
Submitted:
17 November 2023
Posted:
20 November 2023
You are already at the latest version
Abstract
Terpenes are high-value chemicals which can be produced by engineered cyanobacteria from sustainable resources, solar energy, water and CO2. We previously reported that the euryhaline unicellular cyanobacteria Synechocystis PCC 6803 (S.6803) and Synechococcus PCC 7002 (S.7002) produce farnesene and limonene, respectively, more efficiently than other terpenes. In the present study, we attempted at enhancing farnesene production in S.6803 and limonene production in S.7002. Practically, we tested the influence of key cyanobacterial enzymes acting in carbon fixation (RubisCO, PRK, CcmK3 and CcmK4), utilization (CrtE, CrtR and CruF) and storage (PhaA and PhaB) on terpene production in S.6803, and we compared some of the findings with the data obtained in S.7002. We report that the overproduction of RubisCO from S.7002 and PRK from Cyanothece PCC 7425 increased farnesene production in S.6803, but not limonene production in S.7002. The overexpression of the crtE genes (synthesis or terpene precursors) from S.6803 or S.7002 did not increased farnesene production in S.6803. In contrast, the overexpression of the crtE gene from S.6803, but not S.7002, increased farnesene production in S.7002, emphasizing the physiological difference between these two model cyanobacteria. Furthermore, the deletion of the crtR and cruF genes (carotenoid synthesis) and phaAB genes (carbon storage) did not increase the production of farnesene in S.6803. Finally, as a containment strategy of genetically modified (GMO) strains of S.6803 we report that the deletion of the ccmK3K4 genes (carboxysome for CO2 fixation) did not affect the production of limonene but decreased the production of farnesene in S.6803.
Keywords:
Synechocystis PCC 6803
; Synechococcus PCC 7002
; farnesene
; limonene
; RubisCO
; phosphoribulokinase
; carboxysome
; geranylgeranyl pyrophosphate synthase
; β-carotene hydroxylase
; γ-carotene hydroxylase
1. Introduction
Terpenes constitute a large family of chemicals produced by plants in low quantities and consequently at high prices. They are used for the production of fragrances, pharmaceuticals, pesticides, solvents and potentially biofuels [1,2]. All terpenes (general formula (C5H8)n) derive from the five-carbons (C5) building blocks dimethylallyl pyrophosphate (DMAPP) and isopentenyl pyrophosphate (IPP). The head-to-tail covalent linkage of DMAPP and IPP catalyzed by the geranyl diphosphate synthase (GPPS) enzyme, generates the geranyl diphosphate (GPP) precursor of monoterpenes (C10H16), such as limonene. Then, the addition of another IPP unit on GPP catalyzed by the farnesyl diphosphate synthase (FPPS), forms the farnesyl pyrophosphate (FPP) precursor of sesquiterpenes (C15H24), such as farnesene (Figure 1).
Besides plants, which are better used for food, cyanobacteria, the robust photosynthetic prokaryotes colonizing our planet, can be used for the ecologically responsible production of terpenes from sunlight and CO2 (for reviews see [1,2]). Cyanobacteria fix CO2 using the Calvin-Benson-Bassham (CBB) pathway in which the key enzyme ribulose bisphosphate carboxylase/oxygenase (RubisCO) catalyzes the addition of one molecule of CO2 to one molecule of ribulose biphosphate produced by the other key enzyme phosphoribulokinase (PRK). In parallel, they have developed the carboxysome sub-cellular compartment (assembled from various shell proteins such as CcmK3 and CcmK4) to encapsulate RubisCO in a CO2-rich environment that favor its carbon-fixing (carboxylase) activity over its detrimental oxygenase activity (for reviews see [3,4]). A part of the photosynthetically-fixed carbon is used by the methylerythritol-4-phosphate (MEP) pathway to produce DMAPP and IPP (Figure 1), which are transformed by a geranylgeranyl pyrophosphate synthase (CrtE) into GPP, FPP, and GGPP which is used by the carotene hydroxylases (CrtR and CruF) to produce chlorophyll and carotenoids [1,2,5]. GPP and FPP can also be transformed into terpenes, following introduction and expression in cyanobacteria of synthetic terpene-synthase encoding genes adapted to the cyanobacterial codon usage [1,2]. The unicellular euryhaline cyanobacteria Synechocystis PCC 6803 (hereafter S.6803) and Synechococcus PCC 7002 (S.7002, costal organism) are interesting for this purpose. They have a good genetics and they grow well in sea and brackish waters to preserve freshwater resources for agriculture [6].
In the frame of our exploration of the ability of physiologically-different cyanobacteria for the photoproduction of chemically-different terpenes, we previously engineered several cyanobacteria by cloning codon-adapted version of terpene synthase-encoding genes, expressed from the strong lambda-phage pR-promoter, in a RSF1010-derived replicative plasmid or in a neutral chromosome site [7,8,9,10]. The results showed that S.6803 produces farnesene better than bisabolene, limonene, santalene and pinene [8], whereas S.7002 produces limonene more efficiently than bisabolene and pinene [10]. In the present study we used several genetic strategies to address the improvement of farnesene production in S.6803 and limonene production in S.7002.
2. Results and Discussion
2.1. The Overexpression of the RubisCO genes from Synechococcus PCC 7002 Increases Farnesene Production in Synechocystis PCC 6803
Previous studies showed that overexpressing the RubisCO-encoding genes can improve cell growth and/or the photosynthetic production of high value chemicals in S.6803 and/or S.7002 [11,12,13,14]. In this study, we attempted to improve farnesene production in S.6803 by introducing an extra-copy of the RubisCO-encoding genes, in the S.6803-ChrFS expressing the Picea abies α-farnesene synthase gene from a neutral site of its chromosome [8]. To increase the odd of improving RubisCO activity in S.6803, we used extra RubisCO genes from not only S.6803, but also physiologically-diverse cyanobacteria studied in our laboratory. These are namely: Cyanothece PCC 7425 (hereafter C.7425) and S.7002 where their RubisCO genes constitute the rbcLXS operon encoding the large (RbcL) and small (RbcS) RubisCO subunits, and the RbcX chaperon (See Cyanobase). We also used the rbcLS operon of Synechococcus elongatus PCC 7942 (S.7942) where rbcX is located away from rbcLS [15,16].
These RubisCO operons were PCR amplified from their respective cyanobacterial genomes (Figure S1), using oligonucleotides (Table S1) that introduced convenient restriction site upstream of the ATG start codon of rbcL (NdeI for S.6803, S.7002 and S.7942, and XhoI for C.7425) and downstream of the TAA stop codon of rbcS (EcoRI for S.6803, PvuII for both S.7002 and S.7942, and BspEI for C.7425). After restriction, the RubisCO operons were cloned downstream of the strong pR promoter [17] of the RSF1010-derived replicative pC plasmid vector [18] opened with the same enzymes. The resulting SmR/SpR plasmids, pCrbc6803, pCrbc7002, pCrbc7942 and pCrb7425 (Figure S1 and Table S2) and the (empty) pC control vector, were introduced by conjugation in the S.6803-ChrFS engineered strain that produces farnesene [8]. In each case, two independent SmR/SpR clones were selected and analyzed by PCR and DNA sequencing (Figure S1). The data showed that all these pC-derived plasmids, pCrbC7425, pCrbc7942, pCrbc6803 and pCrbc7002, replicate stably in S.6803 with no impact on cell growth, like the pC control vector (Figure 2). The resulting recombinant strains, which also have their indigenous chromosomal RubisCO genes, are represented in Figure S2.
The production of farnesene by the S.6803-chrFS strains propagating the RubisCO-producing plasmids pCrbc6803, pCrbc7002, pCrbc7425 and pCrbc7942 was measured over 21-days periods of photoautotrophic growths, as previously described [8]. GC-MS analysis of the terpene-trapping dodecane overlay samples from all tested strains showed a peak with similar retention time and ion chromatogram than a pure standard of α-farnesene. As compared to the S.6803-chrFS control strain, the level of farnesene production driven by the pCrbc6803, pCrbc7002, pCrbc7425 and pCrbc7942 plasmids showed that high-level expression of the RubisCO genes from S.7002, but not C.7425, S.6803 and S.7942, enhanced farnesene production (about 2 folds at day 21, Figure 2). This finding suggests that the RubisCO of S.7002 is more active in S.6803 than the other tested RubisCO, including the endogenous enzyme of S.6803.
2.2. The Overexpression of the Phosphoribulokinase Gene from Cyanothece PCC 7425 Increases Farnesene Production in Synechocystis PCC 6803
As the elimination of the CP12 negative regulator of the phosphoribulokinase (PRK) enzyme was shown to improve terpene production in S.6803 [9], we decided to overexpress the prk gene as an attempt to enhance farnesene production. To increase the odd of enhancing PRK activity, we decided to clone an extra copy of prk from not only S.6803, but also C.7425, S.7002 and S.7942, in the pC plasmid vector. The prk genes were PCR amplified from their respective cyanobacterial genome, using oligonucleotide primers (Table S1) that embedded their ATG start codon in a NdeI restriction site (CATATG), and introduced an EcoRI site downstream of their TAA (S.6803) or TAG (S.7002, S.7942 and C.7425) stop codons. After restriction with both NdeI and EcoRI, these genes were cloned in pC opened with the same enzymes, and transformed to E. coli. All attempts to clone the prk genes from S.6803, S.7002 and S.7942 in pC were unsuccessful, indicating that cyanobacterial PRKs are toxic to E. coli. Supporting this interpretation, previous cloning in E.coli of the prk gene from S.7942 [19] and S.6803 [20], for in vitro analysis of PRK, employed an inducible promoter system that decoupled E.coli growth from the production of PRK.
