
Submitted:
02 January 2024
Posted:
05 January 2024
You are already at the latest version
Abstract
The ex situ conservation and sustainable exploitation of neglected or underutilized plant species (NUPs) is urgent and paramount. To this end, we focused on Petromarula pinnata (Campanu-laceae), a vulnerable local endemic of Crete (Greece) garnering interest for its agro-alimentary, medicinal, and ornamental value. A GIS ecological profile was established herein based on its natural distribution in Crete containing detailed information on the climatic conditions (mini-mum, maximum, mean temperature, and precipitation) as well as information on 19 bioclimatic variables shaping its natural adaptation. This profiling contributed to better understanding of the species' ecological requirements and facilitated the germination trials employing stored seeds from four distinct populations (two from lowlands and two from semi-mountainous areas) tested at four temperatures (10, 15, 20 and 25oC) and two light condition treatments. The results herein showed that both temperatures and population as well as their interaction significantly affected seed germination rates. Incubation temperatures of 10 and 15oC were the most appro-priate for the successful seed germination of this species (> 81.25% for both temperatures in three out of four populations), with light conditions not affecting seed germination (86% in light and 80% in darkness). The successful germination protocol of P. pinnata seeds opens avenues for fur-ther sustainable exploitation of this valuable yet vulnerable NUP as a new Greek native crop.
Keywords:
Agro-alimentary
; medicinal
; ornamental interest
; neglected and underutilized plants
; phytoge-netic resources
; sustainable exploitation
; Campanulaceae
1. Introduction
Several members of Campanulaceae are commonly appreciated as ornamental plants worldwide and many of them are praised for their attractive flowers which can vary in color from deep violet to the palest milky blue [1]. Consequently, many members of Campanulaceae are often used as garden plants, potted plants, and potentially as cut flowers, thus representing a growing trend in the development of new ornamental plants [2,3]. Within Campanulaceae, some genera and/or species are unique such as the monotypic genus Petromarula, with P. pinnata (L.) A. DC. being exclusively found on the island of Crete, Greece [4,5].
Besides ornamental appreciation, many Campanulaceae species have traditionally been used as a source of food by residents worldwide [6,7]. For example, French residents or Italian and Spanish inhabitants are reported to consume raw roots of Campanula rapunculus L. in salads or in hot soups, while Chinese traditionally use dried roots of Codonopsis pilosula (Franch.) Nannf. in soups, all sourced directly from the wild due to their nutritional benefits [6,7]. The roots are not the only exploitable part of such plants, as previous studies have shown that the aerial parts (mainly leaf rosettes) of several Campanula spp. sourced from the wild are also used as edible greens in salads, i.e., C. rapunculus in Spain [8,9] or C. pelviformis L. in Crete [10] and P. pinnata in Crete [11,12]. Current research has shown that such ethnobotanically used wild food sources can also provide a particular nutritional value combined with medicinal properties as illustrated for example in the case of C. pelviformis [10] or P. pinnata [12,13]. The latter local Cretan species is often among the basic ingredients of traditional ‘kaltsounia’ which are small halfmoon-shaped pastry filled with a mixture of wild edible greens directly sourced from the wild and cultivated vegetables (e.g. spinach and fresh green onions) along with a mixture of local cheeses (tiromalama or malaka and pungent mizithra) [11]. These pastries locally calles ‘kaltsounia’ in western Crete or ‘chortopitakia’ in central and eastern Crete, are prepared in local houses or in Cretan bakeries and are almost daily consumed as a snack. Most importantly, the consumption of only two pieces of Cretan ‘kaltsounia’ with wild and cultivated greens and cheese correspond to 100 g of food covering almost 40% of the estimated daily intake of strong antioxidants (flavonols and flavones) benefiting from the local version of the Mediterranean diet of the Cretan population [11].
Sustainable exploitation of neglected and underutilized plant species (NUPs) into agricultural settings [14] may alleviate the over-harvesting pressure to wild populations of threatened local endemic plants [3,15,16]. In response to this need, propagation, and cultivation practices for NUPs ought to be developed and established, respectively, in the first place prior to sustainable exploitation [3,15,16]. To this end, sexual propagation constitutes a highly effective and low-cost solution in plant production for many plant species [17] as well as an appropriate propagation method for conservation-oriented research due to maintenance of high genetic diversity of the species in concern [18,19,20,21,22]. Seed germination tests provide detailed and species-specific information on the range of environmental conditions required for the germination of fresh or stored mature seeds, thus shedding light on how target species adapt to their natural environment during this critical phase of their life cycle [3,20,21,22,23].
In support of seed germination studies, species-specific ecological profiles provide comprehensive information on the autoecology of the species in concern, including insight into the prevailing climatic conditions at the natural distribution sites of the species throughout the year which may facilitate the design of seed germination experiments [24]. In this direction, identifying and confirming through experimentation the favorable temperature range for high rates of seed germination in a certain species can effectively enhance their reproduction potential, thus facilitating conservation actions or ensuring the production of seedlings for further development of cultivation protocols with the aim to create new value chains even for rare and/or threatened NUPs [3,20,21,23,25]. Moreover, the species-specific germination protocols derived from such studies are also very useful for massive production of newly introduced plant species by commercial nurseries, thus representing basic know-how that can be exploited in the ornamental-floriculture sector or the medicinal and agro-alimentary industries [3,26].
In this context, targeted research has confirmed the presence of various isolated compounds with notable pharmaceutical properties in C. pelviformis [10], a threatened, range-restricted, local Cretan endemic NUP with ornamental, medicinal, and agro-alimentary interest [3,15,16]; the latter coupled with analytical seed germination protocol developed [25] which has recently paved the way for the sustainable exploitation of this NUP in the frame of newly launched national research project employing pilot cultivation in Crete (N. Krigas, pers. comm.). Following up on this framework, similar research lines are triggered to date aiming to investigate other Cretan species of the Campanulaceae family sharing similar characteristics with biological and economic importance. Species-wise, this study is focused on Petromarula pinnata (L.) A.DC. which ranks high among other Mediterranean NUPs, thus indicating a highly valuable potential for sustainable exploitation. With limited data on its seed germination [27,28] and building up on previous studies showing significant value in different economic sectors namely ornamental [3], agro-alimentary [16] and medicinal-cosmetic [15], as well as in parallel with undertaken pharmacognostic investigations [12], D. Lazari, pers. comm. and manuscript in preparation), the present investigation aimed to achieve the following objectives: (i) Using Geographical Information System (GIS), to unveil the climate range and the ideal conditions for seed germination in P. pinnata based on the ecological profiles of the natural sites where P. pinnata wild-growing populations naturally grow, and (ii) To exploit this knowledge in testing seed germination of stored seeds from four populations collected at different altitudes under different temperature levels and light conditions. Such applied research can also contribute to the conservation of this vulnerable species through targeted collections in the natural environment allowing for enhanced ex situ conservation in seed banks and species-specific propagation and cultivation, thus enabling know-how either for re-introduction purposes when needed or pilot production of plant material for novel cultivations as for example [19,20,21,22,29,30].
