
Submitted:
24 July 2025
Posted:
24 July 2025
You are already at the latest version
Abstract
The pathophysiology of hemorrhoids remains elusive; however, recent research has increasingly focused on the role of genetic and epigenetic mechanisms in this condition, offering prospects for targeted treatments. We conducted a review using PubMed, Embase, and Google Scholar, supplemented by citation searching, to summarize current knowledge of the involvement of genetic and epigenetic mechanisms in hemorrhoids. Our review of 250 papers suggests that several genes, including FOXC2, NOX, NOS, and CALM3, may be responsible for predisposing changes leading to the development of hemorrhoids. These genes have been associated with varicose veins, inflammation, and connective tissue alterations. Additionally, epigenetic mechanisms, particularly those mediated by miRNA, have been implicated in the clinical symptoms of hemorrhoids. While epigenetic regulation may influence inflammation, dilated vessels, and connective tissue degeneration, the exact mechanisms involved in these processes remain unclear. Furthermore, certain predisposing factors for hemorrhoids appear to involve both genetic and epigenetic mechanisms. This knowledge contributes to a better understanding of hemorrhoids and holds promise for developing novel therapeutic approaches.
Keywords:
hemorrhoids
; genetics
; epigenetics
; gene expression
; ncRNA
; microRNA
; miRNA
; inflammation
; targeted therapy
; molecular therap
1. Introduction
Around the anus and the lower rectum, there are located anal cushions, which consist of blood vessels and non-vascular components: epithelium (mucosa or anoderm), connective tissue (elastic and collagenous), a layer of muscular tissue (named muscle of Treitz), and an anchoring system which connects them to surrounding tissue [1,2]. Their purpose is to allow the anus to dilate without damage to the tissue and prevent the stool from leaking in case of increased pressure in the abdomen, which they can achieve thanks to rich vascularity that can be filled or drained of blood, depending on the need. In some cases, leading to pathological changes, they can become swollen, inflamed, and displaced, which results in the development of pathological hemorrhoids, in other words, hemorrhoidal disease [1,2,3,4,5,6,7,8].
We can divide the hemorrhoids into two groups: those placed below the dentate line of the anus – internal hemorrhoids, and those placed above that line – external hemorrhoids. In each of those groups, we can differentiate three main anal cushions: left lateral, right anterior, and right posterior, which make up a total of six anal cushions, three internal and three external [9]. Any of those cushions can develop into pathological hemorrhoids. Other types of hemorrhoidal disease are prolapsed hemorrhoids, a subtype of internal hemorrhoids, in which the enlarged veins of the cushion exit the anus together with the tissue; and thrombosed hemorrhoids, a complication that accompanies mainly external hemorrhoids, however, it can occur with internal ones as well. It can develop due to the formation of blood clots in hemorrhoidal veins and can cause painful swelling of the surrounding tissue [10,11].
Currently, there are three main theories regarding the pathogenesis of hemorrhoids: the varicose vein theory, where hemorrhoids are supposed to originate from abnormally dilated and congested veins of the submucous internal rectal venous plexus [2,8,12,13,14,15]; the vascular hyperplasia theory, where the hemorrhoids develop from hyperplasia of the corpus cavernosum recti [2,16]; and the sliding anal lining theory, which implies that hemorrhoids are a result of degradation of the surrounding connective tissue that causes normal anal cushions to displace, possibly because of an increase in the intra-abdominal pressure, caused by constipation and prolonged straining [2,8,12,13,14,15]. This process can lead to the obstruction of veins via enclosing them by constricting anal muscles [2], which creates increased venous pressure and results in widening and convoluting the vessels, as well as thrombosis – all of that via inflammatory processes featuring leukocytes and macrophages that secrete cytokines, matrix metalloproteinases (MMPs), and oxygen radicals [17,18].
The exact pathophysiology of hemorrhoids remains elusive, as there are no non-ambiguous, well-proved theories. The role of genetics and epigenetics is currently suspected and studied; that is why it seems important to pose a crucial question about the role of those factors in the development of hemorrhoidal disease, summarize and review current knowledge in this field, in hopes of deepening our knowledge about this condition.
Hemorrhoids are the most commonly occurring proctological disease, and it is estimated that every fourth person suffers from this condition. Because of the scale of the problem, it is a major medical issue – data from the National Center for Health Statistics suggest that approximately 10 million people in the United States suffer from hemorrhoids [7], with annual visit count higher than for colon cancer, irritable bowel syndrome, inflammatory bowel disease, ulcerative colitis, or Crohn’s disease [19]. With this many cases, new ways of treatment are constantly in demand [20]. While reviewing gathered studies, we have come across various molecular processes that take part in the development and progression of hemorrhoids. So far, the following mechanisms were reported to occur in hemorrhoidal tissues, which we discuss in detail later in the text: inflammation, changes in vesical pathways, vessel dilation, including that involving an enhanced secretion of NO (nitic oxide), with development of varicose veins, angiogenesis, degeneration of supportive tissue, as well as gene mutations that increased the possibility of development of varicose veins, increased NO secretion or mediated changes in components of connective tissue around the area where hemorrhoids are located. Most, if not all, these mechanisms seem to be a potential base for finding new molecular targeted treatments for hemorrhoids. Targeted therapy based on molecular processes that occur in diseases is not a new concept – numerous works that report on new methods of treatment based on discovered molecular characteristics of a particular disease have been published: in use are cell-free DNA [21], mRNA decay inhibitors [22,23], and anti-cytokines [24]. Some of those methods are based on mechanisms that are similar or corresponding to those uncovered in hemorrhoids – inflammatory bowel disease can be treated with anti-cytokine drugs, targeted at cytokines over expressed in this condition [24], influencing the COX2 and PGE2 expression to After research and some possibly needed modifications, those mechanisms – or similar ones – could be used to treat hemorrhoids as well. However, extensive expertise in the subject is required to develop such a treatment, and in most cases, our current understanding is not sufficient – it seems quintessential to regularly compile current information about known conditions, to accelerate and maximize the potential of treatment-related research.
2. Results
2.1. How do Hemorrhoids Develop?
So far, multiple factors have been claimed to be involved in hemorrhoidal development, especially states that cause an increase in the intra-abdominal pressure: prolonged straining during constipation, obesity, pregnancy, and possibly lifting heavy weights [7,8,12,13,15,25,26,27]. These also include internal agents such as genetic and immune factors, alongside aging, pelvic floor dysfunction, chronic diarrhoea, cirrhosis with ascites, and environmental agents such as diet, sedentary lifestyle, hygiene, alcohol consumption, diverse sexual behaviours (e.g., anoreceptive intercourse), and prolonged sitting [7,8,12,13,15,25,26,27].
Those factors are causing stress to tissues, which respond with inflammatory reaction manifested by vein dilation, destruction of extracellular matrix of hemorrhoidal supporting tissue together with anal subepithelial muscle [7], and, in the end, by the promotion of angiogenic and proliferative factors [28,29], which is in line with all three main theories regarding the development of hemorrhoids: the varicose vein theory, the vascular hyperplasia theory, and the sliding anal lining theory. Occurring activities are regulated by, among others, epigenetic mechanisms, such as non-coding RNA (ncRNA) [27,30,31,32,33]. They function as a messenger when the stress signal occurs and regulate intracellular pathways to trigger a response to that signal. In case of deficiency in ncRNAs, a response to a particular stress signal is much weakened [11,12]. NcRNA are engaged in regulating the expression by engaging in splicing and translation and are a critical factor in the post-transcriptional regulation of transcriptome expression [33]. To the group of ncRNA belong: long noncoding RNA (lncRNA), microRNA (miRNA), which is particularly important in hemorrhoids, circular RNA, ribosomal RNA (rRNA), transfer RNA (tRNA), and small nuclear RNA (snRNA) [34].
MiRNAs are small, single-stranded, non-coding RNA particles that can regulate gene transcription – they bind to the 3′ untranslated region (3'-UTR) of the mRNA molecule transcribed from the target gene, which causes inhibition and induces cleavage of the mRNA by dicer enzymes and silencing the gene expression [35,36,37,38,39,40,41]. One molecule of miRNA can target hundreds of genes, and one gene can be targeted by multiple miRNAs, forming a network that can regulate many pathways. The identification of genes crucial in the development of hemorrhoids is a subject of research, and some of these genes remain to be uncovered [42,43,44,45,46,47].
Below, we discuss currently discovered and documented processes regarding molecular mechanisms occurring in hemorrhoids, with their proven or theorised mechanisms of regulation, in hopes of systematising our knowledge.
2.2. Inflammation
In hemorrhoidal tissue, we can observe very intense inflammation, involving both the dense vascular system and the supportive connective tissue. Inflammation can lead to pathological changes we can observe in hemorrhoids: mucosal ulceration, ischemia, thrombosis, further vessel dilation, and distortion of the smooth muscle layer, together with the surrounding supportive tissue [7,25,26]. This process occurs with the activation of T-lymphocytes, macrophages, neutrophils, monocytes, mast cells, and dendritic cells [48,49], which are responsible for secreting pro- and anti-inflammatory cytokines. The balance, however, shifts very soon to the dominance of pro-inflammatory cytokines, which are highly expressed in hemorrhoids. Among them, there are RANTES (Regulated on Activation, Normal T-cell Expressed and Secreted, CCL-5), TNF-α, and VEGF, which, in addition to proangiogenic function is also increase cell permeability, and as a result, contribute to the inflammation process [49,50,51,52], as well as IL-1β, IL-6, IL-8, IL-17, and IFN-γ – those cytokines were found to be significantly overexpressed in hemorrhoids. IL-10, an anti-inflammatory cytokine, is under-expressed [53] (see Table 1.). A study has shown that some substances help to downregulate the expression of those cytokines in fibroblasts and macrophages of hemorrhoid tissues, which leads to a reduction of inflammation, and, due to VEGF’s primary function, to a decrease in vascular density, which is increased in hemorrhoids [48,54].
In addition to that, genes of CGRP (calcitonin gene-related peptide), substance P (SP), together with TRPV1 (transient receptor potential cation channel, vanilloid receptor 1), which are involved in regulating the expression of cytokines in blood cells [55,56,57], were found to take part in regulating that expression in serum in the case of hemorrhoids as well [58]. Some substances were proven to inhibit the secretion of pro-inflammatory cytokines in serum via downregulating those genes, which helped to ease hemorrhoid symptoms, suggesting that those genes are up-regulated in hemorrhoids and take part in their development.
Other than those factors, increased levels of COX-2 were detected in hemorrhoidal tissues, and they were secreted by fibroblasts and macrophages [28,53,59]. COX-2 is a molecule involved in the production of prostaglandins, main pain mediators, as well as supporting the inflammation process. It is likely to be the cause of pain in hemorrhoid disease [48,60].
Zhou et al. (2025) have found increased miR-770 levels in hemorrhoid tissues. This miRNA promotes polarization of MΦ1 macrophages, which results in observed increased infiltration of these macrophages in hemorrhoid-affected tissues. MiR-770 suppresses RYBP, a member of the PRC1 complex – a complex responsible for inhibiting gene expression, and thus weakens the transcriptional repression. A result of this process is upregulation of genes encoding inflammatory genes like nuclear factor-κB-dependent (NF-κB), IL-1β, and TNF-α, further fuelling the inflammation. [61].
In summary, proinflammatory cytokines, especially VEGF, are more expressed in hemorrhoids than in normal cells and play an important part in their pathogenesis, mainly angiogenesis, and maintaining the inflammatory state, which leads to progression of the disease and clinical symptoms.
2.3. The Role of Vesicles
Some studies regarding the role of miRNA in hemorrhoids found that miRNAs were up- or downregulated in hemorrhoids and are possibly targeting genes, based on available databases, involved in various vesicular pathways (see Table 2.) [27,30].
According to Song et al. [30], miRNAs upregulated in hemorrhoid tissues were miR-375, miR-215-5p, miR-192-5p, miR-143-3p, miR-187-3p, miR-194-5p, miR-145-5p, miR-490-3p, miR-145-3p, while downregulated were miRNAs in hemorrhoids were: miR-376b-3p, miR-34a-5p, miR-152-3p, let-7c-5p, miR-107, miR-517a-3p, miR-517b-3p, miR-1307-5p, miR-190a-5p, miR-378a-5p, miR-708-3p, miR-450a-5p, miR-450a-5p, miR-30e-5p, miR-532-5p. Two of these, miRNA-133b and miRNA-133a-3p, shared 32 of the possible target genes, and the rest of the miRNAs had independent potential target genes.
Genes possibly targeted by them, for both up- and downregulated miRNAs, are involved mainly in cell composition and protein binding. The upregulated miRNAs’ most prominent target, with the most changes in gene expression, is the endocytosis pathway, and the downregulated miRNAs focus the most on genes belonging to the synaptic vesicle pathway. This leads to the higher expression of endocytosis pathway genes and the lower expression of synaptic vesicle pathway genes, which was identifiable in tests and suggests that the development of hemorrhoids may be linked to an imbalance between the expression of the endocytosis and synaptic vesicle cycle pathway genes, generated by changes in levels of miRNAs in hemorrhoidal cells. Those two pathways in hemorrhoidal tissue might be responsible for regulating the infiltration of inflammatory cells, proliferation of vascular endothelial cells, oedema of interstitial cells, or other processes, although that is still a subject for further research [30].
Differences in miRNA expression were found in extracellular vesicles as well [27]. Extracellular vesicles are membranous structures originating from cell membranes [62]. They have a role in cancer progression, metastasis [63], wound healing [64], angiogenesis [65], and immunoregulation [66]. They also function as messengers between cells [67], carrying proteins, lipids, and RNAs [68,69]. Hemorrhoidal extracellular vesicles were found to be carrying different molecules than vesicles derived from healthy tissues, with the most difference being in the content of miRNAs. There were 245 upregulated miRNAs found in hemorrhoidal extracellular vesicles, and within them ten most prominent were: miR-6741-3p, miR-6834-3p, miR-4254, miR-6804-3p, miR-744-3p, miR-8485, miR-299-5p, miR-4636, miR-3175, and miR-4658. Those miRNAs seem to influence the transcription process and target genes involved in protein kinase activity, transcriptional activity, and ubiquitin-protein function, and are most active in proximity to cell junctions. Signalling pathways that are upregulated by them the most are the MAPK (mitogen-activated protein kinases) signalling pathway, axon guidance, and the Ras signalling pathway. They also upregulate AMPK, PI3K-Akt, Hippo, and Wnt signalling pathways and autophagy, though at a lower level than the first three mentioned [27].
The most upregulated miRNA found, miR-6741-3p, is highly likely to be responsible for combining the 3′-UTR of UBQLN1, a gene coding ubiquilin-1, a protein participating in the process of ubiquitination in proteasome-dependent protein degradation, via ubiquitin ligases [70]. An enzyme from this family, HERC3, was established to attenuate nuclear factor-κB-dependent signaling, which mediates inflammatory and immune reactions by inducing its ubiquitination and proteasomal degradation [71]. It means that the upregulation of miR-6741-3p results in lowering the inactivation level of NF-κB and increasing its activity, and consequently, in enhancing local and systemic inflammation, which are prominent in hemorrhoids [27,72].
Some of the upregulated miRNAs in hemorrhoidal extracellular vesicles target the MAPK (mitogen-activated protein kinases) signalling pathway, which is active in physiological and pathological cell proliferation, in carcinogenesis [73] and angiogenesis [74,75], and may enhance inflammation as well [76]. The obtained results suggest that the MAPK pathway may be one of the factors increasing angiogenesis and partially inflammation in hemorrhoids.
Number of downregulated miRNAs found in extracellular vesicles from hemorrhoidal tissues was 202, from which the ten most significant were: miR-548t-5p, miR-323b-5p, miR-1322, miR-3928-5p, miR-346, miR-4704-5p, miR-1913, miR-876-3p, miR-4460, miR-892a. These are the molecules that possibly modulate transcriptional activator activity, also by regulating transcription, and they are the most active around the plasma membrane. Pathways targeted by them are mostly Rap1 signalling pathway, and synthesis, secretion, and action of parathyroid hormone, proteoglycans in cancer, as well as, to a lesser extent, MAPK, cAMP, and Wnt signalling pathways.
The expression pattern of miRNA may vary between proper hemorrhoidal tissues and extracellular vesicles emerging from them, since not always the same miRNAs are present at the same levels between them [27].
