
Submitted:
14 November 2024
Posted:
18 November 2024
You are already at the latest version
Abstract
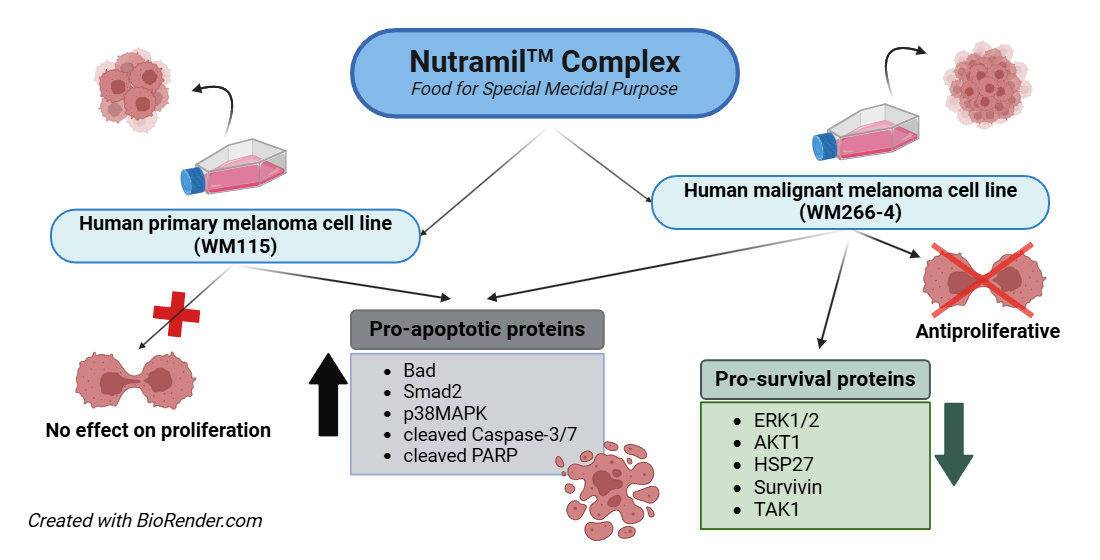
Melanoma malignum is considered the most dangerous form of skin cancer, characterized by the exceptional resistance to many conventional chemotherapies. The aim of the study was to evaluate the effect of NutramilTM Complex (NC) – Food for Special Medical Purpose (FSMP), on two types of melanoma cell lines, primary WM115 and malignant WM266-4. 24h after seeding, growth medium was replaced with a medium containing encoded treatments of NC or NC-CC (NutramilTM Complex without calcium caseinate) at various concentrations. Cells were treated for 24, 48 and 72 h. Our results showed that NutramilTM Complex reduces proliferation of malignant melanoma WM266-4 cells but did not affected the proliferation of WM115 primary melanoma. This was followed by measured downregulation of selected pro-survival proteins expression in WM266-4 cells; specifically ERK1/2, AKT-1, HSP27, Survivin and TAK1. Interestingly, our results showed elevated levels of some pro-apoptotic proteins in both cell lines, including Bad, Smad2, p38MAPK, cleaved forms of Caspase-3/7, as well as cleaved PARP. Taken together, our results indicate that various melanoma cancer cell lines may respond in a different way to the same compound. They also suggest induction of apoptotic pathway by NutramilTM Complex as most likely mechanism of its anticarcinogenic activity.
Keywords:
NutramilTM complex
; FSMP
; melanoma
; apoptosis
; cellular stress response
1. Introduction
Melanoma is one of the most malignant human cancers, with rising incidence rates worldwide, accounted for 1.7% of all cancers in 2020 [1]. In the International Agency for Research on Cancer's latest GLOBOCAN 2022 study, melanoma ranked 17th among the most common cancers worldwide and 22nd in terms of mortality. [2]. Regarding the US, the American Cancer Society estimated 99.780 new cases and mortality at about 7.650 cancer deaths in the United States [3].
Melanoma cells are characterized by exceptional therapy resistance; thus, in addition to research on new pharmacological strategies, studies are carried out on cancer prevention, including the effect of diet. Results of those studies, mainly epidemiological, showed promising benefits of many dietary nutrients in melanoma chemoprevention [4].
Nutramil™ Complex is a Food for Special Medical Purpose (FSMP) composed of all essential nutrients - proteins, carbohydrates, fats, vitamins, micro- and macroelements. It is designed to provide a nutritionally complete and balanced diet for people whose nutritional requirements cannot be met by normal foods. In Europe, the FSMP product composition and placing on the market is regulated by the REGULATION (EU) No 609/2013, supplemented by the COMMISSION DELEGATED REGULATION (EU) 2016/128. Article 9 of the supplement makes each Member State solely responsible for the enforcement of the FSMP legislation. FSMP legislation itself does not impose a registration procedure on the food business operator, as required from medicinal products, but relies on the self – assessment that the product complies with regulations. Therefore, FSMP products are not subjected to preclinical and clinical studies. As a result, available literature is very limited on FSMP. A rare example is our earlier work reporting the effect of Nutramil™ Complex (NC) on breast and prostate cancer cells; specifically, the induction of mitochondrial apoptotic pathway [5]. In the current manuscript we studied the impact of NC on two lines of melanoma cells. Interestingly, our results showed two different responses of these cells to the applied product.
2. Materials and Methods
2.1. Testing Material
NutramilTM Complex (NC) was the testing material and its composition is described in our previously publication [5]. To determine the effect of complete compound on cancer cells, an incomplete NutramilTM Complex without calcium caseinate (NC-CC) was also investigated. We hypothesized that the complete formulation would have the greatest effect on cellular parameters. On the other hand, we would like to check if the protein exclusion would maintain the observed changes, which may be useful in a possible modification of the nutrient composition. All analyses were blanked. All samples were numerically coded and decoded at the end of the study.
2.2. Cell Cultures and Treatments
This research was conducted with use of human primary melanoma cell line WM115 (ATCC® CRL-1675TM), human malignant melanoma cell line WM266-4 (ATCC® CRL-1676TM) and human foreskin fibroblast cell line BJ (ATCC® CRL-2522TM). Cells were cultured in appropriate medium with the addition of 10% FBS (Sigma-Aldrich, MO, USA) and under controlled conditions (temperature 37 °C and 5% CO2 atmosphere) according to the ATCC protocol.
Cancer cells were seeded in a density of 8x103 cells per well in 96-well plates, 9x104 cells per well in 12-well plates and 2x105 per well in 6-well plates. 24h after seeding, growth medium was replaced with a medium containing encoded treatments of NC or NC-CC at various concentrations. The final applied concentrations of NC and NC-CC were 1, 2, 3, 4, 5 and 10%. Then, the cells were incubated for 24, 48 and 72h. Cells cultivated only in complete growth medium were used as a negative control. Staurosporine (Sigma-Aldrich, MO, USA) at final concentration at 1.5 μM was used as positive control for apoptosis assay.
