D.; Weidum, M.; Burnashev, N.; Mansvelder, H.D.J.N. Increased threshold for spike-timing-dependent plasticity is caused by unreliable calcium signaling in mice lacking fragile X gene FMR1. 2007, 54, 627–638. [Google Scholar]
- Martin, H.G.S.; Lassalle, O.; Brown, J.T.; Manzoni, O.J. Age-Dependent Long-Term Potentiation Deficits in the Prefrontal Cortex of the Fmr1 Knockout Mouse Model of Fragile X Syndrome. Cereb Cortex 2016, 26, 2084–2092. [Google Scholar] [CrossRef] [PubMed]
- Hatton, D.D.; Buckley, E.; Lachiewicz, A.; Roberts, J. Ocular status of boys with fragile X syndrome: a prospective study. J AAPOS 1998, 2, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Segal, O.; Kowal, T.; Banet-Levi, Y.; Gabis, L.V. Executive Function and Working Memory Deficits in Females with Fragile X Premutation. Life (Basel, Switzerland) 2023, 13. [Google Scholar] [CrossRef]
- Devaraju, P.; Yu, J.; Eddins, D.; Mellado-Lagarde, M.M.; Earls, L.R.; Westmoreland, J.J.; Quarato, G.; Green, D.R.; Zakharenko, S.S. Haploinsufficiency of the 22q11.2 microdeletion gene Mrpl40 disrupts short-term synaptic plasticity and working memory through dysregulation of mitochondrial calcium. Molecular psychiatry 2017, 22, 1313–1326. [Google Scholar] [CrossRef]
- Pals, M.; Stewart, T.C.; Akyürek, E.G.; Borst, J.P. A functional spiking-neuron model of activity-silent working memory in humans based on calcium-mediated short-term synaptic plasticity. PLoS computational biology 2020, 16, e1007936. [Google Scholar] [CrossRef] [PubMed]
- Zucker, R.S.; Regehr, W.G. Short-term synaptic plasticity. Annual review of physiology 2002, 64, 355–405. [Google Scholar] [CrossRef] [PubMed]
- Christie, S.B.; Akins, M.R.; Schwob, J.E.; Fallon, J.R. The FXG: a presynaptic fragile X granule expressed in a subset of developing brain circuits. The Journal of neuroscience : the official journal of the Society for Neuroscience 2009, 29, 1514–1524. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Gutekunst, C.A.; Eberhart, D.E.; Yi, H.; Warren, S.T.; Hersch, S.M. Fragile X mental retardation protein: nucleocytoplasmic shuttling and association with somatodendritic ribosomes. The Journal of neuroscience : the official journal of the Society for Neuroscience 1997, 17, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Hanson, J.E.; Madison, D.V. Presynaptic FMR1 genotype influences the degree of synaptic connectivity in a mosaic mouse model of fragile X syndrome. The Journal of neuroscience : the official journal of the Society for Neuroscience 2007, 27, 4014–4018. [Google Scholar] [CrossRef] [PubMed]
- Ferron, L.; Nieto-Rostro, M.; Cassidy, J.S.; Dolphin, A.C. Fragile X mental retardation protein controls synaptic vesicle exocytosis by modulating N-type calcium channel density. Nature communications 2014, 5, 3628. [Google Scholar] [CrossRef] [PubMed]
- Bonnycastle, K.; Kind, P.C.; Cousin, M.A. FMRP Sustains Presynaptic Function via Control of Activity-Dependent Bulk Endocytosis. The Journal of neuroscience : the official journal of the Society for Neuroscience 2022, 42, 1618–1628. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A.; Leal, K.; Nanou, E. Calcium channels and short-term synaptic plasticity. The Journal of biological chemistry 2013, 288, 10742–10749. [Google Scholar] [CrossRef] [PubMed]
- Südhof, T.C.; Rothman, J.E. Membrane fusion: grappling with SNARE and SM proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef]
- Wolfes, A.C.; Dean, C. The diversity of synaptotagmin isoforms. Current opinion in neurobiology 2020, 63, 198–209. [Google Scholar] [CrossRef]
- Volynski, K.E.; Krishnakumar, S.S. Synergistic control of neurotransmitter release by different members of the synaptotagmin family. Current opinion in neurobiology 2018, 51, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Bacaj, T.; Wu, D.; Yang, X.; Morishita, W.; Zhou, P.; Xu, W.; Malenka, R.C.; Südhof, T.C. Synaptotagmin-1 and synaptotagmin-7 trigger synchronous and asynchronous phases of neurotransmitter release. Neuron 2013, 80, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Maximov, A.; Südhof, T.C. Autonomous function of synaptotagmin 1 in triggering synchronous release independent of asynchronous release. Neuron 2005, 48, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Satterfield, R.; Young, S.M., Jr.; Jonas, P. Triple Function of Synaptotagmin 7 Ensures Efficiency of High-Frequency Transmission at Central GABAergic Synapses. Cell reports 2017, 21, 2082–2089. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, A.; Tucker, W.C.; Chapman, E.R. Synaptotagmin isoforms couple distinct ranges of Ca2+, Ba2+, and Sr2+ concentration to SNARE-mediated membrane fusion. Molecular biology of the cell 2005, 16, 4755–4764. [Google Scholar] [CrossRef] [PubMed]
- Brandt, D.S.; Coffman, M.D.; Falke, J.J.; Knight, J.D. Hydrophobic contributions to the membrane docking of synaptotagmin 7 C2A domain: mechanistic contrast between isoforms 1 and 7. Biochemistry 2012, 51, 7654–7664. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Trussell, L.O. Inhibitory transmission mediated by asynchronous transmitter release. Neuron 2000, 26, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Jackman, S.L.; Turecek, J.; Belinsky, J.E.; Regehr, W.G. The calcium sensor synaptotagmin 7 is required for synaptic facilitation. Nature 2016, 529, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Sakurai, A.; Littleton, J.T.; Yoshihara, M. Synaptotagmin 7 switches short-term synaptic plasticity from depression to facilitation by suppressing synaptic transmission. Scientific reports 2021, 11, 4059. [Google Scholar] [CrossRef]
- Weyrer, C.; Turecek, J.; Harrison, B.; Regehr, W.G. Introduction of synaptotagmin 7 promotes facilitation at the climbing fiber to Purkinje cell synapse. Cell reports 2021, 36, 109719. [Google Scholar] [CrossRef]
- He, Y.; Kulasiri, D.; Liang, J. A mathematical model of synaptotagmin 7 revealing functional importance of short-term synaptic plasticity. Neural regeneration research 2019, 14, 621–631. [Google Scholar] [CrossRef]
- Djillani, A.; Bazinet, J.; Catterall, W.A. Synaptotagmin-7 Enhances Facilitation of Ca(v)2.1 Calcium Channels. eNeuro 2022, 9. [Google Scholar] [CrossRef]
- Xie, Y.; Zhi, K.; Meng, X. Effects and Mechanisms of Synaptotagmin-7 in the Hippocampus on Cognitive Impairment in Aging Mice. Molecular neurobiology 2021, 58, 5756–5771. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Wang, Q.W.; Liu, Y.N.; Marchetto, M.C.; Linker, S.; Lu, S.Y.; Chen, Y.; Liu, C.; Guo, C.; Xing, Z.; et al. Synaptotagmin-7 is a key factor for bipolar-like behavioral abnormalities in mice. Proceedings of the National Academy of Sciences of the United States of America 2020, 117, 4392–4399. [Google Scholar] [CrossRef]
- Sugita, S.; Han, W.; Butz, S.; Liu, X.; Fernández-Chacón, R.; Lao, Y.; Südhof, T.C. Synaptotagmin VII as a plasma membrane Ca(2+) sensor in exocytosis. Neuron 2001, 30, 459–473. [Google Scholar] [CrossRef]
- Zhang, J.; Hou, L.; Klann, E.; Nelson, D.L. Altered hippocampal synaptic plasticity in the FMR1 gene family knockout mouse models. Journal of neurophysiology 2009, 101, 2572–2580. [Google Scholar] [CrossRef]
