
Submitted:
07 July 2023
Posted:
10 July 2023
You are already at the latest version
Abstract
While human activities and climate change are threatening available natural ecosystems, targeted studies on the biological cycle, propagation, and cultivation of threatened species are crucial both for their conservation and sustainable exploitation. This study was designed to bridge extant research gaps regarding the vulnerable local endemic Campanula pelviformis, a wild edible green traditionally consumed in Crete (Greece) with agro-alimentary and medicinal interest as well as ornamental value. New knowledge was obtained on the abiotic environmental conditions prevailing in its wild habitats (natural distribution mapped in Geographical Information Systems and linked with open-source databases). We collected seeds from wild-growing populations (three lowland, two semi-mountainous), we tested their germination at four different temperatures (10, 15, 20 and 25oC) and under different light conditions (light/dark and darkness), and we performed fertilization trails [integrated nutrient management (INF), chemical fertilization (ChF), control] examining morphological and physiological characteristics, above- and below-ground macro- and micronutrients contents, as well as their phenol content and antioxidant capacity. There were population and temperature effects on seed germination and their interaction was statistically significant. Campanula pelviformis germinated better at 10 and 15 oC (>85% for all populations) with no preference on light conditions (98.75% and 95% in light and dark conditions). The INF application increased root dry mass, chlorophyll content index and chlorophyll fluorescence compared to other treatments and was beneficial for macro- and micronutrients concentrations in above-ground parts compared to previously studied wild-growing material, while below-ground parts were positively impacted by both fertilization types. Total phenols and antioxidant capacity were both increased by ChF fertilization. The data furnished herein led to the re-evaluation and upgrade of the feasibility and the readiness timescale for its sustainable exploitation in different economic sectors.
Keywords:
Endemic plants
; GIS
; integrated nutrient management
; macronutrients
; medicinal interest
; micronutrients
; neglected and underutilized species
; ornamental value
; phytogenetic resources
; sustainable exploitation
; wild edible greens
1. Introduction
The members of the genus Campanula (Campanulaceae) are widely recognized for their ornamental value and many species are frequently used as ornamental plants around the world [1]. In fact, C. glomerata L., C. medium L., C. cochleariifolia Lam. are used as garden plants; C. isophylla Moretti, C. carpatica Jacq., C. rotundifolia L., C. pyramidalis L. are used as pot plants and C. medium, C. persicifolia L., C. pyramidalis, C. glomerata are used for cut flowers. To date, there is an ongoing quest for new Campanula species with interesting or promising ornamental potential in this sector [1,2].
Apart from the horticultural value, species within the genus Campanula are also being exploited for agro-alimentary purposes in different regions [3]. For example, local Spanish residents use raw roots of C. rapunculus L. in salads for their nutritional benefits as well as Italians collect the same plant using its raw roots or aerial parts in salads [3,4]. Furthermore, Campanula pelviformis Lam. has been reported as a wild green traditionally consumed as food in the eastern part of Crete [5,6]. In fact, the phytochemical profile of C. pelviformis has been recently analyzed, and several isolated compounds have been found to possess pharmaceutical properties, thus rendering it as a new medicinal plant [6]. These findings render C. pelviformis as a trustworthy source of diverse phytonutrients and bioactive compounds [6].
The genus Campanula comprises 448 taxa (species and subspecies) worldwide (https://powo.science.kew.org/, accessed on 19 April 2023), most of which are perennials (occasionally annuals or biennials) often thriving in rocky areas, slopes, or grounds; many Campanula members are range-restricted or endemic plants in different regions, thus associated with high biological value [7]. In Greece, there are 95 wild-growing Campanula taxa and most of them are exclusively found in crevices of limestone cliffs growing as chasmophytes [8]. Of these, 11 taxa are to be found in Crete, with 8 being local single-island endemics [8,9]. C. pelviformis is included as a local endemic plant in the extraordinary plant diversity of Crete [9], which involves numerous single-island endemic taxa that are still neglected and underutilized plant species (NUPs) [2,10,11,12]. Previous studies have shown that the exploitation of such unique phytogenetic resources holds great potential for different economic sectors such as in agriculture, medicinal-cosmetics, and ornamental horticulture [1,2,11,12,13]. Furthermore, conservation efforts have been increased to date to achieve effective protection for local endemic NUPs, many of which are threatened with extinction [2]. Significant information on both the economic and biological value of threatened local endemic NUPs has been gathered through species-specific studies [2,11,12,13], and basic research gaps related to their propagation and ex-situ cultivation protocols have been addressed during the last decade [2,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29].
Applied research to develop appropriate propagation methods for local endemic NUPs aiming to re-enforce threatened populations in the wild has primarily been driven by their appreciated biological rarity and priceless ecological value [2,10]; however, this has also been often coupled by potential economic benefits associated with their sustainable utilization and concomitant exploitation [2,11,12]. The effectiveness of these two research directions can be facilitated by information on the environmental conditions associated with targeted NUPs reflecting their adaptation and in natural habitats and ecological preferences [2,15,16,17,18,19,20,22,23,24,25,26,28]. Such data can be utilized to adjust or improve their propagation conditions, for example, through seed propagation [30] which is the most economical and efficient propagation method for many plant species [31]. Furthermore, for the ex-situ conservation of targeted NUPs, it is crucial to achieve successful ex-situ cultivation and production of stock-plants, particularly for threatened species originating from the challenging terrain of the Mediterranean basin [2,32]. Various cultural practices have been employed in the past with reasonable ex-situ cultivation and acclimatization results in terms of morphological and physiological traits, thus leading to the conclusion that ex-situ cultivation can contribute significantly to the production of high-quality plants [33]. The application of fertilizers during the propagation stage in the nursery has been shown to increase the nutrient reserves in propagated plants for population restoration efforts, thus rendering fertilization as an efficient method leading to enhanced establishment success in their natural habitat [34].
Species-wise, this study is focused on Campanula pelviformis, a vulnerable [35] and range-restricted local endemic plant of eastern Crete which is protected by the Greek Presidential Decree 67/1981 although it is traditionally consumed as wild edible green [6], thus meriting conservation concern. Given that many species from genus Campanula are already used for ornamental and ethnobotanical purposes [1,3,4,6], C. pelviformis also takes advantage for its sustainable exploitation since high demand of such unique resources may lead to excessive collection in the wild risking the degradation of genetic pools and local biodiversity. Context-wise, this study was designed to bridge extant research gaps regarding the studied focal species. The objectives of this research were to: (i) Determine the optimal temperature and light conditions required for the successful seed germination of C. pelviformis, examining seed responses of different wild-growing populations from low and semi-mountainous areas of Crete; (ii) Identify the preferred ecological conditions of C. pelviformis based on its natural distribution; (iii) Compare the impact chemical fertilizers and integrated nutrient management on the growth of seedlings and estimate their nutrient content; (iv) Assess the overall phenolic content and antioxidant capacity of its aerial parts; and in the light of all the aforementioned, (v) Reassess the feasibility and readiness for the sustainable utilization of C. pelviformis in different economic sectors.
2. Materials and Methods
2.1. Plant Characteristics
Campanula pelviformis (Figure 1) is a hemicryptophyte with restricted range occurring only in the eastern part of the island of Crete, Greece [8]. This local endemic species thrives in rocky slopes and ledges, stony dry places or walls in villages and rocky road embankments, spanning an elevation range from 0 m to 850 m above sea level [8]. Currently, it is considered as Vulnerable taxon [35]. In Greece, it is considered as a protected plant species due to its inclusion in the Greek Presidential Decree 67/1981 (https://portal.cybertaxonomy.org/flora-greece/cdm_dataportal/taxon/332301ee-177e-4f9f-95b9-62fcea5e9714, accessed 29 June 2023).
2.2. GIS Ecological Profiling
The natural distribution points of C. pelviformis were obtained from a previously published study [36]. To create the ecological profile for this species, historical climate and precipitation data with a pixel size 30 sec were downloaded from the WorldClim database (https://www.worldclim.org/data/worldclim21.html, accessed 15 June 2023). The data encompassed parameters such as minimum, maximum, and average temperatures and precipitation, as well as 19 bioclimatic variables i.e., Annual Mean Temperature (Bio_1), Mean Diurnal Range (Bio_2), Isothermality (Bio_3), Temperature Seasonality (Bio_4), Max Temperature of Warmest Month (Bio_5), Min Temperature of Coldest Month (Bio_6), Temperature Annual Range (Bio_7), Mean Temperature of Wettest Quarter (Bio_8), Mean Temperature of Driest Quarter (Bio_9), Mean Temperature of Warmest Quarter (Bio_10), Mean Temperature of Coldest Quarter (Bio_11), Annual Precipitation (Bio_12), Precipitation of Wettest Month (Bio_13), Precipitation of Driest Month (Bio_14), Precipitation Seasonality (Bio_15),Precipitation of Wettest Quarter (Bio_16), Precipitation of Driest Quarter (Bio_17), Precipitation of Warmest Quarter (Bio_18), and Precipitation of Coldest Quarter (Bio_19). As a result, the following layers were imported in the GIS software:
(a) A raster file including 16 natural distribution points of C. pelviformis and
(b) WorldClim version 2.1 (https://www.worldclim.org/data/worldclim21.html, accessed 15 June 2023), with minimum, maximum, and average temperatures (°C), precipitation values (mm) and data for 19 bioclimatic variables for every month derived from 1970–2000, with a spatial resolution of 1 km2.
2.3. Seed Collection and Storage
Mature seeds from approximately 5-10 wild-growing individuals of five wild-growing populations thriving in different altitudes were collected by hand during the summer of 2019 and the summer of 2020, prior dispersal (Table 1). The seeds were collected using special permission (182336/879 of 16 May 2019 and 64886/2959 of 6 July 2020) issued by the national competent authority, namely the Greek Ministry of Environment and Energy. After collection, the seeds were manually cleaned, dry-stored in glass containers at 3–5oC before experimentation.
