
Submitted:
28 April 2025
Posted:
28 April 2025
You are already at the latest version
Abstract
This study was carried out to investigate the effect of turkey feed supplementation with riboflavin and protease enzyme on rectal temperature and pulse rate as a measure of heat stress during finishing stages. One hundred one-day old British United broiler turkey poults were randomly distributed into four dietary treatments T1, T2, T3, and T4 (T1 as control, T2 containing riboflavin at 6mg/kg, T3 containing protease enzyme at 1000mg/kg, and T4 containing combination of riboflavin and protease at 6mg/kg and 1000mg/kg. During the finishing stages of the birds, the rectal temperature and pulse rate of two birds from each replicate were recorded. They were recorded twice every week during the experimental period. Also, ambient temperature and relative humidity were recorded at the time of taking records on the physical parameters. The data collected were analyzed using SYSTAT. Results of turkey fed supplementation with riboflavin and protease enzyme showed no effect on rectal temperature and pulse rate of finishing turkeys. It was concluded that the addition of riboflavin and protease enzyme at the specified levels did not influence the rectal temperature and pulse rate of finishing turkeys and therefore heat stress.
Keywords:
riboflavin
; protease enzyme
; heat stress
; finishing turkeys
1. Introduction
According to Selye (1976), “stress is the nonspecific response of the body to any demand”, whereas stressor can be defined as “an agent that produces stress at any time”. Therefore, stress represents the reaction of the organism (i.e., a biological response) to stimuli that disturb its normal physiological equilibrium or homeostasis.
Heat stress results from a negative balance between the net amount of energy flowing from the animal’s body to its surrounding environment and the amount of heat energy produced by the animal. This imbalance may be caused by variations of a combination of environmental factors (e.g., sunlight, thermal irradiation, and air temperature, humidity and movement), and characteristics of the animal (e.g., species, metabolism rate, and thermoregulatory mechanisms). Environmental stressors, such as heat stress, are particularly detrimental to animal agriculture (Nienaber and Hahn, 2007; Nardone et al., 2010; Renaudeau et al., 2012; Rostango and Lara, 2013).
Extreme heat stress during the summer is of great interest in the poultry industry (Park et al., 2013). Heat stress is largely a problem in birds selected for high rates of growth or egg production under temperate conditions. A high rate of production usually carries the penalty of higher metabolic heat production and the necessity to dissipate this heat to a warm or hot environment (MacLeod et al., 2004).
For many years, researchers have been investigating the effect of high environmental temperature on the performance of different poultry species, including turkeys (Kohne and Jones, 1976; McKee and Sams, 1997), young chickens (Henken et al., 1983), broilers (Cooper and Washburn, 1998), broiler breeders (McDaniel et al., 1995), and laying hens (Emery et al., 1984; Muiruri and
Harrison, 1991; Whitehead et al., 1998), and have found that high environmental temperatures have deleterious effects on productive performance (Mashaly et al., 2004). Ambient temperatures exceeding the thermal comfort zone increase birds’ heat load, resulting in a decrease in feed consumption, feed efficiency, body weight gain and livability (Borges et al., 2003; Daghir et al., 2008). In addition, meat quality, intestinal microflora and immune system were adversely affected by high ambient temperatures (Borges et al., 2007).
Riboflavin (Vitamin B2) (7, 8-dimethyl-10-ribityl-isoalloxazine) is a water soluble vitamin present in a wide variety of foods. It was initially isolated, although not purified, from milk whey in 1879 and given the name lactochrome. It can be crystallized as orange-yellow crystals and in its pure form is poorly soluble in water. Its most important biologically active forms, flavin adenine dinucleotide (FAD) and flavin mononucleotide (FMN), participate in a range of redox reactions, some of which are absolutely key to the function of aerobic cells (Powers et al., 2003) . Early classic studies identified a riboflavin binding protein in chicken egg white that is induced by estrogen and is essential to fetal survival (White and Merrill, 1988). Also, negative effects caused by heat stress in males have been shown in different studies. Semen volume, sperm concentration, number od live sperm cells and motility decreased when males were subjected to heat stress (Joshi et al., 1980; McDaniel et al., 1995; McDaniel et al., 2004; Lara and Rostango, 2013).
Recent years have witnessed a phenomenal increase in the use of enzymes as industrial catalysts. Proteases (synonymous as peptidase or proteinase) constitute a very large and complex group of enzymes, widely utilized in a host of industries. They differ in properties such as substrate specificity, active site and catalytic mechanism, pH and temperature optima, and stability profiles. Studies relating to such properties are imperative for the successful application of these enzymes in their respective industry (Sumantha et al., 2005). The main sources of the enzymes were from
animals (e.g., calf stomach), plants (e.g., pineapple, fig, and papaya), microbes (e.g., Bacillus spp., Pseudomonas spp.) etc. but the production of enzymes from plant and animal sources is limited due to climatic reasons and ethical issues, respectively (Rao et al., 1998; Shafee et al., 2005).
Proteolytic enzymes are capable of hydrolyzing peptide bonds and are also referred to as peptidases, proteases or proteinases (Barrett and McDonald, 1986). The physiological function of proteases is necessary for all living organisms, from viruses to humans, and proteolytic enzymes can be classified based on their origin: microbial (bacterial, fungal and viral), plant, animal and human enzymes can be distinguished (Mótyán et al., 2013).
In hot climates, periods of high temperatures have a negative effect on the health and performance of domestic animals. The rectum is the final straight portion of the large intestine in some mammals, and the gut in others. Measures of rectal temperature (RT), pulse-rate (PR) and respiratory rate (RR) are some of the most important determinants of the adaptation of poultry to the tropical environment. They also, to a large extent, determine the profitability of the poultry enterprise (Ilori et al., 2012).
It has been established that heat stress is evaluated by measuring the rectal temperature (RT), which is a true reflection of internal body temperature and a reliable index of thermal balance (Bianca, 1976; Mittal and Ghosh, 1979; Ayo et al., 1998).
Data on the variation in rectal temperature and pulse rate of turkeys in response feed supplementation with riboflavin and protease enzyme during the hot-dry season are currently lacking in available literature.
1.1. Justification
Since no single control method has proven most effective in the control of heat stress in poultry production, different studies have been investigating integrated management practices in poultry heat stress. Therefore, this study seeks to investigate the effect of turkey feed supplementation with riboflavin and protease enzyme on rectal temperature and pulse rate which are among the physical parameters of heat stress.
1.2. Objectives
1.2.1. Broad Objective
To determine the effect of turkey feed supplementation with riboflavin and protease enzyme on two physical heat stress parameters during finishing stages of turkey production.
1.2.2. Specific Objectives
- 1)
- To determine the rectal temperature of finishing turkeys fed diets supplemented with riboflavin (vitamin B2) and/or protease enzyme.
- 2)
- To determine the pulse rate of finishing turkeys fed diets supplemented with riboflavin (vitamin B2) and/or protease enzyme.
2. Literature Review
2.1. Poultry Production
Poultry production has been reported to be the fastest growing in the livestock industry, and more particularly in tropical and sub-tropical regions of the world (Daghir et al., 2009; Holik et al., 2009). Today a large percentage of the world’s poultry population is located in regions where heat stress is a major management problem at some particular moments of the bird’s productive lives (Ajakaiye et al., 2011).
The establishment of poultry farms in new areas means that live birds of all ages have to be transported by road, across different ecological zones throughout the year (Ajakaiye et al., 2010). Many elements of the transport process can be harmful to the birds (Knowles and Brown, 1990; Nicol and Saville-Weeks, 1993). These includes handling by humans, air temperature changes, high ambient temperature (AT) and relative humidity (RH), removal of food and water, novelty, confinement, noise, motion, micro-thermal core within the vehicle and the use of inappropriate vehicles (Mitchell and Kettlewell, 1998; Bedonova et al., 2006; Vecerek et al., 2006). The adverse effects of these factors and their combinations may range from mild discomfort and aversion to death (Ajakaiye et al., 2010).
During the early summer season, the turkey industry reports substantial losses in yield due to formed turkey breast products with poor water-holding capacity, poor texture, and pale color. These meat characteristics are consistent with those observed in pale, soft, and exudative (PSE) pork. According to Ogah (2011), Turkey is not common among poultry growers in Nigeria: a number of farms are beginning to breed the bird at commercial level owing to increasing interest as a provider of meat complementing chicken. They are mostly located in urban areas and are gradually spreading even to village farms. The fast growth in the industry requires and intensive research approach to boast its production especially considering the potentials associated with it. The first approach in livestock characterization apart from evaluation of its production performance is the evaluation of body size and conformation (Ibe et al., 1989). Assessment of body weight and linear body measurements have been found useful in quantifying body size and shape (Ibe and Ezekwe, 1994). Linear body measurements have also been used to predict live weight in poultry (Chhabra et al., 1972; Monsi et al., 1992; Gueye et al., 1998). The multitude of different body measurements available has lead several researchers to use multivariate techniques to simultaneously examine the relationship among body measurements and production traits (Brown et al., 1973). Use of principal component analysis to examine the relationship between measurement of size and shape in poultry have been reported in chicken (Ibe et al., 1989; Yakubu et al., 2009) and duck (Shahin et al., 1996; McCracken et al., 2000; Ogah et al., 2009). This multivariate procedure describes the total variation in a large system of body measurements in terms of a few artificial varieties (Ogah et al., 2011).
