
Submitted:
04 June 2025
Posted:
05 June 2025
You are already at the latest version
Abstract
Plastic pollution in marine environments poses a new global threat. Microplastics (MPs) can bioaccumulate in marine organisms, leading to oxidative stress (OS). This study investigates MPs accumulation and associated OS responses in six invertebrate species (Bivalvia, Gastropoda, Malacostraca) and three key fish species of the Bulgarian Black Sea ecosystems. Hydrobionts were collected from representative coastal habitats, MPs were quantified microscopically, and OS biomarkers (lipid peroxidation, glutathione, antioxidant enzymes) were analyzed spectrometrically in fish liver and gills, and invertebrate soft tissues. Results revealed species-specific MPs bioaccumulation, with Palaemon adspersus, Rathke (1837) and Sprattus sprattus (Linnaeus, 1758) having the highest concentrations per gram soft tissue and muscle, respectively. The alien Pacific oyster exhibited higher MPs loads than the native species. OS biomarkers varied markedly among species, reflecting different stress responses. Cluster analyses indicated that MPs were mainly grouped with catalase activity in all species. The specific OS (SOS) index showed that OS levels were linked to both habitat conditions and species-specific antioxidant defense. These findings confirmed the importance of environmental conditions, including MPs pollution, and evolutionarily developed adaptation capacity of organisms for tolerance and resilience to induced stress. The study emphasizes the need for monitoring MPs and OS to better assess the ecological risk.
Keywords:
Black Sea
; ecological risk
; fish
; marine invertebrates
; microplastics
; oxidative stress
1. Introduction
Plastic products are being produced at ever-increasing rates, making plastic pollution one of the most important contemporary environmental issues. Plastic is a very practical material that is long-lasting, resistant to degradation, inert, and easy to shape, with very low production costs. The annual global production of plastic materials is about 450 million tons in 2025, increasing more than 20-fold in the last 60 years [1]. The plastic production is expected to continue rising to approximately 590 million metric tons by 2050. Only a small amount of the plastics is recycled, and a large part is released into the environment as pollutants. About 10% of the plastic produced each year ends up in the oceans and seas [2]. A study reports that between 4 and 75 plastic objects enter the Black Sea every hour through river inflow [3] and account for 60-80% of marine litter [4,5,6].
Contamination of the marine environment with microplastics (MPs; plastics<5 mm) has been identified as an issue of global concern [7] and documented extensively in seawater, marine sediments, and marine biota [8]. The pollution of the marine environment with MPs is particularly high worldwide, and the Black Sea is no exception [9]. Studies on MPs in the Black Sea (including the Bulgarian part) have focused on the sources of pollution [10,11,12,13], their presence in sediments [14,15,16,17,18,19,20] and seawater [15,18,21,22,23,24,25,26], as well as in different species of invertebrates and vertebrates [27,28,29,30,31,32,33,34,35], including bivalves and fish caught along the Bulgarian coast [36,37,38,39,40].
Initially, it was believed that MPs are bioinert, passing through the digestive system of animals without being absorbed and without affecting the organisms. Marine organisms can be exposed to direct ingestion of MPs or indirect ingestion via prey items, or also through respiration. Irrespective of the pathway, intake of MPs by marine organisms can result in adverse physical and chemical impacts [41,42,43].
Currently, there is increasing evidence that MPs can overcome biological membranes and, entering cells and tissues, can have various negative influences on the organism [44,45,46,47,48]. Physical retention of MPs in digestive tracts [49] and chemical leaching of plastic additives into tissues [50] have been reported to have toxic effects. These impacts are often investigated during controlled laboratory exposures using a variety of endpoints such as growth rate [51,52], fecundity [53], and mortality [54].
In wild-caught organisms, however, causality between MPs exposure pathways, accumulation, and observed effects is often difficult to ascertain due to the multitude of stressors present in the marine environment [55,56]. The ecological risks of plastic contamination can be defined as the likelihood of adverse health effects and their ecological consequences occurring as a result of the organisms’ exposure to MPs [57,58]. The small size and variable buoyancy make MPs readily available for uptake by a wide range of organisms across different trophic levels and feeding strategies [59] and thus of particular concern as a pollutant affecting not only organisms but also ecosystems. Indeed, the uptake of MPs has been increasingly confirmed in wild populations of marine organisms across all trophic levels [4]. Hence, understanding endpoints such as MPs bioaccumulation and biomagnification can assist in improving our knowledge of the potential ecological effects associated with exposure pathways in the marine environment [56,60,61,62,63].
The mechanism of MPs' damage may be at least partly due to the induction of oxidative stress (OS) in organisms after their exposure [48]. Consequently, OS has the potential to be a reliable tool to assess the health and metabolic status of marine organisms, being an expression of the unified response of organisms to the multiple effects of the endo- and exogenous factors of their living environment. Nowadays, it is broadly accepted that OS-induced changes at the cellular level can subsequently impact higher hierarchical levels of biological organization - a phenomenon that underpins the theoretical framework of “stress ecology” [64,65].
Experimental studies on OS induced by MPs have been reported for various marine organisms, both invertebrate species (representatives of Annelida, Arthropoda, Cnidaria, Mollusca, Decapoda [40,66,67,68] and vertebrates – predominantly fish [69,70,71,72]. These effects are associated both with MPs direct action by the release of toxic monomers of plastics, but also with the transfer of metals, persistent organic pollutants [48,73,74], and pathogenic microorganisms [75], which can be adsorbed on their surface. As a general end-effect, laboratory studies have shown changes in feeding, growth, reproduction, and survival of aquatic species after exposure to MPs [76,77]. However, limited published evidence still exists for MPs' trophic transfer, biomagnification, and associated additives in marine food webs and the effects on the health of organisms and ecosystems.
The present study aimed to assess MPs transfer, accumulation, and effects in key species from Bulgarian Black Sea habitats and associated OS responses, with their adaptation capacity for tolerance and resilience to induced stress.
2. Materials and Methods
2.1. Species and Sampling Areas
The species for the study were collected from characteristic habitats of the Bulgarian Black Sea coastal areas, both in the northern (larger Varna Bay) and southern (larger Burgas Bay) regions (Figure 1). The invertebrates and fish species selected for this study are key ecosystem components. The invertebrates were Mytilus galloprovincialis Lamarck, 1819; Rapana venosa (Valenciennes, 1846), Bittium reticulatum (da Costa, 1778); Palaemon adspersus Rathke, 1837. The Pacific oyster, Magallana gigas (Thunberg, 1793), is a recent intruder in the Black Sea, and individuals of the species were also collected for comparative purposes. The fish samples of Mullus barbatus Linnaeus, 1758 and Sprattus sprattus (Linnaeus, 1758) were collected during the monitoring pelagic trawl survey of fish stocks, Mesogobius batrachocephalus (Pallas, 1814) individuals were collected using a fishing net cage trap, and the invertebrate species were collected using the diving method.
All sampled individuals from the studied species were shock frozen for best preservation [78] and transported in dry ice to the laboratory. In the laboratory, they were stored at -20°C until the microplastic analyses and at -80°C until the biochemical analyses were performed.
In the analysis of MPs bioaccumulation, available data [38] from our data set for the characteristic species, lagoon cockle (Cerastoderma glaucum (Bruguière, 1789)) and soft-shell clam (Mya arenaria Linnaeus, 1758) from Varna Lake were also included.
2.2. Morphometry
On the day of analysis, the sampled individuals were thawed at room temperature, and the total length (L) and weight (W) of all sampled individuals were measured in centimeters and grams, respectively. In the fish, Fulton’s condition factor (K) was computed according to the formula: K = 100*W/L3, where W is the total body wet weight in grams and L is the total length in cm; the factor 100 is used to bring K close to a value of one [79].
2.3. Determination of Microplastics in Species Samples
To prevent plastic contamination in the laboratory, all surfaces were cleaned with ethanol (EtOH); all liquids were filtered; glass and metal consumables were rinsed thoroughly with filtered water before use and covered with metal foil. Procedural controls for liquids and air were performed and considered in the results of the samples. The filters are inspected and pre-cleaned under a microscope to remove any impurities before analysis. Cotton laboratory clothes with distinguishable colors, nitrile gloves, and cuffs were used. The air in the laboratory was pre-cleansed to minimize self-contamination with an Oberon–520 air purifier and a HEPA filter.
In fish, the number of plastic particles in the gastrointestinal tract (GIT) and muscle tissue was counted and used as an indicator for MPs bioaccumulation and also for possible further transfer to humans. Before tissue digestion, the individuals were rinsed well with a filtered solution of 1:1 EtOH:dH2O, the fish GIT and muscle tissues were carefully separated and the weight was recorded. The chitinous shell and head of the P. adspersus individuals were carefully cut. The surface of the M. galloprovincialis and R. venosa shells was first scraped to clean them of overgrown/attached species, and the soft tissues were carefully separated from the shells. The B. reticulatum shells were carefully broken with curved metal tweezers, and the soft tissue delicately separated.
The tissues were rinsed well with a filtered solution of 1:1 EtOH:dH2O and then digested with filtered 10% potassium hydroxide at 40°C for 72 h in glass containers covered with metal foil. Thereafter, the resulting samples were filtered on glass microfiber filters with a pore size of 2.7 μm and a diameter of 47 mm (Filtratech, FV24A0047) with a stainless steel Rocker MultiVac 300 vacuum system, using a filtered solution of 1:1 EtOH:dH2O. Before filtering, the R. venosa samples were additionally treated with 15-45 ml of 30% hydrogen peroxide (H2O2) for several minutes.
On a ZEISS Stemi 508 stereomicroscope, 435264-9200 (2.0x) objective with ZEISS ZEN core 3.6 software, protected with transparent polymethylmethacrylate plates, the microplastics were determined by type, color, and size directly from the glass Petri dishes, which are opened only to check the structure of the detected particles.
2.4. Tissue Preparation for Biochemical Analyses
Ten specimens of each species were used for the biochemical analyses. Before the analyses, the studied specimens were thawed and dissected. The soft tissues of invertebrates and fish gills and livers were extracted, and each tissue or organ was homogenized with cold 100 mM potassium phosphate (K-PO4) buffer, pH 7.4. The homogenates were centrifuged at 3000 rpm for 10 min at 4°C to obtain a post-nuclear fraction in which lipid peroxidation (LPO), and glutathione concentration (GSH) were measured. A part of the post-nuclear fraction was re-centrifuged at 12 000 rpm for 20 min at 4°C to obtain a post-mitochondrial supernatant in which were assayed the antioxidant enzyme activities: superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), glutathione reductase (GR), glutathione-S-transferase (GST), as well as acetylcholinesterase (AChE).
2.5. Biochemical Analyses
The protein concentrations of the tissue post-nuclear and post-mitochondrial fractions were measured according to Lowry et al. [80] using bovine serum albumin for generating the standard curve. The OS parameters were measured using kits, purchased from Sigma-Aldrich Co. LLC (USA): Lipid Peroxidation (MDA) Assay Kit MAK085, Glutathione Assay Kit CS0260, Superoxide dismutase Determination Kit 19160, Catalase Assay Kit CAT100, Glutathione Peroxidase Cellular Activity Assay CGP1, Glutathione Reductase Assay Kit GRSA, and Glutathione-S-Transferase Assay Kit, CS0410.
The AChE activity was measured using the method described by Ellman et al. [81], which is based on the production of thiocholine. The reaction mixture consisted of 100 mM K-PO₄ buffer at pH 8.0, 0.045 M acetylthiocholine iodide, 0.008 M 5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB), and an appropriate amount of tissue homogenate from either the invertebrates or fish liver and gills. Upon hydrolysis of acetylthiocholine by AChE, the resulting thiocholine reacted with DTNB to produce a yellow compound, 5-thio-2-nitrobenzoic acid. The absorbance of this product was measured at 412 nm, and AChE activity was expressed in units per milligram of protein (U/mg protein).
The Specific Oxidative Stress (SOS) index was calculated as the difference between the pro-oxidative (PrO) score and the antioxidant (AO) score (SOS = PrO – AO), based on oxidative stress markers measured in the studied specimens [82]. The PrO score represents the mean Z-score of the pro-oxidant marker LPO (PrO = mean (zLPO)), while the AO score is the average of the Z-scores of the antioxidant markers GSH, SOD, CAT, GPX, and GST (AO = mean (zGSH, zSOD, zCAT, zGPX, zGR, zGST)). The SOS values near zero indicate a balanced oxidative state.
2.6. Statistical Analysis
Statistical analyses followed Mosaheb et al. [83]. The significance of differences of means between groups was determined using Student’s t-statistic. The correlations between studied variables were assessed using the Spearman rank coefficient, and cluster analysis was applied to organize the observed data into meaningful structures of dependent variables. The calculations were carried out using the STATISTICA 10 package (StatSoft Inc., USA). For detecting the presence of significant thresholds of change in MPs accumulation in species individuals, the Sequential Three-step Analysis of Regime Shifts (STARS 6.3) was used [84,85,86].
