Submitted:
03 July 2025
Posted:
04 July 2025
You are already at the latest version
Abstract
Off-season flowering in Macadamia integrifolia has been observed in high-altitude regions, but the hormonal regulatory mechanisms underlying this phenomenon remain unclear. In this study, the annual dynamics of four key endogenous phytohormones (ZT, GA₃, IAA, and ABA) were quantified in the leaves and branches of trees from both normal and off-season flowering sites using high-performance liquid chromatography (HPLC).Hormonal ratios and correlation network analyses were further performed to investigate their roles in flowering regulation. Floral bud differentiation at the off-season site occurred approximately 1 to 2 months earlier than at the normal site. This advancement was associated with sustained low levels of GA₃ (below 100 μg·g⁻¹ FW), while ZT and ABA levels peaked in September at 108.66 μg·g⁻¹ FW and 24.25 μg·g⁻¹ FW, respectively. The ratios of ABA to GA₃, ABA to IAA, and ZT to GA₃ increased significantly between July and September, indicating the early establishment of a hormonal environment favorable for floral induction. Correlation analysis revealed that IAA, GA₃, and ZT formed a synergistic module promoting flowering, whereas ABA functioned as an antagonistic regulator. These findings provide insight into the hormonal regulation of off-season flowering in macadamia and offer a theoretical basis for precision flowering control in high-altitude cultivation systems.
Keywords:
macadamia
; off-season flowering
; endogenous hormones
; hormone ratio
; floral induction
; phenological regulation
1. Introduction
Macadamia (Macadamia spp.) is an evergreen tree species indigenous to the subtropical rainforests of southeastern Queensland and northeastern New South Wales, Australia [Error! Reference source not found.,Error! Reference source not found.]. Macadamia nuts are rich in unsaturated fatty acids, proteins, amino acids, and various vitamins, rendering them highly valuable for both the food processing and cosmetics industries [Error! Reference source not found.]. In recent years, China has emerged as the world’s leading producer of macadamia, with Yunnan Province alone accounting for nearly 50% of the global cultivation area [4]. However, the rapid expansion of macadamia plantations has been accompanied by an increasing prevalence of off-season flowering in certain production regions. This anomalous phenological behavior disrupts the synchrony of pollination and fertilization, leading to reduced fruit set, compromised fruit quality, and yield fluctuations ranging from 20% to 30% [Error! Reference source not found.]. Such disturbances not only threaten the stability of the supply chain but also pose ecological risks by disrupting the temporal alignment between Macadamia phenology and pollinator dynamics [Error! Reference source not found.]. Moreover, off-season flowering results in asynchronous fruit development within individual trees, thereby complicating harvesting operations and elevating labor costs. When fruits are harvested in a single batch, overall quality often deteriorates, further constraining the sustainable development of the macadamia industry [7].
The flowering process in plants is regulated by a complex interplay of internal and external factors, including photoperiod, temperature, water availability, nutrient status, and endogenous phytohormones [Error! Reference source not found.]. Among these, endogenous hormones function as central integrators, transducing environmental cues into developmental responses. They play critical roles in floral bud differentiation, dormancy release, and the initiation of flowering [Error! Reference source not found.]. The principal classes of phytohormones involved in these processes include gibberellins (GAs), auxins (IAA), cytokinins (CTKs), and abscisic acid (ABA) [10]. Gibberellins are particularly notable for their involvement in multiple regulatory pathways that promote the transition from vegetative to reproductive growth. However, in macadamia, GAs may inhibit floral bud differentiation [Error! Reference source not found.,Error! Reference source not found.], highlighting the species-specific complexity and context-dependent nature of hormonal regulation.
IAA is essential for floral primordium initiation, with auxin deficiency known to impair flower formation. CTKs modulate floral meristem activity by regulating cell division and differentiation [Error! Reference source not found.,13]. ABA, widely recognized for its role in abiotic stress responses, is generally considered a floral repressor. Elevated ABA levels are frequently associated with delayed floral bud development and reduced flowering potential [14]. Emerging evidence indicates that flowering regulation by endogenous hormones is not solely dictated by the absolute concentrations of individual hormones, but rather by the dynamic balance and interaction among them. Hormonal ratios, such as ZT/GA₃, ZT/IAA, ABA/IAA, and ABA/GA₃, have been shown to play decisive roles in determining floral developmental outcomes [15,16]. Higher values of ZT/GA₃, ZT/IAA, ABA/IAA, and ABA/GA₃ ratios are typically associated with the maintenance of buds in a vegetative state, whereas lower values are more conducive to the transition toward reproductive development [17]. Therefore, elucidating the hormonal regulation mechanisms underlying off-season flowering in Macadamia is essential for understanding its phenological plasticity and for developing effective strategies to mitigate production instability. To this end, this study explores hormonal signals underlying off-season flowering in high-altitude Macadamia by comparing hormone dynamics and ratios in contrasting phenological regions over a 12-month period, aiming to inform flowering regulation strategies.
Figure 1.
Off-season flowering in Macadamia, with the co-occurrence of developing.

Fruits and floral inflorescences on the same tree.
2. Results
2.1. Phenological Observation of Macadamia
A systematic observation and comparison of the annual timing of phenological stages were conducted at the two study sites. The results revealed significant differences between the MH and JH sites in the timing of key developmental stages, including floral bud differentiation, flowering, fruit development, and maturation (Table 1). At the MH site, floral bud differentiation primarily occurs from August to September. Initial flowering begins in October and November, with peak flowering extending into January of the following year. Fruit enters the enlargement stage from February to April, followed by oil accumulation from May to July, reaching near maturity by August. In contrast, all corresponding phenophases at the JH site are delayed by approximately 1 to 2 months. Floral bud differentiation occurs from October to December, initial flowering shifts to January–February of the following year, peak flowering occurs in March, fruit enlargement and oil accumulation take place from April–May and June–August, respectively, and maturation and harvest begin in September.
2.2. Dynamics of Endogenous Hormones in Macadamia
2.2.1. Dynamic Changes of Endogenous Hormones in Leaves
The endogenous hormone contents in macadamia leaves were measured monthly from January to December, revealing the dynamic changes over the course of the year (Figure 2).
Zeatin (ZT) content in macadamia leaves exhibited pronounced seasonal variation across both study sites. At the MH site, ZT levels remained consistently low and stable from January to August, followed by a sharp increase beginning in late August, peaking in September at 105.28 µg·g⁻¹ FW. A marked decline occurred in October, with a slight rebound from November to December, maintaining a moderately low level overall. At the JH site, ZT content was similarly low from January to June, then increased sharply in July to 110.32 µg·g⁻¹ FW. After a minor dip in August, ZT levels rose again in September to reach the annual maximum (123.56 µg·g⁻¹ FW), followed by a notable decline in October and a modest recovery during November and December.
Gibberellic acid (GA₃) concentrations showed distinct seasonal dynamics between the two sites. At the MH site, GA₃ content increased steadily from January to March, reaching an early-year peak in March (839.08 µg·g⁻¹ FW). This was followed by a sharp decline in April, and relatively low and stable levels were maintained from April through July. A secondary rise occurred in August and September, followed by a decline in October, a slight rebound in November, and a modest decrease in December. At the JH site, the early-year trend (January to March) was similar, with a peak in March. However, GA₃ levels rose again significantly in May before gradually decreasing to the annual minimum in August. A strong rebound was observed in September, resulting in the highest yearly peak (824.31 µg·g⁻¹ FW), after which values declined rapidly to early-year levels.
Indole-3-acetic acid (IAA) levels at the MH site displayed a “three peaks and three troughs” pattern throughout the year. Concentrations were moderately high from January to February, followed by a sharp drop in March to the first trough. A rapid rebound occurred in April, followed by a slight decline in May, a rise in June, and the highest annual peak in July (464.27 µg·g⁻¹ FW). IAA levels remained relatively elevated through September, then dropped sharply in October to the lowest point of the year (5.25 µg·g⁻¹ FW). A significant increase occurred in November, followed by a minor decline in December. At the JH site, IAA content was also relatively high in January and February but decreased abruptly to 15.40 µg·g⁻¹ FW in March. From April to August, levels remained relatively stable and elevated, followed by a sharp decline from September to October, reaching a second trough in October (1.35 µg·g⁻¹ FW). IAA content then rose sharply in November and stabilized at a moderate level in December.
