
Submitted:
18 June 2025
Posted:
24 June 2025
You are already at the latest version
Abstract
The advent of CRISPR/Cas systems has revolutionized plant genome engineering, transitioning from traditional single-gene edits to sophisticated multiplex genome editing strategies capable of simultaneously targeting multiple loci. This review provides an in-depth examination of CRISPR-mediated multiplexing technologies in plants, emphasizing their molecular mechanisms, delivery systems, and transformative applications in crop improvement. We delineate the evolution of CRISPR systems from early programmable nucleases to diverse Class 2 effectors, including Cas9, Cas12, Cas13, and emerging ultra-compact variants like CasΦ and Cas14. We detail polycistronic gRNA expression platforms—such as tRNA-sgRNA arrays, ribozymes, and Csy4-mediated cleavage—that enable efficient multi-target editing within compact vectors. Furthermore, we explore advanced delivery modalities including Agrobacterium, biolistics, protoplast transfection, and viral vectors, optimized for recalcitrant plant systems. Applications span yield enhancement, disease resistance, abiotic stress tolerance, nutritional fortification, and de novo domestication. Critical challenges including off-target mutagenesis, mosaicism, chromosomal rearrangements, and regulatory constraints are addressed. Finally, we highlight AI-driven sgRNA design, multi-omics integration, and CRISPR libraries as pivotal tools to rationalize and scale multiplex editing. This synthesis underscores multiplex CRISPR as a cornerstone of next-generation plant breeding, with the potential to redefine global agriculture through precision trait stacking and rapid varietal development.
Keywords:
CRISPR/Cas multiplexing
; plant genome editing
; polygenic trait engineering
; sgRNA expression strategies
; precision crop improvement
1. Introduction
Plant breeding has evolved from slow, traditional cross-hybridization and mutation breeding into the era of precise genome editing [1]. Early techniques like zinc-finger nucleases (ZFNs) and TALENs allowed targeted DNA cuts but required re-engineering a new protein for each target [2]. In contrast, CRISPR/Cas systems use a simple guide RNA for target recognition via base complementarity, vastly simplifying multiplexed genome editing [3]. This ease of reprogramming means that multiple genes can be edited simultaneously by introducing single-guide RNAs (sgRNAs) with a single Cas nuclease [4]. The ability to modify several loci at once is transformative for improving polygenic traits (e.g. yield, stress resistance) that typically require the pyramiding of many alleles across breeding cycles [5]. By inducing multiple heritable mutations in one generation, CRISPR multiplexing can bypass extensive multi-generational crossing, accelerating crop improvement to meet global food demands [6].
CRISPR’s impact on plant genome editing builds upon decades of progress in genome engineering. The first programmable nucleases Zinc Finger Nucleases (ZFNs) in the 1990s and Transcription Activator-Like Effector Nucleases (TALENs) in the 2000s demonstrated that targeted mutagenesis could improve traits, even in complex polyploid crops like sugarcane [7]. However, multiplex editing using TALENs was limited, as each TALEN pair required separate construction. In 2012–2013, the CRISPR/Cas9 system from Streptococcus pyogenes was adapted for genome editing [8]. Within a year, multiple studies confirmed its ability to edit plant genomes and induce simultaneous mutations in several genes. Notable early examples included multiplex editing in rice and Arabidopsis in 2013 [9], and the knockout of all three homeologs of a gene in hexaploid wheat by 2014 [10]. Since then, the CRISPR toolbox has rapidly evolved to include diverse Cas nucleases, compact variants, base editors, and more. This review explores the current landscape of CRISPR-mediated multiplex genome editing in plants from its molecular basis to its emerging applications in agriculture.
Multiplex genome editing has become essential for improving complex agronomic traits, such as yield and stress tolerance, which are often regulated by interconnected gene networks rather than single genes [6]. Traditional breeding requires laborious crosses and selection to manipulate these networks, frequently resulting in unwanted linkage drag [11]. In contrast, CRISPR multiplexing enables precise, simultaneous modification of multiple genes or QTLs directly in elite cultivars, eliminating the need for extensive crossing [12]. For example, targeting entire gene families can reveal functional redundancies or generate more robust trait outcomes than single-gene edits alone [13]. This approach is particularly valuable for traits like drought tolerance, which may involve the coordinated regulation of hormonal pathways and transcription factors. With growing pressure from climate change and the need to ensure global food security, multiplex gene editing offers a rapid, efficient strategy to stack multiple traits such as yield, nutrient use efficiency, and stress resilience within a single generation [14]. The following sections review CRISPR/Cas systems, current multiplexing strategies, and their revolutionary impact on plant biology and crop improvement.
2. Evolution and Fundamentals of CRISPR/Cas Systems
Prokaryotic immune origins: CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) and their associated (Cas) proteins originated in bacteria and archaea as an adaptive immune system to defend against viruses (phages) and plasmids [15]. Within a CRISPR locus, short fragments of foreign DNA known as spacers are integrated between palindromic repeat sequences, serving as a molecular record of previous infections [16]. These loci are transcribed and processed into CRISPR RNAs (crRNAs), which guide Cas nucleases to complementary sequences in invading genomes, resulting in their targeted cleavage and neutralization [17]. CRISPR systems are broadly classified into two major classes
Class 1 systems use multi-component effector complexes (multiple Cas proteins) [18]. For example, Type I systems use a Cascade complex to bind DNA and a separate trans-acting nuclease (Cas3) to degrade the DNA. Cas3 is a helicase-nuclease "DNA shredder" that chews target DNA in one direction, causing large deletions rather than precise cuts [19]. Class 1 systems are less harnessed for genome editing due to their complexity, though emerging efforts aim to repurpose them for large integrations [20,21].
Class 2 systems use a single protein effector that performs both target recognition (with the crRNA) and cleavage [22]. Class 2 encompasses the familiar Cas9 and Cas12 nucleases (targeting DNA) and Cas13 (targeting RNA). These single-component systems are more tractable for engineering and have become the principal tools for genome editing [23]. Within Class 2, there are multiple types: Type II (Cas9), Type V (Cas12 variants), Type VI (Cas13), etc., each with distinct properties.
A schematic overview of the CRISPR–Cas system’s prokaryotic origins, classification, and engineered effectors is shown in Figure 1.
Mechanism of Streptococcus pyogenes Cas9 (SpCas9) is the archetypal Class 2 nuclease used in plant genome editing [24]. Cas9 is ~1368 amino acids (~160 kDa) and forms a ribonucleoprotein with a dual RNA guide: the crRNA (target-specific ~20 nt guide) and a trans-activating tracrRNA (or fused as a single sgRNA) [25]. The SpCas9-sgRNA complex recognizes a short PAM (Protospacer Adjacent Motif) sequence (originally NGG) on the target DNA strand, then the sgRNA hybridizes to the complementary sequence [26]. Cas9 introduces a blunt double-strand break (DSB) ~3 bp upstream of the PAM. SpCas9 has been extensively engineered, yielding dozens of variants with altered PAM specificities and improved specificity [27].
One major limitation of wild-type SpCas9 is its strict NGG PAM requirement, which restricts targeting, especially in AT-rich genomic regions. To overcome this, several engineered Cas9 variants have broadened PAM compatibility [28]. SpCas9-NG recognizes NGN PAMs (e.g., NGA, NGT), while xCas9 3.7, developed through phage-assisted evolution, expands targeting to NG, GAA, and GAT sequences, albeit with reduced activity at some sites [29]. SpG efficiently targets NGN sites, and SpRY is nearly PAM-less, recognizing NRN and NYN sequences with high editing efficiency and minimal off-target effects [30]. SaCas9, a compact Cas9 ortholog (~1053 aa) from Staphylococcus aureus, recognizes NNGRRT PAMs and is ideal for delivery via size-constrained vectors, offering an alternative when SpCas9 is limited by size or PAM availability [31]. Other CRISPR systems also broaden editing capabilities. Cas12a (Cpf1), a Type V effector (~1300 aa), introduces 5′ staggered cuts downstream of a T-rich PAM (TTTV), processes its own crRNAs, and enables multiplex editing using a single Pol III promoter—commonly used variants in plants include LbCas12a and AsCas12a [32]. Cas12b (~1100 aa), once limited by temperature sensitivity, now functions efficiently in plants through engineered versions like BhCas12b v4, recognizing TTN PAMs [33]. Cas13, a Type VI effector (e.g., LwaCas13a, Cas13b), targets ssRNA instead of DNA and exhibits collateral RNase activity—useful for diagnostics (e.g., SHERLOCK) but less suited for precise editing [34]. In plants, Cas13 enables RNA knockdown without genome modification, offering virus resistance and high specificity, with the ability to distinguish single-nucleotide changes [35]. Cas7-11, a fusion of Cas7 and Cas11, provides precise RNA editing without collateral activity, reducing toxicity [36]. Recent discoveries of compact CRISPR effectors further expand the genome editing toolkit, particularly for plant systems where vector size is limiting. CasΦ (Cas phi), a ~70 kDa phage-encoded nuclease, performs both crRNA processing and DNA cleavage using a minimal PAM (TBN), functioning effectively in plants [37]. Cas14 (Cas12f), an ultra-small enzyme (400–700 aa), targets ssDNA without PAM dependency and, though used mainly in diagnostics, holds promise for editing ssDNA viruses in plants [38]. CasX and CasY, discovered via metagenomic mining, are compact Type V nucleases (<1000 aa) that cleave dsDNA, with CasX creating 5′ overhangs and targeting T-rich PAMs [39]. Their structural divergence from Cas9 may reduce immune responses. Additionally, novel phage-derived nucleases such as CasΩ, Casμ, and Casλ offer unique, compact editing properties that are still being explored [40,41]. To increase precision, high-fidelity Cas variants like SpCas9-HF1 and eSpCas9 (1.1) have been engineered by introducing alanine substitutions at DNA-contact points, limiting cleavage to perfectly matched sequences [42]. These are widely used in plants, especially during multiplex editing where off-target risks increase. Another approach involves fusing Cas9 with anti-CRISPR proteins like AcrIIA4 linked to cell-cycle regulators, restricting activity to HDR-favorable stages [43]. A summary of representative CRISPR–Cas genome editors and their features is provided in Table 1. While some advanced variants extend beyond this review’s scope, they play a critical role in improving the safety and precision of multiplex genome editing in plants.
3. Mechanisms of Multiplex Genome Editing
3.1. DNA Double-Strand Break Repair Pathways
The indel mutations and targeted modifications achieved by CRISPR ultimately rely on the cell’s DNA repair machinery following a nuclease-induced break. In plant cells, three major DSB repair pathways can be leveraged. The dominant route, non-homologous end joining (NHEJ), directly ligates broken ends, often introducing small insertions or deletions (indels) that disrupt gene function. NHEJ operates throughout the cell cycle and is typically responsible for independent indels at each locus during multiplex editing [44]. Microhomology-mediated end joining (MMEJ) uses short homologous sequences near break sites to guide repair, resulting in predictable deletions. MMEJ can be strategically leveraged—for example, to excise specific gene regions or remove selection markers—though it is generally less efficient than NHEJ [45]. Homology-directed repair (HDR) offers precise modification using a donor template, enabling gene replacements or insertions. HDR is rare in differentiated plant cells but can be enhanced in dividing tissues, via geminiviral replicons, or by inhibiting NHEJ [46].
When multiplex editing (multiple DSBs) is introduced, these pathways can act independently at each break. It’s not uncommon to see different repair outcomes at each target: for instance, one gene may acquire a 1 bp insertion while another undergoes a 20 bp deletion [47]. If two target sites are close on the same chromosome, simultaneous breaks can lead to a larger deletion of the intervening segment or chromosomal rearrangements, an important consideration in designing multiplex experiments to avoid unintended large deletions [48] (see Challenges section).
3.2. Polycistronic Guide RNA Strategies
A central challenge in multiplex editing is how to express multiple gRNAs in plant cells efficiently [49]. Early multiplex attempts simply stacked several U6 promoter–sgRNA cassettes in one vector, which is effective for a handful of targets but becomes unwieldy as the number grows [50]. To overcome this, several elegant strategies have been developed to produce multiple guides from a single transcript (Figure 2):
- Tandem CRISPR array with direct repeats: In their native systems, Cas nucleases process long precursor transcripts containing multiple spacers separated by repeats [51]. This natural mechanism is exploited in Cas12a-based multiplexing, where a single Pol III promoter (e.g., U3 or U6) drives a compact crRNA array formatted as repeat–spacer1–repeat–spacer2––repeat–spacerN. Cas12a’s built-in RNase activity autonomously cleaves the repeats, releasing mature crRNAs for each target [52]. This enables efficient, simultaneous editing at 6–8 loci using a minimal construct. In contrast, Cas9 lacks self-processing ability, requiring a separate tracrRNA for each spacer [53]. Therefore, multiplexing with Cas9 demands more complex synthetic arrays or alternative strategies involving external processing systems.
- tRNA–gRNA system (PTG): Xie et al. (2015) developed the PTG (polycistronic tRNA–gRNA) system, where tRNAs flank sgRNAs in a single transcript under a U6 promoter [54]. Plant RNases process the tRNAs, releasing functional sgRNAs [55]. This strategy enabled efficient multiplex editing in rice and Arabidopsis, with only a slight drop in efficiency as unit number increased [56]. PTG remains popular for its simplicity and native processing.
- Ribozyme-based self-processing: This method uses self-cleaving ribozymes (hammerhead and HDV) flanking each sgRNA, enabling a single Pol II transcript to yield multiple sgRNAs [57]. Tang et al. (2016) showed a maize ubiquitin promoter-driven cassette expressing Cas9 + 6 ribozyme-flanked sgRNAs, achieving high editing efficiency [58]. Ribozymes cleave in cis, work under Pol II or III, and allow compact, all-in-one constructs ideal for viral or single-vector delivery.
- Csy4 cleavage system: Csy4, an RNA endoribonuclease from Pseudomonas, recognizes a 20-nt hairpin and cleaves 3’ of it [59]. When Csy4 hairpins are appended to sgRNAs in a Pol II transcript, co-expressed Csy4 precisely releases individual sgRNAs [60,61]. Tsai et al. and Cermák et al. applied this for plant multiplexing; Cermák found Csy4 outperformed tRNA and ribozyme systems, efficiently processing up to 12 sgRNAs in tomato [62]. The trade-off is the need to co-express the ~600 bp csy4 gene, but its clean cleavage and minimal scar make it well-suited for complex editing [63].
- MicroRNA-based systems (Drosha/Dicer): miRNA-based sgRNA expression leverages the plant’s own miRNA processing machinery. sgRNAs are embedded in native miRNA backbones (e.g., OsMIR528, AtMIR390), allowing Drosha/DCL1 and Dicer to process the pri-miRNA into functional sgRNAs [64]. Multiple sgRNAs can be multiplexed in a single Pol II transcript. Though sometimes less efficient than tRNA or ribozyme systems, this method enables tissue-specific or inducible expression and has been successfully used in Arabidopsis and rice for multi-gene knockouts.
