
Submitted:
25 July 2025
Posted:
28 July 2025
You are already at the latest version
Abstract
Rosa roxburghii Tratt., a fruit crop known for its high vitamin C content and other nutritional compounds, has not yet been studied for its Auxin Responsive Factor (ARF) gene family, which plays a crucial role in plant growth and fruit development. In the present study, we identified 14 ARF genes (designated as RrARFs) in R. roxburghii, which are distributed across seven chromosomes and grouped into four subfamilies. Cis-acting elements analyses revealed that these genes might be involved in various biological processes, including plant development, light responses, cell cycle regulation, phytohormone responses, and responses to abiotic and biotic stresses. Gene expression analysis demonstrated differential expression of RrARF genes across different tissues and stages of fruit development, with four members showing higher expression during the fruit ripening stages. Furthermore, coexpression analysis identified that RrARF5 was highly coexpressed with RrMDHAR1, a key enzyme involved in vitamin C biosynthesis. Moreover, transactivation assays and transient overexpression experiments confirmed that RrARF5 activates the transcription of RrMDHAR1. The outcomes of this study enhance our understanding of the ARF gene family's role in vitamin C accumulation in R. roxburghii and its broader impact on fruit quality regulation.
Keywords:
R. roxburghii
; ARF
; genome-wide identification
; fruit development
; vitamin C
1. Introduction
Auxin is a significant plant hormone that plays a pivotal role in various plant developmental processes, including embryogenesis, vascular bundle formation, flower and fruit development, and root growth (Du et al., 2012). The transcriptional regulation of several gene families, such as Gretchen Hagen 3 (GH3), Aux/IAA, and Auxin Response Factor (ARF) families, has been shown to influence auxin synthesis and transport, thereby affecting overall plant growth and development (Tiwari et al., 2001; Park et al., 2007; El-Sharkawy et al., 2014). Of these families, ARF plays a significant role in regulating plant growth and development by binding to the TGTCTC DNA motif, mediating hormonal responses, and consequently affecting plant developmental processes (Guilfoyle et al., 1998).
A typical ARF consists of three distinct structural domains. The first is the highly conserved N-terminal B3-like DNA-binding domain (DBD), which is present in all ARFs and is essential for binding to target DNA sequences. The second domain is the C-terminal dimerization domain (CTD), which facilitates the formation of heterodimers with proteins from the Aux/IAA family. The third domain is the variable intermediate region (MR), which can act either as an activation domain (AD) or a repression domain (RD) (Guilfoyle and Hagen, 2007; Zhou et al., 2017; Ye et al., 2020). The regulatory activity of ARF depends on the proper presence and interaction of all three domains (Wang et al., 2007).
The identification of ARF gene families has been accomplished in numerous plant species, including Arabidopsis thaliana L. (Yoko et al., 2005), Fragaria vesca L. (Wang et al., 2018), Vitis vinifera L. (Wan et al., 2014), Citrus sinensis L. (Li et al., 2015), Capsicum annuum L. (Zhang et al., 2017), Solanum lycopersicum L. (Kumar et al., 2011) and Oryza sativa L. (Wang et al., 2007). ARF family genes play critical roles in various developmental and physiological processes in plants, such as embryogenesis (Dolf et al., 2014), lateral root growth (Yoko et al., 2007), leaf expansion (Wilmoth et al., 2010), and leaf senescence (Lim et al., 2010). In addition, ARF gene family members have also been shown to function as crucial regulators in fruit development, ripening, and quality metabolism (Juan et al., 2007; Vriezen et al., 2008; Yuan et al., 2018; Yuan et al., 2019; Yi et al., 2022; Gambhir et al., 2024). MdARF5 was reported to activate the expression of ethylene-related genes, thereby initiating apple fruit ripening (Yue et al., 2020). Similarly, CpARF2 is involved in the regulation of fruit ripening in papaya by promoting the transcriptional activity of CpEIL1 (Zhang et al., 2020). In tomato, SlARF10 and SlARF6A have been demonstrated to facilitate chlorophyll accumulation by activating SlGLK1 expression (Yuan et al., 2018; 2019). However, the role and mode of action of ARF in Vc biosynthesis and metabolism remain largely unknown.
The wild fruit crop R. roxburghii, belonging to the genus Rosa within the family Rosaceae, is widely distributed in the mountainous regions of southwestern and south-central China. Its cultivation has expanded significantly, with an estimated planting area exceeding 30,000 hectares and an annual yield of approximately 2,000–3,000 tons (Lu et al., 2016). The fruit is edible and characterized by a unique aroma, crunchy texture, and a slightly acidic and astringent flavor (Yang et al., 2014). Notably, R. roxburghii is recognized for its exceptional nutritional profile, particularly its high content of ascorbic acid (Vitmain C) (Liu et al., 2016). Ascorbic acid is a vital metabolite for most organisms, serving as an antioxidant and cofactor in various biological processes, including stress resistance, phytohormone biosynthesis, cell division, and cell expansion (An et al., 2007). During plant growth, AsA accumulation is primary regulated by its biosynthesis, metabolism and translocation. So far, four major biosynthetic pathways have been identified in plants: the L-galactose pathway, the inositol pathway, the D-galacturonic acid pathway and the L-gulose pathway. AsA metabolism is further divided into degradation and recycling pathways. Within the L-galactose pathway, L-galactose dehydrogenase (GalDH) functions as a key precursor enzyme, while L-galactono-1,4-lactone dehydrogenase (GalLDH) catalyzes the final step, converting L-galactono-1,4-lactone into AsA. Two additional enzymes, GDP-L-galactose phosphorylase (GGP) and GDP-D-mannose 3’, 5’-differential isomerase (GME), are critical regulatory components of this biosynthetic pathway. AsA also plays an essential physiological role in scavenging reactive oxygen species (ROS), serving as a specific electron donor in the reduction of hydrogen peroxide (H2O2) to H2O and O2. The recycling of oxidized AsA is mediated by monodehydroascorbate reductase (MDHAR) and dehydroascorbate reductase (DHAR), which are crucial for maintaining cellular AsA levels. In previous studies of kiwifruit, the enzymatic activities of GalDH, GalLDH, MDHAR, and DHAR showed a similar trend of AsA accumulation, with significantly positive correlations observed between enzyme activity and AsA content, which suggesting a coordinated regulation (Liao et al., 2021). Since humans and many other animals lack the endogenous capacity to synthesize AsA, it must be obtained from dietary sources like fresh fruits and vegetables (Smirnoff 2018). The pattern of AsA accumulation varies among fruit species: in kiwifruit and apple, peak AsA levels are observed during the young fruit stage; in jujube, during fruit expansion stage; and in strawberry and tomato, during fruit ripening stage (Liao et al., 2023). Previous studies have shown that AsA content increases progressively in R. roxburghii throughout fruit development and ripening process (Zong et al., 2023). Due to these characteristics, R. roxburghii is renowned for its exceptionally high AsA concentration, earning it the title "King of Vitamin C" and becoming a commercially significant fruit crop. Therefore, elucidating the regulatory mechanisms underlying AsA accumulation during R. roxburghii fruit development holds substantial significance for both crop improvement and nutritional enhancement.
Recently, genome-wide data for R. roxburghii have been assembled (Zong et al., 2023). However, despite the ARF gene family is widely studied in other species, its role in R. roxburghii remains unexplored. Given the ARF family’s known roles in fruit development and ripening, it is likely that these genes play a similar role in R. roxburghii. Investigating the presence and function of ARF genes in this species could offer critical insights into the mechanism behind its high ascorbic acid accumulation during the ripening stage, potentially leading to strategies for enhancing the fruit quality of R. roxburghii.
2. Materials and Methods
2.1. Identification of ARF Family Genes in R. roxburghii Genome
The R. roxburghii genome sequence was obtained from the CNSA (CNGB Nucleotide Sequence Archive) with the accession number CNP0004212 (https://db.cngb.org/cnsa/) (Zong et al., 2023). To effectively identify the ARF family members in the R. roxburghii genome, candidate protein sequences containing the Auxin Resp (PF06507) and B3 DNA- binding domain (PF02362) structural motifs were downloaded from the Pfam protein family database (http://Pfam.xfam.org/). Candidate ARF genes were identified from the Rosa roxburghii genome (e-value ≤ e-10) using the “Simple HMM search” in the TBtools software (Chen et al. 2020). Using the NCBI-CDD (https://www.ncbi.nlm.nih.gov/cdd) and SMART (http://smartembl.de/) databases to confirm each putative RrARFs. Using the SMART database (http://SMART.embl-heidelberg.de/), Markov model (HMM) files were utilized to confirm each putative RrARFs. The number of amino acid residues (AA), molecular weight (MW), and theoretical isoelectric point (pI) of the RrARFs proteins were calculated using ExPASy software (http://cn.expasy.org/tools).
