
Submitted:
07 May 2025
Posted:
08 May 2025
You are already at the latest version
Abstract
Background/Objectives: Cutaneous melanoma (CM) poses a critical public health concern due to its high recurrence rates and resistance to standard treatments. This study investigated the antineoplastic potential of Curcuma Longa L. extract enriched with Bisdemethoxycurcumin (AC3TM) on human melanoma cell lines A375 and SK-MEL-28. Methods: The A375 and SK-MEL-28 cell lines were cultured according to the manufacturer's recommendations and were subsequently treated with different concentrations of AC3TM. The MTT test and fluorescence microscopy were used to assess cell viability. The wound healing assay was used to verify cell migration after treatment with the compound. To determine reactive oxygen species (ROS) levels, we used the fluorescence assay with 2,7-dichlorodihydrofluorescein diacetate. Subsequently, the molecular pathways associated with apoptosis were evaluated using RT-PCR, including the modulation of the inflammatory profile and the components of purinergic signaling. Results: The results revealed that AC3™ effectively inhibited cell proliferation and migration in both melanoma cell lines. In A375 cells, the compound reduced ROS levels and significantly modulated apoptotic pathways, including upregulation of caspase-3 expression. Similarly, in SK-MEL-28 cells, increased expression of caspase-8 and caspase-3 indicated the activation of both intrinsic and extrinsic apoptotic mechanisms. Additionally, AC3™ influenced the inflammatory response by increasing the expression of Tumor Necrosis Factor Alpha (TNF-α), Interleukin 6 (IL-6) and NLR Family Pyrin Domain Containing 3 (NLRP3) in A375 cells while downregulating NLRP3 expression in SK-MEL-28 cells, highlighting its differential immune modulatory effects. Furthermore, the compound modulates the expression and enzymatic activity of ecto-nucleoside triphosphate diphosphohydrolase 1 (CD39) and ecto-5'-nucleotidase (CD73), key regulators of purinergic signaling, suggesting its ability to impact immune evasion mechanisms in melanoma. Conclusions: These findings position AC3™ as a promising therapeutic candidate for melanoma, targeting critical pathways related to tumor progression, immune regulation, and cellular survival.
Keywords:
Curcuminoids
; Cutaneous melanoma
; Apoptosis
; TNF/TNFR1 axis
; Purinergic signaling
; Antitumor Therapy
1. Introduction
Since ancient times, humans have used natural compounds to prevent and treat various diseases. Over time, several studies have been carried out, and the functional properties of these products have been sought, as well as their route of action [1]. There is currently a wealth of evidence highlighting the potential of natural compounds in the prevention or treatment of aging [2], obesity and non-alcoholic fatty liver disease (NAFLD) [3,4], neurodegenerative diseases [5], inflammatory bowel diseases [6], kidney disease [7], osteoporosis [8], chronic pain [9], and numerous types of cancer [10,11,12]
Polyphenols and flavonoids, such as curcumin, resveratrol, and phenolic acids, stand out among the compounds with the most significant scientific evidence. These compounds have numerous pharmacological properties, including antioxidant, anti-inflammatory, antibacterial, immunomodulatory, anticancer, antiproliferative, antimutagenic, and antithrombotic activities [13,14,15,16,17,18,19,20,21,22,23,24,25,26,27].
Curcumin, derived from Curcuma longa L., a plant belonging to the Zingiberaceae family, stands out as one of the most extensively investigated polyphenols. In addition to curcumin (CUR) (about 75–80%), the plant contains approximately 50 other curcuminoids, including demethoxycurcumin (DMC) and bisdemethoxycurcumin (BDMC), which together comprise the primary bioactive constituents [28].
Despite its potent pharmacological activities, the low bioavailability of curcumin limits its effectiveness which has prompted the researchers to work with other curcuminoids, such as DMC and BDMC [29]. These compounds differ significantly from curcumin in both their chemical composition and biological activity, offering superior pharmacological effects [30]. BDMC, in particular, has been shown to possess more favorable properties than curcumin, in terms of bioavailability, and bioaccessibility, leading to more potent therapeutic effects [31]. Moreover, these curcuminoids support curcumin’s biological activities and physical stability improving its therapeutic potential. The combination of these three curcuminoids was found to be more effective since they act synergistically [28].
Among the therapeutic properties of curcuminoids, their antineoplastic activity has garnered significant attention [32]. Cancer remains a major global health challenge, with certain types, such as cutaneous melanoma, demonstrating resistance to conventional therapies and high recurrence rates, contributing to elevated mortality [33]. To solve this problem, several natural compounds, such as curcuminoids, and the modulation of alternative signaling pathways, such as purinergic signaling, have been studied as adjuvant strategies for treating the disease [34,35,36]. Cutaneous melanoma is characterized by its aggressive nature and ability to evade immune responses, often mediated by purinergic signaling pathways involving ectonucleotidases such as CD39 and CD73 [37]. These enzymes regulate the extracellular levels of ATP and adenosine, which play opposing roles in tumor immunity [38]. Targeting these pathways has emerged as a promising strategy for overcoming therapeutic resistance in melanoma.
Considering the evidence presented above, the objective of the present study was to evaluate the antineoplastic activity of the Advanced Curcumin C3 Complex (AC3TM) (Sami-Sabinsa Group Limited), in two cutaneous melanoma cell lines, A375 and SK-MEL-28. Unlike regular turmeric extracts, it is a specialized formulation derived from Curcuma longa, enriched with DMC and BDMC for enhanced physiological benefits. Additionally, we sought to elucidate the key signaling pathways modulated by AC3TM, focusing on its effects on purinergic signaling, oxidative stress, and apoptotic mechanisms.
2. Materials and Methods
2.1. Chemicals and Reagents
Chemicals and reagents used in this study were high analytical grade purchased from Sigma-Aldrich (Sigma-Aldrich, St. Louis, MO, USA), Merck (Darmstadt, Germany), Thermo Fisher Scientific (Grand Island, NY, USA), Invitrogen Life Technologies (Carlsbad, CA, USA) and Sami-Sabinsa Group Limited (India). The cell lines A375 and SK-MEL-28 were purchased from the Cell Bank of Rio de Janeiro (BCRJ) (Brazil). Cell culture medium was purchased from Vitrocell™ (Brazil), and plates and flasks used for culture procedures were obtained from Kasvi™ (Brazil).
2.2. Cell Culture and Exposure to AC3TM
The A375 and SK-MEL-28 cell lines were cultured following the Bank of Cell of Rio de Janeiro (BCRJ) recommendations. For A375, Dulbecco's Modified Eagle's Medium (DMEM) containing 2mM of L-glutamine, 4500 mg/L glucose, and 10% of fetal bovine serum was used. For SK-MEL-28, we used Dulbecco's Modified Eagle's Medium (DMEM) with 2mM L-glutamine, 1.0 g/L glucose, and 10% of fetal bovine serum. The cells were maintained three times a week and kept in a humidified and controlled atmosphere of 5% carbon dioxide (CO2) at 37°C until reaching the desired confluence. AC3TM was dissolved in 0.2% dimethyl sulfoxide (DMSO); the non- toxic dosages used (0.78 μg/mL, 1.56 μg/mL, 3.12 μg/mL, 6.25 μg/mL) were defined through a previous study [32]. Cells belonging to the negative control (CT) group were treated with a culture medium only. After treatment, all treatment groups were incubated for 24h.
2.3. Cell Viability by MTT Assay
To assess cell viability, the 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay was used as previously described by Mosmann [39]. After the treatment exposure time, the treatment was removed, the cells were washed with phosphate-buffered saline (PBS) (0.1 M, pH 7.4), and 100 µL of MTT dissolved in PBS was added. The cells were incubated at 37°C for 2 hours, during which time the viable cells reduced the MTT to formazan crystals, which were subsequently dissolved in DMSO and read at 570 nm in a microplate reader (Thermo Scientific™ Varioskan™ LUX).
2.4. Cell Viability by Fluorescence Microscopy Assay
Cell viability was also measured using the fluorophore acridine orange (AO), which is taken up only by viable cells and stains both double-stranded (ds) and single-stranded (ss) nucleic acids. Consequently, viable cells emit green fluorescence, which is read under a fluorescence microscope (Nikon® Eclipse TS2-FL) (480–490 nm) in triplicate at 200× magnification and adjusted for brightness and contrast linearly by Imagej® software [40].
2.5. Measurement of Mitochondrial Transmembrane Potential (ΔΨm)
The mitochondrial transmembrane potential was measured using tetramethylrhodamine ethyl ester (TMRE), accumulating in active mitochondria and emitting red fluorescence proportional to the mitochondrial transmembrane potential when excited at 550 nm. All samples were read in triplicate under a fluorescence microscope (Nikon® Eclipse TS2-FL) at 200× magnification and adjusted for brightness and contrast linearly using Imagej® software [41].
2.6. Wound-Healing Migration Assay
Cell migration assay was performed according to Justus et al. [42] cells were seeded in monolayer in 6-well plates, at a density of 1 × 10⁶ cells/well. When they reached 100% confluence, a scratch was made using a sterile 200 μL tip, and an initial image was recorded by optical microscopy at 4× magnification. After treatment, the supernatant was discarded, the cells were washed with saline solution, and a new image was obtained. The final images were linearly adjusted for brightness and contrast using ImageJ® software. The results were expressed as percentage (%) of wound closure compared to the control.
2.7. Detection of Reactive Oxygen Species (ROS)
ROS levels were checked by a fluorescent assay using 2,7-dichlorodihydrofluorescein diacetate (H2DCF-DA), as previously described by Manica et al. [43]. The reading was performed on a fluorescence plate reader (Thermo Scientific™ Varioskan™ LUX) at Ex./Em. = 488/525 nm. All tests were performed in quadruplicate, and results were expressed as a percentage (%) of relative fluorescence compared to the control.
2.8. Gene Expression
To extract total RNA from cells, TRIzol® reagent (Invitrogen) was used according to the manufacturer's instructions. Subsequently, RNA quantification and purity verification were performed by the absorption ratio of 260/280 nm using a μDrop™ spectrophotometer (Varioskan LUXTM, Thermo Fisher Scientific, USA). RNA was treated with DNAse (Thermo Scientific, USA) following the manufacturer's recommendations. cDNA was prepared according to the instructions of the cDNA High-Capacity cDNA Reverse Transcription Kit (Thermo Scientific, USA). To evaluate gene expression, each reaction consisted of 6 µL of cDNA sample, 10 µL of PowerUp™ SYBR™ Green Master Mix for qPCR (Thermo Scientific, USA), 2 µL of primer F (500 nM), and 2 µL of primer R (500 nM) for a final volume of 20 µL. Amplification occurred in the QuantStudio™ 7 Pro Real-Time PCR System (Thermo Scientific, USA). Melting curve analysis was performed to verify product identity. Tests were conducted in quadruplicate, and GAPDH was used for housekeeping. A calibrator was used, and data were calculated by the ΔΔCq method. The forward and reverse (5'-3') oligo sequences used for each gene are described in Table 1.
