
Submitted:
28 July 2025
Posted:
30 July 2025
You are already at the latest version
Abstract
Interference between similar associations poses a core challenge for episodic memory. While retrieval-based accounts have dominated past theories, it remains unclear whether retroactive interference (RI) and proactive interference (PI) can emerge independently of retrieval competition. To address this, we employed a two-alternative forced-choice (2AFC) associative recognition task that minimized retrieval competition by using same-list distractors. Participants studied overlapping (A–B, A–C) and non-overlapping (E–F, G–H) associations, followed by associative recognition and source memory judgments. Results revealed an asymmetric interference pattern: RI impaired recognition accuracy without affecting response times, suggesting degraded recollection due to representational suppression. In contrast, PI was reflected in longer response times without accuracy loss, consistent with increased retrieval demands stemming from encoding-related differentiation. Source judgments showed no integration-induced cost and even faster RTs for overlapping pairs, further supporting the operation of pattern separation during A–C encoding. These findings highlight the critical role of encoding—rather than retrieval alone—in shaping interference. RI and PI reflect distinct processes: suppression-driven weakening of prior traces versus complexity-induced retrieval delays, respectively. Our results challenge retrieval-centric models and underscore the need for encoding-focused frameworks in memory interference research. Future work should examine how encoding strength, contextual differentiation, and individual variability modulate interference effects across memory tasks.
Keywords:
proactive interference
; retroactive interference
; associative recognition
; source memory
; encoding processes
; response time
1. Introduction
1.1. Associative Interference in Episodic Memory
Episodic memories are characterized by the composed of associative connections between items that are spatially and temporally proximate. The associative memories—where items are linked together based on their co-occurrence in an event—enable individuals to recall not only isolated items but also the associations between them—such as a person’s face and name. However, episodic memories often share overlapping elements across different experiences, allowing for generalization (Moscovitch et al., 2016; Renoult et al., 2015)but can also lead to interference (Kim et al., 2017) when shared elements serve as retrieval cues.
This is particularly relevant in AB/AC paradigm, also known as the paired associate learning paradigm (Martin, 1971; Postman & Gray, 1977) which has often been used to investigate associative interference. In the AB/AC paradigm, participants study two lists of cue-target pairs (e.g., picture-word pairs), where cues (A) are paired with different targets in List 1 (A-B) and List 2 (A-C). Retroactive interference (RI) is typically measured by comparing memory for List 1 (A-B pairs) between an experimental group (who also study List 2, A-C pairs) and a control group (who do not study List 2). Similarly, proactive interference (PI) is assessed by comparing memory for List 2 (A-C pairs) between participants who have or have not studied List 1 (A-B pairs). These “pure” control conditions, in which one list is entirely omitted, serve to isolate the effects of interference attributable to the presence of competing associations (Martin, 1971; Postman & Underwood, 1973). However, such control conditions lack ecological validity—real-world memory rarely involves complete absence of prior or subsequent learning. More importantly, these traditional control conditions do not distinguish between interference driven by cue-overlap-based similarity and interference arising from general cognitive load or list-based competition.In the present study, we adopt a similarity-based contrast design in which the control condition consists of non-overlapping associative pairs (e.g., E-F and G-H) drawn from the same lists as the overlapping pairs (A-B and A-C). That is, all participants study both List 1 and List 2, but only some pairs involve shared cues (i.e., A-B/A-C) while others do not (i.e., E-F, G-H). This within-subject, mixed-list paradigm enables a more precise comparison between conditions with and without cue overlap, isolating interference effects specifically attributable to associative similarity, rather than to general learning or list position.This design approach has been increasingly adopted in recent studies examining memory interference and integration, where cue similarity and representational overlap are treated as critical experimental factors (Chanales et al., 2019; Kuhl et al., 2010; Ritvo et al., 2024a)
1.2. Two-Factor Interference Theory: Retrieval Competition and Associative Unlearning
Classic theories of interference attribute memory disruption primarily to retrieval-phase processes. The retrieval competition account (McGeoch, 1932; Watkins & Watkins, 1975) posits that cue overload diminishes retrieval specificity, leading to competition among associated memories
The Search of Associative Memory (SAM) model (Mensink, 1988; Mensink & Raaijmakers, 1989; Raaijmakers & Shiffrin, 1981) formalizes this process, representing memory as a network of interconnected nodes, where a cue activates multiple nodes, and their relative activation strengths determine retrieval success. In the SAM model, retrieval is governed by a probabilistic process where the probability of retrieving a specific memory trace is proportional to its activation strength divided by the sum of the activation strengths of all traces associated with the cue.This competitive process means that stronger traces dominate retrieval, reducing the likelihood of retrieving weaker traces and causing interference (Favila et al., 2016; Gabitov et al., 2019; Sajikumar et al., 2014).
Associative Unlearning posits that retroactive interference occurs during encoding, when new learning (A-C) weakens prior associations (A-B) independent of retrieval competition (Barnes & Underwood, 1959). especially when cue-outcome contingencies are violated (Mujezinović et al., 2024). This view, however, has primarily been supported in overlearning contexts, leaving its applicability to episodic memory less clear.
1.3. Encoding-Based Accounts: Integration, Differentiation, and Similarity-Based Interference
Recent research suggests that encoding overlapping associations (A-C) can reactivate and modify prior memories (A-B), leading to integration or differentiation (Chanales et al., 2019; Kuhl et al., 2010, 2012; Ritvo et al., 2024a). Similarity-based interference during encoding arises when new information (A-C) shares overlapping cues with prior content (A-B), altering the encoding process compared to non-overlapping pairs (E-F, G-H). This alteration, driven by hippocampal pattern completion (Ritvo et al., 2024b; Ritvo et al., 2019; Zeithamova & Preston, 2010, 2017), depends on the reactivation strength of A-B during A-C encoding.
Strong reactivation strengthens shared features, forming cohesive memory representations (Herszage & Censor, 2017; Koen & Rugg, 2016) lead to memory integration. This may enhance A-B retention (retroactive facilitation) but increase PI risks by blurring distinctions between A-B and A-C during retrieval (Chanales et al., 2019; Huijbers et al., 2009; Richter et al., 2016); Moderate reactivation triggers hippocampal pattern separation, weakening overlapping features to reduce interference (Norman et al., 2007; Ritvo et al., 2019) which will result in differentiation. This enhances memory distinctiveness, potentially mitigating RI and PI, but may cause distortions if features are overly suppressed (Chanales et al., 2021); No Interaction: Minimal reactivation results in no modification of either memory trace, avoiding interference but also integration or differentiation benefits.
Similarity-based interference is amplified by cue overlap, making encoding-based effects more pronounced for A-B/A-C pairs than for E-F/G-H pairs. Neural mechanisms, such as hippocampal differentiation or medial prefrontal cortex (mPFC) integration (Schlichting & Preston, 2015), further modulate these outcomes, suggesting that RI and PI may originate from memory reorganization during encoding, not solely retrieval or unlearning.
1.4. The Present Work
While prior research has focused predominantly on retrieval-based interference, it remains unclear whether RI and PI can emerge independently of retrieval competition. The present study addresses this gap by using a non-competitive, two-alternative forced-choice (2AFC) associative recognition task in which targets are selected among same-list distractors, minimizing direct competition between overlapping associations.Participants also make source memory judgments to identify whether recognized associations originated from List 1 or List 2. This dual-task design allows for dissociation of retrieval- and encoding-related mechanisms by testing the following hypotheses:
Presence of Interference: If RI exists, A-B recognition (List 1) will be lower than E-F due to subsequent A-C learning. If PI exists, A-C recognition (List 2) will be lower than G-H due to prior A-B learning. The presence of only RI, only PI, both, or neither will inform the relative contributions of encoding vs. retrieval mechanisms.
Asymmetric Interference: If encoding-based processes such as integration or differentiation dominate, RI and PI effects may differ in magnitude or direction, depending on reactivation strength during A-C encoding.
Source Memory as a Diagnostic: Integration should impair source discrimination (e.g., misattributing A-B to List 2), while differentiation should enhance it. Source memory accuracy and response times (RTs) will serve as behavioral markers of these encoding processes.
In addition to accuracy measures, we examine response times (RTs) during associative recognition and source memory judgments as process-sensitive indices of memory interference. Prior studies have shown that even when retrieval is successful, longer RTs can reveal latent competition or increased cognitive demands during memory access (Anderson & Reder, 1999; Norman et al., 2007). In the context of associative memory, RTs provide insight into the accessibility and resolution difficulty of overlapping traces, which may not be apparent from accuracy alone. For example, Wimber et al. (2009) demonstrated that retrieval-induced forgetting is accompanied by slower RTs when competing items are partially activated (Wimber et al., 2009), and Horner et al. (2015) found that integrated memories result in slower source discrimination due to increased representational overlap (Horner et al., 2015).