2.2. Chromosomal Distribution of RrARFs
The locations of RrARFs on chromosomes were extracted from the genome annotation files. The genes densities across the entire chromosomes were determined and visualized using TBtools software (Chen et al., 2020). For further analysis of conserved motifs and domains, only RrARFs that were anchored to chromosomes were displayed.
2.3. Analysis of Cis-Acting Elements in RrARFs Promoters
The 2000 bp sequences upstream of the initiation codon (ATG) of each R. roxburghii ARF gene (RrARFs) were extracted from the R. roxburghii genome database using TBtools software (Chen et al., 2020). PlantCARE database was used to further analyze the cis-acting regulatory elements presented in the promoter regions of ARF family genes (http://bioinformatics.psb.ugent.be/ webtools/plantcare/html/).
2.4. Phylogenetic Analysis of the R. roxburghii ARF Gene Family
The ARF protein sequences from R. roxburghii (14 sequences), Arabidopsis thaliana (23 sequences), Fragaria vesca (12 sequences), Malus domestica (29 sequences) and Vitis vinifera (19 sequences) were subjected to multiple sequence alignment using the default settings of Clustal W in MEGA 7 software. A phylogenetic tree was then constructed based on the results of the sequence alignment using the neighbor-joining method, in which the bootstrap value was set to 1000 (Kumar et al., 2016). The protein sequences of ARFs from Arabidopsis thaliana, Fragaria vesca, Malus domestica and Vitis vinifera species were obtained from the NCBI database (https://www.ncbi.nlm.nih.gov/). The final phylogenetic tree was edited and visualized using the iTOL online tool (https://itol.embl.de/).
2.5. Gene Structure and Conserved Motifs Characterization
Conserved motifs in RrARFs were identified using the MEME server (https://meme-suite.org/) with the following parameters: a maximum number of 10 motifs, a minimum motif width of 6, and a maximum motif width of 50. We performed structural domain analysis using the NCBI Conserved Domain Database (CDD) to ascertain the types and location of structural domains within all RrARF sequences. In addition, we used TBtools software to visualize the exon/intron structures of the RrARFs genes, as well as the conserved motifs and structural domains of the corresponding RrARF proteins (Chen et al., 2020).
2.6. Gene Expression Analysis of ARF Family Genes
RNA-Seq data from different tissues and organs of R. roxburghii, including root, stem, leaf, flower, and fruits harvested in different developmental and ripening stages were downloaded from the CNSA (https://db.cngb.org/cnsa/) under accession number CNP0004212 (Zong et al., 2023). Data filtering and quality control were performed using fastp software, and the resulting clean data were used for subsequent analyses (Chen et al., 2018). The R. roxburghii genome was used as a reference for read alignment (https://db.cngb.org/cnsa/), and String Tie was used to quantify the aligned reads. Differential expression TPM (Transcripts Per Kilobase of exon model per Million mapped reads) across different tissues was performed using TBtools (Chen et al., 2020). The normalized data were then used to plot expression heat maps with TBtools.
2.7. Weighted Correlation Network Analysis (WGCNA)
The top 50% of genes, ranked according to their expression levels, were selected for WGCNA analysis. After an initial threshold screening, a soft-threshold power of β=7 was selected to strengthen the original proportionality matrix to obtain the derivation of an adjusted neighbor-joining matrix. To better assess the correlation of expression patterns among genes, the neighbor-joining matrices were further transformed into topological overlap matrices (TOMs). Subsequently, gene-shearing analysis was performed using a topological heterogeneity dissimilarity matrix, defined as dissTOM = 1 - TOM. Clustering and module partitioning were then performed, with a minimum module size set at thirty genes (minModuleSize=30). Module merging was based on a similarity threshold of 0.3 (cutHeight= 0.3), and the network type was defined as “signed” (type = “signed” or networkType = “signed”). Visualization of the network was performed using Cytoscape software (Otasek et al., 2019).
2.8. Dual-Luciferase (Dual-LUC) Reporter Assay
To investigate the transcriptional regulation of RrMDHAR1 by RrARF5, the coding sequence of RrARF5 was cloned into the pGreenII 62SK vector to function as the effector construct. The promoter sequence of RrMDHAR1 was cloned into the pGL3 vector as the reporter construct. Leaf protoplasts from N. benthamiana were then prepared following the methodology reported by Pei et al. (2024). The dual luciferase reporter assay kit was used to quantify the transcriptional activity, according to the manufacturer's instructions.
2.9. Accession Numbers
The accession numbers of VvARFs genes in Vitis vinifera were reported in the study of Wan et al. (2014). The accession numbers of AtARFs genes in Arabidopsis thaliana and MdARFs genes in Malus domestica were reported in the study of Luo et al. (2014). The accession numbers of FvARFs genes in Fragaria vesca were reported in the study of Wang et al. (2018).
3. Results
3.1. Identification of ARF Family Genes in R. roxburghii Genome
Following an HMMER search and subsequent analysis using the NCBI Conserved Domain Database (CDD) search tool, 14 ARF genes were identified in R. roxburghii genome (Table 1). Each of the corresponding RrARF proteins contained the characteristic structural domains of ARF proteins (Auxin Response Factor, Auxin Resp). These genes were then sequentially named RrARF1 to RrARF14 based on their chromosomal locations (Figure 1, Table 1).
The amino acid (aa) lengths of the RrARF proteins vary from 570 aa (RrARF11) to 1182 aa (RrARF1), resulting in an average protein length of 791.64 aa. Accordingly, the molecular weights of the RrARF proteins ranged from 62.6 kDa (RrARF11) to 132.8 kDa (RrARF1). The isoelectric points for the 14 RrARF proteins were mainly below 7, except for RrARF6, which exhibited a higher pl of 8.56, suggesting that RrARF proteins may function in diverse cellular microenvironments.
To investigate the evolutionary relationships among RrARF homologous genes, 97 ARF proteins were obtained from five different plants species: R. roxburghii, Arabidopsis thaliana, Malus domestica (apple), Vitis vinifera (grape) and Fragaria vesca (wild strawberry). A phylogenetic tree was constructed using the neighbour-joining method, which is presented in Figure 2. Based on the characteristics of the protein structural domains of the ARF family in Arabidopsis thaliana, our phylogenetic tree classified these ARF homologues into four distinct groups. The distribution of the 14 RrARFs members across these subgroups was as follows: subgroup I (1 member), subgroup II (4 members), subgroup III (5 members) and subgroup IV (4 members).
3.2. Analysis of Conserved Structural Domains and Promoter Sequences of ARF Family Members in R. roxburghii
To gain further insight into the structural characteristics of the RrARF members and to ascertain its potential function, an in-depth examination was conducted on its exon-intron configuration, conserved structural domain, and motifs composition (Figure 3).
To fully confirm the accuracy of the conserved structural domains in RrARF, multiple sequence comparison was performed using ClustalW. The typical ARF structure is characterized by the presence of three conserved structural domains: B3 (DBD), Auxin_responsive, and AUX/IAA superfamily (CTD) (Xing et al., 2011). The results showed that all members of the RrARF family possessed both the B3 structural domain (DBD) and the Auxin-responsive structural domain. However, the AUX/IAA domain (CTD) was lacked in RrARF4, RrARF6, RrARF11, RrARF12, and RrARF14 (Figure 3B; Figure S1).
In addition, the RrARF protein sequence was further analyzed by the online software MEME and 12 motifs ranging from 21 to 50 amino acids in length were identified (Figure 3A; Table S1). Among these motifs, motifs 1, 2, 3, 5 belong to the B3 structural domain (DBD), motifs 6, 8, 11 belong to the auxin response structural domain, and motifs 9, 10 belong to the AUX/IAA structural domain (CTD).
Gene structure analysis revealed that the majority of the coding sequences were interrupted by introns, with members of the same phylogenetic group often sharing similar gene structures (Figure 3C). Interestingly, although RrARF9 and RrARF10 from the group of class III are similar in their protein structures, their exon-intron distributions showed differences. In addition, although RrARF1 and RrARF5 from the Class IV group have the same number of exon-introns, their CDS sizes are different. These differences may indicate that RrARFs have diverse splicing and rich transcripts.
Given the crucial role of cis-acting elements in regulating gene expression, we retrieved the 2-kb promoter sequences of all identified RrARFs genes and submitted them to the PlantCare database for cis-acting elements prediction and analysis. This analysis revealed the presence of hormone-responsive elements (e.g., MeJA, abscisic acid, salicylic acid, gibberellin, etc.), wound-responsive elements, and stress-related elements (e.g., low temperature, drought, etc.) as well as light-responsive elements (Table S2). Notably, the TGA-element and AuxRR-core motifs, which are indicative of a typical auxin response, were only identified in the promoters of RrARF1, RrARF7, RrARF9, and RrARF13, suggesting a putative role for these genes in auxin-mediated growth processes.