However, we were able to clone the prk gene from C.7425 downstream of the strong constitutive pR promoter of the pC plasmid (Table S2), suggesting that PRK7425 is somehow less active in E. coli than the other cyanobacterial PRKs presently tested. The resulting plasmid, pCprk7425 (Table S2), was introduced by conjugation in the S.6803-chrFS strain expressing the farnesene synthase gene from its chromosome [8]. A SmR/SpR clone (Figure S2) was analyzed to show that pCprk7425 does not alter the growth of S.6803 (Figure 3). The production of farnesene during the photoautotrophic growth of the S.6803-chrFS-pCprk7425 reporter strain (Figure S2) was analyzed for 21-days, and compared to that of the S.6803-chrFS control strain. The data showed that increasing the expression of prk enhances (about three-folds) the production of farnesene in S.6803 (Figure 3). This result is consistent with our previous report on CP12 [9] and the above finding that overexpressing RubisCO genes improved farnesene production in S.6803 (Figure 2). Collectively, these data indicate that increasing CO2 fixation enhances terpene production in S.6803.
2.3. The Overexpression of the Genes Encoding Rubisco and PRK Enzymes does not Increase Limonene Production in Synechococcus PCC 7002
We also attempted to improve limonene production in the previously described S.7002 strain, named S.7002-chrLS, which harbors the Mentha spicata 4S-limonene synthase gene in a neutral chromosome site [10]. Therefore, the pC-derived plasmids overexpressing the studied phosphoribulokinase gene (pCprk7425) or RubisCO operons (pCrbc6803, pCrbc7002, pCrbC7425 and pCrbc7942) were introduced by conjugation in S.7002-chrLS. The production of limonene of the resulting reporter strains (Figure S3) appeared to be similar to that of the S.7002-chrLS control strain (Figure 4). Collectively, the present findings show that the standard photoautotrophic growth of both S.6803 and S.7002 is not limited by the natural abundance of their indigenous RubisCO and PRK enzymes. Nevertheless, the production of terpene is limited by the natural abundance of the RubisCO and PRK in S.6803, not in S.7002. This difference reminds us that because a model cyanobacterium is essentially a model of itself, it is therefore important to study several cyanobacteria in parallel (in the same laboratory) to better understand their common and specific properties, and exploit this knowledge for biotechnological purposes.
2.4. The Overexpression of the Crte Genes from Synechocystis PCC 6803 or Synechococcus PCC 7002 Decreases the Production of Both Farnesene and Limonene in Synechocystis PCC 6803
One of the challenges in using cyanobacteria for terpene production is the competition for prenyl pyrophosphates (GPP, FPP and GGPP) between the synthesis of photosynthetic pigments and the intended terpene (Figure 1). As most cyanobacteria, such as S.6803 and S.7002, have a single GGPP synthase (named CrtE) for the sequential synthesis of GPP, FPP and GGPP [5,21,22,23], several authors overexpressed synthetic genes encoding FPPS (IspA) from E. coli [24,25] or GPPS from plants [26,27,28] to improve (2-3 folds) terpene production in S.6803 or S.7942 (S.7002 was not tested). These moderate improvements could result from these heterologous enzymes having a low activity and/or stability in cyanobacteria, an eventuality pleading in favor of testing cyanobacterial CrtE enzymes. In this context, the structural differences in the CrtE enzymes from S.7002 and S.6803 [5,23] prompted us to overexpress the crtE genes from S.6803 (hereafter crtES.6803) and S.7002 (crtES.7002) to attempt at enhancing terpene production in both S.6803 and S.7002. Therefore, the protein coding sequences of the crtES.6803 and crtES.7002 genes were PCR amplified from the genomes of S.6803 or S.7002, using specific oligonucleotides (Table S1) flanking them with a unique restriction site upstream of their start codon, and another unique restriction site behind their stop codon (Table S2). After restriction, these DNA fragments were cloned downstream the Picea abies α-farnesene synthase gene (FS) or Mentha spicata 4S-limonene synthase gene (LS) of our previously described pCFS and pCLS plasmids [7,8] opened with the same enzymes (XhoI and EcoRI for pCFS, and EcoRI and AclI for pCLS). The resulting plasmids (Table S2) pCFScrtE6803 and pCFScrtE7002 (expression of the FScrtE6803 or FScrtE7002 operons), and pCLScrtE6803 and pCLScrtE7002 (expression of the LScrtE6803 or LScrtE7002 operons) were introduced by conjugation in S.6803 (Figure S4) where they appeared to replicate stably (Figure S5). The resulting S.6803 reporter strains (Figure S4), which also expressed their indigenous crtE gene from their chromosome, grew similarly to the control strains propagating the pC, pCFS or pCLS control plasmids (Figure 5). The production of farnesene directed by the pCFScrtE6803 and pCFScrtE7002 plasmids was lower (about three-folds) than the level driven by the pCFS control plasmid (Figure 5). Similarly, the production of limonene directed by the pCLScrtE6803 and pCLScrtE7002 plasmids were lower (about six-folds and three-folds, respectively) than the level driven by the pCLS control plasmid (Figure 5). Collectively, these data indicate that the natural abundance of the CrtE enzyme is not limiting terpene production in S.6803. The present finding that an increased level of CrtE actually decreases terpene production suggests that the additional molecules of the GGPP metabolite synthesized by the extra molecules of CrtE might inhibit the MEP pathway and/or the studied terpene synthases, similarly to the feedback inhibitions caused by the accumulation of IPP [2]. In addition, the increased level of CrtE might negatively influence the hypothetical metabolic pairing of CrtE with the GGPP phosphatase [23], which could somehow participate to terpene synthesis.
2.5. The Overexpression of the Crte Gene from Synechocystis PCC 6803, but not Synechococcus PCC 7002 Increases Farnesene Production in Synechococcus PCC 7002
The expression of extra copy of the crtE gene on terpene production in S.7002 was tested as follows. The pCFScrtE6803 and pCFScrtE7002 plasmids overexpressing the operonic genes encoding the Picea abies α-farnesene synthase and the CrtE enzyme from either S.6803 or S.7002, were introduced by conjugation in S.7002, along with the pCFS control plasmid expressing only the farnesene synthase gene (Figure S6). All plasmids appeared to replicate stably (Figure S7) without affecting cells growth (Figure 5). As compared to pCFS, the pCFScrtE6803 and pCFScrtE7002 reporter plasmids increased and (slightly) decreased farnesene production in S.7002, respectively (Figure 6). These data suggest that the activity of the CrtE6803 and CrtE7002 enzymes might be different in S.7002.
2.6. The Deletion of PHB Synthesis Genes does not Increase the Photoproduction of Terpenes in Synechocystis PCC 6803
In S.6803, most of the photosynthetically-fixed carbon is used for the production of biomass and abundant carbon stores such as glycogen and polyhydroxybutyrates (PHB) [29,30], while only 5% of the carbon is allocated to the MEP pathway for the synthesis of photosynthetic pigments (carotenoids, chlorophyll and quinone) [1,31]. As an attempt to redirect the carbon flux toward the MEP pathway to enhance terpene production in S.6803, we deleted the two adjacent genes phaAB encoding the acetoacetyl-CoA thiolase (PhaA) and acetoacetyl-CoA reductase (PhaB) enzymes, which catalyze the first step in PHB synthesis [32,33,34]. Therefore, a DphaAB::KmR DNA cassette (Table S2) harboring a transcription-terminator less KmR marker in place of most of the phaA and phaB collinear genes (from codon 93 of phaA to codon 141 of phaB) was constructed, as follows (Figure S8). The two 300 bp chromosomal DNA regions flanking the phaA-phaB DNA region to be deleted were synthesized by TWIST Bioscience as a single DNA segment harboring an EcoRV restriction site in its middle where we cloned the KmR marker (a HincII segment from pUC4K) in the same orientation as the phaAB genes that it replaced (Figure S8). This ΔphaAB::KmR DNA cassette was then transformed to S.6803, where DNA recombination occurring in the homologous DNA platforms flanking the KmR gene integrated this marker in place of the phaAB gene, in all copies of the S.6803 chromosome (Figure S8). Then, the SmR/SpR pC-derived plasmids pCFS and pCLS [7,8] were introduced by conjugation in the ΔphaAB::KmR mutant (Figure S8 and (Figure S9). In each case, several SmR/SpR/KmR clones were studied. The data showed that the deletion of the PHB synthesis genes phaAB increase the photoproduction of neither farnesene nor limonene in S.6803 (Figure 7).
This approach of deleting PHB synthesis genes to redirect photosynthetically-fixed carbons toward terpene production could not be tested in S.7002 because it does not synthesize PHB [35].
2.7. The Deletion of the Carotenoid Synthesis Genes crtR and cruF Decreases the Production of Farnesene in Synechocystis PCC6803
Production of sesquiterpenes, such as farnesene, competes with the synthesis of photosynthetic pigments (chlorophyll and carotenoids) because they use the same precursor metabolite FPP [36] (Figure 1). To save FPP for a better farnesene production, we decided to decrease the consumption of the FPP-derived metabolite GGPP used for carotenoids synthesis (Figure 1). Therefore, we deleted the genes crtR (β-carotene hydroxylase) and cruF (γ-carotene hydroxylase) acting in GGPP-consuming synthesis of carotenoids. For crtR deletion, a DcrtR::KmR DNA cassette (Table S2) was constructed by replacing the first 82 codons of crtR by a transcription-terminator-less KmR marker, as follows. The two 300 bp chromosomal DNA regions flanking the first 82 codons of crtR to be deleted were synthesized by TWIST Bioscience as a single DNA segment harboring an EcoRV restriction site in its middle, where we cloned the transcription-terminator-less KmR marker (a HincII segment from the pUC4K plasmid) in the same orientation as crtR (Figure S10). The DcrtR::KmR DNA cassette was transformed to S.6803, where homologous DNA recombination replaced crtR by the KmR marker, in all copies of the S.6803 chromosome (Figure S10). The resulting DcrtR::KmR mutant grew as fit as the WT strain under standard light, showing that crtR is not essential for the photoautotrophic growth of S.6803 as previously observed [37,38].
