
Submitted:
28 November 2024
Posted:
28 November 2024
You are already at the latest version
Abstract
Slaughter weight and feed efficiency are highly dependent on the length of post-hatch feed deprivation, due to the short fattening time. Late feed access has negative consequences on the development of the intestine, while threonine might be a candidate to support epithelial tissue development. The present study aimed to examine the effects of different early nutrition methods, particularly early Thr supplementation on the performance, gut and liver histomorphology, and the number of different leukocyte types in broiler chicken. Control birds were fed immediately post-hatch, while others had access to solid feed with a 48-hour delay. In one group in ovo threonine, administration was applied at d17 of incubation, while other groups of birds received hydrogel with or without Thr fortification in the first 48h. Our results confirmed that the best performance was achieved in birds that had no delay in feed access. Early Thr supplementation could at least partly compensate for the loss attributed to the post-hatch delay in feed access. The higher ability of compensation can be explained by the better architecture of the gut tissue, while there were no significant effects of Thr supplementation on the rate of leukocytes and liver histology parameters.

Keywords:
broilers
; threonine
; in ovo nutrition
; hydrogel
; compensation
; performance
; leukocytes
; gut morphology
1. Introduction
Fast-growing broilers must have a continuous and well-managed nutrient supply to realize the genetically determined slaughter weight at the target age. The nutritional status of the embryo is highly dependent on the nutritional status of the hen, as maternal nutrition can affect early development and the metabolism of the progeny through gene modification [1]. In addition, breed and dietary factors strongly affect egg quality and nutritional profile [2]. The hatching egg contains essential nutrients for embryo development, but the length of the hatching window as well as the time to the first feed post-hatch determines slaughter weight. It has been confirmed that in intensive broilers even 24-hour-delay in feed access post-hatch reduces the body weight by 2-3%, while a longer time of feed deprivation (48-72 hours) results in 5-8% less slaughter weight at 42 days of age (de Jong e al., 2017). During the early period of life, the embryo relies on the antibodies and nutrients of the albumen, however, the dynamic changes of the egg yolk from the continuous metabolism can influence the development of the chick [3]. In addition, other stressors such as limited nutrients in the final period of incubation [4] as well as wide hatching window (12-24h), and delayed first feeding [5] can also harm the immune functions and vitality of broilers. Thus, mortality in the first week is 1.5-2.2 times higher if the access to the first feed is delayed by 48-72 hours compared to birds provided solid feeds within 24-36 hours post-hatch [3].
To avoid these outcomes, different early feeding methods have been developed such as offering hydrogel enriched with various nutrients [6], ensuring immediate feed access post-hatch (Patio system; [7]), or providing nutrients in ovo [8]. Vitamins, probiotics, and amino acids can be supplemented as a complementary feed in a gel form by spraying the chicks in the incubator [6] so consuming the gel is carried out by pecking each other. Another option for early nutrition of hydrated gels is applying them in the chick box during transport. The benefits of jelly masses are avoiding dehydration by prompt water consumption [9] and providing probiotics to boost intestinal microbiota and compensate for environmental stressors. It has been well documented that immediate feed access resulted in higher final weight, better breast meat-to-carcass ratio [10] rapid intestinal development [11], greater duodenal villi surface, favorable gut microbiota, and prevention from pathogenic infections by boosting the immune system [12,13]. However, in practice, immediate post-hatch feeding is difficult to proceed. In ovo intervention was first applied in vaccination research in the late '80s [14], and the technique was used later to provide nutrients, particularly carbohydrates [9]. Over the years, several experiments were carried out to determine the optimal timing and site of injection by introducing exogenous substances to the air sac, yolk sac, and amniotic fluid [15,16,17]. It turned out, that by consumption of the amniotic fluid proteins, minerals, and hormones can be supplied to the late-term embryo, therefore in ovo nutrient provision is a reliable non-invasive method to boost immature digestive systems around the first days of life [16]. Improving chick immunity is also possible by adding vitamins, prebiotics, or amino acids before hatching [18, 19]. From the immunomodulatory point of view, amino acids such as methionine (Met), arginine (Arg), and threonine (Thr) contribute to lymphocyte proliferation and enroll monocytes and heterophils from the bone marrow [20, 21, 22].
According to Sirsat et al [23], modern broiler strains require reevaluation of their nutrient provisions, and specific early-phase nutrition must be applied, particularly, in the pre-hatch and the transitional post-hatch period. Specific feeding strategy that improves the digestive efficiency and/or resilience of birds increases the economics of poultry production. The individual amino acids have a prominent role in this respect. Threonine supplementation may improve the morphological characteristics of the intestine, thereby providing a better and larger absorption surface. This supports the supply of nutrients, which on one hand can result in better growth performance and on the other hand may improve feed efficiency. A better slaughter yield can also accompany a higher live weight if the proportion of valuable meat parts is improved.
Hence, the present study aimed to examine the effects of different early nutrition methods, particularly early Thr supplementation on the performance, gut histomorphology, liver histology, and differentiated leukocyte number of broiler chicken.
