
Submitted:
05 June 2025
Posted:
06 June 2025
You are already at the latest version
Abstract
Aiming to develop an efficient propagation method for the exploitation of Thymelaea hirsuta and T. tartonraira ssp. tartonraira in xeriscaping and pharmaceutical industry, the effect of: i) pretreatment (mechanical and chemical scarification or immersion in hot water, ii) incubation temperature (5-30 oC), iii) incubation light conditions (16 h photoperiod or continuous darkness) and iv) storage period at room temperature and darkness (up to 24 months), was examined on in vitro germination of their seeds. Seeds collected for two years from the same wild plants in Greece were surface sterilized with commercial bleach solution (15% for 15 min) after the abovementioned treatments and placed for germination in Petri dishes with half-strength MS medium in growth chambers. The rate and final percentage of germination were recorded. For both species, scarification after dipping in concentrated H2SO4, preferably for 20 min, was necessary for seed germination, which indicates coat dormancy. Higher germination percentages were observed at temperatures of 10-20 oC, under continuous darkness for T. hirsuta (79-100%) and regardless of photoperiod for T. tartonraira (73-90%). Storage time reduced the germination of only T. tartonraira seeds (54-68%, at optimum temperatures, 23 months after collection). For both species, an effective seed propagation protocol suitable for their exploitation as ornamental and landscape plants was developed.
Keywords:
sexual propagation
; scarification pre-treatment
; sulphuric acid
; incubation temperature
; light conditions
; storage period
; seed coat dormancy
; xerophytic ornamentals
1. Introduction
Thymelaea (F. Thymelaeaceae) is a large genus comprising around 30 species of evergreen xerophyllous shrubs [1,2], most of which are native to the Mediterranean area [2,3]. The species Thymelaea hirsuta (L.) Endl. and Thymelaea tartonraira ssp. tartonraira (L.) All. are small evergreen shrubs resistant to heat and drought, which grow well in poor stony soils. Thus, they are considered suitable for xeriscaping, in urban and peri-urban green spaces, as well as in landscape restoration and archaeological sites, in arid and semi-arid regions with Mediterranean climate [4,5,6]. Sustainable landscaping is now a requirement and can be ensured by the use of native species, as well as by the consideration of their ecological requirements and their response to abiotic and biotic stress factors [7].
Thymelaea hirsuta (Figure 1A) is a dioecious and sometimes monoecious, perennial, small evergreen shrub, up to 1.0 m, erect, spreading or prostrate, with white-downy stems and shiny green small, thick and scale-like leaves (Figure 1B) , densely overlapping along the stem. Plants have very small yellowish flowers, born in clusters (Figure 1C), from October to May. The species grows in garrigue, dry grassy and sandy places of the Mediterranean region. In the past, the tough stem fibers have been used for making ropes [8,9]. Its prolonged inflorescence period including winter, make it a valuable bee-friendly plant, since typical entomophilous adaptations, such as the large amount of pollenkitt and nektar, are present in its flowers and insects have been found to play a major role in the pollination process of this species in the Northern Mediterranean region [10]. The system of sexual tetramorphism was revealed in its natural populations, which combines characteristics of subdioecy (subandroecious and subgynoecious individuals) and heterodichogamy (protogynous and protandrous individuals), supporting the hypothesis that it is an evolutionary pathway from heterodichogamy to dioecy [11]. The increased genetic diversity of T. hirsuta populations leads to increased plant adaptability and resilience to different environmental conditions, while its richness in total lipids and phenols enables the plant to adapt under harsh environmental conditions [12].
Moreover, T. hirsuta is an important medicinal plant, that has been used as traditional medicine in several countries of northern Africa [2,13,14], as a decoction in the treatment of diabetes [14,15,16,17], to treat infertility [16] and skin infections [18], as well as for its antimelanogenesis [19], hepatoprotective [14] and antioxidant [20] properties. It was also recommended as a remedy for asthma, coughing, hemorrhoids, stomach, intestine and sexual diseases [21,22]. Extracts of this plant are natural sources of natural bioactive compounds with high anti-oxidant activity [13,18,23], which could be an alternative to synthetic antioxidants, showing potential anticancer activities as well [13,14]. Its high healing potential, antioxidant and antimicrobial activity [2,16,23] can be devoted to the presence of valuable secondary metabolites in its leaves, such as flavonoids, tannins, phenols, diterpenes, triterpenes, sterols, saponins, alkaloids, cardiac glycosides, carbohydrates, volatile oils, fixed oils and fats [16]. Apart from its use to remedy different ailments traditionally, it is a promising plant for utilization by pharmaceutical industries to prepare medicines for various diseases [16]. A new polysaccharide, named THP, was also extracted from this plant, which is promising as a natural additive in various food and pharmaceutical emulsions, while it demonstrates significant antioxidant properties, surpassing those of synthetic antioxidants [24]. Moreover, it could be a potential source for biodiesel production, due to its fatty acid methyl esters profile and oil content [25].
Thymelaea tartonraira ssp. tartonraira (L.) All. (Figure 1D) is a much-branched, silvery-grayish, native dwarf dioecious shrub, up to 0.50 m, with numerous silkily hairy leaves (Figure 1E) and yellowish flowers, borne in small clusters of 2-5 at the base of the upper leaves (Figure 1F), from February to April, which grows in rocky and sandy habitats of the Mediterranean region, often close to the coast [8,9]. It possesses medicinal properties too, as a decoction of its leaves has been used in the treatment of diabetes [26]. Valuable secondary metabolites, especially flavonoids, were found in its leaves that could serve as antioxidant and specific antihyperglycemic agents [27], while its extracts constitute a valuable source of new natural products to combat leishmaniasis [28]. Moreover, its nitrogen and phosphorous content has been studied, along with various Mediterranean plant species [29], as well as the annual fluctuations of soluble sugars, lipids, starch and nitrogen content have been determined in its tissues [30]. It has also been used in a phylogenetic study aiming to reveal evolutionary history of Thymelaea [1].
The development of an efficient propagation method would facilitate their sustainable exploitation, since their natural populations are gradually declining [31,32]. However, rooting of stem cuttings, which is a popular propagation method in horticultural practice, proved unsuccessful for these two species [32,33]. Regarding micropropagation attempts, although satisfactory in vitro culture establishment and shoot multiplication has been succeeded for both species using node explants excised from young seedlings grown in vitro [4,5,6,34], microshoot rooting was ineffective [32], due to failure of microshoots to root [4,6,34].
Since vegetative reproduction proved difficult to implement, as described above, sexual breeding seems to be the only way to propagate these species and regenerate their natural population, with potential use in conservation strategies [32]. However, their sexual propagation presents difficulties, because of: i) seasonal variation in fruit and seed production [32], ii) plants’ dimorphism that greatly differentiates fruit productivity of each plant type, i.e., high fruit production in females and protandrous plants vs. low fruit production in males and protogynous plants [11] and iii) extremely low seed germination ability [32,33]. Immersion of the seeds in concentrated H2SO4 for 10-20 min, combined with GA3 pretreatment, increased their germination up to 45% [35], which indicates coat seed dormancy. Furthermore, research is required in order to increase its effectiveness, through testing techniques to break seed dormancy and increase efficiency of seed germination. Propagation by seed is an easy method for commercial nurseries, especially when it concerns native species, as it is also suitable for the production of plants intended for landscape regeneration or restoration, due to genetic variability [36]. Nevertheless, seed of native species can vary widely in seed quality factors, such as seed size, purity, dormancy and germination vigor [37]. Propagation by seed can also be used in breeding programs to select genotypes with desirable traits, which is of high importance for the studied Thymelaea species, since they are both promising medicinal plants.
Dormancy and germination are two closely linked physiological traits of seeds, which are influenced by a variety of internal and external environment factors, having a great impact on adaptation and survival of seed plants, and being also important for agricultural production [38,39,40]. Diverse endogenous hormones, such as abscisic acid (ABA) and gibberellins (GAs) are known to act antagonistically regulating seed dormancy, while recent findings demonstrate that also auxin is critical for inducing and maintaining seed dormancy [38]. The determinant role of ABA in seed dormancy and the requirement for GAs for germination has been identified by genetic analysis [41]. Among the environmental factors, light and temperature are the most critical, as they regulate the ABA/GA biogenesis and signaling pathways, as well as affect pathways of auxin and other hormones, such as ethylene, and jasmonic and salicylic acid, during seed germination, acting as a signal input to determine whether seeds stay in a dormant state or start to germinate [38,42,43,44]. Other factors, such as nitrogen availability, water content and bacteria in soil also affect seed dormancy and germination [40,42].
Seed dormancy and germination depend on seed structures surrounding the embryo and/or on factors affecting the growth potential of the embryo, which may be compounds imported from the mother plant or produced by the embryo itself [41]. Therefore, seed dormancy can be caused by hard seed coats, underdeveloped or primitive embryos and inhibitory compounds. In natural conditions, dormancy in seeds is broken by soil melting and freezing, microbial activity, forest fires, soil activity, and animal consumption seeds [45]. Seeds can also be released from their dormancy by scarification, which can be carried out mechanically, either by using sandpaper or a sharp instrument to chip or pierce the seed coat, or chemically by immersion in sulphuric acid, followed by others like stratification, light and heat treatment, dipping in hot water, leaching etc, and the most appropriate method varies according to the plant species [45,46].
Temperature and light conditions are the most important environmental factors in seed germination, affecting both the germination rate and speed [47]. Temperature can also control timing of germination through the temperature range over which germination may occur. In natural seed populations, environmental temperature regulates their dormancy status, allowing the seeds to avoid harsh environmental conditions for seedling establishment [48]. Storage period and conditions during storage also affect the germination process [49], as during the post-harvest phase of after-ripening important physiological and biochemical changes are caused in seeds that modify the seeds internal environment, ultimately breaking dormancy and enabling germination [50]. Unlike other dormancy breaking methods, which requires external factors, after-ripening primarily occurs in dried seeds under dry storage conditions and determines germination potential [50].
In the present work, the effect of breaking dormancy pretreatment, incubation conditions (temperature and photoperiod) and storage period on in vitro germination of T. hirsuta and T. tartonraira ssp. tartonraira seeds was examined, aiming to the development of an efficient propagation method that would facilitate their wider use in commercial floriculture and landscaping, as well as in pharmaceutical industry. Seeds were collected for two years from the same wild plants in Greece and several experiments were conducted, in which various factors were tested, such as: i) pretreatment (mechanical scarification with sandpaper for 2 or 4 min, immersion in concentrated sulphuric acid (H2SO4) for 15 or 20 min, immersion in hot water (100 oC) for 1 or 5 min), ii) incubation temperature (5, 10, 15, 20, 25 or 30 oC), iii) incubation light conditions (16 h photoperiod, continuous darkness) and iv) storage period of seeds at room temperature and darkness (5, 9, 12, 18 or 24 months).