Abscisic acid (ABA) concentrations remained relatively low throughout the observation period compared with other hormones. At the MH site, two distinct peaks were observed in September and November, with maximum values of 24.25 and 20.27 µg·g⁻¹ FW, respectively. In other months, ABA content was consistently low and showed minimal fluctuation. At the JH site, ABA variation was even more constrained, with peak values lower than those at the MH site. The highest annual concentration occurred in November (9.34 µg·g⁻¹ FW), and levels remained consistently lower than those at MH during most of the year.
2.2.2. Dynamic Changes of Endogenous Hormones in Branches
The annual variation in the concentrations of four endogenous hormones, including zeatin (ZT), gibberellic acid (GA₃), indole-3-acetic acid (IAA), and abscisic acid (ABA), in macadamia branches is illustrated in Figure 3, and the corresponding temporal patterns are described below.
Zeatin (ZT) content in macadamia branches exhibited similar bimodal seasonal trends at both sites, though with notable temporal variation. At the MH site, ZT levels remained low from January to May, followed by a sharp increase beginning in June, reaching the first peak in July (74.70 μg·g⁻¹ FW). After a brief decline, levels rose again to the annual maximum in September (108.66 μg·g⁻¹ FW), and then dropped rapidly from October to December. The JH site showed a comparable pattern with two pronounced peaks also in July and September. However, JH exhibited a small additional peak in March (51.26 μg·g⁻¹ FW), not observed at the MH site. Moreover, the increase in ZT content during June was less marked at JH than at MH, indicating possible phase differences in ZT regulation between the two sites.
Gibberellic acid (GA₃) levels in branches demonstrated more pronounced pulse-like fluctuations at the MH site than at JH. At MH, GA₃ content remained low in January and February, followed by a sharp surge in March that resulted in the annual maximum (238.22 μg·g⁻¹ FW). Levels then declined rapidly and remained low from April to August, with a minimum in July (3.02 μg·g⁻¹ FW). A second peak was observed in September (145.62 μg·g⁻¹ FW), followed by a decline in October and a minor rebound in November and December. In contrast, GA₃ levels at JH fluctuated more smoothly. After low levels in January and February, a modest rise in March was followed by the annual peak in May (211.01 μg·g⁻¹ FW). Levels then declined until August, increased again in September (217.62 μg·g⁻¹ FW), and dropped in October, stabilizing in the final two months of the year. Notably, the first peak at JH occurred approximately two months later than at MH.
Indole-3-acetic acid (IAA) in branches displayed a distinct seasonal rhythm with similar trends at both sites. At MH, IAA content was low in January but rose sharply in February, peaking in March at 816.57 μg·g⁻¹ FW, which represented the highest annual value. Levels then declined rapidly and remained relatively stable from April to June. A secondary moderate peak occurred in July (415.47 μg·g⁻¹ FW), followed by a decline to the lowest annual value in October (0.75 μg·g⁻¹ FW). Slight rebounds occurred in November, and a small decrease followed in December. At the JH site, the pattern was comparable but with slightly lower amplitudes. IAA content gradually increased from January to March (peak: 791.78 μg·g⁻¹ FW), then declined sharply through June. A minor rise was observed in July, followed by a steady decrease until October (minimum: 0.35 μg·g⁻¹ FW), with a moderate rebound in November and slight decline in December.
Abscisic acid (ABA) content in branches varied seasonally and showed contrasting peak timings between sites. At the MH site, ABA levels remained low during January and February, then rose sharply in March, forming the first major peak. Levels declined substantially from April to July, with the lowest value recorded in July. A notable increase in August led to a second peak in September, followed by a slight drop in October and another rise in December, forming the third-highest annual value. At the JH site, the pattern was somewhat different: ABA content also remained low from January to April (minimum in April), then began rising in July and continued increasing steadily, reaching the annual maximum in November (35.83 μg·g⁻¹ FW), followed by a sharp decline in December. Overall, the timing of peak ABA levels differed between sites, with the MH site peaking in September and the JH site in November, and an opposing trend observed during July.
2.3. Dynamic Changes in the Endogenous Hormone Balance Ratios in Macadamia
To further assess the regulatory role of hormonal interactions, a heat map was used to visualize the relative monthly changes in four key hormone ratios: ABA/GA₃, ZT/GA₃, ABA/IAA, and ZT/IAA. The color gradient ranged from green to purplish-red, indicating a transition from low to high ratio values. The results (Figure 4) demonstrated significant spatiotemporal variation in hormonal balance across sites and tissue types.
At the MH site (Figure 4a), hormone ratios in both leaves (L) and branches (B) exhibited pronounced seasonal dynamics. In branches, the ABA/IAA and ZT/IAA ratios showed intensified coloration in July and August, indicating significant increases and annual peaks during this period. Additionally, the ZT/GA₃ ratio also increased markedly in July. In leaves, ZT/IAA and ABA/IAA displayed slight increases between April and June, whereas ABA/GA₃ and ZT/GA₃ remained relatively stable throughout the year, with only modest elevations from June to July. Overall, hormone ratio peaks in branches were concentrated in mid-summer, while fluctuations in leaves were more moderate and sustained.
At the JH site (Figure 4b), seasonal dynamics of hormone ratios showed clear differences from those at MH. In branches, ABA/IAA and ZT/IAA ratios increased markedly in October, reaching their highest annual values. By contrast, ZT/GA₃ and ABA/GA₃ ratios exhibited only minor fluctuations across the year, with relatively uniform coloration and small increases in July and November. These patterns suggest a generally stable hormonal balance in branches, with no distinct high-ratio periods. In leaves, fluctuations were minimal; ZT/IAA showed a slight increase and ABA/IAA a marginal rise in October, while changes in other months were negligible. The ZT/GA₃ and ABA/GA₃ ratios in leaves were marked by lighter coloration in July and August, and ABA/GA₃ maintained a relatively uniform distribution throughout the year, indicating minimal seasonal variation.
2.4. Correlation Analysis of Endogenous Hormones and Their Ratios in Macadamia
To elucidate the interrelationships among endogenous hormones and their ratios in Macadamia, Spearman correlation analysis was performed on hormone concentrations from leaves and branches at the two sampling sites (MH and JH) (Figure 5). The results revealed notable differences in correlation patterns between the two sites.
At the MH site (Figure 5a), a significant positive correlation was observed between ABA and GA₃ in leaves. GA₃ levels in branches were significantly positively correlated with ZT in leaves, and GA₃ also showed positive correlations with ABA in both leaves and branches. The leaf ZT/GA₃ ratio was significantly negatively correlated with leaf GA₃, but positively correlated with branch ZT. The leaf ABA/IAA ratio showed a negative correlation with leaf IAA, while exhibiting a highly significant positive correlation with branch GA₃ and a significant positive correlation with branch ABA. The branch ABA/GA₃ ratio was significantly negatively correlated with branch GA₃ and highly significantly negatively correlated with the leaf ZT/IAA ratio. The branch ZT/GA₃ ratio was significantly negatively correlated with leaf GA₃, branch GA₃, branch ABA, and the leaf ABA/IAA ratio, while being significantly positively correlated with leaf IAA and branch ZT. A significant positive correlation was also found between the branch ABA/IAA ratio and the ABA/GA₃ ratio.