3.3. Expression Profiles of RrARFs in Different Tissues and Developmental Stages
To investigate the potential functions of the RrARF gene family in R. roxburghii, we analyzed the spatiotemporal expression profiles using published RNA-seq data from various tissues and different developmental stages of R. roxburghii, including root, stem, leaf, and flower, as well as fruit tissues sampled at five different developmental and ripening stages. The expression profiles of the 14 RrARF family genes in different tissues and during development fruit developmental stages can be divided into four subgroups (Figure 4). The 4 genes of RrARF that make up subgroup I (RrARF5, RrARF11, RrARF12, and RrARF14) were highly expressed in the latter stages of fruit growth and ripening, indicating a potential role in the processes of late fruit maturity and ripening. Two RrARF genes (RrARF1 and RrARF6) that are mostly expressed in leaf and flower tissues make up subgroup II. This suggests that the two genes may have a role in leaf and flower development. Four genes of RrARF (RrARF7, RrARF8, RrARF9, and RrARF10) found in subgroup III displayed strong expression in roots, stems, and young fruits, suggesting that these members may be crucial to the development of both vegetative and early fruit. Subgroup IV contains four RrARF genes (RrARF2, RrARF3, RrARF4, and RrARF13) with specific expression patterns in the root and stem, indicating their vital function in root and stem growth and development.
3.4. Co-Expressed Gene Networks of RrARFs
To investigate the expression patterns of RrARF genes associated with vitamin C synthesis and metabolism, a co-expression network was constructed using weighted gene co-expression network analysis (WGCNA). The expression profiles of 14,310 genes were grouped into 14 modules, with the remaining unclustered genes placed in grey modules (Figure 5A; Figure S5). The number of genes within each module was provided in Table S3. The turquoise module was the most gene-rich, comprising 3,890 genes. In contrast, the module with the lowest number of genes was the cyan module, which contained 49 genes (Table S3).
The correlation between the 14 modules and different periods of fruit development was analyzed (Figure 5B). Among these, the MEblue module showed the highest correlation with fruit-oct stage (r = 0.95, p < 0.000000081), which was a crucial phase for vitamin C synthesis in R. roxburghii. Consequently, the RrARFs genes within the blue module were subsequently identified as the core genes in regulation of Vc biosynthesis. A gene coexpression network was then constructed using these core genes and their coexpression genes related to vitamin C synthesis and metabolism (Figure 5C). As shown in Figure 5C, three ripening-related ARF genes (RrARF5, RrARF11, and RrARF12) showed a high correlation with five genes involved in VC biosynthesis, suggesting a putative regulatory relationship between these ARF and the five VC biosynthesis genes. The weight values between two genes in the gene interaction network are shown in Table S5.
3.5. RrARF5 Involved in VC Biosynthesis by Activating RrMDHAR1 Transcription
The analysis of the public RNA-seq data enabled the examination of the expression of genes associated with vitamin C synthesis and metabolism within the context of a co-expression network. The results demonstrated that the gene Rr602006, named as RrMDHAR1, which encodes monodehydroascorbate reductase (MDHAR) (Figure S2), exhibited high expression levels in R. roxburghii during fruit development (Table S4). Additionally, the expression patterns of RrARF5 and RrMDHAR1 exhibited a high positive correlation during fruit development and ripening (Figure 5C; Figure S4), suggestion a positive regulatory relationship between RrARF5 and RrMDHAR1. Promoter analysis revealed the presence of two TGA elements and an AuxRR core ARF transcription factor regulatory element in the promoter region of RrMDHAR1 (Figure S3). Therefore, it was hypothesized that RrMDHAR1 may be a target of RrARF5. To further investigate their relationship, we performed a dual-luciferase reporter assay in Nicotiana benthamiana leaf protoplasts by co-transfecting the effector construct 35S:RrARF5 with the reporter construct pro:RrMDHAR1 (Figure 6A). The results showed that RrARF5 significantly activated the expression of RrMDHAR1 compared to the control (Figure 6B), thereby supporting the hypothesis that RrARF5 critically involved in Vc biosynthesis by regulating RrMDHAR1 in R. roxburghii fruits.
To further validate the regulation of RrARF5 in Vc accumulation by regulating RrMDHAR1, we constructed 35S:RrMDHAR1-FLAG and 35S:RrARF5-FLAG vectors and transiently expressed these vectors in tomato fruits (Figure 6C). As shown in Figure 6D, ectopic expression of 35S:RrMDHAR1-FLAG resulted in a significant increase in Vc content compared to the control (injected with empty vector). Moreover, ectopic expression of both 35S:RrMDHAR1-FLAG and 35S:RrARF5-FLAG in tomato fruits led to significantly higher content of Vc compared to the fruits only ectopic expression of 35S:RrMDHAR1-FLAG (Figure 6D). These data further indicated that RrARF5 modulates Vc accumulation in fruits by regulation of RrMDHAR1.
4. Discussion
ARF family is a central component of the auxin signaling pathway, regulating a multitude of processes integral to plant development. The ARF transcription factor family has been extensively studied in various plant species, such as Arabidopsis thaliana (Guilfoyle and Hagen, 2007), tomato (Solanum lycopersicum) (Zouine et al., 2014), papaya (Carica papaya L.) (Liu et al., 2015), longan (Dimocarpus longan L.) (Peng et al., 2020), apple (Malus domestica) (Luo et al., 2014), and rice (Oryza sativa) (Wang et al., 2007). Nevertheless, comprehensive investigations of the entire ARF gene family in R. roxburghii have not yet been conducted. In the present study, we identified 14 RrARF transcription factors in R. roxburghii, which were unevenly distributed across seven chromosomes (Figure 1). Phylogenetic analysis of the 14 RrARF proteins showed that they can be classified into four major groups, with each group showing a close evolutionary relationship to the FvARF genes (Figure 2). This observation indicates that ARF genes in these taxa may have originated from a common ancestor, or alternatively, that they may share highly conserved functions (Paterson et al., 2004). Moreover, we identified twelve conserved motifs in RrARF family in R. roxburghii (Figure 3). Although the number of members differs in each phylogenetic group, the motifs within a group are markedly conserved. In addition to the conserved motifs, further research is required to determine whether additional motifs may be associated with novel functions.
The majority of ARFs are comprised of three structural domains with conserved features (Zouine et al., 2014). However, five RrARFs (RrARF4, RrARF6, RrARF11, RrARF12, and RrARF14) lack the Aux/IAA-binding domain (Supplemental Figure S1), which is consistent with the structure of the AtARF3 from the Arabidopsis subfamily that lacks the CTD structural domain. Moreover, the ARF members that lack the Aux/IAA-binding domain can also be found in tomato, rice and orchid species (Kumar et al., 2011; Wu et al., 2011; Bai et al. 2023). Since the Aux/IAA-binding domain is critical for the interaction between ARF and Aux/IAA, the lack of this domain in the five RrARFs may suggest these proteins exert their function in an auxin-dependent manner. It is noteworthy that RrARF7 contains a PABP-1234 superfamily structural domain in addition to the three typical ARF domains (Figure 3). However, the function of this conserved domain and also the function of RrARF7 require further studies. Additionally, the presence of cis-acting elements associated with plant development, abiotic and biotic stresses, light responses, and phytohormone responses in the promoter regions of the RrARF family genes and their distinct expression patterns in different tissues and fruit developmental stages support their diverse functions in plant growth and fruit development. However, the functional significances of these RrARF genes require further investigation.
High vitamin C content is one of the most important quality traits of fruits of R. roxburghii, understanding the regulatory mechanism of vitamin C biosynthesis and metabolism is vital for the improvement of fruit quality of R. roxburghii. In the present study, thorough coexpression analysis, transactivation assays, and transient overexpression assays, we revealed that RrARF5 acts as positive regulator of Vc biosynthesis by activation of the expression of RrMDHAR1, a structure gene which was reported to be correlated with ascorbic acid content (Stevens et al., 2008). These findings contribute to a better understanding of the function of ARF gene family in fruit quality regulation, while also providing a theoretical foundation for further research into the molecular mechanisms underlying ascorbic acid accumulation in R. roxburghii.
It is worth pointing out that although we identified the ARF gene family in R. roxburghii and revealed the role of one ARF member in Vc accumulation, the function of other ARF genes in different plant growth and developmental process remain largely unknown. In the future, we will focus on the development of the plant genetic transformation technology of R. roxburghii and uncovering the biological function of ARF family members.