Thus, if cue overlap triggers interference during encoding or retention, we expect slower RTs for A-B and A-C recognition (compared to E-F and G-H), even in the absence of significant accuracy differences. Similarly, elevated RTs during source judgments would reflect increased contextual ambiguity, especially under integration. By jointly analyzing accuracy and latency, the present design provides a comprehensive behavioral profile of associative interference mechanisms.
By controlling for cue-target familiarity, emotional valence, and output order effects, this study isolates the role of encoding and retention dynamics in associative interference. It thus provides a process-level understanding of how cue overlap and memory reactivation shape the structure and accessibility of episodic memories beyond retrieval competition.
2. Method
Experiments were approved by the University’s Institutional Review Board. All participants read and signed an informed consent form and were compensated appropriately upon completion. The experiment was approved by the Ethics Committee of the host institution.
2.1. Design
We used a 2 (List: List 1 vs. List 2) × 2 (Pair type: Cue overlap vs. Non-overlap) repeated measure design. Participants completed two lists memory tasks: an initial list followed by second list. Both lists contained both cue overlap and non-overlap associations to measure interference effects. The dependent variables were accuracy and reaction time for associative recognition, and accuracy and reaction time for list source judgments, specifically for associative recognition pairs that were successfully identified.
2.2. Participants
Based on prior research and an a priori power analysis (α = .05, power = .80, two-tailed), a sample size of 78 participants was determined to be sufficient to detect a medium effect size (f = 0.25) (Cohen, 2013). Seventy-eight undergraduate students (25 men, 53 women; M = 21.94 years, SD = 2.40) were recruited for the study. All participants were right-handed and reported normal or corrected-to-normal vision. Each participant was tested individually in a quiet, controlled laboratory environment.
2.3. Materials
The experimental materials included both verbs and images, selected and arranged to create conditions of cue overlap and non-overlap in list 1 and list 2. A total of 100 Chinese verbs were selected from the Chinese Affective Words System (CAWS) (Wang et al., 2008). Of these, 4 were reserved for practice trials. The remaining 96 verbs were divided into four sets of 24, matched on emotional valence and familiarity. Mean valence ratings (M ± SD) were: Group 1 (4.92 ± 1.59), Group 2 (4.90 ± 1.50), Group 3 (4.62 ± 1.54), and Group 4 (5.11 ± 1.50). Familiarity ratings were comparable: Group 1 (5.09 ± 1.95), Group 2 (5.07 ± 1.96), Group 3 (5.10 ± 1.93), and Group 4 (5.06 ± 2.07). 76 images were selected from the Bank of Standardized Stimuli (BOSS) (Brodeur et al., 2010), with four images reserved for practice trials. The remaining 72 experimental images were evenly divided into two categories: biotic (e.g., mammals, birds, insects; n = 36) and abiotic (e.g., furniture, clothing, tools, buildings; n = 36). All images were highly familiar to participants. The 36 biotic and 36 abiotic images were further randomized into three groups, each containing 12 biotic and 12 abiotic images. The groups were matched on both familiarity (M ± SD: Group 1 = 4.43 ± 0.78, Group 2 = 4.50 ± 0.80, Group 3 = 4.51 ± 0.75) and visual complexity (Group 1 = 2.62 ± 1.27, Group 2 = 2.56 ± 1.24, Group 3 = 2.61 ± 1.25), with no significant differences across groups (all ps > .05).
2.4. Experimental Pairings Were as Follows:
One set of 24 images was used in both List 1 and List 2. In List 1, each image was paired with a unique verb (A–B); in List 2, the same image was paired with a different, unrelated verb (A–C). These formed the cue-overlap pairs (A-B,A-C), where the image served as the repeated retrieval cue, and B and C were pre-assigned with low semantic relatedness.To verify low semantic overlap between co-paired verbs (B and C), 24 participants rated the association strength of the AB–AC word pairs on a 7-point Likert scale (1 = not at all associated, 7 = very strongly associated). The average rating was low (M = 2.43, SD = 0.48), confirming minimal semantic association.
The remaining 48 images (two sets) were used only once: one set appeared in List 1 and the other in List 2, paired with different sets of verbs (EF or GH). These formed the non-overlap pairs, with no shared cues across lists
2.5. Procedure.
The experiment was programmed in E-Prime 3.0 (Psychology Software Tools) and presented on a 16-inch laptop with 1920 × 1080 resolution (60 Hz refresh rate). Participants responded via keyboard while maintaining a viewing distance of approximately 40 cm from the screen center. Images (500 × 500 pixels) were presented in the upper center of the screen, with words displayed directly below each image.Participants first completed a 4-trial practice block using stimuli not included in the main experiment. The formal experiment consisted of five sequential phases:
Encoding Phase (List 1): Participants studied list 1word–image pairs. Half of these were cue-overlap pairs (AB type)—images that would later be repeated in List 2 with a different verb. The other half were non-overlap pairs (EF type)—images and words that appeared only in List 1. Each trial began with a 500 ms fixation cross, followed by a 3,000 ms presentation of a word–image pair. Pairs were presented in a randomized order;
Distractor Task 1: Participants completed a 60-second Chinese Cancellation Task to prevent rehearsal of List 1 material;
Encoding Phase (List 2): Participants studied another list word–image pairs. For half of the trials, the images from the cue-overlap set in List 1 were repeated and paired with new, unrelated words (AC pairs). The remaining trials featured new word–image pairs (GH type) that did not appear in List 1, forming the non-overlap condition for List 2.
Distractor Task 2: A 3-minute distractor task was used before testing.
Test Phase: Participants completed two tasks. In the Associative Recognition Task, participants were presented with an image and two previously studied words (e.g., Investigate vs. march) and were asked to select the word that had originally been paired with the image. Both the target and the foil were drawn from the same list (either List 1 or List 2), ensuring that associative recognition for AB pairs (List 1) and for AC pairs (List 2) was tested independently.After selecting the correct verb, participants indicated whether the chosen word–image pair had been studied in List 1 or List 2. The order of test pair from List 1 and List 2 was randomized across trials. A schematic illustration of the entire procedure is shown in Figure 1.
3. Results
Data analysis was conducted using R version 4.5.0 (R Core Team, 2025). A two-way repeated-measures analysis of variance (ANOVA) was performed using the afex package (Singmann et al., 2025) to examine the effects of pair type (cue-overlap vs. non-overlap) and list (List 1 vs. List 2) on associative recognition accuracy, with a significance level set at α = .05. Trials with reaction times (RTs) exceeding 5,000 ms (2.36% of total trials) were excluded because the experimental design automatically advanced to the next trial after 5,000 ms, indicating no participant response. Additionally, trials with recognition RTs < 200 ms or > 3SD, and source judgment RTs < 100 ms or > 3SD, were removed to ensure data quality.The data and analysis scripts are available at https://osf.io/pfgca/
3.1. Associative Recognition Results
Descriptive statistics for associative recognition accuracy are presented in Table 1. A 2 (List: List 1 vs. List 2) × 2 (Pair Type: Cue-Overlap vs. Non-Overlap) repeated-measures ANOVA revealed a significant main effect of list, F(1, 77) = 13.31, p < .001, partial η2 = .15, with higher accuracy observed in List 1 (M = 0.78, SD = 0.14) than in List 2 (M = 0.75, SD = 0.14). A significant main effect of pair type was also found, F(1, 77) = 5.01, p = .028, partial η2 = .06, indicating higher accuracy for non-overlap pairs (M = 0.78, SD = 0.15) than for cue-overlap pairs(M = 0.75, SD = 0.13).The interaction between list and pair type was also significant, F(1, 77) = 9.28, p = .003, partial η2 = .11 (see Figure 2).
To explore this interaction, simple effects analyses were conducted using the emmeans package (Lenth et al., 2025). For List 1, accuracy was significantly higher for non-overlap pairs (M = 0.81, SD = 0.13) compared to cue-overlap pairs (M = 0.75, SD = 0.15), t(77) = -4.45, p < .001, Cohen’s d = -0.50. In contrast, for List 2, there was no significant difference between cue-overlap (M = 0.75, SD = 0.11) and non-overlap pairs (M = 0.74, SD = 0.17), t(77) = 0.68, p = .50, Cohen’s d = 0.08. All p-values are two-tailed and Bonferroni-adjusted where applicable.
To complement the accuracy analysis, recognition reaction times (RTs) were analyzed to examine potential interference effects that may not be evident from accuracy alone. RTs were subjected to a 2 (List) × 2 (Pair Type) × 2 (Recognition Accuracy: Correct, Incorrect) repeated-measures ANOVA using the afex package (Singmann et al., 2025). Due to incomplete data, 7 participants were excluded, resulting in a final sample of 71 participants. Descriptive statistics are presented in Table 1, and patterns are illustrated in Figure 3.