3. Results
3.1. Morphological Characteristics of the Studied Species and Accumulated Microplastic Particles in Them
The indicator species selected for this study were both invertebrates (bivalves, gastropods, crustaceans) and vertebrates (fish) that belong to different trophic levels and are key species for different habitats.
The studied invertebrate species were significantly different in size and weight (Table 1). The analyses showed that the percentage of occurrence (%FO) of MPs in the soft tissues of the different invertebrates studied varied between 47% and 83% (Table 1). The lowest percentage was in R. venosa, where MPs were recorded in approximately every second individual (46.67%), and the highest percentage was in B. reticulatum (83.3%).
The accumulated MPs in the soft tissues of the key invertebrate species studied differed significantly. The estimation of the number of MPs per gram of soft tissue showed that they were most abundant in P. adspersus (0.99±1.09 particles/g ST), and least abundant in B. reticulatum (0.0033±0.0025 particles/g ST) (Table 1). Practically the alien species M. gigas had the highest accumulation of MPs.
There was no significant correlation present between the accumulated particles of MPs and the individual length and weight of the invertebrate species. The dependence of size on the accumulation of MPs was analyzed by correlation analyses of MPs content in the soft tissues and the size of the individuals of the different invertebrate species.
The studied fish significantly differed in their size and weight. The biggest individuals were those of M. batrahocephalus and the smallest were the individuals of S. sprattus (Table 2). The frequency of occurrence (%FO) of MPs in the GIT was significantly higher in M. batrachocephalus (53.3%) compared to S. sprattus (33.4%) (Table 2). Conversely, the %FO of MPs in the muscle tissue of M. batrachocephalus (26.7%) was significantly lower compared to S. sprattus (46.5%). The number of particles per gram of muscle in S. sprattus was significantly higher compared to M. batrachocephalus and M. barbatus (Table 2).
The only species where a significant correlation was present between the number of MPs/g muscle and the fish size was S. sprattus.
The transfer, bioaccumulation and the possible biomagnification of MPs in ecosystem food chains (and webs) and the ecological consequences are still not well understood. Due to the high variance of means in our study for the detecting of significant thresholds and shifts in MPs content in the studied species individuals, the Sequential Three-step Analysis of Regime Shifts (STARS 6.3) [84,85,86] was used. In this analysis, our previous data on MPs accumulation in individuals of C. glaucum and M. arenaria from the same regions (Varna Lake) [38] were also included. The results are graphically presented in Figure 2.
The graphic clearly indicated the presence of a significant increase in the MPs accumulation in the individuals of P. adspersus and the fish species studied. These results suggested the presence of differences in MPs accumulation in species from different habitats and different trophic levels.
3.2. Oxidative Stress Levels in the Studied Species
Under natural conditions, it is almost impossible to differentiate the specific ecological effects of MPs, and in this study, the stress-response effects by measuring OS levels were used as the basic component of the stress ecology approach.
The measured values of the OS biomarkers in the soft tissue of the studied invertebrate species are shown in Table 3. The values of the measured OS indicators varied among the studied invertebrate species. In P. adspersus, against the background of the highest content of MPs per gram of tissue, the highest LPO and the lowest values of antioxidants – non-enzymatic GSH and the enzymes SOD, CAT, GPx, and GR – were recorded, which indicate the presence of higher OS. The values of GST and AChE were also low. The lowest LPO and the highest values of GSH, SOD, CAT, GR, and GST were measured in R. venosa, which indicated the presence of low OS level, together with a relatively low accumulation of MPs per gram tissue (0.05±0.06 particles/g). Compared to native invertebrate species, the alien Pacific oyster showed a relatively high LPO, accompanied, however, by markedly higher values of GSH, SOD, and GR; as well as high CAT and GPx activities.
The lowest concentration of MPs per gram tissue was recorded in B. reticulatum, where also a relatively low level of LPO, high concentration of GSH, and high activity of the antioxidant enzymes were present, together with the highest AChE values. In the tissues of M. galloprovincialis, where average MPs values were present, relatively high LPO and low levels of GSH and antioxidant enzyme activities were found. In these mussels, there were statistically significantly lower GST and AChE activities compared to the other invertebrate species studied (Table 3).
For the integrative analysis of the degree of OS present and the specific accumulation of MPs per gram of tissue in the invertebrates, the percentage ratio of the individual indicators in the species studied was measured and is presented in Figure 3.
The graphic clearly shows that the recently introduced foreign species M. gigas was characterized by the highest MPs accumulation and high LPO. But here, high levels of the anti-stress system activity were found (SOD, CAT, GSH). Similarly, P. adspersus was also characterized by high MPs accumulation together with high stress level (high LPO and the lowest GSH, SOD, CAT, GR and GST activities) compared to the other studied species. In B. reticulatum low MPs accumulation was present together with low OS level (lowest LPO, highest GSH and GPx activities, and relatively high SOD, CAT, GR and GST activities) compared to the other studied invertebrates.
In fish, OS indicators were examined in the liver (Table 4a) and gills (Table 4b), as organs that are most susceptible to stress and in which it is possible for a larger amount of MPs to accumulate.

Table 4a.
Biometrics (W – Total weight; L – Total length; K – Fulton’s condition index) and oxidative stress biomarkers (mean±SD) in fish liver.
Table 4a.
Biometrics (W – Total weight; L – Total length; K – Fulton’s condition index) and oxidative stress biomarkers (mean±SD) in fish liver.
| biometrics | K | oxidative stress biomarkers | |||||||||
| W | L | LPO | GSH | SOD | CAT | GPx | GR | GST | AChE | ||
| Species | g | cm | nM/mg protein | ng/mg protein | U/mg protein | U/mg protein | U/mg protein | U/mg protein | U/mg protein | U/mg protein | |
| M. batrachocephalus | 73.0 | 19.5 | 0.98 | 7.45b | 32.9b,c | 80.5b,c | 2.02b | 74.6b,c | - | 225.9b,c | 17.8b,c |
| ±3.62 | ±0.35 | ±0.02 | ±0.83 | ±4.30 | ±1.94 | ±0.16 | ±6.25 | - | ±11.78 | ±0.52 | |
| M. barbatus | 9.60 | 9.44 | 1.04 | 0.93a,c | 135.1a | 86.1c | 8.00a,c | 27.5a | 17.42 | 476.8a,c | 50.1a,c |
| ±4.17 | ±1.40 | ±0.05 | ±0.19 | ±67.45 | ±14.89 | ±4.00 | ±9.15 | ±1.25 | ±16.65 | ±13.02 | |
| S. sprattus | 2.94 | 7.70 | 0.61 | 10.3a | 130.9a | 19.97a,b | 1.87b | 32.2a | 17.10 | 850.4a,b | 32.9a,b |
| ±1.12 | ±0.87 | ±0.04 | ±3.84 | ±13.95 | ±2.64 | ±0.27 | ±10.78 | ±2.62 | ±10.05 | ±8.12 | |
Significant difference from: a - M. batrachocephalus; b - M. barbatus; c - S. sprattus.
In the liver of M. barbatus (Table 4a), the lowest LPO levels were measured. This finding, together with the relatively high levels of antioxidant enzymes (including statistically significant higher CAT activity) compared to the other fish species studied, suggested low levels of OS. In M. batrachocephalus and S. sprattus, LPO levels were high. In M. batrachocephalus, a low concentration of GSH was measured, and the lowest GST activity, probably because GSH is a co-substrate in the enzymatic reaction. On the other hand, the GPx activity of these fish individuals was significantly higher than that of M. barbatus and S. sprattus, which suggests a possible activation of the recovery processes of GSH from GSSG under OS conditions. In S. sprattus, the lowest SOD activity was recorded compared to the other studied fish. In S. sprattus, GST activity was also statistically higher compared to both M. batrachocephalus and M. barbatus, which indicates an increased level of detoxification processes (Table 4a).
In the gills (Table 4b), there were no statistically significant differences in the LPO levels among the fish species. High antioxidant defense was present in M. batrachocephalus, including significantly higher concentration of GSH, and activities of CAT and GST, as well as AChE compared to those in M. barbatus and S. sprattus. In S. sprattus, low levels of GSH together with the lowest activity of SOD, GR, and also of AChE were measured compared to the other studied fish species.
Table 4b.
Biometrics (W – Total weight; L – Total length; K – Fulton’s condition index) and oxidative stress biomarkers (mean±SD) in fish gills.
Table 4b.
Biometrics (W – Total weight; L – Total length; K – Fulton’s condition index) and oxidative stress biomarkers (mean±SD) in fish gills.
| biometrics | K | oxidative stress biomarkers | |||||||||
| W | L | LPO | GSH | SOD | CAT | GPx | GR | GST | AChE | ||
| Species | g | cm | nM/mg protein | ng/mg protein | U/mg protein | U/mg protein | U/mg protein | U/mg protein | U/mg protein | U/mg protein | |
| M. batrachocephalus | 73.0 | 19.5 | 0.98 | 19.2 | 320.1b,c | 19.3c | 3.68b.c | 24.8b,c | - | 121.8b,c | 187.2b,c |
| ±3.62 | ±0.35 | ±0.02 | ±1.29 | ±33.50 | ±2.03 | ±0.58 | ±3.11 | - | ±3.16 | ±18.95 | |
| M. barbatus | 9.60 | 9.44 | 1.04 | 11.3 | 160.5a | 20.2c | 0.50a | 51.7a | 23.7 | 51.8a,c | 57.3a,c |
| ±4.17 | ±1.40 | ±0.05 | ±6.62 | ±62.66 | ±3.90 | ±0.20 | ±23.4 | ±5.2 | ±5.54 | ±16.2 | |
| S. sprattus | 2.94 | 7.70 | 0.61 | 20.5 | 141.7a | 8.49a,b | 0.55a | 47.6a | 13.7 | 86.2a,b | 17.3a,b |
| ±1.12 | ±0.87 | ±0.04 | ±0.47 | ±11.59 | ±1.25 | ±0.12 | ±6.81 | ±1.81 | ±6.08 | ±1.92 | |
Significant difference from: a - M. batrachocephalus; b - M. barbatus; c - S. sprattus.
The visualization of the ratios of OS parameters (means in liver and gills) and MPs accumulation of the studied fish are presented in Figure 4. It is evident from the data that S. sprattus exhibits the highest stress parameters values, as indicated by the highest LPO and the lowest/inhibited SOD and CAT activities, together with highest MPs accumulation in this species. In contrast, M. barbatus had the lowest MPs accumulation and stress, represented by the lowest LPO and the highest SOD and CAT activities compared to the other fish species studied.
In order to identify the possible interactions of MPs and the measured OS parameters with potential ecological consequences, cluster analysis was applied. The results revealed the close grouping of MPs accumulation with OS indicators in the key invertebrate species studied (Figure 5). The data of M. gigas is intentionally not presented as this is not a key species but is a foreign intruder recently beginning to settle down in the Bulgarian Black Sea part.
In M. galloprovicialis, the cluster analysis results (Figure 5a) showed very close relationship between MPs accumulation and CAT activity, which were also closely related to LPO. More distant were the relationships with a group including the glutathione-dependent enzymes GPx and GR. Overall, MPs and OS indicators formed a relatively compact group with different relations with others, where the most distant was the relation with GSH. Similarly, a cluster suggesting close relations between MPs and CAT activity, along with that of GR, was also present in P. adspersus (Figure 5c). These clusters in turn were also closely related to LPO and more distantly to SOD and GPx. The activities of GST and AChE formed a separate cluster. In P. adspersus, as in M. galloprovicialis, the relation with GSH was the most distant (Figure 5c). In R. venosa and B. reticulatum, a cluster suggesting very close relation between the accumulation of MPs and LPO was present, and in B. reticulatum, CAT activity was also involved (Figure 5b,d). In R. venosa, a cluster of close relations was present with the antioxidant enzymes (GPx, CAT, GR and SOD), all forming a common group. A separate cluster of GSH levels and AChE activity was formed, which was most distant from GST and the other studied OS parameters. In B. reticulatum, SOD and GPx activities were closely related to each other and formed a cluster also related to GR activity. A separate cluster was formed by GSH levels and GST activity, also related to MPs accumulation and the other OS indicators. In B. reticulatum, AChE activity had the most distant relationship with the other measured parameters, and also MPs (Figure 5d).
The cluster analyses of the grouping relations of MPs accumulation and the OS indicators measured in the liver of the studied fish species are presented in Figure 6 (a, b). The results show that the estimated accumulation of MPs in the fish species is grouped with the main OS biomarkers, and this suggests the presence of interactions.