2.2. Heat Stress
Heat stress results from a negative balance between the net amount of energy flowing from the animal’s body to its surrounding environment and the amount of heat energy produced by the animal. This imbalance may be caused by variations of a combination of environmental factors (e.g., sunlight, thermal irradiation, and air temperature, humidity and movement), and characteristics of the animal (e.g., species, metabolism rate, and thermoregulatory mechanisms). Environmental stressors, such as heat stress, are particularly detrimental to animal agriculture (Nienaber and Hahn, 2007; Nardone et al., 2010; Renaudeau et al., 2012).
High ambient temperature is a problem in many parts of the world. Heat stress has been associated with decreases in broiler weight gain, feed intake, feed efficiency, nitrogen retention, protein digestibility and total mineral retention (Austic et al., 1985; Sahin and Kucuk, 2003).
The high humidity will aggravate the bad influence of high temperature on the broiler (Yahav et al., 1995; Gu et al., 1999; Lin et al., 2005a, b) reported that humidity could affect the thermoregulation of broiler chickens by redistributing heat within the body at high, low and even thermoneutral temperatures, high humidity above 60% impaired the heat transmission from body core to the surrounding at high temperature. At hot environment, the chemical composition of chicken is changed (Leenstra and Cahaner, 1991; Geraert et al., 1996) and meat sensory quality is decreased by heat stress (Osman et al., 1989; Northcutt et al., 1994; Li et al., 1999). The oxidative damage of tissues induced by heat stress is one of the possible reasons (Sandercock et al., 2001; Lin et al., 2006b).
It has been reported that chronic heat exposure negatively affects fat deposition and meat quality in broilers, in a breed-dependent manner (Lu et al., 2007). In fact, recent studies demonstrated that heat stress is associated with depression of meat chemical composition and quality in broilers (Dai et al., 2012; Imik et al., 2012). Another recent study (Zhang et al., 2012) demonstrated that chronic heat stress decreased the proportion of breast muscle, while increasing the proportion of thigh muscle in broilers. Moreover, the study also showed that protein content was lower and fat deposition higher in birds subjected to heat stress.
Environmental stress causes oxidative stress and impairs antioxidant status in vivo (Halliwell and Gutteridge, 1989; Sahin et al., 2001). Mujahid et al. (2005) shown that superoxide production by the skeletal muscle mitochondria of meat type chickens is significantly enhanced by heat stress.
This in turn was associated with a heat-induced increase in rectal and muscle temperatures, leading to a significant body weight loss.
When the protein level increases under the heat stress, even more metabolic heat is produced, thus it is helpful to increase essential amino acids like methionine and lysine. As well, stimulating feed intake by providing high preference ingredients such as soy oil or molasses, and providing vitamin C are helpful to minimize heat stress (Leeson and Summers, 1991; Park et al., 2013). The benefits of vitamin C, vitamin E, potassium chloride, ammonium chloride, potassium sulphate and sodium bicarbonate in drinking water or feed during the hot period of the day have been reported (Sahin and Kucuk, 2001; Ciftci et al., 2005). For this purpose, vitamin C and vitamin E are used in the poultry diet because of their anti-oxidant properties in the neutralization of the free radicals generated during heat stress (Ramnath et al., 2008). Turkey poults are sensitive to environmental temperature with a lower critical temperature of about 29 °C (Scott et al., 1983). As a consequence of increased environmental temperature, the internal body core and skin temperature of the bird increases. The bird begins to breathe more rapidly, while its heart rate and oxygen consumption increase. The increase in oxygen consumption is a direct reflection of the increased energy demand for maintenance due to panting (Miles et al., 1999). Turkeys cannot tolerate a concurrent high temperature and high humidity. When the surrounding air is moist, it cannot absorb as much moisture from the lungs; consequently the bird must pant faster. Similarly, when the outside temperature is high, respiratory rate is increased. With both high temperature and high humidity the bird may not be able to pant fast enough to remove the heat from its body (El Boushy & Van Marle, 1978). The increase in the breathing rate is accompanied by an increase in the loss of moisture from the body. To compensate for this loss, the bird drinks more water to avoid dehydration. Eventually, the bird drinks more water than it can exhale, and the surplus is excreted through the droppings. The amount of moisture in the ambient air (humidity) also affects the panting rate; the higher the humidity the more rapid the respiration (North et al., 1984). Consequently, either at low or high inside air temperature, the rate of productive performance of turkey poults decreases. Therefore, inside air temperature when brooding turkey should be kept at an optimal level (Konca et al., 2001) by reducing heat loss and using supplementary heating when needed (Anonymous et al., 1987 and Gencoglan et al., 2009).
It has been established that heat stress is evaluated by measuring the rectal temperature (RT), which is a true reflection of internal body temperature and a reliable index of thermal balance (Bianca et al., 1976; Mittal and Ghosh, 1979; Ayo et al., 1998). Changes in reproductive hormone secretion represent the final sequence in the neuroendocrine pathway leading to the diminished reproductive performance associated with stress (Rozenboim et al., 2007).
2.3. Riboflavin (Vitamin B2)
Almost all riboflavin in tissues is enzyme bound, such as FAD covalently bound to succinic dehydrogenase (EC 1.3.5.1) (Singer and Kenney, 1974). Unbound flavins are relatively labile and are rapidly hydrolyzed to free riboflavin, which diffuses from cells and is excreted. The intracellular phosphorylation of riboflavin is therefore a form of metabolic trapping key to riboflavin homeostasis (Gastaldi et al., 2000; Powers et al., 2003).
Riboflavin deficiency is associated with demyelination of peripheral nerves and consequent locomotion difficulty. The classical disease associated with this deficiency is known as curled toe paralysis. It is reported that a condition similar to curled-toe paralysis, which occurred in turkeys fed with 3-Nitrogen, 4-hydroxyphenylarsonic acid. It was found that dietary levels of this substance greater that 50ppm caused demyelization of peripheral nerves (Das et al., 2011).
Milk and dairy products make the greatest contribution to riboflavin intake in Western diets, making riboflavin exceptional among the water-soluble vitamins. National dietary surveys in the United Kingdom report that, on average, milk and dairy products contribute 51% of intake in preschool children, 35% in schoolchildren, 27% in adults, and 36% in the elderly. Cereals, meats (especially offal), and fatty fish are also good sources of riboflavin, and certain fruit and vegetables, especially dark-green vegetables, contain reasonably high concentrations (Powers et al., 2003). A small amount of riboflavin is present in foods as free riboflavin, which is an isoalloxazine ring bound to a ribitol side chain; most is present as the derivative FAD, and a smaller amount occurs as the monophosphorylated form, FMN. FAD and FMN occur predominantly in a non-covalently-bound form to enzymes; flavins that are covalently bound do not appear to be available for absorption (McCormick et al., 1972). In contrast with most foodstuffs, milk and eggs contain appreciable quantities of free riboflavin bound to specific binding proteins (Zanette et al., 1984). The flavocoenzymes such as FMN and FAD comprise the major part of riboflavin in blood plasma (Ohkawa et al., 1982). The bioactive forms of riboflavin are hydrolyzed to riboflavin before they enter into the cells, but riboflavin is accumulated in tisues by resynthesis of flavocoenzymes (A.W et al., 1983; McCormick and Zhang, 1993).
2.4. Protease Enzymes
The use of exogenous enzymes is not a new concept and has been extensively studied and reported (Campbell and Bedford, 1992; Leeson et al., 1996; Seskevicience et al., 1999; Smits and Annison, 1996).
According to the Nomenclature Committee of the International Union of Biochemistry and Molecular Biology, proteases are classified under the subgroup 4 of Group 3 (hydrolases). However, proteases do not comply easily with the general system of enzyme nomenclature due to their huge diversity of action and structure. On the basis of their site of action on protein substrates, proteases are broadly classified as endo- or exo-enzymes (Rao et al., 1998). They are further caegorized as serine proteases, aspartic proteases, cysteine proteases or metallo proteases— depending on their catalytic mechanism. Proteases are also classified into different clans and families depending on their amino acid sequences and evolutionary relationships. Based on the pH optima, they are referred to as acidic, neutral, or alkaline proteases (Rao et al., 1998; and V. N. Jisha et al., 2013).
Proteases are extensively applied enzymes in several sectors of industry and biotechnology, furthermore, numerous research applications require the use of them, including the production of Klenow fragments, peptide synthesis, digestion of unwanted proteins during nucleic acid purification, use of proteases in cell culture experiments and in tissue dissociation, preparation of recombinant antibody fragments for research, diagnostics and therapy, exploration of the structure- function relationships by structural studies, removal of affinity tags from fusion proteins in recombinant protein techniques, peptide sequencing, and proteolytic digestion of proteins in proteomics (Mótyán et al., 2013).
