Submitted:
06 February 2024
Posted:
08 February 2024
You are already at the latest version
Abstract
The AREB/ABF (ABA response element binding) proteins in plants are vital for plant stress responses, but the understanding of AREB/ABFs in orchid species, an important traditional medicinal and ornamental plants, is limited. Here, twelve AREB/ABF genes were identified within three orchids’ complete genome and classified into three groups (groups I, II, and III) via a phylogenetic analysis, which was further supported by the analysis of their conserved motifs and gene structures. The cis-elements analysis showed that hormone response elements followed by light and stress responses were widely rich in AREB/ABFs. A prediction analysis of orchid ABRE/ABF-mediated regulatory network was further constructed through cis-regulatory elements (CREs) analysis of their promoter regions and revealed that several transcriptional factor (TF) gene family were abundant as the potential regulators for orchid AREB/ABFs. Expression profile analysis based on public transcriptomic data showed that most AREB/ABF genes have distinct tissue-specific expression patterns in orchid plants. Additionally, DcaABI5 as the homolog of ABA INSENSITIVE 5 (ABI5) from Arabidopsis was selected for further analysis. The results showed that Arabidopsis transgenic overexpressing DcaABI5 could rescue the ABA insensitive phenotype in the mutant abi5. Collectively, these findings will provide valuable information on AREB/ABF genes in orchids.
Keywords:
Orchid
; gene family
; AREB/ABF genes
; cis-regulatory elements (CREs) analysis
; gene expression
; abscisic acid (ABA)
; ABA INSENSITIVE 5 (ABI5)
1. Introduction
As one of the most species-rich plant families, the orchid family (Orchidaceae) consists of approximately 30,000 orchid species worldwide, which is a diverse and ecologically important plant family [1,2,3]. In addition to its ecological importance, the orchid provides numerous sources for ecology, pharmaceuticals, food, and aesthetics [4,5,6,7]. Many orchid plants mostly grow on shady mountain rocks or forest trunks, and is often threatened by adversity such as drought, light expose [8]. Therefore, it is important to identify stress-related genes from orchid genome and investigate their functions. In recent years, the high-quality chromosome-scale assembly of genome sequence for several orchid species has been achieved [9,10,11,12]. Thus, the valuable sequence of the orchid genome will provide genetic resources for gene functional studies and essential information for the further modification of orchid varieties to increase yield.
Under unfavorable growth conditions, plants will produce elevated levels of stress-related hormone abscisic acid (ABA) [13]. ABA notably reduces stress damage in plant by controlling expression of many stress-related genes involved in signal transduction pathways [14,15]. The ABA-mediated signal network has been extensively studied in model plant Arabidopsis. Generally, ABA binds to the receptor PYR/PYL/RCAR and inhibits the protein phosphatase activity of PP2Cs, which hinder the kinase activity of sucrose non-fermenting2-related protein kinase SnRK2. Subsequently, accumulation of the activated SnRK2 kinases directly target downstream ABA-responsive genes including the ABA-responsive element binding protein/ABRE-binding factor (AREB/ABF) subfamily members, allowing activating expression of target genes [16,17,18,19]. ABA-independent pathway has been also proposed with the participation of particularly important transcriptional factors (TFs) in stress response. For instance, Osmotic stress-responsive gene expression is regulated by ABA-dependent and ABA-independent pathways governed by two key transcription factors: AREB/ABFs and DREB2A, respectively [20,21].
AREB/ABF subfamily members, which belong to a subgroup of the basic leucine zipper (bZIP) TF family, are the most essential representatives of the ABA-responsive regulatory pathway and have important roles in plant responses to abiotic stress [22,23,24]. As one of the largest TF families, the bZIP gene family is the most abundant and evolutionarily conserved gene family in plants. The bZIP gene family is characterized by the highly conserved bZIP domain, and mostly bind to the cis-element with an ACGT core motif of the target genes’ promoters, including TACGTA (A-box), GACGTC (C-box), and CACGTG (G-box) [25,26]. In Arabidopsis, nine members (ABF1, ABF2, ABF3, ABF4, AtBI5, AtDPBF2, AtDPBF3, AtDPBF4, and AtbZIP15) consist of the AREB/ABF subfamily, which belong to group A of bZIP family with ten groups [24]. Numerous ABA and/or stress-regulated genes contain a (C/T)ACGTGGC consensus sequence, known as the ABA responsive element (ABRE), in their promoter regions, which are directly targeted by AREB/ABF genes including AREB1/ABF2 and ABF3 under stressful conditions [27,28,29,30,31]. Many studies have demonstrated that the regulatory effects of AREB/ABF TFs on their target genes are mainly induced by stresses, and thereby contribute to stress tolerance in plants [32,33].
Several studies have reported the AREB/ABF genes from various plants in improving abiotic stress adaptability and resistance, such as in Arabidopsis [24], rice [33], wheat [34,35,36], potato [37,38], cotton [39], apple [40], strawberry [41], rose [42], Lily [43], Kiwifruit [44], and Jute [45]. To date, the AREB/ABF subfamily in orchid plants remains largely unknown. In this study, twelve AREB/ABF genes were identified within three orchids’ complete genome: Dendrobium catenatum (D. catenatum), Apostasia shenzhenica (A. shenzhenica), and Phalaenopsis equestri (P. equestris). The features of these AREB/ABF gene family were characterized using combined bioinformatics methods. And the representative AREB/ABF members DcaABI5 from D. catenatum was demonstrated to play an essential role in ABA signaling response. Collectively, our results will provide valuable information on AREB/ABF genes and provide a perspective for further functional characterization of potentially important ones in orchids.
2. Results
2.1. Identification and Characteristics of Orchid AREB/ABFs
AREB/ABF subfamily members from the three orchid species genome (D. catenatum, A. shenzhenica, and P. equestris) were identified using Arabidopsis AREB/ABF proteins as the query to search for candidate genes by BLASTP. After strict screening and sequence analysis, 5, 4, and 3 AREB/ABF genes were identified from D. catenatum, A. shenzhenica and P. equestris, respectively. These AREB/ABF genes were named according to the similarity to the homologs from Arabidopsis. Sequence analysis revealed that the full-length coding sequences of these AREB/ABF genes ranged from 1278 bp (AshABF2) to 936 bp (DcaDPBF2) with an average of 1146 bp (Table 1). Analysis with ExPasy (https://www.expasy.org) showed that the molecular mass was predicted to range from 34.83 kDa (DcaABI5) to 45.56 kDa (AshABF2), and their isoelectric points (pI) varied widely from 5.47 (AshABI5) to 9.5 (DcaDPBF2). Additionally, the predicted subcellular localizations of these proteins suggested that all AREB/ABF subfamily proteins may be located in the nucleus.
The typical AREB/ABF has a highly conserved protein structure including conserved phosphorylation domains (C1, C2, C3 and C4), and kinase recognition motifs, RXXS/T and S/TXXE/D, as well as the bZIP domain [46]. Multiple sequence alignments analysis revealed that AREB/ABF proteins from the three orchid species contain all these conserved domains and kinase recognition motifs, which provides further support for their identities (Figure 1).
2.2. Phylogenetic Analysis of Orchid AREB/ABF Proteins
To explore the classification and evolutionary characteristics of these AREB/ABF genes from D. catenatum, A. shenzhenica and P. equestris, a non-rooted phylogenetic tree was constructed based on multiple sequence alignments of AREB/ABF proteins with homologs from Arabidopsis thaliana (9) and Oryza sativa (7). The phylogenetic tree showed that the all AREB/ABF proteins from these plant species were mainly divided into three groups (Figure 2). All the ABF proteins were clustered in Group A and the ABF proteins from orchid species exhibited a close relationship. However, ABF proteins from Arabidopsis and rice were individually clustered apart from those homologs from orchids. The ABI5 and DPBF proteins were clustered into two other groups and distributed evenly among all species.
2.3. Conserved Motifs and Gene Structure Analysis of Orchid AREB/ABF genes
In order to understand the conservation and diversification of AREB/ABFs, the putative motifs of all AREB/ABF proteins were predicted by MEME motif analysis according to the full-length phylogenetic relationships. A total of 10 motifs ranging from 17 to 49 amino acids were identified and named as the corresponding motifs 1–10. The number of the conserved motifs for each AREB/ABF protein ranged from 5 to 10, and AREB/ABFs in the same group or subgroup shared highly similar motif compositions (Figure 3A). All AREB/ABF from group I as in phylogenetic analysis had 10 motifs except for AshABF1, which was short of motif 8. The AREB/ABF from the other two groups contained only 5 motifs. Notably, AREB/ABF from group C had no gap between motif 4 and motif 5.
The exon/intron distributions and the intron numbers were analyzed to further detect structural features and evolutionary events of the AREB/ABF genes. The AREB/ABF genes from group I generally had four exons and long introns except for AsABF2, which contained 5 exons and short introns (Figure 3B). The AREB/ABF genes from group II also had four 4 exons but short introns. Additionally, AREB/ABF genes from group III possessed either 3 or 5 exons and variable length of introns.
Collectively, the motif composition and exon/intron numbers of AREB/ABF genes in the same group were closer than that of genes between different groups.
2.4. Cis-element Analysis of orchid AREB/ABF Family
To identify putative cis-elements in the promoter regions of the AREB/ABF genes, 2000 bp upstream sequences of the AREB/ABF genes were investigated. As predicted, the composition of cis-acting elements was detected (Figure 4A). Phytohormone-responsive elements, mainly correlated with ABA (ABRE), ethylene (ERE), GA (GARE-motif, P box, TATC-box), JA (CGTCA-motif), auxin (AuxRR core, TGA-element), and SA (TCA-element), were detected in the promoter regions. Among them, ABRE, ERE and CGTCA-motif were mostly enriched in all AREB/ABF genes. The defense and stress response elements were widely found within these AREB/ABF genes, including MBS (drought response element), LTR (low-temperature responsive element), WUN-motif and WRE3 (wound-responsive element). Notably, the dominant stress response element STRE (stress responsive element) was presented in most AREB/ABF genes except for AsABF1, AsABF2 and AsDEPB2. The most numerous response elements found in the promoter region of AREB/ABFs were related to light response, including Box 4, G-box, GATA-motif, I-box, TCT-motif. Additionally, some elements associated with tissue specific expression were also detected. For instance, three CAT-box related to meristem expression were found in the promoter region of AsABF1. The circadian element related to circadian expression pattern was found in some AREB/ABF genes. Moreover, two other less known elements were detected, including O2-site (zein metabolism regulation) and MBSI (flavonoid biosynthetic genes regulation). These results provide fundamental clues regarding the possible functions and expression patterns of AREB/ABF genes related to the composition of cis-acting elements.