2.3. Cytotoxicity Assay
Cell cytotoxicity was analyzed by Cytotoxicity Detection Kit LDH (Roche, BASEL, Switzerland) which is based on measuring the activity of lactate dehydrogenase (LDH) released into the culture medium during cell death. The assay was carried out according to the manufacturer’s protocol. The experiments were performed in three independent tests and 4-5 technical replicates.
2.4. Cell Proliferation Assay
Cell proliferation was determined with 5’-bromo-2’-deoxy-uridine (BrdU) Labeling and Detection Kit III (Roche, BASEL, Switzerland), according to manufacturer's instructions. The method is based on the ability of BrdU to be incorporated instead of thymidine into the DNA of proliferating cells. All experiments were performed in three independent tests and measured in triplicates. Results were standardized to the negative control (non-treated cells) as 100%.
2.5. RNA Isolation, RT Reaction and Real-Time PCR Analysis
The total RNA was isolated from cell lines using the Total RNA Mini Kit (A&A Biotechnology, Gdansk, Poland), in accordance with the manufacturer's instructions. Concentration, purity and quality of RNA was measured in µDrop Plate (Thermo Fisher Scientific, Waltham, MA, USA). Reverse transcription reaction was performed using the Maxima first Strand cDNA Synthesis kit for RT-qPCR (Thermo Fisher Scientific, Waltham, MA, USA). A quantitative analysis of mRNA gene expression was performed using the panels of TaqMan® Array Human C-MYC and Apoptosis (Thermo Fisher Scientific, Waltham, MA, USA) according to manufacturer’s instructions and carried out in StepOnePlusTM System (Thermo Fisher Scientific, Waltham, MA, USA). Analysed genes included: AKT1, APAF1, BAD, BAX, BCL2, BID, CASP3, CASP8, CDKN2A, CYCS, FADD, FAS, FASLG, HRAS, IGF1, IGF1R, KRAS, MYC, NRAS, RRAS, TP53, YWHAB, YWHAE, YWHAG, YWHAH, YWHAQ, YWHAZ. Results were normalized using at least two reference genes (18S, GAPDH, HPRT1 or GUSB) and were calculated using the 2-∆∆CT method [6].
2.6. Stress and Apoptosis Signaling Assay
Cell extracts were prepared and analyzed using the PathScan® Stress and Apoptosis Signaling Antibody Array Kit (Chemiluminescent Readout #12856, Cell Signaling Technology, MA, USA). Assay target proteins were: P44/42 MAPK (ERK1/2) phosphorylation, Akt phosphorylation, Bad phosphorylation, HSP27 phosphorylation, Smad2 phosphorylation, p53 phosphorylation, p38 MAPK phosphorylation, SAPK/JNK phosphorylation, PARP cleavage, Caspase-3 cleavage, Caspase-7 cleavage, Ikβα total, Chk1 Ser345 phosphorylation, Chk2 phosphorylation, Ikβα phosphorylation, eIF2α phosphorylation, TAK1 phosphorylation, Survivin and α-Tubulin as a reference protein. Images were acquired by briefly exposing the slide to standard chemiluminescent film. Densitometry analysis was performed using ImageJ (http://imagej.nih.gov/ij/). Results are shown as a mean±SD normalized to the internal reference protein (α_Tubulin). Untreated negative control (UC) was set as 100% expression level.
2.7. Statistical Analysis
The statistical analysis was performed by using Statistica ver.12 (StatSoft, Tulsa, OK, USA). All experiments were performed in three independent tests and measured in triplicates. Shapiro-Wilk’s test was applied to assess normality of distribution. Results are expressed as mean ± standard deviation (SD). Statistical analysis of all experiments in melanoma and BJ cells was conducted using an independent samples t-test to compare unpaired means between two groups. p values less than 0.05 were considered as statistically significant.
3. Results
3.1. Cytotoxicity
Nutramil™ Complex showed strong cytotoxic effects on both examined melanoma cell lines (WM266-4 and WM115), as well as non-neoplastic BJ cells (Figure 1).
Cytotoxicity was dose- and time-dependent and showed similar trend for both NC-CC (NutramilTM Complex without calcium caseinate) and NC treatment. Application of Cytotoxicity Detection Kit LDH (Roche, Poland) showed that 4% treatment did not initiate necrosis (Figure 1 A-C). Based on the results, 4% concentration was chosen for further studies as the inhibitory concentration for 10% cytotoxicity (EC10).
3.2. Cell Proliferation
The BrdU test demonstrated that Nutramil™ Complex reduced the proliferation of WM266-4 cell line to approximately 75-80% of the negative control (Figure 2B) but did not affect WM115 melanoma cells (Figure 2A). In contrast, NC stimulated proliferation of normal BJ cell line (Figure 2C). Treatment with NC-CC cells showed similar effects as NC on studied cell lines (Figure 2A-C).
3.3. mRNA Expression of Genes Associated with Cell Cycle and Apoptosis
The expression of genes regulating cell cycle and apoptosis was investigated with TaqMan® Array C-MYC and Apoptosis Kit. Results are presented in Table 1.
For WM-115 primary melanoma cells, results showed up-regulation of many pro-apoptotic genes, including: APAF1, BAD, BAX, BID, CASP3, CASP8, CASP9, CYCS, FADD and down-regulation of pro-survival genes: BCL-2, HRAS, IGF1, KRAS, MYC and YWHA family.
For WM266-4 melanoma cells, results showed up-regulation of pro-apoptotic BAD, BAX, BID, CASP3, CASP9, FADD, FAS and TP53, and down regulation of APAF1 and CYCS. Pro-survival genes, including: AKT1, BCL-2, HRAS, IGF1, IGF1R, KRAS, NRAS, MYC and genes from the YWHA family, showed a reduction in their mRNA levels after NC treatment.
Results for treatment with NC-CC showed similar trends (Table 1).
3.4. Expression of Proteins Involved in Cellular Stress and Apoptosis Signalling
Results of the PathScan® Stress and Apoptosis Signalling Antibody Array analysis for WM115 and WM266-4 are presented in Figure 3A and Figure 3B, respectively.
Treatment of WM115 with NC did not significantly reduce levels of pro-survival proteins, including ERK1/2, AKT-1, HSP27 and Survivin; however, it significantly increased levels of phospho-BAD (128% of UC) as well as cleaved form of Caspase -7 (200% of UC), Smad2 tumor suppressor (211% of UC), P38MAPK (220% of UC) and SAPK/JNK (157% of UC). The level of p53, Caspase-3 and PARP cleavage were not significantly affected. Expression of Chk-1 and Chk-2, total and phosphorylated Iκβα, eIF-2α as well as TAK1 were significantly increased to (148%, 330%, 161%, 132%, 447%, 493% of UC, respectively) (Figure 3A).
