
Submitted:
28 July 2025
Posted:
29 July 2025
You are already at the latest version
Abstract
Glycine is an amino acid that functions as both a carbon and nitrogen source. Glycine mixotrophy enhances microalgal biomass, reduces dependency on light, and optimizes carbon and nitrogen allocation, thereby facilitating the adaptation of microalgae to industrial production environments. This study investigated the physiological and biochemical parameters, as well as gene expression profiles, of Isochrysis zhanjiangensis under varying glycine concentrations to elucidate the effects of glycine on fucoxanthin content and yield, along with its underlying mechanisms. At the physiological and biochemical level, fucoxanthin yield peaked at 4.74 mg/L under an 8 g/L glycine concentration, a 6.36-fold increase compared to the control group. Biomass reached its maximum of 9.38 × 106 cells·mL-1 at a 2 g/L glycine concentration, reflecting a 62.41% increase over the control. Chlorophyll content and fluorescence parameters revealed that glycine treatment increased chlorophyll levels, enhanced the photosynthetic activity of photosystem II (PSII), and mitigated photoinhibition. At the gene expression level, transcriptome analysis indicated that glycine significantly upregulated the expression of genes encoding enzymes in the Calvin cycle, potentially enhancing carbon fixation and providing energy for algal cell proliferation. A portion of the carbon flux was redirected toward fatty acid storage. Additionally, the fucoxanthin biosynthesis pathway gene ZEP1 exhibited elevated expression, collectively contributing to the observed increase in fucoxanthin yield.
Keywords:
Isochrysis zhanjiangensis
; glycine
; mixotrophy
; fucoxanthin
; photosynthesis
; transcriptome analysis
; carbon metabolism
1. Introduction
Fucoxanthin (Fx) is a carotenoid unique to brown algae, diatoms, and golden algae, exhibiting significant potential as a bioactive molecule. It possesses critical physiological functions, including the suppression of cancer cell expression [1], treatment of Alzheimer’s disease [2], anti-obesity effects [3], and anti-diabetic properties [4]. Furthermore, fucoxanthin offers potential protective effects against various inflammation-related diseases. Traditionally, fucoxanthin is extracted from macroalgae such as Undaria pinnatifida and Laminaria japonica, but its low content in these sources renders it extremely costly, far exceeding the price of gold. Microalgae, with their rapid growth rates and elevated fucoxanthin content, have emerged as a focal point for fucoxanthin production research.
Isochrysis zhanjiangensis, a marine unicellular microalga rich in fucoxanthin [5], lacks a cytoderm [6], which facilitates fucoxanthin extraction. This species can withstand temperatures up to 35°C, and thrives across a broad temperature range, underscoring its potential for industrial fucoxanthin production. Recent studies indicate that fucoxanthin content in I. zhanjiangensis is significantly influenced by light intensity and spermidine [7]. Under low light conditions (20 μmol·m-2·s-1) and 150 μM spermidine treatment, both fucoxanthin content and yield reached their highest levels, although the yield remained below 1.0 mg/L. While most microalgae are cultivated autotrophically, some are grown mixotrophically [8] or heterotrophically [9] using carbon or nitrogen sources. Compared to mixotrophic cultivation, autotrophic growth requires more time to achieve equivalent biomass accumulation and is often constrained by light availability in later growth stages. Mixotrophy, however, partially alleviates light limitation, yielding greater biomass. Certain species, such as Nannochloropsis laevis and Cyclotella cryptica, can grow heterotrophically in the dark using organic carbon sources, sometimes exhibiting higher fucoxanthin content than autotrophic cultures [10,11]. Supplementing nutrients and altering microalgal nutritional modes offer an effective strategy for high-density cultivation.
Glycine (Gly), an amino acid with dual carbon and nitrogen source properties, demonstrates high bioavailability as a nitrogen source for microalgae due to its solubility and stability in aquatic environments [12]. Its metabolites contribute to energy metabolism pathways, including glycolysis and the tricarboxylic acid cycle, in algal cells [13]. Glycine mixotrophy has been shown to promote biomass accumulation in I. zhanjiangensis [14], a finding corroborated in other algal species [15,16]. Additionally, glycine enhances secondary metabolite content in plants and algae [16,17], though its impact on fucoxanthin levels in I. zhanjiangensis remains unreported.
Notably, glycine also modulates photosynthesis in plants. For instance, 2.85 mM glycine increases photosynthesis in rice leaves under cold stress by upregulating chlorophyll content, Y(II), and qP [18]. Exogenous 10 mM glycine mitigates drought stress in Pinus bungeana by enhancing Fv/Fm, Y(II), and NPQ [19], while 5 mM glycine boosts the net photosynthetic rate in Caragana korshinskii [20]. Whether glycine similarly enhances photosynthetic efficiency in I. zhanjiangensis, thereby influencing the synthesis of the light-harvesting pigment fucoxanthin, remains unexplored.
Transcriptome analysis has increasingly been applied to investigate algal metabolite metabolism. Liu, et al. [21] utilized transcriptomics to identify key genes and their functions in methyl jasmonate-induced fucoxanthin synthesis in Phaeodactylum tricornutum, suggesting that methyl jasmonate enhances fucoxanthin content via amino acid biosynthesis and carbon metabolism pathways. Li, et al. [22] employed transcriptomics to examine the effects of different light wavelengths on carotenoid biosynthesis in Dunaliella salina. Additionally, transcriptomics has elucidated the influence of nutritional modes on secondary metabolite production within the same species. Pal, et al. [23] used comparative transcriptomics to uncover differences in autotrophic and heterotrophic modes related to secondary metabolite biosynthesis in Swertia chirayita. Given the complex and poorly understood metabolic processes of I. zhanjiangensis, transcriptome analysis represents a viable and comprehensive approach to investigate the mechanisms by which glycine mixotrophy affects fucoxanthin synthesis pathways in this species.
2. Results
2.1. Effects of Glycine Concentration on the Growth of I. zhanjiangensis
The effects of different glycine concentrations on the growth of I. zhanjiangensis are presented in Figure 1. On the 11th day, the 2 g/L Gly treatment group exhibited the highest cell density, reaching 9.38×10⁶ cells·mL⁻¹, which was 62.41% higher than that of the control group. All glycine-treated groups showed a substantial increase in cell density compared to the control group, with all differences being highly significant (P < 0.001). Meanwhile, the control group gradually entered the plateau phase on the sixth day, whereas the glycine-treated groups continued to increase in cell number, with a decline in growth rate observed only on the tenth day, marking the onset of the plateau phase. Although the high-concentration glycine treatment groups (16 g/L and 32 g/L) exhibited evident growth inhibition compared to the control group during the early stage of algal growth (days 1 to 6), their cell densities by the 11th day were still highly significantly greater than that of the control group (P < 0.001).
2.2. Effects of Glycine Concentration on Fucoxanthin Content and Yield in I. zhanjiangensis
The influence of glycine on fucoxanthin content is depicted in Figure 2A. Except for the 2 g/L treatment group, the intracellular fucoxanthin content of I. zhanjiangensis increased highly significantly with increasing glycine concentration (P < 0.001). The intracellular fucoxanthin content exhibited a continuous upward trend with increasing glycine concentration and began to stabilize at 16 g/L; the 16 g/L and 32 g/L treatment groups displayed the highest intracellular fucoxanthin content, reaching 5.44~5.51×10⁻⁴ mg/10⁵ cells, representing an increase of 96.35% to 99.04% compared to the control group.
Fucoxanthin yield (Figure 2B) was significantly higher in all glycine treatments compared to the control (P < 0.0001). The fucoxanthin yield per unit volume of algal culture showed a trend of initially increasing and then decreasing with increasing glycine concentration; at 8 g/L glycine, the optimal treatment concentration, the fucoxanthin yield reached its maximum value of 4.78 mg/L, which was 5.41-fold higher than the control.
In scaled-up cultures (1 L), the fucoxanthin content and yield results for the control group and the optimal glycine treatment concentration (8 g/L) of I. zhanjiangensis are presented in Figure 2C and Figure 2D. Under the 8 g/L glycine treatment, the fucoxanthin yield of I. zhanjiangensis was 2.96 g/L, an increase of 69.16% compared to the control group; the content was 3.50×10⁻⁴ mg/10⁵ cells, an increase of 50.89% compared to the control group, with both differences being highly significant (P < 0.0001).
2.3. Effects of Glycine Concentrations on Chlorophyll Concentration in I. zhanjiangensis
The impact of glycine on chlorophyll content is shown in Figure 3. Glycine-treated groups exhibited higher chlorophyll a, chlorophyll c, and total chlorophyll levels than the control, with concentrations initially increasing and then declining with rising glycine levels. The 8 g/L treatment yielded the highest values: 2.19 mg/L (chlorophyll a), 1.37 mg/L (chlorophyll c), and 3.54 mg/L (total chlorophyll), corresponding to 6.16-fold, 1.44-fold, and 2.72-fold increases over the control, respectively (P < 0.0001). Chlorophyll a showed a more pronounced increase than chlorophyll c.
2.4. Effects of Glycine Concentrations on Chlorophyll Fluorescence Parameters and ROS Levels in I. zhanjiangensis
Changes in chlorophyll fluorescence parameters with glycine concentration are illustrated in Figure 4. The maximum quantum yield (Fv/Fm) decreased at 2 g/L and then increased (Figure 4A). The actual photosynthetic efficiency (Y(II)) rose with concentration, stabilizing at 0.493–0.497 beyond 8 g/L (P < 0.0001 vs. control) (Figure 4B). The photochemical quenching coefficient (qP) increased significantly at higher concentrations (P < 0.01) (Figure 4C), while non-photochemical quenching (NPQ) decreased significantly across all treatments (P < 0.0001) (Figure 4D). Reactive oxygen species (ROS) levels initially declined, reaching a minimum at 16 g/L, then increased (Figure 4E).