Each of these strategies has pros and cons. Pol III-based strategies (U6-driven arrays or tRNA processing) are generally simple and produce high levels of sgRNAs in most cell types, but Pol III promoters are small and lack regulatory elements for tissue control. Pol II strategies (ribozyme, miRNA, Csy4) can be coupled with strong or inducible promoters and even included on the same transcript as Cas9, but they involve more complex design. Table 2 provides a comparative overview of multiplex gRNA expression techniques in plants.
3.3. Ultra-Multiplexing Capacity
Ultra-multiplex CRISPR enables editing of dozens of genes simultaneously, limited mainly by construct size and sgRNA expression. Studies routinely deliver >10 sgRNAs, and a record system in rice (GoGo/pKOCK) assembled 48 sgRNAs targeting 49 cytokinin-related genes using modular U6 promoters [65]. In sugarcane, CRISPR/TALENs mutated 100+ alleles across a gene family [66]. These tools allow large-scale, combinatorial editing of complex traits (yield, metabolism, architecture), enabling single-step rewiring of entire pathways and phenotypic screening at unprecedented scales.
4. Expression Systems for Multiple gRNAs
Delivering multiple guide RNAs into plant cells requires not only clever transcript processing but also robust expression units and vector architectures. Key considerations include the choice of promoter for sgRNAs, how to co-express the Cas nuclease, and regulatory elements like introns and nuclear localization signals to enhance expression.
4.1. Pol III vs. Pol II Promoters
RNA polymerase III promoters like U6 and U3 are commonly used for sgRNA expression due to their short, constitutive nature and ability to produce non-polyadenylated transcripts—crucial for sgRNA functionality [70]. They are effective across plant species but lack tissue or temporal specificity and make constructs bulky and repetitive when multiple U6 cassettes are used. In contrast, Pol II promoters (e.g., CaMV 35S, ZmUbiquitin, EC1.2) offer greater flexibility, enabling tissue-specific, inducible, or polycistronic sgRNA expression when combined with processing elements like ribozyme, Csy4, or miRNA scaffolds [49]. This allows precise control of editing events, reduces mosaicism, and supports germline transmission. Hybrid systems are often adopted: Pol III for 2–6 sgRNAs, and Pol II-based systems for larger arrays or conditional expression [71]. Cas12a, with its self-processing activity, further simplifies multiplexing by requiring only a single Pol III promoter for multiple crRNAs [72]. Figure 3 illustrates comparative architectures of Pol III- and Pol II-driven sgRNA expression systems and modular assembly approaches.
4.2. Single Versus Multiple Transcripts
In multiplex genome editing, Cas9 (or Cas12) and sgRNAs can be expressed from separate transcripts—typically, Cas9 under a Pol II promoter and each sgRNA under individual Pol III promoters [73]. Alternatively, “all-in-one” designs encode both Cas9 and sgRNA arrays in a single Pol II transcript, using ribozyme or Csy4 elements for sgRNA processing and 2A peptides for polyprotein cleavage [74]. This approach ensures co-delivery and synchronized expression, often improving editing efficiency. However, assembling large all-in-one constructs can be technically difficult and may reduce transcript stability or transformation efficiency due to transcript size constraints in planta.
4.3. Codon Optimization, Introns, and NLS
Efficient Cas9/Cas12 expression in plants requires codon optimization to match plant-preferred codons and remove unwanted RNA motifs [75]. Modern constructs often use Arabidopsis- or maize-optimized Cas9, with SV40 nuclear localization signals (NLS) at one or both termini to ensure nuclear targeting—critical for activity [76], as Cas proteins are ineffective if retained in the cytoplasm. Additionally, intron insertion into the Cas9 gene (Intron-Mediated Enhancement, IME) significantly boosts expression and editing efficiency by enhancing mRNA stability, export, or avoiding gene silencing [77]. For example, intron-optimized Cas9 driven by the 35S promoter showed higher mutagenesis than intron-less versions [78]. Many current vectors include synthetic or native introns (e.g., petunia TEF1α intron) to maximize performance in planta [79].
4.4. Promoter choices for Cas9
Cas9 is typically driven by a strong, constitutive Pol II promoter to ensure robust expression across plant tissues. Widely used promoters include CaMV 35S [80], AtUBQ10 (Arabidopsis Ubiquitin-10) [81], ZmUBI (Maize Ubiquitin) [82], and Actin promoters [83]. However, for achieving heritable edits, germline-specific promoters like EC1.2 (egg cell-specific) or SPOROCYTE (pollen-specific) are preferred [84,85], as they confine Cas9 activity to reproductive cells, producing non-chimeric, stably edited progeny. For example, in soybean, egg cell-specific expression of Cas9 transformed somatic mutations into inheritable edits in the T1 generation [86]. An advanced method called HI-Edit [87], which expresses Cas9 in a haploid inducer background (e.g., maize), enables the generation of edited progeny without transgene integration. Ultimately, the choice of promoter for Cas9 can be tailored to the experimental goal ubiquitous promoters for broad editing or tissue-specific promoters to enhance germline editing efficiency and reduce somatic off-target effects.
4.5. Golden Gate and Modular Assembly Systems
Building CRISPR multiplex vectors can be complex, but modular cloning systems like Golden Gate [88], MoClo [89], and GoldenBraid [90] streamline the process. GoldenBraid 4.0 (GB4.0) simplifies plant genome engineering by offering ready-to-use parts (promoters, sgRNA scaffolds, terminators) and one-step assembly. GB4.0 enables quick swapping of promoters or addition of guides, and allows integration of markers like fluorescent tags for Cas9. Using a one-tube reaction, researchers can assemble T-DNA vectors carrying Cas9 and multiple U6-sgRNA modules [91]. GB4.0 was successfully used to edit gene families in tobacco. Similarly, Plant MoClo [92] and CRISPR-ABC [93] kits support efficient multiplex editing in academic labs.
4.6. Vector Delivery Formats
For stable transformation, binary T-DNA vectors (via Agrobacterium) are widely used and can carry Cas9 with up to ~10 sgRNAs (T-DNA < 20 kb) [94]. Larger constructs may reduce transformation efficiency. For transient or virus-based delivery, compact all-in-one designs are preferred, often using smaller Cas variants or transgenic lines expressing Cas9 with sgRNAs delivered by virus [95]. Modern multiplex CRISPR systems—featuring strong promoters, codon-optimized Cas9 with NLS and introns, and efficient gRNA expression—enable high editing efficiency at multiple loci. These advancements have made complex plant genome engineering highly feasible.
5. Delivery Systems for Mulplex-CRISPR Components in Plants
Successfully editing plant genomes requires getting the CRISPR-Cas machinery (Cas proteins or their genes, and guide RNAs) into plant cells. Plants pose additional challenges for delivery compared to animal systems due to the rigid cell wall, need for regenerating whole plants from edited cells, and often longer generation times. A variety of delivery methods have been developed (Figure 4), including both traditional transformation platforms and novel nanotechnology or virus-based approaches.
Agrobacterium-mediated delivery: Agrobacterium tumefaciens, a soil bacterium that naturally transfers T-DNA into plant genomes, is the most widely used method for delivering CRISPR constructs in plants, particularly for stable transformation [96]. Cas9 and sgRNA cassettes are inserted into binary T-DNA vectors, which Agrobacterium transfers into plant cells like embryogenic calli or explants. The integrated T-DNA enables expression of editing components, leading to targeted genome modifications. This method is efficient, relatively simple, and effective across many species—especially dicots (e.g., tomato, tobacco, soybean) and optimized monocots (e.g., rice) [97]. Regenerated T₀ plants often carry heritable edits, and segregation of the T-DNA in the next generation yields transgene-free lines [98]. For instance, CRISPR editing of OsERF922 in rice via Agrobacterium produced ~50% disease-resistant mutants [99]. Agrobacterium’s ability to carry large T-DNA constructs makes it suitable for multiplex editing. However, transformation efficiency remains species- and genotype-dependent, and tissue culture steps are labor-intensive [100].
Biolistics (particle bombardment): Biolistic delivery (gene gun) propels DNA or RNA coated gold/tungsten particles into plant cells and is commonly used for species less responsive to Agrobacterium, such as maize, wheat, and barley [101,102]. For stable transformation, CRISPR plasmids are delivered into embryogenic tissues, where they may integrate and express the transgene. This method accommodates large DNA constructs and co-delivery of elements like Cas and HDR templates [103]. It enabled the first CRISPR edits in maize via Cas9/sgRNA plasmid bombardment [104]. Biolistics also enables transgene-free editing by delivering Cas9–sgRNA ribonucleoproteins (RNPs) directly into cells; for example, Liang et al. (2017) achieved targeted, DNA-free edits in maize [105].
Protoplast transfection: Protoplast-based delivery uses enzymatic digestion to remove plant cell walls, allowing uptake of CRISPR components via PEG or electroporation [106]. Commonly used for transient expression in species like rice or lettuce, it enables rapid edit detection (1–2 days) using plasmid DNA or Cas9–sgRNA RNPs, supporting DNA-free editing. It's valuable for testing CRISPR variants (e.g., SpCas9, LbCas12a). The main limitation is regenerating whole plants, feasible only in species with efficient tissue culture (e.g., lettuce, cabbage, sorghum). In such cases, RNP-edited protoplasts can regenerate into transgene-free plants. For example, editing potato protoplasts with Cas9 RNPs targeting Vlnv and Asn1 reduced acrylamide content without foreign DNA integration [107].
Viral vectors (virus-induced genome editing, VIGE): Editing VIGE uses engineered plant viruses to deliver CRISPR components, avoiding tissue culture [108]. Due to limited cargo capacity, viruses typically carry sgRNAs or compact Cas variants, not full-length SpCas9. The common strategy involves Cas9-transgenic plants infected with viral vectors (e.g., TRV, gemini viruses) delivering sgRNAs for systemic editing. TRV enabled somatic and heritable edits in tobacco, while gemini viruses, with higher replication and cargo capacity, improved HDR efficiency, as shown in tomato gene knock-ins [109]. Viruses facilitate multiplex editing by spreading sgRNAs throughout the plant, even to meristems, often using tRNA-based sgRNA processing [110]. Though mosaicism can hinder uniformity, regenerating from infected tissues or targeting reproductive organs enhances germline edits [111]. Some systems co-deliver compact Cas variants like SaCas9 (~3.2 kb) with sgRNAs, demonstrated in lettuce [112].
Nanoparticle and novel delivery: Nanotechnology-based delivery offers non-viral, non-biological methods to introduce CRISPR components into plants using materials like carbon nanotubes (CNTs), gold nanoparticles (AuNPs), and lipid nanoparticles (LNPs) [113]. Functionalized single-walled CNTs have successfully delivered plasmid DNA and Cas9 RNPs into Eruca sativa and Nicotiana benthamiana leaves, enabling transient, DNA-free editing [114]. AuNPs, proven in mammalian systems (e.g., CRISPR-Gold), and PEGylated liposomes have also induced edits in wheat calli without foreign DNA [115]. These nanoparticles support DNA-free editing with less tissue damage than biolistics, though targeting inner tissues and meristems remains challenging. Innovative techniques like pollen magnetofection (using magnetic Cas9 RNP-coated pollen) and spray-induced gene editing (nanocarrier-bound RNPs applied to leaves) show promise for germline edits and in-field, transgene-free crop editing broadening future applications in plant genome engineering [116].
Tissue culture considerations: Many delivery methods like Agrobacterium, biolistics, and protoplast transfection rely on in vitro culture and plant regeneration, posing challenges for recalcitrant species (e.g., trees, elite maize lines) [117]. One solution is co-expression of morphogenic regulators like BABY BOOM (BBM) and WUSCHEL (WUS) during transformation [118], which promotes regeneration and enables somatic embryogenesis, even in previously untransformable genotypes—used in methods like Fast-Tracc and HI-Edit. Another strategy is in planta transformation, such as the Arabidopsis floral dip [119], where Agrobacterium is applied to flowers to yield transformed seeds. This method efficiently introduces CRISPR constructs and often results in biallelic edits in T1 progeny [120]. Similar tactics, like agro-infiltration of floral tissues or meristems, can bypass tissue culture by editing gametes directly, producing heritable mutations in the next generation [121].
6. CRISPR Multiplexing in Crop Improvement and Trait Engineering
6.1. Yield and Biomass
Improving crop yield often involves targeting multiple genes simultaneously, and multiplex CRISPR has proven transformative in this regard. A landmark study by Zhu’s team in rice demonstrated that knocking out all 13 PYL ABA receptor genes—normally acting as brakes on growth—led to a 25–31% increase in grain yield in field trials without significantly compromising stress tolerance [122]. Another study edited three yield-associated genes (OsPIN5b, GS3, and OsMYB30) in rice, resulting in plants with longer grains, more spikelets, and improved cold tolerance, effectively overcoming the common trade-off between yield and stress resilience [123]. In tomato, multiplex editing of stem cell regulatory genes within the CLV-WUS pathway yielded larger fruits and more per truss in just one generation emulating decades of traditional breeding [124]. Sugarcane, a complex polyploid, also benefited from this approach: researchers used multiplex CRISPR to edit multiple copies of the LG1 gene regulating leaf angle. One edited line (L35) showed a 56% reduction in leaf inclination angle and an 18% boost in biomass yield by optimizing canopy structure for better light interception [125]. Similarly, in Brassica napus, targeting five JAGGED (JAG) gene homoeologs induced significant phenotypic shifts: leaf serration, floral abnormalities, and notably, enhanced resistance to pod shattering via modified dehiscence zone architecture [126]. These studies highlight how multiplex CRISPR can rapidly combine beneficial alleles to create high-yielding ideotypes, offering precision and efficiency far beyond conventional breeding methods (Table 3).
6.2. Disease Resistance
CRISPR multiplexing enables simultaneous targeting of susceptibility genes and stacking of resistance traits. In rice, dual knockout of OsERF922 and OsSEC3A significantly enhanced blast resistance. Wheat lines with all three MLOhomoeologs edited showed strong resistance to powdery mildew, bypassing the need for complex breeding [99]. In citrus, editing the CsLOB1 promoter alongside defense gene insertion conferred canker resistance [130,131]. Tomato plants with JAZrepressor knockouts displayed enhanced fungal immunity [132]. Li et al. (2018) demonstrated the use of multiplex CRISPR in Arabidopsis to dissect the roles of BnWRKY11 and BnWRKY70, identifying BnWRKY70 as key for Sclerotinia resistance [126]. In elite wheat, combined knockout of MLO genes and activation of dormant R genes achieved robust mildew resistance [133]. These examples highlight the potential of multiplex editing for durable, broad-spectrum disease resistance (Table 4).
6.3. Abiotic Stress Tolerance
Abiotic stress tolerance involves complex, multi-genic traits. Multiplex CRISPR enables coordinated edits across key pathways. For instance, knocking out multiple PYL ABA receptors in rice improved drought tolerance and yield by reducing growth suppression under water deficit [137]. In tomato, editing negative regulators in ethylene and jasmonate signaling enhanced salt stress tolerance [138]. In Arabidopsis, quadruple knockout in the proline degradation pathway increased osmoprotectant levels, improving drought resilience [139]. For heat tolerance, combined knockout of heat-susceptibility genes and CRISPRa-driven overexpression of heat shock regulators improved thermotolerance. Abdallah et al. (2025) successfully knocked out five TaSal1 homologs in wheat using multiplex CRISPR, with full knockout lines showing enhanced drought tolerance and improved anatomical traits [140]. These studies show how stacking minor-effect edits can yield significant gains in stress resilience (Table 5).