Treatment of WM266-4 with NC significantly affected pro-survival proteins, including: AKT-1 (57% of UC), Survivin (64% of UC) and TAK1 (69% of UC). Levels of ERK1/2 and HSP27 showed an insignificant reduction. Among the pro-apoptotic proteins, results showed an increase in levels of Bad, Smad2, p38MAPK, cleaved forms of Caspase-3 and -7, as well as cleaved PARP (144%, 160%, 134%, 237%, 111%, 148% of UC, respectively). In addition, Chk-1 levels were significantly reduced to 70% of UC, while Chk-2 remained unchanged. Expression of p53 suppressor was reduced (76% of UC). The SAPK/JNK, Iκβα-phospho and total Iκβα showed a decrease while the eIF-2α protein level was elevated to 126% of UC (Figure 3B).
For NC-CC, results for the majority of tested proteins were consistent with the results observed for NC treatment (Figure 3A-B).
4. Discussion
Available epidemiological research indicates the importance of diet in reducing the risk and progression of melanoma. Anti-cancer properties are primarily expected from products with a strong antioxidant and anti-inflammatory potential, associated with the ability to reverse the oxidative damage caused by UV light [7]. On the other hand, main therapeutic strategies are focused on induction of apoptosis and suppression of survival pathways [8].
Our data showed that NC reduced proliferation of malignant melanoma WM266-4 cells but did not affected the proliferation of WM115 primary melanoma (Figure 2). Initially, both melanoma lines showed a measurable downregulation of most of the analyzed pro-survival genes expression at the mRNA level (Table 1). However, these results were not confirmed for WM115 cells at the protein expression level (Figure 3A), which is consistent with the observed lack of changes in WM115 cell proliferation (Figure 2A). It should be emphasized that changes at the transcription level are not always reflected in protein expression [9].
Measured protein expression in WM266-4 cells confirmed downregulation of pro-survival ERK1/2, AKT-1, HSP27, Survivin and TAK1 (Figure 3) and upregulation of pro-apoptotic Bad, Smad2, p38MAPK, cleaved forms of Caspase-3 and -7, as well as cleaved PARP (Figure 3B). Interestingly, similarly to WM266-4 cells, WM-115 cells showed a significant increase in levels of some pro-apoptotic proteins, including phospho-BAD, cleaved form of Caspase -7, P38MAPK and SAPK/JNK as well as Smad2 tumor suppressor (Figure 3A). The sensitivity of various melanoma cancer cell lines to the same compound may differ, depending on the distinctive mechanism of formation and progression. The speed of cell division and their translational activity are mentioned as the primary determinants [10]. The resistance of tumor may also result from the decreased ability to undergo spontaneous apoptosis. Available research indicates that phenotype changes occurring during cancer development may play a crucial role in developing resistance to applied pro-apoptotic factors [11]. Our proliferation data would confirm that hypothesis as both NC-treated melanoma cell lines showed similar trends in pro-apoptotic protein expression, but they manifested a different response. In addition, our results also highlight the complexity of mechanisms and interactions that must be triggered to regulate growth and death of cancer cells.
Literature suggests that in human melanomas two major signaling pathways RAS/RAF/MEK/ERK and the PI3K/Akt are constitutively activated through genetic mutations. It also suggests the influence of regulatory proteins of the Bcl-2 family [12,13].
The RAS/RAF/MEK/ERK signaling pathway is an important regulator of cell growth and survival and it has been reported to be activated in about 90% of human melanomas [14]. Of all RAS isoforms (HRAS, KRAS, and NRAS in humans), the most common mutation in melanomas occurs in NRAS gene, while in other types of human cancers mutations are more frequent in the KRAS gene [15]. Some melanomas exhibit an excessive activation of heat shock proteins, such as HSP27. HSP27 increase the resistance of melanoma cancer cells to apoptosis by binding cytochrome c and preventing the activation of caspase-9 and caspase-3 [16]. HSP27 also accelerates the proteolysis of p27 (CDK inhibitor), which prevents cell cycle arrest in the G1 phase [17]. HSP27 is also associated with the promotion of the proteasomal degradation of Iκβα – the inhibitor of the NF-κβ transcription factor. On the other hand, the presence of phosphorylated form of Iκβα indicates degradation of the NF-κβ / Iκβ complex and subsequent translocation of NF-κβ to the nucleus, where it can activate many anti-apoptotic genes [5]. Treatment of WM-266-4 cells with the NutramilTM Complex had no significant effect on the level of the phosphorylated form of Iκβα. It also did not increase the level of the non-phosphorylated Iκβα, capable of binding to NF-κβ (Figure 3B). However, our results showed a reduced level of TAK1 protein, which may stimulate the phosphorylation of the Iκβα protein [18].
PI3K/Akt signaling as was found dysregulated in over 50% of melanomas [19]. PI3K/Akt signaling cascade is activated via the IGF growth factors paracrine/autocrine signal. Active RAS induces membrane translocation and activation of PI3K leading to PIP2 phosphorylation to PIP3 and activation of Akt protein. Akt effectors promote cell survival, proliferation and invasion [20], including activation of NF-κB transcription factor (15), promoting expression of pro-survival proteins from Bcl-2 family and inhibiting expression of pro-apoptotic Bad [21,22]. NutramilTM Complex had a downregulating effect on the expression of AKT1 gene and its protein in WM266-4 cells, while it had no significant effect in WM-115 cells (Table 1; Figure 3). However, down-regulation of BCL-2 gene expression and increased expression of pro-apoptotic genes, including BAD, BID and BAX were observed in both cell lines (Table 1). In addition, an increased level of Bad protein was also measured in both cell lines (Figure 3). Our results also showed an elevated level of phosphorylated kinases MAP p38 MAPK and SAPK/JNK MAP in both melanoma cell lines after NC treatment (Figure 3). The literature indicates that activation of p38 MAPK and SAPK/JNK MAP kinases occurs via dual phosphorylation mechanism in response to cellular stressors and leads to the cell-cycle arrest and apoptosis induction. In addition, it is suggested that SAPK/JNK may contribute to the activation of both apoptotic pathways, the extrinsic and the mitochondrial-dependent pathway. In order to induce cell death, these mechanisms may interact or act independently [5], which finds confirmation in measured upregulated expression of FADD, FAS and CASP8 genes (Table 1).