In the liver of all the fish species studied, the MPs belonged to a distinct group, including also CAT, LPO, or GR (Figure 6a). Additionally, in S. sprattus associated with this group was also SOD and in M. barbatus - GPx. The AChE in M. batrahocephalus was also related to these OS indicators and thus belonged to this group (Figure 6a). In the fish species M. batrahocephalus and M. barbatus, the liver GST activity was most distant from the clusters of OS biomarkers, suggesting that the activity of this enzyme seemed not directly related to the presence of MPs. In S. sprattus, liver GSH concentration and AChE activity formed a separate cluster, which was only distantly related to the accumulated MPs and the other OS biomarkers (Figure 6a).
In the gills of the studied fish species, the grouping and the possible relations of MPs with closely associated OS variables were similar to those in the liver (Figure 6b).
In the gills of the three fish species studied, the MPs' presence formed a distinct and very compact cluster with CAT, thus clearly indicating the presence of functional relations. To this compact main group in the fish studied there were also other clusters of specific OS indicators including LPO, SOD, GR which were also more or less close to the MPs-CAT group.
To study the cumulative effect on OS levels, the values of the SOS index in the studied species were calculated (Table 5a,b).
Table 5a.
Calculated values of the SOS index of the studied species.
| M. batrachocephalus | M. barbatus | S. sprattus | M. galloprovincialis | R. venosa | B. reticulatum | P. adspersus | M. gigas | |
|---|---|---|---|---|---|---|---|---|
| PrO | 0.109 | -1.872 | 1.410 | -0.132 | -1.118 | -0.736 | 0.149 | 0.561 |
| AO | 0.804 | 0.863 | -1.335 | -0.891 | -0.681 | 0.357 | -0.457 | 0.627 |
| SOS | -0.694 | -2.736 | 2.745 | 0.758 | -0.437 | -1.093 | 0.606 | -0.066 |
Table 5b.
Classification of the stress level of the studied species according to the SOS index.
| Very low stress (<-2) |
Low stress (-1 to -2) |
Stress actively compensated (-1 to +1) |
Moderate stress (+1 to +2) |
High stress (+2<) |
|---|---|---|---|---|
| B. reticulatum |
R. venosa M. galloprovincialis M. gigas P. adspersus |
- | ||
| M. barbatus | M. batrachocephalus | - | S. sprattus |
Two of the studied species (B. reticulatum and M. barbatus) showed low levels of SOS (Table 5b). Most of the invertebrate species (R. venosa, M. galloprovincialis, M. gigas, P. adspersus) and the fish species M. batrachocephalus were in stress balance (SOS near zero), as the OS was actively compensated by the anti-stress cell system. The only species having a significantly high OS that the anti-stress system could not effectively compensate was S. sprattus (Table 5b). It is indicative that in the studied S. sprattus individuals, this coincides with the high level of accumulated MPs.
4. Discussion
The small size of MPs and their variable buoyancy make them easily accessible for uptake by organisms inhabiting different habitats and with different nutritional characteristics, indicating the “natural” trophic transfer of MPs in marine biota with implications for the broader food web, including humans [56,61].
The present study showed that MPs were present in most of the individuals of the species studied. The high percentage of MPs (%FO) in the soft tissues of B. reticulatum (83.33 %) and P. aspersus (70%) can be related to the number of plastics present in their habitats. Both species inhabit shallow sandy areas in algal and eelgrass beds, often in brackish water [87,88,89], and algae play a significant role in the retention of MPs and their removal from the water column [90]. The MPs' retention is achieved by their adhesion to algae or embedment/encrustation within the epibiont matrix of algae, thereby preventing MPs from migrating with water currents [90]. It was found that vegetated sites had much higher MPs than non-vegetated sites and that filamentous species contain more MPs due to the entanglement [91]. On the other hand, the significantly higher content of MPs in the tissues of P. aspersus compared to B. reticulatum was probably due also to species specific factors, including size, food preferences, metabolism, etc. Furthermore, P. aspersus are omnivorous invertebrates and, as such, they consume organisms that are likely to be highly loaded with MPs in their kelp habitat. Although B. reticulatum also inhabits kelp habitats, it is significantly smaller and thus consumes much smaller amount of food, and is also a microalgae herbivore unlike P. aspersus [92].
Concerning the fish species, the relatively higher percentage of occurrence (%FO) of MPs in the GIT of M. batrachocephalus and M. barbatus (about 50%) compared to S. sprattus was probably also due to their lifestyle. Both M. batrachocephalus and M. barbatus are demersal and less mobile fish, while S. sprattus is pelagic-neritic species [93]. Similar to other studies [94,95], in the present study there was a significant increase of ingested (in GIT) number of MPs related to fish lifestyle and size. The accumulation of MPs was found to be higher in demersal fish species than in the pelagic species [96]. It was assumed that their feeding habits most likely influenced the difference in MPs accumulation. Demersal fish are more stationary than pelagic fish and mostly feed on the prey species present on the sea floor. Specifically, M. batrachocephalus feeds mainly on fish [97], M. barbatus feeds on small benthic crustaceans, worms, and mollusks [98], and S. sprattus feeds on planktonic crustaceans. Furthermore, a study has reported the mean abundance of MPs in herbivorous fish species to be almost two times smaller than the abundance of MPs in carnivores and omnivores [99], and Rasta et al. [100] detected more MPs particles in omnivorous than in carnivorous fish. The established in this study almost 10 times more MPs per g muscle in S. sprattus, compared to the demersal fishes (M. batrachocephalus and M. barbatus), probably depends at least to some extent on the type of muscle. Demersal fish often have white meat, but pelagic fish typically have red flesh, indicative of the powerful swimming muscles they require. Tissues with high blood flow may have a higher likelihood of MPs being transported to them via the bloodstream. It has been found that different tissues and organs of fish have significant differences in MPs accumulation (concentrations of 0.84 ± 0.45 items/g in the digestive tract, 2.56 ± 0.73 items/g in the gills, and 0.3 ± 1.72 items/g in the muscle tissue) [95]. The organs of fish where MPs accumulation is generally higher include the gills and liver. Gills are significant hotspots for MPs accumulation compared to other organs, since the gills are used to filter water for respiration, and MPs can be trapped there [101].
The transfer and accumulation of MPs in species belonging to different levels of food chains (and webs) remains still not well studied. The applied STARS analysis (graphic in Figure 2) showed that in the individuals of P. adspersus and in the individuals of the fish species studied significantly higher numbers of MPs were present but this could hardly be interpreted as some kind of MPs trophic transfer from lower to higher trophic levels which is in agreement with other studies [56].
Irrespective of the pathways, intake of MPs can result in adverse physical and chemical impacts on organisms [41,42,43]. Due to the presence of multiple stressors in the marine environment it is almost impossible to assess specific effects of MPs on organism health [55,56]. In order to reveal the ecological effects of MPs in line with the stress ecology approach in this study, the effects of OS and the organism response were assessed.
Recent data show that MPs can contribute to OS in marine organisms [68,102,103] by generating excess reactive oxygen species (ROS) directly produced by the plastic particles themselves. MPs can release toxic chemicals such as phthalates, bisphenol A, and polycyclic aromatic hydrocarbons, which can induce ROS production [104]. Another mechanism is through the adsorbed pollutants onto the MPs’ surface, acting as a sink for heavy metals, organic compounds, and pesticides [102,105,106,107]. All of these pollutants can lead to ROS formation through various chemical reactions. For instance, it was found that when polycyclic aromatic hydrocarbons (PAHs) adsorb onto MPs surfaces, ROS can be produced via photochemical reactions under UV radiation [106,108]. Another mechanism could be the Fenton reaction, which involves a transition metal ion, such as iron or copper, to produce hydroxyl radicals [102,106].
In this study, quite different ratios of OS parameters (Figure 3 and Figure 4), together with different MPs accumulation in the studied species, were present. In P. adspersus high MPs accumulation was found together with high OS stress level. In B. reticulatum low MPs accumulation was present and OS was low compared to the other studied invertebrates. Interestingly, the recently settled in the Bulgarian Black Sea foreign species M. gigas had the highest number of accumulated MPs, but low OS due to the activity of the anti-stress cell system. In the studied fish species, the ratios of OS parameters (means in liver and gills) and MPs accumulation showed differences, but the highest OS in S. sprattus was accompanied by the highest MPs accumulation. Here, the GST activity was also high indicating an increased level of the detoxification processes, as it is a major phase II detoxification enzyme, detoxifying many endogenous compounds and breaking down xenobiotic substrates through conjugation of reduced glutathione with various substrates [109].
The cluster analyses we performed (Figure 5 and Figure 6) confirmed the presence of relations between MPs' content and the measured OS indicators. In most of the studied species, a closer relation between MPs' accumulation and CAT activity was observed. Thus, our findings support the suggestion that CAT can be used as a model enzyme to assess the biological effects of MPs [110]. Catalase is an essential enzyme that decomposes specifically H2O2, preventing the formation of the most damaging agent - the hydroxyl radicals. A study of the effects of polyvinyl chloride-MPs on hepatic antioxidant enzymes of showed a time-dependent decrease in the activities of SOD, GPx, and CAT and an increase in LPO levels [111]. Evidence also suggests that MPs may directly interact with CAT [110]. Therefore, MPs themselves or through toxic substances or pathogens carried on their surface are really a significant pro-oxidant and, upon prolonged exposure, can lead to depletion or inhibition of antioxidants in organisms [110,111].
The OS and response processes are also specific for different organs and have different effects. The gills and liver are most susceptible to OS. For example, the liver is a major metabolic and detoxifying organ of the body, and during these processes, ROS can be generated as byproducts [112]. The liver also plays a crucial role in immune responses by activating immune cells upon encountering pathogens or toxins [113]. This activation can trigger inflammation and the subsequent excess production of ROS. Disruption of the redox balance causes OS stress and contributes to cellular damage, which affects liver function modulation. The results obtained by us proved that in the gills of the studied fish species, grouping of MPs with closely associated OS variables was present. The gills are in direct contact with the aquatic environment, being the primary target of pollutants, are richly blood-supplied, and have a high surface area, where gas exchange and filtration occur [114]. Research on contamination in marine fishes found that MPs were present in the gills, dorsal muscle, and gastrointestinal tract. MPs contaminated fish had increased LPO levels in the brain, dorsal muscle, and gills, and higher AChE activity in the brain [115].
Recently, efforts to determine the ecological impacts of MPs pollutants have increased because of the increasing plastic contamination of the marine environment. However, recent review papers and reports have identified current research gaps [116,117,118]. The ecological risks of plastic contamination can be defined as the likelihood of adverse health effects on biota and their ecological consequences as a result of the organisms’ exposure to MPs. The changes caused by OS at the cellular level can subsequently affect higher hierarchical levels, which is defined as “stress ecology” [64,65].
In this study, we used a previously introduced SOS index as a general composite risk indicator to measure the complex effects of multiple environmental stressors, including plastics, on the studied species (Table 5). Two of the species (B. reticulatum and M. barbatus) showed low levels of OS. Most of the species (R. venosa, M. galloprovincialis, M. gigas, P. adspersus, M. batrachocephalus) were in stress balance, as the OS was actively compensated by the anti-stress cell system. A really high OS was present only in S. sprattus which the antistress system seems to not effectively compensate. This is also the species with the highest MPs accumulation. The SOS index proved that it can serve as an integral measure of the OS level in the organisms. However, the SOS index cannot differentiate the specific factor/s causing OS changes, which also applies to MPs accumulation.
5. Conclusions
Much remains unknown regarding the environmental consequences of MPs pollution, and more detailed studies are required to assess the ecological and human health risks. The specific OS (SOS) index showed that OS levels were linked to both habitat conditions and species-specific antioxidant defense. These findings confirmed the priority role of environmental conditions (including MPs pollution) in OS, and the evolutionarily developed adaptation capacity and resilience to induce stress in marine organisms. This study emphasizes the increasing need for specific monitoring of MPs and OS of marine organisms to better assess ecological risks. Further research is strongly recommended in order to determine ecosystem-level effects of MPs.
Author Contributions
Conceptualization, N.Ch. and A.A.; methodology, A.A. and S.M.; software, N.Ch.; formal analysis, A.A., S.M., E.Tz., M.A, G.Pr., G.P., V.D., K.S., M.G., H.S. and E.H.; investigation, A.A., S.M., E.Tz., M.A, G.Pr., and G.P.; resources, P.I. and A.A.; data curation, N.Ch.; writing—original draft preparation, N.Ch. and A.A.; writing—review and editing, N.Ch. and A.A.; visualization, A.A.; supervision, N.Ch. and A.A.; project administration, P.I. and A.A.; funding acquisition, P.I., N.Ch and A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Fund, Sofia, Bulgaria, grant number № КP-06-Н81/2.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Researchers wishing to access the data used in this study can make a request to the corresponding authors.