6.4. Nutritional Quality
Improving crop nutrition often requires editing multiple genes. In potato, simultaneous knockout of VInv and Asn1drastically reduced acrylamide formation during frying, enhancing food safety [128]. In tomato, editing sugar-repressing kinases (SlCDPK26 and SlCDPK27) boosted glucose and fructose by 30% without yield loss [144]. Sorghum biofortification involved editing α-kafirin genes to improve protein digestibility and removing anti-nutritional peptides via PTR transporter knockout, increasing lysine availability [145]. In rice, multiplex promoter editing of three NF-YC genes raised grain protein content without yield penalty [146]. Similarly, BnFAD2 and BnFAE1 mutations in rapeseed boosted oleic acid content to 70–80% [147]. Multiplex editing has also enabled gluten reduction in wheat by targeting γ- and ω-gliadin genes, and altered tomato fruit pigmentation through edits in pigment biosynthesis genes—showcasing CRISPR’s potential in tailoring nutritional quality and crop aesthetics [148].
6.5. De Novo Domestication of Wild Plants
CRISPR enables rapid domestication of wild or orphan crops by editing multiple domestication genes simultaneously. Zsögön et al. (2018) edited six loci in wild tomato (S. pimpinellifolium), enhancing fruit size, yield, and lycopene content—achieving traits similar to modern tomato in one generation [124]. Similarly, Lemmon et al. (2018) used multiplex CRISPR to improve groundcherry (Physalis pruinosa) architecture and fruiting traits, making it cultivation-ready [149]. This approach mimics natural domestication but accelerates the process, allowing breeders to transform wild species—rich in stress tolerance or nutrition—into productive crops by stacking key domestication alleles (e.g., larger fruit, loss of seed dormancy, reduced shattering) all at once.
Breeders often aim to improve yield, disease resistance, and quality together. Multiplex CRISPR enables this in a single step. For instance, Chinese researchers edited Gn1a (yield), Pi21 (blast susceptibility), and Wx (grain quality) simultaneously in elite rice, producing high-yielding, blast-resistant lines with preferred grain traits—without linkage drag [150]. In tomato, knocking out DET1 (carotenoid repressor) and IAA9 (fruit initiation repressor) created seedless, deep-red fruits [151]. These examples show how CRISPR accelerates trait stacking, replacing years of breeding with precise, multi-gene edits.
7. Challenges and Limitations
Off-target effects are a major concern in multiplex genome editing, as introducing multiple sgRNAs increases the likelihood of unintended mutations at similar genomic sites due to mismatch tolerance [152]. This risk is heightened when dozens of guides are used, potentially confounding phenotypic analysis and raising regulatory issues. Careful guide design using tools like CRISPR-P [153], CHOPCHOP [154], or CRISPOR [155], along with high-fidelity Cas9 variants such as SpCas9-HF1 [42] and eSpCas9 [156], can significantly reduce off-target activity. In plants, such mutations are generally rare, likely due to chromatin structure, but validation of top predicted off-target sites is still recommended. Transient delivery methods like Cas9 RNPs further reduce risk by shortening the editing window.
Editing efficiency is especially challenging in polyploid crops (e.g., wheat, potato, sugarcane), where multiple gene copies must be edited simultaneously. Conserved sgRNAs or multiple guides can target all alleles, but achieving full edits in a single plant is difficult due to mosaicism and varying sgRNA efficiency [157]. Techniques like using strong promoters, multiple sgRNAs, and haploid induction systems (e.g., HI-Edit in maize) help improve efficiency, though extensive screening and genotyping are often required [158].
Multiplex editing also increases the chance of unintended large deletions or chromosomal rearrangements when multiple cuts occur close together [159]. While sometimes intentional, such events may affect nearby genes or regulatory elements. Rare instances of translocations have been reported in plants, particularly with nucleases like Cas12a [160]. Detection methods include long-read sequencing or PCR across target regions, and risks can be reduced by spacing guide expression or avoiding closely located targets [161].
Mosaicism and chimerism are common in T0 plants, as editing may occur unevenly during regeneration, making early phenotyping unreliable [162]. Stable homozygous lines are typically obtained in T1/T2 generations via selfing or crossing. Using egg cell-specific promoters (e.g., EC1) or delivering CRISPR as RNPs can reduce mosaicism and promote uniform edits [163].
Delivery and tissue culture hurdles remain as a major barriers, especially in woody and recalcitrant crops, with transformation often inducing somaclonal variation and gene silencing due to transgene load. Optimizing expression with viral promoters or intron-containing constructs and exploring novel methods like meristem-targeted transformation or in planta editing may help circumvent tissue culture.
Intellectual property and regulatory frameworks also impact deployment of multiplex-edited crops. Patent restrictions around CRISPR components and delivery platforms necessitate complex licensing for commercial use. Regulations vary globally: countries like the USA, Canada, Brazil, and Japan often exempt DNA-free genome edits from GMO rules, while the EU applies strict GMO regulations. India and China are still shaping their policies. Although public concern is low, demand for safety and compositional analyses is increasing, especially for traits involving metabolic changes. Clear labeling, transparency, and safety validation are vital for gaining public trust and regulatory approval.
The biological complexity of gene networks poses challenges, as multiplex edits can produce unintended phenotypes due to pleiotropy or gene interactions. For example, full knockout of ABA receptors improved yield but increased drought sensitivity [164]. Breeders may prefer partial knockouts or precise edits via promoter, base, or prime editing to fine-tune gene function. Balancing editing breadth with functional precision, supported by systems biology insights, is key to avoiding trade-offs and achieving desired agronomic outcomes.
8. Computational and AI Integration in Multiplex-CRISPR Design
The complexity of multiplex editing – choosing many target sites across the genome – is greatly aided by computational tools. Bioinformatics and artificial intelligence (AI) approaches are now integral to designing effective and specific CRISPR experiments:
In silico sgRNA design tools have significantly advanced CRISPR research in plants. Tools like CRISPR-P, which was specifically developed for plants, allow researchers to input a gene or genomic region and obtain a ranked list of sgRNAs based on predicted on-target efficiency and potential off-target effects. CRISPR-P was first introduced in 2014 and later updated to CRISPR-P 2.0, which includes support for plant-specific repeats and offers designs for multiple Cas variants such as Cas9 and Cas12a. General tools like CHOPCHOP, E-CRISPR, and Benchling also support plant genomes and are widely used. These platforms are particularly beneficial for multiplex genome editing, as they allow batch design of sgRNAs while minimizing overlaps and unintended off-target interactions, especially when targeting gene families.
AI for on-target efficiency has become an essential part of sgRNA design. Since not all sgRNAs perform equally, machine learning models trained on large datasets are now used to predict their cutting efficiency. Tools like DeepCRISPR [165], DeepSpCas9 [166], and SSC (sequence-specific scoring) [167] models analyze sequence features to estimate cleavage potential. Although most training data come from human systems, many plant-based tools incorporate these models. CRISPRon [168], a newer AI-based tool, integrates features like GC content, sequence motifs, and nucleotide preferences to generate high-accuracy activity scores. When designing multiplex sgRNA sets, using these AI-derived scores helps in selecting the top-performing guides with minimal off-target risk. Some tools also predict the nature of indels produced, enabling selection of guides that induce desired frameshift mutations.
Off-target prediction and avoidance is a crucial computational step, particularly in multiplex editing. Tools like CAS-OFFinder [169] and CRISPOR can identify potential off-target sites across the genome by allowing up to 3–4 mismatches. The CRISPRoff platform complements CRISPRon by offering predictions on off-target cleavage probabilities. These tools allow users to customize the PAM sequence and select the genome of interest, ensuring that sgRNAs do not unintentionally hit other gene family members or repetitive sequences. This becomes even more critical in polyploid species, where homeologous genes may be similar enough to be mistaken for off-targets. In such cases, sgRNA design can be fine-tuned depending on whether those genes should be co-targeted or excluded.
Machine learning for prime and base editing is an evolving frontier in CRISPR technology. Prime editing requires the design of pegRNAs, which are structurally complex. Tools like PrimeDesign [170] and PE-Designer [171] use machine learning algorithms trained on experimental data to suggest pegRNA sequences with higher predicted success. Similarly, base editing tools predict the effective editing window and bystander mutations for a given target site, helping researchers design guide RNAs that maximize precision. These AI tools are especially useful when attempting subtle nucleotide edits, such as correcting point mutations or introducing SNPs for trait improvement.
Multi-omics target identification leverages big data and AI to determine which genes to edit for complex traits. By integrating genomics, transcriptomics, proteomics, and metabolomics data, researchers can prioritize targets for multiplex editing. For instance, genome-wide association studies (GWAS) can identify several loci associated with a trait like drought tolerance. AI tools can further refine this list by incorporating expression QTLs and known pathways. A real-world example includes enhancing tomato sweetness by targeting genes like SlCDPK27 and SlCDPK26, identified via co-expression network analysis [144]. This omics-driven approach enables CRISPR validation of multiple candidates simultaneously, accelerating trait improvement.
CRISPR libraries for plants represent large-scale sgRNA design efforts aimed at editing entire gene families or even entire genomes. Tools like CCTop [172] and CHOPCHOP offer batch design capabilities, allowing researchers to generate sgRNAs for hundreds or thousands of targets. These designs can be filtered using AI models for on-target efficiency and off-target avoidance. An example includes the work by Meng et al., who created a comprehensive sgRNA library targeting a large number of genes, enabling high-throughput functional genomics studies [173]. Such libraries are instrumental for forward genetics screens and offer a path to uncover novel gene functions through multiplex editing.
9. Conclusions
CRISPR multiplex genome editing has ushered in a new horizon for plant science and crop breeding. We can now target any set of genes and sculpt plant genomes with a level of control previously unimaginable. As the tools improve – smarter design by AI, better delivery, more precise nucleases – and as regulatory frameworks adapt to these technologies, we will likely witness an acceleration in crop improvement akin to the “green revolution”, but driven by bytes and base edits rather than by crosses and chemicals. The ultimate vision is climate-resilient, high-yield, nutritious crops engineered in a fraction of the time of conventional breeding, helping secure food production in a sustainable manner. Achieving this will require addressing the current challenges and continuing to advance our scientific and technical capabilities, but the rapid progress to date gives confidence that these goals are within reach. Each year brings new CRISPR breakthroughs – by the next decade, multiplex editing could well be a standard tool in every plant breeder’s toolkit, and many of our staple crops might be products of this precise genome surgery, feeding the world with higher efficiency and fewer inputs. The seeds of that future are already being sown today in laboratories and greenhouses across the globe.
Authorship Contributions: Uday Kumar BV: Writing - Original Draft, Writing - Review & Editing, Visualization,Dyumn Dwivedi: Visualization; Uddalak Das: Writing - Review & Editing; Conceptualization; Supervision.
Funding
None.
Conflicts of Interest
The authors report no conflict of interest.
References
- Hamdan, M.F.; Tan, B.C. Genetic modification techniques in plant breeding: A comparative review of CRISPR/Cas and GM technologies. Hortic. Plant J. 2024. [CrossRef]
- Carroll, D. Genome engineering with zinc-finger nucleases. Genetics 2011, 188, 773–782. [CrossRef]
- Sander JD, Joung JK. CRISPR-Cas systems for editing, regulating and targeting genomes. Nat Biotechnol. 2014 Apr;32(4):347–55.
- Xiong, X.; Chen, M.; Lim, W.A.; Zhao, D.; Qi, L.S. CRISPR/Cas9 for Human Genome Engineering and Disease Research. Annu. Rev. Genom. Hum. Genet. 2016, 17, 131–154. [CrossRef]
- Abdul Aziz M, Masmoudi K. Molecular breakthroughs in modern plant breeding techniques. Hortic Plant J. 2025 Jan;11(1):15–41.
- Abdelrahman, M.; Wei, Z.; Rohila, J.S.; Zhao, K. Multiplex Genome-Editing Technologies for Revolutionizing Plant Biology and Crop Improvement. Front. Plant Sci. 2021, 12. [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., III. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2014 31, 397–405. [CrossRef]
- Le Rhun, A.; Escalera-Maurer, A.; Bratovič, M.; Charpentier, E. CRISPR-Cas in Streptococcus pyogenes. RNA Biol. 2019, 16, 380–389. [CrossRef]
- Xing, H.-L.; Dong, L.; Wang, Z.-P.; Zhang, H.-Y.; Han, C.-Y.; Liu, B.; Wang, X.-C.; Chen, Q.-J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 1–12. [CrossRef]
- Abe, F.; Haque, E.; Hisano, H.; Tanaka, T.; Kamiya, Y.; Mikami, M.; Kawaura, K.; Endo, M.; Onishi, K.; Hayashi, T.; et al. Genome-Edited Triple-Recessive Mutation Alters Seed Dormancy in Wheat. Cell Rep. 2019, 28, 1362–1369.e4. [CrossRef]
- Singer, S.D.; Laurie, J.D.; Bilichak, A.; Kumar, S.; Singh, J. Genetic Variation and Unintended Risk in the Context of Old and New Breeding Techniques. Crit. Rev. Plant Sci. 2021, 40, 68–108. [CrossRef]
- Hua K, Zhang J, Botella JR, Ma C, Kong F, Liu B, et al. Perspectives on the Application of Genome-Editing Technologies in Crop Breeding. Mol Plant. 2019 Aug;12(8):1047–59.