2.9. Assessment of CD39 and CD73 Enzymatic Activities
The enzymatic activities of CD39 and CD73 were performed according to Pilla et al. [44] and Lunkes et al. [45] with adaptations. The reactions were performed in quadruplicate, prepared as described [46], and subsequently read at 630 nm using the Varioskan LUX TM spectrophotometer (Thermo Fisher Scientific, USA). A standard curve with KH2PO4 was used to calculate the enzymatic activity, and the results were expressed as nmol/Pi /min/mg of protein.
2.10. Protein Determination
The Bradford method [47] was employed for protein determination, using bovine serum albumin as the standard. The protein samples were adjusted according to each assay in mg/mL.
2.11. Statistical Analysis
All the parameters were statistically evaluated by analysis of variance followed by the appropriate post hoc test using GraphPad Prism 9 software. All data were expressed as mean ± standard deviation. The differences between the groups in terms of the studied variables were evaluated through one-way ANOVA analysis. The differences in the probability of rejecting the null hypothesis as < 5% (P < 0.05) were considered statistically significant. Statistical significance was defined for p values of * (P < 0.05), ** (P < 0.01), *** (P < 0.001) and **** (P < 0.0001).
3. Results
3.1. AC3TM Decreased Viability and Migration of Melanoma Cells
Figure 1 presents the antineoplastic effect of AC3TM on the SK-MEL-28 and A375 cells. AC3TM significantly inhibited the proliferation and migration of A375 and SK-MEL-28 cells. In the A375 cell line, a significant reduction in cell viability was observed at concentrations of 3.12 μg/mL (P = 0.0009) and 6.12 μg/mL (P < 0.0001) (Figure 1A).
The decrease in cell viability was further supported by fluorescence microscopy analysis, where nuclei were stained with the fluorescent agent acridine orange (AO, green), and mitochondria were stained with Tetramethylrhodamine Methyl Ester (TMRE, red), A decrease in fluorescence intensity indicated impaired cellular proliferation.. Moreover, the wound-healing assay demonstrated an inhibition of A375 cell migration following treatment with AC3TM, with significant effects observed at concentrations of 1.56 μg/mL (P < 0.0001), 3.12 μg/mL (P = 0.0003), and 6.25 μg/mL (P < 0.0001) (Figure 1C-D).
Similarly, in the SK-MEL-28 cutaneous melanoma cell line, AC3TM demonstrated a significant reduction in cell proliferation at the highest concentration tested (P = 0.0304) (Figure 1E-F). Furthermore, the wound-healing assay indicated an inhibition of cell migration, as evidenced by a significant reduction in wound closure across all tested concentrations (P < 0.0001) (Figure 1G-H).
3.2. AC3TM Decreases ROS Levels and Modulates Caspase Expression in CM A375 and SK-MEL-28 Cells
Following a 24-h treatment with AC3TM, a significant reduction in reactive oxygen species (ROS) levels was observed across all tested concentrations in both A375 (Figure 2A) and SK-MEL-28 (Figure 2D) melanoma cell lines. To further elucidate the AC3TM mechanism of action, the gene expression of caspases 8 and 3 was evaluated. In SK-MEL-28 cells, treatment with AC3™ at 3.12 µg/mL resulted in a significant upregulation of caspase-8 gene expression (P = 0.0435) (Figure 2E). In contrast, no significant alterations in caspase-8 expression were detected in A375 cells when compared to the control (Figure 2B). Regarding caspase 3, we found a substantial increase in caspase-3 gene expression after treatment with 6.25 µg/mL of the compound in the A375 (P = 0.0150) (Figure 2C) and SK-MEL-28 (P < 0.0001) (Figure 2F) cell lines.
3.3. AC3TM Modulates the Inflammatory Cascade
We used RT-qPCR to evaluate TNF-α, IL-6, and NLRP3 gene expression, as shown in Figure 3. After 24h of treatment with 6.25 µg/mL of AC3™, there was an increase in the gene expression of TNF-α (P = 0.0446), IL-6 (P = 0.0035) and NLRP3 (P = 0.0012) in the A375 cell line (Figure 3A-C). In the SK-MEL-28 cell line, an increase in the gene expression of TNF-α (P < 0.0001) was observed at the highest treatment concentration (Figure 3D). In contrast, in this same cell line and treatment concentration, a reduction in the levels of NLRP3 was observed (P = 0.0006) (Figure 3F).
3.4. AC3TM Modulates Gene Expression of CD39 and CD73
We evaluated the gene expression of ectonucleotidases after 24 h of treatment with AC3TM in A375 and SK-MEL-28 cells, as shown in Figure 04 (A-D). In A375 cells, the treatment concentration of 3.12 µg/mL (P = 0.0011) decreased CD39 expression. In comparison, at the concentration of 6.25 µg/mL (P = 0.0078), there was an increase in CD39 gene expression (Figure 4A). Regarding CD73, the concentration of 6.25 µg/mL increased gene expression in A375 cells (P < 0.0001) (Figure 4B). Differently, for SK-MEL-28 cells, the treatment was able to decrease CD39 gene expression at the concentration of 6.25 µg/mL (P = 0.0016) (Figure 4C). Similarly, treatment with 6.25 µg/mL of AC3TM decreased CD73 gene expression, compared to the control (P = 0.0009) (Figure 4D).
3.5. AC3TM Modulates Enzymatic Activity of CD39 and CD73
Figure 5 shows the results of the enzymatic activity of CD39 and CD73 after 24 h of treatment with AC3TM (Figure 5A-F). In the A375 cell line, an increase in ATP hydrolysis was observed after treatment with 3.12 μg/mL (P = 0.0059) and 6.25 μg/mL (P = 0.0015) (Figure 5A). No significant difference was observed in the levels of ADP (Figure 5B) and AMP (Figure 5C) hydrolysis in this cell line after treatment with the compound.
In SK-MEL-28 cells, after 24 h of treatment with AC3TM, we observed an increase in ATP hydrolysis at the concentration of 3.12 μg/mL (P = 0.0251), followed by a decrease at the concentration of 6.25 μg/mL (P < 0.0001) (Figure 5D). Similar results were obtained in AMP hydrolysis, where an increase in CD39 activity occurred at a concentration of 3.12μg/mL (P < 0.0001), followed by a decrease at a concentration of 6.25 μg/mL (P = 0.0061) (Figure 5E). Similarly to the A375 cell line, CD73 activity did not show significant differences after treatment with AC3TM.
4. Discussion
Curcuminoids have been used for many years as a spice. Among them, the most studied curcuminoid is CUR, which has shown antitumor effects in several types of cancer, such as breast [48] and ovarian [49] and esophagus [50]. In melanoma cells lines, it was proved that curcumin induces apoptosis by activating cell death pathways involving caspases 3, 7, 8, and 9, and AKT/mTOR. It also can decrease the expression of factors associated with angiogenesis and invasion, such as Vascular Endothelial Growth Factor (VEGF), Matrix Metalloproteinase-2 (MMP- 2), Matrix Metalloproteinase-9 (MMP-9), and Cyclooxygenase (COX), which are predictive of metastasis [43,51,52,53]. The others curcuminoids, such as DMC and BDMC, also have antitumor activity and have been shown to act synergistically with curcumin [32,54,55,56]. Considering the previous antineoplastic activity of available commercial curcuminoids, we verified the antineoplastic properties of a mixture of phytonutrients that expounds the benefits of BDMC, as the major ingredient, along with DMC and CUR standardized in ACM™ in two cutaneous melanoma cell lines.
Furthermore, in our study, we chose to use two cutaneous melanoma cell lines, A375 and SK-MEL-28. These cell lines are widely studied and present different levels of aggressiveness. A comparative study between cutaneous melanoma cell lines using an aggressiveness scale (Melanoma Agressiveness Score: MAGS) showed that the A375 cell line was classified as the most aggressive and the SK-MEL-28 cell line the least aggressive [57].
We found that the AC3TM was able to decrease cell proliferation and migration in both melanoma cells lines. According to Huang et al. [32], the three curcuminoid constituents, CUR, DMC, and BDMC, act synergistically. Our results corroborate previous findings, higher doses of curcuminoids showed beneficial effects on cell proliferation and migration of both the cutaneous melanoma cell lines, A375 and SK-MEL-28 [54,58,59]. Similarly, curcumin-loaded nanoparticles have also demonstrated antiproliferative and anti-migratory effects and marked cell viability reduction in breast cancer models [60]. In this study, we observed a significant decrease in cell viability at lower doses (Figure 1).
In addition to reducing cell viability, previous studies have found that curcuminoids increase ROS levels, causing damage to DNA as a way of inducing apoptosis [61,62]. In contrast to the results found here, we found that when in association, these compounds decrease ROS levels in A375 and SK-MEL-28 cells and increase caspases-8 and -3 expression independent of ROS levels (Figure 2). Sandur et al. [28] found similar results, in which, regardless of ROS levels, curcuminoids induced cell death via nuclear factor-κB (NF-kB) and TNF, that is, apoptosis through the extrinsic pathway. Our data suggest that AC3™ exerts its antineoplastic effects, at least in part, by reducing oxidative stress and modulating the apoptotic machinery by regulating caspase gene expression in melanoma cells.
An increase in TNF-α gene expression was observed after treatment with AC3TM in the A375 and SK-MEL-28 cells (Figure 3). In these cases, apoptosis is mediated via TNF and carried out by the TNF receptor 1 (DR1/TNFR1), in which interaction occurs between TNF and TNFR1 leading to the recruitment of the TNFR-associated death domain (TRADD) protein, through its DD [63]. Subsequently, interaction occurs with the Fas-associated death domain (FADD), which recruits pro-caspase-8 and cleaves it, transforming it into active caspase-8, which finally activates caspase-3, an effector of cell death [64]. In A375 cells, Tyciakova et al. [65] demonstrated that TNF overexpression is correlated with an increase in IL-6 and the pro-apoptotic tumor necrosis factor ligand gene (TRAIL) mediated by TNF/TNFR1 signaling. Similar results were found in our study, where we observed an increase in the inflammatory cascade and the overexpression of TNF, IL-6, and NLRP3 in cell line A375.