The analysis revealed a significant main effect of list, F(1, 70) = 6.99, p = .010, partial η2 = .09, with shorter RTs in List 1 (M = 2,491, SD = 353.7) than in List 2 (M = 2,574, SD = 346.9). There was also a significant main effect of pair type, F(1, 70) = 15.12, p < .001, partial η2 = .18, with longer RTs for cue-overlap pairs (M = 2,592, SD = 357.6) than non-overlap pairs (M = 2,473, SD = 341.5). A highly significant main effect of recognition accuracy was found, F(1, 70) = 82.41, p < .001, partial η2 = .54, reflecting faster responses for correct (M = 2,367, SD = 273.3) versus incorrect recognition (M = 2,698, SD = 423.8).
A significant interaction emerged between list and pair type, F(1, 70) = 10.72, p = .002, partial η2 = .13. No significant interactions were observed between recognition accuracy and the other factors, all ps > .05. Follow-up comparisons indicated that for List 1, the RT difference between cue-overlap (M = 2,513, SD = 383.7) and non-overlap pairs (M = 2,469, SD = 368.2) was not significant, t(70) = 1.21, p = .231, Cohen’s d = 0.12. For List 2, RTs were significantly longer for cue-overlap (M = 2,671, SD = 380.4) than for non-overlap pairs (M = 2,477, SD = 369.7), t(70) = 4.86, p < .001, Cohen’s d = 0.51.
3.2. Source Judgement Results
To examine source memory, we analyzed the proportion of correct source judgments for successfully recognized associations. A 2 (Pair Type: Cue-Overlap vs. Non-Overlap) × 2 (List: List 1 vs. List 2) repeated-measures ANOVA revealed a significant main effect of List, F(1, 77) = 4.79, p = .032, partial η2 = .06, with higher source accuracy in List 1 (M = 0.74, SD = 0.14) than in List 2 (M = 0.70, SD = 0.15). The main effect of pair type was not significant, F(1, 77) = 0.77, p = .383, partial η2 = .01, nor was the interaction, F(1, 77) = 0.07, p = .790, partial η2 < .001. These findings suggest that cue overlap did not significantly affect source memory, although source accuracy was generally better for List 1. Descriptive statistics are shown in Table 2 and Figure 4.
A 2 (List) × 2 (Pair Type) × 2 (Source Accuracy: Correct, Incorrect) repeated-measures ANOVA was conducted to examine mean response times during the source judgment task. Six participants were excluded due to missing data, yielding a final sample of 72 participants. Descriptive statistics are reported in Table 2.
There was a significant main effect of source accuracy, F(1, 71) = 39.17, p < .001, partial η2 = .36, indicating faster RTs for correct judgments (M = 616.5, SD = 374.8) than incorrect judgments (M = 700.0, SD = 425.8). The main effect of pair type was marginal, F(1, 71) = 3.36, p = .071, partial η2 = .05, suggesting slightly faster RTs for cue-overlap (M = 645.5, SD = 392.8) compared to non-overlap pairs (M = 671.0, SD = 407.8). The main effect of list was not significant, F(1, 71) = 0.02, p = .880, partial η2 < .01.
Figure 5.
Boxplots of source judgement response time by pair type and learning list, separated by judgement accuracy. Boxes show IQR (25th–75th percentiles), solid lines represent medians, whiskers extend to 1.5×IQR, and white circles indicate condition means. Significant differences between cue-overlap and non-overlap pairs are marked with asterisks (*p <0.05, **p <0.01, ***p < .001); n.s. = not significant (p ≥ .05). Post hoc tests adjusted with Bonferroni correction.
Figure 5.
Boxplots of source judgement response time by pair type and learning list, separated by judgement accuracy. Boxes show IQR (25th–75th percentiles), solid lines represent medians, whiskers extend to 1.5×IQR, and white circles indicate condition means. Significant differences between cue-overlap and non-overlap pairs are marked with asterisks (*p <0.05, **p <0.01, ***p < .001); n.s. = not significant (p ≥ .05). Post hoc tests adjusted with Bonferroni correction.
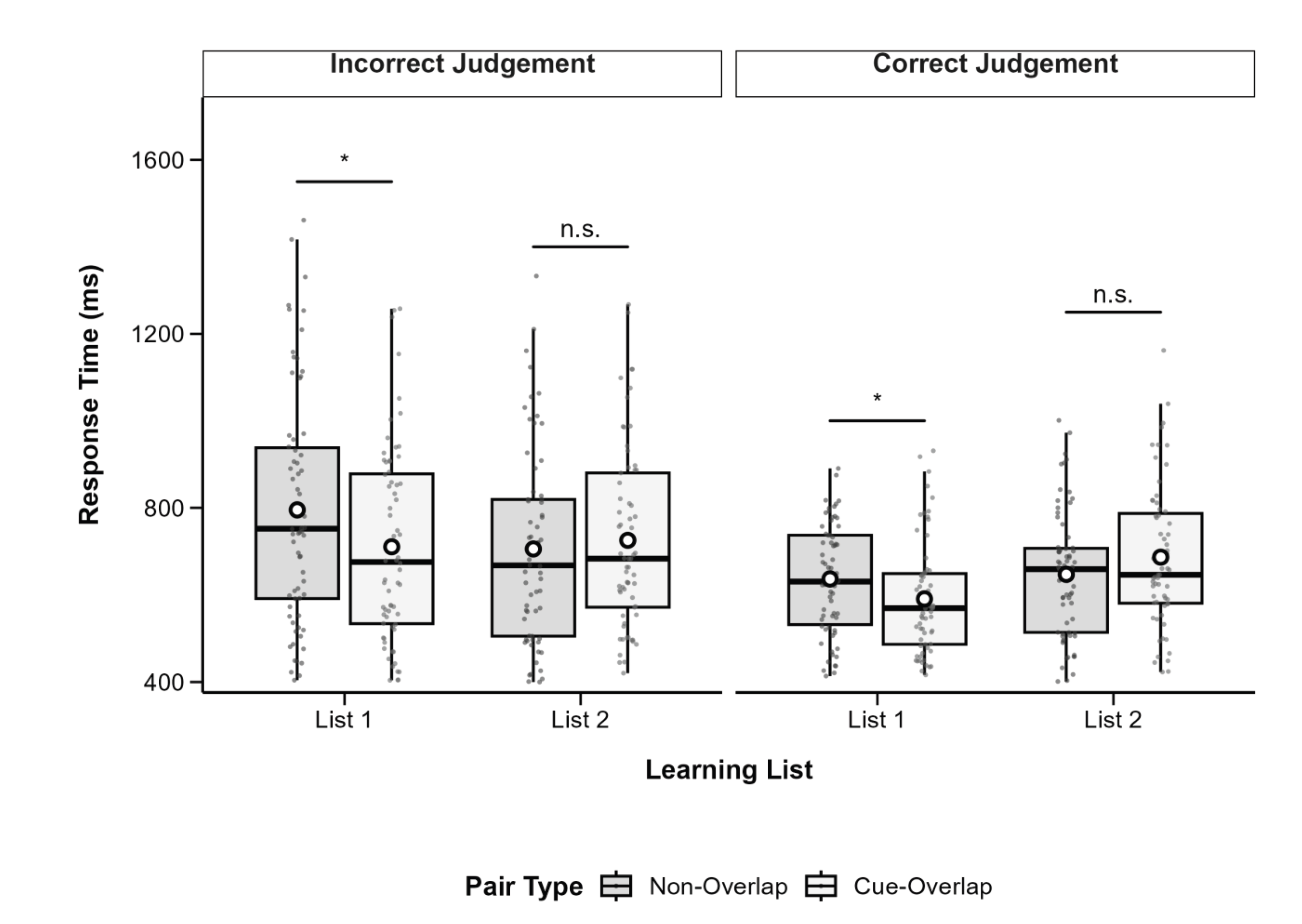
Significant two-way interactions were found between list and pair type, F(1, 71) = 8.96, p = .004, partial η2 = .11, and between list and source accuracy, F(1, 71) = 8.09, p = .006, partial η2 = .10. A significant three-way interaction was also observed among list, pair type, and source accuracy, F(1, 71) = 4.85, p = .031, partial η2 = .06. To follow up on the three-way interaction, we first examined the interaction between list and pair type separately for correct and incorrect source judgments. When source judgments were incorrect, the interaction between list and pair type was significant, F(1, 71) = 8.68, p = .005. Simple effects analyses revealed that in List 1, RTs were significantly slower for non-overlap pairs (M = 764, SD = 443.0) than for cue-overlap pairs (M = 674, SD = 406.0), t(71) = -2.46, p = .017, Cohen’s d = -0.29. In contrast, in List 2, the difference between cue-overlap (M = 710, SD = 427.0) and non-overlap pairs (M = 652, SD = 427.0) was not significant, t(71) = 1.65, p = .104. For correct source judgments, the interaction between list and pair type was not significant, F(1, 71) = 1.02, p = .316. However, a significant simple effect of pair type was observed in List 1, with cue-overlap pairs eliciting faster responses (M = 572, SD = 344) than non-overlap pairs (M = 619, SD = 381), t(71) = -2.55, p = .013, Cohen’s d = -0.30. In List 2, the difference was non-significant, t(71) = -1.30, p = .198, with mean RTs of 626 (SD = 394.0) for cue-overlap and 649 (SD = 380) for non-overlap pairs.