Then, the SmR/SpR pCFS plasmid was introduced by conjugation in the DcrtR::KmR mutant (Figure S10), and the production of farnesene by two SmR/SpR/KmR clones was analyzed during photoautotrophic growth (Figure 8). The data showed that the deletion of crtR does not increase the photoproduction of farnesene in S.6803 (it was decreased), unlike what was expected.
For cruF deletion, a DcruF::KmR DNA cassette (Table S2) was constructed (Figure S12) by replacing an internal part of the cruF coding sequence (CS, from codon 100 to codon 271) by the same KmR marker. The two 300 bp DNA regions flanking the 171 codons of cruF to be deleted were synthesized by TWIST Bioscience as a single DNA segment harboring an EcoRV site in its middle, where we cloned the KmR marker in the same orientation as cruF. The DcruF::KmR DNA cassette was verified by PCR and nucleotide sequencing, and then transformed to S.6803 where homologous DNA recombination replaced the 171 codons of cruF by the KmR marker, in all copies of the S.6803 chromosome (Figure S12). The resulting DcruF::KmR mutant grew as fit as the WT strain under standard light showing, for the first time, that cruF is not crucial for the photoautotrophic growth of S.6803.
Then, the SmR/SpR pCFS plasmid was introduced by conjugation in the DcruF::KmR mutant (Figures S11 and S12), and the farnesene production of two independent SmR/SpR/KmR clones was analyzed (Figure 8). As reported above in the case of crtR, the deletion of cruF did not increase, but decreased, the photoproduction of farnesene in S.6803 (Figure 8).
Since the strategy of deleting the crtR and cruF genes did not increase the production of farnesene in S.6803 it was not tested in S.7002.
2.8. Deletion of Both the ccmK3 and ccmK4 Genes Encoding Carboxysome Shell Proteins does not Alter the Production of Limonene but Decreases the Production of Farnesene in Synechocystis PCC 6803
Besides the engineering of genetically-modified cyanobacterial organisms (GMO) for the photosynthetic production of high-value chemicals, it is important to consider strategies to limit accidental release of these GMO in natural environments. For this purpose, an interesting target is the carbon concentrating mechanism (CCM) that cyanobacteria use to grow in the low CO2 concentration of their natural aquatic biotopes. The CCM system uses the carboxysome sub-cellular compartment, assembled from various Ccm shell proteins, which encapsulates the RubisCO enzyme in a CO2-rich environment favoring its carbon-fixing (carboxylase) activity over its detrimental oxygenase activity (for reviews see [3,4]). As a biocontainment strategy, previous workers have deleted carboxysome genes of S.7002 and S.7942 GMO to impose a high-CO2 requirement phenotype (HCR) preventing them from escaping from their cultivation photobioreactors [39,40]. Such a HCR phenotype did not negatively impact L-lactate production in S.7002 [39], but it decreased (about two fold) farnesene production in S.7942 [40]. Since the HCR containment of GMO has not yet been tested in S.6803, we decided to delete the ccmK3 and ccmK4 adjacent genes encoding the CcmK3 and CcmK4 carboxysome shell proteins which assemble as CcmK3-K4 hetero-hexamers in both S.6803 [41] and S.7942 [42]. These proteins were shown to be required for cell growth at low CO2 in S.7942 [42,43], and for optimal photoautotrophic growth in S.6803 [44].
A DccmK3K4::KmR DNA cassette (Table S2) was constructed to delete both the ccmK3 and ccmK4 colinear genes (from the start of ccmK3 coding sequence (CS) to the stop codon of ccmK4). The 250 bp regions upstream the ccmK3 CS and downstream the ccmK4 CS were synthesized by TWIST Bioscience as a single DNA segment harboring a SwaI and a BamHI restriction sites where we cloned the KmR marker (using the same enzymes) in the same orientation as the ccmK3 and ccmK4 CS it replaced (Figure S13). The DccmK3K4::KmR grew as healthy as the WT strain in the Na2CO3-rich MM medium, but was unable to grow under atmospheric (low) CO2 level (MM0 medium), in agreement with some previous findings in S.7942 [42,43] and S.6803 [44].
Then, the SmR/SpR plasmids, pCFS and pCLS were introduced by conjugation in the DccmK3K4::KmR double mutant, the production of terpenes in the resulting reporter strains (Figure S13) was analyzed during photoautotrophic growth in the Na2CO3-rich MM medium. The data showed that the double deletion of ccmK3K4 did not alter the photoproduction of limonene (monoterpene, C10) in S.6803 (Figure 9), as expected. In contrast, the deletion of ccmK3K4 decreased the higher production of farnesene (sesquiterpene, C15). Interestingly, these data showed that the more carbon atoms is used to produce the observed quantity of the studied terpene (the production of farnesene (C15) is more important than that of limonene (C10)) the more the production is decreased by the DccmK3K4::KmR deletion.
3. Materials and Methods
3.1. Bacterial Strains and Growth Conditions
E. coli strains used for DNA manipulations (TOP10 and NEB10 beta, Table S1) or conjugative transfer to S.6803 and S.7002 (CM404, [17]) of pC-derived replicative plasmids (Table S2) were grown at 37°C (TOP10 and NEB10 beta) or 30°C (CM404), on LB medium containing selective antibiotics: ampicillin (Amp) 100 μg.mL-1, kanamycin (Km) 50 μg.mL-1, streptomycin (Sm) 25 μg.mL-1 or spectinomycin (Sp) 75 μg.mL-1.
S.6803 and S.7002 were grown under continuous white light (2500 lux; 31.25 μE.m-2.s-1) and agitation (140 rpm, Infors rotary shaker) at 30°C, in liquid mineral medium: MM, i.e. BG11 [45] enriched with 3.78 mM Na2CO3 for S.6803 [46], or A+ supplemented with B12 vitamin (4 μg.L-1) for S.7002 [47]. The terpene-producing strains were grown in the presence of the selective antibiotics: Km 50 μg.mL-1 for both S.6803 and S.7002, Sm 5 μg.mL-1 and Sp 5 μg.mL-1 for S.6803, and Sm 50 μg.mL-1 and Sp 50 μg.mL-1 for S.7002. Growth was monitored by regular measurements of optical density at 750 nm (OD750) with a spectrophotometer (Jenway 6700).
3.2. Genetic Manipulations and Gene Transfer Techniques
The studied cyanobacterial genes were amplified by polymerase chain reaction (PCR) from cyanobacterial DNA with specific oligonucleotide primers (Table S1) using Hot start Phusion polymerase (ThermoFisher Science). PCR products were digested with appropriate restriction enzymes and cloned either in the RSF1010-derived autonomously replicating pC vector [18] for high-level gene expression, or a commercial E.coli plasmid for gene deletion. All DNA constructions were verified by PCR and DNA sequencing (Mix2Seq Kit, Eurofins Genomics) using appropriate oligonucleotide primers (Table S1). The pC-derived plasmids (Table S2) were introduced in S.6803 and S.7002 by conjugation [8,10,48], using a 72h co-incubation of E. coli and cyanobacterial cells. The DNA deletion cassettes were introduced in S.6803 by transformation [49]. Cells were then plated on MM containing Km 50 μg.mL-1 or both Sp 5 μg.mL-1 and Sm 5 μg.mL-1 and solidified with 1% Bacto Agar (Difco).
3.3. Terpenes Collection, and Quantification by Gas Chromatography–Mass Spectrometry
S.6803 and S.7002 engineered strains were grown photoautotrophically in the presence of selective antibiotics, in 250 mL erlenmeyers containing 50 mL cell suspensions covered with 20% (vol/vol) dodecane overlay (analytical grade, Sigma-Aldrich) to trap terpenes. At time intervals, 300 μL of these dodecane overlays were collected, and 1 μL of these samples were injected into a GC-MS apparatus (Trace1300 (GC) + ISQ LT (MS), ThermoScientific) using split modes of 10:1 (limonene) or 5:1 (farnesene). Terpenes were quantified as we previously described [7,8,10].