2.5. Prediction analysis of AREB/ABF-mediated regulatory network in orchid plants
To predict the potential roles of AREB/ABFs in orchid plants, the CREs analysis of their promoter regions was constructed. The results showed that hundreds of TFs belonged to over 30 different TF families, such as ERF, MYB, C2H2, ARF, WRKY, bZIP, NAC, LBD, MADS, Dof, etc., were identified as the potential regulators for AREB/ABFs from the three orchid plants (Figure S1). The predicted TFs were abundant in ERF, WRKY, bZIP, MYB, NAC and Dof. Based on the prediction results, DcaABF1 and AshABF1 have the biggest number of regulators among all AREB/ABFs (302 TFs), followed by DcaABI5 (265 TFs), AshABF2 (258 TFs), and PeqABF1 (229 TFs) (Table S1). In addition, we identified the top seven gene families that were predicted to regulate all AREB/ABFs, which included ERF, WRKY, MYB, bZIP, C2H2, Dof, and NAC (Figure 5A). We also compared the regulators for all the AREB/ABFs and found the ERF family being the most enriched except for AshABI5, which was predicted to be mostly targeted by NAC (29 TFs) family members (Figure 5B). Additionally, we studied the common and specific TF regulators for AREB/ABFs in each orchid species. The results showed that 13 and 11 TFs are as the common regulators for AREB/ABFs from D. catenatum and A. shenzhenica, respectively (Figure 5C). Unexpectedly, a total of 55 TFs were found as the common regulators for AREB/ABFs from P. equestris. Individual AREB/ABF was also found targeted by specific TFs. For instance, DcaABI5 was predicted to be specifically targeted by 39 TFs among five AREB/ABFs from D. catenatum and more detailed analysis showed that DcaABI5 was probably regulated by Dehydration-responsive element-binding protein 1C (DREB1C, AT4G25470) and DERB1D (AT5G51990), two proteins involved in freezing tolerance and cold acclimation as well as drought tolerance [47,48,49]. Overall, the predicted TF regulatory network of AREB/ABFs suggests their potential involvement in plant growth, response to stresses, and network associations.
2.6. Distinct Expression Profiles of Orchid AREB/ABF Genes in Different Tissues
To analyze the expression profiles of the orchid AREB/ABF genes, we investigated their transcripts abundance patterns across multiple tissues including flower, leaf, stem and root based on transcriptome data available in public database. The heat map showed that most AREB/ABF genes had tissue-specific expression patterns (Figure 6A-C). For instance, DcaABF1 and DcaABF2 were abundantly expressed in most tissues except for leaf, stem, and pollinium. Instead, DcaABF3 was highly expressed in leaf and stem. DcaDPBF2 was found presented in all tissues except for pollinium, while DcaABI5 was mainly distributed in flower tissues, especially in pollinium. In A. shenzhenica, AshABF1 and AshDPBF2 were mainly expressed flower tissues, leaf, and tuber, while AshABF2 was highly presented in stem. Notably, AshABI5 was dominantly found in pollinium. A similar expression pattern was found in PeqABI5 from P. equestris, which was strongly accumulated in pollinium.
Transcriptome analysis showed that ABI5 in Arabidopsis was dominantly expressed in developing seed (Figures S2). Since the expression pattern of DcaABI5 in seed is not available in public database, we performed RT-qPCR and found the comparable expression of DcaABI5 in fresh seeds (Figure 6D). These findings revealed that ABI5 from these orchid plants presented high expression in pollinium but low in seed, suggesting diversity role of ABI5 in orchid species.
2.7. Functional Analysis of DcaABI5
ABI5 as the typical bZIP protein is well-known in response to ABA signaling [50]. The different expression profiles in DcaABI5 compared with ABI5 from Arabidopsis raised us to investigate the role of ABI5 in D. catenatum. We firstly investigated the subcellular localization of DcaABI5-GFP in tobacco leaves with a fluorescence confocal microscope. The result showed that DcaABI5 was mainly presented in the nucleus (Figure 7A). In controls using the empty transformation vector carrying the GFP gene, GFP was distributed throughout the cell.
To reveal the role of DcaABI5 in ABA signaling, we introduced the T-DNA insertion line abi5 from Arabidopsis and had DcaABI5 overexpressed in this genetic background. Further phenotypic analysis revealed that wild-type Col-0 was sensitive to ABA with reduced seed germination rate and green cotyledon (Figure 7B, C). Notably, The DcaABI5 overexpression led to severe ABA sensitivity. On the contrary, abi5 is insensitive to ABA as previously reported [51], while the ABA insensitive phenotype in abi5 was largely rescued with DcaABI5 overexpressed. We further examine the expression pattern for representative targets of ABI5, including EM1 and RAB18 [52]. The results showed that DcaABI5 overexpression significantly promoted the expression of EM1 and RAB18 upon ABA treatment in Arabidopsis (Figure 7D, E). Additionally, DcaABI5 overexpression also efficiently rescue the transcriptional regulation of these target genes in abi5 mutant.
Collectively, these data suggested that DcaABI5 also played an essential role in ABA singling pathway.
3. Discussion
Many orchid plants grown in the wild forest are usually suffered from environmental stresses including drought, low temperature and light. It’s thus important to learn the fundamental mechanism behind the survival strategies in orchids. A number of studies have shown the importance of ABA perception and signaling for the stress response in plants [50,53,54]. The AREB/ABF subgroup from bZIP gene family are demonstrated to play an essential role in ABA signaling pathway for the plant adaptation to external stresses [18,22]. Nevertheless, detailed information concerning AREB/ABFs characters and functions from orchids, particularly their role in stresses responses, largely remains unclear.
In this study, a total of twelve AREB/ABF genes were identified from three orchid species, and sequence analysis suggested that all AREB/ABF genes contain conserved domains, such as C1-C4 and bZIP (Figure 1). This feature certainly provides full support for the identify for AREB/ABF subgroup family from orchids. Compared to the numbers of AREB/ABF family in Arabidopsis (nine) and rice (seven), the side of this gene family is smaller in the orchid species with five, four, three in D. catenatum, A. shenzhenica, and P. equestris, respectively. It seems that the number of AREB/ABF gene family does not completely correlate with the genome size. As compared to the genome size of Arabidopsis (125 Mb) and rice (480 Mb), the genome of D. catenatum (1.11 Gb) and P. equestris (1.16 Gb) are substantially larger. Gene duplication events have been shown to play an important role in genome expansion and production of large numbers of gene families [55]. Notably, whole genome duplication evens were found occurred in all modern orchids, which may be related to their diversification [10,11,56,57]. For instance, the gene expansion in the SWEET gen family from Dendrobium chrysotoxum may associated with enrichment in polysaccharides [57]. However, the number of genes is smaller, suggesting that AREB/ABF gene duplication might not occur during gene evolution in the three orchid species. The MADX-box gene family in orchids is also smaller than that in Arabidopsis (107 genes) and rice (80 genes) with 51 and 63 putative ones identified in P. equestris, and D. catenatum, respectively [9,10,58]. Despite having fewer MADS-box genes, orchids contain more MADS-box genes related to floral organ production, suggesting that higher MADS-box gene diversity might be connected with highly specific floral morphological traits in orchids. It’s also possible that the gene diversity of AREB/ABF gene in orchids might be associated with specific biological functions. Interestingly, the phylogenetic analysis showed that all the orchid ABF genes were clustered into group A and formed a unique subgroup apart from homologs from Arabidopsis and rice (Figure 2), implying that these genes might share independent functions and functional redundancy in orchids.
As observed in other plants [39,40,41,42,43,44,45], phylogenic closed AREB/ABFs members always have relatively consistent motifs and exon/intron structures (Figure 3), suggesting that those AREB/ABF subgroup proteins might function in the same manner. Of the 10 conserved motifs identified, motif 1 and motif 2-5 were highly conserved in all AREB/ABF proteins (Figure 3), which is composed of the conserved bZIP domain and conserved phosphorylation domains C1-C3, respectively. The bZIP domain is critical for the functional specificities of these transcription factors. The C1-C4 domains are supposed to function in decoding different signals. For instance, the domain C1-C3 was found to be phosphorylated by the SnRK2/SnRK3 or CDPK kinase in response to hyperosmotic/cold stress [59,60]. The C4 domain from AREB/ABF proteins is essential for their protein stability, as deletion of the C4 domain accelerates degradation of Arabidopsis AtABF1 and AtABF3 in vitro [61]. We found that the motif 7 is uniquely presented with C4 domain from ABF proteins. It’s likely that a 14-3-3 interaction site in the motif 7 contributes for the stability of ABF genes [61,62], indicating a gene diversity of AREB/ABF family.