2. Materials and Methods
2.1. Seed Collection and Its Viability
Seeds of T. hirsuta were collected in late May 2013 and 2015 in the Ilioupoli area (37ο95’03.07’’ Β, 23ο75’39.26’’ Ε, altitude 170 m), a suburban municipality southeast of central Athens, at the foot of Mount Hymettus, Greece, while seeds of T. tartonraira ssp. tartonraira were collected in late May 2014 and 2015 a little higher up in the foothills of Mount Hymettus (37ο92’68.44’’ Β, 23ο76’68.52’’ Ε, altitude 396 m). Since both species are dioecious, only female plants with high seed production were carefully chosen and both seed collections for each species were done from the same wild plants. The seed material was left to dry in an open-air shaded place for 15 d and then stored in glass vessels at room temperature (about T = 21 oC and darkness). Before each germination experiment, the dried floral debris were removed from seeds rubbing them together by hand (Figure 2A). The weight of 100 seeds was estimated after removing the floral debris.
Seed viability of T. hirsuta was determined two and twenty-four months after harvesting using seeds collected in May 2013. Seed viability of T. tartonraira was determined two months after harvesting using seeds collected in May 2014. Seeds were submitted to 2,3,5-triphenyl-tetrazolium chloride (TZ) staining (1.0%), at 20 oC, in the dark, for 24 h. For each test, a total of 100 seeds and four vessels containing 25 mL of TZ-solution each were used (25 seeds/Magenta™ glass vessel). The seeds remained in the solution for 24 h, at 20 oC, in the dark. Then, the coloration of the embryo was observed using a portable QS.20200-P digital microscope (Euromex Microscopen, Arnhem, The Netherlands). The embryo of viable seeds was colored red. Embryos were considered non-viable when they had less than 1/2 cotyledon colored red or non-colored hypocotyl [51].
2.2. Seed Surface Sterilization Method
In all germination experiments of both species, a common surface sterilization method of seeds was applied, which had been successfully used previously in other Mediterranean species [52,53]. This stage took place after the application of various pretreatments for breaking seed dormancy and before seeds’ placement in Petri dishes for germination.
Surface sterilization of seeds was done by immersion in a 15% v/v water solution of commercial bleach (4.5% w/v sodium hypochloride) for 15 min, in which 1–2 drops of Tween 20 (polyxyethylenesorbitan monolaurate, Merck KGaA, Darmstadt, Germany) were added. Three 3-min rinses with sterile distilled water were following.
2.3. Preliminary In Vitro Germination Experiments of T. hirsuta
Aiming to investigate the effect of various treatments for breaking seed dormancy and of incubation conditions (temperature and photoperiod) on in vitro germination of T. hirsuta seeds, the following preliminary experiments were conducted with seeds collected in 2013:
A. In October 2013 (5 months after collection), surface sterilized seeds were placed for in vitro germination in Petri dishes (ten seeds per dish) on half-strength (1/2) MS medium [54] (Sigma-Aldrich, St. Louis, MO, USA), with sucrose (20 g L-1) and agar (8 g L-1), at temperatures of 5, 10, 15, 20, 25 oC, and in a photoperiod of 16 h cool white fluorescent light (37.5 mmol·m–2·s–1) and 8 h darkness. Seeds had received: i) either no pretreatment (control) or ii) they had been scarified with sandpaper (suitable for wooden surfaces) for 2 or 4 min or iii) they had been immersed in concentrated H2SO4 (>95%, Fisher Scientific, Loughborough, UK) for 15 min. Sandpaper, which is suitable for iron surfaces, was also tested as a scarification material for the same period of time.
B. In March 2014 (9 months after collection), seeds were subjected to various pretreatments: i) control, ii) scarification using sandpaper (suitable for wooden surfaces) for 2 or 4 min, iii) immersion in H2SO4 for 15 or 20 min, and iv) immersion in boiling water (100 oC) for 1 or 5 min, before surface sterilization and incubation for in vitro germination in Petri dishes with ½MS medium, at 10 oC, and 16 h photoperiod.
C. In early June 2014 (12 months after collection), seeds were either untreated (control) or immersed in H2SO4 for 15 min, subjected to the same surface sterilization and placed in Petri dishes with ½MS medium at temperature 5, 10, 15, 20, 25, 30 oC, under 16 h light or continuous darkness. Seeds immersed in H2SO4 for 20 min were also incubated at the above temperatures, only under 16 h light.
D. In November 2014 and May 2015 (18 and 24 months, respectively, after collection), seeds were only immersed in H2SO4 for 20 min and placed in Petri dishes with ½ MS medium at temperature 5, 10, 15, 20, 25, 30 oC, under 16 h of light or continuous darkness. Ιn the experiments conducted 12 and 18 months after collection, it was observed that mainly seeds whose white endosperm had been partially or fully exposed after scarification germinated. Thus, in May 2015, in seeds (30% of the total seeds) where this had not happened, the coat, which had become thin from the H2SO4 treatment and was fragile, was carefully broken by pressing the seeds one by one with a scalpel in the container used for disinfection.
Five replicates of 20 seeds were used for each treatment. Germination observations were made every 3 days for a period of one month. The radicle should be longer than 1 mm in order a seed to be considered germinated. In the experiments that took place 18 and 24 months after seed collection, apart from seed germination percentage (%), they were also estimated: i) T50, which is the time needed for 50% of the final germination value, and it is calculated from the two germination values closest to median germination, ii) time for full germination and iii) percentage (%) of germinated seeds with well developed root.
2.4. Preliminary In Vitro Germination Experiments of T. tartonraira ssp. Tartonraira
Preliminary experiments of T. tartonraira were conducted using seeds collected in May 2014. The experiments were limited by a lack of seed material. For this reason, the effect of darkness on germination could not be studied.
A. In June 2014 (2 weeks after collection), after surface sterilization, seeds were placed in Petri dishes with 1/2 MS medium at 10 oC, for the first two weeks, and at 15 oC, thereafter, after firstly having been immersed in concentrated H2SO4 for 20 min.
B. In October 2014 and May 2015 (4 and 12 months, respectively, after collection), seeds were immersed in concentrated H2SO4 for 20 min, surface sterilized and placed in dishes with 1/2 MS medium at temperature 5, 10, 15, 20, 25, 30 oC, under 16 h photoperiod. In May 2015, those seeds whose coat had not broken were carefully broken partially to reveal the white endosperm by pressing the seeds one by one with a scalpel in the container used for disinfection.
Observations were taken with the same frequency as in case of T. hirsuta and the same data were estimated.
2.5. Main Experiments on In Vitro Germination of Both Thymelaea Species
Due to the sufficient amount of seed collected for both Thymelaea species in May 2015, a main experiment with the same treatments for both species could be carried out simultaneously, based on the results of the preliminary experiments. So, the effect of three factors, i.e., chemical scarification, incubation temperature and light conditions, could be examined simultaneously in a three-factor experiment, that was repeated twice for each species (9 and 23 months after seed collection).
In February 2016 and April 2017 (9 and 23 months, respectively, after collection), seeds of T. hirsuta and T. tartonraira either received no pretreatment (control) or they were immersed in concentrated H2SO4 for 20 min. Then, they were surface sterilized and placed in Petri dishes with ½ MS medium at temperatures 10, 15, 20 and 25 oC, under 16 h photoperiod or continuous darkness. The temperatures 5 and 30 oC were excluded in main experiments, based on preliminary results. Regarding chemically scarified seeds, firstly seeds whose white endosperm had been partially or completely exposed, were used. In the remaining seeds, the coat was carefully broken by pressing the seeds one by one with a scalpel in the container used for surface sterilization, before their use. In this way, the empty seeds were discarded as well. Based on the results of tetrazolium test for seed viability, in order to have 100 viable seeds for each treatment, about 150 seeds were used to be scarified. Five replicates of 20 seeds were used for each treatment. Germination observations were taken every 3 days for a period of one month. The radicle should be longer than 1 mm in order a seed to be considered germinated. In the main experiments, they were estimated: i) germination percentage (%), ii) germination percentage (%) with well developed seedling (with normal leaf and root development) and iii) T50.
2.6. In Vitro Culture Conditions
In all experiments, seeds were placed on the same substrate for germination, which was 1/2 MS, with the addition of 20 g L-1 sucrose, solidified with 8 g·L–1 agar. Its pH was adjusted to 5.7 before addition of the agar and autoclaving at 121 oC for 20 min. Incubation of seeds took place in Petri dishes with 10 mL substrate, where ten seeds were put. The dishes were sealed on the side with parafilm (roll 5 cm x 15 m, Brand, Germany). The dishes with seeds that received the treatment of 16 h photoperiod were maintained inside incubation chambers, at 25 oC with a 16 h photoperiod/ 8 h darkness, at 37.5 mmol·m–2·s–1 provided by cool-white fluorescent lamps. The dishes with seeds that received the treatment of continuous darkness were maintained in the same incubation chambers, but were wrapped in aluminum foil to create dark conditions.
2.7. Statistical Analysis
In all experiments a completely randomized design was used. The significance of the results was tested by one- or two- or three-way analysis of variance (ANOVA) and the means of the treatments were compared by the Student’s t test at p ≤ 0.05 (JMP 13.0 software, SAS Institute Inc., Cary, NC, USA, 2013). The data on percentage were statistically analyzed after arcsine transformation.
3. Results
3.1. Preliminary In Vitro Germination Experiments
3.1.1. T. hirsuta
The weight of 100 seeds of T. hirsuta was 0.275 g and the tetrazolium test showed that in fresh (2 months after collection) and stored (24 months after collection) seed the percentage of seed viability did not differ, since 74.0% and 72.0% of the seeds, respectively, were viable.
Five months after collection, T. hirsuta seeds that were incubated under 16 h photoperiod and at temperatures of 5 to 25 oC, without any scarification pretreatment, or that were mechanically scarified with sandpaper for 1 min, germinated at percentages lower than 5% and only seeds that had been immersed in concentrated H2SO4 for 15 min germinated at much higher percentages reaching 40% at 10 oC (Figure 3A). Sandpaper, either for wooden or iron surfaces, was not suitable as a scarification material, because it destroyed the seeds by crushing them. At 5 oC, no seeds germinated (Figure 3A).
Nine months after collection, seeds were incubated only at 10 oC, under 16 h photoperiod. Only the seed pretreatment with H2SO4 for 15 or 20 min dramatically increased the percentage of seed germination to 64-74%, in contrast to scarification with sandpaper for 2 or 4 min, immersion in boiling water for 1 or 5 min or the absence of any pretreatment that caused germination at less than 4% of the seeds (Figure 3B).