These results indicate that cue-overlap facilitated faster source decisions relative to non-overlap pairs, particularly in List 1 and more so when the source judgment was incorrect. The pattern suggests that encoding differences between overlapping and non-overlapping associations may differentially affect the fluency of source retrieval depending on list context and judgment accuracy.
4. Discussion
4.1. Memory Interference Persists Beyond Retrieval Competition
The present study investigated whether retroactive interference (RI) and proactive interference (PI) can arise in associative memory even in the absence of retrieval competition. Using a non-competitive two-alternative forced-choice (2AFC) recognition paradigm, in which targets were selected among same-list distractors, we minimized cue-based retrieval conflict between overlapping associations. Despite this control, both RI and PI were observed, demonstrating that interference can emerge independently of retrieval-based competition, which challenge models that localize associative interference solely to retrieval dynamics (e.g., retrieval competition or response selection conflict) and instead align with accounts emphasizing encoding-phase processes, such as trace integration (Shohamy & Wagner, 2008) or interference during consolidation (Dewar et al., 2007).
Specifically, recognition accuracy revealed significant RI (A-B < E-F), while response time data revealed PI effects (A-C > G-H). These findings indicate that memory interference is not limited to retrieval-phase competition, as traditionally emphasized (Anderson et al., 1994; Levy & Anderson, 2002), but can also originate from mechanisms operating during encoding or retention. Importantly, the observed interference was cue-specific, appearing only when associations shared overlapping cues, thereby underscoring the role of cue-based representational similarity in shaping interference even under conditions of minimal output interference.
This result aligns with recent theoretical frameworks suggesting that memory interference is highly dependent on representational overlap and organizational dynamics during encoding (Kuhl et al., 2011; Norman et al., 2007). The findings suggest that interference effects can arise from structural similarity across learning episodes, irrespective of retrieval mode, and manifest in distinct ways depending on when and how such overlap affects memory processing.
Dissociable Mechanisms Underlying Retroactive and Proactive Interference
Although both RI and PI were observed, their behavioral signatures diverged, pointing toward distinct underlying mechanisms. RI was manifested as a decline in recognition accuracy for A-B associations, whereas PI was not detectable in accuracy but revealed through increased response times for A-C recognition. This asymmetry suggests that RI and PI operate via functionally distinct pathways, consistent with prior research proposing differential encoding versus retention vulnerabilities (Polack et al., 2017; Postman & Underwood, 1973).
With respect to RI, the observed reduction in recognition accuracy for A-B associations compared to E-F pairs indicates that subsequent learning of A-C associations interfered with memory for the original A-B pairs. However, source memory results complicate the interpretation that this interference is driven by integrative encoding. If strong integration between A-B and A-C had occurred during the encoding of A-C, we would expect impaired source discrimination for overlapping (A-B and A-C) associations relative to non-overlapping controls. Contrary to this prediction, source accuracy in the cue-overlap condition was not significantly worse than in the non-overlap condition, and in fact, source judgments for overlapping pairs were often faster, particularly when correct. This pattern is inconsistent with the integration account, which posits that reactivated A–B memories are merged with newly encoded A–C representations, leading to increased source confusion and delayed decisions (Horner et al., 2015). Instead, the data favor a differentiation-based encoding mechanism, wherein partial or weak reactivation of A–B during A–C learning prompts representational separation rather than fusion (Hulbert & Norman, 2015; Kim et al., 2017). This process reduces later retrieval competition but may come at the cost of damaging the relational components of the original A–B memory, especially those required for recollection-based recognition.
According to dual-process theories of recognition memory (Wixted & Mickes, 2010; Yonelinas, 2002), associative recognition tasks rely more heavily on recollection than item recognition, particularly when distinguishing between highly familiar associations. Consequently, when participants attempt to recognize previously studied A–B pairs, failure to retrieve specific relational details can lead them to rely predominantly on familiarity-based processes. Although familiarity is generally preserved, it is less diagnostic for associative judgments. This reliance on familiarity rather than recollection likely accounts for why recognition accuracy for A–B pairs declined compared to novel E–F pairs (which maintained both recollection and familiarity). It also explains the absence of differences in reaction times, as familiarity-driven responses tend to be consistently rapid regardless of their accuracy (Ratcliff & McKoon, 2008).
Differentiation mechanisms such as pattern suppression or representational repulsion have been proposed as neural strategies for minimizing interference between overlapping memories (Norman et al., 2007; Kim et al., 2017). These mechanisms act by suppressing or altering previously encoded traces when similar information is encountered again—effectively creating more orthogonal memory representations to prevent future confusion. Although this strategy benefits the encoding of new associations (e.g., A–C), it may concurrently disrupt the relational coherence of prior associations (e.g., A–B), rendering them more difficult to retrieve via recollection.
Such findings are consistent with the view that pattern separation processes in the hippocampus (Favila et al., 2020; Hulbert & Norman, 2015) can support memory specificity by reducing shared representational features. However, by creating distinct and non-overlapping traces, this process may simultaneously weaken original, relational linkages that support recollection in associative recognition tasks (Yonelinas, 2002).
In summary, differentiation-based encoding offers a powerful mechanism to protect new memories from interference, but it does so by compromising the retrievability of prior relational structures, particularly those dependent on recollection. The observed accuracy deficit without RT slowing for A–B recognition reflects this dissociation—a byproduct of strategic reorganization during encoding that prioritizes future distinctiveness over backward compatibility with earlier associations.
While proactive interference did not impair recognition accuracy, the prolonged response times for A–C pairs—relative to non-overlapping controls—suggest increased cognitive demands during retrieval. From an encoding perspective, this effect may be attributed to differentiation-based processes that were engaged to distinguish newly encoded A–C associations from prior A–B pairs sharing the same cue. During A–C encoding, participants likely engaged in comparative and separation-based processing, which not only increased representational distinctiveness but also introduced greater complexity into the resulting memory trace (Hulbert & Norman, 2015). Consequently, although recollection remained intact, retrieval required more time-consuming evidence accumulation, reflecting greater internal decisional uncertainty or representational reconfiguration.
Supporting this interpretation, pattern separation mechanisms in the hippocampus have been shown to increase source discriminability while simultaneously delaying successful pattern completion due to reduced associative overlap (Favila et al., 2020; Kim et al., 2017). According to diffusion models of decision-making, such changes in representational structure can slow the rate of information accumulation, thereby increasing recognition latency even when final accuracy remains unaffected (Ratcliff et al., 2016). Thus, the observed PI effect in RTs reflects the cognitive cost of encoding-based interference resolution, where the mnemonic benefit of reduced interference is offset by increased retrieval effort.
4.2. The Central Role of Encoding in Interference Mechanisms
Although traditional memory models have prioritized retrieval as the primary locus of interference (e.g., SAM: Raaijmakers & Shiffrin, 1981; REM: Shiffrin & Steyvers, 1997), our findings reinforce the emerging view that encoding is not merely a precursor to retrieval, but a dynamic, constructive process that actively shapes the susceptibility of memories to interference. In our study, both retroactive and proactive interference effects emerged under conditions where retrieval competition was minimized, suggesting that the organization and differentiation of overlapping associations during encoding play a pivotal role in determining later accessibility and response efficiency.
Recent neuroscientific and computational theories support this interpretation. For example, the Complementary Learning Systems (CLS) framework (Norman & O’Reilly, 2003) posits that the hippocampus dynamically engages pattern separation to reduce representational overlap between similar episodes, especially during encoding. In this view, the same hippocampal system can later engage in pattern completion during retrieval to reconstruct full memory representations from partial cues—highlighting that the encoding-retrieval processes are two sides of the same computational coin. Importantly, the extent and direction of these operations (separation vs. completion) depend on contextual similarity, retrieval goals, and prior encoding dynamics (Ritvo et al., 2019; Yassa & Stark, 2011).
In our paradigm, source memory RT benefits for cue-overlap pairs, despite increased associative overlap, suggest that encoding-related differentiation mechanisms successfully reduced cross-list interference. This aligns with findings from studies showing that differentiation during encoding leads to increased neural separation and reduced retrieval interference (Hulbert & Norman, 2015; Favila et al., 2020). Moreover, theories like stimulus-encoding theory (Martin, 1968) and encoding variability models (Bower, 1972; Tzeng, 1973) emphasize that the same nominal stimulus (e.g., a repeated cue word) can produce qualitatively different internal traces when encoded under distinct contextual or associative constraints.
However, despite mounting behavioral and neural evidence for the encoding basis of interference, formal models of memory have disproportionately focused on retrieval dynamics, often treating encoding as a fixed or uniform process. While models like SAM and REM simulate forgetting through probabilistic cue competition or noise in retrieval strength, they lack a rich account of how encoding strategies like item integration, contextual reinstatement, or differentiation modulate memory formation and later performance (Cowan, 2019; Ranganath, 2010). This imbalance limits their ability to account for findings such as ours, where memory strength (accuracy) and accessibility (RT) dissociate as a function of encoding manipulations.