Cis-elements play crucial roles in the transcription of genes for particular functions. In this study, many regulatory motifs related to hormone-responsive, stress-responsive, and growth development were predicted in the promoter region of the orchid AREB/ABF genes (Figure 4). Phytohormones play important roles in plant growth and response to various environmental stimulus [63]. We found that cis-elements related to ABA, JA, and ethylene are mostly enriched, suggesting that expression of ABRE/ABF genes in orchid may be closely related to these variable phyhormones under different environmental changes. It’s no doubt that all AREB/ABFs underling ABA regulation have ABRE elements. This is consistent with other AREB/ABF genes reported [28,45,46]. Hence, the AREB/ABF genes are certainly contributing for drought stress tolerance in orchids. JA is also involved in the regulation of important growth and developmental processes and responses against environmental stresses, such as stomatal opening, external damage et al [64,65]. The orchid AREB/ABF genes probably function as the cross-talk between the two hormones in mediating stomatal movement for dehydration and rehydration, or insert/pathogen attack for the attractive flowers. Unexpectedly, numerous cis-elements related to ethylene response were predicted in orchid AREB/ABF genes (Figure 4), which is different to those ones reported in other plant species, such as tomato, mei, jute [37,42,45]. Consistently, the predicted potential TFs binding to AREB/ABFs revealed that ethylene response factors (ERFs) are the most abundant (Figure 5B). It has been reported that ethylene and ABA jointly mediate seed germination, root growth, and fruit ripening [66,67,68]. The Arabidopsis ERF55 and ERF58 were found to directly regulate ABI5 transcription to suppress seed germination [69]. Thus, AREB/ABFs under regulation by ethylene probably play a role in seed germination, which is extremely complex in orchids and largely remains to be illustrated. Additionally, stress-responsive elements related to drought, cold and wound stresses were evenly predicted in the promoter region of AREB/ABF genes. Notably, cis-elements related to light responsive were highly redundant. Overall, the results suggested a close relationship between the expression of AREB/ABFs with growth of orchids under environmental stress conditions.
To better explore the function of AREB/ABFs, the expression levels of them in different tissues and developmental stages were evaluated with publicly available transcriptome data. As these data were generated from different experimental projects, the number and identity of tissues analyzed were not identical among the orchid species. Although there were differences in the expression profiles of the AREB/ABFs from different orchid species, some commonalities were observed. For instance, the ABI5 genes from three orchid species were found dominant expressed in pollinium (Figure 6A-C), indicating that orchid ABI5 gene may play an important and diversity role in the development of pollen. Unexpectedly, the ABI5 genes was relative low in orchid seeds, whereas ABI5 from Arabidopsis is mostly expressed in seed for dormancy and germination [51,52]. Since the ABI5 was conserved, we found that overexpression of the full length of DcaABI5 conferred sensitive to ABA during seed germination and cotyledon greening (Figure 7B). In addition, the ABA insensitive phenotype of abi5 mutant from Arabidopsis could be significantly rescued with DcaABI5 overexpressed, suggesting a conserved role of DcaABI5 in ABA signaling pathway.
4. Materials and Methods
4.1. Identification of the AREB/ABF Genes in Orchid Species
To identify all genes encoding AREB/ABFs in orchids, all the nine Arabidopsis AREB/ABFs were used as queries to search the orchid genome of OrchidBase 4.0 (http://orchidbase.itps.ncku.edu.tw/est/home2012.aspx) for candidate AREB/ABF sequences by using the BLASTP program. All resulting candidate protein sequences were further examined by SMART (http://smart.embl-heidelberg.de/) to confirm the integrity of the C1-C4 and bZIP domains. Finally, non-redundant and confident genes were gathered and assigned as the orchid AREB/ABF genes. The features including isoelectric point (pI) and molecular weight (MW) of AREB/ABFs were analyzed using ExPASy-ProtParam (Expasy 3.0; http://web.expasy.org/protparam/).
4.2. Analyses of Conserved Motifs, Exon-Intron Structures and Cis-regulatory Elements in Orchid AREB/ABF Genes
The exon-intron structures of all orchid AREB/ABF genes were analyzed using the Gene Structure Display Server (GSDS) program by comparing the coding sequences (CDS) and the genomic sequences [70]. The conserved motifs in AREB/ABF proteins were searched using Multiple Em for Motif Elicitation (MEME) v5.5.5) (http://meme-suite.org/tools/meme), and motifs were further analyzed using the InterPro database (https://www.ebi.ac.uk/interpro/). To identify potential cis-elements in the promoters of AREB/ABF genes, 2000bp sequences upstream of the coding regions of the AREB/ABF genes were first obtained using the TBtools-II [71], and then submitted to the PlantCARE database (http://bioinformatics.psb. ugent.be/webtools/plantcare/html/).
4.3. TFs regulatory network analysis
The Plant Transcriptional Regulatory Map (PTRM) (http://plantregmap.gao-lab.org/) was used to predict potential regulatory interactions of TFs in the upstream (2000bp) regions of orchid AREB/ABFs with threshold (P-value ≤1e−7). The Arabidopsis was the selected plant species. The heat map, wordcloud, and Venn were constructed using TBtools-II [71].
4.4. Expression Profiles of AREB/ABF Genes in Different Tissues
The expression levels of AREB/ABF genes from D. catenatum, P. equestris, and A. shenzhenica were estimated according to the published RNA sequencing data OrchidBase 4.0 [72,73]. The expression levels of AREB/ABF genes from Arabidopsis were derived from The Bio-Analytic Resource for Plant Biology (https://www.bar.utoronto.ca/). The expression heat map and cluster analyses were constructed using TBtools-II [71].
4.5. Plant Materials and Treatments
The wild-type Arabidopsis thaliana Col-0 accession was used in this study. The mutant used in this study was abi5 (SALK_013163) as previous reported [74]. The plant growth was carried out in a culture room at 22oC over a long-day photoperiod (16 h:8 h; light:dark, respectively). For assessment of phenotype, seeds of each genotype were washed twice with distilled water, sterilized with 75% ethanol for 10 min, washed more than five times with sterile water. The sterilized seeds were sown one by one on half MS medium plates (with 1% sucrose and 1.5% agar, pH 5.8) or half MS medium plates supplemented with 0.5 μM ABA, and left at 4oC for 2 days before transfer to the growth chamber. Seeds germination (emergence of radicles) was scored and photographed after 3 days of stratification, and cotyledon greening was recorded 7 days after stratification.
4.6. Vectors Construction and Plant Transformation
For subcellular localization, the coding region of DcaABI5 was cloned into the modified gateway-compatible binary vector pGWB414. All binary vectors were sequenced and introduced into Agrobacterium tumefaciens (A. tumefaciens) strain GV3101 cells. Then A. tumefaciens cells containing each construct were prepared to an OD600 of 0.8, and then injected into N. benthamiana leaves. The transient expression of the fusion protein was examined using a confocal laser-scanning microscope (Leica SP8; Leica Microsystems GmbH, Solms, Germany) at 48 h after transformation. Transgenic Arabidopsis plants were generated with the Agrobacterium tumefaciens-mediated floral dip method [75]. All transgenic lines used in this study were homozygous T3 lines. The primers for the vector construction are listed in Table S2.
4.7. Analysis of Gene Expression with RT-qPCR Analysis
The total RNA extraction from the indicated tissues and organs, first-strand cDNA, and RT-qPCR assay were performed following our previous study [76]. The transcript data were calculated with 2-ΔΔCt to quantify the gene expression levels [77]. The Actin2 from Arabidopsis and DcaActin7 from D. catenatum were used as the internal controls. Each experiment was performed with three replicates. The primers for the RT-qPCR are listed in Table S2.
5. Conclusions
The AREB/ABF genes are essential to ABA signaling pathways and plant adaptation to various environmental stresses. Nevertheless, there is no report on AREB/ABFs in orchids. In this study, twelve AREB/ABF genes were identified within three orchids’ genome (D. catenatum, A. shenzhenic, and P. equestris). and classified into three groups via a phylogenetic analysis. The cis-elements analysis suggested that AREB/ABFs were widely involved in hormone response, including ABA, JA and ethylene. The orchid AREB/ABF-mediated regulatory network constructed through cis-regulatory elements (CREs) analysis revealed that ethylene response factor (ERF) gene family was the most abundant as the potential regulators. Expression profile analysis based on public transcriptomic data showed that most AREB/ABF genes have distinct tissue-specific expression patterns in orchid plants. Notably, the representative AREB/ABF member ABI5 from orchid species was found specifically expressed in pollinium. Additionally, overexpression of ABI5 from D. catenatum conferred ABA sensitivity and rescued the ABA deficient mutant abi5 in Arabidopsis. Collectively, our results provide valuable information on AREB/ABF genes and provide a perspective for further functional characterization of potentially important ones in orchids.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: The statistics of predicted TFs binding to AREB/ABFs in orchids. Figure S2: Expression patterns analysis for AREB/ABFs from Arabidopsis. The heat map was constructed from the transcriptome data using TBtools-II with the log2-transformed RPKM values of each gene. The expression level was shown in color as the scale; Table S1: List of TFs binding to AREB/ABFs in orchids; Table S2: List of the primers used in this study.
Author Contributions
Conceptualization, Zhiyong Li; Data curation, Miaoyan Lin and Zhiyong Li; Formal analysis, Xi Xie and Zhiyong Li; Funding acquisition, Xi Xie and Zhiyong Li; Investigation, Xi Xie, Miaoyan Lin and Zhiyong Li; Methodology, Xi Xie and Miaoyan Lin; Project administration, Gengsheng Xiao and Qin Wang; Resources, Miaoyan Lin and Zhiyong Li; Software, Xi Xie, Miaoyan Lin and Zhiyong Li; Supervision, Gengsheng Xiao, Qin Wang and Zhiyong Li; Validation, Xi Xie and Zhiyong Li; Visualization, Xi Xie; Writing – original draft, Xi Xie and Zhiyong Li; Writing – review & editing, Gengsheng Xiao, Qin Wang and Zhiyong Li. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: The project was supported by the Guangdong Provincial Key Laboratory of Lingnan Specialty Food Science and Technology (2021B1212040013) and the National Natural Science Foundation of China (42106130).