2.4. Nitric Oxide and Varicose Veins
Nitric oxide (NO) is a molecule produced in many different cells, and it contributes to vascular dilatation and the development of varicose veins [77]. It was found to be overly present, together with two Nitric Oxide Synthases (NOS): endothelial (eNOS) and neuronal (nNOS) [78] in hemorrhoids, while asymmetric dimethylarginine, a molecule that inhibits NOS, was underexpressed in those tissues [5,8,12,13,14,15,32,79,80]. High expression of NOS in hemorrhoids leads to excessive amounts of NO being synthesised, and that causes an increase in blood flow and twisting of veins, which adds to the varicose veins theory of pathogenesis of hemorrhoids.
2.5. Angiogenesis
Hemorrhoids occur with increased angiogenesis, which leads to high vascular density in the pathological tissue, dilatation of blood vessels (which in the end might contribute to the development of varicose veins), and signs of oedema, sometimes with thrombosis [5,8,12,13,14,15,32,80]. In some works, it was suggested that thrombosis might be one of many factors inducing angiogenesis (neovascularization) [89,90], possibly via the increase in expression of VEGF [91,92], and that might be the case in hemorrhoids as well [80]. Nuclei of endothelial cells appear enlarged, and vascular endothelial cell markers vWF, CD31, and CD34, as well as endoglin, are expressed much more in hemorrhoids than in normal tissues, which further shows that there is intensified vascular proliferation present [31,80]. Endoglin is a glycoprotein overexpressed in endothelial cells, an accessory receptor for TGF-β, and a marker for angiogenesis, due to its appearance in different tissues with increased vascular proliferation [93,94,95,96]. Prominent levels of VEGF and VEGFR2, proangiogenic factors, were also identified in hemorrhoids, and they intensify the angiogenesis process as well [80,97].
On top of that, signalling pathways responsible for inhibiting angiogenesis were found to be significantly less active in hemorrhoid cells, leading to increased vascular proliferation [31,32].
MiR-143-3p was significantly downregulated in hemorrhoid tissues, especially in severe cases. Its downregulation increases expression of vascular markers, such as CD31, vWf, and VEGFR2, cell proliferation and migration, and enhances apoptosis, and it is a potential key regulator in angiogenesis in hemorrhoid progression and postoperative wound healing [98].
On chromosome 14, there is located a genomic imprinted region – DLK1-DIO3 [43,44,45,46]. It contains, among others, microRNAs. Four of those miRNAs, miR-412-5p, miR-422-5p, miR-432-5p, and miR-1185-1-3p, were found to be expressed differently in healthy and hemorrhoidal tissues, and in particular, miR-412-5p and miR-1185-1-3p were found to be significantly underexpressed in hemorrhoidal tissue [32].
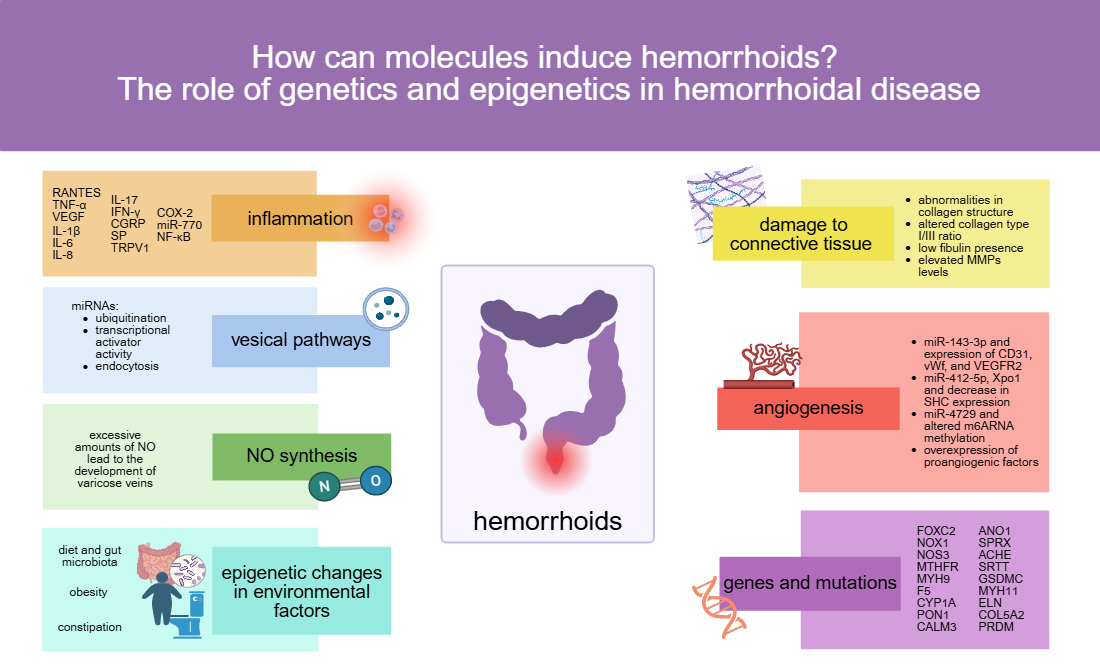
MiR-412-5p targets the gene of Exportin1 (Xpo1) (see Figure 1) [32], a nuclear protein in endothelial cells that is responsible for exporting proteins, such as p53, p21, FOXO, PI3K/AKT, Wnt/β-catenin, AP-1, and NF-κB, from the nucleus to the cytoplasm [91,92,99,100]. Under-expression of miR-412-5p leads to overexpression of Xpo1 in hemorrhoidal endothelial cells [32], since miRNAs silence gene expression, and are the reason for augmented translocation of p53 to the cytoplasm. The p53 protein in the nucleus is responsible for regulating the cell cycle by increasing the expression of certain genes. In case of endothelial cells in hemorrhoids, those genes are the SHC genes: they are divided after transcription into three subtypes: p46SHC, p52SHC, and p66SHC [101,102,103]. The p66SHC has a role in the Ras signalling pathway [101,102,104,105], inducing apoptosis and aging [101,102,103,106,107,108,109,110]. Lack of p53 in the nucleus of hemorrhoidal endothelial cells results in a decrease in SHC expression, especially p66SHC, blocking the p53-p66SHC-p16 pathway. The exact connection between that pathway and hemorrhoids is not clear yet, but it appears to inhibit and aggravate the cell cycle control, causing proliferation of endothelial cells and angiogenesis, which is a factor in the formation of pathological hemorrhoids [32].
Another factor contributing to angiogenesis in hemorrhoids is changes in methylation of RNA N-6 methyladenosine (m6ARNA) (see Figure 2). Enzymes from the m6A methyltransferase complex, described also as m6A writers, METTL14 and METTL3, form a heterodimeric complex, with METTL14 acting as a switch for METTL3’s activity, and they catalyse the methylation of m6A RNA [33,111,112,113,114]. Other components in that complex are auxiliary cofactors such as WTAP, VIRMA, RBM15/15B, ZC3H13, and HAKAI, which help with binding to the target RNA (first three), achieving the correct localization of the whole enzymatic complex (ZC3H1) [33]. For the further fate and function of m6A RNA, the readers are responsible: they improve its stability, regulate splicing, increase its transcriptional and translational activity, and promote carcinogenesis and invasion as well [33,112,114,115,116,117].
Ten miRNAs are predicted to target METTL14 expression, and from these, only one, miR-4729, was significantly low in hemorrhoid tissues. This miRNA, when it is up-regulated, inhibits METTL14 expression in endothelial cells, and, as a result, inhibits m6A RNA methylation as well. It was later observed that in this case, proliferation, as well as overall vascular endothelial cell function, were inhibited. One gene was found to be especially inhibited in cells with miR-4729 overexpression – TIE1. TIE1 is a tyrosine kinase that plays a critical role in angiogenesis and blood vessel stability, as well as tissue remodelling and inflammation; it can be found in endothelial cells [118,119,120,121,122,123]. This gene is related to VEGFA, VEGFR2, vWF, and CD31 and can induce vWF and CD31 expression by activating VEGFA/VEGFR2 receptor ligands, which promotes angiogenesis. They form together a signalling pathway called the TIE1/VEGFA signal molecular loop, in addition to the previously known TIE1/TIE2/VEGFR2 signalling pathway. TIE1 was found to be significantly under-expressed in cells with high miR-4729 content, which suggests that this miRNA is a key factor regulating TIE1 expression, via influencing METTL14 expression, and then, as a result, m6A RNA methylation. In hemorrhoids, miR-4729 is under-expressed, which causes an increase in the synthesis of METTL14 and then an intensification in methylation of m6ARNA, especially TIE1 [31], which was found to be present in those cells at very high levels [123]. As a result, we can observe vascular hyperplasia, together with an increased expression of endothelial markers such as CD31, which is distinct in hemorrhoids.
2.6. A Role of Estrogen in Angiogenesis?
In some cases of hemorrhoids in women, we can detect the presence of estrogen nuclear receptors (ERα) in the tissues of the anal canal [81,82]. They bind estrogens, which activate a signalling pathway leading to the downregulation of miR-424-5p. The higher the level of estrogens, the more ERα are present [83]. Low levels of miR-424-5p and elevated levels of ERα cause upregulation of VEGF, and, as a result, angiogenesis, and edema (see Figure 3). The opposite – decrease in expression of VEGF – happens when miR-424-5p is overexpressed. This miRNA’s predicted target is the ESR1 gene, whose product regulates the transcription of estrogen-inducible genes [84], such as VEGF [85]. Downregulation of miR-424-5p in hemorrhoids, caused by estrogens activating the ERα, leads to an increase in levels of ESR1 and then, as a result, VEGF, which promotes angiogenesis and edema in hemorrhoids in women. There is also some evidence of the role of estrogens in angiogenesis in other tissues – multiple studies across the years have reported that they are involved in the recovery of arterial endothelium after damage [86], possibly via promoting VEGF expression, as it is another function of estrogens, not only in hemorrhoids, but also in other tissues: uterine and vascular altogether [87,88], suggesting that their influence on hemorrhoids is not a solitary case.
2.7. Degeneration of Connective Tissue
The stroma of hemorrhoids consists of connective tissue, with collagen and elastic fibres cross-linked with each other, providing tensile strength, elasticity, integrity, and stability to the tissue; a layer of smooth muscle (Treitz’s muscle) and blood vessels [2,124].
Abnormalities in collagen structure, especially collagen I and III, participate in the development of hemorrhoids – whether due to its degradation or incorrect structure, which leads to poor mechanical endurance and stability. Conducted studies have shown that patients with hemorrhoids had many more defects in collagen structure than healthy patients, even those advanced in age. The same studies reported that collagen levels and collagen type I/III ratio were significantly reduced in hemorrhoids in comparison to normal anal cushions, with significant predominance of collagen III [125], which results in less cross-linking and poor mechanical endurance, since the mature collagen I provides more tensile strength than the less mature collagen III [126,127]. This suggests that the occurrence of hemorrhoids is associated with early and severe collagen degradation, as well as alterations in collagen type ratio. These changes may result from genetic, metabolic, or environmental factors, potentially leading to the early onset of the disease. However, the precise background of these alterations remains unclear and warrants further study [126,127].
Changes in expression of elastic connective tissue components have been reported as well, in the fibulin family of extracellular matrix proteins. Fibulin-3 and fibulin-5 were found to be under-expressed in hemorrhoids [53,128,129,130,131,132]. Fibulin-5 stabilizes and organizes elastic fibres in extracellular matter and inhibits angiogenesis [133], whereas fibulin-3 maintains the stability of the basal membrane, stabilizes the extracellular matrix [134] and inhibits matrix metalloproteinases (MMPs) – hydrolytic enzymes responsible for the degradation of the extracellular matrix [135]. Low levels of those fibulins may result in damage to elastic fibres, and consequently, to the structure of connective tissue, predisposing to hemorrhoids, since the components of that tissue are linked to one another. Some medicines are successful in increasing their expression [53]. This suggests that some epigenetic mechanism may be regulating that expression, although the specific pathways are yet to be determined.
Levels of various MMPs have been found to be elevated in patients with hemorrhoids [26,27,53,136,137,138]. In grade I and II of the disease (early stages), those enzymes were MMP-1, MMP-2, and MMP-3; in grade III – MMP-3, MMP-7, MMP-8 and MMP-9; and in grade IV, MMP-2, MMP-8 and MMP-9 [26,139], with very high expression of MMP-9, together with NGAL (neutrophil gelatinase-associated lipocalin, lipocalin-2) – a marker for neutrophil activation, and consequently for inflammation; NGAL and MMP-9 levels being particularly elevated in thrombosed and prolapsed hemorrhoids [26]. In addition to elastic fibers, these enzymes can target and degrade other components of the extracellular matrix, such as collagen. This process leads to inflammation and severe damage to the supportive tissue of the anal canal, as well as disruption of regenerative processes. Consequently, it contributes to the progression of hemorrhoidal disease, particularly by promoting the development of prolapsed hemorrhoids, as the damaged supportive tissue is no longer able to prevent the prolapse of anal structures [53,137,138,139]. MMP-9 plays an additional role – it promotes angiogenesis by damaging the basal lamina of endothelium, which stimulates the release of VEGF [140,141]. Prominent levels of those enzymes might be a consequence of under-expression of fibulin-3 in hemorrhoidal tissues, as well as activation of macrophages and lymphocytes, which secrete MMPs, but that may not be the only mechanism; further studies must be conducted to determine those mechanisms.
2.8. Genes Significant in the Development of Hemorrhoids
Along with the development of our knowledge about molecular biology, especially in the matter of genetics and epigenetics, the significant role of genes and changes associated with them as risk factors for hemorrhoidal disease has been widely considered. Numerous studies have been conducted across the span of years to research the involvement of genetics in the development of hemorrhoids and a possible genetic susceptibility to their occurrence, and we have aimed to gather and summarize them in our paper. The results are displayed in a table below (see Table 3.). The mentioned genes and their mutations seem to be predisposing to hemorrhoids whether by impairing the function and structure of the epithelium and connective tissue, including the smooth muscles, changing the organisation of the extracellular matter (ECM), limiting neuromuscular motility, as well as enhancing the synthesis of some proteins. Those changes can be and were previously associated with gastrointestinal, neuroaffective, and cardiovascular conditions [142].
2.9. Possible Epigenetic Factors in the Development of Hemorrhoids
There are numerous environmental and lifestyle-dependent factors considered to be connected to the pathogenesis of hemorrhoids. Some of those factors, discussed below, are either known or suspected to cause epigenetic changes in the cells, which can lead to changes that are a direct cause for hemorrhoids (see Figure 4).
2.9.1. Diet and Gut Microbiota
Dietary changes are known to change the composition of gut microbiota. An exemplary, low-fiber diet was reported to induce changes in microbiota [178], as well as it was observed that hemorrhoids occur more often in patients with a deficiency of diet-sourced fiber [7,179]. Although the change of microbiota components is not conditioned by epigenetic mechanisms, it can induce epigenetic modifications in the organism. Diet compounds, together with their bacterial metabolites, can influence gene expression via removing or providing compounds that are substrates for epigenetic modifications [180]. Among bacterial species that influence the levels of such proteins are butyrate-producing species such as Faecalibacterium prausnitzii, Roseburia, and Coprococcus species; species that can produce isothiocyanates, such as Escherichia coli, Bacteroides thetaiotaomicron, Enterococcus, Peptostreptococcus, and Bifidobacterium, or those that metabolize ellagitannins to urolithins – e.g., Clostridium coccoides, Actinobacterium, Lactobacillus, or – again – Bifidobacterium. Those changes were observed to be involved in the pathogenesis of various diseases, such as colon cancer, type 2 diabetes, obesity, and possibly constipation [180,181,182], with the last two being among risk factors for hemorrhoidal disease. Furthermore, there is a theory that implicates a correlation between the alteration of gut microbiota and the development of hemorrhoids [183], although no research has been conducted to prove that theory yet.
2.9.2. Obesity
Across many different studies, obesity has been named as a factor increasing the probability of developing hemorrhoids [184,185,186,187]. It is suspected that the pathogenesis of that process includes the increase in intra-abdominal pressure (that is the case in pregnancies as well) due to enlarged amounts of adipose tissue exerting pressure on veins, especially, which causes them to widen and create varicose veins, present in hemorrhoids [188,189,190]. Adiposity also escalates the inflammatory process in tissues by stimulating the release of the proinflammatory cytokines – among others adipokines, TNF-α, IL-6, and other interleukins – as well as angiotensinogen, PAI-1 (plasminogen activator inhibitor-1), MMPs, TSP-1 (thrombospondin), VEGF, oxygen radicals, reactive lipids, and ECM components with chemotactic activity [191,192,193], which accelerates the inflammation [187]. A study conducted by Huang et al. (2023) has highlighted a causal association between hemorrhoids and high body weight [190], and it was in conformity with observational studies on this subject conducted across different populations [187,194,195,196]. Obesity is known to be conditioned by genetic and epigenetic modifications [197,198,199], so development of hemorrhoids may also be linked to those changes; however, that connection has not been proved yet.