Proteolytic enzymes are capable of hydrolyzing peptide bonds and are also referred to as peptidases, proteases or proteinases (Barrett and McDonald, 1986).
The physiological function of proteases is necessary for all living organisms, from viruses to humans, and proteolytic enzymes can be classified based on their origin: microbial (bacterial, fungal and viral), plant, animal and human enzymes can be distinguished.
Proteolytic enzymes belong to the hydrolase class of enzymes (EC 3) and are grouped into the subclass of the peptide hydrolases or peptidases (EC 3.4). Depending on the site of enzyme action the proteases can also be subdivided into exopeptidases or endopeptidases. Exopeptidases catalyze the hydrolysis of the peptide bonds near the N- or C-terminal ends of the substrate. Aminopeptidases can liberate single amino acids (EC 3.4.11), dipeptides (dipeptidyl peptidases, EC 3.4.14) or tripeptides (tripeptidyl peptidases EC 3.4.14) from the N-terminal end of their substrates. Single amino acids can be released from dipeptide substrates by dipeptidases (EC 3.4.13) or from polypeptides by carboxypeptidases (EC 3.4.16-3.4.18), while peptidyldipeptidases (EC 3.4.15) liberate dipeptides from the C-terminal end of a polypeptide chain. Endopeptidases cleave peptide bonds within and distant from the ends of a polypeptide chain (Rao et al., 1998). Based on the catalytic mechanism and the presence of amino acid residue(s) at the active site the proteases can be grouped as aspartic proteases, cysteine proteases, glutamic proteases, metalloproteases, asparagine proteases, serine proteases, threonine proteases, and proteases with mixed or unknown catalytic mechanism (Rawlings et al., 2012).
The current classification system further classifies the proteases into families based on sequence similarities, furthermore, homologous families are grouped into clans using a structure-based classification (Rawling et al., 2012; Rawlings and Barrett, 1993). Classification and nomenclature of proteolytic enzymes as well as a detailed description of individual proteases is available in the MEROPS database (Rawlings et al., 2012 and Mótyán et al., 2013). Action of the proteolytic enzymes is essential in several physiological processes, e.g., in digestion of food proteins, protein turnover, cell division, blood-clotting cascade, signal transduction, processing of polypeptide hormones, apoptosis and the life-cycle of several disease-causing organisms including the replication of retroviruses (Neurath and Walsh, 1976; Devlin et al., 2002). Due to their key role in the life-cycle of many hosts and pathogens they have great medical, pharmaceutical, and academic importance (Li et al., 2013; Craik et al., 2011; Antonelli and Turriziani, 2012). They are intensively studied to explore their structure-function relationships, to investigate their interactions with the substrates and inhibitors, to develop therapeutic agents for antiviral therapies (Antonelli and Turriziani, 2012) or to improve their thermostability, efficiency and to change their specificity by protein engineering for industrial or therapeutic purposes (Li et al., 2013). Studying proteolytic enzymes is highly justified by their key role in several fields of industry (Rao et al., 1998; Kirk et al., 2002; Rani et al., 2012; Ray et al., 2012), as well.
2.6. Physiological Parameters
According to Altan et al. (2000), exposure of broilers to 39 °C significantly increased rectal temperatures, heterophil and basophil proportions and Heterophil to Lymphocyte (H/L) ratios, and decreased monocyte and lymphocyte proportions.
Rectal temperature (RT), respiratory rate (RR) and heart rate (HR) are important physiological parameters most relevant for on-the-spot evaluation of the health status and adaptability of animals, including poultry species (Bianca, 1976; Ayo et al., 1998). The parameters are easily measured and are of value in the determination of state of stress in birds, especially during the process of transportation in rural areas where laboratory facilities may be lacking.
3. Materials and Methods
3.1. Experimental Location
The experiment was carried out at the turkey unit, College of Animal Science and Livestock Production (COLANIM) Farms, Federal University of Agriculture Abeokuta, Ogun State, Nigeria. The farm is located on latitude 70 10’N and longitude 30 2’E and lies in the southwestern part of Nigeria with prevailing tropical climate and a mean annual rainfall of about 1037mm.The mean monthly ambient temperate ranges from 280C in December to 360C in February with a yearly average relative humidity of about 82%.
3.2. Experimental Birds, Materials and Management
A total of 100 one-day old British United broiler turkey poults were used for the experiment. The poults were bought from a commercial hatchery: O. H. L., Ibadan, Oyo State, Nigeria. The brooding house was prepared and disinfected before the arrival of the poults. The poults were brooded intensively for four weeks at a controlled temperature of about 34.50C for the first two days and reduced gradually every week at 20C to a final ambient temperature of about 270C at the fourth week of brooding. The brooding was carried out on a deep litter system, using wood shavings as the litter materials. Management routine practices were carried out, feeders and drinkers were cleaned daily. The vaccination and medication schedule was strictly adhered to. After the brooding period, the poults were further reared for four weeks (28 days), prior to the commencement of the experiment. Birds were fed with pre-starter diet (0-4 weeks),starter (4-8 weeks),grower (8-12 weeks) and finisher (12-18 weeks). The experiment was commenced at the end of the starter phase.
3.3. Source of Riboflavin and Protease Enzyme
There was an inclusion of riboflavin and protease enzyme in the bird’s diet. Riboflavin (vitamin B2) was procured from a trusted pharmaceutical company in Abeokuta, Ogun State. Also, protease enzyme was procured from Norogen Nigeria Limited, Lagos, Nigeria.
3.4. Experimental Design
Eighty broiler turkeys of similar weight range were selected and allotted on weight equalization basis to four dietary treatment at 56 days. There were four experimental diets formulated for feeding growing and finishing phases of the study. The diets were supplemented with 1000mg/kg of protease enzyme, 6mg/kg of riboflavin (vitamin B2), (1000mg+6mg)/kg of both protease enzyme and riboflavin and a standard basal diet containing no supplement.
3.5. Experimental Diet

Table 1.
Experimental Diets.
| Four experimental diets were used and they are: | |
| Diet 1 | basal diet without riboflavin and protease enzyme. |
| Diet 2 | basal diet with riboflavin 6mg/kg. |
| Diet 3 | basal diet with protease enzyme 1000mg/kg. |
| Diet 4 | basal diet with riboflavin and protease enzyme (6mg+1000mg)/kg. |
3.6. Data Collection
3.6.1. Rectal Temperature
The rectal temperature of the birds was taken by inserting clinical digital thermometer (0.10C accuracy) into the rectum via the cloaca until an alarm sound beeps, indicating the end of the reading. It was done twice every week during the experimental period.
3.6.2. Pulse Rate
The pulse rate of the birds was taken with the aid of a stethoscope. It was read by the number of heart beats per minutes. This also was taken twice every week during the course of the experiment.
3.6.3. Environmental Indicators in the Poultry House
Ambient temperature and relative humidity were taken with the aid of the hygrometer. They were taken twice in a week at the time of taking records on physical parameters of the birds during the experimental period.
2.7. Statistical Design and Analysis
The data collected was analyzed using factorial experiment of SYSTAT. Results were reported at least square means (LS Means).
Table 2.
Gross composition of pre-starter (0-4 weeks) and starter (4-8 weeks) diets.
| Gross composition of diets (%) | ||
| Ingredient | Pre-starter (0-4 weeks) | Starter (4-8 weeks) |
|
Maize |
42.50 |
50.00 |
| Soybean Meal | 40.70 | 36.00 |
| Fish Meal (72%) | 9.00 | 8.90 |
| Bone Meal | 4.50 | 3.00 |
| Limestone | 2.00 | 1.00 |
| *Vitamin/trace mineral Premix | 0.50 | 0.15 |
| Lysine | 0.10 | 0.20 |
| DL Methionine | 0.40 | 0.50 |
| Salt | 0.30 | 0.25 |
| Total | 100.00 | 100.00 |
| Calculated analyses | ||
| Metabolizable Energy (Kcal/kg) | 2,840 | 2878.7 |
| Crude Protein (%) | 28.19 | 26.62 |
| Crude Fibre (%) | 3.00 | 3.97 |
| Ether Extract (%) | 3.78 | 2.89 |
| Calcium (%) | 2.16 | 1.39 |
| Phosphorus (%) | 0.86 | 0.67 |
| Lysine (%) | 1.91 | 1.89 |
| Methionine (%) | 0.88 | 0.68 |
| Arginine (%) | 1.82 | 1.71 |
Vitamin/Mineral Premix composition per Kg diet: vit A: 40, 000IU, vit D3: 4000IU, vit E: 40.0 mg, vit K3: 8mg, vit B1: 1.0mg, vit B2: 8mg, vit B6: 5mg, vit B12: 0.025mg, Niacin: 60mg, Panthothenic acid: 20mg, Folic acid: 2000mg, Biotin: 150mg, , Iron: 32mg,Manganese: 64mg, Zinc: 40mg, Copper: 8mg, Cobalt: 80mg, Iodine:0.15mg, Selenium: 0.2mg, Choline: 300mg.