Data Availability Statement
Publicly available datasets were analyzed in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fay, M.F.; Chase, M.W. Orchid biology: From Linnaeus via Darwin to the 21st century. Preface. Ann. Bot. 2009, 104, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Chase, M.W.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Salazar, G.; van den Berg, C.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef]
- Kindlmann, P.; Kull, T.; McCormick, M. The Distribution and Diversity of Orchids. Diversity 2023, 15, 810. [Google Scholar] [CrossRef]
- Mérillon, J.M.; Kodja, H. Orchids Phytochemistry, Biology and Horticulture; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Zhang, D.; Zhao, X.; Li, Y.; Ke, S.; Yin, W.; Lan, S.; Liu, Z. Advances and prospects of orchid research and industrialization. Hortic. Res. 2022, 9, uhac220. [Google Scholar] [CrossRef]
- Liaqat, F.; Xu, L.; Khazi, M.I.; Ali, S.; Rahman, M.U.; Zhu, D. Extraction, purification, and applications of vanillin: A review of recent advances and challenges. Ind. Crop. Prod. 2023, 204, 117372. [Google Scholar] [CrossRef]
- Das, P.; Chandra, T.; Negi, A.; Jaiswal, S.; Iquebal, M.A.; Rai, A.; Kumar, D. A comprehensive review on genomic resources in medicinally and industrially important major spices for future breeding programs: Status, utility and challenges. Curr. Res. Food Sci. 2023, 7, 100579. [Google Scholar] [CrossRef]
- Zotz, G.; Winkler, U. Aerial Roots of Epiphytic Orchids: The Velamen Radicum and its Role in Water and Nutrient Uptake. Oecologia 2013, 171, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Cai, J., Liu, X., Vanneste, K., Proost, S., Tsai, W. C., Liu, K. W., Chen, L. J., He, Y., Xu, Q., Bian, C., et al. The genome sequence of the orchid Phalaenopsis equestris. Nat. Genet. 2015, 47, 65–72.
- Zhang, G.Q., Xu, Q., Bian, C., Tsai, W.C., Yeh, C.M., Liu, K.W., Yoshida, K., Zhang, L.S., Chang, S.B., Chen, F., et al. The Dendrobium catenatum Lindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution. Sci. Rep. 2016, 6, 19029.
- Zhang, G.Q., Liu, K.W., Li, Z., Lohaus, R., Hsiao, Y.Y., Niu, S.C., Wang, J. Y., Lin, Y. C., Xu, Q., Chen, L.J., et al. The Apostasia genome and the evolution of orchids. Nature 2017, 549, 379–383.
- Chao, Y. T., Chen, W. C., Chen, C. Y., Ho, H. Y., Yeh, C. H., Kuo, Y. T., Su, C. L., Yen, S. H., Hsueh, H. Y., Yeh, J. H., et al. Chromosome-level assembly, genetic and physical mapping of Phalaenopsis aphrodite genome provides new insights into species adaptation and resources for orchid breeding. Plant Biotechnol. J. 2018, 16, 2027–2041.
- Raghavendra, A.S., Gonugunta, V.K., Christmann, A., Grill, E. ABA perception and signalling. Trends Plant Sci. 2010, 15, 395–401.
- Nakashima, K., Yamaguchi-Shinozaki, K. ABA signaling in stress-response and seed development. Plant Cell Reports 2013, 32, 959–70. [CrossRef]
- Fujita, Y., Fujita, M., Shinozaki, K., Yamaguchi-Shinozaki, K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. J. Plant Res. 2011, 124, 509–25. [CrossRef]
- Park, S. Y., Fung, P., Nishimura, N., Jensen, D. R., Fujii, H., Zhao, Y., Lumba, S., Santiago, J., Rodrigues, A., Chow, T. F., et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009, 324, 1068–71. [CrossRef] [PubMed]
- Ma, Y., Szostkiewicz, I., Korte, A., Moes, D., Yang, Y., Christmann, A., Grill, E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 2009, 324, 1064-8.
- Fujita, Y.; Yoshida, T.; Yamaguchi-Shinozaki, K. Pivotal role of the AREB/ABF-SnRK2 pathway in ABRE-mediated transcription in response to osmotic stress in plants. Physiol. Plant 2013, 147, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T. , Fujita, Y., Sayama, H., Kidokoro, S., Maruyama, K., Mizoi, J., Shinozaki, K., Yamaguchi-Shinozaki, K.. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–85. [Google Scholar]
- Qin, F., Sakuma, Y., Tran, L. S., Maruyama, K., Kidokoro, S., Fujita, Y., Fujita, M., Umezawa, T., Sawano, Y., Miyazono, K., et al. Arabidopsis DREB2A-interacting proteins function as RING E3 ligases and negatively regulate plant drought stress-responsive gene expression. Plant Cell 2008, 20, 1693–707.
- Jakoby, M., Weisshaar, B., Dröge-Laser, W., Vicente-Carbajosa, J., Tiedemann, J., Kroj, T., Parcy, F., bZIP Research Group. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–11.
- Kang, J.Y., Choi, H.I., Im, M.Y., Kim, S.Y. Arabidopsis basic leucine zipper proteins that mediate stress-responsive abscisic acid signaling. Plant Cell 2002, 14, 343–57. [CrossRef] [PubMed]
- Landschulz, W.; Johnson, P.; McKnight, S. The leucine zipper: a hypothetical structure common to a new class of DNA binding proteins. Science 1998, 240, 1759–64. [Google Scholar] [CrossRef] [PubMed]
- Foster, R., Izawa, T., Chua, N. Plant bZIP proteins gather at ACGT elements. FASEB J. 1994, 8, 192. [CrossRef] [PubMed]
- Siberil, Y., Doireau, P., Gantet, P. Plant bZIP G-box binding factors. Modular structure and activation mechanisms. Eur. J. Biochem. 2001, 268, 5655–5666. [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K., Shinozaki, K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 1994, 6, 251–64.
- Choi, H., Hong, J., Ha, J., Kang, J., Kim, S.Y. ABFs, a family of ABA-responsive element binding factors. J. Biol. Chem. 2000, 275, 1723–30. [CrossRef]
- Busk, P.K., Pages, M. Regulation of abscisic acid-induced transcription. Plant Mol. Biol. 1998, 37, 425–35. [CrossRef] [PubMed]
- Koornneef, M., Leon-Kloosterziel, K.M., Schwartz, S.H., Zeevaart, J.A.D. The genetic and molecular dissection of abscisic acid biosynthesis and signal transduction in Arabidopsis. Plant Physiol. Biochem. 1998, 36, 83–9. [CrossRef]
- Shinozaki, K., Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 2000, 3, 217–23. [CrossRef]
- Yoshida, T., Fujita, Y., Sayama, H., Kidokoro, S., Maruyama, K., Mizoi, J., Shinozaki, K., Yamaguchi-Shinozaki, K. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–685.
- Miyazono, K.; Koura, T.; Kubota, K.; Yoshida, T.; Fujita, Y.; Yamaguchi-Shinozaki, K.; Tanokura, M. Purification, crystallization and preliminary X-ray analysis of OsAREB8 from rice, a member of the AREB/ABF family of bZIP transcription factors, in complex with its cognate DNA. Acta Crystallogr. Sect. F. Struct. Biol. Cryst. Commun. 2012, 68, 491–494. [Google Scholar] [CrossRef]
- Rikiishi, K.; Matsuura, T.; Maekawa, M. TaABF1, ABA response element binding factor 1, is related to seed dormancy and ABA sensitivity in wheat (Triticum aestivum L.) seeds. J. Cereal Sci. 2010, 52, 236–238. [Google Scholar] [CrossRef]
- Wang, J., Li, Q., Mao, X., Li, A., Jing, R. Wheat Transcription Factor TaAREB3 Participates in Drought and Freezing Tolerances in Arabidopsis. Int. J. Biol. Sci. 2016, 12, 257–69. [CrossRef]
- Li, F., Mei, F., Zhang, Y., Li, S., Kang, Z., Mao, H. Genome-wide analysis of the AREB/ABF gene lineage in land plants and functional analysis of TaABF3 in Arabidopsis. BMC Plant Biol. 2020, 10, 558.
- Liu, T.; Zhou, T.; Lian, M.; Liu, T.; Hou, J.; Ijaz, R.; Song, B. Genome-wide identification and characterization of the AREB/ABF/ABI5 subfamily members from Solanum tuberosum. Int. J. Mol. Sci. 2019, 20, 31135. [Google Scholar] [CrossRef]
- Wang, W.; Qiu, X.; Yang, Y.; Kim, H.S.; Jia, X.; Yu, H.; Kwak, S. Sweet potato bZIP transcription factor IbABF4 confers tolerance to multiple abiotic stresses. Front. Plant Sci. 2019, 10, 630. [Google Scholar] [CrossRef] [PubMed]
- Kerr, T.C.; Abdel-Mageed, H.; Aleman, L.; Lee, J.; Payton, P.; Cryer, D.; Allen, R.D. Ectopic expression of two AREB/ABF orthologs increases drought tolerance in cotton (Gossypium hirsutum). Plant Cell Environ. 2018, 41, 898–907. [Google Scholar] [CrossRef]
- Ma, Q.J.; Sun, M.H.; Lu, J.; Liu, Y.J.; You, C.X.; Hao, Y.J. An apple CIPK protein kinase targets a novel residue of AREB transcription factor for ABA-dependent phosphorylation. Plant Cell Environ. 2017, 40, 2207–2219. [Google Scholar] [CrossRef]
- Li, D.; Mou, W.; Luo, Z.; Li, L.; Limwachiranon, J.; Mao, L.; Ying, T. Developmental and stress regulation on expression of a novel miRNA, Fan-miR73 and its target ABI5 in strawberry. Sci. Rep. 2016, 6, 28385. [Google Scholar] [CrossRef]
- Yong, X.; Zheng, T.; Zhuo, X.; Ahmad, S.; Li, L.; Li, P.; Yu, J.; Wang, J.; Cheng, T.; Zhang, Q. Genome-wide identification, characterization, and evolution of ABF/AREB subfamily in nine Rosaceae species and expression analysis in mei (Prunus mume). PeerJ 2021, 9, e10785. [Google Scholar] [CrossRef]
- eng, Z., Lyu, T., & Lyu, Y. LoSWEET14, a Sugar Transporter in Lily, Is Regulated by Transcription Factor LoABF2 to Participate in the ABA Signaling Pathway and Enhance Tolerance to Multiple Abiotic Stresses in Tobacco. Int. J. Mol. Sci. 2022, 23, 15093.