2.9.3. Constipation
Constipation has been known as one of the crucial factors for hemorrhoids for a long time and is still being recognised in modern research [185,200,201,202]. It seems to be an effect of chronic stress subjected to anal tissues during defecation, causing them to stretch and degenerate, or increasing intra-abdominal pressure, leading to an impairment of venous circulation, promoting dilatation of veins and development of varicose veins and hemorrhoids [2,7]. There are various causes for constipation, and amongst them is irritable bowel syndrome (IBS) – a disorder of the brain-gut axis, resulting in abdominal pain and alteration of frequency of bowel movements. Recently, there have been published some works hinting at the possible association of IBS with hemorrhoids [142,203,204,205]. Development of hemorrhoids was observed in multiple cases of IBS across several independent studies. IBS is multifactorial, and it is mediated, among others, by genetic and epigenetic mechanisms [206]. Although a direct connection has not yet been established, hemorrhoids can likely occur because of constipation during genetic and epigenetic modifications in IBS. It is also theorized that changes in gut microbiota, possibly caused by a low-fiber diet, can participate in inducing constipation [178].
3. Discussion
Constipation has been known as one of the crucial factors for hemorrhoids for a long time and is still being recognised in modern research [185,200,201,202]. It seems to be an effect of chronic stress subjected to anal tissues during defecation, causing them to stretch and degenerate, or increasing intra-abdominal pressure leading to impairment of venous circulation, promoting dilatation of veins and development of varicose veins and hemorrhoids [2,7]. There are various causes for constipation, and amongst them is irritable bowel syndrome (IBS) – a disorder of the brain-gut axis, resulting in abdominal pain and alteration of frequency of bowel movements. Recently, there have been published some works hinting at the possible association of IBS with hemorrhoids [142,203,204,205]. Development of hemorrhoids was observed in multiple cases of IBS across several independent studies. IBS is multifactorial, and it is mediated, among others, by genetic and epigenetic mechanisms [206]. Although a direct connection has not yet been established, hemorrhoids can likely occur because of constipation during genetic and epigenetic modifications in IBS. It is also theorized that changes in gut microbiota, possibly caused by a low-fiber diet, can participate in inducing constipation [178].

Table 4.
A summary table presenting molecular factors involved in the development of hemorrhoids.
| Molecular Components Altered in Hemorrhoids. | |||
| State | Upregulated | Downregulated | Mutated or Altered* Genes |
| Molecule | Pro-inflammatory cytokines, especially VEGF | Anti-inflammatory cytokines | FOXC2 |
| CGRP, SP, and TRPV1 | Inhibitory pathways of angiogenesis | MTHFR | |
| COX-2 | Vesical miRNAs modulating synaptic vesicle pathways and transcriptional activator activity | MYH9 | |
| Vesical miRNAs modulating endocytosis, transcription, protein kinase activity, and ubiquitination | miR-412-5p, leading to deregulation of the cell cycle | CYP1A | |
| NOS3 | miR-4729, leading to overexpression of TIE1 and angiogenesis via METTL14 regulation | PON1 | |
| NOS and NO | miR-424-5p, caused by estrogens, leading to increased expression of VEGF | ANO1 | |
| vWF, CD31, CD34, endoglin | Fibulin-3 and fibulin-5 | SPRX* | |
| VEGF, VEGFR2 | Collagen levels, collagen I/III ratio | SRTT* | |
| MMPs | ·· | GSDMC* | |
| NOX1 | ·· | COL5A2 | |
| CALM3 | ·· | ·· | |
| ACHE | ·· | ·· | |
| MYH11 | ·· | ·· | |
| ELN | ·· | ·· | |
| PRDM | ·· | ·· | |
* The phrase “altered genes” refers to genes that were found to be altered in hemorrhoidal cells, in comparison to normal cells, but the nature of that change is not specified.
As for other factors suspected to influence the development of hemorrhoids, such as a low-fiber diet, obesity, and constipation, it is conceivable that genetic and epigenetic mechanisms may play a role, albeit indirectly. Dietary changes can trigger epigenetic alterations and modifications in gut microbiota, potentially leading to gut motility issues and predisposing individuals to constipation. Constipation, a long-established primary risk factor for hemorrhoids, is not inherently linked to genetic and epigenetic changes, but certain forms, like irritable bowel syndrome (IBS), may be associated with them. Given the higher incidence of hemorrhoids in patients with IBS, there may be an indirect connection between the two conditions. Obesity is also known to be influenced by epigenetic mechanisms, as excessive adipose tissue exerts pathological pressure on veins, initiating the pathological mechanisms underlying hemorrhoids. While these connections have not been conclusively established through scientific methods and are primarily based on observation or association studies, they represent an area that warrants further research. All those mechanisms are possible targets for the treatment of hemorrhoids, which may help in either relieving the symptoms, stopping the aggravation of the current condition, or even preventing its occurrence in patients who are highly likely to develop hemorrhoids, based on their medical record and risk factors. A deeper understanding of the molecular pathological mechanisms of hemorrhoids will allow us to work on more precise ways of pharmacological treatment before the state of the patient requires surgical intervention.
An evident limitation of our study is the lack of validation in a clinical setting. The studies analysed in our review encompass both clinical and in vitro investigations. We refrain from assessing the clinical relevance of the research included in our review concerning the utility of the discussed genes or their products as therapeutic targets. Instead, we view our study as an impetus for researchers to delve further into the molecular aspects of hemorrhoids, aiming to discover new avenues for targeted and much-needed treatment. However, our study is also constrained by the number of databases reviewed, the keywords searched, and the necessity to limit our search on PubMed and Google Scholar to 200 articles per query, owing to the vast quantity of available records. While we believe our selection of databases is adequate, it remains possible that we did not retrieve and review 100% of the available research on this topic. Additionally, there is limited research on this topic, as the molecular components have often only been investigated in a single original study. Moreover, the validation of these results solely within the research team raises the possibility of inconsistency, which we, as reviewers, lack the means to confirm, representing yet another limitation. To conclusively validate the information gathered in this review, further studies are imperative.
4. Materials and Methods
Our narrative review was conducted following the SANRA (Scale for the Assessment of Narrative Review Articles) checklist [207], as it is the standard for narrative reviews. We aimed for our work to meet all six quality criteria of these guidelines to ensure the highest standards of scientific rigor and reliability (see Figure 5).
For the search, we have used PubMed, Embase, and Google Scholar, as this combination of scientific databases was shown to be sufficient for reviews in peer-reviewed studies [208,209]. We searched for data about genetic and epigenetic factors contributing to the development of hemorrhoids. Search was conducted using a combination of keywords, one combination for one search: (“hemorrhoids”) AND (“genetics” OR “epigenetics” OR “genetic factors” OR “epigenetic factors” OR “factors”). Terms were searched as a combination of the word “hemorrhoids” and one word from the second group, e.g., “hemorrhoids genetics”. The initial search revealed the following number of records, combined from all keywords: n=24039 for PubMed, n=887 for Embase, and n=94420 for Google Scholar, finding a total of 119 346 records. Based on the initial search, we have decided to limit the number of publications qualified for screening to 200 per search. This limited the number of results to 2887, from which we identified papers that met our criteria.
In the first screening, we have looked for papers that have mentioned hemorrhoids in the title, keywords, or abstract, together with genetics in a broad sense, gene expression, specific epigenetic mechanisms, or changes in mechanisms that are likely to be mediated by epigenetics. We tried to minimize errors by double screening of papers by two independent researchers. Moreover, after identifying papers, to minimize errors, we have utilized first-pass screening, including only titles and keywords, and second-pass screening, including abstracts, full text was read during the second pass if needed.
Based on the criteria mentioned above, we identified, while also excluding the duplicates, 83 articles. Articles included in the study were only original papers about performed studies, meta-analyses, as well as reviews, which suited the topic of our review. At this point, all types of research papers other than meta-analyses, reviews, and original studies were excluded. We performed a secondary review of articles we first identified, which led to the exclusion of five more articles due to their irrelevance to the topic of this study after reviewing their text, lack of full text in a language understandable to us, or inability to be retrieved. If a paper was deemed irrelevant by one author but relevant by another, it was up for discussion until a unanimous decision was reached. The inclusion criteria stayed the same as during the search for papers.
The next step in our research was citation searching. We have searched previously identified papers and added more articles to our base. Finally, 250 articles were included for data extraction and analysis.
We divided articles from our base into four sub-groups: those concerning only genetics, only epigenetics, possibly epigenetics, and a mixed one, including both genetics and epigenetics. One part of our team was responsible for reviewing articles and writing the text related to the involvement of genetics in hemorrhoids, based on groups: genetics and mixed genetics, and epigenetics, while the other focused solely on epigenetic mechanisms by reviewing articles from groups epigenetics, possibly epigenetics, and mixed genetics and epigenetics.
Author Contributions
Conceptualization, B.P. and M.M.; methodology, B.P., M.M.; data analysis, B.P., O.S.; writing—original draft preparation, B.P., O.S.; writing—review and editing, B.P., O.S., M.M., S.W., Z.D.; figure design, K.D.; supervision, S.W., Z.D.; project administration, B.P., Z.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors would like to express sincere gratitude to Professor Bartosz Kempisty for providing critical feedback on the manuscript.
Graphic abstract: Created in BioRender. Mazurek, M. (2025) https://BioRender.com/5wak9ao.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| FOXC2 | Forkhead Box C2 |
| NOX | NADPH Oxidase |
| NOS | Nitric Oxide Synthase |
| CALM3 | Calmodulin 3 |
| miRNA | microRNA |
| mRNA | messenger RNA |
| DNA | Deoxyribonucleic Acid |
| COX2 | Cyclooxygenase-2 |
| RYBP | Ring1 and YY1-binding protein |
| PRC1 | Polycomb Repressive Complex 1 |
| NF-κB | Nuclear Factor Kappa-light-chain-enhancer of Activated B Cells |
| PGE2 | Prostaglandin E2 |
| ncRNA | non-coding RNA |
| lncRNA | long non-coding RNA |
| rRNA | ribosomal RNA |
| tRNA | transfer RNA |
| snRNA | small nuclear RNA |
| UTR | Untranslated Region |
| MMP | Matrix Metalloproteinase |
| RANTES | Regulated on Activation, Normal T-cell Expressed and Secreted |
| CCL-5 | Chemokine (C-C motif) Ligand 5 |
| TNF-α | Tumor Necrosis Factor Alpha |
| VEGF | Vascular Endothelial Growth Factor |
| IL | Interleukin |
| IFN-γ | Interferon Gamma |
| CGRP | Calcitonin Gene-Related Peptide |
| SP | Substance P |
| TRPV1 | Transient Receptor Potential Vanilloid 1 |
| MAPK | Mitogen-Activated Protein Kinase |
| AMPK | AMP-Activated Protein Kinase |
| PI3K | Phosphoinositide 3-Kinase |
| UBQLN1 | Ubiquilin-1 |
| HERC3 | HECT and RLD Domain Containing E3 Ubiquitin Protein Ligase 3 |
| ECM | Extracellular Matrix |
| vWF | von Willebrand Factor |
| CD | Cluster of Differentiation |
| TGF-β | Transforming Growth Factor Beta |
| Xpo1 | Exportin 1 |
| PI3K/AKT | Phosphoinositide 3-Kinase/Protein Kinase B |
| AP-1 | Activator Protein 1 |
| SHC | Src Homology 2 Domain Containing |
| m6A | N6-methyladenosine |
| METTL14 | Methyltransferase Like 14 |
| METTL3 | Methyltransferase Like 3 |
| WTAP | Wilms Tumor 1 Associated Protein |
| VIRMA | Vir-Like m6A Methyltransferase Associated |
| RBM15/15B | RNA Binding Motif Protein 15/15B |
| ZC3H13 | Zinc Finger CCCH-Type Containing 13 |
| TIE1 | Tyrosine Kinase with Immunoglobulin Like and EGF Like Domains 1 |
| VEGFA | Vascular Endothelial Growth Factor A |
| ESR1 | Estrogen Receptor 1 |
| ERα | Estrogen Receptor Alpha |
| IBS | Irritable Bowel Syndrome |
| PAI-1 | Plasminogen Activator Inhibitor-1 |
| TSP-1 | Thrombospondin-1 |
| NGAL | Neutrophil Gelatinase-Associated Lipocalin |
| SANRA | Scale for the Assessment of Narrative Review Articles |
References
- Margetis, N. Pathophysiology of Internal Hemorrhoids. Ann Gastroenterol 2019, 32, 264. [Google Scholar] [CrossRef] [PubMed]
- Thomson, W.H.F. The Nature of Haemorrhoids. Br J Surg 1975, 62, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Loder, P.B.; Kamm, M.A.; Nicholls, R.J.; Phillips, R.K.S. Haemorrhoids: Pathology, Pathophysiology and Aetiology. Br J Surg 1994, 81, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Pata, F.; Sgró, A.; Ferrara, F.; Vigorita, V.; Gallo, G.; Pellino, G. Anatomy, Physiology and Pathophysiology of Haemorrhoids. Rev Recent Clin Trials 2021, 16, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.; Tietze, P.H.; Ramakrishnan, K. Anorectal Conditions: Hemorrhoids. FP Essent 2014, 419, 11–19. [Google Scholar] [PubMed]
- Burkitt, D.P.; Graham-Stewart, C.W. Haemorrhoids--Postulated Pathogenesis and Proposed Prevention. Postgrad Med J 1975, 51, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Lohsiriwat, V. Hemorrhoids: From Basic Pathophysiology to Clinical Management. World Journal of Gastroenterology : WJG 2012, 18, 2009. [Google Scholar] [CrossRef] [PubMed]
- Sugerman, D.T. olmach Hemorrhoids. JAMA 2014, 312, 2698–2698. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Rivero, S. Article Commentary: Hemorrhoids: Diagnosis and Current Management. https://doi.org/10.1177/000313480907500801 2009, 75, 635–642. [CrossRef]
- Picciariello, A.; Rinaldi, M.; Grossi, U.; Verre, L.; De Fazio, M.; Dezi, A.; Tomasicchio, G.; Altomare, D.F.; Gallo, G. Management and Treatment of External Hemorrhoidal Thrombosis. Front Surg 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Morgado, P.J.; Suárez, J.A.; Gómez, L.G.; Morgado, P.J. Histoclinical Basis for a New Classification of Hemorrhoidal Disease. Dis Colon Rectum 1988, 31, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, D. Clinical Practice. Hemorrhoids. N Engl J Med 2014, 371, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, W.A. Office Management of Hemorrhoids. Am J Gastroenterol 2018, 113, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Lohsiriwat, V. Treatment of Hemorrhoids: A Coloproctologist’s View. World J Gastroenterol 2015, 21, 9245–9252. [Google Scholar] [CrossRef] [PubMed]
- Rakinic, J.; Poola, V.P. Hemorrhoids and Fistulas: New Solutions to Old Problems. Curr Probl Surg 2014, 51, 98–137. [Google Scholar] [CrossRef] [PubMed]
- Stelzner, F. Die Hämorrhoiden Und Andere Krankheiten Des Corpus Cavernosum Recti Und Des Analkanals. DMW - Deutsche Medizinische Wochenschrift 1963, 88, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Ganz, R.A. The Evaluation and Treatment of Hemorrhoids: A Guide for the Gastroenterologist. Clin Gastroenterol Hepatol 2013, 11, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Parish Budiono, B.; Adi Presetyo, S.; Riwanto, I.; Sulistyaningsih, S.; Nugroho, E.A. Graptophyllum Pictum Extract in the Treatment of Experimental Hemorrhoids: Effects on Vascular Leakage and Matrix Metalloproteinase-9 Levels. Open Access Maced J Med Sci 2021, 9, 1785–1789. [Google Scholar] [CrossRef]
- Peery, A.F.; Crockett, S.D.; Barritt, A.S.; Dellon, E.S.; Eluri, S.; Gangarosa, L.M.; Jensen, E.T.; Lund, J.L.; Pasricha, S.; Runge, T.; et al. Burden of Gastrointestinal, Liver, and Pancreatic Diseases in the United States. Gastroenterology 2015, 149, 1731–1741.e3. [Google Scholar] [CrossRef] [PubMed]
- Etzioni, D.A.; Beart, R.W.; Madoff, R.D.; Ault, G.T. Impact of the Aging Population on the Demand for Colorectal Procedures. Dis Colon Rectum 2009, 52, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Műzes, G.; Bohusné Barta, B.; Szabó, O.; Horgas, V.; Sipos, F. Cell-Free DNA in the Pathogenesis and Therapy of Non-Infectious Inflammations and Tumors. Biomedicines 2022, Vol. 10, Page 2853 2022, 10, 2853. [Google Scholar] [CrossRef] [PubMed]
- Carrard, J.; Ratajczak, F.; Elsens, J.; Leroy, C.; Kong, R.; Geoffroy, L.; Comte, A.; Fournet, G.; Joseph, B.; Li, X.; et al. Identifying Potent Nonsense-Mediated MRNA Decay Inhibitors with a Novel Screening System. Biomedicines 2023, 11, 2801. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Mailliot, J.; Schaffitzel, C. Nonsense-Mediated MRNA Decay Factor Functions in Human Health and Disease. Biomedicines 2023, Vol. 11, Page 722 2023, 11, 722. [Google Scholar] [CrossRef] [PubMed]
- Vebr, M.; Pomahačová, R.; Sýkora, J.; Schwarz, J. A Narrative Review of Cytokine Networks: Pathophysiological and Therapeutic Implications for Inflammatory Bowel Disease Pathogenesis. Biomedicines 2023, Vol. 11, Page 3229 2023, 11, 3229. [Google Scholar] [CrossRef] [PubMed]
- Chong, P.S.; Bartolo, D.C.C. Hemorrhoids and Fissure in Ano. Gastroenterol Clin North Am 2008, 37, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.; Gallelli, L.; Grande, R.; Amato, B.; De Caridi, G.; Sammarco, G.; Ferrari, F.; Butrico, L.; Gallo, G.; Rizzuto, A.; et al. Hemorrhoids and Matrix Metalloproteinases: A Multicenter Study on the Predictive Role of Biomarkers. Surgery 2016, 159, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, Y.; Ma, X.; Ge, X.; Deng, Y. Identification of the MicroRNA Alterations in Extracellular Vesicles Derived from Human Haemorrhoids. Exp Physiol 2023, 108, 752–761. [Google Scholar] [CrossRef] [PubMed]
- ASHWIN PORWAL, G.C.K.G.B. and R.B. Herbal Medicine AnoSpray Suppresses Proinflammatory Cytokines COX-2 and RANTES in the Management of Hemorrhoids, Acute Anal Fissures and Perineal Wounds. Available online: https://www.spandidos-publications.com/10.3892/etm.2021.11009 (accessed on 16 February 2024).