4. Conclusions
In this study, we have tested various genetic strategies to attempt at enhancing farnesene production in S.6803 and limonene production in S.7002. We report that the overexpression of the genes encoding the CO2-fixing enzymes RubisCO and phosphoribulokinase increase the production of terpene in S.6803, not in S.7002. Furthermore, the overexpression of the crtE genes encoding the geranylgeranyl pyrophosphate synthase (synthesis or terpene precursors) from S.6803 or S.7002 enhance farnesene production in S.6803. In contrast, the overexpression of the crtE gene from S.6803, but not S.7002, increase farnesene production in S.7002. Collectively these results emphasize the physiological difference between the two model cyanobacteria S.6803 and S.7002, which are often used for biotechnological projects. These differences remind us that because a model cyanobacterium is essentially a model of itself, it is important to study several cyanobacteria in parallel (in the same laboratory) to better understand their common and specific properties, and exploit this knowledge for biotechnological purposes. We also showed that, the deletion of the crtR and cruF genes (carotenoid synthesis) and phaAB genes (carbon storage) did not increase the production of farnesene in S.6803. Finally, we have tested a containment strategy of genetically modified (GMO). We report that the deletion of the ccmK3K4 genes (carboxysome for CO2 fixation) did not impact the production of limonene but decreased the production of farnesene in S.6803. It is clear from these results and other data in the literature that we need to better understand the metabolism of cyanobacteria to engineer strains for an economically-viable photoproduction of terpenes.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Physical map of the pC-derived plasmids expressing the studied rbc operons. Figure S2: Schematic representation of the farnesene-producing Synechocystis PCC 6803 strains overexpressing CO2-fixation genes. Figure S3: Schematic representation of the limonene-producing Synechococcus PCC 7002 strains overexpressing CO2-fixation genes. Figure S4: Schematic representation of the Synechocystis PCC 6803 strains producing farnesene or limonene in the presence, or not, of an extra crtE gene from Synechocystis PCC 6803 or Synechococcus PCC 7002. Figure S5: PCR verification of the pCFS-crtE and the pCLS-crtE replicative plasmids in Synechocystis PCC 6803. Figure S6: Schematic representation of the Synechococcus PCC 7002 strains producing farnesene in the presence, or not, of an extra crtE gene from Synechococcus PCC 7002 or Synechocystis PCC 6803. Figure S7: PCR verification of the pCFS-CrtE and pCLS-CrtE replicative plasmids in Synechococcus PCC 7002. Figure S8: Construction and analysis of the DphaAB::KmR mutant and their derivative reporter strains propagating the pCFS or pCLS plasmids. Figure S9: Schematic representation of the Synechocystis PCC 6803 producing farnesene or limonene in presence or absence of the phaA-phaB genes. Figure S10: Construction and analysis of the DcrtR::KmR mutant and its derivative reporter strain propagating the pCFS plasmid. Figure S11: Schematic representation of the Synechocystis PCC 6803 producing farnesene in presence or absence of the crtR or cruF genes. Figure S12: Construction and analysis of the DcruF::KmR mutant and its derivative reporter strain propagating the pCFS plasmid. Figure S13: Construction and analysis of the DccmK3K4::KmR double mutant. Figure S14: Schematic representation of the Synechocystis PCC 6803 producing farnesene or limonene in presence or absence of the ccmK3-ccmK4 genes. Table S1: Characteristics of the oligonucleotide primers used in this study. Table S2: Characteristics of the bacterial strains and plasmids used in this study.
Author Contributions
Conceptualization, F.C. and C.C.-C.; methodology, F.C. and C.C.-C.; validation, M.V., C.C., V.B.-G., M.C., S.F. and L.F.G.-A.; formal analysis, M.V., C.C., V.B.-G., M.C. and S.F.; investigation, C.C., V.B.-G., M.V., M.C., S.F., C.C.-C. and F.C.; resources, F.C., C.C.-C. and L.F.G.-A.; writing—original draft preparation, F.C. and C.C.-C.; writing—review and editing, F.C., C.C.-C., M.V., C.C., V.B.-G. and L.F.G.-A.; supervision, F.C. and C.C.-C.; project administration, F.C. and C.C.-C.; funding acquisition, F.C., C.C.-C. and L.F.G.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by the CEA program Focus ECC and the ANR projects CalvinDesign (ANR-17-CE05-001) and SpaceHex SpaceHex (ANR-19-CE09-0032-01).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Frédéric Taran for access to GC-MS.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lin, P.C.; Pakrasi, H.B. Engineering cyanobacteria for production of terpenoids. Planta 2019, 249, 145–154. [Google Scholar] [CrossRef]
- Rautela, A.; Kumar, S. Engineering plant family TPS into cyanobacterial host for terpenoids production. Plant Cell Rep. 2022, 41, 1791–1803. [Google Scholar] [CrossRef]
- Veaudor, T.; Blanc-Garin, V.; Chenebault, C.; Diaz-Santos, E.; Sassi, J.F.; Cassier-Chauvat, C.; Chauvat, F. Recent advances in the photoautotrophic metabolism of cyanobacteria: Biotechnological implications. Life 2020, 10. [Google Scholar] [CrossRef]
- Kupriyanova, E.; Pronina, N.A.; Los, D.L. Adapting from Low to High: An Update to CO2-Concentrating Mechanisms of Cyanobacteria and Microalgae. Plants 2023, 12, 1569. [Google Scholar] [CrossRef]
- Feng, Y.; Morgan, R.M.L.; Fraser, P.D.; Hellgardt, K.; Nixon, P.J. Crystal Structure of Geranylgeranyl Pyrophosphate Synthase (CrtE) Involved in Cyanobacterial Terpenoid Biosynthesis. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Cassier-Chauvat, C.; Blanc-Garin, V.; Chauvat, F. Genetic, Genomics, and Responses to Stresses in Cyanobacteria: Biotechnological Implications. Genes 2021, 12, 500. [Google Scholar] [CrossRef]
- Chenebault, C.; Diaz-Santos, E.; Kammerscheit, X.; Görgen, S.; Ilioaia, C.; Streckaite, S.; Gall, A.; Robert, B.; Marcon, E.; Buisson, D.-A.; et al. A Genetic Toolbox for the New Model Cyanobacterium Cyanothece PCC 7425: A Case Study for the Photosynthetic Production of Limonene. Front. Microbiol. 2020, 11, 586–601. [Google Scholar] [CrossRef]
- Blanc-Garin, V.; Chenebault, C.; Diaz-Santos, E.; Vincent, M.; Sassi, J.F.; Cassier-Chauvat, C.; Chauvat, F. Exploring the potential of the model cyanobacterium Synechocystis PCC 6803 for the photosynthetic production of various high-value terpenes. Biotechnol. Biofuels 2022, 15. [Google Scholar] [CrossRef] [PubMed]
- Blanc-Garin, V.; Veaudor, T.; Sétif, P.; Gontero, B.; Lemaire, S.D.; Chauvat, F.; Cassier-Chauvat, C. First in vivo analysis of the regulatory protein CP12 of the model cyanobacterium Synechocystis PCC 6803: Biotechnological implications. Front. Plant Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Chenebault, C.; Blanc-Garin, V.; Vincent, M.; Diaz-Santos, E.; Goudet, A.; Cassier-Chauvat, C.; Chauvat, F. Exploring the Potential of the Model Cyanobacteria Synechococcus PCC 7002 and PCC 7942 for the Photoproduction of High-Value Terpenes. Biomolecules 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Higashide, W.; Liao, J.C. Direct photosynthetic recycling of carbon dioxide to isobutyraldehyde. Nat. Biotechnol. 2009, 27, 1177–1180. [Google Scholar] [CrossRef] [PubMed]
- Ruffing, A.M. Improved free fatty acid production in cyanobacteria with Synechococcus sp. PCC 7002 as host. Front. Bioeng. Biotechnol. 2014, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Englund, E.; Lindberg, P.; Lindblad, P. Engineered cyanobacteria with enhanced growth show increased ethanol production and higher biofuel to biomass ratio. Metab. Eng. 2018, 46, 51–59. [Google Scholar] [CrossRef]
- Tharasirivat, V.; Jantaro, S. Increased Biomass and Polyhydroxybutyrate Production by Synechocystis sp. PCC 6803 Overexpressing RuBisCO Genes. Int. J. Mol. Sci. 2023, 24, 6415. [Google Scholar] [CrossRef]
- Emlyn-Jones, D.; Woodger, F.J.; Price, G.D.; Whitney, S.M. RbcX can function as a Rubisco chaperonin, but is non-essential in Synechococcus PCC7942. Plant Cell Physiol. 2006, 47, 1630–1640. [Google Scholar] [CrossRef]
- Bracher, A.; Whitney, S.M.; Hartl, F.U.; Hayer-Hartl, M. Biogenesis and Metabolic Maintenance of Rubisco. Annu. Rev. Plant Biol. 2017, 68, 29–60. [Google Scholar] [CrossRef]
- Mermet-Bouvier, P.; Chauvat, F. A conditional expression vector for the cyanobacteria Synechocystis sp. strains PCC6803 and PCC6714 or Synechococcus sp. strains PCC7942 and PCC6301. Curr. Microbiol. 1994, 28, 145–148. [Google Scholar] [CrossRef]
- Veaudor, T.; Cassier-Chauvat, C.; Chauvat, F. Overproduction of the cyanobacterial hydrogenase and selection of a mutant thriving on urea, as a possible step towards the future production of hydrogen coupled with water treatment. PLoS ONE 2018, 13, e0198836. [Google Scholar] [CrossRef]