At the JH site (Figure 5b), ZT levels in leaves and branches showed a significant positive correlation. The leaf ABA/GA₃ ratio exhibited a highly significant negative correlation with leaf GA₃ and a significant negative correlation with branch GA₃, but was positively correlated with leaf IAA. The leaf ZT/GA₃ ratio was significantly positively correlated with both leaf ZT and the leaf ABA/GA₃ ratio, and significantly negatively correlated with leaf GA₃. A highly significant positive correlation was found between the leaf ABA/IAA ratio and branch ABA. The branch ABA/GA₃ ratio was significantly positively correlated with leaf ABA and the branch ABA/IAA ratio, and exhibited a highly significant positive correlation with branch ABA levels, while being significantly negatively correlated with branch GA₃. The branch ZT/GA₃ ratio showed significant or highly significant positive correlations with the leaf ABA/IAA ratio and the branch ABA/GA₃ ratio. The branch ABA/IAA ratio exhibited significant or highly significant positive correlations with branch ABA, the ABA/GA₃ ratio, and the leaf ABA/IAA ratio, while showing a significant negative correlation with branch IAA. Similarly, the branch ZT/IAA ratio was highly significantly negatively correlated with branch IAA.
In summary, the two sites exhibited distinct hormone regulatory patterns. At the MH site, coordinated interactions between ABA and GA₃ formed the core of the regulatory network, characterized by significant cross-organ correlations between leaves and branches. In contrast, hormonal regulation at the JH site was primarily governed by ABA-dominated ratios, with significant correlations predominantly confined within the branches. These findings suggest that Macadamia trees exhibit site-specific hormonal response patterns, reflecting distinct regulatory adaptations to contrasting ecological environments.
3. Discussion
3.1. Relationship Between Endogenous Hormone Dynamics and Off-Season Flowering in Macadamia
Flowering in plants is a complex physiological process that is tightly regulated by the interplay of genetic background, environmental conditions, and endogenous signals, among which plant hormones play a central role during key developmental stages such as floral induction, dormancy release, reproductive transition, and floral organ formation [Error! Reference source not found.]. In this study, the temporal dynamics and potential regulatory roles of zeatin (ZT), gibberellic acid (GA₃), indole-3-acetic acid (IAA), and abscisic acid (ABA) were systematically investigated based on twelve months of continuous monitoring at two sampling sites (MH and JH). Cytokinins, particularly ZT, are critical regulators of floral bud initiation and the transition from vegetative to reproductive growth in perennial woody species. These effects are primarily mediated through the promotion of cell division, modulation of source–sink relationships, and optimization of nutrient allocation, thereby facilitating reproductive switching [19-Error! Reference source not found.]. In the present study, ZT levels in branches at both MH and JH exhibited a bimodal distribution from July to September, although the timing and magnitude of the peaks differed significantly between the two sites. At MH, ZT levels began to increase in July and peaked in September, closely corresponding with elevated ZT concentrations in leaves and the period of floral bud sprouting. These findings suggest that ZT may facilitate off-season floral induction by enhancing floral initiation signals and regulating carbon allocation. This observation implies that, even before seasonal cues have fully subsided, ZT-mediated floral initiation pathways are prematurely activated at MH, thereby triggering atypical reproductive development. In contrast, JH not only exhibited two major peaks of ZT but also showed a minor peak in March, which coincided with the expected seasonal floral initiation period. This suggests that ZT is involved not only in floral induction but also in subsequent stages such as floral organ development and anthesis [22,Error! Reference source not found.]. Previous studies in perennial fruit trees, including apple and lychee, have demonstrated that a rapid increase in ZT prior to floral differentiation can activate key transcription factors and hormonal signaling pathways that enhance flowering potential [23]. Moreover, ZT operates within a synergistic regulatory network alongside auxin and gibberellins [25,26]. The advanced timing and sharpened peak of ZT signals observed at MH likely reflect region-specific ecological modulation of ZT biosynthesis, transport, and perception, ultimately contributing to off-season flowering.
Gibberellins (GA₃) play a pivotal role in regulating flowering time and floral bud development. Moderate GA₃ levels promote floral induction, whereas excessive concentrations often suppress flowering, delay reproductive transition, and promote sustained vegetative growth [18,Error! Reference source not found.]. In the present study, GA₃ levels in MH leaves increased rapidly during the early stages, followed by a sharp decline and a prolonged low phase. In contrast, GA₃ concentrations at JH remained generally higher and more stable, with the onset of the low-GA₃ phase occurring later than at MH. Previous studies have shown that GA₃ can accelerate reproductive transition by upregulating flowering-related genes such as SOC1 and LFY [28,Error! Reference source not found.]. In Macadamia, low GA₃ levels have been reported to facilitate floral induction [Error! Reference source not found.]. Therefore, site-specific differences in the timing of the low-GA₃ threshold may be a critical factor contributing to off-season flowering.
Auxin (IAA), a hormone essential for vegetative development, is typically associated with cell elongation and the differentiation of vegetative organs [Error! Reference source not found.]. A progressive reduction in IAA concentration is believed to relieve its inhibitory effect on the reproductive transition, thereby promoting floral induction [32]. In this study, IAA levels in MH leaves exhibited pronounced troughs in March, May, and October, while branch IAA concentrations peaked in March. A similar pattern was observed at JH, with leaf IAA declining in March and October and a corresponding peak in branch IAA in March. This inverse trend between organs during the same period may reflect a regulatory mechanism governing IAA transport and spatial distribution during specific developmental windows [33]. Elevated IAA accumulation in branches during early floral induction may function as a localized signal that promotes floral meristem initiation or reprogramming of vegetative tissues [34]. Such spatiotemporal asynchrony in IAA distribution may play a crucial role in hormonal regulation underlying off-season flowering. In comparison, IAA levels at JH remained relatively stable throughout the year, suggesting a more synchronized and consistent flowering rhythm.
Abscisic acid (ABA), in addition to its well-established role in abiotic stress responses, has also been implicated in floral transition through the regulation of downstream targets such as FT and SOC1. High ABA accumulation during floral bud differentiation has been shown to promote the formation of floral primordia [34,35]. In this study, peak ABA levels at MH occurred approximately two months earlier than those at JH, suggesting that early activation of ABA signaling may play a significant role in promoting off-season floral induction. Moreover, the timing of ABA peaks closely matched the observed period of off-season flowering, indicating that ABA may not only serve as a floral activator but also exert a biphasic regulatory effect, facilitating both the initiation and sustained development of floral buds. In contrast, the delayed and weaker ABA signaling observed at JH may represent an intrinsic physiological constraint that suppresses off-season flowering at this site.
In summary, the occurrence of off-season precocious flowering in macadamia is not driven by a single hormone, but rather results from the coordinated regulation of a multi-hormone signaling network composed of IAA, GA₃, ZT, and ABA, acting in a temporally and spatially integrated manner. The pronounced ZT peaks, reduction in GA₃ levels, periodic decline in IAA, and the earlier peak of ABA collectively form a hormonal profile conducive to disrupting conventional phenological rhythms and promoting the differentiation of off-season floral buds. These hormones act in a complementary fashion across different developmental stages, and their interaction-coupled with a compensatory mechanism involving multiple waves of floral bud differentiation-shapes the regional variations in flowering timing and rhythm in macadamia. This finding provides critical insights into the endogenous regulatory basis underlying off-season flowering in this species.
3.2. Relationship between Endogenous Hormone Balance Ratios in Macadamia and Off-Season Flowering
In plant development, while the absolute concentrations of individual hormones provide essential physiological information, the relative ratios among different hormone types often offer a more accurate indication of signaling strength and the direction of regulatory processes [36,37]. This study examined the seasonal dynamics of key hormonal ratios, specifically ABA/GA₃, ABA/IAA, ZT/GA₃, and ZT/IAA, and their relationship with off-season flowering. The results revealed that at the off-season flowering site (MH), these ratios in shoot tissues exhibited a marked increase from July to September, immediately preceding floral bud differentiation. This temporal pattern suggests the establishment of a hormonal configuration favorable to the initiation of reproductive development.