Twelve months after collection, seeds that had been immersed in H2SO4 for 15 min and then incubated under 16 h photoperiod showed a higher germination percentage at 10 oC. Under continuous darkness, correspondingly high germination percentages were observed in a wider range of temperatures, from 10 to 20 oC. At 5 oC, no seeds germinated, while at 25 and 30 oC seedlings were dehydrated, weak and with reduced growth. Seeds that had not received any pretreatment showed zero germination percentage (Figure 3C). Seeds that had been immersed in H2SO4 for 20 min and then incubated under 16 h light germinated relatively faster than those immersed in H2SO4 for 15 min, at the corresponding temperature (Figure 3D).
Eighteen months after collection, seeds, that had been immersed in H2SO4 for 20 min, germinated at the highest rates at 15 and 20 oC under 16 h photoperiod (Figure 2B-F), and from 10 to 25 oC under continuous darkness (Figure 2G, Figure 4A). At 5 oC, no seeds germinated, while at 30 oC, the seedlings were chlorotic and weak (Figure 2H). The increase of temperature decreased time for T50 and time for full germination, while incubation under continuous darkness also decreased time for full germination compared to 16 h photoperiod (Table of Figure 4A).
Twenty-four months after collection, the highest seed germination rates (90%) of all experiments were observed, after immersion in concentrated H2SO4 for 20 min and incubation at 10 to 20 oC, under continuous darkness (Figure 4B). In this experiment, only seeds whose endosperm had been exposed were used, either after immersion in concentrated H2SO4, or after additional breaking of the coat with a scalpel. In this temperature range, the superiority of incubation under darkness over germination under 16 h of light on seed germination percentage was obvious (Figure 4B). Besides, only seedlings that germinated at temperatures from 10 to 20 oC had normally developed radicles in very high percentages. On the contrary, at 25 and 30 oC, in addition to the fact that the seedlings were dehydrated, the normal development of the radicle was hindered (Figure 2H) and this was more pronounced under conditions of 16 h light compared to continuous darkness (Table of Figure 4B).
3.1.2. T. tartonraira ssp. tartonraira
The weight of 100 seeds of T. tartonraira was 0.265 g and the tetrazolium test showed that 68.0% of the seeds were viable two months after collection.
Two weeks after collection, T. tartonraira seeds, which had been treated with concentrated H2S04 for 20 min, did not germinate during the first two weeks at 10 oC. So, afterwards, they were transferred to 15 oC, where they germinated at a high percentage (78%).
Four months after collection, seeds, which had been immersed in H2SO4 for 20 min and incubated under 16 h photoperiod, germinated at higher rates at temperature from 10 to 20 oC. At 5 oC, no seeds germinated, while at 25 and 30 oC, lower germination percentages were observed (Figure 5Aa). Only seeds whose endosperm had been exposed were used.
Twelve months after collection, seeds, which had been immersed in H2SO4 for 20 min and incubated under 16 h photoperiod, germinated at higher percentages at 15 to 25 oC, followed by 10 oC (Figure 5B). Only seeds whose endosperm had been exposed were used. At 5 oC, no seeds germinated, while at 30 oC lower germination rates were observed and the seeds germinated but did not grow further (Figure 5B).
The increase of temperature decreased time for T50 and time for full germination in this species as well. Higher percentage of seedlings with well developed root were observed at 15 and 20 oC, followed by 10 oC (Table of Figure 5).
3.2. Main In Vitro Germination Experiments
3.2.1. T. hirsuta
Regarding the effect of the main experimental factors, i.e., i) pretreatment with chemical scarification, ii) incubation temperature and iii) photoperiod, on in vitro germination of T. hirsuta seeds, three-way ANOVA showed significant interaction, irrespectively of storage period. So, germination data were analyzed by one-way ANOVA (Table 1).
At both repetitions of the experiment, 9 and 23 months of storage led to the following similar results:
Pretreatment of seeds with concentrated H2SO4 for 20 min was the most important factor for seed germination, since it dramatically increased germination percentages of scarified seeds (up to 100% in some treatments) compared to non-scarified ones, which did not germinate (Table 1). It should be emphasized that only seeds, whose white endosperm was revealed, were used and the non-viable seeds were discarded.
Germination of scarified seeds was also strongly affected by photoperiod. Seeds that were incubated under continuous darkness germinated at higher percentages (up to 100%) than those incubated under 16 h light and 8 h dark (less than 55%) (Table 1).
The effect of temperature differed depending on the photoperiod during incubation. When incubation took place under 16 h photoperiod, higher germination percentages (53-76%) were observed at 10 oC than at 15-25 oC. When incubation took place under continuous darkness, higher germination percentages were recorded at 10-20 oC (71-100%) compared to 25 oC (50-84%) (Table 1). At the optimal temperature range for each light condition, almost all seedlings were well developed. Increasing temperature from 10 oC to 20 oC accelerated seed germination reducing, almost to half, time for T50 (Table 1).
Storage period (up to 23 months) did not have any negative effect on germination capacity. On the contrary, even higher germination percentages were recorded in the optimum treatments (Table 1).
3.2.2. T. tartonraira ssp. tartonraira
In T. tartonraira, nine months after seed collection and storage, three-way ANOVA did not show any interaction of the main experimental factors, i.e., scarification, incubation temperature and photoperiod, regarding germination percentage and germination percentage with well developed seedlings. Only pretreatment with chemical scarification was found to have a significant effect, while incubation conditions had not (Table 2).
Twenty-three months after seed collection and storage, three-way ANOVA, regarding seed germination percentage, showed significant interaction among the three factors. As regards germination percentage with well developed seedlings, there was only interaction between scarification and photoperiod, while temperature had an insignificant effect (Table 2).
In this species as well, scarification with concentrated H2SO4 was the most important factor for germination, since higher percentages were recorded by scarified seeds (44-90%) compared to non-scarified ones (0-2%) (Table 2).
Nine months after seed collection and storage, scarified seeds generally germinated at satisfactory percentages (60-90%), irrespectively of incubation conditions (Table 2), which were equally high to those recorded for T. hirsuta seeds stored for the same period (Table 1). The highest germination percentages were recorded by scarified seeds incubated under 16 h photoperiod at 15-20 oC, as well as those that were incubated under continuous darkness at 10 oC and 20 oC. However, regarding germination percentage with well developed seedlings, this was highest for scarified seeds that were incubated under 16 h photoperiod and 15 oC or under continuous darkness and 10 oC (Table 2).
Twenty-three months after seed collection and storage, in scarified seeds, lower germination percentages (44-68%) were recorded compared to seed stored for nine months (Table 2), which were lower than those recorded for T. hirsuta seeds stored for 23 months (Table 1). The highest germination percentages were recorded by scarified seeds incubated under 16 h photoperiod at 15 oC and 25 oC, as well as those incubated under continuous darkness at 15-25 oC (Table 2). However, regarding germination percentage with well developed seedlings, this was highest for scarified seeds incubated only under continuous darkness at 15-20 oC (Table 2).
In this species as well, increasing temperature from 10 oC to 25 oC reduced time for T50. Besides, seeds stored for 23 months presented less time for T50 than seeds stored for 9 months, mainly at the lower temperatures (Table 2).
Storage period seemed to have a negative effect on germination capacity of T. tartonraira, since lower germination percentages were recorded in seeds stored for 23 months, while germination percentage with well developed seedling were even lower compared to seeds stored for 9 months (Table 2).
4. Discussion
The low germination percentages that have been reported for T. hirsuta [32,33], along with the ineffectiveness of other vegetative propagation techniques [4,6,32,33,34] and the necessity for the development of an efficient method for the sustainable exploitation of T. hirsuta and T. tartonraira led to the investigation of in vitro germination of their seeds in the present study.
Based on all the conducted experiments, it can be said that pretreatment with chemical scarification was the most determinant factor for in vitro germination of both Thymelaea species, while the effect of incubation temperature, photoperiod and storage period depended on the species. Scarification is a main technique to release seeds from their dormancy [45,46], in cases that seed dormancy is caused by hard seed coats [45].
Regarding the effect of various pretreatments on germination of both Thymelaea species, only scarification with H2SO4 for 15 or 20 min significantly increased germination percentages, in verification of previous studies on T. hirsuta [35] and other Mediterranean shrubs, such as Teucrium sp. [52,55], Anthyllis sp. [53,56,57], Salvia sp. [58] and Astragalus sp. [59]. Immersion for 20 min was preferred due to the slightly greater germination percentages and the relatively faster germination. In the experiments where only seeds whose white endosperm had been revealed, either after immersion in concentrated H2SO4 or after breaking the coat of chemically scarified seeds with a scalpel, even higher seed germination percentages were recorded. In this way, all seeds with atrophic endosperm were discarded as well. Although this scarification method is not applicable on a commercial scale, it showed how important removing the seed coat was for germination. On the other hand, non-scarified seeds showed zero germination percentages in most treatments. These observations constitute strong indication of a coat-imposed dormancy, which may be caused by the impermeable coat or the mechanical prevention of radicle extension or inhibitory substances that prevent embryo from germinating [41,60]. Germination ability of seeds of other Thymeleaceae species, such as Thymelaea velutina [61], Daphne sp. [62] and Pimelea arenaria [63] was also considerably low, because of physiological dormancy. Seeds longevity and ecological adaptation is benefited by impermeable coat, since it ensures recolonization of burnt area after fire and resistance of seeds to ingestion by animals and birds [64]. Seeds of other species of Thymeleacea family, such as Pimelea trichostachya, P. simplex and P. elongata, which are endemic to arid regions of Australia, under similar conditions to Mediterranean region, have also been reported to be strongly dormant for years [65], whereas the species Aquilaria malaccensis [66] and Gyrinops walla [67], which grow in tropical regions of Southeastern Asia and Sri Lanka, respectively, possess recalcitrant seeds that have short viability period, germinating efficiently directly after collection.
Moreover, mechanical scarification with sandpaper was not only ineffective in breaking seed dormancy, which has also been reported for Teucrium capitatum [52], but also crushed the seeds destroying the embryo. On the other hand, in Anthyllis hermanniae [53], as well as in Salvia fruticosa and S. officinalis [58], both chemical and mechanical scarification were effective in increasing seed germination, through the facilitation of water entry and gas exchange. Neither pretreatment by immersion in hot water had any effect on the improvement of seed germination of Thymelaea sp., in accordance to what has been reported for Teucrium sp., in which no seed germinated after immersion in boiling water [52,55].