- Kobayashi, D.; Tamoi, M.; Iwaki, T.; Shigeoka, S.; Wanado, A. Molecular characterization and redox regulation of phosphoribulokinase from the cyanobacterium Synechococcus sp. PCC 7942. Plant Cell Physiol. 2003, 44, 269–276. [Google Scholar] [CrossRef]
- Nishiguchi, H.; Liao, J.; Shimizu, H.; Matsuda, F. Novel allosteric inhibition of phosphoribulokinase identified by ensemble kinetic modeling of Synechocystis sp. PCC 6803 metabolism. Metab. Eng. Commun. 2020, 11, e00153. [Google Scholar] [CrossRef] [PubMed]
- Ohto, C.; Ishida, C.; Nakane, H.; Nishiro, T.; Obata, S. A thermophilic cyanobacterium Synechococcus elongatus has three different Class I prenyltransferase genes. Plant Mol. Biol. 1999, 40, 307–321. [Google Scholar] [CrossRef]
- Rodrigues, J.S.; Lindberg, P. Metabolic engineering of Synechocystis sp. PCC 6803 for improved bisabolene production. Metab. Eng. Commun. 2021, 12, e00159. [Google Scholar] [CrossRef] [PubMed]
- Satta, A.; Esquirol, L.; Ebert, B.E.; Newman, J.; Peat, T.S.; Plan, M.; Schenk, G.; Vickers, C.E. Molecular characterization of cyanobacterial short-chain prenyltransferases and discovery of a novel GGPP phosphatase. FEBS J. 2022, 289, 6672–6693. [Google Scholar] [CrossRef]
- Valsami, E.A.; Psychogyiou, M.E.; Pateraki, A.; Chrysoulaki, E.; Melis, A.; Ghanotakis, D.F. Fusion constructs enhance heterologous β-phellandrene production in Synechocystis sp. PCC 6803. J. Appl. Phycol. 2020. [Google Scholar] [CrossRef]
- Choi, S.Y.; Lee, H.J.; Choi, J.; Kim, J.; Sim, S.J.; Um, Y.; Kim, Y.; Lee, T.S.; Keasling, J.D.; Woo, H.M. Photosynthetic conversion of CO2 to farnesyl diphosphate-derived phytochemicals (amorpha-4,11-diene and squalene) by engineered cyanobacteria. Biotechnol. Biofuels 2016, 9. [Google Scholar] [CrossRef]
- Lin, P.-C.; Saha, R.; Zhang, F.; Pakrasi, H.B. Metabolic engineering of the pentose phosphate pathway for enhanced limonene production in the cyanobacterium Synechocystis sp. PCC 6803. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Betterle, N.; Melis, A. Photosynthetic generation of heterologous terpenoids in cyanobacteria. Biotechnol. Bioeng. 2019, 116, 2041–2051. [Google Scholar] [CrossRef]
- Li, M.; Long, B.; Dai, S.Y.; Golden, J.W.; Wang, X.; Yuan, J. Altered Carbon Partitioning Enhances CO2 to Terpene Conversion in Cyanobacteria. BioDesign Res. 2022, 9897425. [Google Scholar] [CrossRef]
- Luan, G.; Zhang, S.; Wang, M.; Lu, X. Progress and perspective on cyanobacterial glycogen metabolism engineering. Biotechnol. Adv. 2019, 37, 771–786. [Google Scholar] [CrossRef]
- Koch, M.; Forchhammer, K. Polyhydroxybutyrate: A Useful Product of Chlorotic Cyanobacteria. Microbiol Physiol. 2021, 31, 67–77. [Google Scholar] [CrossRef]
- Melis, A. Carbon partitioning in photosynthesis. Curr. Opin. Chem. Biol. 2013, 17, 453–456. [Google Scholar] [CrossRef]
- Taroncher-Oldenbourg, G.; Nishina, K.; Stephanopulos, G. Identification and Analysis of the Polyhydroxyalkanoate-Specific β-Ketothiolase and Acetoacetyl Coenzyme A Reductase Genes in the Cyanobacterium Synechocystis sp. Strain PCC6803. Appl. Environ. Microbiol. 2000, 66, 4440–4448. [Google Scholar] [CrossRef]
- Khetkorn, W.; Incharoensakdi, A.; Lindblad, P.; Jantaro, S. Enhancement of poly-3-hydroxybutyrate production in Synechocystis sp. PCC 6803 by overexpression of its native biosynthetic genes. Bioresour. Technol. 2016, 214, 761–768. [Google Scholar] [CrossRef]
- Velmurugan, R.; Incharroensakdi, A. Disruption of Polyhydroxybutyrate Synthesis Redirects Carbon Flow towards Glycogen Synthesis in Synechocystis sp. PCC 6803 Overexpressing glgC/glgA. Plant Cell Physiol. 2018, 59, 2020–2029. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Y.; Bryant, D.A. Metabolic engineering of Synechococcus sp. PCC 7002 to produce poly-3-hydroxybutyrate and poly-3-hydroxybutyrate-co-4-hydroxybutyrate. Metab. Eng. 2015, 32, 174–183. [Google Scholar] [CrossRef]
- Pagels, F.; Vasconcelos, V.; Guedes, A.C. Carotenoids from Cyanobacteria: Biotechnological Potential and Optimization Strategies. Biomolecules 2021, 11, 735. [Google Scholar] [CrossRef]
- Lagarde, D.; Vermaas, W. The zeaxanthin biosynthesis enzyme beta-carotene hydroxylase is involved in myxoxanthophyll synthesis in Synechocystis sp. PCC 6803. FEBS Lett. 1999, 454, 247–251. [Google Scholar] [CrossRef]
- Schafer, L.; Vioque, A.; Sandmann, G. Functional in situ evaluation of photosynthesis-protecting carotenoids in mutants of the cyanobacterium Synechocystis PCC6803. J. Photochem. Photobiol. B Biol. 2005, 78, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.L.; Gordon, G.C.; Bennett, N.R.; Lyu, H.; Root, T.W.; Pfleger, B.F. High-CO2 Requirement as a Mechanism for the Containment of Genetically Modified Cyanobacteria. ACS Synth. Biol. 2018, 7, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Choi, J., II; Woo, H.M. Biocontainment of Engineered Synechococcus elongatus PCC 7942 for Photosynthetic Production of α-Farnesene from CO2. J. Agric. Food Chem. 2021, 69, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alles, L.; Root, K.; Maveyraud, L.; Aubry, N.; Lesniewska, E.; Mourey, L.; Zenobi, R.; Truan, G. Occurrence and stability of hetero-hexamer associations formed by β-carboxysome CcmK shell components. PLoS ONE 2019, 14, e0223877. [Google Scholar] [CrossRef]
- Sommer, M.; Sutter, M.; Gupta, S.; Kirst, H.; Turmo, A.; Lechno-Yossef, S.; Burton, R.L.; Saechao, C.; Sloan, N.B.; Cheng, X.; et al. Heterohexamers formed by CcmK3 and CcmK4 increase the complexity of beta carboxysome shells. Plant Physiol. 2019, 179, 156–167. [Google Scholar] [CrossRef]
- Rae, B.D.; Long, B.M.; Badger, M.R.; Price, G.D. Structural determinants of the outer shell of β-carboxysomes in Synechococcus elongatus PCC 7942: Roles for CcmK2, K3-K4, CcmO, and CcmL. PLoS ONE 2012, 7, e43871. [Google Scholar] [CrossRef]
- Zhang, S.; Laborde, S.M.; Frankel, L.K.; Bricker, T.M. Four Novel Genes Required for Optimal Photoautotrophic Growth of the Cyanobacterium Synechocystis sp. Strain PCC 6803 Identified by In Vitro Transposon Mutagenesis. J. Bacteriol. 2004, 186, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef] [PubMed]
- Domain, F.; Houot, L.; Chauvat, F.; Cassier-Chauvat, C. Function and regulation of the cyanobacterial genes lexA, recA and ruvB: LexA is critical to the survival of cells facing inorganic carbon starvation. Mol. Microbiol. 2004, 53, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Stevens, S.E.; Pat Paterson, C.O.; Myers, J. The production of hydrogen peroxide by blue-green algae: A survey. J. Phycol. 1973, 9, 427–430. [Google Scholar] [CrossRef]
- Mermet-Bouvier, P.; Cassier-Chauvat, C.; Marraccini, P.; Chauvat, F. Transfer and replication of RSF1010-derived plasmids in several cyanobacteria of the genera Synechocystis and Synechococcus. Curr. Microbiol. 1993, 27, 323–327. [Google Scholar] [CrossRef]
- Labarre, J.; Chauvat, F.; Thuriaux, P. Insertional mutagenesis by random cloning of antibiotic resistance genes into the genome of the cyanobacterium Synechocystis strain PCC 6803. J. Bacteriol. 1989, 171, 3449–3457. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the metabolic pathway and key compounds involved in the synthesis of terpenes from CO2. Abbreviations. CA carbonic anhydrase; CBB: Calvin Benson Bassham cycle (consuming 1 NADPH, 1 ATP); CrtE: geranylgeranyl pyrophosphate synthase; DMAPP: dimethylallyl pyrophosphate; DXP: 1-deoxy-D-xylulose-5-phosphate; Fed: ferredoxin; FPP: farnesyl pyrophosphate; FS: farnesene synthase; GAPDH: glyceraldehyde-3P-dehydrogenase; GPP: geranyl pyrophosphate; GPP: geranyl pyrophosphate; GGPP: geranylgeranyl pyrophosphate; G3P: glyceraldehyde-3-phosphate; 3PGA: 3-phosphoglycerate; IPP: isopentenyl pyrophosphate; IDI: isopentenyl-diphosphate isomerase; LS: limonene synthase; MEP: methylerythritol 4-phosphate (it consumes 2 NADPH, 1 ATP & 1 CTP); phaA: acetyl-CoA acetyltransferase; phaB: Acetoacetyl-CoA reductase; phaC: PHB synthase; PRK: phosphoribulokinase; Rbc: ribulose biphosphate carboxylase; RuBP: ribulose-1,5-bisphosphate.
Figure 1.
Schematic representation of the metabolic pathway and key compounds involved in the synthesis of terpenes from CO2. Abbreviations. CA carbonic anhydrase; CBB: Calvin Benson Bassham cycle (consuming 1 NADPH, 1 ATP); CrtE: geranylgeranyl pyrophosphate synthase; DMAPP: dimethylallyl pyrophosphate; DXP: 1-deoxy-D-xylulose-5-phosphate; Fed: ferredoxin; FPP: farnesyl pyrophosphate; FS: farnesene synthase; GAPDH: glyceraldehyde-3P-dehydrogenase; GPP: geranyl pyrophosphate; GPP: geranyl pyrophosphate; GGPP: geranylgeranyl pyrophosphate; G3P: glyceraldehyde-3-phosphate; 3PGA: 3-phosphoglycerate; IPP: isopentenyl pyrophosphate; IDI: isopentenyl-diphosphate isomerase; LS: limonene synthase; MEP: methylerythritol 4-phosphate (it consumes 2 NADPH, 1 ATP & 1 CTP); phaA: acetyl-CoA acetyltransferase; phaB: Acetoacetyl-CoA reductase; phaC: PHB synthase; PRK: phosphoribulokinase; Rbc: ribulose biphosphate carboxylase; RuBP: ribulose-1,5-bisphosphate.