2. Materials and Methods
2.1. Characteristics of the focal plant species
Petromarula pinnata (Figure 1) belongs to the monotypic genus Petromarula of Campanulaceae, and it is exclusively found on the island of Crete, Greece [4,5]. P. pinnata thrives as chasmophyte in many crevices of limestone cliffs, usually in semi-shade places across the island of Crete, and it may also be found frequently on stonewalls of old buildings and fortresses [5]. Its distribution ranges from sea level up to 800 or even up to 1300 m occasionally, being in flower during April and May or later, depending on altitude and slope [5]. Hence, P. pinnata is considered as a widely scattered hemicryptophyte in Crete [4,5]. It is a local single-island endemic plant which is currently assessed as ‘Vulnerable’ [31].
2.2. GIS Ecological Profiling
The ecological profile of P. pinnata was developed in GIS, following the methodology applied previously to other species with similar characteristics and conservation concern [18,20,21,23]. In summary, the natural distribution points of P. pinnata (n = 51) as derived from a previous study [32] were herein linked with climate and precipitation data (minimum, maximum, and average values) as well as with 19 bioclimatic variables, all based on climate historical data of 30 years (1970-2000) with pixel size 30 sec and spatial resolution of 1 km2 (https://www. worldclim.org/data/worldclim21.html, accessed on 15 June 2023).
2.3. Seed Collection and Storage
All mature seeds of P. pinnata were collected on the same day (29 August 2018) from up to five individuals per accessed wild-growing populations found in different locations in Crete (Table 1). The seeds were collected using a special collection permit (182336/879 of 16 May 2019 and 64886/2959 of 6 July 2020) issued by the Greek Ministry of Environment and Energy. Seed storage conditions (three years storage) were as those mentioned in previous studies [25].
2.4. Germination Tests
Prior to germination experiments, morphological measurements such as weight of 1000 seeds and average length/width of 30 seeds were obtained for every population of P. pinnata [25]. The weight of 1000 seeds was 0.041 g for GR-BBGK-1-19,97; 0,044 g for GR-BBGK-1-19,126; 0,034 g for GR-BBGK-1-19,124; and 0.0432 g for GR-BBGK-1-19,130. The average length/width ranged from 0.206/0.107 to 0.231/0.114 mm for lowland populations, while semi-mountainous populations had relatively larger seeds ranging from 0.217/0.117 to 0.258/0.126 mm.
Germination tests were conducted following the germination protocol described in previous study ([25] and all trials took place at the Laboratory of Floriculture, School of Agriculture, Aristotle University of Thessaloniki (Thermi, Greece) in April 2022. In brief, seed responses per incubation temperature were examined at four different temperatures (10, 15, 20 and 25oC) using four replications of 20 seeds in populations GR-BBGK-1-19,97 and GR-BBGK-1-19,126, and four replications of 25 seeds in populations GR-BBGK-1-19,124 and GR-BBGK-1-19,130.
The light requirements of P. pinnata were investigated after the end of germination experiments [25]. Only seeds from population GR-BBGK-1-19,124 were used for this experiment conducted in a controlled-temperature chamber set at 15oC.
2.5. Statistical Analysis
In the germination tests, the experimental design was a completely randomized factorial design, with the population and incubation temperature as separate factors. The germination data in incubation temperature of 25°C was not analyzed, as none of the seeds germinated or the percentages of germinated seeds were very low (< 3%). Therefore, in the statistical analysis, the levels for the population factor were four combined with three for the incubation temperature factor (4 x 3 factorial design). The data was analyzed using the ANOVA method in the frame of the GLM (General Linear Model) [33]. The germination percentage data was transformed into arc-sine square root values, before analysis [34]. The transformed data was checked for normality and homogeneity of variances and then was analyzed by ANOVA, while the comparisons of the means were performed using the LSD test at a significance level of p ≤ 0.05. In the comparisons between the two light conditions (alternating light/dark and continuous dark conditions), the T test was used [35]. All statistical analyses were carried out using SPSS 27.0 (SPSS, ΙΒΜ, Inc., USA).
3. Results
3.1. Ecological profiling
The ecological profile of P. pinnata was generated in GIS based on its natural distribution occurrences in Crete to illustrate the favorable climate conditions of its wild-growing populations (Figure 2). According to historical climate data, the lowest average temperatures were recorded during the first annual quarter (9.49±1.11oC, 9.52±1.12oC, 10.95±1.08oC in January, February, and March, respectively). When such data were combined with the mean temperature of coldest quarter (9.98±1.10oC) and minimum temperature of coldest month (6.48±1.06oC), it was concluded that P. pinnata wild-growing populations thrive in areas with mild winters. From April (14.05±1.03oC) to June (22.25±0.93oC), the average temperature in these areas gradually rose until July (24.28±0.94oC) and August (24.09±0.94oC), showing the highest average temperatures. This period coincides with the natural flowering and fruiting of P. pinnata wild-growing populations. Despite the latter being the warmest period for P. pinnata, the average temperature was shown to remain around 25oC (maximum temperature of warmest month = 28.39±0.87oC). After the summer season, the average temperature in these areas were shown to decrease from September (21.64±0.97oC) to December (11.05±1.10oC). Considering all the above-mentioned data as well as the lowest and highest temperature limits (Tmin of Tmin = 4.15oC in February, Tmax of Tmax = 29.90oC in July) and the mean diurnal range (7.22±0.13oC), it seems that P. pinnata wild-growing populations thrive in environments with no extreme climate conditions. These temperature limits may showcase the natural adaptation of the wild-growing populations of P. pinnata.
P. pinnata wild-growing populations thrive in habitats with a notable rainy season, as indicated by the precipitation-related attributes in Figure 2. The pattern indicated a rainy season starting in mid-October (73.16±8.88 mm) and lasting until mid-March (86.72±8.63 mm), with January being the wettest month (142.40±14.44 mm). From April (37.77±5.43 mm) and onwards while P. pinnata populations are in flower [5], the precipitation values were shown to decrease significantly until June (5.75±1.63 mm), marking the onset of the dry season (precipitation of driest quarter = 10.81±3.31 mm) and the fruit setting period during which wild-growing plant individuals are dried. From September (18.06 ± 1.77 mm), rainfall patterns were shown to rise again until the onset of the rainy season. These precipitation patterns may showcase the natural adaptation of the wild-growing populations of P. pinnata allowing them to fulfill their biological cycle.
3.2. Seed Germination Success
3.2.1. Effect of incubation temperature
According to the results of the statistical analysis, both the main effects of population and temperature incubation as well as their interaction significantly affected the germination of P. pinnata seeds (Table 2). In detail, the seeds of population GR-BBGK-1-19,97 collected from the highest altitude exhibited the lowest germination percentage (Figure 3). Furthermore, the germination percentage of population GR-BBGK-1-19,124 was higher than that of population GR-BBGK-1-19,126 (both collected at lower altitudes). The germination percentage of seeds incubated at 20oC was the lowest, while no statistical difference was observed between the germination percentage of seeds incubated at 10 and 15oC (Figure 4).
