
Submitted:
26 July 2025
Posted:
28 July 2025
You are already at the latest version
Abstract
Increasing dietary nitrate (NO3-) through beetroot juice (BRJ) supplementation elicits acute ergogenic benefits. However, it is unknown whether chronic NO3- supplementation can enhance resistance training (RT) adaptations in middle-aged and older individuals. Therefore, we sought to determine whether 12 weeks of combined RT and NO3- supplementation enhanced hypertrophic, vascular, strength, and skeletal muscle angiogenesis adaptations in this population. Twenty-eight apparently healthy, untrained men and women (56±7 years old, 29.1±5.3 kg/m2 body mass index) completed 12 weeks of supervised full-body RT (2x/week) while ingesting either BRJ (140 mL daily, providing 800 mg NO3-; n=14 with 7M/7W) or NO3--depleted BRJ placebo (PLA; n=14 with 7M/7W). Participants underwent a whole-body dual-energy x-ray absorptiometry scan, right mid-thigh ultrasonography for muscle imaging, right leg popliteal artery flow-mediated dilation (FMD) assessments, a biopsy of the right mid-thigh vastus lateralis, and strength testing prior to and following the 12-week intervention. Biopsy analyses included a NO3-/nitrite (NOx) fluorometric assay, immunoblotting for proteins involved in angiogenesis, and immunohistochemistry to quantify fiber type-specific capillaries and cross-sectional areas. Muscle NOx values did not significantly change: +15.4% in BRJ (p=0.073) and +7.8% (p=0.514) in PLA. Both groups significantly improved measures related to muscle hypertrophy, strength, and FMD. However, no significant group × time interactions were observed for whole-body lean mass, mid-thigh muscle cross-sectional area, popliteal artery FMD outcomes, or histological or molecular markers. In conclusion, BRJ supplementation does not enhance RT adaptations in middle-aged and older adults.
Keywords:
dietary nitrates
; resistance training
; older adults
; skeletal muscle
1. Introduction
Maintaining or increasing skeletal muscle mass is imperative among middle-aged and older adults for reducing the risk of life-threatening injuries from falls, cardiometabolic and cardiovascular disease, and, ultimately, enhancing their quality of life [1,2,3]. Although resistance training (RT) is a viable strategy to maintain or increase skeletal muscle mass, RT interventions among middle-aged and older adults produce inconsistent or attenuated hypertrophic responses, especially when compared to younger adults [4,5,6,7,8]. This phenomenon may be explained by anabolic resistance, which is the impaired ability of skeletal muscle to grow in response to a typically anabolic stimulus such as ingesting dietary protein and/or RT, and which adults may begin to experience around the fifth decade of life [2,9,10]. Age-induced anabolic resistance has been studied extensively [9,11,12,13], and many lines of research have indicated that this issue is primarily due to dampened skeletal muscle protein synthesis.
Aging is associated with progressive declines in macrovascular (large conduit arteries) and microvascular (arterioles and capillaries) function [9]. Specifically, reductions in macrovascular function begin around the fifth decade of life [14]. There is evidence suggesting that reduced nutrient delivery and blood flow may contribute to anabolic resistance [15,16]. For instance, a negative correlation has been reported between arterial stiffness and skeletal muscle mass among middle-aged and older adults [17]. Similarly, reduced endothelial function is associated with lower grip strength in older women [18]. At the microvascular level, decreased skeletal muscle capillarization is associated with attenuated hypertrophic responses to RT in older adults [19,20]. At the molecular level, exercise-induced alterations in vascular endothelial growth factor (VEGF), which mechanistically triggers angiogenesis, are diminished in older compared to younger adults [21,22,23,24]. These findings highlight the need for interventions that can improve vascular function among aging populations to enhance skeletal muscle hypertrophic response of RT.
Commercially available dietary supplements are commonly used to enhance health and exercise outcomes [25,26,27]. Dietary nitrate (NO3-) exhibits health benefits, particularly in relation to vascular function and blood flow [28,29,30,31,32]. While dietary NO3- can be ingested from various foods, it is commonly delivered as a supplement in concentrated beetroot juice (BRJ) [28,29,30,31,32]. When consumed as a pre-exercise supplement, certain studies report BRJ provides ergogenic benefits including enhanced muscular endurance, improved strength-endurance performance, and reduced fatigue [29,31,33,34,35]. The proposed mechanism involves the sequential reduction of ingested NO3- to nitrite (NO2-) by oral bacteria, followed by further reduction to nitric oxide (NO) in tissues, ultimately resulting in vascular smooth muscle relaxation, vasodilation, and enhanced blood flow [29,36,37,38]. Although dietary NO3- improves vascular health and acute exercise performance [31,32,33,39], only one study to our knowledge has examined how chronic NO3- supplementation during a period of RT affects functional outcomes [40]. Carter and colleagues assigned a cohort of postmenopausal women to either a BRJ + training group or training-only group whereby training consisted of 24 bouts of circuit style RT and cardiovascular training exercises over eight weeks. Functional tests were performed prior to the intervention and during week 9, after BRJ participants had ceased supplementation for one week. Interestingly, BRJ supplementation significantly improved distance covered during a 6-minute walk test greater than training alone, but neither group improved body composition. Additionally, while not measured, it was posited that type II myofiber adaptations with BRJ supplementation could lead to positive functional alterations given that NO3- has been posited to provide more benefit to these muscle fibers [28,41].
Thus, a mechanistic knowledge gap remains regarding whether NO3- supplementation can augment RT adaptations in middle-aged and older individuals. Evidence from older adults shows that greater skeletal muscle capillarization is associated with enhanced hypertrophic responses to RT, and some data indicate that NO3- may mechanistically enhance angiogenesis [42,43]. However, whether NO3- supplementation can enhance RT-induced angiogenesis in a middle-aged and older population has not been examined. Therefore, the purpose of this study was to determine whether NO3- supplementation via BRJ during a 12-week RT intervention enhances outcomes related to skeletal muscle hypertrophy, vascular function, myofiber characteristics, proteins involved in angiogenesis, and/or strength in middle-aged and older adults. Based on the literature reviewed above, we hypothesized that BRJ supplementation would enhance skeletal muscle hypertrophy and strength outcomes from RT compared to placebo supplementation, and that this would be accompanied by enhancements in vascular function and skeletal muscle angiogenesis.
2. Methods
2.1. Ethical Approval, Consent, and Experimental Design
Ethical approval and consent. This study was approved by the Auburn University Institutional Review Board (Protocol # 24-863 MR 2405) and adhered to the standards set by the latest revisions of the Declaration of Helsinki except for being pre-registered as a clinical trial. Thirty men and women between the ages of 40–70 years old were recruited from the local community. Potential participants were excluded if they: i) had prior exercise habits (>1 day/week) for one year prior to enrollment, ii) consumed supplemental BRJ (or other NO3-rich supplements), L-arginine, L-citrulline, or creatine-containing supplements one month prior to the initiation of the study, iii) had blood clotting issues precluding the collection of skeletal muscle biopsies, were allergic to supplement contents and/or lidocaine, or were diagnosed with cardio-metabolic or orthopedic issues precluding study participation. We aimed to enroll postmenopausal women due to vascular fluctuations that influence flow-mediated dilation, which occur throughout the menstrual cycle in premenopausal women [44,45,46]. Participants who met the inclusion criteria were informed of the study procedures and provided verbal and written consent to participate.
Overview of PRE/POST testing, intervention, and supplementation. The intervention consisted of 12 weeks of supervised full-body RE while supplementing daily with either BRJ in the form of commercially available Beet-It (n=14 participants; 140 mL concentrated providing 800 mg NO3-) or NO3--depleted BRJ PLA (n=14 participants; 140 mL providing less than 50 mg NO3-); both supplements (termed BRJ or PLA herein) were purchased from James White Drinks, Ipswich, UK. Prior to (PRE) and following the 12-week intervention (POST), participants underwent flow-mediated dilation (FMD) of the right leg popliteal artery, a full body dual-energy x-ray absorptiometry (DXA) scan, an ultrasound of the right mid-thigh for vastus lateralis (VL) muscle cross sectional area (mCSA) assessments, a biopsy of the right mid-thigh VL, and various measures of functional and strength testing. For logistical purposes, PRE testing assessments consisted of two days which were 1-5 days apart (day 1: FMD; day 2: DXA scan, mid-thigh ultrasound, VL biopsy, and strength testing). After completing all PRE testing, participants were allocated into the BRJ or PLA groups by a single investigator (ADF), who performed block randomization according to age, sex, and PRE DXA-derived whole-body lean tissue mass.
Approximately 2-3 days following day 2 of the PRE assessments, participants began the 12-week training intervention. During the first training day, participants were provided with a one-week supply of their respective supplement and were instructed to consume 140 mL 2.5 hours prior to workouts on exercise days and in the morning on non-exercise days, per previous recommendations [47,48]. Throughout the entirety of the study, participants were provided with weekly allocations of their respective supplements and adherence was documented based on verbal confirmation. Data collection and analysis (chiefly by MCM and DTB) was completed in a double-bind fashion throughout the entirety of the study.
The participants’ last training session was approximately 72-96 hours prior to POST testing and participants were instructed to consume their last supplement 24 hours prior to POST testing. POST testing included FMD, DXA scan, mid-thigh ultrasound, VL biopsy, and strength testing (within a two-hour period), identical to day 2 of PRE testing. Participants were asked to abstain from the use of alcohol-based mouthwash throughout the study to avoid altering oral microbiota that facilitate NO3- reduction to NO2- [49,50]. A schematic of the study design is presented in Figure 1 below, and more in-depth details about testing procedures follow.
2.2. PRE- and POST-Intervention Blood Pressure and Right Leg Popliteal Artery FMD
Twenty-four hours prior to PRE and POST FMD assessments, participants were instructed to avoid a list of high NO3- foods and caffeine intake [45,51,52]. Participants arrived at the School of Kinesiology at Auburn University following an overnight fast for FMD assessments (matched for time of day at PRE and POST ± 1 hour). After ~15-minute dark room supine rest on an athletic training table, blood pressure was obtained in triplicate using an automated oscillometer blood pressure monitor with the cuff placed on the right arm (OMRON model BP785N, Lake Forest, IL, USA). Participants were then repositioned in a prone position on an athletic training table and a high-resolution B-mode and doppler ultrasound (Logiq S7 R2 Expert; General Electric, Fairfield, CT USA) was used to assess right leg popliteal artery blood flow with a multifrequency linear-array transducer (10-12 MHz scanning frequency) for three minutes by a single investigator (DTB). Following baseline artery diameter and blood velocity readings, a pre-placed cuff (Rapid Cuff Inflation System, D.E. Hokanson, Inc., Bellevue, WA, USA) was rapidly inflated and remained inflated for five minutes to 225 mmHg (cuff location was on the proximal right calf, 2-4 cm below the popliteal fossa). Three minutes into occlusion the ultrasound probe was placed in the same position to record the arterial response prior to deflation and up to five minutes post-occlusion.