Another essential component of the inflammatory cascade is the NLRP3 inflammasome, considered a double-edged sword in several types of cancer, being able to exert a protective antitumorigenic effect and a pro-tumorigenic role [66]. In CM, the NLRP3 inflammasome is highly expressed, acting on tumor progression through activating and cleavage of caspases and IL-1β, decreasing the immune response and consequently suppressing anti-melanoma T cell responses [64,67,68]. In our previous study on the SK-MEL-28 line treated with CUR [36], we observed that the substance could decrease the expression of NLRP3 and consequently increase the efficacy of anti-melanoma therapies, particularly those targeting the anti-programmed cell death 1 (anti-PD-1) pathway. Contrary to what was found in A375 cells, in the SK-MEL-28 cell line, we observed a decrease in the gene expression of the NLRP3 inflammasome, supporting the hypothesis of the previous study that in this type of melanoma, treatment with CUR or a mix of curcuminoids CUR, DMC and BDMC, in this case, AC3TM can improve the response to therapies, acting mainly in the inhibition of tumor suppression, that is, positively regulating the immune response [69].
Another important therapeutic target for the treatment of CM, capable of modulating the immune response, are the components of purinergic signaling, in particular, the purinergic mediators of the extracellular environment, adenosine triphosphate (ATP) and adenosine (Ado) [70,71]. ATP present in the extracellular environment can be released in different ways. One of them is cell lysis, a process that is directly related to apoptosis or by non-lytic mechanisms such as exocytosis of ATP-containing vesicles via nucleotide-permeable channels (connexin/pannexin hemichannels, maxi-anionic channels, volume-regulated anion channels, and P2X7 receptor channels), through transport vesicles that deliver proteins to the cell membrane or through lysosomes [72,73]. Ado comes from the dephosphorylation of ATP by the ectonucleotidases CD39 and CD73, the former being responsible for the conversion of ATP into adenosine monophosphate (AMP) and the latter converts the AMP resulting from CD39 activity into Ado [74]. High levels of ATP in the tumor microenvironment act as a Danger-Associated Molecular Pattern (DAMP), inducing the immune response. Otherwise, Ado inhibits the immune response, causing immunosuppression [75,76]. Antitumor immunity induced by Ado is one of the significant challenges encountered in the treatment of CM since the newly available therapies that act by inhibiting immune checkpoints (ICIs) and represent a more attractive prognosis depend on the negative regulation of the CD39/CD73/Ado axis [77,78].
Patients with CM have higher levels of CD39 and CD73 expression, which are associated with resistance to therapies and worse prognosis [37,79,80]. In this study, we found that treatment with 6.25 μg/mL of the AC3TM complex was able to decrease the gene expression of the ectonucleotidases CD39 and CD73in the SK-MEL-28 cell line, considered the least aggressive (Figure 4). Furthermore, in this same lineage we observed a decrease in the activity of the CD39 enzyme (Figure 5). In contrast, in the A375 cell line, there was an increase in the expression of the CD39 and CD73 enzymes and in the enzymatic activity of CD39 (Fig 4-5), demonstrating that the compound can act in different ways in cell lines, possibly due to the different levels of aggressiveness of the tested strains [81].
Lower levels of adenosine in the extracellular environment, due to lower expression of CD73, can decrease immunosuppression, increasing the response to therapies [82]. One of the therapies for treating cutaneous melanoma with the highest levels of efficacy inhibits an immune checkpoint via PD-1/PD-L1 blockade [83]. However, high resistance rates are still observed, so recent studies support associated therapies [84]. This association can be observed in the study by Tu et al., who found that the combination of anti-PD-L1 and anti-CD73 antibodies improved the response to treatment of non-small cell lung cancer [85].
In melanoma, we observed that treatment with AC3™ decreased CD73 expression in SK-MEL-28 cells. This agrees with our previous study, in which CUR exerted similar activity in SK-MEL-28 cells. Thus, both CUR and the curcuminoid mixture are strong candidates for adjuvant therapy of MC (17).
5. Conclusions
In this study we showed that AC3™ reduces cell viability, inhibits cell migration, increases caspase-3 levels, and modulates the levels of pro-inflammatory markers and components of the purinergic system in melanoma cell models A375 and SK-MEL-28. This complex compound positively regulated the TNF pathway, promoting apoptosis of melanoma cells. In addition, regulating the inflammatory cascade results in lower levels of immunosuppression, a key factor contributing to therapeutic resistance in melanoma. Finally, AC3™ emerges as a promising candidate for further investigation in CM, given its ability to modulate purinergic signaling through the CD39/CD73 axis, which controls extracellular ATP levels, and through the antagonistic action on P1 receptors. Taken together, our findings position AC3™ as a multifaceted and innovative therapeutic candidate capable of enhancing the biological and therapeutic activity of curcuminoids, while overcoming the limitations associated with curcumin’s poor bioavailability. By targeting critical mechanisms such as tumor cell survival, immune evasion, and purinergic signaling, AC3™ offers a promising strategy to improve treatment outcomes and address therapeutic resistance in melanoma.
Author Contributions
Conceptualization: DM, AZ and MDB. Methodology: DM, GBS, MDB, and AZ. Investigation: DM, GBS, BO, VC, VAR. Visualization: DM, GBS, MDB and AZ. Funding acquisition: CO, IDB, AM, SM, MDB and AZ. Project administration: DM, GBS, MDB, and AZ. Supervision: MDB and AZ. Writing – original draft: DM. Writing – review, and editing: DM, GBS, CO, IDB, AM, SM, APK, MDB and AZ.
Funding
This study was funded by grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (MDB, Grant: 310606/2021-7; AZ, Grant: #313311/2021-8), Fundação de Amparo à Pesquisa e Inovação do Estado de Santa Catarina (FAPESC) (MDB, Grant: 2021TR001726; AZ, Grant: TO#2024TR002196) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) (DM; Grant: Financial Code 001). MDB, AZ, and APK are 2 CNPq Researchers Fellows (PQ2).All authors reviewed and approved the final version.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
All data are available in the main text or supplementary materials.
Conflicts of Interests
CO, IDB, AM and SM are affiliated with Sami-Sabinsa Group Limited, which develops the product AC3TM. This commercial affiliation is disclosed as a potential conflict of interest. The remaining authors report no relevant financial or non-financial interests to disclose.
Abbreviations
The following abbreviations are used in this manuscript:
| CM | Cutaneous melanoma |
| AC3TM | Advanced Curcumin C3 Complex |
| ROS | Reactive oxygen species |
| TNF-α | Tumor Necrosis Factor Alpha |
| IL-6 | Interleukin 6 |
| NLRP3 | NLR Family Pyrin Domain Containing 3 |
| CD39 | Ecto-nucleoside triphosphate diphosphohydrolase 1 |
| CD73 | Ecto-5'-nucleotidase |
| CASP-8 | Caspase-8 |
| CASP-3 | Caspase-3 |
| NAFLD | Non-alcoholic fatty liver disease |
| CUR | Curcumin |
| DMC | Demethoxycurcumin |
| BDMC | Bisdemethoxycurcumin |
| BCRJ | Bank of Cell of Rio de Janeiro |
| DMEM | Dulbecco's Modified Eagle's Medium |
| MTT | 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide |
| AO | Acridine orange |
| TMRE | Tetramethylrhodamine ethyl ester |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| ADA | Adenosine deaminase |
| VEGF | Vascular Endothelial Growth Factor |
| MMP-2 | Matrix Metalloproteinase-2 |
| MMP-9 | Matrix Metalloproteinase-9 |
| COX | Cyclooxygenase |
| MAGS | Melanoma Agressiveness Score |
| NF-kB | Nuclear factor-κB |
| DR1/TNFR1 | TNF receptor 1 |
| TRADD | TNFR-associated death domain |
| FADD | Fas-associated death domain |
| TRAIL | Tumor necrosis factor ligand gene |
| anti-PD-1 | Anti-programmed cell death 1 |
| ATP | Adenosine triphosphate |
| Ado | Adenosine |
| AMP | Adenosine monophosphate |
| DAMP | Danger-Associated Molecular Pattern |
| ICIs | Inhibiting immune checkpoints |
References
- Pan, S.-Y.; Litscher, G.; Gao, S.-H.; Zhou, S.-F.; Yu, Z.-L.; Chen, H.-Q.; Zhang, S.-F.; Tang, M.-K.; Sun, J.-N.; Ko, K.-M. Historical Perspective of Traditional Indigenous Medical Practices: The Current Renaissance and Conservation of Herbal Resources. Evid. Based Complement. Alternat. Med. 2014, 2014, 525340. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, L.; Qu, L.; Yu, W.; Liu, T.; Ning, F.; Li, J.; Guo, X.; Sun, F.; Sun, B.; et al. Potential Implications of Natural Compounds on Aging and Metabolic Regulation. Ageing Res. Rev. 2024, 101, 102475. [Google Scholar] [CrossRef]
- Merenda, T.; Juszczak, F.; Ferier, E.; Duez, P.; Patris, S.; Declèves, A.-É.; Nachtergael, A. Natural Compounds Proposed for the Management of Non-Alcoholic Fatty Liver Disease. Nat. Prod. Bioprospecting 2024, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Pezzino, S.; Sofia, M.; Mazzone, C.; Litrico, G.; Greco, L.P.; Gallo, L.; La Greca, G.; Latteri, S. Innovative Treatments for Obesity and NAFLD: A Bibliometric Study on Antioxidants, Herbs, Phytochemicals, and Natural Compounds. Heliyon 2024, 10, e35498. [Google Scholar] [CrossRef]