- Jin, M., Gan, S., Jiao, J., He, Y., Liu, H., Yin, X., Zhu, Q., Rao, J. Genome-wide analysis of the bZIP gene family and the role of AchnABF1 from postharvest kiwifruit (Actinidia chinensis cv. Hongyang) in osmotic and freezing stress adaptations. Plant Sci. 2021, 308, 110927. [CrossRef]
- Fiallos-Salguero, M. S., Li, J., Li, Y., Xu, J., Fang, P., Wang, Y., Zhang, L., Tao, A.. Identification of AREB/ABF Gene Family Involved in the Response of ABA under Salt and Drought Stresses in Jute (Corchorus olitorius L.). Plants (Basel) 2023, 12, 1161.
- Uno, Y., Furihata, T., Abe, H., Yoshida, R., Shinozaki, K., Yamaguchi-Shinozaki, K. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc. Natl. Acad. Sci. U. S. A. 2000, 97, 11632-7. [CrossRef] [PubMed]
- Haake, V., Cook, D., Riechmann, J.L., Pineda, O., Thomashow, M.F., Zhang, J.Z. Transcription factor CBF4 is a regulator of drought adaptation in Arabidopsis. Plant Physiol. 2002, 130, 639–48. [CrossRef]
- Lee, B.H., Henderson, D.A., Zhu, J.K. The Arabidopsis cold-responsive transcriptome and its regulation by ICE1. Plant Cell 2005, 17, 3155–75. [CrossRef] [PubMed]
- Vonapartis, E., Mohamed, D., Li, J., Pan, W., Wu, J., Gazzarrini, S. CBF4/DREB1D represses XERICO to attenuate ABA, osmotic and drought stress responses in Arabidopsis. Plant J. 2022, 110, 961–977. [CrossRef] [PubMed]
- Li, Z., Luo, X., Wang, L., Shu, K. ABSCISIC ACID INSENSITIVE 5 mediates light-ABA/gibberellin crosstalk networks during seed germination. J. Exp. Bot. 2022, 73, 4674–4682. [CrossRef]
- Carles, C., Bies-Etheve, N., Aspart, L., Léon-Kloosterziel, K. M., Koornneef, M., Echeverria, M., Delseny, M.. Regulation of Arabidopsis thaliana Em genes: role of ABI5. Plant J. 2002, 30, 373–83.
- Finkelstein, R.R., Lynch, T.J. The Arabidopsis abscisic acid response gene ABI5 encodes a basic leucine zipper transcription factor. Plant Cell 2000, 12, 599–609. [CrossRef]
- Raghavendra, A.S., Gonugunta, V.K., Christmann, A., Grill, E. ABA perception and signalling. Trends Plant Sci. 2010, 15, 395–401.
- Lee, S.C., Luan, S. ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant Cell Environ. 2012, 35, 53–60. [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Yuan, Y., Jin, X., Liu, J., Zhao, X., Zhou, J., Wang, X., et al. (2018). The gastrodia elata genome provides insights into plant adaptation to heterotrophy. Nat. Commun. 9, 1–11.
- Zhang, Y., Zhang, G. Q., Zhang, D., Liu, X. D., Xu, X. Y., Sun, W. H., et al. Chromosome-scale assembly of the dendrobium chrysotoxum genome enhances the understanding of orchid evolution. Hortic. Res. 2021, 8.
- Chao, Y. T., Chen, W. C., Chen, C. Y., Ho, H. Y., Yeh, C. H., Kuo, Y. T., et al. Chromosome-level assembly, genetic and physical mapping of phalaenopsis aphrodite genome provides new insights into species adaptation and resources for orchid breeding. Plant Biotechnol. J. 2018, 16, 2027–2041. [CrossRef]
- Fujita, Y., Nakashima, K., Yoshida, T., Katagiri, T., Kidokoro, S., Kanamori, N., Umezawa, T., Fujita, M., Maruyama, K., Ishiyama, K., et al. Three SnRK2 protein kinases are the main positive regulators of abscisic acid signaling in response to water stress in Arabidopsis. Plant Cell Physiol. 2009, 50, 2123–32.
- Lynch, T., Erickson, B.J. Finkelstein, R.R. Direct interactions of ABA-insensitive(ABI)-clade protein phosphatase(PP)2Cs with calcium-dependent protein kinases and ABA response element-binding bZIPs may contribute to turning off ABA response. Plant Mol. Biol. 2012, 80, 647–658. [CrossRef]
- Chen, Y.T., Liu, H., Stone, S., Callis, J. ABA and the ubiquitin E3 ligase KEEP ON GOING affect proteolysis of the Arabidopsis thaliana transcription factors ABF1 and ABF3. Plant J. 2013, 75, 965–76. [CrossRef]
- Vysotskii, D. A., de Vries-van Leeuwen, I. J., Souer, E., Babakov, A. V., de Boer, A. H.. ABF transcription factors of Thellungiella salsuginea: Structure, expression profiles and interaction with 14-3-3 regulatory proteins. Plant Signal Behav. 2013, 8, e22672. [CrossRef]
- Yu, Z., Duan, X., Luo, L., Dai, S., Ding, Z., Xia, G. How Plant Hormones Mediate Salt Stress Responses. Trends Plant Sci. 2020, 25, 1117–1130. [CrossRef]
- Li, C., Xu, M., Cai, X., Han, Z., Si, J., Chen, D. Jasmonate Signaling Pathway Modulates Plant Defense, Growth, and Their Trade-Offs. Int. J. Mol. Sci. 2022, 23, 3945. [CrossRef] [PubMed]
- Li, M., Yu, G., Cao, C., Liu, P. Metabolism, signaling, and transport of jasmonates. Plant Commun. 2021, 2, 100231. [CrossRef] [PubMed]
- Kou, X., Zhou, J., Wu, C.E., Yang, S., Liu, Y., Chai, L., Xue, Z. The interplay between ABA/ethylene and NAC TFs in tomato fruit ripening: a review. Plant Mol. Biol. 2021, 106, 223–238. [CrossRef] [PubMed]
- Müller, M. Foes or Friends: ABA and Ethylene Interaction under Abiotic Stress. Plants (Basel) 2021, 10, 448. [Google Scholar] [CrossRef] [PubMed]
- Huang, G., Kilic, A., Karady, M., Zhang, J., Mehra, P., Song, X., Sturrock, C. J., Zhu, W., Qin, H., Hartman, S., et al. Ethylene inhibits rice root elongation in compacted soil via ABA- and auxin-mediated mechanisms. Proc. Natl. Acad. Sci. U. S. A. 2022, 119, e2201072119. [CrossRef] [PubMed]
- Li, Z., Sheerin, D.J., von Roepenack-Lahaye E., Stahl, M., Hiltbrunner, A. The phytochrome interacting proteins ERF55 and ERF58 repress light-induced seed germination in Arabidopsis thaliana. Nat Commun. 2022, 13, 1656. [CrossRef] [PubMed]
- Hu, B., Jin, J., Guo, A.Y., Zhang, H., Luo, J., Gao, G. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296-7. [CrossRef]
- Chen, C., Wu, Y., Li, J., Wang, X., Zeng, Z., Xu, J., Liu, Y., Feng, J., Chen, H., He, Y., Xia, R. TBtools-II: A "one for all, all for one" bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [CrossRef]
- Fu, C. H., Chen, Y. W., Hsiao, Y. Y., Pan, Z. J., Liu, Z. J., Huang, Y. M., Tsai, W. C., Chen, H. H. OrchidBase: a collection of sequences of the transcriptome derived from orchids. Plant Cell Physiol. 2011, 52, 238–243. [CrossRef]
- Tsai, W. C., Fu, C. H., Hsiao, Y. Y., Huang, Y. M., Chen, L. J., Wang, M., Liu, Z. J., Chen, H. H. OrchidBase 2.0: comprehensive collection of Orchidaceae floral transcriptomes. Plant Cell Physiol. 2013, 54, e7. [CrossRef] [PubMed]
- Guo, C., Jiang, Y., Shi, M., Wu, X., Wu, G. ABI5 acts downstream of miR159 to delay vegetative phase change in Arabidopsis. New Phytol. 2021, 231, 339–350. [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Fu, Y.; Wang, Y.; Liang, J. Scaffold protein RACK1 regulates BR signaling by modulating the nuclear localization of BZR1. New Phytol. 2023, 239, 1804–1818. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Multiple sequence alignment of AREB/ABF members from D. catenatum, A. shenzhenica, and P. equestris. The positions of C1 to C4 are conserved domains and basic regions are represented with different colors above the protein sequences. Potential phosphorylated residues (R-S-SX/T) of the characteristic phosphorylation sites are indicated by black boxes.
Figure 1.
Multiple sequence alignment of AREB/ABF members from D. catenatum, A. shenzhenica, and P. equestris. The positions of C1 to C4 are conserved domains and basic regions are represented with different colors above the protein sequences. Potential phosphorylated residues (R-S-SX/T) of the characteristic phosphorylation sites are indicated by black boxes.