Image analysis was performed using specialized software (Cardiovascular Suite, QUIPU srl, Pisa, Italy) by a single investigator (DTB). Popliteal artery diameters were determined by measuring the distance between the near and far wall of the intima, and blood velocity was determined via selection of a region of interest around the Doppler waveform. Popliteal FMD (%) was expressed as a percentage increase from baseline diameter to peak diameter following cuff deflation. FMD measurements were also normalized to the mean shear rate calculated from the first 10 seconds following cuff deflation (normalized FMD expressed as FMD(%) ÷ sec-1).
2.3. Pre- and Post-Intervention DXA Scans for Body Composition
Participants arrived at our laboratory following an overnight fast for PRE and POST DXA scans. Prior to DXA scans, participants provided a ≥5ml urine sample that was assessed for urine specific gravity using a handheld refractometer (ATAGO; Bellevue, WA, USA). All participants’ urine specific gravity was <1.020, which was used as a threshold of sufficient hydration to continue testing. Height and body mass were measured using a digital column scale (Seca 769; Hanover, MD, USA). Participants were then asked to lie in a supine position for five minutes on the DXA scanner (Lunar Prodigy; GE Corporation, Fairfield, CT, USA) prior to the assessment. Following scans, whole-body lean tissue and fat mass were automatically segmented by the software and are reported in kilograms (kg). Estimated total body skeletal muscle mass was also calculated using appendicular lean mass per the equation provided by Kim et al. [53]. Though test-retest DXA scans were not performed on these participants, prior data from our laboratory on test-calibrate-immediate retest on 10 participants produced an intraclass correlation coefficient of 0.998 for total body lean tissue mass [54].
2.4. Pre- and Post-Intervention Right Mid-Thigh Ultrasonography Assessment
Following DXA scans, real-time B-mode ultrasonography (NextGen LOGIQe R8, GE Healthcare) with a multifrequency linear-array transducer (L4-12T, 4–12 MHz, GE Healthcare) was used to capture right leg mid-thigh VL images in the transverse plane for measurement of mCSA values. The panoramic function of the device (LogicView, GE Healthcare) was used to capture the entire image of the musculature. All ultrasound settings were held constant across participants and laboratory visits (frequency: 10 MHz, gain: 50 dB, dynamic range: 75). Images were obtained and analyzed by a single investigator who was blinded to group allocations (DLP), and mCSA values were calculated by manual tracing of the border of the VL along the fascia using the polygon function in ImageJ software (National Institutes of Health, Bethesda, MD, USA). DLP’s previously determined test-retest reliability for VL mCSA in 10 non-involved participants yielded an intraclass correlation coefficient of 0.984.
2.5. Pre- and Post-Intervention Right Mid-Thigh VL Biopsies
VL biopsies were obtained from the mid-belly of the right leg in the same plane as ultrasound assessments. Following ultrasound assessments, the participants remained in a supine position on an athletic training table while the upper thigh was shaved and cleaned with 70% isopropanol. A subcutaneous injection of 1% lidocaine (0.8 mL) was administered, and the area was cleaned with chlorhexidine solution after a 5-minute wait period. Thereafter, a 7-mm pilot incision was made through the dermis with a sterile No. 11 surgical blade (AD Surgical; Sunnyvale, CA., USA.) A 5-gauge biopsy needle was inserted into the pilot incision, through the muscle fascia, and ~2 cm into the muscle (for a total depth of ~4-7 cm) where a 40-80 mg sample was collected while applying suction. Following the biopsy, the tissue was rapidly teased of blood and connective tissue. A portion of tissue (~50 mg) was preserved in freezing media (OCT; Tissue-Tek, Sakura Finetek Inc; Torrence, CA, USA), slowly frozen in liquid nitrogen-cooled isopentane, placed in a box floating atop liquid nitrogen for ~1-3 hours, and subsequently stored at −80°C for immunohistochemical analysis. Another portion of tissue (~30 mg) was placed in prelabeled foil, placed directly in liquid nitrogen for 1-3 hours, and subsequently stored at −80°C for molecular analysis. This tissue triage process (performed by JMM) took ~2 minutes per participant.
Strength and strength-endurance outcomes At PRE and POST, each participant’s estimated one-repetition maximum (est. 1-RM) was assessed using a 5-repetition maximum test for the hex bar deadlift exercise based on National Strength and Conditioning Association testing guidelines [55] (testing performed by KH, BMP, and DAA). Briefly, participants were escorted to an exercise training room following VL biopsies. Participants were then instructed to perform a series of dynamic stretches of the lower body musculature, which took approximately 5 minutes. Following 10 minutes of gait assessments (not reported herein), participants were coached through the form of the hex bar deadlift by study staff, a warm-up set of 10 repetitions was performed prior to 5-RM attempts, and up to five 5-RM attempts were performed with each attempt being separated by 2-3 minutes. Following this assessment, participants rested for 3-5 minutes. A hex bar deadlift repetitions-to failure test was then performed using 60% of each participant’s est. 1-RM results. During this test, participants completed as many hex bar deadlift repetitions as possible with proper exercise technique. The primary outcome was volume-load (total weight lifted), calculated by multiplying load by number of repetitions performed.
2.6. Resistance Training Program
Full-body RT sessions were performed on non-consecutive days at the same time-of-day in a dedicated research training facility (two non-consecutive days/week for 12 weeks) under the supervision of a National Strength and Conditioning Association certified strength and conditioning specialist (MCM) and other research staff using progressive overload that emphasized skeletal muscle hypertrophy. Each training session lasted ~60 minutes beginning with ~10-minute standardized warm-up and warm-up sets of the allocated exercises. Each session included hex bar deadlifts, pin-loaded chest press machine, 45º plate-loaded leg press, pin-loaded cable pulldown machine, pin-loaded leg extensor machine, and pin-loaded prone leg curl machine.
Intensity was monitored using repetitions in reserve (RIR) as described by Zourdos et al. [56]. Participant RIR was recorded after every set of each exercise and adjustments in load were made to achieve an RIR of 0-2. If RIR was >2, the load was increased. If participants were unable to complete the prescribed number of repetitions, the load was decreased. The training program utilized a linear progression where load was increased and volume was decreased throughout the 12-week period. Note that no participant was excluded from the study due to exercise adherence, which was predetermined to be ≥4 workouts over the 12-week intervention. For the first four weeks, load was assigned with the intent of participants performing 3 sets of ~12-15 repetitions per exercise. For weeks 5-8, load was assigned with the intent of participants performing 3 sets of 10-12 repetitions for each exercise. For the final four weeks, the load was assigned with the intent of participants performing 3 sets of 8-10 repetitions for each exercise. Following the conclusion of the study, total intervention volume-load (total sets x total repetitions x load per set) as well as lower body volume-load were calculated for each participant and expressed in kg.
2.7. Analyses on Biopsy Specimens
Immunohistochemistry for type I and II fCSA and capillary quantification. Biopsy samples preserved in OCT were sectioned at a thickness of 7-12 µm at -21℃ using a cryotome (Leica Biosystems; Buffalo Grove, IL, USA). Sections were adhered to positively charged glass slides (VWR; Radnor, PA, USA) and stored at -80℃ until batch processing for immunohistochemical analyses to detect capillaries, type I fibers and type II fibers, and dystrophin. PRE and POST samples from the same participant were placed on the same slide to limit slide-to-slide variation. During batch-processing, sections were removed from -80ºC storage, air-dried at room temperature for ≥2 hours and fixed with acetone at -20°C for five minutes. Slides were then incubated with 3% hydrogen peroxide for 10 minutes at room temperature, followed by one-minute incubation with autofluorescence quenching reagent (TrueBlack, Cat. No. 23007; Biotium, Fremont, CA, USA) and blocked for one hour in 2.5% horse serum in phosphate-buffered saline (PBS) at room temperature. After blocking, slides were incubated overnight at 4°C with a primary antibody cocktail in PBS containing 2.5% horse serum and 1:100 v/v dilutions of anti-PECAM-1, anti-type I myosin heavy chain, and anti-dystrophin antibodies listed in Table 1. The following day, sections were washed with PBS for 3x5 minutes and incubated for 60 minutes in a secondary antibody cocktail containing 1:250 v/v dilutions of the fluorophore-conjugated secondary antibodies listed in Table 1. Slides were then mounted with glass coverslips using 1:1 PBS and glycerol as mounting medium and stored in the dark at 4°C until imaging was conducted. Multiple digital 10× images per sample were captured to visualize the entire section, then stitched together using the “Acquisition” function, selecting “Tiles”, then selecting multiple “Support Points” setting with a fluorescence microscope and motorized stage (Zeiss Axio Imager.M2) [57]. All images were captured in 10×. Approximately 388 myofibers were analyzed per sample. Type I and II fiber cross-sectional area (fCSA) quantification was conducted using Myovision [58], and analyzed images were manually checked to remove faulty myofibers (e.g., oblong or freeze-fractured) from the final analysis. Capillary contacts to type I and type II fibers were manually quantified by a single investigator (MCM) using a tally counter in the Zeiss Axio Imager.M2 software under the “Points” and “Events” function [59]. Capillaries that contacted multiple fibers were tallied for each fiber. Capillary number was reported as capillary per fiber type (type I or type II) as previously reported [59].
Immunoblotting. Per a recent publication from our laboratory [59], the immunoblotting targets of interest were related to increased and inhibited angiogenesis and responsiveness. Specifically, primary targets included vascular endothelial growth factor (VEGF), VEGF receptor 2 (VEGFR2), and thrombospondin-1 (TSP-1), an inhibitor of angiogenesis [60]. Transcription factor EB (TFEB) was assayed given its role in regulating angiogenesis [61]. Finally, total endothelial nitric oxide synthase (eNOS) and bioactive phosphorylated eNOS (p-eNOS, Ser1177) were assayed given the interaction of this enzyme with NO3- in regulating nitric oxide production [62].
Muscle tissue (~20 mg) was lysed using a general cell/tissue lysis buffer (Cell Signaling Technology, Danvers, MA, USA; Cat. No. 9803) and tight-fitting pestles. Lysates were then centrifuged at 500 g for five minutes and supernatants were placed into new 1.7 mL tubes. A commercially available BCA protein assay kit (Thermo Fisher, Waltham, MA, USA; Cat. No. A55864) was used to determine supernatant protein concentrations. Thereafter, supernatants were prepared for western blotting at equal protein concentrations (1 µg/µL) using 4x Laemmli buffer and deionized water. Western blot preps (15 µL) were pipetted onto SDS gels (4-15% Criterion TGX Stain-free gels, Bio-Rad Laboratories; Hercules, CA, USA), and proteins were separated by electrophoresis at 180 V for 50 minutes. Proteins were then transferred to methanol-preactivated PVDF membranes (Bio-Rad Laboratories) for 2 hours at 200 mA. Following transfers, the membranes were Ponceau stained for 10 minutes, washed with distilled water for ~30 seconds, dried, and digitally imaged (ChemiDoc Touch, Bio-Rad). Following Ponceau imaging, membranes were reactivated in methanol, blocked with 5% non-fat bovine milk in tris-buffered saline with Tween-20 (TBST) for one hour, and washed 3x5 minutes in TBST. Membranes were then incubated with primary antibodies diluted 1:1000 in TBST containing 5% bovine serum albumin (BSA) on a rocker overnight at 4°C (antibodies listed in Table 1).