Further analysis showed that the IAA/ABA and GA₃/ABA ratios at the MH site exhibited more pronounced temporal fluctuations, highlighting their potential importance in regulating the onset of off-season floral induction. Specifically, the IAA/ABA ratio reached its peak in August, closely coinciding with the seasonal maximum of IAA. This pattern implies that the ratio may serve as a potential regulatory switch that facilitates dormancy release and activates pathways involved in floral differentiation. At the same time, the GA₃/ABA ratio exhibited both an earlier rise and greater amplitude, suggesting a priming effect during the initial phase of floral bud induction and contributing to the physiological foundation for off-season flowering. Similar regulatory patterns have been reported in other fruit trees, such as apple (Malus domestica) and citrus (Citrus spp.), where a sharp increase in the IAA/ABA ratio prior to floral differentiation alleviates ABA-mediated repression and promotes the transition to reproductive growth [38,39].
The elevation of these hormone ratios not only reflects a shift toward promotive hormonal dominance but also demonstrates the plant’s ability to reconfigure its internal physiological state through the integration of its hormonal regulatory network. This type of ratio-based regulation enhances the temporal coordination and systemic alignment of hormonal signals, thereby strengthening the plant’s adaptive capacity to cope with environmental challenges such as seasonal mismatches and resource variability [40-Error! Reference source not found.]. Accordingly, the intrinsic mechanism underlying off-season flowering in Macadamia may be attributed to the synergistic regulation of specific hormone ratios during key developmental periods. This mechanism offers an effective physiological compensation strategy when normal seasonal rhythms are disrupted.
3.3. Synergistic Effects of Endogenous Hormones in Macadamia and the Mechanism of Flowering Regulation
Multiple endogenous plant hormones interact through complex signaling crosstalk and integrated regulatory networks to coordinate the transition from vegetative to reproductive development [43]. In this study, correlation network analysis of hormone concentrations revealed significant positive associations among IAA, GA₃, and ZT. This pattern suggests the presence of a synergistic hormone module that promotes flowering by jointly facilitating the activation of reproductive developmental pathways during floral bud induction. In contrast, ABA exhibited significant negative correlations with these promotive hormones and showed relatively low network connectivity, indicating its role as a limiting or inhibitory signal that constrains excessive activation within the hormonal regulatory system.
Comparative analysis of hormone interaction networks between sites further revealed that the off-season flowering site (MH) exhibited higher network density and stronger hormonal synergism. In this network, GA₃ and IAA occupied central and highly connected positions, which underscores their pivotal roles in signal integration[44,45]. As a key hormone promoting flowering, GA₃ not only upregulates the expression of genes involved in reproductive development but also accelerates floral bud differentiation and organogenesis by activating essential floral regulators such as LEAFY and SOC1 [Error! Reference source not found.]. IAA likely contributes to floral morphogenesis by regulating cell division and elongation through signaling crosstalk with GA₃ and synergistic interactions with ZT [46].
Although ZT did not exhibit marked fluctuations in absolute concentration, its strong positive correlations with both IAA and GA₃ suggest a functional role as a signal tuning factor that contributes to the stability and robustness of the flower-inducing hormonal network. ABA, as a classical antagonistic hormone, maintained a negative correlation structure, which supports its role in fine-tuning floral induction by modulating the timing and amplitude of hormonal activation. Collectively, these results support a synergistic and antagonistic regulatory framework in which hormones achieve functional balance and signaling specificity through dynamic network coordination. The coordinated upregulation of IAA, GA₃, and ZT not only counteracts ABA-mediated repression but also establishes an internal hormonal environment that favors floral bud initiation. This composite regulatory mechanism challenges the conventional model of single-hormone dominance and instead emphasizes the highly integrated nature of hormonal network regulation in plants. These findings are consistent with hormone interaction patterns reported in both model and crop species, including Arabidopsis thaliana, Oryza sativa (rice), and Vitis vinifera (grapevine) [47,48,49].
4. Materials and Methods
4.1. Site Description
Two field sites were selected for this study: an experimental site (MH) exhibiting off-season flowering, and a control site (JH) with typical flowering phenology. The MH site is located in a commercial macadamia orchard in Nannuoshan Village, Menghai County, Xishuangbanna Dai Autonomous Prefecture, Yunnan Province (21°58′52.66″N, 100°37′8.40″E), at an elevation of approximately 1300 m. This plantation was established nine years ago with a tree spacing of 4.0 m × 6.0 m, and is characterized by a uniform stand structure and vigorous tree growth. The JH site is situated in the Macadamia Germplasm Resource Garden under the Ministry of Agriculture in Jinghong City (22°00′52.30″N, 100°46′48.41″E), at an elevation of approximately 550 m. This site functions as a conservation orchard and is managed under standardized horticultural protocols, ensuring consistently healthy tree development. Both sites are located within the northern tropical humid monsoon climate zone, which is typified by warm, humid conditions and distinct wet and dry seasons. The rainy season generally extends from May to October. The MH site has a mean annual temperature of 21.5 °C and receives approximately 1250 mm of precipitation annually. In contrast, the JH site exhibits a higher mean annual temperature of 24.3 °C, with annual precipitation ranging from 948 to 1515 mm.
4.2. Field Experiment Layout and Sample Collection
The field experiment was conducted in 2024 using macadamia nut trees (variety O.C.) of uniform age and growth status. Fifteen trees were randomly selected from each experimental plot, with one replicate assigned to every three trees, resulting in five biological replicates per treatment. Sampling was carried out on the 10th day of each month between 08:00 and 10:00. For each tree, fully expanded second leaves from the apex of mature shoots, as well as one-year-old branches, were collected from the eastern, southern, western, and northern quadrants of the canopy. The total mass of each tissue type per replicate was no less than 15 g. All samples were immediately flash-frozen in liquid nitrogen and stored at −80 °C in an ultra-low temperature freezer. Endogenous hormone concentrations were determined using high-performance liquid chromatography (HPLC). Sample preparation and analytical procedures followed the plant hormone quantification protocol described in reference [50]. Target hormones included zeatin (ZT), gibberellic acid (GA₃), indole-3-acetic acid (IAA), and abscisic acid (ABA).
4.3. Field Experiment Layout and Sample Collection
4.3.1. Sample Preparation Method
After branch or leaf tissues were excised, 3.0 g of each sample was accurately weighed and finely ground into powder in liquid nitrogen. Subsequently, 20 mL of pre-chilled 80% methanol and 2 mL of antioxidant solution were added. The mixture was subjected to ultrasonic extraction for 30 minutes in an ice bath and then stored at 4 °C overnight. On the following day, the samples were centrifuged at 6000 rpm for 20 minutes, and the resulting supernatant was transferred into a 50 mL centrifuge tube for further analysis. The residue was extracted again with five volumes of pre-chilled methanol, thoroughly mixed, and sonicated in an ice bath for 30 minutes. The mixture was then kept at 4 °C in the dark for 4 hours, followed by centrifugation at 6000 rpm for 15 minutes. The resulting supernatant was combined with the first extract.
Next, 0.2 g of cross-linked polyvinylpyrrolidone (PVP-P) was added to the pooled supernatant, thoroughly mixed, and shaken on a rotary shaker for 20 minutes. The mixture was then centrifuged at 6000 rpm for 15 minutes to remove impurities. The combined supernatant was concentrated at 40 °C using a rotary evaporator to remove methanol. The remaining solution was extracted three times with twice the volume of petroleum ether and once with an equal volume of ethyl acetate. The organic phases were pooled and evaporated to dryness at 40 °C. The residue was redissolved and brought to volume with 2 mL of chromatographic-grade methanol, then filtered through a 0.22 μm microporous membrane. The resulting filtrate was used for subsequent endogenous hormone analysis.
4.3.2. Measurement Method
(1) Preparation of Standard Solutions
Accurately weigh appropriate amounts of the standard substances zeatin (ZT), gibberellic acid (GA₃), indole-3-acetic acid (IAA), and abscisic acid (ABA). Dissolve each in methanol and dilute to a final volume of 10 mL to prepare stock standard solutions. The concentrations of the stock solutions are 0.505 mg/mL for ZT, 0.505 mg/mL for GA₃, 0.512 mg/mL for IAA, and 0.508 mg/mL for ABA, respectively.