- Iohannes, S.D.; Jackson, D. Tackling redundancy: genetic mechanisms underlying paralog compensation in plants. New Phytol. 2023, 240, 1381–1389. [CrossRef]
- Chavhan, R.L.; Jaybhaye, S.G.; Hinge, V.R.; Deshmukh, A.S.; Shaikh, U.S.; Jadhav, P.K.; Kadam, U.S.; Hong, J.C. Emerging applications of gene editing technologies for the development of climate-resilient crops. Front. Genome Ed. 2025, 7, 1524767. [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [CrossRef]
- Karginov, F.V.; Hannon, G.J. The CRISPR System: Small RNA-Guided Defense in Bacteria and Archaea. Mol. Cell 2010, 37, 7–19. [CrossRef]
- Zhu, Y.; Klompe, S.E.; Vlot, M.; van der Oost, J.; Staals, R.H. Shooting the messenger: RNA-targetting CRISPR-Cas systems. Biosci. Rep. 2018, 38. [CrossRef]
- Liu, T.Y.; Doudna, J.A. Chemistry of Class 1 CRISPR-Cas effectors: Binding, editing, and regulation. J. Biol. Chem. 2020, 295, 14473–14487. [CrossRef]
- Kim, D.Y.; Lee, S.Y.; Ha, H.J.; Park, H.H. Structural basis of Cas3 activation in type I-C CRISPR-Cas system. Nucleic Acids Res. 2024, 52, 10563–10574. [CrossRef]
- Ebrahimi, S.; Khosravi, M.A.; Raz, A.; Karimipoor, M.; Parvizi, P. CRISPR-Cas Technology as a Revolutionary Genome Editing tool: Mechanisms and Biomedical Applications. Iran. Biomed. J. 2023, 27, 219–246. [CrossRef]
- Das, U.; Chandramouli, L.; Uttarkar, A.; Kumar, J.; Niranjan, V. Discovery of natural compounds as novel FMS-like tyrosine kinase-3 (FLT3) therapeutic inhibitors for the treatment of acute myeloid leukemia: An in-silico approach. Asp. Mol. Med. 2024, 5. [CrossRef]
- Shmakov, S.; Abudayyeh, O.O.; Makarova, K.S.; Wolf, Y.I.; Gootenberg, J.S.; Semenova, E.; Minakhin, L.; Joung, J.; Konermann, S.; Severinov, K.; et al. Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems. Mol. Cell 2015, 60, 385–397. [CrossRef]
- Tang, Y.; Fu, Y. Class 2 CRISPR/Cas: an expanding biotechnology toolbox for and beyond genome editing. Cell Biosci. 2018, 8, 1–13. [CrossRef]
- Agudelo, D.; Carter, S.; Velimirovic, M.; Duringer, A.; Rivest, J.-F.; Levesque, S.; Loehr, J.; Mouchiroud, M.; Cyr, D.; Waters, P.J.; et al. Versatile and robust genome editing with Streptococcus thermophilus CRISPR1-Cas9. Genome Res. 2020, 30, 107–117. [CrossRef]
- Asmamaw M, Zawdie B. Mechanism and Applications of CRISPR/Cas-9-Mediated Genome Editing. Biol Targets Ther. 2021;15:353–61.
- Karvelis, T.; Gasiunas, G.; Young, J.; Bigelyte, G.; Silanskas, A.; Cigan, M.; Siksnys, V. Rapid characterization of CRISPR-Cas9 protospacer adjacent motif sequence elements. Genome Biol. 2015, 16, 1–13. [CrossRef]
- Kleinstiver, B.P.; Prew, M.S.; Tsai, S.Q.; Topkar, V.V.; Nguyen, N.T.; Zheng, Z.; Gonzales, A.P.W.; Li, Z.; Peterson, R.T.; Yeh, J.-R.J.; et al. Engineered CRISPR-Cas9 nucleases with altered PAM specificities. Nature 2015, 523, 481–485. [CrossRef]
- Li, J.; Xu, R.; Qin, R.; Liu, X.; Kong, F.; Wei, P. Genome editing mediated by SpCas9 variants with broad non-canonical PAM compatibility in plants. Mol. Plant 2021, 14, 352–360. [CrossRef]
- Hu, J.H.; Miller, S.M.; Geurts, M.H.; Tang, W.; Chen, L.; Sun, N.; Zeina, C.M.; Gao, X.; Rees, H.A.; Lin, Z.; et al. Evolved Cas9 variants with broad PAM compatibility and high DNA specificity. Nature 2018, 556, 57–63. [CrossRef]
- Yang, C.; Zhou, Z.; Sun, X.; Ju, H.; Yue, X.; Rao, S.; Xue, C. PAMless SpRY exhibits a preference for the seed region for efficient targeting. Cell Rep. 2024, 43, 114225. [CrossRef]
- Kaya, H.; Ishibashi, K.; Toki, S. A Split Staphylococcus aureus Cas9 as a Compact Genome-Editing Tool in Plants. Plant Cell Physiol. 2017, 58, 643–649. [CrossRef]
- Bandyopadhyay, A.; Kancharla, N.; Javalkote, V.S.; Dasgupta, S.; Brutnell, T.P. CRISPR-Cas12a (Cpf1): A Versatile Tool in the Plant Genome Editing Tool Box for Agricultural Advancement. Front. Plant Sci. 2020, 11. [CrossRef]
- Ming, M.; Ren, Q.; Pan, C.; He, Y.; Zhang, Y.; Liu, S.; Zhong, Z.; Wang, J.; Malzahn, A.A.; Wu, J.; et al. CRISPR–Cas12b enables efficient plant genome engineering. Nat. Plants 2020, 6, 202–208. [CrossRef]
- Kellner MJ, Koob JG, Gootenberg JS, Abudayyeh OO, Zhang F. SHERLOCK: nucleic acid detection with CRISPR nucleases. Nat Protoc. 2019 Oct;14(10):2986–3012.
- Kavuri, N.R.; Ramasamy, M.; Qi, Y.; Mandadi, K. Applications of CRISPR/Cas13-Based RNA Editing in Plants. Cells 2022, 11, 2665. [CrossRef]
- Kato, K.; Zhou, W.; Okazaki, S.; Isayama, Y.; Nishizawa, T.; Gootenberg, J.S.; Abudayyeh, O.O.; Nishimasu, H. Structure and engineering of the type III-E CRISPR-Cas7-11 effector complex. Cell 2022, 185, 2324–2337.e16. [CrossRef]
- Pausch, P.; Al-Shayeb, B.; Bisom-Rapp, E.; Tsuchida, C.A.; Li, Z.; Cress, B.F.; Knott, G.J.; Jacobsen, S.E.; Banfield, J.F.; Doudna, J.A. CRISPR-CasΦ from huge phages is a hypercompact genome editor. Science 2020, 369, 333–337. [CrossRef]
- Xin, C.; Yin, J.; Yuan, S.; Ou, L.; Liu, M.; Zhang, W.; Hu, J. Comprehensive assessment of miniature CRISPR-Cas12f nucleases for gene disruption. Nat. Commun. 2022, 13, 1–10. [CrossRef]
- Urbaitis T, Gasiunas G, Young JK, Hou Z, Paulraj S, Godliauskaite E, et al. A new family of CRISPR -type V nucleases with C-rich PAM recognition. EMBO Rep. 2022 Dec 6;23(12):e55481.
- Al-Shayeb, B.; Skopintsev, P.; Soczek, K.M.; Stahl, E.C.; Li, Z.; Groover, E.; Smock, D.; Eggers, A.R.; Pausch, P.; Cress, B.F.; et al. Diverse virus-encoded CRISPR-Cas systems include streamlined genome editors. Cell 2022, 185, 4574–4586.e16. [CrossRef]
- Das, U.; Banerjee, S.; Sarkar, M.; L, F.M.; Soni, T.K.; Saha, M.; Pradhan, G.; Chatterjee, B. Circular RNA vaccines: Pioneering the next-gen cancer immunotherapy. Cancer Pathog. Ther. 2024. [CrossRef]
- Casini, A.; Olivieri, M.; Petris, G.; Montagna, C.; Reginato, G.; Maule, G.; Lorenzin, F.; Prandi, D.; Romanel, A.; Demichelis, F.; et al. A highly specific SpCas9 variant is identified by in vivo screening in yeast. Nat. Biotechnol. 2018, 36, 265–271. [CrossRef]
- Matsumoto D, Kishi K, Matsugi E, Inoue Y, Nigorikawa K, Nomura W. CAS9-GEMININ and Cdt1-fused ANTI-CRISPR protein synergistically increase editing accuracy. FEBS Lett. 2023 Apr;597(7):985–94.
- Stinson, B.M.; Loparo, J.J. Repair of DNA Double-Strand Breaks by the Nonhomologous End Joining Pathway. Annu. Rev. Biochem. 2021, 90, 137–164. [CrossRef]
- Sfeir A, Symington LS. Microhomology-Mediated End Joining: A Back-up Survival Mechanism or Dedicated Pathway? Trends Biochem Sci. 2015 Nov;40(11):701–14.
- Chen, J.; Li, S.; He, Y.; Li, J.; Xia, L. An update on precision genome editing by homology-directed repair in plants. Plant Physiol. 2022, 188, 1780–1794. [CrossRef]
- Xue, C.; Greene, E.C. DNA Repair Pathway Choices in CRISPR-Cas9-Mediated Genome Editing. Trends Genet. 2021, 37, 639–656. [CrossRef]
- Zhou, H.; Liu, B.; Weeks, D.P.; Spalding, M.H.; Yang, B. Large chromosomal deletions and heritable small genetic changes induced by CRISPR/Cas9 in rice. Nucleic Acids Res. 2014, 42, 10903–10914. [CrossRef]
- Hsieh-Feng, V.; Yang, Y. Efficient expression of multiple guide RNAs for CRISPR/Cas genome editing. aBIOTECH 2020, 1, 123–134. [CrossRef]
- Nakanishi, T.; Maekawa, A.; Tabata, H.; Yoshioka, T.; Pei, Z.; Sato, K.; Mori, M.; Kato, M.; Saito, I. Highly multiplex guide RNA expression units of CRISPR/Cas9 were completely stable using cosmid amplification in a novel polygonal structure. J. Gene Med. 2019, 21, e3115. [CrossRef]
- Saito, M.; Ladha, A.; Strecker, J.; Faure, G.; Neumann, E.; Altae-Tran, H.; Macrae, R.K.; Zhang, F. Dual modes of CRISPR-associated transposon homing. Cell 2021, 184, 2441–2453.e18. [CrossRef]
- Zhang, Y.; Ren, Q.; Tang, X.; Liu, S.; Malzahn, A.A.; Zhou, J.; Wang, J.; Yin, D.; Pan, C.; Yuan, M.; et al. Expanding the scope of plant genome engineering with Cas12a orthologs and highly multiplexable editing systems. Nat. Commun. 2021, 12, 1–11. [CrossRef]
- Ng WA, Reed BH. A method to estimate the frequency of chromosomal rearrangements induced by CRISPR/Cas9 multiplexing in Drosophila [Internet]. 2019 [cited 2025 Apr 17]. Available from: http://biorxiv.org/lookup/doi/10.1101/815431.
- Xie K, Minkenberg B, Yang Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proc Natl Acad Sci U S A. 2015 Mar 17;112(11):3570–5.
- Safari, F.; Zare, K.; Negahdaripour, M.; Barekati-Mowahed, M.; Ghasemi, Y. CRISPR Cpf1 proteins: structure, function and implications for genome editing. Cell Biosci. 2019, 9, 1–21. [CrossRef]
- Stuttmann J, Barthel K, Martin P, Ordon J, Erickson JL, Herr R, et al. Highly efficient multiplex editing: one-shot generation of 8× Nicotiana benthamiana and 12× Arabidopsis mutants. Plant J. 2021 Apr;106(1):8–22.
- He, Y.; Zhang, T.; Yang, N.; Xu, M.; Yan, L.; Wang, L.; Wang, R.; Zhao, Y. Self-cleaving ribozymes enable the production of guide RNAs from unlimited choices of promoters for CRISPR/Cas9 mediated genome editing. J. Genet. Genom. 2017, 44, 469–472. [CrossRef]
- Tang, X.; Zheng, X.; Qi, Y.; Zhang, D.; Cheng, Y.; Tang, A.; Voytas, D.F.; Zhang, Y. A Single Transcript CRISPR-Cas9 System for Efficient Genome Editing in Plants. Mol. Plant 2016, 9, 1088–1091. [CrossRef]
- Lee, H.Y.; Haurwitz, R.E.; Apffel, A.; Zhou, K.; Smart, B.; Wenger, C.D.; Laderman, S.; Bruhn, L.; Doudna, J.A. RNA–protein analysis using a conditional CRISPR nuclease. Proc. Natl. Acad. Sci. 2013, 110, 5416–5421. [CrossRef]
- Huang, S.; Zhang, Z.; Tao, W.; Liu, Y.; Li, X.; Wang, X.; Harati, J.; Wang, P.-Y.; Huang, X.; Lin, C.-P. Broadening prime editing toolkits using RNA-Pol-II-driven engineered pegRNA. Mol. Ther. 2022, 30, 2923–2932. [CrossRef]
- Das U, Banerjee S, Sarkar M. Bibliometric analysis of circular RNA cancer vaccines and their emerging impact. Vacunas. 2025 Mar;500391.
- ermák, T.; Curtin, S.J.; Gil-Humanes, J.; Čegan, R.; Kono, T.J.Y.; Konečná, E.; Belanto, J.J.; Starker, C.G.; Mathre, J.W.; Greenstein, R.L.; et al. A Multipurpose Toolkit to Enable Advanced Genome Engineering in Plants. Plant Cell 2017, 29, 1196–1217. [CrossRef]
- Naeem, M.; Majeed, S.; Hoque, M.Z.; Ahmad, I. Latest Developed Strategies to Minimize the Off-Target Effects in CRISPR-Cas-Mediated Genome Editing. Cells 2020, 9, 1608. [CrossRef]
- Xie, C.; Chen, Y.-L.; Wang, D.-F.; Wang, Y.-L.; Zhang, T.-P.; Li, H.; Liang, F.; Zhao, Y.; Zhang, G.-Y. SgRNA Expression of CRIPSR-Cas9 System Based on MiRNA Polycistrons as a Versatile Tool to Manipulate Multiple and Tissue-Specific Genome Editing. Sci. Rep. 2017, 7, 1–12. [CrossRef]
- Wu, S.; Kyaw, H.; Tong, Z.; Yang, Y.; Wang, Z.; Zhang, L.; Deng, L.; Zhang, Z.; Xiao, B.; Quick, W.P.; et al. A simple and efficient CRISPR/Cas9 system permits ultra-multiplex genome editing in plants. Crop. J. 2024, 12, 569–582. [CrossRef]
- Kannan, B.; Jung, J.H.; Moxley, G.W.; Lee, S.; Altpeter, F. TALEN-mediated targeted mutagenesis of more than 100 COMT copies/alleles in highly polyploid sugarcane improves saccharification efficiency without compromising biomass yield. Plant Biotechnol. J. 2017, 16, 856–866. [CrossRef]
- Yang, T.; Ali, M.; Lin, L.; Li, P.; He, H.; Zhu, Q.; Sun, C.; Wu, N.; Zhang, X.; Huang, T.; et al. Recoloring tomato fruit by CRISPR/Cas9-mediated multiplex gene editing. Hortic. Res. 2022, 10, uhac214. [CrossRef]
- Oh, Y.; Kim, S.-G. RPS5A Promoter-Driven Cas9 Produces Heritable Virus-Induced Genome Editing in Nicotiana attenuata. Mol. Cells 2021, 44, 911–919. [CrossRef]
- Kim, H.K.; Lee, S.; Kim, Y.; Park, J.; Min, S.; Choi, J.W.; Huang, T.P.; Yoon, S.; Liu, D.R.; Kim, H.H. High-throughput analysis of the activities of xCas9, SpCas9-NG and SpCas9 at matched and mismatched target sequences in human cells. Nat. Biomed. Eng. 2020, 4, 111–124. [CrossRef]
- Kor, S.D.; Chowdhury, N.; Keot, A.K.; Yogendra, K.; Chikkaputtaiah, C.; Reddy, P.S. RNA Pol III promoters—key players in precisely targeted plant genome editing. Front. Genet. 2023, 13, 989199. [CrossRef]
- Kuzmin, A.A.; Tomilin, A.N. Building Blocks of Artificial CRISPR-Based Systems beyond Nucleases. Int. J. Mol. Sci. 2022, 24, 397. [CrossRef]
- Kleinstiver BP, Sousa AA, Walton RT, Tak YE, Hsu JY, Clement K, et al. Engineered CRISPR-Cas12a variants with increased activities and improved targeting ranges for gene, epigenetic and base editing. Nat Biotechnol. 2019 Mar;37(3):276–82.