2. Materials and Methods
2.1 Hatching protocol and treatment groups
The experiment was carried out at the Hungarian University of Agricultural and Life Sciences (MATE) Kaposvár Campus, Department of Farm Animal Nutrition by the Declaration of the Hungarian National Scientific Ethical Committee of Animal Experimentation for studies involving animals, protocol license number is SO/31/00444-2/2021.
A total of 1120 Ross 308 broiler eggs were involved in the study. Prior to hatch, the eggs were held in transport boxes at 17-18oC for 6 days without rotation or extra humidification due to the short storage time. A PLM B1350 two-staged incubator was used for the hatching process with 9 tray levels. Each level was equipped with a built-in measurement system for ventilation, humidity, and temperature between the levels. The hatching protocol was carried out according to the recommendations of the Aviagen Hatching Management Guide (2019): the dry bulb temperature and humidity were set at 37.9 ± 0.1°C and 65 ± 3%, respectively (Table 1). The eggs were candled on days 10 and 17 to exclude infertile eggs or dead embryos. The fertility of eggs was calculated after candling by the following formula:
Fertility rate % = number of fertile eggs/total number of eggs set [25].
At the beginning of the study, the eggs were assigned to 7 experimental groups. In the two control treatments, chickens were fed immediately after hatch (Int_0 and IoS_0), while chickens emerged from the rest of the eggs and received solid feed only 48 hours post-hatch. Two groups of eggs were injected with 0.5 mL of physiological saline (0.9 g/mL concentration of NaCl), either from the immediate (IoS_0) or from the delayed fed groups (IoS_48). This intervention was needed to evaluate the possible stress caused by the needle puncture. Another group was treated with in ovo Thr using 0.5% concentration solution dissolved in physiological saline and the birds had 48 hours of feed deprivation post-hatch (IoT_48). There was no intervention in four groups of eggs, but the hatched chicks in those treatments had different feeding management. Chicken in group Int_0 and Int_48 had immediate or delayed feed access, Int_48G and Int_48GT had no solid feed in the first 48 hours but immediate access to Hydrogel® (Bábolna Feed Ltd.) without Threonine or with 5 g/kg Thr enrichment, respectively, in the transport boxes. The basal Hydrogel® composed of corn starch (30%), probiotic lactic acid bacteria (Pediococcus acidilactici (E1712) 1 x1010 CFU/g), and vitamin C (5000 mg/kg) and contained 5,4 MJ AMEn/kg. The rationale of the experimental treatments is shown in Table 2.
2.2 In ovo intervention
The in-ovo injection was carried out following the protocol of Uni and Ferket [24] with a 2 mL syringe and a 21-gauge needle. The injection procedure was performed in a sterile cabinet (ScanLaf, LaboGen Inc., Denmark) to prevent any microbiological contamination. Before the intervention all eggs were cleaned with cotton wool dipped in an iodine solution. The eggs were then carefully drilled on the blunt side through the air chamber without reaching the shell membrane. Before the intervention, the position of the embryo was checked, and a 0.5 ml solution (either 0.9% (w/V) NaCl solution or 0.5% Thr solved in 0.9% (w/V) NaCl solution) was transferred to the amniotic fluid without injuring the embryo afterward. To avoid entry of pathogens, a sterile, plastic tape was applied, and the eggs were placed back into the incubator until day 21 of hatching.
2.3 Identification, feeding management, and housing
The birds were weighted with gram precision at hatching day right after harvesting, and wing tagged for individual identification. Birds in treatment Int_0 and IoS_0 were immediately placed in the barn where a mash starter feed was offered ad libitum. The rest of the birds were stored in a transporter paper box (25 birds/box) and kept at 32ᵒC for 48 hours in a separate room. Birds assigned to Int_48, InS_48, and IoT_48 received no feed supplementation, while in Int_G48 and Int_GT48 groups the birds had access to Hydrogel with or without Thr fortification. On day 2 all birds were weighed with gram precision and placed in the barn where the starter feed was freely available.
The broilers were randomly placed into floor pens (16 birds/pen, 8 pen/treatment). Each pen represented an individual treatment group and was equipped with one feeder and one drinker. The housing management was set in compliance with Aviagen's (2019) recommendation for temperature, humidity, hours, and intensity of light.
A three-phase feeding program was applied as follows: between days 1 and 10 starter feed (mashed feed), between days 11 and 21 pelleted grower feed, and between days 22 and 35 pelleted finisher feed produced by the Department of Farm Animal Nutrition was offered to all birds. Each feed was formulated on a corn-soybean meal basis. Nutritional content (dry matter, crude protein, fat, ash, calcium, and phosphorus) was determined by the University Lab Center of MATE according to the recommendations of the Association of Official Analytical Chemists (AOAC, [25]).
The birds were fed ad libitum from self-feeders during the trial. One feeder was presented per pen. Drinking water was also available ad libitum. The analyzed composition of the feed is presented in Table 3.
2.4 Experimental procedure
The live weight of the birds was measured at hatch (d1), on days 3, 10, 21, and 35. The average daily gain (ADG) was calculated individually, while feed intake (FI) was recorded per pen for the time intervals by measuring the offered and remaining feed for each phase. The feed conversion ratio (FCR) was also calculated per pen basis.