Acknowledgments
This work was supported by Grant № КP-06-Н81/2, National Science Fund, Sofia, Bulgaria.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| AChE | Acetylcholinesterase |
| AO | Antioxidant score |
| CAT | Catalase |
| FO | Frequency of occurrence |
| GIT | Gastrointestinal tract |
| GPx | Glutathione peroxidase |
| GR | Glutathione Reductase |
| GSH | Glutathione |
| GST | Glutathione-S-Transferase |
| L | Total length |
| LPO | Lipid Peroxidation |
| MPs | Microplastics |
| OS | Oxidative Stress |
| PrO | Pro-oxidative score |
| ROS | Reactive Oxygen Species |
| SOD | Superoxide Dismutase |
| SOS | Specific Oxidative Stress Index |
| ST | Soft tissues |
| W | Total weight |
References
- Statista. Available online: https://www.statista.com.
- Plastic Pollution Coalition. Available online: https://plastic-pollution.org/.
- González-Fernández, D.; Pogojeva, M.; Hanke, G.; Machitadze, N.; Kotelnikova, Y.; Tretiak, I.; Savenko, O.; Gelashvili, N.; Bilashvili, K.; Kulagin, D.; et al. Anthropogenic Litter Input through Rivers in the Black Sea. In Marine Litter in the Black Sea; Aytan, Ü., Pogojeva, M., Simeonova, A., Eds.; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2020; Publication No: 56. [Google Scholar]
- Moncheva, S.; Stefanova, K.; Krastev, A.; Apostolov, A.; Bat, L.; Sezgin, M.; Sahin, F.; Timofte, F. Marine Litter Quantification in the Black Sea: A Pilot Assessment. Turk. J. Fish. Aquat. Sci. 2016, 16, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Uzer, U.; Yildiz, T.; Karakulak, F.S. Distribution and Composition of Marine Litter on Seafloor in the Western Black Sea, Turkey. In Marine Litter in the Black Sea; Aytan, Ü., Pogojeva, M., Simeonova, A., Eds.; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2020; Publication No: 56. [Google Scholar]
- Kasapoğlu, N.; Dağtekin, M.; İlhan, S.; Erik, G.; Özsandikçi, U.; Büyükdeveci, F. Distribution and Composition of Seafloor Marine Litter in the Southeastern Black Sea. In Marine Litter in the Black Sea; Aytan, Ü., Pogojeva, M., Simeonova, A., Eds.; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2020; Publication No: 56. [Google Scholar]
- GESAMP. Guidelines for the Monitoring and Assessment of Plastic Litter in the Ocean; GESAMP Reports Stud. 2019, 99; GESAMP: London, UK, 2019; p. 130. [Google Scholar]
- Lusher, A. Microplastics in the Marine Environment: Distribution, Interactions and Effects. In Marine Anthropogenic Litter; Springer: Cham, Switzerland, 2015; pp. 245–307. [Google Scholar]
- Savuca, A.; Nicoara, M.N.; Faggio, C. Comprehensive Review Regarding the Profile of the Microplastic Pollution in the Coastal Area of the Black Sea. Sustainability 2022, 14, 14376. [Google Scholar] [CrossRef]
- Simeonova, A.; Chuturkova, R. Marine Litter Accumulation Along the Bulgarian Black Sea Coast: Categories and Predominance. Waste Manag. 2019, 84, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Simeonova, A.; Chuturkova, R.; Toneva, D.; Tsvetkov, M. Plastic pollution along the Bulgarian Black Sea coast: Current status and trends. In Marine Litter in the Black Sea; Aytan, Ü., Pogojeva, M., Simeonova, A., Eds.; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2020; Publication No: 56. [Google Scholar]
- Miladinova, S.; Macias, D.; Stips, A.; Garcia-Gorriz, E. Identifying Distribution and Accumulation Patterns of Floating Marine Debris in the Black Sea. Mar. Pollut. Bull. 2020, 153, 110964. [Google Scholar] [CrossRef] [PubMed]
- Strokal, M.; Vriend, P.; Bak, M.P.; et al. River Export of Macro- and Microplastics to Seas by Sources Worldwide. Nat. Commun. 2023, 14, 4842. [Google Scholar] [CrossRef]
- Săvucă, A.; Nicoara, M.N.; Faggio, C. Comprehensive review regarding the profile of the microplastic pollution in the coastal area of the Black Sea. Sustainability 2022, 14, 14376. [Google Scholar] [CrossRef]
- Aytan, Ü.; Şentürk, Y.; Esensoy, F. B.; Öztekin, A.; Ağırbaş, E.; Valente, A. Microplastic pollution along the southeastern Black Sea. In book: Aytan, Ü., Pogojeva, M., Simeonova, A. (Eds.) 2020, Marine Litter in the Black Sea. Publisher: Turkish Marine Research Foundation (TUDAV) Publication No: 56, Istanbul, Turkey.
- Cincinelli, A.; Scopetani, C.; Chelazzi, D.; Martellini, T.; Pogojeva, M.; Slobodnik, J. Microplastics in the Black Sea Sediments. Sci. Total Environ. 2021, 760, 143898. [Google Scholar] [CrossRef]
- Pojar, I.; Stănică, A.; Stock, F.; Kochleus, C.; Schultz, M.; Bradley, C. Sedimentary microplastic concentrations from the Romanian Danube River to the Black Sea. Sci. Rep. 2021, 11, 2000. [Google Scholar] [CrossRef]
- Terzi, Y.; Gedik, K.; Eryaşar, A.R.; Öztürk, R.Ç.; Şahin, A.; Yılmaz, F. Microplastic contamination and characteristics spatially vary in the southern Black Sea beach sediment and sea surface water. Mar. Pollut. Bull. 2022, 174, 113228. [Google Scholar] [CrossRef]
- Mutlu, T.; Minaz, M.; Baytaşoğlu, H.; Gedik, K. Microplastic pollution in stream sediments discharging from Türkiye’s eastern Black Sea basin. Chemosphere 2024, 352, 141496. [Google Scholar] [CrossRef]
- Bobchev, N.; Berov, D.; Klayn, S.; Karamfilov, V. High microplastic pollution in marine sediments associated with urbanised areas along the SW Bulgarian Black Sea coast. Mar. Pollut. Bull. 2024, 209, 117150. [Google Scholar] [CrossRef] [PubMed]
- Berov, D.; Klayn, St. Microplastics and floating litter pollution in Bulgarian Black Sea coastal waters. Mar. Pollut. Bull. 2020, 156, 111225. [Google Scholar] [CrossRef]
- Pojar, I.; Kochleus, C.; Dierkes, G.; Ehlers, S.M.; Reifferscheid, G.; Stock, F. Quantitative and qualitative evaluation of plastic particles in surface waters of the Western Black Sea. Environ. Pollut. 2021, 268, 115724. [Google Scholar] [CrossRef] [PubMed]
- Bat, L.; Öztekin, A. Microplastic Pollution in the Black Sea: An Overview of the Current Situation. In Microplastic Pollution. Emerging Contaminants and Associated Treatment Technologies; Hashmi, M., Ed.; Springer: Cham, Switzerland, 2022; pp. 149–171. [Google Scholar] [CrossRef]
- Georgieva, S.K.; Peteva, Z.V.; Stancheva, M.D. Evaluation of abundance of microplastics in the Bulgarian coastal waters. In Actual Problems of Ecology; Chankova, S., Danova, K., Beltcheva, M., Radeva, G., Petrova, V., Vassilev, K., Eds.; BioRisk 2023, 20, 59–69. [CrossRef]
- Öztekin, A.; Bat, L. Microlitter Pollution in Sea Water: A Preliminary Study from Sinop Sarikum Coast of the Southern Black Sea. In Proceedings of the International Symposium on Marine Sciences; 2017. [Google Scholar]
- Öztekin, A.; Üstün, F.; Bat, L.; Tabak, A. Microplastic Contamination of the Seawater in the Hamsilos Bay of the Southern Black Sea. Water Air Soil Pollut. 2024, 235, 325. [Google Scholar] [CrossRef]
- Aytan, U.; Valente, A.; Senturk, Y.; Usta, R.; Esensoy Sahin, F.B.; Mazlum, R.E.; Agırbas, E. First evaluation of neustonic microplastics in Black Sea waters. Mar. Environ. Res. 2016, 119, 22–30. [Google Scholar] [CrossRef]
- Aytan, U.; Esensoy, F.B.; Senturk, Y.; Arifoğlu, E.; Karaoğlu, K.; Ceylan, Y.; Valente, A. Plastic Occurrence in Commercial Fish Species of the Black Sea. Turk. J. Fish. Aquat. Sci. 2022, 22, TRJFAS20504. [Google Scholar] [CrossRef]
- Aytan, U.; Esensoy, F.B.; Senturk, Y. Microplastic ingestion and egestion by copepods in the Black Sea. Sci. Total Environ. 2022, 806, 150921. [Google Scholar] [CrossRef]
- Aytan, U.; Esensoy, F.B.; Senturk, Y.; Güven, O.; Karaoğlu, K.; Erbay, M. Plastic occurrence in fish caught in the highly industrialized Gulf of İzmit (Eastern Sea of Marmara, Türkiye). Chemosphere 2023, 324, 138317. [Google Scholar] [CrossRef]
- Gedik, K.; Eryaşar, A.R. Microplastic pollution profile of Mediterranean mussels (Mytilus galloprovincialis) collected along the Turkish coasts. Chemosphere 2020, 260, 127570. [Google Scholar] [CrossRef]
- Şentürk, Y.; Esensoy, F.B.; Öztekin, A.; Aytan, Ü. Microplastics in bivalves in the southern Black Sea. In Marine Litter in the Black Sea; Aytan, Ü., Pogojeva, M., Simeonova, A., Eds.; Turkish Marine Research Foundation (TUDAV), Publication No. 56: Istanbul, Turkey, 2020; p. 303. [Google Scholar]
- Şentürk, Y.; Emanet, M.; Ceylan, Y.; Aytan, U. The First Evidence of Microplastics Occurrence in Greater Pipefish (Syngnathus acus Linnaeus, 1758) in the Black Sea. Turk. J. Fish. Aquat. Sci. 2023, 23, TRJFAS23764. [Google Scholar] [CrossRef]
- Gedik, K.; Gozler, A.M. Hallmarking microplastics of sediments and Chamelea gallina inhabiting Southwestern Black Sea: A hypothetical look at consumption risks. Mar. Pollut. Bull. 2022, 174, 113252. [Google Scholar] [CrossRef]
- Filimon, A.; Ciucă, A.-M.; Harcotă, G.-E.; Stoica, E. Preliminary Study on Microplastic Contamination in Black Sea Cetaceans: Gastrointestinal Analysis of Phocoena phocoena relicta and Tursiops truncatus ponticus. Animals 2024, 14, 886. [Google Scholar] [CrossRef] [PubMed]
- Alexandrova, A.; Ignatova-Ivanova, T.V.; Bachvarova, D.G.; Toschkova, S.G.; Doichinov, A.H.; Ibryamova, S.F.; Chipev, N.H. Pilot Screening and Assessment of Microplastic Bioaccumulation in Wedge Clams Donax trunculus Linnaeus, 1758 (Bivalvia) from the Bulgarian Black Sea Coast. Acta Zool. Bulg. 2022, 74, 569–577. [Google Scholar]
- Ibryamova, S.; Toschkova, S.; Bachvarova, D.; Lyatif, A.; Stanachkova, E.; Ivanov, R.; Natchev, N.; Ignatova-Ivanova, T. Assessment of the bioaccumulation of microplastics in the Black Sea mussel Mytilus galloprovincialis L., 1819. J. IMAB 2022, 28, 4676–4682. [Google Scholar] [CrossRef]
- Mihova, M.; Toschkova, S.; Bachvarova, D.; Ibryamova, S.; Ignatova-Ivanova, T.; Natchev, N. Microplastic Uptake by Mya arenaria Linnaeus, 1758, Mytilus galloprovincialis Lamarck, 1819 and Cerastoderma glaucum (Bruguière, 1789) (Bivalvia) from Varna Lake, Bulgaria. Acta Zool. Bulg. 2024, 76, 367–374. [Google Scholar] [CrossRef]
- Toschkova, S.; Ibryamova, S.; Bachvarova, D.Ch.; Koynova, T.; Stanachkova, E.; Ivanov, R.; Natchev, N.; Ignatova-Ivanova, T. The assessment of the bioaccumulation of microplastics in key fish species from the Bulgarian aquatory of the Black Sea. BioRisk 2024, 22, 17–31. [Google Scholar] [CrossRef]
- Pramatarov, G.I.; Tsvetanova, E.R.; Ilinkin, V.M.; Andreeva, M.N.; Alexandrova, A.V.; Chipev, N.H. Effects of Microplastics and Metal Pollution on Bivalves from the Bulgarian Black Sea Sublittoral, with Comments on their Adaptive Capacity. Acta Zool. Bulg. 2025, 77, 107–119. [Google Scholar] [CrossRef]