2.5. Potential Mechanisms of Glycine Affecting Cell Proliferation and Fucoxanthin Synthesis in I. zhanjiangensis
Figure 5 reveals the potential mechanisms by which glycine affects cell proliferation and fucoxanthin accumulation in I. zhanjiangensis. Transcriptome results showed that the expression of the glycine decarboxylase gene (GLDC), a key enzyme in the glycine cleavage system (GCS), was upregulated by 1.10, suggesting that after entering the cell, glycine may be catalyzed by GCS to form ammonia, carbon dioxide, and serine. The expression of glutamine synthetase (GS), a key enzyme in nitrogen assimilation, was upregulated by 2.27, indicating that the ammonia produced may be linked to the glutamine synthetase-glutamate synthase (GS-GOGAT) cycle and participate in nitrogen assimilation. As shown in Figure 5, the expression of genes involved in catalyzing the interconversion of various substances in the Calvin cycle, namely GAPDH, TK, PGK, PRK, and rbcL, was upregulated by 3.05, 3.07, 2.7, 2.26, and 1.57, respectively. Simultaneously, the gene transcription level of ferredoxin-NADP⁺ reductase (FNR) was observed to increase by 4.78.
The expression of genes related to the fucoxanthin chlorophyll a/c-binding protein (FCP) subunits, which are closely associated with algal photosynthetic efficiency and composed of fucoxanthin, also underwent significant upregulation. A total of 29 FCP subunit genes were significantly upregulated, with Lhcq11, Lhcr2, and Lhcf1 upregulated by 4.42, 4.03, and 3.72, respectively. In the fucoxanthin biosynthesis pathway, PDS, ZEP, and VDE were upregulated by 1.51, 2.18, and 1.33 times, respectively, while ZDS and LCYb were downregulated by 0.83 and 0.72 times, respectively (Figure 5).
In the carbon source metabolism process, the transcription level of the ACS gene, which encodes the key enzyme acetyl-CoA carboxylase (ACCase), increased by 2.95. ACCase has been confirmed as a critical enzyme in fatty acid synthesis, and its expression is closely related to fatty acid synthesis. Fatty acids are the primary form of energy storage and transfer in cells and may serve as the metabolic direction for the carbon flow produced by carbon fixation.
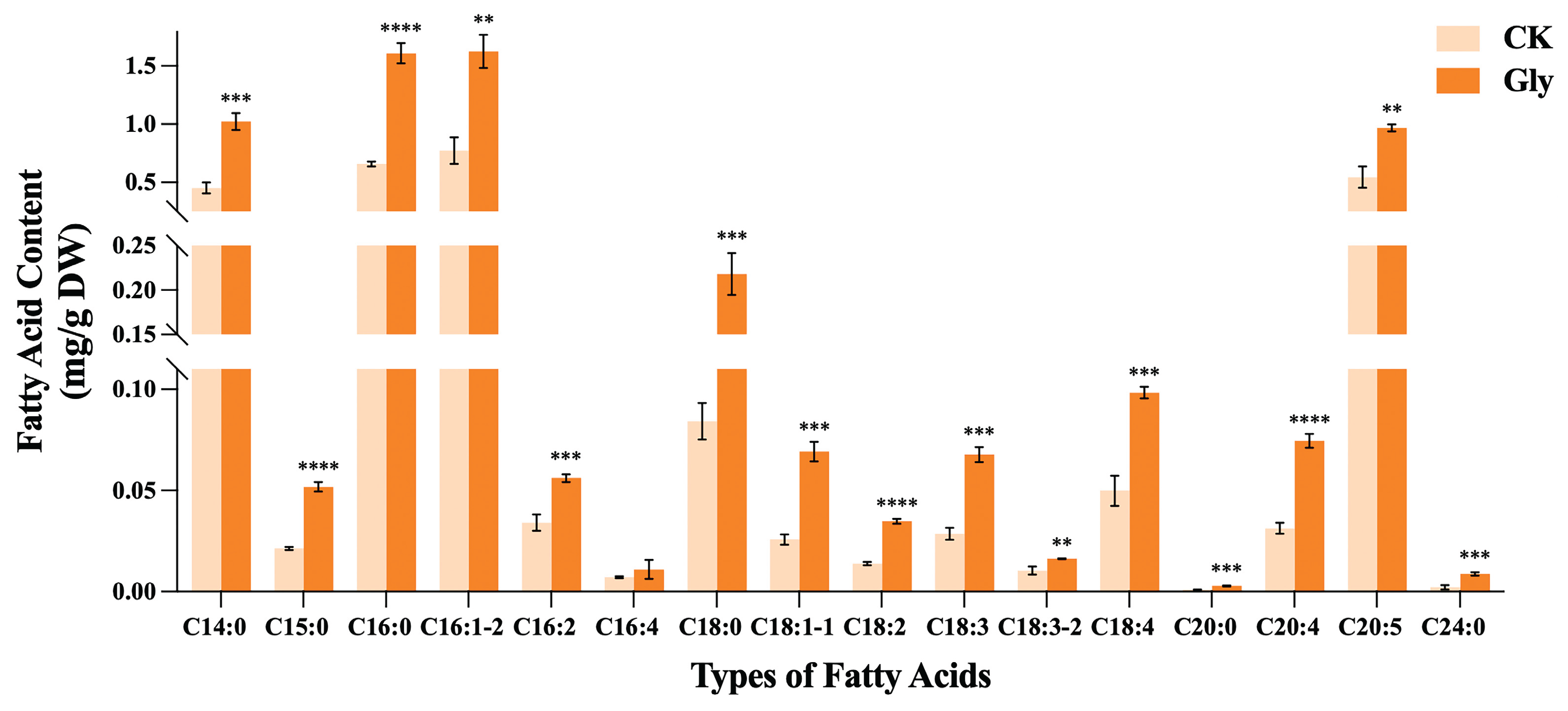
To verify the direction of carbon flow, we measured the fatty acid profiles of I. zhanjiangensis in an 8 g/L glycine treatment group and a control group (Figure 6). Following the addition of glycine, the overall fatty acid content was higher than that of the control group, with all individual fatty acids except hexadecatetraenoic acid(C16:4) exhibiting a highly significant increase (P < 0.01). Specifically, the content of eicosapentaenoic acid (EPA, C20:5) was relatively high and increased by 77.60% compared to the control group. Additionally, in the glycine-treated I. zhanjiangensis, oleic acid (C18:1), linoleic acid (C18:2), and arachidonic acid (C20:4) also showed substantial increases, with elevations of 168.45%, 151.25%, and 138.04%, respectively, compared to the control group.
3. Discussion
3.1. Glycine Significantly Promotes the Accumulation of Biomass in I. zhanjiangensis
Growth results indicate that the cell number in the glycine treatment group was significantly higher than that in the control group. This effect is not exclusive to I. zhanjiangensis; when certain microalgae utilize glycine as a nitrogen source, they also exhibit higher cell densities [24-26]. Concurrently, studies have found that Tetraselmis sp. under glycine treatment demonstrates a higher cell growth rate [27]. Research confirms that glycine can form dissolved free amino acids in the culture medium, making it more readily utilizable by microalgae [24]. Within cells, glycine can be catalyzed by glycine decarboxylase (GDC) and serine hydroxymethyltransferase (SHMT) to convert two glycine molecules into one serine molecule, releasing one molecule of CO₂ and NH₃ [28]. The NH₃ can be reutilized to form various amino acids required by the cell, while CO₂ can participate in numerous metabolic activities within the cell. Additionally, glycine can assist algal cells in alleviating photoinhibition during the mid-to-late growth stages, potentially by reducing photosynthetic metabolic stress and regulating energy metabolism to promote algal cell proliferation. Woo et al. [29] suggest that glycine, through oxidative decarboxylation in mitochondria, provides NADH for NO₃⁻ reduction. The increase in intracellular NH₃ and CO₂ levels may supply fundamental substances for cell metabolism, enhance its reaction capacity, and accelerate energy production processes, thereby resulting in faster cell growth and increased density.
However, ammonia also exerts an inhibitory effect on cells, which varies depending on the microalgal species. Reports indicate that the marine microalga Dunaliella tertiolecta exhibits cell growth inhibition when ammonia concentrations exceed 10 mM [30]. Since glycine is enzymatically decomposed into CO₂ and NH₃ within cells, higher glycine concentrations may lead to excessively high intracellular ammonia levels. This excessive ammonia concentration may be one of the reasons why the cell concentrations in the 16 g/L and 32 g/L treatment groups showed growth inhibition in the early growth stage compared to the control group, while the overall cell number remained lower than that in the 2 g/L, 4 g/L, and 8 g/L treatment groups.