Following overnight primary antibody incubations, antibody solutions were decanted, and membranes were washed for 3x5 minutes in TBST. The membranes were then incubated at room temperature for 60 minutes in TBST containing 5% BSA and a 1:2000 v/v dilution of HRP-conjugated antibody against the host species of the primary antibody (antibodies listed in Table 1). The secondary antibody solution was decanted, and the membranes were washed for 3x5 minutes in TBST. The membranes were then developed in a gel documentation system (ChemiDoc Touch, Bio-Rad) with enhanced chemiluminescent reagent (Luminata Forte HRP substrate; Millipore Sigma, Burlington, MA, USA), and band densitometry was performed using associated software. For non-phosphorylated targets, target band densities were obtained and divided by Ponceau densitometry values and fold-change values were derived by dividing Ponceau-normalized band density values by the aggregate PRE mean value of the PLA group. Band density values for phosphorylated eNOS were divided by corresponding pan band density values, and again fold-change values were derived by dividing these values by the aggregate PRE mean value of the PLA group.
Skeletal muscle nitrate/nitrite measurement. General skeletal muscle lysates were assayed in duplicate using a commercially available fluorometric assay kit (Cayman Chemical; Nitrate/Nitrite Fluorometric Assay Kit; Item No. 780051, Ann Arbor, MI, USA) according to manufacturer’s instructions. First, lysates were spun in 30 kD molecular weight cutoff spin filter tubes at 12,000 g for 10 minutes. Filtrates (10 µL) were then placed in a white 96-well plate; 90 µL of kit assay buffer was added to each well, and 20 µL of nitrate reductase/enzyme co-factor solution was added to each well. Following a 120-minute room temperature incubation in the dark, 10 µL of 2,3-diaminonaphthalene solution was added to each well. The plate was then incubated at room temperature for 10 minutes and 10 µL of sodium hydroxide provided in the kit was added to each well. The plate was then read using a microplate fluorometer (BioTek Synergy H1, Winooski, VT, USA) with an excitation wavelength of 360 nm and emission wavelength of 430 nm. Sample NOx concentrations were extrapolated using a standard curve, and these data were normalized to lysate protein concentrations yielding nmol NOx/milligram of protein values.
2.8. Statistical Analyses
Statistical analyses were completed using GraphPad Prism (Version 10.4.2). Except for 12-week training volume-load and adherence data (which were compared between groups using independent samples t-tests), outcome variables were analyzed using two-way mixed ANOVA (supplement group × time) with repeated measures on the time factor. Additional exploratory analyses were performed using jamovi v2.3.28 and included examining potential sex interactions for FMD and other key outcome variables, which was performed using three-way (supplement group × sex × time) repeated measures ANOVAs. Data are presented as mean ± standard deviation values, and statistical significance was set at p<0.05 throughout.
3. Results
3.1. Participants, Protocol Adherence, and Missing Data
Of the 30 participants initially enrolled in the study, 28 (14 male; 14 female) participants completed the intervention. Two participants did not return following PRE assessments due to time constraints and, thus, were not randomized into groups. Table 2 contains pre-intervention participant characteristics for the 28 completers; notably, no significant pre-intervention differences existed for age, height, body mass, body mass index, or hex bar deadlift strength between groups.
Supplementation adherence was excellent, with BRJ participants reporting 98±2% and PLA participants 99±2% compliance (p=0.313). Exercise adherence was also excellent as twenty-six of the 28 participants completed all 24 training sessions; one BRJ participant missed three sessions, and one PLA participant missed one session. Due to poor image quality, complete DXA data was obtained for 13 PLA and 13 BRJ participants, and complete muscle ultrasound data was obtained for 14 PLA and 12 BRJ participants. Technical issues with biopsies or tissue processing (low protein yield or damaged sections) resulted in varying sample sizes: 13 PLA and 13 BRJ for immunoblotting, 11 PLA and 13 BRJ for fCSA, 10 PLA and 11 BRJ for capillary data, and 12 PLA and 13 BRJ for muscle NOx data. Additionally, 10 PLA and 11 BRJ participants provided adequate quality images for FMD. No data were manually excluded from any analysis.
3.2. Whole Body and Mid-Thigh Skeletal Muscle Hypertrophic Outcomes
Main effects of time, but no significant group × time interactions (p>0.05), were observed for whole-body lean/soft tissue mass (time: p=0.002; Figure 2a), estimated total body skeletal muscle mass (time: p=0.001; Figure 2b), and VL mCSA values (time: p<0.001; Figure 2c). Each of these measures was significantly greater at POST than PRE.
3.3. Strength and Strength-Endurance Outcomes
An independent t-test determined no statistical differences between groups for 12-week total training volume-load (p=0.294) and 12-week lower body training volume-load (p=0.268) between PLA and BRJ (Figure 3a). Main effects of time were observed for hex bar estimated 1-RM (p<0.001; Figure 3b) and hex bar strength-endurance test volume-load completed (p<0.001; Figure 3c) whereby POST values were significantly greater than PRE, but no significant group × time interaction was found for either measure (p>0.05).
3.4. Popliteal Artery FMD Outcomes
Main effects of time were observed for relative FMD (p<0.001; Figure 4a) and normalized FMD (p<0.001; Figure 4b) whereby POST values were significantly greater than PRE, but no significant group × time interactions were present (p>0.05). Table 3 has all FMD-related variables (including Figure 4 data) and three-way ANOVA statistics.
3.5. Fiber Cross-Sectional, Fiber Type, and Skeletal Muscle Capillarization Outcomes
Type I fCSA displayed a significant main effect of time (p=0.017; Figure 5a) as PRE was greater than POST, but no significant group × time interaction existed. No significant main effects of time or group × time interactions (p>0.05) existed for type IIA/X fCSA (Figure 5b), type IIA/X percentage (Figure 5c), type I myofiber capillary contacts per fiber (Figure 5d), or type II myofiber capillary contacts per fiber (Figure 5e).
3.6. Immunoblotting Markers and Muscle NOx
TSP-1 protein content displayed a significant main effect of time (POST>PRE, p=0.031; Figure 6b), but no significant group × time interaction was present. VEGF protein content (Figure 6a), VEGFR2 protein content (Figure 6c), TFEB protein content (Figure 6d), pan eNOS protein content (Figure 6e), and phosphorylated-eNOS protein content (Figure 6f) did not display significant main effects of time or group × time interactions. VL muscle NOx (not graphed) did not display a significant main effect of time (p=0.141) or a group × time interaction (p=0.741). Levels non-significantly increased 15.4% in the BRJ group from PRE to POST (7.7±2.1 to 8.9±1.6 ng/mg protein, p=0.073) and 7.8% in the PLA group from PRE to POST (9.6±2.8 to 10.4±3.4 ng/mg protein, p=0.514) though these changes were not statistically significant.
3.7. Exploratory Analysis of Sex Effects for Non-FMD Variables
Table 4 contains group × sex × time interaction p-values for the other (non-FMD) outcome variables demonstrating significant training effects. Three-way ANOVA analyses revealed no significant group × sex × time interactions for RT-responsive outcome variables, suggesting that supplementation effects were comparable between males and females.
4. Discussion
To our knowledge, this is the first study to comprehensively examine how chronic NO3- supplementation affects RT adaptations in middle-aged and older adults. The primary finding was that 12 weeks of BRJ supplementation (800 mg NO3-/day) did not enhance skeletal muscle hypertrophy, strength, or vascular adaptations compared to placebo during the supervised RT program. Additionally, our three-way ANOVA results did not reveal sex-specific effects with supplementation. These null findings were contrary to our hypotheses and warrant further discussion.
Several factors may explain why BRJ supplementation did not enhance chronic training adaptations despite other literature reporting acute performance benefits. First, while muscle NOx levels numerically increased in the BRJ group (+15.4% vs +7.8% in placebo), this was not statistically significant. As our hypothesis was predicated on BRJ supplementation increasing muscle nitrate content to elicit beneficial training effects, this may not have been accomplished with the 800 mg/day supplementation protocol. It is also possible that the 24-hour washout period prior to POST testing may have allowed tissue NO3- levels to return toward baseline, potentially explaining the lack of significant muscle NOx accumulation. This aspect of the study design was chosen to ensure that FMD outcomes were not influenced by the consumption of BRJ during the morning of POST testing. However, it is notable that the timing of NO3- consumption (e.g., 1-2 hours prior to exercise) likely influences exercise performance outcomes as well as plasma and tissue NO3- concentrations [28]. Second, the 800 mg/day dose, while effective for acute performance benefits, may be insufficient for chronic adaptations in this population. In this regard, several acute supplementation studies have reported NO3- supplementation increases repetitions to failure [33], oxygen and nutrient delivery to skeletal muscle during exercise [34], power-related outcomes [63], and exercise tolerance [29]. Additionally, a study by Benjamin et al. [50] reported six days 800 mg NO3- supplementation improves cardiovascular parameters in postmenopausal women following acute submaximal exercise, and others have reported four weeks of L-citrulline supplementation (a supplement that also increases nitric oxide bioavailability [64]) improves cardiovascular measures in postmenopausal women [65,66]. However, our null findings agree with other published reports in older adults examining longer-term supplementation paradigms. For instance, Caballero-Garcia et al. [67] reported that six weeks of L-citrulline supplementation did not affect various strength or endurance outcomes in older individuals. Carter et al. [40] reported eight weeks of BRJ (providing the same dose of NO3- as our study) combined with circuit-style training did not significantly increase knee extensor power in postmenopausal women. Collectively, the disconnect between acute improvements in exercise performance with acute peri-exercise NO3- ingestion and chronic adaptations with long-term supplementation remains unclear.
At the molecular level, we posited that BRJ supplementation may also affect markers of angiogenesis (VEGF, VEGFR, TSP-1, and eNOS) or skeletal muscle capillarization since increased nitric oxide can alter vascular function and shear stress to potentially affect these outcomes [68]. However, these markers were also not affected by supplementation. In contrast with other studies reporting RT to increase skeletal muscle VEGF, VEGFR, and skeletal muscle capillarization [19,59,69,70,71], we also did not observe training effects in spite of participants experiencing increases in outcomes related to skeletal muscle hypertrophy. It is worth noting that findings from several of the aforementioned studies were in younger adults and these adaptations (VEGF expression in particular) are seemingly blunted in older adults [21,22,23,24]. Our results also suggest that BRJ supplementation does not augment markers of angiogenesis from RT among middle-aged and older adults. In contrast, we found that RT improved FMD and the shear rate during reactive hyperemia (both peak shear and the shear area under the curve), indicating an improvement in applied measures of microvascular and macrovascular endothelial function, although there was no effect of supplementation. These findings suggest that the RT intervention led to favorable adaptations that could reduce cardiovascular and cardiometabolic disease risk in this population.