At hatch, 48 hours later, and at d21 of the experiment, 10 birds from each treatment were used to collect blood, liver, and intestinal samples. For that purpose, the birds were slaughtered following carbon dioxide stunning in compliance with the relevant legal regulations (Council Regulation 1099/2009/EC). In the case of day-old and 3-day-old chicks, capillary blood samples were collected from the dorsal metatarsal vein and direct blood smears were prepared immediately from 10 birds/treatment. Similarly, to day-old chickens, the three-week-old birds were sampled for bloodwork, but this time blood samples were collected from the jugular vein into 5 ml tubes containing EDTA for further evaluation. Blood smears were prepared immediately after the collection process.
To determine gut morphology through villus height (VH), crypt depth (CD), and villus/crypt ratio (VH/CD) tissue samples were collected from duodenum, ileum, and colon (10 birds/treatment). Liver samples were also collected, and all samples were placed into a 10% neutral buffered formalin solution and processed in an Epredia™ Citadel 2000 Tissue Processor (Thermo Fischer Scientific, Waltham, MA, USA) to make paraffin-embedded blocks. Two-micrometer-thick paraffin sections were obtained in a rotary microtome unit (HistoCore BIOCUT, Leica Biosystems, Nussloch, Germany). These sections were de-waxed, rehydrated, and transferred to glass slides, and hematoxylin-eosin staining was performed according to standard histological techniques [26]. Each sample from each broiler was cross-sectioned three times, and three measurements were carried out on each section. Villus sizes were measured at 400X magnification [27]. The measurement method is demonstrated in Figure 1.
Histological injuries on the liver were classified as none, mild, moderate, or marked lesions according to their severity with scores from 0 to 3.
2.4. Statistical analysis
Data were analyzed according to a completely randomized block design with 7 treatments. A Shapiro–Wilk normality test was carried out on the base data. The outliers were checked and excluded from the statistical analysis. Outliers were defined as values being more than 2 times of standard deviation away from the mean, Levene’s test was used to examine group homogeneity among treatment groups. A one-way ANOVA was applied to performance data and other results considering the effect of treatment as the main factor of variation in body weight at a different age, feed intake (FI), feed conversion ratio (FCR), and the number of white blood cells, as well as morphometry parameters. In case of statistically significant difference (P<0.05) Tukey’s post-hoc test was applied to check the differences between treatment groups. Frequency distributions were calculated for the histological injuries of liver tissue samples along with a Kurksal-Wallis test to the ordinal scale of the injuries.
3. Results
3.1. Growth performance and feed efficiency
Hatchability (hatched chicken/total number of eggs) was in the range of 0.74-0.79 in eggs having no intervention (intact eggs), and 0.77-0.78 in groups applied physiological saline solution and 0.81 when in ovo threonine supplementation was applied at day 17 of the incubation. The live weight of birds in different experimental treatments at hatch and 48 hours later is shown in Figure 2. The ANOVA confirmed a significant difference in hatching weight, as birds in one of in ovo saline-treated group assigned to IoS_48 group was heavier than all others (P<0.05). Live weight on the 3rd day of the trial was much higher in birds fed immediately post-hatch (Int_0 and IoS_0 vs. other groups).
Bars represent treatment means and SDa, b different letters indicate a significant difference (P<0.05)
The effect of dietary treatments including early threonine supplementation on LW, FI, FCR, and ADG of broilers is shown in Table 4 and Table 5. Birds with immediate feed access reached 240 g on average by day 10, while birds with a 48-hour delay in access to the first solid feed had more than 20% lower body weight (194 g on average). At the end of the starter phase, eggs supplemented with in ovo saline (IoS_0) had higher LW, than the intact group (Int_0), while treatments with 48 hours of delayed feeding did not differ from each other. LW at day 21 was the highest in birds fed immediately post-hatch and differed significantly from the different groups (Int_0 and IoS_0 vs others, P<0.05). The lowest LW was recorded in the Int_48 group, and from the delayed groups the in ovo Thr-treated birds had the highest body weight at the end of the grower phase. At the end of the trial, on day 35, the immediately-fed birds still had the highest LW. Compared to the immediate fed counterparts there was no statistically confirmed difference between the LW of the delayed and in-ovo Thr or Hydrogel ® supplemented birds. Group IoT_48 significantly did not differ either from the IoS_48 and Int_48GT or from the Int_48 which had the lowest final LW.
The FI of the birds showed significant differences in all of the examined time intervals, however, the advantage of groups Int_0 and IoS_0 remained constant. Statistics confirmed a significant treatment effect on FCR in the starter, grower, and finisher phases. In ovo Thr-supplemented birds had more efficient feed utilization in the starter phase than birds who received their diet immediately post-hatch and were treated with in ovo saline (IoS_0, P=0.04). The average daily gain was significantly affected by dietary treatments in all phases and the whole trial. In the starter phase, the 48-hour delay in FI resulted in a lower growth rate (Int_0 and IoS_0 vs others, P<0.05). In the grower and finisher phases, the advantage of the immediate fed birds was obvious, while supplementation of in ovo Thr and the hydrogel with or without Thr enrichment could compensate at least partly the loss caused by the late feed access.