In contrast to many other cancers, the p53 suppressor gene is rarely mutated in melanoma [19] but its functional attenuation is needed for tumor development [23]. Our results showed an increased level of TP53 gene mRNA in both NC-treated melanoma cell lines (Table 1); however, WM266-4 also showed a reduced level of p53 protein (Figure 3). This indicates potentially p53-independent induction of apoptosis in this cell line. These results are consistent with other studies on human melanoma cells that showed potential degradation of p53 protein rather than its accumulation (11). In addition, recently, the essential p53 inhibitor Mdmx was determined frequently over-expressed in melanoma. Mdmx displays both p53-dependent and p53-independent oncogenic effects needed for melanoma growth [23].
APAF-1 gene dysregulation is often indicated (42%) as one of the factors inhibiting the apoptotic process in melanoma cells; namely, by directly preventing caspase-9 activation and initiation of protease cascade [24,25]. Our results showed that treatment of both melanoma lines with NutramilTM Complex increased the expression of APAF-1 as well as CASP 9 and CASP 3 (Table 1). However, simultaneous increase in the protein levels of apoptotic markers, including the cleaved form of caspase-3 and its substrate PARP, was observed only in WM266-4 cells (Figure 3). Similarly, only WM266-4 cell line showed a reduction in the level of Survivin (Figure 3), a protein highly expressed in most cancer cells. Survivin inhibits the caspase activation in tumor cells, thereby suppressing their apoptosis and at promoting cell proliferation [26,27,28]. It has been shown that by inhibiting Survivin expression via various mechanism, including blocking its transcription, increased the sensitivity of various cancer cells, including melanoma, to some chemotherapeutic agents [29,30]. For this reason, Survivin is considered an independent marker associated with poor prognosis [11,31]. Taken together, measured gene and protein expression supports presented proliferation results in the examined cell lines (Figure 2). In the WM115 cells, despite the observed activation of some pro-apoptotic genes, NC did not cause the death of these cells. On the other hand, the reduction of WM266-4 cell line proliferation was associated both with the downregulation of pro-survival genes and induction of apoptosis. Available literature indicates that dysregulation of apoptotic processes in melanoma cells are primarily associated with impaired mitochondrial-dependent pathway. Although our results do not include the full set of proteins involved in this process, measured changes expression of BCL-2 genes, upregulation of APAF-1 and reduced Survivin expression (Table 1, Figure 3) support this hypothesis. In addition, in our previous study showed a similar effect of this FSMP on the induction of the intrinsic pathway of apoptosis in breast and prostate carcinoma cells [5]. Taken together, NutramilTM Complex, as a food for special medical purposes, may support the nutritional treatment of oncological patients. In addition, in the current and previous study, we presented its cytotoxic effect against cancer cells. This effect on reducing the growth of cancer cells, described in detail in our earlier publication, may be associated with the product composition, including the contribution and chemical form of the particular macro and microelements [5].
5. Conclusions
In conclusion, it should be emphasized that the results presented in current paper, are one of the very few investigating the effect of the selected FSMP product, NutramilTM Complex, on the proliferation of melanoma cells. Results obtained from WM266-4 line cells are consistent with our previous findings in breast and prostate cancer lines, which confirms beneficial effect of the product. On the other hand, the observed differences in response to the NC treatment between two studied melanoma line cells may result from their different sensitivity. Although our experiments are limited to the in vitro model and require verification in vivo, we find those tests necessary according to the 3R principle (Replacement, Reduction and Refinement). Together, they serve as a valuable and multifaceted source of information and provide justification for further research.
Author Contributions
Conceptualization, A.K. and T.L; formal analysis, A.K.; funding acquisition, A.K. and T.L.; investigation, M.D., E.PS., A.K; methodology, A.K.; project administration, A.K; Visualization, K.K., and A.S; supervision, A.K.; writing – original draft, A.K., K.K.; AS; writing – review & editing, T.L., K.K., A.K.
Funding
This work was supported by the Polish Ministry of Science and Higher Education under Grant DS 3710/ KŻCz/2018.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomatarm I, Jemal A, Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Indicence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2022;71:209-249. [CrossRef]
- Globocan 2022 Global Cancer Observatory. Available online: https://gco.iarc.who.int/media/globocan/factsheets/cancers/16-melanoma-of-skin-fact-sheet.pdf (accessed on 12 August 2024).
- Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer statistics. CA Cancer J Clin 2022;72:7-33. [CrossRef]
- Tong LX, Young LC. Nutrition: the future of melanoma prevention? J Am Aca. Dermatol. 2014;71:151-160. [CrossRef]
- Koronowicz AA, Drozdowska M, Wielgos B, Piasna-Słupecka E, Domagała D, Dulińska-Litewka J, Leszczyńska T. The effect of “NutramilTM Complex”, food for special medical purpose, on breast and prostate carcinoma cells. PLOS ONE 2018;13: e0192860. [CrossRef]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001;25:402-408. [CrossRef]
- Yang K, Fung TT, Nan H. An Epidemiological Review of Diet and Cutaneous Malignant Melanoma. Cancer Epidemiol Biomarkers Prev. 2018;27:1115-1122. [CrossRef]
- Eberle J, Kurbanov BM, Hossini AM, Trefzer U, Fecker LF. Overcoming apoptosis deficiency of melanoma-hope for new therapeutic approaches. Drug Resist Updat. 2007;10: 218-234. [CrossRef]
- Master A, Nauman A. Molecular mechanisms of protein biosynthesis initiation-biochemical and biomedical implications of a new model of translation enhanced by the RNA hypoxia response element (rHRE). Postepy Biochem. 2014;60:39-54.
- McConkey DJ, Zhu K. Mechanisms of proteasome inhibitor action and resistance in cancer. Drug Resist Updat. 2008;11:164-179. [CrossRef]
- Cichorek M, Kozłowska K, Wachulska M, Zielińska K. Spontaneous apoptosis of melanotic and amelanotic melanoma cells in different phases of cell cycle: relation to tumor growth. Folia Histochem Cytobiol. 2006;44: 31-36.
- Pokrywka M, Litynska A. Targeting the melanoma. Postepy Biol. Komorki 2012;39:3-24.