A final noteworthy and unanticipated finding unrelated to supplementation was the decrease in type I fCSA from PRE to POST as well as no change in type II fCSA values with training. This contrasts with several studies indicating that RT increases type I and II fCSA values [72]. Both groups experienced significant increases in other skeletal muscle hypertrophy outcomes (e.g., VL mCSA and DXA lean tissue mass metrics) following the 12-week RT intervention. However, this disconnect is not entirely surprising given that our laboratory has previously reported that different measures of skeletal muscle hypertrophy do not always correlate [57,73]. Moreover, our laboratory and others have reported that RT interventions in older individuals do not significantly affect type I or II fCSA outcomes despite other tissue-level hypertrophy variables significantly increasing [4,74]. It is also notable that magnitudes of skeletal muscle hypertrophy, vascular health, and plasma NO3- also differ between younger and older adults [6,75,76]. Hence, a younger cohort serving as a comparator group could have provided additional insight into the chronic effects of NO3- supplementation, and future studies should consider examining similar outcomes in a younger population.
4.1. Limitations and Future Considerations
Though well-designed and executed, several limitations, in addition to sample size, are noted. There were no measures of skeletal muscle blood flow indicators during training sessions. Future studies assessing skeletal muscle blood flow or proxy measurements such as near-infrared spectroscopy to evaluate blood flow measures during exercise could be insightful [77]. Diet influences hypertrophic outcomes [2], but dietary habits were not recorded and may have affected our results. Another limitation is that we did not assess peri-ingestion plasma NOx to determine whether the dose of BRJ we provided lead to substantial increases in NO3- and NO2- within our participants. Achieving large increases in circulating NOx would presumably be needed to facilitate an increase in muscle NOx, which we did not observe. Nonetheless, the dose of NO3- we administered was consistent with several prior studies.
4.2. Conclusions
Our findings indicate that 12 weeks of NO3- rich BRJ supplementation does not enhance various RT adaptations in middle-aged and older adults. Future studies are needed to understand the disconnect between acute ergogenic findings that are widely reported and the null findings regarding chronic NO3-supplementation in this and other studies.
Author Contributions
This study served as a dissertation project for MCM. MCM. primarily conceived the study idea with significant input from MDR, DTB, LBG, ATR, and ANK. MCM, MDR, and DTB applied for and were awarded grant funding for the study. MCM and MDR primarily drafted the manuscript and prepared figures. MCM primarily carried out laboratory-based assays. BJM, DRT, DAA, AA-B, GK Jr., DLP, MLM, NJK, JMM, KH, BMP, DL, MKC, and HK provided crucial assistance with original data collection procedures and/or training. All co-authors assisted with revising and editing the manuscript, and all co-authors approved the final version.
Funding
MCM was supported by the Porter Fellowship (American Physiological Society), DLP and DL were supported by Auburn University Presidential Research Fellowships, DAA was supported by an Auburn University Presidential Opportunity Fellowship, and DRT was partially supported by Auburn University’s College of Education Dean Fellowship. Funding for most direct costs for this project was provided by a REAP Grant awarded to DTB and MDR by the Edward Via College of Osteopathic Medicine. Some reagents were purchased via discretionary laboratory funds from ADF.
Institutional Review Board Statement
This study was not registered as a clinical trial. Study protocols were carried out in accordance with the most recent version of the declaration of Helsinki. All study procedures were approved by Auburn University’s Institutional Review Board (approval number: Protocol # 24-863 MR 2405). All participants in this study provided verbal and written consent in accordance with the above IRB approval.
Data Availability Statement
Data for this study are available from one of the co-corresponding authors (mdr0024@auburn.edu) upon reasonable request.
Acknowledgments
We would like to acknowledge the participants who completed this study.
Conflicts of Interest
The authors declare they have no competing interests in relation to these data.
References
- Troen BR. The biology of aging. Mt Sinai J Med 70: 3-22, 2003.
- Breen L, and Phillips SM. Skeletal muscle protein metabolism in the elderly: Interventions to counteract the 'anabolic resistance' of ageing. Nutr Metab (Lond) 8: 68, 2011. [CrossRef]
- Bell KE, von Allmen MT, Devries MC, and Phillips SM. Muscle Disuse as a Pivotal Problem in Sarcopenia-related Muscle Loss and Dysfunction. J Frailty Aging 5: 33-41, 2016. [CrossRef]
- Ruple BA, Mattingly ML, Godwin JS, McIntosh MC, Kontos NJ, Agyin-Birikorang A, Michel JM, Plotkin DL, Chen SY, Ziegenfuss TN, Fruge AD, Gladden LB, Robinson AT, Mobley CB, Mackey AL, and Roberts MD. The effects of resistance training on denervated myofibers, senescent cells, and associated protein markers in middle-aged adults. FASEB J 38: e23621, 2024. [CrossRef]
- Michel JM, Lievense KK, Norton SC, Costa JV, Alphin KH, Bailey LA, and Miller GD. The Effects of Graded Protein Intake in Conjunction with Progressive Resistance Training on Skeletal Muscle Outcomes in Older Adults: A Preliminary Trial. Nutrients 14: 2022. [CrossRef]
- Kosek DJ, Kim JS, Petrella JK, Cross JM, and Bamman MM. Efficacy of 3 days/wk resistance training on myofiber hypertrophy and myogenic mechanisms in young vs. older adults. J Appl Physiol (1985) 101: 531-544, 2006. [CrossRef]
- Phillips BE, Williams JP, Greenhaff PL, Smith K, and Atherton PJ. Physiological adaptations to resistance exercise as a function of age. JCI Insight 2: 2017. [CrossRef]
- Shad BJ, Thompson JL, and Breen L. Does the muscle protein synthetic response to exercise and amino acid-based nutrition diminish with advancing age? A systematic review. Am J Physiol Endocrinol Metab 311: E803-E817, 2016. [CrossRef]
- Banks NF, Rogers EM, Church DD, Ferrando AA, and Jenkins NDM. The contributory role of vascular health in age-related anabolic resistance. J Cachexia Sarcopenia Muscle 13: 114-127, 2022. [CrossRef]
- Morton RW, Traylor DA, Weijs PJM, and Phillips SM. Defining anabolic resistance: implications for delivery of clinical care nutrition. Curr Opin Crit Care 24: 124-130, 2018. [CrossRef]
- Burd NA, Gorissen SH, and van Loon LJ. Anabolic resistance of muscle protein synthesis with aging. Exerc Sport Sci Rev 41: 169-173, 2013. [CrossRef]
- Haran PH, Rivas DA, and Fielding RA. Role and potential mechanisms of anabolic resistance in sarcopenia. J Cachexia Sarcopenia Muscle 3: 157-162, 2012. [CrossRef]
- Aragon AA, Tipton KD, and Schoenfeld BJ. Age-related muscle anabolic resistance: inevitable or preventable? Nutr Rev 81: 441-454, 2023.
- Holder SM, Bruno RM, Shkredova DA, Dawson EA, Jones H, Hopkins ND, Hopman MTE, Bailey TG, Coombes JS, Askew CD, Naylor L, Maiorana A, Ghiadoni L, Thompson A, Green DJ, and Thijssen DHJ. Reference Intervals for Brachial Artery Flow-Mediated Dilation and the Relation With Cardiovascular Risk Factors. Hypertension 77: 1469-1480, 2021. [CrossRef]
- Xu X, Wang B, Ren C, Hu J, Greenberg DA, Chen T, Xie L, and Jin K. Age-related Impairment of Vascular Structure and Functions. Aging Dis 8: 590-610, 2017. [CrossRef]
- Verdijk LB, Snijders T, Holloway TM, J VANK, and LJ VANL. Resistance Training Increases Skeletal Muscle Capillarization in Healthy Older Men. Med Sci Sports Exerc 48: 2157-2164, 2016. [CrossRef]
- Dvoretskiy S, Lieblein-Boff JC, Jonnalagadda S, Atherton PJ, Phillips BE, and Pereira SL. Exploring the Association between Vascular Dysfunction and Skeletal Muscle Mass, Strength and Function in Healthy Adults: A Systematic Review. Nutrients 12: 2020. [CrossRef]
- Yoo JI, Kim MJ, Na JB, Chun YH, Park YJ, Park Y, Hah YS, Ha YC, and Park KS. Relationship between endothelial function and skeletal muscle strength in community dwelling elderly women. J Cachexia Sarcopenia Muscle 9: 1034-1041, 2018. [CrossRef]
- Snijders T, Nederveen JP, Joanisse S, Leenders M, Verdijk LB, van Loon LJ, and Parise G. Muscle fibre capillarization is a critical factor in muscle fibre hypertrophy during resistance exercise training in older men. J Cachexia Sarcopenia Muscle 8: 267-276, 2017. [CrossRef]
- Moro T, Brightwell CR, Phalen DE, McKenna CF, Lane SJ, Porter C, Volpi E, Rasmussen BB, and Fry CS. Low skeletal muscle capillarization limits muscle adaptation to resistance exercise training in older adults. Exp Gerontol 127: 110723, 2019. [CrossRef]
- Olfert IM, Baum O, Hellsten Y, and Egginton S. Advances and challenges in skeletal muscle angiogenesis. Am J Physiol Heart Circ Physiol 310: H326-336, 2016. [CrossRef]
- Ryan NA, Zwetsloot KA, Westerkamp LM, Hickner RC, Pofahl WE, and Gavin TP. Lower skeletal muscle capillarization and VEGF expression in aged vs. young men. J Appl Physiol (1985) 100: 178-185, 2006. [CrossRef]
- Croley AN, Zwetsloot KA, Westerkamp LM, Ryan NA, Pendergast AM, Hickner RC, Pofahl WE, and Gavin TP. Lower capillarization, VEGF protein, and VEGF mRNA response to acute exercise in the vastus lateralis muscle of aged vs. young women. J Appl Physiol (1985) 99: 1872-1879, 2005. [CrossRef]
- Endo Y, Hwang CD, Zhang Y, Olumi S, Koh DJ, Zhu C, Neppl RL, Agarwal S, and Sinha I. VEGFA Promotes Skeletal Muscle Regeneration in Aging. Adv Biol (Weinh) 7: e2200320, 2023. [CrossRef]