- Nguyen-Thi, P.-T.; Vo, T.K.; Pham, T.H.T.; Nguyen, T.T.; Van Vo, G. Natural Flavonoids as Potential Therapeutics in the Management of Alzheimer’s Disease: A Review. 3 Biotech 2024, 14, 68. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wu, Q.; Li, S.; Lin, X.; Yang, S.; Zhu, R.; Fu, C.; Zhang, Z. Harnessing Nature’s Pharmacy: Investigating Natural Compounds as Novel Therapeutics for Ulcerative Colitis. Front. Pharmacol. 2024, 15, 1394124. [Google Scholar] [CrossRef]
- Natesan, V.; Kim, S.-J. Natural Compounds in Kidney Disease: Therapeutic Potential and Drug Development. Biomol. Ther. 2025, 33, 39–53. [Google Scholar] [CrossRef]
- Zhang, Y.; Qu, Z.; Zhao, Y.; Zhang, B.; Gong, Y.; Wang, X.; Gao, X.; Wang, D.; Yan, L. The Therapeutic Effect of Natural Compounds on Osteoporosisthrough Ferroptosis. Curr. Med. Chem. 2024, 31, 2629–2648. [Google Scholar] [CrossRef]
- Faheem, M.; Khan, A.; Shah, F.A. Pharmacological Investigation of Natural Compounds for Therapeutic Potential in Neuropathic Pain. Nat. Prod. Res. 2024, 1–25. [Google Scholar] [CrossRef]
- Mazurakova, A.; Koklesova, L.; Samec, M.; Kudela, E.; Kajo, K.; Skuciova, V.; Csizmár, S.H.; Mestanova, V.; Pec, M.; Adamkov, M.; et al. Anti-Breast Cancer Effects of Phytochemicals: Primary, Secondary, and Tertiary Care. EPMA J. 2022, 13, 315–334. [Google Scholar] [CrossRef]
- Mitsiogianni, M.; Koutsidis, G.; Mavroudis, N.; Trafalis, D.T.; Botaitis, S.; Franco, R.; Zoumpourlis, V.; Amery, T.; Galanis, A.; Pappa, A.; et al. The Role of Isothiocyanates as Cancer Chemo-Preventive, Chemo-Therapeutic and Anti-Melanoma Agents. Antioxidants 2019, 8, 106. [Google Scholar] [CrossRef] [PubMed]
- Nagar, A.; Dubey, A.; Sharma, A.; Singh, M. Exploring Promising Natural Compounds for Breast Cancer Treatment: In Silico Molecular Docking Targeting WDR5-MYC Protein Interaction. J. Biomol. Struct. Dyn. 2024, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Asma, S.T.; Acaroz, U.; Imre, K.; Morar, A.; Shah, S.R.A.; Hussain, S.Z.; Arslan-Acaroz, D.; Demirbas, H.; Hajrulai-Musliu, Z.; Istanbullugil, F.R.; et al. Natural Products/Bioactive Compounds as a Source of Anticancer Drugs. Cancers 2022, 14, 6203. [Google Scholar] [CrossRef] [PubMed]
- Saberi-Karimian, M.; Katsiki, N.; Caraglia, M.; Boccellino, M.; Majeed, M.; Sahebkar, A. Vascular Endothelial Growth Factor: An Important Molecular Target of Curcumin. Crit. Rev. Food Sci. Nutr. 2019, 59, 299–312. [Google Scholar] [CrossRef]
- Juturu, V.; Sahin, K.; Pala, R.; Tuzcu, M.; Ozdemir, O.; Orhan, C.; Sahin, N. Curcumin Prevents Muscle Damage by Regulating NF-kB and Nrf2 Pathways and Improves Performance: An in Vivo Model. J. Inflamm. Res. 2016, Volume 9, 147–154. [Google Scholar] [CrossRef]
- Zhang, C.; He, L.; Ye, H.; Liu, D.; Zhu, Y.; Miao, D.; Zhang, S.; Chen, Y.; Jia, Y.; Shen, J.; et al. Nrf2 Is a Key Factor in the Reversal Effect of Curcumin on Multidrug Resistance in the HCT-8/5-Fu Human Colorectal Cancer Cell Line. Mol. Med. Rep. 2018. [Google Scholar] [CrossRef]
- Javed, Z.; Sadia, H.; Iqbal, M.J.; Shamas, S.; Malik, K.; Ahmed, R.; Raza, S.; Butnariu, M.; Cruz-Martins, N.; Sharifi-Rad, J. Apigenin Role as Cell-Signaling Pathways Modulator: Implications in Cancer Prevention and Treatment. Cancer Cell Int. 2021, 21, 189. [Google Scholar] [CrossRef]
- Topçu-Tarladaçalışır, Y.; Sapmaz-Metin, M.; Mercan, Z.; Erçetin, D. Quercetin Attenuates Endoplasmic Reticulum Stress and Apoptosis in TNBS-Induced Colitis by Inhibiting the Glucose Regulatory Protein 78 Activation. Balk. Med. J. 2024, 30–37. [Google Scholar] [CrossRef]
- Benayad, S.; Wahnou, H.; El Kebbaj, R.; Liagre, B.; Sol, V.; Oudghiri, M.; Saad, E.M.; Duval, R.E.; Limami, Y. The Promise of Piperine in Cancer Chemoprevention. Cancers 2023, 15, 5488. [Google Scholar] [CrossRef]
- Murlimanju, B.V. Neuroprotective Effects of Resveratrol in Alzheimer Rsquo s Disease. Front. Biosci. 2020, 12, 139–149. [Google Scholar] [CrossRef]
- Da Silva, G.B.; Manica, D.; Da Silva, A.P.; Marafon, F.; Moreno, M.; Bagatini, M.D. Rosmarinic Acid Decreases Viability, Inhibits Migration and Modulates Expression of Apoptosis-Related CASP8/CASP3/NLRP3 Genes in Human Metastatic Melanoma Cells. Chem. Biol. Interact. 2023, 375, 110427. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-Y.; Hsu, T.-W.; Chen, Y.-R.; Kao, S.-H. Rosmarinic Acid Potentiates Cytotoxicity of Cisplatin against Colorectal Cancer Cells by Enhancing Apoptotic and Ferroptosis. Life 2024, 14, 1017. [Google Scholar] [CrossRef]
- Anand, P.; Thomas, S.G.; Kunnumakkara, A.B.; Sundaram, C.; Harikumar, K.B.; Sung, B.; Tharakan, S.T.; Misra, K.; Priyadarsini, I.K.; Rajasekharan, K.N.; et al. Biological Activities of Curcumin and Its Analogues (Congeners) Made by Man and Mother Nature. Biochem. Pharmacol. 2008, 76, 1590–1611. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Gilani, A. Therapeutic Potential of Turmeric in Alzheimer’s Disease: Curcumin or Curcuminoids? Phytother. Res. 2014, 28, 517–525. [Google Scholar] [CrossRef]
- Ammon, H.; Wahl, M. Pharmacology of Curcuma Longa. Planta Med. 1991, 57, 1–7. [Google Scholar] [CrossRef]
- Lu, K.-H.; Lu, P.W.-A.; Lu, E.W.-H.; Lin, C.-W.; Yang, S.-F. Curcumin and Its Analogs and Carriers: Potential Therapeutic Strategies for Human Osteosarcoma. Int. J. Biol. Sci. 2023, 19, 1241–1265. [Google Scholar] [CrossRef]
- Pei, H.; Yang, Y.; Cui, L.; Yang, J.; Li, X.; Yang, Y.; Duan, H. Bisdemethoxycurcumin Inhibits Ovarian Cancer via Reducing Oxidative Stress Mediated MMPs Expressions. Sci. Rep. 2016, 6, 28773. [Google Scholar] [CrossRef]
- Sandur, S.K.; Pandey, M.K.; Sung, B.; Ahn, K.S.; Murakami, A.; Sethi, G.; Limtrakul, P.; Badmaev, V.; Aggarwal, B.B. Curcumin, Demethoxycurcumin, Bisdemethoxycurcumin, Tetrahydrocurcumin and Turmerones Differentially Regulate Anti-Inflammatory and Anti-Proliferative Responses through a ROS-Independent Mechanism. Carcinogenesis 2007, 28, 1765–1773. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.-Y.; Majeed, A.; Ho, C.-T.; Pan, M.-H. Bisdemethoxycurcumin and Curcumin Alleviate Inflammatory Bowel Disease by Maintaining Intestinal Epithelial Integrity and Regulating Gut Microbiota in Mice. J. Agric. Food Chem. 2025, 73, 3494–3506. [Google Scholar] [CrossRef]
- Thanawala, S.; Shah, R.; Alluri, K.V.; Somepalli, V.; Vaze, S.; Upadhyay, V. Comparative Bioavailability of Curcuminoids from a Water-Dispersible High Curcuminoid Turmeric Extract against a Generic Turmeric Extract: A Randomized, Cross-over, Comparative, Pharmacokinetic Study. J. Pharm. Pharmacol. 2021, 73, 816–823. [Google Scholar] [CrossRef]
- Mohankumar, K.; Sridharan, S.; Pajaniradje, S.; Singh, V.K.; Ronsard, L.; Banerjea, A.C.; Somasundaram, D.B.; Coumar, M.S.; Periyasamy, L.; Rajagopalan, R. BDMC-A, an Analog of Curcumin, Inhibits Markers of Invasion, Angiogenesis, and Metastasis in Breast Cancer Cells via NF-κB Pathway—A Comparative Study with Curcumin. Biomed. Pharmacother. 2015, 74, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Lu, H.-F.; Chen, Y.-H.; Chen, J.-C.; Chou, W.-H.; Huang, H.-C. Curcumin, Demethoxycurcumin, and Bisdemethoxycurcumin Induced Caspase-Dependent and –Independent Apoptosis via Smad or Akt Signaling Pathways in HOS Cells. BMC Complement. Med. Ther. 2020, 20, 68. [Google Scholar] [CrossRef]
- Garbe, C.; Amaral, T.; Peris, K.; Hauschild, A.; Arenberger, P.; Basset-Seguin, N.; Bastholt, L.; Bataille, V.; del Marmol, V.; Dréno, B.; et al. European Consensus-Based Interdisciplinary Guideline for Melanoma. Part 1: Diagnostics: Update 2022. Eur. J. Cancer 2022, 170, 236–255. [Google Scholar] [CrossRef]
- Passarelli, A.; Tucci, M.; Mannavola, F.; Felici, C.; Silvestris, F. The Metabolic Milieu in Melanoma: Role of Immune Suppression by CD73/Adenosine. Tumor Biol. 2019, 41, 101042831983713. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho Braga, G.; Coiado, J.V.; De Melo, V.C.; Loureiro, B.B.; Bagatini, M.D. Cutaneous Melanoma and Purinergic Modulation by Phenolic Compounds. Purinergic Signal. 2024, 20, 581–593. [Google Scholar] [CrossRef]
- Manica, D.; Da Silva, G.B.; Narzetti, R.A.; Dallagnoll, P.; Da Silva, A.P.; Marafon, F.; Cassol, J.; De Souza Matias, L.; Zamoner, A.; De Oliveira Maciel, S.F.V.; et al. Curcumin Modulates Purinergic Signaling and Inflammatory Response in Cutaneous Metastatic Melanoma Cells. Purinergic Signal. 2024. [Google Scholar] [CrossRef]
- Monteiro, I.; Vigano, S.; Faouzi, M.; Treilleux, I.; Michielin, O.; Ménétrier-Caux, C.; Caux, C.; Romero, P.; De Leval, L. CD73 Expression and Clinical Significance in Human Metastatic Melanoma. Oncotarget 2018, 9, 26659–26669. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Di Virgilio, F. Purinergic Signalling and Cancer. Purinergic Signal. 2013, 9, 491–540. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- McGahon, A.J.; Martin, S.J.; Bissonnette, R.P.; Mahboubi, A.; Shi, Y.; Mogil, R.J.; Nishioka, W.K.; Green, D.R. Chapter 9 The End of the (Cell) Line: Methods for the Study of Apoptosis in Vitro. In Methods in Cell Biology; Elsevier, 1995; Vol. 46, pp. 153–185 ISBN 978-0-12-564147-0.