Acknowledgments
The authors gratefully acknowledge the financial support by the Regional First-class Discipline of Ecology in Guizhou Province (XKTJ[2020]22), the Technology Projects of Guizhou Province, China (No.ZK[2021]090), the Outstanding Young Scientific and Technological Talents of Sichuan Province (2022JDJQ0057).
References
- An, H.; Fan, W.; Chen, L.; Asghar, S.; Liu, Q. Molecular characterisation and expression of L-galactono-1,4-lactone dehydrogenase and L-ascorbic acid accumulation during fruit development in Rosa roxburghii. The Journal of Horticultural Science and Biotechnology 2007, 82, 627–635. [Google Scholar] [CrossRef]
- Bai, Y.; Ma, Y.; Chang, Y.; Zhang, W.; Deng, Y.; Zhang, N.; Zhang, X.; Fan, K.; Hu, X.; Wang, S.; Jiang, Z.; Hu, T. Identification and transcriptome data analysis of ARF family genes in five Orchidaceae species. Plant molecular biology 2023, 112, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas Hannah, R.; Frank Margaret, H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Molecular Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Weijers, D.; Benkova, E.; Jäger, K.E.; Schlereth, A.; Hamann, T.; Kientz, M.; Wilmoth, J.C.; Reed, J.W.; Jürgens, G. Developmental specificity of auxin response by pairs of ARF and Aux/IAA transcriptional regulators. EMBO J. 2005, 24, 1874–1885. [Google Scholar] [CrossRef]
- Du, H.; Wu, N.; Fu, J.; Wang, S.; Li, X.; Xiao, J.; Xiong, L. A GH3 family member, OsGH3-2, modulates auxin and abscisic acid levels and differentially affects drought and cold tolerance in rice. Journal of Experimental Botany 2012, 63, 6467–6480. [Google Scholar] [CrossRef]
- El-Sharkawy, I.; Sherif Sherif, M.; Jones, B.; Mila, I.; Kumar Prakash, P.; Bouzayen, M.; Jayasankar, S. TIR1-like auxin-receptors are involved in the regulation of plum fruit development. Journal of Experimental Botany 2014, 65, 5205–5215. [Google Scholar] [CrossRef] [PubMed]
- Gambhir, P.; Raghuvanshi, U.; Kumar, R.; Sharma Arun, K. Transcriptional regulation of tomato fruit ripening. Physiology and Molecular Biology of Plants 2024, 30, 289–303. [Google Scholar] [CrossRef]
- Guilfoyle Tom, J.; Hagen, G. Auxin response factors. Current Opinion in Plant Biology 2007, 10, 453–460. [Google Scholar] [CrossRef]
- Guilfoyle Tom J, Ulmasov T, Hagen, G. The ARF family of transcription factors and their role in plant hormone-responsive transcription. Cellular and Molecular Life Sciences 1998, 54, 619–627. [CrossRef]
- Juan Carlos, S.; Mariano, F.; Alejandro, A.; José, L. García-Martínez. Effect of gibberellin and auxin on parthenocarpic fruit growth induction in the cv Micro-Tom of tomato. Journal of Plant Growth Regulation 2007, 26, 211–221. [Google Scholar]
- Kumar, R.; Tyagi Akhilesh, K.; Sharma, A.K. Genome-wide analysis of auxin response factor (ARF) gene family from tomato and analysis of their role in flower and fruit development. Molecular Genetics and Genomics 2011, 285, 245–260. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; OuYang, W.; Hou, X.; Xie, L.; Hu, C.; Zhang, J. Genome-wide identification, isolation and expression analysis of auxin response factor (ARF) gene family in sweet orange (Citrus sinensis). Frontiers in Plant Science 2015, 6, 119. [Google Scholar] [CrossRef]
- Liao, G.; Chen, L.; He, Y.; Li, X.; Lv, Z.; Yi, S.; Zhong, M.; Huang, C.; Jia, D.; Qu, X.; Xu, X. Three metabolic pathways are responsible for the accumulation and maintenance of high AsA content in kiwifruit (Actinidia eriantha). BMC Genomics 2021, 22, 13. [Google Scholar] [CrossRef]
- Liao, G.; Xu, Q.; Allan Andrew, C.; Xu, X. L-Ascorbic acid metabolism and regulation in fruit crops. Plant physiology 2023, 192, 1684–1695. [Google Scholar] [CrossRef]
- Lim Pyung, O.; Lee In, C.; Kim, J.; Kim Hyo, J.; Ryu Jong, S.; Woo Hye, R.; Nam Hong, G. Auxin response factor 2 (ARF2) plays a major role in regulating auxin-mediated leaf longevity. Journal of Experimental Botany 2010, 61, 1419–1430. [Google Scholar] [CrossRef]
- Liu, K.; Yuan, C.; Li, H.; Lin, W.; Yang, Y.; Shen, C.; Zheng, X. Genome-wide identification and characterization of auxin response factor (ARF) family genes related to flower and fruit development in papaya (Carica papaya L.). BMC Genomics 2015, 16, 901. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, Q.; Zhang, Y.; Lu, X.; Fu, W.; He, J. Chemical analysis of dietary constituents in Rosa roxburghii and Rosa sterilis fruits. Molecules 2016, 21, 1204. [Google Scholar] [CrossRef]
- Liu, X.; Dinh Thanh, T.; Li, D.; Shi, B.; Li, Y.; Cao, X.; Guo, L.; Pan, Y.; Jiao, Y.; Chen, X. AUXIN RESPONSE FACTOR 3 integrates the functions of AGAMOUS and APETALA2 in floral meristem determinacy. The Plant journal: for cell and molecular biology 2014, 80, 629–641. [Google Scholar] [CrossRef]
- Lu, M.; An, H.; Li, L. Genome survey sequencing for the characterization of the genetic background of Rosa roxburghii Tratt and leaf ascorbate metabolism genes. PLoS One 2016, 11, e0147530. [Google Scholar] [CrossRef]
- Luo, X.; Sun, M.; Xu, R.; Shu, H.; Wang, J.; Zhang, S. Genomewide identification and expression analysis of the ARF gene family in apple. Journal of Genetics 2014, 93, 785–797. [Google Scholar] [CrossRef]
- Otasek, D.; Morris John, H.; Bouças, J.; Pico Alexander, R.; Demchak, B. Cytoscape Automation: empowering workflow-based network analysis. Genome Biology 2019, 20, 185. [Google Scholar] [CrossRef] [PubMed]
- Park Jung-Eun, Park Ju-Young, Kim Youn-Sung, Staswick Paul, E.; Jeon, J.; Yun, J.; Kim Sun-Young, Kim, J.; Lee Yong-Hwan, Park Chung-Mo. Gh3-mediated auxin homeostasis links growth regulation with stress adaptation response in Arabidopsis. The Journal of Biological Chemistry 2007, 282, 10036–10046. [CrossRef] [PubMed]
- Paterson Andrew H, Bowers John E, Chapman, B.A. Ancient polyploidization predating divergence of the cereals, and its consequences for comparative genomics. Proceedings of the National Academy of Sciences 2004, 101, 9903–9908. [CrossRef]
- Pei, Y.; Xue, Q.; Shu, P.; Xu, W.; Du, X.; Wu, M.; Liu, K.; Pirrello, J.; Bouzayen, M.; Hong, Y.; Liu, M. Bifunctional transcription factors SlERF.H5 and H7 activate cell wall and repress gibberellin biosynthesis genes in tomato via a conserved motif. Developmental Cell 2024, 59, 1345–1359. [Google Scholar] [CrossRef]
- Peng, Y.; Fang, T.; Zhang, Y.; Zhang, M.; Zeng, L. Genome–wide identification and expression analysis of auxin response factor (ARF) gene family in Longan (Dimocarpus longan L.). Plants 2020, 9, 221. [Google Scholar] [CrossRef]
- Shen, C.; Yue, R.; Sun, T.; Zhang, L.; Xu, L.; Tie, S.; Wang, H.; Yang, Y. Genome-wide identification and expression analysis of auxin response factor gene family in Medicago truncatula. Frontiers in Plant Science 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Smirnoff, N. Ascorbic acid metabolism and functions: a comparison of plants and mammals. Free Radical Biology and Medicine 2018, 122, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.; Page, D.; Gouble, B.; Garchery, C.; Zamir, D.; Causse, M. Tomato fruit ascorbic acid content is linked with monodehydroascorbate reductase activity and tolerance to chilling stress. Plant, Cell & Environment 2008, 31, 1086–1096. [Google Scholar] [CrossRef]
- Tiwari Shiv, B.; Wang, X.; Gretchen, H.; Guilfoyle Tom, J. AUX/IAA proteins are active repressors, and their stability and activity are modulated by auxin. The Plant Cell 2001, 13, 2809–2822. [Google Scholar] [CrossRef]
- Vriezen Wim, H.; Feron, R.; Maretto, F.; Keijman, J.; Mariani, C. Changes in tomato ovary transcriptome demonstrate complex hormonal regulation of fruit set. The New Phytologist 2008, 17, 60–76. [Google Scholar] [CrossRef]
- Wan, S.; Li, W.; Zhu, Y.; Liu, Z.; Huang, W.; Zhan, J. Genome-wide identification, characterization and expression analysis of the auxin response factor gene family in Vitis vinifera. Plant Cell Reports 2014, 33, 1365–1375. [Google Scholar] [CrossRef]
- Wang, D.; Pei, K.; Fu, Y.; Sun, Z.; Li, S.; Liu, H.; Tang, K.; Han, B.; Tao, Y. Genome-wide analysis of the auxin response factors (ARF) gene family in rice (Oryza sativa). Gene 2007, 394, 13–24. [Google Scholar] [CrossRef]
- Wang, S.; Shi, F.; Dong, X.; Li, Y.; Zhang, Z.; Li, H. Genome-wide identification and expression analysis of auxin response factor (ARF) gene family in strawberry (Fragaria vesca). Journal of Integrative Agriculture 2018, 18, 1587–1603. [Google Scholar] [CrossRef]
- Wilmoth Jill, C.; Wang, S.; Tiwari Shiv B, Joshi Atul D, Hagen, G.; Guilfoyle Thomas J, Alonso Jose M, Ecker Joseph R, Reed, J.; W. NPH4/ARF7 and ARF19 promote leaf expansion and auxin-induced lateral root formation. The Plant Journal 2010, 43, 118–130.