3.2. Intestinal morphometry
Table 6, Table 7 and Table 8 present the gut morphology results affected by dietary treatments at hatch, 48 hours post-hatch, and 21 days of age, respectively.
The intestinal morphological results on villus height measured at hatch (Table 6) show a significant difference in the duodenum comparing IoS_0 vs Int_48 groups, in the ileum comparing Int_0 and IoS_0 vs. IoS_48 and Int_G48 groups, and between IoS_48 and IoT_48 vs. Int_GT48 in the colon region. Crypt depth was affected in the duodenum and the colon. IoS_48 group had the lowest crypt depth as it differed significantly from Int_0, IoS_0, Int_48, and IoT_48 groups in the duodenum. In the architecture of the colon, birds assigned to in ovo saline (IoS_0 and IoS_48) and Int_GT48 group showed the lowest crypt depth, while that of birds in Int_48 and IoT_48 treatments had the highest.
The villus height was affected by treatments in two examined intestinal sections on day 3 (48 hours post-hatch) (Table 7). It was the highest in the Int_0 group and the lowest in the IoS_0 group both in the duodenum and colon. Crypt depth was statistically the same in all groups. VH/CD ratio was affected only in the colon, showing the lowest rate in the Int_48 group significantly differing from that of birds in Int_0, IoS_0, Int_G48, and Int_GT48.
Significant differences were found on day 21 regarding villus height in the duodenum and the ileum, but not in the colon (Table 8). A statistically confirmed difference was obtained in duodenal villus height between Int_48 and Int_G48 birds, while all others did not differ. The ileal villi were the longest in birds assigned to IoT_48 treatments, and differed from almost all other groups, except for birds in IoS_0. Crypt depth was not affected by dietary treatments. Villus height/crypt depth ratio was different between InT_0 and Int_G48, and between IoT and Int_GT48, in the duodenum and ileum, respectively.
3.3. Liver histology and blood cell results
Results of the liver histology demonstrate that most of the alterations regarding the heterophil granulocyte infiltration within the periportal regions occurred right after hatch, and remained constant by day 21. Interestingly it showed the worst results in the in ovo treated groups by Day 3. From day 3 another pathological change, mononuclear infiltration appeared in the liver, mostly in the periportal areas, but sometimes as multifocal lesions scattered in the parenchyma. This lymphohistiocytic invasion increased and gradually became predominant by day 21 in almost all groups. In addition, hepatocytes showed moderate to marked lipid accumulation in every section. This hepatic lipidosis gradually reduced from day 1 to day 21, and completely disappeared in most of the groups. Only in groups Int_G48 and Int_GT48, a mild lipidosis had been detected on day 21, and it remained as a faint vacuolization in group IoS_48 by this time.
Due to some technical failure determination of leukocytes’ ratio was not successful in Int_0 birds and the data of that treatment was discarded from statistical analysis. With blood smear evaluation moderate lymphocyte depletion and a mild increase in monocyte numbers were detected in all groups and ages (Table 10). Heterophil and eosinophil granulocyte numbers did not show remarkable alterations to dietary treatments (P>0.05).
4. Discussion
Our study aimed to evaluate the efficiency of different early feeding methods by revealing some physiological changes that might be induced by post-hatch feed deprivation. It has been investigated whether the provided early threonine supplementation either before or after hatch could alleviate the negative impact of delay in solid feed access. In ovo feeding was first reported several decades ago, and it is still not a common practice in hatcheries. However, an increasing number of studies evaluate the effect of specific nutrient supplementation of the poultry embryo on the growth performance of meat-type poultry and reveal the mode of action of the supplementation.
In the present study in ovo intervention did not reduce the hatchability of the eggs. Some studies reported a lower hatchability rate when in ovo feeding [28, 29,30], however, it seems that not the intervention itself but the circumstances like the amount, concentration, and osmolarity of the injected solution, etc. were responsible for the lower hatch rate in different studies [31] Statistically confirmed difference was observed between hatching weight of IoS_48 and that of the other groups, however, that result is ambiguous. The in ovo saline solution was used as a control of the in ovo intervention per se. The treatments IoS_0 and IoS_48 can be considered as identical groups at the time of emergence, since in their case in ovo manipulation took place on day 17 of incubation, and at hatch, even the intake of feed did not cause any difference, since the Int_0 birds were settled after the weighing. Therefore, we can conclude that eggs in groups of IoS_0 and IoS_48 were treated equally. In agreement with our results, Kadam et al., [32] confirmed in a meta-analysis that the effect of physiological saline is not consequent and the treated birds do not always have higher weight at hatch. The beneficial effects of in-ovo feeding on hatching weight are majorly shown when carbohydrates are used as nutrient supplementation [8, 10, 33]. In line with other results [32; 34] threonine supplementation on day 17 of incubation did not shift hatching weight in the present study. It is well documented that birds at emergence need energy and thus, carbohydrates can be promptly used as a fuel to compensate for the energy deficit at hatch. In the case of an amino acid supplementation, the aim was not to support the energy status but to provide nutrients, as building blocks for specific proteins like epithelia or functional proteins involved in defense mechanisms. We hypothesized that early Thr supplementation provided either during late embryo development or in the post-hatch period might contribute to better gut tissue development, and resulted in a better growth rate and more efficient birds. That has been at least partly confirmed. Live weight and the average daily gain of broilers were supported by early threonine supplementation when the birds had a 48-hour delay to solid feed access. However, it has to be admitted that the performance may still be slightly compromised, and the feed deprivation in the first two days of life may be hardly compensated completely by either of the early methods of Thr supplementation. Immediate post-hatch feeding is an ideal situation, but it is hard to proceed in practice. Birds get their first feed with a 36-72-hour delay very often. Therefore, in our study, the immediately-fed group represents the genetic potential of the birds. In a recent work of [34] early (within 2 or 24 hours) post-hatch feeding compared to a 48-hour delay to the first feed, the growth performance of broilers improved, most likely due to an improved hormone secretion (T3, T4, and IGF-1), and also by enhancing the intestinal health and modulating the microbiota, especially at day 21. Results of de Jong et al. [3] also confirmed that even a 24-36-hour delay in post-hatch feed access reduces the body weight of chicks to a statistically verifiable extent. It has to be noted, however, that the longer the fattening period is (e.g. 50 days), the more ability the birds have to compensate for the early perturbation.