- Joshi, D.C.; Bakowska, J.C. Determination of Mitochondrial Membrane Potential and Reactive Oxygen Species in Live Rat Cortical Neurons. J. Vis. Exp. 2011, 2704. [Google Scholar] [CrossRef]
- Justus, C.R.; Leffler, N.; Ruiz-Echevarria, M.; Yang, L.V. In Vitro Cell Migration and Invasion Assays. J. Vis. Exp. 2014, 51046. [Google Scholar] [CrossRef]
- Manica, D.; Silva, G.B.D.; Silva, A.P.D.; Marafon, F.; Maciel, S.F.V.D.O.; Bagatini, M.D.; Moreno, M. Curcumin Promotes Apoptosis of Human Melanoma Cells by Caspase 3. Cell Biochem. Funct. 2023, 41, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Pilla, C.; Emanuelli, T.; Frassetto, S.S.; Battastini, A.M.O.; Dias, R.D.; Sarkis, J.J.F. ATP Diphosphohydrolase Activity (Apyrase, EC 3.6.1.5) in Human Blood Platelets. Platelets 1996, 7, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Lunkes, G.I.; Lunkes, D.; Stefanello, F.; Morsch, A.; Morsch, V.M.; Mazzanti, C.M.; Schetinger, M.R.C. Enzymes That Hydrolyze Adenine Nucleotides in Diabetes and Associated Pathologies. Thromb. Res. 2003, 109, 189–194. [Google Scholar] [CrossRef]
- Manica, D.; Da Silva, G.B.; De Lima, J.; Cassol, J.; Dallagnol, P.; Narzetti, R.A.; Moreno, M.; Bagatini, M.D. Caffeine Reduces Viability, Induces Apoptosis, Inhibits Migration and Modulates the CD39/CD73 Axis in Metastatic Cutaneous Melanoma Cells. Purinergic Signal. 2023. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Li, M.; Guo, T.; Lin, J.; Huang, X.; Ke, Q.; Wu, Y.; Fang, C.; Hu, C. Curcumin Inhibits the Invasion and Metastasis of Triple Negative Breast Cancer via Hedgehog/Gli1 Signaling Pathway. J. Ethnopharmacol. 2022, 283, 114689. [Google Scholar] [CrossRef]
- Mohamadian, M.; Bahrami, A.; Moradi Binabaj, M.; Asgharzadeh, F.; Ferns, G.A. Molecular Targets of Curcumin and Its Therapeutic Potential for Ovarian Cancer. Nutr. Cancer 2022, 74, 2713–2730. [Google Scholar] [CrossRef]
- Zheng, B.-Z.; Liu, T.-D.; Chen, G.; Zhang, J.-X.; Kang, X. The Effect of Curcumin on Cell Adhesion of Human Esophageal Cancer Cell. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 551–560. [Google Scholar] [CrossRef]
- Zhao, G.; Han, X.; Zheng, S.; Li, Z.; Sha, Y.; Ni, J.; Sun, Z.; Qiao, S.; Song, Z. Curcumin Induces Autophagy, Inhibits Proliferation and Invasion by Downregulating AKT/mTOR Signaling Pathway in Human Melanoma Cells. Oncol. Rep. 2016, 35, 1065–1074. [Google Scholar] [CrossRef]
- Qiu, Y.; Yu, T.; Wang, W.; Pan, K.; Shi, D.; Sun, H. Curcumin-Induced Melanoma Cell Death Is Associated with Mitochondrial Permeability Transition Pore (mPTP) Opening. Biochem. Biophys. Res. Commun. 2014, 448, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Song, E.; Hu, D.-N.; Chen, M.; Xue, C.; Rosen, R.; McCormick, S.A. Curcumin Induces Cell Death in Human Uveal Melanoma Cells through Mitochondrial Pathway. Curr. Eye Res. 2010, 35, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.-J.; Kim, K.; Kim, C.; Lee, S.-E. Antimelanogenic Effects of Curcumin and Its Dimethoxy Derivatives: Mechanistic Investigation Using B16F10 Melanoma Cells and Zebrafish (Danio Rerio) Embryos. Foods 2023, 12, 926. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-J.; Yang, Z.-X.; Dai, X.-T.; Chen, Y.-F.; Yang, H.-P.; Zhou, X.-D. Bisdemethoxycurcumin Sensitizes Cisplatin-Resistant Lung Cancer Cells to Chemotherapy by Inhibition of CA916798 and PI3K/AKT Signaling. Apoptosis 2017, 22, 1157–1168. [Google Scholar] [CrossRef]
- Liao, C.-L.; Chu, Y.L.; Lin, H.-Y.; Chen, C.-Y.; Hsu, M.-J.; Liu, K.-C.; Lai, K.-C.; Huang, A.-C.; Chung, J.-G. Bis Demethoxycurcumin Suppresses Migration and Invasion of Human Cervical Cancer HeLa Cells via Inhibition of NF-ĸB, MMP-2 and -9 Pathways. Anticancer Res. 2018, 38, 3989–3997. [Google Scholar] [CrossRef]
- Rossi, S.; Cordella, M.; Tabolacci, C.; Nassa, G.; D’Arcangelo, D.; Senatore, C.; Pagnotto, P.; Magliozzi, R.; Salvati, A.; Weisz, A.; et al. TNF-Alpha and Metalloproteases as Key Players in Melanoma Cells Aggressiveness. J. Exp. Clin. Cancer Res. 2018, 37, 326. [Google Scholar] [CrossRef]
- Lunkes, V.L.; Palma, T.V.; Mostardeiro, V.B.; Mastella, M.H.; Assmann, C.E.; Pillat, M.M.; Cruz, I.B.M. da; Morsch, V.M.M.; Chitolina, M.R.; Andrade, C.M. de Curcumin and Vinblastine Induce Apoptosis and Impair Migration in Human Cutaneous Melanoma Cells. Res. Soc. Dev. 2022, 11, e20511225611. [Google Scholar] [CrossRef]
- Szlasa, W.; Supplitt, S.; Drąg-Zalesińska, M.; Przystupski, D.; Kotowski, K.; Szewczyk, A.; Kasperkiewicz, P.; Saczko, J.; Kulbacka, J. Effects of Curcumin Based PDT on the Viability and the Organization of Actin in Melanotic (A375) and Amelanotic Melanoma (C32) – in Vitro Studies. Biomed. Pharmacother. 2020, 132, 110883. [Google Scholar] [CrossRef] [PubMed]
- Schlichting, L.; Valentini, G.; Santos, B.M.; Sanches, M.P.; Soares, R.V.; Saatkamp, R.H.; Kviecinski, M.R.; Zamoner, A.; Parize, A.L. Chitosan/Hydroxypropyl Methylcellulose Acetate Succinate Nanoparticles as a Promising Delivery System for Curcumin in MDA-MB-231 Breast Cancer Cells. J. Mol. Liq. 2025, 429, 127600. [Google Scholar] [CrossRef]
- Kocyigit, A.; Guler, E.M. Curcumin Induce DNA Damage and Apoptosis through Generation of Reactive Oxygen Species and Reducing Mitochondrial Membrane Potential in Melanoma Cancer Cells. Cell. Mol. Biol. 2017, 63, 97–105. [Google Scholar] [CrossRef]
- Liao, W.; Xiang, W.; Wang, F.-F.; Wang, R.; Ding, Y. Curcumin Inhibited Growth of Human Melanoma A375 Cells via Inciting Oxidative Stress. Biomed. Pharmacother. 2017, 95, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, Z.; Shukla, Y. Death Receptors: Targets for Cancer Therapy. Exp. Cell Res. 2010, 316, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Kiraz, Y.; Adan, A.; Kartal Yandim, M.; Baran, Y. Major Apoptotic Mechanisms and Genes Involved in Apoptosis. Tumor Biol. 2016, 37, 8471–8486. [Google Scholar] [CrossRef]
- Tyciakova, S.; Valova, V.; Svitkova, B.; Matuskova, M. Overexpression of TNFα Induces Senescence, Autophagy and Mitochondrial Dysfunctions in Melanoma Cells. BMC Cancer 2021, 21, 507. [Google Scholar] [CrossRef] [PubMed]
- Hamarsheh, S.; Zeiser, R. NLRP3 Inflammasome Activation in Cancer: A Double-Edged Sword. Front. Immunol. 2020, 11, 1444. [Google Scholar] [CrossRef]
- Van Deventer, H.W.; Burgents, J.E.; Wu, Q.P.; Woodford, R.-M.T.; Brickey, W.J.; Allen, I.C.; McElvania-Tekippe, E.; Serody, J.S.; Ting, J.P.-Y. The Inflammasome Component Nlrp3 Impairs Antitumor Vaccine by Enhancing the Accumulation of Tumor-Associated Myeloid-Derived Suppressor Cells. Cancer Res. 2010, 70, 10161–10169. [Google Scholar] [CrossRef]
- Dunn, J.H.; Ellis, L.Z.; Fujita, M. Inflammasomes as Molecular Mediators of Inflammation and Cancer: Potential Role in Melanoma. Cancer Lett. 2012, 314, 24–33. [Google Scholar] [CrossRef]