Table 3.
Gross composition of experimental grower diet (8-12 weeks).
|
T1 (Control ) |
T2 (Riboflavin ) |
T3 (Protease) |
T4 (Riboflavin and Protease) |
|
|
Supplemental levels of feed additives |
0 | 6mg/kg | 1000mg/k g |
6mg+1000mg/kg |
| Ingredient composition | ||||
| Maize | 555 | 555 | 555 | 555 |
| Soya Bean Meal | 260 | 260 | 260 | 260 |
| Fish meal (72% CP) | 65 | 65 | 65 | 65 |
| Wheat offal | 50 | 50 | 50 | 50 |
| Palm oil | 17 | 17 | 17 | 17 |
| Bone Meal | 20 | 20 | 20 | 20 |
| Limestone | 15 | 15 | 15 | 15 |
| *Vitamin/mineral Premix | 5 | 5 | 5 | 5 |
| Methaonine | 2 | 2 | 2 | 2 |
| Lysine | 1.5 | 1.5 | 1.5 | 1.5 |
| Salt (NaCl) | 2.5 | 2.5 | 2.5 | 2.5 |
| Total | 1000.00 | 1000.00 | 1000.00 | 1000.00 |
| Calculated analysis | ||||
| Metabolisable Energy (Kcal/kg) | 2952.92 | 2952.92 | 2952.92 | 2952.92 |
| Crude Protein (%) | 21.90 | 21.90 | 21.90 | 21.90 |
| Crude Fibre (%) | 3.70 | 3.70 | 3.70 | 3.70 |
| Ether Extract (%) | 3.39 | 3.39 | 3.39 | 3.39 |
| Phosphorus (%) | 0.90 | 0.90 | 0.90 | 0.90 |
| Calcium (%) | 2.38 | 2.38 | 2.38 | 2.38 |
| Lysine (%) | 1.29 | 1.29 | 1.29 | 1.29 |
| Methionine (%) | 0.62 | 0.62 | 0.62 | 0.62 |
Vitamin/Mineral Premix composition per Kg diet: vit A: 40, 000IU, vit D3: 4000IU, vit E: 40.0 mg, vit K3: 8mg, vit B1: 1.0mg, vit B2: 8mg, vit B6: 5mg, vit B12: 0.025mg, Niacin: 60mg, Panthothenic acid: 20mg, Folic acid: 2000mg, Biotin: 150mg, Iron:g, Manganese: 64mg, Zinc: 40mg, Copper: 8mg, Cobalt: 80mg, Iodine: 0.15mg, Selenium: 0.2mg, Choline: 300mg.
Table 4.
Gross composition of experimental finisher diet (12-18 weeks).
|
T1 (Control ) |
T2 (Riboflavin ) |
T3 (Protease) |
T4 (Riboflavin and Protease) |
|
|
Supplemental levels of feed additives |
0 | 6mg/kg | 1000mg/k g |
6mg+1000mg/kg |
| Ingredient composition | ||||
| Maize | 645 | 645 | 645 | 645 |
| Soya Bean Meal | 205 | 205 | 205 | 205 |
| Fish meal (72% CP) | 45 | 45 | 45 | 45 |
| Wheat offal | 37 | 37 | 37 | 37 |
| Palm oil | 15 | 15 | 15 | 15 |
| Bone Meal | 27 | 27 | 27 | 27 |
| Limestone | 15 | 15 | 15 | 15 |
| *Vitamin/mineral Premix | 5 | 5 | 5 | 5 |
| Methaonine | 2 | 2 | 2 | 2 |
| Lysine | 1.5 | 1.5 | 1.5 | 1.5 |
| Salt (NaCl) | 2.5 | 2.5 | 2.5 | 2.5 |
| Total | 1000.00 | 1000.00 | 1000.00 | 1000.00 |
| Calculated analysis | ||||
| Metabolisable Energy (Kcal/kg) |
3030.64 | 3030.64 | 3030.64 | 3030.64 |
| Crude Protein (%) | 18.31 | 18.31 | 18.31 | 18.31 |
| Crude Fibre (%) | 3.45 | 3.45 | 3.45 | 3.45 |
| Ether Extract (%) | 2.84 | 2.84 | 2.84 | 2.84 |
| Phosphorus (%) | 0.86 | 0.86 | 0.86 | 0.86 |
| Calcium (%) | 2.19 | 2.19 | 2.19 | 2.19 |
| Lysine (%) | 1.08 | 1.08 | 1.08 | 1.08 |
| Methionine (%) | 0.56 | 0.56 | 0.56 | 0.56 |
*Vitamin/Mineral Premix composition per Kg diet: vit A: 40, 000IU, vit D3: 4000IU, vit E: 40.0 mg, vit K3: 8mg, vit B1: 1.0mg, vit B2: 8mg, vit B6: 5mg, vit B12: 0.025mg, Niacin: 60mg, Panthothenic acid: 20mg, Folic acid: 2000mg,. Biotin: 150mg, Iron:g, Manganese: 64mg, Zinc: 40mg, Copper: 8mg, Cobalt: 80mg, Iodine: 0.15mg, Selenium: 0.2mg, Choline: 300mg.
4. Results and Discussion
4.1. Results
4.1.1. Effect of Feed Supplementation with Riboflavin and Protease Enzyme on Rectal Temperature of Finishing Turkeys
The effect of feed supplementation with riboflavin and protease enzyme on rectal temperatures of finishing turkeys is presented in Table 5. The analysis of variance showing effect of riboflavin and protease enzyme on rectal temperature of finishing turkeys is presented on Table 6. There was no significant (P<0.05) difference in the rectal temperatures of birds offered the four experimental diets within age. The range of rectal temperature observed was between 40.4500C and 41.1310C. However, age had effect on rectal temperature. Rectal temperature decreased significantly at the age of 16 and 17 weeks compared to earlier ages.
4.1.2. Effect of Feed Supplementation with Riboflavin and Protease Enzyme on Pulse Rate of Finishing Turkeys
The effect of feed supplementation with riboflavin and protease enzyme on pulse rate of finishing turkeys is presented on Table 7. The analysis of variance showing effect of riboflavin and protease enzyme on pulse rate of finishing turkeys is presented on Table 8. There was no significant (P<0.05) difference in pulse rate within age of birds offered the four experimental diets. The range of pulse rate observed was between 150.250 and 171.250. However, age had effect on pulse rate. Pulse rate decreased significantly at the age of 16 and 17 weeks compared to earlier ages.
4.1.3. Result of Ambient Temperature and Relative Humidity of the Poultry House
4.1.3.1. Ambient Temperature
Ambient temperature is shown on Figure 1. The range of ambient temperature observed was between 29.20C and 29.80C. Ambient temperature decreased significantly with the poultry house during week 16 and 17 of the experiment.
4.1.3.2. Relative Humidity
Relative humidity is shown on Figure 2. The range of relative humidity observed was between 72% and 74%. Relative humidity decreased significantly during week 17 compared to earlier periods of the experiment.
4.2. Discussion
In this current study, it was observed that there was no significant difference in the rectal temperatures and pulse rate of birds offered the four experimental diets within age. The addition of riboflavin and protease enzyme at the specified levels did not influence these physiological parameters. This suggest that supplementation of riboflavin and protease enzyme did not effect any changes in metabolic heat generated during the finishing stage of turkey birds. However, the benefits of vitamin C, vitamin E, potassium chloride, ammonium chloride, potassium sulphate and sodium bicarbonate in drinking water or feed during hot period of the day have been reported (Sahin and Kucuk, 2001; Cifti et al., 2005).
However, age had effect on rectal temperature and pulse rate of the birds. The rectal temperature and pulse rate decreased significantly at the age of 16 and 17 weeks compared to earlier ages. In consonance, the ambient temperature and relative humidity decreased significantly at week 16 and 17 compared to earlier periods of the experiment. The simultaneous decrease in body temperature, pulse rate and the environmental parameters suggest interdependence. As the environmental queues, changes in the physiological parameters will be influenced. The rectal temperature and pulse rate of the birds decreased significantly at week 16 and 17 compared to earlier periods of the experiment. This suggest that, high humidity will aggravate the bad influence of high temperature on the turkey. Yahav et al. (1995) and Lin et al. (2005a,b) reported that humidity could affect the thermoregulation of broiler chickens by redistributing heat within the body at high, low and even thermoneutral temperatures. High humidity above 60% will impair the heat transmission from body core to the surrounding at high temperature. The ambient temperature and relative humidity had effect on rectal temperature and pulse rate in reduction of metabolic heat of the birds.
5. Conclusions
It is concluded that the addition of riboflavin and protease enzyme at the specified levels did not influence the rectal temperatures and pulse rate of finishing turkeys and therefore suggest that the two types of supplements at the levels used are not likely to be effective in controlling heat stress in turkey.
5.1. Recommendation
Addition of riboflavin and protease enzyme at 6mg/kg and 1000mg/kg may not be effective on these two physiological parameters. However, further studies are required to ascertain whether other levels of supplementation may be effective.
