
Submitted:
20 June 2025
Posted:
09 July 2025
You are already at the latest version
Abstract
Medicinal plants are gaining attention as natural alternatives to chemical additives in poultry production. This study evaluated the antioxidant and anticoccidial activities of both aqueous (AQ) and hydromethanolic (HM) extracts of Mentha aquatica (ME), Rosmarinus officinalis (RO), and Thymus algeriensis (TH). In vitro assays were conducted against Eimeria oocysts, while in vivo trials involved 450 Cobb 500 broiler chicks randomly distributed into nine groups, including untreated and infected controls, more specifically, a group treated with Toltrazuril, and six groups treated with plant extracts at two concentrations (15 and 25 mL/L in drinking water). Administration began on day 7, and all animals except uninfected controls were inoculated orally with 10,000 sporulated Eimeria oocysts on day 10. Growth performance, feed efficiency, oocyst output, lesion scores, and histopathological changes were assessed until day 32. HM extracts showed higher antioxidant activity, particularly ME-HM. RO-HM, which demonstrated the strongest in vitro anticoccidial effect. The in vivo results revealed that TH-AQ and RO-AQ significantly improved body weight, feed conversion efficiency, and intestinal integrity. Anticoccidial indices (ACI) confirmed their efficacy, with values exceeding 180 for TH and RO, indicating excellent anticoccidial activity comparable to that of Toltrazuril. These findings support the potential of these phytogenic extracts as practical natural tools in poultry health management.
Keywords:
Eimeria sp.
; natural additives
; phytogenic compounds
; anticoccidial activity
; antioxidant capacity
; broiler chickens
; feed conversion
; growth performance
; anticoccidial index
1. Introduction
Coccidiosis is one of the most common and economically devastating diseases in poultry farming, particularly in broiler production systems [1,2,3,4]. It is caused by several species of protozoan parasites from the genus Eimeria, which invade the intestinal epithelium of chickens, leading to symptoms such as diarrhoea, dehydration, weight loss, poor feed conversion, and increased susceptibility to secondary infections [3,5,6,7]. According to Blake et al. (2020), the global economic cost of coccidiosis in chickens was estimated at approximately EUR 11.91 billion in 2016, considering both performance losses and the expenses of prevention and treatment [8]. This financial burden reflects the widespread occurrence of the disease and the high cost of its control [4,9,10].
The parasite responsible for coccidiosis belongs to the phylum Apicomplexa, family Eimeriidae. It completes its life cycle within 4 to 6 days in the host's intestinal epithelium, causing lesions that range in severity depending on the species and infection load [3,5,11]. These lesions can limit nutrient absorption, result in weight loss and poor feed efficiency, and may cause bloody diarrhoea or even death in severe cases [12,13,14]. Complicating eradication efforts is the formation of resilient oocysts, which are environmentally persistent and resistant to many disinfectants, making effective biosecurity and sanitation difficult [15,16]. In Algeria, coccidiosis remains one of the most frequently encountered parasitic diseases in poultry farms, with significant repercussions on flock health and production yields [17]. Several studies have identified Eimeria tenella, Eimeria acervulina, and Eimeria maxima as the predominant species affecting broiler flocks across various regions of the country [17,18,19,20,21,22]. The disease is exacerbated by factors such as poor hygiene practices, overcrowding, and inadequate vaccination protocols [18,20,23,24].
Moreover, surveys and field reports have highlighted increasing resistance to commonly used anticoccidial drugs, particularly monensin and diclazuril, which further complicates disease management [17,23]. This context underscores the pressing need to develop alternative, sustainable control strategies tailored to the specific epidemiological conditions prevalent in Algerian poultry production systems. Traditionally, the poultry industry has relied on various anticoccidial drugs, such as monensin and amprolium, which act by disrupting the parasite's metabolism or reproduction. However, the extensive and prolonged use of these drugs has led to the development of resistance, and diminishing their efficacy [25,26,27,28]. Resistance can arise through several mechanisms, including changes in membrane permeability, drug efflux, modifications to the target site, or the activation of alternative biochemical pathways [26,28,29,30,31]. Reports have highlighted total resistance to drugs like monensin and robenidine, and partial resistance to others such as salinomycin and toltrazuril [26,27,31]. More concerning is the potential for cross-resistance to emerge in other pathogens. For example, the use of coccidiostats has been linked to an increase in antimicrobial resistance in Campylobacter spp., raising significant public health concerns [32]. This situation underscores the need for integrated and sustainable approaches to manage coccidiosis and associated infections in poultry [33,34].
In response to these challenges, a wide array of alternative strategies has been explored, including acidifiers, probiotics, prebiotics, symbiotics, yeast-based products, and botanical additives [11,35]. Among these, botanicals and their secondary metabolites, such as essential oils, polyphenols, tannins, and alkaloids, have attracted attention for their antimicrobial, antiparasitic, antioxidant, and immunomodulatory properties [36,37,38].
Unlike conventional anticoccidial drugs, which often target specific stages of the parasite's life cycle, medicinal plant products act through a variety of mechanisms due to their diverse chemical constituents [39,40]. Their efficacy has been demonstrated under controlled experimental conditions, both through the administration of whole dried and ground plants and through the use of plant extracts [39]. They exhibit multiple biological effects, including antimicrobial, antioxidant, and anti-inflammatory properties, which are particularly relevant in the context of intestinal parasitic infections, such as coccidiosis [37,41,42]. By modulating oxidative stress, enhancing immune responses, and stabilizing gut function, medicinal plant products may offer a multifaceted approach to disease management [43,44,45,46].
In poultry, a growing body of evidence suggests that dietary inclusion of phytogenic supplements can lead to improvements in body weight gain, feed intake, and feed conversion ratio [47,48,49]. These benefits are believed to arise, not only, from direct effects on pathogenic organisms, but also, from improvements in intestinal barrier integrity and nutrient absorption [50,51,52]. Furthermore, medicinal plants have demonstrated the ability to modulate the gut microbiota, thereby reducing the incidence of dysbiosis and supporting a more resilient immune system [53,54,55]. They contribute to a reduction in oxidative stress and increase the antioxidant capacity in various tissues, including the intestinal epithelium, liver, and immune organs. These effects are especially valuable in the context of coccidiosis, where the host organism experiences high levels of oxidative damage due to parasite metabolism and the inflammatory immune response [38,54].
Plants from the Lamiaceae family, such as Mentha aquatica, Rosmarinus officinalis, and Thymus algeriensis are rich in diverse groups of bioactive compounds, including polyphenols (e.g., phenolic acids and flavonoids), terpenoids (such as monoterpenes and diterpenes), alkaloids, saponins, tannins, and essential oils [56]. These phytochemicals have been widely documented for their antioxidant, anti-inflammatory, antimicrobial, immunomodulatory, and antiparasitic properties [38,56,57,58]. Despite their widespread traditional use, the potential of these plants as anticoccidial agents remains underexplored in controlled experimental settings. In this context, our study was designed to evaluate the antiparasitic potential of Lamiaceae plant extracts against Eimeria species, the protozoan parasites responsible for avian coccidiosis. The investigation included both in vitro and in vivo approaches. In vitro assays were conducted on the sporulated oocysts of Eimeria to assess the extracts' disinfectant and antiparasitic efficacy. In vivo trials were then performed in broiler chickens to determine the impact of these extracts on parasite burden, gut integrity, and zootechnical performance indicators. By integrating both laboratory and animal-based assessments, this study aims to provide a scientific basis for the development of sustainable phytogenic alternatives to synthetic anticoccidial drugs in modern poultry production systems.
2. Materials and Methods
2.1. Plant Preparation
The aerial parts of Mentha aquatica (ME), Rosmarinus officinalis (RO) and Thymus algeriensis (TA) were collected in the district of Bordj Bou Arréridj, north-eastern Algeria. Specimens of ME and TA were harvested in the Oued Sayed area (Medjana), at 1050 m altitude, whereas RO was gathered in the Hammam El Biban area (Mansoura). The geographic coordinates of each site are listed in Table 1. After taxonomic verification, voucher specimens were deposited in the Herbarium of the VALCOR Laboratory, Faculty of Science, University M’Hamed Bougara, Boumerdes, Algeria.
For extraction, the air-dried aerial parts were ground into a fine powder (mean particle size lower than 910 µm), and 10 g of each sample was macerated at room temperature in 100 mL of pure water (aqueous extract, AQ) and a methanol: water (80:20, v/v) solution (hydromethanolic extract, HM) for 24 hours. The macerates were filtered, and the solvents were removed under reduced pressure with a rotary evaporator at controlled temperature. The concentrates were then lyophilised to give the crude extracts; the corresponding extraction yields are reported in Table 1.
2.2. Total Polyphenol Content
Total polyphenols were quantified with the Folin–Ciocalteu (FC) assay, following Singleton et al. (1999), and expressed as gallic-acid equivalents (GAE). Briefly, 20 µL of each plant extract (1000 ppm in methanol) was mixed with 100 µL of 10 % (v/v) Folin–Ciocalteu reagent [59]. After 5 min, 75 µL of 7.5 % (w/v) Na₂CO₃ were added. A reagent blank was prepared under identical conditions, substituting methanol for the extract. The mixtures were kept in the dark at room temperature for 2h, and absorbance was then read at 765 nm. All measurements were performed in triplicate, and results are reported as µg GAE/mg extract (mean ± SD).
2.3. Total Flavonoid Content
Flavonoid content was quantified by the aluminium-chloride colourimetric assay described by Türkoğlu et al. (2007). The method relies on the formation of a yellow Al³⁺–flavonoid complex, which shows a maximum absorbance at 415 nm. In each well of a 96-well microplate, 50 µL of plant extract, 130 µL of methanol, 10 µL of 1 M potassium acetate, and 10 µL of 10 % (w/v) AlCl₃·6H₂O were mixed [60]. A reagent blank was prepared by replacing the extract with methanol. After incubation for 40 min at room temperature in the dark, absorbance was measured at 415 nm. A quercetin calibration curve (0–200 µg/mL) was used for quantification, and results were expressed as milligrams of quercetin equivalents per milligram of extract (mg QE/mg).
2.4. Antioxidant Activity
2.4.1. DPPH Assay
The hydrogen- - or electron-donating capacity of the samples was assessed using the DPPH (2,2-diphenyl-1-picrylhydrazyl) assay, following the method described by Blois [61]. A 0.04% (w/v) DPPH solution was prepared in methanol (4 mg DPPH/100 mL), stored in the dark at 4 °C, and then brought to room temperature before use [61]. In a 96-well plate, 40 µL of each sample at the desired concentration was mixed with 160 µL of the DPPH solution (0.1 mM). A negative control consisted of 40 µL of methanol and 160 µL of DPPH. The plate was incubated for 30 min in the dark at room temperature, after which absorbance was read at 517 nm. Butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT) served as positive standards for antioxidants.
A Control is the initial concentration of the DPPH.
A Sample is the absorbance of the remaining concentration of DPPH in the presence of the extract and positive control.
Results are expressed as IC₅₀ values (µg/mL), the sample concentration required to scavenge 50% of DPPH radicals under the assay conditions.
2.4.2. CUPRAC Assay
The cupric-ion reducing capacity of each fraction was evaluated using the CUPRAC assay, as described by Apak et al. (2004). In a test tube, 1.0 mL of 10 mM CuCl₂·2H₂O, 1.0 mL of ammonium-acetate buffer (1 M, pH 7.0), and 1.0 mL of 7.5 mM neocuproine were mixed with 0.5 mL of plant extract or reference standard at the desired concentration [62]. The reaction volume was brought to 4.1 mL by adding 0.6 mL of distilled water. All samples were prepared in triplicate. The mixtures were incubated for 60 min at room temperature, after which absorbance was read at 450 nm against a reagent blank (buffer replacing the extract) [62]. A CUPRAC solution containing 50 µg/mL of standard antioxidant served as the assay control; BHT and BHA were used as positive references. An increase in absorbance reflected a greater reducing (antioxidant) capacity. Results are expressed as IC₅₀ values (µg/mL), i.e., the extract concentration required to produce an absorbance of 0.50 under the assay conditions.
2.5. Anticoccidial Activity
2.5.1. Parasite Source
Field sampling was carried out on deceased broiler chickens. Farms were selected in coordination with the Wilaya of Boumerdes' Agricultural Services Department (DSA); the selection criteria included prior veterinary reports indicating a suspected presence of coccidiosis. A total of ≈ 80 carcasses were collected. After necropsy, the entire intestinal tract was removed, and both external and internal surfaces were inspected. Intestinal contents were recovered by gentle scraping at multiple sites along the tract to ensure comprehensive parasite collection [63]. The resulting material was pooled in conical tubes, placed in insulated containers, and transported immediately to the VALCOR laboratory at the Faculty of Science, University M’hamed Bougara, Boumerdes. Samples were stored at 5–10 °C until further processing and preparation of parasite suspensions. For species identification by morphometry, 100 sporulated oocysts were measured in triplicate (n = 300) under a microscope equipped with a calibrated ocular micrometre [64,65]. Length, width, and the length/width ratio were recorded for each oocyst. These values were then compared to established morphometric references to determine species. The resulting species composition was as follows: Eimeria maxima (37.0%), Eimeria brunetti (19.4%), Eimeria necatrix (15.7%), Eimeria praecox (13.9%), Eimeria tenella (8.3%), and Eimeria mitis (5.6%).
2.5.2. In Vitro Anticoccidial Activity
The anticoccidial activity of the plant extracts against Eimeria spp. Oocysts were assessed using the procedure described by Remmal et al. (2011), with minor modifications. A sterile 96-well microplate was first filled with 140 µL per well of an agar-supplemented HBSS medium (Hank’s balanced salt solution lacking sodium bicarbonate and phenol red) [66]. The liquid phase was fortified with penicillin (100 IU/mL) and fluconazole (17 mg/mL) and then sterilised by filtration through a 0.2 µm membrane. Each plant extract was dissolved in phosphate-buffered saline (PBS) to give stock solutions ranging from 0.2 to 20 mg/mL. For every concentration tested, 40 µL of the extract was added to a well, followed by 20 µL of a parasite suspension containing 2 × 10⁵ sporulated oocysts per millilitre. Two commercial anticoccidials, Sulfaquinoxaline/trimethoprim (Vetacox) and Toltrazuril (Baycox ND), were included at the same concentrations, while untreated wells served as negative controls. Plates were left uncovered to ensure adequate aeration and incubated for 24 hours at 25–30°C under 60–80% relative humidity with gentle shaking. After incubation, oocysts in each well were enumerated with a Malassez hemacytometer, and both the percentage of viable oocysts and any wall deformations were recorded. The LC₅₀, the extract concentration required to eliminate or deform 50 % of the oocysts present in the negative control, was calculated for each treatment. All tests were performed in triplicate at each concentration. The lytic effect of the plant extracts and reference anticoccidials on Eimeria oocysts was further evaluated by monitoring the release of intracellular material at 273 nm [66]. Aliquots (1 mL) were prepared in sterile Eppendorf tubes containing a washed and filtered oocyst suspension, 700 µL of PBS (pH 7.4), and 200 µL of each plant extract at concentrations ranging from 0.2 to 20 mg/mL [66]. Tubes were incubated for 24h at 25°C. Sulfaquinoxaline/trimethoprim (SUS) and Toltrazuril (TOTRA) were included as positive controls at the same concentration levels. After incubation, samples were centrifuged at 3200 rpm for 5 min at 4°C. Supernatants (500 µL) were collected, and their absorbance was measured at 273 nm using a UV spectrophotometer [66]. Baseline correction was performed using PBS containing the corresponding extract concentration, but without the addition of oocysts. The resulting pellets were resuspended, and oocysts were counted in a Malassez hemacytometer to correlate absorbance changes with oocyst lysis. Data are presented as the change in absorbance at 273 nm and the corresponding oocyst viability versus extract concentration. Each assay was tested in triplicate.
2.5.3. In Vivo Evaluation of Plant-Extract Efficacy Against Coccidiosis
- Farm characteristics and animal husbandry
The study was conducted in a broiler facility located within the AVIGA complex (Industrial Poultry Accumulation and Fattening) in the Wilaya of Boumerdes. The farm is privately owned and specialises in broiler production. Although the selected house has a total surface area of 940 m², only a portion, stocked at roughly 10 birds m-², was used for the entire 42-day trial. A total of 450 Cobb 500 (Gallus gallus domesticus) chicks were placed on straw bedding for the first ten days; they were then transferred to 21 floor pens (100 × 120 cm, wire-mesh sides, no cover) containing wood shavings, at 10 birds per pen. Each pen was equipped with a feeder, a drinker and a butane-gas radiant heater, all of which had been disinfected with bleach. Two exhaust fans provided ventilation. Chicks were weighed on day 0 to establish a uniform average hatch weight and again on day 7, immediately before being randomly allocated to pens. All birds were vaccinated against Newcastle disease, infectious bronchitis and infectious bursal disease (Gumboro) and received an anti-stress supplement before and after each vaccination or transfer. Any chick that died during the first two weeks was weighed and replaced with one of similar weight. The feed was provided as mash, formulated with a multivitamin premix and without anticoccidial additives. Starter and grower diets were offered on days 0-21 and 22-42, respectively (Table 2), with both feed and water supplied ad libitum. A work schedule was established to record body weight at the end of each feeding phase.
- Safety dose assessment
Before the main experimental trial, a preliminary study (10 days) was conducted to assess the safety of the different plant extracts on broiler chickens (Not published). The animals used in this study were 10 days old (10 × 3 subjects per group), clinically healthy, and free of coccidial infection, as confirmed by parasitological examination of faecal samples. The aqueous extract stock solutions were prepared by suspending 100 g of dried, ground plant material in 1 L of distilled water and allowing the mixtures to stand at room temperature for 24 hours. From these stock solutions, four working concentrations: 5 mL, 15 mL, 50 mL, 100 mL, and 200 mL were diluted in 1 L of drinking water and freshly prepared each day. The hydro-methanolic extract was initially prepared, dried, and then reconstituted in water before being administered orally via gavage. However, at all tested concentrations, the hydro-methanolic extract induced an unacceptably high mortality rate during the first 24 hours (60%). Clinical signs of toxicity were observed, including lethargy, ruffled feathers, reduced feed and water intake, loss of balance, and severe prostration, indicating acute systemic effects. As a result, only the aqueous extract was retained for the in vivo study. It was administered through drinking water to ensure consistent intake and to minimise stress related to handling. Based on the safety outcomes, only the concentrations of 15 mL/L and 50 mL/L of the aqueous extract were selected for continuation in the experimental phase. All procedures were conducted in accordance with current European regulations and ethical standards for animal experimentation, including the EMA-CVMP guidelines for target animal safety [67,68,69,70].
- Experimental design
From day 7 onward, the 450 chicks were allocated to nine experimental groups based on the plant species and administered dose (Table 3). Each group consisted of 30 birds, housed in three replicate pens of 10 birds. Plant treatments also began on day 7. The aqueous and hydroethanolic extract stock solutions were prepared by suspending 100 g of dried, ground plant material in 1 L of distilled water and allowing the mixtures to stand at room temperature for 24 hours. Two working concentrations, 15 mL/L and 50 mL/L of extract preparation, were mixed in drinking water and prepared fresh each day. On day 10, birds in the six challenged groups received 10,000 sporulated oocysts of Eimeria spp. by oral gavage (1 mL per bird). Unchallenged groups were given 1 mL of sterile 0.9 % saline. The toltrazuril group (I) was treated when clinical signs of coccidiosis first appeared (25 mg toltrazuril/200 L drinking water for five consecutive days, equivalent to 7 mg/kg body weight). Groups G (uninfected, untreated) and H (infected, untreated) received no anticoccidial medication. To rule out pre-existing infections, all chicks underwent routine health checks, including coprological screening, during a pre-trial adaptation period.
- Studied parameters
- 1.
- Growth Performance Assessment
Throughout the trial, growth was monitored by weighing all birds in each pen at placement and then weekly, specifically on days 0, 10, 20, and 32 post-infection (PI). The difference between consecutive weighings provided body weight gain (BWG) for each interval. In contrast, feed intake for the same intervals was calculated by subtracting residual feed and the feed estimated for any birds that died from the total feed offered. Feed-conversion ratio (FCR) was expressed as feed intake divided by weight gain and adjusted for mortality. Daily checks were made to record mortality, and every deceased bird was weighed and examined to establish the cause of death. Together, body weight (BW), feed intake (FI), feed conversion ratio (FCR), and mortality provided an overview of growth performance and overall flock health [71,72,73]. To summarise these variables in single metrics of techno-economic efficiency, two composite indices were computed using the method of Marcu et al. [74] : the European Production Efficiency Factor (EPEF) and the European Broiler Index (EBI). Both indices combine growth rate, feed efficiency and livability, offering an integrated appraisal of each treatment’s overall performance across the 32-day experimental period.
- 2.
- Survival and Weight-Gain Indices
A comprehensive evaluation was performed on day 32 PI. Clinical symptoms and mortality were assessed and recorded each day after infection. Survival rate for each treatment was calculated from the proportion of birds alive relative to the initial number placed. The weight-gain rate for each bird corresponded to its net weight increase between the time of infection and day 32. Relative weight-gain (RWG) rate expressed this increase as a percentage of the animal’s pre-infection weight. These indices allowed direct comparison of post-infection growth and viability among the various experimental groups.
- 3.
- Oocyst Enumeration
The parasitic load was tracked by counting oocysts at several time points. Faecal samples (500g) were collected from each pen on days 10, 20, and 32 post-inoculation (PI) (n = 6 replicates), homogenised, and examined microscopically using a Malassez hemacytometer to determine oocysts per gram of faeces (OPG). Thereafter, the OPG was converted into the oocyst value of oocysts (OV% %). The reduction rate of oocyst production and oocyst value was calculated as indicated by Chauhan et al. (2017) [75]The results were expressed as OPG output [76].
- 4.
- Lesion Scoring
At day 10 post-infection (PI), a standardised sampling protocol was carried out on fifteen broiler chickens (five birds per treatment group) [75,77,78]. Before sampling, these chickens were fasted for 24 hours to clear their gastrointestinal tract, while water was provided ad libitum to maintain hydration. On day 10 PI, the birds were humanely slaughtered by approved animal welfare guidelines. Slaughter was performed by exsanguination via the jugular vein, and immediately afterwards, each carcass was de-feathered and eviscerated. The entire intestinal tract was then carefully removed and collected for gross examination of lesions. Gross intestinal lesions were scored using the established Johnson & Reid scoring system, which assigns a numerical severity grade from 0 to 4. A score of 0 indicates no visible lesions, whereas a score of 4 reflects the most severe intestinal damage, characterised by extensive haemorrhage and large caseous cores. Intermediate scores (1, 2, and 3) denote progressively increasing levels of lesion severity, as defined by the standard criteria [79]. This grading method provides a consistent and objective assessment of lesion severity across all treatment groups, allowing for reliable comparisons of disease impact between different experimental conditions.
- 5.
- Anticoccidial Index (ACI)
The ACI was calculated for each treatment group as an integrated measure of anticoccidial efficacy, incorporating four parameters: survival rate (SR), relative weight-gain rate (RWG), oocyst value (OV), and lesion score (LS) [80]. This index provides a comprehensive assessment of how effectively the treatment controls coccidiosis. Interpretation of ACI values followed standard thresholds: ACI < 120 indicates no anticoccidial effect; values of 120–159 reflect low efficacy; 160–179 denotes moderate efficacy; and ACI ≥ 180 is considered indicative of excellent anticoccidial efficacy.
- 6.
- Histopathological Analysis
Intestinal tissue sampled at day 10 PI was fixed in neutral-buffered 10 % formalin, processed routinely, embedded in paraffin, sectioned at 5 µm and stained with haematoxylin–eosin [81]. Microscopic examination focused on mucosal integrity, inflammatory infiltrates, haemorrhage and the presence of parasitic stages within epithelial cells. Histopathology thus complemented gross lesion scoring by revealing cellular-level alterations induced by infection and by any protective effects of the administered treatments.
2.6. Statistical Analysis
Statistical analyses were performed on data obtained from triplicate measurements for each assay. GraphPad Prism (version 8) was used to conduct a one-way ANOVA (α = 0.05). Post-hoc comparisons among treatments were carried out with Tukey’s and Dunnett’s multiple-comparison tests at the 95 % confidence level. Correlation analyses employed Pearson correlation coefficients (r), simple linear regression, and analysis of covariance (ANCOVA).