- Johnson AL, and Webster M. Dark Chocolate Elevates Resting Energy Expenditure in Postmenopausal Women. Int J Exerc Sci 18: 316-328, 2025. [CrossRef]
- Wageh M, Fortino SA, Pontello R, Maklad A, McGlory C, Kumbhare D, Phillips SM, and Parise G. The Effect of Multi-Ingredient Protein versus Collagen Supplementation on Satellite Cell Properties in Males and Females. Med Sci Sports Exerc 56: 2125-2134, 2024. [CrossRef]
- McIntosh MC, Ruple BA, Kontos NJ, Mattingly ML, Lockwood CM, and Roberts MD. The effects of a sugar-free amino acid-containing electrolyte beverage on 5-kilometer performance, blood electrolytes, and post-exercise cramping versus a conventional carbohydrate-electrolyte sports beverage and water. J Int Soc Sports Nutr 21: 2296888, 2024. [CrossRef]
- Mueller BJ, Roberts MD, Mobley CB, Judd RL, and Kavazis AN. Nitric oxide in exercise physiology: past and present perspectives. Front Physiol 15: 1504978, 2024. [CrossRef]
- Jones AM. Dietary nitrate supplementation and exercise performance. Sports Med 44 Suppl 1: S35-45, 2014. [CrossRef]
- Lansley KE, Winyard PG, Bailey SJ, Vanhatalo A, Wilkerson DP, Blackwell JR, Gilchrist M, Benjamin N, and Jones AM. Acute dietary nitrate supplementation improves cycling time trial performance. Med Sci Sports Exerc 43: 1125-1131, 2011. [CrossRef]
- Stanaway L, Rutherfurd-Markwick K, Page R, and Ali A. Performance and Health Benefits of Dietary Nitrate Supplementation in Older Adults: A Systematic Review. Nutrients 9: 2017. [CrossRef]
- Gee LC, and Ahluwalia A. Dietary Nitrate Lowers Blood Pressure: Epidemiological, Pre-clinical Experimental and Clinical Trial Evidence. Curr Hypertens Rep 18: 17, 2016. [CrossRef]
- Williams TD, Martin MP, Mintz JA, Rogers RR, and Ballmann CG. Effect of Acute Beetroot Juice Supplementation on Bench Press Power, Velocity, and Repetition Volume. J Strength Cond Res 34: 924-928, 2020. [CrossRef]
- Zamani H, de Joode M, Hossein IJ, Henckens NFT, Guggeis MA, Berends JE, de Kok T, and van Breda SGJ. The benefits and risks of beetroot juice consumption: a systematic review. Crit Rev Food Sci Nutr 61: 788-804, 2021. [CrossRef]
- McMahon NF, Leveritt MD, and Pavey TG. The Effect of Dietary Nitrate Supplementation on Endurance Exercise Performance in Healthy Adults: A Systematic Review and Meta-Analysis. Sports Med 47: 735-756, 2017. [CrossRef]
- Hobbs DA, George TW, and Lovegrove JA. The effects of dietary nitrate on blood pressure and endothelial function: a review of human intervention studies. Nutr Res Rev 26: 210-222, 2013. [CrossRef]
- Hobbs DA, Goulding MG, Nguyen A, Malaver T, Walker CF, George TW, Methven L, and Lovegrove JA. Acute ingestion of beetroot bread increases endothelium-independent vasodilation and lowers diastolic blood pressure in healthy men: a randomized controlled trial. J Nutr 143: 1399-1405, 2013. [CrossRef]
- Clifford PS, and Hellsten Y. Vasodilatory mechanisms in contracting skeletal muscle. J Appl Physiol (1985) 97: 393-403, 2004. [CrossRef]
- Tan R, Pennell A, Price KM, Karl ST, Seekamp-Hicks NG, Paniagua KK, Weiderman GD, Powell JP, Sharabidze LK, Lincoln IG, Kim JM, Espinoza MF, Hammer MA, Goulding RP, and Bailey SJ. Effects of Dietary Nitrate Supplementation on Performance and Muscle Oxygenation during Resistance Exercise in Men. Nutrients 14: 2022. [CrossRef]
- bold>Carter SJ, Blechschmid TH, Baranauskas MN, Long EB, Gruber AH, Raglin JS, Lim K, and Coggan AR. Preworkout dietary nitrate magnifies training-induced benefits to physical function in late postmenopausal women: a randomized pilot study. Am J Physiol Regul Integr Comp Physiol 327: R534-R542, 2024. [CrossRef]
- Jones AM, Ferguson SK, Bailey SJ, Vanhatalo A, and Poole DC. Fiber Type-Specific Effects of Dietary Nitrate. Exerc Sport Sci Rev 44: 53-60, 2016. [CrossRef]
- Kolluru GK, Sinha S, Majumder S, Muley A, Siamwala JH, Gupta R, and Chatterjee S. Shear stress promotes nitric oxide production in endothelial cells by sub-cellular delocalization of eNOS: A basis for shear stress mediated angiogenesis. Nitric Oxide 22: 304-315, 2010. [CrossRef]
- Rammos C, Luedike P, Hendgen-Cotta U, and Rassaf T. Potential of dietary nitrate in angiogenesis. World J Cardiol 7: 652-657, 2015. [CrossRef]
- Hashimoto M, Akishita M, Eto M, Ishikawa M, Kozaki K, Toba K, Sagara Y, Taketani Y, Orimo H, and Ouchi Y. Modulation of endothelium-dependent flow-mediated dilatation of the brachial artery by sex and menstrual cycle. Circulation 92: 3431-3435, 1995. [CrossRef]
- Thijssen DHJ, Bruno RM, van Mil A, Holder SM, Faita F, Greyling A, Zock PL, Taddei S, Deanfield JE, Luscher T, Green DJ, and Ghiadoni L. Expert consensus and evidence-based recommendations for the assessment of flow-mediated dilation in humans. Eur Heart J 40: 2534-2547, 2019. [CrossRef]
- Thijssen DH, Black MA, Pyke KE, Padilla J, Atkinson G, Harris RA, Parker B, Widlansky ME, Tschakovsky ME, and Green DJ. Assessment of flow-mediated dilation in humans: a methodological and physiological guideline. Am J Physiol Heart Circ Physiol 300: H2-12, 2011.
- Wylie LJ, Kelly J, Bailey SJ, Blackwell JR, Skiba PF, Winyard PG, Jeukendrup AE, Vanhatalo A, and Jones AM. Beetroot juice and exercise: pharmacodynamic and dose-response relationships. J Appl Physiol (1985) 115: 325-336, 2013.
- Siervo M, Lara J, Ogbonmwan I, and Mathers JC. Inorganic nitrate and beetroot juice supplementation reduces blood pressure in adults: a systematic review and meta-analysis. J Nutr 143: 818-826, 2013. [CrossRef]
- Silva KVC, Costa BD, Gomes AC, Saunders B, and Mota JF. Factors that Moderate the Effect of Nitrate Ingestion on Exercise Performance in Adults: A Systematic Review with Meta-Analyses and Meta-Regressions. Adv Nutr 13: 1866-1881, 2022. [CrossRef]
- Benjamim CJR, da Silva LSL, Sousa YBA, Rodrigues GDS, Pontes YMM, Rebelo MA, Goncalves LDS, Tavares SS, Guimaraes CS, da Silva Sobrinho AC, Tanus-Santos JE, Gualano B, and Bueno Junior CR. Acute and short-term beetroot juice nitrate-rich ingestion enhances cardiovascular responses following aerobic exercise in postmenopausal women with arterial hypertension: A triple-blinded randomized controlled trial. Free Radic Biol Med 211: 12-23, 2024. [CrossRef]
- Walker MA, Bailey TG, McIlvenna L, Allen JD, Green DJ, and Askew CD. Acute Dietary Nitrate Supplementation Improves Flow Mediated Dilatation of the Superficial Femoral Artery in Healthy Older Males. Nutrients 11: 2019. [CrossRef]
- Hord NG, Tang Y, and Bryan NS. Food sources of nitrates and nitrites: the physiologic context for potential health benefits. Am J Clin Nutr 90: 1-10, 2009. [CrossRef]
- Kim J, Wang Z, Heymsfield SB, Baumgartner RN, and Gallagher D. Total-body skeletal muscle mass: estimation by a new dual-energy X-ray absorptiometry method. Am J Clin Nutr 76: 378-383, 2002. [CrossRef]
- Kephart WC, Wachs TD, Thompson RM, Mobley CB, Fox CD, McDonald JR, Ferguson BS, Young KC, Nie B, Martin JS, Company JM, Pascoe DD, Arnold RD, Moon JR, and Roberts MD. Correction to: Ten weeks of branched-chain amino acid supplementation improves select performance and immunological variables in trained cyclists. Amino Acids 50: 1495, 2018.
- Strength N-N, and Association C. Essentials of strength training and conditioning. Human kinetics, 2021.
- Zourdos MC, Klemp A, Dolan C, Quiles JM, Schau KA, Jo E, Helms E, Esgro B, Duncan S, Garcia Merino S, and Blanco R. Novel Resistance Training-Specific Rating of Perceived Exertion Scale Measuring Repetitions in Reserve. J Strength Cond Res 30: 267-275, 2016. [CrossRef]
- Michel JM, Godwin JS, Plotkin DL, McIntosh MC, Mattingly ML, Agostinelli PJ, Mueller BJ, Anglin DA, Kontos NJ, Berry AC, Vega MM, Pipkin AA, Stock MS, Graham ZA, Baweja HS, Mobley CB, Bamman MM, and Roberts MD. Effects of leg immobilization and recovery resistance training on skeletal muscle-molecular markers in previously resistance-trained versus untrained adults. J Appl Physiol (1985) 138: 450-467, 2025. [CrossRef]
- Wen Y, Murach KA, Vechetti IJ, Jr., Fry CS, Vickery C, Peterson CA, McCarthy JJ, and Campbell KS. MyoVision: software for automated high-content analysis of skeletal muscle immunohistochemistry. J Appl Physiol (1985) 124: 40-51, 2018. [CrossRef]
- McIntosh MC, Michel JM, Godwin JS, Plotkin DL, Anglin DA, Mattingly ML, Agyin-Birikorang A, Kontos NJ, Baweja HS, Stock MS, Mobley CB, and Roberts MD. Disuse and subsequent recovery resistance training affect skeletal muscle angiogenesis related markers regardless of prior resistance training experience. J Appl Physiol (1985) 2025. [CrossRef]
- Lawler PR, and Lawler J. Molecular basis for the regulation of angiogenesis by thrombospondin-1 and -2. Cold Spring Harb Perspect Med 2: a006627, 2012. [CrossRef]
- Doronzo G, Astanina E, Cora D, Chiabotto G, Comunanza V, Noghero A, Neri F, Puliafito A, Primo L, Spampanato C, Settembre C, Ballabio A, Camussi G, Oliviero S, and Bussolino F. TFEB controls vascular development by regulating the proliferation of endothelial cells. EMBO J 38: 2019. [CrossRef]
- Carlstrom M, Liu M, Yang T, Zollbrecht C, Huang L, Peleli M, Borniquel S, Kishikawa H, Hezel M, Persson AE, Weitzberg E, and Lundberg JO. Cross-talk Between Nitrate-Nitrite-NO and NO Synthase Pathways in Control of Vascular NO Homeostasis. Antioxid Redox Signal 23: 295-306, 2015. [CrossRef]
- Coggan AR, Hoffman RL, Gray DA, Moorthi RN, Thomas DP, Leibowitz JL, Thies D, and Peterson LR. A Single Dose of Dietary Nitrate Increases Maximal Knee Extensor Angular Velocity and Power in Healthy Older Men and Women. J Gerontol A Biol Sci Med Sci 75: 1154-1160, 2020. [CrossRef]
- Barkhidarian B, Khorshidi M, Shab-Bidar S, and Hashemi B. Effects of L-citrulline supplementation on blood pressure: A systematic review and meta-analysis. Avicenna J Phytomed 9: 10-20, 2019.