Together, these results advocate for a reconceptualization of memory interference as an emergent property of encoding-retrieval interactions, not retrieval alone. Encoding is not a passive imprinting of associations but a strategic, sometimes competitive process that actively sculpts the structure of memory representations, with downstream consequences for how these memories are accessed, discriminated, or forgotten. Future models should incorporate computational mechanisms for representational modulation during encoding, including those sensitive to similarity, context, and retrieval history.
4.3. Limitations and Future Directions
While the present study offers important insights into the asymmetric mechanisms of retroactive and proactive interference and the central role of encoding processes, several limitations warrant consideration. First, the use of weakly associated word–picture pairs and brief encoding durations may have constrained the engagement of recollection-based retrieval, potentially amplifying familiarity-driven recognition effects. Given that associative recognition often relies on recollection, future studies should systematically manipulate associative strength and encoding duration to disentangle the relative contributions of familiarity and recollection under varying memory demands (Yonelinas, 2002; Wixted, 2007).
Second, although the two-alternative forced-choice (2AFC) paradigm employed here effectively minimized retrieval competition, it may not fully capture the complexity of naturalistic memory interference, where multiple associative traces may simultaneously compete during recall. Incorporating complementary paradigms—such as free recall, cued recall, or retrieval-induced forgetting tasks—could provide a richer characterization of how encoding- and retrieval-based mechanisms jointly shape interference effects (Anderson, 2003; Hulbert & Norman, 2015).
Third, while we highlight the pivotal role of encoding dynamics (e.g., pattern separation and differentiation) in shaping interference, this study did not directly measure neural correlates of encoding processes, such as hippocampal activity patterns or neural similarity metrics. Future research could integrate neuroimaging or computational modeling approaches (e.g., CLS models, Norman & O’Reilly, 2003) to test how encoding operations like pattern suppression or representational repulsion (Favila et al., 2020) mediate the trade-off between reduced interference and potential costs to recollection.
Finally, individual differences in encoding strategies, cognitive control, and working memory capacity may systematically modulate the degree of interference observed. Our findings suggest that adaptive encoding processes (e.g., differentiation) can either mitigate or exacerbate interference depending on individual strategy use. Future studies could examine these factors by including executive function measures or strategy assessments as covariates or moderators, which would deepen our understanding of variability in interference effects across individuals.
In summary, while this study advances the theoretical account of memory interference by emphasizing encoding as a critical locus of both retroactive and proactive effects, future research should adopt multi-method and multi-level approaches—combining behavioral paradigms, neural measures, and computational modeling—to further clarify the interplay between encoding and retrieval processes in shaping associative memory.
5. Conclusion
The present study investigated the mechanisms of retroactive (RI) and proactive interference (PI) in associative memory using a non-competitive two-alternative forced-choice (2AFC) paradigm. By minimizing retrieval-based competition and incorporating a source memory judgment task, we dissociated the effects of encoding and retention processes on memory interference. Our findings revealed an asymmetric pattern of interference: RI impaired associative recognition accuracy, whereas PI primarily manifested as slower response times, with no significant effect on accuracy. This dissociation suggests that RI and PI arise from distinct cognitive operations—RI reflecting degraded recollection due to representational suppression, and PI reflecting increased cognitive load from encoding complexity, likely arising from differentiation-based processes.The observed variability in interference effects suggests that memory is not only shaped by what is retrieved, but fundamentally by how and under what conditions it is encoded.
Authors Contributions
Yahui Zhang: Conceptualization, Data Curation, Formal analysis, Writing – original draft. Xiping Liu: Supervision, Methodology, Writing – review & editing Weihai Tang: Supervision, Methodology, Formal analysis Mei Peng: Visualization, Formal analysis, Data Curation.
Funding
No funds: grants, or other support was received.
Institutional Review Board Statement
This study was approved by the Ethics Committee of Tianjin Normal University (Approval No. 2024052105).
Informed Consent Statement
All subjects signed informed consent prior to the experiment.
Consent for publication: All authors approved the final manuscript and the submission to this journal.
Data Availability Statement
Data for the experiments are available https://osf.io/pfgca/. Code availability: Code for the experiments is available https://osf.io/pfgca/. Open Practices Statements: Data or materials for the experiments are available https://osf.io/pfgca/, and none of the experiments was preregistered.
Conflicts of Interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
References
- Anderson, J. R. , & Reder, L. M. (1999). The fan effect: New results and new theories. Journal of Experimental Psychology: General. [CrossRef]
- Anderson, M. C. , Bjork, R. A., & Bjork, E. L. (1994). Remembering can cause forgetting: Retrieval dynamics in long-term memory. Journal of Experimental Psychology: Learning, Memory, and Cognition, 1087. [Google Scholar] [CrossRef]
- Brodeur, M. B. , Dionne-Dostie, E., Montreuil, T., & Lepage, M. (2010). The Bank of Standardized Stimuli (BOSS), a New Set of 480 Normative Photos of Objects to Be Used as Visual Stimuli in Cognitive Research. ( 5(5), e10773. [CrossRef] [PubMed]
- Bower, G. H. (1972). Stimulus-sampling theory of encoding variability. In A. W. Melton & E. Martin (Eds.), Coding Processes in Human Memory (pp. 85–121). Washington, DC: Winston.
- Chanales, A. J. H. , Dudukovic, N. M., Richter, F. R., & Kuhl, B. A. (2019). Interference between overlapping memories is predicted by neural states during learning. A. ( 10(1), 5363. [CrossRef]
- Cohen, J. (2013). Statistical Power Analysis for the Behavioral Sciences. [CrossRef]
- Cowan, N. (2001). The magical number 4 in short-term memory: A reconsideration of mental storage capacity. Behavioral and Brain Sciences. [CrossRef]
- Cowan, N. (2019). Short-term memory based on activated long-term memory: A review in response to Norris (2017). Psychological Bulletin. [CrossRef]
- Dewar, M. T. , Cowan, N., & Sala, S. D. (2007). Forgetting Due to Retroactive Interference: A Fusion of Müller and Pilzecker’s (1900) Early Insights into Everyday Forgetting and Recent Research on Anterograde Amnesia. Cortex. [CrossRef]
- Favila, S. E. , Chanales, A. J. H., & Kuhl, B. A. (2016). Experience-dependent hippocampal pattern differentiation prevents interference during subsequent learning. Nature Communications. [CrossRef]
- Favila, S. E. , Lee, H., & Kuhl, B. A. (2020). Transforming the Concept of Memory Reactivation. A. ( 43(12), 939–950. [CrossRef]
- Gabitov, E. , Lungu, O., Albouy, G., & Doyon, J. (2019). Weaker Inter-hemispheric and Local Functional Connectivity of the Somatomotor Cortex During a Motor Skill Acquisition Is Associated With Better Learning. Frontiers in Neurology. [CrossRef]