3. Results and Discussion
3.1. Polyphenols and Flavonoid Contents
The total phenolic content (TPC) and total flavonoid content (TFC) of Mentha aquatica (ME), Rosmarinus officinalis (RO), and Thymus algeriensis (TH) extracts are summarised in Table 4. Overall, the hydromethanolic (HM) extracts yielded higher total phenolic content (TPC) than the aqueous (AQ) extracts for all three species (Figure 1). For instance, TH showed a TPC of 220.35 ± 5.92 mg GAE/g DW in the HM extract, compared to 128.11 ± 10.56 mg GAE/g in the AQ extract, representing a nearly 1.7-fold increase. RO similarly exhibited a higher TPC in HM (215.50 ± 17.19 mg GAE/g) compared to water extract (160.96 ± 6.55 mg/g). MA had the highest TPC among aqueous extracts (167.13 ± 26.48 mg/g), which further rose to 199.26 ± 12.26 mg/g in the HM extract. Statistical analysis confirmed that TPC was significantly higher (p < 0.05) in HM extracts compared to AQ extracts for each plant (Figure 1). Thus, adding 80% methanol markedly enhanced polyphenol extraction efficiency, consistent with the general notion that polar organic solvents recover more phenolics than water alone; these results are in agreement with previous works [82,83]. Among the species, TH and RO-HM extracts exhibited the highest phenolic levels (~215–220 mg GAE/g), while ME was only slightly lower (~199 mg GAE/g) (Figure 1). In the aqueous preparations, ME retained the highest total phenolic content (TPC), followed by RO and TH. These results indicate that both species identity and solvent type influenced the polyphenol yield [84,85].
The trends in flavonoid content (TFC) were more variable among the extracts. ME stood out with the highest flavonoid levels in its HM extract (TFC = 59.77 ± 4.07 mg QE/g DW), which was roughly double that of its water extract (29.56 ± 2.58 mg/g) (Figure 1). TH also showed a substantially higher TFC in the HM extract (28.05 ± 1.31 mg/g) compared to the AQ extract (15.20 ± 2.30 mg/g) (Figure 1). In contrast, RO displayed the opposite pattern: its AQ extract had a notably greater TFC (49.26 ± 0.40 mg/g) than the HM extract (33.26 ± 1.40 mg/g). This anomaly suggests that certain flavonoids in rosemary may be more efficiently extracted by water (perhaps due to highly water-soluble glycosides) or that some compounds in the methanol-water mixture could interfere with flavonoid yield [86]. Despite this species-specific divergence, the differences in TFC between extract types were statistically significant (p < 0.05) for each plant (Figure 1). Overall, ME had the richest flavonoid content in the HM form, whereas RO led in flavonoid content for the aqueous extracts. These data suggest that the choice of extraction solvent can differentially affect flavonoid recovery, depending on the plant's phytochemistry. Hydromethanol (80% MeOH) tends to extract more flavonoids in ME and TH, whereas pure water is surprisingly effective for RO flavonoids. Such variations could be linked to the distinct profiles of flavonoid compounds in each species (e.g. rosemary may contain more polar flavonoid glycosides, while mint and thyme have flavonoids that are better solubilised by aqueous methanol [87,88,89].
Across the three Lamiaceae herbs, comparative work keeps converging on two consistent messages: (i) extraction and post-harvest handling can swing phenolic and flavonoid yields by an order of magnitude, and (ii) even modest quantitative differences matter, because antioxidant capacity tracks very closely with the totals. Mentha aquatica illustrates the first point vividly. In Safaiee et al.'s (2019) study, which systematically varied drying strategies and ultrasound conditions, freeze-drying followed by a short 5-minute sonication at 60 °C delivered the highest recovery. However, this peak still reached only 0.245 mg gallic acid equivalents (GAE) per gram of dry leaf, equivalent to approximately 245 µg GAE/g. Microwave- and oven-dried samples came in at lower levels, indicating that water-mint phenolics are delicate once moisture is removed. Sonication time itself, interestingly, had almost no statistical weight on the final yield [90]. Our yields eclipse or at least equal the best figures reported so far. In Benabdallah et al. (2016), six wild Mentha species were recorded at 43.21 ± 1.09 mg GAE/g [91], while Fidan et al. (2023) confirmed a comparable value of 43.36 mg GAE/g; both are roughly one-quarter of our hydromethanolic value [92].
Rosmarinus officinalis behaves quite differently. Afonso et al (2013) prepared a gentle aqueous infusion (INF) and a non-esterified phenolic fraction (NEPF) from dried rosemary leaves. The infusion retained a respectable 16.67 ± 0.40 mg GAE/g, while the more selective NEPF held 8.59 ± 0.31 mg GAE/g. Although that study did not report a separate total-flavonoid figure, its chromatographic profile confirmed rosmarinic acid and carnosic acid as dominant compounds, and the authors linked the higher phenolic load of the INF directly to stronger in vivo antioxidant endpoints (lower TBARS and boosted catalase/GPx activities) [93]. Our results, therefore, sit an order of magnitude above the infusion and about double the strongest alcoholic benchmarks, reflecting both our late-summer harvest and the higher affinity of methanol for diterpenic acids such as rosmarinic and carnosic .
Thymus algeriensis has long been considered modest in its soluble polyphenol content. A recent work optimised ultrasound-assisted extraction with a Box–Behnken design. Under the best conditions (60 °C, 36.7 min, 32.8 mL solvent), flowering-stage material yielded 2.961 g GAE 100/g (≈ 29.6 mg GAE/g) and 0.887 g quercetin equivalents (QE) 100/g (≈ 8.9 mg QE/g). Those totals, coupled with IC₅₀ values of 0.016 mg/mL for DPPH and 0.426 mg/mL for FRAP assays, underscore how ultrasound can coax out a chemically potent but extraction-sensitive phenolic pool in this North-African thyme [94]. The leap very likely comes from the exceptionally arid provenance of our stand (a known trigger for caffeic-acid biosynthesis) combined with a 24-h maceration that penetrates both leaf and woody bract tissues. Ziani et al (2019) data on Thymus algeriensis fit the same pattern, topping out near 34 mg GAE/g again, well below our 220 mg plateau [95].
These comparisons show that extraction chemistry and ecological stress can outweigh species identity when the aim is to maximise antioxidant density. Where the literature typically quotes double-digit milligram values, our hydromethanolic protocol routinely climbs past 200 mg GAE/g. That potency, however, comes with trade-offs: more bitter diterpenes in rosemary, a darker hue in mint, and resinous notes in thyme that formulators will have to balance against shelf-life or health-claim targets.
3.2. Antioxidant Activity
Oxidative stress is known to exacerbate intestinal damage and immune dysfunction in poultry affected by coccidiosis, a major parasitic disease that compromises gut integrity, growth performance, and overall welfare in broiler chickens [96,97,98]. Given the increasing restrictions on synthetic anticoccidials and growing consumer demand for natural alternatives, plant-based extracts with antioxidant and anti-inflammatory properties are gaining interest as supportive agents in poultry health management. In this context, evaluating the antioxidant potential of selected medicinal plants can provide valuable insights into their capacity to mitigate oxidative damage associated with Eimeria infections.
To this end, we assessed the radical-scavenging activity of aqueous (AQ) and hydromethanolic (HM) extracts prepared from three aromatic species. Both extraction methods reflect realistic applications, infusions mimic traditional use, and methanol–water mixtures enhance bioactive recovery. Two well-established in vitro assays were used to quantify the antioxidant activity of the extracts, providing a dual perspective on their ability to quench free radicals and reduce oxidative burden in broiler models exposed to coccidial stress.
Table 5 compares the radical-scavenging power of aqueous (AQ) and hydromethanolic (HM) extracts prepared from Mentha aquatica (MA), Thymus algeriensis (TH) and Rosmarinus officinalis (RO). Activity was quantified with the well-established DPPH assay, which measures the capacity of an antioxidant to reduce the purple DPPH• radical, and with the CUPRAC test, which monitors the reduction of the Cu(II)–neocuproine complex at physiological pH. In both methods, a lower IC₅₀ denotes a more potent antioxidant, i.e. less extract is required to quench 50 % of the radicals [99,100].
Hydromethanolic extraction enhanced antioxidant potency. The ME-HM extract was the most powerful of all plant samples (IC50ᴰᴾᴾᴴ = 10.19 ± 0.51; IC50CUPRAC = 9.47 ± 0.19 µg/mL), outperforming every other extract and even surpassing the synthetic antioxidant BHT in the DPPH test, while approaching BHT in CUPRAC. Only BHA remained markedly stronger (IC50ᴰᴾᴾᴴ = 5.73 ± 0.41; IC50CUPRAC = 3.64 ± 0.19 µg/mL). The second-best sample was the ME-AQ extract (12.79 ± 0.05; 12.06 ± 0.60 µg/mL), followed by TH-HM (15.56 ± 0.63; 11.72 ± 0.85 µg/mL) and RO-HM (15.74 ± 0.69; 14.50 ± 0.14 µg/mL). In every species, the HM fraction showed lower IC50s than its aqueous counterpart, confirming that a 50 % MeOH/H₂O solvent retrieves a broader spectrum of redox-active metabolites.
The concordant ranking delivered by DPPH and CUPRAC suggests that the extracts contain both hydrogen-donating and electron-donating antioxidants, mostly polyphenols. Mixed methanol–water systems are known to extract these compounds more efficiently than water alone, because methanol disrupts cell walls and solubilises moderately polar phenolics while still keeping highly polar compounds in solution [101]. Therefore, the superior activity of ME-HM likely reflects a richer load of flavonoids, rosmarinic acid derivatives and other phenolics contributing additionally to radical scavenging [102,103].
The antioxidant ranking obtained in our study (ME-HM > ME-AQ > TH-HM ≈ RO-HM > RO-AQ > TH-AQ) is broadly consistent with the recent literature on the same Lamiaceae species, while highlighting how solvent choice, chemotype and processing can modulate absolute IC50 values. For Mentha aquatica, Pereira et al. (2019) reported a DPPH EC₅₀ of 8.1 ± 1.3 µg/mL for an 80 % hydroethanolic extract, very close to the 10.2 µg mL⁻¹ we measured for the hydromethanolic fraction and likewise identified the mint as the most potent antioxidant among the plants they compared [104]. Another study comparing an infusion (INF) with a 70 % hydroethanolic extract (HE) showed the DPPH IC₅₀ of the HE to be approximately six-fold lower than the INF, mirroring the solvent-driven gain we observed for the HM fraction over the aqueous one [85]. A more recent study encapsulating a pure methanolic extract of M. aquatica in alginate hydrogels showed that the entrapped extract retained low-micromolar DPPH IC₅₀ values and superior oxidative stability, confirming that the strong radical-scavenging power of this species is preserved across formulation types [105]. Elsewhere, a drying-method investigation (microwave, oven, shade) demonstrated that essential oil–rich fractions lose up to fourfold antioxidant capacity compared with the corresponding ethanolic extracts, reinforcing the solvent-polarity effect we observed (HM < AQ).
A Tunisian survey of Thymus algeriensis wild populations found that methanolic leaf extracts scavenged DPPH radicals with an IC₅₀ of ≈ approximately 7 µg/mL, outperforming BHT (11.5 µg/mL) and confirming the strong radical-quenching capacity of this species when extracted with polar organic solvents [106]. An investigation of Algerian populations reported that a phenolic-rich hydromethanolic fraction (50% MeOH) scavenged DPPH radicals with an IC₅₀ of ≈ approximately 7 µg/mL and markedly enhanced plasma antioxidant status in vivo [107]. Our hydromethanolic extract (IC₅₀ = 15.6 µg/mL) retains the same activity range. However, the slightly higher value is expected because we analysed the whole aerial parts instead of the phenolic-rich leaves alone and used a milder 50% MeOH/H₂O system. By contrast, the hydro-distilled essential oil (rich in terpenoids but almost devoid of polyphenols) displayed very weak radical-scavenging power (DPPH IC₅₀ = 83.8 mg/mL), roughly four orders of magnitude higher than our HM extract (15.6 µg/mL) [108]. The result highlights how removing the phenolic fraction in favour of volatile monoterpenes can drastically reduce antioxidant efficacy.
For Rosmarinus officinalis, a multi-approach study on Algerian rosemary reported IC₅₀ values of 6.82 µg/mL (DPPH) and 1.59 µg/mL (CUPRAC) for an 80% methanolic extract, which is substantially stronger than our HM fraction (15.7 and 14.5 µg/mL, respectively). The gap highlights the impact of chemotype (Mediterranean vs. local ecotype) and intensification techniques (sonication, higher MeOH content) on polyphenol yield; nevertheless, both studies concur in ranking rosemary as a top natural antioxidant under CUPRAC conditions [109]. Sahunie (2024) showed that switching from 80 % methanol to ethyl-acetate (low-polarity) extraction more than doubled the IC₅₀ (DPPH and CUPRAC), confirming that semi-polar solvents recover fewer redox-active diterpenes and phenolic acids than hydro(methanol) systems [110]. Conversely, the study by Doozakhdarreh et al. (2022) on rosemary essential oils harvested and dried under different regimes reported IC₅₀ values as low as 6–10 µg/mL for shade-dried material, comparable to our HM extract, and showing that an optimised terpene profile can rival polyphenol-rich solvent extracts [111]. A recent study using super-critical CO₂, coupled with natural deep-eutectic solvents (NADES), demonstrated that cleaner, terpene-enriched fractions still display measurable radical-quenching activity. However, they remain less potent than hydro-methanolic counterparts because key phenolic acids (rosmarinic, carnosic) partition poorly into CO₂ [112]. Martín-Sampedro et al. (2016) demonstrated that super-critical CO₂ antisolvent fractionation can further concentrate carnosic and rosmarinic acids; however, DES or hydro-methanol still yield the lowest IC₅₀s overall [113]. According to Bejenaru et al. (2024), ethyl-acetate, ethanolic and aqueous extracts gave DPPH IC₅₀ values of 272, 387 and 534 µg/mL, respectively, far weaker than our HM fraction (15.7 µg/mL) and illustrating the limited solubility of carnosic acid/carnosol in those solvents [114]. Conversely, an agrifood study found that the essential oil scavenged DPPH with an IC₅₀ of 15.0 µg/mL, which essentially matches our HM value and underscores that diterpenes in the oil can, when sufficiently concentrated, rival polyphenol-rich solvent extracts [115].
Taken together, these external datasets reinforce the qualitative hierarchy observed here and demonstrate that hydromethanolic extraction systematically boosts the recovery of redox-active phenolics, thereby lowering IC₅₀ values. Variations across studies primarily reflect geographic chemotypes, plant parts, and extraction intensity; yet, the core message remains consistent: Mentha aquatica and Thymus algeriensis possess exceptional electron- and hydrogen-donating capacities, while rosemary extracts remain highly effective, mainly when richer methanolic systems or process intensification are employed. The MeOH/H₂O system we used thus remains a robust, food-compatible strategy for maximising antioxidant efficacy in Mentha aquatica, Thymus algeriensis and Rosmarinus officinalis.
3.3. In Vitro Anticoccidial Activity
Maintaining effective biosecurity in broiler production is essential to controlling coccidiosis, a parasitic disease that remains one of the most economically damaging and welfare-compromising conditions in poultry farming. The infective oocysts of Eimeria are incredibly resilient in the environment, resisting standard cleaning and surviving for extended periods in poultry housing, where they can rapidly reinfect flocks. Traditional chemical disinfectants often fail to destroy the fully sporulated, infective form of the parasite, and their repeated use raises concerns about environmental and health risks.
In light of this, we explored sustainable, plant-based alternatives that can target Eimeria oocysts outside the host. The aim was to evaluate the direct oocysticidal activity of selected plant extracts as potential natural disinfectants. To this end, fully sporulated oocysts, the most resistant developmental stage, were incubated for 24 hours with aqueous and hydromethanolic (HM) extracts of Rosmarinus officinalis (RO), Mentha aquatica (ME), and Thymus algeriensis (TH). According to Table 6, the most effective treatment was RO-HM, with an LC₅₀ of 233.14 ± 3.56 µg/mL, followed by ME-HM (967.11 ± 2.58 µg/mL) and RO-AQ (699.11 ± 4.61 µg/mL). The aqueous extract ME-AQ also showed relevant activity (329.86 ± 2.78 µg/mL). In comparison, both extracts of Thymus algeriensis were markedly less active, TH-HM (2621.62 ± 2.90 µg/mL) and TH-AQ (3449.56 ± 2.53 µg/mL), indicating limited oocysticidal potential.
Conventional anticoccidial drugs, sulfaquinoxaline/trimethoprim and toltrazuril, exhibited no measurable activity under these experimental conditions, underscoring the importance of developing effective and environmentally friendly alternatives (Table 6). They are engineered to act inside the bird, where Eimeria schizonts or gamonts are actively synthesising folate or generating ATP; they therefore leave entirely sporulated oocysts, metabolically quiescent and shielded by a thick, dityrosine-cross-linked wall, completely untouched [116,117]. The impermeability and biochemical inertia of that wall mean that even high drug concentrations cannot penetrate or find a vulnerable target [117,118]. As a result, the environmental reservoir of infective oocysts persists despite in-host medication, underscoring the need for complementary strategies that break the cycle outside the bird, rigorous litter management, and the use of alkaline or cresolic disinfectants, as well as novel agents that block sporulation altogether [116,119]. In this context, plant-derived phenolics, such as thymol and carvacrol, have demonstrated the ability to deform or even lyse sporulated oocysts and inhibit new sporulation in vitro, suggesting phytogenic additives may be promising adjuncts to conventional treatments [120,121,122].
Marked differences in oocysticidal efficacy were observed among the tested extracts. The hydromethanolic extracts of RO-HM and ME-HM exhibited the most vigorous activity, reducing oocyst viability by more than 65% compared to the untreated control (Figure 2). These two extracts demonstrated comparable potency, suggesting similar effectiveness in disrupting oocyst integrity. In contrast, the hydromethanolic extract of TH-HM was significantly less effective, with viability reductions approximately 25% lower than those achieved by ME-HM, indicating limited antiparasitic activity (Figure 2). Among aqueous extracts, ME-AQ showed the highest efficacy, significantly decreasing oocyst viability and surpassing the effects observed with both RO-AQ and TH-AQ. TH-AQ, in particular, exhibited the weakest activity, with oocyst viability remaining relatively high even at the highest concentrations tested (Figure 2).
Our findings accord with earlier in vitro studies. Remmal et al. (2011) demonstrated that four essential oils (thyme, artemisia, tea tree, and clove) effectively destroyed sporulated Eimeria oocysts within hours, accompanied by the release of 273-nm-absorbing compounds [66]. Jitviriyanon et al. later reported Thai essential oils of Ocimum basilicum and Boesenbergia pandurata with IC₅₀ values of 0.101 and 0.134 mg/mL, respectively, against E. tenella sporulation [123]. More recently, Gadelhaq et al. [124] found that aqueous garlic and moringa extracts blocked chicken oocyst sporulation almost completely (up to 100% at 70% v/v), whereas plain hypochlorite achieved only ~50% inhibition under identical conditions [124]. Maodaa et al. [125] observed that a methanolic extract of Thymus daenensis inhibited E. papillata sporulation by >90 % at 300 mg/mL [125]. Orange-peel essential oil, an agro-industrial co-product, damaged both sporulated and unsporulated E. tenella oocysts, causing up to 64 % structural loss at 30 % v/v [126]. Pomegranate peel, another processing by-product, blocked oocyst sporulation and reduced shedding in vivo [127]. Compared with conventional disinfectants, these extracts perform favourably. Lee et al. determined that ≥ 7.5% hydrogen peroxide solutions had to act for three hours to inactivate Eimeria acervulina oocysts, fully and H₂O₂ gas plasma was required for rapid eradication [128]. Against that benchmark, the LC₅₀s of RO-HM (0.23 mg/mL) and ME-AQ (0.33 mg/mL) place the botanical extracts among the most potent agents, rivalling reference oxidants while being less corrosive and safer for operators.
RO-HM’s superiority stems from its high content of phenolic diterpenes (carnosic acid, carnosol) and rosmarinic acid, which can permeabilise parasite membranes and inhibit calcium-dependent kinases essential to the oocyst life-cycle [66,128]. The intense activity of ME-AQ highlights the role of highly hydrophilic polyphenols (rosmarinic acid glycosides, flavonoids) that, like essential oil components, destabilise the oocyst’s double chitinous wall. Thymus extract is weaker, reflecting its lower thymol/carvacrol content; studies on thyme and oregano oils show that boosting these monoterpenes can increase oocysticidal efficacy by an order of magnitude [129].
Mortality was paralleled by a rise in supernatant absorbance at 273 nm, indicating leakage of nucleotides and aromatic amino acids and confirming a membrane-disruptive mode of action (Figure 3). The lysis of the Eimeria population was described by a linear regression equation (Table 7) and revealed uniformly negative slopes (p ≤ 0.003), confirming a dose-dependent coccidiocidal action for all samples tested. Among aqueous extracts, the potency hierarchy was RO > ME > TH, with rosemary displaying the steepest decline (slope = −0.024 ± 0.005), whereas the hydromethanolic series favoured Mentha aquatica (−0.018 ± 0.002). Goodness-of-fit values (R² = 0.77–0.90) indicated that linear models accounted for most of the variance, although ME-AQ showed hints of non-linearity at sub-inhibitory doses (R² = 0.70) (Figure 3, Table 7). The divergence between chemical richness (highest TPC in HM extracts) and biological potency (often higher in AQ extracts) suggests that rapidly diffusing polar metabolites, rather than bulk phenolics, dominate early oocyst inactivation. Because each concentration was assayed once, these slopes should be viewed as provisional potency indices pending full biological replication.
Linear-regression analysis leaves no doubt that oocyst viability decreases as extract concentration increases (Figure 3, Table 7): every slope is negative, and, according to the F-tests (p ≤ 0.003), each differs significantly from zero. The magnitude of the slope is a direct measure of potency on a mass basis. The aqueous rosemary extract (RO-AQ) shows the steepest slope in absolute terms (−0.024 % mL/mg) and is therefore the most aggressive, followed by the aqueous mint extract (ME-AQ) and, lastly, the aqueous thyme extract (TH-AQ) (Figure 3).
In the hydromethanolic series, the pattern is reversed: mint (ME-HM) is now the most active, while thyme (TH-HM) and rosemary (RO-HM) bring up the rear with slopes of roughly −0.012 % mL/mg. This ranking does not accurately reflect the total phenolic content, which is typically higher in the hydromethanolic fractions; instead, it suggests that rapid oocysticidal activity is driven by highly polar metabolites that diffuse readily through the oocyst wall. Model quality supports this view: coefficients of determination range from 0.77 to 0.90 (with the sole exception of ME-AQ, R² = 0.70) (Figure 3, Table 7), meaning that a straight line explains between 77% and 90% of the observed variance, while the standard error of the regression (Sy. x) remains below 20%. The y-intercept, which varies between 44% and 94% depending on the batch, remains close to the expected spontaneous mortality and does not compromise slope comparisons.
From an application standpoint, aqueous rosemary stands out as the leading candidate because it combines rapid action, technological simplicity and regulatory acceptability. The other aqueous extracts also display appreciable potential, whereas the hydromethanolic fractions, despite their richer chemistry, are less effective on a per-unit-mass basis. Taken together, the results call for biological replication, targeted identification of the active water-soluble small molecules, and optimisation of an aqueous formulation suited to veterinary or agri-food use.
3.4. In Vivo Experimentation
3.4.1. Growth Performance and Feed Efficiency
The inclusion of extracts from Lamiaceae plants has emerged as a promising nutritional strategy for broiler chickens. Beyond their well-known antioxidant and anti-inflammatory properties, many of these botanicals exhibit direct anticoccidial effects, resulting in faster growth, improved feed conversion efficiency, and enhanced gut health in commercial flocks [130,131,132,133,134]. Systematic reviews and controlled experiments demonstrate that incorporating these phytogenic supplements, either alone or in combination with standard anticoccidials, consistently yields heavier finishing weights and more efficient feed conversion. In this context, our experimental trial aimed to assess both the prophylactic and therapeutic efficacy of aqueous extracts from Rosmarinus officinalis, Thymus algeriensis, and Mentha aquatica against avian coccidiosis in broiler chickens. To evaluate their potential, we conducted a controlled trial using aqueous extracts of the plants at two concentrations (15 and 50 mL/L). From day 7, 450 chicks were distributed into nine groups (n = 30), with extract administration via drinking water (Figure 4). On day 10, six groups were challenged with Eimeria spp. (10,000 oocysts/bird), while others remained uninfected or untreated. A toltrazuril-treated group served as a positive control (Figure 4). The aim was to monitor anticoccidial and performance-related effects in vivo. Table 8 summarises the post-infection evolution of performance parameters BWG (Body Weight Gain), TWG (Total Weight Gain), ADG (Average Daily Gain), FI, FCR, EPEF, EBI).