- Wang, X.S.; Peng, C.Z.; Cai, W.J.; Xia, J.; Jin, D.; Dai, Y.; Luo, X.G.; Klyachko, V.A.; Deng, P.Y. Activity-dependent regulation of release probability at excitatory hippocampal synapses: a crucial role of fragile X mental retardation protein in neurotransmission. The European journal of neuroscience 2014, 39, 1602–1612. [Google Scholar] [CrossRef] [PubMed]
- Dahlmanns, M.; Valero-Aracama, M.J.; Dahlmanns, J.K.; Zheng, F.; Alzheimer, C. Tonic activin signaling shapes cellular and synaptic properties of CA1 neurons mainly in dorsal hippocampus. iScience 2023, 26, 108001. [Google Scholar] [CrossRef] [PubMed]
- Tidball, P.; Burn, H.V.; Teh, K.L.; Volianskis, A.; Collingridge, G.L.; Fitzjohn, S.M. Differential ability of the dorsal and ventral rat hippocampus to exhibit group I metabotropic glutamate receptor-dependent synaptic and intrinsic plasticity. Brain and neuroscience advances 2017, 1. [Google Scholar] [CrossRef]
- Milior, G.; Castro, M.A.; Sciarria, L.P.; Garofalo, S.; Branchi, I.; Ragozzino, D.; Limatola, C.; Maggi, L. Electrophysiological Properties of CA1 Pyramidal Neurons along the Longitudinal Axis of the Mouse Hippocampus. Scientific reports 2016, 6, 38242. [Google Scholar] [CrossRef]
- Babiec, W.E.; Jami, S.A.; Guglietta, R.; Chen, P.B.; O’Dell, T.J. Differential Regulation of NMDA Receptor-Mediated Transmission by SK Channels Underlies Dorsal-Ventral Differences in Dynamics of Schaffer Collateral Synaptic Function. The Journal of neuroscience : the official journal of the Society for Neuroscience 2017, 37, 1950–1964. [Google Scholar] [CrossRef]
- Dubovyk, V.; Manahan-Vaughan, D. Less means more: The magnitude of synaptic plasticity along the hippocampal dorso-ventral axis is inversely related to the expression levels of plasticity-related neurotransmitter receptors. Hippocampus 2018, 28, 136–150. [Google Scholar] [CrossRef]
- Maruki, K.; Izaki, Y.; Nomura, M.; Yamauchi, T. Differences in paired-pulse facilitation and long-term potentiation between dorsal and ventral CA1 regions in anesthetized rats. Hippocampus 2001, 11, 655–661. [Google Scholar] [CrossRef]
- Papatheodoropoulos, C.; Kostopoulos, G. Dorsal-ventral differentiation of short-term synaptic plasticity in rat CA1 hippocampal region. Neuroscience letters 2000, 286, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Papatheodoropoulos, C. Striking differences in synaptic facilitation along the dorsoventral axis of the hippocampus. Neuroscience 2015, 301, 454–470. [Google Scholar] [CrossRef] [PubMed]
- Papaleonidopoulos, V.; Trompoukis, G.; Koutsoumpa, A.; Papatheodoropoulos, C. A gradient of frequency-dependent synaptic properties along the longitudinal hippocampal axis. BMC neuroscience 2017, 18, 79. [Google Scholar] [CrossRef]
- Koutsoumpa, A.; Papatheodoropoulos, C. Short-term dynamics of input and output of CA1 network greatly differ between the dorsal and ventral rat hippocampus. BMC neuroscience 2019, 20, 35. [Google Scholar] [CrossRef]
- Koutsoumpa, A.; Papatheodoropoulos, C. Frequency-dependent layer-specific differences in short-term synaptic plasticity in the dorsal and ventral CA1 hippocampal field. Synapse 2021, e22199. [Google Scholar] [CrossRef] [PubMed]
- Samara, M.; Oikonomou, G.D.; Trompoukis, G.; Madarou, G.; Adamopoulou, M.; Papatheodoropoulos, C. Septotemporal variation in modulation of synaptic transmission, paired-pulse ratio and frequency facilitation/depression by adenosine and GABA(B) receptors in the rat hippocampus. Brain and neuroscience advances 2022, 6, 23982128221106315. [Google Scholar] [CrossRef]
- Trompoukis, G.; Tsotsokou, G.; Koutsoumpa, A.; Tsolaki, M.; Vryoni, G.; Papatheodoropoulos, C. Age-dependent modulation of short-term neuronal dynamics in the dorsal and ventral rat hippocampus. The International journal of developmental biology 2022, 66, 285–296. [Google Scholar] [CrossRef]
- Strange, B.A.; Witter, M.P.; Lein, E.S.; Moser, E.I. Functional organization of the hippocampal longitudinal axis. Nat Rev Neurosci 2014, 15, 655–669. [Google Scholar] [CrossRef]
- Leontiadis, L.J.; Trompoukis, G.; Felemegkas, P.; Tsotsokou, G.; Miliou, A.; Papatheodoropoulos, C. Increased Inhibition May Contribute to Maintaining Normal Network Function in the Ventral Hippocampus of a Fmr1-Targeted Transgenic Rat Model of Fragile X Syndrome. Brain sciences 2023, 13, 1598. [Google Scholar] [CrossRef]
- Leontiadis, L.J.; Trompoukis, G.; Tsotsokou, G.; Miliou, A.; Felemegkas, P.; Papatheodoropoulos, C. Rescue of sharp wave-ripples and prevention of network hyperexcitability in the ventral but not the dorsal hippocampus of a rat model of fragile X syndrome. Frontiers in cellular neuroscience 2023, 17. [Google Scholar] [CrossRef]
- Napolitano, A.; Schiavi, S.; La Rosa, P.; Rossi-Espagnet, M.C.; Petrillo, S.; Bottino, F.; Tagliente, E.; Longo, D.; Lupi, E.; Casula, L.; et al. Sex Differences in Autism Spectrum Disorder: Diagnostic, Neurobiological, and Behavioral Features. Frontiers in psychiatry 2022, 13, 889636. [Google Scholar] [CrossRef] [PubMed]
- Werling, D.M.; Geschwind, D.H. Sex differences in autism spectrum disorders. Current opinion in neurology 2013, 26, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Calderoni, S. Sex/gender differences in children with autism spectrum disorder: A brief overview on epidemiology, symptom profile, and neuroanatomy. Journal of neuroscience research 2023, 101, 739–750. [Google Scholar] [CrossRef]
- Jeon, S.J.; Gonzales, E.L.; Mabunga, D.F.N.; Valencia, S.T.; Kim, D.G.; Kim, Y.; Adil, K.J.L.; Shin, D.; Park, D.; Shin, C.Y. Sex-specific Behavioral Features of Rodent Models of Autism Spectrum Disorder. Experimental neurobiology 2018, 27, 321–343. [Google Scholar] [CrossRef] [PubMed]
- Koutsoumpa, A.; Papatheodoropoulos, C. Frequency-dependent layer-specific differences in short-term synaptic plasticity in the dorsal and ventral CA1 hippocampal field. Synapse 2021, 75, e22199. [Google Scholar] [CrossRef]
- Regehr, W.G. Short-term presynaptic plasticity. Cold Spring Harbor perspectives in biology 2012, 4, a005702. [Google Scholar] [CrossRef]
- Kouvaros, S.; Papatheodoropoulos, C. Theta burst stimulation-induced LTP: Differences and similarities between the dorsal and ventral CA1 hippocampal synapses. Hippocampus 2016, 26, 1542–1559. [Google Scholar] [CrossRef]
- Papaleonidopoulos, V.; Papatheodoropoulos, C. beta-adrenergic receptors reduce the threshold for induction and stabilization of LTP and enhance its magnitude via multiple mechanisms in the ventral but not the dorsal hippocampus. Neurobiology of learning and memory 2018, 151, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Kenney, J.; Manahan-Vaughan, D. NMDA receptor-dependent synaptic plasticity in dorsal and intermediate hippocampus exhibits distinct frequency-dependent profiles. Neuropharmacology 2013, 74, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.D.; Jones, L.S.; Wilson, M.A. Sex differences in hippocampal slice excitability: role of testosterone. Neuroscience 2002, 109, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Monfort, P.; Gomez-Gimenez, B.; Llansola, M.; Felipo, V. Gender differences in spatial learning, synaptic activity, and long-term potentiation in the hippocampus in rats: molecular mechanisms. ACS chemical neuroscience 2015, 6, 1420–1427. [Google Scholar] [CrossRef] [PubMed]