A wide range of temperatures, from 5 to 30 oC, was tested in seed germination, revealing that temperatures from 10 to 20 oC were the most appropriate for T. hirsuta and those from 10 to 25 oC for T. tartonraira. At 5 oC no seed germinated, similarly to T. capitatum [52] and A. hermanniae [53], whereas at 30 oC germination percentages were significantly lower and seedlings were dehydrated and abnormal, since radicle development was hindered, similarly to T. capitatum [52] as well. The increase of temperature from 10 oC to 20 oC accelerated seed germination, reducing time for T50, as temperature is the most important environmental factors in seed germination, affecting germination speed as well, apart from germination percentage [47]. For other Mediterranean species, the optimum temperature to reach maximum germination has also been found to be in the range of 10-25 oC [52,53,68,69,70,71], although the influences of the temperature incubation in germination behavior depended strongly on species [68].
As regards the effect of photoperiod conditions, in T. hirsuta, seed germination was greatly favored by continuous darkness, since not only higher germination percentages were recorded but also seeds germinated efficiently at a wider range of temperature (10-20 oC) compared to 10 oC, that was the optimum temperature under 16 h photoperiod. In T. tartonraira, no photoperiodical requirement appeared. Similarly to T. hirsuta, germination of T. velutina was highest when seeds were left in the dark [61], whereas T. capitatum [52], Anthyllis sp. [53,57] and Coridothymus capitatus [69] responded as T. tartonraira, since photoperiod had no significant effect on their germination. An intermediate response towards photoperiod was found for some Lamiaceae of Crete [69,70.], as seeds partially germinated in darkness, but their germination was significantly enhanced by light. An absolute requirement of light for germination was found in Origanum vulgare subsp. hirtum [69].
Seed germination of the studied species also differed in response to storage period. Increasing storage period (up to two years) reduced the germination capacity only of T. tartonraira seeds in opposition to T. hirsuta seeds, which germinated at even higher percentages at optimum temperatures 23 months after collection and storage. Preservation of germination ability or even higher germination percentages in older seeds compared to younger ones have been found for T. velutina [61], T. capitatum [52] and other Lamiaceae, possibly as a result of the volatization of essential oils present on their coat [69]. Faster germination of older seeds (up two years) than of younger ones (two to six months) have also been reported [52,61]. On the other hand, similarly to T. tartonraira, scarified seeds of A. hermanniae, stored for 18 months showed slightly reduced germination percentage, in opposition to unscarified ones, which showed stable, low germinability throughout the storage period [53]. In A. barba-jovis as well, storage for more than 24 months reduced germination, mostly of unscarified seeds [57].
5. Conclusions
In both T. hirsuta and T. tartonraira ssp. tartonraira, chemical scarification of the seeds by immersion in concentrated H2SO4 for 20 min was necessary for in vitro germination, since it resulted in a significant increase in germination percentages (up to 73-100% at optimum treatments) compared to almost zero germination in non-scarified seeds, which indicates dormancy caused by impermeable coat.
In T. hirsuta, the highest germination percentages (79-100%) were observed after H2SO4 pretreatment and incubation at 10-20 oC, under continuous darkness, regardless of seed storage time.
In T. tartonraira ssp. tartonraira, no large variations were observed in germination percentages of scarified seeds (60-90%) regarding tested temperature range, i.e., 10–25 oC, and photoperiod conditions, while increasing the seed storage time, up to almost two years, reduced their germination capacity (44-68%).
Higher germination percentages (73-100%) were observed at temperatures of 10-20 oC, under continuous darkness for T. hirsuta and regardless of photoperiod conditions for T. tartonraira.
Storage time (up to two years) reduced the germination of only T. tartonraira seeds, whereas in T. hirsuta older seeds germinated at even higher percentages.
To sum up, chemical scarification was the most determinant factor for in vitro germination of T. hirsuta and T. tartonraira ssp. tartonraira, due to the successful breaking of seed coat dormancy, while the effect of incubation temperature, photoperiod and storage period on germination depended on the species. For both species, an effective seed propagation protocol suitable for their exploitation as ornamental and landscape plants was developed.
Author Contributions
Conceptualization, M.P.; methodology, A.N.M. and M.P.; validation, A.N.M. and M.P.; formal analysis, A.N.M.; investigation, A.N.M.; resources, M.P.; data curation, A.N.M.; writing—original draft preparation, A.N.M. and M.P.; writing—review and editing, A.N.M. and M.P.; visualization, A.N.M.; supervision, M.P.; project administration, M.P.; funding acquisition, M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NSRF 2007-2013, Operational Program “Education & Lifelong Learning” – THALES – ARCHAEOSCAPE, MIS code 380 237.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Galicia-Herbada, D. Origin and diversification of Thymelaea (Thymelaeaceae): inferences from phylogenetic study based on ITS (rDNA) sequences. Plant Syst. Evol. 2006, 257(3-4), 159–187. [Google Scholar] [CrossRef]
- Marmouzi, I.; Bouchmaa, N.; Kharbach, M.; Ezzat, S.M.; Merghany, R.M.; Berkiks, I.; El Jemli, M. Thymelaea genus: Ethnopharmacology, chemodiversity, and bioactivities. South African Journal of Botany 2021, 142, 175–192. [Google Scholar] [CrossRef]
- Boudjelal, A.; Henchiri, C.; Sari, M.; Sarri, D.; Hendel, N.; Benkhaled, A.; Ruberto, G. Herbalists and wild medicinal plants in M’Sila (North Algeria): An ethnopharmacology survey. J. Ethnopharmacol. 2013, 148(2), 395–402. [Google Scholar] [CrossRef] [PubMed]
- Papafotiou, M.; Martini, A.N.; Vlachou, G. In vitro propagation as a tool to enhance the use of native ornamentals in archaeological sites of Greece. Acta Hortic. 2017, 1155, 301–308. [Google Scholar] [CrossRef]
- Martini, A.N.; Papafotiou, M. Effect of cytokinins on in vitro blastogenesis of Thymelaea tartonraira ssp. tartonraira (L.) All. Acta Hortic. 2019, 1242, 511–514. [Google Scholar] [CrossRef]
- Martini, A.N.; Papafotiou, M. Investigation of micropropagation of the Mediterranean xerophyte Thymelaea hirsuta (L.) Endl. (Thymelaeaceae). Acta Hortic. 2020, 1298, 335–340. [Google Scholar] [CrossRef]
- Mircea, D.-M.; Boscaiu, M.; Sestras, R.E.; Sestras, A.F.; Vicente, O. Abiotic Stress Tolerance and Invasive Potential of Ornamental Plants in the Mediterranean Area: Implications for Sustainable Landscaping. Agronomy 2025, 15, 52. [Google Scholar] [CrossRef]
- Huxley, A.; Taylor, W. Flowers of Greece and the Aegean; Publisher: Chatto and Windus: London, UK, 1977; pp. 104–105. [Google Scholar]
- Blamey, M.; Grey-Wilson, C. Mediterranean wild flowers; Publisher: Harper Collins Publishers: London, UK, 1993; p. 138. [Google Scholar]
- Cornara, L.; Borghesi, B.; Caporali, E.; Casazza, G.; Roccotiello, E.; Troiano, G.; Minuto, L. Floral features and reproductive ecology in Thymelaea hirsuta (L.) Endl. Plant Syst. Evol. 2005, 250, 157–172. [Google Scholar] [CrossRef]
- Dommée, B.; Bompar, J.L.; Denelle, N. Sexual tetramorphism in Thymelaea hirsuta (Thymelaeaceae): evidence of the pathway from heterodichogamy to dioecy at the infraspecific level. American Journal of Botany 1990, 77(11), 1449–1462. [Google Scholar] [CrossRef]
- Abd El-Maboud, M.M..; El-Zayat, M.A. Genetic and biochemical studies of Thymelaea hirsuta L. growing naturally at the North western coast of Egypt. Int. J. Plant & Soil Sci. 2020, 32, 28–37. [Google Scholar] [CrossRef]
- Yahyaoui, M.; Bouajila, J.; Cazaux, S.; Abderrabba, M. The impact of regional locality on chemical composition, anti-oxidant and biological activities of Thymelaea hirsuta L. extracts. Phytomedicine 2018, 41, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.M. Review article on chemical constituents and biological activity of Thymelaea hirsuta. Rec. Pharm. Biomed. Sci. 2019, 3(2), 28–32. [Google Scholar] [CrossRef]
- Ziyyat, A.; Legssyer, A.; Mekhfi, H.; Dassouli, A.; Serhrouchni, M.; Benjelloun, W. Phytotherapy of hypertension and diabetes in oriental Morocco. J Ethnopharmacol. 1997, 58(1), 45–54. [Google Scholar] [CrossRef] [PubMed]
- Alafid, F.; Edrah, S.M.; Meelad, F.M.; Belhaj, S.; Altwair, K.; Maizah, N.R. Evaluation of phytochemical constituents and antibacterial activity of Thymelaea hirsuta (L.) Endl, and that utilised as a conventional treatment of infertility and diabetic in Libya. World J. Pharm. Res. 2019, 8(11), 72–88. [Google Scholar] [CrossRef]
- Abid, S.; Mekhfi, H.; Ziyyat, A.; Legssyer, A.; Aziz, M.; Bnouham, M. Beneficial effect of Thymelaea hirsuta on pancreatic islet degeneration, renal fibrosis, and liver damages as demonstrated in streptozotocin-induced diabetic rat. The Scientific World Journal 2021, 2021(1), 6614903. [Google Scholar] [CrossRef]
- Amari, N. O.; Razafimandimby, B.; Auberon, F.; Azoulay, S.; Fernandez, X.; Berkani, A.; Bouchara, J.P.; Landreau, A. Antifungal and antiaging evaluation of aerial part extracts of Thymelaea hirsuta (L.) endl. Nat. Prod. Com. 2021, 16(2), 1–10. [Google Scholar] [CrossRef]
- Kawano, M.; Matsuyama, K.; Miyamae, Y.; Shinmoto, H.; Kchouk, M.E.; Morio, T.; Shigemori, H.; Isoda, H. Antimelanogenesis effect of Tunisian herb Thymelaea hirsuta extract on B16 murine melanoma cells. Exp. Dermatol. 2007, 16(12), 977–984. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some Algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2005, 97(4), 654–660. [Google Scholar] [CrossRef]
- Borris, R.P.; Blaskó, G.; Cordell, G.A. Ethnopharmacologic and phytochemical studies of the Thymelaeaceae. J. Ethnopharmacol. 1988, 24(1), 41–91. [Google Scholar] [CrossRef]