- Blokx WA, van Dijk MC, Ruiter DJ. Molecular cytogenetics of cutaneous melanocytic lesions - diagnostic, prognostic and therapeutic aspects. Histopathology. 2010;56:121-132. [CrossRef]
- Gray-Schopfer V, Wellbrock C, Marais R. Melanoma biology and new targeted therapy. Nature 2007;445:851-857. [CrossRef]
- Sullivan RJ, Atkins MB. Molecular-targeted therapy in malignant melanoma. Expert Rev Anticancer Ther. 2009;9:567-581. [CrossRef]
- Sidor-Kaczmarek J, Cichorek M, Spodnik JH, Wójcik S, Moryś J. Proteasome inhibitors against amelanotic melanoma. Cell Biol Toxicol. 2017;33:557-573. [CrossRef]
- Parcellier A, Brunet M, Schmitt E, Col E, Didelot C, Hammann A, Nakayama K, Nakayama KI, Khochbin S, Solary E, Garrido C. HSP27 favors ubiquitination and proteasomal degradation of p27Kip1 and helps S-phase re-entry in stressed cells. FASEB J. 2006;20:1179-1181. [CrossRef]
- Okada M, Matsuzawa A, Yoshimura A, Ichijo H. The lysosome rupture-activated TAK1-JNK pathway regulates NLRP3 inflammasome activation. J. Biol Chem. 2014; 289:32926-32936. [CrossRef]
- Dahl C, Guldberg P. The genome and epigenome of malignant melanoma. APMIS 2007;115:1161-1176. [CrossRef]
- Yajima I, Kumasaka MY, Thang ND, Goto Y, Takeda K, Yamanoshita O, Iida M, Ohgami N, Tamura H, Kawamoto Y, Kato M. RAS/RAF/MEK/ERK and PI3K/PTEN/AKT Signaling in Malignant Melanoma Progression and Therapy. Dermatol Res Pract 2012;2012. [CrossRef]
- Krześlak A. Akt kinase: a key regulator of metabolism and progression of tumors. Postepy Hig Med Dosw (Online) 2010;64:490-503.
- Koronowicz AA, Banks P, Domagała D, Master A, Leszczyńska T, Piasna E, Marynowska M, Laidler P. Fatty acid extract from CLA-enriched egg yolks can mediate transcriptome reprogramming of MCF-7 cancer cells to prevent their growth and proliferation. Genes Nutr 2016;11:1-12. [CrossRef]
- Jochemsen AG. Reactivation of p53 as therapeutic intervention for malignant melanoma. Curr Opin Oncol. 2014;26:114-119. [CrossRef]
- Kyrgidis A, Tzellos TG, Triaridis, S. Melanoma: Stem cells, sun exposure and hallmarks for carcinogenesis, molecular concepts and future clinical implications. J Carcinog. 2010;9:3. [CrossRef]
- Ko JM, Velez NF, Tsao H. Pathways to melanoma. Semin Cutan Med Surg. 2010;29: 210-217. [CrossRef]
- Wheatley SP, McNeish IA. Survivin: a protein with dual roles in mitosis and apoptosis. Int Rev Cytol. 2005;247:35-88. [CrossRef]
- O’Driscoll L, Linehan R, Clynes M. Survivin: role in normal cells and in pathological conditions. Curr Cancer Drug Targets. 2003;3:131-152. [CrossRef]
- Tamm I, Wang Y, Sausville E, Scudiero DA, Vigna N, Oltersdorf T, Reed JC. IAP-family protein survivin inhibits caspase activity and apoptosis induced by Fas (CD95), Bax, caspases, and anticancer drugs. Cancer Res. 1998;58:5315-5320.
- Ikeguchi M, Hirooka Y, Kaibara N. Quantitative analysis of apoptosis-related gene expression in hepatocellular carcinoma. Cancer. 2002;95:1938-1945. [CrossRef]
- Grossman D, Altieri DC. Drug resistance in melanoma: mechanisms, apoptosis, and new potential therapeutic targets. Cancer Metastasis Rev. 2001;20:3-11. [CrossRef]
- Gradilone A, Gazzaniga P, Ribuffo D, Scarpa S, Cigna E, Vasaturo F, Bottoni U, Innocenzi D, Calvieri S, Scuderi N, Frati L, Aglianò AM. Survivin, bcl-2, bax, and bcl-X gene expression in sentinel lymph nodes from melanoma patients. J Clin Oncol. 2003;21:306-312. [CrossRef]
Figure 1.
Cytotoxycity of NutramilTM Complex and NutramilTM Complex without calcium caseinate in human melanoma cell lines: A) WM-115, B) WM-266-4 and C) BJ normal fibroblast cell line. Cells were treated for 24, 48 and 72h with 1-10% concentration of NutramilTM Complex (NC) or NutramilTM Complex without calcium caseinate (NC-CC ). Cytotoxicity was measured with Cytotoxicity Detection Kit LDH (Roche, Poland). Values are expressed as mean ± SD for n=15, standardized to untreated control (UC) as 100%.
Figure 1.
Cytotoxycity of NutramilTM Complex and NutramilTM Complex without calcium caseinate in human melanoma cell lines: A) WM-115, B) WM-266-4 and C) BJ normal fibroblast cell line. Cells were treated for 24, 48 and 72h with 1-10% concentration of NutramilTM Complex (NC) or NutramilTM Complex without calcium caseinate (NC-CC ). Cytotoxicity was measured with Cytotoxicity Detection Kit LDH (Roche, Poland). Values are expressed as mean ± SD for n=15, standardized to untreated control (UC) as 100%.