- Harlow, E.G.; Till, S.M.; Russell, T.A.; Wijetunge, L.S.; Kind, P.; Contractor, A.J.N. Critical period plasticity is disrupted in the barrel cortex of FMR1 knockout mice. 2010, 65, 385–398. [Google Scholar]
- Galvez, R.; Greenough, W.T.J.A.j.o.m.g.P.A. Sequence of abnormal dendritic spine development in primary somatosensory cortex of a mouse model of the fragile X mental retardation syndrome. 2005, 135, 155–160. [Google Scholar]
- Nimchinsky, E.J.A.d.o.d.s.i.F.k.-o.m.J.N. Oberlander AM, and Svoboda K. 2001, 21, 5139–5146. [Google Scholar]
- Berry-Kravis, E.; Filipink, R.A.; Frye, R.E.; Golla, S.; Morris, S.M.; Andrews, H.; Choo, T.H.; Kaufmann, W.E. Seizures in Fragile X Syndrome: Associations and Longitudinal Analysis of a Large Clinic-Based Cohort. Frontiers in pediatrics 2021, 9, 736255. [Google Scholar] [CrossRef] [PubMed]
- Sabaratnam, M.; Vroegop, P.G.; Gangadharan, S.K. Epilepsy and EEG findings in 18 males with fragile X syndrome. Seizure 2001, 10, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Spencer, D.D.; Spencer, S.S.; Mattson, R.H.; Williamson, P.D.; Novelly, R.A. Access to the posterior medial temporal lobe structures in the surgical treatment of temporal lobe epilepsy. Neurosurgery 1984, 15, 667–671. [Google Scholar] [CrossRef]
- Babb, T.L.; Brown, W.J.; Pretorius, J.; Davenport, C.; Lieb, J.P.; Crandall, P.H. Temporal lobe volumetric cell densities in temporal lobe epilepsy. Epilepsia 1984, 25, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Bódi, V.; Májer, T.; Kelemen, V.; Világi, I.; Szűcs, A.; Varró, P. Alterations of the Hippocampal Networks in Valproic Acid-Induced Rat Autism Model. Frontiers in neural circuits 2022, 16, 772792. [Google Scholar] [CrossRef]
- Singer, W.; Gray, C.M. Visual feature integration and the temporal correlation hypothesis. Annual review of neuroscience 1995, 18, 555–586. [Google Scholar] [CrossRef]
- Steyaert, J.; Legius, E.; Borghgraef, M.; Fryns, J.P. A distinct neurocognitive phenotype in female fragile-X premutation carriers assessed with visual attention tasks. Am J Med Genet A 2003, 116a, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Norris, J.E.; Schmitt, L.M.; De Stefano, L.A.; Pedapati, E.V.; Erickson, C.A.; Sweeney, J.A.; Ethridge, L.E. Neuropsychiatric feature-based subgrouping reveals neural sensory processing spectrum in female FMR1 premutation carriers: A pilot study. Frontiers in integrative neuroscience 2023, 17, 898215. [Google Scholar] [CrossRef] [PubMed]
- Shanks, N.F.; Savas, J.N.; Maruo, T.; Cais, O.; Hirao, A.; Oe, S.; Ghosh, A.; Noda, Y.; Greger, I.H.; Yates, J.R., 3rd; et al. Differences in AMPA and kainate receptor interactomes facilitate identification of AMPA receptor auxiliary subunit GSG1L. Cell reports 2012, 1, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Dobrunz, L.E.; Stevens, C.F. Heterogeneity of release probability, facilitation, and depletion at central synapses. Neuron 1997, 18, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Papatheodoropoulos, C. Electrophysiological evidence for long-axis intrinsic diversification of the hippocampus. Frontiers in Bioscience (Landmark Ed.). 2018, 23, 109–145. [Google Scholar] [CrossRef] [PubMed]
- Trompoukis, G.; Papatheodoropoulos, C. Dorsal-Ventral Differences in Modulation of Synaptic Transmission in the Hippocampus. Frontiers in synaptic neuroscience 2020, 12, 24. [Google Scholar] [CrossRef] [PubMed]
- Katz, B.; Miledi, R. The role of calcium in neuromuscular facilitation. The Journal of physiology 1968, 195, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Geppert, M.; Goda, Y.; Hammer, R.E.; Li, C.; Rosahl, T.W.; Stevens, C.F.; Südhof, T.C. Synaptotagmin I: a major Ca2+ sensor for transmitter release at a central synapse. Cell 1994, 79, 717–727. [Google Scholar] [CrossRef]
- McDowell, N.; Zorn, A.M.; Crease, D.J.; Gurdon, J.B. Activin has direct long-range signalling activity and can form a concentration gradient by diffusion. Current biology : CB 1997, 7, 671–681. [Google Scholar] [CrossRef]
Figure 1.
Genotype does not affect STSP in the adult male DH and VH. Results are shown for DH (A,B) and VH (C,D). A & C: Representative traces of synaptic responses at 5 Hz, 20 Hz, 40 Hz, and 100 Hz stimulation trains (top panels), and the corresponding percent change of responses during stimulation trains (bottom panels) are shown for WT and KO male rats. B & D. Average paired-pulse ratio (PPR) at four different inter-pulse intervals (IPIs) (left), percentage of steady-state response (average of 8th -10th responses) plotted as a function of stimulus frequency (middle), and percentage of all conditioned responses (average of 2th -10th responses) plotted as a function of stimulus frequency (right). Superimposed sample traces of paired-pulse responses from WT and KO are shown in inserts; scale bars: 25 ms, 1 mV. The results of UNIANOVA are shown on the bottom of graphs in B and D.
Figure 1.
Genotype does not affect STSP in the adult male DH and VH. Results are shown for DH (A,B) and VH (C,D). A & C: Representative traces of synaptic responses at 5 Hz, 20 Hz, 40 Hz, and 100 Hz stimulation trains (top panels), and the corresponding percent change of responses during stimulation trains (bottom panels) are shown for WT and KO male rats. B & D. Average paired-pulse ratio (PPR) at four different inter-pulse intervals (IPIs) (left), percentage of steady-state response (average of 8th -10th responses) plotted as a function of stimulus frequency (middle), and percentage of all conditioned responses (average of 2th -10th responses) plotted as a function of stimulus frequency (right). Superimposed sample traces of paired-pulse responses from WT and KO are shown in inserts; scale bars: 25 ms, 1 mV. The results of UNIANOVA are shown on the bottom of graphs in B and D.
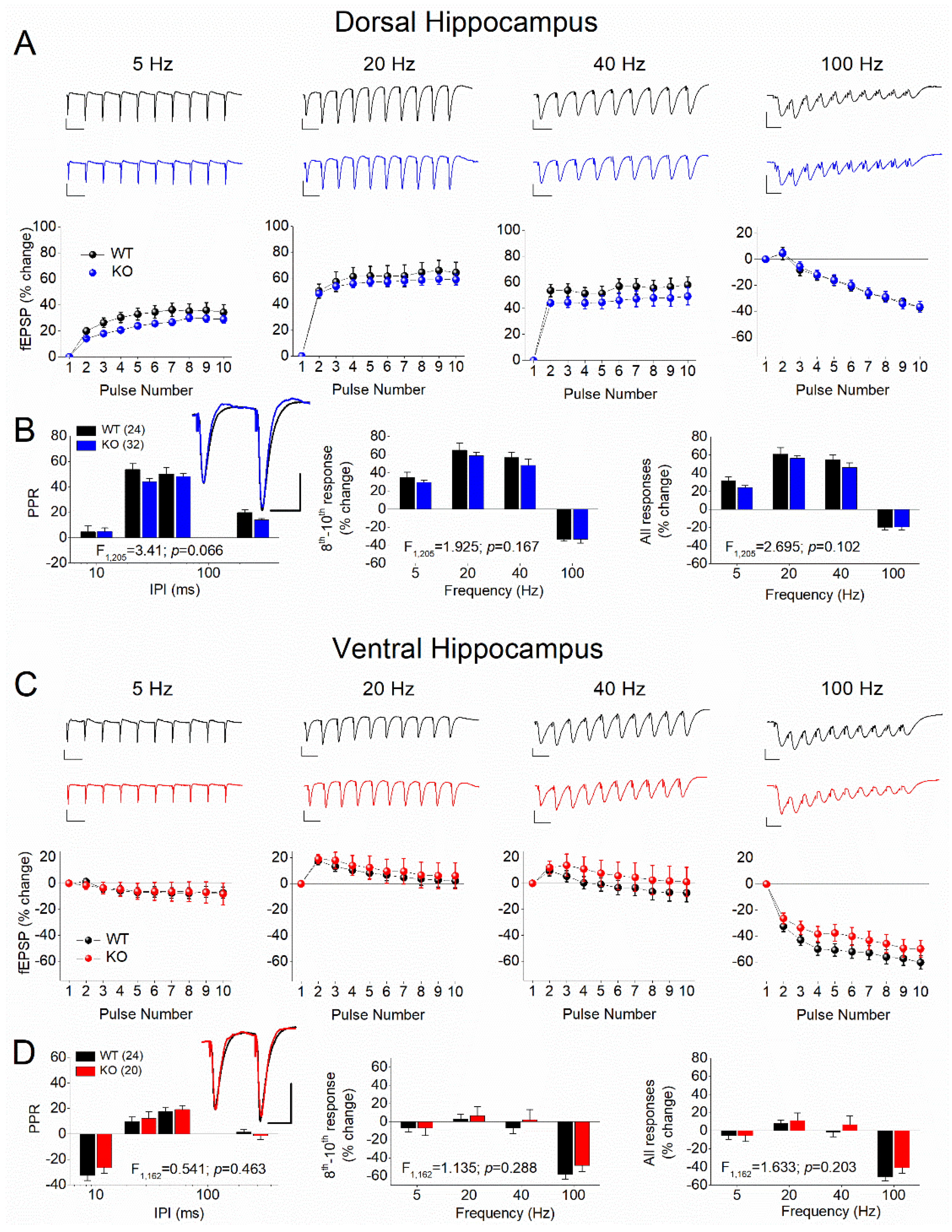
Figure 2.
Genotype affects STSP in the adult female VH but not DH. Results are shown for DH (A,B) and VH (C,D). A & C: Representative traces of synaptic responses at 5 Hz, 20 Hz, 40 Hz, and 100 Hz stimulation trains (top panels), and the corresponding percent change of responses during stimulation trains (bottom panels) are shown for WT and KO female rats. B & D. Average paired-pulse ratio (PPR) at four different inter-pulse intervals (IPIs) (left), percentage of steady-state response (average of 8th -10th responses) plotted as a function of stimulus frequency (middle), and percentage of all conditioned responses (average of 2th -10th responses) plotted as a function of stimulus frequency (right). Superimposed sample traces of paired-pulse responses from WT and KO are shown in inserts; scale bars: 25 ms, 1 mV. The results of UNIANOVA are shown on the bottom of graphs in B and D. Asterisk in D denotes statistically significant difference between WT and KO.