- Herszage, J. , & Censor, N. (2017). Memory Reactivation Enables Long-Term Prevention of Interference. Current Biology, 1534. [Google Scholar] [CrossRef]
- Horner, A. J. , Bisby, J. A., Bush, D., Lin, W.-J., & Burgess, N. (2015). Evidence for holistic episodic recollection via hippocampal pattern completion. ( 6(1), 7462. [CrossRef] [PubMed]
- Huijbers, W. , Pennartz, C. M., Cabeza, R., & Daselaar, S. M. (2009). When Learning and Remembering Compete: A Functional MRI Study. PLoS Biology, 0001. [Google Scholar] [CrossRef]
- Hulbert, J. C. , & Norman, K. A. (2015). Neural Differentiation Tracks Improved Recall of Competing Memories Following Interleaved Study and Retrieval Practice. A. ( 25(10), 3994–4008. [CrossRef]
- Kim, G. , Norman, K. A., & Turk-Browne, N. B. (2017). Neural Differentiation of Incorrectly Predicted Memories. B. ( 37(8), 2022–2031. [CrossRef]
- Koen, J. D. , & Rugg, M. D. (2016). Memory Reactivation Predicts Resistance to Retroactive Interference: Evidence from Multivariate Classification and Pattern Similarity Analyses. The Journal of Neuroscience, 4399. [Google Scholar] [CrossRef]
- Kuhl, B. A. , Bainbridge, W. A., & Chun, M. M. (2012). Neural Reactivation Reveals Mechanisms for Updating Memory. The Journal of Neuroscience, 3461. [Google Scholar] [CrossRef]
- Kuhl, B. A. , Rissman, J., Chun, M. M., & Wagner, A. D. (2011). Fidelity of neural reactivation reveals competition between memories. D. ( 108(14), 5903–5908. [CrossRef]
- Kuhl, B. A. , Shah, A. T., DuBrow, S., & Wagner, A. D. (2010). Resistance to forgetting associated with hippocampus-mediated reactivation during new learning. Nature Neuroscience. [CrossRef]
- Levy, B. J. , & Anderson, M. C. (2002). Inhibitory processes and the control of memory retrieval. C. ( 6(7), 299–305. [CrossRef]
- Luck, S. J. , & Vogel, E. K. (1997). The capacity of visual working memory for features and conjunctions. K. ( 390(6657), 279–281. [CrossRef]
- Martin, E. (1968). Stimulus meaningfulness and paired-associate transfer: An encoding variability hypothesis. Psychological Review. [CrossRef]
- Martin, E. (1971). Verbal learning theory and independent retrieval phenomena. Psychological Review. [CrossRef]
- McGeoch, J. A. (1932). Forgetting and the law of disuse. Psychological Review. [CrossRef]
- Mensink. (1988). A Model for Interference and Forgetting. ( 95(4), 434–455. [CrossRef]
- Mensink,; Raaijmakers, J. G. W. (1989). A model for contextual fluctuation. G. W. ( 33(2), 172–186. [CrossRef]
- Moscovitch, M. , Cabeza, R., Winocur, G., & Nadel, L. (2016). Episodic Memory and Beyond: The Hippocampus and Neocortex in Transformation. Annual Review of Psychology. [CrossRef]
- Norman, K. A. , Newman, E. L., & Detre, G. (2007). A neural network model of retrieval-induced forgetting. Psychological Review. [CrossRef]
- Norman, K. A. , & O’Reilly, R. C. (2003). Modeling hippocampal and neocortical contributions to recognition memory: A complementary-learning-systems approach. Psychological Review. [CrossRef]
- Polack, C. W. , Jozefowiez, J., & Miller, R. R. (2017). Stepping back from ‘persistence and relapse’ to see the forest: Associative interference. Behavioural Processes. [CrossRef]
- Postman, L. , & Gray, W. (1977). Maintenance of Prior Associations and Proactive Inhibition. Journal of Experimental Psychology: Human Learning and Memory. [CrossRef]
- Postman, L. , & Underwood, B. J. (1973). Critical issues in interference theory. Memory & Cognition. [CrossRef]
- Raaijmakers,; Shiffrin, R. M. (1981). Search of associative memory. M. ( 88(2), 93–134. [CrossRef]
- Ranganath, C. (2010). Binding items and contexts: The cognitive neuroscience of episodic memory. Current Directions in Psychological Science. [CrossRef]
- Ratcliff, R. , & McKoon, G. (2008). The diffusion decision model: Theory and data for two-choice decision tasks. Neural Computation. [CrossRef]
- Renoult, L. , Davidson, P. S. R., Schmitz, E., Park, L., Campbell, K., Moscovitch, M., & Levine, B. (2015). Autobiographically Significant Concepts: More Episodic than Semantic in Nature? An Electrophysiological Investigation of Overlapping Types of Memory. Journal of Cognitive Neuroscience. [CrossRef]
- Richter, F. R. , Chanales, A. J. H., & Kuhl, B. A. (2016). Predicting the integration of overlapping memories by decoding mnemonic processing states during learning. A. ( 124, 323–335. [CrossRef]
- Ritvo, V. J. H. , Nguyen, A., Turk-Browne, N. B., & Norman, K. A. (2024a). A neural network model of differentiation and integration of competing memories. A. ( 12, RP88608. [CrossRef]
- Ritvo, V. J. H. , Nguyen, A., Turk-Browne, N. B., & Norman, K. A. (2024b). Differentiation and Integration of Competing Memories: A Neural Network Model, 5352. [Google Scholar] [CrossRef]
- Ritvo, V. J. H. , Turk-Browne, N. B., & Norman, K. A. (2019). Nonmonotonic Plasticity: How Memory Retrieval Drives Learning. Trends in Cognitive Sciences. [CrossRef]
- Sajikumar, S. , Morris, R. G. M., & Korte, M. (2014). Competition between recently potentiated synaptic inputs reveals a winner-take-all phase of synaptic tagging and capture. Proceedings of the National Academy of Sciences, 1221. [Google Scholar] [CrossRef]
- Schlichting, M. L. , & Preston, A. R. (2015). Memory integration: Neural mechanisms and implications for behavior. Current Opinion in Behavioral Sciences. [CrossRef]
- Shohamy, D. , & Wagner, A. D. (2008). Integrating Memories in the Human Brain: Hippocampal-Midbrain Encoding of Overlapping Events. Neuron. [CrossRef]
- Tzeng, O. J. (1973). Stimulus meaningfulness, encoding variability, and the spacing effect. Journal of Experimental Psychology, 99(2), 162–166. [CrossRef]
- Victoria, J.H. Ritvo, Nicholas B. Turk-Browne, & Kenneth A. Norman. (2019). Nonmonotonic Plasticity: How Memory Retrieval Drives Learning. Trends in Cognitive Sciences.
- Wang, Y. N. , Zhou, L. M., & Luo, Y. J. (2008). The Pilot Establishment and Evaluation of Chinese Affective Words System. Chinese Mental Health Journal. [CrossRef]
- Watkins, O. C. , & Watkins, M. J. (1975). Buildup of proactive inhibition as a cue-overload effect. Journal of Experimental Psychology: Human Learning and Memory. [CrossRef]
- Wimber, M. , Rutschmann, R. M., Greenlee, M. W., & Bäuml, K.-H. T. (2009). Retrieval from Episodic Memory: Neural Mechanisms of Interference Resolution. Journal of Cognitive Neuroscience. [CrossRef]
- Wixted, J. T. , & Mickes, L. (2010). A continuous dual-process model of remember/know judgments. Psychological Review, 1054. [Google Scholar] [CrossRef]
- Yassa, M. A. , & Stark, C. E. L. (2011). Pattern separation in the hippocampus. E. L. ( 34(10), 515–525. [CrossRef]
- Yonelinas, A. P. (2002). The nature of recollection and familiarity: A review of 30 years of research. Journal of Memory and Language. [CrossRef]
- Zeithamova, D. , & Preston, A. R. (2010). Flexible Memories: Differential Roles for Medial Temporal Lobe and Prefrontal Cortex in Cross-Episode Binding. The Journal of Neuroscience, 4676. [Google Scholar] [CrossRef]
- Zeithamova, D. , & Preston, A. R. (2017). Temporal Proximity Promotes Integration of Overlapping Events. R. ( 29(8), 1311–1323. [CrossRef]
Figure 1.
Experimental procedure. Participants studied two lists of word-picture pairs, each containing half cue overlap pairs (AB or AC condition, shown in red frames for illustrative purposes) and half non-overlap pairs (EF or GH condition, shown in green frames for illustrative purposes).
Figure 1.
Experimental procedure. Participants studied two lists of word-picture pairs, each containing half cue overlap pairs (AB or AC condition, shown in red frames for illustrative purposes) and half non-overlap pairs (EF or GH condition, shown in green frames for illustrative purposes).
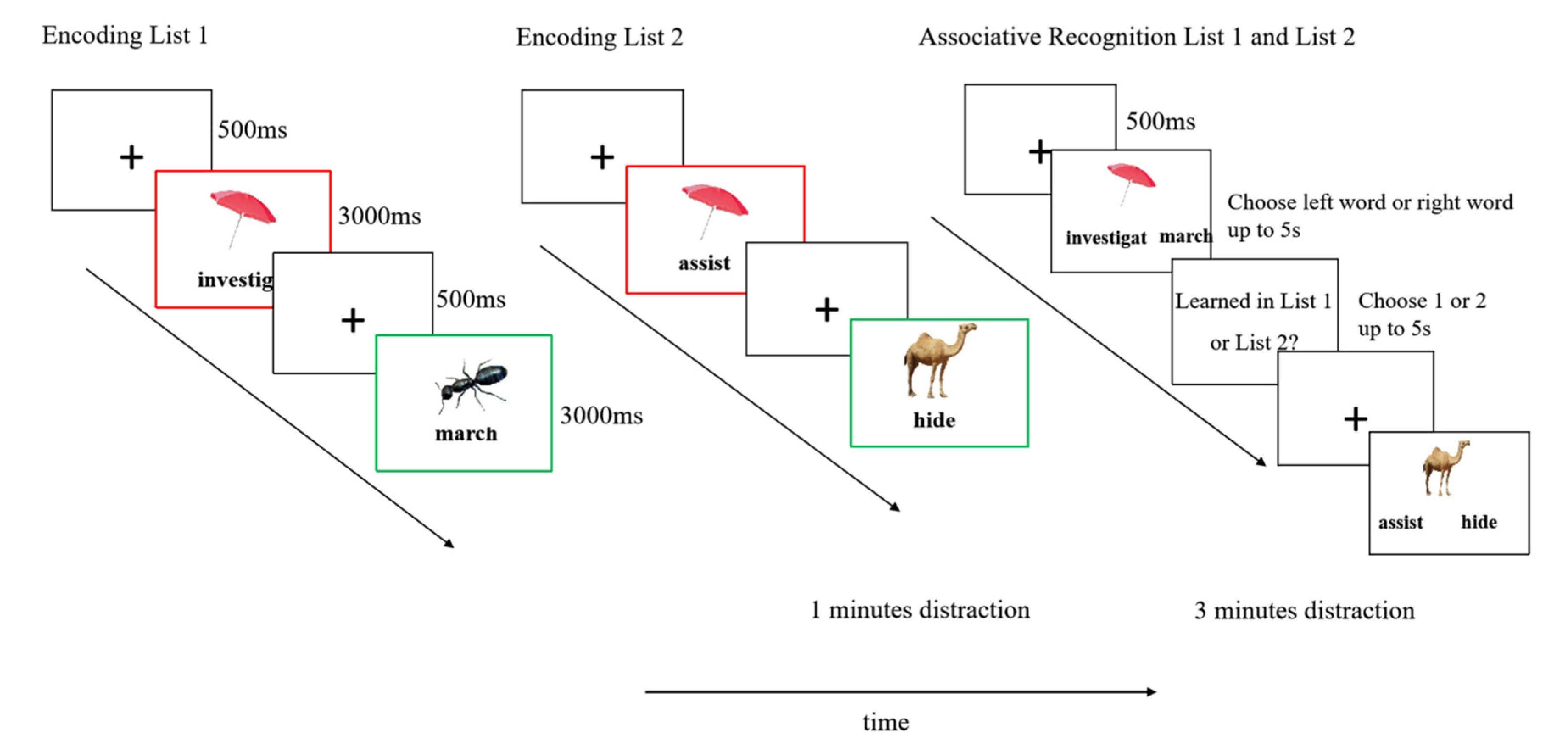
Figure 2.