2.4. Germination Tests
The germination experiments were conducted in April 2022 at the Laboratory of Floriculture, School of Agriculture, Aristotle University of Thessaloniki (Thermi, Greece). The experiments included five distinct populations from various altitudes and evaluated their seed responses to four constant temperatures of 10, 15, 20 and 25oC within four different temperature-controlled growth chambers. Each temperature treatment consisted of four replications of 20 seeds. The experiment employed 9 cm sterile plastic Petri dishes with seeds allowed to germinate on two filter papers moistened with distilled water. The Petri dishes were arranged randomly on the shelves of the growth chambers, under a 12 h light/12 h dark photoperiod. The filter papers were kept moist as required throughout the whole experimental period. The germinated seeds were counted and removed every five days throughout a period of 45 days. An individual seed was considered germinated upon the emergence of radicle protrusion from the seed coat.
Furthermore, after completion of the temperature-related germination experiments, new experiments were carried out to investigate the light requirements for C. pelviformis to germinate. The population GR-BBGK-1-19,1194 with the highest gemination percentages and the temperature of 10oC for seed germination was selected for these experiments. The same germination protocol was followed, and Petri dishes were chambered under alternating light/dark (12 h light/ 12 h dark photoperiod) and continuous dark conditions. For the treatment in the dark, each Petri dish was wrapped with aluminum foil. For each treatment (light/dark, dark), the mean germination time (MGT) was calculated as the average of four replications. The MGT was calculated for each replication per treatment according to the following equation:
where n is the number of seeds which germinated on day D, and D is the number of days counted from the beginning of the test [37].
MGT = Σ(Dn) / Σn
2.5. Plant Production
Seeds of C. pelviformis GR-BBGK-1-19,1056 (Table 1) were used for a pilot germination experiment. The experiment was conducted in December 2020 and the germinated seeds were used for plant production. Specifically, the germinated seeds were sown in plastic pots (6 x 6 x 6.5 cm) filled with a 3:1 ratio (v/v) substrate of enriched peat (TS2, Klassmann) and perlite which were carefully covered with sand before the containers were placed on a greenhouse bench. Irrigation was applied periodically to maintain the proper moisture conditions for seedling growth. The seedlings were grown in the above pots until developing a good root system. In the middle of March, the seedlings of each species were carefully transplanted into larger pots (8.5 x 8.5 x 9.5 cm). The new pots were filled with a substrate of soil, enriched peat (TS2, Klassmann) and perlite that was prepared in a ratio of 4:5:1 (v/v). To assess the soil fertility before the substrate preparation, a soil sample of approximately 1.5 kg was taken and transferred to the laboratory for chemical analysis. The results of the chemical analysis as well as the mechanical analysis of the soil are presented in Table 2.
After transplanting, 36 seedlings were randomly divided into three groups. In the seedlings of the first group, an integrated nutrient management (INM) was applied; in the the second group, a chemical fertilization (ChF) was applied while no fertilization was applied in the last group (control). Both types of fertilizers were foliar spray applications. The INM fertilization consisted of a nutrient solution of THEOCOPPER at 7 ml/L, THEOCAL at 1.5 g/L, THEOFAST at 5 ml/L, 10-47-10 (AGRI.FE.M. LTD Fertilizers, Greece) at 3.2 g/L, K2SO4 (0-0-52, AGRI.FE.M. LTD Fertilizers, Greece) at 2.07 g/L, micronutrients (Plex Mix, AGRI.FE.M. LTD Fertilizers, Greece) at 1.5 ml/L and MgSO4 (Mg 25.6%, AGRI.FE.M. LTD Fertilizers, Greece) at 0.6 g/L [21,27]. For the conventional fertilization (ChF), the nutrient solution consisted of NH4NO3 (34,4-0-0, Neofert®, Neochim PLC, Bulgaria) at 2.7 g/L, Ca(NO3)2 (NITROCAL, Agrohimiki, Greece) at 1.7 g/L, 10-47-10 at 3.2 g/L, K2SO4 (0-0-52) at 2.27 g/L, micronutrients Plex Mix at 1.5 ml/L and MgSO4 (Mg 25.6 %) at 0.6 g/L [21,27].The application of fertilization treatments were initiated in late March and were performed every week until the middle of June. The plants were grown inside a greenhouse of the laboratory of Floriculture, School of Agriculture, in the farm of Aristotle University of Thessaloniki (Thermi, Greece). During the experimental period, the plants were irrigated every three days.
2.6. Morphological and Physiological Measurements of Seedlings
At the end of June, the effect of fertilization treatment on morphological and physiological variables was evaluated. In each plant per treatment, the main shoot height (SH), root collar diameter (RCD) and the shoot number were measured, using a metal ruler and a digital caliper, respectively. In addition, for the measurement of the root dry biomass (RDB) and the above ground dry biomass (AGDB), four plants per treatment were randomly sampled. Dry weights were determined after drying in the oven at 74℃ for 48 h.
At the same time, gas exchange parameters, chlorophyll and chlorophyll fluorescence were determined in the fertilized and control seedlings of C. pelviformis. A chlorophyll meter CCM 200 (Opti-sciences, Tyngsboro, Massachusetts, USA) was used calculating chlorophyll content index (CCI) based on the ratio of transmittance measurement at 660 and 940 nm [38]. Chlorophyll fluorescence parameter Fv/Fm was measured by OS30p+ Rapid Plant Stress Screening Device (Opti-sciences, Tyngsboro, Massachusetts, USA). Generally, Fv/Fm is the ratio of variable (v) to maximum (m) fluorescence after dark-adaptation, representing the maximum photochemical efficiency of photosystem II which is commonly used for detection of various stress conditions [39]. For each fertilization treatment, 13 measurements with CCM200 and ten measurements with OS30p+ were measured on fully expanded young leaves of the C. pelviformis plant individuals.
2.7. Plant Tissue Analyses of Seedlings
After being dried in the oven at 74℃ for 48 h, the tissue samples of the C. pelviformis plant individuals were finely ground to pass a 40-mesh sieve. More precisely, for each treatment the above-ground part (leaves and shoots) and the below-ground part (roots) of the dried plants were separately grounded to determine the respective tissue nutrient concentration. Three samples of fine powder were formed for each plant part (above-ground and below-ground), thus corresponding to the three fertilization treatments (ChF, INM, control) examined. Subsequently, from each fine powdered and homogenized sample, three subsamples of ca. 0.25 g each were randomly taken, and each subsample was disorganized by the method of wet oxidation until a transparent solution was obtained using a triple acid mixture of H2SO4, HNO3 and HClO4 in a ratio of 5:1:1 at 80°C [40]. The solutions resulted from the digested samples, were analyzed colorimetrically for the total P (phosphorus) determination according to the Molybdenum Blue Method by using a Shimadzu spectrophotometer model UV-1201V [41]. The potassium (K), calcium (Ca), magnesium (Mg), sodium (Na), copper (Cu), iron (Fe), zinc (Zn) and manganese (Mn) concentrations were determined by atomic absorption spectroscopy (Perkin-Elmer Analyst 300). Total nitrogen (N) was determined by the Kjeldahl method [42].
2.8. Determination of Total Phenol Content and Antioxidant Capacity
Fresh leaves from C. pelviformis plant individuals per fertilization treatment were selected at the end of the experiment and were grounded to powder by liquid nitrogen. The amount of 0.1 g lyophilized leaves was extracted by adding 1 mL methanol, was vortexed for 20 min and was centrifuged at 10,000 g at room temperature for 5 min. The upper liquid portion was separated from the solid residue, was transferred to a new vial, and was stored under -20°C until analyzed.
The determination of total phenol content (TPC) in samples was conducted by the Folin–Ciocalteu method [43]. Each sample (0.5 mL) was diluted in MeOH (1:50 v/v) with the addition of 2.5 mL of the Folin–Ciocalteu solution (1:10 v/v). Two (2) mL of Na2CO3 (7.5% w/v) solution were also added after 6 min, and the whole mixture was vortexed. Finally, the prepared sample mixtures were placed in a water bath for 5 min at 50°C (LabTech Digital Water Bath, Gurgaon, India). Analyses of all samples were conducted by recording their absorbance at 760 nm by a Helios Alpha spectrophotometer (Thermo Spectronic, Cambridge, UK). TPC calculations were based on a gallic acid calibration curve (100, 50, 25, 12.5 and 0 g/mL) and were expressed in mg of gallic acid equivalents per g of fresh weight (mg GAE/g FW).
For the determination of antioxidant capacity (AC), the 2,2-diphenyl-1-picrylhydrazyl (DPPH) method was applied [43]. In 0.2 mL of samples dissolved in 80% v/v MeOH (1:20 v/v), the amount of 2.8 mL 4% w/v DPPH was added, vortexed and incubated in darkness for 30 min at room temperature. The AC of the samples was recorded at 517 nm in a spectrophotometer, was compared to an ascorbic acid calibration curve (0.1, 0.05, 0.025, 0.01 and 0 g/mL), and was expressed in mg of ascorbic acid equivalents per g of fresh weight (mg AAE/g FW).
The final readings of TPC and AC determination of each C. pelviformis sample represented the mean of three separate measurements, and they were shown as mean ± standard error. Analysis of variation (ANOVA) was used for comparisons of means using SPSS 27 and Duncan’s multiple range test at p < 0.05.
2.9. Statistical Analysis
In the germination experiment, the experimentation employed a completely randomised design with two factors (different populations, and incubation temperature). The germination data at 25°C was not further analysed since none of the seeds germinated for populations GR-BBGK-1-19,1194; GR-BBGK-1-19,1187A; GR-BBGK-1-19,1187B, or the percentages of the germinated seeds were very low, i.e., < 6% in populations GR-BBGK-1-20,365 and GR-BBGK-1-19,1056. Therefore, in the statistical analysis, the levels of the factors were: five for the populations and three for the incubation temperature (5 x 3 factorial design). The data were analysed using ANOVA in the frame of the procedure GLM (General Linear Model) [44] and the means were compared using the LSD test at a significance level p ≤ 0.05. In the remaining statistical analyses, the comparisons of the means were made using the Duncan’s test at a significance level p ≤ 0.05 [45]. All statistical analyses were carried out using SPSS 21.0 (SPSS, Inc., USA).