Figure 2.
Genotype affects STSP in the adult female VH but not DH. Results are shown for DH (A,B) and VH (C,D). A & C: Representative traces of synaptic responses at 5 Hz, 20 Hz, 40 Hz, and 100 Hz stimulation trains (top panels), and the corresponding percent change of responses during stimulation trains (bottom panels) are shown for WT and KO female rats. B & D. Average paired-pulse ratio (PPR) at four different inter-pulse intervals (IPIs) (left), percentage of steady-state response (average of 8th -10th responses) plotted as a function of stimulus frequency (middle), and percentage of all conditioned responses (average of 2th -10th responses) plotted as a function of stimulus frequency (right). Superimposed sample traces of paired-pulse responses from WT and KO are shown in inserts; scale bars: 25 ms, 1 mV. The results of UNIANOVA are shown on the bottom of graphs in B and D. Asterisk in D denotes statistically significant difference between WT and KO.
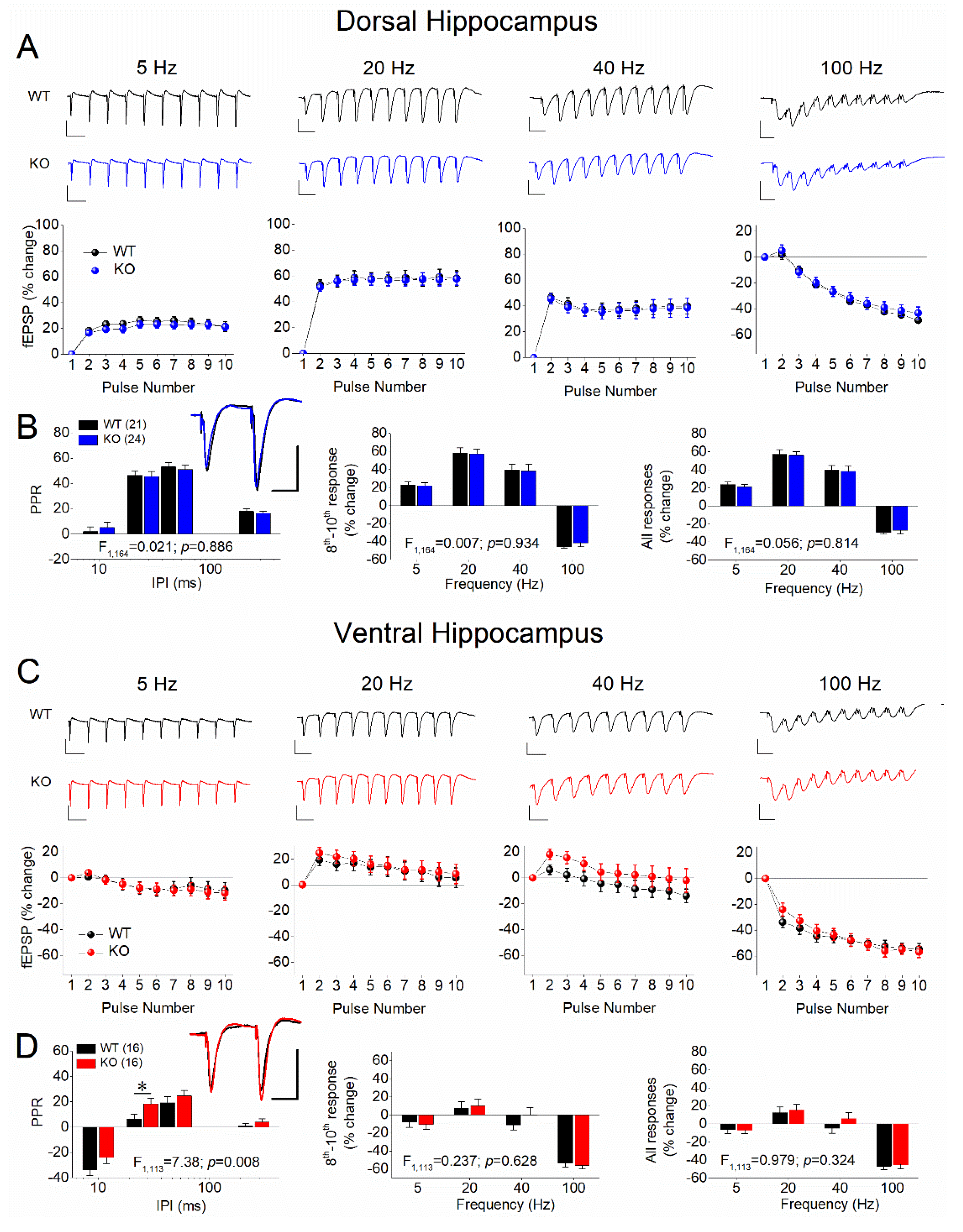
Figure 3.
STSP strongly differs between DH and VH. Results are shown for males (A,B) and females (C,D), as well as for WT (A,C) and KO (B,D) rats. In each of the four panels (A,B,C,D) are shown: the average PPR (left), the percentage of steady-state response (average of 8th -10th responses), and the percentage of all conditioned responses (average of 2th -10th responses). The results of UNIANOVA, comparing STSP between the two segments of the hippocampus are shown on the bottom of each graph. Data presented here are replotted from Figure 1 and Figure 2 to illustrate the dorsoventral differences in STSP.
Figure 3.
STSP strongly differs between DH and VH. Results are shown for males (A,B) and females (C,D), as well as for WT (A,C) and KO (B,D) rats. In each of the four panels (A,B,C,D) are shown: the average PPR (left), the percentage of steady-state response (average of 8th -10th responses), and the percentage of all conditioned responses (average of 2th -10th responses). The results of UNIANOVA, comparing STSP between the two segments of the hippocampus are shown on the bottom of each graph. Data presented here are replotted from Figure 1 and Figure 2 to illustrate the dorsoventral differences in STSP.
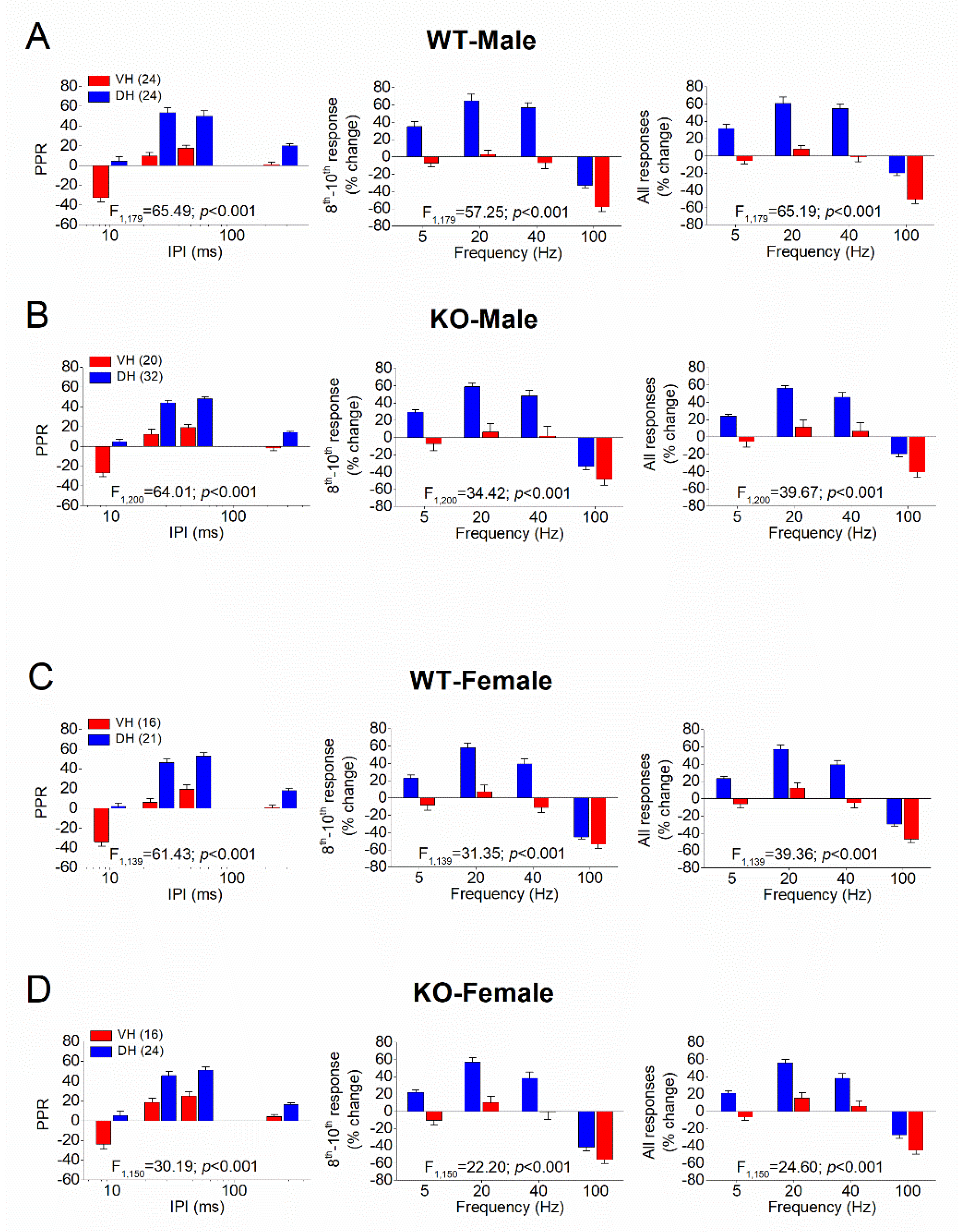
Figure 4.
