
Submitted:
18 July 2025
Posted:
21 July 2025
You are already at the latest version
Abstract
Background: Wildfires represent a recurrent disturbance in the Patagonia Andean region with increasing frequency in recent decades. Austrocedrus chilensis is an arbuscular-mycorrhizal (AM) endemic conifer particularly vulnerable to fire, a situation that will worsen with climate change. In February 2015, a wildfire affected 5700ha of Austrocedrus chilensis forests with varying severities (WFS). The aim of this study was to determine and compare the density of AM spores (AMS) in soil affected by different WFS and non-affected sites, considering site features. Methods: Ninety soil samples were collected from three sites 10 months and 5 years after fire. AMS’ density was determined, a bioassay was set, and soil physicochemical features were evaluated. Results: after the wildfire, spores were significantly more abundant in sites affected by moderate severity fires. After 5 years, burned sites showed no differences between severities, but the driest sites showed no changes since fire occurred. Seedlings from the bioassay showed less than 25% mycorrhizal colonization growing in soil from burned sites, regardless fire severity compared with unburned soils. Conclusions: For restoration strategies, it must be considered that wildfires reduce mycorrhizal spores and mycelium, alter soil properties, and that drier conditions have less resilience, requiring active restoration.
Keywords:
AM inocula
; restoration
; native forests
; forests fire
1. Introduction
Wildfire has been a recurrent disturbance in the Patagonia Andean region, causing significant environmental, social and economic impacts similar to those in other countries around the world [1,2]. In these forests, the natural fire regime varies according to the precipitation regime. In general, fire frequency increases from the humid sites in the west, towards the xeric sites in the East [3]. In the past, fire frequency increased in the majority of A. chilensis stands after 1850, coincidentally with an increase in number of indigenous people that inhabited the region and the beginning of European settlement. This frequency peaked at the end of the 19th century, and started to decline from then on because indigenous population decreased, and the creation of several National Parks in the region brought about the policy of fire suppression [1]., while during the last three decades, the number of wildfire ignitions has risen in the whole central Patagonia Andean region [4,5]. As rains are markedly limited to autumn-winter in the region, summer provides favorable conditions for wildfires occurrence, combining the greatest water deficit and the highest temperatures [6]. Moreover, in a climate change scenario, with predictions of 1°C to 3°C temperature increase, and a 10% to 30% precipitation decrease for West Patagonia [7,8], wildfire frequency is expected to rise [9,10]
Austrocedrus chilensis (D. Don) Pic. Sern et Bizarri is an endemic and emblematic conifer of northern Patagonia forests, one of the most affected by wildfires [11]. It grows in pure or in mixed stands interspersed with Nothofagus dombeyi Mirb. Oerst. and other tree species between 37°07′and 43°44′ S latitude [12]. Austrocedrus chilensis is an obligate seeder [13,14] that does not resprout after fires [15], highly vulnerable to fire given the thin bark and persistent, dry, branches that favor fire access to the crown [16], and the high resin foliage contents that burns explosively [17]. It forms extensive stands valuable for forestry and tourism, and of significant ecological importance, covering an area of 100.000 ha [18]. However, fire and diseases [19,20,21], invasions by other plant species [22]as well as herbivory on saplings after fires [23] have generated severe degradation, resulting in great economic and environmental losses [24]. It has been declared as a ‘near threatened’ species by The IUCN (International Union for the Conservation of Nature) since 2013.
Regarding its mycorrhizal status, A. chilensis is an obligated AM species [25,26]. This mutualism is of great ecological importance, involved in seedlings establishment and tree growth, with major roles in absorption, transport and translocation of nutrients (P, N, C, K and Ca) [27], increased water availability and tolerance to drought [28,29], and resistance to pathogen attack [30]. On the other hand, AM Fungi (AMF) improves soil physicochemical properties [31] and plays a role in nutrient cycling [32], serving as a key element of ecosystem health and supporting vegetation recovery following degradation [33]. Nevertheless, AMF are highly susceptible to fire [34,35,36], especially mycelial structures and colonized AM root tips. Although, AM spores (resistance structures) have been reported as not drastically affected by fire [37,38,39]; fire heat could even break spore dormancy [40]. Anyway, fire can reduce AMF diversity compared with unburned areas [34,41,42] but, if edaphic, weather and vegetation conditions are favorable, AM community could recover in a few decades [43,44], contributing to a quick ecosystem recovery after fire [45,46]. Although mycorrhizas are crucial for maintaining biodiversity and ecosystem function, there is surprisingly limited knowledge regarding the landscape-scale biogeographical patterns of AMF species and the environmental factors that affect their distributions, including their response to fire severity. Empirical work has linked certain edaphic properties (e.g., soil texture, pH, nitrogen, phosphorus) to AMF distributions [47,48,49]. Dispersal capabilities of AMF are likely species-specific and environmentally dependent [50], although the mechanisms are poorly understood and have not been addressed for A. chilensis forest.
During February of 2015, near to Cholila (Chubut, Argentina), an extreme behavior wildfire, the most severe in the past century, occurred. From the total 27,101 ha burned, approximately 5700 ha corresponded to pure and mixed A. chilensis forests [5]. Therefore, it constituted a suitable scenario to assess the AM spore bank´s post fire condition, understand its evolution over time to properly manage restoration actions. In this sense, the aims of this study were 1) determine and compare over time the abundance of AMS in A. chilensis forest areas affected by different WFS, in relation to non-affected patches, and considering sites features and 2) analyze the behavior of the AM spore bank through its infectivity on A. chilensis seedlings analyze the behavior of the AM spore bank of those burned forest areas on A. chilensis seedlings.
2. Materials and Methods
2.1. Study Area
This study was carried out in three pure stands of A. chilensis forest affected by 2015 wildfire, located near “Las Horquetas” in Villegas Valley (A), Tigre River (B) and Cholila lake (C) in Chubut province (Figure 1). Three different fire severities were selected as treatments in each location. Fire severity was categorized considering the damage in trees, following [51]: unburned forests (green areas of forest), moderately affected forests (areas with partial tree burned), and severely affected forests (areas with total trees burned) (Figure 1). The slope ranged between 10 % and 35 % in all sites, while the aspect was mostly southeast, except for unburned site A and severely affected site B (northeast) and unburned site B (southwest). Site A Annual precipitation ≅ 2000 mm; Site B annual precipitation ≅ 1500 mm; Site C Annual precipitation ≅ 800 mm [52].
2.2. Sampling Design
During January and February 2016, a sampling plot of ≅ 100 m2 was set and sampled for each treatment / site and revisited and sampled in February 2021. In each plot five soil subsamples of approximately 500 g, from the first 15 cm of soil, were collected and placed in new plastic bags and stored in a refrigerator at 4 ° C until the AMS’ evaluation. A total of 90 soil samples were obtained (2016 and 2021) which were processed in duplicate (180 subsamples in total).
In addition, three composite soil samples (~630 cm3 each) of the upper 5 cm of mineral soil were collected at each sampling site in 2016 (27 soil samples) to determine physicochemical parameters.
2.3. Sample Processing; Spore and Soil Analyses
For the extraction of soil AMS, the wet sieving and sucrose gradient method (adapted from [53]) was used. Soil subsamples of 50 to 75 g per treatment/site were placed in a 20% sodium pyrophosphate solution for 60 minutes, sieved with distilled water using a 25 μ sieve; the obtained fraction was centrifuged at 2000 rpm for 5 minutes to precipitate the AMS. After discarding the supernatant, 50 ml of 50% sucrose solution was added, homogenized and centrifuged at 2000 rpm for 5 minutes. The supernatant was filtered on filter paper (103 slow, XinXing®), placed in a Petri dish and counted under stereomicroscope. Density of AMS was expressed as the amount of AMS per 100 g of dry soil by treatment [54].
Soil analyses were carried out at CIEFAP Soil Laboratory with dried 2-mm sieved fraction of soil. Physicochemical parameters analyzed were: (1) current pH 1:2.5 soil/distilled water ratio [55]; (2) electrical conductivity (EC, ds/m) [56]; (3) organic matter (OM%) [57]; soil organic carbon (C) was estimated from the ratio between organic matter and the van Bemmelen factor of 1.724 [58]; (4) total nitrogen (N, %) by obtained by Kjeldahl method [59]; (5) available phosphorus (P, mg/kg) estimated by the Olsen method [60], recommended for slightly acidic soils such as Andisols [61]; (6) cation exchange capacity (CEC) [62]; and (7) exchangeable bases, to determine calcium (Ca, meq/100g) and magnesium (Mg, meq/100g), sodium (Na, meq/100g) and potassium (K, meq/100g) [63].