- Yoon, S.O.; Park, S.J.; Yun, C.H.; Chung, A.S. Roles of Matrix Metalloproteinases in Tumor Metastasis and Angiogenesis. J Biochem Mol Biol 2003, 36, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Zhou, H.; Lu, H.; Luo, C.; Wang, C.; Wang, Q.; Peng, Y.; Xin, Y.; Liu, T.; Yang, W. Aberrant Expression for MicroRNA Is Potential Crucial Factors of Haemorrhoid. Hereditas 2020, 157, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhou, H.; Lu, H.; Luo, C.; Wang, Q.; Peng, Y.; Yang, W.; Xin, Y. MiR-4729 Regulates TIE1 MRNA M6A Modification and Angiogenesis in Hemorrhoids by Targeting METTL14. Ann Transl Med 2021, 9, 232–232. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lu, H.; Luo, C.; Song, C.; Wang, Q.; Peng, Y.; Xin, Y.; Liu, T.; Yang, W. MiR-412-5p Targets Xpo1 to Regulate Angiogenesis in Hemorrhoid Tissue. Gene 2019, 705, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wei, W.; Wang, H.; Dong, J. N6-Methyladenosine-Sculpted Regulatory Landscape of Noncoding RNA. Front Oncol 2021, 11, 743990. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and Their Integrated Networks. J Integr Bioinform 2019, 16. [Google Scholar] [CrossRef] [PubMed]
- Van Rooij, E. The Art of MicroRNA Research. Circ Res 2011, 108, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.K.L.; Sharp, P.A. MicroRNA Functions in Stress Responses. Mol Cell 2010, 40, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA Expression Profiles Classify Human Cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Borghini, A.; Andreassi, M.G. Genetic Polymorphisms Offer Insight into the Causal Role of MicroRNA in Coronary Artery Disease. Atherosclerosis 2018, 269, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Singh, D.; Ganju, L.; Kumar, B. MicroRNA in Gastrointestinal Cell Signalling. Inflammopharmacology 2018, 26. [Google Scholar] [CrossRef] [PubMed]
- Creugny, A.; Fender, A.; Pfeffer, S. Regulation of Primary MicroRNA Processing. FEBS Lett 2018, 592, 1980–1996. [Google Scholar] [CrossRef] [PubMed]
- Mendell, J.T.; Olson, E.N. MicroRNAs in Stress Signaling and Human Disease. Cell 2012, 148, 1172–1187. [Google Scholar] [CrossRef] [PubMed]
- Yen, Y.P.; Hsieh, W.F.; Tsai, Y.Y.; Lu, Y.L.; Liau, E.S.; Hsu, H.C.; Chen, Y.C.; Liu, T.C.; Chang, M.; Li, J.; et al. Dlk1-Dio3 Locus-Derived LncRNAs Perpetuate Postmitotic Motor Neuron Cell Fate and Subtype Identity. Elife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zheng, J.; Lin, J.; Chen, J.; Yu, Z.; Chen, C.; Liu, T. MiR-758 Mediates OxLDL-Dependent Vascular Endothelial Cell Damage by Suppressing the Succinate Receptor SUCNR1. Gene 2018, 663, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zheng, J.; Shen, H.; Huang, Y.; Liu, T.; Xi, H.; Chen, C. Curcumin Suppresses In Vitro Proliferation and Invasion of Human Prostate Cancer Stem Cells by Modulating DLK1-DIO3 Imprinted Gene Cluster MicroRNAs. Genet Test Mol Biomarkers 2018, 22, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, T.; Huang, Y. MicroRNA-134 Suppresses Endometrial Cancer Stem Cells by Targeting POGLUT1 and Notch Pathway Proteins. FEBS Lett 2015, 589, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Liu, T.; Huang, Y.; Qin, W.; Yang, H.; Chen, J. MicroRNA-134-3p Is a Novel Potential Inhibitor of Human Ovarian Cancer Stem Cells by Targeting RAB27A. Gene 2017, 605, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.; Sun, Y.; Ouyang, Q.; Hu, L.; Tan, Y.; Zhou, X.; Xiong, B.; Zhang, Q.; Yuan, D.; Pan, Y.; et al. Physiological Oxygen Prevents Frequent Silencing of the DLK1-DIO3 Cluster during Human Embryonic Stem Cells Culture. Stem Cells 2014, 32, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Ashwin Porwal Gopal, C. Kundu Gajanan Bhagwat Ramesh Butti Polyherbal Formulation Anoac-H Suppresses the Expression of RANTES and VEGF for the Management of Bleeding Hemorrhoids and Fistula. Available online: https://www.spandidos-publications.com/10.3892/mmr.2021.12376# (accessed on 12 February 2024).
- Shrivastava, L.; Silva Borges, G. da; Shrivastava, R. Clinical Efficacy of a Dual Action, Topical Anti-Edematous and Antiinflammatory Device for the Treatment of External Hemorrhoids. Clin Exp Pharmacol 2018, 08. [Google Scholar] [CrossRef]
- Ray, A. Cytokines and Their Role in Health and Disease: A Brief Overview. MOJ Immunol 2016, 4. [Google Scholar] [CrossRef]
- Srivastava, A.; Yadav, S.K.; Yachha, S.K.; Thomas, M.A.; Saraswat, V.A.; Gupta, R.K. Pro-Inflammatory Cytokines Are Raised in Extrahepatic Portal Venous Obstruction, with Minimal Hepatic Encephalopathy. J Gastroenterol Hepatol 2011, 26, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Szukiewicz, D. The Role of Inflammatory and Anti-Inflammatory Cytokines in the Pathogenesis of Osteoarthritis. Mediators Inflamm 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Jin, W.; Li, P.; Wang, R.; Guo, X. Traditional Chinese Medicine in the Treatment of Hemorrhoids—a Review of Preparations Used and Their Mechanism of Action. Front Pharmacol 2023, 14, 1270339. [Google Scholar] [CrossRef] [PubMed]
- Porwal, A.; Kundu, G.; Bhagwat, G.; Nimma, R.; Chowdhury, J. Turmocin Plus Suppresses Vascular Endothelial Growth Factor (VEGF) and Macrophage Infiltration in the Management of Perineal Wounds, Anal Fistula, Acute Anal Fissures and Haemorrhoids. Journal of Natural Remedies 2024, 24, 283–291. [Google Scholar] [CrossRef]
- Cuesta, M.C.; Quintero, L.; Pons, H.; Suarez-Roca, H. Substance P and Calcitonin Gene-Related Peptide Increase IL-1β, IL-6 and TNFα Secretion from Human Peripheral Blood Mononuclear Cells. Neurochem Int 2002, 40, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Khalil, M.; Alliger, K.; Weidinger, C.; Yerinde, C.; Wirtz, S.; Becker, C.; Engel, M.A. Functional Role of Transient Receptor Potential Channels in Immune Cells and Epithelia. Front Immunol 2018, 9, 174. [Google Scholar] [CrossRef] [PubMed]
- Torii, H.; Hosoi, J.; Beissert, S.; Xu, S.; Fox, F.E.; Asahina, A.; Takashima, A.; Rook, A.H.; Granstein, R.D. Regulation of Cytokine Expression in Macrophages and the Langerhans Cell-like Line XS52 by Calcitonin Gene-Related Peptide. J Leukoc Biol 1997, 61, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zhao, Y.; Zhang, X.; Li, W.; Zhang, H.; Piao, S.; Li, G.; Yan, M. The Beneficial Effect of Sanhuang Ointment and Its Active Constituents on Experimental Hemorrhoids in Rats. J Ethnopharmacol 2024, 319, 117173. [Google Scholar] [CrossRef] [PubMed]
- Klink, C.; Binnebösel, M.; Kämmer, D.; Willis, S.; Prescher, A.; Klinge, U.; Schumpelick, V. Haemorrhoids Are Related to Changes of Cell Function in Mucosa and Submucosa. Int J Colorectal Dis 2009, 24, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.S. Role and Regulation of Cyclooxygenase-2 during Inflammation. Am J Med 1999, 106, 37S–42S. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Liu, T.; Zhong, S.; Yang, W. MicroRNA-770 Promotes Polarization of Macrophages and Hemorrhoids by Suppressing RYBP Expression and Monoubiquitination of Histone H2A on Lys119 Modification. Mol Immunol 2025, 182, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Nieuwland, R.; Falcon-Perez, J.M.; Soekmadji, C.; Boilard, E.; Carter, D.; Buzas, E.I. Essentials of Extracellular Vesicles: Posters on Basic and Clinical Aspects of Extracellular Vesicles. J Extracell Vesicles 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Tkach, M.; Théry, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Cabral, J.; Ryan, A.E.; Griffin, M.D.; Ritter, T. Extracellular Vesicles as Modulators of Wound Healing. Adv Drug Deliv Rev 2018, 129, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Todorova, D.; Simoncini, S.; Lacroix, R.; Sabatier, F.; Dignat-George, F. Extracellular Vesicles in Angiogenesis. Circ Res 2017, 120, 1658. [Google Scholar] [CrossRef] [PubMed]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol 2017, 27, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu Rev Cell Dev Biol 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Crewe, C.; Joffin, N.; Rutkowski, J.M.; Kim, M.; Zhang, F.; Towler, D.A.; Gordillo, R.; Scherer, P.E. An Endothelial-to-Adipocyte Extracellular Vesicle Axis Governed by Metabolic State. Cell 2018, 175, 695–708.e13. [Google Scholar] [CrossRef] [PubMed]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell Mol Neurobiol 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Gene UBQLN1 ubiquilin 1 [ Homo sapiens (human) ]. Bethesda (MD): National Library of Medicine (US), N.C. for B.I. 2004 – cited 11/03/2024 UBQLN1 Ubiquilin 1 [ Homo Sapiens (Human) ]. Available online: https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=ShowDetailView&TermToSearch=29979 (accessed on 11 March 2024).
- Hochrainer, K.; Pejanovic, N.; Olaseun, V.A.; Zhang, S.; Iadecola, C.; Anrather, J. The Ubiquitin Ligase HERC3 Attenuates NF-ΚB-Dependent Transcription Independently of Its Enzymatic Activity by Delivering the RelA Subunit for Degradation. Nucleic Acids Res 2015, 43, 9889–9904. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Meng, Q.; Wei, L.; Liu, J.; Li, M.; Liang, X.; Lin, F.; Zhang, Y.; Li, Y.; Liu, Z.; et al. Myocardial Ischemia-reperfusion Induced Cardiac Extracellular Vesicles Harbour Proinflammatory Features and Aggravate Heart Injury. J Extracell Vesicles 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O.; et al. A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer. Cancers (Basel) 2019, 11, 1618. [Google Scholar] [CrossRef] [PubMed]
- Sahún-Español, Á.; Clemente, C.; Jiménez-Loygorri, J.I.; Sierra-Filardi, E.; Herrera-Melle, L.; Gómez-Durán, A.; Sabio, G.; Monsalve, M.; Boya, P.; Arroyo, A.G. P38 MAPK Priming Boosts VSMC Proliferation and Arteriogenesis by Promoting PGC1α-Dependent Mitochondrial Dynamics. Scientific Reports 2022 12:1 2022, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, L.; Wang, S.; Cheng, H.; Xu, L.; Pei, G.; Wang, Y.; Fu, C.; Jiang, Y.; He, C.; et al. Signaling Pathways and Targeted Therapy for Myocardial Infarction. Signal Transduct Target Ther 2022, 7. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.Y.; Koh, M.S.; Moon, A. The P38 MAPK Inhibitors for the Treatment of Inflammatory Diseases and Cancer. Expert Opin Investig Drugs 2009, 18, 1893–1905. [Google Scholar] [CrossRef] [PubMed]
- Haviarová, Z.; Janegová, A.; Janega, P.; Durdík, S.; Kováč, P.; Stvrtinová, V.; Mráz, P. Expression of Constitutive Nitric Oxide Synthase Isoforms in Varicose Vein Wall; Preliminary Results. Int J Vasc Med 2011, 2011, 204723. [Google Scholar] [CrossRef] [PubMed]
- Gokce, A.H.; Gokce, F.S.; Durmus, S.; Hajiyeva, R.; Ersoz, F.; Gelisgen, R.; Uzun, H. The Effect of Nitric Oxide, Endothelial Nitric Oxide Synthetase, and Asymmetric Dimethylarginine in Hemorrhoidal Disease. Rev Assoc Med Bras 2020, 66, 1128–1133. [Google Scholar] [CrossRef] [PubMed]
- Lohsiriwat, V.; Wilson, V.G.; Scholefield, J.H.; Dashwood, M.R. Regional Distribution of Nitric Oxide Synthase in Human Anorectal Tissue: A Pilot Study on the Potential Role for Nitric Oxide in Haemorrhoids. Curr Vasc Pharmacol 2018, 18, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Hou, Y.C.; Pan, A.C.H. Endoglin (CD105) Expression in the Development of Haemorrhoids. Eur J Clin Invest 2004, 34, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Chen, X.; Liu, H.; Zhang, L.; Yu, T.; Xie, M. Fulvestrant, an Estrogen Receptor Inhibitor, Relieves Postoperative Hemorrhoid Edema via up-Regulation of MiR- 424-5p. Tropical Journal of Pharmaceutical Research 2022, 21, 1101–1107. [Google Scholar] [CrossRef]
- Parés, D.; Iglesias, M.; Pera, M.; Pascual, M.; Torner, A.; Baró, T.; Alonso, S.; Grande, L. Expression of Estrogen and Progesterone Receptors in the Anal Canal of Women According to Age and Menopause. Dis Colon Rectum 2010, 53, 1687–1691. [Google Scholar] [CrossRef] [PubMed]
- Iorga, A.; Cunningham, C.M.; Moazeni, S.; Ruffenach, G.; Umar, S.; Eghbali, M. The Protective Role of Estrogen and Estrogen Receptors in Cardiovascular Disease and the Controversial Use of Estrogen Therapy. Biol Sex Differ 2017, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Gene [Internet]. Bethesda (MD): National Library of Medicine (US), N.C. for B.I. 2004 – cited 12/03/2024ESR1 estrogen receptor 1 [ H. sapiens (human) ] ESR1 Estrogen Receptor 1 [ Homo Sapiens (Human) ].