(2) Chromatographic Conditions
Chromatographic analysis was carried out using an InertSustain AQ-C18 column (4.6 × 250 mm, 5.0 μm; GL Sciences, Japan). The column temperature was maintained at 30 °C, with a flow rate of 1.0 mL/min. The detection wavelength was set at 210 nm, and the injection volume was 10 μL. Gradient elution was performed according to the program detailed in Table 2.
(3) Assessment of Linearity
Stock solutions of each reference standard were serially diluted according to predetermined concentration gradients to prepare a series of standard solutions. Each solution was injected and analyzed under the chromatographic conditions described above. Standard calibration curves were generated by plotting the reference standard concentrations (X, μg/mL) against the corresponding peak areas (Y). The linear regression equations for each hormone component were then derived (Table 3).
(4) Determination of Endogenous Hormone Content
The extracted solutions from flower or leaf tissues were injected and analyzed under the chromatographic conditions described above. The peak areas corresponding to each hormone were recorded and applied to the respective linear regression equations to calculate their concentrations. The endogenous hormone content in each sample was then determined using the following formula:
where C is the concentration of the hormone in the sample (μg/mL), V is the final volume of the extract (mL), and m is the fresh weight of the tissue sample used for extraction (g).
4.4. Data Processing
Data were analyzed using one-way analysis of variance (ANOVA) in SPSS 27.0 to assess significant differences among treatments. Statistical analysis and visualization of endogenous hormone concentrations and their ratios were performed in R software (version 4.2.2). Pearson correlation coefficients were calculated using the cor() function, and correlation heatmaps were generated with the corrplot package to visualize the relationships among variables. For data that did not meet the assumptions of normality, Spearman’s rank correlation was calculated using the cor.test() function. Data reshaping and figure enhancement were conducted using the ggplot2 and reshape2 packages to improve clarity and presentation.
5. Conclusions
This study systematically elucidated the annual dynamics of key endogenous hormones and their coordinated regulatory mechanisms associated with off-season flowering in Macadamia integrifolia cultivated in high-altitude regions. Clear differences in phenological progression were observed between off-season and normally flowering sites, accompanied by region-specific and temporally variable patterns in both hormone concentrations and hormonal ratios. Compared with the normal-flowering site, floral bud differentiation at the off-season site occurred significantly earlier and was associated with persistently low GA₃ levels as well as elevated concentrations of ZT and ABA. Furthermore, critical hormonal ratios, including ABA/GA₃, ABA/IAA, and ZT/GA₃, increased markedly during the early stages of floral induction. These changes suggest the establishment of an internal hormonal environment that disrupts conventional phenological rhythms and promotes the initiation of reproductive development. Collectively, these findings provide important theoretical insights into the endogenous regulatory networks underlying flowering phenology in Macadamia, and offer a scientific basis for the development of targeted, hormone-based strategies for flowering regulation. Such approaches hold considerable potential for enhancing environmental adaptability and stabilizing yield in high-altitude Macadamia production systems.
Author Contributions
Conceptualization, H.Y.; Software, Y.N.; Validation, Y.N.; Formal Analysis, Y.N.; Investigation, Y.C.; Resources, T.Y.; Data Curation, Y.N.; Writing - Original Draft Preparation, Y.N.; Writing - Review & Editing, Y.N. and X.H.; Visualization, Y.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the following programs: the Project on Flowering Regulation in Macadamia (RF2025-33); the Special Program for the Construction of Tropical Crop Science and Technology Innovation System in Yunnan Province (RF2025-13); the Innovation Guidance and Technology-based Enterprise Incubation Program (202404BP090014); and the Xingdian Talents Support Program (2022-0068).
Data Availability Statement
Data will be made available on request.
Conflicts of Interest
he authors declare no conflicts of interest.
References
- Rahman, A.; Wang, S.; Yan, J. S.; Xu, H. R. Intact macadamia nut quality assessment using near-infrared spectroscopy and multivariate analysis. J. Food Compos. Anal. 2021, 102, 104033. [CrossRef]
- Appleby, N.; Edwards, D.; Batley, J. New technologies for ultrahigh throughput genotyping in plants. Methods Mol. Biol. 2009, 513, 19–39. [CrossRef]
- Malvestiti, R.; Borges, L.S.; Weimann, E.; et al. The effect of macadamia oil intake on muscular inflammation and oxidative profile kinetics after exhaustive exercise. Eur. J. Lipid Sci. Technol. 2017, 119, 1600382. [CrossRef]
- He, X.Y.; Tao, L.; Liu, J.; et al. Overview and development trends of the global macadamia industry. South China Fruits. 2015, 44(4), 5. [CrossRef]
- Sedgley, M.; Olesen, T. Flowering and fruit-set in macadamias (Macadamia integrifolia): Studies on temporal patterns and sources of yield fluctuation. Scientia Horticulturae 1992, 50(1–2), 107–118.
- Trueman, S.J. The reproductive biology of macadamia. Sci. Hortic. 2013, 150, 354–359. [CrossRef]
- Ning, Y.; Chen, Y.C.; Yue, H.; et al. Investigation of flowering rhythm and preliminary study on flowering period reg-ulation in macadamia in Yunnan. Tropical Agricultural Science and Technology. 2024, 47(1), 1–5. [CrossRef]
- Maple, R.; Zhu, P.; Hepworth, J.; Wang, J.-W.; Dean, C. Flowering time: From physiology, through genetics to mechanism. Plant Physiol. 2024, 195(1), 190–212. [CrossRef]
- Yuan, C.Q.; Ahmad, S.; Cheng, T.R.; et al. Red to far-red light ratio modulates hormonal and genetic control of axillary bud outgrowth in Chrysanthemum. Int. J. Mol. Sci. 2018, 19(6), 1590. [CrossRef]
- Kaur, A.; Maness, N.; Ferguson, L.; et al. Role of plant hormones in flowering and exogenous hormone application in fruit/nut trees: A review of pecans. Fruit Res. 2021, 1(1), 1–9. [CrossRef]
- Chen, X.; Qi, S.; Zhang, D.; Li, Y.; An, N.; et al. Comparative RNA-sequencing–based transcriptome profiling of buds from profusely flowering ‘Qinguan’ and weakly flowering ‘Nagafu no. 2’ apple varieties reveals novel insights into the regulatory mechanisms underlying floral induction. BMC Plant Biol. 2018, 18, 370. [CrossRef]
- Gangwar, S.; Singh, V.P.; Tripathi, D.K.; et al. Plant responses to metal stress: the emerging role of plant growth hormones in toxicity alleviation. In: Ahmad, P.; Rasool, S., Eds.; Emerging Technologies and Management of Crop Stress Tolerance; Academic Press: Cambridge, MA, USA, 2014; pp. 215–248. [CrossRef]
- O Okada, K.; Ueda, J.; Komaki, M.K.; Bell, C.J.; Shimura, Y. Requirement of the auxin polar transport system in early stages of Arabidopsis floral bud formation. Plant Cell 1991, 3(7), 677–684. [CrossRef]