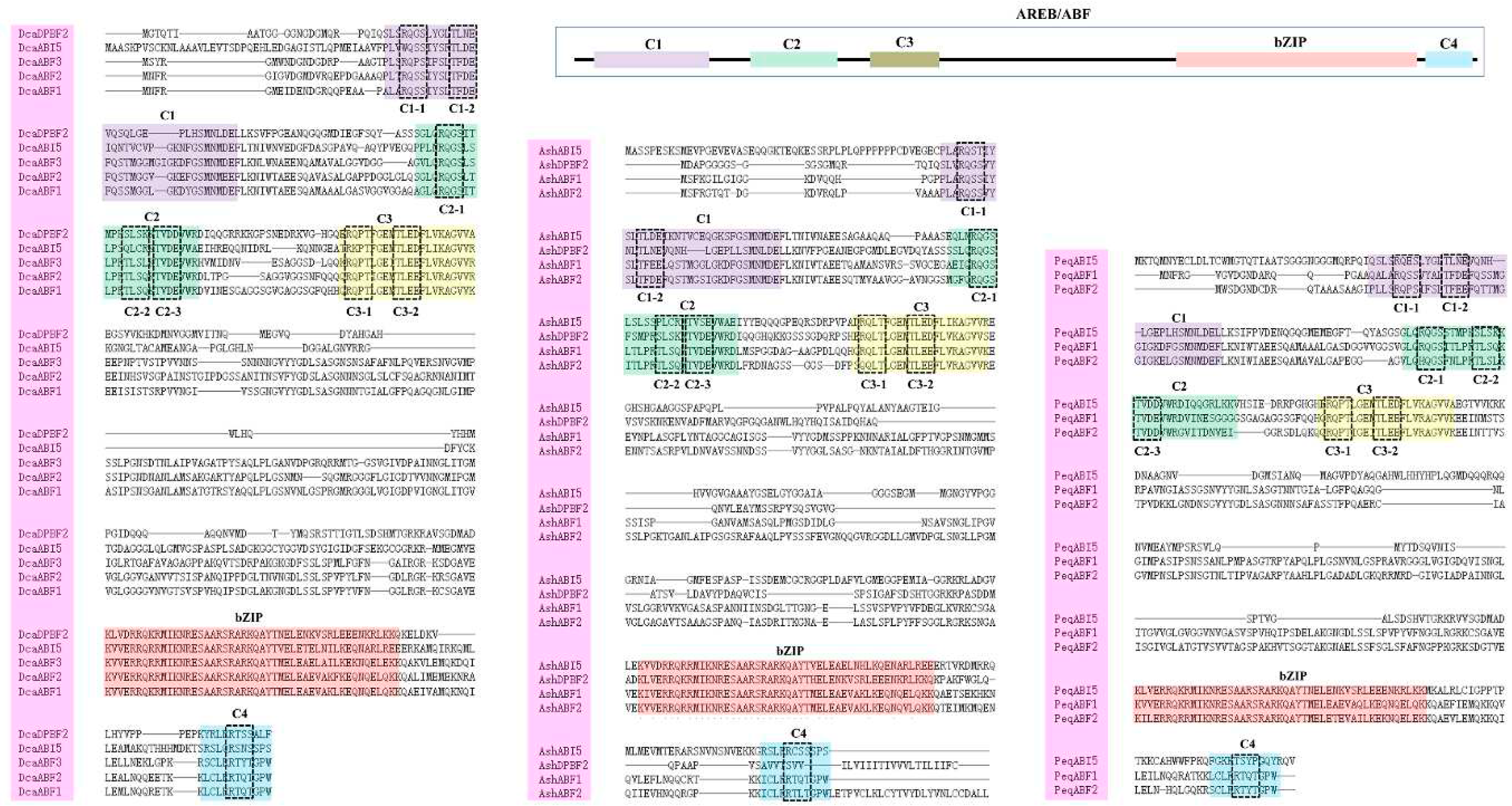
Figure 2.
Phylogenetic analysis and family classification of the AREB/ABF proteins. The different colored arcs indicate the diverse groups of the AREB/ABF proteins. Protein sequences from D. catenatum, A. shenzhenica, P. equestris, rice (Oryza sativa), and Arabidopsis are indicated by black stars, blue triangle, yellow squares, purple circles, and red stars, respectively.
Figure 2.
Phylogenetic analysis and family classification of the AREB/ABF proteins. The different colored arcs indicate the diverse groups of the AREB/ABF proteins. Protein sequences from D. catenatum, A. shenzhenica, P. equestris, rice (Oryza sativa), and Arabidopsis are indicated by black stars, blue triangle, yellow squares, purple circles, and red stars, respectively.
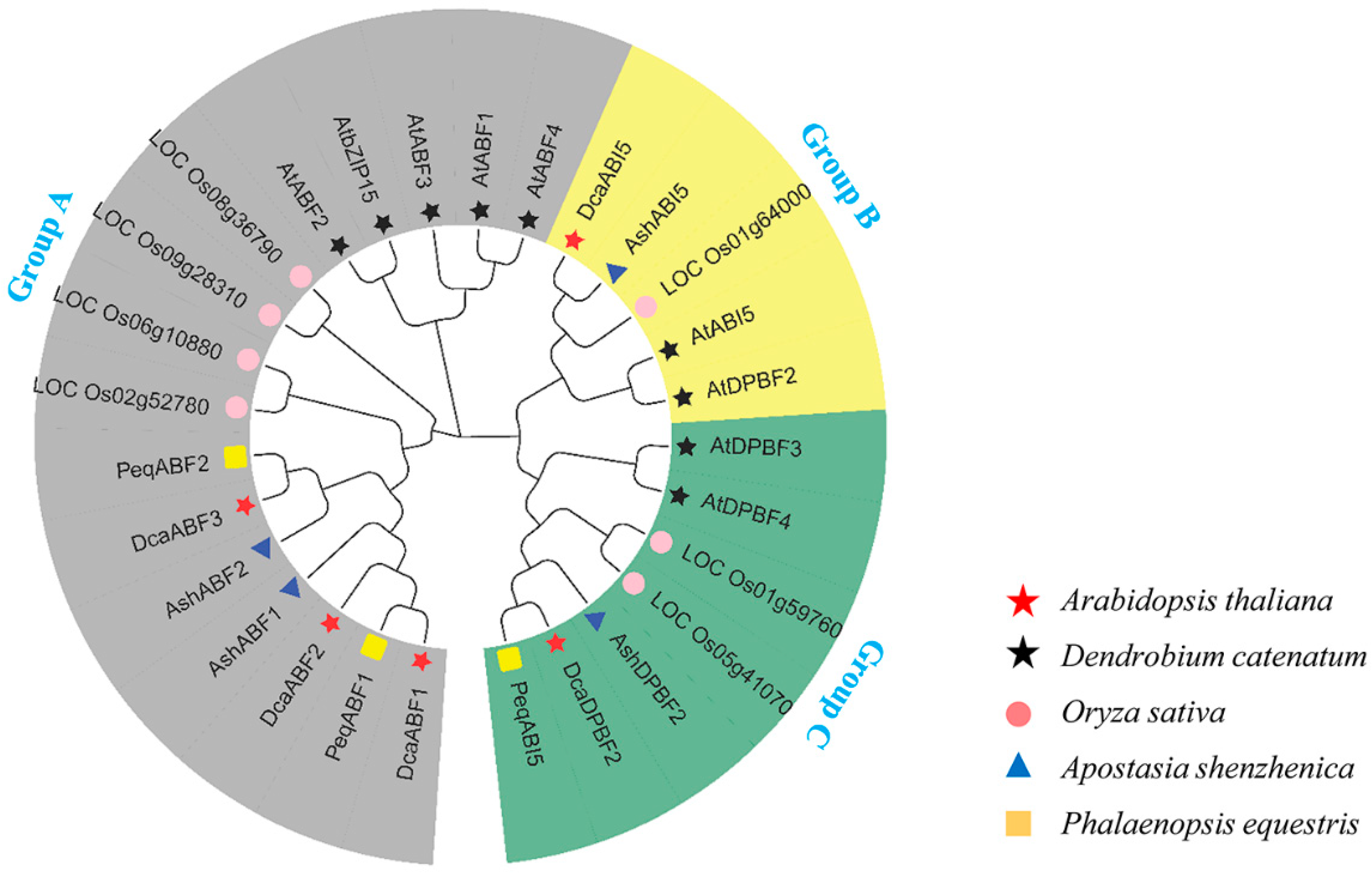
Figure 3.
Architecture of conserved motifs and gene structures of AREB/ABFs. (A) The neighbor-joining phylogenetic tree was produced by MEGA using the neighbor-joining method with 1000 bootstrap replicates. Schematic represent the conserved motifs of the AREB/ABFs identified by MEME. Each motif is indicated by a colored box numbered and sequences. (B) Intron/exon structures of AREB/ABF genes. Exon(s), intron(s), and UTR(s) are represented by yellow boxes, black lines, and blue boxes, respectively.
Figure 3.
Architecture of conserved motifs and gene structures of AREB/ABFs. (A) The neighbor-joining phylogenetic tree was produced by MEGA using the neighbor-joining method with 1000 bootstrap replicates. Schematic represent the conserved motifs of the AREB/ABFs identified by MEME. Each motif is indicated by a colored box numbered and sequences. (B) Intron/exon structures of AREB/ABF genes. Exon(s), intron(s), and UTR(s) are represented by yellow boxes, black lines, and blue boxes, respectively.

Figure 4.
Analysis of cis-elements in the promoter regions of AREB/ABF genes from orchids. (A) The different colored blocks represent the different types of cis-elements and their locations in each AREB/ABF gene. (B) Evaluation of cis-elements of each AREB/ABF gene. The number of the individual elements is indicated with the colorful circle.
Figure 4.
Analysis of cis-elements in the promoter regions of AREB/ABF genes from orchids. (A) The different colored blocks represent the different types of cis-elements and their locations in each AREB/ABF gene. (B) Evaluation of cis-elements of each AREB/ABF gene. The number of the individual elements is indicated with the colorful circle.