- Wu, J.; Wang, F.; Cheng, L.; Kong, F.; Peng, Z.; Liu, S.; Yu, X.; Lu, G. Identification, isolation and expression analysis of auxin response factor (ARF) genes in Solanum lycopersicum. Plant cell reports 2011, 30, 2059–2073. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Pudake, R.N.; Guo, G.; Xing, G.; Hu, Z.; Zhang, Y.; Sun, Q.; Ni, Z. Genome-wide identification and expression profiling of auxin response factor (ARF) gene family in maize. BMC Genomics 2011, 12, 178. [Google Scholar] [CrossRef]
- Yang, H.; Hu, J.; Huang, X.; Zhou, C.; Li, L.; Fan, M. Risk Assessment of Heavy Metals Pollution for Rosa sterilis and Soil from Planting Bases Located in Karst Areas of Guizhou Province. Applied Mechanics and Materials 2014, 700, 475–481. [Google Scholar] [CrossRef]
- Ye, Y.; Wang, J.; Wang, W.; Xu, L. ARF family identification in Tamarix chinensis reveals the salt responsive expression of TcARF6 targeted by miR167. PeerJ 2020, 8, e8829. [Google Scholar] [CrossRef]
- Yi, S.; Mao, J.; Zhang, X.; Li, X.; Zhang, Z.; Li, H. FveARF2 negatively regulates fruit ripening and quality in strawberry. Frontiers in Plant Science 2022, 13, 1023739–1023739. [Google Scholar] [CrossRef] [PubMed]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 Regulate Lateral Root Formation via Direct Activation of LBD/ASL Genes inArabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Yoko, O.; Overvoorde Paul, J.; Arima, K.; Alonso Jose, M.; Chan, A.; Chang, C.; Ecker Joseph, R.; Hughes, B.; Lui, A.; Nguyen, D.; Onodera, C.; Quach, H.; Smith, A.; Yu, G.; Theologis, A. Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: unique and overlapping functions of ARF7 and ARF19. The Plant Cell 2005, 17, 444–463. [Google Scholar]
- Yuan, Y.; Mei, L.; Wu, M.; Wei, W.; Shan, W.; Gong, Z.; Zhang, Q.; Yang, F.; Yan, F.; Zhang, Q.; Luo, Y.; Xu, X.; Zhang, W.; Miao, M.; Lu, W.; Li, Z.; Deng, W. SlARF10, an auxin response factor, is involved in chlorophyll and sugar accumulation during tomato fruit development. Journal of Experimental Botany 2018, 69, 5507–5518. [Google Scholar] [CrossRef]
- Yuan, Y.; Xu, X.; Gong, Z.; Tang, Y.; Wu, M.; Yan, F.; Zhang, X.; Zhang, Q.; Yang, F.; Hu, X.; Yang, Q.; Luo, Y.; Mei, L.; Zhang, W.; Jiang, C.; Lu, W.; Li, Z.; Deng, W. Auxin response factor 6A regulates photosynthesis, sugar accumulation, and fruit development in tomato. Horticulture Research 2019, 6, 1–16. [Google Scholar] [CrossRef]
- Yue, P.; Lu, Q.; Liu, Z.; Lyu, T.; Li, X.; Bu, H.; Liu, W.; Xu, Y.; Yuan, H.; Wang, A. Auxin-activated MdARF5 induces the expression of ethylene biosynthetic genes to initiate apple fruit ripening. The New Phytologist 2020, 226, 1781–1795. [Google Scholar] [CrossRef]
- Zhang, H.; Cao, N.; Dong, C.; Shang, Q. Genome-wide identification and expression of ARF gene family during adventitious root development in hot pepper (Capsicum annuum). Horticultural Plant Journal 2017, 3, 151–164. [Google Scholar] [CrossRef]
- Zhang, T.; Li, W.; Xie, R.; Xu, L.; Zhou, Y.; Li, H.; Yuan, C.; Zheng, X.; Xiao, L.; Liu, K. CpARF2 and CpEIL1 interact to mediate auxin-ethylene interaction and regulate fruit ripening in papaya. The Plant Journal 2020, 103, 1318–1337. [Google Scholar] [CrossRef]
- Zhou, Z.; Schenke, D.; Miao, Y.; Cai, D. Investigation of the crosstalk between the flg22 and the UV-B-induced flavonol pathway in Arabidopsis thaliana seedlings. Plant, Cell & Environment 2017, 40, 453–458. [Google Scholar]
- Zong, D.; Liu, H.; Gan, P.; Ma, S.; Liang, H.; Yu, J.; Li, P.; Jiang, T.; Sahu Sunil, K.; Yang, Q.; Zhang, D.; Li, L.; Qiu, X.; Shao, W.; Yang, J.; Li, Y.; Guang, X.; He, C. Chromosomal-scale genomes of two Rosa species provide insights into genome evolution and ascorbate accumulation. The Plant Journal 2023, 117, 1264–1280. [Google Scholar] [CrossRef]
- Zouine, M.; Fu, Y.; Chateigner-Boutin Anne-Laure, Mila, I.; Frasse, P.; Wang, H.; Audran Corinne , Roustan Jean-Paul, Bouzayen, M. Characterizationof the tomato ARF gene family uncovers a multi–levels post-transcriptional regulation including alternative splicing. Plos One 2014, 9, e84203.
Figure 1.
Distribution of ARF family genes on Rosa roxburghii chromosomes. The gene density was visualized with the color intensity.
Figure 1.
Distribution of ARF family genes on Rosa roxburghii chromosomes. The gene density was visualized with the color intensity.