Both the plant species and the extract dosage markedly influenced broiler performance. The group treated with Thymus algeriensis at 15 mL/L (TH-AQ1) exhibited the highest cumulative body weight gain (BWG) at 32 days post-infection (2319.55 ± 43.56 g), significantly outperforming all other groups (p < 0.05) (Table 8). This superior growth was further reflected in total weight gain (TWG) (2098.40 ± 35.05 g) and average daily gain (ADG) (65.58 ± 1.10 g), underscoring the efficacy of this treatment in sustaining growth even under parasitic pressure (Table 8). These trends are visually reinforced in Figure 4, which tracks the evolution of BWG over time. The graph clearly shows that broilers receiving Thymus algeriensis extracts (both TH-AQ1 and TH-AQ2) maintained a consistent upward trajectory in weight gain post-infection, exceeding both the positive control group (TOTRA) and the infected untreated group (INF-NT). By contrast, broilers treated with Mentha aquatica, especially at 50 mL/L (ME-AQ2), exhibited a notable stagnation in growth following parasite inoculation, indicating poor resilience and possible toxicity or suboptimal efficacy. FI and FCR results mirrored the growth patterns (Table 6). Birds from the TH-AQ1 group consumed 2933.52 ± 60.22 g of feed during the whole rearing period (0–32 days) with an FCR of 1.40 ± 0.01, indicating efficient feed utilisation. The TOTRA group, while recording a slightly higher FI (3073.28 ± 30.07 g), exhibited a less efficient FCR (1.60 ± 0.01). Meanwhile, the INF-NT group presented very low feed intake (469.90 ± 5.38 g) and artificially favourable FCR (0.69 ± 0.00), which were associated with severe growth inhibition and increased mortality (Table 8).
Performance indices confirmed these observations (Table 8). The European Production Efficiency Factor (EPEF) and European Broiler Index (EBI) were highest in the TH-AQ1 group (395.08 ± 6.06 and 469.10 ± 7.84, respectively), surpassing even the toltrazuril-treated group (TOTRA: EPEF 319.16 ± 0.93; EBI 375.68 ± 1.39). These results suggest that Thymus algeriensis may not only serve as a viable natural alternative but could also complement or enhance the efficacy of conventional anticoccidial agents or probiotics, particularly by reducing inflammation and improving resilience in broiler chickens. Although further targeted research is necessary to elucidate its mechanisms of action fully, current findings support the potential integration of Thymus algeriensis into poultry health management strategies.
While specific studies on the effects of Thymus algeriensis in broilers remain limited, evidence from related Lamiaceae species indicates that their phytochemicals may modulate the gut microbiota, preserve intestinal barrier integrity, and reduce inflammatory responses during coccidial infections [135]. For instance, supplementation with essential oil blends containing Thymus vulgaris has been shown to significantly enhance growth performance, reduce oocyst shedding, and improve intestinal health in broilers infected with Eimeria spp., often surpassing the efficacy of conventional anticoccidials, such as amprolium [136]. The mode of action is usually attributed to antimicrobial and anti-inflammatory properties of thymol and carvacrol, which improve gut morphology and microbial balance.
Furthermore, studies on Thymus daenensis have shown increased weight gain and enhanced villus height and crypt depth ratios in the jejunum, indicating improved nutrient assimilation [137]. Similarly, Thymus vulgaris supplementation was shown to enhance immunity markers, such as antibody titers post-vaccination against Newcastle Disease, as well as reduce mortality under viral challenge, suggesting a potential immunostimulatory effect of thyme-based phytogenics [138]. Supplementation with thyme at 2–5 g/kg of feed improves body weight gain, feed efficiency, and production indices [139,140]. Even in low-protein diets, thyme helps maintain growth and litter quality by enhancing protein and energy utilisation [141].
Thyme also supports health and immunity. It boosts immunoglobulin levels (IgY, IgM, IgG), increases lymphocyte counts, and improves humoral immunity [142,143]. Its antioxidant properties are notable, enhancing SOD and GPx activity and lowering malondialdehyde (MDA) levels. It reduces inflammatory cytokine expression and upregulates mucin expression in the intestine, improving gut integrity [144]. Carcass and metabolic quality are also enhanced. Thyme reduces abdominal fat, liver weight, and improves carcass yield [145]. It improves lipid profiles by reducing total cholesterol, LDL, VLDL, and triglycerides, while increasing HDL levels. In laying hens, thyme lowers yolk cholesterol and enriches the egg in omega-3 fatty acids without impairing productivity [143]. Importantly, thyme and its active compounds (thymol and carvacrol) present antimicrobial, antiviral, and anti-inflammatory properties, offering a natural alternative to antibiotics in poultry production [146,147]. When combined with other herbs such as garlic or rosemary, or with probiotics, these effects may be amplified [140,141]. Compared to probiotics, which have well-documented benefits on gut morphology, microbial balance, lesion reduction, and immunity [148,149], thyme exhibits promising yet indirect evidence of similar effects. Combining thyme with probiotics could offer a powerful synergy for improving gut health, performance, and disease resistance under coccidial pressure [150], a promising avenue for future research.
Studies suggest Thymus algeriensis contains antioxidant and antimicrobial compounds that can support gut integrity, modulate the gut microbiota, and enhance immune function, especially under parasitic or microbial stress [151]. This plant has been identified as a rich source of bioactive compounds, including flavonoids, phenolic acids, and monoterpenes such as thymol and carvacrol that may help maintain tight junction integrity, reduce oxidative stress, and suppress the growth of pathogenic bacteria, all of which are critical in mitigating the damaging effects of Eimeria infections on the intestinal barrier [95,107,152].
Conversely, groups treated with Mentha aquatica and Rosmarinus officinalis demonstrated inferior performance outcomes, both in terms of growth and feed efficiency. The ME-AQ2 group, which received a high dose of Mentha aquatica aqueous extract, showed significantly reduced performance indices, including the lowest European Production Efficiency Factor (EPEF) and European Broiler Index (EBI) among all treatments (EPEF = 91.71 ± 6.56; EBI = 109.97 ± 7.86). These poor scores aligned with diminished body weight gain and elevated feed conversion ratio, suggesting that high concentrations of entha. aquatica may compromise growth under coccidial stress. Several studies on closely related species, such as Mentha pulegium, have reported similar patterns. For example, Ghalamkari et al. [153] found that supplementation with pennyroyal did not significantly enhance broiler growth performance compared to antibiotic controls[151]. Similarly, Goodarzi and Nanekarani [154] noted that while Mentha. pulegium could improve some carcass traits, higher doses did not lead to better growth or feed conversion efficiency[152]. Other research by Ölmez and Yörük [155] demonstrated that although Mentha pulegium had antimicrobial and antioxidant effects, its impact on performance metrics was inconsistent and dose-dependent[153].
Likewise, birds treated with Rosmarinus officinalis extracts (RO-AQ1 and RO-AQ2) exhibited moderate performance, with complete survival and some reduction in oocyst shedding and lesion severity; however, their EPEF and EBI remained substantially lower than those of both Thymus algeriensis and the toltrazuril-treated group. For example, RO-AQ2 recorded an EPEF of 227.80 ± 2.74 and EBI of 272.24 ± 3.27, in contrast to TH-AQ1 (EPEF = 395.08; EBI = 469.10). These findings align with those of Rasouli et al. (2021), who reported that Rosmarinus officinalis exhibits partial anticoccidial effects, reducing intestinal lesion scores, but is less effective than conventional anticoccidials or certain essential oils in improving feed efficiency and growth performance [156]. Additionally, studies have shown that although rosemary possesses potent antioxidant and antimicrobial properties, its influence on performance indicators such as EPEF or FCR remains modest compared to synthetic drugs or more potent phytogenics [157].
3.4.2. Clinical Protection, Lesion Scores and Anticoccidial Index
The evaluation of anticoccidial efficacy in broiler chickens relies on a series of standardised clinical performance indicators. The survival rate directly reflects the severity of infection and the protective potential of the treatment. In contrast, the relative weight-gain percentage indicates the product's capacity to maintain growth under pathogenic stress. Oocyst output (oocysts per gram of faeces, OPG) quantifies intestinal replication of the parasite, and the lesion score measures intestinal damage. These four variables are consolidated in the Anticoccidial Index (ACI); values > 160 denote adequate protection, whereas lower scores indicate partial or poor efficacy [158,159]. Together, these parameters, summarised in Table 9 and Figure 5, offer a comprehensive assessment of the clinical protection conferred by the tested plant extracts against Eimeria infection in broilers.
As expected, the uninfected, untreated group (NINF-NT) maintained optimal parameters across all metrics, serving as the physiological benchmark (100 % survival and relative weight gain, zero oocysts, and no lesions) (Figure 5). Conversely, the infected untreated group (INF-NT) showed typical signs of severe coccidiosis, with a reduced survival rate (90 %), significantly lowered relative weight gain (74.82 %), maximal oocyst output (1.0639 × 10⁵), and the highest lesion score (2.85 ± 0.57), resulting in a low ACI of 28.50 ± 5.71.
The aqueous extract of Thymus algeriensis showed the most robust protection. At the lower dose (TH-AQ1) (Figure 5), birds achieved 100 % survival, a remarkable 230.5 % relative weight gain, minimal oocyst shedding (median ≈ 1.9 × 10⁴ OPG; box-plot whiskers remain below 3 × 10⁴) (Table 9) and the lowest lesion score (1.43 ± 0.52). These metrics translate into the highest ACI (342.1), surpassing even toltrazuril (TOTRA), which recorded an ACI of 263.6 despite a strong performance (211.2% weight gain and 3.19 × 10⁴ OPG) (Figure 5). The higher thyme dose (TH-AQ2, light green) produced slightly more oocyst output (median ≈ 3.7 × 10⁴ OPG) but still delivered excellent overall protection (100% survival, ACI = 298.2), indicating dose flexibility without loss of efficacy (Figure 5).
Rosmarinus officinalis provided moderate, dose-dependent control (Table 9). The high dose (RO-AQ2) lowered median oocyst counts to ~6 × 10⁴ OPG (Figure 5), improved relative weight gain to 170.2 % and raised the ACI to 200.3, whereas the low dose (RO-AQ1, orange) clustered near the infected controls in both the box-plot and clinical metrics (75.6 % weight gain, ACI = 83.7). Although both rosemary doses achieved 100% survival, their growth suppression and higher shedding rates confirm weaker anticoccidial potency compared to thyme and toltrazuril.
Mentha aquatica was the least effective. Both doses (ME-AQ1, ME-AQ2) generated broad, right-skewed OPG distributions (medians 7–8 × 10⁴) (Figure 5), substantial lesion scores (up to 2.65 ± 0.56), and pronounced growth depression. The higher dose (ME-AQ2) yielded the worst ACI (55.4), despite a 90% survival rate; the lower dose improved slightly (ACI = 73.6) but remained far below the protection afforded by the other treatments (Table 9).
Figure 5 reinforces the numerical conclusions: TH-AQ1 is not only statistically but also visually superior, demonstrating the tightest and lowest oocyst count distribution, better even than toltrazuril. Meanwhile, rosemary offers intermediate, dose-responsive benefits, and mint provides only marginal suppression of parasite shedding. Overall, aqueous Thymus algeriensis emerges as the most promising botanical anticoccidial in vivo, capable of delivering consistent, high-level protection that rivals or surpasses a standard chemotherapeutic drug.
Similar high levels of efficacy have been reported with other botanicals; however, most do not surpass that of toltrazuril. For instance, a tannin-rich herbal blend comprising Artemisia annua, Quercus infectoria, and Allium sativum demonstrated lesion reduction and oocyst suppression comparable to toltrazuril, whilst also improving feed conversion and weight gain in broilers experimentally infected with Eimeria spp. [160]. Likewise, Artemisia sieberi extract reduced oocyst output and enhanced growth performance to a degree comparable to that of monensin, particularly when combined with other plant components [161]. The methanolic extract of Artemisia vestita, a traditional herb from the Himalayas, significantly decreased oocyst output and enhanced feed efficiency, suggesting the presence of highly active compounds [162].
Several studies have explored the efficacy of essential oils and their active compounds. Oils derived from Thymus vulgaris, Origanum vulgare, and Allium sativum have been shown to disrupt early intracellular development of Eimeria tenella, thereby reducing parasite pathogenicity [129]. In a study involving garlic, sage, thyme, echinacea, and oregano, dietary supplementation with this herbal blend restored growth performance to levels comparable to those observed in birds treated with coccidiostats or in uninfected birds. However, lesion scores remained moderately improved [163].
In contrast, rosemary and mint exhibited limited protective effects in our study. These results are supported by other studies, which show that while Artemisia annua improved weight gain, it did not significantly reduce oocyst output or lesion severity [164]. Other plant-based solutions, such as Citrus sinensis essential oil or artemisinin-containing extracts, have demonstrated efficacy in vitro or as litter treatments; however, their in vivo performance against systemic infection remains inferior to that of leading botanical candidates like Thymus algeriensis [165,166].
The findings of the current study align with broader research on the Lamiaceae family, which comprises several herbs recognised for their antiparasitic properties. For example, a comprehensive review by Lahlou et al. [38] documented the efficacy of Rosmarinus officinalis and Thymus vulgaris, two other Lamiaceae members, in improving weight gain, reducing lesion severity, and supporting overall gut health in broiler chickens experimentally infected with Eimeria spp. These plants contain essential oils, phenolic acids, and flavonoids that provide antioxidant, immunomodulatory, and antimicrobial effects, contributing to their anticoccidial action [38].
The current data, when considered in light of these findings, suggest that Thymus algeriensis may not only share but also exceed the bioactivity reported in other Lamiaceae plants. While rosemary and thyme are effective natural growth promoters and moderately anticoccidial, Thymus algeriensis distinguishes itself through its ability to provide consistent protection, even surpassing synthetic drugs under similar experimental conditions. The bioefficacy of Lamiaceae species appears to be closely linked to the presence and concentration of secondary metabolites, which vary between species and chemotypes.
3.4.3. Intestinal Histopathology
Histopathology is the “gold standard” read-out in experimental coccidiosis. Because Eimeria spp. Develop intracellularly, and each species targets a characteristic intestinal segment. Microscopic examination reveals not only the extent of tissue destruction but also the identity and developmental stage of the parasite. Lesion scoring systems such as Johnson & Reid [79] (0 – 4 per segment) have therefore become indispensable for (i) benchmarking new anticoccidials, (ii) explaining performance losses that gross necropsy may miss, and (iii) linking field indicators (weight gain, oocyst output, litter quality) to concrete epithelial damage. Typical coccidial lesions include villus blunting or collapse, crypt hyper-regeneration (the “honey-comb” pattern), fibrino-purulent exudate, heterophil swarms, and the presence of schizonts, gamonts, or developing oocysts within enterocytes [167,168,169]. When these microscopic features are mapped along the gut, they mirror the tropism of the challenge inoculum, thus providing a high-resolution measure of how well each treatment interrupts the parasite's life cycle [167,168].
In our study, across all experimental groups (Figure 6, Table 10), the caecum-to-duodenum histology traced a clear continuum that mirrored the clinical endpoints and the known intestinal tropism of the six Eimeria species in the challenge inoculum.
The negative control (Figure 6A, NINF-NT) displayed an entirely normal mucosa: tall, tapering villi, orderly crypts, and a virtually acellular lamina propria; the global Johnson & Reid score was 0.00 ± 0.00. By contrast, untreated infection (Figure 6C and D, INF-NT) produced the classic picture of uncontrolled coccidiosis. Villi had collapsed into mats of necrotic debris; crypts were ballooned and lake-like, with diffuse coagulative necrosis filling the lamina propria. Heterophils formed dense carpets, and all parasite stages, schizonts, gamonts, and oocysts, were abundant. The composite score climbed to 2.85 ± 0.57, the highest of the study, explaining the poorest weight gain and the heaviest faecal shedding (Table 10). Toltrazuril (Figure 6B, TOTRA) almost normalised the tissue. Only mild villus blunting and a sparse mononuclear sprinkling hinted at prior infection; no intracellular parasites could be detected. The mean lesion index of 1.75 ± 0.62 aligned with the drug’s high anticoccidial index and near-physiological growth. Low-dose aqueous Thymus algeriensis (Figure 6 J–K, TH-AQ1) achieved a similar outcome: villi and crypts were preserved, inflammation was mild, and parasite forms were exceedingly rare, yielding a score of 1.43 ± 0.52 and the lowest oocyst output among the plant preparations. Doubling the thyme dose (Figure 6 L–M, TH-AQ2) did not enhance protection; crypt hyperplasia became slightly more pronounced, but parasites remained sporadic, and the score plateaued at 1.08 ± 0.22, indicating that efficacy had already reached its ceiling at the lower concentration (Table 10).
Aqueous Rosmarinus officinalis produced an intermediate picture. At dose 1 (Figure 6 N–P, RO-AQ1), large necrotic fields and abundant macrogamonts persisted, giving a mean score of 1.17 ± 0.73. Increasing the dose (Figure 6 Q–R, RO-AQ2) halved the parasite burden and restricted damage to patchy villus stumps supported by hyperplastic crypts; however, the score remained moderate at 2.18 ± 0.33, and growth performance was only partly restored. Mentha aquatica was the least effective extract. The low dose (Figure 6 E–F, ME-AQ1) produced heterogeneous damage, characterised by alternating residual crypts and carpets of necrotic, fusiform enterocytes; the score was fixed at 2.50 ± 0.00 (Table 10). The high dose (Figure 6 G–I, ME-AQ2) worsened the picture: villi were obliterated, crypts formed a honey-comb of hyperplasia, fibrino-purulent pockets dissected the lamina propria and macrogamonts or early oocysts were still plentiful, raising the score to 2.65 ± 0.56 and leaving performance indices scarcely better than the untreated infection (Table 10).
Across groups, the microscopic findings directly translated into improved digestive function. Toltrazuril and both thyme doses preserved absorptive villus surface and barrier integrity, sustaining almost normal weight gain. Rosemary allowed for partial villus loss, and the mint extract resulted in extensive loss, diverting nutrients into costly crypt regeneration and provoking heterophil-rich inflammation that correlates with wet litter and increased bacterial risk. The prevalence of macrogamonts in the tissue paralleled the faecal oocyst curve, ranging from <2 × 10⁴ OPG under TH-AQ1 to >1 × 10⁵ OPG in INF-NT. Segment-specific damage mirrored the tropism of the six challenge species: duodenal blunting marked Eimeria praecox, massive jejunal crypt carpets flagged Eimeria maxima, mid-gut necrosis betrayed Eimeria necatrix, and caecal gamonts revealed Eimeria tenella. When the lesion matrix is read alongside the slides, protection ranks unambiguously as follows: toltrazuril ≈ TH-AQ1 > TH-AQ2 > RO-AQ2 > RO-AQ1 > ME-AQ1 ≈ ME-AQ2 ≫ INF-NT. Low-dose aqueous Thymus algeriensis, therefore, emerges as the most convincing botanical substitute for toltrazuril, conferring near-chemotherapeutic mucosal protection. In contrast, rosemary affords only partial rescue and mint extracts, regardless of dose, failing to prevent severe lesions along the digestive tract.
Infection with Eimeria species is known to disrupt the intestinal ecosystem's equilibrium, causing significant damage to the mucosal barrier. The parasite invades epithelial cells, triggering inflammation, villus atrophy, crypt hyperplasia, and disruption of tight junction proteins, which in turn increase intestinal permeability and reduce nutrient absorption [170,171,172]. These pathological changes also disrupt the gut microbial composition, leading to a decline in beneficial bacteria, such as Lactobacillus and Faecalibacterium, while promoting the proliferation of opportunistic pathogens, including Clostridium perfringens, Campylobacter, and Escherichia coli [173,174,175,176]. During an acute infection, there is a marked reduction in short-chain fatty acid-producing bacteria, which are crucial for gut health. At the same time, facultative anaerobes and potential pathogens become more abundant [173,174,176]. The disruption of the gut barrier and microbial balance can persist beyond the acute phase, leading to prolonged adverse effects on growth, nutrient utilisation, and overall animal health [177,178]. These findings highlight the complex interplay between Eimeria, the intestinal barrier, and the microbiota, emphasising the importance of strategies that protect or restore gut integrity and microbial balance to mitigate the impact of coccidiosis [170,172,173,174,175,176].
Phytogenic plant extracts, rich in polyphenols, essential oils, and flavonoids, can counteract the intestinal disturbances caused by Eimeria infection through several complementary mechanisms. These bioactive compounds have been shown to enhance the expression of tight junction proteins, such as occludin and ZO-1, thereby restoring barrier integrity and reducing intestinal permeability [179,180]. They also upregulate antioxidant enzymes, such as superoxide dismutase and glutathione peroxidase, which help mitigate oxidative stress and tissue damage induced by the parasite [179,180]. By modulating key inflammatory pathways, including NF-κB and MAPK, phytogenics reduce the production of pro-inflammatory cytokines and limit the inflammatory response in the gut [181,182].
Many plant extracts also exhibit direct antimicrobial activity, suppressing the overgrowth of pathogenic bacteria, such as Clostridium perfringens, while promoting beneficial microbes and the production of short-chain fatty acids, which are crucial for gut health [183,184,185]. In vivo studies confirm that these effects translate into reduced oocyst shedding, improved gut histology, increased goblet cell numbers, and better overall animal performance during Eimeria infection [179,180,181,185]. Collectively, phytogenic extracts help restore the balance of the gut microbiota, reinforce the mucosal barrier, and reduce inflammation, making them promising natural alternatives or adjuncts to conventional anticoccidial drugs [179,180,181,182,183,185].
In this context, the present study demonstrates that low-dose aqueous Thymus algeriensis extract offers remarkable mucosal protection against Eimeria infection, with performance metrics closely resembling those of toltrazuril. This is particularly notable given its natural origin and low dosage, suggesting that specific phytoconstituents in T. algeriensis possess strong anticoccidial properties capable of modulating host–parasite interactions, maintaining epithelial integrity, and limiting parasite replication [151]. These effects are likely attributable to the presence of thymol, carvacrol, and other bioactive terpenoids, which are known to exert antimicrobial, antioxidant, and immunomodulatory effects [151].
In contrast, rosemary extract exhibited only partial efficacy. While it modestly improved weight gain and reduced lesion severity in a dose-dependent manner, it failed to match the consistent protection observed with Thymus algeriensis. These findings align with those of Peng et al. (2024), who reported that rosemary increased the abundance of Lactobacillus and improved cecal health. However, it did not entirely suppress oocyst shedding or epithelial damage [186]. The partial rescue effect of rosemary may stem from lower concentrations or less potent ratios of active constituents, such as rosmarinic acid or 1,8-cineole, which may not be sufficient to disrupt the complex life cycle of Eimeria in vivo.
Mint extracts, on the other hand, showed negligible protective effects, with high lesion scores and oocyst outputs observed across both tested doses. The limited efficacy of mint may be due to the predominance of menthol and menthone, which lack the broad-spectrum antiparasitic mechanisms found in other Lamiaceae plants, such as Thymus or Origanum [187,188,189].
Phytogenic feed additives, including essential oils, polyphenols, and flavonoid-rich plant extracts, have demonstrated strong potential in mitigating these adverse effects and restoring gut microbial balance. In a recent study, a mixed botanical formula enhanced growth performance and reduced oocyst shedding, while increasing Lactobacillus abundance in the ceca of coccidia-infected broilers [160]. Similarly, dietary supplementation with rosemary extract helped modulate the cecal microbiota and alleviate inflammatory damage by increasing beneficial bacteria and enhancing the production of short-chain fatty acids [186]. Further supporting this, polyherbal formulations containing Artemisia annua and Allium sativum not only reduced Eimeria lesions and oocyst output but also increased microbial diversity and improved the Firmicutes-to-Bacteroidetes ratio, suggesting enhanced gut resilience [160]. Additionally, thyme and oregano essential oils have been shown to restore tight-junction integrity, inhibit the growth of pathogenic bacteria, and stimulate the secretion of mucins and antimicrobial peptides, thereby supporting mucosal recovery [129,190].