References
- A.w TY, Jones DP, McCormick DB. (1983). Uptake of riboflavin by isolated rat liver cells. Journal Nutrition 113:1249-54. [CrossRef]
- Ajakaiye J.J., Ayo J.O. and Ojo S.A. (2010). Effects of heat stress on some blood parameters and egg production of Shika Brown layer chickens transported by road. Biological Research 43:183-189. [CrossRef]
- Ajakaiye, J.J; Perez-Bello, A; Mollineda-Trujillo, A. (2011). Impact of heat stress on egg quality hens supplemented with I-ascorbic and dl-tocopherol acetate. Veterinrski Arhiv 81 (1), 119-132.
- Altan, O.; Altan A.; Oguz I.; Pabuccuoglu, A.; and Konyalioglu, S. (2000). Effects of heat stress on growth, some blood variables and lipid oxidation in broilers exposed to high temperature at an early age. British Poultry Science, 41:489-493. [CrossRef]
- Anonymous, (1987). Animal Houses - rules for isolation and heating TS 5087, Turk Standards Institute. NecatiBeyEvanue No: 112, Bakanliklar-Ankara. pp. 1-12 (in Turkish).
- Antonelli, G.; Turriziani, O. 2012. Antiviral therapy: Old and current issues. International Journal of Antimicrobial Agents 40, 95–102. [CrossRef]
- Austic, R.E., (1985). Feeding poultry in hot and cold climates. In: Stress Physiology in Livestock.Vol. 3. Yousef, M.K. (Ed.). CRC Press, Boca Raton, FL., pp: 123-136.
- Ayo, J. O., Oladele, S. B., Fayomi, A., Jumbo, S.D., and Hambolu, J.O. (1998). Body temperature, respiration and heart rate in Red Sokoto goat during harmattan season. Bulletin of Animal Health Production of Africa, 46: 161-166.
- Barrett, A.J., and McDonald, J.K. (1986). Nomenclature: Protease, proteinase and peptidase. Biochemical Journal 237, 935. [CrossRef]
- Barrett, A.J.; McDonald, J.K. (1986). Nomenclature: Protease, proteinase and peptidase. Biochemical Journal, 237, 935. [CrossRef]
- Bedanova I., Voslarova E., Vecerek V., Pistekova V., Chloupek P. (2006). Effects of reduction in floor space during crating on haematological indices in broilers. Berlin und Munchener Tierarztliche Wochenschrift 119: 17-21.
- Bianca, W. K., (1976). The Significance of Meteorology in Animal Production. International Journal of Biometeorology, 20: 139-156. [CrossRef]
- Borges, S.A., da Silva A.V.F., and Mairoka A., (2007). Acid-base balance in broilers. World’s Poultry Science Journal, 63: 73-81.
- Borges, S.A., da Silva A.V.F., Ariki J., Hooge D.M. and Cummings K.R., (2003). Dietary Electrolyte Balance for Broiler Chickens Exposed to Thermoneutral or Heat-Stress Environments. Poultry Science 82: 428-435. [CrossRef]
- Brown, C. J., Brown, J. E., and Butts, W. T. (1973). Evaluating relationship among immatured measure of size, shape and performance of beef bulls.11. The relationships between immature measures of size shape and feedlot traits in young bulls. Journal of Animal Science, vol. 36, p. 1021-1023.
- Campbell, G. L., Bedford M. R. (1992). Enzyme application for monogastric feeds: a review. Canadian Journal of Animal Science, 72, 449-466. [CrossRef]
- Chhabra, A. D., Sapra, K.L., and Sharma R. K. (1972). Shank length, growth and carcass quality in broiler breeds of poultry. Industrial and Veterinary Journal, 1972, vol. 49, no. ,p. 506-511.
- Ciftci, M., Nihatertas, O., and Guler, T. (2005). Effects of vitamin E and vitamin C dietary supplementation on egg production and egg quality of laying hens exposed to a chronic heat stress. Revue de Médecine Véterinaire 156 (Suppl. II), 107-111.
- Cooper, M. A., and Washburn, K. W. (1998). The relationships of body temperature to weight gain, feed consumption, and feed utilization in broilers under heat stress. Poultry Science 77:237–242. [CrossRef]
- Craik, C.S.; Page, M.J.; Madison, E.L. (2011). Proteases as therapeutics. Biochemical Journal 435, 1–16.
- Daghir, N. J. (2009). Nutritional strategies to reduce heat stress in broilers and broiler breeders. Lohmann information 44: 6-15.
- Daghir, N.J. (2008). Poultry Production in Hot Climates. 2nd Edition, CAB International, Wallingford, Oxfordshire, UK. Pages: 387.
- Dai, S.F.; Gao, F.; Xu, X.L.; Zhang, W.H.; Song, S.X.; Zhou, G.H. (2012). Effects of dietary glutamine and gamma-aminobutyric acid on meat colour, pH, composition, and water- holding characteristic in broilers under cyclic heat stress. British Poultry Science 53, 471–481. [CrossRef]
- Das, S.; Palai, T. K.; Mishra, S. R.; Das, D.; Jena, B. (2011). Nutrition in relation to diseases and heat stress in poultry. Veterinary World, 4:429-432. [CrossRef]
- Devlin, T.M. (2002). Textbook of Biochemistry with Clinical Correlations, 5th ed.; Wiley & Sons: New York, NY, USA.
- El Bousy, A.R. & Van Marle, A.L. (1978). The effects of climate on poultry physiology in tropics and their improvement. World’s Poultry Science Journal 34, 155-170. [CrossRef]
- Emery, D. A., Vohra P., Ernst, R. A., and Morrison, S. R. (1984). The effect of cyclic and constant ambient temperatures on feed consumption, egg production, egg weight, and shell thickness of hens. Poultry Science 63:2027–2035. [CrossRef]
- Gastaldi G, Ferrari G, Verri A, Casirola D, Orsenigo M.N, Laforenza U. (2000). Riboflavin phosphorylation is the crucial event in riboflavin transport by isolated rate enterocytes. Journal of Nutrition; 130:2556-61. [CrossRef]
- Gencoglan, S., Gencoglan C., and Akyuz, A. (2009) Supplementary heat requirements when brooding tom turkey poults. South African Journal of Animal Science, 39 (1). [CrossRef]
- Geraert, P. A., J. C. F. Padilha and S. Guillaumin. (1996). Metabolic and endocrine changes induced by chronic heat exposure in broiler chickens: biological and endocrinological variables. British Journal of Nutrition, 75:205-216. [CrossRef]
- Gu, X. H., R. Du and L. Fang. (1999). Effect of humidity on rectal temperature, plasma t3 and insulin level in broilers under high ambient temperature. Scientia Agricultura Sinica, 32:105-107.
- Gueye, E. F., Ndiaye, A., and Branckaert, R. D. S. (1998). Prediction of body weight on the basis of body measurement in mature indigenous chickens in Senegal. Livestock Research for Rural Development, vol. 10, no. 3.
- Halliwell, B.E. and J.M.C. Gutteridge, (1989). Lipid peroxidation: A radical chain reaction. In: Free Radicals in Biology and Medicine. 2nd Edn., Oxford University Press, New York, pp: 188-218.
- Henken, A. M., Groote Schaarsberg A.M. J., and Nieuwland M. G. B. (1983). The effect of environmental temperature on immune response and metabolism of the young chicken. Effect of environmental temperature on the humoral immune response following injection of sheep red blood cells. Poultry Science 62:51–58.
- Holik, V. (2009). Management of laying hens to minimize heat stress. Lohmann Information, 44:16-29.
- Ibe, S. N. (1989). Measurement of size and confirmation in commercial broilers. Journal of Animal Breeding and Genetics, vol. 106, p. 461-469. [CrossRef]
- Ibe, S. N., and Ezekwe, A. G. (1994). Quantifying size and shape differences between Muturu and N‘Dama breeds of cattle. Nigerian Journal of Animal Production, vol. 21, p.51-58. [CrossRef]
- Ilori, B.M, Peters, S.O, Yakubu, A, Imumorin, I.G, Adeleke, M.A, Ozoje, M.O, Ikeobi, C.O.N, and Adebambo, O.A. (2012). Physiological adaptation of local, exotic and crossbred turkeys to the hot and humid tropical environment of Nigeria. ActnAgriculturaeScandinavica,Section A-Animal Science.
- Imik, H.; Atasever, M.A.; Urgar, S.; Ozlu, H.; Gumus, R.; Atasever, M. (2012). Meat quality of heat stress exposed broilers and effect of protein and vitamin E. British Poultry Science, 53, 689-698. [CrossRef]
- Jisha, V. N.; Smitha, R. B.; Pradeep, S. (2013). Versatility of microbial proteases. Advances in Enzyme Research, 1(3): 39-51. [CrossRef]
- Joshi, P .C., Panda, B., and Joshi, B. C. (1980). Effect of ambient temperature on semen characteristics of White Leghorn male chickens. Indian Veterinary Journal, 57, 52–56.