3.2. Glycine Induces the Production of Fucoxanthin in Algae Through Carbon and Nitrogen Supplementation
Glycine can serve as both a nitrogen source and a carbon source to promote the accumulation of fucoxanthin in microalgae. Studies confirm that, compared to culture media without a carbon source, supplementation with a carbon source increased the fucoxanthin content in Pavlova sp. OPMS 30543 by 0.84 times [31]. The fucoxanthin content obtained from mixotrophic Nitzschia laevis was higher than that from autotrophic growth [32]. High nitrogen concentrations (≥300 mg/L) also facilitate greater fucoxanthin accumulation in microalgae (0.54% to 2.33% Dry Weight) [33-35]. As a light-harvesting pigment that absorbs light energy and transfers it to chlorophyll a, fucoxanthin can be utilized to synthesize fucoxanthin-chlorophyll protein complexes (FCP), enhancing the efficiency of light energy utilization in algal cells, as demonstrated in Phaeodactylum tricornutum [36]. Limited by nitrogen availability, the number of FCPs may be influenced by nitrogen levels [37]. Alleviating nitrogen limitation in algal cells may promote FCP formation, indirectly facilitating fucoxanthin production. Research shows that under low light conditions, an increased nitrogen supply directly promotes the formation of FCP complexes, yielding 2.78 times more fucoxanthin than in autotrophic cells [38]. [39] reported that in Tisochrysis lutea, nitrogen supplementation significantly enhanced FCP complex synthesis and increased fucoxanthin accumulation; furthermore, the gene expression levels of FCP complexes were markedly affected by nitrogen supply, suggesting that nitrogen is a critical factor in the synthesis of FCPs and fucoxanthin. High nitrogen supplementation may explain why the fucoxanthin content in the 16 g/L and 32 g/L treatment groups was significantly higher than in other groups. However, the 2 g/L glycine treatment group, which exhibited the highest cell density, and the 16 g/L and 32 g/L groups, with the highest fucoxanthin content, did not achieve the highest yield, possibly due to the mutual exclusivity between cell growth and pigment storage. After scaling up the culture, glycine maintained a significant advantage in promoting fucoxanthin production, providing a theoretical foundation for subsequent large-scale industrial production.
3.3. Glycine Promotes the Synthesis of Chlorophyll as a Nitrogen Source
Chlorophyll, a nitrogen-rich compound, serves as an intracellular nitrogen pool to support further cell growth and biomass production. Studies demonstrate that when nitrogen (70 and 700 mg L⁻¹ N) was added to nitrogen-starved Chlorella, the contents of chlorophyll a and chlorophyll b increased. This suggests that when nitrogen is sufficient, it accumulates within chlorophyll molecules [40]. Meanwhile, insufficient nitrogen stresses microalgae, impairing their chlorophyll and protein biosynthesis; algal cells grown without nitrogen typically adjust their metabolism to accumulate lipids and carotenoids, favoring respiration over photosynthesis [41,42]. On this basis, supplementing nitrogen mitigates stress, reduces respiration (including photorespiration), enhances algal photosynthesis, and increases photosynthetic pigment content. Thus, adding glycine as a nitrogen source is beneficial to the synthesis of photosynthetic pigments. Fathy et al. [16] found that glycine increased chlorophyll a, chlorophyll b, and carotenoids in Synechocystis sp. by 19.4, 44.3, and 55.3 times, respectively, compared to untreated strains. However, more nitrogen is not always better. At higher glycine treatment concentrations, the chlorophyll content in I. zhanjiangensis began to decline, possibly due to nitrogen stress. Ahmad et al. [43] observed that both low and high nitrogen treatments (0 and 300 kg ha⁻¹) resulted in lower chlorophyll a content in sorghum compared to a medium nitrogen treatment (150 kg ha⁻¹), with reductions of 26.2% and 27.7%, respectively. Excessive nitrogen supply does not enhance wheat growth, yield, or fertilizer use efficiency but instead reduces chlorophyll content and net photosynthetic rate [44]. Therefore, an appropriate nitrogen treatment concentration is crucial for increasing chlorophyll content.
3.4. Glycine Enhances the Photosynthetic Activity of I. zhanjiangensis
After adding glycine, the Fv/Fm ratio slightly increased compared to the control group, indicating that under mixotrophic conditions with glycine, photoinhibition in I. zhanjiangensis decreased, unlike other mixotrophic sources that may induce PSII photoinhibition [32]. Additionally, under high glycine concentration treatment, Y(II) significantly increased (P < 0.0001), reaching up to 1.80 times that of the control group. This suggests that glycine enables I. zhanjiangensis to utilize more energy for photosynthesis, rendering PSII more active and enhancing photosynthetic activity. Furthermore, qP results indicate an increase in energy used for photochemical reactions. Cao et al. [18] reported a similar phenomenon, where glycine treatment significantly improved Y(II) and qP values in rice seedlings, suggesting that glycine increases the proportion of open PSII reaction centers and maximizes light capture efficiency. This may, to some extent, promote the synthesis of the light-harvesting pigment fucoxanthin. Moreover, culturing with glycine significantly reduces energy loss due to non-photochemical quenching in I. zhanjiangensis. The reduction in NPQ indicates that most of the light energy captured by I. zhanjiangensis is utilized, decreasing the over-reduction of the photosynthetic electron transport chain caused by excess photons, reducing ROS production, and preventing damage to photosynthetic components, thereby ensuring the effective operation of the photosynthetic electron transport chain. Badran et al. [45] also confirmed in salt-stressed wheat that glycine facilitates more efficient ROS scavenging.
3.5. Potential Mechanisms by Which Glycine Affects Cell Proliferation and Fucoxanthin Synthesis in I. zhanjiangensis
The addition of glycine upregulates the expression of a series of enzymes related to carbon fixation, among which the rbcL gene encodes the large subunit of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO), a key enzyme in the Calvin cycle that catalyzes the incorporation of inorganic carbon (CO₂) into carbon fixation and the formation of energy-rich carbohydrates [46]. Part of the CO₂ generated from glycine decomposition may be transferred to the chloroplast stroma for carbon fixation. The elevated transcription level of the rbcL gene facilitates I. zhanjiangensis in fixing more inorganic carbon released from glycine, aiding in the accumulation of additional biomass. Photorespiration involving glycine has been shown to enhance carbon fixation in the Calvin cycle by reducing carbon loss [47], which may explain the upregulation of genes related to Calvin cycle enzymes. The exogenous addition of glycine may enhance photorespiratory metabolism, with downstream serine production serving as a carbon skeleton for further metabolism into PGA, participating in the Calvin cycle and increasing its carbon fixation capacity. According to Li et al. [48], under low light conditions, the carbon fixation capacity of I. zhanjiangensis is weak. Exogenous glycine addition strengthens carbon fixation within algal cells, enabling I. zhanjiangensis to synthesize more carbohydrates for central carbon metabolism, providing additional energy for cell growth, division, and the conversion of other metabolites. The increased transcription level of the FNR gene suggests that electron transfer between ferredoxin and NADPH may accelerate in glycine-treated algal cells, supplying sufficient reducing power for the Calvin cycle. The elevated transcription level of the GAPDH gene may indicate that more reducing power enters the Calvin cycle, enhancing the carbon fixation level in microalgal cells.
The addition of glycine also contributes to improved photosynthetic efficiency. Excessive reactive oxygen species (ROS) production is detrimental to algal cells, causing oxidative damage to photosynthetic systems and reducing photosynthetic efficiency. Li et al. [48] found that high light conditions (HL) generate more ROS, leading to attacks on photosynthetic components by ROS and inhibiting fucoxanthin formation. Glycine addition effectively reduces ROS content in I. zhanjiangensis, thereby mitigating oxidative damage to the photosynthetic system by ROS and allowing the electron transport process in photosynthesis to proceed efficiently. Additionally, low light conditions produce less ROS than high light conditions and induce algae to synthesize more carotenoids, such as lutein, to capture and transfer light energy, potentially promoting the formation of the light-harvesting complex FCP. Simultaneously, nitrogen, an essential nutrient for protein synthesis, influences FCP synthesis. In Phaeodactylum tricornutum, FCP and general photosynthetic pathways are downregulated during nitrogen stress [49]. Glycine treatment provides a nitrogen-rich growth environment for I. zhanjiangensis, resulting in significant upregulation of 29 FCP subunit genes. The upregulation of FCP protein subunit gene expression following nitrogen supplementation may induce more fucoxanthin to bind with them, promoting fucoxanthin synthesis to some extent.
The key enzyme genes in the fucoxanthin synthesis pathway, ZEP and VDE, have been proven critical for fucoxanthin synthesis in Phaeodactylum tricornutum, with diatom mutants lacking VDL2 or ZEP1 showing no fucoxanthin [50]. Additionally, zeaxanthin, an upstream precursor in fucoxanthin synthesis, increases in content to favor conversion to downstream products, promoting fucoxanthin synthesis [51] The ZEP gene exhibits a greater fold increase in expression compared to VDE, facilitating the conversion of zeaxanthin to violaxanthin and thus promoting fucoxanthin synthesis. Therefore, glycine treatment enhances the upregulation of genes in the fucoxanthin synthesis pathway in I. zhanjiangensis, increasing fucoxanthin content and yield.
During carbon source metabolism, the transcription level of the ACS gene, encoding the key enzyme acetyl-CoA carboxylase (ACCase), increased by 2.95. ACCase is confirmed as a critical enzyme in fatty acid synthesis, with its expression closely tied to fatty acid production. Studies show that 2-oxoglutarate (2-OG) enhances fatty acid synthesis efficiency by alleviating inhibition of ACCase [52]. The carbon fixation process also provides numerous precursors for fatty acid synthesis, such as PGA and G3P. Fatty acids, the primary form of energy storage and transfer within cells, may represent the metabolic fate of carbon flow from carbon fixation.
The measured fatty acid profile (Figure 6) indicates that the overall fatty acid content following glycine treatment was significantly higher than that of the control group, partially validating the direction of carbon flow, where excess carbon is converted into fatty acids for cellular storage. Fatty acids participate in multiple metabolic pathways and are closely linked to carbon, nitrogen, and energy metabolism. As a mixotrophic source with both carbon and nitrogen properties, glycine provides a carbon- and nitrogen-rich environment for algal cells. Fathy et al. [16] found that glycine promotes photosynthesis in Synechocystis sp. PAK13 and Chlorella variabilis DT025, inducing sugar production that supports fatty acid biosynthesis by providing intermediates such as acetyl-CoA, G3P, and the energy required for synthesis. Glycine may regulate carbon source allocation in microalgae, promoting the conversion of starch to lipids and pigments, thereby facilitating the synthesis of fatty acids and fucoxanthin, as also observed in Lu et al. [32]. As a mixotrophic source with dual carbon and nitrogen properties, glycine regulates carbon and nitrogen allocation within I. zhanjiangensis, promoting the metabolic conversion to fucoxanthin and achieving increased fucoxanthin production.