2.4. Bioassay Setup
A soil bioassay, to know AMS real colonization capacity, was conducted with soil sampled 10 months after fire. Austrocedrus chilensis seeds were surface sterilized in 10 % sodium hypochlorite for 10 min, sown in sterile distilled water [64], and then hydrated for 24 h in sterile distilled water before storing them at 4 C in a refrigerator for 45 days to stratify (adapted from [65]). Non-AM seedlings were obtained in a growth chamber at 17-19°C, 48–55% relative air humidity, and 16 h photoperiod with 1400 lux radiation with sterilized pumicite (autoclave 3 times at 120 C for 30 min; adapted from [66]) for 30 d. Three weeks later, 2 seedlings were transplanted to a clean, 250 cm3 pot filled with a 1:1 (v/v) mix of soil obtained from each sampling unit and sterilized pumicite (as mentioned above). As a control, 10 pots were filled with mixed (1:1, v/v) sterilized soil (mixing soil from each treatment from all sites) and sterilized pumicite, both autoclaved as previously mentioned. Seedlings were randomly arranged and grown for 18 months in a growing chamber, under the same set up described for germination.
The root system of each seedling was cut into 10 mm lengths portions (approx. 600 mg per seedling) to fit in Tissue-Tek plastic capsules (Fisher Scientific Co., Pittsburgh, PA), cleared in 10% KOH for 30 min at 100°C under water bath and 15% H2O2 overnight at room temperature. Cleared samples were immersed 60 min at 4 C in a staining solution of 0.05% trypan-blue in lactoglycerol, rinsed with tap water and stored in lactoglycerol at 4 C until microscopic examination [67]. Arbuscular mycorrhizal colonization percentage for each seedling root system (AM%) was estimated following [68], using the complete root system, as:
AM%= (number mycorrhizal intersects/total intersects)*100
All AM structures (arbuscules, coils, vesicles and hyphae) were count separately and considered as mycorrhizal intersects for AM%.
2.5. Statistical Analysis
Spore density and AM% were analyzed using a two-way generalized linear mixed model (2W-GLMM) with the restricted maximum likelihood estimation method (RELM) with posterior comparisons with the test DGC (test of exclusionary groups). Sites (A, B, C) were treated as blocks (incorporated as a random effect) and different treatments (HIGH, MODERATE and UNBURNED wildfire severity) as fixed-effects [69] with R-DCOM in Infostat [69]. Arbuscular mycorrhizal structures (arbuscules, coils, vesicles and hyphae) were analyzed with a hierarchical clustering analysis using pheatmap, ggplot2 packages for RStudio. Soil physicochemical differences between sites and treatments were analyzed using two-way ANOVA, with treatments nested within each site, and site x treatment interactions. When variables showed significant interaction, treatments were analyzed within each site. Prior to the analysis, the assumptions of normality and homogeneity of variance were verified. To further analyze the relationships between seedlings mycorrhizal status and soil features, Pearson correlation tests and PCA were conducted, including AM%, AM structures, pH, OM%, electrical conductivity, content of N, Mg, P, K, Ca, Na.
3. Results
3.1. Analyses of Soil AMS Densities and Soil Variables
In total, 24.434 spores belonging to AMF were recovered from 180 soil samples (Table S1). Ten months after wildfire, AM spores’ density was significantly and consistently more abundant in moderately affected sites, compared both with the unburned or high affected sites (Figure 2A, 2W-GLMM, F= 5.14; p=0.0115). Also, the driest site (C) tripled the AMS density compared with the other sites for the moderately affected condition (Figure 2A).
After 5 years (2021), burned sites showed no differences in AMS densities between wildfire severities for sites A and B (Figure 2B). Unburned sites presented higher densities from the initial situation (Table S1), and significantly higher than burned treatments, except in the driest site (C) (2W-GLMM, F= 7.39; p=0.0022). Also, the AMS density patterns between treatment differed considering the site (Figure 2A, B). Interestingly, the driest site (C) did not show changes in AMS’ density after 5 years in burned sites.
Regarding soil physicochemical properties, pH was lower in Site A compared to B and C; high severity treatment presented significantly higher pH than the moderate, and both significantly higher than the unburned. Nitrogen was only significantly lower in site B compared to C, while both did not differ with site A; burned treatments from all sites consistently presented lower N than the unburned ones. The EC was significantly higher for the high severity compared to the unburned treatment, while those of moderate severity showed intermediate values. The OM, Na and C/N were consistently higher in the unburned treatment compared to both burned ones. Potassium and Mg evidenced differences only in site B, where the unburned treatments had a significantly higher concentration than the burned ones. Calcium concentration was significantly higher in unburned treatment, compared to both severities in sites B y C, while only to moderate severity in site A (Table 1).
3.2. Seedlings AM Colonization from Soil Bioassay
The AM % of 100 A. chilensis seedling from the bioassay showed significant differences between wildfire severities treatments, and both significantly lower values compared to the unburned treatment (Figure 3B, Table S2). The AM% for the moderate severity treatment represents 33,8% of the unburned treatment value, while the high severity treatment only 12 %. Control seedlings showed no AM colonization (Figure 3, Table S2), and lead to non-significant differences with the high severity treatment. Consequently, seedlings from the unburned treatment showed the greatest amounts of AM nutrient and photosynthate exchange structures (arbuscules, coils), while seedlings grown in soil affected by wildfire showed mostly explorations and storage of nutrient and photosynthate structures (hyphae and vesicles) but in small amounts especially in wet sites (A= Villegas valley) (Figure 3A, Table S2).
3.3. Relationships Between Seedlings AM Colonization, Spore Density and Soil Features
Arbuscular mycorrhizal colonization in A. chilensis seedlings was negatively correlated with soil pH and EC and positively with OM% and C/N (Pearson correlation coefficient, r= -0.7, p= 0.05; r= -0.7, p= 0.05; r= 0,8, p= 0,02; r= 0.8, p=0.02; respectively). Arbuscles and hyphae abundance were positively correlated with OM% and C/N and negatively correlated with soil pH (Pearson correlation coefficient, r= 0.9, p=0.003; r= 0.8, p=0.01; r= 0.8, p= 0.01; r= 0.8, p= 0.01; r= -0.7, p= 0.03; r= -0.8, p= 0.02, respectively). Coils’ abundance was positively correlated with OM%, Mg content and C/N (Pearson correlation coefficient, r=0.92, p=0,0005; r= 0.7, p= 0.03; r= 0.95, p= 0.0001, respectively). Finally, the vesicles’ abundance was negatively correlated with soil electrical conductivity and positively with OM% and C/N (Pearson correlation coefficient, r= 0,7, p=0,03; r= -0.7, p=0.05; r= 0.7, p=0.05, respectively). On the other hand, spore density (2016) was not correlated with any soil feature (Table S3).
Principal component analysis (PCA) showed that the combination of AM structures AM% and soil features (Figure 4) could be explained by two components, including 69.6% of variance (Table S4, S5); PC1 and PC2 explained 43.1% and 26.5% of the variance, respectively. PC1 discriminates between burned treatments from the unburned, while PC2 discriminates between sites (A= Villegas valley, B= Tigre River and C= Cholila lake).
The N, K, Ca, Mg content (EV=0.715; -0.701; -0.818; -0.680, respectively), arbuscles, vesicles, coils, hyphae abundance (EV= -0.732, -0.788, -0.798, -0.881) AM% (EV=-0.816) were spatial variables that contributed to PC1, grouping variables affected by fire. On the other hand, pH (EV=-0.657) OM % (EV=0.751) and Na content (EV=0.736) contribute to PC2, grouping site dependent variables.
4. Discussion
Arbuscular mycorrhizal colonization and spore-based studies are a gap of knowledge for South American Austrocedrus chilensis forests, except for few works about plant mycorrhizal status from last century [25,26,72]. For this reason, this study represents the first analysis regarding bank evolution and infectivity after wildfires for these native ecosystems. Our results show that wildfire events had direct effects on the AMS communities associated with A. chilensis forests, and that these effects are affected and vary according to fire severity and site conditions. It has been already stated that fire reduce root AM colonization [73,74] but increase AMS density in the soil [75]. In our study, immediately after wildfire occurs, AMS density was significantly more abundant in the moderately affected treatment in the dry site (C), compared both with unburned to highly affected treatment at the same site and with all treatments from the other wetter sites. On the other hand, seedlings growing on soils affected by wildfires showed low or null AM colonization regardless of WFS. After a wildfire, the surviving soil microecosystem and their ability to recover are essential to forest succession and restoration [76,77,78]. For those microorganisms beneath the surface, soil is a good insulator, especially when it is dry, and the temperature at 2.5 cm depth may be 50°C when the surface is 100°C [79]. In this sense, the results of this work showed that soon after fire, moderate severity wildfire were associated with higher AMS densities, and that this effect was more pronounced in dryer site conditions. In those xeric sites, spore abundance does not recover even 5 years after the event. [37] found that AM communities are resilient to wildfire on decadal timescales; this resilience appears to depend on the post-fire regrowth of understory vegetation and the subsequent recovery of soil chemical properties. However, regarding the direct effects of wildfire on AM fungi, results have been contradictory, reporting either negative e.g., [80,81,82,83,84,85], neutral e.g., [34,45,86,87,88], or positives [89].