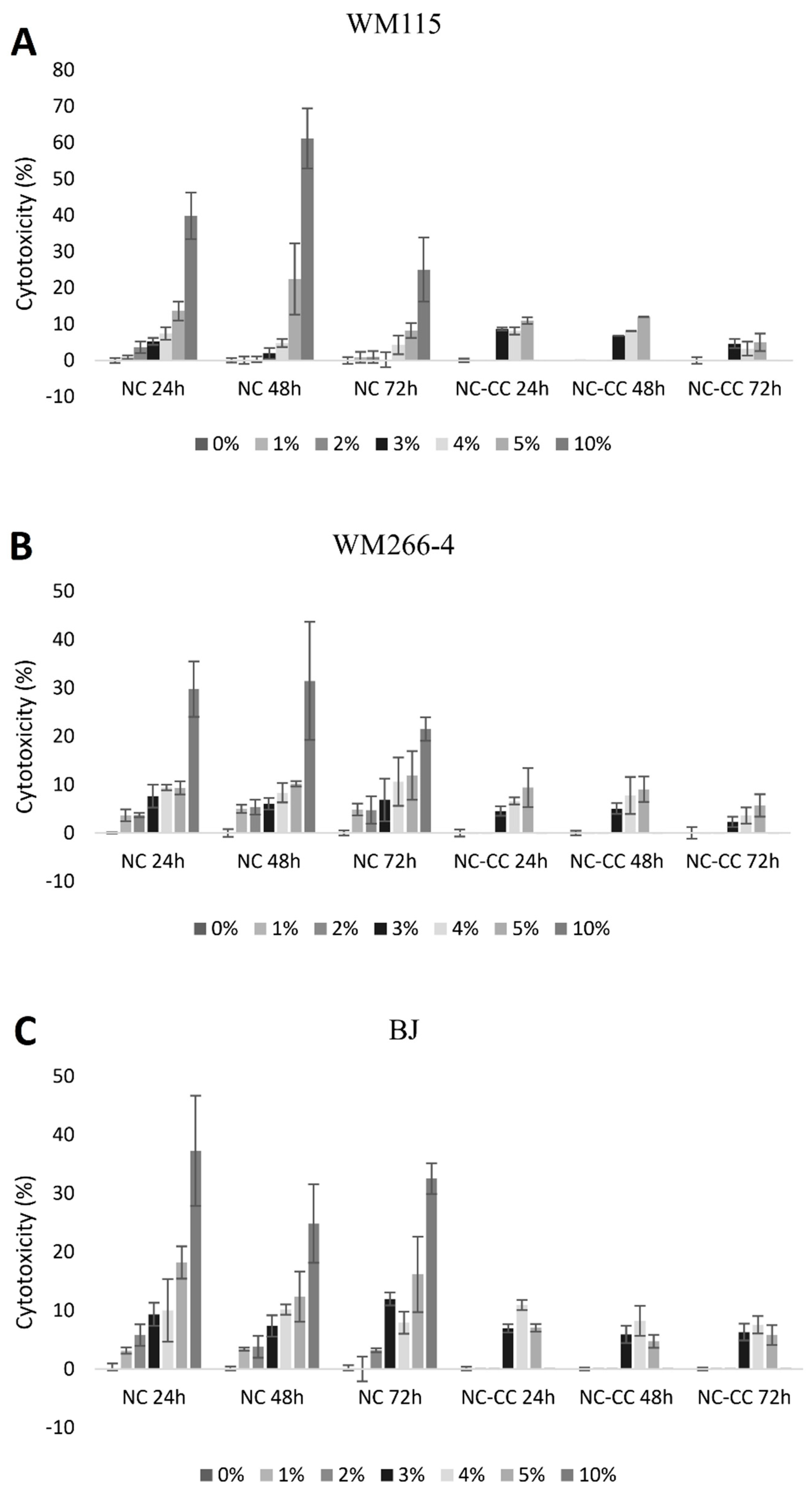
Figure 2.
The effect of NutramilTM Complex and NutramilTM Complex without calcium caseinate on proliferation in human melanoma cell lines (WM-115, WM-266-4) and normal fibroblast cell line (BJ). Cells WM-115 (A); WM266-4 (B) and BJ (C) were treated with NutramilTM Complex (NC) or NutramilTM Complex without calcium caseinate (NC-CC ) at concentration 0, 3, 4% for 24, 48 and 72h. Cell proliferation was measured using Cell Proliferation ELISA, BrdU (Roche, Poland). Values are expressed as mean ± SD for n=12, standardized to untreated control (UC) as 100%. Statistical significance was based on T-test; * vs. UC when p < 0.05.
Figure 2.
The effect of NutramilTM Complex and NutramilTM Complex without calcium caseinate on proliferation in human melanoma cell lines (WM-115, WM-266-4) and normal fibroblast cell line (BJ). Cells WM-115 (A); WM266-4 (B) and BJ (C) were treated with NutramilTM Complex (NC) or NutramilTM Complex without calcium caseinate (NC-CC ) at concentration 0, 3, 4% for 24, 48 and 72h. Cell proliferation was measured using Cell Proliferation ELISA, BrdU (Roche, Poland). Values are expressed as mean ± SD for n=12, standardized to untreated control (UC) as 100%. Statistical significance was based on T-test; * vs. UC when p < 0.05.
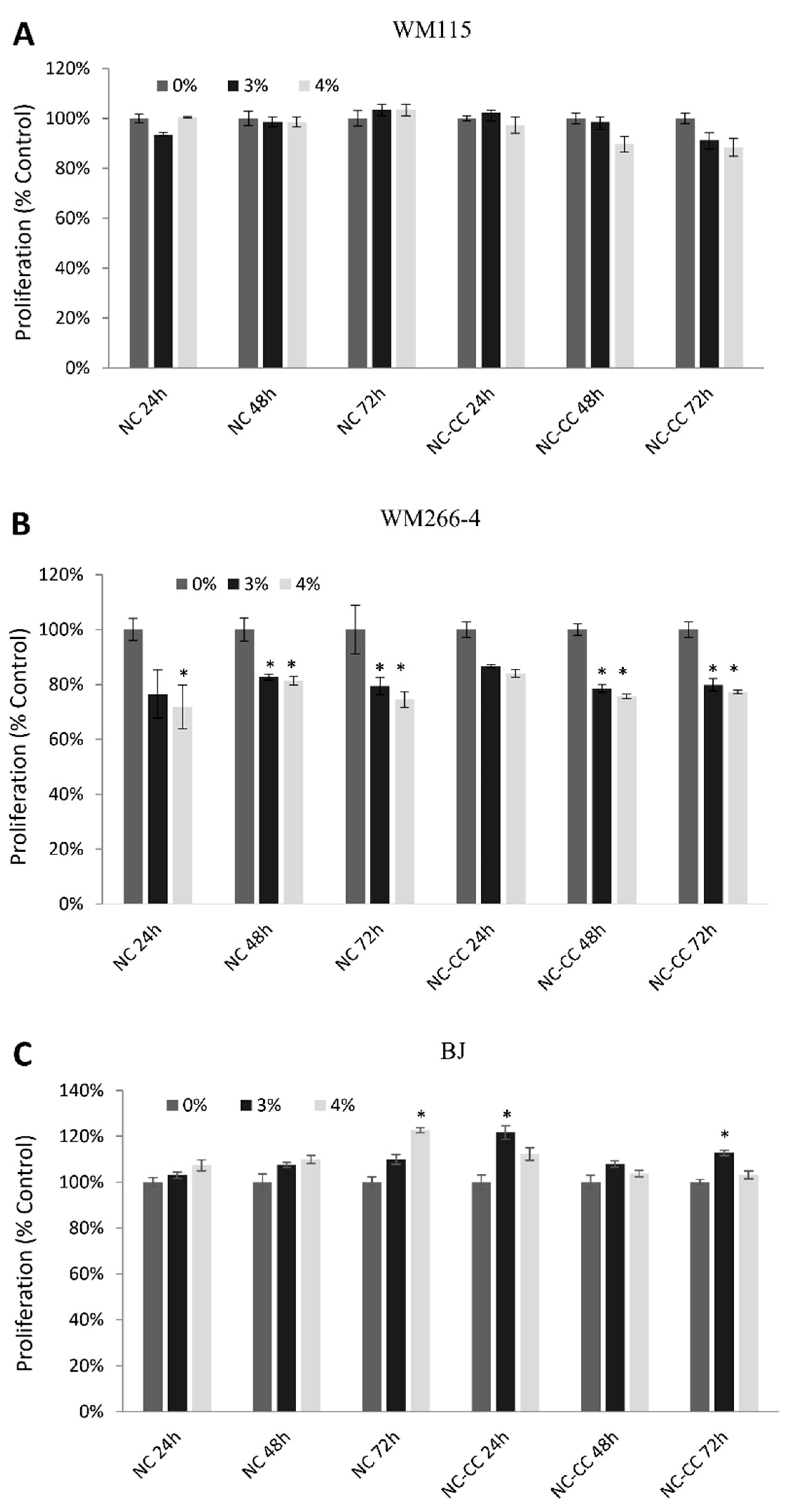
Figure 3.