4. Materials and Methods
4.1. Algal Strains and Culture Conditions
The I. zhanjiangensis strain used in this study was obtained from the Laboratory of Marine College, Ningbo University. Algal cells were inoculated at an initial concentration of 1×106 cells·mL-1 into F/2 culture medium prepared with seawater, supplemented with varying concentrations of glycine (Gly) as an organic carbon source. The medium formulation adhered to DB3302/T 162-2018 (Technical Specifications for Large-Scale Cultivation of Marine Microalgal Feed). Cultures were maintained in an illuminated incubator under a light intensity of 55 μmol·m-2·s-1, a photoperiod of 12 h light : 12 h dark, and a temperature of 25°C. The initial algal cell concentration was 1×106 cells·mL-1, with a culture volume of 100 mL. Glycine was added at seven concentrations: 0 g/L, 2 g/L, 4 g/L, 8 g/L, 16 g/L, and 32 g/L, with 0 g/L serving as the control. Each treatment was conducted in triplicate. Cultures were manually shaken several times daily to prevent cell adhesion to the flask walls.
4.2. Determination of Cell Parameters
From 0 h, 3 mL of algal suspension was sampled every 24 h until the stationary phase was reached. The optical density at 680 nm was measured using a UV-5200 UV-Vis spectrophotometer (Shanghai Metash Instruments Co., Ltd.). Cell density of I. zhanjiangensis was calculated using the regression equation:
Y = 94.446 × X - 6.6741 (R2 = 0.9934)
Y: cell number (105 cells·mL-1); X: absorbance at 680 nm; R2: coefficient of determination.
4.3. Determination of Fucoxanthin Content and Yield
On day 7 of cultivation(the start of stationary phase), 5 mL of algal culture was collected from each flask and centrifuged at 6,000 rpm for 10 min. The supernatant was discarded, and the resulting pellet was resuspended in 2 mL of absolute ethanol. The suspension was mixed thoroughly and incubated in a 45 ℃ water bath in the dark for 1 h. Following incubation, the mixture was centrifuged again at 6,000 rpm for 10 min, and the supernatant was collected. The extraction was repeated once under identical conditions, and the two supernatants were combined. The absorbance of the pooled extract was measured at 445 nm using a spectrophotometer. Fucoxanthin content and yield were calculated based on the absorbance values according to the following equations. All treatments were performed in triplicate with three biological replicates.
The equation for calculating fucoxanthin content is as follows:
D = (1000 × A445 × N × V) / (100 × A1%1cm × Y)
D: fucoxanthin content (mg/105 cells); A445: absorbance of the extract at 445 nm; N: dilution factor; V: volume of the crude extract (mL); A1%1cm: theoretical absorbance of fucoxanthin at 1 g/L in a 1 cm cuvette (1600); Y: cell density (105 cells/mL).
Fucoxanthin yield (mg/L) was calculated as:
Yield = (1000 × D × Y) / V1
D: fucoxanthin content (mg/105 cells); Y: cell density (105 cells/mL); V1: volume of algal suspension used for centrifugation.
4.4. Determination of Fucoxanthin Content and Yield in Scaled-Up Cultures
Algal cells were inoculated at 1×105 cells·mL-1 into F/2 medium supplemented with glycine at the optimal concentration, determined from prior experiments. Cultures were maintained under the same conditions as in section 2.1, with an initial cell concentration of 1×106 cells·mL-1 and a volume of 1 L. The control group was cultured without glycine. Each treatment was performed in triplicate, and cultures were shaken daily to prevent cell adhesion. Cell density, fucoxanthin content, and yield were measured following the methods outlined in sections 2.2 and 2.3.
4.5. Effects of Glycine on Photosynthetic Parameters in I. zhanjiangensis
4.5.1. Determination of Chlorophyll Content
Take 10 ml from each bottle of algal culture that has grown to the stationary phase. Use a high-speed refrigerated centrifuge (Eppendorf, Germany) to centrifuge at 4°C, 4000 rpm for 10 min, discard the supernatant. Add an equal volume of methanol (10 ml) to the pellet, and incubate in the dark for 24 h. Then centrifuge again at 6000 rpm for 10 min, take the supernatant, and use a UV spectrophotometer to measure the absorbance at 663 nm and 645 nm. The concentrations of chlorophyll a, chlorophyll c, and total chlorophyll in the extract can be calculated using standard formulas:
Chla=12.7×A663-2.59×A645
Chlc= 22.9×A645-4.68×A663
ChlT=20.2×A645+8.02×A663
A663 = absorbance at 663 nm; A645 = absorbance at 645 nm.
4.5.2. Chlorophyll Fluorescence Parameters
On the 7th day post-glycine treatment, 2 mL of algal suspension was sampled from each treatment group. Chlorophyll fluorescence parameters were measured using AquaPen and FluorPen software, following the manufacturer’s instructions. This technique evaluates the photosynthetic efficiency of photosystem II (PSII) in microalgal cells. Light intensities at 540 nm were set to 10, 20, 50, 100, 300, 500, and 1000 μmol·m-2·s-1 for 60 s. An induction curve was measured with a 20 s delay before illumination (300 μmol·m-2·s-1), followed by saturation pulses (3000 μmol·m-2·s-1) to assess non-photochemical quenching (NPQ) and quantum yield. Saturation pulses were applied every 21 s for 2 s until photoinactivation occurred after 4 min. The minimal fluorescence (F0) was maintained between 200~500 au to ensure measurement accuracy. Following 15 minutes of dark adaptation, Fv/Fm, Y(II), NPQ, and qP were recorded.
4.6. Determination of ROS Content
Cells from cultures grown for 7 days under 0 g/L and 8 g/L Gly conditions (15 mL each) were harvested by low-temperature centrifugation at 500 × g for 10 min. The supernatant was discarded, and the pellet was washed and resuspended in phosphate-buffered saline (0.1 mol/L PBS, pH 7.2). The cell suspension was incubated with 10 μM 2’,7’-dichlorofluorescein diacetate (DCFH-DA) at 37℃ in the dark for 40 min, with gentle mixing every 5 min. After incubation, cells were washed with PBS and resuspended for fluorescence measurement. Fluorescence intensity was detected using a multifunctional microplate reader (SpectraMax iD3) at an excitation wavelength of 488 nm and emission wavelength of 525 nm. As values may vary across instruments, reactive oxygen species (ROS) levels were expressed in arbitrary units (AU).
4.7. Transcriptome Assay
After culturing I. zhanjiangensis with 8 g/L glycine for 24 h, 50 mL of algal suspension was centrifuged at 5000 rpm for 10 min at 4°C. The supernatant was discarded, and the pellet was resuspended in 1 mL PBS buffer, transferred to a 1.5 mL RNA-free centrifuge tube, and centrifuged again under the same conditions. After removing the supernatant, the tube containing the algal pellet was flash-frozen in liquid nitrogen for 5 min and stored at -80°C. Samples were labeled and shipped under low-temperature conditions to Suzhou Anshengda Company for sequencing. Differential gene expression analysis was conducted using DESeq2 (v1.6.3) from the Bioconductor package, with significance defined as a fold change ≥ 2 and q-value (FDR, padj) ≤ 0.05. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed using the clusterProfiler package in R (v3.4.3). Metabolic pathways related to fucoxanthin synthesis were investigated using KEGG mapper and clusterProfiler.
4.8. Esterification and Analysis of Fatty Acids
On the seventh day of cultivation, the algae culture was centrifuged to concentrate the cells, which were then vacuum-dried for 24 hours using a vacuum pump. From each sample, 35 mg of dried algae powder was taken and subjected to lipid extraction using a chloroform–methanol mixture (chloroform:methanol = 2:1, v/v). The resulting supernatant was collected and evaporated to dryness under a stream of nitrogen. For methylation, 1.5 mL of methanolic sulfuric acid solution (sulfuric acid:methanol = 5:95, v/v) was added to the residue and incubated in a water bath at 70 °C for 4 hours to convert fatty acids into fatty acid methyl esters (FAMEs). Following methylation, 1.5 mL of n-hexane and 1.5 mL of ultrapure water were added to the mixture, which was then vortexed thoroughly and allowed to stand for 30 minutes. The upper organic phase was carefully collected and filtered through a 0.22 μm organic membrane filter into a GC vial for subsequent analysis. FAMEs were analyzed using gas chromatography–mass spectrometry (GC-MS) on an Agilent 5977C GC/MSD system.
4.9. Data Statistics and Analysis
Three groups of experimental data were collected under identical conditions and analyzed in parallel. Statistical analysis was performed using Microsoft Excel 2019, and the standard deviation (SD) was calculated for each group. All error bars in the figures represent the mean±standard deviation (mean±SD), reflecting the variability among biological replicates. Statistical significance was assessed using SPSS Statistics 19 with one-way ANOVA and post-hoc LSD tests, with a significance level of P < 0.05.
5. Conclusions
This study investigated the effects of exogenous glycine on cell proliferation and fucoxanthin synthesis in I. zhanjiangensis. Fucoxanthin production reached its highest level under the treatment with 8 g/L glycine. Glycine enhances photosynthetic efficiency and reduces reactive oxygen species (ROS) levels, thereby minimizing oxidative damage to the photosynthetic apparatus. It upregulates genes encoding FCP subunits and ZEP, promoting fucoxanthin formation. Additionally, glycine strengthens the Calvin cycle, enabling greater fixation of inorganic carbon and production of carbohydrates, which supports biomass accumulation and cell proliferation. These combined effects ultimately maximize fucoxanthin yield.