Associative recognition accuracy as a function of pair type and study list. Boxes show IQR (25th–75th percentiles), solid lines represent medians, whiskers extend to 1.5×IQR, and white circles indicate condition means. Significant differences between cue-overlap and non-overlap pairs are marked with asterisks (*p <0.05, **p <0.01, ***p < .001); n.s. = not significant (p ≥ .05). Post hoc tests adjusted with Bonferroni correction.
Figure 2.
Associative recognition accuracy as a function of pair type and study list. Boxes show IQR (25th–75th percentiles), solid lines represent medians, whiskers extend to 1.5×IQR, and white circles indicate condition means. Significant differences between cue-overlap and non-overlap pairs are marked with asterisks (*p <0.05, **p <0.01, ***p < .001); n.s. = not significant (p ≥ .05). Post hoc tests adjusted with Bonferroni correction.
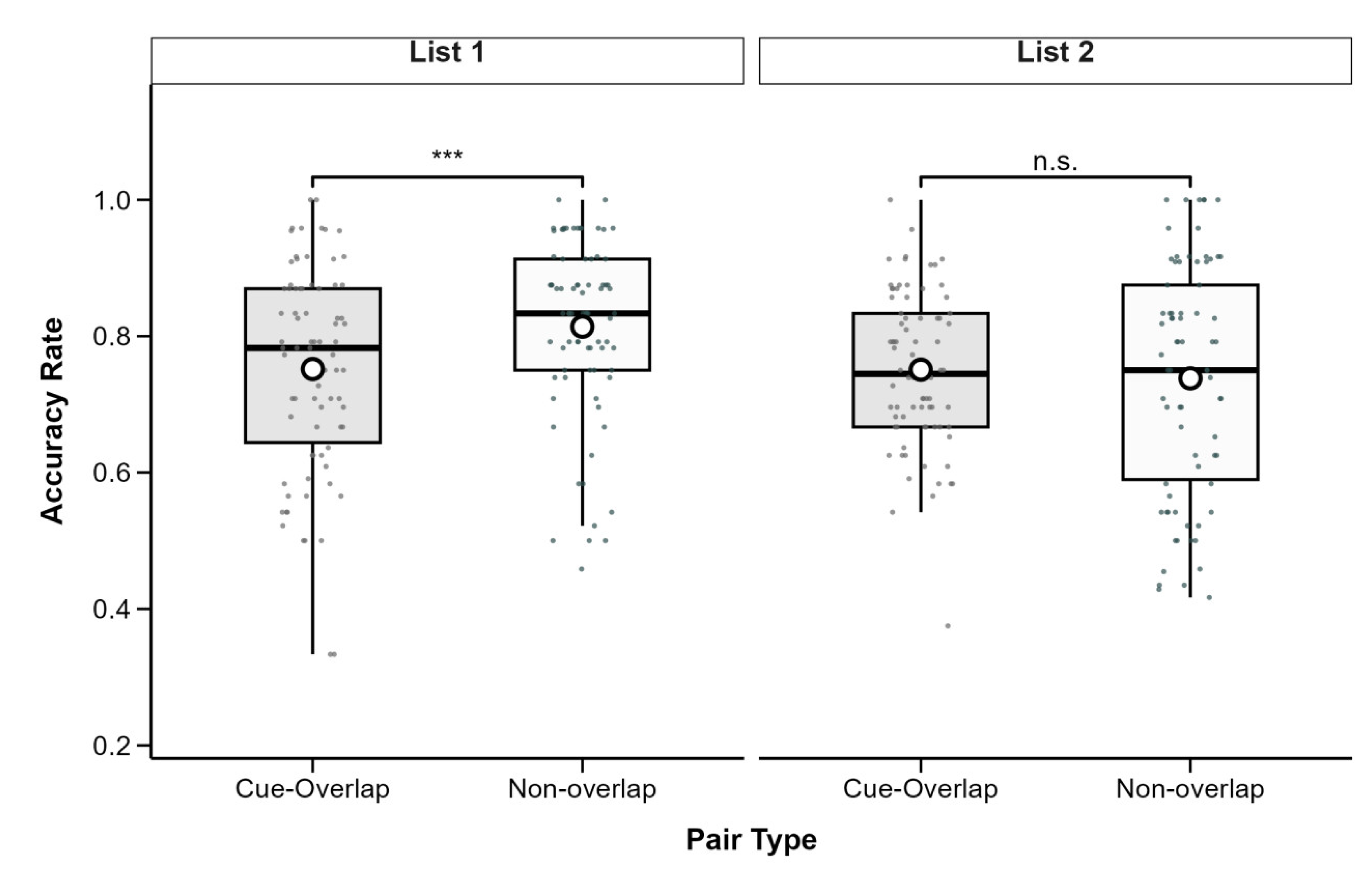
Figure 3.
Boxplots of recognition response time by pair type and learning list, separated by recognition accuracy. Boxes show IQR (25th–75th percentiles), solid lines represent medians, whiskers extend to 1.5×IQR, and white circles indicate condition means. Significant differences between cue-overlap and non-overlap pairs are marked with asterisks (*p <0.05, **p <0.01, ***p < .001); n.s. = not significant (p ≥ .05). Post hoc tests adjusted with Bonferroni correction.
Figure 3.
Boxplots of recognition response time by pair type and learning list, separated by recognition accuracy. Boxes show IQR (25th–75th percentiles), solid lines represent medians, whiskers extend to 1.5×IQR, and white circles indicate condition means. Significant differences between cue-overlap and non-overlap pairs are marked with asterisks (*p <0.05, **p <0.01, ***p < .001); n.s. = not significant (p ≥ .05). Post hoc tests adjusted with Bonferroni correction.