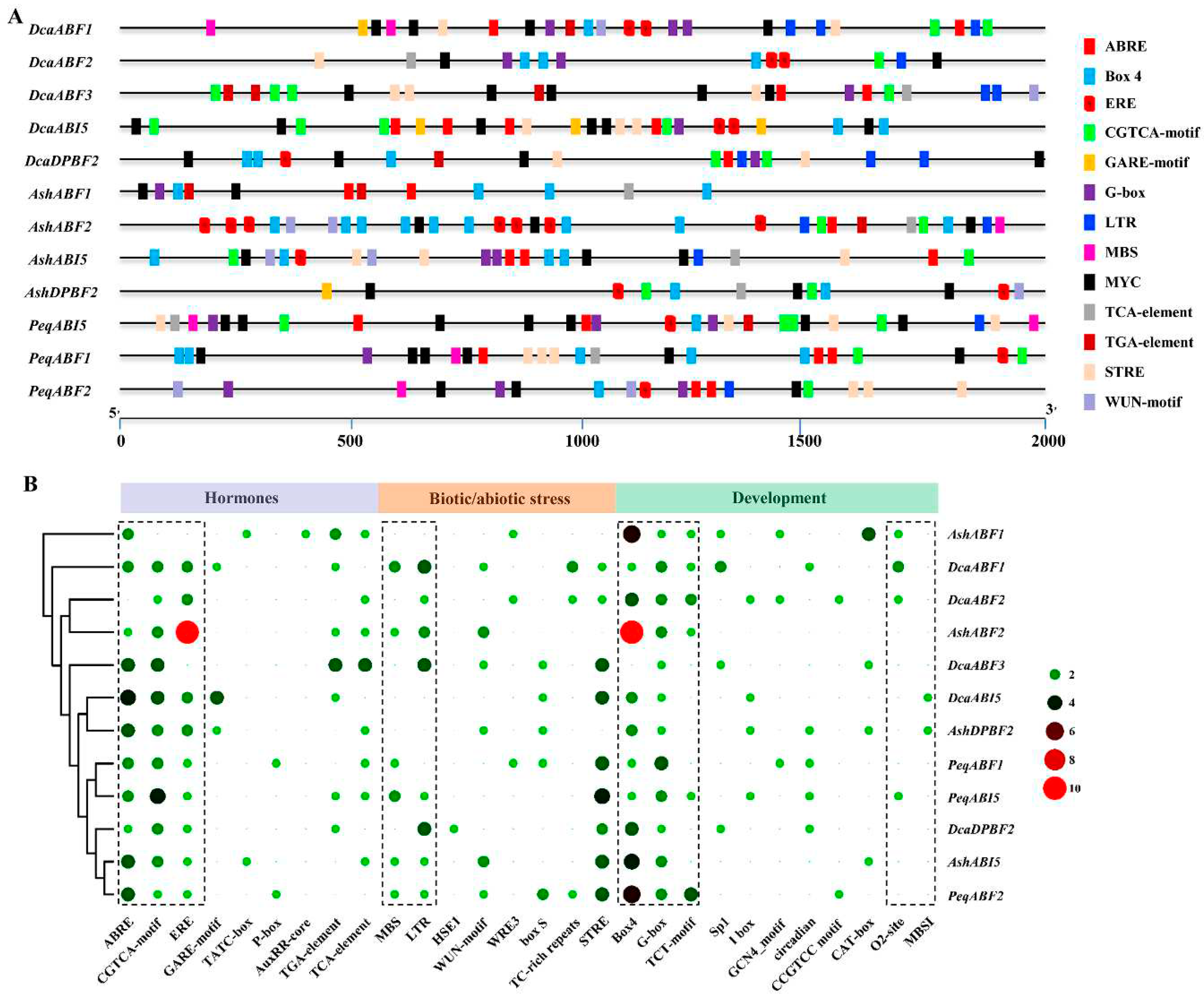
Figure 5.
The putative TFs regulatory network analysis of AREB/ABFs from orchid plants. (A) An overview of the cluster of enriched TF regulators for all AREB/ABF genes. The highly enriched TFs were indicated with dash boxes. (B) The distribution of TF regulators for individual AREB/ABF gene with pie chart and wordcloud. The font size is positively correlated with the number of corresponding TF regulators. (C) Venn diagram showing the overlapping TF regulators among AREB/ABFs from three orchid genomes.
Figure 5.
The putative TFs regulatory network analysis of AREB/ABFs from orchid plants. (A) An overview of the cluster of enriched TF regulators for all AREB/ABF genes. The highly enriched TFs were indicated with dash boxes. (B) The distribution of TF regulators for individual AREB/ABF gene with pie chart and wordcloud. The font size is positively correlated with the number of corresponding TF regulators. (C) Venn diagram showing the overlapping TF regulators among AREB/ABFs from three orchid genomes.
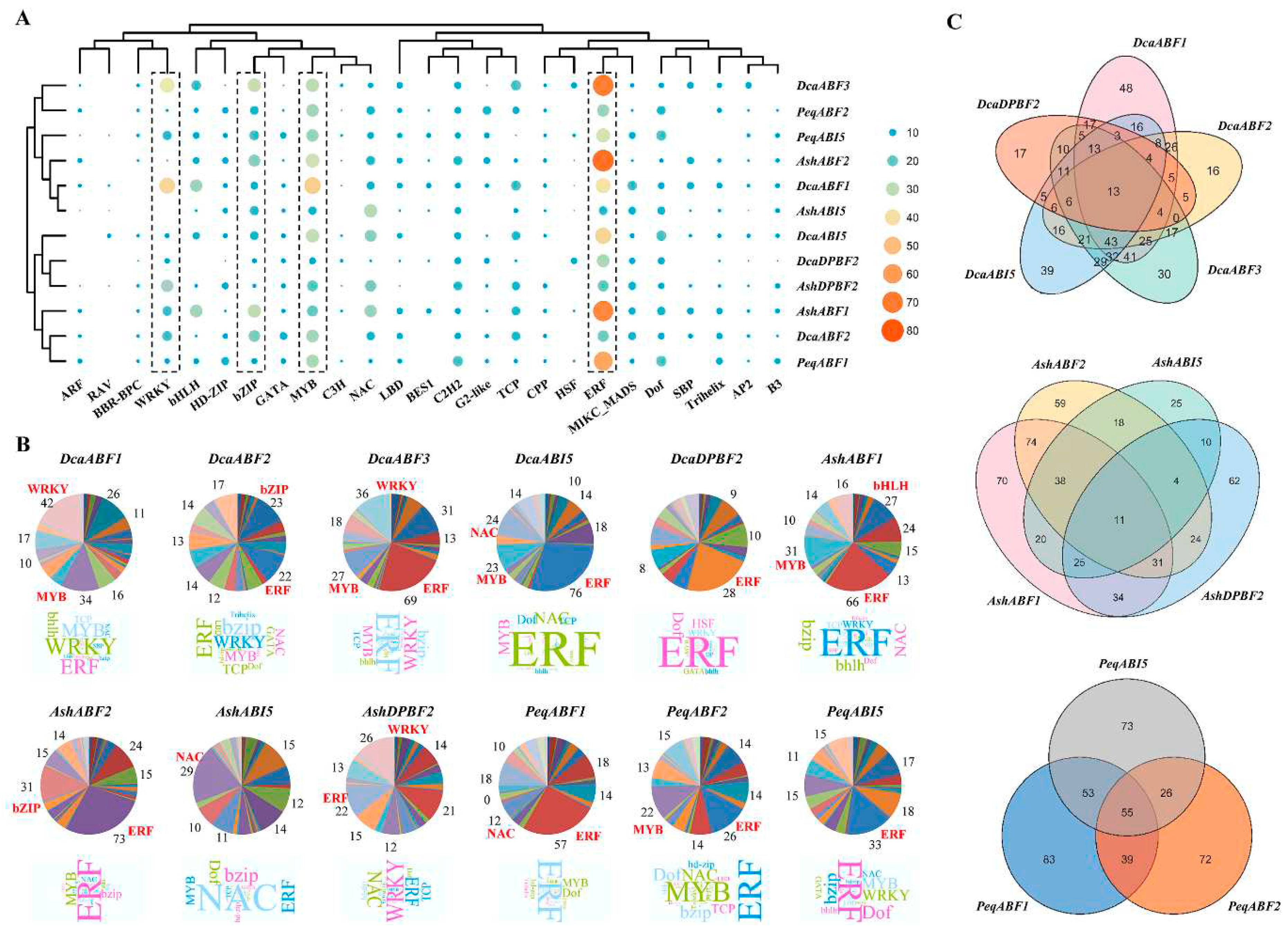
Figure 6.
Expression patterns analysis for orchid AREB/ABF gene family. (A-C) Expression profiles of the AREB/ABF genes in different tissues from indicated orchid species: D. catenatum (A), A. shenzhenica (B), and P. equestris (C). The heat map was constructed from the transcriptome data using TBtools-II with the log2-transformed RPKM values of each gene. The expression level was shown in color as the scale. (D) Expression patterns of DcaABI5 in various tissues. DcaActin7 was used as the control. Three independent biological experiments, each with three technical replicates were performed.
Figure 6.
Expression patterns analysis for orchid AREB/ABF gene family. (A-C) Expression profiles of the AREB/ABF genes in different tissues from indicated orchid species: D. catenatum (A), A. shenzhenica (B), and P. equestris (C). The heat map was constructed from the transcriptome data using TBtools-II with the log2-transformed RPKM values of each gene. The expression level was shown in color as the scale. (D) Expression patterns of DcaABI5 in various tissues. DcaActin7 was used as the control. Three independent biological experiments, each with three technical replicates were performed.