- Dar, N.A.; Amin, I.; Wani, W.; et al. Abscisic acid: A key regulator of abiotic stress tolerance in plants. Plant Gene 2017, 11, 106–111. [CrossRef]
- Zeng, H.; Chen, H.B.; Du, L.Q.; et al. Effect of spraying gibberellin on the flower bud formation of Macadamia (Macadamia integrifolia). J. Fruit Sci. 2008, 25(2), 203–208. [CrossRef]
- Zhang, M.; Han, M.; Ma, F.; Shu, H. Effect of bending on the dynamic changes of endogenous hormones in shoot terminals of ‘Fuji’ and ‘Gala’ apple trees. Acta Physiol. Plant. 2015, 37, 1–9. [CrossRef]
- Cao, S.Y.; Zhang, J.C.; Wei, L.H. Changes in endogenous hormones during floral bud development in apple. Journal of Fruit Science. 2000, 17(4), 244–248. [CrossRef]
- Mutasa-Göttgens, E.; Hedden, P. Gibberellin as a factor in floral regulatory networks. Journal of Experimental Botany, Volume 60, Issue 7, May 2009, Pages 1979–1989. [CrossRef]
- Werner, T.; Motyka, V.; Laucou, V.; et al. Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell 2001, 13(11), 2539–2551. [CrossRef]
- Zwack, P.J.; Rashotte, A.M. Interactions between cytokinin signalling and abiotic stress responses. Journal of Experimental Botany, Volume 66, Issue 16, August 2015, Pages 4863–4871. [CrossRef]
- Ramirez, J.; Hoad, G.V. Cytokinin application induces flowering on defoliated pear spurs: evidence from Zeatin experiments. J. Exp. Bot. 2008, 59, 3215–3222. [CrossRef]
- Takei, K.; Ueda, N.; Aoki, K.; et al. AtIPT3 is a key determinant of nitrate-dependent cytokinin biosynthesis in Arabidopsis. Plant and Cell Physiology, Volume 45, Issue 8, 15 August 2004, Pages 1053–1062. [CrossRef]
- Yang, C.; Xu, Z.; Wang, X.; et al. Hormone profiling and ratio dynamics define floral fate in apple. Plant Physiol. 2021, 186, 1782–1796. [CrossRef]
- Leibfried, A.; To, J.P.C.; Busch, W.; et al. WUSCHEL controls meristem function by direct regulation of cytokinin-inducible response regulators. Nature 2005, 438(7071), 1172–1175. [CrossRef]
- Hwang, I.; Sheen, J.; Müller, B. Cytokinin signaling networks. Annu. Rev. Plant Biol. 2012, 63, 353–380. [CrossRef]
- Richter, R.; Behringer, C.; Zourelidou, M.; Schwechheimer, C. Convergence of auxin and gibberellin signaling on regulation of GNC and GNL in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 13192–13197. [CrossRef]
- de Lucas, M.; Davière, J.M.; Rodríguez Fernández, M.; et al. A molecular framework for gibberellin control of cell division and expansion in Arabidopsis. Curr. Biol. 2008, 18, 2156–2165. [CrossRef]
- Lee, J.O.; Park, H.M.; Lee, H.I. SOC1 translocated to the nucleus by interaction with AGL24 directly regulates LEAFY.The Plant Jorrnal. 2008, 55(5), 832–843. [CrossRef]
- Moon, J.; Zhu, L.; Shen, L.; et al. Gibberellin regulates Arabidopsis floral initiation through modulation of SOC1 and LEAFY expression. Plant Cell 2003, 15, 1553–1565.
- Zeng, H.; Du, L.Q.; Zou, M.H.; et al. Changes in endogenous hormone levels during floral bud differentiation in macadamia. J. Anhui Agric. Sci. 2008, 36(34), 14949–14953. [CrossRef]
- Kotake, T.; Nakagawa, N.; Takeda, K.; Sakurai, N. Auxin-induced elongation growth and expressions of cell wall-bound exo- and endo-β-glucanases in barley coleoptiles. Plant Cell Physiol. 2000, 41, 1272–1278. [CrossRef]
- Jung, C. Flowering time regulation: Agrochemical control of flowering. Nat. Plants 2017, 3(4), 17045. [CrossRef]
- Brunoud, G.; Wells, D.; Oliva, M.; et al. A novel sensor to map auxin response and distribution at high spatio-temporal resolution. Nature 2012, 482, 103–107. [CrossRef]
- Wang, W.F.; Chen, X.; et al. Transcriptomic analysis of topping-induced axillary shoot outgrowth in Nicotiana tabacum. Gene 2018, 646, 169–180. [CrossRef]
- Wang, Y.; Zhang, Y.; Wang, L.; et al. Abscisic acid modulates floral transition by interacting with flowering pathway integrators FT and SOC1 in Arabidopsis. Front. Plant Sci. 2021, 12, 638123.
- Shu, K.; Zhou, W.; Chen, F.; Luo, X.; Yang, W. Abscisic acid and gibberellins antagonistically mediate plant development and abiotic stress responses. Front. Plant Sci. 2018, 9, 416. [CrossRef]
- Fischer, U.; et al. Hormonal crosstalk: a central regulator in plant stress signaling. Plant Physiol. 2017, 174(2), 106–121.
- Zhang, S.; Han, M.; Ma, F.; et al. Effect of exogenous GA₃ and its inhibitor paclobutrazol on floral formation, endogenous hormones, and flowering-associated genes in ‘Fuji’ apple. Plant Physiol. Biochem. 2016, 107, 178–186. [CrossRef]
- Shalom, L.; Samuels, S.; Zur, N.; et al. Fruit load induces changes in global gene expression and in abscisic acid (ABA) and indole acetic acid (IAA) homeostasis in citrus buds. J. Exp. Bot. 2014, 65(12), 3029–3044. [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; et al. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23(10), 680–694. [CrossRef]
- Altmann, M.; Altmann, S.; Rodriguez, P.A.; et al. Extensive signal integration by the phytohormone protein network. Nat. Commun. 2020, 11(1), 1–11. [CrossRef]
- Guo, W.W.; Wu, X.M.; Deng, X.X.; et al. Advances in Citrus Flowering: A Review. Front. Plant Sci. 2022, 13, 868831. [CrossRef]
- Vanstraelen, M.; Benková, E. Hormonal interactions in the regulation of plant development. Annu. Rev. Cell Dev. Biol. 2012, 28, 463–487. [CrossRef]
- Nguyen, T.P.; Setiawan, S.; Omega, E.; et al. Correlation-based hormone network analysis reveals IAA and GA₃ as central inte-grators of floral induction. Sci. Hortic. 2021, 281, 109976. [CrossRef]
- Li, W.; Chen, Z.; Wei, D.; et al. Hormonal interactions in grapevine flowering regulation: insights from IAA–GA synergy. BMC Plant Biol. 2020, 20, 114.
- Moubayidin, L.; Di Mambro, R.; Sabatini, S. Cytokinin–auxin crosstalk. Trends Plant Sci. 2009, 14(10), 557–562. [CrossRef]
- Yamaguchi, N.; Winter, C.M.; Wu, M.F.; et al. A molecular framework for auxin-mediated initiation of flower primordia. Dev. Cell 2009, 16(5), 765–777. [CrossRef]
- Zhang, Z.; Wang, Y.; Zhang, J.; Wang, P. Hormone crosstalk during flower development: Insights from transcriptome analysis in grapevine. BMC Plant Biol. 2021, 21, 421.
- Li, H.; Chen, X.; Liu, Y.; Wang, Y. Hormonal crosstalk regulates flowering time under stress conditions: A comparative study in rice. Plant Physiol. Biochem. 2023, 199, 107795. [CrossRef]
- Huang, F.Y. Study on tissue culture and endogenous hormone content of Lagerstroemia ‘Zijingling’. Master’s Thesis, Central South University of Forestry and Technology. 2022. [CrossRef]
Figure 2.
Dynamic changes in endogenous hormone contents in Macadamia leaves at MH (purple line) and JH (green line) sites. (a) Zeatin (ZT); (b) Gibberellic acid (GA₃); (c) Indole-3-acetic acid (IAA); (d) Abscisic acid (ABA). Error bars represent standard error (n = 5). Different lowercase letters indicate significant differences (p < 0.05) among months within the same site based on one-way ANOVA followed by the LSD test.
Figure 2.
Dynamic changes in endogenous hormone contents in Macadamia leaves at MH (purple line) and JH (green line) sites. (a) Zeatin (ZT); (b) Gibberellic acid (GA₃); (c) Indole-3-acetic acid (IAA); (d) Abscisic acid (ABA). Error bars represent standard error (n = 5). Different lowercase letters indicate significant differences (p < 0.05) among months within the same site based on one-way ANOVA followed by the LSD test.