3. Results
3.1. Seed Germination Success
3.1.1. Effects of temperature treatments
The population (p = 0.002) and temperature (p = 0.000) factors as well as their interaction (p = 0.000) had significant effect on seed germination of C. pelviformis. However, in the presence of a significant interaction, it was considered that the interpretation of the main individual effects was of less importance.
The most successful germination was achieved at 10 and 15oC for populations GR-BBGK-1-19,1194 and GR-BBGK-1-19,1187A (97.5% and 92.5%, respectively), while populations GR-BBGK-1-20,365 (96.25%) and GR-BBGK-1-19,1187B (95%) exhibited higher germination percentage at 15 and 10oC, respectively, than at 20oC (Figure 2). For population GR-BBGK-1-19,1056, temperature had no statistically significant impact on seed germination (Figure 2).
Regarding the germination speed of C. pelviformis seeds, it was observed that in all populations the seeds germinated faster at 15oC in comparison with the rest of the temperature treatments (Figure 2). In fact, population GR-BBGK-1-20,365, GR-BBGK-1-19,1056 and GR-BBGK-1-19,1187B recorded higher rates of germination (>60%) on the 10th day (Figure 2). Furthermore, the germination of seeds at 20oC was quite gradual compared to the rest of the examined temperatures for all populations (Figure 2).
The temperature level of 10oC had a significant impact on seed germination of C. pelviformis, with seeds of GR-BBGK-1-19,1194 population exhibiting higher germination percentage compared to those of population GR-BBGK-1-19,1056 (Table 3). At 15oC, the seeds of the lowland populations GR-BBGK-1-20,365 and GR-BBGK-1-19,1194 achieved higher germination percentages than those of the semi-mountainous populations GR-BBGK-1-19,1187A and GR-BBGK-1-19,1187B (Table 3). The temperature level of 20oC notably reduced all seed germination percentages of the examined populations, particularly in GR-BBGK-1-19,1187A and GR-BBGK-1-19,1194. However, in the same temperature the seeds of the lowland population GR-BBGK-1-19,1056 germinated at their highest percentage and those of population GR-BBGK-1-19,1187B still showed 81.25% germination (see Table 3).
3.1.2. Effects of light treatments
No statistically significant results were observed in terms of mean germination percentage and mean germination time (MGT) between C. pelviformis seeds germinated under alternating light/dark and continuous dark conditions. Germination percentages were high for both treatments (98.75% in light/dark and 95% in dark) (Table 4), and the examined seeds showed similar behaviour regarding MGT under different light conditions (10.95 days in light/dark and 11.32 days in dark conditions).
3.1.3. Ecological profiling
To better understand the biological circle of C. pelviformis, an ecological profile was created in GIS depicting the temperature and precipitation needs of the wild-growing populations (Supplementary material Table S1).
The results of the temperature-related attributes showed that C. pelviformis populations are naturally adapted to a relatively warm winter season (mean temperature of coldest quarter = 11.12 ± 0.87oC; minimum temperature of coldest month = 7.63 ± 0.86oC), with the lowest average temperatures recorded in January (10.77 ± 0.89oC) and February (10.69 ± 0.86oC). The average temperature is shown to rise gradually from March (11.96 ± 0.84oC) until June (22.71 ± 0.70oC). Onwards in mid-summer (July-August), the average temperature steadily rises to 26oC. After the summer season, the average temperature decreases from 22.72 ± 0.79oC in September to 12.25 ± 0.88oC in December (winter season). The aforementioned factors, combined with the absence of extreme temperatures in its natural distribution sites (Tmin of Tmin = 5.71oC in February, Tmax of Tmax = 30.29oC in July) and the low mean diurnal range (6.86 ± 0.13oC), may indicate that C. pelviformis is naturally adapted in relatively warm environments with regards to temperature.
Historical precipitation data depicted a seasonality of rainfall in the natural range of C. pelviformis. Specifically, high precipitation was recorded from December (120.09 ± 11.06 mm) till February (98.31 ± 8.99 mm), with the highest precipitation values recorded in January (134.03 ± 12.20 mm). From March (75.16 ± 7.74 mm) onwards, the mean of precipitation was shown to decrease until May (16.90 ± 2.75 mm), leading to summer season, which is the driest period with scarce precipitation (4.54 ± 1.22 mm in June, 1.68 ± 0.60 mm in July and 2.00 ± 0.52 mm in August). After summer, the mean of precipitation was shown to rise again from 17.42 ± 1.20 mm in September to 72.80 ± 4.80 mm in November, initiating the wet season.
3.2. Seedlings’ Growth
The parameters main shoot height (p = 0.085), root collar diameter (p = 0.827), number of shoots (p = 0.146), above-ground dry biomass (p = 0.086) were not affected by the applications of fertilizers (Table 5), whereas fertilizing had impact on root dry biomass (p = 0.005), chlorophyll content index (p = 0.000), and chlorophyll fluorescence (p = 0.017) of C. pelviformis plant individuals (Table 5). As far as the root dry biomass is concerned, control plants showed significantly lower values compared to INM fertilized plants, whereas no significant difference was observed between plants fertilized with INM and ChF (Table 5). The unfertilized plants recorded the lowest values of chlorophyll content, and the plants fertilized with INM had the highest values (Table 5). The chlorophyll fluorescence of fertilized plants regardless of the type of fertilizer was significantly higher than that of the unfertilized plants of C. pelviformis (Table 5).
3.3. Leaf Nutrient Concentration
Macronutrient concentrations in leaves and shoots of C. pelviformis (Table 6) were in general affected by fertilization (p = 0.000 for N; p = 0.002 for P; p = 0.000 for K; p = 0.000 for Ca; p = 0.000 for magnesium; p = 0.000 for Na). The macronutrient content of C. pelviformis leaves and shoots showed a significantly increased concentration of N, K, Ca, Mg and Na in plants treated with INM as compared with unfertilized plants (control) or ChF, while P was detected in the highest concentration in the leaves and shoots of untreated plants (Table 6).
In the roots of the plant, fertilization affected all macronutrient concentrations (p = 0.000 for N; p = 0.004 for Pl; p = 0.000 for K; p = 0.000 for Ca; p = 0.005 for Mg) except Na (p = 0.198). INM fertilization significantly increased the contents of N, Ca and Mg compared to ChF and control plants, while the application of INF significantly decreased P and K compared to the other two treatments.
The fertilization affected the concentration of micronutrients (Table 7) in the above-ground parts (p = 0.000 for Fe; p = 0.008 for Mn; p = 0.000 for Zn; p = 0.000 for Cu) of cultivated C. pelviformis. A remarkable 10-fold increase was detected in the concentration of Cu in plants treated with INM compared to control plants (Table 7). A high increase was also noticed in the concentration of Zn, but no statistical differences were found regarding Mn between the plants treated with INM and ChF. The concentration of Fe was increased in ChF plants compared to the other two treatments.
Similarly, root micronutrients were also affected by INF fertilization (p = 0.052 for Fe; p = 0.025 for Mn; p = 0.001 for Cu; p = 0.525 for Zn). INF fertilization significantly increased the concentration of Cu, while ChF and control plants had significantly higher concentration for Mn (Table 7).
3.4. Total Phenol Content and Antioxidant Capacity
The presence of total phenols was detected in all samples of C. pelviformis, but it was more prominent and significantly higher in the fertilized samples, especially those treated with ChF (Table 8). The same trend was observed for the AC of the samples, where the one with the most potent antioxidant capacity was again the samples from fertilized plants with ChF.
4. Discussion
4.1. Seed Germination Protocols
The results of this study suggested a strong temperature effect (p = 0.0001) in the germination of C. pelviformis seeds in line with previous studies examining other members of the genus Campanula [32]. The highest percentages (>85%) of seed germination were observed at 10 and 15oC, while high rates were even recorded at 20oC (>70%). Similar results have been recently reported for another chasmophytic local endemic plants of Crete, namely C. saxatilis L. subsp. saxatilis [43]. Previous studies have indicated that various taxa of the genus Campanula can germinate even at 25°C under specific pretreatment methods [32], i.e., 99% for C. merxmuelleri Phitos, 91% for C. pangea Hatvig, and 80% for C. glomerata. However, no pretreatments were tested herein for C. pelviformis, and at this temperature C. pelviformis seed germination was quite low and was not included in the analysis.
The investigation herein attempted for the first time to examine the germination of seeds collected from different populations of C. pelviformis from lowlands (three populations) and semi-mountainous areas (two populations) of Eastern Crete, Greece. This study reports for the first time on the combined influence of altitude and temperature in seed germination behavior of C. pelviformis as recorded for the different populations examined. Although individual effects were not considered herein, however, this study provides evidence on variation in seed germination of lowland C. pelviformis populations; for example, the seeds of population GR-BBGK-1-19,1194 achieved higher germination percentage compared to population GR-BBGK-1-19,1056 at 10oC, while the latter showed its highest seed germination at 20oC. Some variation was also detected between the two semi-mountainous populations of C. pelviformis examined herein from Mt. Thrypti (the second highest mountain in Lasithi region of Crete), resulting in different germination percentages but similar decrease rates. In general, it is well established that differences may exist in the germination characteristics (rate and percentage of germination) of a taxon’s populations [30]. Previous studies have indicated that other Balkan endemic Campanula species may have quite different germination responses related to temperatures depending on the altitude of original collections; for example: (i) 10%, 11.1%, and 61.1% at 5, 15 and 25oC, respectively, in C. spatulata Sm. seeds from intermediate (900 m) altitudes compared to 0%, 21.1% and 2.2% at 5, 15 and 25oC, respectively, in seeds collected from high (1850 m) altitudes [46], or (ii) 0%, 64.4% and 65.6% at 5, 15 and 25oC, respectively, in C. lingulata Waldst. & Kit. seeds collected from lowlands (450 m) compared to 0%, 54.4% and 30% at 5, 15 and 25oC, respectively, for seeds originating from plants of intermediate (850 m) altitudes and 0%, 56.7% and 16.7% at 5, 15 and 25oC, respectively, for seeds of plants from higher (1300 m) altitudes as well as 0%, 36.7% and 4.4% at 5, 15 and 25oC, respectively, for seeds of subalpine altitudes (1700 m) close to timberline [46]. It should be noted, however, that the above-mentioned examples refer to rather common and widespread in Greece Campanula members which are Balkan endemics, with broad ecological adaptations and different habitat preferences [46]. As such, these findings should not be directly compared with other range-restricted chasmophytic Campanula members with narrow niche confined in small geographical territories such as C. pelviformis studied herein (single-island endemic of Crete). Undoubtedly, it would be useful to conduct further experimentation in freshly collected seeds to draw safer conclusions in relation to the germination requirements of the species in concern and in fact this presents another research line that we will soon follow.