Author Contributions
Conceptualization, Y.C. and Y.G.; methodology, Y.C. and Y.G.; validation, Y.C. and C.C.; formal analysis, Y.C. and C.C.; investigation, Y.C., C.C., X.W., S.S. and Z.W.; resources, Y.G. and H.L.; data curation, Y.C. and C.C.; writing—original draft preparation, Y.C. and C.C.; writing—review and editing, Y.C. and Y.G.; visualization, Y.C.; supervision, Y.G. and H.L.; project administration, Y.C. and Y.G.; funding acquisition, Y.G. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Crosswise tasks of Ningbo University (No. HX2022000115), the K.C.Wong Magna Fund in Ningbo University.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data will be made available on request.
Acknowledgments
This study was supported by the Crosswise tasks of Ningbo University (No. HX2022000115), the K.C.Wong Magna Fund in Ningbo University.
Conflicts of Interest
Hao Liu is employed by Institute of Bioengineering, Biotrans Technology Co., Ltd. and United New Drug Research and Development Center, Biotrans Technology Co., Ltd. All other authors declare that there are no potential conflicts of interest. Institute of Bioengineering, Biotrans Technology Co., Ltd. and United New Drug Research and Development Center, Biotrans Technology Co., Ltd. have no role in the study design, collection, analysis, interpretation of data, the writing of this article or the decision to submit it for publication.
Abbreviations
The following abbreviations are used in this manuscript:
| ACS | Acetyl-CoA Synthetase |
| BCH | β-Carotene Hydroxylase |
| CCM | Central Carbon Metabolism |
| CRTISO | Carotenoid Isomerase |
| DDE | Diadinoxanthin De-epoxidase |
| DEP | Diatoxanthin Epoxidase |
| DMAPP | Dimethylallyl Pyrophosphate |
| FCP | Fucoxanthin-Chlorophyll a/c-binding Protein |
| FNR | Ferredoxin–NADP+ Reductase |
| GAPDH | Glyceraldehyde 3-phosphate Dehydrogenase |
| GCS | Glycine Cleavage System |
| GLDC | Glycine Decarboxylase |
| GS | Glutamine Synthetase |
| IPP | Isopentenyl Pyrophosphate |
| LCYb | Lycopene β-Cyclase |
| Lhcr, Lhcf, Lhcq | Fucoxanthin-Chlorophyll a/c-binding Protein subfamilies |
| NSY | Neoxanthin Synthase |
| PDS | Phytoene Desaturase |
| PGK | Phosphoglycerate Kinase |
| PRK | Phosphoribulokinase |
| PYS | Phytoene Synthase |
| rbcL | Rubisco Large Subunit |
| TK | Transketolase |
| VDE | Violaxanthin De-epoxidase |
| ZDS | ζ-Carotene Desaturase |
| ZEP | Zeaxanthin Epoxidase |
References
- Peng, J.; Yuan, J.-P.; Wu, C.-F.; Wang, J.-H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Marine drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Gammone, M.A.; D’Orazio, N. Anti-obesity activity of the marine carotenoid fucoxanthin. Marine drugs 2015, 13, 2196–2214. [Google Scholar] [CrossRef]
- Miyashita, K. Function of marine carotenoids. Food factors for health promotion 2009, 61, 136–146. [Google Scholar]
- Nomura, T.; Kikuchi, M.; Kubodera, A.; Kawakami, Y. Proton-donative antioxidant activity of fucoxanthin with 1, 1-diphenyl-2-picrylhydrazyl (DPPH). IUBMB Life 1997, 42, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Wang, S.; Wang, X.; Wang, L.; Wang, Q.; He, Y. Analysis and Identification of Major Carotenoids in Isochrysis zhanjiangensis. Journal of Food Safety & Quality 2018, 9, 1901–1905. [Google Scholar]
- Hu, H.; Lü, S.; Liu, H. A New Species of the Genus Isochrysis (Isochrysidales)—Isochrysis zhanjiangensis sp. nov. and Observations on Its Ultrastructure. Acta Oceanologica Sinica 2007, 29, 111–119. [Google Scholar]
- Bo, Y.; Wang, S.; Ma, F.; Manyakhin, A.Y.; Zhang, G.; Li, X.; Zhou, C.; Ge, B.; Yan, X.; Ruan, R. The influence of spermidine on the build-up of fucoxanthin in Isochrysis sp. Acclimated to varying light intensities. Bioresource Technology 2023, 387, 129688. [Google Scholar] [CrossRef]
- Garcıa, M.C.; Mirón, A.S.; Sevilla, J.F.; Grima, E.M.; Camacho, F.G. Mixotrophic growth of the microalga Phaeodactylum tricornutum: influence of different nitrogen and organic carbon sources on productivity and biomass composition. Process Biochemistry 2005, 40, 297–305. [Google Scholar] [CrossRef]
- Ano, A.; Funahashi, H.; Nakao, K.; Nishizawa, Y. Effect of glycine on 5-aminolevulinic acid biosynthesis in heterotrophic culture of Chlorella regularis YA-603. Journal of bioscience and bioengineering 1999, 88, 57–60. [Google Scholar] [CrossRef]
- Guo, B.; Liu, B.; Yang, B.; Sun, P.; Lu, X.; Liu, J.; Chen, F. Screening of diatom strains and characterization of Cyclotella cryptica as a potential fucoxanthin producer. Marine Drugs 2016, 14, 125. [Google Scholar] [CrossRef]
- Lu, X.; Sun, H.; Zhao, W.; Cheng, K.-W.; Chen, F.; Liu, B. A hetero-photoautotrophic two-stage cultivation process for production of fucoxanthin by the marine diatom Nitzschia laevis. Marine Drugs 2018, 16, 219. [Google Scholar] [CrossRef]
- Bowden, N.A.; Sanders, J.P.; Bruins, M.E. Solubility of the proteinogenic α-amino acids in water, ethanol, and ethanol–water mixtures. Journal of Chemical & Engineering Data 2018, 63, 488–497. [Google Scholar]
- Neilson, A.; Lewin, R. The uptake and utilization of organic carbon by algae: an essay in comparative biochemistry. Phycologia 1974, 13, 227–264. [Google Scholar] [CrossRef]
- Huang, Z.; Duan, S.; Zhang, X.; Hu, Z. Effects of Organic Carbon Compounds on the Growth of Isochrysis zhanjiangensis. Ecological Sciences 2007, 26, 14–17. [Google Scholar]
- Berland, B.; Bonin, D.; Guerin-Ancey, O.; Antia, N. Concentration requirement of glycine as nitrogen source for supporting effective growth of certain marine microplanktonic algae. Marine biology 1979, 55, 83–92. [Google Scholar] [CrossRef]
- Fathy, W.A.; AbdElgawad, H.; Essawy, E.A.; Tawfik, E.; Abdelhameed, M.S.; Hammouda, O.; Korany, S.M.; Elsayed, K.N. Glycine differentially improved the growth and biochemical composition of Synechocystis sp. PAK13 and Chlorella variabilis DT025. Frontiers in Bioengineering and Biotechnology 2023, 11, 1161911. [Google Scholar] [CrossRef]
- Yang, X.; Feng, L.; Zhao, L.; Liu, X.; Hassani, D.; Huang, D. Effect of glycine nitrogen on lettuce growth under soilless culture: A metabolomics approach to identify the main changes occurred in plant primary and secondary metabolism. Journal of the Science of Food and Agriculture 2018, 98, 467–477. [Google Scholar] [CrossRef]
- Cao, X.; Zhong, C.; Zhu, L.; Zhang, J.; Sajid, H.; Wu, L.; Jin, Q. Glycine increases cold tolerance in rice via the regulation of N uptake, physiological characteristics, and photosynthesis. Plant Physiology and Biochemistry 2017, 112, 251–260. [Google Scholar] [CrossRef]
- Yang, N.; Wang, C.-L.; He, W.-P.; Qu, Y.-Z.; Li, Y.-S. Photosynthetic characteristics and effects of exogenous glycine of Chorispora bungeana under drought stress. Photosynthetica 2016, 54, 459–467. [Google Scholar] [CrossRef]
- Kang, T.; Wu, H.; Lu, B.; Luo, X.; Gong, C.; Bai, J. Low concentrations of glycine inhibit photorespiration and enhance the net rate of photosynthesis in Caragana korshinskii. Photosynthetica 2018, 56, 512–519. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Y.; Wang, H.; Huang, Y.; Hu, Y.; Zhao, Y.; Gong, Y. Identification of Potential Factors for the Promotion of Fucoxanthin Synthesis by Methyl Jasmonic Acid Treatment of Phaeodactylum tricornutum. Marine Drugs 2023, 22, 7. [Google Scholar] [CrossRef]
- Li, Y.; Cai, X.; Gu, W.; Wang, G. Transcriptome analysis of carotenoid biosynthesis in Dunaliella salina under red and blue light. Journal of Oceanology and Limnology 2020, 38, 177–185. [Google Scholar] [CrossRef]