- Xu, X.Y.; Lee, W.T.; Chan, A.K.Y.; Lo, H.S.; Shin, P.K.S.; Cheung, S.G. Microplastic ingestion reduces energy intake in the clam Atactodea striata. Mar. Pollut. Bull. 2017, 124, 798–802. [Google Scholar] [CrossRef]
- Barboza, L.G.A.; Vieira, L.R.; Branco, V.; Carvalho, C.; Guilhermino, L. Microplastics increase mercury bioconcentration in gills and bioaccumulation in the liver, and cause oxidative stress and damage in Dicentrarchus labrax juveniles. Sci. Rep. 2018, 8, 15655. [Google Scholar] [CrossRef]
- Gardon, T.; Reisser, C.; Soyez, C.; Quillien, V.; Le Moullac, G. Microplastics affect energy balance and gametogenesis in the pearl oyster Pinctada margaritifera. Environ. Sci. Technol. 2018, 52, 5277–5286. [Google Scholar] [CrossRef] [PubMed]
- von Moos, N.; Burkhardt-Holm, P.; Köhler, A. Uptake and effects of microplastics on cells and tissue of the blue mussel Mytilus edulis L. after an experimental exposure. Environ. Sci. Technol. 2012, 46, 11327–11335. [Google Scholar] [CrossRef] [PubMed]
- Avio, C.G.; Gorbi, S.; Milan, M.; Benedetti, M.; Fattorini, D.; d’Errico, G.; Pauletto, M.; Bargelloni, L.; Regoli, F. Pollutants bioavailability and toxicological risk from microplastics to marine mussels. Environ. Pollut. 2015, 198, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Avio, C.G.; Gorbi, S.; Regoli, F. Plastics and microplastics in the oceans: From emerging pollutants to emerged threat. Mar. Environ. Res. 2017, 128, 2–11. [Google Scholar] [CrossRef]
- Abbasi, S.; Soltani, N.; Keshavarzi, B.; Moore, F.; Turner, A.; Hassanaghaei, M. Microplastics in different tissues of fish and prawn from the Musa Estuary, Persian Gulf. Chemosphere 2018, 205, 80–87. [Google Scholar] [CrossRef]
- Kadac-Czapska, K.; Ośko, J.; Knez, E.; Grembecka, M. Microplastics and Oxidative Stress—Current Problems and Prospects. Antioxidants 2024, 13, 579. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, Y.; Deng, Y.; Jiang, W.; Zhao, Y.; Geng, J.; et al. Uptake and accumulation of polystyrene microplastics in zebrafish (Danio rerio) and toxic effects in liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef]
- Karami, A.; Romano, N.; Galloway, T.; Hamzah, H. Virgin microplastics cause toxicity and modulate the impacts of phenanthrene on biomarker responses in African catfish (Clarias gariepinus). Environ. Res. 2016, 151, 58–70. [Google Scholar] [CrossRef]
- Lo, H.K.A.; Chan, K.Y.K. Negative effects of microplastic exposure on growth and development of Crepidula onyx. Environ. Pollut. 2018, 233, 588–595. [Google Scholar] [CrossRef]
- Chapron, L.; Peru, E.; Engler, A.; Ghiglione, J.F.; Meistertzheim, A.L.; Pruski, A.M.; et al. Macro- and microplastics affect cold-water corals growth, feeding and behaviour. Sci. Rep. 2018, 8, 15299. [Google Scholar] [CrossRef]
- Cole, M.; Lindeque, P.; Fileman, E.; Halsband, C.; Galloway, T.S. The impact of polystyrene microplastics on feeding, function and fecundity in the marine copepod Calanus helgolandicus. Environ. Sci. Technol. 2015, 49, 1130–1137. [Google Scholar] [CrossRef] [PubMed]
- Besseling, E.; Wegner, A.; Foekema, E.M.; van den Heuvel-Greve, M.J.; Koelmans, A.A. Effects of microplastic on fitness and PCB bioaccumulation by the lugworm Arenicola marina (L.). Environ. Sci. Technol. 2013, 47, 593–600. [Google Scholar] [CrossRef]
- Koelmans, A.A.; Besseling, E.; Foekema, E.; Kooi, M.; Mintenig, S.; Ossendorp, B.C.; et al. Risks of plastic debris: Unravelling fact, opinion, perception, and belief. Environ. Sci. Technol. 2017, 51, 11513–11519. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.E.; Hamann, M.; Kroon, F.J. Bioaccumulation and biomagnification of microplastics in marine organisms: A review and meta-analysis of current data. PLoS ONE 2020, 15, e0240792. [Google Scholar] [CrossRef] [PubMed]
- USEPA. Microplastics Expert Workshop Report; U.S. Environmental Protection Agency: Washington, DC, USA, 2017; Available online: https://www.epa.gov/sites/default/files/2018-03/documents/microplastics_expert_workshop_report_final_12-4-17.pdf(accessed on [Insert Date]).
- GESAMP. Proceedings of the GESAMP International Workshop on Assessing the Risks Associated with Plastics and Microplastics in the Marine Environment; Kershaw, P.J., Carney Almroth, B., Villarrubia-Gómez, P., Koelmans, A.A., Gouin, T., Eds.; IMO/FAO/UNESCO-IOC/UNIDO/WMO/IAEA/UN/UNEP/UNDP/ISA Joint Group of Experts on the Scientific Aspects of Marine Environmental Protection: London, UK, 2020; GESAMP Reports and Studies No. 103, 68 pp. [Google Scholar]
- Hermabessiere, L.; Dehaut, A.; Paul-Pont, I.; Lacroix, C.; Jezequel, R.; Soudant, P.; Duflos, G. Occurrence and Effects of Plastic Additives on Marine Environments and Organisms: A Review. Chemosphere 2017, 182, 781–793. [Google Scholar] [CrossRef]
- USEPA. Guidelines for Exposure Assessment; Risk Assessment Forum, U.S. Environmental Protection Agency: Washington, DC, USA, 1992; EPA/600/Z-92/001. [Google Scholar]
- Farrell, P.; Nelson, K. Trophic Level Transfer of Microplastic: Mytilus edulis (L.) to Carcinus maenas (L.). Environ. Pollut. 2013, 177, 1–3. [Google Scholar] [CrossRef]
- Nelms, S.E.; Galloway, T.S.; Godley, B.J.; Jarvis, D.S.; Lindeque, P.K. Investigating Microplastic Trophic Transfer in Marine Top Predators. Environ. Pollut. 2018, 238, 999–1007. [Google Scholar] [CrossRef]
- Zhao, S.; Ward, J.E.; Danley, M.; Mincer, T.J. Field-Based Evidence for Microplastic in Marine Aggregates and Mussels: Implications for Trophic Transfer. Environ. Sci. Technol. 2018, 52, 11038–11048. [Google Scholar] [CrossRef]
- Van Straalen, N.M. Ecotoxicology Becomes Stress Ecology. Environ. Sci. Technol. 2003, 37, 324A–330A. [Google Scholar] [CrossRef]
- Steinberg, C.E.W. Stress Ecology: Environmental Stress as Ecological Driving Force and Key Player in Evolution; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar] [CrossRef]
- Hu, M.; Palić, D. Micro- and Nano-Plastics Activation of Oxidative and Inflammatory Adverse Outcome Pathways. Redox Biol. 2020, 37, 101620. [Google Scholar] [CrossRef]
- D’Costa, A.H. Microplastics in Decapod Crustaceans: Accumulation, Toxicity and Impacts, a Review. Sci. Total Environ. 2022, 832, 154963. [Google Scholar] [CrossRef] [PubMed]
- Dovzhenko, N.V.; Slobodskova, V.V.; Mazur, A.A.; Kukla, S.P.; Istomina, A.A.; Chelomin, V.P.; Beskhmelnov, D.D. Oxidative Stress in Mussel Mytilus trossulus Induced by Different-Sized Plastics. J. Xenobiotics 2024, 14, 1826–1835. [Google Scholar] [CrossRef] [PubMed]
- Alomar, C.; Sureda, A.; Capó, X.; Guijarro, B.; Tejada, S.; Deudero, S. Microplastic Ingestion by Mullus surmuletus and Its Potential for Causing Oxidative Stress. Environ. Res. 2017, 159, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, C.; Esteban, M.Á.; Cuesta, A. Dietary Administration of PVC and PE Microplastics Produces Histological Damage, Oxidative Stress and Immunoregulation in European Sea Bass (Dicentrarchus labrax L.). Fish Shellfish Immunol. 2019, 95, 574–583. [Google Scholar] [CrossRef]
- Solomando, A.; Capó, X.; Alomar, C.; Álvarez, E.; Compa, M.; Valencia, J.M.; Pinya, S.; Deudero, S.; Sureda, A. Long-Term Exposure to Microplastics Induces Oxidative Stress and a Pro-Inflammatory Response in the Gut of Sparus aurata. Environ. Pollut. 2020, 266, 115295. [Google Scholar] [CrossRef]
- Kim, J.H.; Yu, Y.B.; Choi, J.H. Toxic Effects on Bioaccumulation, Hematological Parameters, Oxidative Stress, Immune Responses and Neurotoxicity in Fish Exposed to Microplastics: A Review. J. Hazard. Mater. 2021, 413, 125423. [Google Scholar] [CrossRef]
- Bowmer, T.; Kershaw, P. Proceedings of the GESAMP International Workshop on Micro-Plastic Particles as a Vector in Transporting Persistent, Bio-Accumulating and Toxic Substances in the Oceans; GESAMP Reports & Studies; UNESCO-IOC: Paris, France, 2010; 68 pp. [Google Scholar]
- Andrady, A.L. Microplastics in the Marine Environment. Mar. Pollut. Bull. 2011, 62, 1596–1605. [Google Scholar] [CrossRef]
- Bowley, J.; Baker-Austin, C.; Porter, A.; Hartnell, R.; Lewis, C. Oceanic Hitchhikers—Assessing Pathogen Risks from Marine Microplastic. Trends Microbiol. 2021, 29, 107–116. [Google Scholar] [CrossRef]
- Foley, C.J.; Feiner, Z.S.; Malinich, T.D.; Höök, T.O. A Meta-Analysis of the Effects of Exposure to Microplastics on Fish and Aquatic Invertebrates. Sci. Total Environ. 2018, 631–632, 550–559. [Google Scholar] [CrossRef]
- Guzzetti, E.; Sureda, A.; Tejada, S.; Faggio, C. Microplastic in Marine Organism: Environmental and Toxicological Effects. Environ. Toxicol. Pharmacol. 2018, 64, 164–171. [Google Scholar] [CrossRef]
- Secci, G.; Parisi, G. From Farm to Fork: Lipid Oxidation in Fish Products. A Review. Ital. J. Anim. Sci. 2016, 15, 124–136. [Google Scholar] [CrossRef]
- Nash, R.D.; Valencia, A.; Geffen, A.J. The Origin of Fulton’s Condition Factor: Setting the Record Straight. Fisheries 2006, 31, 236–238. [Google Scholar]
- Lowry, O.; Rosebrough, N.; Farr, A.; Randall, R. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Zlateva, I.; Raykov, V.; Alexandrova, A.; Ivanova, P.; Chipev, N.; Stefanova, K.; Dzhembekova, N.; Doncheva, V.; Slabakova, V.; Stefanova, E.; et al. Effects of anthropogenic and environmental stressors on the current status of red mullet (Mullus barbatus L., 1758) populations inhabiting the Bulgarian Black Sea waters. Nature Conserv. 2023, 54, 55–79. [Google Scholar] [CrossRef]
- Mosaheb, I.; Hiroaki, T.; Ruby, R.M.; Teeluck, S.; Kawasaki, H.; Chineah, V. Basic Biostatistics for Marine Biologists; Coastal Fisheries Resources & Environment Conservation Project: 2001. ISBN 99903-22-12-0.
- Rodionov, S. A brief overview of the regime shift detection methods. In Large-Scale Disturbances (Regime Shifts) and Recovery in Aquatic Ecosystems: Challenges for Management Toward Sustainability; Velikova, V., Chipev, N., Eds.; UNESCO-ROSTE/BAS Workshop on Regime Shifts: Varna, Bulgaria, 2005; pp. 17–24. [Google Scholar]
- Rodionov, S. A Sequential Method of Detecting Abrupt Changes in the Correlation Coefficient and Its Application to Bering Sea Climate. Climate 2015, 3, 474–491. [Google Scholar] [CrossRef]
- Regime Shift Test Website. Available online: https://sites.google.com/view/regime-shift-test (accessed on 26 May 2025).