Significant differences were observed among the germination percentage of seeds incubated at 10, 15 and 20oC (Figure 5). Specifically, the seeds of populations GR-1-BBGK-19,126 and GR-1-BBGK-19,130 which were incubated at 10 and 15oC germinated at higher percentages (81.25% and 85% for first one and 90% and 89% for the second one, respectively) compared to those incubated at 20oC (65% and 79%, respectively). In population GR-1-BBGK-19,97, the highest germination percentage was achieved at 15oC (75%) whereas in population GR-1-BBGK-19,124 the seeds which were incubated at 10oC germinated at higher percentages (91% and 84%, respectively) compared to those incubated at 20oC (79%) (Figure 5, p<0.05). According to Figure 5, in all populations, the seeds incubated at 15oC germinated earlier than those incubated at 10oC. Specifically, germination of the seeds was recorded on the 10th day and seed germination completion was achieved on the 30th day.
Furthermore, the incubation temperature showed significant impact on the seed germination of four wild populations of P. pinnata (Table 3, p<0.05). Specifically, at 10 oC, the seeds from population GR-1-BBGK-19,97 exhibited the lowest germination percentage (48.75%), while no statistically significant difference was observed among the remaining three populations. At incubation temperature of 15oC, the seeds of population GR-1-BBGK-19,130 germinated at higher percentages (89%) than those of population GR-1-BBGK-19,97 (75%). The incubation temperature of 20oC generally reduced the seed germination percentages. In general, the seeds of population GR-1-BBGK-19,97 exhibited the lowest germination percentage (48.75% at 10o C), whereas the seeds of population GR-1-BBGK-19,124 germinated at higher percentages than those of population GR-1-BBGK-19,126 (Table 3).
3.2.2. Effects of light treatments
Regarding the light requirements for germination at 15°C of P. pinnata seeds, no statistically significant results were observed in terms of mean germination percentage (MGP) between seeds germinated under alternating light/dark and continuous dark conditions (Table 4). In both treatments, the germination percentages were high (86% in alternating light/dark conditions and 80% in continuous dark conditions). Furthermore, the seeds showed similar germination speed under different light conditions. Specifically, the mean germination time (MGT) was 18.59 days for seeds exposed to light/dark conditions and 20.05 days for seeds exposed to dark conditions (Table 4).
4. Discussion
In general, the development of seed propagation protocols for threatened species is necessary either for effective either ex situ or in situ conservation actions [3,36,37] or to enable the sustainable utilization of NUPs due to significant value in different economic sectors namely ornamental [3], agro-alimentary [16] and medicinal-cosmetic [15].
Although seed gemination of several Campanulaceae members has been previously investigated, the seed biology of this large family rather remains understudied [28]. Petromarula pinnata has been previously subjected to multi-species in vivo [27] and in vitro seed trials [28] with promising general results. The study herein, however, investigated for the first time the light- and temperature-dependance of P. pinnata seed germination facilitated by its GIS-derived ecological profiling, thus furnished insight into the most appropriate treatments in terms of temperature and light requirements.
In general, several members of the genus Campanula appear to have specific temperature preferences for their seed germination as suggested by previous studies [28]; this fact has also been confirmed in the current study dealing with P. pinnata showing a strong temperature effect (p = 0.0001) in its seed germination trials. The incubation temperature of 10 and 15oC presented the highest impact on seed germination of P. pinnata, except for population GR-1-BBGK-19,97 in which the highest percentage was observed at 15oC (75%). These temperature levels also improved the germination of Cretan local endemic members of genus Campanula, i.e., C. saxatilis L. subsp. saxatilis [23] and C. pelviformis [25]. The incubation temperature of 20oC resulted in relatively high percentages (>50%), while the temperature level of 25oC dramatically reduced germination percentages (<3%). However, previous studies have shown that the temperature of 25oC can benefit seed germination with appropriate pretreatment methods in many members of the Campanulaceae, such as Asyneuma chinense D.Y.Hong (88%), Cyananthus inflatus Hook f. & Thompson (85%) or Lobelia oahuensis Rock (95%) [28].
The study herein investigated for the first time the germination of P. pinnata seeds collected from lowland and semi-mountainous populations of Central and Eastern Crete, Greece. The examination of the population effect (p = 0.0001) investigated whether different altitudes in conjunction with four incubation temperatures can affect the seed germination behavior of different P. pinnata wild-growing populations from two semi-mountainous and two lowlands areas originating from Central and Eastern Crete. Although previous studies report that some Balkan endemic Campanula species may have quite different germination responses related to temperatures depending on the altitude of original collections [38], no significant variation was observed in the seed germination percentages of P. pinnata populations from different collection altitudes. In particular, the lowland populations had similar behavior, with the highest germination percentages at 10 and 15oC, while the same was true for the semi-mountainous populations, achieving their successful germination at 15oC. However, it is well known that such fluctuations may be expected across different populations of the same species [24]. Moreover, germination variations have been detected between populations originating from similar altitudes for C. pelviformis as shown in a recent study [25]. Undoubtedly, conducting experiments with freshly collected seeds (seeds with no storage) will lead to a more complete picture of the seed germination behavior and preferences of the focal species in concern, including the determination of after-ripening effect and possible dormancy type; therefore, further research is suggested with the aim among others to verify previously published in vivo seed germination results [27]. In fact, the latter outlines another research line which is aimed to follow shortly.
P. pinnata was shown herein to germinate without any light preference, showing 86% germination in alternating light/dark conditions and 80% in darkness at 15°C. Our results contrast with a previous study that observed different behavior of seeds of this species in constant light (76%) and constant darkness (3%) at 20°C, probably since the above-mentioned trial was conducted with fresh seeds at 20oC [28] and not with stored seeds at 15oC as the trial herein. However, it is worth noting that many members of the Campanulaceae family may exhibit light preferences as observed in other local Greek endemics of genus Campanula such as C. cretica (A.DC.) A.Dietr. (99% in light and 24% in darkness) and C. goulimyi Turrill (96% in light and 0% in darkness) [28]. In terms of seed dormancy, temperatures above 15oC are known to allow the embryo development in the seeds of several Campanulaceae members with morphological or morphopysiological dormancy [39].
The GIS-derived ecological profile of P. pinnata indicated the temperature limits and precipitation patterns of its wild-growing populations. When the ecological profile of P. pinnata was combined with the germination results obtained herein, it appeared that natural seed germination of P. pinnata probably takes place from the last quarter of the year with prevailing mean temperatures equaling to 17.98, 14.09 and 11.05oC during October, November, and December, respectively. During this period, temperatures were shown to reach 10 and 15oC in the natural habitats of this species in Crete and, simultaneously, moisture levels in these periods were shown to increase due to the augmented precipitation patterns, starting with 73.16 mm in October and peaking at 142.40 mm in January (Figure 2). Most probably, the appropriate temperatures of April (14.05oC) combined to decreased precipitation (37.77 mm) in early spring may not favor seed germination (Figure 2).
The knowledge furnished herein for this valuable, yet vulnerable local endemic plant of Crete can be exploited consecutively either for conservation purposes or to facilitate the sustainable exploitation of P. pinnata due to its ornamental value [3] and agro-alimentary interest [16] associated also with medicinal value [12]. Notably, this species is traditionally used across Crete as an ingredient of the widely consumed ‘kaltsounia’ in western Crete or ‘chortopitakia’ in central and eastern Crete of the local Mediterranean (Cretan) diet, i.e., the local savory staffed dough with wild-sourced and cultivated greens associated with a strong antioxidant potential [11]. To this end, a newly launched research project will readily apply this knowledge to produce enough plant material for a pilot cultivation to be established in the next growing season in its place of origin, namely Crete (Greece).