Regarding the light requirements for C. pelviformis seed germination, the results of this study suggest that this taxon has no specific preference, as high percentages of germination were achieved in both light treatments (98.75% in alternating light/dark conditions and 95% in darkness), thus aligning with previous studies found in grey literature (Antonidaki-Giatromanolaki as cited by Koutsouvoulou et al. [32]). On the other hand, the seeds of several Campanula taxa clearly exhibit a preference for light for their germination [32], i.e., C. andrewsii A. DC. subsp. andrewsii (93% in light and 26% in darkenss), C. cretica (A. DC.) A. Dietr. (99% in light and 24% in darkness), C. spatulata subsp. spruneriana (Hampe) Hayek (90% in light 60% in darkness), C. spatulata subsp. spatulata (92% in light and 63% in darkness), and C. versicolor Andrews (85% in light and 38% in darkness).
Based on the available data, it appears that the seeds of C. pelviformis germinate best at temperatures of 10 and 15oC. Consequently, its natural germination in the wild habitats is expected to occur between late autumn and early winter, where the mean monthly air temperature may range between 10 and 15°C as indicated by the GIS-derived ecological profile. During this period, the high soil moisture due to increased amount of precipitation probably creates the optimum conditions for seed germination (Supplementary material Table S1). Furthermore, this timing of seed germination probably allows the growth of seedlings through winter to develop into fresh rosettes of C. pelviformis by February when local inhabitants of Crete traditionally collect this wild edible green from the wild [6].
C. pelviformis is widely used in the eastern part of Crete (Lasithi) and young rosettes of leaves are sourced directly from the wild habitats at the end of winter or early in spring together with other wild edible plants. This represents a very localized old tradition in Lasithi area; for example, C. pelviformis is not mentioned as an edible wild green by Pieroni et al. [47] in their study in Central Crete although it occurs in Heraklion prefecture [8]. This wild gathering of C. pelviformis is mainly being performed to prepare the typical local culinary preparation called “tsigarolachana” (meaning to stir in olive oil). The “tsigarolachana” can be served in everyday life with other basic food ingredients (fish, other seafood, or meat), can be cooked in casserole with all the latter and/or with eggs and other common vegetables. Ultimately, “tsigarolachana” is also the basic filling (together with other wild greens) for pot pies with hand-made dough [6]. The daily use and consumption of wild ‘chórta’ (or hórta) including C. pelviformis is linked with the traditional Mediterranean (Cretan) Diet, which is associated with high adult life expectancy (highest in the world) and decreased rates of coronary heart disease, certain cancers, and other diet-related chronic diseases [47]. Among other wild greens, C. pelviformis is considered as a medicinal plant of high nutritional value [6]. In Lasithi area, there are many regular open markets selling at very high prices equivalent to high-quality meat wild herbs and chórta in bunches, especially at the end of winter and beginning of spring. It should be noted, however, that the high demand of such localized products even at local scales can trigger over-collection from range-restricted wild populations, thus jeopardizing local genetic pools of the local Cretan endemic C. pelviformis. Therefore, the ex-situ propagation and cultivation with fertilization regimes as presented herein for the first time may offer a sustainable exploitation strategy able to satisfy any future demand regarding the traditionally consumed nutritious rosettes of C. pelviformis.
4.2. Fertilization and morphological-physiological traits
Previous studies have indicated that fertilization applications can enhance the plant photosynthetic rate [48]. The results of this study indicated that the photosynthetic rate such as chlorophyll content index and chlorophyll fluorescence had positive impact from the INF treatment. Similar outcomes had been observed in other rare local endemic species such as the Tunisian endemics Teucrium luteum (Mill.) Degen subsp. gabesianum (S. Puech) Greuter or Marrubium aschersonii Magnus, where fertilization treatments led to an increased photosynthetic rate [22,28]. Additionally, another three Cretan endemic species namely Lomelosia minoana (P. H. Davis) Greuter & Burdet subsp. minoana and subsp. asterusica (Greuter) Greuter & Burdet, and Eryngium ternatum Poir. were affected in the same direction regarding these physiological characteristics from the application of fertilization regimes [43]. More specifically, the INF treatment that resulted in this study in increased root dry biomass in C. pelviformis in a similar pattern to the observations related with M. aschersonii, while L. minoana subsp. asterusica and subsp. minoana showed a negative impact following the INF treatment [28].
4.3. Nutrient Content and Absorption
The effect of fertilization on ex-situ cultivated C. pelviformis plants is the first study to date, describing the content and the absorption of macro- and micronutrients. Both INM and ChF fertilization schemes resulted positively regarding most of the measured parameters compared to the control. The INM fertilization showed statistically significant higher values in chlorophyll content index (CCI) and root dry biomass (RDB) compared to the ChF and control; however, ChF resulted in higher values for the rest of the parameters measured but these were not statistically significant (Table 5).
Tissue analysis of the above-ground parts (leaves, stems) indicated that INM fertilization presented statistically significant higher nutrient content and absorption for N, K, Ca, Mg, Na, Zn and Cu compared to the ChF and control; however, ChF and control plants resulted in statistically significant higher values only for Fe and P, respectively (Table 6 and Table 7).
Tissue analysis for the below-ground part (roots) indicated that INM fertilization presented statistically significant higher nutrient content and absorption for N, Ca, Mg, and Cu compared to the ChF and control; however, both ChF and control resulted in statistically significant higher values for P, K, and Mn compared to INM (Table 6 and Table 7).
Measurements conducted on the leaves of other local endemic plants of Crete revealed that L. minoana subsp. minoana benefited from the INF application in terms of all macronutrients examined except for Na [43]. E. ternatum showed benefits in terms of N and K, while L. minoana subsp. asterusica exhibited high N and P levels. Additionally, in terms of micronutrient measurements in plant leaves of these taxa, the INF fertilization resulted in increased Cu levels for L. minoana subsp. minoana, Mn and Cu levels for E. ternatum, as well as in all micronutrients except Mn for L. minoana subsp. asterusica [43]. It is well known that Cu is an important micronutrient in photosynthesis and in carbon metabolism [49,50] but is also toxic to plants when in excess [51]. In the current study, it is worth noting a 10-fold increase and 58.6-64.7% higher amounts of Cu through INM foliar application for the tissue analysis of above-ground parts and below-ground parts of C. pelviformis, respectively, as compared to the ChF and control plants. The latter means that Cu moves systemically in plant tissues and moreover is translocated downstream to the root system of C. pelviformis plants, without showing any toxicity effects. Since Cu plays an important role in controlling fungal diseases in plants [52], the high amounts of Cu in C. pelviformis leaves, stems and roots probably act as protective agent against fungal pathogens. Undoubtedly, more experiments are needed to investigate the beneficial properties of Cu absorption in different developmental stages of C. pelviformis prior to further conclusions.
By analyzing the concentrations of nutrients in different plant organs, valuable insights are obtained that enhance the understanding of plant health, ecological adaptation as well as specific needs in terms of fertilization [53]. At the same time, identifying nutrient deficiencies in plants can contribute significantly to the optimization of precision agricultural technology, forming appropriate strategies for applying nutrients to plants through fertilization [54,55]. This approach ensures both economic and environmental benefits which are the keys in modern agriculture [56].
This study also showcased the beneficial effect of fertilization and the ease of responsiveness of the cultivated germplasm to fertilization, and it can provide evidence on the nutritional superiority of the cultivated germplasm. The latter, if sustainably exploited and managed, reveals a noteworthy economic potential for new product development with high added value which can be marketed in an exclusive way to address the commercial demand for C. pelviformis rosettes at least at local (Crete) or national (Greece) scales [2]. In the light of the above-mentioned, INF fertilization appears to offer promising prospects for enhancing cultivation practices for C. pelviformis when preparing nursery stocks either for population re-introduction purposes or sustainable exploitation strategies.
4.4. Total Phenol Content, Antioxidant Capacity and Fertilization Protocols
This study reports for the first time on the macro- and micronutrient profiles of the ex-situ cultivated C. pelviformis and on the effect of fertilization regimes in TPC and AC of C. pelviformis. The presence of total phenols was evidenced in all samples of ex-situ cultivated C. pelviformis regardless fertilization, but they were prominent and significantly higher in fertilized samples, especially those treated with ChF. The same trend was observed for the AC of the cultivated samples of C. pelviformis.
Considering other Cretan endemic plant species examined to date, the ChF application has been shown to have a positive impact on both total phenols and antioxidant capacity for L. minoana subsp. minoana and E. ternatum similarly to the patterns observed in C. pelviformis, while total phenol content is reported to increase in plants without any fertilization application and antioxidant capacity is reported as negatively affected by the INF application for L. minoana subsp. asterusica [43]. Previous research on other Campanula species has shown that the presence of total phenolics and significant antioxidant activity is a fact. For instance, previous studies [57] demonstrated strong antioxidant activity in methanol extracts of whole plants of C. alliariifolia Willd. Similarly, other studies [58] investigated in a comparative way the antioxidant activity of 15 Campanula species from Palestine concluding that C. sulphurea Boiss. and C. sidoniensis Boiss. & C.I.Blanche show comparatively the best DPPH radical scavenging activity. In a separate study, the phytochemical analysis of C. latifolia L. has reported very high levels of DPPH radical scavenging activity in both the aerial parts and roots of the species [59]; however, comparatively lower concentrations of total phenols have been detected in the aerial parts (0.053 mg Gallic acid/g extract) and root extracts (0.037 mg Gallic acid/g extract) of C. latifolia compared to the concentrations found in fresh leaves of C. pelviformis in the present study.