- Pazhouhesh Far, N.; Hajiheidari Varnousafaderani, M.; Faghihkhorasani, F.; Etemad, S.; Abdulwahid, A.R.R.; Bakhtiarinia, N.; Mousaei, A.; Dortaj, E.; Karimi, S.; Ebrahimi, N.; et al. Breaking the Barriers: Overcoming Cancer Resistance by Targeting the NLRP3 Inflammasome. Br. J. Pharmacol. 2025, 182, 3–25. [Google Scholar] [CrossRef]
- Yegutkin, G.G. Nucleotide- and Nucleoside-Converting Ectoenzymes: Important Modulators of Purinergic Signalling Cascade. Biochim. Biophys. Acta BBA - Mol. Cell Res. 2008, 1783, 673–694. [Google Scholar] [CrossRef]
- Wu, L.; Xie, W.; Li, Y.; Ni, Q.; Timashev, P.; Lyu, M.; Xia, L.; Zhang, Y.; Liu, L.; Yuan, Y.; et al. Biomimetic Nanocarriers Guide Extracellular ATP Homeostasis to Remodel Energy Metabolism for Activating Innate and Adaptive Immunity System. Adv. Sci. 2022, 9, 2105376. [Google Scholar] [CrossRef]
- Fredholm, B.B.; IJzerman, A.P.; Jacobson, K.A.; Linden, J.; Müller, C.E. International Union of Basic and Clinical Pharmacology. LXXXI. Nomenclature and Classification of Adenosine Receptors—An Update. Pharmacol. Rev. 2011, 63, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Boison, D.; Yegutkin, G.G. Adenosine Metabolism: Emerging Concepts for Cancer Therapy. Cancer Cell 2019, 36, 582–596. [Google Scholar] [CrossRef] [PubMed]
- Faas, M.M.; Sáez, T.; De Vos, P. Extracellular ATP and Adenosine: The Yin and Yang in Immune Responses? Mol. Aspects Med. 2017, 55, 9–19. [Google Scholar] [CrossRef]
- Ahmed, A.; Tait, S.W.G. Targeting Immunogenic Cell Death in Cancer. Mol. Oncol. 2020, 14, 2994–3006. [Google Scholar] [CrossRef]
- Vijayan, D.; Young, A.; Teng, M.W.L.; Smyth, M.J. Targeting Immunosuppressive Adenosine in Cancer. Nat. Rev. Cancer 2017, 17, 709–724. [Google Scholar] [CrossRef]
- Ziogas, D.C.; Theocharopoulos, C.; Koutouratsas, T.; Haanen, J.; Gogas, H. Mechanisms of Resistance to Immune Checkpoint Inhibitors in Melanoma: What We Have to Overcome? Cancer Treat. Rev. 2023, 113, 102499. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Yin, S.; To, K.K.W.; Fu, L. CD39/CD73/A2AR Pathway and Cancer Immunotherapy. Mol. Cancer 2023, 22, 44. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, M.; Calaf, G.M. Curcumin Inhibits Invasive Capabilities through Epithelial Mesenchymal Transition in Breast Cancer Cell Lines. Int. J. Oncol. 2016, 49, 1019–1027. [Google Scholar] [CrossRef]
- Giraulo, C.; Orlando, L.; Morretta, E.; Voli, A.; Plaitano, P.; Cicala, C.; Potaptschuk, E.; Müller, C.E.; Tosco, A.; Monti, M.C.; et al. High Levels of Soluble CD73 Unveil Resistance to BRAF Inhibitors in Melanoma Cells. Biomed. Pharmacother. 2024, 177, 117033. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, A.; Gupte, A.A.; Hamilton, D.J. Plumbagin Elicits Cell-Specific Cytotoxic Effects and Metabolic Responses in Melanoma Cells. Pharmaceutics 2021, 13, 706. [Google Scholar] [CrossRef]
- Soleimani, A.; Farshchi, H.K.; Mirzavi, F.; Zamani, P.; Ghaderi, A.; Amini, Y.; Khorrami, S.; Mashayekhi, K.; Jaafari, M.R. The Therapeutic Potential of Targeting CD73 and CD73-Derived Adenosine in Melanoma. Biochimie 2020, 176, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Carlino, M.S.; Larkin, J.; Long, G.V. Immune Checkpoint Inhibitors in Melanoma. The Lancet 2021, 398, 1002–1014. [Google Scholar] [CrossRef] [PubMed]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.-J.; Rutkowski, P.; Lao, C.D.; Cowey, C.L.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Five-Year Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2019, 381, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Tu, E.; McGlinchey, K.; Wang, J.; Martin, P.; Ching, S.L.K.; Floc’h, N.; Kurasawa, J.; Starrett, J.H.; Lazdun, Y.; Wetzel, L.; et al. Anti–PD-L1 and Anti-CD73 Combination Therapy Promotes T Cell Response to EGFR-Mutated NSCLC. JCI Insight 2022, 7, e142843. [Google Scholar] [CrossRef] [PubMed]
Figure 01.
Antiproliferative and migratory activity of AC3TM in CM A375 and SK-MEL-28 cells. In the A375 cell line, the MTT assay indicated that treatment concentrations with 3.12μg/mL and 6.12μg/mL of AC3TM reduced cell viability after 24 hours of treatment (A); similar results were found in fluorescence microscopy (B). In the cell migration assay, we observed a reduction in cell migration at concentrations of 1.56μg/mL, 3.12μg/mL, and 6.25μg/mL (C, D). In the SK-MEL-28 cell line, we found that treatment with 6.25μg/mL of AC3 decreased cell viability (E, F). Furthermore, all treatment concentrations effectively reduced CM cell migration (G, H). Data are presented as mean ± SD. Statistical analysis: ANOVA. Values with P<0.05 were considered statistically significant. *(P<0.05) **(P<0.01) ***(P<0.001) ****(P<0.0001).
Figure 01.
Antiproliferative and migratory activity of AC3TM in CM A375 and SK-MEL-28 cells. In the A375 cell line, the MTT assay indicated that treatment concentrations with 3.12μg/mL and 6.12μg/mL of AC3TM reduced cell viability after 24 hours of treatment (A); similar results were found in fluorescence microscopy (B). In the cell migration assay, we observed a reduction in cell migration at concentrations of 1.56μg/mL, 3.12μg/mL, and 6.25μg/mL (C, D). In the SK-MEL-28 cell line, we found that treatment with 6.25μg/mL of AC3 decreased cell viability (E, F). Furthermore, all treatment concentrations effectively reduced CM cell migration (G, H). Data are presented as mean ± SD. Statistical analysis: ANOVA. Values with P<0.05 were considered statistically significant. *(P<0.05) **(P<0.01) ***(P<0.001) ****(P<0.0001).
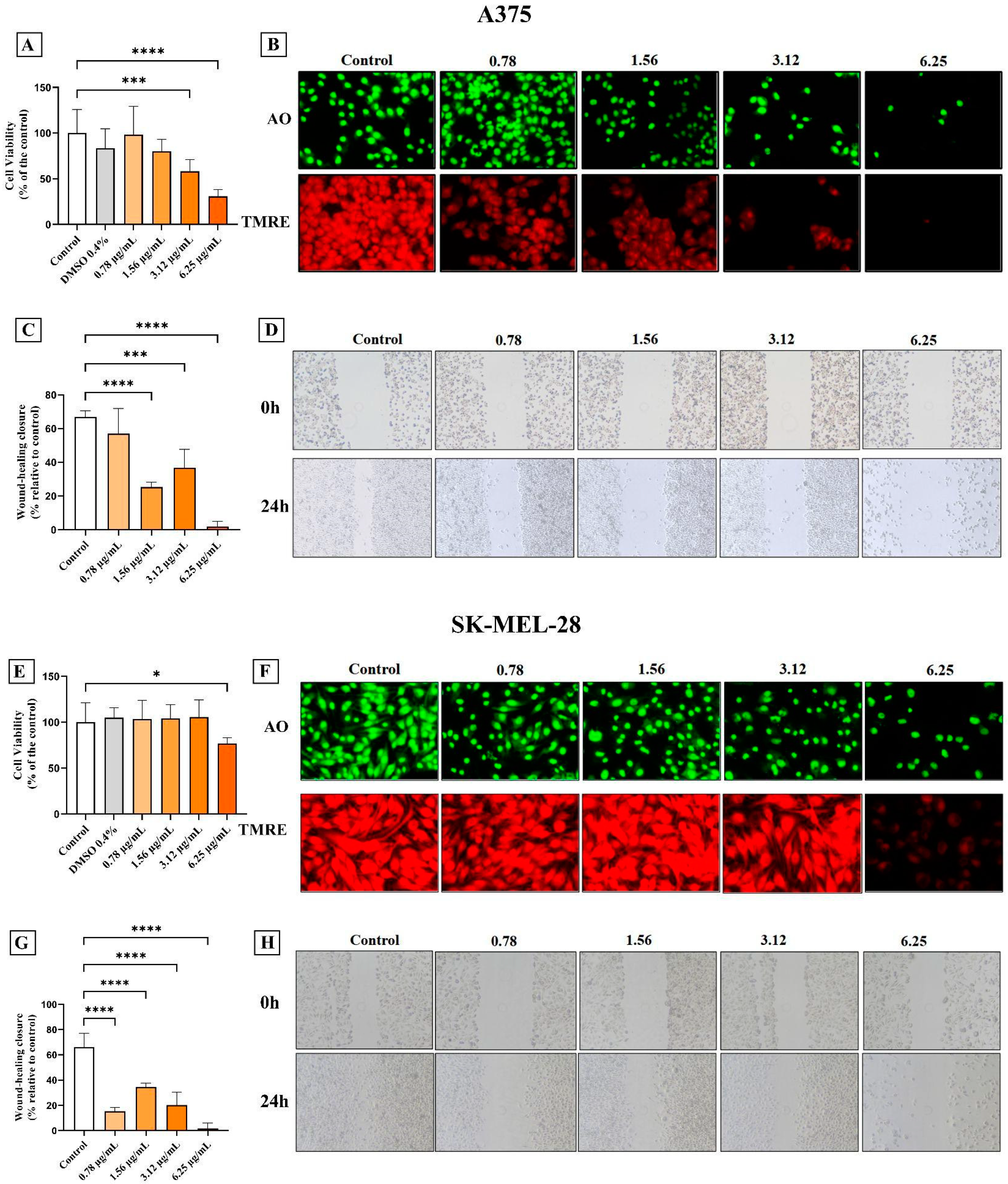
Figure 02.
AC3TM modulates ROS levels and caspase expression in CM A375 and SK-MEL-28 cells. Treatment with all AC3TM concentrations reduced ROS levels in A375 (A) and SK-MEL-28 (D) cell lines. We did not observe a significant change in Caspase-8 gene expression levels in A375 (B) cells, whereas, in SK-MEL-28 cells, treatment with 3.12μg/mL of AC3TM increased gene expression (E). Caspase-3 gene expression significantly increased after treatment with 6.25μg/mL of AC3 in A375 (C) and SK-MEL-28 (F) cells. Data are presented as mean ± SD. Statistical analysis: ANOVA. Values with P<0.05 were considered statistically significant. *(P<0.05) **(P<0.01) ***(P<0.001) ****(P<0.0001).
Figure 02.
AC3TM modulates ROS levels and caspase expression in CM A375 and SK-MEL-28 cells. Treatment with all AC3TM concentrations reduced ROS levels in A375 (A) and SK-MEL-28 (D) cell lines. We did not observe a significant change in Caspase-8 gene expression levels in A375 (B) cells, whereas, in SK-MEL-28 cells, treatment with 3.12μg/mL of AC3TM increased gene expression (E). Caspase-3 gene expression significantly increased after treatment with 6.25μg/mL of AC3 in A375 (C) and SK-MEL-28 (F) cells. Data are presented as mean ± SD. Statistical analysis: ANOVA. Values with P<0.05 were considered statistically significant. *(P<0.05) **(P<0.01) ***(P<0.001) ****(P<0.0001).