[90] found that natural Austrocedrus chilensis regeneration could only be expected to occur in patches that have been unaffected by fire, or in areas affected by low severity wildfires because of low seeds availability. The potential for natural regeneration of this species following large-scale disturbances that eliminate or drastically reduce the forest canopy is limited [11]. The emergence of Austrocedrus chilensis seedling pattern varied with site conditions and forest management practices [91]. Additionally, A. chilensis seedling survival improves if early successional herbaceous species and the burned canopy are not removed [11]. In this sense, herbaceous and shrubs species from A. chilensis understory tended to increase their presence with wildfire [17,92]; as most of them harbor AMF [26,93]they could improve AM inoculum availability and colonization of new A. chilensis seedling, although the specificity of A. chilensis-associated AM fungi in relation to understory AM fungal species has not yet been studied. We also found low seedling colonization, mostly with explorations and storage structures (hyphae and vesicles), in treatments with soils of the severely burned forests. Despite the fact that very little is known in this regard, considering that AM symbiosis is crucial to new plant establishment [94], our findings indicate that inoculation with AM fungi should be recommended for the ecological restoration of forests affected by high WFS. Further research should examine its effects on seedling establishment in post-fire plantations.
Post-fire impacts include significant alteration of soil properties, such as pH, OM%, EC, N, C/N ratio, and CEC [51]. It was stated that soil pH increases after wildfires [92] due to the combustion of organic matter [95]. The direct impact of lower pHs on AMF is the limited sporulation and germination of spores in the rhizosphere; soils with neutral pH revealed greater amounts of AMF spores compared with acidic, pH 5.5 soils [96]. When pH rises, as found in burned soils in this work, then a stimulation of AMF sporulation can occur, increasing spore densities as found in sites moderated affected by fire. Arbuscular mycorrhizal fungi can also be stimulated by organic matter contents [27,97]. In this sense, our seedlings revealed AM structures colonization consistently associated with OM soil content, even when wildfire, regardless of WFS, showed significantly lower percentage of OM.
Arbuscular mycorrhizal spore density in natural soils/ environments is highly variable [27] and experimental studies have shown that several factors could be involved in AMS sporulating, such as climatic, geological and/or edaphic features, plant associated phenology, human activities, soil legacy [27,98,99,100]. In this sense, unburned sites in this study showed a noticeable increase in AMS density after 5 years. Although our sampling design does not allow us to establish causalities, considering sampling plots and methodology were not changed, it could be hypothesized that these differences could be related to previous climatic conditions to each sampling period (January-February 2016 and 2021). In average, the four autumns prior to the first sampling were warmer and drier, with less days with frost in 2016 than in 2021 (Table S6). Other studies have found that warming [101,102] and drought conditions [103] significantly decrease AMS abundance. More local studies are needed to unravel this issue, understand its dynamic and predict evapotranspiration effects on the AMS bank.
Thinking of restoration strategies, it must be considered that wildfire would also cause extramatrical mycelium loss, although some stimulus occurs in moderately affected soil that increased spores’ abundance. Moderately burned areas have better chances of natural restoration considering the abundant AM spores bank, while artificial inoculation must be considered for severely affected areas or in burned dry sites.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, M.E.S.S., M.F.U. and C.B.; methodology, M.E.S.S., validation, MESS and C.B.; formal analysis, M.E.S.S. and MFU; investigation, M.E.S.S., S.S.T., S.G., A.D.E.; resources, MESS and M.F.U.; data curation, M.E.S.S., A.D.E.; writing—original draft preparation, M.E.S.S.; writing—review and editing, M.E.S.S., M.F.U., C.B.; visualization, M.E.S.S.; supervision, M.E.S.S.; project administration, M.E.S.S. and MFU.; funding acquisition, M.E.S.S. and MFU. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially supported by PICT Startup Nº 0704/2016. Author MESS has received research support from Dirección Nacional del Fondo para la Investigación Científica y Tecnológica (FONCYT), Agencia Nacional de Promoción de la Investigación, el Desarrollo Tecnológico y la Innovación. This work was partially supported by Project PE7A11703. Author MFU has received research support from Centro de Investigación y Extensión Forestal Andino Patagónico (CIEFAP).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data was created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
We thank the residents and farms keepers for allowing us to carry out the field work, and Virginia Alonso CIEFAP Soil laboratory chief. ADE and SG are Technician, MESS, MFU and CB are researchers for the National Research Council of Argentina (CONICET).
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| AMF | Arbuscular mycorrhizal fungi |
| AMS | Arbuscular mycorrhizal spores |
| AM% | Arbuscular mycorrhizal colonization percentaje |
| WFS | Wildfire severity |
| AM | Arbuscular mycorrhiza |
| OM | Soil organic matter |
| N | Soil Nitrogen content |
| EC | Soil Electrical conductivity |
| C/N | Soil carbon nitrogen ratio |
| 2W-GLMM | Two-way general linear mix model |
| PCA | Principal component analysis |
References
- Veblen, T.T., Kitzberger, T., Villalva, R., Donnegan, J. Fire History in Northern Patagonia: The Roles of Humans and Climatic Variation. Ecol Monogr, 1999, 69(1), 47–67. [CrossRef]
- Kalogiannidis, S., Chatzitheodoridis, F., Kalfas, D., Patitsa, C., Papagrigoriou, A. Socio-Psychological, Economic and Environmental Effects of Forest Fires. Fire, 2023, 6, 280. [CrossRef]
- Kitzberger, T., Veblen, T.T., Fire-induced changes in northern Patagonian landscapes. Landsc Ecol, 1999, 14, 1–15 http://link.springer.com/article/10.1023/A:1008069712826. [CrossRef]
- de Torres Curth, M.I., Ghermandi, L., Pfister, G. Los incendios en el noroeste de la Patagonia: su relación con las condiciones meteorológicas y la presión antrópica a lo largo de 20 años. Ecología Austral, 2008, 18, 153–167. [CrossRef]
- Mohr Bell, D. Superficies afectadas por incendios en la región Bosque Andino Patagónico durante los veranos de 2013-2014 y 2014-2015. Nodo Regional Bosque Andino Patagónico (SAyDS -CIEFAP). 2015.
- Kitzberger, T., Tiribelli, F., Barberá, I., Gowda, J.H., Morales, J.M., Zalazar, L., Paritsis, J. Projections of fire probability and ecosystem vulnerability under 21st century climate across a trans-Andean productivity gradient in Patagonia. Sci Total Environ, 2022, 839, 156303. [CrossRef]
- Barros. V.R., Boninsegna, J.A., Camilloni, I.A., Chidiak, M., Magrín, G.O., Rusticucci, M. Climate change in Argentina: trends, projections, impacts and adaptation. WIREs Clim Change, 2015, 6(2), 151–169. [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Masson-Delmotte, V., P. Zhai, A. Pirani, S. L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M. I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J.B.R. Matthews, T. K. Maycock, T. Waterfield, O. Yelekçi, R. Yu and B. Zhou (eds.). Cambridge University Press. 2021.
- González, M.E., Lara, A., Urrutia, R., Bosnich, J. Cambio climático y su impacto potencial en la ocurrencia de incendios forestales en la zona centro-sur de Chile (33º-42º S). Bosque, 2011, 32(3), 215–219. [CrossRef]
- Mundo, I.A., Villalba, R., Veblen, T.T., Kitzberger, T., Holz, A., Paritsis, J., Ripalta, A. Fire history in southern Patagonia: human and climate influences on fire activity in Nothofagus pumilio forests. Ecosphere, 2017, 8(9), e01932. [CrossRef]
- Urretavizcaya, M.F., Defossé, G.E. Restoration of burned and post-fire logged Austrocedrus chilensis stands in Patagonia: effects of competition and environmental conditions on seedling survival and growth. Int. J. Wildland Fire, 2019, 28(5), 365. [CrossRef]
- Pastorino, M.J., Fariña, M.M., Bran, D., Gallo, L.A. Extremos geográficos de la distribución natural de Austrocedrus chilensis (Cupressaceae). Boletín de la SAB, 2006, 41, 307–311.
- Urretavizcaya, M.F.; Defossé, G. Soil seed bank of Austrocedrus chilensis (D. Don) Pic. Serm. et Bizarri related to different degrees of fire disturbance in two sites of southern Patagonia, Argentina. Forest Ecol Manag, 2004, 187, 361-372. [CrossRef]
- López Bernal, P.M., Urretavizcaya, M.F., Defossé, G.E. Seedling dynamics in an environmental gradient of Andean Patagonia, Argentina. In From seed germination to young plants: ecology, growth and environmental influences; Busso, C.A., Ed.; Publisher: Nova Science Publishers, Hauppauge, NY, 2013, pp. 189–210.