- Lev, E. Reconstructed materia medica of the Medieval and Ottoman al-Sham. J. Ethnopharmacol. 2002, 80(2-3), 167–179. [Google Scholar] [CrossRef]
- Trigui, M.; Ben Hsouna, A.; Tounsi, S.; Jaoua, S. Chemical composition and evaluation of antioxidant and antimicrobial activities of Tunisian Thymelaea hirsuta with special reference to its mode of action. Ind. Crops Prod. 2013, 41, 150–157. [Google Scholar] [CrossRef]
- Ghamgui, H.; Jarboui, R.; Jeddou, K.B.; Torchi, A.; Siala, M.; Cherif, S.; Trigui, M. Polysaccharide from Thymelaea hirsuta L. leaves: Structural characterization, functional properties and antioxidant evaluation. Inter. J. Biol. Macromol. 2024, 262(2), 129244. [Google Scholar] [CrossRef]
- Helal, N.M.; Alharby, H.F.; Alharbi, B.M.; Bamagoos, A.A.; Hashim, A.M. Thymelaea hirsuta and Echinops spinosus: Xerophytic plants with high potential for first- generation biodiesel production. Sustainability 2020, 12, 1137. [Google Scholar] [CrossRef]
- Tahraoui, A.; El-Hilaly, J.; Israili, Z.H.; Lyoussi, B. Ethnopharmacological survey of plants used in the traditional treatment of hypertension and diabetes in south-eastern Morocco (Errachidia province). J. Ethnopharmacol. 2007, 110(1), 105–117. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Koubaa, I.; Dhouib, I.; Khemakhem, B.; Marchand, P.; Allouche, N. New specific α-glucosidase inhibitor flavonoid from Thymelaea tartonraira leaves: structure elucidation, biological and molecular docking studies. Chem. Biodiv. 2023, 20(3), e202200944. [Google Scholar] [CrossRef]
- Soltani, S.; Koubaa, I.; Cojean, S.; Picot, C.; Marchand, P.; Allouche, N. Phytochemical, antileishmanial, antifungal and cytotoxic profiles of Thymelaea tartonraira (L.) All. extracts. Nat. Prod. Res. 2024, 38(20), 3481–3487. [Google Scholar] [CrossRef] [PubMed]
- Margaris, N.S.; Adamandiadou, S.; Siafaca, L.; Diamantopoulos, J. Nitrogen and phosphorous content in plant species of Mediterranean ecosystems in Greece. Vegetatio 1984, 55(1), 29–35. [Google Scholar] [CrossRef]
- Meletiou-Christou, M.S.; Banilas, G.P.; Diamantoglou, S. (1998). Seasonal trends in energy conents and storage substances of the Mediterranean species Dittrichia viscosa and Thymelaea tartonraira. Environ. Exp. Bot. 1998, 39, 21–32. [Google Scholar] [CrossRef]
- Airò, M.; Fascella, G.; Zizzo, G.; Ruffoni, B. Preliminary studies on introduction and in vitro cultivation of Thymelaea hirsuta (L.) Endl. Italus Hortus 2004, 11(4), 144–146. [Google Scholar]
- Minuto, L.; Casazza, G.; Profumo, P. Population decrease of Thymelaea hirsuta (L.) Endl. in Liguria: conservation problems for the North Tyrrhenian sea. Plant Biosystems - An International Journal Dealing with All Aspects of Plant Biology 2004, 138(1), 11–19. [Google Scholar] [CrossRef]
- Papafotiou, M.; Triandaphyllou, N.; Chronopoulos, J. Studies on propagation of species of the xerophytic vegetation of Greece with potential floricultural use. Acta Hortic. 2000, 541, 269–272. [Google Scholar] [CrossRef]
- Papafotiou, M. In vitro propagation of temperate zone woody plants with potential ornamental use. Acta Hortic. 2010, 885, 255–262. [Google Scholar] [CrossRef]
- Shaltout, K.H.; El-Shourbagy, M.N. Germination requirements and seedling growth of Thymelaea hirsuta (L.) Endl. Flora 1989, 183(5-6), 429–436. [Google Scholar] [CrossRef]
- Vrijmoed, P. Collection, propagation and use of native plants. In Southern forest nursery association conference; Publisher: Southern Research Station: Asheville, USA, 1999; p. 156. [Google Scholar]
- Vogel, K.P. The challenge: High quality seed of native plants to ensure successful establishment. Seed Technology 2002, 24(1), 9–15. [Google Scholar]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two faces of one seed: hormonal regulation of dormancy and germination. Mol. Plant 2016, 9(1), 34–45. [Google Scholar] [CrossRef]
- Carrera-Castaño, G.; Calleja-Cabrera, J.; Pernas, M.; Gómez, L.; Oñate-Sánchez, L. An Updated Overview on the Regulation of Seed Germination. Plants 2020, 9, 703. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. The control of seed dormancy and germination by temperature, light and nitrate. Bot. Rev. 2020, 86(1), 39–75. [Google Scholar] [CrossRef]
- Koornneef, M.; Bentsink, L.; Hilhorst, H. Seed dormancy and germination. Current Opinion in Plant Biology 2002, 5(1), 33–36. [Google Scholar] [CrossRef]
- Shu, K.; Meng, Y.J.; Shuai, H.W.; Liu, W.G.; Du, J.B.; Liu, J.; Yang, W.Y. Dormancy and germination: How does the crop seed decide? Plant biology 2015, 17(6), 1104–1112. [Google Scholar] [CrossRef]
- Yang, L.; Liu, S; Lin, R. The role of light in regulating seed dormancy and germination. Journal of Integrative Plant Biology 2020, 62(9), 1310–1326. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ma, W.; Shen, S.; Gu, A. Underlying Biochemical and Molecular Mechanisms for Seed Germination. Int. J. Mol. Sci. 2022, 23, 8502. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, P. (2023). Review on dormancy, causes, uses, and measures of overcoming it. Science 2023, 7(2), 24–27. [Google Scholar] [CrossRef]
- Wang, Y.R.; Hanson, J.; Mariam, Y. Effect of sulfuric acid pretreatment on breaking hard seed dormancy in diverse accessions of five wild Vigna species. Seed Sci. Technol. 2007, 35, 550–559. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Seeds-physiology of development and germination, 2nd ed.; Publisher: Plenum Press: New York, NY, USA, 1994; p. 445. [Google Scholar]
- Probert, R.J. The role of temperature in the regulation of seed dormancy and germination. In Seeds. The ecology of regeneration in plant communities, 2nd ed.; Fenner, M., Ed.; Publisher: CAB International: Wallingford, UK, 2000; pp. 261–292. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Breaking seed dormancy during dry storage: A useful tool or major problem for successful restoration via direct seeding? Plants 2020, 9, 636. [Google Scholar] [CrossRef]
- Chandra, R. J.; Masilamani, P.; Suthakar, B.; Rajkumar, P.; Sivakumar, S.D.; Manonmani, V. Seed dormancy and after-ripening mechanisms in seed germination: A comprehensive review. International Journal of Plant & Soil Science 2024, 36(10), 68–92. [Google Scholar] [CrossRef]
- Moore, R.P.E. Handbook on Tetrazolium Testing, 1st ed.; Publisher: The International Seed Testing Association (ISTA): Bassersdorf, Switzerland, 1985; p. 99. [Google Scholar]
- Papafotiou, M.; Martini, A.N. In vitro seed and clonal propagation of the Mediterranean aromatic and medicinal plant Teucrium capitatum. HortScience 2016, 51(4), 403–411. [Google Scholar] [CrossRef]
- Martini, A.N.; Papafotiou, M. In vitro seed and clonal propagation of the Mediterranean bee friendly plant Anthyllis hermanniae L. Sustainability 2023, 15(5), 4025. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Nadjafi, F.; Bannayan, M.; Tabrizi, L.; Rastgoo, M. Seed germination and dormancy breaking techniques for Ferula gummosa and Teucrium polium. J. Arid Environ. 2006, 64, 542–547. [Google Scholar] [CrossRef]
- Ibañez, A.; Passera, C. Factors affecting the germination of albaida (Anthyllis cytisoides L.), a forage legume of the Mediterranean coast. J. Arid. Environ. 1997, 35, 225–231. [Google Scholar] [CrossRef]
- Bertsouklis, K.; Vlachou, G.; Trigka, M.; Papafotiou, M. In vitro studies on seed germination of the Mediterranean species Anthyllis barbajovis to facilitate its introduction into the floriculture industry. Horticulturae 2022, 8, 889. [Google Scholar] [CrossRef]
- Vlachou, G.; Martini, A.Ν.; Dariotis, E.; Papafotiou, Μ. Comparative evaluation of seed germination of five Mediterranean sage species (Salvia sp.) native to Greece. Acta Hortic. 2020, 1298, 593–598. [Google Scholar] [CrossRef]
- Soltani, E.; Baskin, J.M.; Baskin, C.C.; Benakashani, F. A meta-analysis of the effects of treatments used to break dormancy in seeds of the megagenus Astragalus (Fabaceae). Seed Sci. Res. 2020, 30, 224–233. [Google Scholar] [CrossRef]
- Kelly, K.M.; Van Staden, J.; Bell, W.E. Seed coat structure and dormancy. Plant Growth Regul 1992, 11, 201–209. [Google Scholar] [CrossRef]
- Bandera, M.C.d.l.; Traveset, A. Reproductive ecology of Thymelaea velutina (Thymelaeaceae)-Factors contributing to the maintenance of heterocarpy. Plant Syst. Evol. 2005, 256, 97–112. [Google Scholar] [CrossRef]
- Di Sacco, A.; Gajdošová, Z.; Slovák, M; Turisová, I.; Turis, P.; Jaromír Kučera, J.; Müller, J.V. Daphne arbuscula (Thymelaeaceae) compared to the more widespread Daphne cneorum. Folia Geobot 2021, 656, 13–25. [Google Scholar] [CrossRef]
- Dawson, P.A.C.; Rapson, G.L.; Robertson, A.W.; Fordham, R.A. Limitations on recruitment of the rare sand daphne Pimelea arenaria (Thymelaeaceae), lower North Island, New Zealand. N. Z. J. Bot. 2005, 43(3), 619–630. [Google Scholar] [CrossRef]
- Rolston, M.P. Water impermeable seed dormancy. Bot. Rev. 1978, 44, 365–396. [Google Scholar] [CrossRef]
- Silcock Richard, G.; Mann Michael, B. (2014) Germinating the seeds of three species of Pimelea sect. Epallage (Thymelaeaceae). Austr. J. Bot. 2014, 62, 74–83. [Google Scholar] [CrossRef]
- Tabin, T.; Shrivastava, K. (2014). Factors affecting seed germination and establishment of critically endangered Aquilaria malaccensis (Thymelaeaceae). Asian J. Plant Sci. Res. 2014, 4(6), 41–46. [Google Scholar]
- De Alwis, H.N.; Subasinghe, S.M.C.U.P.; Hettiarachchi, D. S. Effect of storage time and temperature on Gyrinops walla Gaertn. seed germination. J. Envir. Prof. Sri Lanka 2016, 5(2), 16–24. [Google Scholar]
- Galmés, J.; Medrano, H.; Flexas, J. Germination capacity and temperature dependence in Mediterranean species of the Balearic Islands. Forest Systems 2006, 15(1), 88–95. [Google Scholar] [CrossRef]
- Thanos, C.A.; Kadis, C.C.; Skarou, F. Ecophysiology of germination in the aromatic plants thyme, savory and oregano (Labiatae). Seed Sci. Res. 1995, 5, 161–170. [Google Scholar] [CrossRef]
- Thanos, C.A.; Doussi, M.A. Ecophysiology of seed germination in endemic Labiates of Crete. Israel J. Plant Sci. 1995, 43, 227–237. [Google Scholar] [CrossRef]
- Kanellou, E.; Vlachou, G.; Martini, A.N.; Bertsouklis, K.; Papafotiou, M. Seed germination of five sage species (Salvia sp.) of populations native to Greece. Acta Hortic. 2022, 1345, 439–444. [Google Scholar] [CrossRef]
Figure 1.