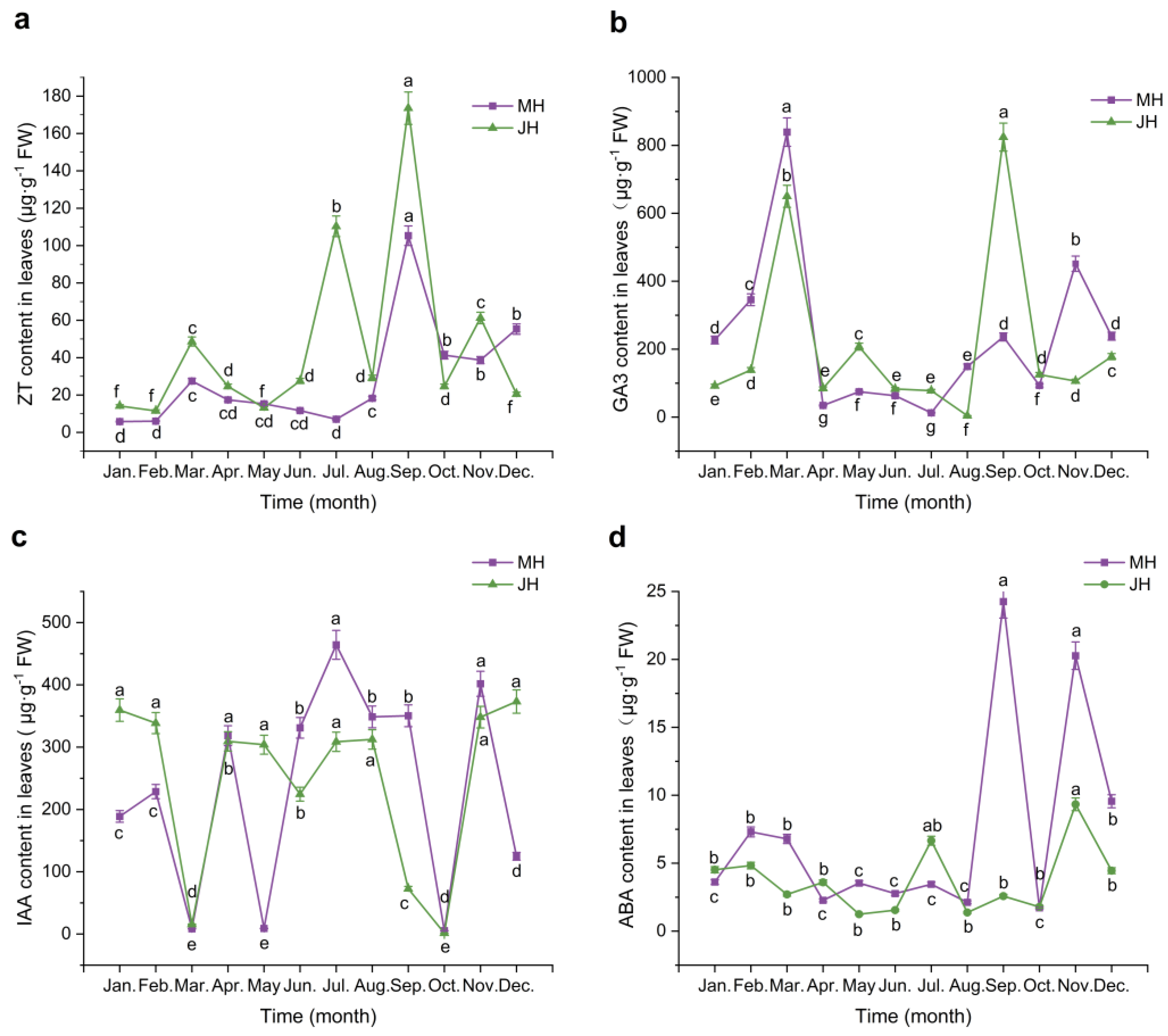
Figure 3.
Dynamic changes in endogenous hormone contents in Macadamia branches at MH (purple line) and JH (green line) sites. (a) Zeatin (ZT); (b) Gibberellic acid (GA₃); (c) Indole-3-acetic acid (IAA); (d) Abscisic acid (ABA). Error bars represent standard error (n = 5). Different lowercase letters indicate significant differences (p < 0.05) among months within the same site based on one-way ANOVA followed by the LSD test.
Figure 3.
Dynamic changes in endogenous hormone contents in Macadamia branches at MH (purple line) and JH (green line) sites. (a) Zeatin (ZT); (b) Gibberellic acid (GA₃); (c) Indole-3-acetic acid (IAA); (d) Abscisic acid (ABA). Error bars represent standard error (n = 5). Different lowercase letters indicate significant differences (p < 0.05) among months within the same site based on one-way ANOVA followed by the LSD test.
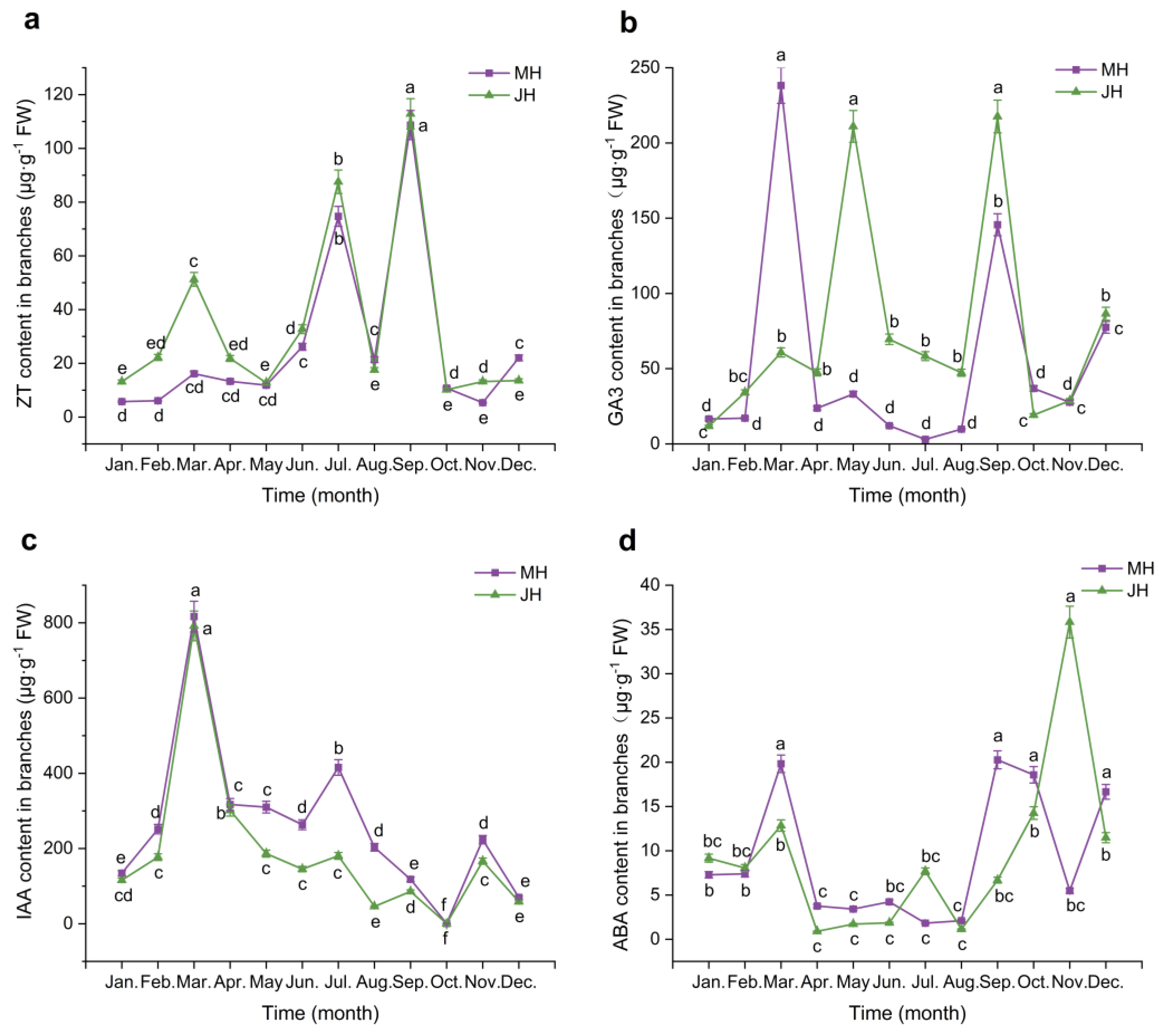
Figure 4.