5. Conclusions
The development of seed propagation protocols for threatened species is necessary for effective ex situ conservation and enables their sustainable utilization. The investigation herein furnished for the first time a detailed profiling of the non-biotic environmental conditions experienced evolutionary by P. pinnata wild-growing populations based on its natural distribution occurrences in rocky habitats of Crete, Greece. The GIS ecological profile developed herein may facilitate the successful ex situ conservation and further cultivation of this vulnerable local endemic species, also offering guidance for its adaptation in man-made settings for conservation or sustainable exploitation purposes. The germination trials performed in this investigation using seeds sourced from four wild-growing populations revealed no significant variation in seed germination but identified the most appropriate incubation temperature for the effective germination of this species. These results can pave the way for the successful ex situ conservation of the natural species diversity in seed banks and may further serve the current attempts undertaken for the sustainable exploitation of this valuable, yet vulnerable plant with ornamental, medicinal and agro-alimentary potential.
Author Contributions
Conceptualization, S.H., N.K., Ι.A., and G.T.; data curation, N.K., I.A., S.K., E.P., E.D., and G.T.; formal analysis, I.A., E.P., N.K., S.K., and S.H.; investigation, I.A., N.K., S.H., E.D., E.P., E.K., S.K., and G.T.; methodology, I.A., N.K., S.H., E.P., S.K., E.D., E.K., V.P., V.G., and G.T.; project administration, S.H., N.K. and G.T.; resources, N.K., G.T., S.H., V.G., and S.K.; software, I.A.; supervision, N.K., S.H., and G.T.; validation, N.K., I.A., E.K., E.D., V.G., and G.T.; visualization, I.A., E.P., V.G., and N.K.; writing—original draft, I.A., E.P., S.H., N.K., E.K., and G.T.; writing—review and editing, all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All data supporting the results of this study are included in the manuscript and the datasets are available upon request.
Acknowledgments
This investigation was performed as part of the PhD of Ioannis Anestis and in support of the project “Indigenous edible plants of Crete as alternative new crops contributing to biodiversity preservation, protection from soil degradation and mitigation of climate change impacts” (acronym: Cretan Greens 4 Clima Pro, Μ16ΣΥΝ-01106).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Seglie, L.; Scariot, V.; Larcher, F.; Devecchi, M.; Chiavazza, P.M. In vitro seed germination and seedling propagation in Campanula spp. Plant Biosyst. 2011, 146, 15–23. [Google Scholar] [CrossRef]
- Scariot, V.; Seglie, L.; Caser, M.; Devecchi, M. Evaluation of ethylene sensitivity and postharvest treatments to improve the vase life of four Campanula species. Eur. J. Horticult. Sci. 2008, 73, 166–170. [Google Scholar]
- Krigas, N.; Tsoktouridis, G.; Anestis, I.; Khabbach, A.; Libiad, M.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Lamchouri, F.; Tsiripidis, I.; Tsiafouli, M.A.; El Haissoufi, M.; Bourgou, S. Exploring the potential of neglected local endemic plants of three Mediterranean regions in the ornamental sector: Value chain feasibility and readiness timescale for their sustainable exploitation. Sustainability 2021, 13, 2539. [Google Scholar] [CrossRef]
- Cellinese, N.; Smith, S.A.; Edwards, E.J.; Kim, S.T.; Haberle, R.C.; Avramakis, M.; Donoghue, M.J. Historical biogeography of the endemic Campanulaceae of Crete. J. Biogeogr. 2009, 36, 1253–1269. [Google Scholar] [CrossRef]
- Strid, A. Atlas of the Aegean Flora Part 1 (Text & Plates) & Part 2 (Maps), 1st ed.; Englera 33 (1 & 2); Botanic Garden and Botanical Museum: Berlin, Germany; Freie Universität: Berlin, Germany, 2016; ISSN 0170-4818. [Google Scholar]
- González-Tejero, M.R.; Casares-Porcel, M.; Sánchez-Rojas, C.P.; Ramiro-Gutiérrez, J.M.; Molero-Mesa, J.; Pieroni, A.; Giusti, M.E.; Censorii, E.; de Pasquale, C.; Della, A.; et al. Medicinal plants in the Mediterranean area: Synthesis of the results of the project Rubia. J. Ethnopharmacol. 2008, 116, 341–357. [Google Scholar] [CrossRef]
- Lim, T.K. Edible Medicinal and Non-Medicinal Plants; Springer International Publishing: Gewerbestr, Switzerland, 2016; ISBN 9789401772761. [Google Scholar]
- Heinrich, M.; Müller, W.E.; Galli, C. Local Mediterranean Food Plants and Nutraceuticals; Karger Medical and Scientific Publishers: Basel, Switzerland, 2006; Volume 59, ISSN 1660-0347. [Google Scholar]
- Hadjichambis, A.CH.; Hadjichambi, D.P.; Della, A.; Giusti, M. E.; De Pasquale, C.; Lenzarini, Censorii, E.; Reyes Gonzales Tejero, M.; Sanchez-Rojas, C.P.; Ramiro Gutierrez, J.M; Skoula, M.; Johnson, Sarpaki, A.; Hmamouchi, M.; Jorhi, S.; El-Demerdash, M.; El-Zayat, M.; Pieroni, A. Wild and semi-domesticated food plant consumption in seven circum-Mediterranean areas. Int. J. Food Sci. Nutr. 2008, 59, 383–414. [CrossRef]
- Tsiftsoglou, O.S.; Lagogiannis, G.; Psaroudaki, A.; Vantsioti, A.; Mitić, M.N.; Mrmošanin, J.M.; Lazari, D. Phytochemical analysis of the aerial parts of Campanula pelviformis Lam. (Campanulaceae): Documenting the dietary value of a local endemic plant of Crete (Greece) traditionally used as wild edible green. Sustainability 2023, 15, 7404. [Google Scholar] [CrossRef]
- Vasilopoulou, E.; Trichopoulou, A. Green pies: The flavonoid rich Greek snack. Food Chem. 2011, 126, 855–858. [Google Scholar] [CrossRef]
- Dimitriadis, K.M.; Karavergou, S.; Hadjipavlou-Litina, D.; Krigas, N.; Lazari, D. Phytochemical and antioxidant evaluation of the ex-situ cultivated species Petromarula pinnata (L.) A. DC. and Campanula cretica (A.DC.) Dietr. (Campanulaceae) from Crete (Greece). Planta Medica 2022, 88, 1523. [Google Scholar]
- Kalpoutzakis, E.; Chatzimitakos, T.; Athanasiadis, V.; Mitakou, S.; Aligiannis, N.; Bozinou, E.; Gortzi, O.; Skaltsounis, L.A.; Lalas, S.I. Determination of the total phenolics content and antioxidant activity of extracts from parts of plants from the Greek Island of Crete. Plants 2023, 12, 1092. [Google Scholar] [CrossRef]
- Padulosi, S.; Thompson, J.; Rudebjer, P. Fighting Poverty, Hunger and Malnutrition with Neglected and Underutilized Species (NUS): Needs, Challenges and the Way Forward; Biodiversity International: Rome, Italy, 2013. [Google Scholar]
- Bourgou, S.; Ben Haj Jilani, I.; Karous, O.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; Greveniotis, V.; Avramakis, M.; Hatzilazarou, S.; Anestis, I.; Tsoktouridis, G.; Krigas, N. Medicinal-cosmetic potential of the local endemic plants of Crete (Greece), Northern Morocco and Tunisia: Priorities for conservation and sustainable exploitation of neglected and underutilized phytogenetic resources. Biology 2021, 10, 1344. [Google Scholar] [CrossRef] [PubMed]