- Gallo, L., Pastorino, M.J., Donoso, C. Variación en Austrocedrus chilensis (D. Don) Pic. Ser et Bizzarri (Ciprés de la Cordillera). In Variación intraespecífica en las especies arbóreas de los bosques templados de Chile y Argentina; Donoso, C., Premoli, A., Gallo, L., Ipinza, R., Eds.; Publisher: Marisa Cuneo, Valdivia, Chile, 2004, pp. 233–250.
- La Manna, L., Bava, J., Collantes, M., Rajchenberg, M. Características estructurales de los bosques de Austrocedrus chilensis afectados por “mal del ciprés” en Patagonia, Argentina. Bosque, 2006, 27(2), 135–145.
- Gobbi, M., Sancholuz, L. Regeneración pos-incendio del ciprés de la cordillera (Austrocedrus chilensis) en los primeros años. Bosque, 1992, 13(2), 25–32.
- CIEFAP, MAyDS. Actualización de la Clasificación de Tipos Forestales y Cobertura del Suelo de la Región Bosque Andino Patagónico. Informe Final https://drive.google.com/open?id=0BxfNQUtfxxeaUHNCQm9lYmk5RnM CIEFAP. 2016, 111 p.
- Bava, J., Lencinas, J.D., Haag, A. Determinación de la materia prima disponible para proyectos de inversión forestales en cuencas de la provincia del Chubut. Informe Parcial. Consejo Federal de Inversiones. 2006, 85 pp.
- Morales, D., Rostagno, C.M., La Manna, L. Runoff and erosion from volcanic soils affected by fire: The case of Austrocedrus chilensis forests in Patagonia. Argentina. Plant Soil, 2013, 370(1), 367–380. [CrossRef]
- Vélez, M.L., La Manna, L., Tarabini, M., Gomez, F., Elliott, M., Hedley, P.E., Cock, P., Greslebin, A. Phytophthora austrocedri in Argentina and co-inhabiting Phytophthoras: Roles of anthropogenic and abiotic factors in species distribution and diversity. Forests, 2020, 11(11), 1223. [CrossRef]
- Orellana, I.A., Raffaele, E. The spread of the exotic conifer Pseudotsuga menziesii in Austrocedrus chilensis forests and shrublands in northwestern Patagonia, Argentina. N Z J For Sci, 2020, 40, 199–209.
- Urretavizcaya, M.F. Propiedades del suelo en bosques quemados de Austrocedrus chilensis en Patagonia, Argentina. Bosque, 2010, 31(2), 140–149. [CrossRef]
- Loguercio, G.A., Urretavizcaya, M.F., Caselli, M., Defossé, G.E. Propuestas silviculturales para el manejo de bosques de Austrocedrus chilensis sanos y afectados por el mal del ciprés de Argentina. In Silvicultura en bosques nativos. Experiencias en silvicultura y restauración en Chile, Argentina y el oeste de Estados Unidos; Donoso, P.J., Promis, A., Soto, D.P., Eds.; Publisher: Imprenta América, Valdivia, Chile, 2018, pp. 117–134. () ISBN 978–0-692–09238–5.
- Godoy, R., Mayr, R. Caracterización morfológica de micorrizas vesículo-arbusculares en coníferas endémicas del sur de Chile. Bosque, 1989, 10(2), 89–98.
- Fontenla, S., Godoy, R., Rosso, P., Havrylenko, M. Root associations in Austrocedrus forests and seasonal dynamics of arbuscular mycorrhizas. Mycorrhiza, 1998, 8, 29–33. [CrossRef]
- Smith, S.E., Read, D.J. Mycorrhizal symbiosis. 3th ed. Publisher: Academic Press, Cambridge, UK, 2008, 145–187.
- Sendek, A., Karakoç, C., Wagg, C., Domínguez-Begines, J., Martucci do Couto, G., van der Heijden, M.G.A., Ahmad Naz, A., Lochner, A., Chatzinotas, A., Klotz, S., Gómez-Aparicio, L., Eisenhauer, N. Drought modulates interactions between arbuscular mycorrhizal fungal diversity and barley genotype diversity. Sci Rep, 2019, 9, 9650. [CrossRef]
- Abdalla, M., Bitterlich, M., Jansa, J., Püschel, D., Ahmed, M.A. The role of arbuscular mycorrhizal symbiosis in improving plant water status under drought. J Exp Bot, 2023, 74(16), 4808–4824. [CrossRef]
- Veresoglou, S. D., Rillig, M. C. Suppression of fungal and nematode plant pathogens through arbuscular mycorrhizal fungi. Biol Lett, 2012, 8(2), 214–217. [CrossRef]
- Willis, A., Rodrigues, B. F., Harris, P. J. C. The ecology of arbuscular mycorrhizal fungi. Crit Rev Plant Sci, 2013, 32(1), 1–20. [CrossRef]
- Powell, J. R., Rillig, M. C. Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol, 2018, 220(4), 1059–1075. [CrossRef]
- Neuenkamp, L., Prober, S. M., Price, J. N., Zobel, M., Standish, R. J. Benefits of mycorrhizal inoculation to ecological restoration depend on plant functional type, restoration context and time. Fungal Ecol, 2019, 40, 140–149. [CrossRef]
- Longo, S., Nouhra, E., Goto, B. T., Berbara, R. L., Urcelay, C. Effects of fire on arbuscular mycorrhizal fungi in the Mountain Chaco Forest. For Ecol Manage, 2014, 315, 86–94. [CrossRef]
- Cofré, N., Urcelay, C., Wall, L. G., Domínguez, L., Becerra, A. El potencial de colonización micorrícico-arbuscular varía entre prácticas agrícolas y sitios en diferentes áreas geográficas de la región Pampeana. Ecología Austral, 2018, 28(3), 581–592.
- Day, N.J., Dunfield, K.E., Johnstone, J.F., Mack, M.C., Turetsky, M.R., Walker, X.J., Baltzer, J.L. Wildfire severity reduces richness and alters composition of soil fungal communities in boreal forests of western Canada. Global Change Biol, 2019, 25(7), 2310–2324. [CrossRef]
- Xiang, X., Gibbons, S.M., Yang, J., Kong, J., Sun, R., Chu, H. Arbuscular mycorrhizal fungal communities show low resistance and high resilience to wildfire disturbance. Plant Soil, 2015, 397, 347–356. [CrossRef]
- Sun, J., Miller, J.B., Granqvist, E., Wiley-Kalil, A., Gobbato, E., Maillet, F., Maillet, F., Cottaz, S., Samain, E., Venkateshwaran, M., Fort, S., Morris, R. J., Ané, J., Dénarié, J., Oldroyd, G.E. Activation of symbiosis signaling by arbuscular mycorrhizal fungi in legumes and rice. Plant Cell, 2015, 27(3), 823–838. [CrossRef]
- Holden, S.R., Rogers, B.M., Treseder, K.K., Randerson, J.T. Fire severity influences the response of soil microbes to a boreal forest fire. Environ Res Lett, 2016, 11(3), 035004. [CrossRef]
- Claridge, A. W., Trappe, J. M., Hansen, K. Do fungi have a role as soil stabilizers and remediators after forest fire? For Ecol Manage, 2009, 257(3), 1063–1069. [CrossRef]
- Rashid, A., Ahmed, T., Ayub, N., Khan, A.G. Effect of forest fire on number, viability and post-fire re-establishment of arbuscular mycorrhizae. Mycorrhiza, 1997, 7, 217–220. [CrossRef]
- de Assis, D.M.A., De Melo, M.A.C., da Silva, D.K.A., Oehl, F., da Silva, G.A. Assemblages of arbuscular mycorrhizal fungi in tropical humid and dry forests in the Northeast of Brazil. Botany, 2018, 96(12), 859–871. [CrossRef]
- Orgiazzi, A., Bardgett, R.D., Barrios, E., Behan-Pelletier, V., Briones, M.J.I., Chotte, J-L., De Deyn, G.B., Eggleton, P., Fierer, N., Fraser, T., Hedlund, K., Jeffery, S., Johnson, N.C., Jones, A., Kandeler, E., Kaneko, N., Lavelle, P., Lemanceau, P., Miko, L., Montanarella, L., Moreira, F. M. S., Ramírez, K.S., Scheu, S., Singh, B.K., Six, J., van der Putten, W.H., Wall, D. H. (Eds.) Global soil biodiversity atlas. European Commission. European Commission, Publications Office of the European Union, Luxembourg, 2016, 176 pp.