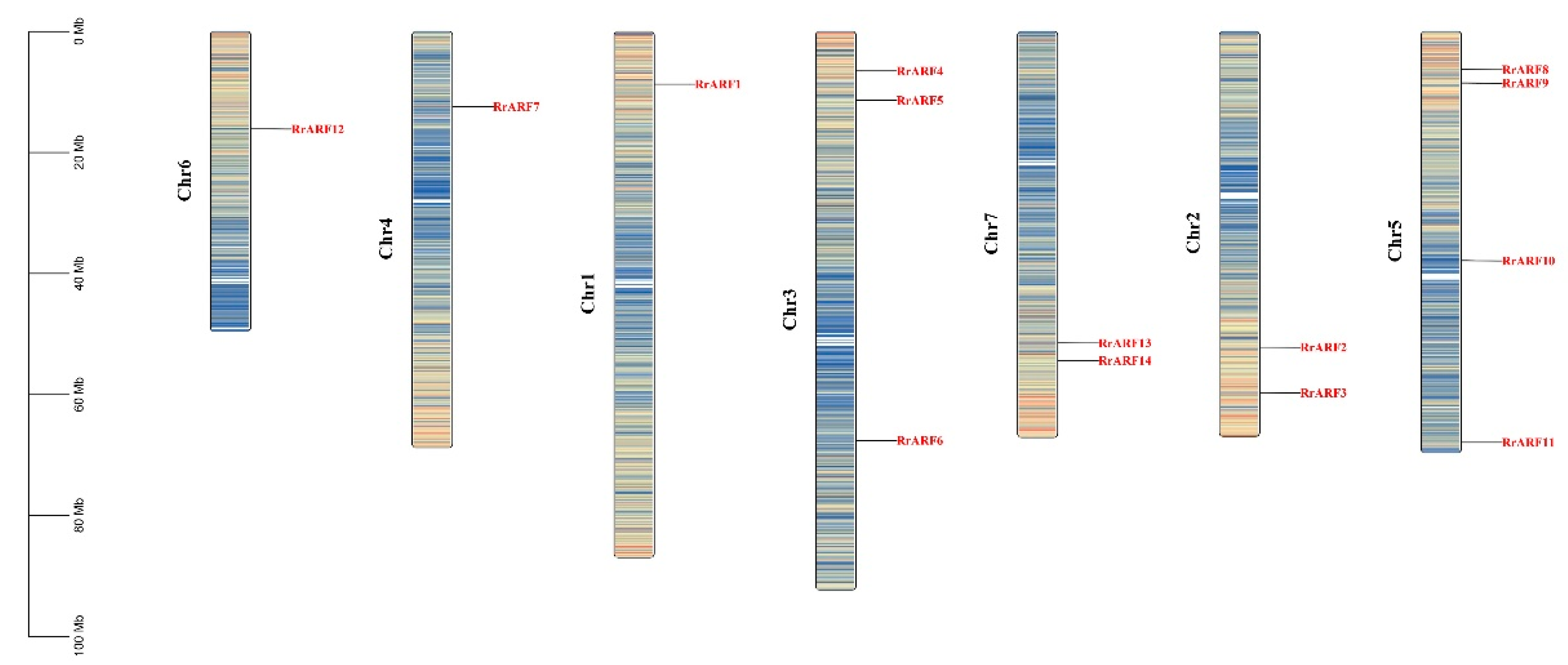
Figure 2.
Phylogenetic analysis of ARF family genes in different plant species. The ARF family genes are divided into four groups, represented by different colors. At, Arabidopsis thaliana; Fv, Fragaria vesca; Md, Malus domestica; Rr, Rosa roxburghii; and Vv, Vitis vinifera.
Figure 2.
Phylogenetic analysis of ARF family genes in different plant species. The ARF family genes are divided into four groups, represented by different colors. At, Arabidopsis thaliana; Fv, Fragaria vesca; Md, Malus domestica; Rr, Rosa roxburghii; and Vv, Vitis vinifera.
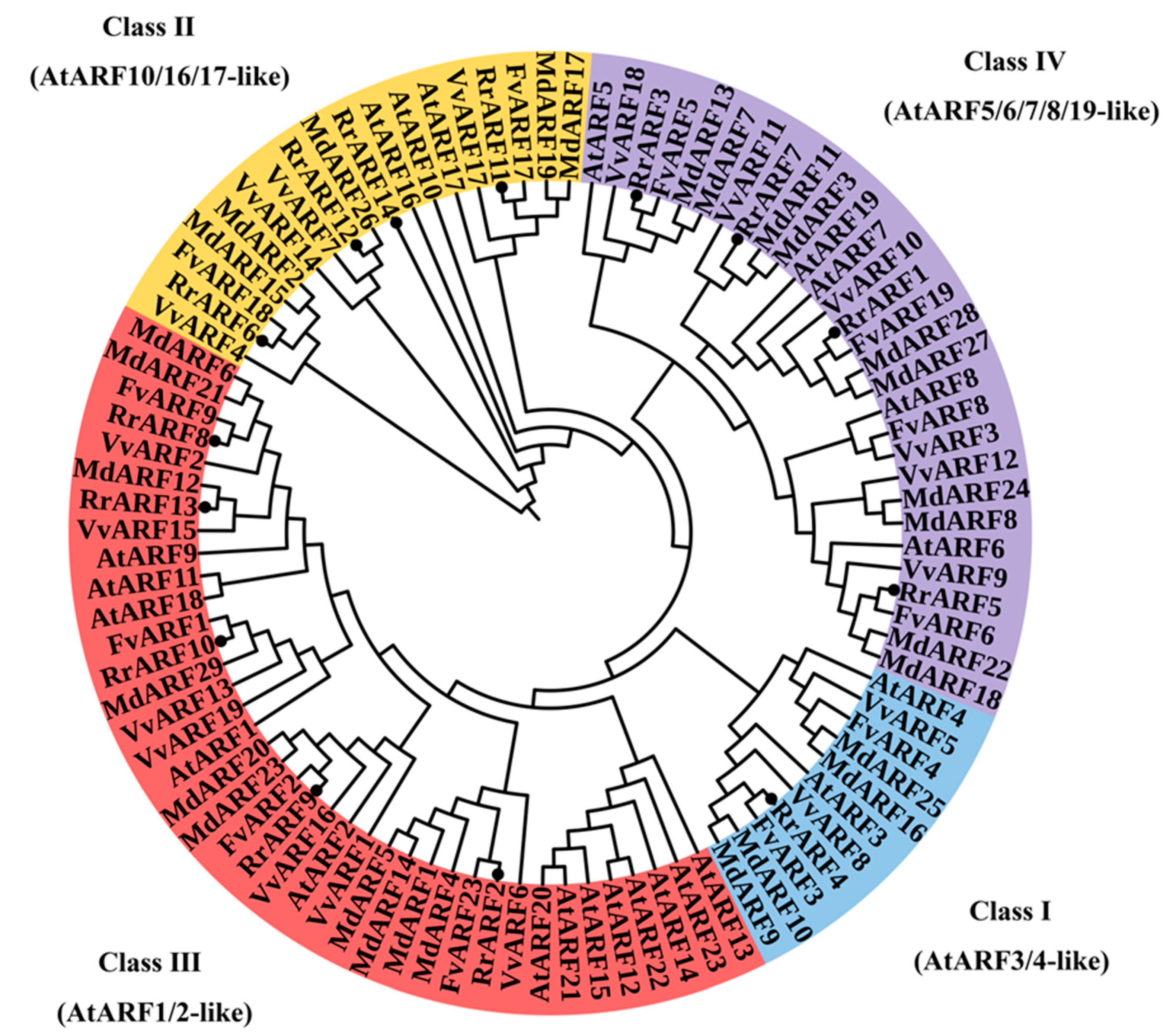
Figure 3.
Conserved motifs, typical ARF structures, and gene structure analysis of Rosa roxburghii ARF family genes. (A) Distribution of conserved motifs in RrARF proteins. (B) Analysis of RrARFs protein domains. (C) Exon/intron structure of the RrARF members. Different colored boxes represent different themes. The scale bars are shown at the bottom.
Figure 3.
Conserved motifs, typical ARF structures, and gene structure analysis of Rosa roxburghii ARF family genes. (A) Distribution of conserved motifs in RrARF proteins. (B) Analysis of RrARFs protein domains. (C) Exon/intron structure of the RrARF members. Different colored boxes represent different themes. The scale bars are shown at the bottom.
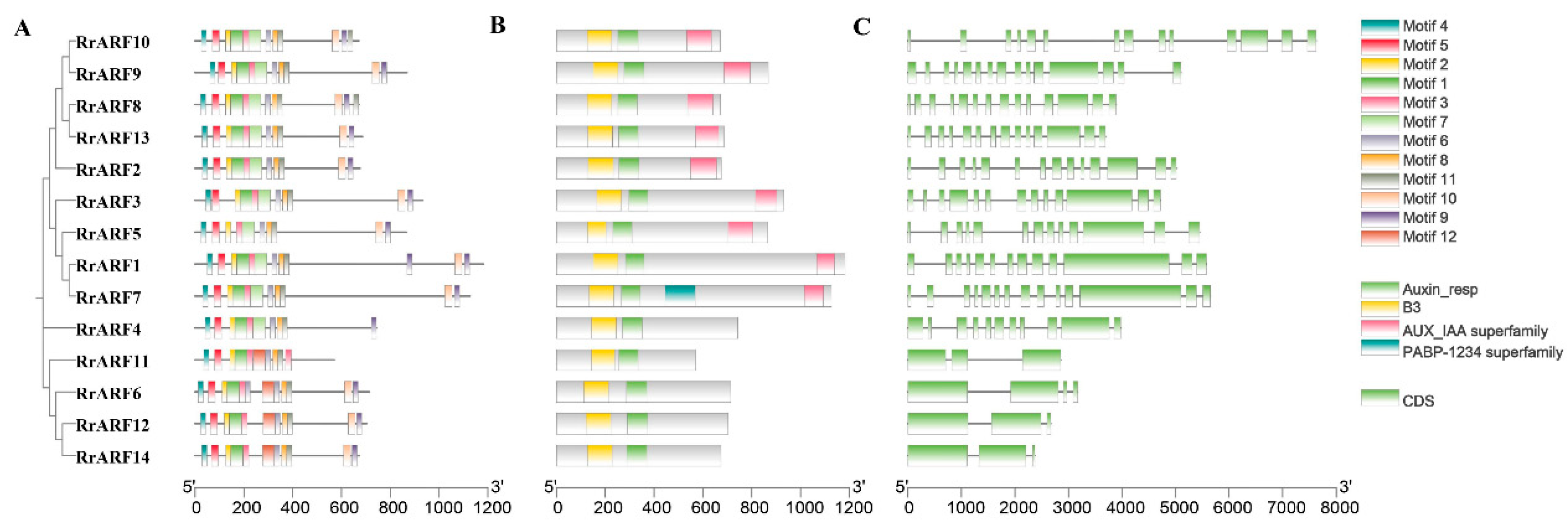
Figure 4.