The provision of gel supplements is the most widely used in practice among early feeding methods. Several studies report the beneficial effects of hydrogels such as hydrating the day-old-birds or adding their probiotic benefits [35, 36]. Even though it is a high supplement with high moisture content, hydrogel could not compensate for the two-day feed deprivation, as the body weight of birds was lower on day 3 than on day 1 of the trial. This result emphasizes, that the energy supply from the yolk sac is not enough even for maintenance purposes in intensive genotypes. As reviewed by Al-Huwaizi [37], the yolk sac nutrients-particularly fatty acids- can be more efficiently utilized if birds receive solid feed post-hatch. Among delayed-fed birds the early access to hydrogel, particularly with Thr enrichment resulted in the highest body weight at the end of the experiment numerically, but not in the starter and grower phases.
In the first 3 weeks, the in ovo Thr supplementation provided in ovo improved the growth performance like ADG and FCR of birds compared to the non-supplemented counterparts. It has been repeatedly confirmed that in ovo supplementation of Thr alone or in combination with carbohydrates or other amino acids like arginine can improve the growth rate of broilers [38,39,40,41,42]. Those positive results were almost completely explained by the improvements of the morphology of the gut.
It is well-documented that dietary threonine has a key role in intestinal development from both structural and functional points of view [43, 44]. Thr -being an essential amino acid-, plays a vital role in the maintenance of intestinal barrier integrity and mucin synthesis (45, 46). It is the most abundant essential amino acid in the endogenous protein secreted in the intestine, particularly in the mucins in which Thr represents 16% of total amino acids [47]. In conventional feeding trials, it has been confirmed that insufficient threonine supplementation has a negative effect on the morphological state of the intestine [48, 49]. Moreover, feeding Thr supplementation above the recommended level resulted in higher villus length, lower crypt depth, and improved VH/CD ratio in broilers, compared to the control group [43]. In contrast with Ospina-Rojas et al. [50], however, they did not experience a statistically reliable difference in gut morphology when feeding diets with an increased threonine content.
Earlier studies reported, that under commercial conditions a higher level of Thr that exceeds the current NRC recommendation [49] is required to achieve maximum immune function and health status for poultry (51, 52, 53). Thus, it is an interesting issue whether a short-term but targeted Thr supplementation has benefits for broilers.
The intestinal morphological results measured at hatch can hardly be explained by consequent dietary treatments. Overall, we could not confirm any positive effect of the provided threonine supplementation during the embryonic stage on brush border development, at hatch. However, it seems that early Thr supplementation may have a positive effect on the architecture of the intestinal epithelium, particularly on the villus height/crypt depth ratio that is correlated with the absorptive surface of the gut later on. In agreement with other studies [35, 54; 55, 56] our results show that post-hatch feed deprivation compromises the ideal trajectory of gut tissue maturation and there is a significant difference between brush border architecture when birds fed immediately or with 48 hours delay post-hatch. Moreover, data from Proszkowiec-Weglarz et al. [57] suggest, that delay in feeding may indirectly affect the gut barrier function of the small intestine as well as possibly reduce the absorption and utilization of nutrients such as carbohydrates in the intestinal tract of broilers. Those negative effects can be at least partly compensated if gel supplements or early Thr supplementation is provided, as confirmed by our results. In line with these findings, numerous studies [40, 58,] reported higher villi and more intensive mucin secretion in birds received in ovo Thr.
Periportal heterophilic infiltration in day-old chickens and the following mononuclear invasion later at the same histological sites might indicate acute inflammatory response in the young birds that developed a chronic process thereafter in the older animals [59]. The origin of acute inflammation remained unrevealed, although the nature of the lesions may indicate microbial infection before or on the occasion of hatching [60, 61]. As no significant connections were detected between the pathological alterations and the different groups, we can conclude that in ovo treatment neither could induce the histological changes nor had any impact on their further behavior. Hepatic lipidosis in young poultry is generally a natural process that can be connected to lipid transportation from the yolk sac and the slower lipid metabolism in day-old chickens. Usually, these processes induce only mild fat accumulation in a few days after hatching [61.], although more severe and long-lasting lesions were observed in our study. The inflammation resulting in hepatocellular damage could be in the background of prolonged and more prominent lipidosis.