STSP is similar in males and females. Results are shown for WT (A,B) and KO (C,D) rats, as well as for DH (A,C) and VH (B,D). In each of the four panels (A,B,C,D) are shown: the average PPR (left), the percentage of steady-state response (average of 8th -10th responses), and the percentage of all conditioned responses (average of 2th -10th responses). The results of UNIANOVA, comparing STSP between the two segments of the hippocampus are shown on the bottom of each graph. Data presented here are replotted from Figure 1 and Figure 2 to illustrate the similarity in STSP between the two sexes.
Figure 4.
STSP is similar in males and females. Results are shown for WT (A,B) and KO (C,D) rats, as well as for DH (A,C) and VH (B,D). In each of the four panels (A,B,C,D) are shown: the average PPR (left), the percentage of steady-state response (average of 8th -10th responses), and the percentage of all conditioned responses (average of 2th -10th responses). The results of UNIANOVA, comparing STSP between the two segments of the hippocampus are shown on the bottom of each graph. Data presented here are replotted from Figure 1 and Figure 2 to illustrate the similarity in STSP between the two sexes.
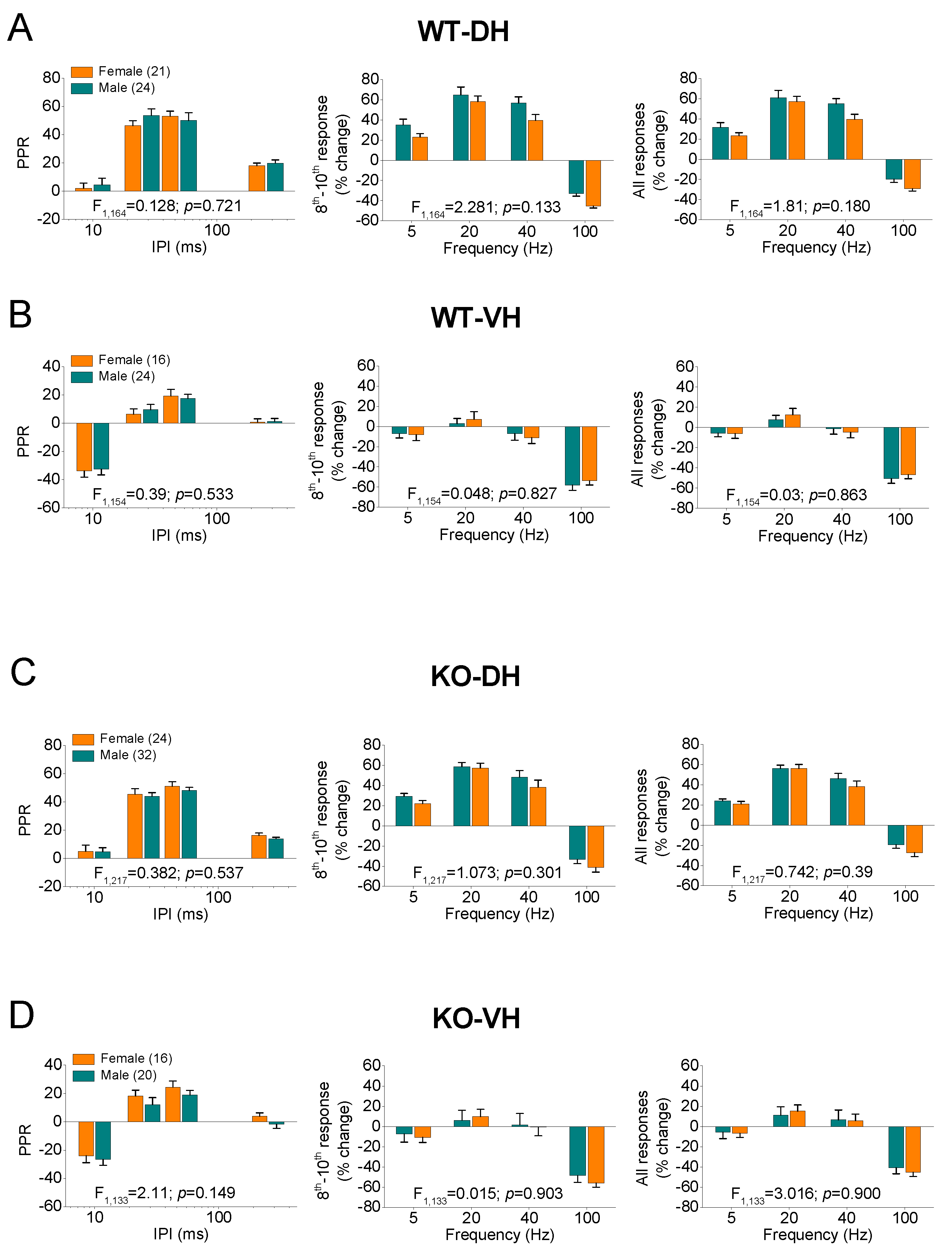
Figure 5.
Expression of Synaptotagmin-1 (Syt-1) and Synaptotagmin-7 (Syt-7) in the CA1 hippocampal region of male rats. Syt-7 but not Syt-1 levels differ between DH and VH but not between WT and KO male rats. A. Protein expression of Syt-1 is similar between WT and KO and between DH and VH male rats. B. Protein expression of Syt-7 significantly differ between DH and VH in both genotypes but does not differ between the two genotypes in either segment of the hippocampus. Asterisks denote statistically significant differences at p < 0.05 (independent t-test).
Figure 5.
Expression of Synaptotagmin-1 (Syt-1) and Synaptotagmin-7 (Syt-7) in the CA1 hippocampal region of male rats. Syt-7 but not Syt-1 levels differ between DH and VH but not between WT and KO male rats. A. Protein expression of Syt-1 is similar between WT and KO and between DH and VH male rats. B. Protein expression of Syt-7 significantly differ between DH and VH in both genotypes but does not differ between the two genotypes in either segment of the hippocampus. Asterisks denote statistically significant differences at p < 0.05 (independent t-test).
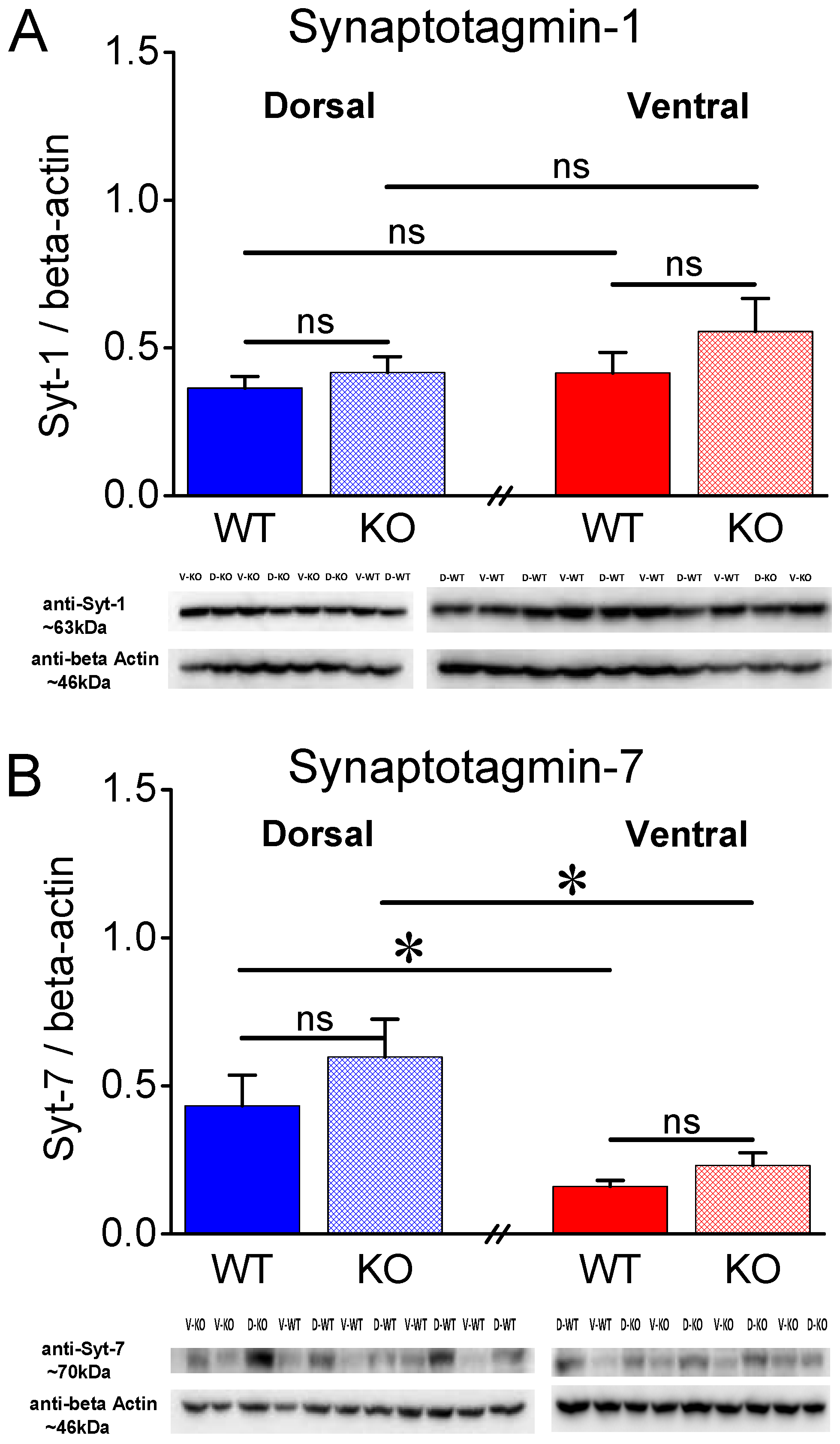
Figure 6.