- Whitman, T., Whitman, E., Woolet, J., Flannigan, M.D., Thompson, D.K., Parisien, M.A. Soil bacterial and fungal response to wildfires in the Canadian boreal forest across a burn severity gradient. Soil Biol Biochem, 2019, 138, 107571. [CrossRef]
- Dove, N.C., Hart, S.C. Fire reduces fungal species richness and in situ mycorrhizal colonization: a meta-analysis. Fire Ecol, 2017, 13(2), 37–65. [CrossRef]
- Saini, R., Sharma, S. Climate resilient microbes in sustainable crop production. In Contaminants in Agriculture and Environment: Health Risks and Remediation; Kumar, V., Kumar, R., Singh, J., Kumar, P, Eds.; Publisher: Agro Environ Media, Publication Cell, Agriculture and Environmental Science Academy, Haridwar (Uttarakhand), India, 2019, 265–283pp.
- Johnson, N.C., Tilman, D., Wedin, D. Plant and soil controls on mycorrhizal fungal communities. Ecology, 1992, 73, 2034–2042.
- Egerton-Warburton, L.M., Johnson, N.C., Allen, E.B. Mycorrhizal community dynamics following nitrogen fertilization: A cross-site test in five grasslands. Ecol Monogr, 2007, 77, 527–544. [CrossRef]
- Lekberg, Y., Gibbons, S.M., Rosendahl, S., Ramsey, P.W. Severe plant invasions can increase mycorrhizal fungal abundance and diversity. ISME J, 2013, 7, 1424–1433. [CrossRef]
- Egan, C., Li, D.-W., Klironomos, J. Detection of arbuscular mycorrhizal fungal spores in the air across different biomes and ecoregions. Fungal Ecol, 2014, 12, 26–31. [CrossRef]
- Urretavizcaya, M.F., Rago, M.M., Caselli, M., Ríos Campano, F., Gianolini, S., Alonso, V. Effect of fire severity and presence of bamboo (Chusquea culeou) on soil chemical properties in Andean Patagonian forests of Argentina. Int J Wildland Fire, 2025, 34, WF24011. [CrossRef]
- SUIE 2002 Sistema Unificado de Información Energética (SUIE). Secretaría de Energía de la Nación Argentina. 2002. Mapa de Isohietas República Argentina. Available in: https://sig.energia.gob.ar/visor/visorsig.php. Visited in December of 2023.
- Ianson, D.C., Allen, M.F. The Effects of soil texture on extraction of vesicular-arbuscular mycorrhizal fungal spores from arid sites. Mycologia, 1986, 78(2),164–168. [CrossRef]
- Allen, M.F., Moore, Jr., T.S., Christensen, M., Stanton, N. America growth of Vesicular-Arbuscular-Mycorrhizal and Nonmycorrhizal Bouteloua gracilis in a defined medium. Mycologia, 1979, Vol. 71(3), 666–669. [CrossRef]
- Bailey W. Soil Science. In Análisis químico de suelos; Jackson M.L., Ed.; Publisher: Ediciones Omega S.A. Barcelona, 1943, 55-143.
- Allison, L.E. Diagnóstico y rehabilitación de suelos salinos y sódicos. Publisher: Editorial Limusa, México, 1980.
- Davies, B E. Loss-on ignition as an estimate of soil organic matter. Soil Sci Proc, 1974, 38, 150. [CrossRef]
- Tabatabai, M.A. Soil Organic Matter Testing: An Overview. In Soil organic matter: analysis and interpretation; Magdoff, F.R.; Tabatabai, M.A., Hanlon Jr., E.A-, Eds.; Publisher: Soil Science Society of America, Madison, USA, 1996, 1–9.
- Bremmer JM. Determination of Nitrogen in soil by the Kjeldahl method. J Agr Sci, 1960, 55, 1–23. [CrossRef]
- Olsen, S.R., Cole, D.V., Watanabe, F.S., Dean, L.A. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. U.S.D.A. 939, USA, 1954, pp 19.
- Elrashidi, M.A. Soil and Water Conservation Advances in the United States: A review. J Soil Water Conserv, 2011, 66(1), 5A. [CrossRef]
- Richter, M., Conti, M., Maccarini, G. Mejoras en la determinación de cationes intercambiables y capacidad de intercambio catiónico en los suelos. Rev Fac Agronomía, 1982, 3(2), 145–155.
- Schollenber, C.J., Simon, R.H. Determination of exchange capacity and exchangeable bases in soil - ammnium acetate method. Soil Sci, 1945, 59,13–25.
- Sauer, D.B., Burroughs, R. Disinfection of seed surfaces with sodium hypochlorite. Phytopathology, 1986, 76(7),745–749.
- Schinelli Casares T. Producción de Nothofagus bajo condiciones controladas. Publisher: Ediciones INTA, Bariloche, Argentina, 2012, 52pp.
- van der Heijden, M.G.A., Klironomos, J.N., Ursic, M., Moutoglis, P., Streitwolf-Engel, R., Boller, T., Wiemken, A., Sanders, I.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Letter to Nature, 1998, 396, 69–72. [CrossRef]
- Cázares, E., Smith, J.E. Ocurrence of vesicular-arbuscular mycorrhizae in Pseudotsuga menziesii and Tsuga heterophylla seedlings grown in Oregon coast range soil. Mycorrhiza, 1996, 6, 65–67. [CrossRef]
- Brundrett, M., Bougher, N., Grove, T., Malajczuk, N. Working with mycorrhizas in forestry and agriculture. Australian Center for International Agricultural Reserch, Monograph 32. Canberra, Australia, 1996, 374pp.
- Di Rienzo, J.A., Macchiavelli, R.E., Casanoves, F. Modelos Mixtos en InfoStat. Manual del Usuario; Publisher: Editorial Brujas, Córdoba, Argentina, 2010.
- Di Rienzo, J.A., Casanoves, F., Balzarini, M.G., Gonzalez, L., Tablada, M., Robledo, C.W. InfoStat versión 2020. Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba, Argentina, 2020. URL: http://www.infostat.com.ar.
- RStudio Team. Posit team RStudio: Integrated Development Environment for R. Posit Software, PBC, Boston, MA. 2022, URL http://www.posit.co/.
- Carrillo, R., Godoy, R., Peredo, H. Simbiosis micorrícica en comunidades boscosas del Valle Central en el sur de Chile. Bosque, 1992, 13(2), 57–67. [CrossRef]
- Dhillion, S.S., Anderson, DC. Seasonal dynamics of dominant species of arbuscular mycorrhizae in burned and unburned sand prairies. Can J Bot, 1993, 71, 1625–1630. [CrossRef]
- Pattinson, G.S., Hammill, K.A., Sutton, B.G., McGee, P.A. Simulated fire reduces the density of arbuscular mycorrhizal fungi at the soil surface. Mycol Res, 1999, 103(4), 491–496. [CrossRef]
- Eom, A.-H., Hartnett, D.C., Wilson, G.W.T., Figge, D.A.H. The effect of fire, mowing and fertilizer amendment on arbuscular mycorrhizas in Tallgrass Prairie. Am Midl Nat, 1999, 142, 55–70. [CrossRef]
- Allen, E.B., Steers, R.J., Dickens, S.J. Impacts of fire and invasive species on desert soil ecology. Rangeland Ecol Manage, 2011, 64(5), 450–462. [CrossRef]
- Bouffaud, M.-L., Bragalini, C., Berruti, A., Peyret-Guzzon, M., Voyron, S., Stockinger, H., van Tuinen, D., Lumini, E., Wipf, D., Plassart, P., Lemanceau, P., Bianciotto, V., Redecker, D., Girlanda, M. Arbuscular mycorrhizal fungal community differences among European long-term observatories. Mycorrhiza, 2017, 27,331–343. [CrossRef]
- Soudzilovskaia, N.A., Vaessen, S., Barcelo, M., He, J., Rahimlou, S., Abarenkov, K., Brundrett, M.C., Gomes, S.I.F., Merckx, V., Tedersoo, L. FungalRoot: Global online database of plant mycorrhizal associations. New Phytol, 2020, 227, 955–966. [CrossRef]