These findings suggest that not all Lamiaceae species are equally effective as anticoccidials. The superior performance of Thymus algeriensis in this study highlights the importance of its phytochemical composition and concentration, as well as the synergistic interactions between its active compounds. It also reinforces the need for targeted screening of botanical candidates based not just on traditional use or taxonomic relatedness, but on demonstrated efficacy in relevant in vivo models.
4. Conclusions
In conclusion, our work demonstrates that the judicious use of botanical extracts from Mediterranean Lamiaceae can play a highly effective role in controlling avian coccidiosis. After showing in vitro that these extracts possess potent antioxidant activity and can lyse sporulated oocysts, we validated their in vivo efficacy using a challenge model with six Eimeria species and a comprehensive set of clinical, parasitological, and histological endpoints. Aqueous Thymus algeriensis extracts, even at low doses, protected nearly as well as toltrazuril, preserving villus architecture, limiting inflammation, and reducing parasite stages to almost zero. Rosmarinus officinalis extracts provided an intermediate, dose-dependent level of protection, while Mentha aquatica, despite its antioxidant profile, showed only partial and highly variable anticoccidial effects. Histopathology confirmed these findings, revealing a gradient of lesions that faithfully mirrored the segmental replication patterns of each Eimeria species. Beyond the improvements in weight gain and oocyst reduction, our results underscore that both the concentration of phenolic and flavonoid compounds and the choice of extraction solvent are critical determinants of antiparasitic efficacy. Moving forward, it will be essential to standardise the active-compound profiles of these extracts, explore synergistic combinations, elucidate their molecular modes of action, and undertake large-scale field trials to ensure consistent efficacy and safety. These efforts will pave the way for integrating optimised phytogenic alternatives into sustainable coccidiosis control programs.
Author Contributions
Conceptualization, R.A.L and M.B.; methodology, R.A.L; software, R.A.L; validation, M.B.; A.M; G.A. and L.R.S; formal analysis, M.B.; investigation, R.A.L; A.C.C; K.H; M.B.J resources, A.C.C; K.H; M.BJ data curation, R.A.L; writing—original draft preparation, R.A.L; writing—review and editing, R.A.L and L.R.S; visualization, M.B. and L.R.S; supervision, M.B.; A.M; G.A. and L.R.S.; project administration, G.A and L.R.S; funding acquisition, G.A and L.R.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Programa Operacional Regional do Centro (CEN-TRO-04-3559-FSE-000162) within the European Social Fund (ESF), CICS-UBI (UIDP/00709/2020) financed by National Funds from Fundação para a Ciência e a Tecnologia (FCT), Community Funds (UIDB/00709/2020), by Fundação La Caixa and Fundação para a Ciência e Tecnologia (FCT) under the Programa Promove Project PD21-00023 (PharmaStar).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board of the Department of Agricultural Services of the Wilaya of Boumerdes (protocol code 03570, dated February 16, 2017).
Informed Consent Statement
Not applicable.
Acknowledgements
We gratefully acknowledge the AVIGA Complex and its staff for providing exceptional facilities, expert technical assistance, and seamless logistical support that were essential to our experimental work. We also thank the Department of Agricultural Services of the Wilaya of Boumerdès for their prompt coordination of field sites, provision of agronomic data, and guidance on local practices, which substantially enriched our trials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Parker, C.D.; Lister, S.A.; Gittins, J. Impact Assessment of the Reduction or Removal of Ionophores Used for Controlling Coccidiosis in the UK Broiler Industry. Veterinary Record 2011, 189, no. [Google Scholar] [CrossRef]
- Blake, D.P.; Tomley, F.M. Securing Poultry Production from the Ever-Present Eimeria Challenge. Trends Parasitol 2014, 30, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Pineda, C.; Navarro-Ruíz, J.L.; López-Osorio, S.; Chaparro-Gutiérrez, J.J.; Gómez-Osorio, L.M. Chicken Coccidiosis: From the Parasite Lifecycle to Control of the Disease. Front Vet Sci 2021, 8, 787653. [Google Scholar] [CrossRef]
- Attree, E.; Sanchez-Arsuaga, G.; Jones, M.; Xia, D.; Marugan-Hernandez, V.; Blake, D.; Tomley, F. Controlling the Causative Agents of Coccidiosis in Domestic Chickens; an Eye on the Past and Considerations for the Future. CABI Agriculture and Bioscience 2021, 2, 37. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.D. Coccidiosis in Egg Laying Poultry. Egg Innovations and Strategies for Improvements 2017, 571–579. [Google Scholar] [CrossRef]
- Latha, M.J.; Srikanth, M.K. Occurrence of Caecal Coccidiosis in Poultry. Indian Veterinary Journal 2022, 99, 55–56. [Google Scholar]
- Mesa-Pineda, C.; Navarro-Ruíz, J.L.; López-Osorio, S.; Chaparro-Gutiérrez, J.J.; Gómez-Osorio, L.M. Chicken Coccidiosis: From the Parasite Lifecycle to Control of the Disease. Front Vet Sci 2021, 8, 787653. [Google Scholar] [CrossRef]
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D.; et al. Re-Calculating the Cost of Coccidiosis in Chickens. Vet Res 2020, 51, 115. [Google Scholar] [CrossRef]
- Zaheer, T.; Abbas, R.Z.; Imran, M.; Abbas, A.; Butt, A.; Aslam, S.; Ahmad, J. Vaccines against Chicken Coccidiosis with Particular Reference to Previous Decade: Progress, Challenges, and Opportunities. Parasitol Res 2022, 121, 2749–2763. [Google Scholar] [CrossRef]
- Blake, D.P. Eimeria of Chickens: The Changing Face of an Old Foe. Avian Pathology 2025, 54, 267–278. [Google Scholar] [CrossRef]
- Avi, R.; Reperant, J.M.; Bussiere, F.I.; Silvestre, A.; Le Roux, J.F.; Moreaud, D.; Gonzalez, J. Coccidiosis in Domestic Chickens: A Review of Prevention and Control Strategies[La Coccidiose Chez Les Poulets Domestiques: Revue Sur Les Stratégies de Prévention et de Contrôle]. Inra Productions Animales 2023, 36, 7558. [Google Scholar] [CrossRef]
- Kogut, M.; Miska, K.B.; Antonio Casarin Penha Filho, R.; Gómez-Osorio, L.M.; López-Osorio, S.; Chaparro-Gutiérrez, J.J. Overview of Poultry Eimeria Life Cycle and Host-Parasite Interactions. Front Vet Sci 2020, 7, 384. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Salem, H.M.; Attia, M.M.; Yehia, N.; Abdelkader, A.H.; Mawgod, S.A.; Kamel, N.M.; Alkafaas, S.S.; Alsulami, M.N.; Ahmed, A.E.; et al. Alternatives to Antibiotics against Coccidiosis for Poultry Production: The Relationship between Immunity and Coccidiosis Management – a Comprehensive Review. Annals of Animal Science 2025. [Google Scholar] [CrossRef]
- Lee, Y.; Lu, M.; Lillehoj, H.S. Coccidiosis: Recent Progress in Host Immunity and Alternatives to Antibiotic Strategies. Vaccines (Basel) 2022, 10, 215. [Google Scholar] [CrossRef] [PubMed]
- Bora, C.A.F.; Kumar, V.J.A.; Mathivathani, C. Prevalence of Avian Coccidiosis in India: A Review. Journal of Parasitic Diseases 2024, 48, 181–188. [Google Scholar] [CrossRef]
- Avi, R.; Reperant, J.M.; Bussiere, F.I.; Silvestre, A.; Le Roux, J.F.; Moreaud, D.; Gonzalez, J. Coccidiosis in Domestic Chickens: A Review of Prevention and Control Strategies[La Coccidiose Chez Les Poulets Domestiques: Revue Sur Les Stratégies de Prévention et de Contrôle]. Inra Productions Animales 2023, 36, 7558. [Google Scholar] [CrossRef]
- Rahmani, A.; Ahmed Laloui, H.; Kara, R.; Dems, M.A.; Cherb, N.; Klikha, A.; Blake, D.P. The Financial Cost of Coccidiosis in Algerian Chicken Production: A Major Challenge for the Poultry Sector. Avian Pathology 2024, 53, 368–379. [Google Scholar] [CrossRef]
- Debbou-Iouknane, N.; Benbarek, H.; Ayad, A. Prevalence and Aetiology of Coccidiosis in Broiler Chickens in Bejaia Province, Algeria. Onderstepoort Journal of Veterinary Research 2018, 85, a1590. [Google Scholar] [CrossRef]
- Djemai, S.; Ayadi, O.; Khelifi, D.; Bellil, I.; Hide, G. Prevalence of Eimeria Species, Detected by ITS1-PCR, in Broiler Poultry Farms Located in Seven Provinces of Northeastern Algeria. Trop Anim Health Prod 2022, 54, 250. [Google Scholar] [CrossRef]
- Benabdelhak, A.C.; Derbak, H.; Titouah, H.; Aissanou, S.; Debbou-Iouknane, N.; Ayad, A. Epidemiological Survey on Post Mortem Coccidiosis in Broiler Chicken in Bejaia Province, Northern Algeria. Acta Parasitol 2024, 69, 791–802. [Google Scholar] [CrossRef]
- Ayadi, O.; Djemai, S.; Hide, G. Prevalence of Eimeria Species, Detected by ITS1-PCR Immobilized on FTA Cards, in Future Laying Hens and Breeding Hens in Six Provinces in Northeastern Algeria. Acta Parasitol 2024, 69, 439–445. [Google Scholar] [CrossRef]
- Amina, K.; Bachene, M.S.; Mustapha, O.; Hamdi, T.M. Prevalence of Coccidiosis in Broiler Chickens in Medea, Algeria. Comp Immunol Microbiol Infect Dis 2025, 118, 102323. [Google Scholar] [CrossRef]
- Djemai, S.; Mekroud, A.; Jenkins, M.C. Evaluation of Ionophore Sensitivity of Eimeria Acervulina and Eimeria Maxima Isolated from the Algerian to Jijel Province Poultry Farms. Vet Parasitol 2016, 224, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Peek, H.W.; Landman, W.J.M. Higher Incidence of Eimeria Spp. Field Isolates Sensitive for Diclazuril and Monensin Associated with the Use of Live Coccidiosis Vaccination with ParacoxTM-5 in Broiler Farms. Avian Dis 2006, 50, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, L.; Ren, G.; Si, H.; Song, X.; Liu, X.; Suo, X.; Hu, D. Forward Genetic Analysis of Monensin and Diclazuril Resistance in Eimeria Tenella. Int J Parasitol Drugs Drug Resist 2023, 22, 44–51. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, L.; Si, H.; Liu, X.; Suo, X.; Hu, D. Early Transcriptional Response to Monensin in Sensitive and Resistant Strains of Eimeria Tenella. Front Microbiol 2022, 13, 934153. [Google Scholar] [CrossRef]
- Zhao, H.; Dong, H.; Zhao, Q.; Zhu, S.; Jia, L.; Zhang, S.; Feng, Q.; Yu, Y.; Wang, J.; Huang, B.; et al. Integrated Application of Transcriptomics and Metabolomics Provides Insight into the Mechanism of Eimeria Tenella Resistance to Maduramycin. Int J Parasitol Drugs Drug Resist 2024, 24, 100526. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Huang, B.; Xu, L.; Zhao, Q.; Zhu, S.; Zhao, H.; Dong, H.; Han, H. Comparative Transcriptome Analyses of Drug-Sensitive and Drug-Resistant Strains of Eimeria Tenella by RNA-Sequencing. Journal of Eukaryotic Microbiology 2020, 67, 406–416. [Google Scholar] [CrossRef]
- Schwarz, S.; Shen, J.; Wendlandt, S.; Feßler, A.T.; Wang, Y.; Kadlec, K.; Wu, C.-M. Plasmid-Mediated Antimicrobial Resistance in Staphylococci and Other Firmicutes. Microbiol Spectr 2014, 2, 2014. [Google Scholar] [CrossRef]
- Schwarz, S.; Feßler, A.T.; Loncaric, I.; Wu, C.; Kadlec, K.; Wang, Y.; Shen, J. Antimicrobial Resistance among Staphylococci of Animal Origin. Microbiol Spectr 2018, 6. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, L.; Ren, G.; Si, H.; Song, X.; Liu, X.; Suo, X.; Hu, D. Forward Genetic Analysis of Monensin and Diclazuril Resistance in Eimeria Tenella. Int J Parasitol Drugs Drug Resist 2023, 22, 44–51. [Google Scholar] [CrossRef]
- Zhang, Q.; Plummer, P.J. Mechanisms of Antibiotic Resistance in Campylobacter. In Campylobacter: Third Edition; ASM Press, 2014; pp. 263–276 ISBN 9781683671442.
- Bukari, Z.; Emmanuel, T.; Woodward, J.; Ferguson, R.; Ezughara, M.; Darga, N.; Lopes, B.S. The Global Challenge of Campylobacter: Antimicrobial Resistance and Emerging Intervention Strategies. Trop Med Infect Dis 2025, 10, 25. [Google Scholar] [CrossRef]
- Lopez-Chavarrias, V.; Ugarte-Ruiz, M.; Barcena, C.; Olarra, A.; Garcia, M.; Saez, J.L.; de Frutos, C.; Serrano, T.; Perez, I.; Moreno, M.A.; et al. Monitoring of Antimicrobial Resistance to Aminoglycosides and Macrolides in Campylobacter Coli and Campylobacter Jejuni From Healthy Livestock in Spain (2002–2018). Front Microbiol 2021, 12, 689262. [Google Scholar] [CrossRef] [PubMed]
- Attree, E.; Sanchez-Arsuaga, G.; Jones, M.; Xia, D.; Marugan-Hernandez, V.; Blake, D.; Tomley, F. Controlling the Causative Agents of Coccidiosis in Domestic Chickens; an Eye on the Past and Considerations for the Future. CABI Agriculture and Bioscience 2021, 2, 37. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Yu, Y.H.; Hua, K.F.; Chen, W.J.; Zaborski, D.; Dybus, A.; Hsiao, F.S.H.; Cheng, Y.H. Management and Control of Coccidiosis in Poultry — A Review. Anim Biosci 2024, 37, 1–15. [Google Scholar] [CrossRef]
- Al-Shaebi, E.M.; Al-Quraishy, S.; Maodaa, S.N.; Abdel-Gaber, R. In Vitro Studies for Antiparasitic Activities of Punica Granatum Extract. Microsc Res Tech 2023, 86, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Aitfella Lahlou, R.; Bounechada, M.; Mohammedi, A.; Silva, L.R.; Alves, G. Dietary Use of Rosmarinus Officinalis and Thymus Vulgaris as Anticoccidial Alternatives in Poultry. Anim Feed Sci Technol 2021, 273, 114826. [Google Scholar] [CrossRef]
- Aljohani, A.S.M. Phenolics of Botanical Origin for the Control of Coccidiosis in Poultry. Pak Vet J 2024, 44, 222–228. [Google Scholar] [CrossRef]
- Ismaeil, H.; Mira, N.M.; El-Ashram, S.; Dkhil, M.A.; Abdel-Gaber, R.; Kasem, S.M. Utilizing Coriander as a Natural Remedy for Combating Eimeria Papillata Infection and Oxidative Stress in Mice. Journal of Taibah University for Science 2025, 19, 2467478. [Google Scholar] [CrossRef]
- Blomstrand, B.M.; Enemark, H.L.; Steinshamn, H.; Aasen, I.M.; Johanssen, J.R.E.; Athanasiadou, S.; Thamsborg, S.M.; Sørheim, K.M. Administration of Spruce Bark (Picea Abies) Extracts in Young Lambs Exhibits Anticoccidial Effects but Reduces Milk Intake and Body Weight Gain. Acta Vet Scand 2022, 64, 10. [Google Scholar] [CrossRef]
- Ghaniei, A.; Tohidi, E.; Vafaei, A. Efficacy of a Commercial Mixed Botanical Formula in Treatment and Control of Coccidiosis in Poultry[Djelotvornost Komercijalne Biljne Mješavine u Liječenju i Kontroli Kokcidioze Peradi]. Vet Arh 2022, 92, 723–734. [Google Scholar] [CrossRef]
- Yang, W.C.; Yang, C.Y.; Liang, Y.C.; Yang, C.W.; Li, W.Q.; Chung, C.Y.; Yang, M.T.; Kuo, T.F.; Lin, C.F.; Liang, C.L.; et al. Anti-Coccidial Properties and Mechanisms of an Edible Herb, Bidens Pilosa, and Its Active Compounds for Coccidiosis. Sci Rep 2019, 9, 2896. [Google Scholar] [CrossRef] [PubMed]
- Felici, M.; Tugnoli, B.; De Hoest-Thompson, C.; Piva, A.; Grilli, E.; Marugan-Hernandez, V. Thyme, Oregano, and Garlic Essential Oils and Their Main Active Compounds Influence Eimeria Tenella Intracellular Development. Animals 2024, 14, 77. [Google Scholar] [CrossRef] [PubMed]
- Elshershaby, R.E.; Dkhil, M.A.; Dar, Y.; Abdel-Gaber, R.; Delic, D.; Helal, I.B. Cassia Alata’s Dual Role in Modulating MUC2 Expression in Eimeria Papillata-Infected Jejunum and Assessing Its Anti-Inflammatory Effects. Microsc Res Tech 2024, 87, 2437–2446. [Google Scholar] [CrossRef]
- Jamil, M.; Mansoor, M.; Ali, A.; Shahzad, H.; Rizwan-Ul-Haq; Awan, A. A.; Gul, J. Role of Herbal Immunomodulators in Control of Coccidiosis Disease. Pakistan Journal of Scientific and Industrial Research Series B: Biological Sciences 2017, 60, 176–182. [Google Scholar] [CrossRef]
- Saleh, A.A.; Ebeid, T.A.; Abudabos, A.M. Effect of Dietary Phytogenics (Herbal Mixture) Supplementation on Growth Performance, Nutrient Utilization, Antioxidative Properties, and Immune Response in Broilers. Environmental Science and Pollution Research 2018, 25, 14606–14613. [Google Scholar] [CrossRef]
- Zhao, Y.R.; Chen, Y.P.; Cheng, Y.F.; Qu, H.M.; Li, J.; Wen, C.; Zhou, Y.M. Effects of Dietary Phytosterols on Growth Performance, Antioxidant Status, and Meat Quality in Partridge Shank Chickens. Poult Sci 2019, 98, 3715–3721. [Google Scholar] [CrossRef]
- Sadek, K.M.; Ahmed, H.A.; Ayoub, M.; Elsabagh, M. Evaluation of Digestarom and Thyme as Phytogenic Feed Additives for Broiler Chickens[Prüfung von Digestarom Und Thymian Als Phytogene Futterzusatzstoffe Bei Broilern]. European Poultry Science 2014, 78, 1–12. [Google Scholar] [CrossRef]
- Hashemipour, H.; Kermanshahi, H.; Golian, A.; Veldkamp, T. Metabolism and Nutrition: Effect of Thymol and Carvacrol Feed Supplementation on Performance, Antioxidant Enzyme Activities, Fatty Acid Composition, Digestive Enzyme Activities, and Immune Response in Broiler Chickens. Poult Sci 2013, 92, 2059–2069. [Google Scholar] [CrossRef]
- Al-Garadi, M.A.; Alhotan, R.A.; Hussein, E.O.; Qaid, M.M.; Suliman, G.M.; Al-Badwi, M.A.; Fazea, E.H.; Olarinre, I.O. Effects of a Natural Phytogenic Feed Additive on Broiler Performance, Carcass Traits, and Gut Health under Diets with Optimal and Reduced Energy and Amino Acid Density. Poult Sci 2025. [Google Scholar] [CrossRef]
- Amad, A.A.; Männer, K.; Wendler, K.R.; Neumann, K.; Zentek, J. Effects of a Phytogenic Feed Additive on Growth Performance and Ileal Nutrient Digestibility in Broiler Chickens. Poult Sci 2011, 90, 2811–2816. [Google Scholar] [CrossRef] [PubMed]
- Saleem, K.; Hayat, Z.; Latif, H.R.A.; Tariq, Z.; Riaz, T.; Ullah, S.; Jamil, S.; Rehman, S.; Azam, M. Growth Performance and Gut Microbiota of Broilers Administered Different Levels of Mango Seed Kernel Extract. Revista Brasileira de Ciencia Avicola / Brazilian Journal of Poultry Science 2024, 26, 1847. [Google Scholar] [CrossRef]
- Pliego, A.B.; Tavakoli, M.; Khusro, A.; Seidavi, A.; Elghandour, M.M.M.Y.; Salem, A.Z.M.; Márquez-Molina, O.; Rene Rivas-Caceres, R. Beneficial and Adverse Effects of Medicinal Plants as Feed Supplements in Poultry Nutrition: A Review. Anim Biotechnol 2022, 33, 369–391. [Google Scholar] [CrossRef]
- Murugesan, G.R.; Syed, B.; Haldar, S.; Pender, C. Phytogenic Feed Additives as an Alternative to Antibiotic Growth Promoters in Broiler Chickens. Front Vet Sci 2015, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Fraternale, D.; Ricci, D. Essential Oils of Lamiaceae Family Plants as Antifungals. Biomolecules 2020, Vol. 10, Page 103 2020, 10, 103. [Google Scholar] [CrossRef]
- Uritu, C.M.; Mihai, C.T.; Stanciu, G.D.; Dodi, G.; Alexa-Stratulat, T.; Luca, A.; Leon-Constantin, M.M.; Stefanescu, R.; Bild, V.; Melnic, S.; et al. Medicinal Plants of the Family Lamiaceae in Pain Therapy: A Review. Pain Res Manag 2018, 2018. [Google Scholar] [CrossRef]
- Bekut, M.; Brkić, S.; Kladar, N.; Dragović, G.; Gavarić, N.; Božin, B. Potential of Selected Lamiaceae Plants in Anti(Retro)Viral Therapy. Pharmacol Res 2018, 133, 301–314. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Türkoğlu, A.; Duru, M.; analytical, N.M.-E. journal of; 2007, undefined Antioxidant and Antimicrobial Activity of Russula Delica Fr: An Edidle Wild Mushroom. researchgate.net 2007, 2. [Google Scholar] [CrossRef]
- Blois, M. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Apak, R.; Güçlü, K.; Özyürek, M.; Esin Karademir, S.; Erçǧ, E. The Cupric Ion Reducing Antioxidant Capacity and Polyphenolic Content of Some Herbal Teas. Int J Food Sci Nutr 2006, 57, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Conway, D.P.; McKenzie, M.E.C.N.-S. 6. C.C. 2007 Poultry Coccidiosis: Diagnostic and Testing Procedures; 3rd ed.; Blackwell Pub: Ames, Iowa, 2007; ISBN 978-0-8138-2202-0. [Google Scholar]
- MW REID; BW CALNEK; LR Mc DOUGALD Protozoa-Coccidiosis. In (783-814); Iowa State University Press: Aimes Iowa (USA), 1978; p. 949.
- Reperant JM. Taille Comparée Des Oocystes de Coccidies Du Poulet [Image]. ANSES – Laboratoire de Ploufragan-Plouzané; Ploufragan, 2018.
- Remmal, A.; Achahbar, S.; Bouddine, L.; Chami, N.; Chami, F. In Vitro Destruction of Eimeria Oocysts by Essential Oils. Vet Parasitol 2011, 182, 121–126. [Google Scholar] [CrossRef]
- Medicines Agency, E. Adoption by the Committee for Veterinary Medicinal Products (CVMP) for Guideline on Quality Data Requirements for Applications for Biological Veterinary Medicinal Products Intended for Limited Markets; 2022.
- Medicines Agency, E. Guideline on Safety and Residue Data Requirements for Applications for Non-Immunological Veterinary Medicinal Products Intended for Limited Markets but Not Eligible for Authorisation under Article 23 of Regulation (EU) 2019/6; 2024.
- FDA CVM GFI #185 (VICH GL43) Target Animal Safety for Veterinary Pharmaceutical Products | FDA; 2009.