Characteristic plant (A), shiny green scale-like leaves (B) and small yellowish flowers (C) of Thymelaea hirsuta; Typical plant (D), silvery-grayish silkily hairy leaves (E) and yellowish flowers (F) of Thymelaea tartonraira ssp. tartonraira.
Figure 1.
Characteristic plant (A), shiny green scale-like leaves (B) and small yellowish flowers (C) of Thymelaea hirsuta; Typical plant (D), silvery-grayish silkily hairy leaves (E) and yellowish flowers (F) of Thymelaea tartonraira ssp. tartonraira.

Figure 2.
Seeds of Thymelaea hirsuta after removal of dried floral debris (A); Typical germination of seeds scarified by concentrated H2SO4 for 20 min and incubated under 16 h photoperiod for 30 days at: 10 oC (B); 15 oC (C); 20 oC (D); 25 oC (E) and 30 oC (F), as well as incubated at 25 oC, under 16 h photoperiod or continuous darkness (G); Abnormal radicle development in seedlings germinated at 30 oC (H).
Figure 2.
Seeds of Thymelaea hirsuta after removal of dried floral debris (A); Typical germination of seeds scarified by concentrated H2SO4 for 20 min and incubated under 16 h photoperiod for 30 days at: 10 oC (B); 15 oC (C); 20 oC (D); 25 oC (E) and 30 oC (F), as well as incubated at 25 oC, under 16 h photoperiod or continuous darkness (G); Abnormal radicle development in seedlings germinated at 30 oC (H).

Figure 3.
Preliminary observations on the effect of seed pretreatments, as well as of incubation temperature and photoperiod on in vitro germination of Thymelaea hirsuta seeds, 5 months (A), 9 months (B) and 12 months (C and D) after harvesting and storage at room temperature. Mean values (n= 5 repetitions of 20 seeds) in each figure followed by the same lower-case letter do not differ significantly at p ≤ 0.05 by Student’s t test. § **, significant at p ≤ 0.01.
Figure 3.
Preliminary observations on the effect of seed pretreatments, as well as of incubation temperature and photoperiod on in vitro germination of Thymelaea hirsuta seeds, 5 months (A), 9 months (B) and 12 months (C and D) after harvesting and storage at room temperature. Mean values (n= 5 repetitions of 20 seeds) in each figure followed by the same lower-case letter do not differ significantly at p ≤ 0.05 by Student’s t test. § **, significant at p ≤ 0.01.
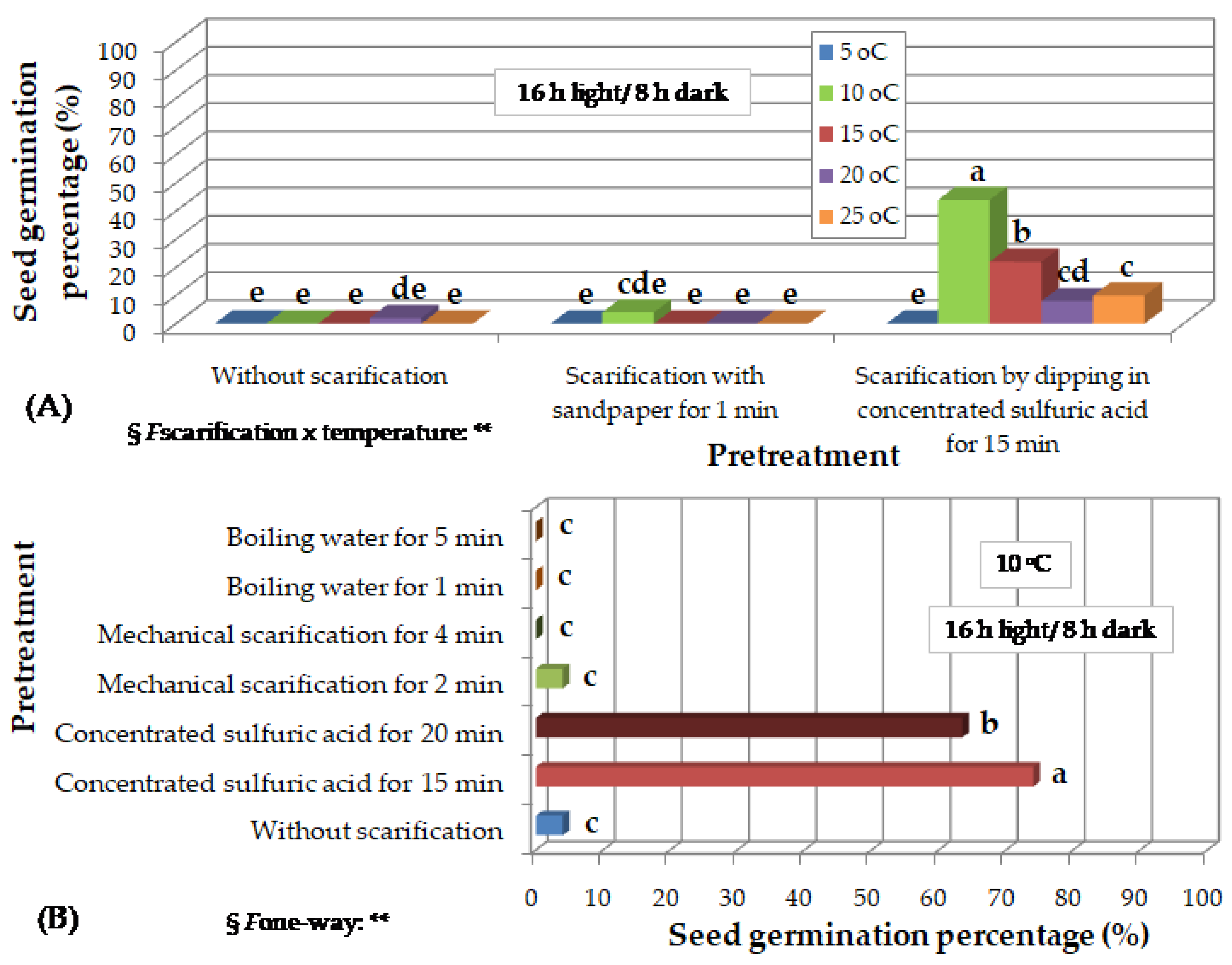
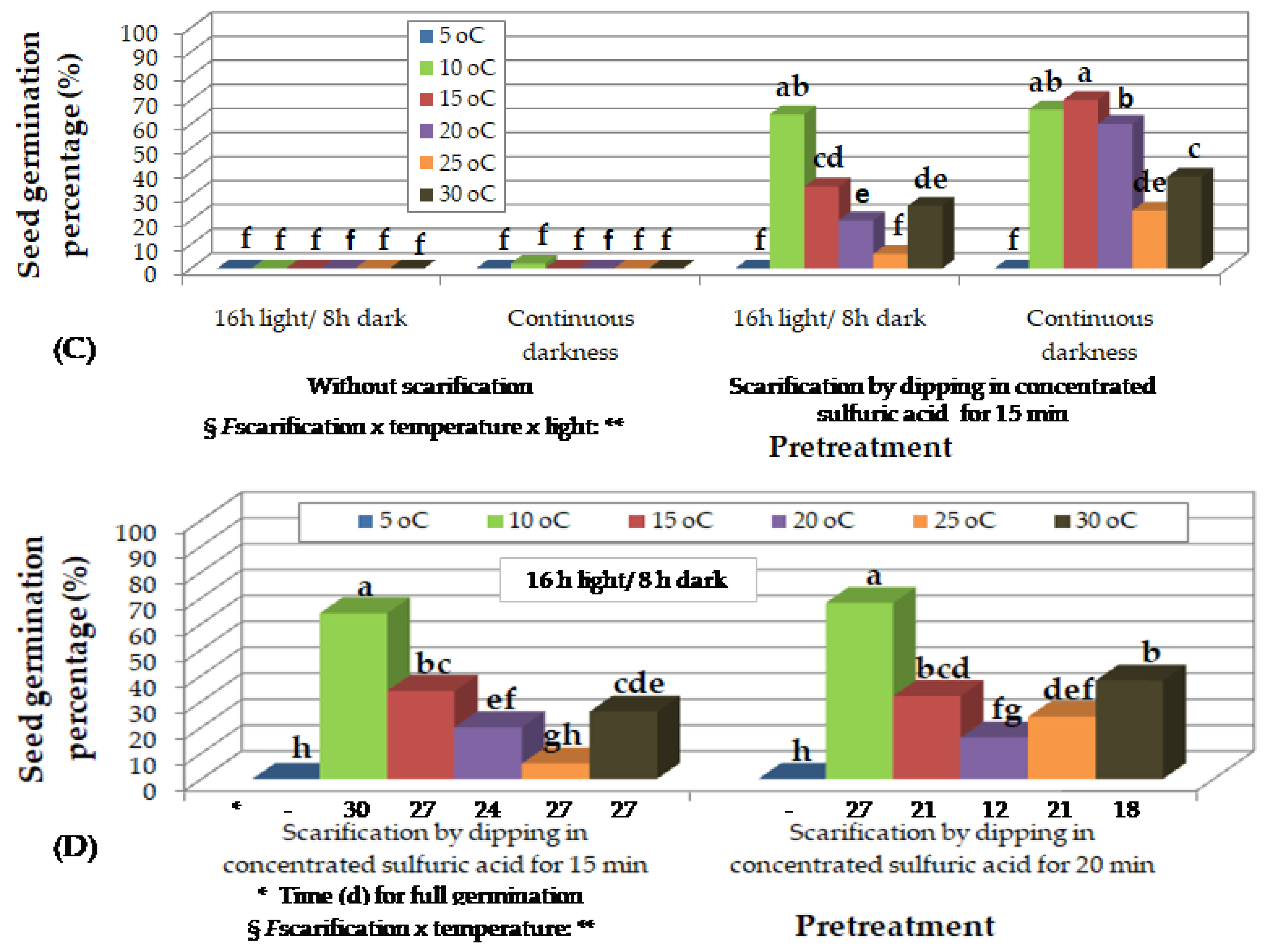
Figure 4.