Expression of Syt-1 and Syt-7 in the CA1 hippocampal region of female rats. A. Syt-1 exhibits a higher expression in KO than WT DH but not VH of female rats. B. Protein expression levels of Syt-7 are significantly lower in VH than DH WT but not in KO female rats. Furthermore, Syt-7 is similarly expressed in the two genotypes. Asterisks denote statistically significant differences at p < 0.05 (independent t-test).
Figure 6.
Expression of Syt-1 and Syt-7 in the CA1 hippocampal region of female rats. A. Syt-1 exhibits a higher expression in KO than WT DH but not VH of female rats. B. Protein expression levels of Syt-7 are significantly lower in VH than DH WT but not in KO female rats. Furthermore, Syt-7 is similarly expressed in the two genotypes. Asterisks denote statistically significant differences at p < 0.05 (independent t-test).
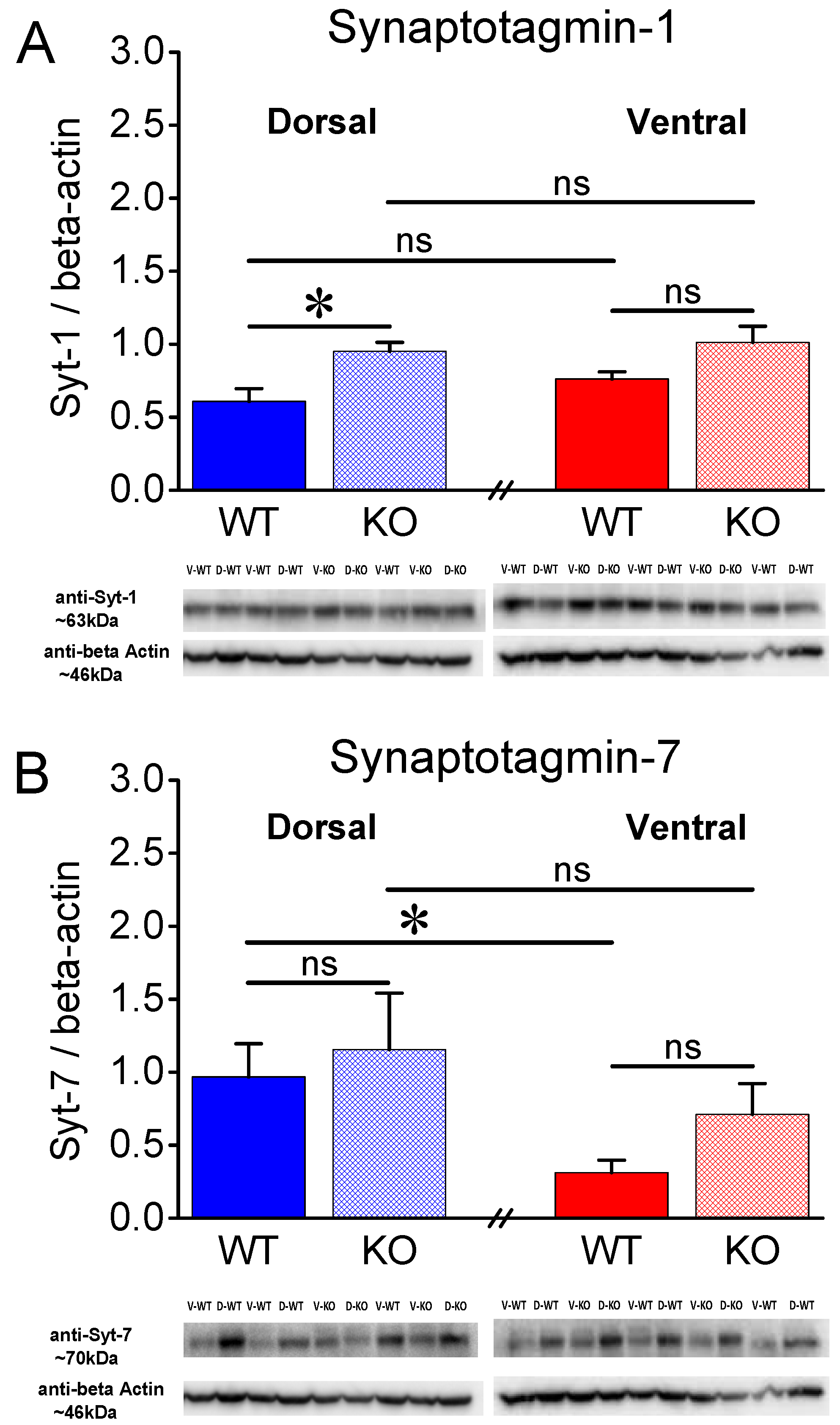
Figure 7.
Expression of AMPA receptor subunits GluA1 and GluA2 in the CA1 region of male rats. A. The GluA1 subunit was similarly expressed in WT and KO dorsal (t3.56 = -0.818, p = 0.465, WT=5 and KO=4) and ventral hippocampus (t6 = -0.878, p = 0.414, WT=5 and KO=3) of WT rats. Also, the expression of GluA1 is similar in DH and VH of WT rats (t8 = 1.78, p = 0.113), and in DH and VH of KO rats (t3.95 = 1.081, p = 0.341). B. The GluA2 subunit was similarly expressed in WT and KO dorsal (t4.43 = 1.503, p = 0.2, WT=5 and KO=4) and ventral hippocampus (t6 = 0.41, p = 0.696, WT=5 and KO=3) of WT rats. Also, the expression of GluA2 is similar in DH and VH of WT rats (t8 = 1.101, p = 0.303), and in DH and VH of KO rats (t5 = 0.841, p = 0.439).
Figure 7.
Expression of AMPA receptor subunits GluA1 and GluA2 in the CA1 region of male rats. A. The GluA1 subunit was similarly expressed in WT and KO dorsal (t3.56 = -0.818, p = 0.465, WT=5 and KO=4) and ventral hippocampus (t6 = -0.878, p = 0.414, WT=5 and KO=3) of WT rats. Also, the expression of GluA1 is similar in DH and VH of WT rats (t8 = 1.78, p = 0.113), and in DH and VH of KO rats (t3.95 = 1.081, p = 0.341). B. The GluA2 subunit was similarly expressed in WT and KO dorsal (t4.43 = 1.503, p = 0.2, WT=5 and KO=4) and ventral hippocampus (t6 = 0.41, p = 0.696, WT=5 and KO=3) of WT rats. Also, the expression of GluA2 is similar in DH and VH of WT rats (t8 = 1.101, p = 0.303), and in DH and VH of KO rats (t5 = 0.841, p = 0.439).