- Kirk, O.; Borchert, T.V.; Fuglsang, C.C. (2002). Industrial enzyme applications. Current Opinion in Biotechnolonogy, 13, 345–351.
- Knowles T.G., and Broom D.M. (1990). The handling and transport of broilers and spent hens. Applied Animal Behaviour Science 28: 75-91. [CrossRef]
- Kohne, H. J., and Jones, J. E. (1976). The relationship of circulating levels of estrogens, corticosterone and calcium to production performance of adult turkey hens under conditions of increasing ambient temperature. Poultry Science 55:277–285. [CrossRef]
- Konca, Y. (2001). Turkey Brooding. Agricultural Research and Education Coordination (TAYEK/TYUAP), Proc. Animal Working Group Meeting in 2001. Ege Agricultural Research Institute Directorate, 27-29 March, Izmir, Publ. No: 100. pp. 21-31. (in Turkish).
- Lara L. J., RostagnoM. H. (2013). Impact of Heat Stress on Poultry Production. Animals 3, 356-369.
- Leenstra, F. and A. Cahaner. (1991). Genotype by environment interactions using fast growing, lean or fat broiler chickens, originating from the Netherlands and Israel, raised at normal or low temperature. Poultry Science 70:2028-2039.
- Leeson, S. and Summers, J.D. (1991). Commercial Poultry Nutrition. University books, Guelph, Ontario, Canada.
- Leeson S.; Caston L. J.; Yungblat D. (1996). Adding roenzyme to wheat diets of chickens and turkey broilers. Journal of Applied Poultry Research, vol. 5, 167-172. [CrossRef]
- Li, Q.; Yi, L.; Marek, P.; Iverson, B.L. (2013). Commercial proteases: Present and future. FEBS Letters 587, 1155–1163. [CrossRef]
- Li, S. Y. (1999). Studies on the effect of heat stress on performance and meat quality of broilers and anti-stress effect of riboflavin. Ph. D. Thesis, Chinese Academy of Agricultural Sciences, Beijing.
- Lin, H., E. Decuypere and J. Buyse. (2006). Acute heat stress induces oxidative stress in broiler chickens. Comparative Biochemistry and Physiology, Part A. 144:11-17. [CrossRef]
- Lin, H., H. F. Zhang, H. C. Jiao, T. Zhao, S. J. Sui, X. H. Gu, Z. Y. Zhang, J. Buyse and E. Decuypere. (2005a). The thermoregulation response of broiler chickens to humidity at different ambient temperatures I. One-week-age. Poultry Science 84:1166-1172.
- Lin, H., H. F. Zhang, R. Du, X. H. Gu, Z. Y. Zhang, J. Buyse and E. Decuypere. (2005b). The thermoregulation response of broiler chickens to humidity at different ambient temperatures I. Fourweek- age. Poultry Science 84:1173-1178.
- Lu, Q. Wen, J. Zhang, H. (2007). Effect of chronic heat exposure on fat deposition and meat quality in two genetic types of chicken. Poultry Science 86, 1059–1064.
- MacLeod, M.G. (2004). Climate-Nutrition Interactions in Poultry.1rst Annual Confrence , FVM., Moshtohor. Roslin Institute (Edinburgh), Scotland EH25 9PS 1-21.
- Mashaly, M. M., Hendricks, G. L., Kalama, M. A., Gehad A. E., Abbas, A. O., and Patterson, P. H. (2004). Effect of Heat Stress on Production Parameters and Immune Responses of Commercial Laying Hens1. Poultry Science 83:889–894. [CrossRef]
- McCormick D. B., Zhang Z. (1993). Cellular assimilation of water soluble vitamin in mammal: riboflavin, B6, biotin and C. Proceedings of the Society for Experimental Biologic and Medicine 202:265-70.
- McCormick DB. The fate of riboflavin in the mammal. Nutrition Reviews 1972;30:75–9. [CrossRef]
- McCracken, K. G., Paton, D. C., and Afton, A. D. (2000). Sexual size dimorphism of the musk duck. Wilson Bulletin, vol. 112, no. 4, p. 457-466. [CrossRef]
- McDaniel, C. D., Bramwell, R.K., Wilson, J. L., and Howarth, B. Jr. (1995). Fertility of male and female broiler breeders following exposure to elevated ambient temperatures. Poultry Science 74:1029–1038. [CrossRef]
- McDaniel, C.D.; Bramwell, R.K.; Wilson, J.L.; and Howarth, B. (1995). Fertility of male and female broiler breeders following exposure to an elevated environmental temperature. Poultry Science 74, 1029–1038. [CrossRef]
- McDaniel, C.D.; Hood, J.E.; and Parker, H.M. (2004). An attempt at alleviating heat stress infertility in male broiler breeder chickens with dietary ascorbic acid. International Journal of Poultry Science 3, 593–602.
- McKee, S. R., and Sams, A. R. (1997). The Effect of Seasonal Heat Stress on Rigor Development and the Incidence of Pale, Exudative Turkey Meat. Poultry Science 76:1616–1620. [CrossRef]
- Miles, R.D. (1999). Understanding heat stress in poultry and strategies to improve production through good management and maintaining nutrient and energy intake. Nutrition and Management Department of Dairy and Poultry Sciences University of Florida Gainesville. pp. 1-20.
- Mitchell M. A., Kettlewell P. J. (1998). Physiological stress and welfare of broiler chickens in transit: Solutions not problems! Poultry Science 77: 1803-1814.
- Mittal, J. P. and Ghosh, P. K. (1979). Body temperature, respiration and pulse rate in Corriedale, Marwari and Magra Sheep in the Rajasthan desert. Journal of Agricultural Science (Cambridge), 95: 587-591. [CrossRef]
- Monsi, A. (1992). Appraisal of interrelationships among live measurements at different ages in meat type chickens. Nigerian Journal of Anim.al Production, vol. 19, no.1&2, p.15- 24.
- Mótyán, J.A; Tóth, F.,and Tőzsér, J. (2013). Research Applications of Proteolytic Enzymes in Molecular Biology Biomolecules, 3, 923-942. [CrossRef]
- Muiruri, H. K., and Harrison, P. C. (1991). Effect of roost temperature on performance of chickens in hot ambient environments. Poultry Science. 70:2253–2258. [CrossRef]
- Mujahid, A., Y. Yoshiki, Y. Akiba and M. Toyomizu, (2005). Superoxide radical production in chicken skeletal muscle induced by acute heat stress. Poultry Science, 84: 307-314. Nardone, A.; Ronchi, B.; Lacetera, N.; Ranieri, M. S.; and Bernabucci, U. (2010). Effects of climate changes on animal production and sustainability of livestock systems. Livestock Science 130, 57–69.
- Neurath, H.; Walsh, K.A. (1976). Role of proteolytic enzymes in biological regulation (a review). Proceedings of National Academy of Science, USA, 73, 3825–3832. [CrossRef]
- Nicol, C.J., Saville-Weeks C. (1993). Poultry handling and transport. In: Livestock handling and transport. GRANDINT (ed) Wallingford Oxon UK: CAB International pp 273-287.
- Nienaber, J. A., and Hahn, G. L. (2007). Livestock production system management responses to thermal challenges. International Journal of Biometeorology. 52, 149–157. [CrossRef]
- North, M.O., (1984). Commercial Chicken Production Manual: Poultry Housing. AVI Publishing Company, Inc. Westport, Connecticut. pp. 148-177.
- Northcutt, J. K., E. A. Foegeding and F. W. Edens. (1994). Waterholding properties of thermally preconditiond chicken breast and leg meat. Poultry Science 73:308-316. [CrossRef]
- Ogah, D. M. (2011). Assessing Size and Conformation of the Body of Nigerian Indigenous Turkey. Slovak Journal of Animal Science, 44 (1): 21-27.
- Ogah, D.M., Musa, I. S., Yakubu, A., Momoh, M.O. and Dim, N. I. (2009). Variation in morphological traits of geographical separated population of indigenous muscovy duck (Cairina moschata) in Nigeria. Proceeding of 5th International Poultry Conference Taba Egypt, p. 46-52.
- Ohkawa H. Ohishi N. Yagi K.A. (1982). Simple method for microdetermination of flavin in human serum and whole blood by high performance liquid chromatography. Biochemistry International 4:187 94.
- Osman, A. M. A., E. S. Tawfik, F. W. Klein and W. Hebeler. (1989). Effect of environmental temperature on growth, carcass traits and meat quality of broilers of both sexes and different ages. 1. Growth. Archiv fur Geflugelkunde 53:168-175.
- Park, S.O., Hwangbo, J., Ryu, C.M., Park, B.S., Chae, H.S., Choi, H.C., Kang, H.K., Seo, O.S., and Choi, Y.H. (2013). Effects of extreme heat stress on growth performance, lymphoid organ, IgG and cecum microflora of broiler chickens. International Journal of Agricultural Biology, 15: 1204–1208.
- Powers, H. J. (2003). Riboflavin (vitamin B2) and health. American Journal of Clinical Nutrition 77:1352–60.