- Neary, D.G., Klopatek, C.C., DeBano, L.F., Folliott, P.F. Fire effects on belowground sustainability: a review and synthesis. For. Ecol. Manag, 1999, 122, 51–71. [CrossRef]
- Chimal-Sánchez, E., Araiza-Jacinto, M.L., Román-Cárdenas, V.J. El efecto del fuego en la riqueza de especies de hongos micorrizógenos arbusculares asociada a plantas de matorral xerófilo en el Parque Ecológico “Cubitos”. Revista Especializada en Ciencias Químico-Biológicas, 2015, 18(2),107–115. [CrossRef]
- Barraclough, A.D., Olsson, P.A. Slash-and-burn practices decrease arbuscular mycorrhizal fungi abundance in soil and the roots of Didierea madagascariensis in the dry tropical forest of Madagascar. Fire, 2018, 1, 37. [CrossRef]
- Chávez, D., Machuca, A., Fuentes-Ramirez, A., Fernandez, N., Cornejo, P. Shifts in soil traits and arbuscular mycorrhizal symbiosis represent the conservation status of Araucaria araucana forests and the effects after fire events. For Ecol Manage, 2020, 458, 117806. [CrossRef]
- Kivlin, S.N., Harpe. V.R., Turner, J.H., Moore, J.A.M., Moorhead, L.C., Beals, K.K., Hubert, M.M., Papes, M., Schweitzer, J.A. Arbuscular mycorrhizal fungal response to fire and urbanization in the Great Smoky Mountains National Park. Elem Sci Anth, 2021, 9, 1–12. [CrossRef]
- Mino, L., Kolp, M.R., Fox, S., Reazin C., Zeglin, L., Jumpponen, A. Watershed and fire severity are stronger determinants of soil chemistry and microbiomes than within-watershed woody encroachment in a tallgrass prairie system. FEMS Microbiol Ecol, 2021, 97(12), fiab154. [CrossRef]
- Hopkins, J.R., McKenna, T.P., Bennett, AE. 2024. Fire season and time since fire determine arbuscular mycorrhizal fungal trait responses to fire. Plant Soil, 2024, 503, 231–245. [CrossRef]
- Docherty, K.M., Balser, T.C., Bohannan, B.J.M., Gutknecht, J.L.M. Soil microbial responses to fire and interacting global change factors in a California annual grassland. Biogeochemistry, 2012, 109,63–83. [CrossRef]
- Longo, S., Nouhra, E., Tecco, P.A., Urcelay, C. Functional stability of mycorrhizal interactions in woody natives and aliens facing fire disturbance. Plan Ecol, 2020, 221, 321–331. [CrossRef]
- Stürmer, S.L., Hackbarth Heinz, K.G., Marascalchi, M.N., Giongo, A., Siqueira, J.O. Wildfire does not affect spore abundance, species richness, and inoculum potential of arbuscular mycorrhizal fungi (Glomeromycota) in ferruginous Canga ecosystems. Acta Bot Brasil, 2022, 36, e2021abb0218. [CrossRef]
- Moreira, M., Baretta, D., Tsai, S.M., Nogueira Cardoso, E.J.B. Spore density and root colonization by arbuscular mycorrhizal fungi in preserved or disturbed Araucaria angustifolia (Bert.) O. Ktze. Ecosystems. Sci. Agric. (Piracicaba, Braz.), 2006, 63(4), 380-385. [CrossRef]
- Urretavizcaya, M.F., Defossé, G.E., Gonda, H.E. Effect of sowing season, plant cover, and climatic variability on seedling emergence and survival in burned Austrocedrus chilensis forests. Restor Ecol, 2012, 20(1), 131–140. [CrossRef]
- Gobbi, M., Schlichter, T. Survival of Austrocedrus chilensis seedlings in relation to microsite conditions and forest thinning. For Ecol Manage, 1998, 111, 137–146. [CrossRef]
- La Manna, L., Barroetaveña, C. Propiedades químicas del suelo en bosques de Nothofagus antarctica y Austrocedrus chilensis afectados por fuego. Rev. FCA UNCUYO, 2011, 43(1), 41–55.
- Marín, C., Aguilera, P., Oehl, F., Godoy, R. Factors affecting arbuscular mycorrhizal fungi of Chilean temperate rainforests. J Soil Sci Plant Nutr, 2017, 17 (4), 966–984. [CrossRef]
- Begum, N., Qin, C., Ahanger, M.A., Raza, S., Khan, M.I., Ashraf, M., Ahmed, N., Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front Plant Sci, 2019, 10, 1068. [CrossRef]
- Alcañiz, M., Outeiro, L., Francos, M., Úbeda, X. Effects of prescribed fires on soil properties: a review. Sci Total Environ, 2018, 613–614, 944–957. [CrossRef]
- Jamiołkowska, A., Księżniak, A., Gałązka, A., Hetman, B., Kopacki, M., Skwaryło-Bednarz, B. Impact of abiotic factors on development of the community of arbuscular mycorrhizal fungi in the soil: a Review. Int Agrophys, 2018, 32, 133–140. [CrossRef]
- Gavito, M.E., Olsson, P.A. Foraging for resources in arbuscular mycorrhizal fungi: What is an obligate symbiont searching for and how is it done? In Mycorrhiza, A. Varma (ed.), Springer-Verlag Berlin Heidelberg, 2008, pp.73–88. [CrossRef]
- Mohammadi Goltapeh, E., Rezaee Danesh, Y., Prasad, R., Varma, A. Mycorrhizal fungi: What we know and What should we know? In Mycorrhiza, A. Varma (ed.), Springer-Verlag Berlin Heidelberg, 2008, pp. 3–27. [CrossRef]
- Öpik, M., Saks, Ü., Kennedy, J., Daniell, T. Global diversity patterns of arbuscular mycorrhizal fungi–community composition and links with functionality. In Mycorrhiza, A. Varma (ed.), Springer-Verlag Berlin Heidelberg, 2008, pp.89–111. [CrossRef]
- Kytöviita, M-M., Vestberg, M. Soil legacy determines arbuscular mycorrhizal spore bank and plant performance in the low Arctic. Mycorrhiza, 2020, 30, 623–634. [CrossRef]
- Sun, X.F., Su, Y.Y., Zhang, Y., Wu, M.Y., Zhang, Z., Pei, K.Q., Sun, L.F., Wan, S.Q., Liang, Y. Diversity of arbuscular mycorrhizal fungal spore communities and its relations to plants under increased temperature and precipitation in a natural grassland. Chin Sci Bull, 2013, 58(33), 4109–4119. [CrossRef]
- Zhang, T., Yang, X., Guo, R., Guo, J. Response of AM fungi spore population to elevated temperature and nitrogen addition and their influence on the plant community composition and productivity. Sci Rep, 2015, 6, 24749. [CrossRef]
- Kilpeläinen, J., Aphalo, P.J., Lehto, T. Temperature affected the formation of arbuscular mycorrhizas and ectomycorrhizas in Populus angustifolia seedlings more than a mild drought. Soil Biol Biochem, 2020, 146, 107798. [CrossRef]
Figure 1.
Study area. Sampling sites: Villegas valley (A), Tigre River (B) and Cholila lake (C) (Map from [5]). In the zoomed-in map, the yellow line indicates the wildfire perimeter, green dots unburned sites, yellow dots moderately affected sites and red dots severely affected sites.
Figure 1.
Study area. Sampling sites: Villegas valley (A), Tigre River (B) and Cholila lake (C) (Map from [5]). In the zoomed-in map, the yellow line indicates the wildfire perimeter, green dots unburned sites, yellow dots moderately affected sites and red dots severely affected sites.
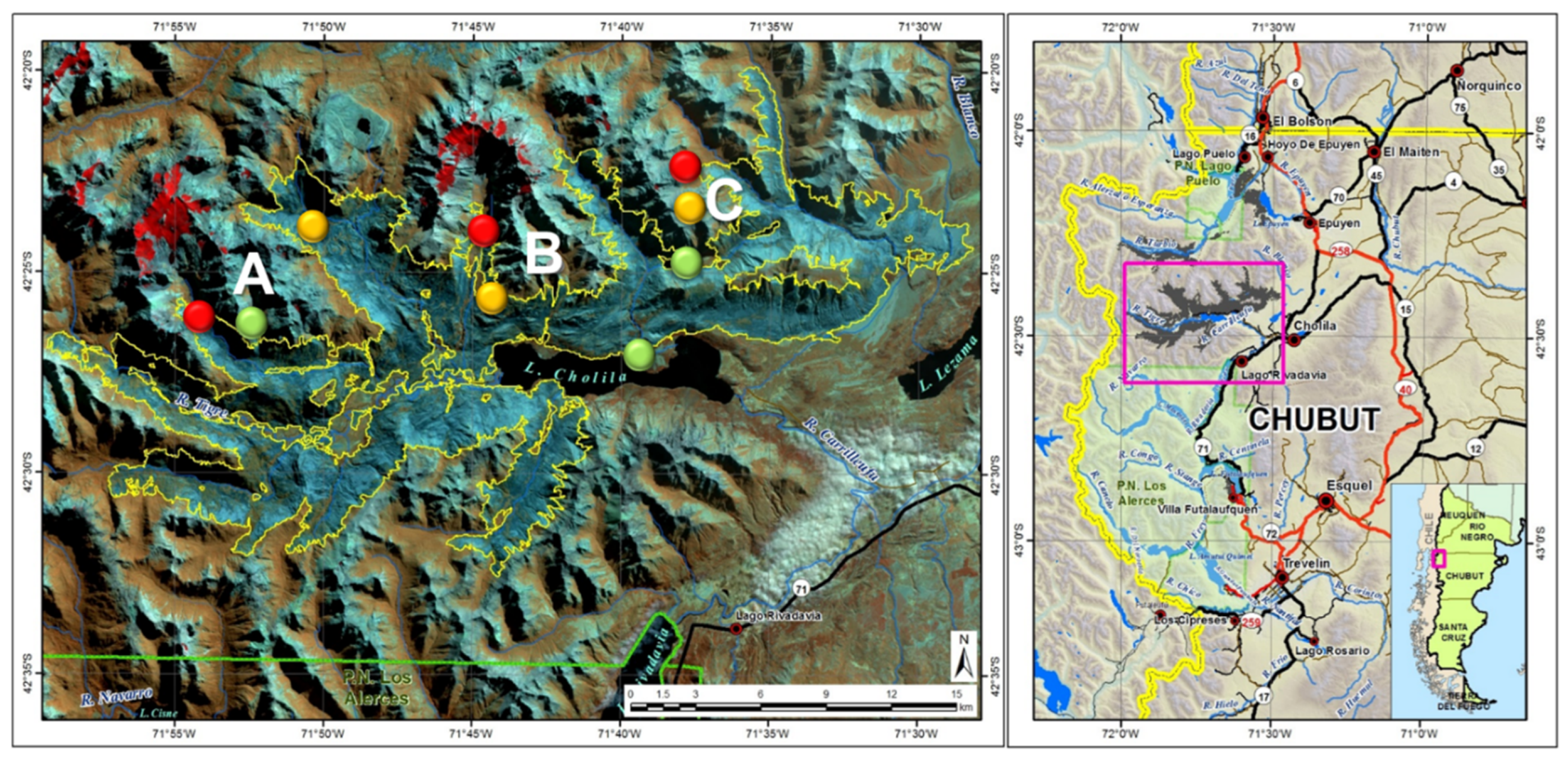
Figure 2.