- Holthuis, L.B. FAO Species Catalogue: Shrimps and Prawns of the World. An Annotated Catalogue of Species of Interest to Fisheries; FAO Fisheries Synopsis, 1980; Volume 1, p. 271. [Google Scholar]
- Berglund, A. Different reproductive success at low salinity determines the estuarine distribution of two Palaemon prawn species. Ecol. Ser. 1985, 8, 49–52. [Google Scholar] [CrossRef]
- Gonzalez-Ortegon, E.; Sargent, P.; Pohle, G.; Martinez-Lage, A. The Baltic prawn Palaemon adspersus Rathke, 1837 (Decapoda, Caridea, Palaemonidae): First record, possible establishment, and illustrated key of the subfamily Palaemoninae in northwest Atlantic waters. Aquat. Invasions 2015, 10, 299–312. [Google Scholar] [CrossRef]
- Jiang, Y.; Niu, S.; Wu, J. The role of algae in regulating the fate of microplastics: A review for processes, mechanisms, and influencing factors. Sci. Total Environ. 2024, 949, 175227. [Google Scholar] [CrossRef]
- Huang, S.; Jiang, R.; Craig, N.J.; Deng, H.; He, W.; Li, J.Y.; Su, L. Accumulation and re-distribution of microplastics via aquatic plants and macroalgae—A review of field studies. Mar. Environ. Res. 2023, 187, 105951. [Google Scholar] [CrossRef]
- SeaLifeBase: Bittium reticulatum. Available online: https://www.sealifebase.se/summary/Bittium-reticulatum.html (accessed on 26 May 2025).
- FishBase: Sprattus sprattus. Available online: https://www.fishbase.se/summary/sprattus-sprattus.html (accessed on 26 May 2025).
- Filgueiras, A.V.; Preciado, I.; Cartón, A.; Gago, J. Microplastic ingestion by pelagic and benthic fish and diet composition: A case study in the NW Iberian shelf. Mar. Pollut. Bull. 2020, 160, 111623. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.B.; Pingki, F.H.; Azad, M.A.S.; Nur, A.-A.U.; Banik, P.; Paray, B.A.; Arai, T.; Yu, J. Microplastics in different tissues of a commonly consumed fish, Scomberomorus guttatus, from a large subtropical estuary: Accumulation, characterization, and contamination assessment. Biology 2023, 12, 1422. [Google Scholar] [CrossRef] [PubMed]
- Keerthika, K.; Padmavathy, P.; Rani, V.; Jeyashakila, R.; Aanand, S.; Kutty, R.; Tamilselvan, R.; Subash, P. Microplastics accumulation in pelagic and benthic species along the Thoothukudi coast, South Tamil Nadu, India. Mar. Pollut. Bull. 2023, 189, 114735. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J. Gobiidae. In Fishes of the Northeastern Atlantic and the Mediterranean, Volume 3; Whitehead, P.J.P., Bauchot, M.-L., Hureau, J.-C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; pp. 1019–1085. [Google Scholar]
- Hureau, J.-C. Mullidae. In Fishes of the Northeastern Atlantic and the Mediterranean, Volume 2; Whitehead, P.J.P., Bauchot, M.-L., Hureau, J.-C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; pp. 877–882. [Google Scholar]
- Bora, G.; Phukan, B.; Talukdar, A.; Ahmed, I.; Chutia, S.J.; Gogoi, R.; Sarma, J.; Ali, A.; Gowala, T.; Xavier, K.A.M. Assessment and quantification of microplastic contamination in fishes with different food habits from Beel wetlands. J. Hazard. Mater. Adv. 2025, 17, 100618. [Google Scholar] [CrossRef]
- Rasta, M.; Sattari, M.; Taleshi, M.S.; Namin, J.I. Microplastics in Different Tissues of Some Commercially Important Fish Species from Anzali Wetland in the Southwest Caspian Sea, Northern Iran. Mar. Pollut. Bull. 2021, 169, 112479. [Google Scholar] [CrossRef]
- Yin, X.; Wu, J.; Liu, Y.; Chen, X.; Xie, C.; Liang, Y.; Li, J.; Jiang, Z. Accumulation of Microplastics in Fish Guts and Gills from a Large Natural Lake: Selective or Non-Selective? Environ. Pollut. 2022, 309, 119785. [Google Scholar] [CrossRef]
- Geremia, E.; Muscari Tomajoli, M.T.; Murano, C.; Petito, A.; Fasciolo, G. The Impact of Micro- and Nanoplastics on Aquatic Organisms: Mechanisms of Oxidative Stress and Implications for Human Health—A Review. Environments 2023, 10, 161. [Google Scholar] [CrossRef]
- Subaramaniyam, U.; Allimuthu, R.S.; Vappu, S.; Ramalingam, D.; Balan, R.; Paital, B.; Panda, N.; Rath, P.K.; Ramalingam, N.; Sahoo, D.K. Effects of Microplastics, Pesticides and Nano-Materials on Fish Health, Oxidative Stress and Antioxidant Defense Mechanism. Front. Physiol. 2023, 14, 1217666. [Google Scholar] [CrossRef]
- Urli, S.; Corte Pause, F.; Crociati, M.; Baufeld, A.; Monaci, M.; Stradaioli, G. Impact of Microplastics and Nanoplastics on Livestock Health: An Emerging Risk for Reproductive Efficiency. Animals 2023, 13, 1132. [Google Scholar] [CrossRef]
- Hu, L.; Zhao, Y.; Xu, H. Trojan Horse in the Intestine: A Review on the Biotoxicity of Microplastics Combined Environmental Contaminants. J. Hazard. Mater. 2022, 439, 129652. [Google Scholar] [CrossRef]
- Napolitano, G.; Fasciolo, G.; Venditti, P. The Ambiguous Aspects of Oxygen. Oxygen 2022, 2, 382–409. [Google Scholar] [CrossRef]
- Rai, P.K.; Sonne, C.; Brown, R.J.C.; Younis, S.A.; Kim, K.H. Adsorption of Environmental Contaminants on Micro- and Nano-Scale Plastic Polymers and the Influence of Weathering Processes on Their Adsorptive Attributes. J. Hazard. Mater. 2022, 427, 127903. [Google Scholar] [CrossRef]
- Duan, J.; Li, Y.; Gao, J.; Cao, R.; Shang, E.; Zhang, W. ROS-Mediated Photoaging Pathways of Nano- and Micro-Plastic Particles under UV Irradiation. Water Res. 2022, 216, 118320. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D.; Meade, G.; Foley, V.M.; Dowd, C.A. Structure, Function and Evolution of Glutathione Transferases: Implications for Classification of Non-Mammalian Members of an Ancient Enzyme Superfamily. Biochem. J. 2001, 360, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Samal, R.R.; Navani, H.S.; Saha, S.; Kisan, B.; Subudhi, U. Evidence of Microplastics Release from Polythene and Paper Cups Exposed to Hot and Cold: A Case Study on the Compromised Kinetics of Catalase. J. Hazard. Mater. 2023, 454, 131496. [Google Scholar] [CrossRef]
- Iheanacho, S.C.; Odo, G.E. Dietary Exposure to Polyvinyl Chloride Microparticles Induced Oxidative Stress and Hepatic Damage in Clarias gariepinus (Burchell, 1822). Environ. Sci. Pollut. Res. 2020, 27, 21159–21173. [Google Scholar] [CrossRef]
- Allameh, A.; Niayesh-Mehr, R.; Aliarab, A.; Sebastiani, G.; Pantopoulos, K. Oxidative Stress in Liver Pathophysiology and Disease. Antioxidants 2023, 12, 1653. [Google Scholar] [CrossRef]
- Zhou, Z.; Xu, M.J.; Gao, B. Hepatocytes: A Key Cell Type for Innate Immunity. Cell Mol. Immunol. 2016, 13, 301–315. [Google Scholar] [CrossRef]
- Sweidan, A.H.; El-Bendary, N.; Hegazy, O.M.; Hassanien, A.E.; Snasel, V. Water Pollution Detection System Based on Fish Gills as a Biomarker. Procedia Comput. Sci. 2015, 65, 601–611. [Google Scholar] [CrossRef]
- Barboza, L.G.A.; Lopes, C.; Oliveira, P.; Bessa, F.; Otero, V.; Henriques, B.; Raimundo, J.; Caetano, M.; Vale, C.; Guilhermino, L. Microplastics in Wild Fish from North East Atlantic Ocean and Its Potential for Causing Neurotoxic Effects, Lipid Oxidative Damage, and Human Health Risks Associated with Ingestion Exposure. Sci. Total Environ. 2020, 717, 134625. [Google Scholar] [CrossRef]
- GESAMP. Sources, Fate and Effects of Microplastics in the Marine Environment: A Global Assessment; Kershaw, P.J., Ed.; Rep. Stud. GESAMP No. 90, 96 pp; 2015. [Google Scholar]
- Syberg, K.; Khan, F.R.; Selck, H.; Palmqvist, A.; Banta, G.T.; Daley, J.; Sano, L.; Duhaime, M.B. Microplastics: Addressing Ecological Risk through Lessons Learned. Environ. Toxicol. Chem. 2015, 34, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Watson-Wright, W.M.; Wells, P.G.; Duce, R.A.; Gilardi, K.V.; Girvan, A.S.T.; Huber, M.E.; Kershaw, P.J.; Linders, J.B.H.J.; Luit, R.J.; Vivian, C.M.G.; Vousden, D.H. The UN Joint Group of Experts on the Scientific Aspects of Marine Environmental Protection (GESAMP)—An Ocean Science-Policy Interface Standing the Test of Time. Mar. Pollut. Bull. 2024, 199, 115917. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sampling areas.
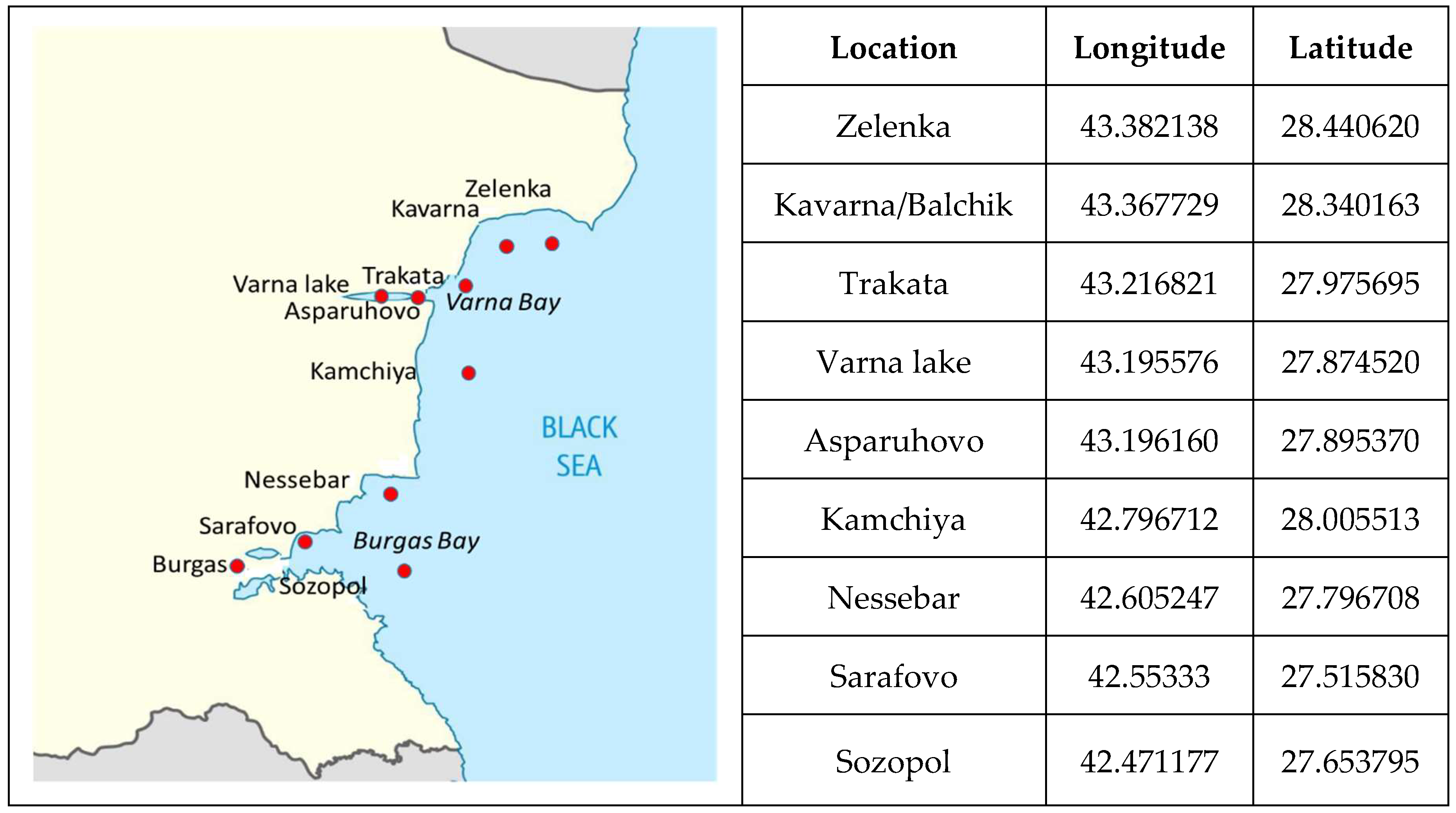
Figure 2.