- Ramnath, V., Rekha, P. S., and Sujatha, K. S. (2008). Amelioration of heat stress induced disturbances of antioxidant defense system in chicken by Brahma Rasayana. Evidence- Based Complementary and Alternative Medecine 5 (Suppl. I), 77-84. [CrossRef]
- Rani, K.; Rana, R.; Datt, S. (2012). Review on latest overview of proteases. International Journal Current Life Science 2, 12–18.
- Rao, M. B., Tanksale, A. M., Ghatge, M. S. and Deshpande, V. V. (1998). Molecular and biotechnological aspects of microbial proteases. Microbiology and Molecular Biology Reviews, 62, 597-635. [CrossRef]
- Rao, M.B.; Tanksale, A.M.; and Ghatge, M.S.; Deshpande, V.V. (1998). Molecular and biotechnological aspects of microbial proteases. Microbiology and Molecular Biology Reviews 62, 597–635. [CrossRef]
- Rao, M.B.; Tanksale, A.M.; Ghatge, M.S.; Deshpande, V.V. (1998). Molecular and biotechnological aspects of microbial proteases. Microbiology and Molecular Biology Reviews, 62, 597–635. [CrossRef]
- Rawlings, N.D.; and Barrett, A.J. (1993). Evolutionary families of peptidases. Biochemical Journal 290, 205–218. [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; and Bateman, A. (2012). MEROPS: The database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Research 40, D343–D350. [CrossRef]
- Ray, A. (2012). Protease enzyme- potential industrial scope. International Journal Technology 2, 1–4.
- Renaudeau, D.; Collin, A.; Yahav, S.; de Basilio, V.; Gourdine, J. L.; and Collier, R.J. (2012). Adaptation to hot climate and strategies to alleviate heat stress in livestock production. Animal, 6, 707–728. [CrossRef]
- Rozenboim, I., Tako, E., Gal-Garber, O., Proudman, J. A and Uni, Z. (2007). The effect of heat stress on ovarian function of laying hens Poultry Science 86:1760–1765.
- Sahin, K. and O. Kucuk, (2003). Heat stress and dietary vitamin supplementation of poultry diets. Nutr. Abstr. Rev. Ser. B. Livestock Feeds Feeding, 73: 41R-50R. [CrossRef]
- Sahin, K., and Kucuk O. (2001). Effects of vitamin C and vitamin E on performance, digestion of nutrients and carcass characteristics of Japanese quails reared under chronic heat stress (34 °C). Journal of Animal Physiology and Animal Nutrition 85, 335-341.
- Sahin, K., N. Sahin, M. Onderci, S. Yaralioglu and O.Kucuk, (2001). Protective role of supplemental vitamin E on lipid peroxidation, vitamins E, A and some mineral concentrations of broilers reared under heat stress. Veterinary Medicine, Czech Republic, 46: 140-144. [CrossRef]
- Sandercock, D. A., R. R. Hunter, G. R. Nute, M. A. Mitchell and P. M. Hocking. (2001). Acute heat stress-induced alterations in blood acid/base status and skeletal muscle membrane integrity in broiler chickens at two ages: implications for meat quality. PoultryScience 80:418-425. [CrossRef]
- Scott, N.R., DeShazer, J.A., and Roller, W.L. (1983). Effect of thermal and gaseous environment on livestock. In: Ventilation of Agricultural Structures. EdsHellickson, M. A. and Walker, J.N., The American Society of Agricultural Engineers, 2950 Niles Road St. Joseph, Michigan 49085- 9659, USA. pp.121-165.
- Selye, H. (1976). Forty years of stress research: principal remaining problems and misconceptions.
- Seskevicience, J.; Jeroch, H.; Danieke, S.; Gruzauskas, R.; Volker, L.; Broz, J. (1999). Feeding value of wheat and wheat-based diets with different content of soluble pentosans when fed to broiler chickens without or with enzyme supplementation. Archive Furgeflugelkunde, vol. 63, pp. 129-132. [CrossRef]
- Shafee, N., Aris, S. N., Rahman, R. Z. A., Basri, M. and Salleh, A. B. (2005). Optimization of Environmental and Nutritional Conditions for the Production of Alkaline Protease by a Newly Isolated Bacterium Bacillus cereus Strain 146. Journal of Applied Sciences Research, 1, 1-8.
- Shahin, K.A. (1996). Analysis of muscle and bone weight variation in an Egypt strain of Peking ducklings. Annual Zootechnology, vol. 45, no. 2, p. 173-184.
- Singer, T. P.; Kenny, W. C. (1974). Biochemistry of covalently-bound flavins. Vitamins and Hormones, 32:1-45.
- Smits, C. H. M.; Annison G. (1996). Non-starch plant polysaccharide in broiler nutrition towards a physiologically valid approach to their determination. World’s Poultry Science Journal, vol. 52, pp. 203-206. [CrossRef]
- Sumantha, A., Sandhya, C., Szakacs, G., Soccol, C.R. and Pandey, A. (2005). Production and partial purification of a neutral metalloprotease by fungal mixed substrate fermentation. Food Technology and Biotechnology, 43, 313-319.
- Vecerek V., Grbalova S., Voslarova E., Janackova B., and Melena M. (2006). Effects of travel distance and the season of the year on death rates of broilers transported to poultry processing plants. Poultry Science 85: 1881-1884. [CrossRef]
- White HB III, Merrill AH Jr. Riboflavin-binding proteins. Annual Review of Nutrition 1988;8: 279–99.
- Whitehead, C. C., Bollengier-Lee, S., Mitchell, M. A., and Williams, P. E.V. (1998). Alleviation of depression in egg production in heat stressed laying hens by vitamin E. In Proceedings of 10th European Poultry Conference, Jerusalem, Israel. Pages 576– 578.
- Yakubu, A., Ogah, D. M., and Idahor, K. O. (2009). Principal component analysis of the morphostructural indices of White Fulani cattle. Trakia Journal Science, vol. 7, p. 67- 73.
- Zanette D, Monaco HL, Zanotti G, Spadon P. (1984). Crystallisation of hen egg white riboflavin-binding protein. Journal of Molecular Biology 180:1185–7.
- Zhang, Z.Y.; Jia, G.Q.; Zuo, J.J.; Zhang, Y.; Lei, J.; Ren, L.; Feng, D.Y. (2012). Effects of constant and cyclic heat stress on muscle metabolism and meat quality of broiler breast fillet and thigh meat. Poultry Science 91, 2931–2937. [CrossRef]
Figure 1.
Bar chart showing ambient temperature per week.
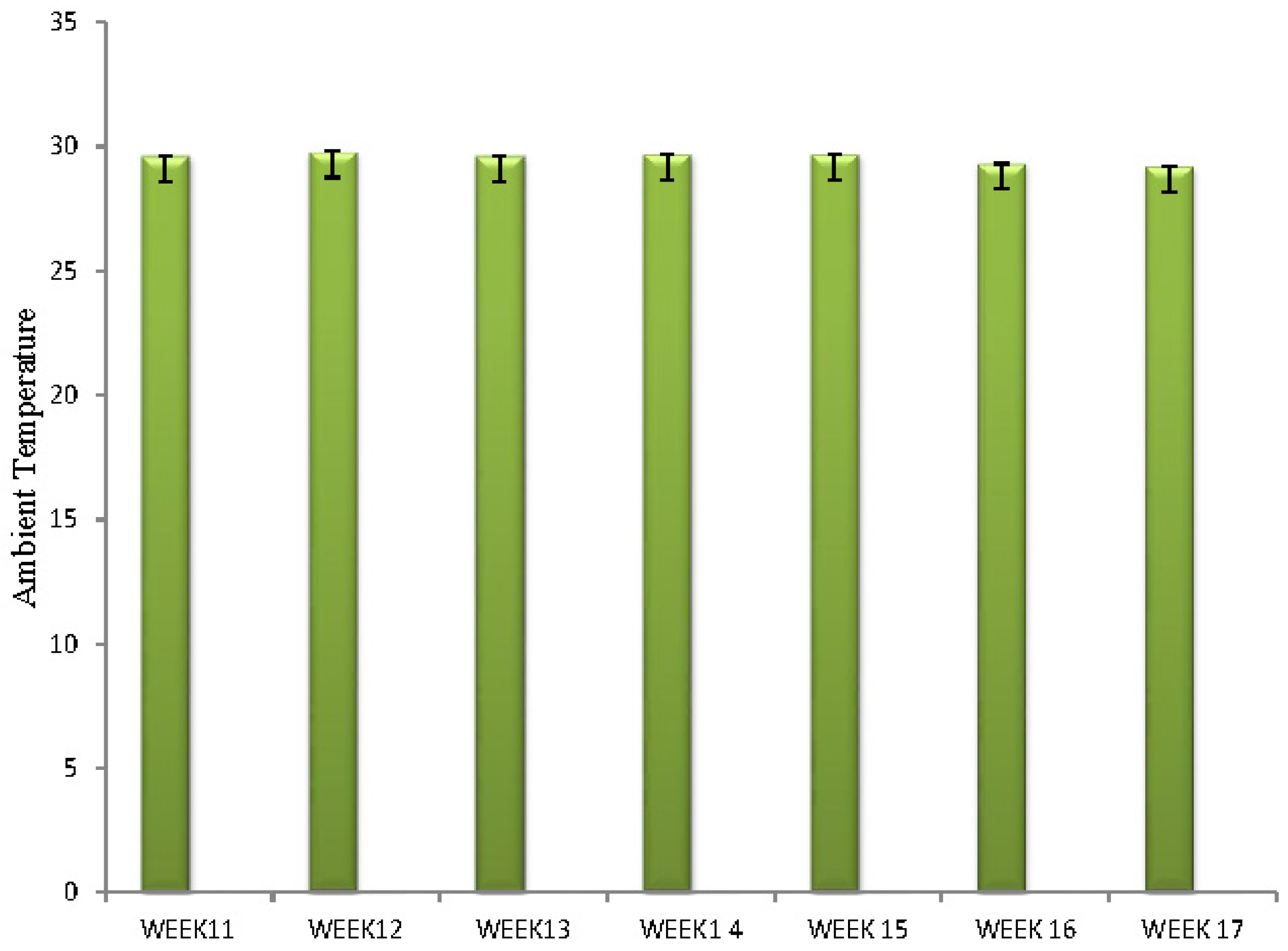
Figure 2.