The effect of NutramilTM Complex and NutramilTM Complex without calcium caseinate on expression of stress and apoptosis proteins in melanoma cells (WM-115, WM-266-4).Cells WM-115 (A) and WM266-4 (B) were treated for 48h with 4% of Nutramil TM Complex (NC) or Nutramil TM Complex without calcium caseinate (NC-CC). Staurosporine (ST; 1,5 μM concentration) was used as positive control. Cell extracts were analyzed using the PathScan® Stress and Apoptosis Signaling Antibody Array Kit (Chemiluminescent Readout) #12856, Cell Signaling Technology, MA, USA. Images were acquired by briefly exposing the slide to standard chemiluminescent film. Densitometry analysis was performer using ImageJ. Results are shown as a mean±SD normalized to the internal reference protein (α_Tubulin). Untreated negative control (UC) was set as 100% expression level. Statistical significance was based on T-test; * vs. UC when p < 0.05. Gene symbols and names: P44/42 MAPK (ERK1/2) phosphorylation (Thr202/Tyr204), Akt-1 phosphorylation (Ser473), Bad phosphorylation (Ser136), HSP27 phosphorylation (Ser82), Smad2 phosphorylation (Ser465/467), p53 phosphorylation (Ser15), p38 MAPK phosphorylation (Thr180/Tyr182), SAPK/JNK phosphorylation (Thr183/Tyr185), PARP cleavage (Asp214), Caspase-3 cleavage (Asp175), Caspase-7 cleavage (Asp198), IkB total, Chk-1 phosphorylation (Ser345), Chk-2 phosphorylation (Thr68), IkBα phosphorylation (Ser32/36), eIF2a phosphorylation (Ser51), TAK1 phosphorylation (Ser412), Survivin total.
Figure 3.
The effect of NutramilTM Complex and NutramilTM Complex without calcium caseinate on expression of stress and apoptosis proteins in melanoma cells (WM-115, WM-266-4).Cells WM-115 (A) and WM266-4 (B) were treated for 48h with 4% of Nutramil TM Complex (NC) or Nutramil TM Complex without calcium caseinate (NC-CC). Staurosporine (ST; 1,5 μM concentration) was used as positive control. Cell extracts were analyzed using the PathScan® Stress and Apoptosis Signaling Antibody Array Kit (Chemiluminescent Readout) #12856, Cell Signaling Technology, MA, USA. Images were acquired by briefly exposing the slide to standard chemiluminescent film. Densitometry analysis was performer using ImageJ. Results are shown as a mean±SD normalized to the internal reference protein (α_Tubulin). Untreated negative control (UC) was set as 100% expression level. Statistical significance was based on T-test; * vs. UC when p < 0.05. Gene symbols and names: P44/42 MAPK (ERK1/2) phosphorylation (Thr202/Tyr204), Akt-1 phosphorylation (Ser473), Bad phosphorylation (Ser136), HSP27 phosphorylation (Ser82), Smad2 phosphorylation (Ser465/467), p53 phosphorylation (Ser15), p38 MAPK phosphorylation (Thr180/Tyr182), SAPK/JNK phosphorylation (Thr183/Tyr185), PARP cleavage (Asp214), Caspase-3 cleavage (Asp175), Caspase-7 cleavage (Asp198), IkB total, Chk-1 phosphorylation (Ser345), Chk-2 phosphorylation (Thr68), IkBα phosphorylation (Ser32/36), eIF2a phosphorylation (Ser51), TAK1 phosphorylation (Ser412), Survivin total.
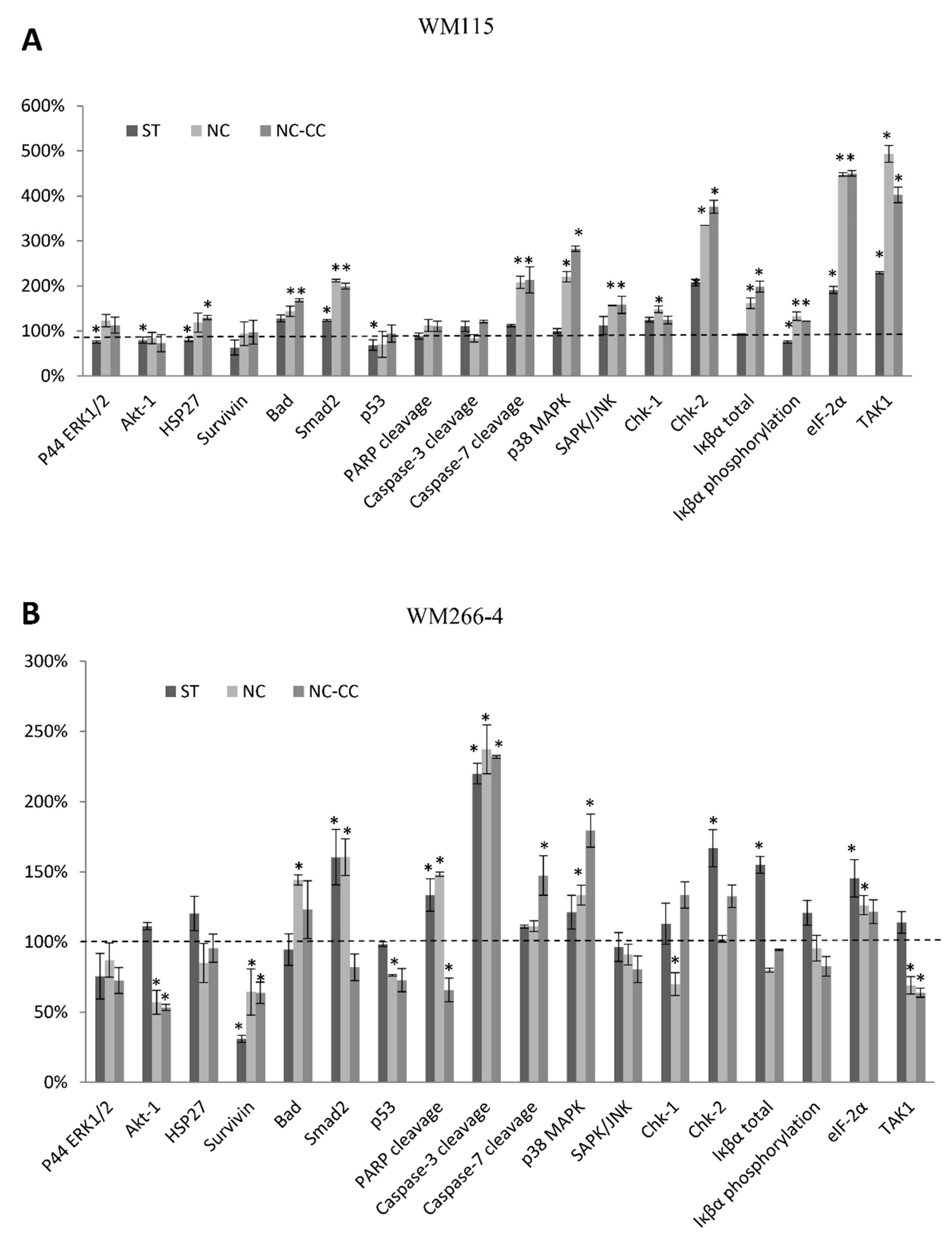

Table 1.