- Maharaj A, Fischer SM, Dillon KN, Kang Y, Martinez MA, and Figueroa A. Effects of L-Citrulline Supplementation on Endothelial Function and Blood Pressure in Hypertensive Postmenopausal Women. Nutrients 14: 2022. [CrossRef]
- Kang Y, Dillon KN, Martinez MA, Maharaj A, Fischer SM, and Figueroa A. L-Citrulline Supplementation Improves Arterial Blood Flow and Muscle Oxygenation during Handgrip Exercise in Hypertensive Postmenopausal Women. Nutrients 16: 2024. [CrossRef]
- Caballero-Garcia A, Pascual-Fernandez J, Noriega-Gonzalez DC, Bello HJ, Pons-Biescas A, Roche E, and Cordova-Martinez A. L-Citrulline Supplementation and Exercise in the Management of Sarcopenia. Nutrients 13: 2021. [CrossRef]
- Santos-Parker JR, Strahler TR, Bassett CJ, Bispham NZ, Chonchol MB, and Seals DR. Curcumin supplementation improves vascular endothelial function in healthy middle-aged and older adults by increasing nitric oxide bioavailability and reducing oxidative stress. Aging (Albany NY) 9: 187-208, 2017.
- Holloway TM, Snijders T, J VANK, LJC VANL, and Verdijk LB. Temporal Response of Angiogenesis and Hypertrophy to Resistance Training in Young Men. Med Sci Sports Exerc 50: 36-45, 2018. [CrossRef]
- Gavin TP, Drew JL, Kubik CJ, Pofahl WE, and Hickner RC. Acute resistance exercise increases skeletal muscle angiogenic growth factor expression. Acta Physiol (Oxf) 191: 139-146, 2007. [CrossRef]
- Kim HB, Seo MW, and Jung HC. Effects of Aerobic vs. Resistance Exercise on Vascular Function and Vascular Endothelial Growth Factor in Older Women. Healthcare (Basel) 11: 2023. [CrossRef]
- Fry AC. The role of resistance exercise intensity on muscle fibre adaptations. Sports Med 34: 663-679, 2004. [CrossRef]
- Ruple BA, Mesquita PHC, Godwin JS, Sexton CL, Osburn SC, McIntosh MC, Kavazis AN, Libardi CA, Young KC, and Roberts MD. Changes in vastus lateralis fibre cross-sectional area, pennation angle and fascicle length do not predict changes in muscle cross-sectional area. Exp Physiol 107: 1216-1224, 2022.
- Toth MJ, Miller MS, VanBuren P, Bedrin NG, LeWinter MM, Ades PA, and Palmer BM. Resistance training alters skeletal muscle structure and function in human heart failure: effects at the tissue, cellular and molecular levels. J Physiol 590: 1243-1259, 2012. [CrossRef]
- Babcock MC, DuBose LE, Witten TL, Brubaker A, Stauffer BL, Hildreth KL, and Moreau KL. Assessment of macrovascular and microvascular function in aging males. J Appl Physiol (1985) 130: 96-103, 2021. [CrossRef]
- Capper TE, Siervo M, Clifford T, Taylor G, Iqbal W, West D, and Stevenson EJ. Pharmacokinetic Profile of Incremental Oral Doses of Dietary Nitrate in Young and Older Adults: A Crossover Randomized Clinical Trial. J Nutr 152: 130-139, 2022. [CrossRef]
- McManus CJ, Collison J, and Cooper CE. Performance comparison of the MOXY and PortaMon near-infrared spectroscopy muscle oximeters at rest and during exercise. J Biomed Opt 23: 1-14, 2018. [CrossRef]
Figure 1.
Study design. A schematic of the study design of the 12-week intervention. Abbreviations: BRJ, nitrate-containing beetroot juice; PLA, placebo; FMD, flow-mediated dilation; VL, vastus lateralis; DXA, dual-energy x-ray absorptiometry.
Figure 1.
Study design. A schematic of the study design of the 12-week intervention. Abbreviations: BRJ, nitrate-containing beetroot juice; PLA, placebo; FMD, flow-mediated dilation; VL, vastus lateralis; DXA, dual-energy x-ray absorptiometry.
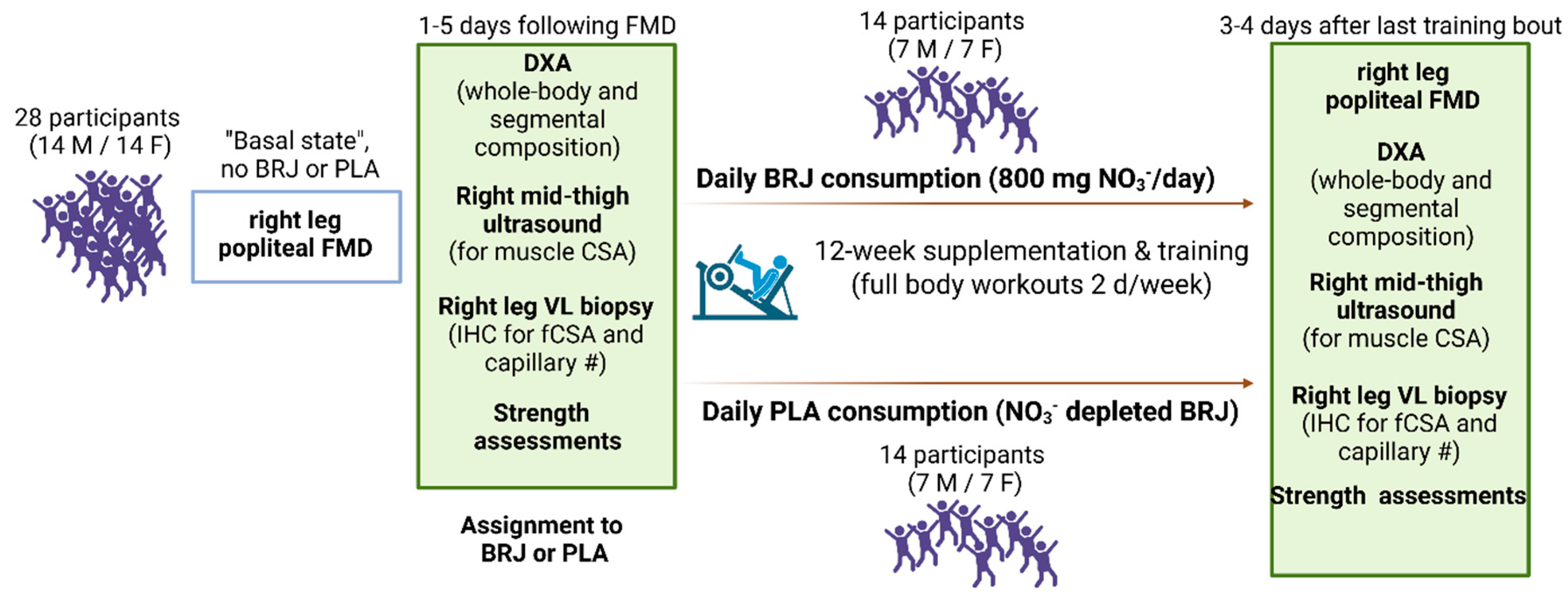
Figure 2.
DXA and mid-thigh ultrasound outcomes. Legend: Data for whole-body lean/soft tissue mass (a), calculated skeletal muscle (SkMus) mass when only considering dual-arm and dual-leg lean/soft tissue mass values (b), and vastus lateralis (VL) muscle cross-sectional area (mCSA, c) prior to (PRE) and following the 12-week intervention (POST). Bar graphs depict mean ± standard deviation bars, individual respondent data are superimposed on bar graphs, and mean values are shown at the bottom of each bar.
Figure 2.
DXA and mid-thigh ultrasound outcomes. Legend: Data for whole-body lean/soft tissue mass (a), calculated skeletal muscle (SkMus) mass when only considering dual-arm and dual-leg lean/soft tissue mass values (b), and vastus lateralis (VL) muscle cross-sectional area (mCSA, c) prior to (PRE) and following the 12-week intervention (POST). Bar graphs depict mean ± standard deviation bars, individual respondent data are superimposed on bar graphs, and mean values are shown at the bottom of each bar.
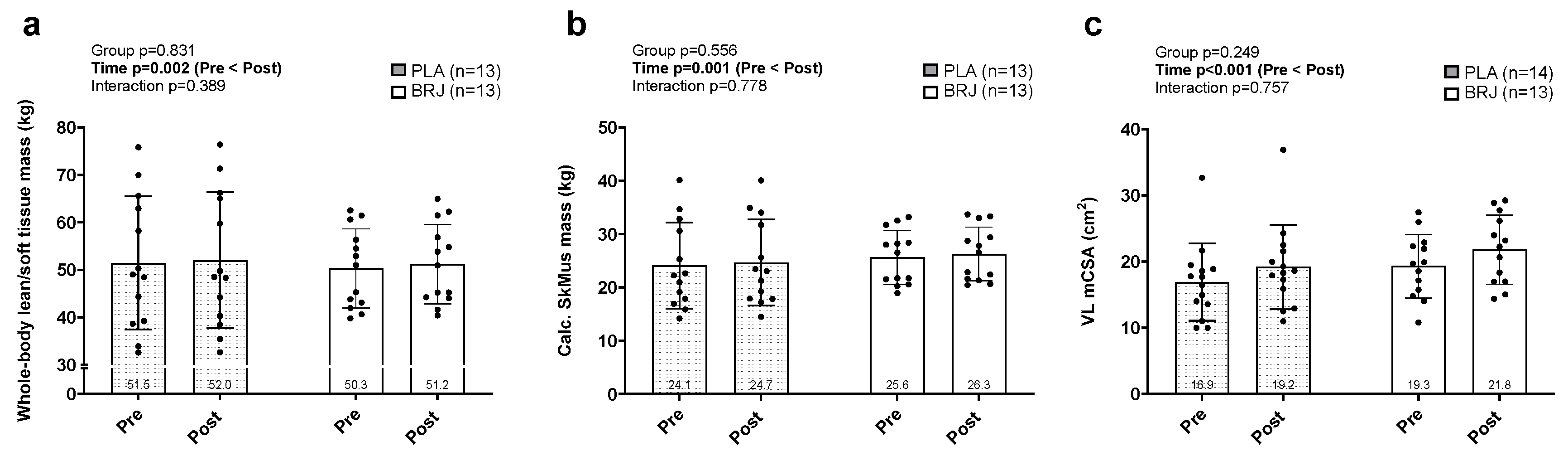
Figure 3.
Training volume and strength outcomes. Legend: Data for 12-week lower-body and total volume-load (a), hex bar estimated one repetition maximal lifts (est. 1-RM, b), volume performed based on 60% of hex bar est. 1-RM (c) prior to (PRE) and following the 12-week intervention (POST). Bar graphs depict mean ± standard deviation bars, individual respondent data are superimposed on bar graphs, and mean values are shown at the bottom of each bar.