AMS’ density for site (A, B, C), year (2016, 2021), and WFS treatment (unburned, moderate, high) (Spores/ 100 g of soil). Sites: A= Villegas valley, B= Tigre River and C= Cholila lake. Bars represent SE. Different letters indicate significant differences between treatments (P<0.05, 2W-GLMM).
Figure 2.
AMS’ density for site (A, B, C), year (2016, 2021), and WFS treatment (unburned, moderate, high) (Spores/ 100 g of soil). Sites: A= Villegas valley, B= Tigre River and C= Cholila lake. Bars represent SE. Different letters indicate significant differences between treatments (P<0.05, 2W-GLMM).
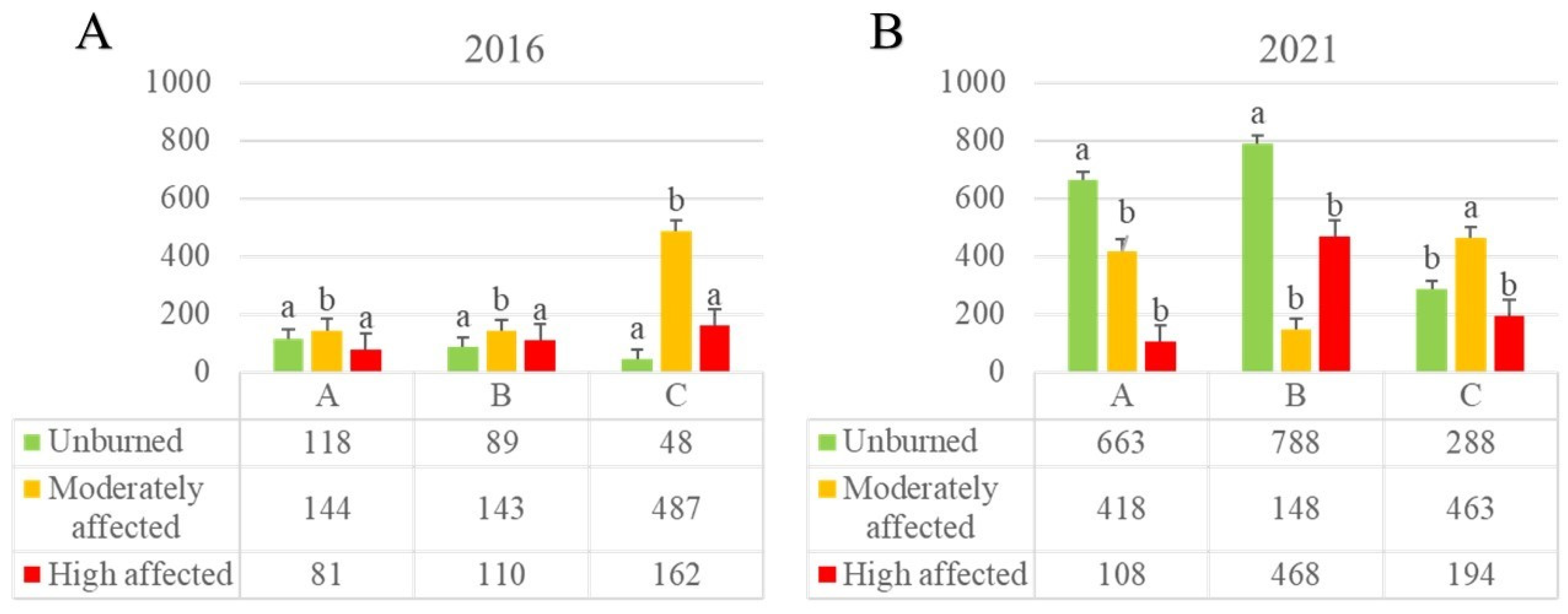
Figure 3.
Bioassay results. A) Heatmap showing AM structures per seedling per WFS treatment (unburned, moderate, high) and sites (A= Villegas Valley, B= Tigre River and C= Cholila lake). Different colors refer to different quantity of AM structures per seedling according to color bar (dark red=80 structures/seedling to Blue=0 structures/ seedling). B) Seedling AM colonization per treatment. Bars represent SE. Different letters indicate significant differences between treatments (P<0.05, 2W-GLMM).
Figure 3.
Bioassay results. A) Heatmap showing AM structures per seedling per WFS treatment (unburned, moderate, high) and sites (A= Villegas Valley, B= Tigre River and C= Cholila lake). Different colors refer to different quantity of AM structures per seedling according to color bar (dark red=80 structures/seedling to Blue=0 structures/ seedling). B) Seedling AM colonization per treatment. Bars represent SE. Different letters indicate significant differences between treatments (P<0.05, 2W-GLMM).
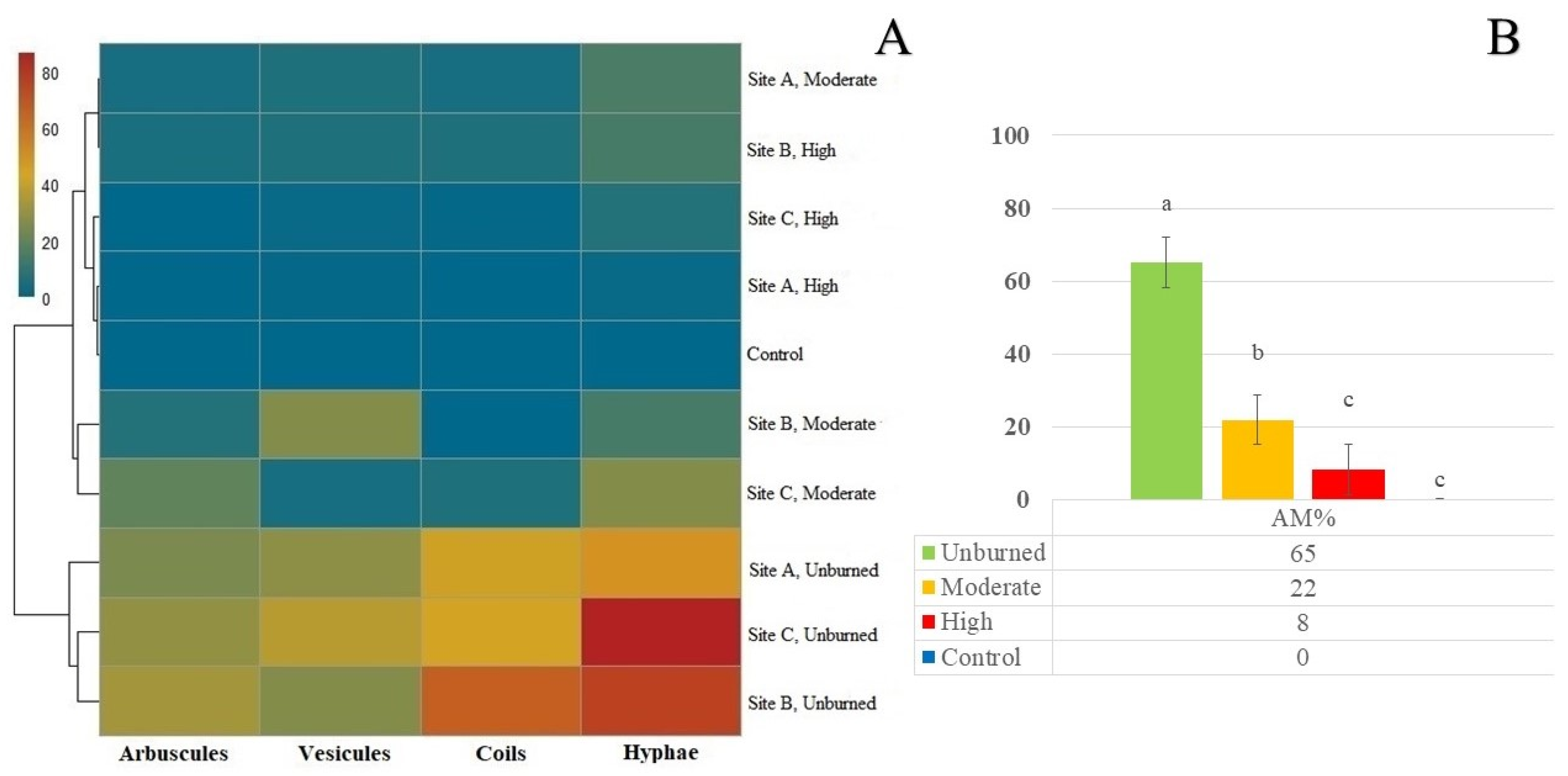
Figure 4.