4.5. Re-Evaluation of Feasibility and Readiness Timescale for Sustainable Exploitation
The data generated in this study may facilitate future conservation efforts and population re-introduction actions in the wild habitats but also pave the way for the sustainable utilization of C. pelviformis. In this sense, the amount of data furnished in this study shed light to the urgent need to re-evaluate the feasibility and readiness timescale of value chain establishment for the species in concern. The medicinal potential (Level I evaluation) of C. pelviformis has been increased considerably to above average to high potential (68.2%) due to newly furnished data on isolated phytochemical compounds with medicinal properties [6] coupled with evidence on its nutritive value in terms of macro- and micronutrients (both Tsiftsoglou et al. [6] and this study), thus rendering C. pelviformis as a noteworthy medicinal plant of agro-alimentary interest.
Furthermore, the feasibility of value chain establishment for C. pelviformis (Level II evaluation in Krigas et al. [2]) was improved from 43.06% [2] to 66.67% after re-evaluation of related attributes in the light of newly furnished data. This upgrade affected the readiness timescale for its sustainable exploitation (Level III evaluation) from achievable in long-term to achievable in medium-term (see Table 9). This re-evaluation was based on the change of threat status for C. pelvifromis (Vulnerable according to Kougioumtzis et al. [35]) which resulted in a score of four compared to zero; the successful seed germination resulting to a score of six compared to zero; the definition of its cultivation needs in terms of watering and fertilization (score six from four) resulting in full cultivation protocols (score 6 instead of two); and the documentation of its extant commercial prices as wild edible green gathered from the wild [6].
5. Conclusions
This study provided for the first time comprehensive information on the abiotic environmental conditions of the C. pelviformis based on its distribution in natural habitats of Crete, Greece. The GIS ecological profile developed offers valuable insights into the successful ex-situ conservation and cultivation of this vulnerable local endemic species, as well as guidance for its adaptation in anthropogenic environments for conservation purposes. The results of the seed trials clearly demonstrated the effective seed germination of C. pelviformis achieved herein for the first time, thus marking a significant step towards its in-situ conservation and its potential sustainable utilization for noteworthy ornamental, agro-alimentary and medicinal properties. The experimental cultivation achieved in this study for the first time provided valuable insights into the beneficial effects of different types of fertilization in morphological-physiological characteristics, in phytochemicals such as total phenols and antioxidant capacity as well as in terms of increased contents of macro- and micro-nutrients contents as documented in the above-ground compared to wild-growing plants as well as in the below-ground part of C. pelviniformis which was studied herein for the first time. All the data furnished in this multidisciplinary investigation built to bridge extant applied research gaps was finally exploited to provide an update on the feasibility and readiness timescale for the sustainable exploitation of C. pelviformis, thus further facilitating the creation of a new value chain for the ex-situ cultivated C. pelviformis. The latter is expected to alleviate the collection pressure exerted to the traditionally harvested wild-growing populations of this remarkable yet vulnerable local endemic plant species of Crete.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Ecological profile across the natural distribution range of Campanula pelviformis wild-growing populations in Crete, Greece.
Author Contributions
Conceptualization, S.H., N.K., Ι.A. and G.T.; data curation, N.K., I.A., S.K., E.Pip., E.Pap., E.D., and G.T.; formal analysis, I.A., E.Pip., E.Pap., N.K., S.K., and S.H.; investigation, I.A., N.K., S.H., E.D., E.Pip., E.Pap., E.Ka., S.K., P.T., E.Kou., E.C. and G.T.; methodology, I.A., N.K., S.H., E.Pip., E.Pap., S.K., E.D., E.Ka., P.T., E.Kou., E.C. and G.T.; project administration, S.H., N.K. and G.T.; resources, N.K., S.H. and S.K.; software, I.A.; supervision, N.K., S.H. and G.T.; validation, N.K., I.A., E.Ka., E.D., and G.T.; visualization, I.A., E.Pip., and N.K.; writing—original draft, I.A., E.Pip., S.H., N.K., E.Ka., and G.T.; writing—review and editing, all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All data supporting the results of this study are included in the manuscript, and the datasets are available upon request.
Acknowledgments
The fieldwork of N.K., G.T., E.D., and I.A. has been partially supported by the MULTI-VAL-END project (2018-2022) entitled “Multifaceted Valorisation of single-country Endemic plants of Crete, Greece, Tunisia and Rif, Morocco for sustainable exploitation in the agro-alimentary, horticultural-ornamental and medicinal-cosmetic sectors” (ARIMNet2 2017 Transnational Joint Call).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Scariot, V.; Seglie, L.; Caser, M.; Devecchi, M. Evaluation of ethylene sensitivity and postharvest treatments to improve the vase life of four Campanula species. Eur. J. Hortic. Sci. 2008, 73, 4,166–170. Available online: https://www.researchgate.net/publication/230691067_Evaluation_of_ethylene_sensitivity_and_postharvest_treatments_to_improve_the_vase_life_of_four_Campanula_species (accessed 3 July 2023).
- Krigas, N.; Tsoktouridis, G.; Anestis, I.; Khabbach, A.; Libiad, M.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Lamchouri, F.; Tsiripidis, I.; Tsiafouli, M.A.; El Haissoufi, M.; Bourgou, S. Exploring the potential of neglected local endemic plants of three Mediterranean regions in the ornamental sector: Value chain feasibility and readiness timescale for their sustainable exploitation. Sustainability 2021a, 13, 2539. [CrossRef]
- González-Tejero, M.R.; Casares-Porcel, M.; Sánchez-Rojas, C.P.; Ramiro-Gutiérrez, J.M.; Molero-Mesa, J.; Pieroni, A.; Giusti, M.E.; Censorii, E.; de Pasquale, C.; Della, A.; Paraskeva-Hadijchambi, D. Medicinal plants in the Mediterranean area: Synthesis of the results of the project Rubia. J. Ethnopharmacol. 2008, 116, 341–357. [CrossRef]
- Hadjichambis, A.C.; Paraskeva-Hadjichambi, D.; Della, A.; Elena Giusti, M.; De Pasquale, C.; Lenzarini, C.; Censorii, E.; Reyes Gonzales-Tejero, M.; Patricia Sanchez-Rojas, C.; Ramiro-Gutierrez, J.M.; Skoula, M. Wild and semi-domesticated food plant consumption in seven circum-Mediterranean areas. Int. J. Food Sci. 2008, 59, 5, 383-414. [CrossRef]
- Heinrich, M.; Müller; W.E.; Galli, C. Local Mediterranean food plants and nutraceuticals, Vol. 59.; Karger Medical and Scientific Publishers: Basel, Switzerland, 2006. ISSN 1660–0347.
- Tsiftsoglou, O.S.; Lagogiannis, G.; Psaroudaki, A.; Vantsioti, A.; Mitić, M.N.; Mrmošanin, J.M.; Lazari, D. Phytochemical analysis of the aerial parts of Campanula pelviformis Lam. (Campanulaceae): Documenting the dietary value of a local endemic plant of Crete (Greece) traditionally used as wild edible green. Sustainability 2023, 15, 9, 7404. [CrossRef]
- Cellinese, N.; Smith, S.A.; Edwards, E.J.; Kim, S.T.; Haberle, R.C.; Avramakis, M.; Donoghue, M.J. Historical biogeography of the endemic Campanulaceae of Crete. J. Biogeogr. 2009, 36, 7, 1253-1269. [CrossRef]
- Strid, A. Atlas of the Aegean Flora Part 1 (Text & Plates) & Part 2 (Maps), 1st ed.; Englera 33 (1 & 2); Botanic Garden and Botanical Museum: Berlin, Germany; Freie Universität: Berlin, Germany, 2016. ISSN 0170-4818.
- Menteli, V.; Krigas, N.; Avramakis, M.; Turland, N.; Vokou, D. Endemic plants of Crete in electronic trade and wildlife tourism: Current patterns and implications for conservation. J. Biol. Res. Thessalon. 2019, 26, 10. [CrossRef]
- Padulosi, S.; Thompson, J.; Rudebjer, P. Fighting Poverty, Hunger and Malnutrition with Neglected and Underutilized Species (NUS): Needs, Challenges and the Way Forward; Biodiversity International: Rome, Italy, 2013. Available online: https://hdl.handle.net/10 568/68927 (accessed 3 July 2023).