Figure 3.
Training volume and strength outcomes. Legend: Data for 12-week lower-body and total volume-load (a), hex bar estimated one repetition maximal lifts (est. 1-RM, b), volume performed based on 60% of hex bar est. 1-RM (c) prior to (PRE) and following the 12-week intervention (POST). Bar graphs depict mean ± standard deviation bars, individual respondent data are superimposed on bar graphs, and mean values are shown at the bottom of each bar.
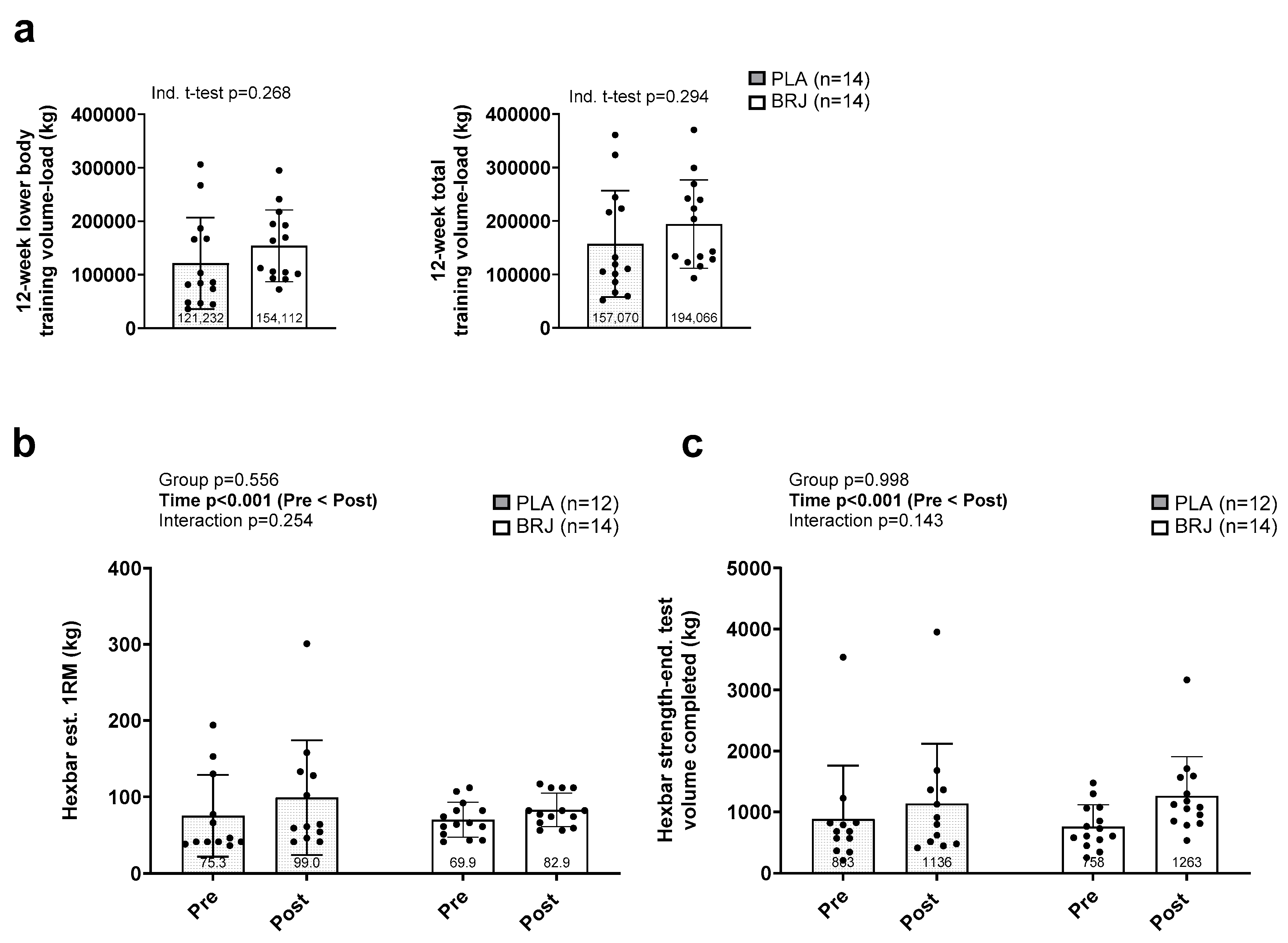
Figure 4.
Popliteal artery flow-mediated dilation outcomes. Legend: Data for relative flow-mediated dilation (FMD, a), and FMD normalized to shear stress (b) prior to (PRE) and following the 12-week intervention (POST). Bar graphs depict mean ± standard deviation bars, individual respondent data are superimposed on bar graphs, and mean values are shown at the bottom of each bar.
Figure 4.
Popliteal artery flow-mediated dilation outcomes. Legend: Data for relative flow-mediated dilation (FMD, a), and FMD normalized to shear stress (b) prior to (PRE) and following the 12-week intervention (POST). Bar graphs depict mean ± standard deviation bars, individual respondent data are superimposed on bar graphs, and mean values are shown at the bottom of each bar.
Figure 5.
Vastus lateralis immunohistochemistry outcomes. Legend: Data for type I fiber cross-sectional area (a), type IIA/X fiber cross-sectional area (b), fiber type percentage (c), capillary contacts with type I fibers (d), and capillary contacts with type II fibers (e) prior to (PRE) and following the 12-week intervention (POST). Panel f displays representative 10x microscopy images; note, images are pseudo-colored from original fluorescence colors for enhanced clarity (dystrophin: gray, MYHC1: purple, capillaries: yellow). Bar graphs depict mean ± standard deviation bars, individual respondent data are superimposed on bar graphs, and mean values are shown at the bottom of each bar.
Figure 5.
Vastus lateralis immunohistochemistry outcomes. Legend: Data for type I fiber cross-sectional area (a), type IIA/X fiber cross-sectional area (b), fiber type percentage (c), capillary contacts with type I fibers (d), and capillary contacts with type II fibers (e) prior to (PRE) and following the 12-week intervention (POST). Panel f displays representative 10x microscopy images; note, images are pseudo-colored from original fluorescence colors for enhanced clarity (dystrophin: gray, MYHC1: purple, capillaries: yellow). Bar graphs depict mean ± standard deviation bars, individual respondent data are superimposed on bar graphs, and mean values are shown at the bottom of each bar.
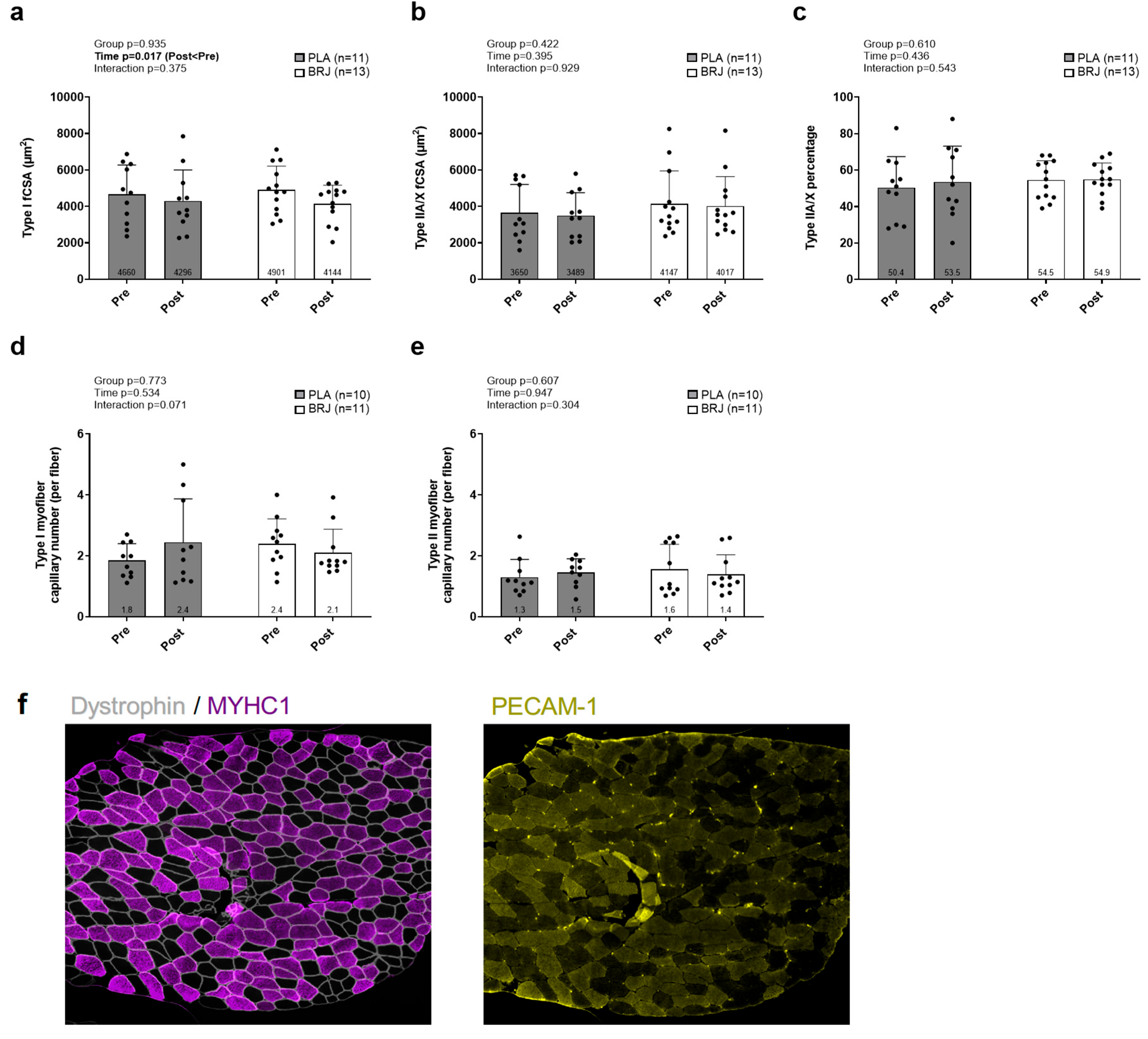
Figure 6.
Vastus lateralis immunoblotting outcomes for markers of angiogenesis. Legend: Protein levels for vascular endothelial growth factor (VEGF, a), Thrombospondin-1 (TSP-1, b), VEGF receptor 2 (VEGFR2, c), Transcription Factor EB (TFEB, d), endothelial nitric oxide synthase (eNOS, e), and phosphorylated eNOS (Ser1177, f) prior to (PRE) and following the 12-week intervention (POST). Panel g displays representative immunoblots. Bar graphs depict mean ± standard deviation bars, individual respondent data are superimposed on bar graphs, and mean values are shown at the bottom of each bar.
Figure 6.