The reduced lymphocyte counts together with increased monocyte numbers in blood work means the so-called stress leukogram both in mammal and bird species that is the result of glucocorticoid effects on blood cells and bone marrow [62]. Our finding indicates that chickens had some degree of stress during their development, although its origin was probably not the nature of experimental treatments because stress leukogram patterns could be observed in all groups and ages.
5. Conclusions
Our results confirmed that the best performance was achieved in birds without delay in feed access. Early Thr supplementation could at least partly compensate for the loss,- attributed to the 48h delay in feed access- post-hatch. The higher compensation ability can be explained by the better architecture of the gut tissue. In ovo Thr supplementation supported the growth and resulted in favorable villus height/crypt depth ratio in the first 3 weeks. The use of Thr fortified Hydrogel® at the post-hatch period according to the performance results, thus it can be recommended for practice for broilers with delayed feed access.
Author Contributions
Conceptualization, writing, and validation V.H.; methodology., Sz., Á., Ö.P., and J.T. J.E J.N; data curation, statistics, visualization V.Á.; formal analysis, V.Á., A.T.; investigation ., N.K. Sz., Á.N., O.Cs.; project administration, V.Á. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by GINOP-2.2.1-18-2018-00031 project and the Flagship Research Groups Programme of the Hungarian University of Agriculture and Life Sciences.
Acknowledgments
A Special thanks goes to János Tossenberger for providing the professional conditions for this study.
Institutional Review Board Statement
The animal study protocol was approved by the Declaration of the Hungarian National Scientific Ethical Committee of Animal Experimentation for studies involving animals, protocol license number is SO/31/00444-2/2021 on 2020.09.29.
Conflicts of Interest
“The authors declare no conflict of interest.”
References
- Fan H, Zengpeng L.V, Gan L, Guo Y: Transcriptomics-related mechanisms of supplementing laying broiler breeder hens with dietary daidzein to improve the immune function and growth performance of offspring J. Agric. Food Chem., 2018, 66, 2049–2060.
- Franco D, Rois D, Arias A, Justo JR, Marti-Quijal FJ, Khubber S, Barba FJ, López-Pedrouso M, Manuel Lorenzo J. Effect of Breed and Diet Type on the Freshness and Quality of the Eggs: A Comparison between Mos (Indigenous Galician Breed) and Isa Brown Hens. Foods. 2020, 9, 342 .
- de Jong, I.C., & van Emous, R.A. (2017): Broiler breeding flocks: Management and animal welfare. In T. Applegate (Ed.), Achieving sustainable production of poultry meat. 2017, 3, 1–19. Burleigh Dodds Science Publishing. https://doi.org/10.19103/AS.2016.0011.26.
- Liu H, Ding P, Tong Y, He X, Yin Y, Zhang H, Song Z, Metabolomic analysis of the egg yolk during the embryonic development of broilers, Poult. Sci., 2021, 100, 101014. [CrossRef]
- Gonçalves FM, Santos VL, Contreira CL, Farina G, Kreuz BS, Gentilini FP, Anciuti MA, Rutz F.In-ovo nutrition: Strategy for precision nutrition in poultry industry. Arch Zootec. 2013, 62, 45–55.
- Batal AB, Parsons CM: Effect of fasting versus feeding oasis after hatching on nutrient utilization in chicks. Poult. Sci. 2002, 81, 853–859.
- Jha R, Singh AK, Yadav S, Berrocoso JFD, Mishra B. Early Nutrition Programming (in ovo and Post-hatch Feeding) as a Strategy to Modulate Gut Health of Poultry. Front Vet Sci. 2019, 21, 82. [CrossRef]
- Riva S, Monjo TP, The importance of early nutrition in broiler chickens: Hydrated gels enriched with nutrients, an innovative feeding system. Anim Husb Dairy Vet Sci, 2020, 4, 1–4. [CrossRef]
- Jha R, Singh AK, Yadav S, Berrocoso JFD, Mishra B. Early Nutrition Programming (in ovo and Post-hatch Feeding) as a Strategy to Modulate Gut Health of Poultry. Front Vet Sci. 2019, 21, 82. [CrossRef]
- Uni Z, Ferket RP. Methods for early nutrition and their potential. Worlds Poult Sci J. 2004, 60, 101–111.
- Noy, Y. and Sklan, D. Yolk utilization in the newly hatched poult. British Poult Sci. 2001, 39, 446–451.
- Geyra,A., Uni, Z. and Sklan, D. Enterocyte dynamics and mucosal development in the posthatch chick. Poult Sci. 2001, 80, 776–782. [CrossRef]
- Vieira SL, Moran ET, Effects of delayed placement and used litter on broiler yields. J App Poult Res, 1999, 8, 75-81. [CrossRef]
- Sharma J and Burmester B, Resistance of Marek's disease at hatching in chickens vaccinated as embryos with the turkey herpesvirus. Avian Dis, 1982, 134-149.