- Zhang, J.; Song, H.; Lu, Y.; Chen, H.; Jiang, S.; Li, L. Effects of Estradiol on VEGF and BFGF by Akt in Endometrial Cancer Cells Are Mediated through the NF-ΚB Pathway. Oncol Rep 2016, 36, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Losordo, D.W.; Isner, J.M. Estrogen and Angiogenesis: A Review. Arterioscler Thromb Vasc Biol 2001, 21, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Karas, R.H.; Gauer, E.A.; Bieber, H.E.; Baur, W.E.; Mendelsohn, M.E. Growth Factor Activation of the Estrogen Receptor in Vascular Cells Occurs via a Mitogen-Activated Protein Kinase-Independent Pathway. J Clin Invest 1998, 101, 2851–2861. [Google Scholar] [CrossRef] [PubMed]
- Shifren, J.L.; Tseng, J.F.; Zaloudek, C.J.; Ryan, I.P.; Meng, Y.G.; Ferrara, N.; Jaffe, R.B.; Taylor, R.N. Ovarian Steroid Regulation of Vascular Endothelial Growth Factor in the Human Endometrium: Implications for Angiogenesis during the Menstrual Cycle and in the Pathogenesis of Endometriosis. J Clin Endocrinol Metab 1996, 81, 3112–3118. [Google Scholar] [CrossRef] [PubMed]
- Tamura, M. Neovascularization in Experimental Retinal Venous Obstruction in Rabbits. Jpn J Ophthalmol 2001, 45, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.G.; Chung, H.; Hyon, J.Y. Experimental Preretinal Neovascularization by Laser-Induced Thrombosis in Albino Rats. Korean J Ophthalmol 1999, 13, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Gravina, G.L.; Senapedis, W.; McCauley, D.; Baloglu, E.; Shacham, S.; Festuccia, C. Nucleo-Cytoplasmic Transport as a Therapeutic Target of Cancer. J Hematol Oncol 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Camacho, S.C.; Silvers, T.R.; Razak, A.R.A.; Gabrail, N.Y.; Gerecitano, J.F.; Kalir, E.; Pereira, E.; Evans, B.R.; Ramus, S.J.; et al. Inhibition of the Nuclear Export Receptor XPO1 as a Therapeutic Target for Platinum-Resistant Ovarian Cancer. Clinical Cancer Research 2017, 23, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Wikström, P.; Lissbrant, I.F.; Stattin, P.; Egevad, L.; Bergh, A. Endoglin (CD105) Is Expressed on Immature Blood Vessels and Is a Marker for Survival in Prostate Cancer. Prostate 2002, 51, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Krupinski, J.; Kaluza, J.; Kumar, P.; Kumar, S.; Wang, J.M. Role of Angiogenesis in Patients with Cerebral Ischemic Stroke. Stroke 1994, 25, 1794–1798. [Google Scholar] [CrossRef] [PubMed]
- Marcovich, A.L.; Morad, Y.; Sandbank, J.; Huszar, M.; Rosner, M.; Pollack, A.; Herbert, M.; Bar-Dayan, Y. Angiogenesis in Pterygium: Morphometric and Immunohistochemical Study. Curr Eye Res 2002, 25, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Altomonte, M.; Montagner, R.; Fonsatti, E.; Colizzi, F.; Cattarossi, I.; Brasoveanu, L.I.; Nicotra, M.R.; Cattelan, A.; Natali, P.G.; Maio, M. Expression and Structural Features of Endoglin (CD105), a Transforming Growth Factor Beta1 and Beta3 Binding Protein, in Human Melanoma. Br J Cancer 1996, 74, 1586. [Google Scholar] [CrossRef] [PubMed]
- LIANG, W.; CAO, J.; YANG, P.; LI, W.; SUN, Z.; CHEN, X.; WANG, Q.; LIU, F. Clinical Significance and Distribution Characteristics of Expression of Vascular Endothelial Growth Factor Receptor 2 in Hemorrhoid Mucosa. The Journal of Practical Medicine 2015, 2830–2832. [Google Scholar] [CrossRef]
- Liu, Q.; Han, W.; Wang, L.; Shang, W.; Cao, X. Role of MiR-143-3p in the Development of Hemorrhoids and Postoperative Wound Healing. Journal of Investigative Surgery 2025, 38, 2480799. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, M.; Ishizawa, J.; Ruvolo, V.; Dilip, A.; Quintás-Cardama, A.; Mcdonnell, T.J.; Neelapu, S.S.; Kwak, L.W.; Shacham, S.; Kauffman, M.; et al. Induction of P53-Mediated Transcription and Apoptosis by Exportin-1 (XPO1) Inhibition in Mantle Cell Lymphoma. Cancer Sci 2014, 105, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Camus, V.; Miloudi, H.; Taly, A.; Sola, B.; Jardin, F. XPO1 in B Cell Hematological Malignancies: From Recurrent Somatic Mutations to Targeted Therapy. J Hematol Oncol 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Fabbrocini, G.; Kisslinger, A.; Iannelli, P.; Vitale, N.; Procaccini, C.; Sparaneo, G.; Chieffi, P.; Ayala, F.; Mancini, F.P.; Tramontano, D. Resveratrol Regulates P66Shc Activation in HaCaT Cells. Exp Dermatol 2010, 19, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, E.; Giogio, M.; Mele, S.; Pelicci, G.; Reboldi, P.; Pandolfi, P.P.; Lanfrancone, L.; Pelicci, P.G. The P66shc Adaptor Protein Controls Oxidative Stress Response and Life Span in Mammals. Nature 1999, 402, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Hu, Y.; Tang, H.; Hu, H.; Pang, L.; Xing, J.; Liu, Z.; Luo, Y.; Jiang, B.; Liu, T.; et al. RNA Methyltransferase NSUN2 Promotes Stress-Induced HUVEC Senescence. Oncotarget 2016, 7, 19099. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liu, Z.; Wu, R.F.; Terada, L.S. P66Shc Restrains Ras Hyperactivation and Suppresses Metastatic Behavior. Oncogene 2010, 29, 5559. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.S.; Anand, D.; Khanday, F.A. P66Shc as a Switch in Bringing about Contrasting Responses in Cell Growth: Implications on Cell Proliferation and Apoptosis. Mol Cancer 2015, 14. [Google Scholar] [CrossRef] [PubMed]
- Camici, G.G.; Schiavoni, M.; Francia, P.; Bachschmid, M.; Martin-Padura, I.; Hersberger, M.; Tanner, F.C.; Pelicci, P.G.; Volpe, M.; Anversa, P.; et al. Genetic Deletion of P66Shc Adaptor Protein Prevents Hyperglycemia-Induced Endothelial Dysfunction and Oxidative Stress. Proc Natl Acad Sci U S A 2007, 104, 5217. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Martin-Padura, I.; De Nigris, F.; Giorgio, M.; Mansueto, G.; Somma, P.; Condorelli, M.; Sica, G.; De Rosa, G.; Pelicci, P.G. Deletion of the P66Shc Longevity Gene Reduces Systemic and Tissue Oxidative Stress, Vascular Cell Apoptosis, and Early Atherogenesis in Mice Fed a High-Fat Diet. Proc Natl Acad Sci U S A 2003, 100, 2112–2116. [Google Scholar] [CrossRef] [PubMed]
- Francia, P.; Delli Gatti, C.; Bachschmid, M.; Martin-Padura, I.; Savoia, C.; Migliaccio, E.; Pelicci, P.G.; Schiavoni, M.; Lüscher, T.F.; Volpe, M.; et al. Deletion of P66shc Gene Protects against Age-Related Endothelial Dysfunction. Circulation 2004, 110, 2889–2895. [Google Scholar] [CrossRef] [PubMed]
- Yamamori, T.; White, A.R.; Mattagajasingh, I.; Khanday, F.A.; Haile, A.; Qi, B.; Byeong, H.J.; Bugayenko, A.; Kasuno, K.; Berkowitz, D.E.; et al. P66shc Regulates Endothelial NO Production and Endothelium-Dependent Vasorelaxation: Implications for Age-Associated Vascular Dysfunction. J Mol Cell Cardiol 2005, 39, 992–995. [Google Scholar] [CrossRef] [PubMed]
- Trinei, M.; Giorgio, M.; Cicalese, A.; Barozzi, S.; Ventura, A.; Migliaccio, E.; Milia, E.; Padura, I.M.; Raker, V.A.; Maccarana, M.; et al. A P53-P66Shc Signalling Pathway Controls Intracellular Redox Status, Levels of Oxidation-Damaged DNA and Oxidative Stress-Induced Apoptosis. Oncogene 2002, 21, 3872–3878. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 Complex Mediates Mammalian Nuclear RNA N6-Adenosine Methylation. Nat Chem Biol 2014, 10, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wei, L.; Law, C.T.; Tsang, F.H.C.; Shen, J.; Cheng, C.L.H.; Tsang, L.H.; Ho, D.W.H.; Chiu, D.K.C.; Lee, J.M.F.; et al. RNA N6-Methyladenosine Methyltransferase-like 3 Promotes Liver Cancer Progression through YTHDF2-Dependent Posttranscriptional Silencing of SOCS2. Hepatology 2018, 67, 2254–2270. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-Methyladenosine in Nuclear RNA Is a Major Substrate of the Obesity-Associated FTO. Nat Chem Biol 2011, 7, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Klukovich, R.; Peng, H.; Wang, Z.; Yu, T.; Zhang, Y.; Zheng, H.; Klungland, A.; Yan, W. ALKBH5-Dependent M6A Demethylation Controls Splicing and Stability of Long 3’-UTR MRNAs in Male Germ Cells. Proc Natl Acad Sci U S A 2018, 115, E325–E333. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-Methyladenosine-Dependent Regulation of Messenger RNA Stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, I.; Tzelepis, K.; Pandolfini, L.; Shi, J.; Millán-Zambrano, G.; Robson, S.C.; Aspris, D.; Migliori, V.; Bannister, A.J.; Han, N.; et al. Promoter-Bound METTL3 Maintains Myeloid Leukaemia by M6A-Dependent Translation Control. Nature 2017, 552, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hao, Y.J.; Zhang, Y.; Li, M.M.; Wang, M.; Han, W.; Wu, Y.; Lv, Y.; Hao, J.; Wang, L.; et al. M(6)A RNA Methylation Is Regulated by MicroRNAs and Promotes Reprogramming to Pluripotency. Cell Stem Cell 2015, 16, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Savant, S.; La Porta, S.; Budnik, A.; Busch, K.; Hu, J.; Tisch, N.; Korn, C.; Valls, A.F.; Benest, A. V.; Terhardt, D.; et al. The Orphan Receptor Tie1 Controls Angiogenesis and Vascular Remodeling by Differentially Regulating Tie2 in Tip and Stalk Cells. Cell Rep 2015, 12, 1761–1773. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.; Sandi, M.J.; Cordelier, P.; Binétruy, B.; Pouysségur, J.; Iovanna, J.L.; Tournaire, R. Tie1 Deficiency Induces Endothelial–Mesenchymal Transition. EMBO Rep 2012, 13, 431. [Google Scholar] [CrossRef] [PubMed]
- Torigata, M.; Yamakawa, D.; Takakura, N. Elevated Expression of Tie1 Is Accompanied by Acquisition of Cancer Stemness Properties in Colorectal Cancer. Cancer Med 2017, 6, 1378. [Google Scholar] [CrossRef] [PubMed]
- La Porta, S.L.; Roth, L.; Singhal, M.; Mogler, C.; Spegg, C.; Schieb, B.; Qu, X.; Adams, R.H.; Scott Baldwin, H.; Savant, S.; et al. Endothelial Tie1-Mediated Angiogenesis and Vascular Abnormalization Promote Tumor Progression and Metastasis. J Clin Invest 2018, 128, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Tiainen, L.; Korhonen, E.A.; Leppänen, V.M.; Luukkaala, T.; Hämäläinen, M.; Tanner, M.; Lahdenperä, O.; Vihinen, P.; Jukkola, A.; Karihtala, P.; et al. High Baseline Tie1 Level Predicts Poor Survival in Metastatic Breast Cancer. BMC Cancer 2019, 19, 732. [Google Scholar] [CrossRef] [PubMed]
- Gene [Internet]. Bethesda (MD): National Library of Medicine (US), N.C. for B.I. 2004 – cited 12/03/2024Gene [Internet]. B. (MD): N.L. of M. (US), N.C. for B.I. 2004 – [cited Y.M.D. TIE1 Tyrosine Kinase with Immunoglobulin like and EGF like Domains 1 [ Homo Sapiens (Human) ].
- Haas, P.A.; Fox, T.A.; Haas, G.P. The Pathogenesis of Hemorrhoids. Dis Colon Rectum 1984, 27, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Sardiñas, C.; Arreaza, D.D.; Osorio, H. Changes in the Proportions of Types I and III Collagen in Hemorrhoids: The Sliding Anal Lining Theory. Journal of Coloproctology 2016, 36, 124–129. [Google Scholar] [CrossRef]
- Willis, S.; Junge, K.; Ebrahimi, R.; Prescher, A.; Schumpelick, V. Haemorrhoids - a Collagen Disease? Colorectal Dis 2010, 12, 1249–1253. [Google Scholar] [CrossRef] [PubMed]
- Nasseri, Y.Y.; Krott, E.; Van Groningen, K.M.; Berho, M.; Osborne, M.C.; Wollman, S.; Weiss, E.G.; Wexner, S.D. Abnormalities in Collagen Composition May Contribute to the Pathogenesis of Hemorrhoids: Morphometric Analysis. Tech Coloproctol 2015, 19, 83. [Google Scholar] [CrossRef] [PubMed]
- Sun, F. The Effect of Tonifying Qi Method on the Expression of Anti-Aging FIbulin Proteins Family of Internal Prolapsed Hemorrhoids. 2014.
- Sun, F., X.T., C.H., and L.Y. Study on Effect of Buzhong Yiqi Tang on Expression of FIbulin-5 in Grade-Iii Prolapsed Hemorrhoid Tissues. 2018.
- Jin, X., W.X., H.D., L.Y., and S.F. Therapeutic Efficacy of Buzhong Yiqi Decoction Containing Different Doses of Radix Astragali for Stage i Internal Hemorrhoids Patients with Spleen Deficiency and Sinking of Qi and Its Effect on FIbulin-3 Expression in Hemorrhoid Tissues. 2017.
- Zhong, P. Study on Clinical Efficiency and FIbulin-3’ Expression of Buzhong-Yiqi-Decoction Containing Different Dose Astragalus in Hemorrhoids’ Patient Belongs to Spleen-Type Gas Trap. 2016.
- Xiao, T. The Study of the Effect of Buzhong Yiqi Decoction on Expression of FIbulin-5 in Iii Degree of Prolapsed Internal Hemorrhoids Tissues. 2016.
- Albig, A.R.; Schiemann, W.P. Fibulin-5 Antagonizes Vascular Endothelial Growth Factor (VEGF) Signaling and Angiogenic Sprouting by Endothelial Cells. DNA Cell Biol 2004, 23, 367–379. [Google Scholar] [CrossRef] [PubMed]
- De Vega, S.; Iwamoto, T.; Yamada, Y. Fibulins: Multiple Roles in Matrix Structures and Tissue Functions. Cell Mol Life Sci 2009, 66, 1890–1902. [Google Scholar] [CrossRef] [PubMed]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Rosa, C.C.D. La; Ramirez-Acuña, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int J Mol Sci 2020, 21, 1–53. [Google Scholar] [CrossRef] [PubMed]
- Kisli, E.; Kemik, A.; Sümer, A.; Kemik, Ö. Matrix Metalloproteinases in Pathogenesis of Hemorrhoidal Disease. https://doi.org/10.1177/000313481307901122 2013, 79, 1181–1184. [CrossRef]
- Xie, D. Effects of Mmp-7 and Mmp-9 on the Pathogenesis of Hemorrhoids. 2008.