Expression patterns of RrARFs genes in different tissues. The heatmap showed the expression patterns of RrARFs in various tissues and fruit developmental stages, as derived from transcriptomic data.
Figure 4.
Expression patterns of RrARFs genes in different tissues. The heatmap showed the expression patterns of RrARFs in various tissues and fruit developmental stages, as derived from transcriptomic data.
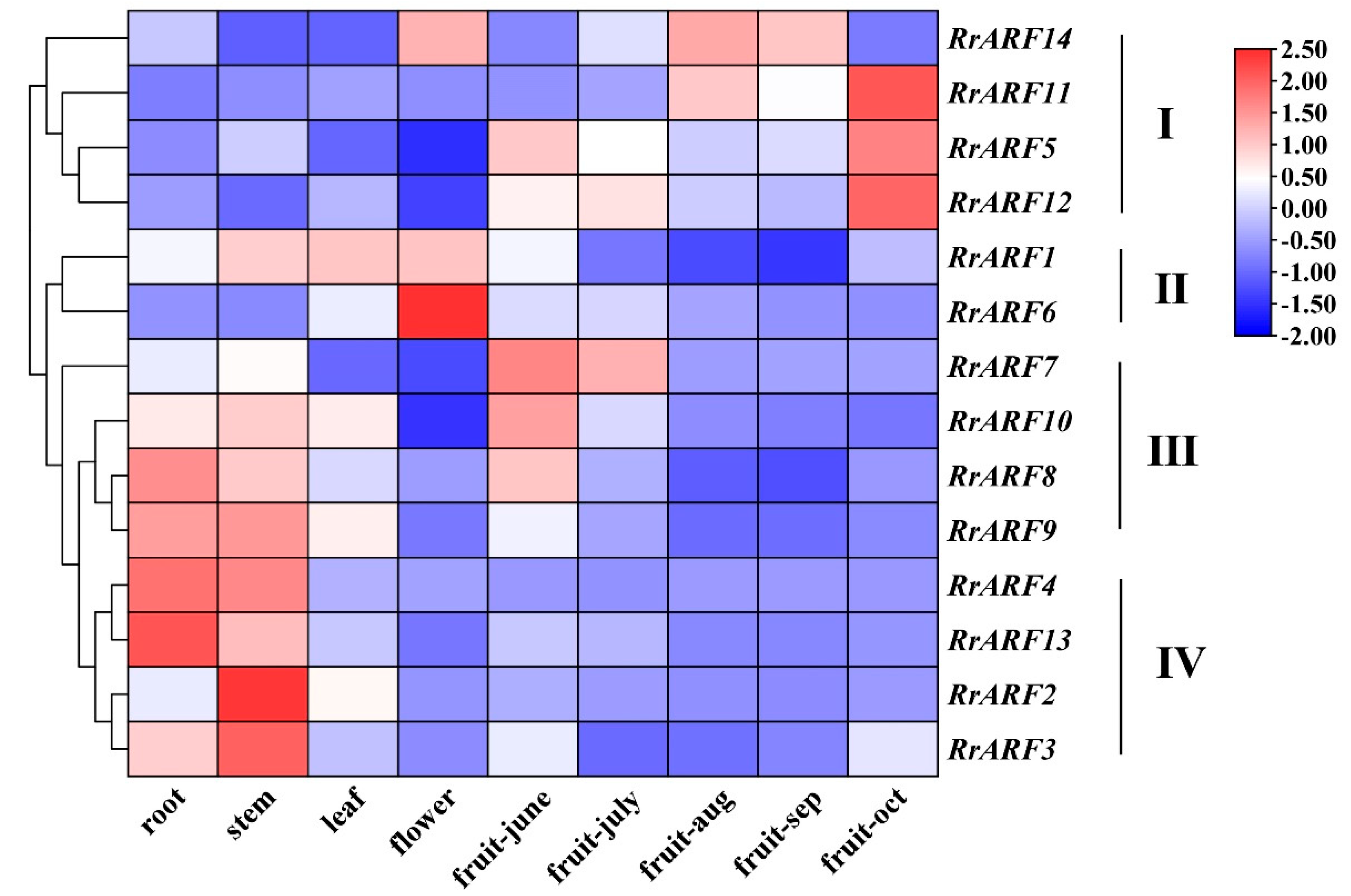
Figure 5.
WGCNA analysis of RrARFs genes and genes related to vitamin C synthesis and metabolism in R. roxburghii (A) Gene clustering dendrogram in WGCNA. The upper part shows the gene clustering dendrogram and the lower part shows the assigned modules with the same colour for the same modules. (B) The relationships between the identified modules and samples at different stages of fruit development were visualised using heatmaps. The numbers displayed within the boxes represent the correlation coefficients and corresponding p-values between the identified modules and the relevant samples. (C) The co-expression network of RrARFs genes and genes related to vitamin C synthesis and metabolism in R. roxburghii within the blue module.
Figure 5.
WGCNA analysis of RrARFs genes and genes related to vitamin C synthesis and metabolism in R. roxburghii (A) Gene clustering dendrogram in WGCNA. The upper part shows the gene clustering dendrogram and the lower part shows the assigned modules with the same colour for the same modules. (B) The relationships between the identified modules and samples at different stages of fruit development were visualised using heatmaps. The numbers displayed within the boxes represent the correlation coefficients and corresponding p-values between the identified modules and the relevant samples. (C) The co-expression network of RrARFs genes and genes related to vitamin C synthesis and metabolism in R. roxburghii within the blue module.
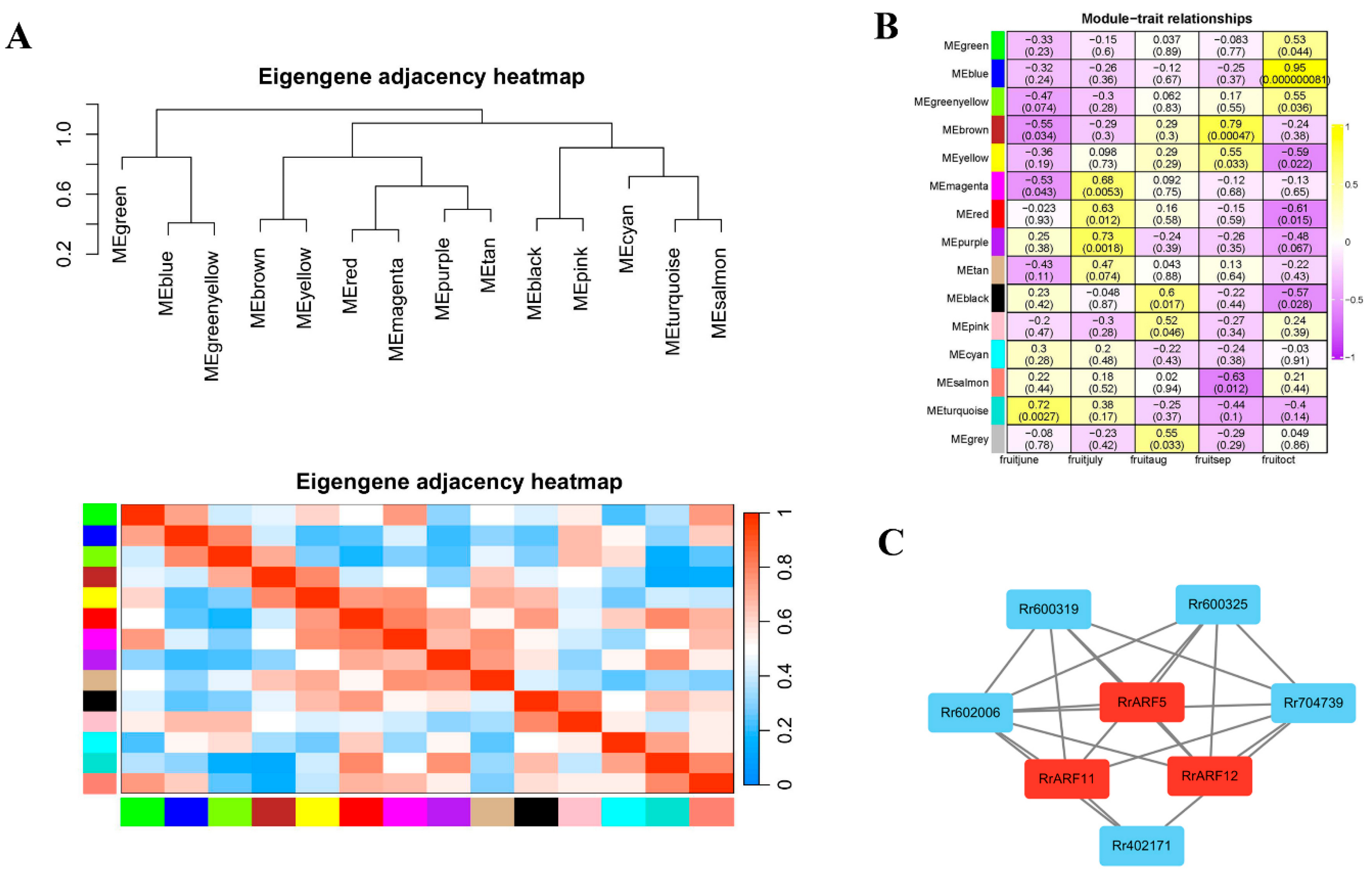
Figure 6.