- Libiad, M.; Khabbach, A.; El Haissoufi, M.; Anestis, I.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Greveniotis, V.; Tsiripidis, I.; Dariotis, E.; Tsiafouli, M.A.; Krigas, N. Agro-alimentary potential of the neglected and underutilized local endemic plants of Crete (Greece), Rif-Mediterranean coast of Morocco and Tunisia: Perspectives and challenges. Plants 2021, 10, 1770. [CrossRef]
- Bourgou, S.; Ben Haj Jilani, I.; Karous, O.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; Greveniotis, V.; Avramakis, M.; Hatzilazarou, S.; Anestis, I.; Tsoktouridis, G.; Krigas, N. Medicinal-cosmetic potential of the local endemic plants of Crete (Greece), Northern Morocco and Tunisia: Priorities for conservation and sustainable exploitation of neglected and underutilized phytogenetic resources. Biology 2021, 10, 1344. [CrossRef]
- Rivera, D.; Obón, C.; Heinrich, M.; Inocencio, C.; Verde, A.; Fajardo, J. Gathered Mediterranean food plants—Ethnobotanical investigations and historical development. In Local Mediterranean Food Plants and Nutraceuticals; Heinrich, M., Müller, W.E., Galli, C., Eds.; Karger Publishing: Basel, Switzerland, 2006; Volume 59, pp. 18–74. [CrossRef]
- Scariot, V.; Seglie, L.; Gaino, W.; Devecchi, M. Evaluation of European native bluebells for sustainable floriculture. Acta Hortic. 2012, 937, 273–280. [CrossRef]
- Gkika, P.I.; Krigas, N.; Menexes, G.; Eleftherohorinos, I.G.; Maloupa, E. Conservation of the rare Erysimum naxense Snogerup and the threatened Erysimum krendlii Polatschek: Effect of temperature and light on seed germination. Open Life Sci. 2013, 8, 1194–1203. [CrossRef]
- Grigoriadou, K.; Krigas, N.; Maloupa, E. GIS-facilitated in vitro propagation and ex situ conservation of Achillea occulta. Plant Cell Tiss. Organ Cult. 2011, 107, 531–540. [CrossRef]
- Grigoriadou, K.; Krigas, N.; Sarropoulou, V.; Papanastasi, K.; Tsoktouridis, G.; Maloupa, E. In vitro propagation of medicinal and aromatic plants: The case of selected Greek species with conservation priority. In Vitro Cell. Dev. Biol. Plant 2019, 55, 635–646. [CrossRef]
- Grigoriadou, K.; Sarropoulou, V.; Krigas, N.; Maloupa, E.; Tsoktouridis, G. GIS-facilitated effective propagation protocols of the Endangered local endemic of Crete Carlina diae (Rech. f.) Meusel and A. Kástner (Asteraceae): Serving ex-situ conservation needs and its future sustainable exploitation as an ornamental. Plants 2020, 9, 1465. [CrossRef]
- Grigoriadou, K.; Krigas, N.; Sarropoulou, V.; Maloupa, E.; Tsoktouridis, G. Propagation and ex-situ conservation of Lomelosia minoana subsp. minoana and Scutellaria hirta-two ornamental and medicinal Cretan endemics (Greece). Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12168. [CrossRef]
- Hatzilazarou, S.; El Haissoufi, M.; Pipinis, E.; Kostas, S.; Libiad, M.; Khabbach, A.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Aslanidou, V.; Greveniotis, V.; Sakellariou, M.A.; Anestis, I.; Tsoktouridis, G.; Krigas, N. GIS-Facilitated seed germination and multifaceted evaluation of the Endangered Abies marocana Trab. (Pinaceae) enabling conservation and sustainable exploitation. Plants 2021, 10, 2606. [CrossRef]
- Fanourakis, D.; Paschalidis, K.; Tsaniklidis, G.; Tzanakakis, V.A.; Bilias, F.; Samara, E.; Liapaki, E.; Jouini, M.; Ipsilantis, I.; Maloupa, E.; Tsoktouridis, G; Matsi, T.; Krigas, N. Pilot cultivation of the local endemic Cretan marjoram Origanum microphyllum (Benth.) Vogel (Lamiaceae): Effect of fertilizers on growth and herbal quality features. Agronomy 2021, 12, 94. [CrossRef]
- Kostas, S.; Hatzilazarou, S.; Pipinis, E.; Bourgou, S.; Ben Haj Jilani, I.; Ben Othman, W.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; Koundourakis, E.; Greveniotis, V.; Papaioannou, E.; Sakellariou, M.A.; Anestis, I.; Tsoktouridis, G.; Krigas, N. DNA Barcoding, GIS-Facilitated seed germination and pilot cultivation of Teucrium luteum subsp. gabesianum (Lamiaceae), a Tunisian local endemic with potential medicinal and ornamental value. Biology 2022, 11, 462. [CrossRef]
- Khabbach, A.; Libiad, M.; El Haissoufi, M.; Bourgou, S.; Megdiche-Ksouri, W.; Lamchouri, F.; Ghrabi-Gammar, Z.; Menteli, V.; Vokou, D.; Tsoktouridis, G.; Krigas, N. Electronic commerce of the endemic plants of Northern Morocco (Mediterranean Coast-Rif) and Tunisia over the Internet. Bot. Sci. 2021, 100, 139–152. [CrossRef]
- Krigas, N.; Mouflis, G.; Grigoriadou, K. Conservation of important plants from the Ionian Islands at the Balkan Botanic Garden of Kroussia, N Greece: Using GIS to link the in situ collection data with plant propagation and ex situ cultivation. Biodivers. Conserv. 2010, 19, 3583–3603. [CrossRef]
- Krigas, N.; Lykas, C.; Ipsilantis, I.; Matsi, T.; Weststrand, S.; Havström, M.; Tsoktouridis, G. Greek tulips: Worldwide electronic trade over the internet, global ex situ conservation and current sustainable exploitation challenges. Plants 2021b, 10, 580. [CrossRef]
- Krigas, N.; Karapatzak, E.; Panagiotidou, M.; Sarropoulou, V.; Samartza, I.; Karydas, A.; Damianidis, C.K.; Najdovski, B.; Teofilovski, A.; Mandzukovski, D.; Stipanović, V.D.; Papanastasi, K.; Kapagianni, P.D.; Fotakis, D.; Grigoriadou, K.; Tsoktouridis, G.; Andonovski, V.; Maloupa, E. Prioritizing plants around the cross-border area of Greece and the Republic of North Macedonia: Integrated conservation actions and sustainable exploitation potential. Diversity 2022, 14, 570. [CrossRef]
- Paschalidis, K.; Fanourakis, D.; Tsaniklidis, G.; Tzanakakis, V.A.; Bilias, F.; Samara, E.; Kalogiannakis, K.; Debouba, F.J.; Ipsilantis, I.; Tsoktouridis, G.; Matsi, T.; Krigas, N. Pilot cultivation of the Vulnerable Cretan endemic Verbascum arcturus L. (Scrophulariaceae): Effect of fertilization on growth and quality features. Sustainability 2021, 13, 14030. [CrossRef]
- Pipinis, E.; Hatzilazarou, S.; Kostas, S.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; Koundourakis, E.; Greveniotis, V.; Papaioannou, E.; Sakellariou , M.A.; Anestis, I.; Tsoktouridis, G.; Krigas, N. Facilitating conservation and bridging gaps for the sustainable exploitation of the Tunisian local endemic plant Marrubium aschersonii (Lamiaceae). Sustainability 2022, 14, 1637. [CrossRef]
- Sefi, O.; Bourgou, S.; Megdiche-Ksouri, W.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; Krigas, N.; Ghrabi-Gammar, Z. Bioactivities and phenolic composition of Limonium boitardii Maire and L. cercinense Brullo & Erben (Plumbaginaceae): Two Tunisian strict endemic plants. Inter. J. Environ. Health Res. 2022, 32, 2496–2511. [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. ISBN: 978-0-12-416677-6.
- Macdonald, B. Practical Woody Plant Propagation for Nursery Growers; Timber Press Inc.: Portland, OR, USA, 2006; p. 669. ISBN: 9780881920628.
- Koutsovoulou, K.; Daws, M.I.; Thanos, C.A. Campanulaceae: A family with small seeds that require light for germination. Ann. Bot. 2014, 113, 1, 135–143. [CrossRef]
- O’Reill, C.; Arnott, J.T.; Owens, J.N. Effects of photoperiod and moisture availability on shoot growth, seedling morphology, and cuticle and epicuticular wax features of container-grown western hemlock seedlings. Can. J. Forest Res. 1989, 19, 122–131. [CrossRef]
- Buendia-Velazquez, M.V.; Lopez-Lopez, M.A.; Cetina-Alcala, V.M.; Diakite, L. Substrates and nutrient addition rates affect morphology and physiology of Pinus leiophylla seedlings in the nursery stage. iForest 2016, 10, 115–120. [CrossRef]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Strid, A.; Dimopoulos, P. Extinction risk assessment of the Greek endemic flora. Biology 2021, 10, 195. [CrossRef]
- Lazarina, M.; Kallimanis, A.S.; Dimopoulos, P.; Psaralexi, M.; Michailidou, D.E.; Sgardelis, S.P. Patterns and drivers of species richness and turnover of neo-endemic and palaeo-endemic vascular plants in a Mediterranean hotspot: the case of Crete, Greece. J. Biol. Res.-Thessaloniki 2019, 26, 12. [CrossRef]
- Ellis, R.H. and Roberts, E.H. The quantification of ageing and survival in orthodox seeds. Seed Sci. Technol. 1981, 9, 373-409.
- Richardson, A.D.; Duigan, S.P.; Berlyn, G.P. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol. 2002, 153, 185–194. [CrossRef]
- Cessna, S.; Demmig-Adams, B.; Adams, W.W., III. Exploring photosynthesis and plant stress using inexpensive chlorophyll fluorometers. J. Nat. Resourc. Life Sci. Educ. 2010, 39, 22–30. [CrossRef]
- Allen, S.E.; Grimshaw, H.M.; Rowland, A.P. Chemical analysis. In Methods in Plant Ecology; Moore, P.D., Chapman, S.B., Eds.; Blackwell Scientific Publication: Oxford, UK, 1986; pp. 285–344. ISBN: 9780632003211.
- Olsen, S.R.; Sommers, L.E. Total phosphorus. In Methods of Soil Analysis Part 2: Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; pp. 403–430. Available online: https://acsess.onlinelibrary.wiley.com/doi/pdf/10.2134/agronmonogr9.2.2ed. frontmatter (accessed 3 July 2023).
- Bremmer, J.M.; Mulvaney, C.S. Nitrogen - Total, In Methods of Soil Analysis Part 2: Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; pp. 595–624. Available online: https://acsess.onlinelibrary.wiley.com/doi/pdf/10.2134/agronmonogr9.2.2ed. frontmatter (accessed 3 July 2023).
- Hatzilazarou, S.; Pipinis, E.; Kostas, S.; Stagiopoulou, R.; Gitsa, K.; Dariotis, E.; Avramakis, M.; Samartza, I.; Plastiras, I.; Kriemadi, E.; Bareka, P.; Lykas, C.; Tsoktouridis, G.; Krigas, N. Influence of temperature on seed germination of five wild-growing Tulipa species of Greece associated with their ecological profiles: Implications for conservation and cultivation. Plants 2023, 12, 1574. [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical procedures in agricultural research, 2nd ed. Wiley: New York, USA, 1984. ISBN 0-471-87092-7.