- Albretsen, J. Target Animal Safety for Veterinary Pharmaceutical Products. Drug Discovery and Evaluation: Safety and Pharmacokinetic Assays 2024, 2515–2528. [Google Scholar] [CrossRef]
- Hafeez, A.; Piral, Q.; Naz, S.; Almutairi, M.H.; Alrefaei, A.F.; Ayasan, T.; Khan, R.U.; Losacco, C. Ameliorative Effect of Pomegranate Peel Powder on the Growth Indices, Oocysts Shedding, and Intestinal Health of Broilers under an Experimentally Induced Coccidiosis Condition. Animals 2023, Vol. 13, Page 3790 2023, 13, 3790. [Google Scholar] [CrossRef]
- Villar-Patiño, G.; Camacho-Rea, M.d.C.; Olvera-García, M.E.; Baltazar-Vázquez, J.C.; Gómez-Verduzco, G.; Téllez, G.; Labastida, A.; Ramírez-Pérez, A.H. Effect of an Alliaceae Encapsulated Extract on Growth Performance, Gut Health, and Intestinal Microbiota in Broiler Chickens Challenged with Eimeria Spp. Animals 2023, 13, 3884. [Google Scholar] [CrossRef] [PubMed]
- Hascoët, A.S.; Torres-Celpa, P.; Riquelme-Neira, R.; Hidalgo-Olate, H. Evaluation of the Effectiveness of a Phytogenic Supplement (Alkaloids and Flavonoids) in the Control of Eimeria Spp. in Experimentally Challenged Broiler Chickens. Animals 2025, 15, 847. [Google Scholar] [CrossRef]
- Marcu, A.; Văcaru-Opriş, I.; Dumitrescu, G.; Petculescu, L.; Marcu, A.; Nicula, M.; Dronca, D.; Kelciov, B. The Influence of Genetics on Economic Efficiency of Broiler Chickens Growth. 2013.
- Chauhan, S.; Singh, V.S.; Thakur, V. Effect of Calotropis Procera (Madar) and Amprolium Supplementation on Parasitological Parameters of Broilers during Mixed Eimeria Species Infection. Vet World 2017, 10, 864–868. [Google Scholar] [CrossRef]
- Holdsworth, P.A.; Conway, D.P.; McKenzie, M.E.; Dayton, A.D.; Chapman, H.D.; Mathis, G.F.; Skinner, J.T.; Mundt, H.C.; Williams, R.B. World Association for the Advancement of Veterinary Parasitology (WAAVP) Guidelines for Evaluating the Efficacy of Anticoccidial Drugs in Chickens and Turkeys. Vet Parasitol 2004, 121, 189–212. [Google Scholar] [CrossRef]
- Johnson, J.; parasitology, W.R.-E. ; 1970, undefined Anticoccidial Drugs: Lesion Scoring Techniques in Battery and Floor-Pen Experiments with Chickens. Elsevier 1970, 28, 30–36. [Google Scholar] [CrossRef]
- Chang, L. yun; Di, K. qian; Xu, J.; Chen, Y. fan; Xi, J. zhong; Wang, D.H.; Hao, E. ying; Xu, L. jun; Chen, H.; Zhou, R. yan Effect of Natural Garlic Essential Oil on Chickens with Artificially Infected Eimeria Tenella. ElsevierL Chang, K Di, J Xu, Y Chen, J Xi, DH Wang, E Hao, L Xu, H Chen, R ZhouVeterinary Parasitology, 2021•Elsevier 2021, 300. [CrossRef]
- Johnson, J.; Reid, W.M. Anticoccidial Drugs: Lesion Scoring Techniques in Battery and Floor-Pen Experiments with Chickens. Exp Parasitol 1970, 28, 30–36. [Google Scholar] [CrossRef]
- Chang, L. yun; Di, K. qian; Xu, J.; Chen, Y. fan; Xi, J. zhong; Wang, D.H.; Hao, E. ying; Xu, L. jun; Chen, H.; Zhou, R. yan Effect of Natural Garlic Essential Oil on Chickens with Artificially Infected Eimeria Tenella. Vet Parasitol 2021, 300, 109614. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Chand, N.; Naz, S.; Alrefaei, A.F.; Albeshr, M.F.; Losacco, C.; Khan, R.U. Response to Dietary Methionine and Organic Zinc in Broilers against Coccidia under Eimeria Tenella-Challenged Condition. Livest Sci 2023, 276. [Google Scholar] [CrossRef]
- Petreska Stanoeva, J.; Balshikevska, E.; Stefova, M.; Tusevski, O.; Simic, S.G. Comparison of the Effect of Acids in Solvent Mixtures for Extraction of Phenolic Compounds From Aronia Melanocarpa. Nat Prod Commun 2020, 15. [Google Scholar] [CrossRef]
- Stanoeva, J.P.; Stefova, M.; Bogdanov, J. Systematic HPLC/DAD/MS n Study on the Extraction Efficiency of Polyphenols from Black Goji: Citric and Ascorbic Acid as Alternative Acid Components in the Extraction Mixture. J Berry Res 2021, 11, 611–630. [Google Scholar] [CrossRef]
- Cabana, R.; Silva, L.R.; Valentão, P.; Viturro, C.I.; Andrade, P.B. Effect of Different Extraction Methodologies on the Recovery of Bioactive Metabolites from Satureja Parvifolia (Phil.) Epling (Lamiaceae). Ind Crops Prod 2013, 48, 49–56. [Google Scholar] [CrossRef]
- Lahlou, R.A.; Gonçalves, A.C.; Bounechada, M.; Nunes, A.R.; Soeiro, P.; Alves, G.; Moreno, D.A.; Garcia-Viguera, C.; Raposo, C.; Silvestre, S.; et al. Antioxidant, Phytochemical, and Pharmacological Properties of Algerian Mentha Aquatica Extracts. Antioxidants 2024, 13, 1512. [Google Scholar] [CrossRef]
- Sultana, B.; Anwar, F.; Ashraf, M. Effect of Extraction Solvent/Technique on the Antioxidant Activity of Selected Medicinal Plant Extracts. Molecules 2009, 14, 2167–2180. [Google Scholar] [CrossRef] [PubMed]
- Cheigh, C.I.; Yoo, S.Y.; Ko, M.J.; Chang, P.S.; Chung, M.S. Extraction Characteristics of Subcritical Water Depending on the Number of Hydroxyl Group in Flavonols. Food Chem 2015, 168, 21–26. [Google Scholar] [CrossRef]
- Howard, L.; Pandjaitan, N. Pressurized Liquid Extraction of Flavonoids from Spinach. J Food Sci 2008, 73 3, 151–157. [Google Scholar] [CrossRef]
- El Houda Lezoul, N.; Belkadi, M.; Habibi, F.; Guillén, F. Extraction Processes with Several Solvents on Total Bioactive Compounds in Different Organs of Three Medicinal Plants. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Safaiee, P.; Taghipour, A.; Vahdatkhoram, F.; Movagharnejad, K. Extraction of Phenolic Compounds from Mentha Aquatica: The Effects of Sonication Time, Temperature and Drying Method. Chemical Papers 2019, 73, 3067–3073. [Google Scholar] [CrossRef]
- Benabdallah, A.; Rahmoune, C.; Boumendjel, M.; Aissi, O.; Messaoud, C. Total Phenolic Content and Antioxidant Activity of Six Wild Mentha Species (Lamiaceae) from Northeast of Algeria. Asian Pac J Trop Biomed 2016, 6, 760–766. [Google Scholar] [CrossRef]
- Fidan, H.; Stankov, S.; Petkova, N.; Dincheva, I.; Stoyanova, A.; Dogan, H. Evaluation of the Phytochemical Profile of Water Mint (Mentha Aquatica L.) from Bulgaria. AIP Conf Proc 2023, 2889. [Google Scholar] [CrossRef]
- Afonso, M.S.; De O Silva, A.M.; Carvalho, E.B.; Rivelli, D.P.; Barros, S.B.; Rogero, M.M.; Lottenberg, A.M.; Torres, R.P.; Mancini-Filho, J. Phenolic Compounds from Rosemary (Rosmarinus Officinalis L.) Attenuate Oxidative Stress and Reduce Blood Cholesterol Concentrations in Diet-Induced Hypercholesterolemic Rats. Nutr Metab (Lond) 2013, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Souadia, A.; Djemoui, A.; Souli, L.; Haiouani, K.; Atoki, A.V.; Djemoui, D.; Messaoudi, M.; Hegazy, S.; Alsaeedi, H.; Barhoum, A. Ultrasound-Assisted Extraction for Maximizing Total Phenolics, Flavonoid Content, and Antioxidant Activity in Thymus Algeriensis: Box-Behnken Experimental Design. Biomass Convers Biorefin 2025, 1–16. [Google Scholar] [CrossRef]
- Ziani, B.E.C.; Heleno, S.A.; Bachari, K.; Dias, M.I.; Alves, M.J.; Barros, L.; Ferreira, I.C.F.R. Phenolic Compounds Characterization by LC-DAD- ESI/MSn and Bioactive Properties of Thymus Algeriensis Boiss. & Reut. and Ephedra Alata Decne. Food Research International 2019, 116, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, Y.H.; Choi, J.; Teng, P.Y.; Yamada, M.; Sugiyama, T.; Kim, W.K. Reduced Bone Formation and Increased Bone Resorption Drive Bone Loss in Eimeria Infected Broilers. Sci Rep 2023, 13, 616. [Google Scholar] [CrossRef]
- Griss, L.G.; Galli, G.M.; Fracasso, M.; Silva, A.D.; Fortuoso, B.; Schetinger, M.R.C.; Morch, V.M.; Boiago, M.M.; Gris, A.; Mendes, R.E.; et al. Oxidative Stress Linked to Changes of Cholinesterase and Adenosine Deaminase Activities in Experimentally Infected Chicken Chicks with Eimeria Spp. Parasitol Int 2019, 71, 11–17. Parasitol Int 2019, 71, 11–17. [Google Scholar] [CrossRef]
- Ghareeb, A.F.A.; Schneiders, G.H.; Richter, J.N.; Foutz, J.C.; Milfort, M.C.; Fuller, A.L.; Yuan, J.; Rekaya, R.; Aggrey, S.E. Heat Stress Modulates the Disruptive Effects of Eimeria Maxima Infection on the Ileum Nutrient Digestibility, Molecular Transporters, and Tissue Morphology in Meat-Type Chickens. PLoS One 2022, 17, e0269131. [Google Scholar] [CrossRef]
- Apak, R.; Güçlü, K.; Özyürek, M.; Karademir, S.E. Novel Total Antioxidant Capacity Index for Dietary Polyphenols and Vitamins C and E, Using Their Cupric Ion Reducing Capability in the Presence of Neocuproine: CUPRAC Method. J Agric Food Chem 2004, 52, 7970–7981. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT - Food Science and Technology 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Babbar, N.; Oberoi, H.S.; Sandhu, S.K.; Bhargav, V.K. Influence of Different Solvents in Extraction of Phenolic Compounds from Vegetable Residues and Their Evaluation as Natural Sources of Antioxidants. J Food Sci Technol 2012, 51, 2568. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Jain, D.C.; Gupta, M.M.; Bhakuni, R.S.; Mishra, H.O.; Sharma, R.P. High-Performance Thin-Layer Chromatographic Analysis of Hepatoprotective Diterpenoids from Andrographis Paniculata. Phytochemical Analysis 2000, 11, 34–36. [Google Scholar] [CrossRef]
- Bicchi, C.; Binello, A.; Rubiolo, P. Determination of Phenolic Diterpene Antioxidants in Rosemary (Rosmarinus Officinalis L.) with Different Methods of Extraction and Analysis. Phytochemical Analysis 2000, 11, 236–242. [Google Scholar] [CrossRef]
- Pereira, O.R.; Macias, R.I.R.; Domingues, M.R.M.; Marin, J.J.G.; Cardoso, S.M. Hepatoprotection of Mentha Aquatica L., Lavandula Dentata L. and Leonurus Cardiaca L. Antioxidants 2019, 8, 267. [Google Scholar] [CrossRef]
- Solhtalab, E.; Nikokar, I.; Mojtahedi, A.; Shokri, R.; Karimian, P.; Mahdavi, E.; Faezi, S. Encapsulation of Mentha Aquatica Methanol Extract in Alginate Hydrogel Promotes Wound Healing in a Murine Model of Pseudomonas Aeruginosa Burn Infection. Int J Biol Macromol 2024, 280, 135920. [Google Scholar] [CrossRef]
- Megdiche-Ksouri, W.; Saada, M.; Soumaya, B.; Snoussi, M.; Zaouali, Y.; Ksouri, R. Potential Use of Wild Thymus Algeriensis and Thymus Capitatus as Source of Antioxidant and Antimicrobial Agents. Journal of New Sciences 2015, 23. [Google Scholar]
- Righi, N.; Boumerfeg, S.; Fernandes, P.A.R.; Deghima, A.; Baali, F.; Coelho, E.; Cardoso, S.M.; Coimbra, M.A.; Baghiani, A. Thymus Algeriensis Bioss & Reut: Relationship of Phenolic Compounds Composition with in Vitro/in Vivo Antioxidant and Antibacterial Activity. Food Research International 2020, 136. [Google Scholar] [CrossRef]
- Hamza, O.; Fahima, A.; Hassani, A. Chemical Composition, Antioxidant Activity of the Essential Oil of Thymus Algeriensis Boiss, North Algeria. International Letters of Chemistry, Physics and Astronomy 2015, 59, 72–80. [Google Scholar] [CrossRef]
- Bencharif-Betina, S.; Benhamed, N.; Benabdallah, A.; Bendif, H.; Benslama, A.; Negro, C.; Plavan, G.; Abd-Elkader, O.H.; De Bellis, L. A Multi-Approach Study of Phytochemicals and Their Effects on Oxidative Stress and Enzymatic Activity of Essential Oil and Crude Extracts of Rosmarinus Officinalis. Separations 2023, 10, 394. [Google Scholar] [CrossRef]
- Sahunie, A. Effect of Rosmarinus Officinalis and Origanum Majorana Extracts on Stability of Sunflower Oil during Storage and Repeated Heating. Oil Crop Science 2024, 9, 29–37. [Google Scholar] [CrossRef]
- Doozakhdarreh, K.; Fatemeh, S.; Khorshidi, J.; Morshedloo, M.R. Essential Oil Content and Components, Antioxidant Activity and Total Phenol Content of Rosemary (Rosmarinus Officinalis L.) as Affected by Harvesting Time and Drying Method. Bulletin of the National Research Centre 2022, 46, 1–12. [Google Scholar] [CrossRef]
- Ayyildiz, S.S.; Pelvan, E.; Karadeniz, B. Optimization of Accelerated Solvent Extraction, Ultrasound Assisted and Supercritical Fluid Extraction to Obtain Carnosol, Carnosic Acid and Rosmarinic Acid from Rosemary. Sustain Chem Pharm 2024, 37, 101422. [Google Scholar] [CrossRef]
- Sánchez-Camargo, A.P.; Mendiola, J.A.; Valdés, A.; Castro-Puyana, M.; García-Cañas, V.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Supercritical Antisolvent Fractionation of Rosemary Extracts Obtained by Pressurized Liquid Extraction to Enhance Their Antiproliferative Activity. J Supercrit Fluids 2016, 107, 581–589. [Google Scholar] [CrossRef]
- Bejenaru, L.E.; Biţă, A.; Mogoşanu, G.D.; Segneanu, A.-E.; Radu, A.; Ciocîlteu, M.V.; Bejenaru, C. Polyphenols Investigation and Antioxidant and Anticholinesterase Activities of Rosmarinus Officinalis L. Species from Southwest Romania Flora. Molecules 2024, 29, 4438. [Google Scholar] [CrossRef]
- Gokbulut, I.; Karaman, Y.; Tursun, A.O. Chemical Composition Phenolic, Antioxidant, and Bio-Herbicidal Properties of the Essential Oil of Rosemary (Rosmarinus Officinalis L.). Acta Scientiarum Polonorum Hortorum Cultus 2022, 21, 21–29. [Google Scholar] [CrossRef]
- Chapman, H.D.; Rathinam, T. Focused Review: The Role of Drug Combinations for the Control of Coccidiosis in Commercially Reared Chickens. Int J Parasitol Drugs Drug Resist 2022, 18, 32. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.A.; Niepceron, A.; Ramakrishnan, C.; Sedano, L.; Hehl, A.B.; Brossier, F.; Smith, N.C. Discovery of a Tyrosine-Rich Sporocyst Wall Protein in Eimeria Tenella. Parasit Vectors 2016, 9, 124. [Google Scholar] [CrossRef]
- Bushkin, G.G.; Motari, E.; Carpentieri, A.; Dubey, J.P.; Costello, C.E.; Robbins, P.W.; Samuelson, J. Evidence for a Structural Role for Acid-Fast Lipids in Oocyst Walls of Cryptosporidium, Toxoplasma, and Eimeria. mBio 2013, 4, e00387–13. [Google Scholar] [CrossRef]
- Mai, K.; Sharman, P.A.; Walker, R.A.; Katrib, M.; de Souza, D.; McConville, M.J.; Wallach, M.G.; Belli, S.I.; Ferguson, D.J.P.; Smith, N.C. Oocyst Wall Formation and Composition in Coccidian Parasites. Mem Inst Oswaldo Cruz 2009, 104, 281–289. [Google Scholar] [CrossRef]
- Arafa, W.M.; Abolhadid, S.M.; Moawad, A.; Abdelaty, A.S.; Moawad, U.K.; Shokier, K.A.M.; Shehata, O.; Gadelhaq, S.M. Thymol Efficacy against Coccidiosis in Pigeon (Columba Livia Domestica). Prev Vet Med 2020, 176, 104914. [Google Scholar] [CrossRef] [PubMed]
- El-Shall, N.A.; Abd El-Hack, M.E.; Albaqami, N.M.; Khafaga, A.F.; Taha, A.E.; Swelum, A.A.; El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; AbuQamar, S.F.; et al. Phytochemical Control of Poultry Coccidiosis: A Review. Poult Sci 2021, 101, 101542. [Google Scholar] [CrossRef] [PubMed]
- Felici, M.; Tugnoli, B.; Ghiselli, F.; Massi, P.; Tosi, G.; Fiorentini, L.; Piva, A.; Grilli, E. In Vitro Anticoccidial Activity of Thymol, Carvacrol, and Saponins. Poult Sci 2020, 99, 5350–5355. [Google Scholar] [CrossRef]
- Jitviriyanon, S.; Phanthong, P.; Lomarat, P.; Bunyapraphatsara, N.; Porntrakulpipat, S.; Paraksa, N. In Vitro Study of Anti-Coccidial Activity of Essential Oils from Indigenous Plants against Eimeria Tenella. Vet Parasitol 2016, 228, 96–102. [Google Scholar] [CrossRef]
- Gadelhaq, S.M.; Arafa, W.M.; Abolhadid, S.M. In Vitro Activity of Natural and Chemical Products on Sporulation of Eimeria Species Oocysts of Chickens. Vet Parasitol 2018, 251, 12–16. [Google Scholar] [CrossRef]
- Maodaa, S.N.; Al-Shaebi, E.M.; Qaid Hailan, W.A.; Abdel-Gaber, R.; Alatawi, A.; Alawwad, S.A.; Al-Quraishy, S. In Vitro Sporulation, Oocysticidal Sporulation Inhibition of Eimeria Papillate and Cytotoxic Efficacy of Methanolic Extract of Thymus Daenensis Leaves. Indian J Anim Res 2024, 58, 99–106. [Google Scholar] [CrossRef]
- Kalita, A. In Vitro Evaluation of Oocysticidal and Sporulation Inhibition Effects of Essential Oil of Orange (Citrus Sinensis) Against Eimeria Tenella. Journal of Animal Research 2022, 12. [Google Scholar] [CrossRef]
- Dkhil, M.A. Anti-Coccidial, Anthelmintic and Antioxidant Activities of Pomegranate (Punica Granatum) Peel Extract. Parasitol Res 2013, 112, 2639–2646. [Google Scholar] [CrossRef]
- Sidiropoulou, E.; Skoufos, I.; Marugan-Hernandez, V.; Giannenas, I.; Bonos, E.; Aguiar-Martins, K.; Lazari, D.; Blake, D.P.; Tzora, A. In Vitro Anticoccidial Study of Oregano and Garlic Essential Oils and Effects on Growth Performance, Fecal Oocyst Output, and Intestinal Microbiota in Vivo. Front Vet Sci 2020, 7. [Google Scholar] [CrossRef]
- Felici, M.; Tugnoli, B.; Ghiselli, F.; Baldo, D.; Ratti, C.; Piva, A.; Grilli, E. Investigating the Effects of Essential Oils and Pure Botanical Compounds against Eimeria Tenella in Vitro. Poult Sci 2023, 102, 102898. [Google Scholar] [CrossRef]
- Pop, L.M.; Varga, E.; Coroian, M.; Nedișan, M.E.; Mircean, V.; Dumitrache, M.O.; Farczádi, L.; Fülöp, I.; Croitoru, M.D.; Fazakas, M.; et al. Efficacy of a Commercial Herbal Formula in Chicken Experimental Coccidiosis. Parasit Vectors 2019, 12, 343. [Google Scholar] [CrossRef] [PubMed]
- Arczewska-Wlosek, A.; Swiatkiewicz, S.; Ognik, K.; Jozefiak, D. Effect of Dietary Crude Protein Level and Supplemental Herbal Extract Blend on Selected Blood Variables in Broiler Chickens Vaccinated against Coccidiosis. Animals 2018, 8, 208. [Google Scholar] [CrossRef]
- Arczewska-Włosek, A.; Świątkiewicz, S.; Kowal, J.; Józefiak, D.; Długosz, J. The Effect of Increased Crude Protein Level and/or Dietary Supplementation with Herbal Extract Blend on the Performance of Chickens Vaccinated against Coccidiosis. Anim Feed Sci Technol 2017, 229, 65–72. [Google Scholar] [CrossRef]
- Shah, S.A.A.; Qureshi, N.A.; Qureshi, M.Z.; Alhewairini, S.S.; Saleem, A.; Zeb, A. Characterization and Bioactivities of M. Arvensis, V. Officinalis and P. Glabrum: In-Silico Modeling of V. Officinalis as a Potential Drug Source. Saudi J Biol Sci 2023, 30. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, S.; Shariatmadari, F.; Karimi Torshizi, M.A.; Ahmadi, H.; Scholey, D. Plectranthus Amboinicus and Rosemary (Rosmarinus Officinalis L.) Essential Oils Effects on Performance, Antioxidant Activity, Intestinal Health, Immune Response, and Plasma Biochemistry in Broiler Chickens. Food Sci Nutr 2023, 11, 3939–3948. [Google Scholar] [CrossRef]
- Madlala, T.; Okpeku, M.; Adeleke, M.A. Understanding the Interactions between Eimeria Infection and Gut Microbiota, towards the Control of Chicken Coccidiosis: A Review. Parasite 2021, 28, 48. [Google Scholar] [CrossRef]
- Abou-Elkhair, R.; Gaafar, K.; Helal, M.; Sameh, G. Bioactive Effect of Dietary Supplementation with Essential Oils Blend of Oregano, Thyme and Garlic Oils on Performance of Broilers Infected with Eimeria Species. Glob Vet 2014, 13, 977–985. [Google Scholar]