- Libiad, M.; Khabbach, A.; El Haissoufi, M.; Anestis, I.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Greveniotis, V.; Tsiripidis, I.; Dariotis, E; Tsiafouli, M.A.; Krigas, N. Agro-alimentary potential of the neglected and underutilized local endemic plants of Crete (Greece), Rif-Mediterranean coast of Morocco and Tunisia: Perspectives and challenges. Plants 2021, 10, 1770. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, B. Practical Woody Plant Propagation for Nursery Growers; Timber Press Inc.: Portland, OR, USA, 2006; p. 669. ISBN 9780881920628. [Google Scholar]
- Krigas, N.; Mouflis, G.; Grigoriadou, K. Conservation of important plants from the Ionian Islands at the Balkan Botanic Garden of Kroussia, N Greece: Using GIS to link the in situ collection data with plant propagation and ex situ cultivation. Biodivers. Conserv. 2010, 19, 3583–3603. [Google Scholar] [CrossRef]
- Krigas, N.; Lykas, C.; Ipsilantis, I.; Matsi, T.; Weststrand, S.; Havström, M.; Tsoktouridis, G. Greek tulips: Worldwide electronic trade over the internet, global ex situ conservation and current sustainable exploitation challenges. Plants 2021, 10, 580. [Google Scholar] [CrossRef] [PubMed]
- Hatzilazarou, S.; El Haissoufi, M.; Pipinis, E.; Kostas, S.; Libiad, M.; Khabbach, A.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Aslanidou, V.; Greveniotis, V.; Sakelariou, M.A.; Anestis, I.; Tsoktouridis, G.; Krigas, N. GIS-Facilitated seed germination and multifaceted evaluation of the Endangered Abies marocana Trab. (Pinaceae) enabling conservation and sustainable exploitation. Plants 2021, 10, 2606. [Google Scholar] [CrossRef] [PubMed]
- Kostas, S.; Hatzilazarou, S.; Pipinis, E.; Bourgou, S.; Ben Haj Jilani, I.; Ben Othman, W.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; Koundourakis, E.; Greveniotis, V.; Papaioannou, E.; Sakellariou, M.A.; Anestis, I.; Tsoktouridis, G.; Krigas, N. DNA Barcoding, GIS-Facilitated seed germination and pilot cultivation of Teucrium luteum subsp. gabesianum (Lamiaceae), a Tunisian local endemic with potential medicinal and ornamental value. Biology 2022, 11, 462. [Google Scholar]
- Pipinis, E.; Hatzilazarou, S.; Kostas, S.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; et al. Facilitating conservation and bridging gaps for the sustainable exploitation of the Tunisian local endemic plant Marrubium aschersonii (Lamiaceae). Sustainability 2022, 14, 1637. [Google Scholar] [CrossRef]
- Hatzilazarou, S.; Kostas, S.; Pipinis, E.; Anestis, I.; Papaioannou, E.; Aslanidou, V.; Tsoulpha, P.; Avramakis, M.; Krigas, N.; Tsoktouridis, G. GIS-Facilitated seed germination, fertilization effects on growth, nutrient and phenol contents and antioxidant potential in three local endemic plants of Crete (Greece) with economic interest: Implications for conservation and sustainable exploitation. Horticulturae 2023, 9, 335. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014; ISBN 978-0-12-416677-6. [Google Scholar]
- Anestis, I.; Pipinis, E.; Kostas, S.; Papaioannou, E.; Karapatzak, E.; Dariotis, E.; Tsoulpha, P.; Koundourakis, E.; Chatzileontari, E.; Tsoktouridis, G.; Hatzilazarou, S.; Krigas, N. GIS-facilitated germination of stored seeds from five wild-growing populations of Campanula pelviformis Lam. and fertilization effects on growth, nutrients, phenol content and antioxidant potential. Horticulturae 2023, 9, 877. [Google Scholar] [CrossRef]
- O’Reill, C.; Arnott, J.T.; Owens, J.N. Effects of photoperiod and moisture availability on shoot growth, seedling morphology, and cuticle and epicuticular wax features of container-grown western hemlock seedlings. Can. J. Forest Res. 1989, 19, 122–131. [Google Scholar] [CrossRef]
- Sarropoulou, V.; Krigas, N.; Tsoktouridis, G.; Maloupa, E.; Grigoriadou, K. Seed Germination Trials and Ex Situ Conservation of Local Prioritized Endemic Plants of Crete (Greece) with Commercial Interest. Seeds 2022, 1, 279–302. [Google Scholar] [CrossRef]
- Koutsovoulou, K.; Daws, M.I.; Thanos, C.A. Campanulaceae: A family with small seeds that require light for germination. Ann. Bot. 2014, 113, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Fanourakis, D.; Paschalidis, K.; Tsaniklidis, G.; Tzanakakis, V.A.; Bilias, F.; Samara, E.; Liapaki, E.; Jouini, M.; Ipsilantis, I.; Maloupa, E.; Tsoktouridis, G.; Matsi, T.; Krigas, N. Pilot cultivation of the local endemic Cretan marjoram Origanum microphyllum (Benth.) Vogel (Lamiaceae): Effect of fertilizers on growth and herbal quality features. Agronomy 2021, 12, 94. [Google Scholar] [CrossRef]
- Paschalidis, K.; Fanourakis, D.; Tsaniklidis, G.; Tzanakakis, V.A.; Bilias, F.; Samara, E.; Kalogiannakis, K.; Debouba, F.J.; Ipsilantis, I.; Tsoktouridis, G.; et al. Pilot cultivation of the Vulnerable Cretan endemic Verbascum arcturus L. (Scrophulariaceae): Effect of fertilization on growth and quality features. Sustainability 2021, 13, 14030. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Strid, A.; Dimopoulos, P. Extinction risk assessment of the Greek endemic flora. Biology 2021, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Lazarina, M.; Kallimanis, A.S.; Dimopoulos, P.; Psaralexi, M.; Michailidou, D.E.; Sgardelis, S.P. Patterns and drivers of species richness and turnover of neo-endemic and palaeo-endemic vascular plants in a Mediterranean hotspot: The case of Crete, Greece. J. Biol. Res.-Thessaloniki 2019, 26, 12. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures in Agricultural Research, 2nd ed.; Wiley: New York, NY, USA, 1984; ISBN 0-471-87092-7. [Google Scholar]
- Snedecor, G.W.; Cochran, W.C. Statistical Methods., 7th ed.; The Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- Klockars, A.; Sax, G. Multiple Comparisons; Sage Publications: Newbury Park, CA, USA, 1986; p. 87. ISBN 0-8039-2051-2. [Google Scholar]
- Maunder, M.; Higgens, S.; Culham, A. The effectiveness of botanic garden collections in supporting plant conservation: A European case study. Biodivers. Conserv. 2001, 10, 383–401. [Google Scholar] [CrossRef]
- Bunn, E.; Turner, S.R.; Dixon, K.W. Biotechnology for saving rare and threatened flora in a biodiversity hotspot. In Vitro Cell Dev. Biol. Plant 2011, 47, 188–200. [Google Scholar] [CrossRef]
- Blionis, G.; Vokou, D. Reproductive attributes of Campanula populations from Mt Olympos, Greece. Plant Ecol. 2005, 178, 77–88. [Google Scholar] [CrossRef]
- Baskin, C. C.; Baskin, J. M.; Yoshinaga, A.; Wolkis, D. Seed dormancy in Campanulaceae: Morphological and morphophysiological dormancy in six species of Hawaiian lobelioids. Botany 2020, 98(6), 327–332. [Google Scholar] [CrossRef]
Figure 1.