- Pal, T.; Padhan, J.K.; Kumar, P.; Sood, H.; Chauhan, R.S. Comparative transcriptomics uncovers differences in photoautotrophic versus photoheterotrophic modes of nutrition in relation to secondary metabolites biosynthesis in Swertia chirayita. Molecular biology reports 2018, 45, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Muthuraj, M.; Palabhanvi, B.; Ghoshal, A.K.; Das, D. High cell density lipid rich cultivation of a novel microalgal isolate Chlorella sorokiniana FC6 IITG in a single-stage fed-batch mode under mixotrophic condition. Bioresource technology 2014, 170, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Muthuraj, M.; Kumar, V.; Palabhanvi, B.; Das, D. Evaluation of indigenous microalgal isolate Chlorella sp. FC2 IITG as a cell factory for biodiesel production and scale up in outdoor conditions. Journal of Industrial Microbiology and Biotechnology 2014, 41, 499–511. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Z.; Yan, S. Effects of various amino acids as organic nitrogen sources on the growth and biochemical composition of Chlorella pyrenoidosa. Bioresource technology 2015, 197, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Mujtaba, G.; Lee, K. Effects of nitrogen sources on cell growth and biochemical composition of marine chlorophyte Tetraselmis sp. for lipid production. Algae 2016, 31, 257–266. [Google Scholar] [CrossRef]
- Zhong, X.; Li, B.; Li, M.; Zhang, Z.; Peng, X. Research Progress on Photorespiration. Journal of Tropical and Subtropical Botany 2022, 30, 782–790. [Google Scholar]
- Woo, K.; Jokinen, M.; Canvin, D.T. Reduction of nitrate via a dicarboxylate shuttle in a reconstituted system of supernatant and mitochondria from spinach leaves. Plant Physiology 1980, 65, 433–436. [Google Scholar] [CrossRef]
- Chen, M.; Tang, H.; Ma, H.; Holland, T.C.; Ng, K.S.; Salley, S.O. Effect of nutrients on growth and lipid accumulation in the green algae Dunaliella tertiolecta. Bioresource technology 2011, 102, 1649–1655. [Google Scholar] [CrossRef]
- Kanamoto, A.; Kato, Y.; Yoshida, E.; Hasunuma, T.; Kondo, A. Development of a method for fucoxanthin production using the Haptophyte marine microalga Pavlova sp. OPMS 30543. Marine Biotechnology 2021, 23, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Liu, B.; He, Y.; Guo, B.; Sun, H.; Chen, F. Novel insights into mixotrophic cultivation of Nitzschia laevis for co-production of fucoxanthin and eicosapentaenoic acid. Bioresource Technology 2019, 294, 122145. [Google Scholar] [CrossRef]
- Premaratne, M.; Liyanaarachchi, V.C.; Nimarshana, P.; Ariyadasa, T.U.; Malik, A.; Attalage, R.A. Co-production of fucoxanthin, docosahexaenoic acid (DHA) and bioethanol from the marine microalga Tisochrysis lutea. Biochemical Engineering Journal 2021, 176, 108160. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Chen, L.; Cheng, W.; Liu, T. Combined production of fucoxanthin and EPA from two diatom strains Phaeodactylum tricornutum and Cylindrotheca fusiformis cultures. Bioprocess and biosystems engineering 2018, 41, 1061–1071. [Google Scholar] [CrossRef]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, characterization, and antioxidant activity of fucoxanthin from the marine diatom Odontella aurita. Marine drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef] [PubMed]
- Durnford, D.; Deane, J.; Tan, S.; McFadden, G.; Gantt, E.; Green, B. A phylogenetic assessment of the eukaryotic light-harvesting antenna proteins, with implications for plastid evolution. Journal of Molecular Evolution 1999, 48, 59–68. [Google Scholar] [CrossRef]
- Xia, S.; Gao, B.; Fu, J.; Xiong, J.; Zhang, C. Production of fucoxanthin, chrysolaminarin, and eicosapentaenoic acid by Odontella aurita under different nitrogen supply regimes. Journal of bioscience and bioengineering 2018, 126, 723–729. [Google Scholar] [CrossRef]
- Truong, T.Q.; Park, Y.J.; Winarto, J.; Huynh, P.K.; Moon, J.; Choi, Y.B.; Song, D.-G.; Koo, S.Y.; Kim, S.M. Understanding the Impact of Nitrogen Availability: A Limiting Factor for Enhancing Fucoxanthin Productivity in Microalgae Cultivation. Marine Drugs 2024, 22, 93. [Google Scholar] [CrossRef]
- Pajot, A.; Lavaud, J.; Carrier, G.; Garnier, M.; Saint-Jean, B.; Rabilloud, N.; Baroukh, C.; Bérard, J.-B.; Bernard, O.; Marchal, L. The fucoxanthin chlorophyll a/c-binding protein in tisochrysis lutea: influence of nitrogen and light on fucoxanthin and chlorophyll a/c-binding protein gene expression and fucoxanthin synthesis. Frontiers in Plant Science 2022, 13, 830069. [Google Scholar] [CrossRef] [PubMed]
- Ördög, V.; Stirk, W.A.; Bálint, P.; van Staden, J.; Lovász, C. Changes in lipid, protein and pigment concentrations in nitrogen-stressed Chlorella minutissima cultures. Journal of Applied Phycology 2012, 24, 907–914. [Google Scholar] [CrossRef]
- Berges, J.A.; Charlebois, D.O.; Mauzerall, D.C.; Falkowski, P.G. Differential effects of nitrogen limitation on photosynthetic efficiency of photosystems I and II in microalgae. Plant Physiology 1996, 110, 689–696. [Google Scholar] [CrossRef]
- Cakmak, T.; Angun, P.; Demiray, Y.E.; Ozkan, A.D.; Elibol, Z.; Tekinay, T. Differential effects of nitrogen and sulfur deprivation on growth and biodiesel feedstock production of Chlamydomonas reinhardtii. Biotechnology and bioengineering 2012, 109, 1947–1957. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Zhu, G.; Zhou, G.; Song, X.; Hussein Ibrahim, M.E.; Ibrahim Salih, E.G. Effect of N on growth, antioxidant capacity, and chlorophyll content of sorghum. Agronomy 2022, 12, 501. [Google Scholar] [CrossRef]
- Lam, S.K.; Han, X.; Lin, E.; Norton, R.; Chen, D. Does elevated atmospheric carbon dioxide concentration increase wheat nitrogen demand and recovery of nitrogen applied at stem elongation? Agriculture, ecosystems & environment 2012, 155, 142–146. [Google Scholar]
- Badran, E.G.; Abogadallah, G.M.; Nada, R.M.; Nemat Alla, M.M. Role of glycine in improving the ionic and ROS homeostasis during NaCl stress in wheat. Protoplasma 2015, 252, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Hong, H.; Li, W.-C.; Yang, L.; Huang, J.; Xiao, Y.-L.; Chen, X.-Y.; Chen, G.-Y. Downregulation of rubisco activity by non-enzymatic acetylation of RbcL. Molecular plant 2016, 9, 1018–1027. [Google Scholar] [CrossRef]
- Fu, X.; Walker, B.J. Photorespiratory glycine contributes to photosynthetic induction during low to high light transition. Scientific Reports 2024, 14, 19365. [Google Scholar] [CrossRef]
- Li, Y.; Sun, H.; Wang, Y.; Yang, S.; Wang, J.; Wu, T.; Lu, X.; Chu, Y.; Chen, F. Integrated metabolic tools reveal carbon alternative in Isochrysis zhangjiangensis for fucoxanthin improvement. Bioresource Technology 2022, 347, 126401. [Google Scholar] [CrossRef]
- Longworth, J.; Wu, D.; Huete-Ortega, M.; Wright, P.C.; Vaidyanathan, S. Proteome response of Phaeodactylum tricornutum, during lipid accumulation induced by nitrogen depletion. Algal research 2016, 18, 213–224. [Google Scholar] [CrossRef]
- Bai, Y.; Cao, T.; Dautermann, O.; Buschbeck, P.; Cantrell, M.B.; Chen, Y.; Lein, C.D.; Shi, X.; Ware, M.A.; Yang, F.; et al. Green diatom mutants reveal an intricate biosynthetic pathway of fucoxanthin. Proceedings of the National Academy of Sciences 2022, 119, e2203708119. [Google Scholar] [CrossRef]
- Dambek, M.; Eilers, U.; Breitenbach, J.; Steiger, S.; Büchel, C.; Sandmann, G. Biosynthesis of fucoxanthin and diadinoxanthin and function of initial pathway genes in Phaeodactylum tricornutum. Journal of Experimental Botany 2012, 63, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Keereetaweep, J.; Liu, H.; Xu, C.; Shanklin, J. The role of sugar signaling in regulating plant fatty acid synthesis. Frontiers in Plant Science 2021, 12, 643843. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of glycine on the growth of I. zhanjiangensis.