- McCarty, N.S.; Graham, A.E.; Studená, L.; Ledesma-Amaro, R. Multiplexed CRISPR technologies for gene editing and transcriptional regulation. Nat. Commun. 2020, 11, 1–13. [CrossRef]
- Geilenkeuser, J.; Armbrust, N.; Steinmaßl, E.; Du, S.W.; Schmidt, S.; Binder, E.M.H.; Li, Y.; Warsing, N.W.; Wendel, S.V.; von der Linde, F.; et al. Engineered nucleocytosolic vehicles for loading of programmable editors. Cell 2025. [CrossRef]
- Wang, L.; Han, H. Strategies for improving the genome-editing efficiency of class 2 CRISPR/Cas system. Heliyon 2024, 10, e38588. [CrossRef]
- Shui, S.; Wang, S.; Liu, J. Systematic Investigation of the Effects of Multiple SV40 Nuclear Localization Signal Fusion on the Genome Editing Activity of Purified SpCas9. Bioengineering 2022, 9, 83. [CrossRef]
- Tanaka, T.; Maeda, Y.; Suhaimi, N.; Tsuneoka, C.; Nonoyama, T.; Yoshino, T.; Kato, N.; Lauersen, K.J. Intron-mediated enhancement of transgene expression in the oleaginous diatom Fistulifera solaris towards bisabolene production. Algal Res. 2021, 57. [CrossRef]
- Grützner, R.; Martin, P.; Horn, C.; Mortensen, S.; Cram, E.J.; Lee-Parsons, C.W.; Stuttmann, J.; Marillonnet, S. High-efficiency genome editing in plants mediated by a Cas9 gene containing multiple introns. Plant Commun. 2021, 2, 100135. [CrossRef]
- Khakhar, A.; Voytas, D.F. RNA Viral Vectors for Accelerating Plant Synthetic Biology. Front. Plant Sci. 2021, 12. [CrossRef]
- Amack, S.C.; Antunes, M.S. CaMV35S promoter – A plant biology and biotechnology workhorse in the era of synthetic biology. Curr. Plant Biol. 2020, 24. [CrossRef]
- Wang, X.; Xu, L.; Liu, X.; Xin, L.; Wu, S.; Chen, X. Development of potent promoters that drive the efficient expression of genes in apple protoplasts. Hortic. Res. 2021, 8, 211. [CrossRef]
- Ren, Q.; Zhong, Z.; Wang, Y.; You, Q.; Li, Q.; Yuan, M.; He, Y.; Qi, C.; Tang, X.; Zheng, X.; et al. Bidirectional Promoter-Based CRISPR-Cas9 Systems for Plant Genome Editing. Front. Plant Sci. 2019, 10, 1173. [CrossRef]
- Collonnier C, Epert A, Mara K, Maclot F, Guyon-Debast A, Charlot F, et al. CRISPR-Cas9-mediated efficient directed mutagenesis and RAD51-dependent and RAD51-independent gene targeting in the moss Physcomitrella patens. Plant Biotechnol J. 2017 Jan;15(1):122–31.
- Mao Y, Zhang Z, Feng Z, Wei P, Zhang H, Botella JR, et al. Development of germ-line-specific CRISPR-Cas9 systems to improve the production of heritable gene modifications in Arabidopsis. Plant Biotechnol J. 2016 Feb;14(2):519–32.
- Das, U.; Chanda, T.; Kumar, J.; Peter, A. Discovery of natural MCL1 inhibitors using pharmacophore modelling, QSAR, docking, ADMET, molecular dynamics, and DFT analysis. Comput. Biol. Chem. 2025, 117, 108427. [CrossRef]
- Zheng, N.; Li, T.; Dittman, J.D.; Su, J.; Li, R.; Gassmann, W.; Peng, D.; Whitham, S.A.; Liu, S.; Yang, B. CRISPR/Cas9-Based Gene Editing Using Egg Cell-Specific Promoters in Arabidopsis and Soybean. Front. Plant Sci. 2020, 11, 800. [CrossRef]
- Kelliher, T.; Starr, D.; Su, X.; Tang, G.; Chen, Z.; Carter, J.; Wittich, P.E.; Dong, S.; Green, J.; Burch, E.; et al. One-step genome editing of elite crop germplasm during haploid induction. Nat. Biotechnol. 2019, 37, 287–292. [CrossRef]
- Bird, J.E.; Marles-Wright, J.; Giachino, A. A User’s Guide to Golden Gate Cloning Methods and Standards. ACS Synth. Biol. 2022, 11, 3551–3563. [CrossRef]
- Weber, E.; Engler, C.; Gruetzner, R.; Werner, S.; Marillonnet, S.; Peccoud, J. A Modular Cloning System for Standardized Assembly of Multigene Constructs. PLOS ONE 2011, 6, e16765. [CrossRef]
- Sarrion-Perdigones, A.; Falconi, E.E.; Zandalinas, S.I.; Juárez, P.; Fernández-Del-Carmen, A.; Granell, A.; Orzaez, D.; Peccoud, J. GoldenBraid: An Iterative Cloning System for Standardized Assembly of Reusable Genetic Modules. PLOS ONE 2011, 6, e21622. [CrossRef]
- Lowder, L.G.; Zhang, D.; Baltes, N.J.; Paul, J.W.; Tang, X.; Zheng, X.; Voytas, D.F.; Hsieh, T.-F.; Zhang, Y.; Qi, Y. A CRISPR/Cas9 Toolbox for Multiplexed Plant Genome Editing and Transcriptional Regulation. Plant Physiol. 2015, 169, 971–985. [CrossRef]
- Vasudevan, R.; Gale, G.A.; Schiavon, A.A.; Puzorjov, A.; Malin, J.; Gillespie, M.D.; Vavitsas, K.; Zulkower, V.; Wang, B.; Howe, C.J.; et al. CyanoGate: A Modular Cloning Suite for Engineering Cyanobacteria Based on the Plant MoClo Syntax. Plant Physiol. 2019, 180, 39–55. [CrossRef]
- Fulco, C.P.; Nasser, J.; Jones, T.R.; Munson, G.; Bergman, D.T.; Subramanian, V.; Grossman, S.R.; Anyoha, R.; Doughty, B.R.; Patwardhan, T.A.; et al. Activity-by-contact model of enhancer–promoter regulation from thousands of CRISPR perturbations. Nat. Genet. 2019, 51, 1664–1669. [CrossRef]
- Costa, L.D.; Piazza, S.; Pompili, V.; Salvagnin, U.; Cestaro, A.; Moffa, L.; Vittani, L.; Moser, C.; Malnoy, M. Strategies to produce T-DNA free CRISPRed fruit trees via Agrobacterium tumefaciens stable gene transfer. Sci. Rep. 2020, 10, 1–14. [CrossRef]
- Knopp, Y.; Geis, F.K.; Heckl, D.; Horn, S.; Neumann, T.; Kuehle, J.; Meyer, J.; Fehse, B.; Baum, C.; Morgan, M.; et al. Transient Retrovirus-Based CRISPR/Cas9 All-in-One Particles for Efficient, Targeted Gene Knockout. Mol. Ther. - Nucleic Acids 2018, 13, 256–274. [CrossRef]
- Thompson, M.G.; Moore, W.M.; Hummel, N.F.; Pearson, A.N.; Barnum, C.R.; Scheller, H.V.; Shih, P.M. Agrobacterium tumefaciens: A Bacterium Primed for Synthetic Biology. 2020, 2020. [CrossRef]
- Tang, Y.; Zhang, Z.; Yang, Z.; Wu, J.; Manavella, P. CRISPR/Cas9 and Agrobacterium tumefaciens virulence proteins synergistically increase efficiency of precise genome editing via homology directed repair in plants. J. Exp. Bot. 2023, 74, 3518–3530. [CrossRef]
- He, Y.; Mudgett, M.; Zhao, Y. Advances in gene editing without residual transgenes in plants. Plant Physiol. 2021, 188, 1757–1768. [CrossRef]
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Liu, Y.-G.; Zhao, K.; A Wilson, R. Enhanced Rice Blast Resistance by CRISPR/Cas9-Targeted Mutagenesis of the ERF Transcription Factor Gene OsERF922. PLOS ONE 2016, 11, e0154027–e0154027. [CrossRef]
- Zhong, H.; Li, C.; Yu, W.; Zhou, H.-P.; Lieber, T.; Su, X.; Wang, W.; Bumann, E.; Castro, R.M.L.; Jiang, Y.; et al. A fast and genotype-independent in planta Agrobacterium-mediated transformation method for soybean. Plant Commun. 2024, 5, 101063. [CrossRef]
- Lacroix, B.; Citovsky, V. Biolistic Approach for Transient Gene Expression Studies in Plants. Methods Mol Biol Clifton NJ. 2020;2124:125–39.
- Das, U. Emerging trends and research landscape of the tumor microenvironment in head-and-neck cancer: A comprehensive bibliometric analysis. Cancer Plus 2025, 7, 64. [CrossRef]
- Vollen, K.; Alonso, J.M.; Stepanova, A.N. Beyond a few bases: methods for large DNA insertion and gene targeting in plants. Plant J. 2025, 121, e70099. [CrossRef]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Cigan, A.M. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [CrossRef]
- Jiao, Y.; Peluso, P.; Shi, J.; Liang, T.; Stitzer, M.C.; Wang, B.; Campbell, M.S.; Stein, J.C.; Wei, X.; Chin, C.-S.; et al. Improved maize reference genome with single-molecule technologies. Nature 2017, 546, 524–527. [CrossRef]
- Reed, K.M.; Bargmann, B.O.R. Protoplast Regeneration and Its Use in New Plant Breeding Technologies. Front. Genome Ed. 2021, 3. [CrossRef]
- Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.; Ohlsson, P.; Gonzalez, M.N.; Samuelsson, M.; Hofvander, P. Genome editing in potato via CRISPR-Cas9 ribonucleoprotein delivery. Physiol. Plant. 2018, 164, 378–384. [CrossRef]
- Wu X, Zhang Y, Jiang X, Ma T, Guo Y, Wu X, et al. Considerations in engineering viral vectors for genome editing in plants. Virology. 2024 Jan;589:109922.
- Oh, Y.; Nagalakshmi, U.; Dahlbeck, D.; Koehler, N.; Cho, M.; Dinesh-Kumar, S.P.; Staskawicz, B.J. Heritable virus-induced germline editing in tomato. Plant J. 2025, 122, e70115. [CrossRef]
- Zhang, C.; Liu, S.; Li, X.; Zhang, R.; Li, J. Virus-Induced Gene Editing and Its Applications in Plants. Int. J. Mol. Sci. 2022, 23, 10202. [CrossRef]
- Brokowski, C.; Adli, M. CRISPR Ethics: Moral Considerations for Applications of a Powerful Tool. J. Mol. Biol. 2019, 431, 88–101. [CrossRef]
- Bertier LD, Ron M, Huo H, Bradford KJ, Britt AB, Michelmore RW. High-Resolution Analysis of the Efficiency, Heritability, and Editing Outcomes of CRISPR/Cas9-Induced Modifications of NCED4 in Lettuce ( Lactuca sativa ). G3 GenesGenomesGenetics. 2018 May 1;8(5):1513–21.
- Vats, S.; Kumawat, S.; Brar, J.; Kaur, S.; Yadav, K.; Magar, S.G.; Jadhav, P.V.; Salvi, P.; Sonah, H.; Sharma, S.; et al. Opportunity and challenges for nanotechnology application for genome editing in plants. Plant Nano Biol. 2022, 1, 100001. [CrossRef]
- Liu, S.; Zheng, Y.; Pan, L.; Wang, W.; Li, Y.; Liu, Z.; Zhang, X. Nanodelivery of nucleic acids for plant genetic engineering. Nanoscale Res. Lett. 2025, 20, 1–15. [CrossRef]
- Zhi, H.; Zhou, S.; Pan, W.; Shang, Y.; Zeng, Z.; Zhang, H. The Promising Nanovectors for Gene Delivery in Plant Genome Engineering. Int. J. Mol. Sci. 2022, 23, 8501. [CrossRef]
- Zhao, X.; Meng, Z.; Wang, Y.; Chen, W.; Sun, C.; Cui, B.; Cui, J.; Yu, M.; Zeng, Z.; Guo, S.; et al. Pollen magnetofection for genetic modification with magnetic nanoparticles as gene carriers. Nat. Plants 2017, 3, 956–964. [CrossRef]
- Babaeianjelodar, N.; Trivedi, J.; Uhde-Stone, C. Eliminating Tissue Culture from Plant Gene Editing in the Near Future: A Wish or Reality?. Curr. Plant Biol. 2025, 41. [CrossRef]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.-J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic Regulators Baby boom and Wuschel Improve Monocot Transformation. Plant Cell 2016, 28, 1998–2015. [CrossRef]
- Bent, A. Arabidopsis thaliana Floral Dip Transformation Method. In: Agrobacterium Protocols [Internet]. New Jersey: Humana Press; 2006 [cited 2025 Apr 17]. p. 87–104. Available from: http://link.springer.com/10.1385/1-59745-130-4:87.
- Saini, H.; Thakur, R.; Gill, R.; Tyagi, K.; Goswami, M. CRISPR/Cas9-gene editing approaches in plant breeding. GM Crop. Food 2023, 14, 1–17. [CrossRef]
- Quiroz, L.F.; Khan, M.; Gondalia, N.; Lai, L.; McKeown, P.C.; Brychkova, G.; Spillane, C. Tissue culture-independent approaches to revolutionizing plant transformation and gene editing. Hortic. Res. 2024, 12, uhae292. [CrossRef]
- Miao, C.; Xiao, L.; Hua, K.; Zou, C.; Zhao, Y.; Bressan, R.A.; Zhu, J.-K. Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proc. Natl. Acad. Sci. 2018, 115, 6058–6063. [CrossRef]
- Zeng, Y.; Wen, J.; Zhao, W.; Wang, Q.; Huang, W. Rational Improvement of Rice Yield and Cold Tolerance by Editing the Three Genes OsPIN5b, GS3, and OsMYB30 With the CRISPR–Cas9 System. Front. Plant Sci. 2020, 10, 1663. [CrossRef]
- Zsögön, A.; Čermák, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.F.; Kudla, J.; Peres, L.E.P. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018, 36, 1211–1216. [CrossRef]
- Brant EJ, Eid A, Kannan B, Baloglu MC, Altpeter F. The extent of multiallelic, co-editing of LIGULELESS1 in highly polyploid sugarcane tunes leaf inclination angle and enables selection of the ideotype for biomass yield. Plant Biotechnol J. 2024 Oct;22(10):2660–71.