- Klockars, A.; Sax, G. Multiple Comparisons; Sage Publications: Newbury Park, CA, USA, 1986; p. 87. ISBN 0-8039-2051-2.
- Blionis, G.; Vokou, D. Reproductive attributes of Campanula populations from Mt Olympos, Greece. Plant Ecol. 2005, 178, 77–88. [CrossRef]
- Pieroni, A.; Sulaiman, N.; Sõukand, R. Chorta (wild greens) in Central Crete: The bio-cultural heritage of a hidden and resilient ingredient of the Mediterranean Diet. Biology 2022, 11, 673. [CrossRef]
- Zou, T.X.; Dai, T.B.; Jiang, D. Effects of nitrogen and potassium application levels on flag leaf photosynthetic characteristics after anthesis in winter wheat. Acta Agron. Sin. 2007, 33, 1667–1673.
- Yruela, I. Transition metals in plant photosynthesis. Metallomics 2013, 5, 9, 1090-109. [CrossRef]
- Dalcorso, G.; Manara, A.; Piasentin, S.; Furini, A. Nutrient metal elements in plants. Metallomics 2014, 10, 1770-1788. [CrossRef]
- Marschner, H. 8 - Functions of Mineral Nutrients: Macronutrients. In Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 1995; pp. 229-312.
- Zhu, Q.; Zhang, M.; Ma, Q. Copper-based foliar fertilizer and controlled release urea improved soil chemical properties, plant growth and yield of tomato. Sci. Hortic. 2012, 143, 109-114. [CrossRef]
- Liu, Y.; Wang, X.; Luo, F.; Wu, L.; Zhang, Y.; Lan, T. Determination and analysis of leaf P and K concentrations of several plant species in Jinan city. E3S Web Conf. 2018, 53, 03055. [CrossRef]
- Burns, I.G. Influence of plant nutrient concentration on growth rate: Use of a nutrient interruption technique to determine critical concentrations of N, P and K in young plants. Plant Soil 1992, 142, 221–233. [CrossRef]
- See, C.R.; Yanai, R.D.; Fisk, M.C.; Vadeboncoeur, M.A.; Quintero, B.A.; Fahey, T.J. Soil nitrogen affects phosphorus recycling: Foliar resorption and plant-soil feedbacks in a northern hardwood forest. Ecology 2015, 96, 2488–2498. [CrossRef]
- Siedliska, A.; Baranowski, P.; Pastuszka-Wozniak, J.; Zubik, M.; Krzyszczak, J. Identification of plant leaf phosphorus content at different growth stages based on hyperspectral reflectance. BMC Plant Biol. 2021, 21, 28. [CrossRef]
- Dumlu, M.U.; Gurkan, E.; Tuzlaci, E. Chemical composition and antioxidant activity of Campanula alliariifolia, Nat. Prod. Res. 2008, 22, 6, 477-482. [CrossRef]
- Jaradat, N.A.; Abualhasan, M. Comparison in vitro of Antioxidant Activity between Fifteen Campanula Species (Bellflower) from Palestinian Flora. Pharmacogn. J. 2015, 7, 5, 276-279. [CrossRef]
- Moosavi, S.R.; Ardekani, M.R.S.; Vazirian, M.; Lamardi, S.N.S. Campanula latifola, giant bellflower: Ethno-botany, phytochemical and antioxidant evaluation. Tradit. Integr. Med. 2018, 3, 113-119. Available online: https://jtim.tums.ac.ir/index.php/jtim/article/view/168 (accessed 3 July 2023).
Figure 1.
Plant individual of Campanula pelviformis as chasmophyte on vertical rocky crevices of Kalo Chorio (Lasithi, Greece) in full flowering (A), ripe fruits collected and Thrypti (B), seed germination in Petri dishes (C) and individual germinated seeds (D).
Figure 1.
Plant individual of Campanula pelviformis as chasmophyte on vertical rocky crevices of Kalo Chorio (Lasithi, Greece) in full flowering (A), ripe fruits collected and Thrypti (B), seed germination in Petri dishes (C) and individual germinated seeds (D).


Figure 2.
Cumulative germination percentage diagrams of Campanula pelviformis seeds of the five populations (A: lowland populations; B: Semi-mountainous populations, see Table 1) incubated at 10 (●), 15 (■) and 20 (□). The comparisons were made using the LSD test. 1 – In each population, means were statistically different at p<0.05 when not sharing a common letter; ns2: Non-significant differences.
Figure 2.
Cumulative germination percentage diagrams of Campanula pelviformis seeds of the five populations (A: lowland populations; B: Semi-mountainous populations, see Table 1) incubated at 10 (●), 15 (■) and 20 (□). The comparisons were made using the LSD test. 1 – In each population, means were statistically different at p<0.05 when not sharing a common letter; ns2: Non-significant differences.
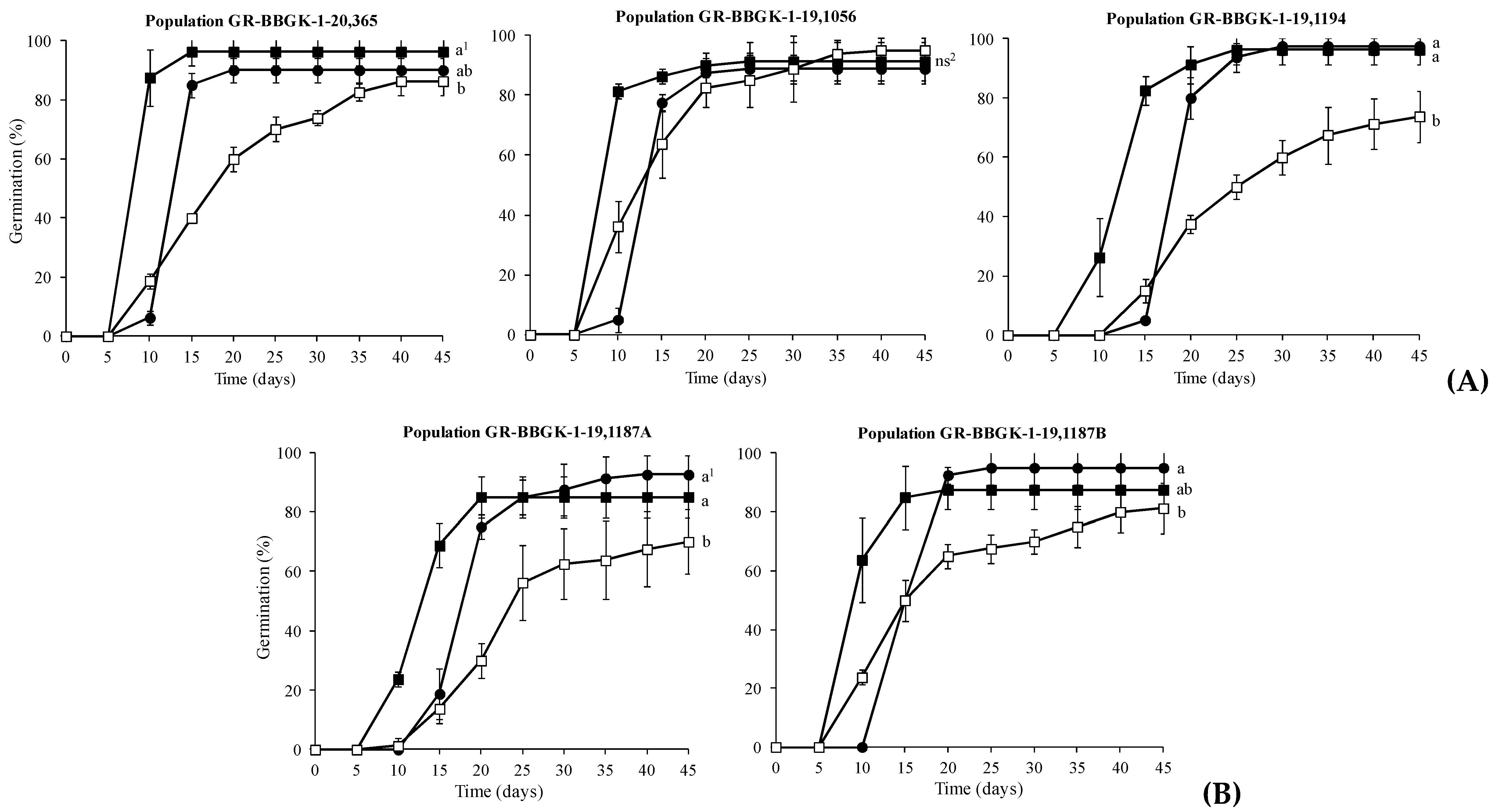

Table 1.
Seed collection details and IPEN (International Plant Exchange Network) accession numbers of Campanula pelviformis seed lots collected from lowland and semi-mountainous areas of Eastern Crete, Greece.
Table 1.
Seed collection details and IPEN (International Plant Exchange Network) accession numbers of Campanula pelviformis seed lots collected from lowland and semi-mountainous areas of Eastern Crete, Greece.
| IPEN Accession number | Collection date | Collection site in Lasithi, Eastern Crete | Latidude (North) | Longitude (East) | Altitude (m) |
| GR-BBGK-1-20,365 | 26 July 2020 | After Kalo Chorio | 35.1242 | 25.7490 | 45 |
| GR-BBGK-1-19,1056 | 6 August 2019 | Istron | 35.1248 | 25.7444 | 25 |
| GR-BBGK-1-19,1194 | 29 July 2019 | Mochlos area |
35.1725 | 25.9051 | 105 |
| GR-BBGK-1-19,1187A | 14 June 2019 | Thrypti northern outskirts |
35.0931 | 25.8639 | 835 |
| GR-BBGK-1-19,1187B | 14 June 2019 | Central Thrypti | 35.0903 | 25.8658 | 870 |
Table 2.
Chemical and physical properties of the soil used for Campanula pelviformis seedling development.
Table 2.