Preliminary observations on the effect of incubation temperature and photoperiod on in vitro germination of Thymelaea hirsuta seeds, scarified by dipping in concentrated sulfuric acid for 20 min, 18 months (A) and 24 months (Β), after harvesting and storage at room temperature. Mean values (n= 5 repetitions of 20 seeds) in each figure followed by the same lower-case letter do not differ significantly at p ≤ 0.05 by Student’s t test. § NS or * or **, non-significant at p ≤ 0.05 or significant at p ≤ 0.05 or p ≤ 0.01, respectively.
Figure 4.
Preliminary observations on the effect of incubation temperature and photoperiod on in vitro germination of Thymelaea hirsuta seeds, scarified by dipping in concentrated sulfuric acid for 20 min, 18 months (A) and 24 months (Β), after harvesting and storage at room temperature. Mean values (n= 5 repetitions of 20 seeds) in each figure followed by the same lower-case letter do not differ significantly at p ≤ 0.05 by Student’s t test. § NS or * or **, non-significant at p ≤ 0.05 or significant at p ≤ 0.05 or p ≤ 0.01, respectively.
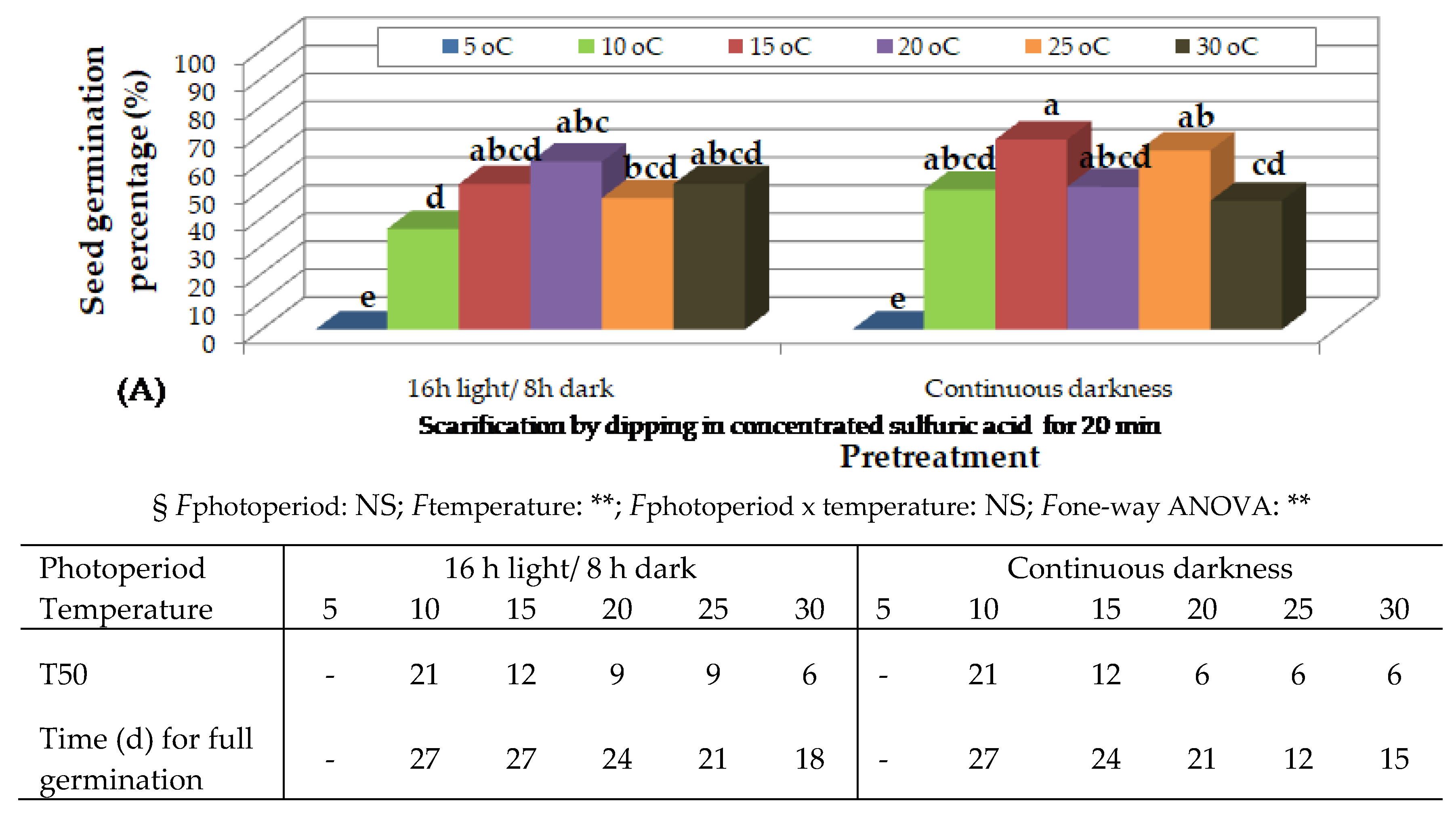
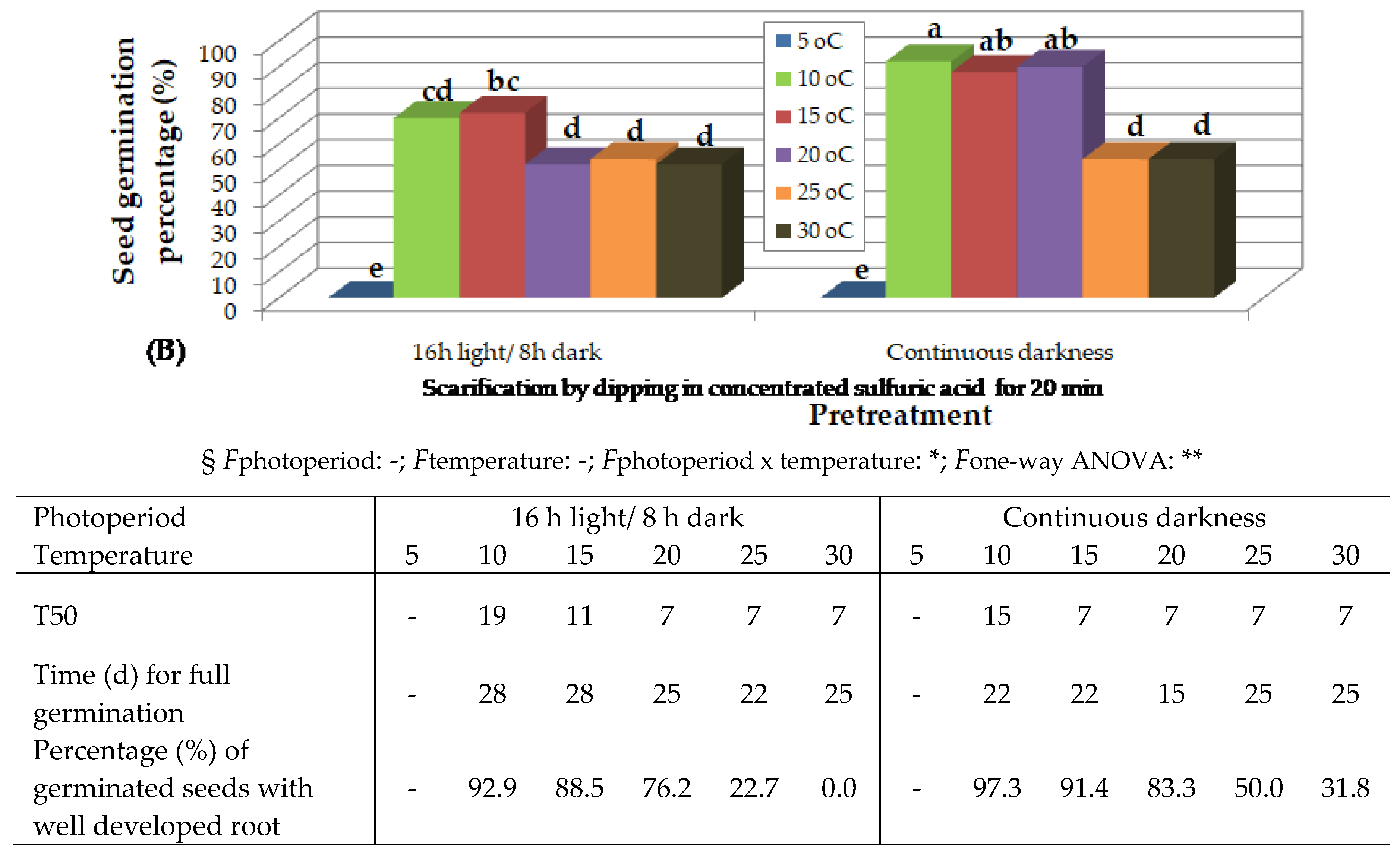
Figure 5.
Preliminary observations on the effect of temperature on in vitro germination of Thymelaea tartonraira seeds, scarified by dipping in concentrated sulfuric acid for 20 min and incubated under 16 h photoperiod, 4 months (A) and 12 months (B), after harvesting and storage at room temperature. Mean values (n= 5 repetitions of 20 seeds) in each figure followed by the same lower-case letter do not differ significantly at p ≤ 0.05 by Student’s t test. § NS or * or **, non-significant at p ≤ 0.05 or significant at p ≤ 0.05 or p ≤ 0.01, respectively.
Figure 5.
Preliminary observations on the effect of temperature on in vitro germination of Thymelaea tartonraira seeds, scarified by dipping in concentrated sulfuric acid for 20 min and incubated under 16 h photoperiod, 4 months (A) and 12 months (B), after harvesting and storage at room temperature. Mean values (n= 5 repetitions of 20 seeds) in each figure followed by the same lower-case letter do not differ significantly at p ≤ 0.05 by Student’s t test. § NS or * or **, non-significant at p ≤ 0.05 or significant at p ≤ 0.05 or p ≤ 0.01, respectively.