- Zaman, Q.U.; Chu, W.; Hao, M.; Shi, Y.; Sun, M.; Sang, S.-F.; Mei, D.; Cheng, H.; Liu, J.; Li, C.; et al. CRISPR/Cas9-Mediated Multiplex Genome Editing of JAGGED Gene in Brassica napus L.. Biomolecules 2019, 9, 725. [CrossRef]
- Shi, J.; Ni, X.; Huang, J.; Fu, Y.; Wang, T.; Yu, H.; Zhang, Y. CRISPR/Cas9-Mediated Gene Editing of BnFAD2 and BnFAE1 Modifies Fatty Acid Profiles in Brassica napus. Genes 2022, 13, 1681. [CrossRef]
- Zhu, X.; Gong, H.; He, Q.; Zeng, Z.; Busse, J.S.; Jin, W.; Bethke, P.C.; Jiang, J. Silencing of vacuolar invertase and asparagine synthetase genes and its impact on acrylamide formation of fried potato products. Plant Biotechnol. J. 2015, 14, 709–718. [CrossRef]
- Biswas S, Ibarra O, Shaphek M, Molina-Risco M, Faion-Molina M, Bellinatti-Della Gracia M, et al. Increasing the level of resistant starch in ‘Presidio’ rice through multiplex CRISPR–Cas9 gene editing of starch branching enzyme genes. Plant Genome. 2023 Jun;16(2):e20225.
- Peng A, Chen S, Lei T, Xu L, He Y, Wu L, et al. Engineering canker-resistant plants through CRISPR /Cas9-targeted editing of the susceptibility gene Cs LOB 1 promoter in citrus. Plant Biotechnol J. 2017 Dec;15(12):1509–19.
- Das, U.; Uttarkar, A.; Kumar, J.; Niranjan, V. In silico exploration natural compounds for the discovery of novel DNMT3A inhibitors as potential therapeutic agents for acute myeloid leukemia. Silico Res. Biomed. 2025, 1. [CrossRef]
- Macioszek, V.K.; Jęcz, T.; Ciereszko, I.; Kononowicz, A.K. Jasmonic Acid as a Mediator in Plant Response to Necrotrophic Fungi. Cells 2023, 12, 1027. [CrossRef]
- Li, M.; Yang, Z.; Chang, C. Susceptibility Is New Resistance: Wheat Susceptibility Genes and Exploitation in Resistance Breeding. Agriculture 2022, 12, 1419. [CrossRef]
- Sun, Q.; Lin, L.; Liu, D.; Wu, D.; Fang, Y.; Wu, J.; Wang, Y. CRISPR/Cas9-Mediated Multiplex Genome Editing of the BnWRKY11 and BnWRKY70 Genes in Brassica napus L.. Int. J. Mol. Sci. 2018, 19, 2716. [CrossRef]
- Kang, Y.; Zhou, M.; Merry, A.; Barry, K. Mechanisms of powdery mildew resistance of wheat – a review of molecular breeding. Plant Pathol. 2020, 69, 601–617. [CrossRef]
- Shawky, A.; Hatawsh, A.; Al-Saadi, N.; Farzan, R.; Eltawy, N.; Francis, M.; Abousamra, S.; Ismail, Y.Y.; Attia, K.; Fakhouri, A.S.; et al. Revolutionizing Tomato Cultivation: CRISPR/Cas9 Mediated Biotic Stress Resistance. Plants 2024, 13, 2269. [CrossRef]
- Xie, Z.; Jin, L.; Sun, Y.; Zhan, C.; Tang, S.; Qin, T.; Liu, N.; Huang, J. OsNAC120 balances plant growth and drought tolerance by integrating GA and ABA signaling in rice. Plant Commun. 2023, 5, 100782. [CrossRef]
- Ding, F.; Wang, C.; Xu, N.; Wang, M. The ethylene response factor SlERF.B8 triggers jasmonate biosynthesis to promote cold tolerance in tomato. Environ. Exp. Bot. 2022, 203. [CrossRef]
- Chen, S.; Zhang, N.; Zhou, G.; Hussain, S.; Ahmed, S.; Tian, H.; Wang, S. Knockout of the entire family of AITR genes in Arabidopsis leads to enhanced drought and salinity tolerance without fitness costs. BMC Plant Biol. 2021, 21, 1–15. [CrossRef]
- Abdallah, N.A.; Elsharawy, H.; Abulela, H.A.; Thilmony, R.; Abdelhadi, A.A.; Elarabi, N.I. Multiplex CRISPR/Cas9-mediated genome editing to address drought tolerance in wheat. GM Crop. Food 2022, 16, 1–17. [CrossRef]
- Patel-Tupper, D.; Kelikian, A.; Leipertz, A.; Maryn, N.; Tjahjadi, M.; Karavolias, N.G.; Cho, M.-J.; Niyogi, K.K. Multiplexed CRISPR-Cas9 mutagenesis of rice PSBS1 noncoding sequences for transgene-free overexpression. Sci. Adv. 2024, 10, eadm7452. [CrossRef]
- Rahim, A.A.; Uzair, M.; Rehman, N.; Fiaz, S.; Attia, K.A.; Abushady, A.M.; Yang, S.H.; Khan, M.R. CRISPR/Cas9 mediated TaRPK1 root architecture gene mutagenesis confers enhanced wheat yield. J. King Saud Univ. - Sci. 2024, 36. [CrossRef]
- Khan MS, Ahmad D, Khan MA. Utilization of genes encoding osmoprotectants in transgenic plants for enhanced abiotic stress tolerance. Electron J Biotechnol. 2015 Jul;18(4):257–66.
- Zhang, J.; Lyu, H.; Chen, J.; Cao, X.; Du, R.; Ma, L.; Wang, N.; Zhu, Z.; Rao, J.; Wang, J.; et al. Releasing a sugar brake generates sweeter tomato without yield penalty. Nature 2024, 635, 647–656. [CrossRef]
- Li, A.; Jia, S.; Yobi, A.; Ge, Z.; Sato, S.J.; Zhang, C.; Angelovici, R.; Clemente, T.E.; Holding, D.R. Editing of an Alpha-Kafirin Gene Family Increases, Digestibility and Protein Quality in Sorghum. Plant Physiol. 2018, 177, 1425–1438. [CrossRef]
- Wang L, O’Conner S, Tanvir R, Zheng W, Cothron S, Towery K, et al. CRISPR /Cas9-based editing of NF-YC4 promoters yields high-protein rice and soybean. New Phytol. 2025 Mar;245(5):2103–16.
- Liu, H.; Lin, B.; Ren, Y.; Hao, P.; Huang, L.; Xue, B.; Jiang, L.; Zhu, Y.; Hua, S. CRISPR/Cas9-mediated editing of double loci of BnFAD2 increased the seed oleic acid content of rapeseed (Brassica napus L.). Front. Plant Sci. 2022, 13, 1034215. [CrossRef]
- Sánchez-León, S.; Marín-Sanz, M.; Guzmán-López, M.H.; Gavilán-Camacho, M.; Simón, E.; Barro, F.; Trafford, K. CRISPR/Cas9-mediated multiplex gene editing of gamma and omega gliadins: paving the way for gliadin-free wheat. J. Exp. Bot. 2024, 75, 7079–7095. [CrossRef]
- Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Van Eck, J.; Lippman, Z.B. Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 2018, 4, 766–770. [CrossRef]
- Feng, X.; Lin, K.; Zhang, W.; Nan, J.; Zhang, X.; Wang, C.; Wang, R.; Jiang, G.; Yuan, Q.; Lin, S. Improving the blast resistance of the elite rice variety Kongyu-131 by updating the pi21 locus. BMC Plant Biol. 2019, 19, 1–12. [CrossRef]
- Wang H, Jones B, Li Z, Frasse P, Delalande C, Regad F, et al. The Tomato Aux / IAA Transcription Factor IAA9 Is Involved in Fruit Development and Leaf Morphogenesis. Plant Cell. 2005 Oct 3;17(10):2676–92.
- Guo, C.; Ma, X.; Gao, F.; Guo, Y. Off-target effects in CRISPR/Cas9 gene editing. Front. Bioeng. Biotechnol. 2023, 11, 1143157. [CrossRef]
- Lei, Y.; Lu, L.; Liu, H.-Y.; Li, S.; Xing, F.; Chen, L.-L. CRISPR-P: A Web Tool for Synthetic Single-Guide RNA Design of CRISPR-System in Plants. Mol. Plant 2014, 7, 1494–1496. [CrossRef]
- Labun, K.; Montague, T.G.; Krause, M.; Torres Cleuren, Y.N.; Tjeldnes, H.; Valen, E. CHOPCHOP v3: expanding the CRISPR web toolbox beyond genome editing. Nucleic Acids Res. 2019, 47, W171–W174. [CrossRef]
- Concordet, J.-P.; Haeussler, M. CRISPOR: intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic Acids Res. 2018, 46, W242–W245. [CrossRef]
- Slaymaker, I.M.; Gao, L.; Zetsche, B.; Scott, D.A.; Yan, W.X.; Zhang, F. Rationally engineered Cas9 nucleases with improved specificity. Science 2016, 351, 84–88. [CrossRef]
- Son, S.; Park, S.R. Challenges Facing CRISPR/Cas9-Based Genome Editing in Plants. Front. Plant Sci. 2022, 13, 902413. [CrossRef]
- Zhou, J.; Luan, X.; Liu, Y.; Wang, L.; Wang, J.; Yang, S.; Liu, S.; Zhang, J.; Liu, H.; Yao, D. Strategies and Methods for Improving the Efficiency of CRISPR/Cas9 Gene Editing in Plant Molecular Breeding. Plants 2023, 12, 1478. [CrossRef]
- Wen, W.; Zhang, X.-B. CRISPR–Cas9 gene editing induced complex on-target outcomes in human cells. Exp. Hematol. 2022, 110, 13–19. [CrossRef]
- Yin, J.; Lu, R.; Xin, C.; Wang, Y.; Ling, X.; Li, D.; Zhang, W.; Liu, M.; Xie, W.; Kong, L.; et al. Cas9 exo-endonuclease eliminates chromosomal translocations during genome editing. Nat. Commun. 2022, 13, 1–14. [CrossRef]
- Iyer, S.V.; Goodwin, S.; McCombie, W.R. Leveraging the power of long reads for targeted sequencing. Genome Res. 2024, 34, 1701–1718. [CrossRef]
- Santelices, B. Mosaicism and chimerism as components of intraorganismal genetic heterogeneity. J. Evol. Biol. 2004, 17, 1187–1188. [CrossRef]
- Wang, Z.-P.; Xing, H.-L.; Dong, L.; Zhang, H.-Y.; Han, C.-Y.; Wang, X.-C.; Chen, Q.-J. Egg cell-specific promoter-controlled CRISPR/Cas9 efficiently generates homozygous mutants for multiple target genes in Arabidopsis in a single generation. Genome Biol. 2015, 16, 1–12. [CrossRef]
- Tenorio Berrío R, Nelissen H, Inzé D, Dubois M. Increasing yield on dry fields: molecular pathways with growing potential. Plant J Cell Mol Biol. 2022 Jan;109(2):323–41.
- Chuai, G.; Ma, H.; Yan, J.; Chen, M.; Hong, N.; Xue, D.; Zhou, C.; Zhu, C.; Chen, K.; Duan, B.; et al. DeepCRISPR: optimized CRISPR guide RNA design by deep learning. Genome Biol. 2018, 19, 1–18. [CrossRef]
- Kim HK, Kim Y, Lee S, Min S, Bae JY, Choi JW, et al. SpCas9 activity prediction by DeepSpCas9, a deep learning–based model with high generalization performance. Sci Adv. 2019 Nov;5(11):eaax9249.
- Xu, H.; Xiao, T.; Chen, C.-H.; Li, W.; Meyer, C.A.; Wu, Q.; Wu, D.; Cong, L.; Zhang, F.; Liu, J.S.; et al. Sequence determinants of improved CRISPR sgRNA design. Genome Res. 2015, 25, 1147–1157. [CrossRef]
- Anthon C, Corsi GI, Gorodkin J. CRISPRon/off: CRISPR/Cas9 on- and off-target gRNA design. Valencia A, editor. Bioinformatics. 2022 Dec 13;38(24):5437–9.
- Bae, S.; Park, J.; Kim, J.S. Cas-OFFinder: a fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 2014, 30, 1473–1475. [CrossRef]
- Hsu, J.Y.; Grünewald, J.; Szalay, R.; Shih, J.; Anzalone, A.V.; Lam, K.C.; Shen, M.W.; Petri, K.; Liu, D.R.; Joung, J.K.; et al. PrimeDesign software for rapid and simplified design of prime editing guide RNAs. Nat. Commun. 2021, 12, 1–6. [CrossRef]
- Hwang, G.-H.; Jeong, Y.K.; Habib, O.; Hong, S.-A.; Lim, K.; Kim, J.-S.; Bae, S. PE-Designer and PE-Analyzer: web-based design and analysis tools for CRISPR prime editing. Nucleic Acids Res. 2021, 49, W499–W504. [CrossRef]
- Stemmer M, Thumberger T, Del Sol Keyer M, Wittbrodt J, Mateo JL. CCTop: An Intuitive, Flexible and Reliable CRISPR/Cas9 Target Prediction Tool. Maas S, editor. PLOS ONE. 2015 Apr 24;10(4):e0124633.
- Meng, X.; Noyes, M.B.; Zhu, L.J.; Lawson, N.D.; A Wolfe, S. Targeted gene inactivation in zebrafish using engineered zinc-finger nucleases. Nat. Biotechnol. 2008, 26, 695–701. [CrossRef]
Figure 1.
Overview of CRISPR–Cas systems: origin, classification, and engineered variants for genome editing. (A) Prokaryotic immune origins of CRISPR systems: the CRISPR locus consists of alternating repeat and spacer sequences derived from invading nucleic acids (e.g., phages), which are transcribed and processed into CRISPR RNAs (crRNAs) to guide Cas nucleases for specific cleavage of foreign DNA. (B) Classification of CRISPR–Cas systems into Class 1 (multi-component complexes such as Cascade–Cas3) and Class 2 (single-effector proteins including Cas9, Cas12, and Cas13). Class 2 systems are more tractable for genome editing due to their simplicity. (C) Engineered SpCas9 and other Cas variants for broadened PAM recognition, RNA targeting, and compact delivery. Shown are widely used variants including SpCas9-NG (NGN PAM), xCas9 3.7 (expanded targeting), SpRY (near-PAMless), SaCas9 (compact form), Cas12a (staggered DSB), and Cas13 (RNA-targeting), along with hypercompact systems like CasΦ and Cas14. These tools are being extensively harnessed in plant genome engineering.
Figure 1.