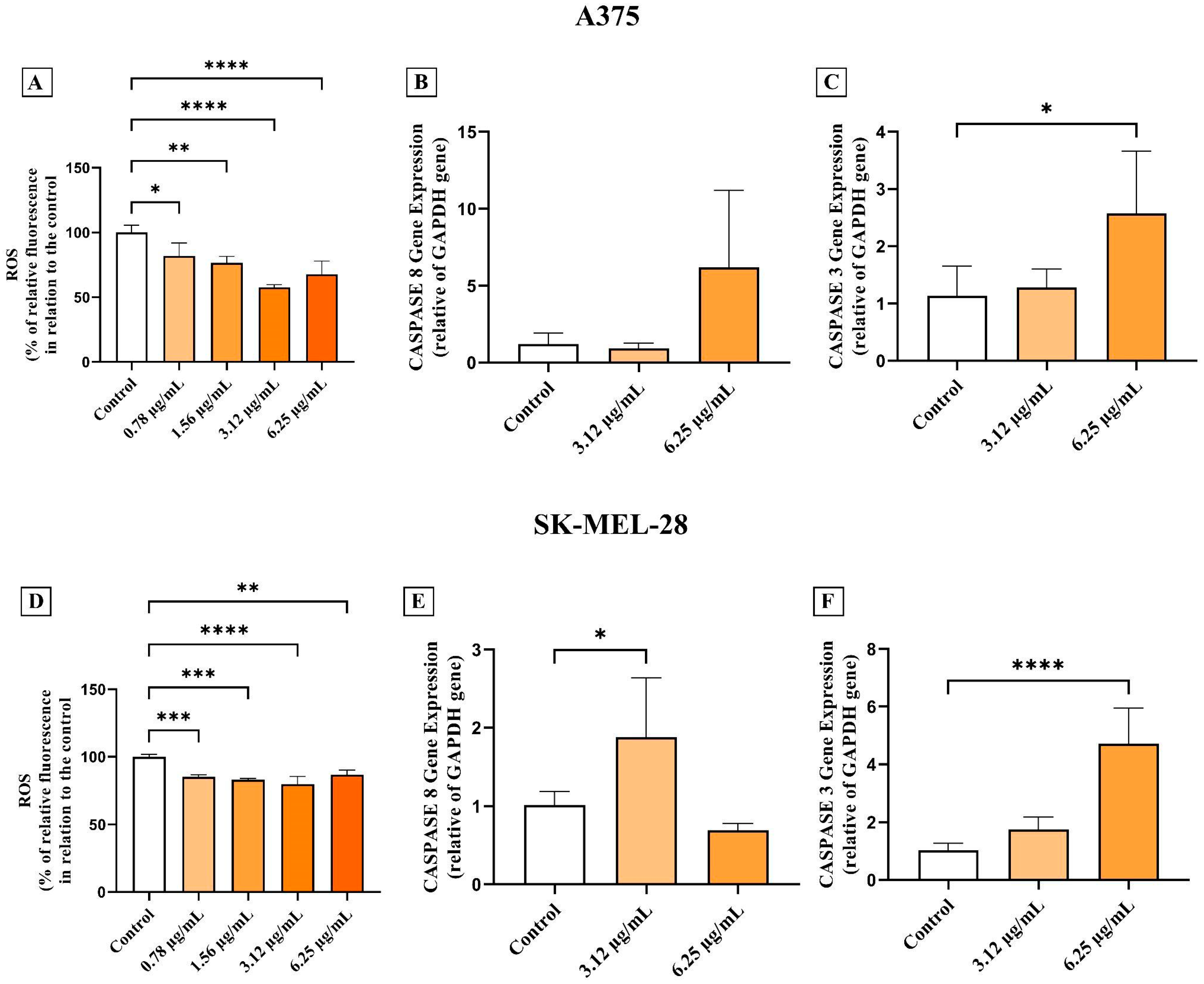
Figure 03.
AC3TM modulates the levels of pro-inflammatory markers in CM A375 and SK-MEL-28 cell lines. In A375 cells, treatment with 6.25 μg/mL of AC3TM increased the gene expression of TNF (A), IL-6 (B), and NLRP3 (C). In SK-MEL-28 cells, we observed similar results in TNF levels (D). In contrast, IL-6 levels (E) did not show significant differences, while there was a reduction in NLRP3 gene expression levels after 24 hours of treatment with 6.25 μg/mL of AC3TM (F). Data are presented as mean ± SD. Statistical analysis: ANOVA. Values with P<0.05 were considered statistically significant. *(P<0.05) **(P<0.01) ***(P<0.001) ****(P<0.0001).
Figure 03.
AC3TM modulates the levels of pro-inflammatory markers in CM A375 and SK-MEL-28 cell lines. In A375 cells, treatment with 6.25 μg/mL of AC3TM increased the gene expression of TNF (A), IL-6 (B), and NLRP3 (C). In SK-MEL-28 cells, we observed similar results in TNF levels (D). In contrast, IL-6 levels (E) did not show significant differences, while there was a reduction in NLRP3 gene expression levels after 24 hours of treatment with 6.25 μg/mL of AC3TM (F). Data are presented as mean ± SD. Statistical analysis: ANOVA. Values with P<0.05 were considered statistically significant. *(P<0.05) **(P<0.01) ***(P<0.001) ****(P<0.0001).
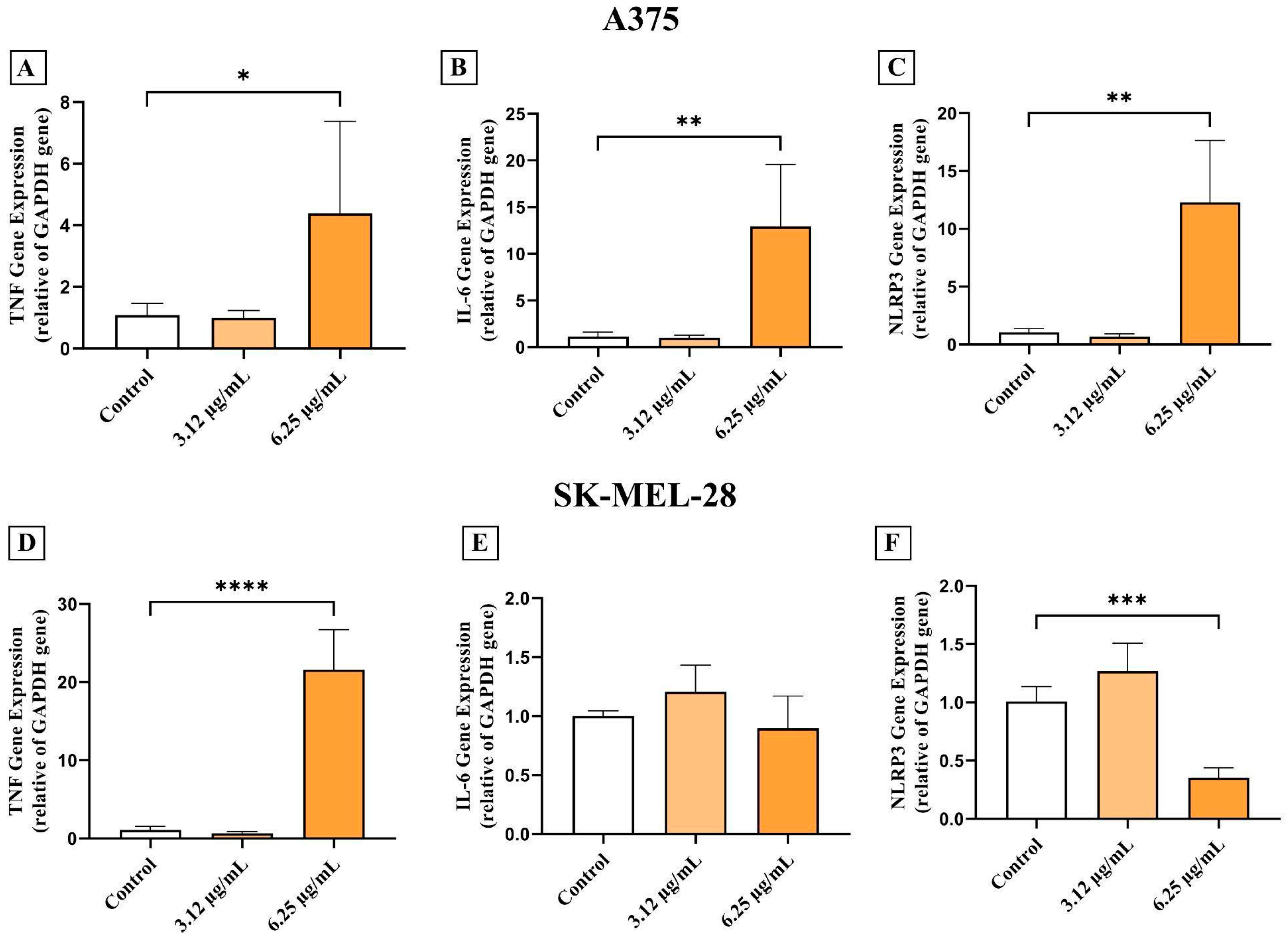
Figure 04.
AC3TM modulates the levels of pro-inflammatory markers in CM A375 and SK-MEL-28 cell lines. CD39 gene expression levels increased after AC3TM (A) treatment in A375 cells. The same occurred in CD73 levels after treatment with 6.25 μg/mL of the compound (B). The highest concentration tested in the SK-MEL-28 cell line decreased CD39 (C) and CD73 (D) gene expression. Data are presented as mean ± SD. Statistical analysis: ANOVA. Values with P<0.05 were considered statistically significant. *(P<0.05) **(P<0.01) ***(P<0.001) ****(P<0.0001).
Figure 04.
AC3TM modulates the levels of pro-inflammatory markers in CM A375 and SK-MEL-28 cell lines. CD39 gene expression levels increased after AC3TM (A) treatment in A375 cells. The same occurred in CD73 levels after treatment with 6.25 μg/mL of the compound (B). The highest concentration tested in the SK-MEL-28 cell line decreased CD39 (C) and CD73 (D) gene expression. Data are presented as mean ± SD. Statistical analysis: ANOVA. Values with P<0.05 were considered statistically significant. *(P<0.05) **(P<0.01) ***(P<0.001) ****(P<0.0001).