Chemical and physical properties of the soil used for Campanula pelviformis seedling development.
| Chemical analysis | Mechanical analysis | ||||||
| pH | Organic matter (%) | Soluble salts (mS/cm) | CaCO3 (%) |
Sand (%) |
Silt (%) |
Clay (%) |
|
| 8.12 | 0.36 | 0.35 | 5.50 | 56.00 | 28.00 | 16.00 | |
| Macronutrient concentrations (ppm) | |||||||
| N-NO3 | P | K | Mg | Ca | |||
| 8.00 | 8.00 | 104.00 | 842.00 | >2000 | |||
| Micronutrient concentrations (ppm) | |||||||
| Fe | Zn | Mn | Cu | ||||
| 4.7 | 2.00 | 7.06 | 0.77 | ||||
Table 3.
Effect of Campanula pelviformis populations from lowland and semi-mountainous areas on the seed germination percentage for each incubation temperature with high seed germination. Means ± standard deviation values are given.
Table 3.
Effect of Campanula pelviformis populations from lowland and semi-mountainous areas on the seed germination percentage for each incubation temperature with high seed germination. Means ± standard deviation values are given.
|
Area |
Population | Incubation Temperatures | ||
| 10°C | 15°C | 20°C | ||
|
Lowland |
GR-BBGK-1-20,365 | 90.00 ± 4.08 ab1 | 96.25 ± 4.79 a | 86.25 ± 4.79 b |
| GR-BBGK-1-19,1056 | 88.75 ± 4.79 b | 91.25 ± 2.50 ab | 95.00 ± 4.08 a | |
| GR-BBGK-1-19,1194 | 97.50 ± 2.89 a | 96.25 ± 4.79 a | 73.75 ± 8.54 cd | |
|
Semi-mountainous |
GR-BBGK-1-19,1187A | 92.50 ± 2.89 ab | 85.00 ± 4.08 b | 70.00 ± 10.80 d |
| GR-BBGK-1-19,1187B | 95.00 ± 4.08 ab | 87.50 ± 5.00 b | 81.25 ± 8.54 bc | |
1Values, in the same column, followed by the same letter are not significantly different (p > 0.05) according to LSD test.
Table 4.
Effect of light conditions on germination of Campanula pelviformis seeds. Means and standard deviation values are given.
Table 4.
Effect of light conditions on germination of Campanula pelviformis seeds. Means and standard deviation values are given.
| Germination percentage (%) | Mean germination time (days) | |
| Light/dark | 98.75 ± 2.50 a1 | 10.95 ± 0.14 a |
| Dark | 95.00 ± 4.08 a | 11.32 ± 0.53 a |
1 In each column, means followed by the same letter are not significantly different at p < 0.05. The comparisons were made using T-test.
Table 5.
Effect of fertilization (ΙΝΜ: Integrated Nutrient Management; ChF: Chemical Fertilization) on morphological and physiological characteristics of Campanula pelviformis plant individuals. Means ± standard deviation values are provided.
Table 5.
Effect of fertilization (ΙΝΜ: Integrated Nutrient Management; ChF: Chemical Fertilization) on morphological and physiological characteristics of Campanula pelviformis plant individuals. Means ± standard deviation values are provided.
| Attribute (Unit) | Control | INM | ChF |
| Shoot height (cm) | 18.58 ± 2.23 a* | 17.75 ± 2.30 a | 19.92 ± 2.43 a |
| Root collar diameter (mm) | 8.94 ± 1.27 a | 9.27 ± 1.51 a | 9.28 ± 1.80 a |
| Number of shoots | 11.58 ± 1.73 a | 11.08 ± 2.74 a | 13.25 ± 3.49 a |
| Root dry biomass (g) | 1.00 ± 0.12 b | 1.72 ± 0.41 a | 1.39 ± 0.42 ab |
| Above ground dry biomass (g) | 1.89 ± 0.29 a | 1.85 ± 0.30 a | 2.32 ± 0.26 a |
| Chlorophyll content index (CCI) | 17.72 ± 2.96 c | 50.59 ± 7.66 a | 29.94 ± 7.44 b |
| Chlorophyll fluorescence (Fv/Fm) | 0.748 ± 0.006 b | 0.763 ± 0.022 a | 0.768 ± 0.009 a |
*Values in the same row followed by the same letter are not significantly different (p > 0.05) according to Duncan’s test.
Table 6.
Macronutrient concentrations in above-ground (leaves and shoots) and below-ground (roots) parts of cultivated Campanula pelviformis plant individuals in greenhouse with regards to fertilization (ΙΝΜ: Integrated Nutrient Management; ChF: Chemical Fertilization; Control: absence of INM or CHF, only water).
Table 6.
Macronutrient concentrations in above-ground (leaves and shoots) and below-ground (roots) parts of cultivated Campanula pelviformis plant individuals in greenhouse with regards to fertilization (ΙΝΜ: Integrated Nutrient Management; ChF: Chemical Fertilization; Control: absence of INM or CHF, only water).
| Treatment | Nitrogen (N) (%) | Phosphorus (P) (mg/g) | Potassium (K) (mg/g) | Calcium (Ca) (mg/g) | Magnesium (Mg) (mg/g) | Sodium (Na) (mg/g) |
| Above-ground part (leaves, stems) | ||||||
| Control | 1.25 c* | 3.20 a | 26.39 b | 15.18 c | 4.88 b | 8.35 c |
| INM | 2.12 a | 2.99 b | 30.17 a | 18.85 a | 5.52 a | 14.63 a |
| ChF | 1.65 b | 3.01 b | 25.45 c | 16.19 b | 4.79 b | 11.06 b |
| Below-ground part (roots) | ||||||
| Control | 1.11 c | 1.23 a | 5.27 a | 2.48 b | 1.93 c | 3.04 a |
| INM | 2.06 a | 0.87 b | 4.41 b | 3.43 a | 2.37 a | 3.28 a |
| ChF | 1.34 b | 1.24 a | 5.27 a | 2.30 b | 2.17 b | 2.98 a |
*In the same column, the means are statistically different at p < 0.05 when not sharing a common letter. The comparisons were made using the Duncan’s test.
Table 7.
Micronutrient concentration in above-ground (leaves and shoots) and below-ground (roots) parts of Campanula pelviformis plant individuals with regards to fertilization (ΙΝΜ: Integrated Nutrient Management; ChF: Chemical Fertilization; Control: Only water).
Table 7.
Micronutrient concentration in above-ground (leaves and shoots) and below-ground (roots) parts of Campanula pelviformis plant individuals with regards to fertilization (ΙΝΜ: Integrated Nutrient Management; ChF: Chemical Fertilization; Control: Only water).
| Treatment | Iron (Fe) (ppm) | Manganese (Mn) (ppm) | Zinc (Zn) (ppm) | Copper (Cu) (ppm) |
| Above-ground part (leaves, stems) | ||||
| Control | 633.08 c* | 52.71 b | 12.22 c | 15.65 b |
| INM | 721.71 b | 64.84 a | 19.80 a | 152.79 a |
| ChF | 797.25 a | 66.94 a | 15.43 b | 2.96 c |
| Below-ground part (root) | ||||
| Control | 553.60 a | 27.05 a | 17.00 a | 13.77 b |
| INM | 452.77 a | 20.52 b | 17.14 a | 22.68 a |
| ChF | 527.01 a | 26.47 a | 15.06 a | 14.30 b |
*In the same column, the means are statistically different at p < 0.05 when not sharing a common letter. The comparisons were made using the Duncan’s test.
Table 8.
Effect of fertilization (ΙΝΜ: Integrated Nutrient Management; ChF: Chemical Fertilization; Control: Only water) on the concentration of total phenols and antioxidant capacity of Campanula pelviformis young leaves, expressed in mg gallic acid equivalents (GAE) and ascorbic acid equivalents (AAE) per g of fresh (FW), respectively.
Table 8.
Effect of fertilization (ΙΝΜ: Integrated Nutrient Management; ChF: Chemical Fertilization; Control: Only water) on the concentration of total phenols and antioxidant capacity of Campanula pelviformis young leaves, expressed in mg gallic acid equivalents (GAE) and ascorbic acid equivalents (AAE) per g of fresh (FW), respectively.
| Treatment | Total Phenols (mg GAE/g FW) |
Antioxidant Capacity (mg AAE/g FW) |
|---|---|---|
| Control | 0.993 ± 0.037 b* | 0.094 ± 0.0004 c |
| INM | 1.022 ± 0.009 b | 0.102 ± 0.0008 b |
| ChF | 1.195 ± 0.021 a | 0.106 ± 0.0006 a |
*Values are means ± standard error of the mean. Means followed by a different letter are statistically significantly different (p < 0.05).
Table 9.
Overview of the multifaceted evaluation (Level I: General and special interest in different economic sectors; Level II: feasibility, and Level III: Readiness timescale for sustainable exploitation after Krigas et al. [2]) regarding Campanula pelviformis revisited in the light of the data furnished in this study. Previous assessments and upgrades of individual scores after the data presented herein are discerned with right arrow (→).
Table 9.
Overview of the multifaceted evaluation (Level I: General and special interest in different economic sectors; Level II: feasibility, and Level III: Readiness timescale for sustainable exploitation after Krigas et al. [2]) regarding Campanula pelviformis revisited in the light of the data furnished in this study. Previous assessments and upgrades of individual scores after the data presented herein are discerned with right arrow (→).
| General Ornamental Interest [2] |
* Special Ornamental Interest [2] |
Agro- Alimentary Interest [11] |
Medicinal Interest [12] |
Feasibility and Readiness Timescale for Sustainable Exploitation (Level II and III Assessments**) |
| Below average | Average | Below average | Low → Above average to high | 43.06% → 66.67% |
| 41.67% | 53.13% / 8.05% / 52.15% / 59.29% | 42.86% | 38.89% → 68.52% | Achievable in long-term → in medium-term |
*1st percentage: as pot/patio plant; 2nd percentage: for home gardening; 3rd percentage: for landscaping; 4th percentage: for xeriscaping. ** Assessments regarding the feasibility (Level II evaluation) and readiness timescale (Level III evaluation) for the sustainable exploitation as estimated in [2].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