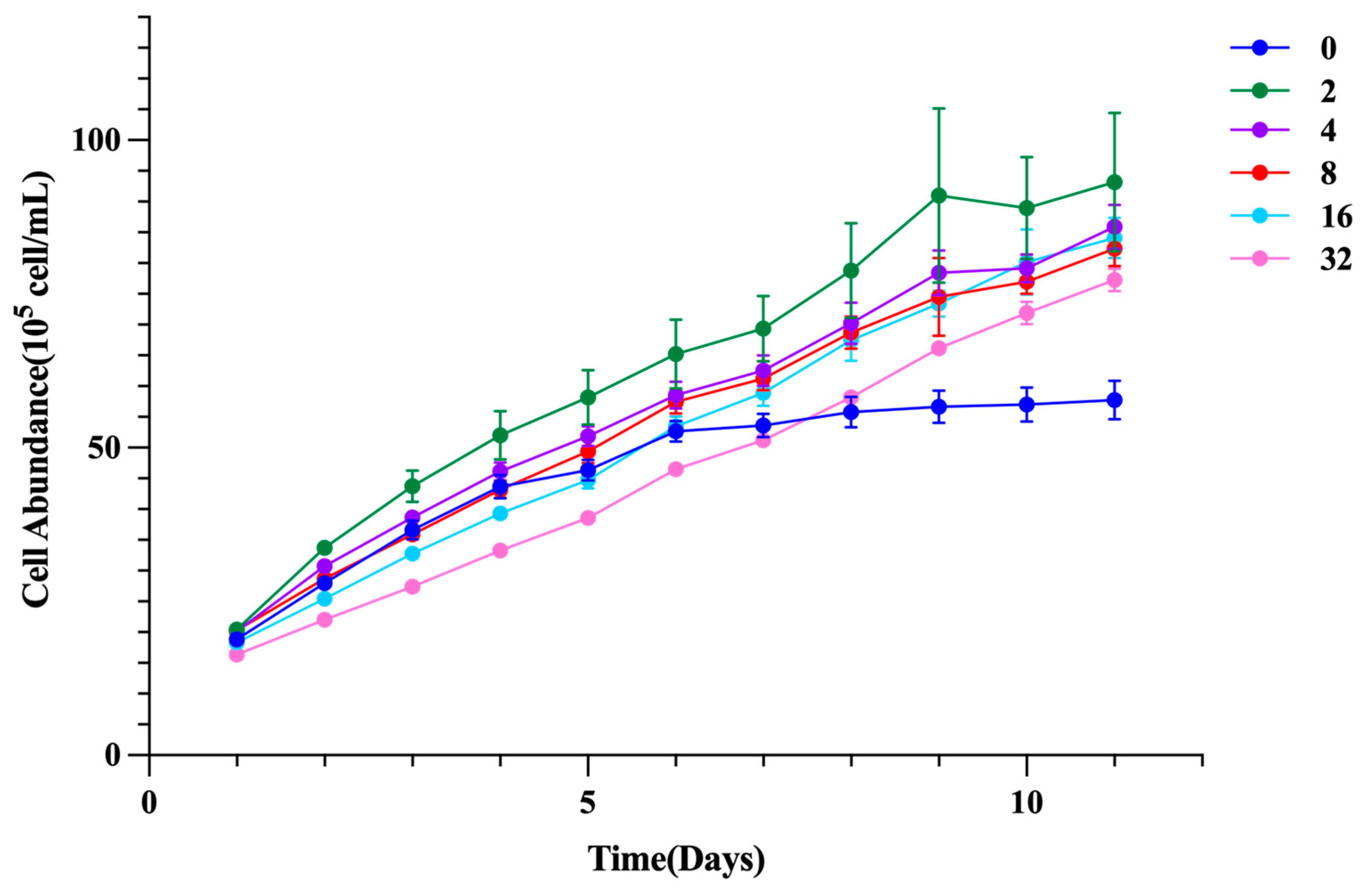
Figure 2.
Effects of glycine on fucoxanthin content and yield in I. zhanjiangensis. (A) Fucoxanthin content, (B) Fucoxanthin yield, (C) Fucoxanthin content in scaled-up cultures, (D) Fucoxanthin yield in scaled-up cultures.
Figure 2.
Effects of glycine on fucoxanthin content and yield in I. zhanjiangensis. (A) Fucoxanthin content, (B) Fucoxanthin yield, (C) Fucoxanthin content in scaled-up cultures, (D) Fucoxanthin yield in scaled-up cultures.
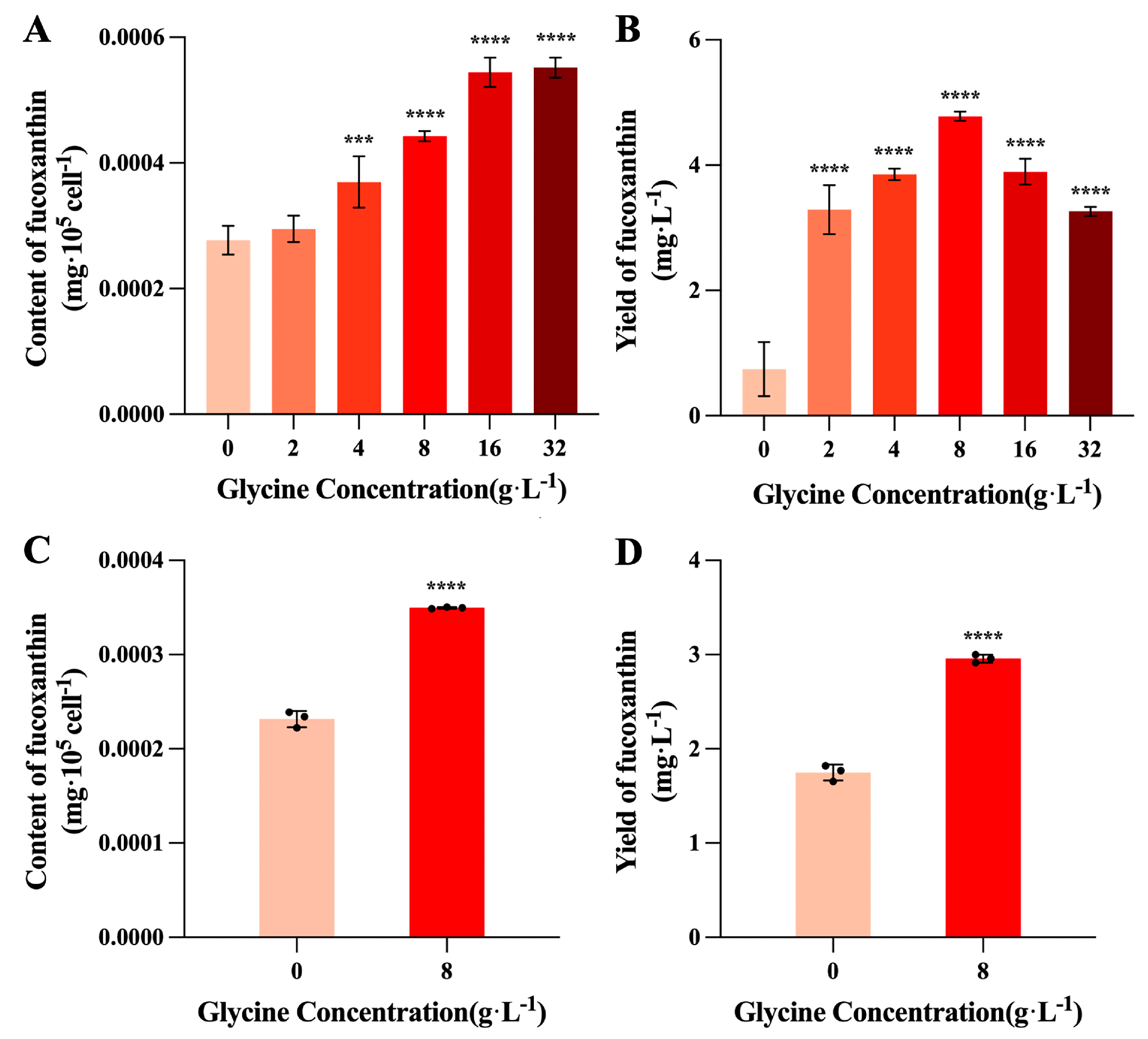
Figure 3.
Effects of glycine on chlorophyll content in I. zhanjiangensis.
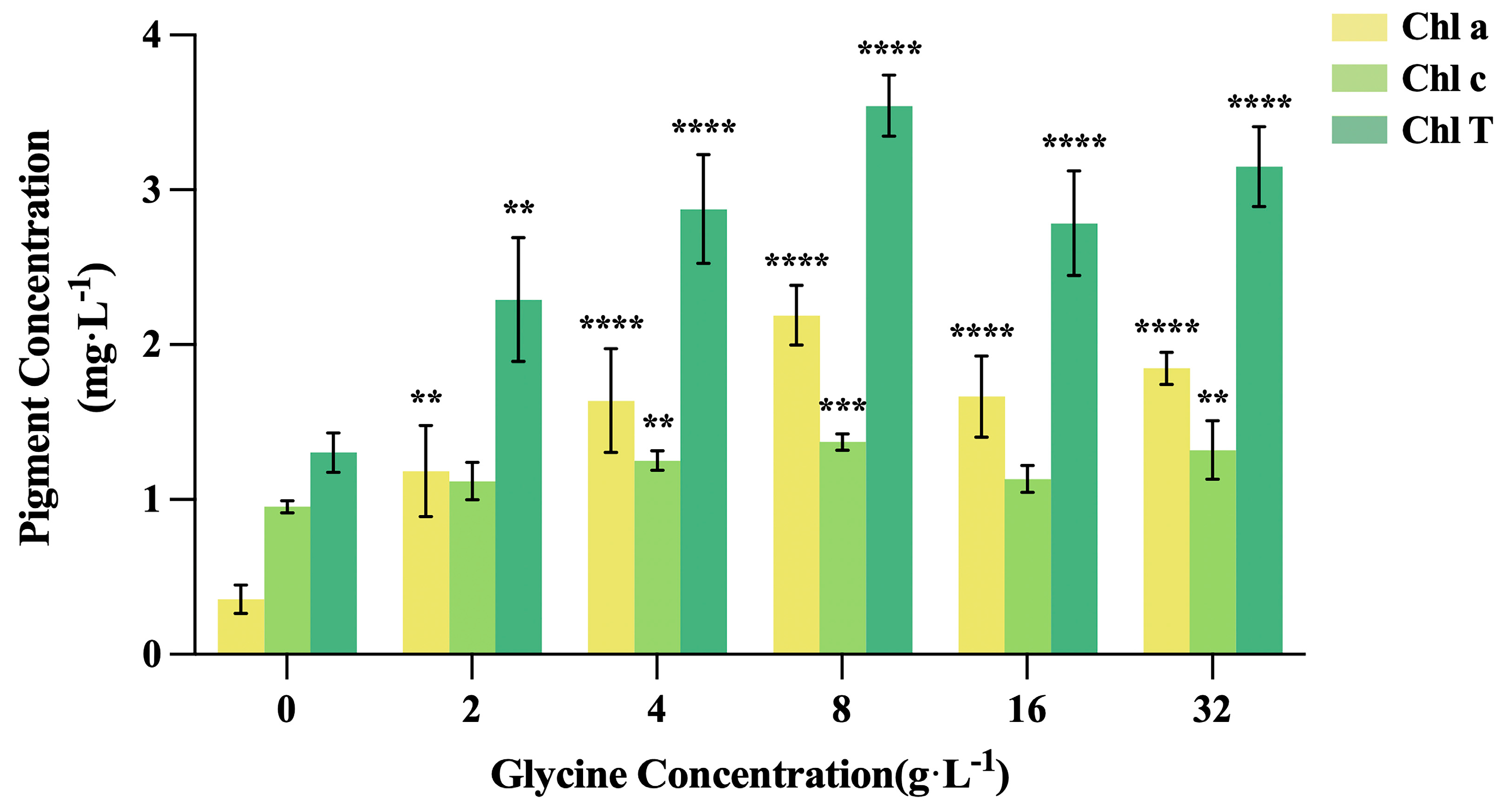
Figure 4.