- Malekzadeh, M.; Shakouri, M.D.; Abdi Benamar, H. Effect of Thyme Species Extracts on Performance, Intestinal Morphometry, Nutrient Digestibility and Immune Response of Broilers. Kafkas Univ Vet Fak Derg 2018, 24, 783–790. [Google Scholar] [CrossRef]
- Hassanin, O.; El-Sebai, A.; El-Motaal, S.A.; Khalifa, H.A. Experimental Trials to Assess the Immune Modulatory Influence of Thyme and Ginseng Oil on NDV-Vaccinated Broiler Chickens. Open Vet J 2024, 14, 398–406. [Google Scholar] [CrossRef]
- Hassan, F.A.M.; Awad, A. Impact of Thyme Powder (Thymus Vulgaris L.) Supplementation on Gene Expression Profiles of Cytokines and Economic Efficiency of Broiler Diets. Environmental Science and Pollution Research 2017, 24, 15816–15826. [Google Scholar] [CrossRef]
- Ashour, E.A.; Aldhalmi, A.K.; Elolimy, A.A.; Madkour, M.; Elsherbeni, A.I.; Alqhtani, A.H.; Khan, I.M.; Swelum, A.A. Optimizing Broiler Performance, Carcass Traits, and Health: Evaluating Thyme and/or Garlic Powders as Natural Growth Promoters in Antibiotic-Free Diets. Poult Sci 2025, 104. [Google Scholar] [CrossRef] [PubMed]
- Golshahi, A.; Shams Shargh, M.; Dastar, B.; Rahmatnejad, E. The Effect of Thymus Vulgaris Extract and Probiotic on Growth Performance, Blood Parameters, Intestinal Morphology, and Litter Quality of Broiler Chickens Fed Low-Protein Diets. Poult Sci 2025, 104. [Google Scholar] [CrossRef]
- Hashemipour, H.; Kermanshahi, H.; Golian, A.; Veldkamp, T. Effect of Thymol and Carvacrol Feed Supplementation on Performance, Antioxidant Enzyme Activities, Fatty Acid Composition, Digestive Enzyme Activities, and Immune Response in Broiler Chickens. Poult Sci 2013, 92 8, 2059–2069. [Google Scholar] [CrossRef]
- Yalçin, S.; Eser, H.; Onbaşilar, İ.; Yalçin, S. Effects of Dried Thyme (Thymus Vulgaris L.) Leaves on Performance, Some Egg Quality Traits and Immunity in Laying Hens. Ankara Üniversitesi Veteriner Fakültesi Dergisi 2020, 67, 303–311. [Google Scholar] [CrossRef]
- Hassan, F.A.M.; Awad, A. Impact of Thyme Powder (Thymus Vulgaris L.) Supplementation on Gene Expression Profiles of Cytokines and Economic Efficiency of Broiler Diets. Environmental Science and Pollution Research 2017, 24, 15816–15826. [Google Scholar] [CrossRef] [PubMed]
- Abdulkarimi, R.; Daneshyar, M.; Aghazadeh, A. Thyme (Thymus Vulgaris) Extract Consumption Darkens Liver, Lowers Blood Cholesterol, Proportional Liver and Abdominal Fat Weights in Broiler Chickens. Ital J Anim Sci 2011, 10, e20. [Google Scholar] [CrossRef]
- Khan, R.U.; Naz, S.; Nikousefat, Z.; Tufarelli, V.; Laudadio, V. Thymus Vulgaris: Alternative to Antibiotics in Poultry Feed. Worlds Poult Sci J 2012, 68, 401–408. [Google Scholar] [CrossRef]
- Gholami-Ahangaran, M.; Ahmadi-Dastgerdi, A.; Azizi, S.; Basiratpour, A.; Zokaei, M.; Derakhshan, M. Thymol and Carvacrol Supplementation in Poultry Health and Performance. Vet Med Sci 2022, 8, 267–288. [Google Scholar] [CrossRef]
- Mohsin, M.; Zhang, Z.; Yin, G. Effect of Probiotics on the Performance and Intestinal Health of Broiler Chickens Infected with Eimeria Tenella. Vaccines 2022, Vol. 10, Page 97 2022, 10, 97. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, X.; Li, X.; Zhao, J.; Zhang, K.; Hao, X.; Liu, K.; Liu, H. Protective Effect of Lactobacillus Plantarum P8 on Growth Performance, Intestinal Health, and Microbiota in Eimeria-Infected Broilers. Front Microbiol 2021, 12, 705758. [Google Scholar] [CrossRef]
- Elbaz, A.M.; Ashmawy, E.S.; Mourad, D.M.; Amin, S.A.; Khalfallah, E.K.M.; Mohamed, Z.S. Effect of Oregano Essential Oils and Probiotics Supplementation on Growth Performance, Immunity, Antioxidant Status, Intestinal Microbiota, and Gene Expression in Broilers Experimentally Infected with Eimeria. Livest Sci 2025, 291, 105622. [Google Scholar] [CrossRef]
- Lahlou, R.A.; Samba, N.; Soeiro, P.; Alves, G.; Gonçalves, A.C.; Silva, L.R.; Silvestre, S.; Rodilla, J.; Ismael, M.I. Thymus Hirtus Willd. Ssp. Algeriensis Boiss. and Reut: A Comprehensive Review on Phytochemistry, Bioactivities, and Health-Enhancing Effects. Foods 2022, 11, 3195. [Google Scholar] [CrossRef] [PubMed]
- Ben El Hadj Ali, I.; Chaouachi, M.; Bahri, R.; Chaieb, I.; Boussaïd, M.; Harzallah-Skhiri, F. Chemical Composition and Antioxidant, Antibacterial, Allelopathic and Insecticidal Activities of Essential Oil of Thymus Algeriensis Boiss. et Reut. Ind Crops Prod 2015, 77, 631–639. [Google Scholar] [CrossRef]
- Ghalamkari, G.; Toghyani, M.; Landy, N.; Tavalaeian, E. Investigation the Effects Using Different Levels of Mentha Pulegium L. (Pennyroyal) in Comparison with an Antibiotic Growth Promoter on Performance, Carcass Traits and Immune Responses in Broiler Chickens. Asian Pac J Trop Biomed 2012, 2, S1396–S1399. [Google Scholar] [CrossRef]
- Goodarzi, M.; Nanekarani, S. Effects of Feeding Mentha Pulegium L. as an Alternative to Antibiotics on Performance of Broilers. APCBEE Procedia 2014, 8, 53–58. [Google Scholar] [CrossRef]
- Ölmez, Μ.; Yörük, Μ. Effects of Dietary Pennyroyal (Mentha Pulegium L.) Dietary Supplementation on Performance, Carcass Quality, Biochemical Parameters and Duodenal Histomorphology of Broilers. Journal of the Hellenic Veterinary Medical Society 2021, 72, 3213. [Google Scholar] [CrossRef]
- Rasouli, B.; Movahhedkhah, S.; Seidavi, A.; Paz, E.; Laudadio, V.; Ayasan, T.; Mail, V.T. Effect of Pennyroyal (Mentha Pulegium L.) Extract on Performance, Blood Constitutes, Immunity Parameters and Intestinal Microflora in Broiler Chickens. Indian J Anim Sci 2021, 90, 1638–1643. [Google Scholar] [CrossRef]
- Kadhim, M.J. Impact of Adding Aqueous Extract of Mentha Pulegium and Lemon and Its Mixture to Drinking Water on Some Growth Performance and Intestine Organs of Broiler Chicken. IOP Conf Ser Earth Environ Sci 2023, 1262. [Google Scholar] [CrossRef]
- Morisawa, Y.; Kataoka, M.; Kitano, N.; Matsuzawa, T. Studies on Anticoccidial Agents. 10. Synthesis and Anticoccidial Activity of 5-Nitronicotinamide and Its Analogs. ACS PublicationsY Morisawa, M Kataoka, N Kitano, T MatsuzawaJournal of Medicinal Chemistry, 1977•ACS Publications 1977, 20, 129–133. [Google Scholar] [CrossRef]
- Lan, L.; Zuo, B.; Ding, H.; Huang, Y.; Chen, X.; Du, A. Anticoccidial Evaluation of a Traditional Chinese Medicine—Brucea Javanica—in Broilers. Poult Sci 2016, 95, 811–818. [Google Scholar] [CrossRef]
- Ghafouri, S.A.; Ghaniei, A.; Sadr, S.; Amiri, A.A.; Tavanaee Tamannaei, A.E.; Charbgoo, A.; Ghiassi, S.; Dianat, B. Anticoccidial Effects of Tannin-Based Herbal Formulation (Artemisia Annua, Quercus Infectoria, and Allium Sativum) against Coccidiosis in Broilers. Journal of Parasitic Diseases 2023, 47, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Mousavinasab, S.S.; Bozorgmehrifard, M.H.; Kiaei, S.M.M.; Rahbari, S.; Charkhkar, S. Comparison of the Effects of Herbal Compounds and Chemical Drugs for Control of Coccidiosis in Broiler Chickens. Bulg J Vet Med 2022, 25, 69–79. [Google Scholar] [CrossRef]
- Ahad, S.; Tanveer, S.; Nawchoo, I.A.; Malik, T.A. Anticoccidial Activity of Artemisia Vestita (Anthemideae, Asteraceae) - a Traditional Herb Growing in the Western Himalayas, Kashmir, India. Microb Pathog 2017, 104, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Arczewska-Włosek, A.; Świątkiewicz, S. The Efficacy of Selected Feed Additives in the Prevention of Broiler Chicken Coccidiosis under Natural Exposure to Eimeria Spp. Annals of Animal Science 2015, 15, 725–735. Annals of Animal Science 2015, 15, 725–735. [Google Scholar] [CrossRef]
- Pop, L.; Györke, A.; Pop, I.; Friss, Z.; Bărburaș, D.; Andra; Toma-Naic; Zsuzsa; Kálmar; Virgínia; et al. Artemisia Annua Improves Chickens Performances but Has Little Anticoccidial Effect in Broiler Chickens. Animals 2021, 11. [Google Scholar] [CrossRef]
- Sprenger, L.K.; Campestrini, L.H.; Yamassaki, F.T.; Buzatti, A.; Maurer, J.B.B.; Baggio, S.F.Z.; Magalhães, P.M. de; Molento, M.B. Efeito anticoccidiano de extrato hidroalcoólico de Artemisia annua em camas de aves contaminadas com Eimeria sp. Pesquisa Veterinária Brasileira 2015, 35, 649–651. [Google Scholar] [CrossRef]
- Imran, A.; Alsayeqh, A. Anticoccidial Efficacy of Citrus Sinensis Essential Oil in Broiler Chicken. The Pakistan Veterinary Journal 2022, 42, 461–466. [Google Scholar] [CrossRef]
- Hafeez, A.; Piral, Q.; Naz, S.; Almutairi, M.H.; Alrefaei, A.F.; Ayasan, T.; Khan, R.U.; Losacco, C. Ameliorative Effect of Pomegranate Peel Powder on the Growth Indices, Oocysts Shedding, and Intestinal Health of Broilers under an Experimentally Induced Coccidiosis Condition. Animals (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Alhotan, R.A.; Abudabos, A. Anticoccidial and Antioxidant Effects of Plants Derived Polyphenol in Broilers Exposed to Induced Coccidiosis. Environmental Science and Pollution Research 2019, 26, 14194–14199. [Google Scholar] [CrossRef]
- Maratea, K.A.; Miller, M.A. Abomasal Coccidiosis Associated with Proliferative Abomasitis in a Sheep. Journal of Veterinary Diagnostic Investigation 2007, 19, 118–121. [Google Scholar] [CrossRef]
- Sharma, M.K.; Liu, G.; White, D.L.; Kim, W.K. Graded Levels of Eimeria Infection Linearly Reduced the Growth Performance, Altered the Intestinal Health, and Delayed the Onset of Egg Production of Hy-Line W-36 Laying Hens When Infected at the Prelay Stage. Poult Sci 2024, 103. [Google Scholar] [CrossRef] [PubMed]
- Schneiders, G.H. Ontogeny of Intestinal Permeability in Chickens Infected with Eimeria Maxima: Implications for Intestinal Health. 2020.
- Chen, P.; Rehman, M.U.; He, Y.; Li, A.; Jian, F.; Zhang, L.; Huang, S. Exploring the Interplay between Eimeria Spp. Infection and the Host: Understanding the Dynamics of Gut Barrier Function. Vet Q 2025, 45, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Campos, P.M.; Miska, K.B.; Jenkins, M.C.; Yan, X.; Proszkowiec-Weglarz, M. Effects of Eimeria Acervulina Infection on the Luminal and Mucosal Microbiota of the Cecum and Ileum in Broiler Chickens. Sci Rep 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Zhang, S.; Zhou, C.; Tang, X.; Li, C.; Wang, C.; Tang, X.; Suo, J.; Jia, Y.; El-Ashram, S.; et al. Influence of Eimeria Falciformis Infection on Gut Microbiota and Metabolic Pathways in Mice. Infect Immun 2018, 86. [Google Scholar] [CrossRef]
- Madlala, T.; Okpeku, M.; Adeleke, M.A. Understanding the Interactions between Eimeria Infection and Gut Microbiota, towards the Control of Chicken Coccidiosis: A Review. Parasite 2021, 28. [Google Scholar] [CrossRef]
- Liu, J.; Guo, J.; Whitmore, M.A.; Tobin, I.; Kim, D.M.; Zhao, Z.; Zhang, G. Dynamic Response of the Intestinal Microbiome to Eimeria Maxima-Induced Coccidiosis in Chickens. Microbiol Spectr 2024, 12. [Google Scholar] [CrossRef]
- Sharma, M.K.; Liu, G.; White, D.L.; Kim, W.K. Graded Levels of Eimeria Infection Linearly Reduced the Growth Performance, Altered the Intestinal Health, and Delayed the Onset of Egg Production of Hy-Line W-36 Laying Hens When Infected at the Prelay Stage. Poult Sci 2024, 103. [Google Scholar] [CrossRef]
- Choi, J.; Goo, D.; Sharma, M.K.; Ko, H.; Liu, G.; Paneru, D.; Choppa, V.S.R.; Lee, J.; Kim, W.K. Effects of Different Eimeria Inoculation Doses on Growth Performance, Daily Feed Intake, Gut Health, Gut Microbiota, Foot Pad Dermatitis, and Eimeria Gene Expression in Broilers Raised in Floor Pens for 35 Days. Animals (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Murshed, M.; Al-Tamimi, J.; Aljawdah, H.M.A.; Al-Quraishy, S. Pharmacological Effects of Grape Leaf Extract Reduce Eimeriosis-Induced Inflammation, Oxidative Status Change, and Goblet Cell Response in the Jejunum of Mice. Pharmaceuticals 2023, 16. [Google Scholar] [CrossRef]
- Maodaa, S.; Al-Shaebi, E.M.; Abdel-Gaber, R.; Alatawi, A.; Alawwad, S.; Alhomoud, D.; Al-Quraishy, S. Anticoccidial and Antioxidant Activities of an Ethanolic Extract of Teucrium Polium Leaves on Eimeria Papillate-Infected Mice. Vet Sci 2024, 11. [Google Scholar] [CrossRef]
- Qasem, M.A.A.; Dkhil, M.A.; Al-Shaebi, E.M.; Murshed, M.; Mares, M.; Al-Quraishy, S. Rumex Nervosus Leaf Extracts Enhance the Regulation of Goblet Cells and the Inflammatory Response during Infection of Chickens with Eimeria Tenella. J King Saud Univ Sci 2020, 32, 1818–1823. [Google Scholar] [CrossRef]
- Jiao, J.Y.; Yang, Y.Q.; Liu, M.J.; Li, J.G.; Cui, Y.; Yin, S.J.; Tao, J.P. Artemisinin and Artemisia Annua Leaves Alleviate Eimeria Tenella Infection by Facilitating Apoptosis of Host Cells and Suppressing Inflammatory Response. Vet Parasitol 2018, 254, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Muthamilselvan, T.; Kuo, T.F.; Wu, Y.C.; Yang, W.C. Herbal Remedies for Coccidiosis Control: A Review of Plants, Compounds, and Anticoccidial Actions. Evid Based Complement Alternat Med 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Lillehoj, H.S.; Lee, Y.; Adetunji, A.O.; Omaliko, P.C.; Kang, H.W.; Fasina, Y.O. Use of Selected Plant Extracts in Controlling and Neutralizing Toxins and Sporozoites Associated with Necrotic Enteritis and Coccidiosis. Applied Sciences 2024, 14. [Google Scholar] [CrossRef]
- Han, M.; Hu, W.; Chen, T.; Guo, H.; Zhu, J.; Chen, F. Anticoccidial Activity of Natural Plants Extracts Mixture against Eimeria Tenella: An in Vitro and in Vivo Study. Front Vet Sci 2022, 9. [Google Scholar] [CrossRef]
- Peng, F.; Duan, J.; He, X.; Xie, K.; Song, Z. Effects of Dietary Water-Soluble Extract of Rosemary Supplementation on Growth Performance and Intestinal Health of Broilers Infected with Eimeria Tenella. J Anim Sci 2024, 102. [Google Scholar] [CrossRef] [PubMed]
- Tjitraresmi, A.; Moektiwardoyo, M.; Susilawati, Y.; Shiono, Y. Antimalarial Activity of Lamiaceae Family Plants: Review. Systematic Reviews in Pharmacy 2020, 11, 324–333. [Google Scholar] [CrossRef]
- Bailén, M.; Illescas, C.; Quijada, M.; Martínez-Díaz, R.A.; Ochoa, E.; Gómez-Muñoz, M.T.; Navarro-Rocha, J.; González-Coloma, A. Anti-Trypanosomatidae Activity of Essential Oils and Their Main Components from Selected Medicinal Plants. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Maciel, M.S.P.; Reis, A.S. dos; Fidelis, Q.C. Antileishmanial Potential of Species from the Family Lamiaceae: Chemical and Biological Aspects of Non-Volatile Compounds. Acta Trop 2022, 228. [Google Scholar] [CrossRef]
- Felici, M.; Tugnoli, B.; De Hoest-Thompson, C.; Piva, A.; Grilli, E.; Marugan-Hernandez, V. Thyme, Oregano, and Garlic Essential Oils and Their Main Active Compounds Influence Eimeria Tenella Intracellular Development. Animals 2023, 14, 77. [Google Scholar] [CrossRef]
Figure 1.
Total polyphenol content (TPC) expressed as gallic acid equivalents (µg GAE/mg DW) and the total flavonoids (TFC) content as quercetin equivalents (µg QE/g DW) of hydromethanolic (HM) and aqueous (AQ) extracts of Mentha aquatica (ME), Rosmarinus officinalis (RO), Thymus algeriensis (TH). Values show means ± standard deviation (SEM) of three independent assays. Statistical analysis: A one-way ANOVA with Tukey's multiple comparisons test was performed for each extract compared to the others. Different letters above the bars indicate significant differences between groups at p < 0.05. Each letter denotes a homogeneous group; bars sharing the same letter are not significantly different, while bars with different letters indicate statistically significant differences.
Figure 1.
Total polyphenol content (TPC) expressed as gallic acid equivalents (µg GAE/mg DW) and the total flavonoids (TFC) content as quercetin equivalents (µg QE/g DW) of hydromethanolic (HM) and aqueous (AQ) extracts of Mentha aquatica (ME), Rosmarinus officinalis (RO), Thymus algeriensis (TH). Values show means ± standard deviation (SEM) of three independent assays. Statistical analysis: A one-way ANOVA with Tukey's multiple comparisons test was performed for each extract compared to the others. Different letters above the bars indicate significant differences between groups at p < 0.05. Each letter denotes a homogeneous group; bars sharing the same letter are not significantly different, while bars with different letters indicate statistically significant differences.
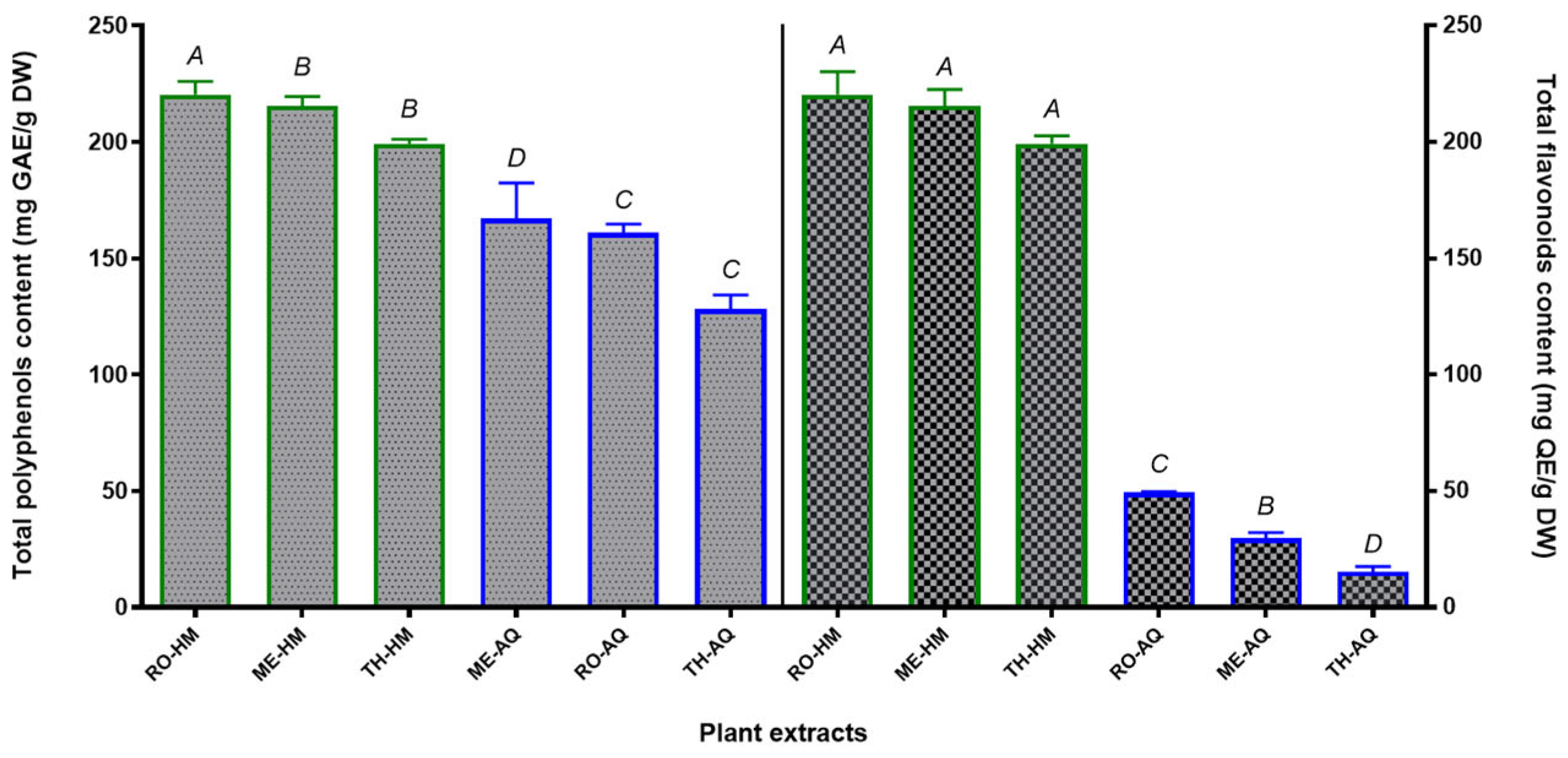
Figure 2.
Viability of Eimeria oocysts after exposure to hydro-methanolic (HM) and aqueous (AQ) extracts of Rosmarinus officinalis (RO), Thymus algeriensis (TH), and Mentha aquatica (ME), across a concentration gradient (C1 to C10). Oocyst viability (%) was assessed following incubation with plant extracts at increasing concentrations ranging from 0.2 to 20 mg/mL. The bar plots show mean viability ± standard error (n = 3). Black bars indicate untreated control. Hydro-methanolic and aqueous extracts are represented by orange and green (RO), red and magenta (TH), and beige and brown (ME). Statistical differences between groups were determined using two-way ANOVA followed by Tukey’s multiple comparisons test. For each concentration point (C1–C10), treatments not sharing a common letter differ significantly (p < 0.05). The analysis revealed significant effects of concentration, extract type, and their interaction on oocyst viability (p < 0.0001 for all factors).
Figure 2.
Viability of Eimeria oocysts after exposure to hydro-methanolic (HM) and aqueous (AQ) extracts of Rosmarinus officinalis (RO), Thymus algeriensis (TH), and Mentha aquatica (ME), across a concentration gradient (C1 to C10). Oocyst viability (%) was assessed following incubation with plant extracts at increasing concentrations ranging from 0.2 to 20 mg/mL. The bar plots show mean viability ± standard error (n = 3). Black bars indicate untreated control. Hydro-methanolic and aqueous extracts are represented by orange and green (RO), red and magenta (TH), and beige and brown (ME). Statistical differences between groups were determined using two-way ANOVA followed by Tukey’s multiple comparisons test. For each concentration point (C1–C10), treatments not sharing a common letter differ significantly (p < 0.05). The analysis revealed significant effects of concentration, extract type, and their interaction on oocyst viability (p < 0.0001 for all factors).