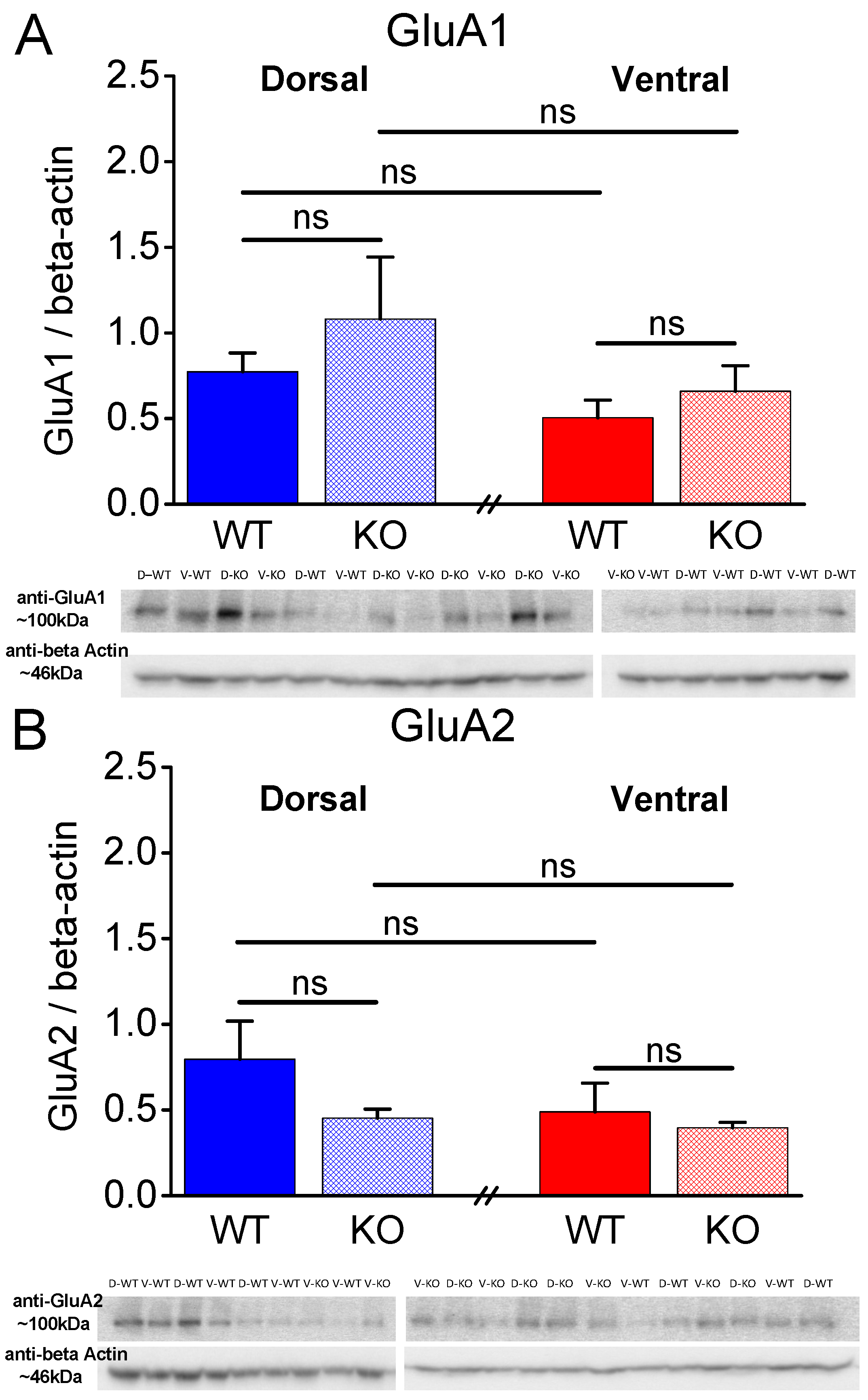
Figure 8.
Expression of AMPA receptor subunits GluA1 and GluA2 in the CA1 hippocampal region of female rats. A. The GluA1 subunit was similarly expressed in WT and KO dorsal (t8 = -0.431, p = 0.678, WT=5 and KO=5) and ventral hippocampus (t6 = -0.812, p = 0.448, WT=4 and KO=4) of WT rats. Also, the expression of GluA1 is similar in DH and VH of WT rats (t7 = 1.923, p = 0.096), and in DH and VH of KO rats (t7 = 1.434, p = 0.195). B. The GluA2 subunit was similarly expressed in WT and KO dorsal (t8 = -0.626, p = 0.549, WT=5 and KO=5) and ventral hippocampus (t6 = -0.003, p = 0.998, WT=4 and KO=4) of WT rats. Also, the expression of GluA2 is similar in DH and VH of WT rats (t7 = 0.977, p = 0.361), and in DH and VH of KO rats (t7 = 1.075, p = 0.318).
Figure 8.
Expression of AMPA receptor subunits GluA1 and GluA2 in the CA1 hippocampal region of female rats. A. The GluA1 subunit was similarly expressed in WT and KO dorsal (t8 = -0.431, p = 0.678, WT=5 and KO=5) and ventral hippocampus (t6 = -0.812, p = 0.448, WT=4 and KO=4) of WT rats. Also, the expression of GluA1 is similar in DH and VH of WT rats (t7 = 1.923, p = 0.096), and in DH and VH of KO rats (t7 = 1.434, p = 0.195). B. The GluA2 subunit was similarly expressed in WT and KO dorsal (t8 = -0.626, p = 0.549, WT=5 and KO=5) and ventral hippocampus (t6 = -0.003, p = 0.998, WT=4 and KO=4) of WT rats. Also, the expression of GluA2 is similar in DH and VH of WT rats (t7 = 0.977, p = 0.361), and in DH and VH of KO rats (t7 = 1.075, p = 0.318).
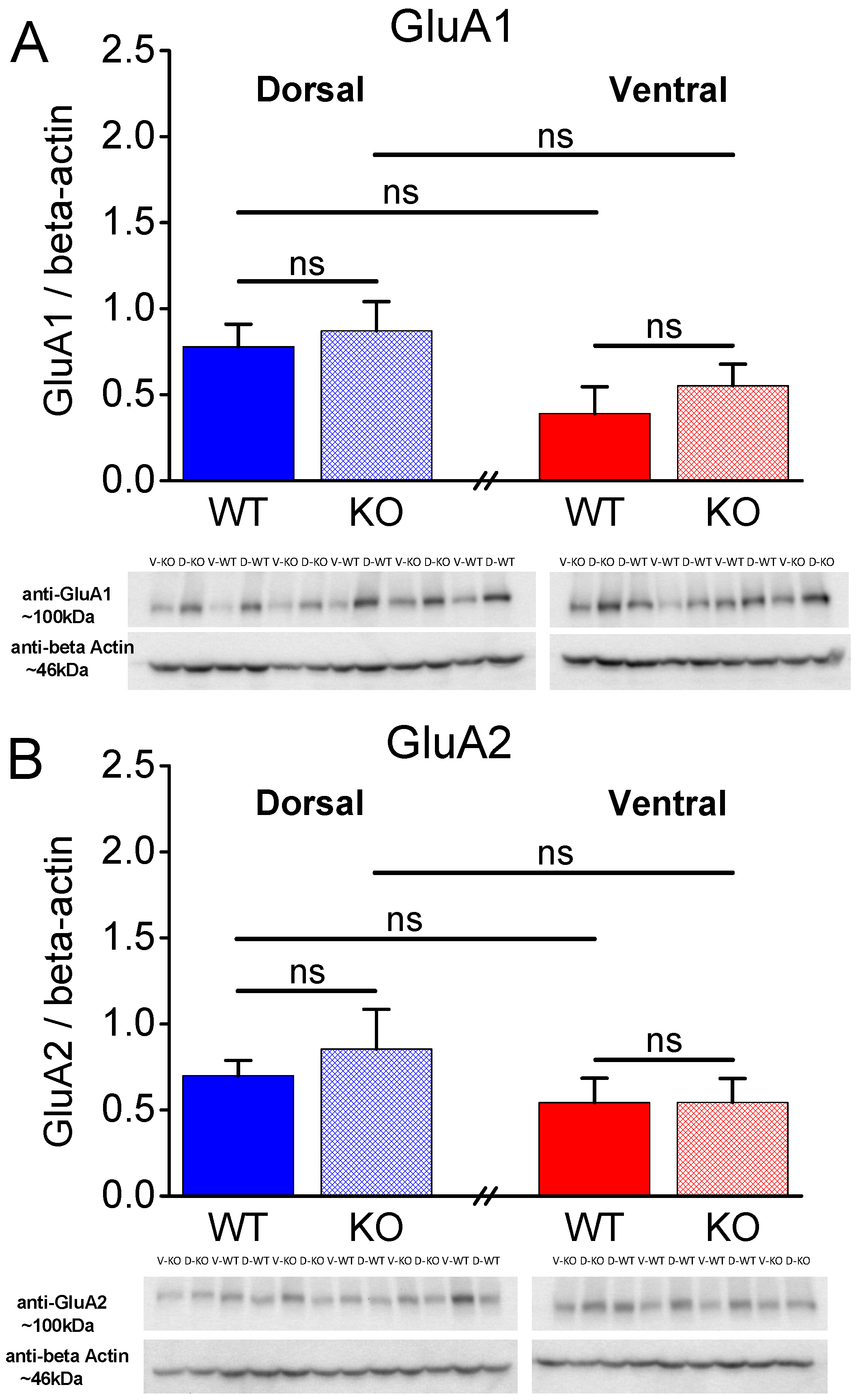
Figure 9.
FXS does not affect excitatory synaptic transmission in either male or female dorsal and ventral hippocampus. Sample traces of synaptic responses evoked with increasing stimulation current intensity (left panels) and input-output functions between stimulation current and fEPSP (middle and right panels) obtained from the dorsal and ventral hippocampus of male (A,B) and female (C,D) wild type (WT) and Fmr1-knockout (KO) rats. Graphs in the middle panels show input-output curves obtained by either dorsal or ventral hippocampus, while the same data are replotted in right panel graphs to compare input-output relationship between dorsal and ventral hippocampus.