- Qin, L., and Q.X. Expression and Significance of Mmp-9 and Vegfr2 in Patients with Internal Hemorrhoids. . 2020.
- Wei Han, Z.W.B.Z.X.Y.D.W.J.W.X.T.F.Z.Y.H. Pathologic Change of Elastic Fibers with Difference of Microvessel Density and Expression of Angiogenesis-Related Proteins in Internal Hemorrhoid Tissues. 2005.
- Di, Y.; Nie, Q.Z.; Chen, X.L. Matrix Metalloproteinase-9 and Vascular Endothelial Growth Factor Expression Change in Experimental Retinal Neovascularization. Int J Ophthalmol 2016, 9, 804. [Google Scholar] [CrossRef] [PubMed]
- Hollborn, M.; Stathopoulos, C.; Steffen, A.; Wiedemann, P.; Kohen, L.; Bringmann, A. Positive Feedback Regulation between MMP-9 and VEGF in Human RPE Cells. Invest Ophthalmol Vis Sci 2007, 48, 4360–4367. [Google Scholar] [CrossRef] [PubMed]
- Tenghao Zheng, D.E.S.J. Genome-Wide Analysis of 944 133 Individuals Provides Insights into the Etiology of Haemorrhoidal Disease - Record Details - Embase. Available online: https://www-1embase-1com-1qtgcbczf0003.han.bg.umw.edu.pl/records?subaction=viewrecord&id=L634938305 (accessed on 15 February 2024).
- Ng, M.Y.M.; Andrew, T.; Spector, T.D.; Jeffery, S. Linkage to the FOXC2 Region of Chromosome 16 for Varicose Veins in Otherwise Healthy, Unselected Siblings Pairs. J Med Genet 2005, 42, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Surendran, S.; Girijamma, A.; Nair, R.; Ramegowda, K.S.; Nair, D.H.; Thulaseedharan, J. V.; Lakkappa, R.B.; Kamalapurkar, G.; Kartha, C.C. Forkhead Box C2promoter Variant C<512C.T Is Associated with Increased Susceptibility to Chronic Venous Diseases. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Brice, G.; Mansour, S.; Bell, R.; Collin, J.R.O.; Child, A.H.; Brady, A.F.; Sarfarazi, M.; Burnand, K.G.; Jeffery, S.; Mortimer, P.; et al. Analysis of the Phenotypic Abnormalities in Lymphoedema-Distichiasis Syndrome in 74 Patients with FOXC2 Mutations or Linkage to 16q24. J Med Genet 2002, 39, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Kume, T. The Cooperative Roles of Foxcl and Foxc2 in Cardiovascular Development. Adv Exp Med Biol 2009, 665, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.; Andreucci, M.; Ielapi, N.; Serraino, G.F.; Mastroroberto, P.; Bracale, U.M.; Serra, R. Molecular Determinants of Chronic Venous Disease: A Comprehensive Review. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Murk, W.; de Wan, A.T. Exhaustive Genome-Wide Search for SNP-SNP Interactions across 10 Human Diseases. G3: Genes, Genomes, Genetics 2016, 6, 2043–2050. [Google Scholar] [CrossRef] [PubMed]
- Qar, J.; Al Zoubi, M.S.; Baydoun, I.M.A.; Aljabali, A.A.A.; Al-Trad, B.; Rabi, F.; Al Batayneh, K.M. Truncating Mutation in FOXC2 Gene in Familial Hemorrhoids and Varicose Veins. International Journal of Biology and Biomedical Engineering 2020, 14, 32–38. [Google Scholar] [CrossRef]
- Petrova, T. V.; Karpanen, T.; Norrmén, C.; Mellor, R.; Tamakoshi, T.; Finegold, D.; Ferrell, R.; Kerjaschki, D.; Mortimer, P.; Ylä-Herttuala, S.; et al. Defective Valves and Abnormal Mural Cell Recruitment Underlie Lymphatic Vascular Failure in Lymphedema Distichiasis. Nat Med 2004, 10, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Bharath, V.; Kahn, S.R.; Lazo-Langner, A. Genetic Polymorphisms of Vein Wall Remodeling in Chronic Venous Disease: A Narrative and Systematic Review. Blood 2014, 124, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Adikusuma, W.; Firdayani, F.; Irham, L.M.; Darmawi, D.; Hamidy, M.Y.; Nopitasari, B.L.; Soraya, S.; Azizah, N. Integrated Genomic Network Analysis Revealed Potential of a Druggable Target for Hemorrhoid Treatment. Saudi Pharmaceutical Journal 2023, 31. [Google Scholar] [CrossRef] [PubMed]
- Schwerd, T.; Bryant, R. V.; Pandey, S.; Capitani, M.; Meran, L.; Cazier, J.B.; Jung, J.; Mondal, K.; Parkes, M.; Mathew, C.G.; et al. NOX1 Loss-of-Function Genetic Variants in Patients with Inflammatory Bowel Disease. Mucosal Immunology 2018 11:2 2017, 11, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Dubois, E.A.; Cohen, A.F. Sapropterin: New Drug Mechanisms. Br J Clin Pharmacol 2010, 69, 576–577. [Google Scholar] [CrossRef] [PubMed]
- Stanhewicz, A.E.; Alexander, L.M.; Kenney, W.L. Oral Sapropterin Acutely Augments Reflex Vasodilation in Aged Human Skin through Nitric Oxide-Dependent Mechanisms. J Appl Physiol 2013, 115, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. International Journal of Biochemistry and Cell Biology 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Xue, M.; Kang, C.; Gao, L.; Zhang, M.; Ma, C.; Jia, W.; Zheng, Y.; Cao, L.; Chen, P.; et al. Increased NOX1 and DUOX2 Expression in the Colonic Mucosa of Patients with Chronic Functional Constipation. Medicine (United States) 2022, 101, E30028. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.W.; Mason, J.B. Folate Status: Effects on Pathways of Colorectal Carcinogenesis. Journal of Nutrition 2002, 132. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, I.; Tran, P.; Christensen, B.; Sibani, S.; Rozen, R. A Second Genetic Polymorphism in Methylenetetrahydrofolate Reductase (MTHFR) Associated with Decreased Enzyme Activity. Mol Genet Metab 1998, 64, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Promthet, S.; Pientong, C.; Ekalaksananan, T.; Songserm, N.; Poomphakwaen, K.; Chopjitt, P.; Wiangnon, S.; Tokudome, S. Risk Factors for Rectal Cancer and Methylenetetrahydrofolate Reductase Polymorphisms in a Population in Northeast Thailand. Asian Pacific Journal of Cancer Prevention 2012, 13, 4017–4023. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.; Ssempijja, L.; Provenzano, M.; Andreucci, M. Genetic Biomarkers in Chronic Venous Disease. Biomark Med 2020, 14, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Abuoğlu, H.H.; Gunay, E.; Uzunoğlu, H. Effect of Genetic Factors on the Etiopathogenesis of Thrombosed Hemorrhoidal Disease. Chirurgia (Romania) 2019, 114, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Shi, H.; Zhou, H.; Song, X.; Yuan, S.; Luo, Y. The Angiogenic Function of Nucleolin Is Mediated by Vascular Endothelial Growth Factor and Nonmuscle Myosin. Blood 2006, 107, 3564–3571. [Google Scholar] [CrossRef] [PubMed]
- Salnikova, L.E.; Khadzhieva, M.B.; Kolobkov, D.S. Biological Findings from the PheWAS Catalog: Focus on Connective Tissue-Related Disorders (Pelvic Floor Dysfunction, Abdominal Hernia, Varicose Veins and Hemorrhoids). Hum Genet 2016, 135, 779–795. [Google Scholar] [CrossRef] [PubMed]
- Pecci, A.; Klersy, C.; Gresele, P.; Lee, K.J.D.; De Rocco, D.; Bozzi, V.; Russo, G.; Heller, P.G.; Loffredo, G.; Ballmaier, M.; et al. MYH9-Related Disease: A Novel Prognostic Model to Predict the Clinical Evolution of the Disease Based on Genotype-Phenotype Correlations. Hum Mutat 2014, 35, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Hillarp, A.; Dahlbäck, B.; Zöller, B. Actuated Protein C Resistance: From Phenotype to Genotype and Clinical Practice. Blood Rev 1995, 9, 201–212. [Google Scholar] [CrossRef] [PubMed]
- McGrath, I.M.; Montgomery, G.W.; Mortlock, S. Genomic Characterisation of the Overlap of Endometriosis with 76 Comorbidities Identifies Pleiotropic and Causal Mechanisms Underlying Disease Risk. Hum Genet 2023, 142, 1345–1360. [Google Scholar] [CrossRef] [PubMed]
- Tsatsakis, A.M.; Zafiropoulos, A.; Tzatzarakis, M.N.; Tzanakakis, G.N.; Kafatos, A. Relation of PON1 and CYP1A1 Genetic Polymorphisms to Clinical Findings in a Cross-Sectional Study of a Greek Rural Population Professionally Exposed to Pesticides. Toxicol Lett 2009, 186, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Humbert, R.; Adler, D.A.; Disteche, C.M.; Hassett, C.; Omiecinski, C.J.; Furlong, C.E. The Molecular Basis of the Human Serum Paraoxonase Activity Polymorphism. Nat Genet 1993, 3, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Hassett, C.; Omiecinski, C.J.; Richter, R.J.; Humbert, R.; Humbert, R.; Furlong, C.E.; Furlong, C.E.; Omiecinski, C.J.; Richter, R.J.; Humbert, R.; et al. Characterization of CDNA Clones Encoding Rabbit and Human Serum Paraoxonase: The Mature Protein Retains Its Signal Sequence. Biochemistry 1991, 30, 10141–10149. [Google Scholar] [CrossRef] [PubMed]
- Blatter Garin, M.C.; James, R.W.; Dussoix, P.; Blanché, H.; Passa, P.; Froguel, P.; Ruiz, J. Paraoxonase Polymorphism Met-Leu54 Is Associated with Modified Serum Concentrations of the Enzyme. A Possible Link between the Paraoxonase Gene and Increased Risk of Cardiovascular Disease in Diabetes. Journal of Clinical Investigation 1997, 99, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Isbell, H.M.; Fressart, V.; Denjoy, I.; Debbiche, A.; Itoh, H.; Poinsot, J.; George, A.L.; Coulombe, A.; Shea, M.A.; et al. Novel CALM3 Variant Causing Calmodulinopathy With Variable Expressivity in a 4-Generation Family. Circ Arrhythm Electrophysiol 2022, 15, E010572. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Ni, Z.; Li, Z. CALM3 Affects the Prognosis of Leukemia and Hemorrhoids. Medicine 2023, 102, e36027. [Google Scholar] [CrossRef] [PubMed]
- Naba, A.; Clauser, K.R.; Whittaker, C.A.; Carr, S.A.; Tanabe, K.K.; Hynes, R.O. Extracellular Matrix Signatures of Human Primary Metastatic Colon Cancers and Their Metastases to Liver. BMC Cancer 2014, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.W.; Johnson, G. Acetylcholinesterase in Hirschsprung’s Disease. Pediatr Surg Int 2005, 21, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Kwartler, C.S.; Chen, J.; Thakur, D.; Li, S.; Baskin, K.; Wang, S.; Wang, Z. V.; Walker, L.; Hill, J.A.; Epstein, H.F.; et al. Overexpression of Smooth Muscle Myosin Heavy Chain Leads to Activation of the Unfolded Protein Response and Autophagic Turnover of Thick Filament-Associated Proteins in Vascular Smooth Muscle Cells. Journal of Biological Chemistry 2014, 289, 14075–14088. [Google Scholar] [CrossRef] [PubMed]
- Plackett, T.P.; Kwon, E.; Ronald A Gagliano, J.; Oh, R.C. Ehlers-Danlos Syndrome—Hypermobility Type and Hemorrhoids. Case Rep Surg 2014, 2014, 171803. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Liu, W.; Alkhouri, R.; Baker, R.D.; Bard, J.E.; Quigley, E.M.; Baker, S.S. Structural Changes in the Gut Microbiome of Constipated Patients. Physiol Genomics 2014, 46, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Brisinda, G. How to Treat Haemorrhoids. Prevention Is Best; Haemorrhoidectomy Needs Skilled Operators. BMJ 2000, 321, 582–583. [Google Scholar] [CrossRef] [PubMed]
- Hullar, M.A.J.; Fu, B.C. Diet, the Gut Microbiome, and Epigenetics. Cancer Journal (United States) 2014, 20, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Watson, M.M.; van der Giezen, M.; Søreide, K. Gut Microbiome Influence on Human Epigenetics, Health, and Disease. Handbook of Epigenetics: The New Molecular and Medical Genetics, Third Edition 2023, 669–686. [CrossRef]
- Sharma, M.; Li, Y.; Stoll, M.L.; Tollefsbol, T.O. The Epigenetic Connection Between the Gut Microbiome in Obesity and Diabetes. Front Genet 2020, 10, 453795. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, V.D.; Tutino, R.; Messina, M.; Santarelli, M.; Nigro, C.; Lo Secco, G.; Piceni, C.; Montanari, E.; Barletta, G.; Venturelli, P.; et al. Altered Gut Microbic Flora and Haemorrhoids: Could They Have a Possible Relationship? J Clin Med 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Kibret, A.A.; Oumer, M.; Moges, A.M. Prevalence and Associated Factors of Hemorrhoids among Adult Patients Visiting the Surgical Outpatient Department in the University of Gondar Comprehensive Specialized Hospital, Northwest Ethiopia. PLoS One 2021, 16. [Google Scholar] [CrossRef] [PubMed]
- Peery, A.F.; Sandler, R.S.; Galanko, J.A.; Bresalier, R.S.; Figueiredo, J.C.; Ahnen, D.J.; Barry, E.L.; Baron, J.A. Risk Factors for Hemorrhoids on Screening Colonoscopy. PLoS One 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Riss, S.; Weiser, F.A.; Schwameis, K.; Riss, T.; Mittlböck, M.; Steiner, G.; Stift, A. The Prevalence of Hemorrhoids in Adults. Int J Colorectal Dis 2012, 27, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, H.E.; Kang, J.H.; Shin, J.Y.; Song, Y.M. Factors Associated with Hemorrhoids in Korean Adults: Korean National Health and Nutrition Examination Survey. Korean J Fam Med 2014, 35, 227. [Google Scholar] [CrossRef] [PubMed]
- Frezza, E.E.; Shebani, K.O.; Robertson, J.; Wachtel, M.S. Morbid Obesity Causes Chronic Increase of Intraabdominal Pressure. Dig Dis Sci 2007, 52, 1038–1041. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.S.; Holmes, M. V.; Zheng, J.; Sanderson, E.; Carter, A.R. The Impact of Education Inequality on Rheumatoid Arthritis Risk Is Mediated by Smoking and Body Mass Index: Mendelian Randomization Study. Rheumatology (Oxford) 2022, 61, 2167–2175. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Gui, Y.; Qin, H.; Xie, Y. Causal Association between Adiposity and Hemorrhoids: A Mendelian Randomization Study. Front Med (Lausanne) 2023, 10. [Google Scholar] [CrossRef] [PubMed]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Frühbeck, G. Role of Extracellular Matrix Remodelling in Adipose Tissue Pathophysiology: Relevance in the Development of Obesity. Histol Histopathol 2012, 27, 1515–1528. [Google Scholar] [CrossRef] [PubMed]
- Karaskova, E.; Velganova-veghova, M.; Geryk, M.; Foltenova, H.; Kucerova, V.; Karasek, D. Role of Adipose Tissue in Inflammatory Bowel Disease. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Kessler, C. Pathophysiology of Obesity. Nurs Clin North Am 2021, 56, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Seidell, J.C.; de Groot, L.C.P.G.M.; van Sonsbeek, J.L.A.; Deurenberg, P.; Hautvast, J.G. Associations of Moderate and Severe Overweight with Self-Reported Illness and Medical Care in Dutch Adults. Am J Public Health 1986, 76, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Negri, E.; Pagano, R.; Decarli, A.; La Vecchia, C. Body Weight and the Prevalence of Chronic Diseases. J Epidemiol Community Health (1978) 1988, 42, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.S.; Jung, K.U.; Rampal, S.; Zhao, D.; Guallar, E.; Ryu, S.; Chang, Y.; Kim, H.O.; Kim, H.; Chun, H.K.; et al. Risk Factors for Hemorrhoidal Disease among Healthy Young and Middle-Aged Korean Adults. Sci Rep 2022, 12, 129. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, S.J.; Molloy, P.L.; Varinli, H.; Morrison, J.L.; Muhlhausler, B.S.; Buckley, M.; Clark, S.J.; McMillen, I.C.; Noakes, M.; Samaras, K.; et al. Epigenetics and Human Obesity. International Journal of Obesity 2015 39:1 2014, 39, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Herrera, B.M.; Keildson, S.; Lindgren, C.M. Genetics and Epigenetics of Obesity. Maturitas 2011, 69, 41–49. [Google Scholar] [CrossRef]
- Rohde, K.; Keller, M.; la Cour Poulsen, L.; Blüher, M.; Kovacs, P.; Böttcher, Y. Genetics and Epigenetics in Obesity. Metabolism 2019, 92, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Pigot, F.; Siproudhis, L.; Allaert, F.A. Risk Factors Associated with Hemorrhoidal Symptoms in Specialized Consultation. Gastroenterol Clin Biol 2005, 29, 1270–1274. [Google Scholar] [CrossRef] [PubMed]
- Delcò, F.; Sonnenberg, A. Associations between Hemorrhoids and Other Diagnoses. Dis Colon Rectum 1998, 41, 1534–1541. [Google Scholar] [CrossRef] [PubMed]
- Mitra, D.; Davis, K.L.; Baran, R.W. Healthcare Costs and Clinical Sequelae Associated with Constipation in a Managed Care Population. American Journal of Gastroenterology 2007, 102, S432. [Google Scholar] [CrossRef]
- Johannsson, H.Ö.; Graf, W.; Påhlman, L. Bowel Habits in Hemorrhoid Patients and Normal Subjects. Am J Gastroenterol 2005, 100, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Helvaci, M.R.; Algin, M.C.; Kaya, H. Irritable Bowel Syndrome and Chronic Gastritis, Hemorrhoid, Urolithiasis. Eurasian J Med 2009, 41, 158. [Google Scholar] [PubMed]
- Rusu, F.; Dumitraşcu, D.L. Four Years Follow-up of Patients with Irritable Bowel Syndrome. Rom J Intern Med 2015, 53, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Dothel, G.; Barbaro, M.R.; Di Vito, A.; Ravegnini, G.; Gorini, F.; Monesmith, S.; Coschina, E.; Benuzzi, E.; Fuschi, D.; Palombo, M.; et al. New Insights into Irritable Bowel Syndrome Pathophysiological Mechanisms: Contribution of Epigenetics. Journal of Gastroenterology 2023 58:7 2023, 58, 605–621. [Google Scholar] [CrossRef] [PubMed]
- Baethge, C.; Goldbeck-Wood, S.; Mertens, S. SANRA—a Scale for the Quality Assessment of Narrative Review Articles. Res Integr Peer Rev 2019, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bramer, W.M.; Rethlefsen, M.L.; Kleijnen, J.; Franco, O.H. Optimal Database Combinations for Literature Searches in Systematic Reviews: A Prospective Exploratory Study. Syst Rev 2017, 6, 245. [Google Scholar] [CrossRef] [PubMed]
- Higgins JPT; Thomas J; Chandler J; Cumpston M; Li T; Page MJ; Welch VA (editors) Cochrane Handbook for Systematic Reviews of Interventions Version 6.4; version 6.4.; Cochrane, 2023.