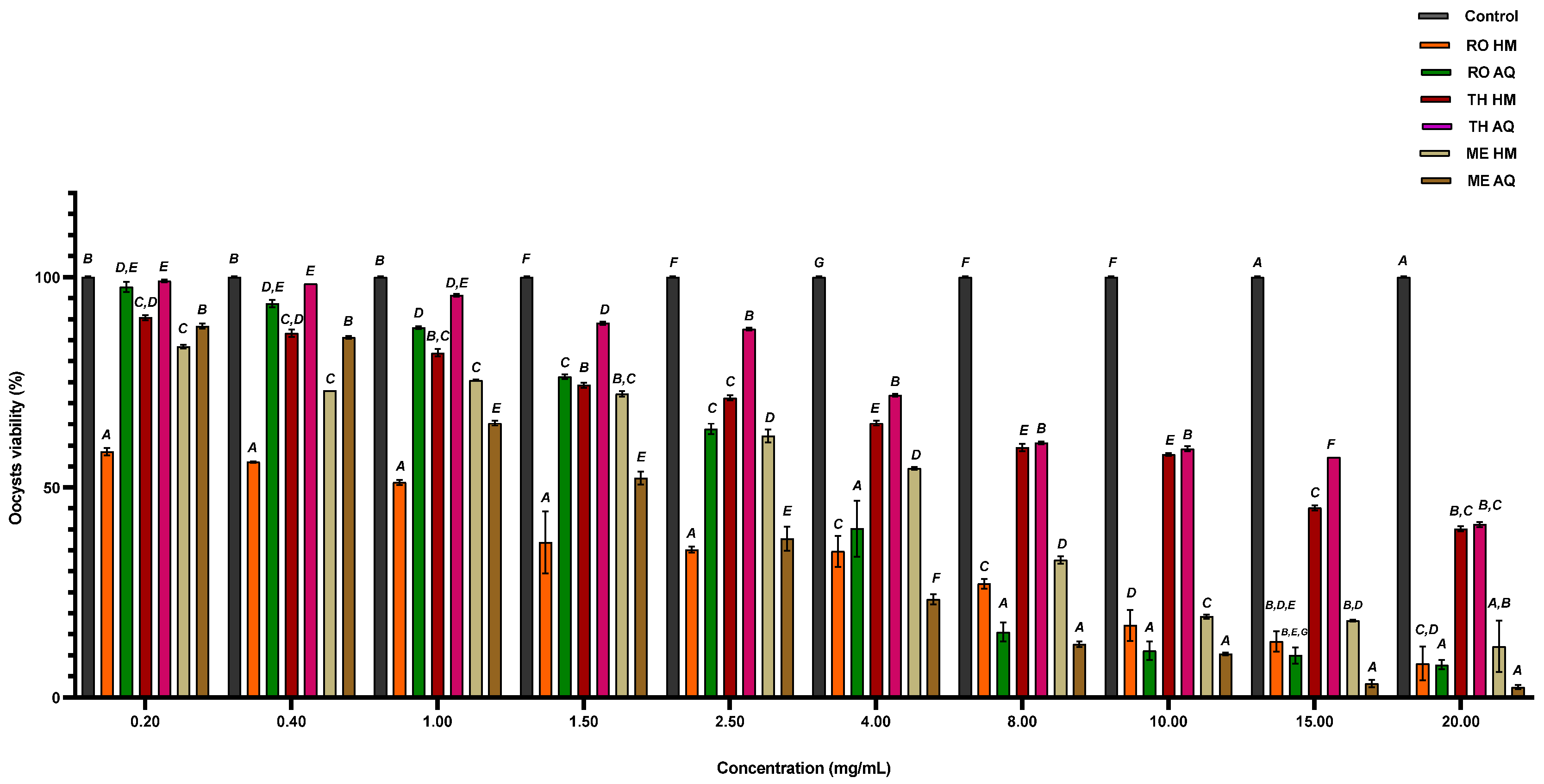
Figure 3.
Dose-response profiles of aqueous (AQ) and hydromethanolic (HM) extracts from Rosmarinus officinalis, Thymus algeriensis and Mentha aquatica showing oocyst inhibition (%) and corresponding UV absorbance at 273 nm.
Figure 3.
Dose-response profiles of aqueous (AQ) and hydromethanolic (HM) extracts from Rosmarinus officinalis, Thymus algeriensis and Mentha aquatica showing oocyst inhibition (%) and corresponding UV absorbance at 273 nm.
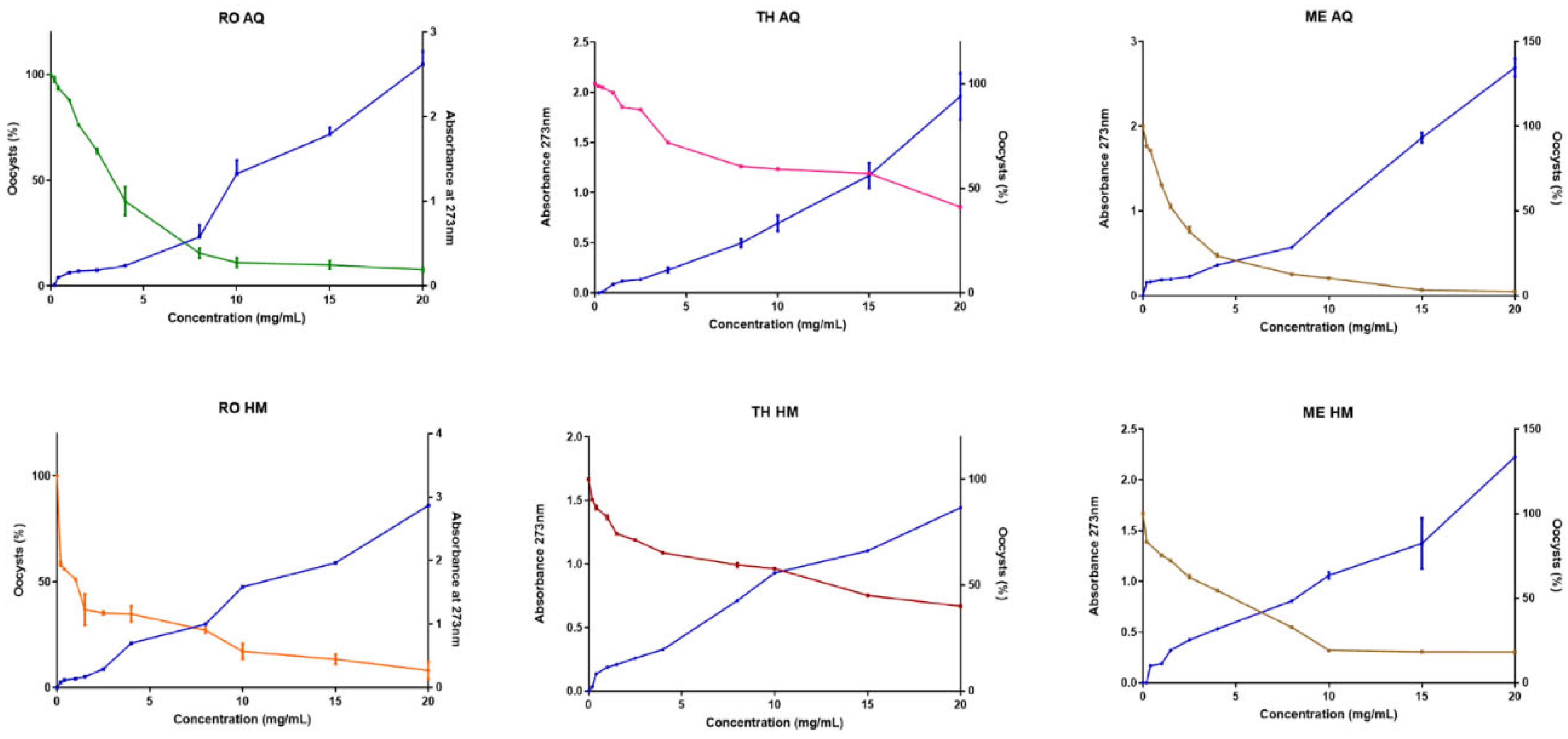
Figure 4.
Evolution of body weight gain (BWG) in broiler chickens from pre-infection (day -10) to the end of the experiment (day 32) in response to treatment with aqueous plant extracts. Arrows indicate the day of Eimeria inoculation. THAQ1 and THAQ2 = Thymus algeriensis; ROAQ1 and ROAQ2 = Rosmarinus officinalis; MEAQ1 and MEAQ2 = Mentha aquatica; INF-NT = infected untreated; NINF-NT = uninfected untreated; TOTRA = toltrazuril-treated group.
Figure 4.
Evolution of body weight gain (BWG) in broiler chickens from pre-infection (day -10) to the end of the experiment (day 32) in response to treatment with aqueous plant extracts. Arrows indicate the day of Eimeria inoculation. THAQ1 and THAQ2 = Thymus algeriensis; ROAQ1 and ROAQ2 = Rosmarinus officinalis; MEAQ1 and MEAQ2 = Mentha aquatica; INF-NT = infected untreated; NINF-NT = uninfected untreated; TOTRA = toltrazuril-treated group.
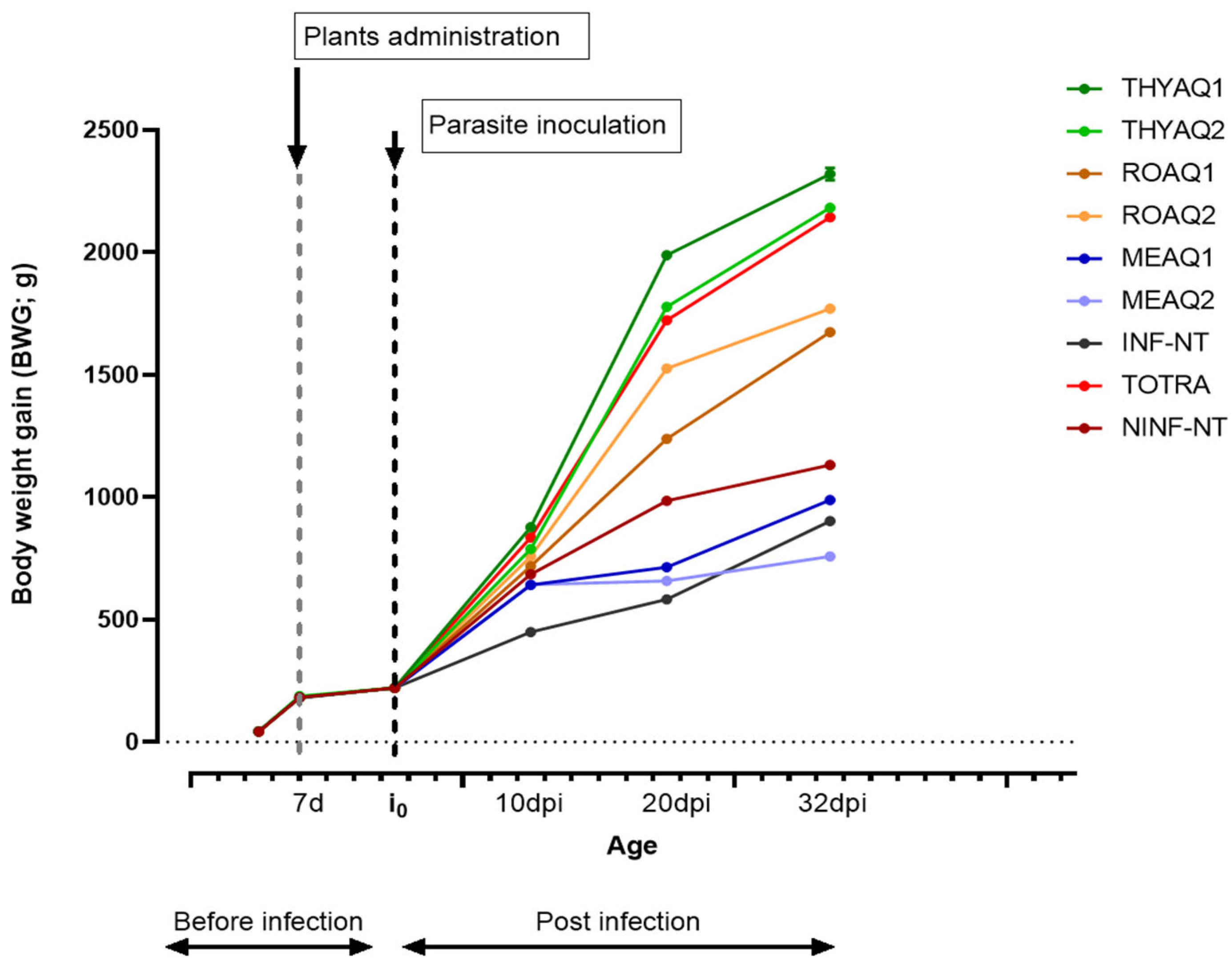
Figure 5.
Longitudinal shedding of Eimeria oocysts in broiler chickens following different treatments. Box-and-whisker plots present faecal oocyst counts (oocysts per gram, OPG) measured every two days from 10 to 32 days post-infection (i.e., 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30 and 32 dpi). For each treatment group (n = 6), all time-point values were pooled to depict overall shedding pressure during the patent period. Boxes represent inter-quartile ranges (25th–75th percentiles); horizontal lines mark medians; whiskers indicate minima and maxima. NINF-NT (non-infected, non-treated), TH-AQ1 and TH-AQ2 (low and high doses of aqueous Thymus algeriensis, TOTRA (toltrazuril; red), RO-AQ2 and RO-AQ1 (high and low doses of aqueous Rosmarinus officinalis, ME-AQ1 and ME-AQ2 (low and high doses of aqueous Mentha aquatica), and INF-NT (infected, non-treated; grey). Lower, more compact boxes denote more potent suppression of oocyst shedding across the 10–32 dpi window; TH-AQ1 achieves the most significant overall reduction, surpassing toltrazuril, whereas mint extracts and low-dose rosemary show limited control.
Figure 5.
Longitudinal shedding of Eimeria oocysts in broiler chickens following different treatments. Box-and-whisker plots present faecal oocyst counts (oocysts per gram, OPG) measured every two days from 10 to 32 days post-infection (i.e., 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30 and 32 dpi). For each treatment group (n = 6), all time-point values were pooled to depict overall shedding pressure during the patent period. Boxes represent inter-quartile ranges (25th–75th percentiles); horizontal lines mark medians; whiskers indicate minima and maxima. NINF-NT (non-infected, non-treated), TH-AQ1 and TH-AQ2 (low and high doses of aqueous Thymus algeriensis, TOTRA (toltrazuril; red), RO-AQ2 and RO-AQ1 (high and low doses of aqueous Rosmarinus officinalis, ME-AQ1 and ME-AQ2 (low and high doses of aqueous Mentha aquatica), and INF-NT (infected, non-treated; grey). Lower, more compact boxes denote more potent suppression of oocyst shedding across the 10–32 dpi window; TH-AQ1 achieves the most significant overall reduction, surpassing toltrazuril, whereas mint extracts and low-dose rosemary show limited control.
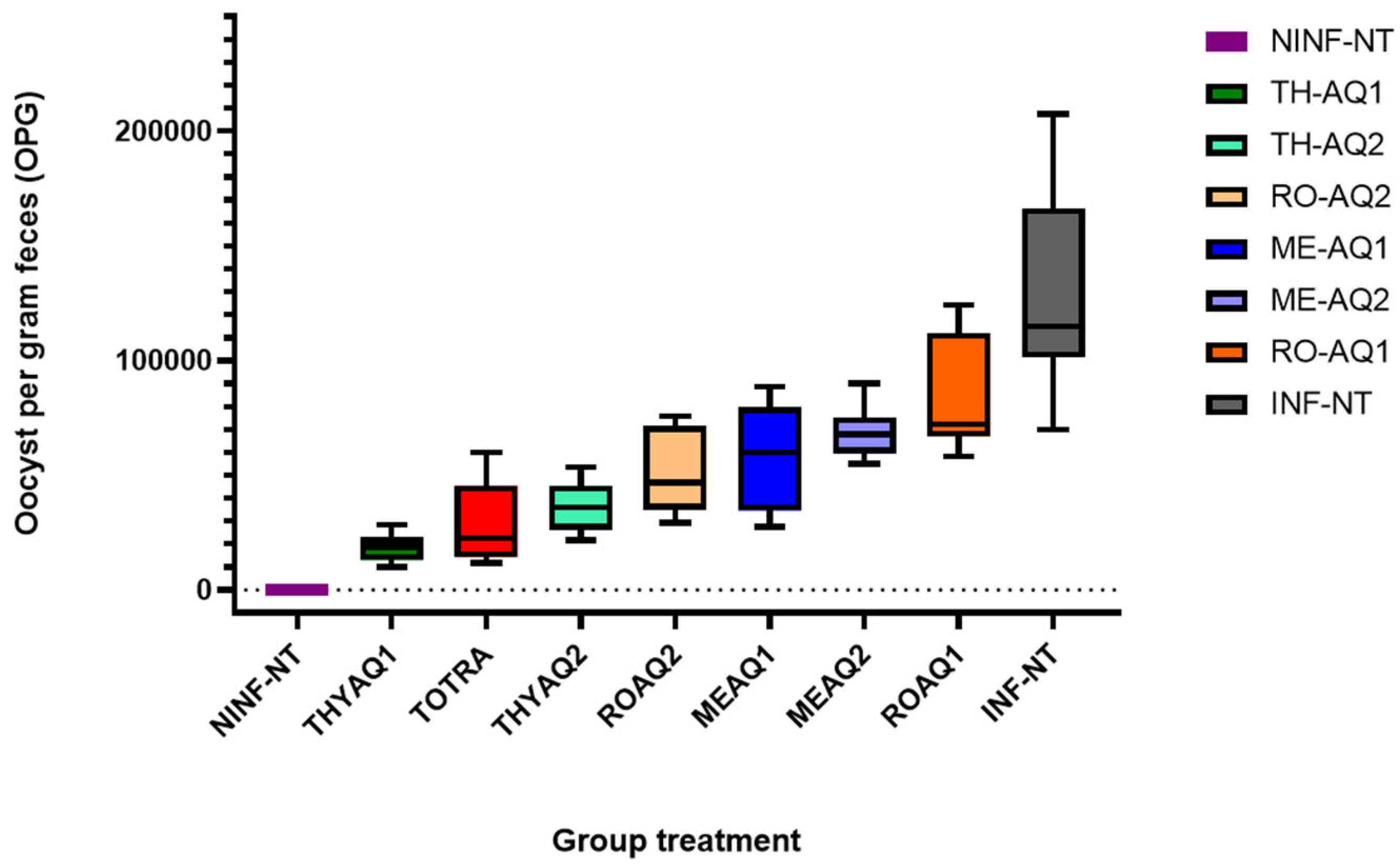
Figure 6.
Representative intestinal histological sections for all groups (A–R). A—NINF-NT (non-infected, non-treated): intact villi and crypts, score 0. B—TOTRA (toltrazuril): slight villus blunting, no parasites, score ≈ 1.7. C, D—INF-NT (infected, non-treated): villus collapse, necrotic crypt lakes, abundant schizonts/gamonts; score ≈ 2.9. E, F—ME-AQ1 (Mentha aquatica): mixed fields, residual crypts and carpets of necrotic fusiform enterocytes, score ≈ 2.5. G, H, I—ME-AQ2 (Mentha aquatica): honeycomb crypt hyperplasia (G), fibrino-purulent pocket (H) and numerous macrogamonts/oocysts (red arrows, I); score ≈ 2.6. J, K—TH-AQ1 (Thymus algeriensis): largely preserved architecture, mild lamina propria infiltrate, parasites rare; score ≈ 1.4. L, M—TH-AQ2 (Thymus algeriensis): similar to TH-AQ1 with slightly greater crypt hyperplasia, score ≈ 1.1. N, P—RO-AQ1 (Rosmarinus officinalis): marked villus loss, focal necrosis, abundant macrogamonts (yellow arrows), score ≈ 3. O (inset, RO-AQ1, higher power): degenerating enterocytes and inflammatory debris. Q, R—RO-AQ2 (Rosmarinus officinalis): patchy villus stumps with regenerating crypts (Q) and scattered parasite stages (R); score ≈ 2.2.
Figure 6.
Representative intestinal histological sections for all groups (A–R). A—NINF-NT (non-infected, non-treated): intact villi and crypts, score 0. B—TOTRA (toltrazuril): slight villus blunting, no parasites, score ≈ 1.7. C, D—INF-NT (infected, non-treated): villus collapse, necrotic crypt lakes, abundant schizonts/gamonts; score ≈ 2.9. E, F—ME-AQ1 (Mentha aquatica): mixed fields, residual crypts and carpets of necrotic fusiform enterocytes, score ≈ 2.5. G, H, I—ME-AQ2 (Mentha aquatica): honeycomb crypt hyperplasia (G), fibrino-purulent pocket (H) and numerous macrogamonts/oocysts (red arrows, I); score ≈ 2.6. J, K—TH-AQ1 (Thymus algeriensis): largely preserved architecture, mild lamina propria infiltrate, parasites rare; score ≈ 1.4. L, M—TH-AQ2 (Thymus algeriensis): similar to TH-AQ1 with slightly greater crypt hyperplasia, score ≈ 1.1. N, P—RO-AQ1 (Rosmarinus officinalis): marked villus loss, focal necrosis, abundant macrogamonts (yellow arrows), score ≈ 3. O (inset, RO-AQ1, higher power): degenerating enterocytes and inflammatory debris. Q, R—RO-AQ2 (Rosmarinus officinalis): patchy villus stumps with regenerating crypts (Q) and scattered parasite stages (R); score ≈ 2.2.
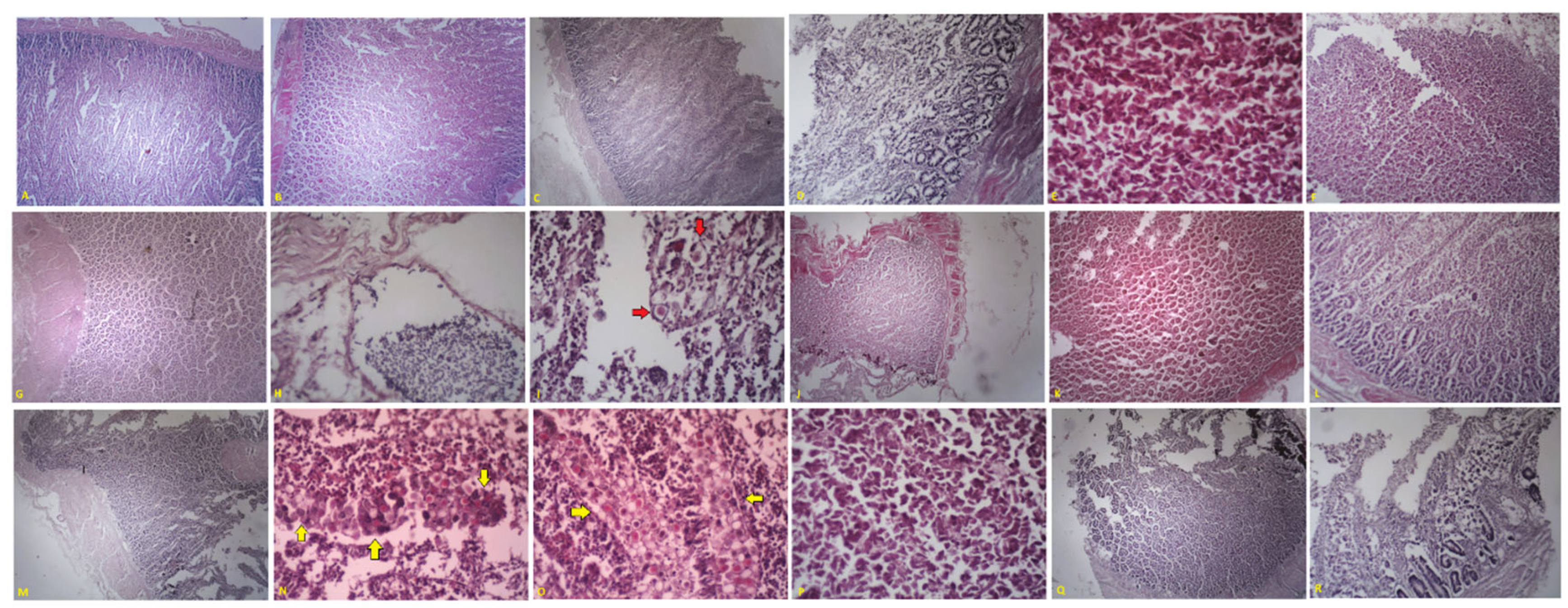

Table 1.