Overview of CRISPR–Cas systems: origin, classification, and engineered variants for genome editing. (A) Prokaryotic immune origins of CRISPR systems: the CRISPR locus consists of alternating repeat and spacer sequences derived from invading nucleic acids (e.g., phages), which are transcribed and processed into CRISPR RNAs (crRNAs) to guide Cas nucleases for specific cleavage of foreign DNA. (B) Classification of CRISPR–Cas systems into Class 1 (multi-component complexes such as Cascade–Cas3) and Class 2 (single-effector proteins including Cas9, Cas12, and Cas13). Class 2 systems are more tractable for genome editing due to their simplicity. (C) Engineered SpCas9 and other Cas variants for broadened PAM recognition, RNA targeting, and compact delivery. Shown are widely used variants including SpCas9-NG (NGN PAM), xCas9 3.7 (expanded targeting), SpRY (near-PAMless), SaCas9 (compact form), Cas12a (staggered DSB), and Cas13 (RNA-targeting), along with hypercompact systems like CasΦ and Cas14. These tools are being extensively harnessed in plant genome engineering.
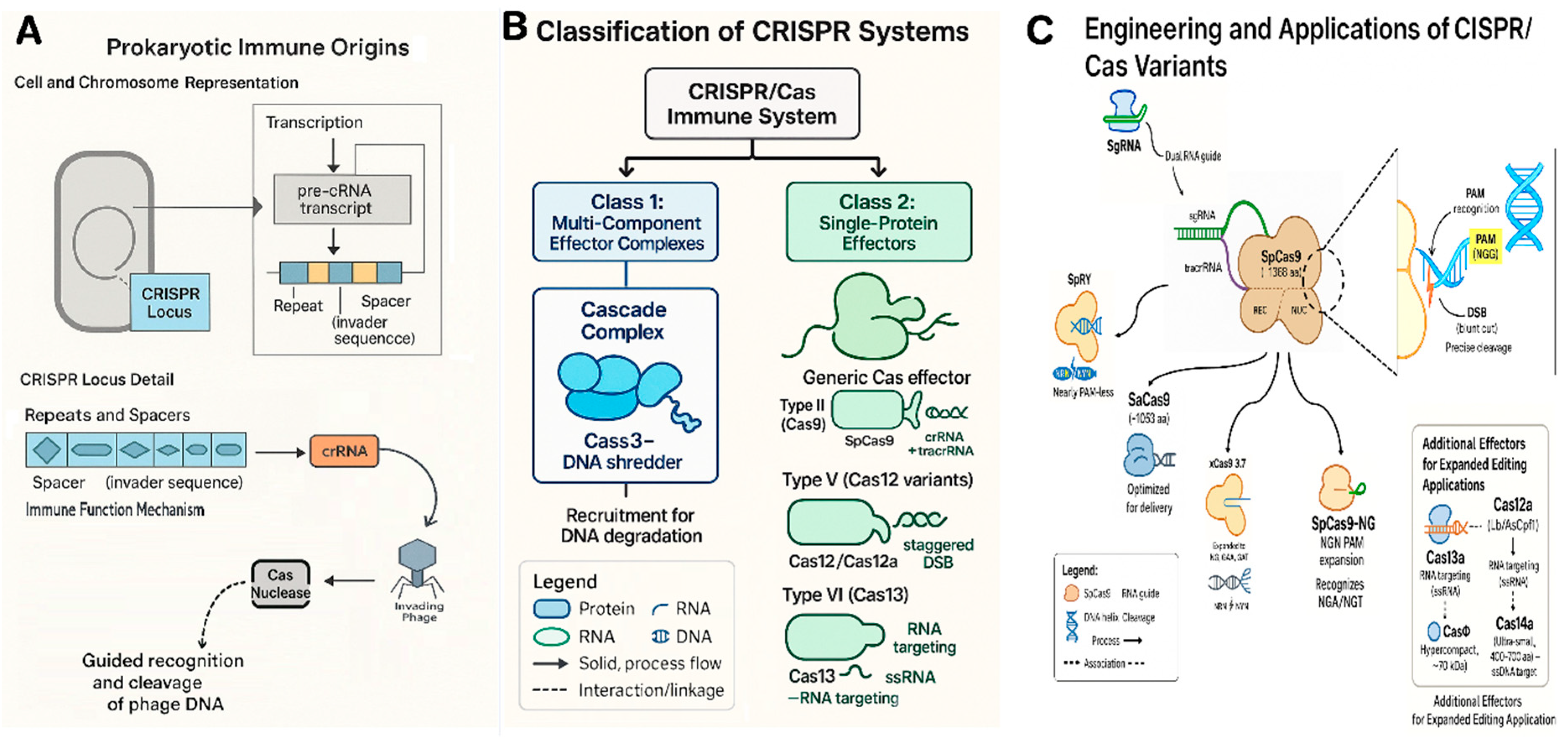
Figure 2.
DNA repair pathways following CRISPR-induced double-strand breaks (DSBs) and strategies for multiplex guide RNA (gRNA) expression. (A) Three major DNA DSB repair mechanisms in plant cells: non-homologous end joining (NHEJ), the dominant and error-prone repair pathway; homology-directed repair (HDR), which uses a donor template for precise edits; and microhomology-mediated end joining (MMEJ), which leverages short homologous sequences to mediate predictable deletions. (B) Overview of polycistronic gRNA expression strategies used for multiplex CRISPR editing in plants. These include multiple U6 promoters, the tRNA–gRNA (PTG) system, ribozyme-based self-processing, Csy4-mediated cleavage, miRNA-based expression (Drosha/Dicer), and the Cas12a array approach. Each method facilitates the expression of multiple sgRNAs or crRNAs from a single construct using distinct RNA processing mechanisms.
Figure 2.
DNA repair pathways following CRISPR-induced double-strand breaks (DSBs) and strategies for multiplex guide RNA (gRNA) expression. (A) Three major DNA DSB repair mechanisms in plant cells: non-homologous end joining (NHEJ), the dominant and error-prone repair pathway; homology-directed repair (HDR), which uses a donor template for precise edits; and microhomology-mediated end joining (MMEJ), which leverages short homologous sequences to mediate predictable deletions. (B) Overview of polycistronic gRNA expression strategies used for multiplex CRISPR editing in plants. These include multiple U6 promoters, the tRNA–gRNA (PTG) system, ribozyme-based self-processing, Csy4-mediated cleavage, miRNA-based expression (Drosha/Dicer), and the Cas12a array approach. Each method facilitates the expression of multiple sgRNAs or crRNAs from a single construct using distinct RNA processing mechanisms.
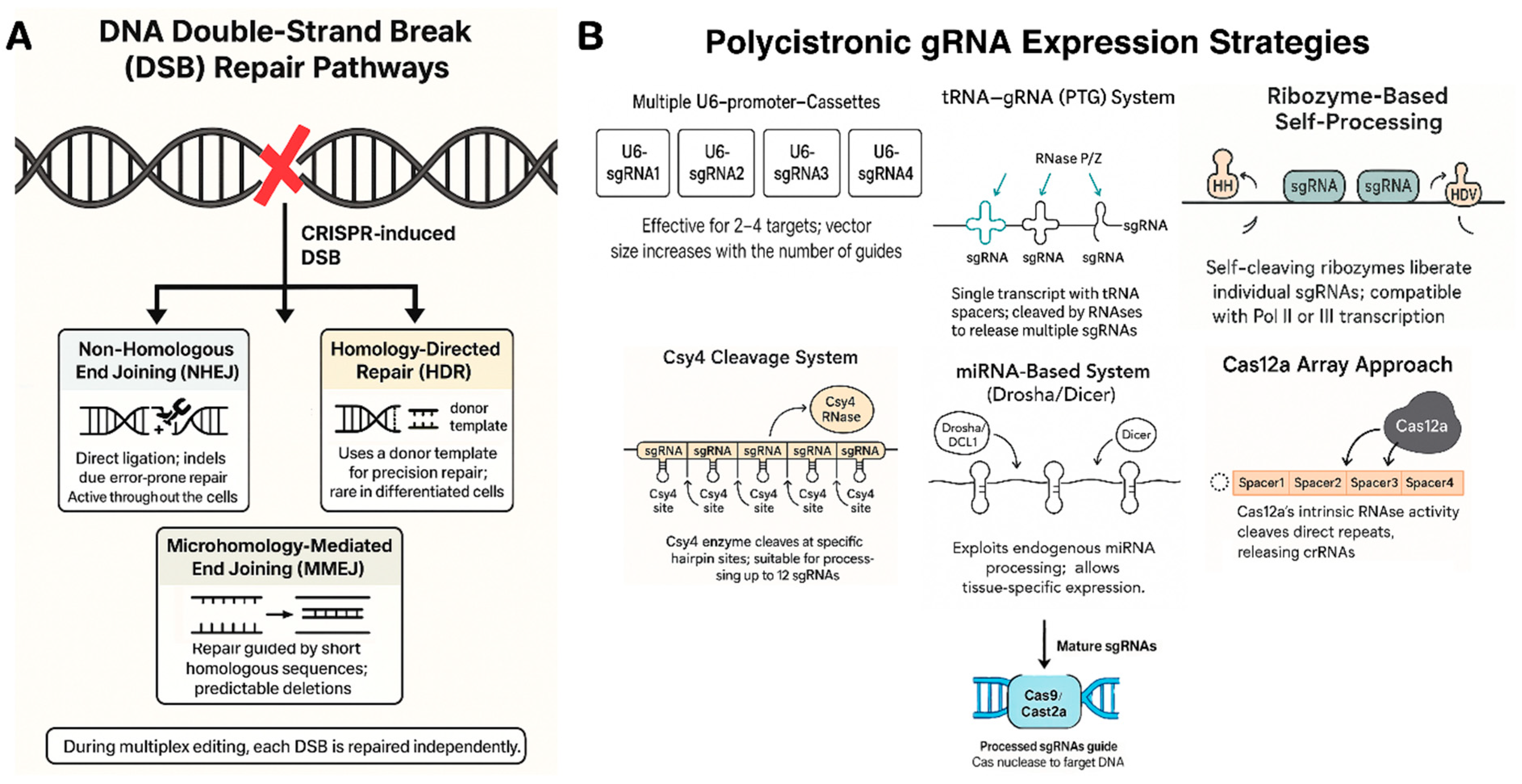
Figure 3.
Expression strategies and modular assembly for multiplexed CRISPR/Cas delivery in plants. (A) Pol III-driven expression of sgRNAs using U6 promoters, suitable for constitutive, non-polyadenylated guide expression, commonly paired with T-DNA vectors. (B) Pol II-driven all-in-one constructs encoding Cas9 and sgRNA arrays within a single transcript, leveraging 2A peptides and ribozyme or Csy4-based processing for efficient guide maturation. (C) Modular assembly of CRISPR vectors using systems like Golden Gate or MoClo, enabling combinatorial cloning from standard parts (promoters, sgRNA scaffolds, terminators) into T-DNA vectors for delivery. Delivery formats include stable Agrobacterium-mediated transformation or transient/virus-based expression for flexibility across plant systems.
Figure 3.
Expression strategies and modular assembly for multiplexed CRISPR/Cas delivery in plants. (A) Pol III-driven expression of sgRNAs using U6 promoters, suitable for constitutive, non-polyadenylated guide expression, commonly paired with T-DNA vectors. (B) Pol II-driven all-in-one constructs encoding Cas9 and sgRNA arrays within a single transcript, leveraging 2A peptides and ribozyme or Csy4-based processing for efficient guide maturation. (C) Modular assembly of CRISPR vectors using systems like Golden Gate or MoClo, enabling combinatorial cloning from standard parts (promoters, sgRNA scaffolds, terminators) into T-DNA vectors for delivery. Delivery formats include stable Agrobacterium-mediated transformation or transient/virus-based expression for flexibility across plant systems.
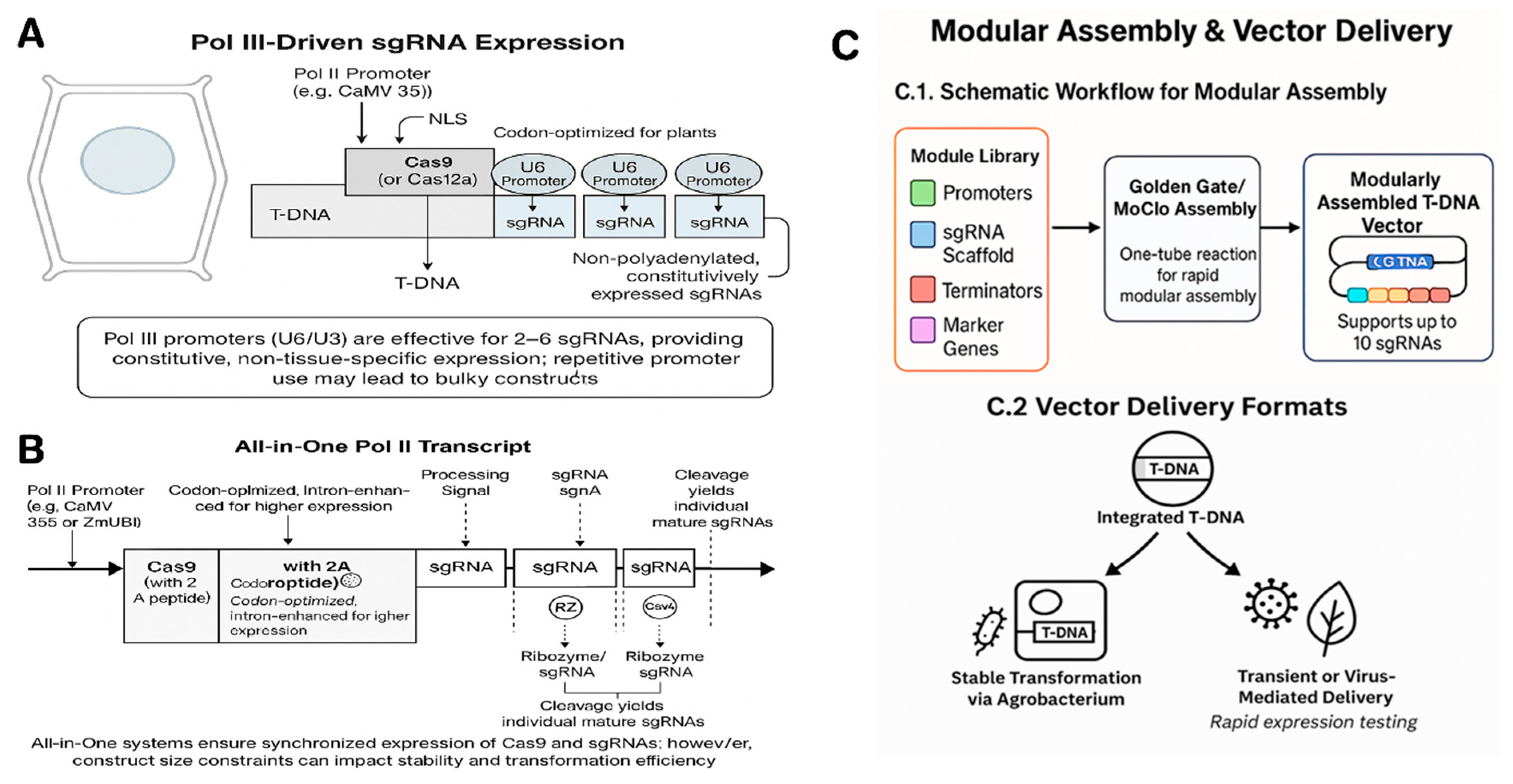
Figure 4.