Dynamic changes in the balance ratios of endogenous hormones in Macadamia at MH (a) and JH (b) sites. Heatmaps present monthly variations in four hormone ratios (ABA/GA₃, ZT/GA₃, ABA/IAA, and ZT/IAA) in leaves (L) and branches (B) from January to December. Color intensity indicates the relative magnitude of the ratios (unitless), as shown by the scale bar.
Figure 4.
Dynamic changes in the balance ratios of endogenous hormones in Macadamia at MH (a) and JH (b) sites. Heatmaps present monthly variations in four hormone ratios (ABA/GA₃, ZT/GA₃, ABA/IAA, and ZT/IAA) in leaves (L) and branches (B) from January to December. Color intensity indicates the relative magnitude of the ratios (unitless), as shown by the scale bar.
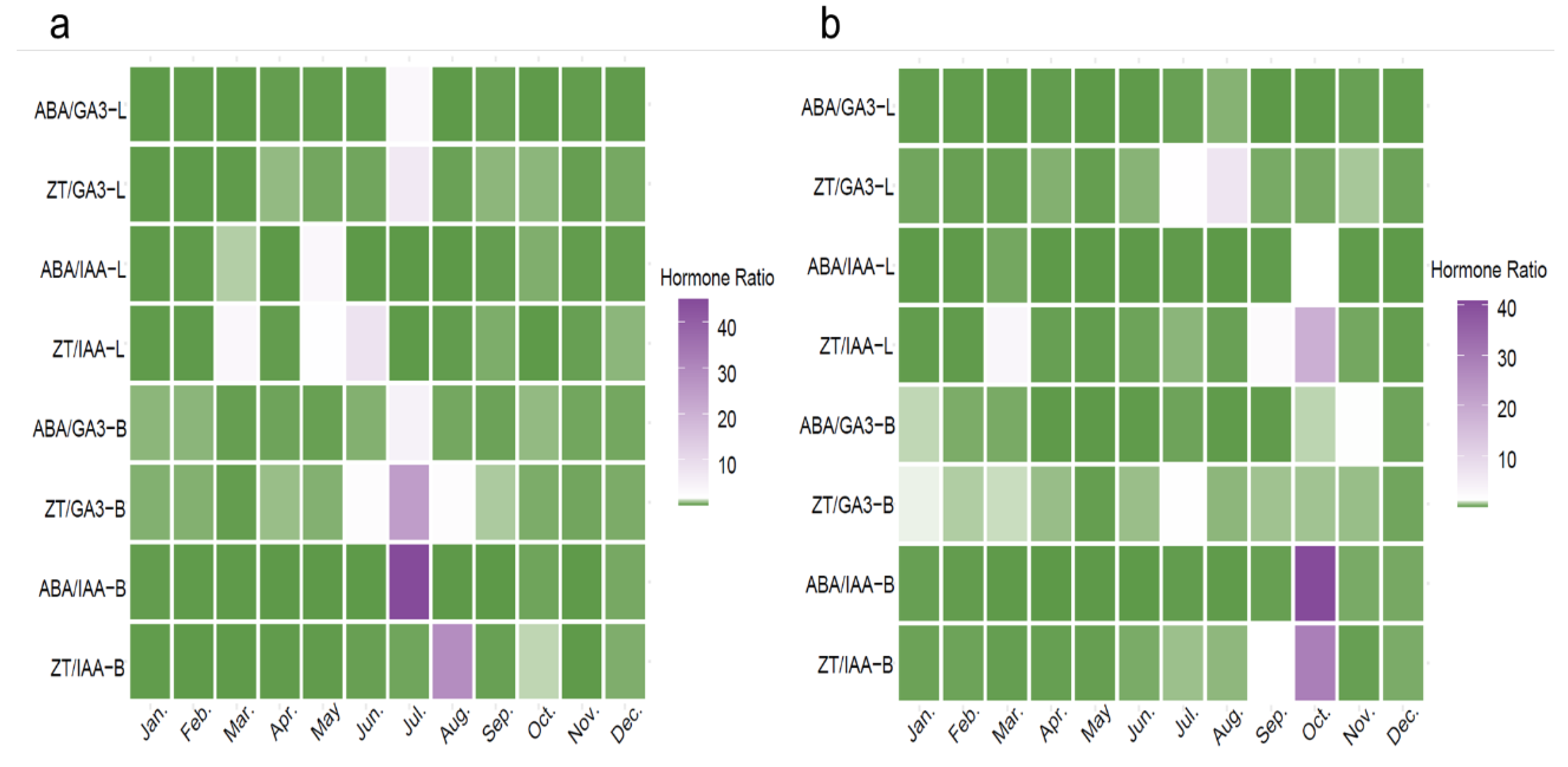
Figure 5.
Correlation analysis of endogenous hormones and their ratios in Macadamia at MH (a) and JH (b) sites. The size and color intensity of the circles indicate the strength of the correlation. Red circles represent positive correlations and blue circles represent negative correlations. An asterisk (*) indicates statistically significant correlations (p < 0.05), and two asterisks (**) indicate highly significant correlations (p < 0.01).
Figure 5.
Correlation analysis of endogenous hormones and their ratios in Macadamia at MH (a) and JH (b) sites. The size and color intensity of the circles indicate the strength of the correlation. Red circles represent positive correlations and blue circles represent negative correlations. An asterisk (*) indicates statistically significant correlations (p < 0.05), and two asterisks (**) indicate highly significant correlations (p < 0.01).

Table 1.
Phenological Survey of Two Macadamia Planting Sites.
| Sampling Site | Initial Flowering Period | Peak Flowering Period | Fruit Enlargement Period | Fruit Oil Accumulation Period | Maturity & Harvest | Floral Bud Dormancy Break Period |
|---|---|---|---|---|---|---|
| MH | Oct–Nov | Jan | Feb–Apr | May–Jul | Aug | Aug–Sep |
| JH | Jan–Feb | Mar | Apr–May | Jun–Aug | Sep | Oct–Dec |
Table 2.
HPLC Gradient Elution Procedure of Endogenous Hormones in Macadamia.
| Time (min) |
Acetonitrile (%) |
Methanol (%) |
0.1% Phosphoric Acid (%) |
|---|---|---|---|
| 3 | 5 | 5 | 90 |
| 10 | 12 | 12 | 76 |
| 20 | 22 | 22 | 56 |
| 30 | 20 | 20 | 60 |
| 35 | 5 | 5 | 90 |
| 40 | 5 | 5 | 90 |
Table 3.
Linear Regression Equations for Endogenous Hormone Standards.
| Component | Regression Equation | Correlation Coefficient | Linear Range (μg/mL) |
|---|---|---|---|
| ZT | y = 43743x + 249867 | 0.999 | 0.203–208.077 |
| GA₃ | y = 9769.6x + 85522 | 0.996 | 0.202–206.848 |
| IAA | y = 75525x + 17728 | 0.999 | 0.202–206.848 |
| ABA | y = 18902x - 18636 | 0.999 | 0.204–209.715 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