Figure 1.
When miR-412-5p is under-expressed, Xpo1 is up-regulated, which results in excessive export of p53 and FOXO from the nucleus. The lack of p53 and FOXO in the nucleus reduces the expression of SHC genes – p46SHC, p52SHC, and p66SHC, which inhibits the p53–p66SHC–p16 pathway. This leads to uncontrolled proliferation and angiogenesis, contributing to the formation of pathological hemorrhoids (Xpo1 – Exportin-1, p53 – tumor suppressor protein, FOXO – Forkhead box O transcription factor, SHC – Src homology 2 domain-containing (p46SHC, p52SHC, p66SHC), p16 – cyclin-dependent kinase inhibitor involved in cell cycle arrest). Created in BioRender. Mazurek, M. (2025) https://BioRender.com/c8w1axi.
Figure 1.
When miR-412-5p is under-expressed, Xpo1 is up-regulated, which results in excessive export of p53 and FOXO from the nucleus. The lack of p53 and FOXO in the nucleus reduces the expression of SHC genes – p46SHC, p52SHC, and p66SHC, which inhibits the p53–p66SHC–p16 pathway. This leads to uncontrolled proliferation and angiogenesis, contributing to the formation of pathological hemorrhoids (Xpo1 – Exportin-1, p53 – tumor suppressor protein, FOXO – Forkhead box O transcription factor, SHC – Src homology 2 domain-containing (p46SHC, p52SHC, p66SHC), p16 – cyclin-dependent kinase inhibitor involved in cell cycle arrest). Created in BioRender. Mazurek, M. (2025) https://BioRender.com/c8w1axi.
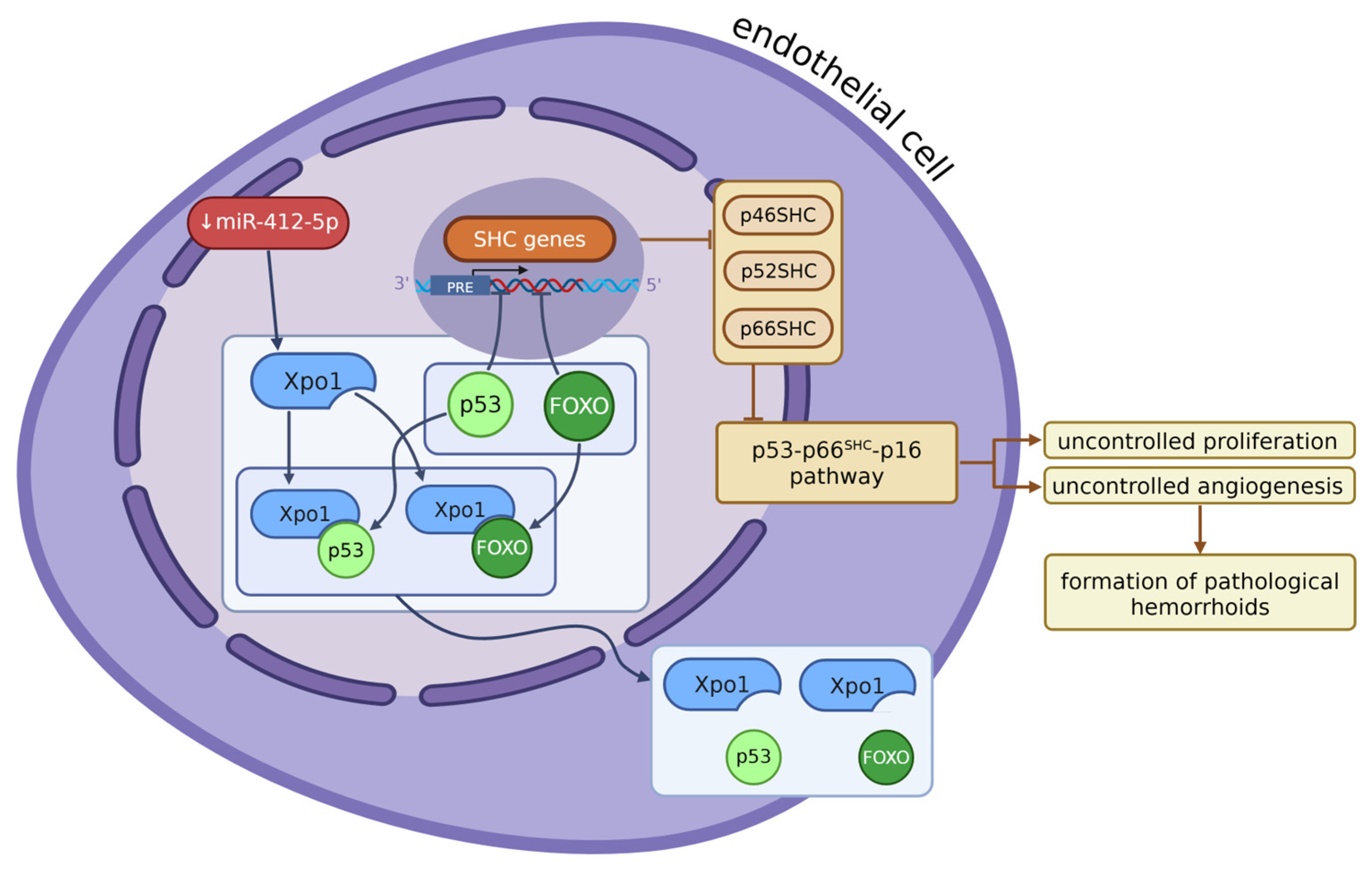
Figure 2.
In hemorrhoidal vascular endothelial cells, miR-4729 is significantly down-regulated, leading to over-expression of METTL14, a key m6A RNA methyltransferase. Elevated METTL14 increases m6A methylation via activating METTL3 at the 3′ UTR of, enhancing TIE1 mRNA stability and translation. As a result, TIE1 protein is overexpressed and activates the TIE1/VEGFA signaling loop, involving VEGFA, VEGFR2, vWF, and CD31, which promotes endothelial cell proliferation and angiogenesis, prominent in hemorrhoids (METTL3/14 – methyltransferase-like 3/14, m6A – N6-methyladenosine, TIE1 – tyrosine kinase receptor, VEGFA – vascular endothelial growth factor, VEGFR2 – vascular endothelial growth factor receptor 2, vWF – von Willebrand factor, CD31 – platelet endothelial cell adhesion molecule, tip/stalk cells – specialized endothelial cells involved in new vessel formation). Created in BioRender. Mazurek, M. (2025) https://BioRender.com/fcihnmu.
Figure 2.
In hemorrhoidal vascular endothelial cells, miR-4729 is significantly down-regulated, leading to over-expression of METTL14, a key m6A RNA methyltransferase. Elevated METTL14 increases m6A methylation via activating METTL3 at the 3′ UTR of, enhancing TIE1 mRNA stability and translation. As a result, TIE1 protein is overexpressed and activates the TIE1/VEGFA signaling loop, involving VEGFA, VEGFR2, vWF, and CD31, which promotes endothelial cell proliferation and angiogenesis, prominent in hemorrhoids (METTL3/14 – methyltransferase-like 3/14, m6A – N6-methyladenosine, TIE1 – tyrosine kinase receptor, VEGFA – vascular endothelial growth factor, VEGFR2 – vascular endothelial growth factor receptor 2, vWF – von Willebrand factor, CD31 – platelet endothelial cell adhesion molecule, tip/stalk cells – specialized endothelial cells involved in new vessel formation). Created in BioRender. Mazurek, M. (2025) https://BioRender.com/fcihnmu.
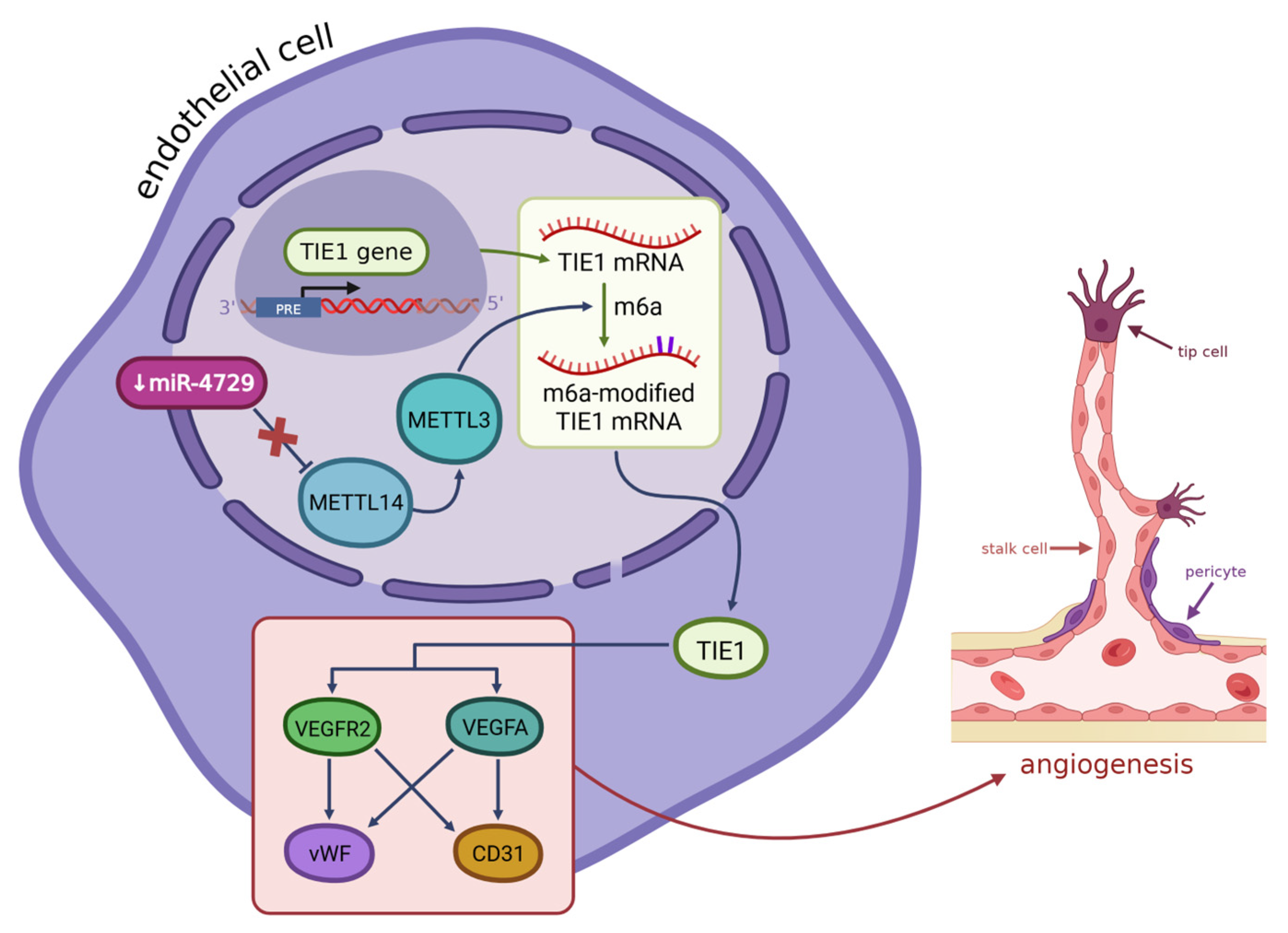
Figure 3.
Estrogens activate ERα in the anal canal epithelium of women with hemorrhoids via genomic and non-genomic pathways, leading to Src activation and downstream signaling through PI3K/Akt, MAPK, and p38. This downregulates miR-4729, which results in the overexpression of ESR1 and enhances the VEGF transcription. Increased VEGF promotes angiogenesis and edema in hemorrhoidal tissue (ERα – estrogen receptor alpha, ERβ – estrogen receptor beta, ESR1 – estrogen receptor 1 gene, VEGF – vascular endothelial growth factor, PI3K – phosphoinositide 3-kinase, MAPK – mitogen-activated protein kinase, miR-4729 – microRNA-4729, SH2/SH3 – Src homology 2/3 domains). Created in BioRender. Mazurek, M. (2025) https://BioRender.com/158e9db.
Figure 3.
Estrogens activate ERα in the anal canal epithelium of women with hemorrhoids via genomic and non-genomic pathways, leading to Src activation and downstream signaling through PI3K/Akt, MAPK, and p38. This downregulates miR-4729, which results in the overexpression of ESR1 and enhances the VEGF transcription. Increased VEGF promotes angiogenesis and edema in hemorrhoidal tissue (ERα – estrogen receptor alpha, ERβ – estrogen receptor beta, ESR1 – estrogen receptor 1 gene, VEGF – vascular endothelial growth factor, PI3K – phosphoinositide 3-kinase, MAPK – mitogen-activated protein kinase, miR-4729 – microRNA-4729, SH2/SH3 – Src homology 2/3 domains). Created in BioRender. Mazurek, M. (2025) https://BioRender.com/158e9db.