Thresholds of significant shifts in MPs accumulation in the studied species individuals.
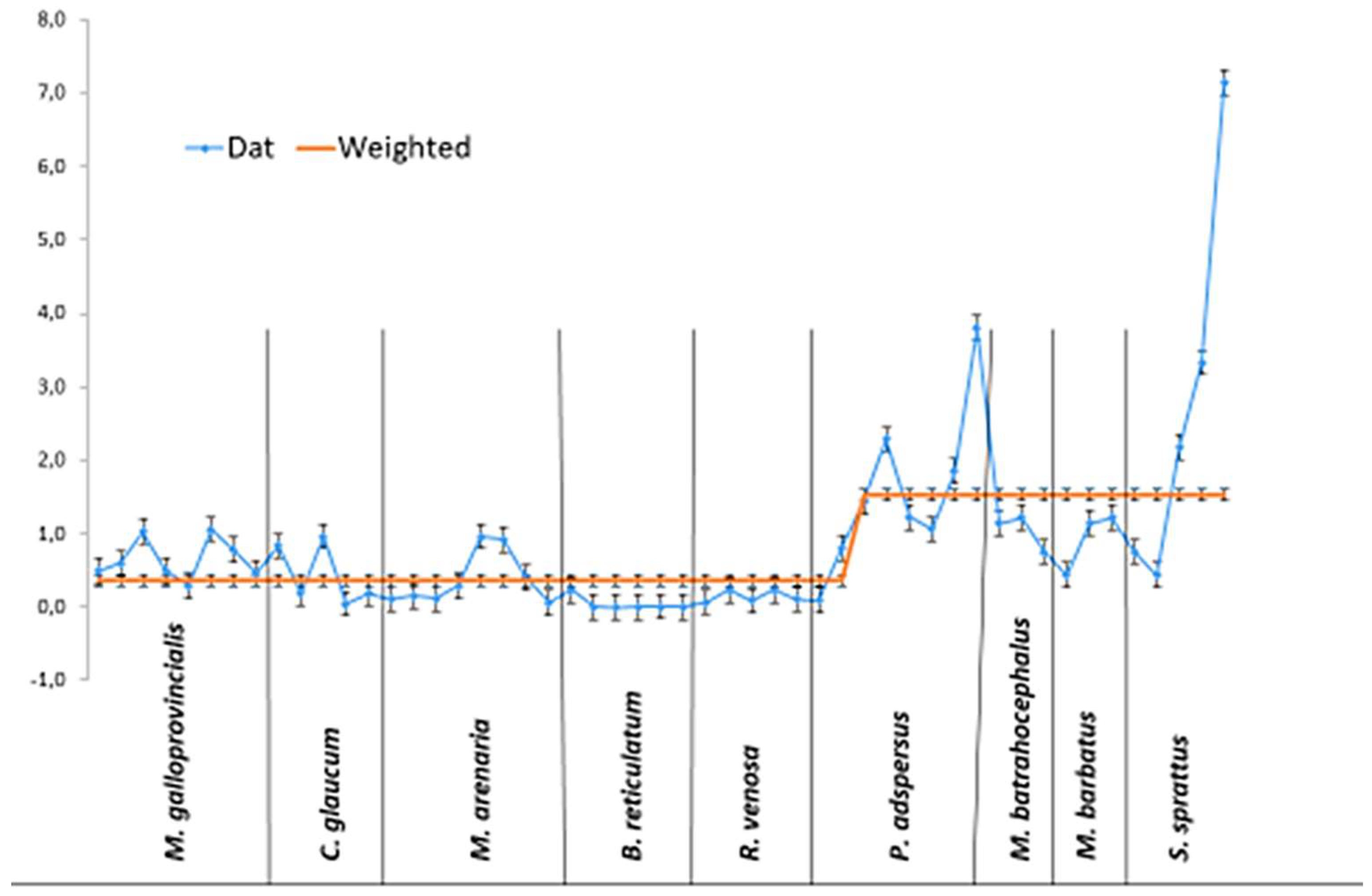
Figure 3.
Percentage ratio of measured OS parameters and MPs accumulation in the tissues of the studied invertebrates.
Figure 3.
Percentage ratio of measured OS parameters and MPs accumulation in the tissues of the studied invertebrates.
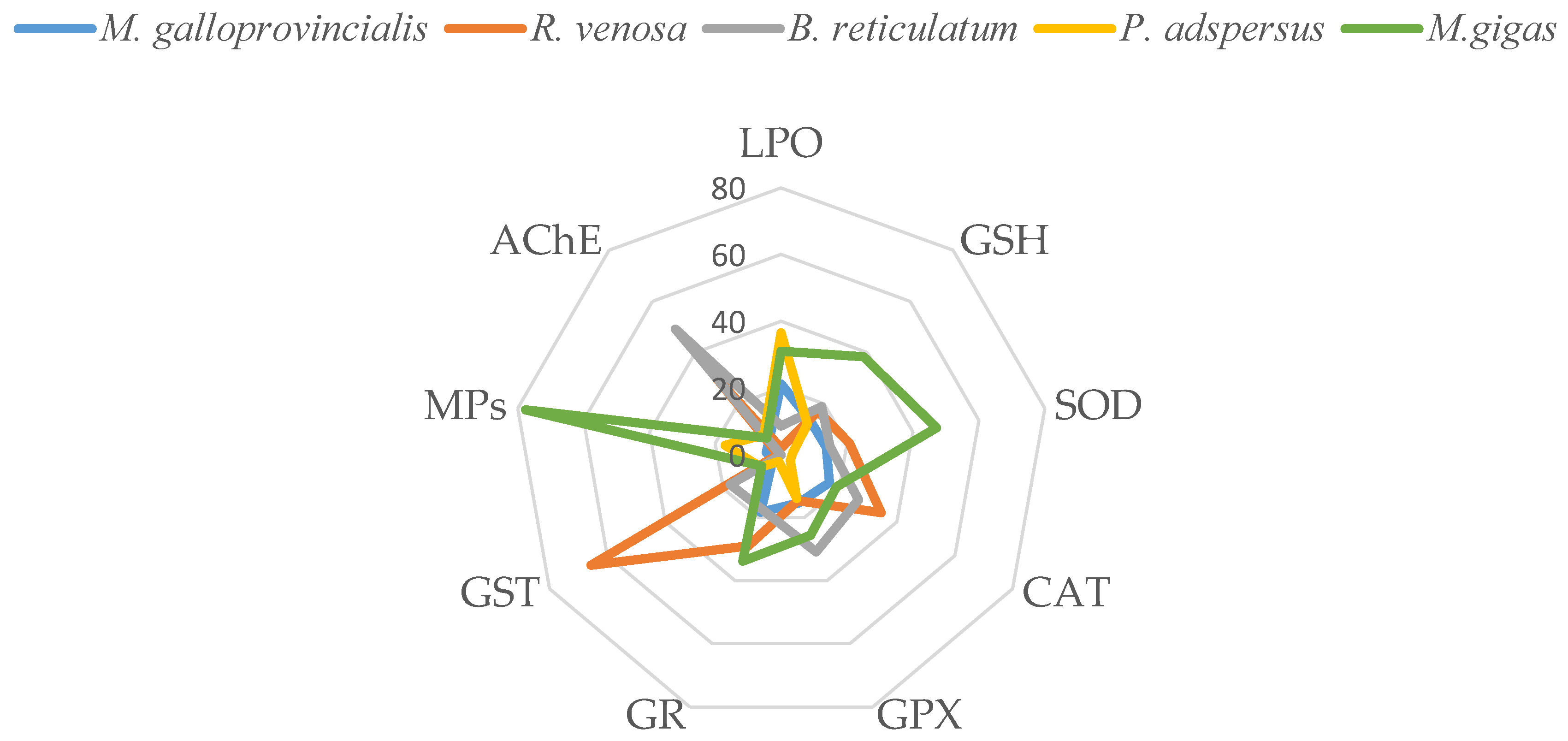
Figure 4.
Percentage ratio of MPs accumulation (particles/g muscle) and the OS parameters in gills (a) and liver (b) of the studied fish species.
Figure 4.
Percentage ratio of MPs accumulation (particles/g muscle) and the OS parameters in gills (a) and liver (b) of the studied fish species.
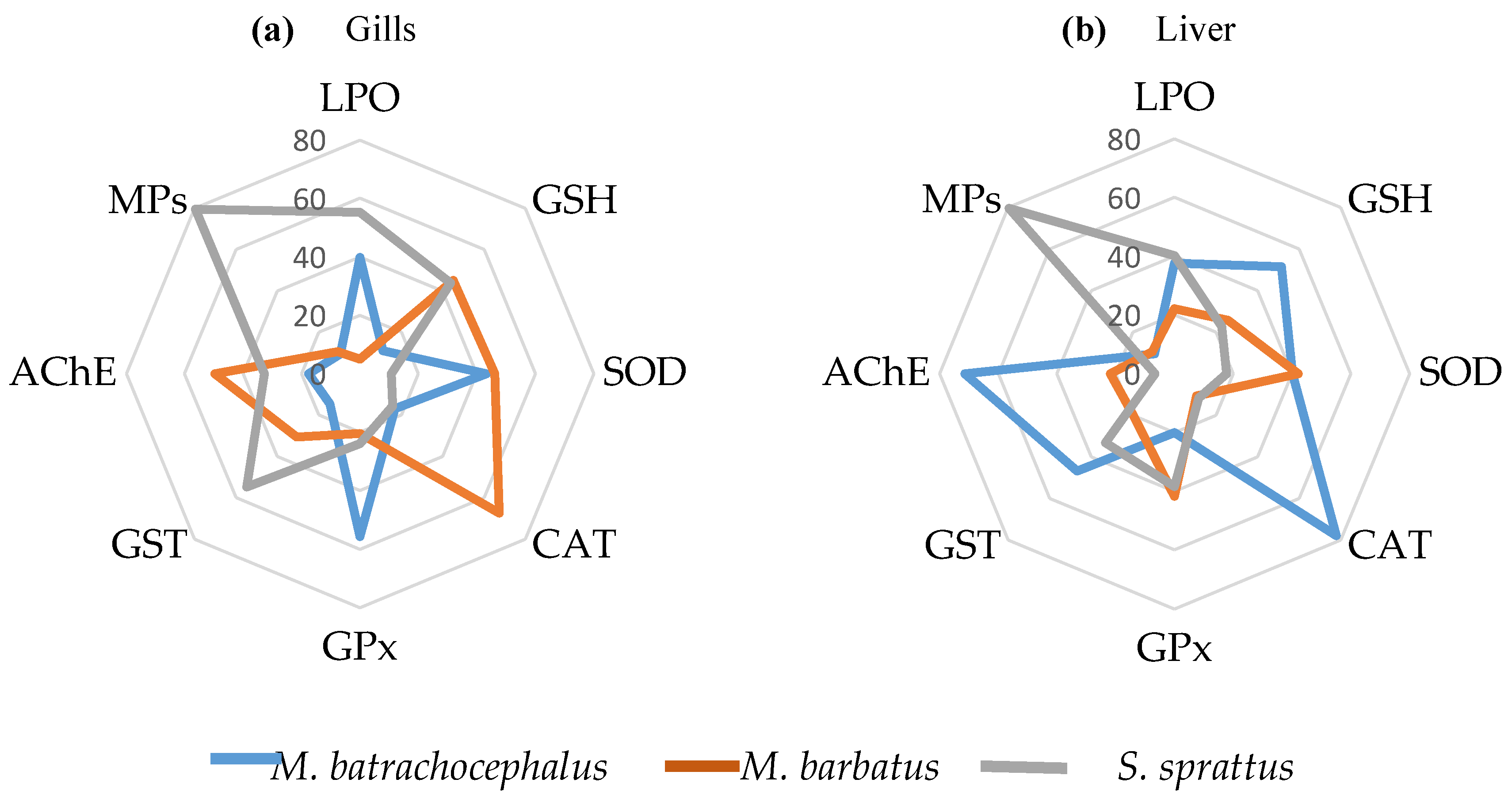
Figure 5.
Results of cluster analysis of the oxidative stress biomarkers and microplastics accumulated in the invertebrate species: (a) M. galloprovicialis, (b) R. venosa, (c) P. adspersus, and (d) B. reticulatum.
Figure 5.
Results of cluster analysis of the oxidative stress biomarkers and microplastics accumulated in the invertebrate species: (a) M. galloprovicialis, (b) R. venosa, (c) P. adspersus, and (d) B. reticulatum.