Wild-growing individual of Petromarula pinnata in full flowering during spring in its natural rocky habitat in Crete (A), bearing ripe fruits during summer months (B) with mature seeds (C) able to germinate upon evident radicle protrusion (D).
Figure 1.
Wild-growing individual of Petromarula pinnata in full flowering during spring in its natural rocky habitat in Crete (A), bearing ripe fruits during summer months (B) with mature seeds (C) able to germinate upon evident radicle protrusion (D).

Figure 2.
Ecological profile across the natural distribution range of Petromarula pinnata wild-growing populations in Crete (n = 51) linked in GIS with geodatabases (WorldClim version 2.1), providing values for: (A) minimum temperatures per month (°C), (B) maximum temperatures per month (°C), (C) average temperatures per month (°C), (D) precipitation per month (mm), and (E) calculated values for 19 bioclimatic variables. For (A–E) minimum, maximum, average, and standard deviation is shown based on data from 1970–2000. The colors of the plotted lines illustrate the minimum (blue), maximum (orange), and mean (grey) monthly values for temperature (°C) and precipitation (mm).
Figure 2.
Ecological profile across the natural distribution range of Petromarula pinnata wild-growing populations in Crete (n = 51) linked in GIS with geodatabases (WorldClim version 2.1), providing values for: (A) minimum temperatures per month (°C), (B) maximum temperatures per month (°C), (C) average temperatures per month (°C), (D) precipitation per month (mm), and (E) calculated values for 19 bioclimatic variables. For (A–E) minimum, maximum, average, and standard deviation is shown based on data from 1970–2000. The colors of the plotted lines illustrate the minimum (blue), maximum (orange), and mean (grey) monthly values for temperature (°C) and precipitation (mm).
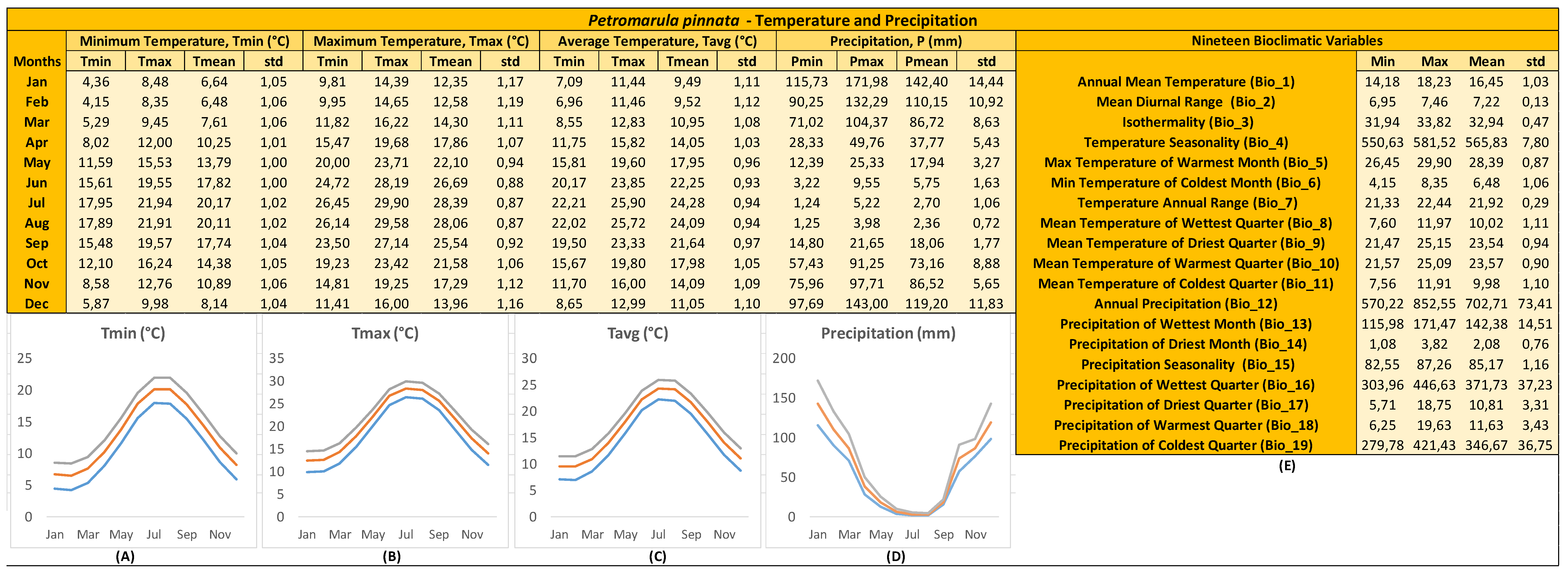
Figure 3.
Effect of population on germination percentage (standard deviation) of Petromarula pinnata seeds regardless the incubation temperature. Columns accompanied by the same letter do not differ significantly. The comparisons were made using LSD test.
Figure 3.
Effect of population on germination percentage (standard deviation) of Petromarula pinnata seeds regardless the incubation temperature. Columns accompanied by the same letter do not differ significantly. The comparisons were made using LSD test.