RrARF5 regulates Vc accumulation by activating the expression of RrMDHAR1 gene. (A) Schematic diagram of effector and reporter constructs used in the transient expression assays. (B) The activation effect of RrARF5 on the transcription of RrMDHAR1. (C) Transcript accumulation of the chimeric RrMDHAR1-FLAG and RrARF5-FLAG in the transient expression in fruits of tomato. Transcript levels were assessed by qRT-PCR in pericarp of injected tomato fruits. Sl-Actin gene as used as internal control. (D) The content of the vitamin C in fruits of control, injected with 35S:RrMDHAR1-FLAG construct, and injected with both 35S:RrMDHAR1-FLAG and 35S:RrARF5-FLAG tomato. Control, injected with empty vector. FW, fresh weight. Values are means ± SD of three replicates.
Figure 6.
RrARF5 regulates Vc accumulation by activating the expression of RrMDHAR1 gene. (A) Schematic diagram of effector and reporter constructs used in the transient expression assays. (B) The activation effect of RrARF5 on the transcription of RrMDHAR1. (C) Transcript accumulation of the chimeric RrMDHAR1-FLAG and RrARF5-FLAG in the transient expression in fruits of tomato. Transcript levels were assessed by qRT-PCR in pericarp of injected tomato fruits. Sl-Actin gene as used as internal control. (D) The content of the vitamin C in fruits of control, injected with 35S:RrMDHAR1-FLAG construct, and injected with both 35S:RrMDHAR1-FLAG and 35S:RrARF5-FLAG tomato. Control, injected with empty vector. FW, fresh weight. Values are means ± SD of three replicates.
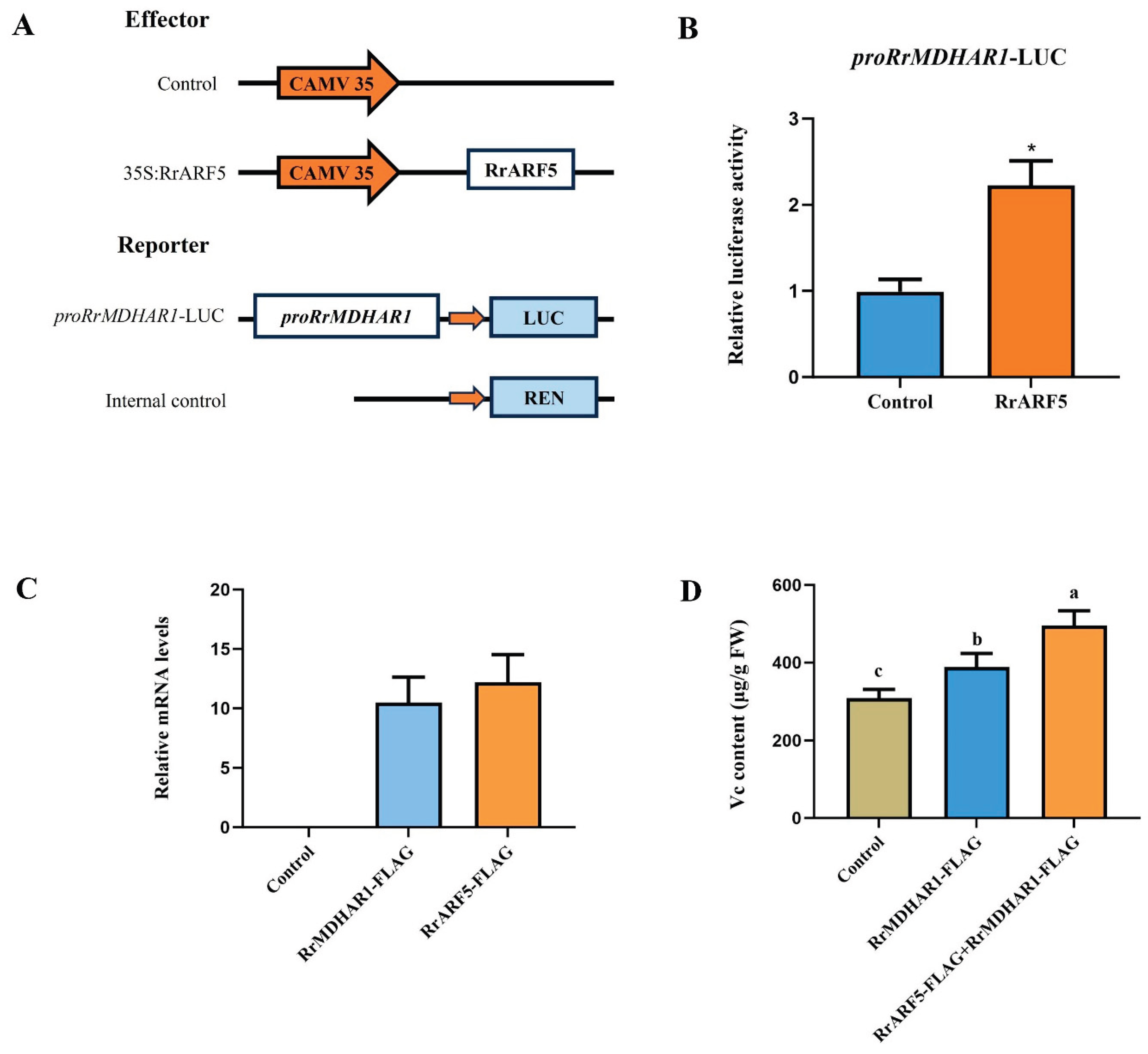

Table 1.
Characterization of RrARF genes in Rosa roxburghii.
| Gene Name |
Gene ID |
CDS Size |
Chromosome Location |
Protein | ||
| Length(aa) | MW(KDa) | pI | ||||
| RrARF1 | Rr100998 | 3549 | Chr1 | 1182 | 132.8 | 6.15 |
| RrARF2 | Rr203848 | 2031 | Chr2 | 676 | 75.5 | 5.83 |
| RrARF3 | Rr204729 | 2799 | Chr2 | 932 | 102.6 | 5.25 |
| RrARF4 | Rr300761 | 2232 | Chr3 | 743 | 81.5 | 6.31 |
| RrARF5 | Rr301243 | 2598 | Chr3 | 865 | 95.3 | 6.09 |
| RrARF6 | Rr304962 | 2145 | Chr3 | 714 | 78.7 | 8.56 |
| RrARF7 | Rr400968 | 3384 | Chr4 | 1127 | 124.7 | 6.24 |
| RrARF8 | Rr500747 | 2019 | Chr5 | 672 | 74.6 | 6.23 |
| RrARF9 | Rr500973 | 2607 | Chr5 | 868 | 96.5 | 6.07 |
| RrARF10 | Rr503508 | 2016 | Chr5 | 671 | 74.4 | 6.10 |
| RrARF11 | Rr505360 | 1713 | Chr5 | 570 | 62.6 | 6.13 |
| RrARF12 | Rr601745 | 2112 | Chr6 | 703 | 77.3 | 6.57 |
| RrARF13 | Rr703521 | 2061 | Chr7 | 686 | 76.0 | 6.56 |
| RrARF14 | Rr703808 | 2025 | Chr7 | 674 | 74.4 | 6.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