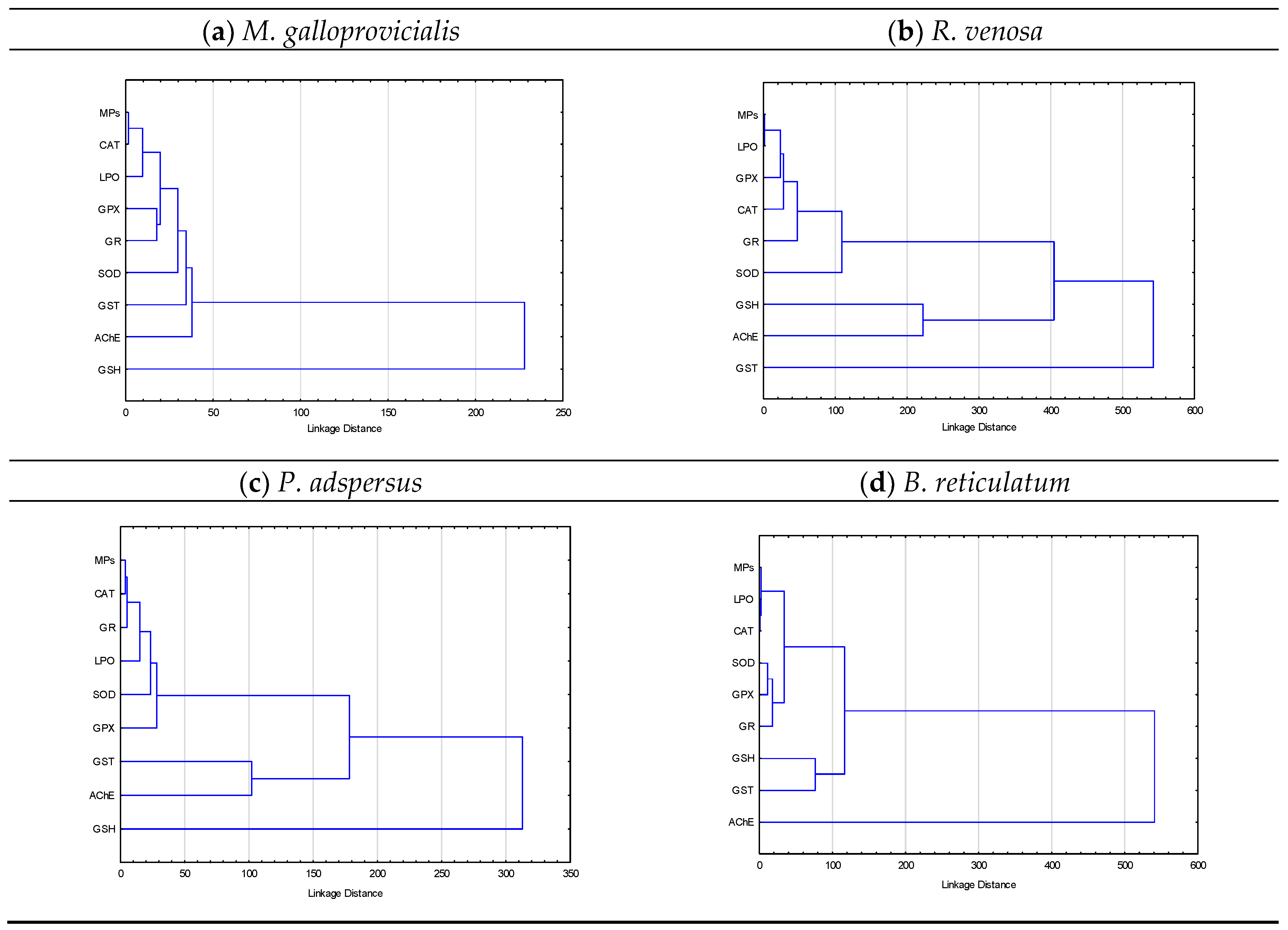
Figure 6.
Results of cluster analysis with the meaningful structures of accumulated microplastics and oxidative stress biomarkers in fish (a) liver and (b) gills.
Figure 6.
Results of cluster analysis with the meaningful structures of accumulated microplastics and oxidative stress biomarkers in fish (a) liver and (b) gills.
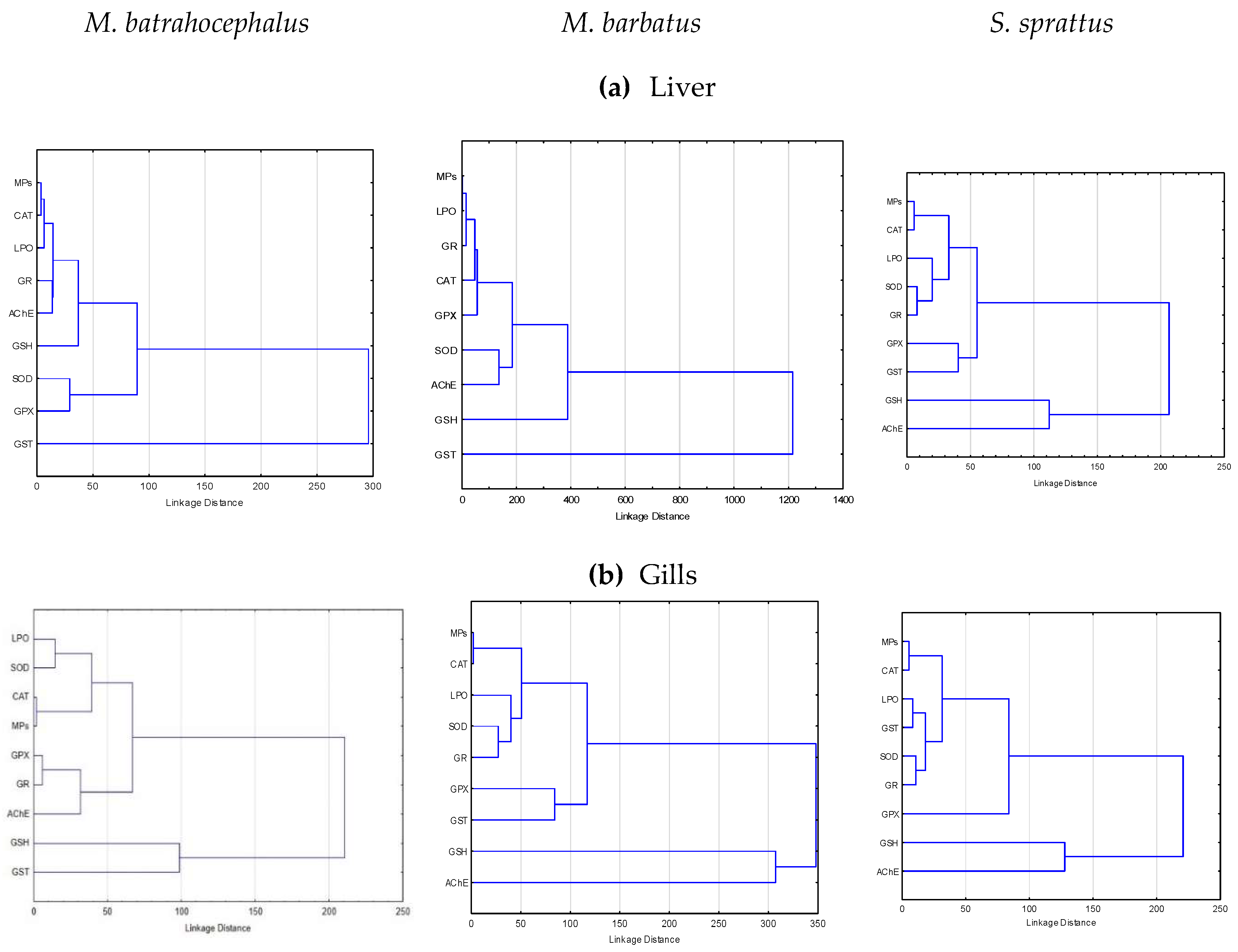
Table 1.
Morphological characteristics of the studied invertebrate species (mean±SD) and accumulated microplastic particles in soft tissues.
Table 1.
Morphological characteristics of the studied invertebrate species (mean±SD) and accumulated microplastic particles in soft tissues.
| Species | W | L | ST weight | MPs | ||
| gr | Cm | gr | %FO in ST | particles/ST | particles/g ST | |
| M. galloprovincialis | 13.62b,c,d,e ±3.30 |
6.04c ±0.42 |
4.04b,c,d,e ±0.95 |
63.33 | 1.13e ±1.11 |
0.320 ±0.31 |
| R. venosa | 37.87a,c,d,e ±6.31 |
5.78c ±0.35 |
11.87a,c,e ±2.33 |
46.67 | 0.60e ±0.83 |
0.050 ±0.07 |
| B. reticulatum | 0.051a,b,d,e ±0.010 |
1.09a,b,d ±0.098 |
0.01a,b,e ±0.03 |
83.33 | 0.30e ±0.19 |
0.0033 ±0.0025 |
| P. adspersus | 1.54a,b,c,e ±0.66 |
5.86c ±1.02 |
- | 70.00 | 1.37e ±1.27 |
0.990 ±1.09 |
| M. gigas | 136.21a,b,c,,d ±19.72 |
11.8 ±0.76 |
22.2 a,b,c,d ±4.54 |
66.67 | 100.5 ±87.36 |
4.530 ±11.08 |
W=Total weight; L=Total length; ST=Soft tissues; FO=Frequency of occurrence. Significant difference from: a - M. galloprovincialis; b - R. venosa; c - B. reticulatum; d - P. adspersus; e – M. gigas.
Table 2.
Morphological characteristics of the studied fish and accumulated microplastic particles in the gastrointestinal tract and muscle (mean±SD).
Table 2.
Morphological characteristics of the studied fish and accumulated microplastic particles in the gastrointestinal tract and muscle (mean±SD).
| W | L | GIT weight | MPs | ||||
|---|---|---|---|---|---|---|---|
| Species | gr | cm | gr | %FO in GIT |
%FO in muscle | particles/ GIT |
particles/g muscle |
| M. batrachocephalus | 90.5b,c ±24.37 |
21.32b,c ±1.83 |
7.57b,c ±4.18 |
53.3c | 26.7c | 0.80 ±1.08 |
0.24c ±0.44 |
| M. barbatus | 21.64a,c ±6.68 |
12.55a ±1.11 |
1.21a,c ±0.34 |
46.7 | 33.4 | 0.93 ±1.16 |
0.27c ±0.48 |
| S. sprattus | 4.19a,b ±0.80 |
9.12a ±0.47 |
0.17a,b ±0.07 |
33.4a | 46.5a | 0.40 ±0.63 |
2.01a,b ±2.56 |
W=Total weight; L=Total length; GIT=Gastrointestinal tract; FO=Frequency of occurrence. Significant difference from: a - M. batrachocephalus; b - M. barbatus; c - S. sprattus.
Table 3.
Values of oxidative stress biomarkers in the soft tissue of the studied marine invertebrate species (mean±SD).
Table 3.
Values of oxidative stress biomarkers in the soft tissue of the studied marine invertebrate species (mean±SD).
| LPO | GSH | SOD | CAT | GPX | GR | GST | AChE | |
|---|---|---|---|---|---|---|---|---|
| Species | nM/mg protein | ng/mg protein | U/mg protein | U/mg protein | U/mg protein | U/mg protein | U/mg protein | U/mg protein |
| M. galloprovincialis | 3.69b,c,d | 93.39c | 21.72b,d | 0.63a,b,c,d | 8.92c | 12.78b,d,e | 13.86a,b,c,d | 33.72b,c,d |
| ±0.33 | ±9.75 | ±1.05 | ±0.07 | ±0.43 | ±1.23 | ±2.00 | ±1.60 | |
| R. venosa | 0.37a,c,d | 118.5 | 32.81a,c,d | 1.30a,b,d | 8.48c | 20.40a,b,d | 323.62a,b,c,d,e | 181.56a,d,e |
| ±0.04 | ±69.60 | ±2.86 | ±0.23 | ±1.57 | ±5.17 | ±10.72 | ±22.67 | |
| B. reticulatum | 1.51a,b,d | 130.9a,d | 23.56b,d | 1.01a,c,d | 17.92ab,d | 12.12d,e | 86.70a,b,c,e | 293.52a,d,e |
| ±0.03 | ±2.74 | ±0.48 | ±0.12 | ±1.20 | ±1.27 | ±1.73 | ±16.31 | |
| P. adspersus | 6.32a,b,c | 84.24c | 5.31a,b,c | 0.12a,b,c,e | 8.03c | 1.48a,b,c,d,e | 34.65a,b,c | 47.51a,b,c |
| ±1.13 | ±4.69 | ±0.55 | ±0.01 | ±0.99 | ±0.15 | ±4.00 | ±5.70 | |
| M. gigas |
5.38b,c ±1.22 |
266.55a,b,c,d ±62.79 |
74.60a,b,c,d ±7.82 |
0.72d ±0.09 |
14.90a,b,d ±1.56 |
23.81a,c,d ±1.61 |
33.10a,b,c, ±3.44 |
40.00b,c ±6.45 |
Significant difference from: a - M. galloprovincialis; b - R. venosa; c - B. reticulatum; d - P. adspersus; e – M. gigas.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