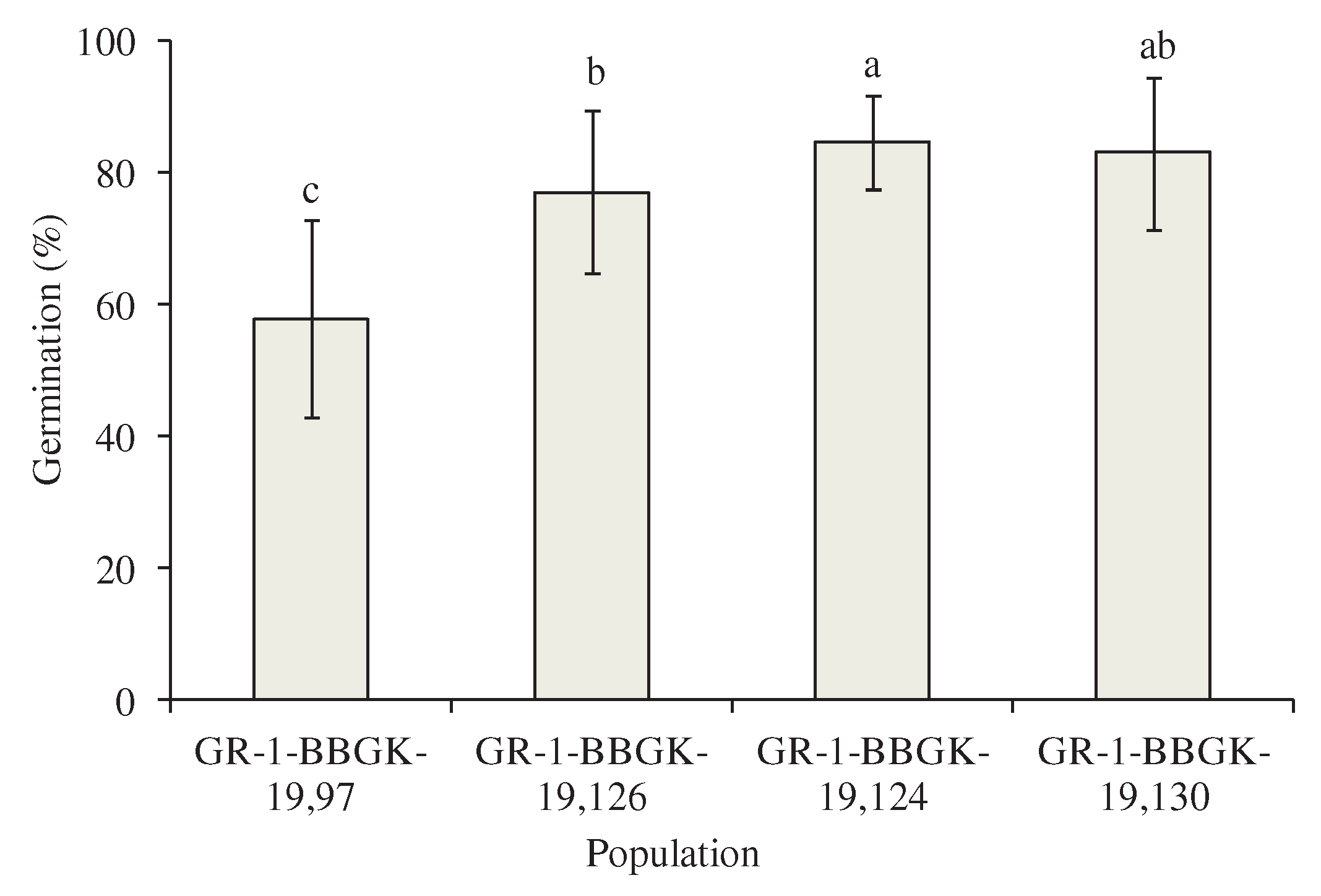
Figure 4.
Effect of incubation temperature on germination percentage (standard deviation) of Petromarula pinnata seeds of four wild-growing populations. Columns accompanied by the same letter do not differ significantly. The comparisons were made using LSD test.
Figure 4.
Effect of incubation temperature on germination percentage (standard deviation) of Petromarula pinnata seeds of four wild-growing populations. Columns accompanied by the same letter do not differ significantly. The comparisons were made using LSD test.
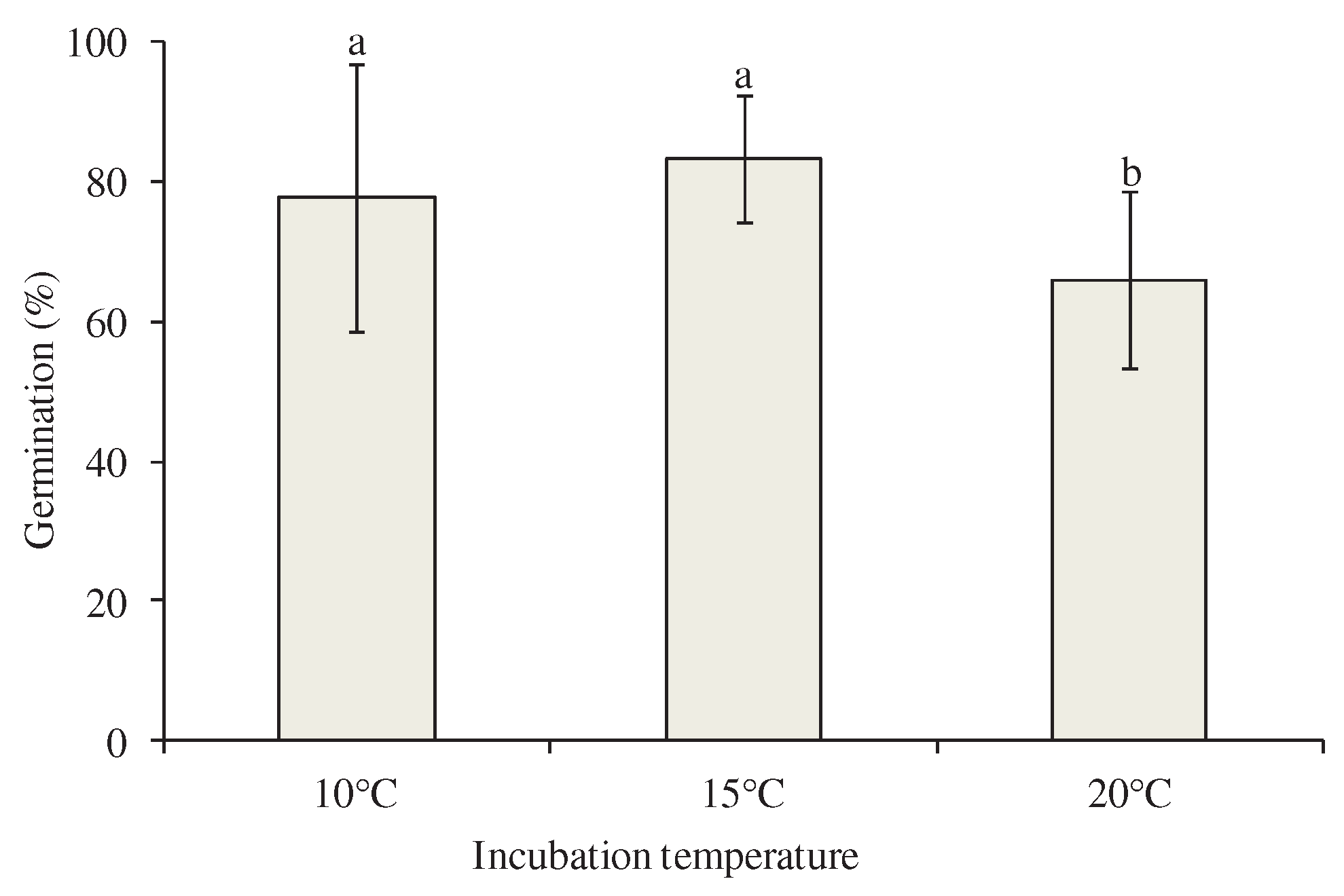
Figure 5.
Cumulative germination percentage diagrams (± standard deviation) of Petromarula pinnata seeds sourced from four populations incubated at 10 (●), 15 (■) and 20 (□). 1In each population, means are statistically different at p≤0.05, when they do not share a common letter. The comparisons were made using the LSD test (n=4).
Figure 5.
Cumulative germination percentage diagrams (± standard deviation) of Petromarula pinnata seeds sourced from four populations incubated at 10 (●), 15 (■) and 20 (□). 1In each population, means are statistically different at p≤0.05, when they do not share a common letter. The comparisons were made using the LSD test (n=4).