PCA for AM structures and seedling colonization (AM%) and soil features considering sites and WFS (unburned, moderate, high). Sites: A= Villegas Valley, B= Tigre River and C= Cholila lake.
Figure 4.
PCA for AM structures and seedling colonization (AM%) and soil features considering sites and WFS (unburned, moderate, high). Sites: A= Villegas Valley, B= Tigre River and C= Cholila lake.
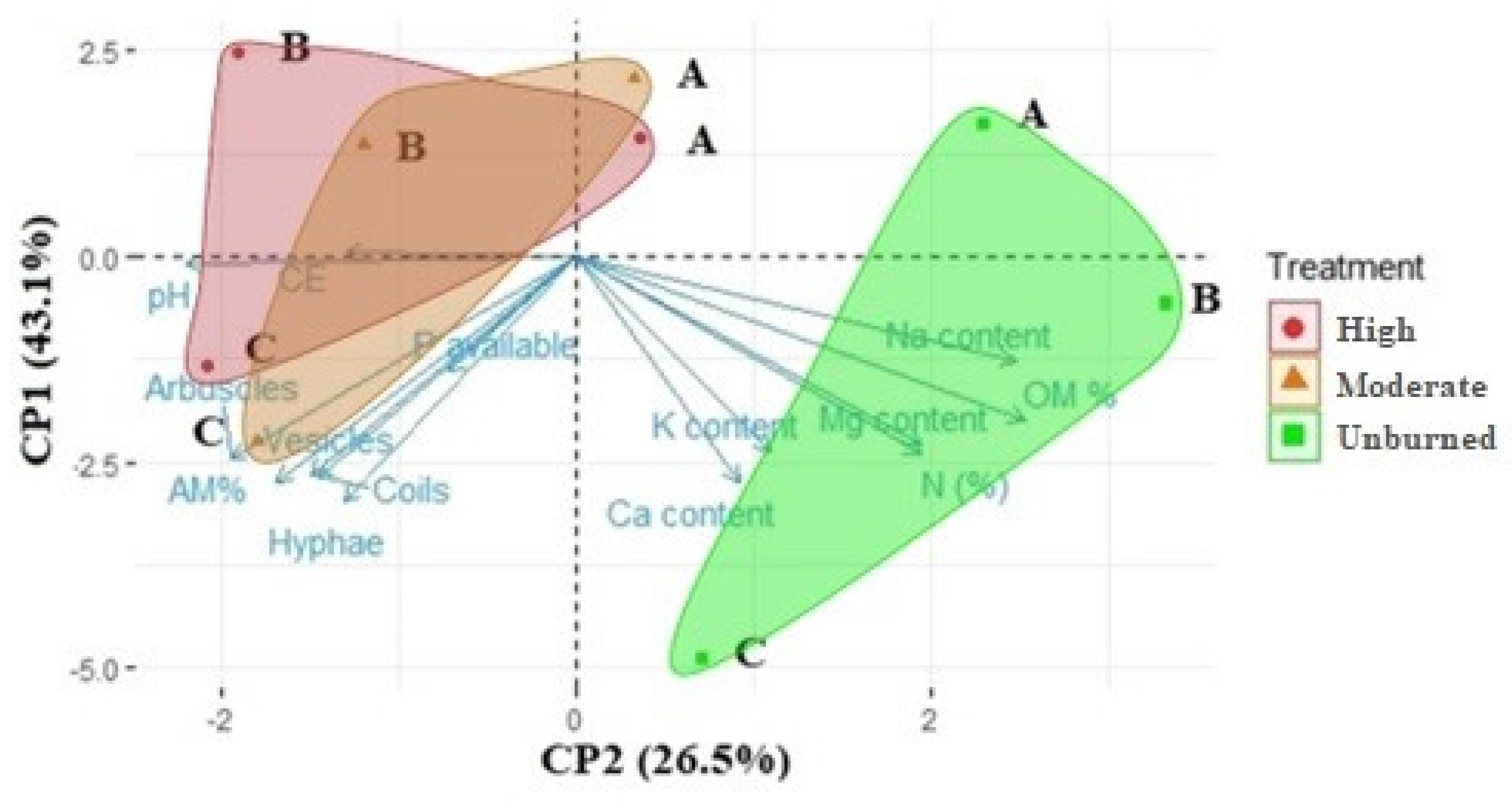

Table 1.
Physicochemical properties by sites and wildfire severity. Different letters indicate significant differences (p< 0.05). Capital letters between sites (*S) and lower-case letters between treatments or severity of fire (*T). Where interaction (*SxT) was detected, lower case letters show differences between treatment within each site [Villegas valley (A), Tigre River (B) and Cholila lake (C)]. Variables without letters showed no significant differences.
Table 1.
Physicochemical properties by sites and wildfire severity. Different letters indicate significant differences (p< 0.05). Capital letters between sites (*S) and lower-case letters between treatments or severity of fire (*T). Where interaction (*SxT) was detected, lower case letters show differences between treatment within each site [Villegas valley (A), Tigre River (B) and Cholila lake (C)]. Variables without letters showed no significant differences.
| Sampling sites | A | B | C | ||||||
| Fire Severity | Unb* | Mod | High | Unb | Mod | High | Unb | Mod | High |
| pH | 6.05 | 6.71 | 7.15 | 6.85 | 7.31 | 7.18 | 6.76 | 6.8 | 7.63 |
| *S*T | Aa | Ab | Ac | Ba | Bb | Bc | Ba | Bb | Bc |
| OM | 24.38 | 11.04 | 11.02 | 28.72 | 8.95 | 7.46 | 34.76 | 12.95 | 8.51 |
| *T | b | a | a | b | a | a | b | a | a |
| N | 0.72 | 0.41 | 0.46 | 0.66 | 0.39 | 0.29 | 0.91 | 0.64 | 0.39 |
| *S*T | Ab | ABa | ABa | Ab | Aa | Aa | Bb | Ba | Ba |
| P available | 40.66 | 21.65 | 55.23 | 24.01 | 47.29 | 30.45 | 54.03 | 44.48 | 39.68 |
| Na | 0.88 | 0.88 | 1.06 | 1.12 | 0.76 | 0.52 | 1.07 | 0.71 | 0.85 |
| *T | b | a | a | b | a | a | b | a | a |
| K | 0.96 | 1.37 | 0.94 | 1.71 | 0.81 | 0.55 | 1.69 | 1.21 | 1.68 |
| S*T* | b | a | a | ||||||
| Ca | 16.91 | 7.08 | 17.42 | 38.16 | 18.42 | 14.83 | 43.33 | 20 | 32.5 |
| *S*T | b | a | b | b | a | a | b | a | a |
| Mg | 4.08 | 2.08 | 5.58 | 9.33 | 1.83 | 1.33 | 7.5 | 5.67 | 4.5 |
| *S*T | b | a | a | ||||||
| EC | 0.13 | 0.13 | 0.21 | 0.08 | 0.09 | 0.13 | 0.12 | 0.13 | 0.22 |
| *T | a | ab | b | a | ab | b | a | ab | b |
| C/N | 16.7 | 13.1 | 11.8 | 24.4 | 11.5 | 12.5 | 19.2 | 9.9 | 10.8 |
| *T | b | a | a | b | a | a | b | a | a |
*Unb= Unburned, Mod= Moderate. OM=%, N= %, Available P= mg/kg, Na= meq/100g, K= meq/100g, Ca= meq/100g, Mg= meq/100g, EC= ds/m, C/N= %.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