Effects of glycine on chlorophyll fluorescence parameters in I. zhanjiangensis. (A) Fv/Fm, (B) Y(II), (C) qP, (D) NPQ, (E) ROS.
Figure 4.
Effects of glycine on chlorophyll fluorescence parameters in I. zhanjiangensis. (A) Fv/Fm, (B) Y(II), (C) qP, (D) NPQ, (E) ROS.
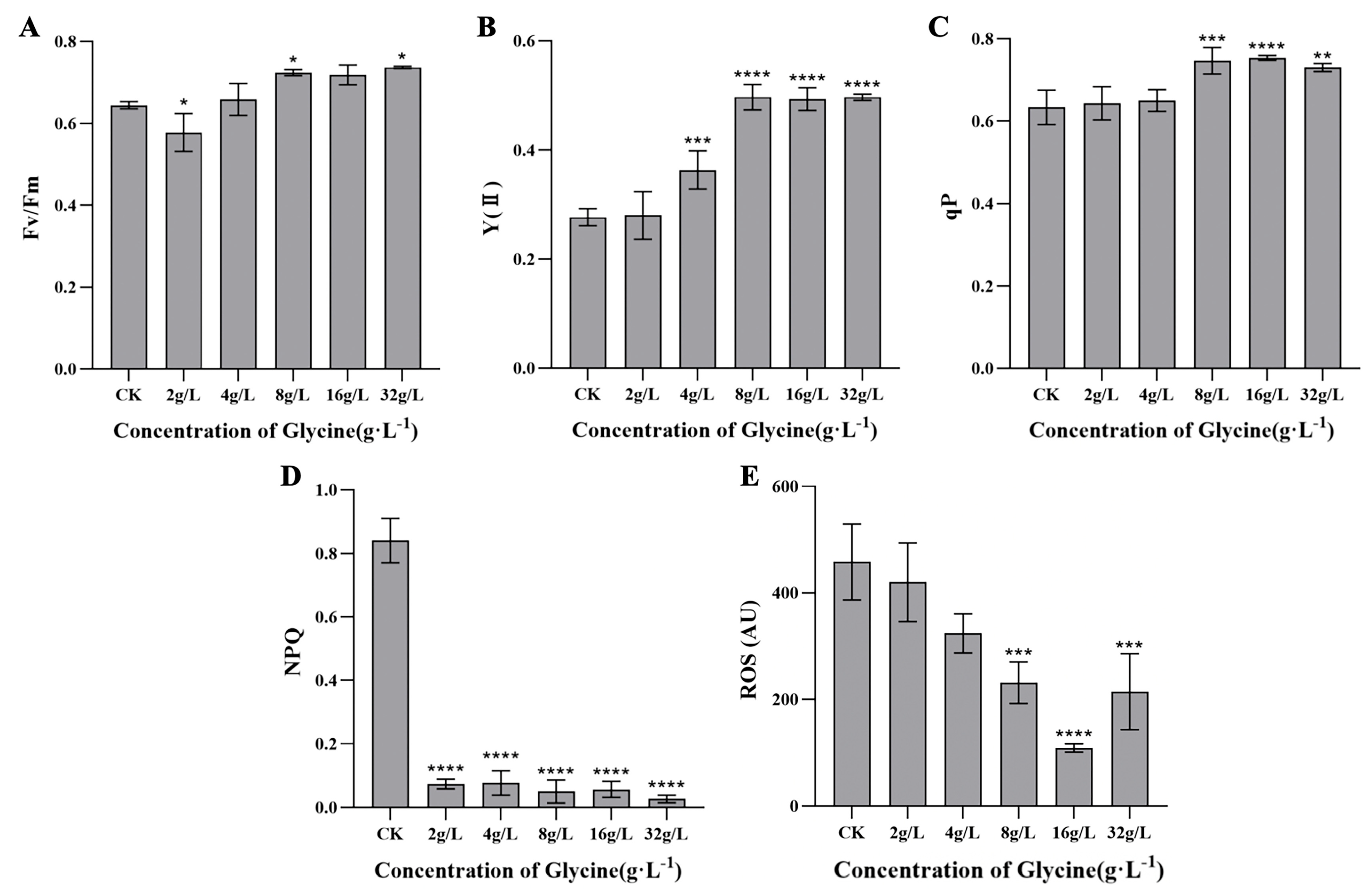
Figure 5.
Exogenous glycine influences cell proliferation and fucoxanthin synthesis in I. zhanjiangensis. Note: Transcriptome sequencing was conducted at 8 g/L glycine. Red indicates upregulated gene transcription; blue indicates downregulated gene transcription. Purple represents decreased content.
Figure 5.
Exogenous glycine influences cell proliferation and fucoxanthin synthesis in I. zhanjiangensis. Note: Transcriptome sequencing was conducted at 8 g/L glycine. Red indicates upregulated gene transcription; blue indicates downregulated gene transcription. Purple represents decreased content.
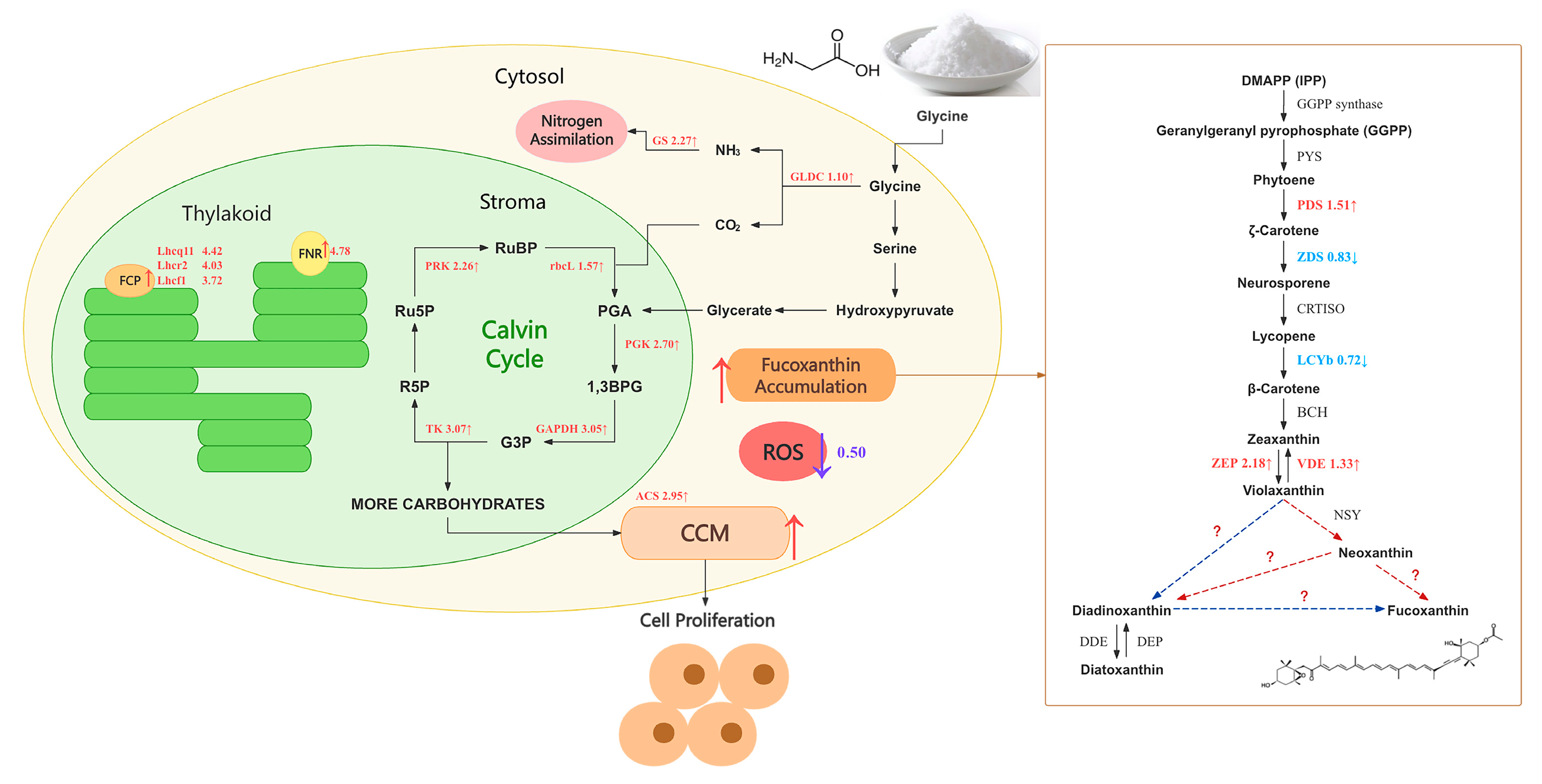
Figure 6.
Effects of glycine (8 g/L) on fatty acid content in I. zhanjiangensis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