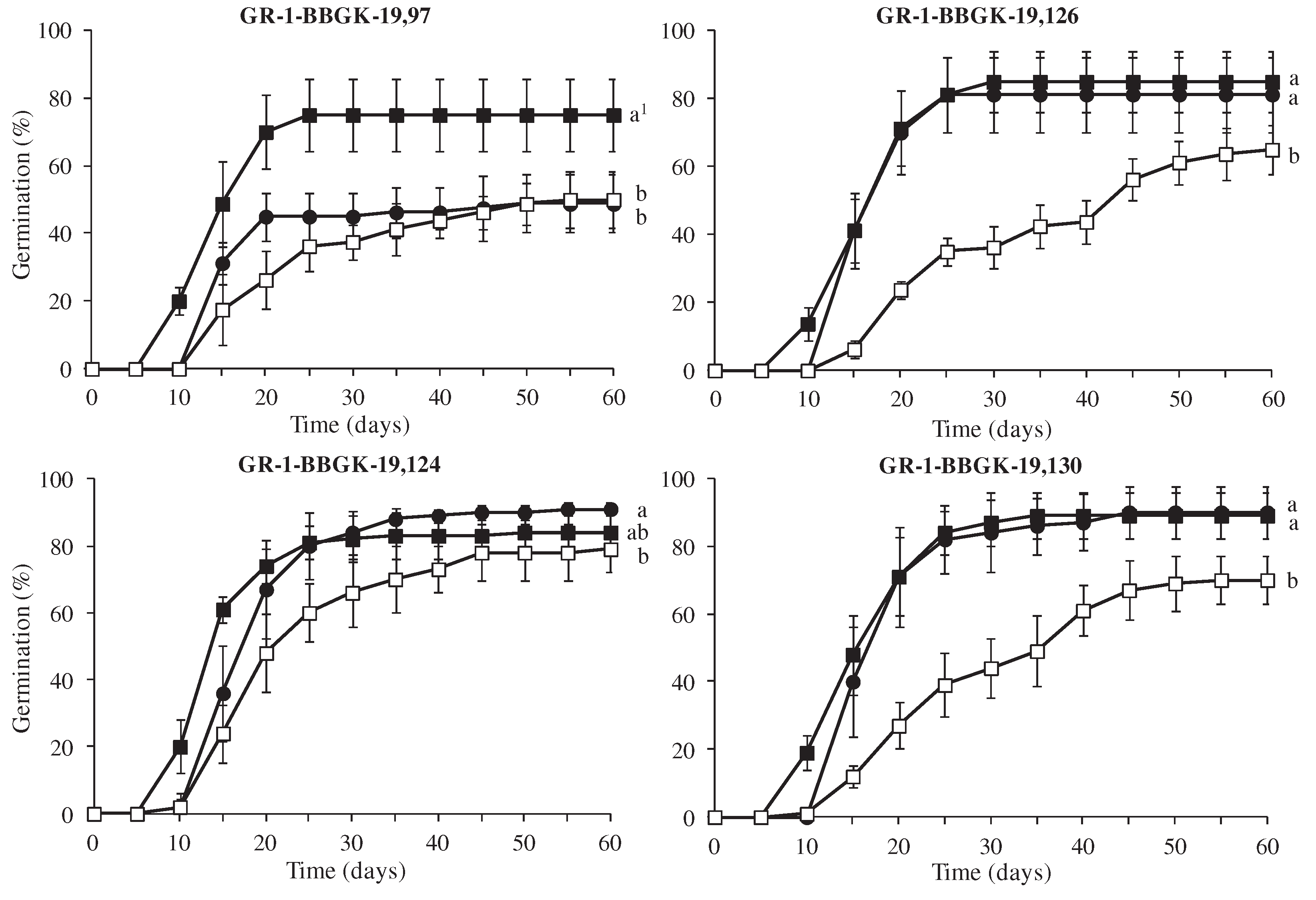

Table 1.
Collection data with geographical information and IPEN (International Plant Exchange Network) accession numbers of the investigated Petromarula pinnata wild-growing populations.
Table 1.
Collection data with geographical information and IPEN (International Plant Exchange Network) accession numbers of the investigated Petromarula pinnata wild-growing populations.
| IPEN Accession number | Collection Site | Latitude (North) | Longitude (East) | Altitude (m) |
|---|---|---|---|---|
| GR-BBGK-1-19,97 | Viannos, Heraklion | 35.0471 | 25.4174 | 632 |
| GR-BBGK-1-19,126 | Agios Georgios Selinaris, Lasithi | 35.2753 | 25.5549 | 225 |
| GR-BBGK-1-19,124 | Agia Eirini gorge, Heraklion | 35.2844 | 25.1653 | 140 |
| GR-BBGK-1-19,130 | Tylissos gorge, Heraklion | 35.2874 | 24.9716 | 395 |
Table 2.
Significance of separate factors (population, temperature) and their interaction on germination percentage of Petromarula pinnata seeds from different wild-growing populations as estimated by ANOVA.
Table 2.
Significance of separate factors (population, temperature) and their interaction on germination percentage of Petromarula pinnata seeds from different wild-growing populations as estimated by ANOVA.
| Source | Sum of Squares | df | Mean Square | F | Sig. |
|---|---|---|---|---|---|
| Population | 2314.13 | 3 | 771.38 | 20.36 | 0.000 |
| Temperature | 1240.85 | 2 | 620.43 | 16.38 | 0.000 |
| `Population × Temperature | 701.35 | 6 | 116.89 | 3.09 | 0.015 |
Table 3.
Effect of three incubation temperatures on the germination percentage of four wild-growing populations of Petromarula pinnata. Means ± standard deviation values are given.
Table 3.
Effect of three incubation temperatures on the germination percentage of four wild-growing populations of Petromarula pinnata. Means ± standard deviation values are given.
| Area | Population | Incubation temperature | ||
|---|---|---|---|---|
| 10°C | 15°C | 20°C | ||
| Semi-mountainous | GR-1-BBGK-19,97 | 48.75 b 1 ± 8.54 | 75.00 b ± 10.80 | 50.00 c ± 8.16 |
| GR-1-BBGK-19,130 | 90.00 a ± 7.66 | 89.00 a ± 6.83 | 70.00 ab ± 6.93 | |
| Lowland | GR-1-BBGK-19,126 | 81.25 a ± 11.09 | 85.00 ab ± 9.13 | 65.00 b ± 7.07 |
| GR-1-BBGK-19,124 | 91.00 a ± 2.00 | 84.00 ab ± 5.66 | 79.00 a ± 6.83 | |
1Values in the same column followed by the same letter are not significantly different (p≤0.05) according to LSD test (n=4).
Table 4.
Effect of light conditions on seed germination of Petromarula pinata. Means and standard deviation values are provided.
Table 4.
Effect of light conditions on seed germination of Petromarula pinata. Means and standard deviation values are provided.
| Germination percentage (%) | Mean Germination Time (Days) | |
|---|---|---|
| Light/dark | 86.00 ± 6.93 a 1 | 18.59 a ± 1.02 a |
| Dark | 80.00 ± 8.64 a | 20.06 a ± 1.72 a |
1In each column, the means are statistically different at p≤0.05 (n=4) when they do not share a common small letter. The comparisons were made using T-test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