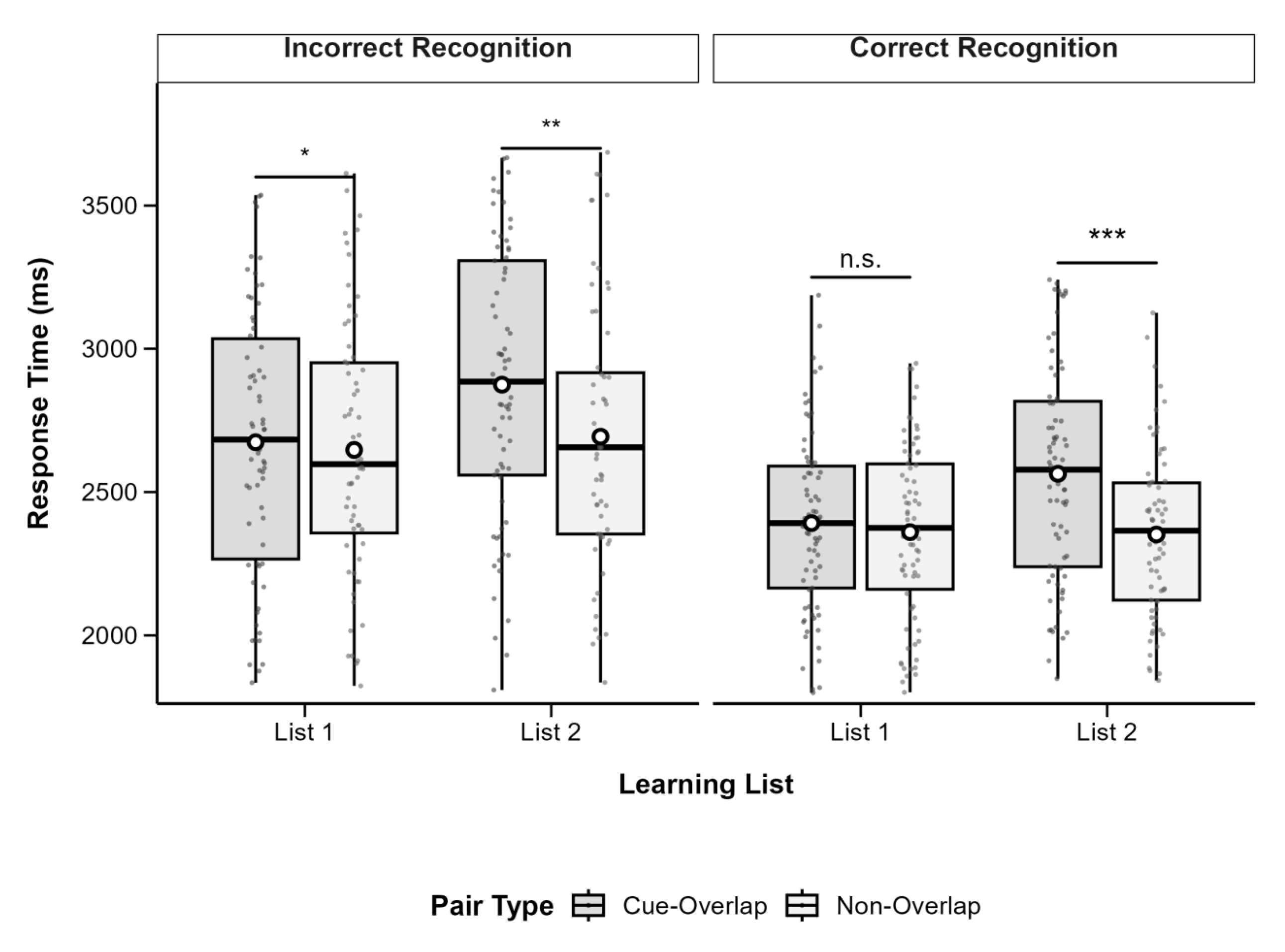
Figure 4.
Boxplots of source judgement accuracy by pair type and learning list. Boxes show IQR (25th–75th percentiles), solid lines represent medians, whiskers extend to 1.5×IQR, and white circles indicate condition means. Significant differences between cue-overlap and non-overlap pairs are marked with asterisks (*p <0.05, **p <0.01, ***p < .001); n.s. = not significant( p ≥ .05). Post hoc tests adjusted with Bonferroni correction.
Figure 4.
Boxplots of source judgement accuracy by pair type and learning list. Boxes show IQR (25th–75th percentiles), solid lines represent medians, whiskers extend to 1.5×IQR, and white circles indicate condition means. Significant differences between cue-overlap and non-overlap pairs are marked with asterisks (*p <0.05, **p <0.01, ***p < .001); n.s. = not significant( p ≥ .05). Post hoc tests adjusted with Bonferroni correction.
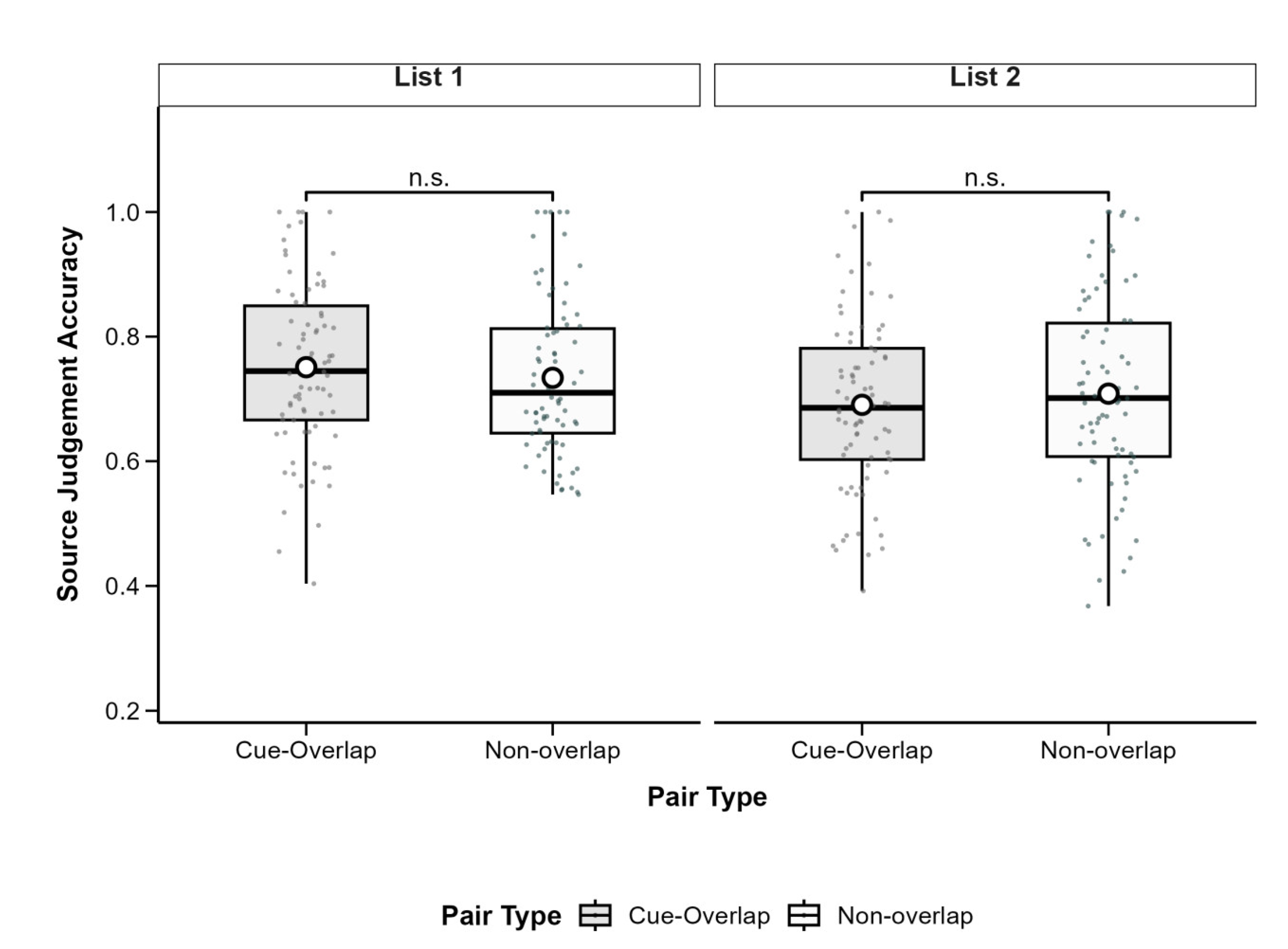

Table 1.
Mean (±SD) recognition accuracy and reaction times (ms) by pair type, list, and response accuracy.
Table 1.
Mean (±SD) recognition accuracy and reaction times (ms) by pair type, list, and response accuracy.
| List | Pair Type | Recognition Accuracy (M ± SD) | Correct Reaction Time(ms) | Incorrect Reaction Time(ms) |
| List 1 | Cue-overlap | 0.75 ± 0.15 | 2339.94 ± 363.04 | 2667.74 ± 647.66 |
| Non-overlap | 0.81 ± 0.13 | 2319.70 ± 346.73 | 2631.99 ± 645.72 | |
| List 2 | Cue-overlap | 0.75 ± 0.11 | 2513.32 ± 427.81 | 2867.40 ± 572.10 |
| Non-overlap | 0.74 ± 0.17 | 2273.00 ± 354.35 | 2668.13 ± 639.50 |
Note. RT = reaction time (in milliseconds). Accuracy is reported as proportion correct.
Table 2.
Mean Accuracy and Reaction Times for Source Judgments by Pair Type in List 1 and List 2.
| List | Pair Type | Source Judgement Accuracy (M ± SD) | Correct Reaction Time(ms) | Incorrect Reaction Time(ms) |
| List 1 | Cue-overlap | 0.75 ± 0.15 | 561.45± 343.75 | 680.074 ± 406.29 |
| Non-overlap | 0.73 ± 0.14 | 608.35 ± 381.49 | 750.08 ± 442.67 | |
| List 2 | Cue-overlap | 0.70 ± 0.15 | 620.62 ± 394.04 | 662.81 ± 427.18 |
| Non-overlap | 0.69 ± 0.16 | 628.75 ± 379.66 | 667.05 ± 427.20 |
Note. RT = reaction time (in milliseconds). Accuracy is reported as proportion correct.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