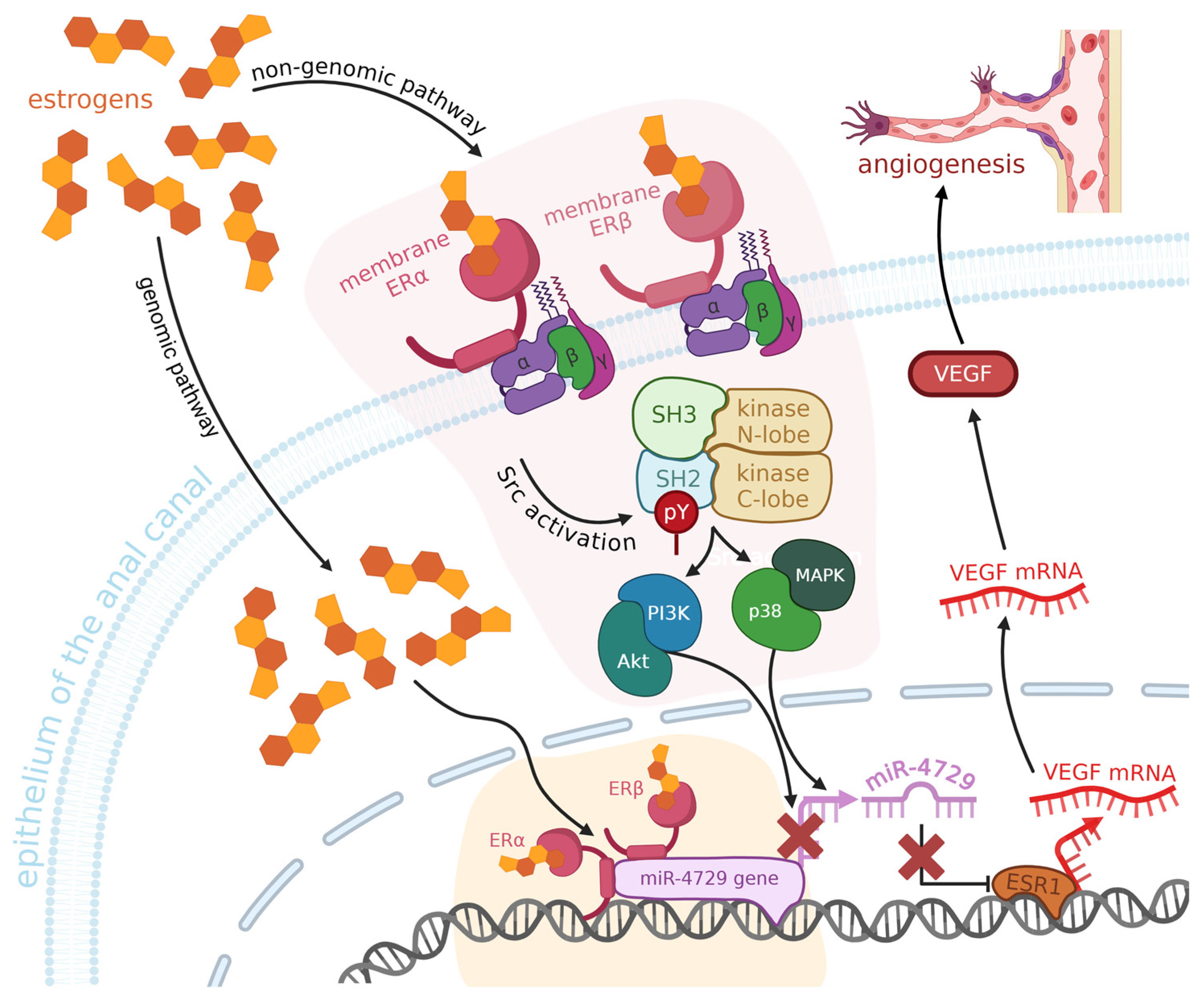
Figure 4.
Hemorrhoid development is influenced by epigenetic and lifestyle factors. A low-fiber diet alters gut microbiota, affecting gene expression. Obesity increases intra-abdominal pressure and inflammation via cytokines, while constipation, present in IBS and IBD, raises venous pressure and degrades connective tissue, promoting varicose vein formation (TNF-α – tumor necrosis factor alpha, IL – interleukin, VEGF – vascular endothelial growth factor, IBS – irritable bowel syndrome, IBD – inflammatory bowel disease). Created in BioRender. Mazurek, M. (2025) https://BioRender.com/rnrh8xd.
Figure 4.
Hemorrhoid development is influenced by epigenetic and lifestyle factors. A low-fiber diet alters gut microbiota, affecting gene expression. Obesity increases intra-abdominal pressure and inflammation via cytokines, while constipation, present in IBS and IBD, raises venous pressure and degrades connective tissue, promoting varicose vein formation (TNF-α – tumor necrosis factor alpha, IL – interleukin, VEGF – vascular endothelial growth factor, IBS – irritable bowel syndrome, IBD – inflammatory bowel disease). Created in BioRender. Mazurek, M. (2025) https://BioRender.com/rnrh8xd.
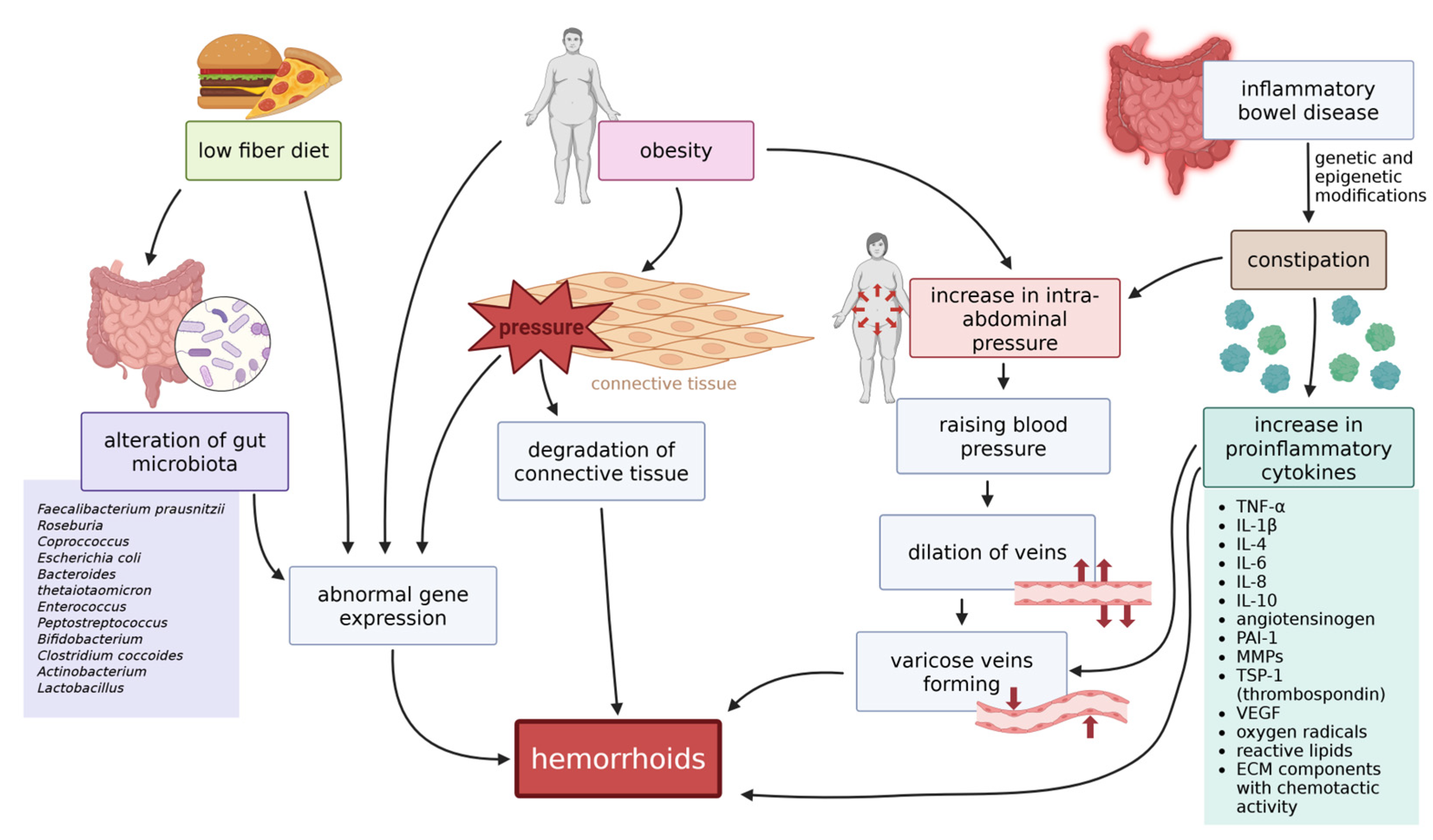
Figure 5.
The diagram above demonstrates the process of gathering evidence while performing this review.
Figure 5.
The diagram above demonstrates the process of gathering evidence while performing this review.
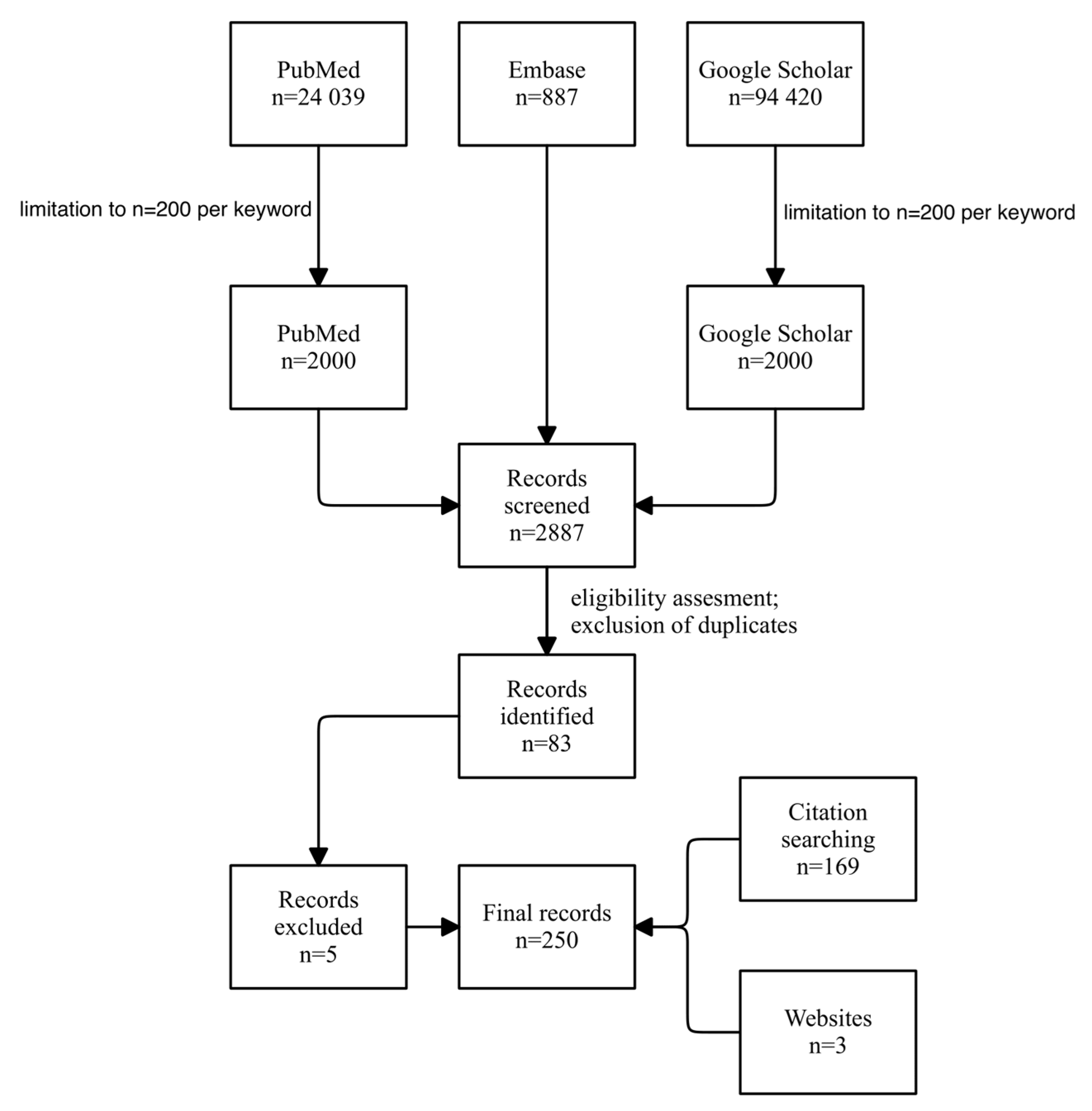
Table 1.
Cytokines presented in the table were identified in hemorrhoidal tissues.
| Cytokines Expressed in Hemorrhoids | |
|---|---|
| Pro-inflammatory Cytokines | Anti-inflammatory Cytokines |
| RANTES | IL-10 |
| TNF-α | ·· |
| VEGF | ·· |
| IL-1β | ·· |
| IL-6 | ·· |
| IL-8 | ·· |
| IL-17 | ·· |
| IFN-γ | ·· |
Table 2.
MiRNAs differently expressed in hemorrhoids seem to target genes that influence various vesical pathways.
Table 2.
MiRNAs differently expressed in hemorrhoids seem to target genes that influence various vesical pathways.
| MiRNAs in vesical pathways in hemorrhoids | MiRNAs in extracellular vesical pathways in hemorrhoids | |||
|---|---|---|---|---|
| Probable targeted processes * | Endocytosis | Synaptic vesicle pathways | Transcription, protein kinase activity, and ubiquitination | Transcriptional activator activity |
| Status | Upregulated | Downregulated | Upregulated | Downregulated |
| Type of miRNA | miR-375 | miR-376b-3p | miR-6741-3p | miR-548t-5p |
| miR-215-5p | miR-34a-5p | miR-6834-3 | miR-323b-5p | |
| miR-192-5p | miR-152-3p | miR-425 | miR-1322 | |
| miR-143-3p | let-7c-5p | miR-6804-3 | miR-3928-5p | |
| miR-187-3p | miR-107 | miR-744-3 | miR-346 | |
| miR-194-5p | miR-517a-3p | miR-848 | miR-4704-5p | |
| miR-145-5p | miR-517b-3p | miR-299-5 | miR-1913 | |
| miR-490-3p | miR-1307-5p | miR-463 | miR-876-3p | |
| miR-145-3p | miR-190a-5p | miR-317 | miR-4460 | |
| ·· | miR-378a-5p | miR-465 | miR-892a | |
| ·· | miR-708-3p | ·· | ·· | |
| ·· | miR-450a-5p | ·· | ·· | |
| ·· | miR-30e-5p | ·· | ·· | |
| ·· | miR-532-5p | ·· | ·· | |
* These are processes associated with genes that the listed miRNAs possibly target, according to available databases.
Table 3.
Genes displayed in the table were proposed or reported to contribute to the pathogenesis of hemorrhoids (“—” applies to unknown, unavailable values).
Table 3.
Genes displayed in the table were proposed or reported to contribute to the pathogenesis of hemorrhoids (“—” applies to unknown, unavailable values).
| Genes Significant to the Development of Hemorrhoids | ||||||
|---|---|---|---|---|---|---|
| Gene | Product | Function | Mechanism of Action | Mutations | Reported Associated Conditions | |
| FOXC2 | FOXC2 (forkhead box protein) [143,144] |
|
— | |||
| NOX1 and NOS3 | NOX1 (NADPH oxidase) NOS3 (nitric oxide synthase) |
|
— | |||
| MTHFR | methylenetetrahydrofolate reductase |
|
— |
|
||
| MYH9 | heavy chain of non-muscle myosin IIA |
|
— |
|
||
| F5 | coagulation factor V |
|
— |
|
||
| CYP1A | aryl hydrocarbon hydroxylase in hepatic and extrahepatic cytochrome P450 | — | — |
|
|
|
| PON1 | serum paraoxonase 1 | — | — |
|
||
| CALM3 | calmodulin 3 |
|
— | — |
|
|
| ANO1 | anoctamin-1 (voltage-gated calcium-activated anion channel) |
|
|
|
— | |
| SPRX | — |
|
|
|
— | |
| ACHE | acetylcholinesterase |
|
— |
|
||
| SRTT | capped-RNA binding protein | — | — | — |
|
|
| GSDMC | gasdermin C |
|
— |
|
|
|
| MYH11 | muscle myosin heavy chain 11 |
|
|
|
|
|
| ELN | elastin |
|
— | — |
|
|
| COL5A2 | type V collagen (regulatory) | — | — | — | ||
| PRDM | histone methyltransferase |
|
— | — |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