- Berrocoso JD, Kida R, Singh AK, Kim YS, Jha R. Effect of in ovo injection of raffinose on growth performance and gut health parameters of broiler chicken. Poult Sci. 2017, 96, 1573–1580. [CrossRef]
- Bhanja SK, Mandal AB. Effect of in ovo injection of critical amino acids on pre and post-hatch growth, immunocompetence and development of digestive organs in broiler chickens. Asian-Australas J Anim Sci. 2005, 18, 524–531.
- Ohta Y, Yoshida T, Tsushima N, Kidd MT. The needle bore diameter for in ovo amino acid injection has no effect on hatching performance in broiler breeder eggs. J Poult Sci. 2002, 39, 194–197. [CrossRef]
- Jeurissen SH, Lewis F, van der Klis JD, Mroz Z, Rebel JM, and ter Huurne AA. Parameters and techniques to determine intestinal health of poultry as constituted by immunity, integrity, and functionality. Curr Issues Intest Microbiol. 2002, 3, 1–14.
- Gore AB, Qureshi MA. Enhancement of humoral and cellular immunity by vitamin E after embryonic exposure. Poult Sci. 1997, 76, 984–991. [CrossRef]
- Kidd MT. Nutritional modulation of immune function in broilers. Poult Sci. 2004, 83, 650–657. [CrossRef]
- Kidd MT, Peebles ED, Whitmarsh SK, Yeatman JB, Wideman RF. Growth and immunity of broiler chicks as affected by dietary arginine. Poult Sci. 2001, 80, 1535–1542. [CrossRef]
- Kogut MH, Klasing K. An immunologist’s perspective on nutrition, immunity, and infectious diseases: Introduction and overview. J Appl Poult Res. 2009, 18, 103–110. [CrossRef]
- Sisrat S.D., Chaitrashree A. R., Ramteke B.N. Early post-hatch feeding chicks and practical constrains- A review. Agricultural Reviews. 2018, 39, 226–233.
- Uni Z; Ferket P.R. Enhancement of development of oviparous species by in ovo feeding. US Regular Patent US, 2003, 6, B2.
- Official Methods of Analysis of AOAC International. 19th Ed., AOAC INTERNATIONAL, Gaithersburg, MD, USA, Official Method 2008.01. Revised October 2013.
- Fischer AH, Jacobson KA, Rose J, Zeller R. Hematoxylin and eosin staining of tissue and cell sections. CSH Protoc. 2008, 1, 2008. [CrossRef]
- Acharya, M., Arsi, K., Donoghue, A.M.. Production and characterization of avian crypt-villus enteroids and the effect of chemicals. BMC Vet Res 2020, 16, 179. [CrossRef]
- Pedroso, A.A., Chaves, L.S., Lopes, K.L. Leandro A.M, Café N.S.M, & Stringhini, J. H. Inoculação de nutrientes em ovos de matrizes pesadas. Revista Brasileira de Zootecnia, 2006, 35, 2018–2026.
- Jia, C. L., Wei, Z. H., Yu, M., Wang, X. Q., & Yu, F. Effect of in-ovo feeding maltose on the embryo growth and intestine development of broiler chicken. Ind Jour Anim Sci, 2011, 81, 503–506.
- Zhai, W., Gerard, P. D., Pulikanti, R., & Peebles, E. Effects of in ovo injection of carbohydrates on embryonic metabolism, hatchability, and subsequent somatic characteristics of broiler hatchlings. Poult Sci, 2011, 90, 2134–2143. [CrossRef]
- Retes, P. L. In ovo feeding of carbohydrates for broilers-a systematic review. J Anim Phys and Anim Nutr, 2018, 102, 2,361–369.
- Kadam, M. M., Bhanja, S. K., Mandal, A. B., Thakur, R., Vasan, P., Bhattacharyya, A., & Tyagi, J. S., Effect of in ovo threonine supplementation on early growth, immunological responses and digestive enzyme activities in broiler chickens. British Poult Sci, 2008, 49(6), 736–741. [CrossRef]
- Kadam M., Barekatain R., Bhanja Kumar S., Iji P.,Prospects of in ovo feeding and nutrient supplementation for poultry: The science and commercial applications - a review. In: Journal of the Science of Food and Agriculture 2013, 93(15). 1-2.
- Alabi, J.O, Fafiolu, A.O, Bhanja, S.K, Oluwatosin, O.O, Onagbesan, O.M, Dada, I.D, Goel, A., Mehra, M., Gopi, M., Rokade, J.J., Kolluri, G. and Pearlin, B.V. 2022. In ovo L-Threonine feeding modulates the blood profile and liver enzymes activity of CARIBRO Vishal broiler chickens. Indian Journal Poultry Science, 2020, 57, 241-249. [CrossRef]
- Li D.L, Wang J.S, Liu L.J, Li K, Xu Y.B, Ding X.Q, Wang Y.Y, Zhang Y.F, Xie L.Y, Liang S, Wang Y.X, Zhan X.A, Effects of early post-hatch feeding on the growth performance, hormone secretion, intestinal morphology, and intestinal microbiota structure in broilers,Poult Sci, 2022, 101, 102133.