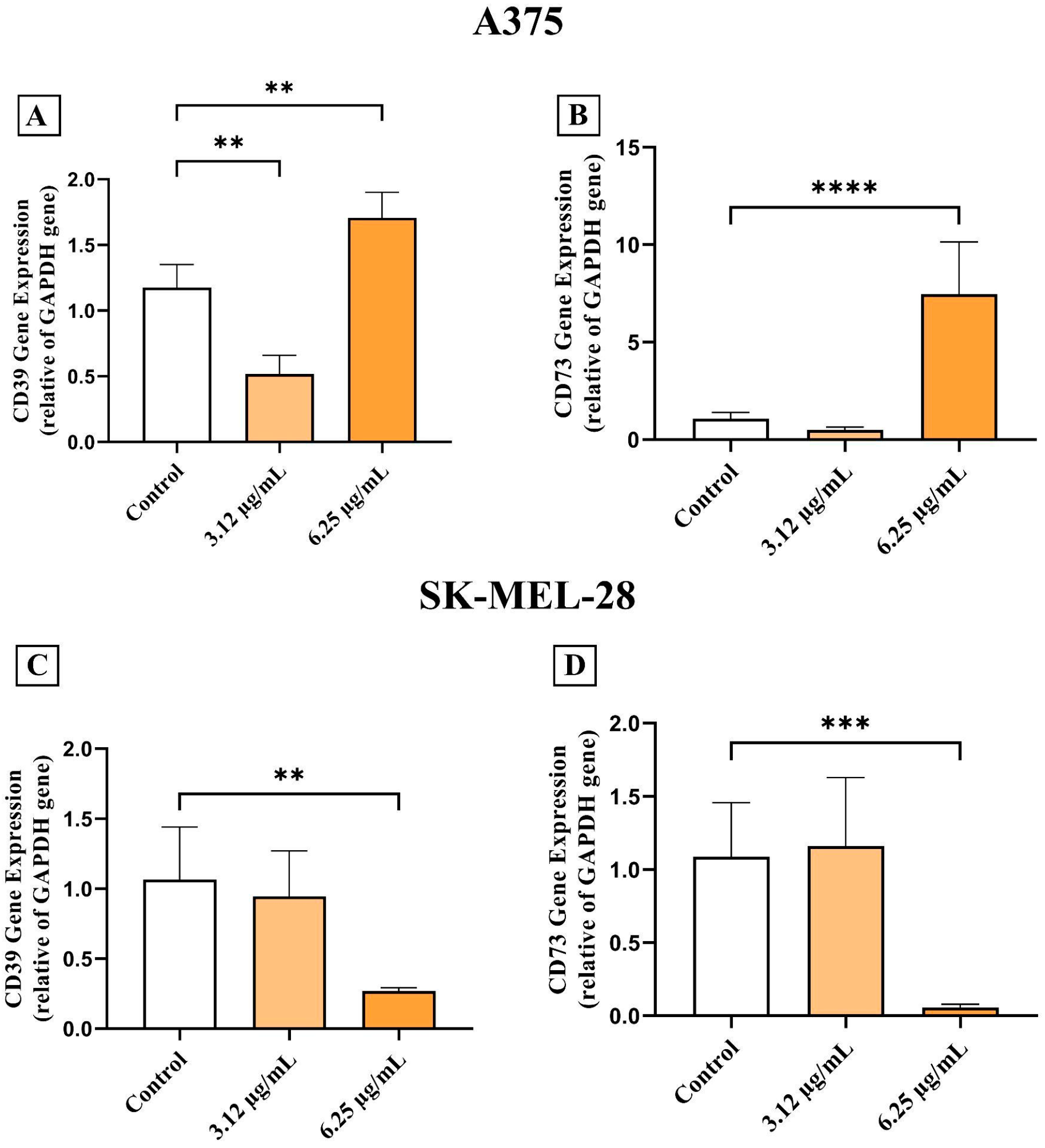
Figure 05.
AC3TM modulates the enzymatic activity of CD39 and CD73 in CM A375 and SK-MEL-28 cell lines. Regarding enzymatic activity, we observed an increase in ATP hydrolysis after treatment with AC3TM in the A375 cell line (A). In contrast, in SK-MEL-28 cells, we observed an increase in ATP hydrolysis at a concentration of 3.12 μg/mL, followed by a decrease in the levels of 6.12 μg/mL (D). The same occurred in the levels of ADP hydrolysis (E). We did not observe significant differences in the levels of ADP (B) and AMP (C) hydrolysis in the A375 cell line or AMP hydrolysis (F) in SK-MEL-28 cells. Data are presented as mean ± SD. Statistical analysis: ANOVA. Values with P<0.05 were considered statistically significant. *(P<0.05) **(P<0.01) ***(P<0.001) ****(P<0.0001).
Figure 05.
AC3TM modulates the enzymatic activity of CD39 and CD73 in CM A375 and SK-MEL-28 cell lines. Regarding enzymatic activity, we observed an increase in ATP hydrolysis after treatment with AC3TM in the A375 cell line (A). In contrast, in SK-MEL-28 cells, we observed an increase in ATP hydrolysis at a concentration of 3.12 μg/mL, followed by a decrease in the levels of 6.12 μg/mL (D). The same occurred in the levels of ADP hydrolysis (E). We did not observe significant differences in the levels of ADP (B) and AMP (C) hydrolysis in the A375 cell line or AMP hydrolysis (F) in SK-MEL-28 cells. Data are presented as mean ± SD. Statistical analysis: ANOVA. Values with P<0.05 were considered statistically significant. *(P<0.05) **(P<0.01) ***(P<0.001) ****(P<0.0001).
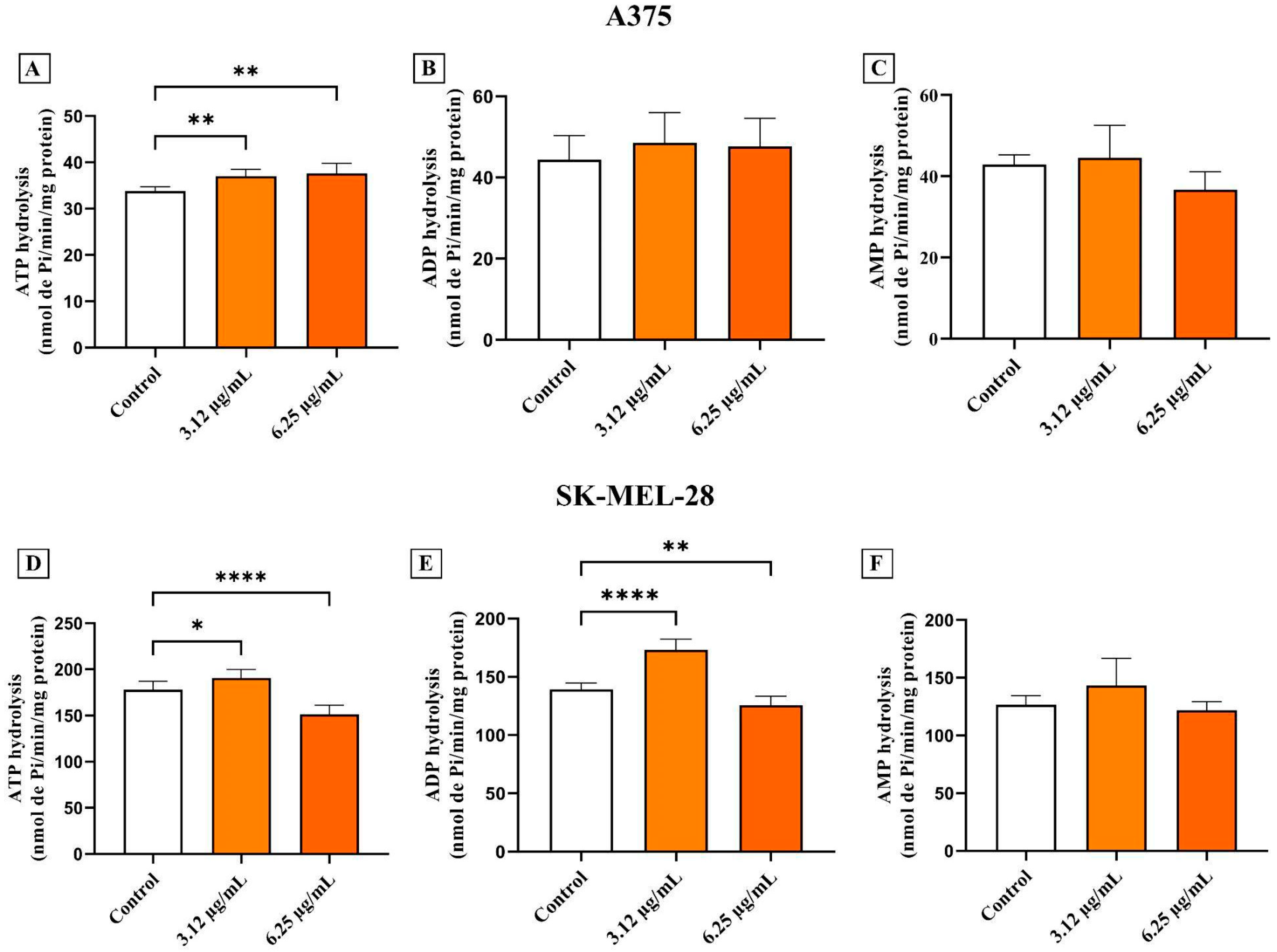

Table 1.
Primer sequences for RT-qPCR.
| Gene | Forward | Reverse |
|---|---|---|
| GAPDH | CTCCTCACAGTTGCCATGTA | GTTGAGCACAGGGTACTTTATTG |
| CASP-8 | AGGAGCTGCTCTTCCGAATT | CCCTGCCTGGTGTCTGAAGT |
| CASP-3 | TTTGAGCCTGAGCAGAGACATG | TACCAGTGCGTATGGAGAAATGG |
| TNF-α | CAGGCAGTCAGATCATCTTC | GCTTGAGGGTTTGCTACAAC |
| IL-6 | TCATCCCATAGCCCAGAGCA | CTGGCATTTGTGGTTGGGTC |
| NLRP3 | AACATGCCCAAGGAGGAAGA | GGCTGTTCACCAATCCATGA |
| CD39 | GCCCTGGTCTTCAGTGTATTAG | CTGGCATAACCTACCTACTCTTTC |
| CD73 | GTGCCTTTGATGAGTCAGGTAG | TTCCTTTCTCTCGTGTCCTTTG |
Abbreviations: Glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Caspase-8 (CASP-8). Caspase-3 (CASP-3). Tumor Necrosis Factor Alpha (TNF-α). Interleukin 6 (IL-6). NLR Family Pyrin Domain Containing 3 (NLRP3). Ectonucleoside triphosphate diphosphohydrolase 1 (CD39). Ecto-5′-nucleotidase (CD73). Adenosine deaminase (ADA).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