Figure 9.
FXS does not affect excitatory synaptic transmission in either male or female dorsal and ventral hippocampus. Sample traces of synaptic responses evoked with increasing stimulation current intensity (left panels) and input-output functions between stimulation current and fEPSP (middle and right panels) obtained from the dorsal and ventral hippocampus of male (A,B) and female (C,D) wild type (WT) and Fmr1-knockout (KO) rats. Graphs in the middle panels show input-output curves obtained by either dorsal or ventral hippocampus, while the same data are replotted in right panel graphs to compare input-output relationship between dorsal and ventral hippocampus.
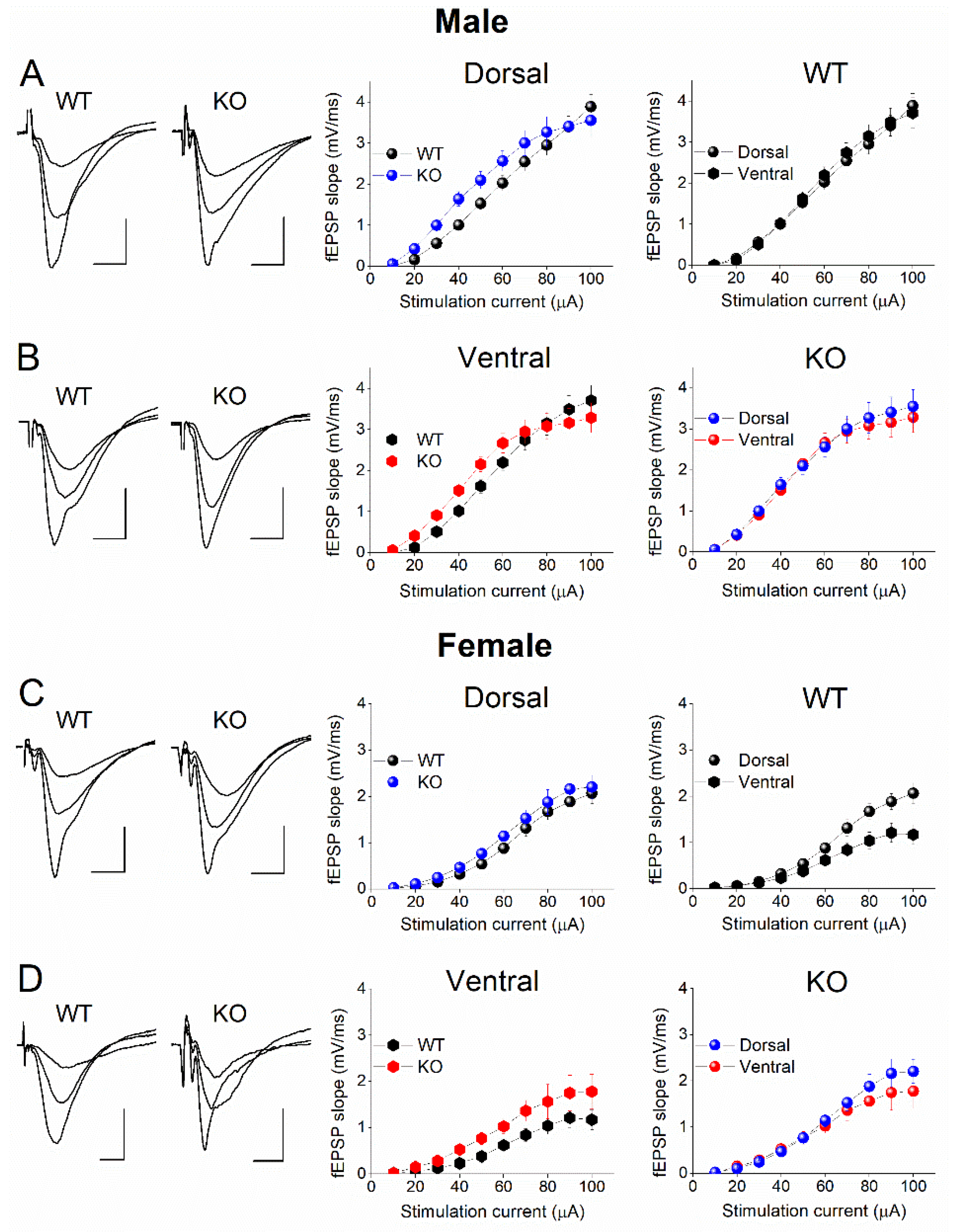
Figure 10.
Excitatory synaptic transmission differs between male and female in both DH and VH, in either WT or KO rats. Data presented here are replotted from Figure 7 to illustrate the gender-associated differences in excitatory synaptic transmission. The results of two-way ANOVA used to compare I-O curves are provided in each graph.
Figure 10.
Excitatory synaptic transmission differs between male and female in both DH and VH, in either WT or KO rats. Data presented here are replotted from Figure 7 to illustrate the gender-associated differences in excitatory synaptic transmission. The results of two-way ANOVA used to compare I-O curves are provided in each graph.
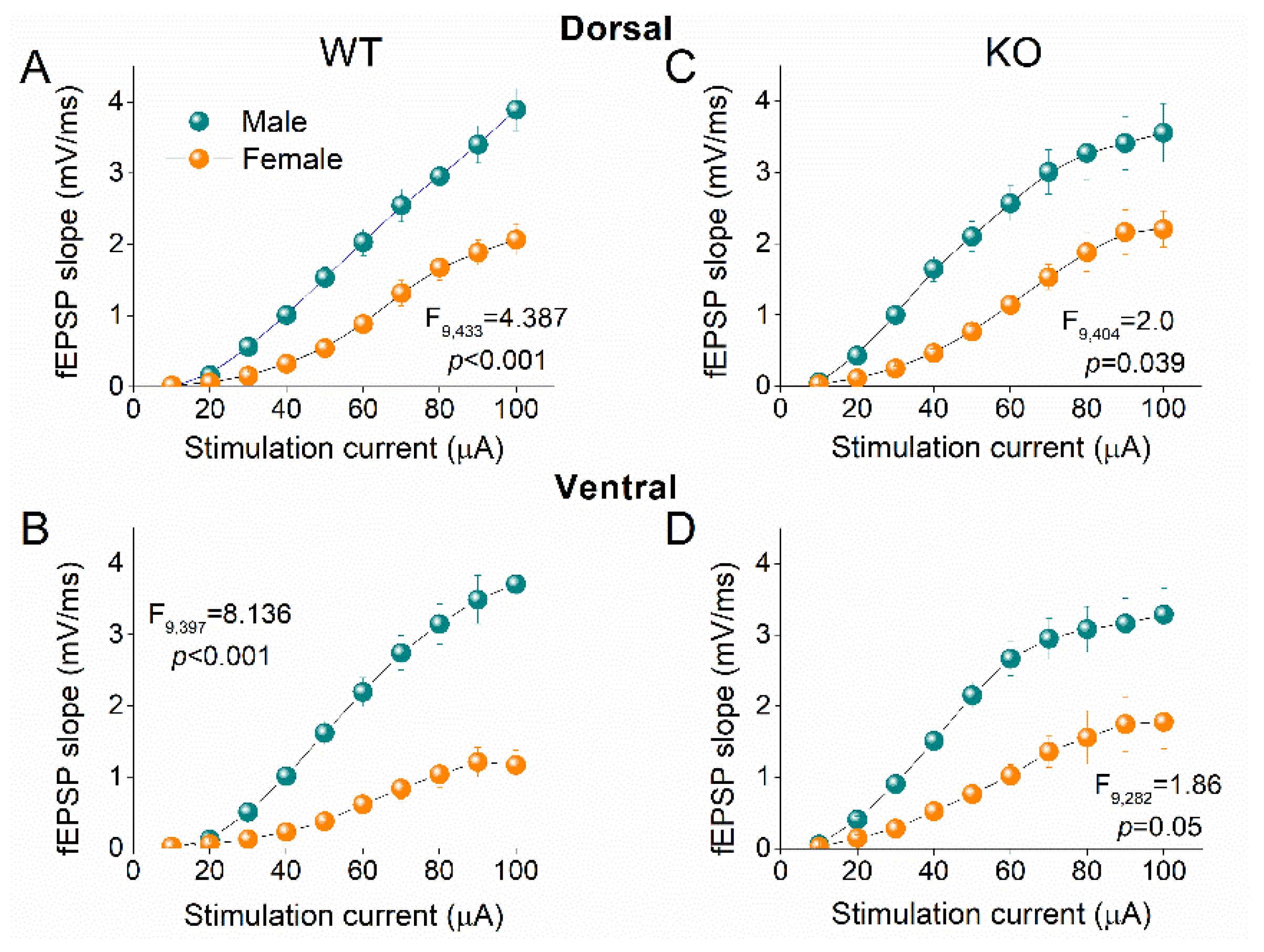
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