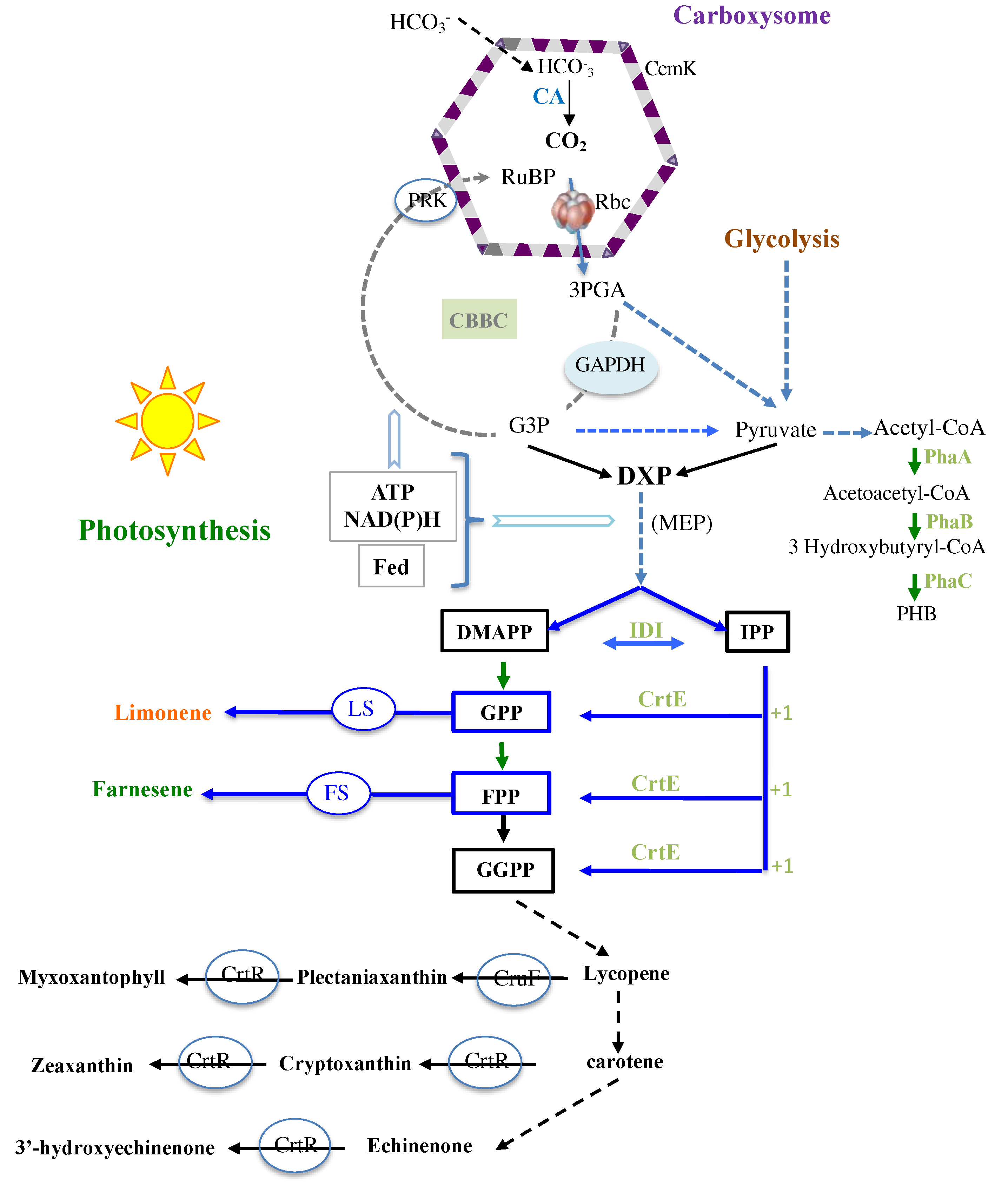
Figure 2.
Simultaneous analysis of growth (A) and farnesene production (B) of the Synechocystis PCC 6803 strains harboring the strongly-expressed genes encoding farnesene-synthase (FS) in a neutral chromosomal site (chrFS) and the RubisCO (rbc) genes, in a pC-derived replicative plasmid (pCrbc). These plasmids encode the RubisCO enzymes from the distantly-related unicellular cyanobacteria Cyanothece PCC 7425 (pCrbc7425), Synechococcus PCC 7002 (pCrbc7002), Synechococcus PCC 7942 (pCrbc7942) and Synechocystis PCC 6803 (pCrbc6803). Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 21 days. Error bars represent standard deviation from biological triplicates.
Figure 2.
Simultaneous analysis of growth (A) and farnesene production (B) of the Synechocystis PCC 6803 strains harboring the strongly-expressed genes encoding farnesene-synthase (FS) in a neutral chromosomal site (chrFS) and the RubisCO (rbc) genes, in a pC-derived replicative plasmid (pCrbc). These plasmids encode the RubisCO enzymes from the distantly-related unicellular cyanobacteria Cyanothece PCC 7425 (pCrbc7425), Synechococcus PCC 7002 (pCrbc7002), Synechococcus PCC 7942 (pCrbc7942) and Synechocystis PCC 6803 (pCrbc6803). Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 21 days. Error bars represent standard deviation from biological triplicates.
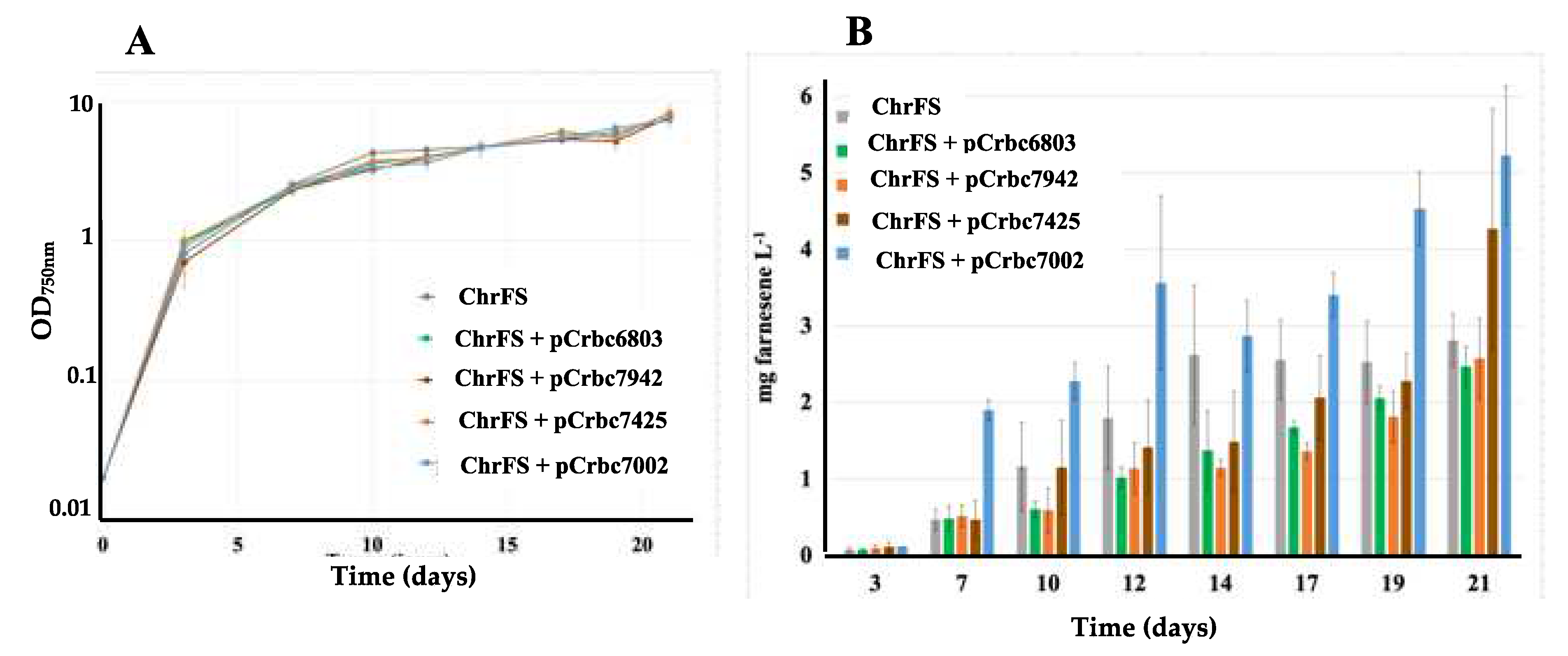
Figure 3.
Simultaneous analysis of growth (A) and farnesene production (B) of the Synechocystis PCC 6803 strain harboring the strongly expressed genes encoding farnesene synthase (FS) and phoshoribulokinase, in a neutral chromosomal site (chrFS) and a pC-derived replicative plasmid, respectively. This pCprk7425 plasmid encodes the PRK enzyme from Cyanothece PCC 7425, a unicellular cyanobacterium distantly related to both Synechocystis PCC 6803 and Synechococcus PCC 7002. Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 21 days. Error bars represent standard deviation from biological triplicates.
Figure 3.
Simultaneous analysis of growth (A) and farnesene production (B) of the Synechocystis PCC 6803 strain harboring the strongly expressed genes encoding farnesene synthase (FS) and phoshoribulokinase, in a neutral chromosomal site (chrFS) and a pC-derived replicative plasmid, respectively. This pCprk7425 plasmid encodes the PRK enzyme from Cyanothece PCC 7425, a unicellular cyanobacterium distantly related to both Synechocystis PCC 6803 and Synechococcus PCC 7002. Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 21 days. Error bars represent standard deviation from biological triplicates.
Figure 4.