Vastus lateralis immunoblotting outcomes for markers of angiogenesis. Legend: Protein levels for vascular endothelial growth factor (VEGF, a), Thrombospondin-1 (TSP-1, b), VEGF receptor 2 (VEGFR2, c), Transcription Factor EB (TFEB, d), endothelial nitric oxide synthase (eNOS, e), and phosphorylated eNOS (Ser1177, f) prior to (PRE) and following the 12-week intervention (POST). Panel g displays representative immunoblots. Bar graphs depict mean ± standard deviation bars, individual respondent data are superimposed on bar graphs, and mean values are shown at the bottom of each bar.
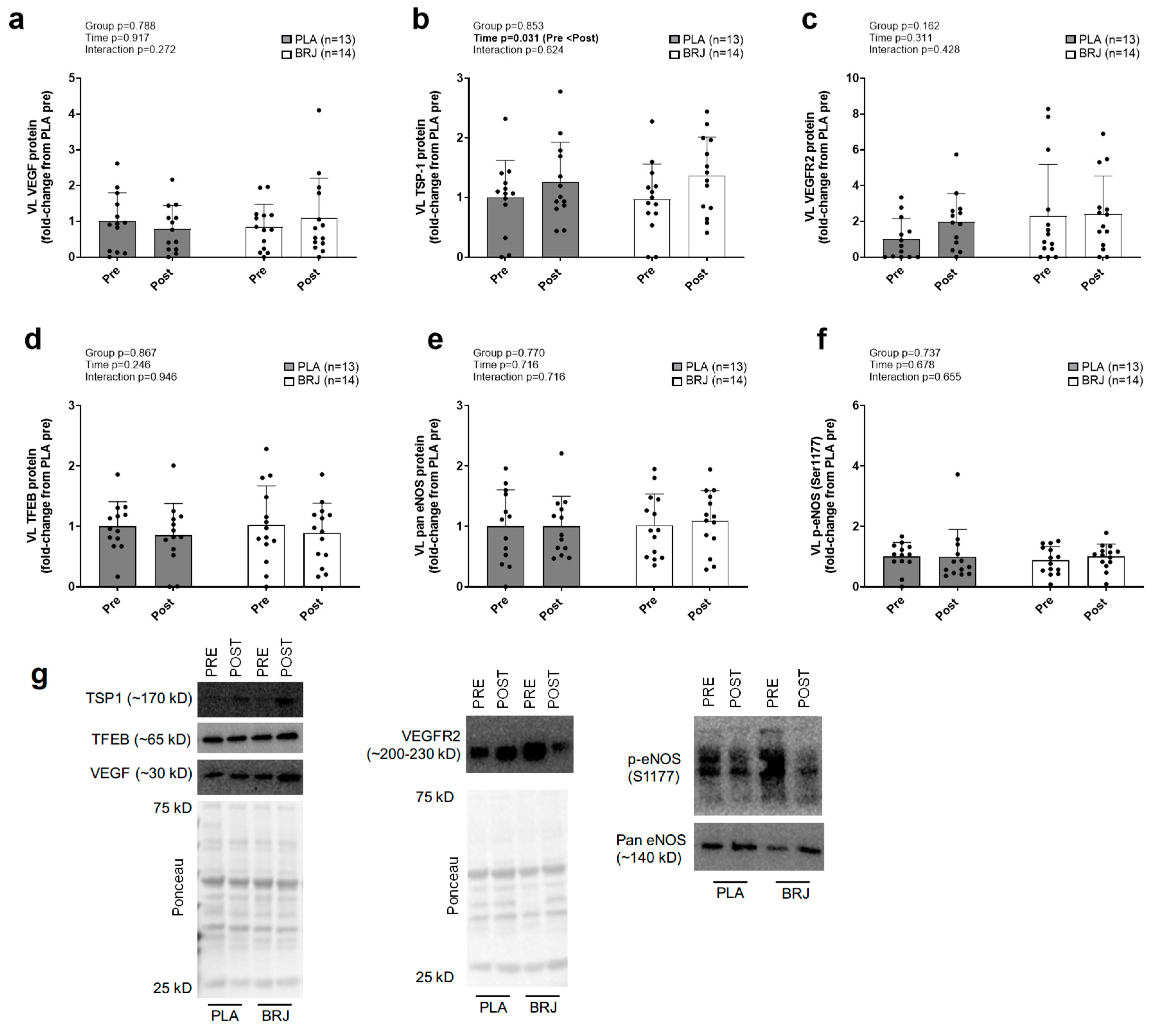

Table 1.
Antibodies used for immunoblotting and immunohistochemistry.
| Antibody | Host species (isotype) | Company (Cat. No.) |
| Western blotting | ||
| VEGF | Rb | Cell Signaling (65373) |
| VEGFR2 | Rb | Cell Signaling (9698) |
| TSP-1 | Rb | Cell Signaling (37879) |
| TFEB | Rb | Cell Signaling (83010) |
| eNOS | Rb | Cell Signaling (32027) |
| Phospho-eNOS (Ser1177) | Rb | Cell Signaling (9507) |
| Anti-Rb IgG, HRP-conjugated | G | Cell Signaling (7074) |
| Immunohistochemistry | ||
| CD31/PECAM-1 | M (IgG1) | DSHB (P2B1) |
| Type I myosin heavy chain | M (IgG2b) | DSHB (BA-D5) |
| Dystrophin | Rb (IgG) | Abcam (ab218198) |
| Anti-M IgG1, AF555-conjugated | G | Thermo Fisher (A-21127) |
| Anti-M IgG2b, AF488-conjugated | G | Thermo Fisher (A-21141) |
| Anti-Rb IgG, AF647-conjugated | G | Thermo Fisher (A-21245) |
Abbreviations: DSHB, Developmental Studies Hybridoma Bank (Iowa City, IA, USA); G, goat; M, mouse; Rb, rabbit. Other note: protein acronyms are defined in text.
Table 2.
Pre-intervention characteristics between supplementation groups.
| Variable | BRJ (n=14) | PLA (n=14) | Ind. t-test p-value |
| Age (years) | 56±6 | 56±7 | 1.000 |
| Sex | 7 M / 7 F | 7 M / 7 F | 1.000 |
| Height (cm) | 174±8 | 172±13 | 0.685 |
| Body mass (kg) | 87.3±16.9 | 85.1±25.1 | 0.788 |
| BMI (kg/m2) | 29.4±5.6 | 28.9±5.1 | 0.788 |
| Hex bar est. 1-RM | 70.0±22.8 | 75.2±53.7 | 0.741 |
Legend: independent samples t-tests were used to compare data between groups except for a Chi-square test being used to compare sex differences. Abbreviations: BMI, body mass index; Hex bar est. 1-RM, estimated maximal strength assessed using a five-repetition test for the hex bar exercise.
Table 3.
Exploratory group × sex × time statistics for popliteal artery flow-mediated dilation variables.
Table 3.
Exploratory group × sex × time statistics for popliteal artery flow-mediated dilation variables.
| Variable | BRJ | PLA | p-values | |
| Heart rate (bpm) | Pre: 61 ± 6 Post: 66 ± 9 |
Pre: 65 ± 7 Post: 73 ± 25 |
Time Sex × time Supplement × time Sex × supplement × time |
0.061 0.472 0.587 0.317 |
| SBP (mmHg) | Pre: 126 ± 13 Post: 131 ± 17 |
Pre: 128 ± 19 Post: 128 ± 19 |
Time Sex × time Supplement × time Sex × supplement × time |
0.400 0.460 0.420 0.795 |
| DBP (mmHg) | Pre: 80 ± 6 Post: 81 ± 8 |
Pre: 83 ± 10 Post: 83 ± 13 |
Time Sex × time Supplement × time Sex × supplement × time |
0.681 0.082 0.506 0.918 |
| MAP (mmHg) | Pre: 111 ± 10 Post: 114 ± 13 |
Pre: 113 ± 16 Post: 113 ± 17 |
Time Sex × time Supplement × time Sex × supplement × time |
0.424 0.356 0.442 0.837 |
| Baseline Diam. (mm) | Pre: 6.28 ± 1.31 Post: 6.25 ± 1.31 |
Pre: 6.19 ± 1.42 Post: 5.82 ± 1.49 |
Time Sex × time Supplement × time Sex × supplement × time |
0.723 0.738 0.488 0.476 |
| Average Shear Rate (s⁻¹) |
Pre: 44.4 ± 19.0 Post: 39.1 ± 17.6 |
Pre: 49.4 ± 22.8 Post: 44.3 ± 17.3 |
Time Sex × time Supplement × time Sex × supplement × time |
0.210 0.950 0.805 0.944 |
| Peak Shear Rate (s⁻¹) |
Pre: 67.1 ± 22.1 Post: 77.8 ± 22.8 |
Pre: 62.8 ± 9.7 Post: 65.9 ± 13.8 |
Time Sex × time Supplement × time Sex × supplement × time |
0.004 0.659 0.599 0.762 |
| Shear Rate Area Under the Curve (AU) | Pre: 6275 ± 2046 Post: 7000 ± 2054 |
Pre: 5228 ± 1858 Post: 5929 ± 1238 |
Time Sex × time Supplement × time Sex × supplement × time |
0.005 0.959 0.987 0.494 |
| FMD (%) | Data presented in Figure 4 | Time Sex × time Supplement × time Sex × supplement × time |
0.001 0.844 0.327 0.151 |
|
| Normalized FMD (%) |
Data presented in Figure 4 | Time Sex × time Supplement × time Sex × supplement × time |
0.001 0.699 0.279 0.211 |
|
Legend: bold p-values indicate significance. Abbreviations: BRJ, nitrate-rich beetroot juice group; PLA, placebo group; F, females; M, males; MAP, mean arterial pressure; Diam., vessel diameter; FMD, flow-mediated dilation; AU, arbitrary units.
Table 4.
Exploratory group × sex × time statistics for non-FMD variables demonstrating training effects.
Table 4.
Exploratory group × sex × time statistics for non-FMD variables demonstrating training effects.
| Variable | BRJ & PLA F/M | Stats |
| DXA lean tissue mass | BRJ: 7F/6M PLA: 7F/6M |
Sex p<0.001 (M>F) Sex × time p=0.478 Sex × supplement × time p = 0.615 |
| VL mCSA | BRJ: 6F/7M PLA: 7F/7M |
Sex p=0.021 (M>F) Sex × time p=0.074 Sex × supplement × time p = 0.456 |
| Hex bar est. 1-RM | BRJ: 6F/7M PLA: 7F/7M |
Sex p<0.001 (M>F) Sex × time p=0.119 Sex × supplement × time p = 0.106 |
| Hex bar strength-end. test | BRJ: 6F/7M PLA: 7F/5M |
Sex p<0.001 (M>F) Sex × time p=0.176 Sex × supplement × time p = 0.492 |
Legend: data are from three-way repeated measures ANOVAs on outcomes demonstrating significant training effects. Although sex differences were noted (M>F, p<0.05), no significant group × sex × time interactions were observed. Abbreviations: BRJ, nitrate-rich beetroot juice group; PLA, placebo group; F, females; M, males; DXA, dual-energy x-ray absorptiometry; VL mCSA, vastus lateralis muscle cross-sectional area; est. 1-RM, estimated maximal strength.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