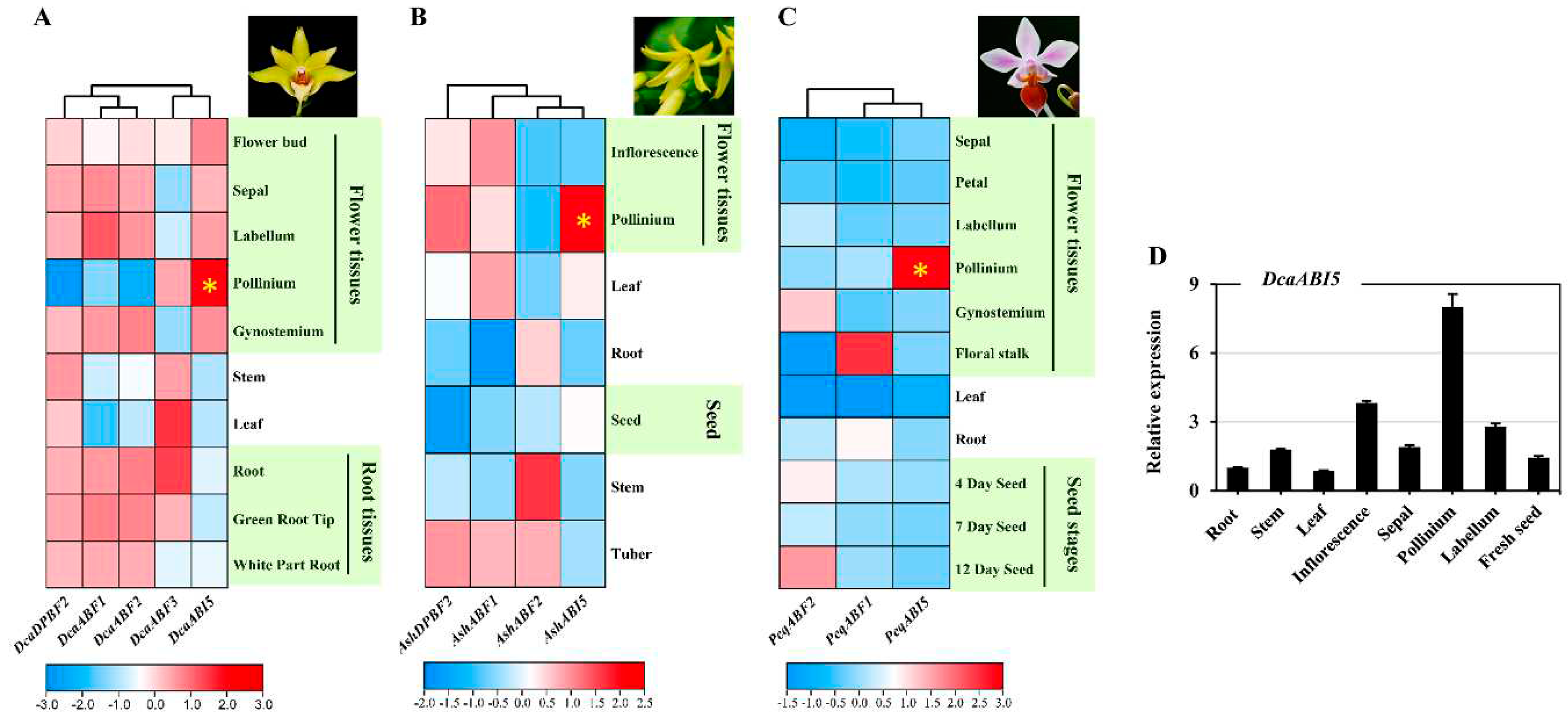
Figure 7.
DcaABI5 is able to rescue ABI5 mutation in Arabidopsis. (A) Confocal microscopy images for the subcellular localization of DcaABI5 in tobacco leaf. (B) Representative images for seedlings of 7-day-old Col-0, abi5, DcaABI5 overexpression lines in WT (DcaABI5OE) and abi5 (DcaABI5OE/abi5) germinated on half MS medium supplemented without or with 0.5 μM ABA. (C) Germination and greening cotyledon percentages of the various genotypes in response to ABA. Seed germination was recorded after 3 days of stratification and cotyledon greening was recorded 7 days after stratification on half MS medium supplemented without or with 0.5 μM ABA. Data indicate mean ± SD (n = 3). Bars with different letters indicate significant differences from the control as determined by one-way ANOVA, P-value <0.05. (D) DcaABI5 regulated expression of stress-responsive genes. RT-qPCR analysis was performed to examine the relative transcript levels of EM1 and RAB18 in indicated plants treated without or with 10 μM ABA. Actin2 was used as the control. Three independent biological experiments were carried out, each with three technical replicates.
Figure 7.
DcaABI5 is able to rescue ABI5 mutation in Arabidopsis. (A) Confocal microscopy images for the subcellular localization of DcaABI5 in tobacco leaf. (B) Representative images for seedlings of 7-day-old Col-0, abi5, DcaABI5 overexpression lines in WT (DcaABI5OE) and abi5 (DcaABI5OE/abi5) germinated on half MS medium supplemented without or with 0.5 μM ABA. (C) Germination and greening cotyledon percentages of the various genotypes in response to ABA. Seed germination was recorded after 3 days of stratification and cotyledon greening was recorded 7 days after stratification on half MS medium supplemented without or with 0.5 μM ABA. Data indicate mean ± SD (n = 3). Bars with different letters indicate significant differences from the control as determined by one-way ANOVA, P-value <0.05. (D) DcaABI5 regulated expression of stress-responsive genes. RT-qPCR analysis was performed to examine the relative transcript levels of EM1 and RAB18 in indicated plants treated without or with 10 μM ABA. Actin2 was used as the control. Three independent biological experiments were carried out, each with three technical replicates.
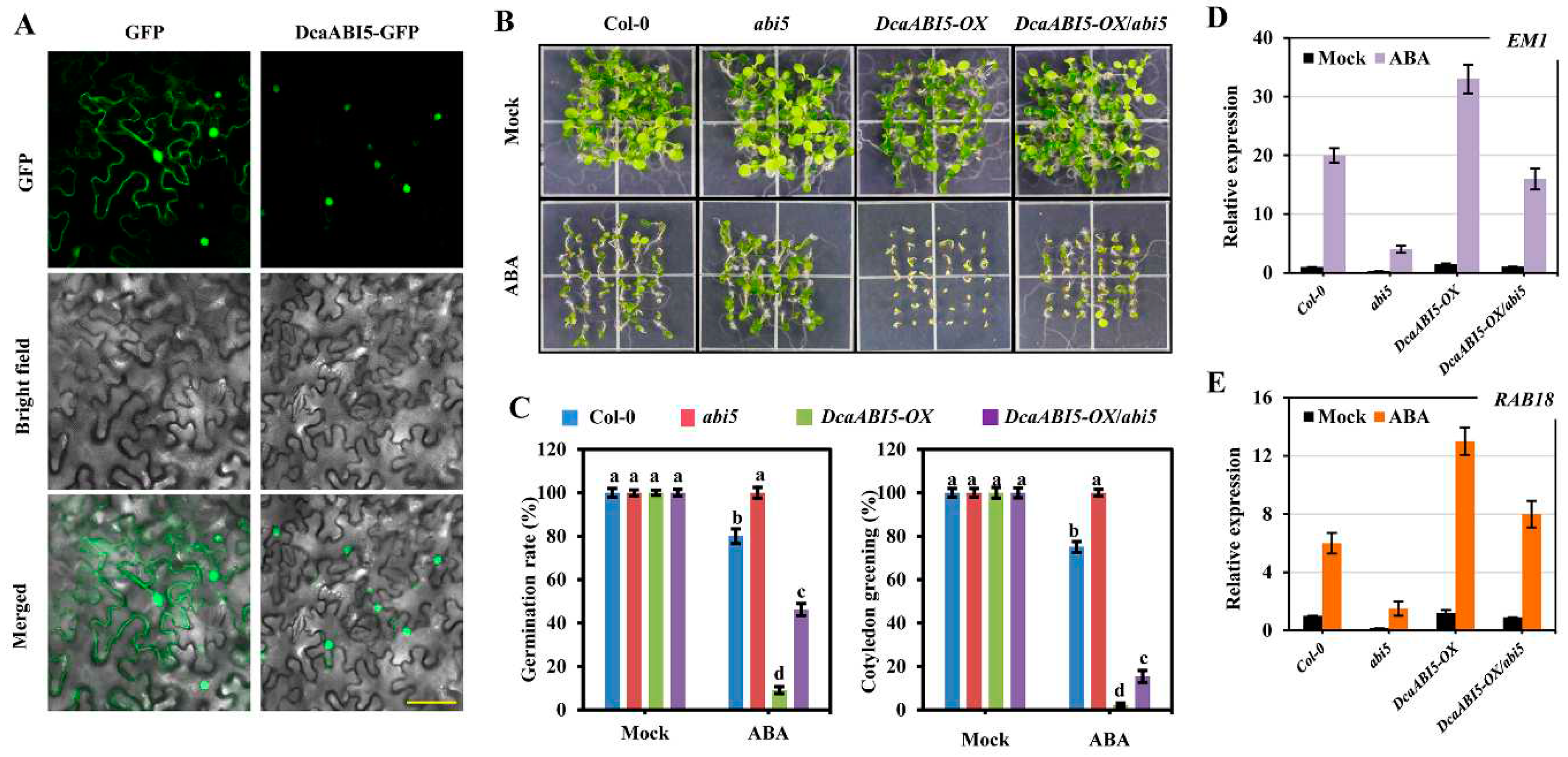

Table 1.
Characteristics of AREB/ABF subfamily members in three orchid species.
| Species | Gene | Gene Id | Coding Sequence (CDS) Length | Protein Length (aa) | Molecular Mass (kDa) | Theoretical pI | Sub-Cellular Location |
| Dendrobium catenatum | DcaABF1 | Dca012913 | 1224 | 407 | 43.50 | 7.7 | Nucleus |
| DcaABF2 | Dca011277 | 1221 | 406 | 42.69 | 9.13 | Nucleus | |
| DcaABF3 | Dca006042 | 1191 | 396 | 42.67 | 9.19 | Nucleus | |
| DcaABI5 | Dca002027 | 1074 | 357 | 38.95 | 7.60 | Nucleus | |
| DcaDPBF2 | Dca016354 | 936 | 311 | 34.83 | 9.50 | Nucleus | |
| Apostasia shenzhenica | AshABF1 | Ash014915 | 1143 | 380 | 40.72 | 8.71 | Nucleus |
| AshABF2 | Ash016767 | 1278 | 425 | 45.56. | 9.00 | Nucleus | |
| AshABI5 | Ash004480 | 1188 | 395 | 42.51 | 5.47 | Nucleus | |
| AshDPBF2 | Ash013161 | 1023 | 340 | 37.18 | 8.74 | Nucleus | |
| Phalaenopsis equestris | PeqABF1 | Peq004088 | 1221 | 406 | 42.60 | 9.39 | Nucleus |
| PeqABF2 | Peq011139 | 1191 | 396 | 42.57. | 8.65 | Nucleus | |
| PeqABI5 | Peq013682 | 1068 | 355 | 39.91 | 9.42 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