Bar chart showing relative humidity per week.
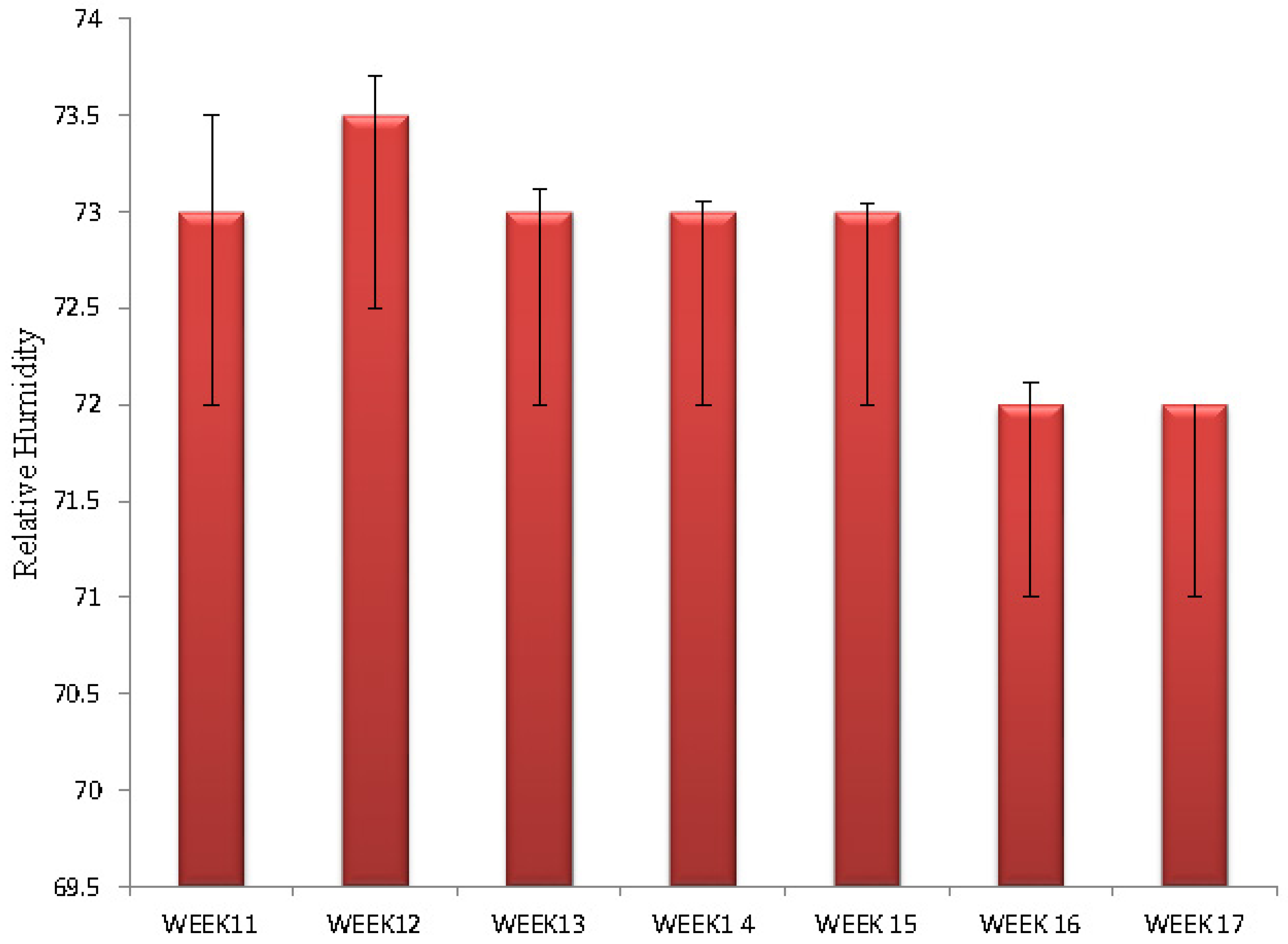
Table 5.
Effect of riboflavin and protease enzyme on rectal temperature of finishing turkeys during hot-dry season.
Table 5.
Effect of riboflavin and protease enzyme on rectal temperature of finishing turkeys during hot-dry season.
| Age (weeks) | Control | Basal diet with riboflavin (6mg/kg) | Basal diet with protease enzyme (1000mg/kg) |
Basal diet with riboflavin and protease enzyme (6mg/kg+1000mg /kg) |
| 11 | 40.931 ± 0.099 | 41.081 ± 0.099 | 40.975 ± 0.099 | 40.875 ± 0.099 |
| 12 | 41.131 ± 0.099 | 40.919 ± 0.099 | 40.794 ± 0.099 | 40.869 ± 0.099 |
| 13 | 40.931 ± 0.099 | 40.938 ± 0.099 | 40.950 ± 0.099 | 40.788 ± 0.099 |
| 14 | 40.969 ± 0.099 | 41.113 ± 0.099 | 40.931 ± 0.099 | 41.063 ± 0.099 |
| 15 | 40.925 ± 0.099 | 40.875 ± 0.099 | 40.919 ± 0.099 | 41.119 ± 0.099 |
| 16 | 40.525 ± 0.099 | 40.594 ± 0.099 | 40.644 ± 0.099 | 40.588 ± 0.099 |
| 17 | 40.350 ± 0.099 | 40.431 ± 0.099 | 40.419 ± 0.099 | 40.425 ± 0.099 |
Table 6.
Analysis of variance showing effect of riboflavin and protease enzyme on rectal temperature during hot-dry season.
Table 6.
Analysis of variance showing effect of riboflavin and protease enzyme on rectal temperature during hot-dry season.
| Source | Degree of Freedom | Mean-square | * |
| Treatment | 3 | 0.854 | NS |
| Week | 6 | 0.000 | *** |
| Treatmeat * Week | 18 | 0.552 | NS |
| Error | 420 |
*** P<0.000, NS: Not Significant.
Table 7.
Effect of riboflavin and protease enzyme on pulse rate of turkeys during hot-dry season.
| Age (week) | Control | Diet with riboflavin (6mg/kg) |
Diet with protease enzyme (1000mg/kg) |
Diet with riboflavin and protease enzyme (6mg/kg+100 0mg/kg) |
| 11 | 167.50ab±1.708 | 162.500bc±1.708 | 164.750b±1.708 | 171.250defg±1. 708 |
| 12 | 153.25defg±1.708 | 157.750cd±1.708 | 157.500cde±1.708 | 156.000defg±1. 708 |
| 13 | 154.50defg±1.708 | 151.000fg±1.708 | 156.000defg±1.708 | 156.000defg±1. 708 |
| 14 | 156.75def±1.708 | 157.250de±1.708 | 155.750defg±1.708 | 153.500defg±1. 708 |
| 15 | 155.000defg±1.708 | 156.313def±1.70 8 |
155.000efg±1.708 | 154.500defg±1. 708 |
| 16 | 154.250defg±1.708 | 152.750defg±1.70 8 |
151.750g±1.708 | 155.500defg±1. 708 |
| 17 | 154.750defg±1.708 | 154.500defg±1.70 8 |
150.250a±1.708 | 153.500defg±1. 708 |
Table 8.
Analysis of variance showing effect of riboflavin and protease enzyme on pulse rate of turkeys during hot-dry Season.
Table 8.
Analysis of variance showing effect of riboflavin and protease enzyme on pulse rate of turkeys during hot-dry Season.
| Source | Degree of Freedom | Standard Error of Mean |
| Treatment | 3 | 40.437 NS |
| Week | 6 | 1343.336 *** |
| Treatment * week | 18 | 85.062 * |
| Error | 420 | 46.703 |
*** P<0.001, * P<0.05 and NS: Not Signicant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