Geographic coordinates and yields of aqueous and hydromethanolic extracts of three Lamiaceae species (Mentha aquatica, Rosmarinus officinalis, Thymus algeriensis.
Table 1.
Geographic coordinates and yields of aqueous and hydromethanolic extracts of three Lamiaceae species (Mentha aquatica, Rosmarinus officinalis, Thymus algeriensis.
| Plants | Geographic coordinates | Yield (%) |
|---|---|---|
| Mentha aquatica | 36°10'49.1"N 4°36'43.6"E | AQ: 21.640 ± 1.619 |
| HM: 19.300 ± 0.080 | ||
| Rosmarinus officinalis | 36°11'00.4"N 4°25'53.7"E | AQ: 18.577 ± 3.696 |
| HM: 15.913 ± 4.110 | ||
| Thymus algeriensis | 36°10'49.1"N 4°36'43.6"E | AQ: 11.675 ± 0.625 |
| HM: 11.455 ± 0.010 |
Table 2.
Ingredients and nutrient composition of the basal diets administered during the trial.
| Category | Substance | Quantity | |
|---|---|---|---|
| Starter (1 to 21 days) | Grower (22 to 42 d) | ||
| Trace Elements | Iron | 5000 mg | 5000 mg |
| Copper | 1000 mg | 1000 mg | |
| Zinc | 5000 mg | 5000 mg | |
| Manganese | 7500 mg | 7500 mg | |
| Iodine | 150 mg | 150 mg | |
| Selenium | 20 mg | 20 mg | |
| Vitamins | Vitamin A | 15000 IU | 10000 IU |
| Vitamin D3 | 3000 IU | 2000 IU | |
| Vitamin E | 100 mg | 100 mg | |
| Vitamin K3 | 150 mg | 150 mg | |
| Vitamin B1 (Thiamine) | 125 mg | 125 mg | |
| Vitamin B2 (Riboflavin) | 400 mg | 400mg | |
| Vitamin PP (Nicotinic acid) | 2500 mg | 2500 mg | |
| Vitamin B5 (Calcium pantothenate) | 825 mg | 825 mg | |
| Vitamin B6 (Pyridoxine) | 200 mg | 200 mg | |
| Vitamin H (Biotin) | 5 mg | 5 mg | |
| Vitamin B9 (Folic acid) | 55 mg | 55 mg | |
| Vitamin B12 (Cyanocobalamin) | 1.25 mg | 1.25mg | |
| Technical Parameters | Choline chloride | 30000 mg | 30000 mg |
| Crude ash | 12% | 8% | |
| Calcium | 1.20% | 1% | |
| Sodium | 0.20% | 0.25% | |
| Crude protein | 21.70% | 20% | |
| Methionine + Betaine | 180000 mg | 100000 mg | |
| Metabolisable energy | 2850 kcal/kg | 3100 kcal/kg | |
| Additional Nutrients | Lysine | 1.30% | 1.10% |
| Threonine | 0.80% | 0.75% | |
| Tryptophan | 0.20% | 0.20% | |
| Arginine | 1.30% | 1.20% | |
| Phosphorus | 0.50% | 0.45% | |
| Fat content | 3.50% | 5% | |
| Crude fiber | 4% | 4% | |
| Ingredients | Corn | 55% | 55% |
| Soybean | 25% | 0.25 | |
| Wheat Bran | 10% | 10% | |
Table 3.
Broiler group treatments for botanical natural products and anticoccidial tests.
| Group | Infection status | Treatment description | Parasite dosage1 |
|---|---|---|---|
| A (TH-AQ1) | Infected | Thymus algeriensis aqueous extract, 15 mL/L (1.5g/L) | 10000 |
| B (TH-AQ2) | Infected | Thymus algeriensis aqueous extract, 50 mL/L (5g/L) | 10000 |
| C (RO-AQ1) | Infected | Rosmarinus officinalis methanolic extract, 15 mL/L (1.5g/L) | 10000 |
| D (RO-AQ2) | Infected | Rosmarinus officinalis methanolic extract, 50 mL/L (5g/L) | 10000 |
| E (ME-AQ1) | Infected | Mentha aquatica methanolic extract, 15 mL/L (1.5g/L) | 10000 |
| F (ME-AQ2) | Infected | Mentha aquatica methanolic extract, 50 mL/L (5g/L) | 10000 |
| G (NINT) | Uninfected | Untreated (blank control) | - |
| H (INT) | Infected | Untreated (negative control) | 10000 |
| I (Toltrazuril) | Infected | Toltrazuril, 7 mg/kg BW for 5 days (positive control) | 10000 |
1 Sporulated oocysts/mL.
Table 4.
Total phenolic and flavonoid content in different plant extracts.
| Samples | Aqueous extracts | Hydromethanolic extracts | ||
|---|---|---|---|---|
| Parameters | TPC1 | TFC1 | TPC1 | TFC1 |
| Rosmarinus officinalis | 160.96 ± 6.55 | 49.26 ± 0.40 | 215.50 ± 17.19 | 33.26 ± 1.40 |
| Thymus algeriensis | 128.11 ± 10.56 | 15.20 ± 2.30 | 220.35 ± 5.92 | 28.05 ± 1.31 |
| Mentha aquatica | 167.139 ± 26.48 | 29.56 ± 2.58 | 199.26 ± 12.26 | 59.77 ± 4.07 |
1 All values are significant at (p<0.05). TPC: Total phenolic content (mg GAE/g DW), TFC: Total flavonoid content (mg QE/g DW).
Table 5.
Antioxidant capacities (IC50 µg/ml) of aqueous and hydromethanolic extracts by CUPRAC and DPPH methods.
Table 5.
Antioxidant capacities (IC50 µg/ml) of aqueous and hydromethanolic extracts by CUPRAC and DPPH methods.
| Extracts | Aqueous extracts* | Hydromethanolic extracts* | ||
|---|---|---|---|---|
| DPPH | CUPRAC | DPPH | CUPRAC | |
| Rosmarinus officinalis | 16.37 ± 0.01 | 18.80 ± 0.02 | 15.74 ± 0.69 | 14.50 ± 0.14 |
| Thymus algeriensis | 17.18 ± 0.09 | 15.65 ± 0.58 | 15.56 ± 0.63 | 11.72 ± 0.85 |
| Mentha aquatica | 12.79 ± 0.05 | 12.06 ± 0.60 | 10.19± 0.51 | 9.47 ± 0.19 |
| BHA | 5.73 ± 0.41 | 3.64 ± 0.19 | 5.73 ± 0.41 | 3.64 ± 0.19 |
| BHT | 22.32 ± 1.19 | 9.62 ± 0.87 | 22.32 ± 1.19 | 9.62 ± 0.87 |
* All values are Significant at (p<0.05). IC50 (µg/mL) indicates the concentration with 50% scavenging intensity.
Table 6.
Anticoccidial activities (IC50 µg/ml) of plant extracts and anticoccidial drugs.
| Plants extracts | Aqueous extracts* | Hydromethanolic extracts* |
|---|---|---|
| Rosmarinus officinalis | 699.11 ± 4.61 | 233.14 ± 3.56 |
| Thymus algeriensis | 3449.56 ± 2.53 | 2621.62 ± 2.90 |
| Mentha aquatica | 329.86 ± 2.78 | 967.11 ± 2.58 |
| Sulfaquinoxalin sodic trimethoprim | NA | |
| Toltrazuril | NA | |
* All values are significant (p< 0.0001).
Table 7.
Linear-regression statistics describing the concentration-dependent fall in Eimeria oocyst viability produced by aqueous (AQ) and hydromethanolic (HM) extracts of Rosmarinus officinalis (RO), Thymus algeriensis (TH) and Mentha aquatica (ME). Ten concentrations (0–20 mg/mL) were tested once each; simple linear fits were generated. DF = 1, 8 for all regressions. Sy.x = standard error of the regression. All slopes differ significantly from zero.
Table 7.
Linear-regression statistics describing the concentration-dependent fall in Eimeria oocyst viability produced by aqueous (AQ) and hydromethanolic (HM) extracts of Rosmarinus officinalis (RO), Thymus algeriensis (TH) and Mentha aquatica (ME). Ten concentrations (0–20 mg/mL) were tested once each; simple linear fits were generated. DF = 1, 8 for all regressions. Sy.x = standard error of the regression. All slopes differ significantly from zero.
| Extract | Slope ± SE (10-³ % mL/mg) | 95 % CI slope | Y-intercept ± SE (%) | R² | Sy.x | Equation | p (slope) |
|---|---|---|---|---|---|---|---|
| RO-AQ | −24.07 ± 4.63 | −34.74 – −13.40 | 80.56 ± 8.35 | 0.772 | 19.03 | y = -4.814301x + 80.56027 | 0.0008 |
| TH-AQ | −14.24 ± 1.73 | −18.22 – −10.26 | 93.82 ± 3.12 | 0.895 | 7.10 | y = -2.848292x + 93.82187 | <0.0001 |
| ME-AQ | −20.11 ± 4.71 | −30.97 – −9.25 | 63.32 ± 8.50 | 0.695 | 19.36 | y = -4.022062x + 63.31536 | 0.0027 |
| RO-HM | −11.83 ± 1.84 | −16.08 – −7.58 | 48.63 ± 3.32 | 0.838 | 7.57 | y = -2.365649x + 48.63178 | 0.0002 |
| TH-HM | −11.70 ± 1.37 | −14.85 – −8.55 | 81.90 ± 2.47 | 0.902 | 5.62 | y = -2.340540x + 81.90387 | <0.0001 |
| ME-HM | −18.64 ± 2.39 | −24.14 – −13.14 | 73.66 ± 4.31 | 0.884 | 9.81 | y = -3.727003x + 73.66176 | <0.0001 |
Table 8.
Growth performance parameters of broiler chickens during the experimental period.
| Period | TH-AQ1 | TH-AQ2 | RO-AQ1 | RO-AQ2 | ME-AQ1 | ME-AQ2 | NINF-NT | INF-NT | TOTRA | |
|---|---|---|---|---|---|---|---|---|---|---|
| BWG | 1d | 44.39 ± 7.60 | 43.37 ±5.51 | 43.38 ± 6.78 | 43.36 ± 7.20 | 43.35 ± 0.07 | 43.38 ± 0.06 | 42.66 ± 2.21 | 43.31 ± 0.58 | 43.14± 0.39 |
| 7dA | 182.72 ± 0.04 | 187.80 ± 6.98 | 183.76 ± 1.36 | 183.39 ± 0.52 | 182.67 ± 0.90 | 182.95 ± 0.09 | 182.68 ± 0.07 | 180.75 ± 6.14 | 182.58± 0.73 | |
| 10dB | 221.15 ± 1.25 | 221.15 ± 1.26 | 221.15 ± 1.27 | 221.15 ± 1.28 | 221.15 ± 1.29 | 221.15 ± 1.30 | 221.15 ± 1.31 | 221.15 ± 1.32 | 221.15± 1.33 | |
| 10d PI | 877.97 ± 3.25a | 788.35 ± 1.70b | 718.20 ± 3.84c | 757.82 ± 1.99d | 641.50 ± 8.00e | 643.38 ± 5.53e | 685.12 ± 1.25f | 448.75 ± 4.18g | 835.74± 1.25h | |
| 20d PI | 1987.74 ± 2.21a | 1777.37 ± 2.17b | 1237.94 ± 3.91c | 1525.97 ± 0.17d | 714.48 ± 2.57e | 658.59 ± 4.34f | 985.43 ± 1.67g | 583.02 ± 3.09h | 1722.06± 1.78i | |
| 32d PI | 2319.55 ± 43.56a | 2182.15 ± 2.52b | 1674.40 ± 0.75c | 1770.24 ± 7.93 | 987.97 ± 12.26e | 757.82 ± 1.99f | 1131.39 ± 5.41g | 902.17 ± 9.12h | 2143.27± 7.64i | |
| TWG | C | 656.82 ± 3.13a | 567.20 ± 1.53b | 497.05 ± 2.62c | 536.68 ± 2.38b | 421.02 ± 7.92d | 421.83 ± 4.62d | 463.98 ± 1.69e | 227.89 ± 2.89f | 614.59 ± 1.39g |
| D | 1109.77 ± 1.18a | 989.02 ± 3.14b | 519.74 ± 1.30c | 768.14 ± 1.59d | 72.32 ± 8.03e | 15.60 ± 7.46f | 300.30 ± 2.16g | 133.98 ± 5.87h | 886.32 ± 1.28i | |
| E | 331.81 ± 37.35b | 404.79 ± 0.91a | 436.46 ± 3.80a | 244.27 ± 6.37c | 273.49 ± 11.28c | 99.24 ± 5.17d | 145.96 ± 4.98e | 319.15 ± 5.36b | 421.20 ± 5.92a | |
| F | 2098.40 ± 35.05a | 1961.01 ± 2.29b | 1453.26 ± 1.57c | 1549.09 ± 7.00d | 766.82 ± 9.20e | 536.68 ± 2.38f | 910.24 ± 5.06g | 681.02 ± 8.46h | 1922.12 ± 7.09i | |
| ADG | C | 65.68 ± 0.31a | 56.72 ± 0.15b | 49.71 ± 0.26c | 53.67 ± 0.24d | 42.10 ± 0.79e | 42.18 ± 0.46e | 46.40 ± 0.17f | 22.79 ± 0.29g | 61.46± 0.14h |
| D | 110.98 ± 0.12a | 98.90 ± 0.31b | 51.97 ± 0.13c | 76.81 ± 0.16d | 7.23 ± 0.80e | 1.56 ± 0.75f | 30.03 ± 0.22g | 13.40 ± 0.59h | 88.63± 0.13i | |
| E | 27.65 ± 3.11a | 33.73 ± 0.08b | 36.37 ± 0.32b | 20.36 ± 0.53c | 22.79 ± 0.94d | 8.27 ± 0.43e | 12.16 ± 0.41f | 26.60 ± 0.45a | 35.10± 0.49b | |
| F | 65.58 ± 1.10b | 61.28 ± 0.07a | 45.41 ± 0.05c | 48.41 ± 0.22d | 23.96 ± 0.29e | 16.77 ± 0.07f | 28.45 ± 0.16g | 21.28 ± 0.26h | 60.07± 0.22a | |
| FI | C | 826.63 ± 15.39a | 901.86 ± 6.80a | 606.37 ± 3.87b | 658.34 ± 12.16b | 362.13 ± 11.35c | 293.58 ± 1.75d | 685.33 ± 16.15b | 130.37± 2.72e | 885.01± 5.16a |
| D | 1489.30 ± 12.76a | 1549.49 ± 19.63a | 751.89 ± 0.61c | 979.64 ± 11.49d | 69.38 ± 7.54b | 10.96 ± 5.24b | 489.49 ± 3.71e | 91.24± 4.00b | 1503.80± 5.65a | |
| E | 528.52 ± 58.92c | 659.76 ± 18.69a | 704.17 ± 7.80a | 354.97 ± 20.60d | 269.48 ± 11.24b | 98.05 ± 5.82e | 256.62 ± 14.76b | 260.75± 4.35b | 699.19± 20.46a | |
| F | 2933.52 ± 60.22d | 3128.91 ± 44.48a | 2073.32 ± 9.08b | 2041.83 ± 38.04b | 717.00 ± 10.86e | 426.78 ± 3.69c | 1475.83 ± 30.85f | 469.90± 5.38c | 3073.28± 30.07a | |
| FCR | C | 1.26 ± 0.03a | 1.59 ± 0.01c | 1.22 ± 0.01a | 1.23 ± 0.02a | 0.86 ± 0.02d | 0.70 ± 0.00e | 1.48 ± 0.03b | 0.57± 0.01f | 1.44± 0.01b |
| D | 1.34 ± 0.01b | 1.57 ± 0.02c | 1.45 ± 0.00d | 1.28 ± 0.01e | 0.96 ± 0.00f | 0.70 ± 0.00a | 1.63 ± 0.01g | 0.68± 0.00a | 1.70± 0.00h | |
| E | 1.59 ± 0.00a | 1.63 ± 0.05a | 1.61 ± 0.01a | 1.45 ± 0.05c | 0.99 ± 0.00b | 0.99 ± 0.01b | 1.76 ± 0.04d | 0.82± 0.00e | 1.66± 0.04f | |
| F | 1.40 ± 0.01a | 1.60 ± 0.02b | 1.43 ± 0.00a | 1.32 ± 0.02c | 0.94 ± 0.01d | 0.80 ± 0.00e | 1.62 ± 0.03b | 0.69± 0.00f | 1.60± 0.01b | |
| EPEF | - | 395.08 ± 6.06a | 325.63 ± 0.31b | 279.44 ± 0.10b | 319.79 ± 1.17c | 251.58 ± 2.55d | 226.90 ± 0.49e | 166.16 ± 0.65f | 311.31 ± 2.57g | 319.16 ± 0.93g |
| EBI | - | 469.10 ± 7.84a | 384.08 ± 0.45c | 318.32 ± 0.34b | 367.29 ± 1.66c | 179.40 ± 2.15d | 189.81 ± 0.84d | 175.45 ± 0.98d | 277.59 ± 3.45e | 375.68 ± 1.39c |
Values in a row with different letters are significantly different at P < 0.05. Groups sharing the same letter are not significantly different. A: 3d before parasite inoculation; B: Inoculation Day of the parasite; C: 0–10 d PI; D: 11–20 d PI; E: 21–32 d PI; F: 0-32 d PI.
Table 9.
Survival Rate (SR), Relative Weight Gain (RWG), Oocyst Shedding, Lesion Severity, and Anticoccidial Index (ACI) in broiler chickens experimentally infected with Eimeria spp. and treated with medicinal plant extracts or toltrazuril: comparison between NINF-NT (Non-Infected, Non-Treated), INF-NT (Infected, Non-Treated), TH-AQ1/2 (Thymus algeriensis aqueous extract, low/high dose), RO-AQ1/2 (Rosmarinus officinalis aqueous extract, low/high dose), ME-AQ1/2 (Mentha aquatica aqueous extract, low/high dose), and TOTRA (Toltrazuril-treated).
Table 9.
Survival Rate (SR), Relative Weight Gain (RWG), Oocyst Shedding, Lesion Severity, and Anticoccidial Index (ACI) in broiler chickens experimentally infected with Eimeria spp. and treated with medicinal plant extracts or toltrazuril: comparison between NINF-NT (Non-Infected, Non-Treated), INF-NT (Infected, Non-Treated), TH-AQ1/2 (Thymus algeriensis aqueous extract, low/high dose), RO-AQ1/2 (Rosmarinus officinalis aqueous extract, low/high dose), ME-AQ1/2 (Mentha aquatica aqueous extract, low/high dose), and TOTRA (Toltrazuril-treated).
| Group | SR (%) | RWG rate (%) | Oocyst number x104 | Oocyst value | Lesion score | Lesion value | ACI |
|---|---|---|---|---|---|---|---|
| NINF-NT | 100 | 100 | 0 | - | - | - | - |
| INF-NT | 90 | 74.818 | 10.639 | 100 | 2.85± 0.57 | 28.50± 5.71 | - |
| TH-AQ1 | 100 | 230.532 | 1.917 | 18.016 | 1.43± 0.52 | 14.33± 5.23 | 298.183 |
| TH-AQ2 | 100 | 287.951 | 3.722 | 34.987 | 1.08± 0.22 | 10.83± 2.24 | 342.130 |
| RO-AQ1 | 100 | 75.607 | 8.500 | 79.896 | 1.17± 0.73 | 11.67± 7.32 | 83.711 |
| RO-AQ2 | 100 | 170.184 | 5.111 | 48.042 | 2.18± 0.33 | 21.83± 3.31 | 200.309 |
| ME-AQ1 | 70 | 84.243 | 5.917 | 55.614 | 2.50± 0.00 | 25.00± 0.00 | 73.630 |
| ME-AQ2 | 90 | 58.960 | 7.139 | 67.102 | 2.65± 0.56 | 26.50± 5.61 | 55.358 |
| TOTRA | 100 | 211.166 | 3.194 | 30.026 | 1.75± 0.62 | 17.50± 6.21 | 263.639 |
Table 10.
Mean intestinal lesion scores and key microscopic findings for each treatment group. Scores are Johnson & Reid composites (duodenum, jejunum, ileum, caeca). The table also lists the segment most affected, provides a concise histological summary, indicates the presence or absence of parasite stages, and offers an interpretation in the context of the mixed Eimeria challenge.
Table 10.
Mean intestinal lesion scores and key microscopic findings for each treatment group. Scores are Johnson & Reid composites (duodenum, jejunum, ileum, caeca). The table also lists the segment most affected, provides a concise histological summary, indicates the presence or absence of parasite stages, and offers an interpretation in the context of the mixed Eimeria challenge.
| Group | Mean composite lesion score ± SD * | Segment(s) most affected † | Concise histological description | Parasite stages seen | Interpretation vs. mixed Eimeria challenge ‡ |
|---|---|---|---|---|---|
| NINF-NT | 0.00 ± 0.00 | – | Long slender villi, orderly crypts, intact epithelium, minimal lamina propria cells. | None | True physiological baseline. Confirms sampling artefacts are negligible. |
| INF-NT | 2.85 ± 0.57 | Duo > Jej ≈ Ile > Cae | Massive villus loss, dilated crypts packed with necrotic debris, extensive coagulative necrosis, dense heterophil infiltrate. | Numerous schizonts, gamonts, and oocysts | Unrestricted replication of all challenge species: E. praecox (duodenum), E. maxima (jejunum), E. necatrix (mid-gut), E. tenella (caeca) etc. |
| TOTRA | 1.75 ± 0.62 | Jej > Duo | Slight villus blunting, crypts intact, continuous epithelium, sparse mononuclear infiltrate; parasites absent. | None | Near-complete protection, matching the broad anti-Eimeria spectrum of toltrazuril. |
| TH-AQ1 | 1.43 ± 0.52 | Duodenum | Largely intact architecture, mild lamina-propria infiltrate, very occasional vacuolated parasite residua. | Very rare | Best-performing plant extract, strong suppression of E. praecox/maxima and reasonable control of E. tenella. |
| TH-AQ2 | 1.08 ± 0.22 | Duo ≈ Jej | Blunted villi, moderate crypt hyperplasia, light mixed infiltrate, parasites extremely sparse. | Sporadic | Higher dose offers no meaningful histological gain over TH-AQ1—efficacy already near plateau. |
| RO-AQ1 | 1.17 ± 0.73 | Duo ≈ Jej | Marked villus loss, crypt destruction, focal necrosis, dense inflammation; many gamonts/oocysts. | Abundant gamonts and oocysts | Incomplete protection: only moderate impact on E. praecox/maxima; caeca relatively spared. |
| RO-AQ2 | 2.18 ± 0.33 | Duo > Jej ≈ Ile | Patchy villus absence, crypt hyperplasia, focal fibrin, parasites fewer than RO-AQ1. | Scattered gamonts | A higher dose lowers the parasite load somewhat, yet leaves moderate lesions, resulting in intermediate efficacy. |
| ME-AQ1 | 2.50 ± 0.00 | All segments (even) | Carpets of necrotic exfoliated enterocytes, partial crypt preservation, diffuse inflammation, and scattered parasites. | Moderate gamonts/oocysts | Variable, only partial protection, limited activity versus E. maxima and E. brunetti. |
| ME-AQ2 | 2.65 ± 0.56 | All, esp. Jej/Ile | Villus obliteration, “honey-comb” hyperplastic crypts, fibrino-purulent exudate, oocysts still numerous. | Numerous macrogamonts/oocysts | A higher dose fails to improve outcome; lesions approach INF-NT severity, explaining the poor ACI. |
* Composite average of Johnson & Reid lesion scores for duodenum, jejunum, ileum and caeca, derived from the spreadsheet you supplied. † Rank order of median segmental scores (Duo = duodenum, Jej = jejunum, Ile = ileum, Cae = caeca). ‡ Lesion distribution reflects the known tropism of species in the oral inoculum (37 % Eimeria maxima, 19 % Eimeria brunetti, 16 % Eimeria necatrix, 14 % Eimeria praecox, 8 % Eimeria tenella, 6 % Eimeria mitis).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