Simultaneous analysis of growth (A) and limonene production (B) of the Synechococcus PCC 7002 strains harboring the strongly-expressed limonene-synthase gene (LS) in a neutral chromosomal site (chrLS) and a pC-derived plasmid expressing either the phoshoribulokinase gene from Cyanothece PCC 7425 (pCprk7425) or the RubisCO genes from Cyanothece PCC 7425 (pCrbc7425), Synechococcus PCC 7002 (pCrbc7002), Synechococcus PCC 7942 (pCrbc7942) or Synechocystis PCC 6803 (pCrbc6803). Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 21 days. Error bars represent standard deviation from biological triplicates.
Figure 4.
Simultaneous analysis of growth (A) and limonene production (B) of the Synechococcus PCC 7002 strains harboring the strongly-expressed limonene-synthase gene (LS) in a neutral chromosomal site (chrLS) and a pC-derived plasmid expressing either the phoshoribulokinase gene from Cyanothece PCC 7425 (pCprk7425) or the RubisCO genes from Cyanothece PCC 7425 (pCrbc7425), Synechococcus PCC 7002 (pCrbc7002), Synechococcus PCC 7942 (pCrbc7942) or Synechocystis PCC 6803 (pCrbc6803). Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 21 days. Error bars represent standard deviation from biological triplicates.
Figure 5.
Simultaneous analysis of growth (A,C) and terpene production (B,D) of Synechocystis PCC 6803 strains harboring a pC-derived plasmid strongly expressing either the farnesene synthase gene (pCFS; A,B) alone or transcriptionally fused to the crtE gene from either Synechocystis PCC 6803 (pCFScrtE6803) or Synechococcus PCC 7002 (pCFScrtE7002), or the limonene synthase gene (pCLS; C,D) alone or transcriptionally fused to a crtE gene (pCLScrtE6803 or pCLScrtE7002). Several clones were studied, as indicated by C1-C8. Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 21 days. Error bars represent standard deviation from biological triplicates.
Figure 5.
Simultaneous analysis of growth (A,C) and terpene production (B,D) of Synechocystis PCC 6803 strains harboring a pC-derived plasmid strongly expressing either the farnesene synthase gene (pCFS; A,B) alone or transcriptionally fused to the crtE gene from either Synechocystis PCC 6803 (pCFScrtE6803) or Synechococcus PCC 7002 (pCFScrtE7002), or the limonene synthase gene (pCLS; C,D) alone or transcriptionally fused to a crtE gene (pCLScrtE6803 or pCLScrtE7002). Several clones were studied, as indicated by C1-C8. Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 21 days. Error bars represent standard deviation from biological triplicates.
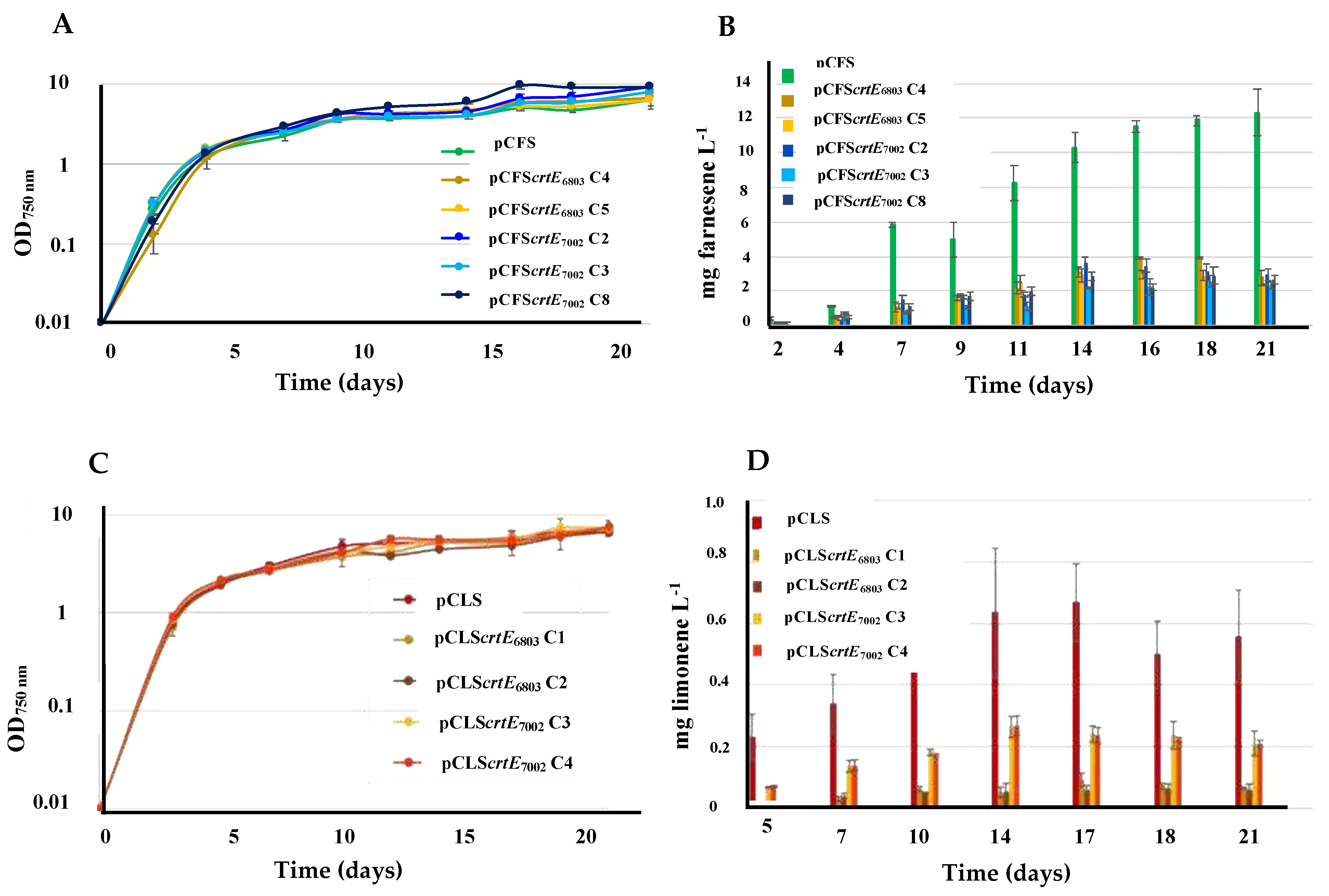
Figure 6.
Simultaneous analysis of growth (A) and farnesene production (B) of Synechococcus PCC 7002 strains harboring a pC-derived plasmid strongly expressing either the farnesene synthase gene (pCFS) alone or transcriptionally fused to the crtE gene from either Synechocystis PCC 6803 (pCFScrtE6803) or Synechococcus PCC 7002 (pCFScrtE7002). Three clones were studied, as indicated by C3, C6 and C9. Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 21 days. Error bars represent standard deviation from biological triplicates.
Figure 6.
Simultaneous analysis of growth (A) and farnesene production (B) of Synechococcus PCC 7002 strains harboring a pC-derived plasmid strongly expressing either the farnesene synthase gene (pCFS) alone or transcriptionally fused to the crtE gene from either Synechocystis PCC 6803 (pCFScrtE6803) or Synechococcus PCC 7002 (pCFScrtE7002). Three clones were studied, as indicated by C3, C6 and C9. Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 21 days. Error bars represent standard deviation from biological triplicates.
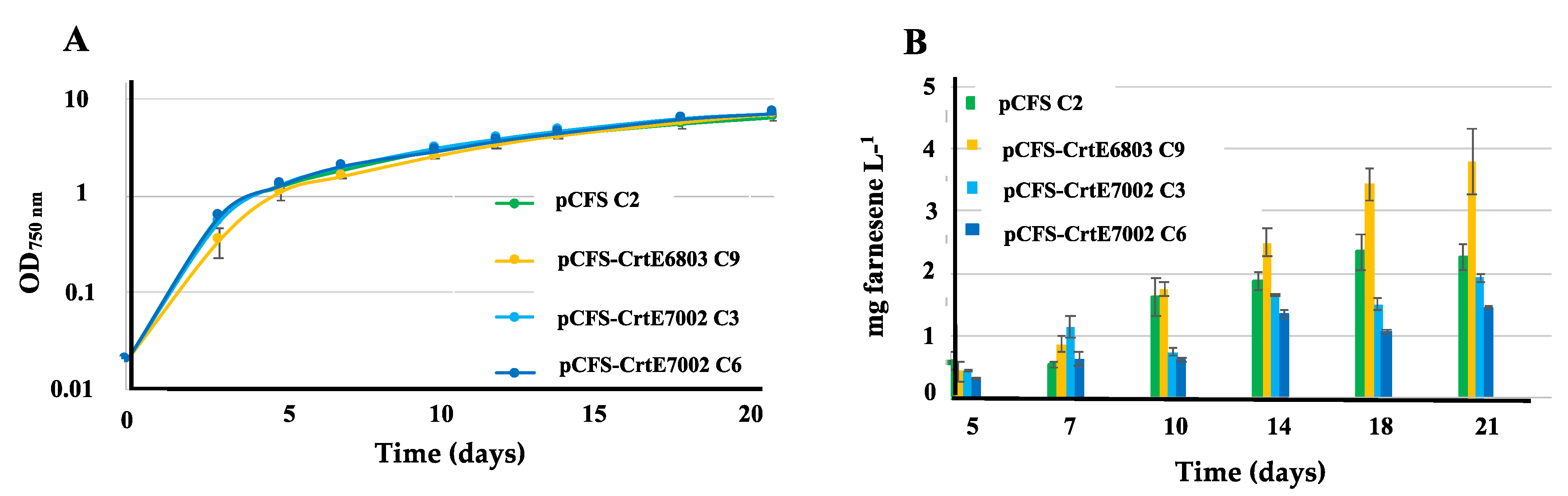
Figure 7.