Delivery platforms for CRISPR-Cas genome editing in plants. A variety of methods are employed to introduce CRISPR components into plant cells, each suited to different species, tissues, and experimental objectives. (A) Agrobacterium tumefaciens-mediated delivery of T-DNA vectors carrying Cas9 and sgRNA cassettes enables stable transformation in many dicots and optimized monocots. (B) Biolistic (particle bombardment) delivery propels CRISPR plasmids or RNPs into embryogenic tissues using a gene gun, offering flexibility for species like maize and wheat. (C) Protoplast transfection through PEG-mediated uptake or electroporation of plasmids or RNPs supports DNA-free editing but is constrained by regeneration efficiency. (D) Viral vectors (e.g., TRV, geminivirus) allow systemic delivery of CRISPR components, particularly sgRNAs or compact Cas variants, and nanoparticle-based delivery (e.g., carbon nanotubes, lipid or gold nanoparticles) offers minimally invasive, DNA-free options. (E) Tissue culture and regeneration considerations, including morphogenic regulators (e.g., BBM, WUS), play a critical role in achieving successful regeneration of edited plants, particularly in recalcitrant species.
Figure 4.
Delivery platforms for CRISPR-Cas genome editing in plants. A variety of methods are employed to introduce CRISPR components into plant cells, each suited to different species, tissues, and experimental objectives. (A) Agrobacterium tumefaciens-mediated delivery of T-DNA vectors carrying Cas9 and sgRNA cassettes enables stable transformation in many dicots and optimized monocots. (B) Biolistic (particle bombardment) delivery propels CRISPR plasmids or RNPs into embryogenic tissues using a gene gun, offering flexibility for species like maize and wheat. (C) Protoplast transfection through PEG-mediated uptake or electroporation of plasmids or RNPs supports DNA-free editing but is constrained by regeneration efficiency. (D) Viral vectors (e.g., TRV, geminivirus) allow systemic delivery of CRISPR components, particularly sgRNAs or compact Cas variants, and nanoparticle-based delivery (e.g., carbon nanotubes, lipid or gold nanoparticles) offers minimally invasive, DNA-free options. (E) Tissue culture and regeneration considerations, including morphogenic regulators (e.g., BBM, WUS), play a critical role in achieving successful regeneration of edited plants, particularly in recalcitrant species.
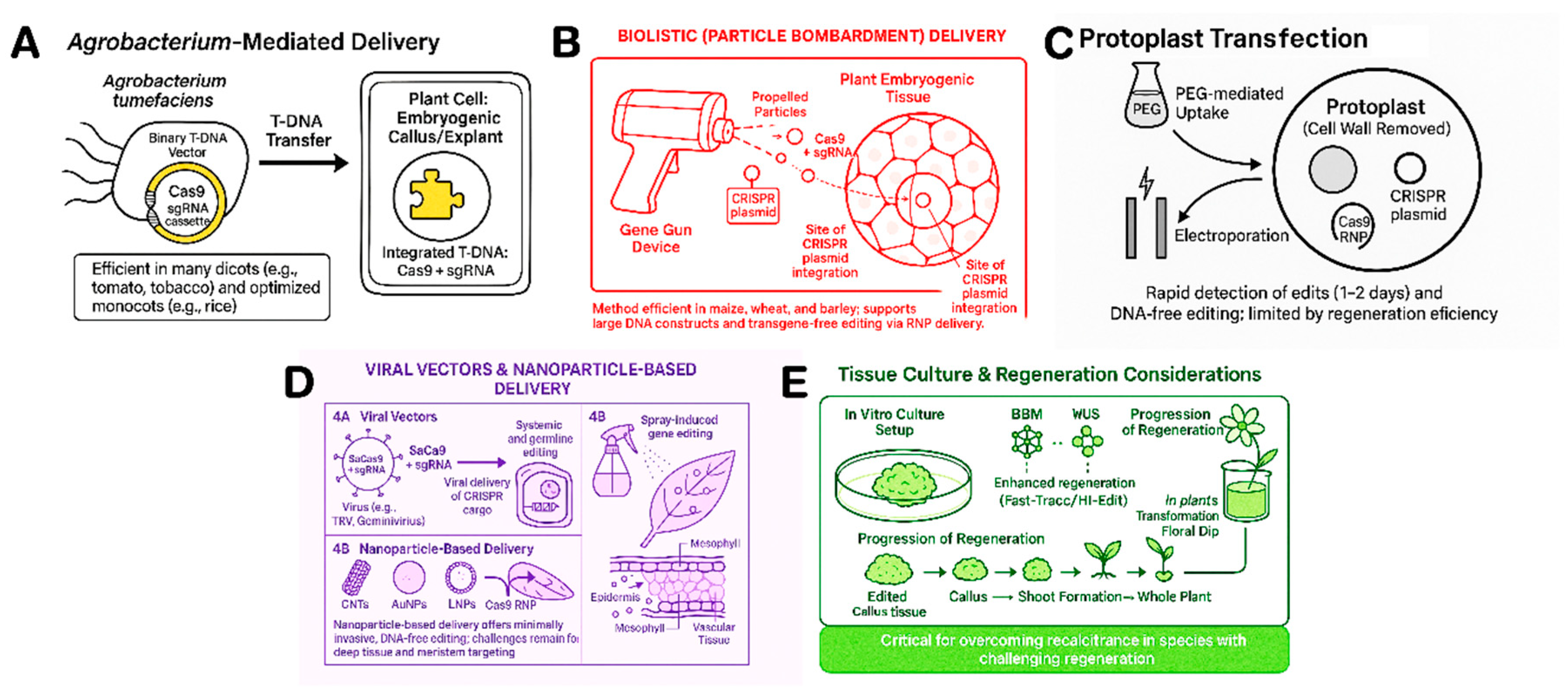

Table 1.
Representative CRISPR-Cas genome editors and their features.
| Cas Protein | Type (Class) | Size (aa) / Complex | Target & Cut | PAM / Target Motif | Notable Features |
| SpCas9 (wild-type) | II-A (Class 2) | ~1368 aa, sgRNA:crRNA+tracrRNA | DNA, blunt DSB (3 bp from PAM) | NGG (also NAG at lower efficiency) | Most widely used; numerous variants (e.g. SpCas9-NG, SpRY) expand PAM to NRN/NYN. Requires tracrRNA. |
| SaCas9 | II-A (Class 2) | ~1053 aa, sgRNA | DNA, blunt DSB | NNGRRT | ~25% smaller than SpCas9, useful for delivery; different PAM. Efficient in some plants after codon optimization. |
| Cas12a (Lb/AsCpf1) | V-A (Class 2) | ~1300 aa, crRNA (42 nt) | DNA, staggered DSB (5’ overhang) | TTTV (e.g. TTTA, TTTG) | No tracrRNA needed; processes its own array of crRNAs. Cuts distal to PAM (~, allowing large fragment deletions or repeat cutting. High AT-targeting ability and specificity. |
| Cas12b (BhCas12b) | V-B (Class 2) | ~1100 aa, crRNA | DNA, staggered DSB | TBD (e.g. TTN) | Smaller type V; engineered variants (e.g. BhCas12b v4) active at 37°C for plants. Potential for compact multiplex tools. |
| CasΦ | V? (Class 2, phage) | ~70 kDa (~600 aa), crRNA | DNA, DSB (overhang unclear) | Minimal PAM (e.g. TBN) | Hypercompact single-effector from phages; single active site for crRNA processing & cleavage. Active in plant cells; easier delivery due to small size. |
| Cas14a (Cas12f) | V-F (Class 2) | ~400–700 aa, crRNA | ssDNA, multiple cuts (collateral activity) | None (no strict PAM) | Extremely small; cleaves single-stranded DNA with high single-base mismatch sensitivity. Useful for diagnostics; potential to target ssDNA viruses. |
| CasX | V-E (Class 2) | ~980 aa, crRNA | DNA, DSB (5’ overhang) | TTCN (for DxCasX variant) | Discovered in Duggariela bacteria; independently evolved from Cas9/Cas12. Compact and non-immunogenic (from non-human microbes). Efficient editing in E. coli and human cells; being adapted for plants. |
| Cas13a (LwaCas13a) | VI (Class 2) | ~1250 aa, crRNA | ssRNA, multiple cleavage (collateral RNase) | Requires protospacer-flanking A/U/C (no DNA PAM) | RNA-specific editing (transient knockdowns). Collateral RNase used for viral RNA detection. Cas13 variants (a–d) can be repurposed for virus resistance in plants without genomic changes. |
| dCas9 or dCas12a | (engineered) | As above but catalytically dead | DNA (no cleavage) | NGG (SpCas9) or TTTV (Cas12a) | DNA-binding platform for base editors (e.g. CBE, ABE) and epigenome modifiers. dCas9 fused to deaminase yields precise single-base changes without DSB. Fusions to activators (CRISPRa) or repressors (CRISPRi) enable gene regulation without mutation. |
Table 2.
Strategies for expressing multiple guide RNAs in plants.
| Strategy | Mechanism & Components | Example Usage and Performance |
| Multiple U6 cassettes(Pol III promoters) | Each sgRNA is expressed from a separate U6/U3 small RNA promoter. Simple but increases vector size linearly with guide number. | Common in early multiplex experiments (2–4 targets). E.g., 4 U6-driven sgRNAs in tomato to mutate four loci [67]. Efficient for few targets, but >6 cassettes becomes cumbersome. |
| tRNA–sgRNA array(Pol III, PTG) | Tandem tRNA–sgRNA units transcribed as one RNA. Endogenous RNase P/Z cleave at tRNA ends, releasing sgRNAs. | Xie et al. (2015) edited 4 genes in rice with one PTG transcript [54]. High editing rates, though very large arrays see some efficiency drop-off. Supports >10 guides. |
| Ribozyme flanks(Pol II or Pol III) | Each sgRNA is flanked by self-cleaving ribozymes (HH and HDV). After transcription, ribozymes auto-cleave to yield individual sgRNAs. | Tang et al. (2016) achieved 6 multiplex edits in rice from a single Pol II transcript (UBI promoter) [58]. Very efficient (up to 100% mutant frequency per target). Modular and portable to any promoter. |
| Csy4-mediated(Pol II) | sgRNAs separated by Csy4 recognition sequences. Co-express Csy4 RNase which cleaves and releases sgRNAs. Requires adding Csy4 protein. | Cermák et al. (2017) multiplexed 8 sgRNAs in tomato; Csy4 system gave highest editing efficiency vs tRNA or ribozymes. Also used for large deletions with up to 12 guides. Extra burden of expressing Csy4, but highly effective. |
| miRNA-based(Pol II, Drosha/Dicer) | sgRNAs embedded in endogenous miRNA stem-loops. The pri-miRNA transcript is processed by Drosha/DCL1, releasing sgRNAs (as faux miRNAs). | Used in Arabidopsis: 2–3 sgRNAs designed into AtMIR390 backbone, driven by RPS5a promoter, yielded heritable edits without transgenes (in a haploid inducer system) [68]. Efficiency can be lower, but allows tissue-specific promoters for sgRNA expression. |
| Cas12a array(Pol III, self-process) | A single CRISPR array (repeats and spacers) transcribed; Cas12a cleaves the repeats to release crRNAs. No tracrRNA needed. | FnCas12a edited 4 targets in soybean from one array (driven by LjU6) with 20–50% efficiency per locus [69]. Very compact – one promoter for many guides – but limited to Cas12a or Cas12j systems. |
Table 3.
Case Studies of Multiplex CRISPR for Yield and Quality Improvement.
| Plant Species | Target Genes | Trait Improved | Specific Improvement | Key Outcomes | Reference |
| Sugarcane (Saccharum) | LG1 | Yield | Adjusted leaf angle | Mutated line showed a 56% decrease in leaf inclination angle and an 18% increase in yield. | [125] |
| Brassica napus(Rapeseed) | BnFAD2, BnFAE1 | Oil Quality | Increased oleic acid, reduced erucic acid | Significant increase in oleic acid content (70-80%) and dramatic reduction in erucic acid levels in seed oil. | [127] |
| Potato (Solanum tuberosum) | Vacuolar Invertase (VInv), Asparagine Synthetase 1 (AS1) | Processing Quality & Safety | Reduced browning and acrylamide formation | Tubers from edited events showed reduced fructose and glucose concentrations after cold storage. Crisps made from these tubers were lighter in color and contained significantly reduced levels of acrylamide. Multiplex CRISPR-Cas9 technology demonstrated its ability to generate improved potato cultivars for healthier processed products. | [128] |
| Rice (Oryza sativa) | Starch Branching Enzyme (SBE) genes (four subunits) | Nutritional Value | Increased resistant starch content | Knockout mutations were identified at all four SBE genes across multiple transgenic rice plants. This multiplex editing led to an increase in the level of resistant starch in the rice grains, enhancing their nutritional value for human consumption. | [129] |
Table 4.
Case Studies of Multiplex CRISPR for Biotic Stress Resistance.
| Plant Species | Target Genes | Pathogen/Pest | Edited Trait (Resistance/Susceptibility) | Key Outcomes | Reference |
| Brassica napus(Rapeseed) | BnWRKY11, BnWRKY70 | Sclerotinia sclerotiorum | Enhanced Resistance (BnWRKY70 mutants) | BnWRKY70 mutants showed enhanced resistance; BnWRKY11 mutants showed no significant difference. Overexpression of BnWRKY70 increased sensitivity. | [134] |
| Wheat (Triticum) | MLO gene family, dormant gene | Powdery Mildew | Resistance | Full resistance to powdery mildew achieved by simultaneously knocking out multiple MLO susceptibility genes and activating a dormant resistance gene in elite wheat varieties. | [135] |
| Tomato (Solanum lycopersicum) | Various | Various | Resistance | Potential for developing lines resistant to common bacterial, fungal, and viral diseases by inducing resistance genes or knocking out susceptibility genes. Multiplexing can further enhance this by targeting multiple factors. | [136] |
Table 5.
Case Studies of Multiplex CRISPR for Abiotic Stress Tolerance.
| Plant Species | Target Genes | Edited Trait | Key Outcomes | Reference |
| Wheat (Triticum) | TaSal1 gene family | Drought Tolerance | Improved germination and growth under simulated drought conditions. | [140] |
| Rice (Oryza sativa) | PsbS regulatory DNA | Water-Use Efficiency | Increased expression of PsbS, leading to enhanced water-use efficiency without foreign DNA integration. | [141] |
| Wheat (Triticum) | TaRPK1 | Water Absorption | Potential for enhanced water absorption through deeper root system development. | [142] |
| Various Plant Species | Sal1 | Osmo-protectant Production | Improved production of osmoprotectants like proline, enhancing drought resistance through precise modulation of gene expression. | [143] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