The effect of NutramilTM Complex and NutramilTM Complex without calcium caseinate on mRNA expression of genes in human melanoma WM-115 and WM-266-4 cells lines.
Table 1.
The effect of NutramilTM Complex and NutramilTM Complex without calcium caseinate on mRNA expression of genes in human melanoma WM-115 and WM-266-4 cells lines.
| Gene Symbol |
WM-115 | WM266-4 | ||||||
|---|---|---|---|---|---|---|---|---|
| NC vs UC | NC-CC vs UC | NC vs UC | NC-CC vs UC | |||||
| FC value | Adjusted p-values |
FC value | Adjusted p-values |
FC value | Adjusted p-values |
FC value | Adjusted p-values |
|
| Pro-apoptotic genes | ||||||||
| APAF1 | ↑4,52* | 1,1E-07 | ↑4,73* | 0,00003 | ↓-4,56* | 0,00001 | ↓-2,69* | 0,00010 |
| BAD | ↑1,59* | 0,01212 | ↑1,25* | 0,00007 | ↑6,20* | 0,01623 | ↑3,62* | 0,00128 |
| BAX | ↑1,37* | 0,00064 | 1,44 | 0,07656 | ↑2,12* | 0,00005 | 1,29 | 0,05086 |
| BID | ↑1,78* | 0,02290 | ↑1,25* | 0,00552 | ↑2,58* | 0,00013 | 1,74 | 0,12706 |
| CASP3 | ↑2,42* | 0,00017 | ↑2,79* | 5,0E-07 | ↑4,00* | 0,00017 | ↑3,27* | 0,00005 |
| CASP8 | ↑4,01* | 0,00015 | ↑5,53* | 0,00007 | ↓-1,55* | 0,00001 | ↓-2,28* | 0,00013 |
| CASP9 | ↑2,91* | 0,00148 | ↑1,40* | 0,00752 | ↑4,80* | 0,00148 | ↑2,93* | 0,00007 |
| CYCS | ↑1,73* | 0,03390 | ↑1,48* | 0,00641 | ↓-2,06* | 0,00295 | ↓-2,76* | 0,00006 |
| FADD | 1,20 | 0,08572 | ↑1,47* | 0,01262 | ↑3,16* | 0,00008 | 1,87 | 0,163556 |
| FAS | 1,01 | 0,3740 | ↑1,12* | 2,80E-05 | 1,12 | 0,37390 | ↑1,34* | 0,00006 |
| TP53 | 1,02 | 0,09595 | ↑1,49* | 0,02467 | ↑1,72* | 0,00001 | 1,28 | 0,06596 |
| Pro-survival genes | ||||||||
| AKT1 | 1,05 | 0,28798 | 1,18 | 0,12187 | ↓-1,97* | 0,00014 | ↓-1,42* | 0,02336 |
| BCL2 | -1,36 | 0,19346 | ↓-1,37* | 0,00859 | ↓-1,57* | 0,00022 | ↓-2,53* | 0,00023 |
| HRAS | ↓-1,82* | 0,02360 | -1,69 | 0,51894 | ↓-2,38* | 0,00033 | ↓-1,64* | 0,00358 |
| IGF1 | ↓-3,04* | 0,04336 | 1,01 | 0,11020 | ↓-7,70* | 0,00003 | -2,16 | 0,08021 |
| IGF1R | ↑1,56* | 0,00003 | ↑1,27* | 0,00040 | -2,01 | 0,43357 | ↓-1,43* | 0,00005 |
| KRAS | ↓-2,28* | 0,00004 | ↓-1,18* | 0,00008 | ↓-2,67* | 0,00004 | ↓-3,29* | 0,00009 |
| MYC | ↓-2,55* | 0,00366 | ↑1,12* | 0,00006 | ↓-1,10* | 0,00006 | ↓-1,23* | 0,03746 |
| NRAS | ↑1,16* | 0,00015 | ↑1,18* | 0,00018 | -3,43* | 0,00001 | ↓-3,70* | 0,00001 |
| RRAS | ↑1,34* | 0,01637 | -1,60 | 0,60253 | ↑1,17* | 0,00007 | -1,15 | 0,05788 |
| YWHA family genes | ||||||||
| YWHAB | ↓-2,27* | 0,00027 | ↑1,04* | 0,00038 | ↓-2,46* | 0,00001 | ↓-1,27* | 0,00014 |
| YWHAE | ↓-1,57* | 0,00001 | ↓-1,29* | 0,00003 | ↓-2,92* | 0,00001 | ↓-3,86* | 0,00001 |
| YWHAG | ↓-1,85* | 0,01506 | ↓-1,29* | 0,00280 | ↓-1,08* | 0,00008 | ↓-1,56* | 0,00496 |
| YWHAH | ↓-1,73* | 0,00376 | ↓-1,38* | 0,00007 | ↓-1,88* | 0,00009 | ↓-1,55* | 0,00754 |
| YWHAQ | 1,01 | 0,10424 | ↓-1,10* | 0,00007 | ↓-2,35* | 0,00001 | ↓-4,10* | 0,00001 |
| YWHAZ | ↓-1,10* | 0,00029 | ↑1,10* | 0,00081 | ↓-2,38* | 0,00002 | ↓-4,10* | 0,00082 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