Simultaneous analysis of growth (A,C) and terpene production (B,D) of Synechocystis PCC 6803 strains WT or DphaAB mutant harboring a pC-derived plasmid strongly expressing either the limonene synthase gene (pCLS; A,B) or the farnesene synthase gene (pCFS; C,D). Several clones were studied, as indicated by C1-C5. Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 17 days. Error bars represent standard deviation from biological triplicates.
Figure 7.
Simultaneous analysis of growth (A,C) and terpene production (B,D) of Synechocystis PCC 6803 strains WT or DphaAB mutant harboring a pC-derived plasmid strongly expressing either the limonene synthase gene (pCLS; A,B) or the farnesene synthase gene (pCFS; C,D). Several clones were studied, as indicated by C1-C5. Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 17 days. Error bars represent standard deviation from biological triplicates.
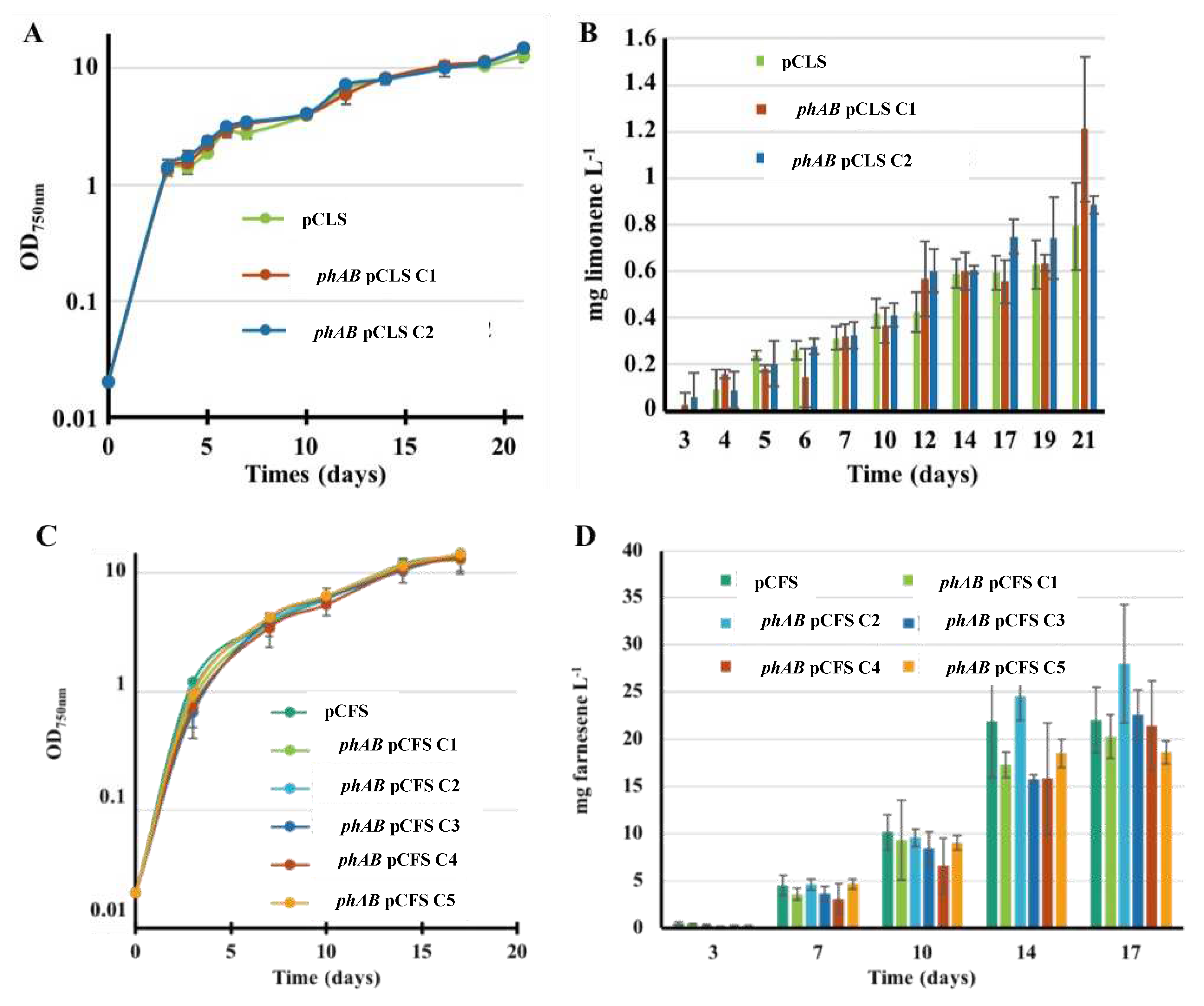
Figure 8.
Simultaneous analysis of growth (A) and farnesene production (B) of Synechocystis PCC 6803 WT strain and two clones (1 and 2) of each of the DcrtR and DcruF mutants harboring the farnesene synthase production pCFS plasmid. Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay farnesene production (measured at day 10). Error bars represent standard deviation from biological triplicates.
Figure 8.
Simultaneous analysis of growth (A) and farnesene production (B) of Synechocystis PCC 6803 WT strain and two clones (1 and 2) of each of the DcrtR and DcruF mutants harboring the farnesene synthase production pCFS plasmid. Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay farnesene production (measured at day 10). Error bars represent standard deviation from biological triplicates.
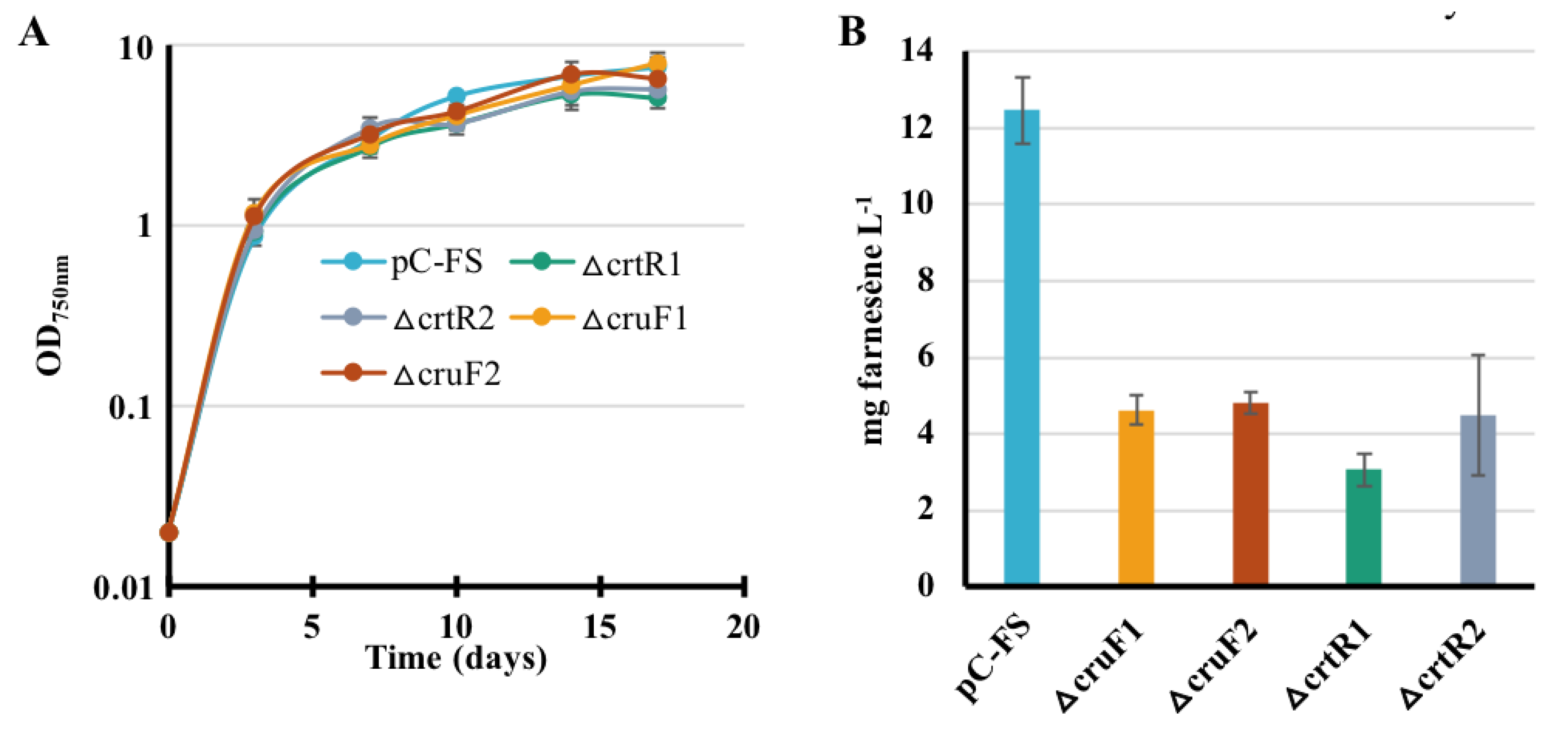
Figure 9.
Simultaneous analysis of growth (A,C) and terpene production (B,D) of Synechocystis PCC 6803 strains WT or DccmK3K4 mutant harboring a pC-derived plasmid strongly expressing either the limonene synthase gene (pCLS; A,B) or the farnesene synthase gene (pCFS; C,D). Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 17 or 21 days. Error bars represent standard deviation from biological triplicates.
Figure 9.
Simultaneous analysis of growth (A,C) and terpene production (B,D) of Synechocystis PCC 6803 strains WT or DccmK3K4 mutant harboring a pC-derived plasmid strongly expressing either the limonene synthase gene (pCLS; A,B) or the farnesene synthase gene (pCFS; C,D). Cells were grown under standard photoautotrophic conditions in the presence of a dodecane overlay (20% vol/vol) to assay terpene production for 17 or 21 days. Error bars represent standard deviation from biological triplicates.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