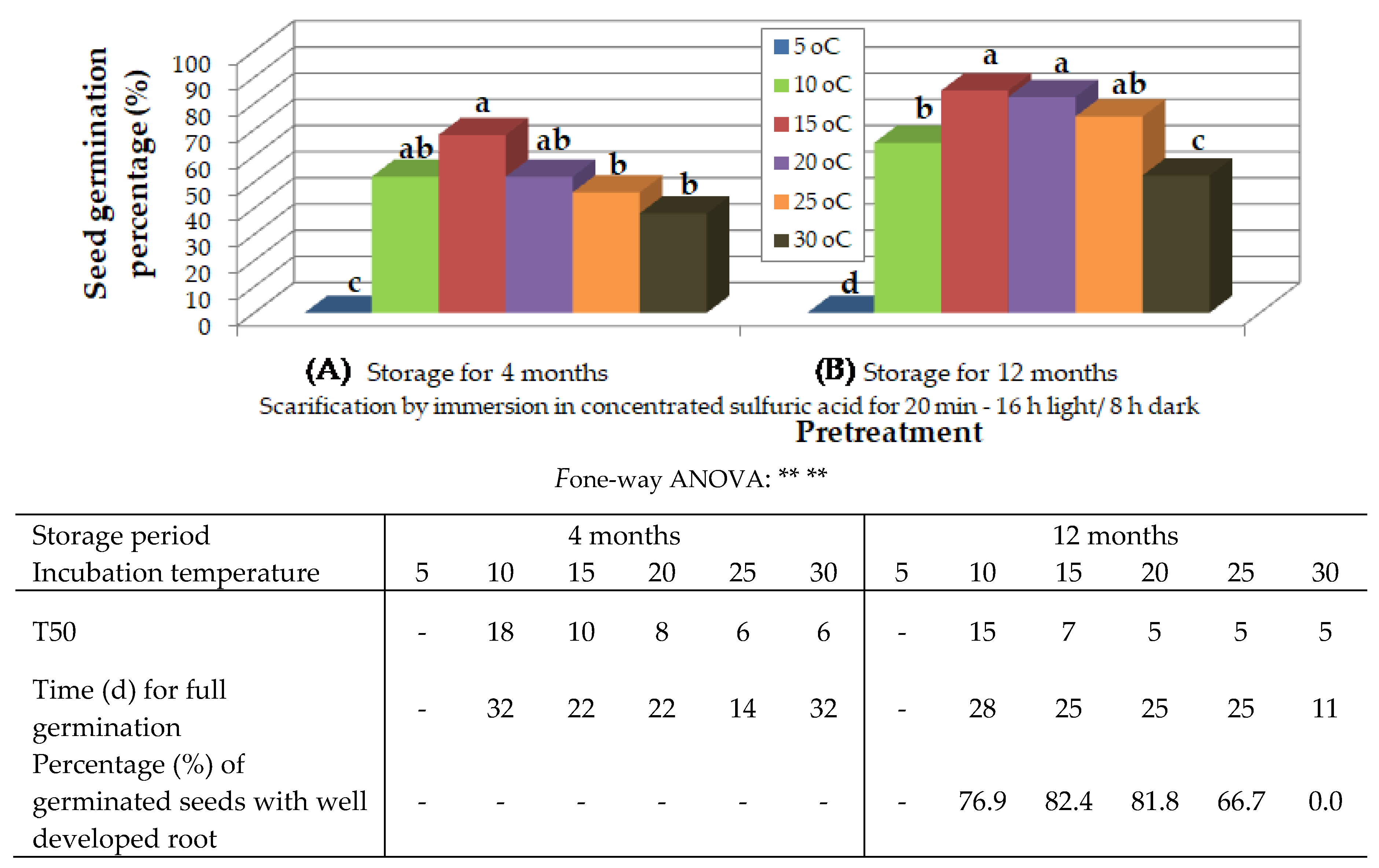

Table 1.
Effect of scarification, incubation temperature and photoperiod on in vitro germination of T. hirsuta, nine and twenty-three months (9M/23M) after seed collection and storage at room temperature.
Table 1.
Effect of scarification, incubation temperature and photoperiod on in vitro germination of T. hirsuta, nine and twenty-three months (9M/23M) after seed collection and storage at room temperature.
| Sarification | Light conditions | Incubation temperature (oC) | Seed germination percentage (%) | Germination (%) with well developed seedling | T50 |
| (9M/23M) | (9M/23M) | (9M/23M) | |||
| Without scarification | 16 h light/ 8 h dark | 10 | 0.0 e§/ 0.0 f | 0.0 g/ 0.0 d | -/ - |
| 15 | 0.0 e/ 0.0 f | 0.0 g/ 0.0 d | -/ - | ||
| 20 | 0.0 e/ 0.0 f | 0.0 g/ 0.0 d | -/ - | ||
| 25 | 0.0 e/ 0.0 f | 0.0 g/ 0.0 d | -/ - | ||
| Continuous darkness | 10 | 2.0 e/ 0.0 f | 2.0 fg/ 0.0 d | 19/ - | |
| 15 | 2.0 e/ 0.0 f | 2.0 fg/ 0.0 d | 18/ - | ||
| 20 | 0.0 e/ 0.0 f | 0.0 g/ 0.0 d | -/ - | ||
| 25 | 0.0 e/ 0.0 f | 0.0 g/ 0.0 d | -/ - | ||
| Scarification by immersing in concentrated sulfuric acid for 20 min | 16 h light/ 8 h dark | 10 | 55.3 c/ 76.0 c | 52.8 c/ 76.0 b | 22/ 20 |
| 15 | 33.8 d/ 22.0 e | 33.8 d/ 22.0 c | 17/ 18 | ||
| 20 | 32.8 d/ 16.0 ef | 19.4 ef/ 12.0 cd | 12/ 11 | ||
| 25 | 31.1 d/ 50.0 d | 11.1 fg/ 20.0 cd | 13/ 13 | ||
| Continuous darkness | 10 | 93.1 a/ 96.4 a | 88.1 a/94.0 a | 18/ 17 | |
| 15 | 91.3 a/ 100.0 a | 88.8 a/ 96.0 a | 11/ 10 | ||
| 20 | 78.6 b/ 100.0 a | 70.6 b/ 96.0 a | 7/ 8 | ||
| 25 | 50.0 c/ 84.0 b | 31.1 d/ 64.0 b | 8/ 7 | ||
| Fscarification | -/ - | -/ - | |||
| Fphotoperiod | -/ - | -/ - | |||
| Ftemperature | -/ - | -/ - | |||
| Fscarification x photoperiod | **/ ** | **/ ** | |||
| Fscarification x temperature | **/ ** | **/ ** | |||
| Fphotoperiod x temperature | */ ** | */ ** | |||
| Fscarification x photoperiod x temperature | **/ ** | **/ ** | |||
| Fone-way ANOVA | **/ ** | **/ ** | |||
| Mean values (n= 5 repetitions of 20 seeds) in each column followed by the same lower-case letter do not differ significantly at p ≤ 0.05 by Student’s t test. § * or **, significant at p ≤ 0.05 or p ≤ 0.01, respectively. | |||||
Table 2.
Effect of scarification, incubation temperature and light conditions on in vitro seed germination of T. tartonraira, nine and twenty-three months (9M/23M) after seed collection and storage at room temperature.
Table 2.
Effect of scarification, incubation temperature and light conditions on in vitro seed germination of T. tartonraira, nine and twenty-three months (9M/23M) after seed collection and storage at room temperature.
| Sarification | Light conditions | Incubation temperature (oC) | Seed germination percentage (%) | Germination (%) with well developed seedling | T50 |
| (9M/23M) | (9M/23M) | (9M/23M) | |||
| Without scarification | 16 h light/ 8 h dark | 10 | 0.0 d§/ 0.0 e | 0.0 d/ 0.0 d | -/ - |
| 15 | 0.0 d/ 0.0 e | 0.0 d/ 0.0 d | -/ - | ||
| 20 | 2.0 d/ 0.0 e | 0.0 d/ 0.0 d | 23/ - | ||
| 25 | 0.0 d/ 0.0 e | 0.0 d/ 0.0 d | -/ - | ||
| Continuous darkness | 10 | 0.0 d/ 0.0 e | 2.0 d/ 0.0 d | -/ - | |
| 15 | 2.0 d/ 2.0 e | 2.0 d/ 2.0 d | 25/ 12 | ||
| 20 | 0.0 d/ 0.0 e | 0.0 d/ 0.0 d | -/ - | ||
| 25 | 0.0 d/ 0.0 e | 0.0 d/ 0.0 d | -/ - | ||
| Scarification by immersing in concentrated sulfuric acid for 20 min | 16 h light/ 8 h dark | 10 | 60.0 c/ 44.0 d | 51.9 b/ 34.0 bc | 23/ 18 |
| 15 | 90.0 a/ 54.0 abcd | 77.5 a/ 28.0 c | 13/ 12 | ||
| 20 | 72.5 abc/ 46.4 cd | 52.5 b/ 28.2 c | 9/ 7 | ||
| 25 | 67.5 bc/ 58.0 abc | 25.0 c/ 24.0 c | 6/ 6 | ||
| Continuous darkness | 10 | 75.0 ab/ 52.0 bcd | 72.5 a/38.0 bc | 18/ 15 | |
| 15 | 70.0 bc/ 68.0 a | 52.5 b/ 56.0 a | 12/ 10 | ||
| 20 | 80.0 ab/ 62.0 ab | 57.5 b/ 50.0 ab | 8/ 9 | ||
| 25 | 70.0 bc/ 60.0 abc | 40.4 bc/ 38.0 bc | 7/ 7 | ||
| Fscarification | **/ - | **/ - | |||
| Flight conditions | NS/ - | NS/ - | |||
| Ftemperature | NS/ - | NS/ NS | |||
| Fscarification x photoperiod | NS/ ** | NS/ * | |||
| Fscarification x temperature | NS/ ** | NS/ NS | |||
| Fphotoperiod x temperature | NS/ ** | NS/ NS | |||
| Fscarification x photoperiod x temperature | NS/ ** | NS/ NS | |||
| Fone-way ANOVA | **/ ** | **/ ** | |||
| Mean values (n= 5 repetitions of 20 seeds) in each column followed by the same lower-case letter do not differ significantly at p ≤ 0.05 by Student’s t test. § NS or * or **, non-significant at p ≤ 0.05 or significant at p ≤ 0.05 or p ≤ 0.01, respectively. | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