- S.I. Boersma S.I, Robinson F.E, Renema R.A, Fasenko G.M, Administering Oasis Hatching Supplement Prior to Chick Placement Increases Initial Growth with No Effect on Body Weight Uniformity of Female Broiler Breeders After Three Weeks of Age,J App Poult Res,2003, 12, 3, 428-434. [CrossRef]
- Al-Huwaizi H. J. N, Ammar H.A. The effect of fasting and early feeding after hatching with the nutritional supplement gel 95 and the safmannan prebiotic gel and the mixture between them on the productive performance of broiler chicks. IOP Conference Series. Earth and Environmental Science, 2021, 735(1).
- van der Wagt I, de Jong I.C, Mitchell M.A, Molenaar R, van den Brand H. A review on yolk sac utilization in poultry Poult. Sci., 2020, 99, 162–2175.
- Mousavi, S., Foroudi, F., Baghi, F., Shivazad, M., & Ghahri, H., The effects of in ovo feeding of threonine and carbohydrates on growth performance of broiler chickens. In: Proceedings of the British Society of Animal Science, 2009, 198-198. [CrossRef]
- Tahmasebi, S., & Toghyani, M. Effect of arginine and threonine administeredin ovoon digestive organ developments and subsequent growth performance of broiler chickens. J of Anim Phys and Anim Nutr, 2015, 100(5). 947–956.
- Kermanshahi, H., Golian, A., Khodambashi Emami, N., Daneshmand, A., Ghofrani Tabari, D., & Ibrahim, S. A. Effects of in ovo injection of threonine on hatchability, intestinal morphology, and somatic attributes in Japanese quail (Coturnix japonica). J of App Anim Res, 2016, 45(1), 437–441.
- Filho A, Ferket, P, Malheiros R, Bruno de Oliveira C.J, Aristimunha P, Wilsmann, D , Patricia G. Enrichment of the amnion with threonine in chicken embryos affects the small intestine development, ileal gene expression and performance of broilers between 1 and 21 days of age. Poult Sci. 2018, 98. 10.3382/ps/pey461. [CrossRef]
- Y.P. Chen, Y.F. Cheng, X.H. Li, W.L. Yang, C. Wen, S. Zhuang, Y.M. Zhou, Effects of threonine supplementation on the growth performance, immunity, oxidative status, intestinal integrity, and barrier function of broilers at the early age, Poult Sci, 2017, 96, 2,. [CrossRef]
- Law G.K, Bertolo R.F, Adjiri-Awere A, Pencharz P.B, Ball R.O, Adequate oral threonine is critical for mucin production and gut function in neonatal piglets Am. J. Physiol. Gastrointest. Liver Physiol., 2007,292,G1293-G1301. [CrossRef]
- Wang W, Zeng X, Mao X, Wu G, Qiao S. Optimal dietary true ileal digestible threonine for supporting the mucosal barrier in small intestine of weanling pigs J. Nutr., 2010, 140, 981.
- Lien K.A, Sauer W.C, Mosenthin R., Souffrant W.B, Dugan M.E. Evaluation of the 15N-isotope dilution technique for determining the recovery of endogenous protein in ileal digestion of pigs: Effect of dilution in the precursor pool for endogenous nitrogen secretion. J Anim. Sci., 1997, 75,148. [CrossRef]
- Rangel-Lugo M, Su C-L, and Austic R.E, Threonine Requirement and Threonine Imbalance in Broiler Chickens. Poult Sci, 1994, 73, 670–681. [CrossRef]
- National Research Council (NRC) Nutrient requirements of poultry. 9th Edition, National Academy Press, Washington DC. 1994.
- Ospina-Rojas I., Murakami A., Oliveira C., Guerra A. Supplemental glycine and threonine effects on performance, intestinal mucosa development, and nutrient utilization of growing broiler chickens. Poult Sci. 2013, 92, 2724–2731. [CrossRef]
- Corzo A, Kidd M.T, Dozier W.A, Pharr G.T, Koutsos D.A. Dietary threonine needs for growth and immunity of broilers raised under different litter conditions J. Appl. Poult. Res., 2007, 16, 574582.
- Azzam M.M.M, Dong X.Y, Xie P, Zou X.T. Influence of L-threonine supplementation on goblet cell numbers, histological structure and antioxidant enzyme activities of laying hens reared in a hot and humid climate Br. Poult. Sci., 2012, 53, 640.
- Star, M. Rovers, E. Corrent, J.D. Van der Klis. Threonine requirement of broiler chickens during subclinical intestinal Clostridium infection Poult. Sci., 2012, 91, 643.
- Geyra A, Uni Z, Sklan D. The effect of fasting at different ages on growth and tissue dynamics in the small intestine of the young chick. British J of Nutr. 2001, 86, 53–61. [CrossRef]
- Bigot K, Mignon-Grasteau S, Picard M, Tesseraud S. Effects of delayed feed intake on body, intestine, and muscle development in neonate broilers. Poult Sci. 2003, 82, 781–788. [CrossRef]
- Lamot DM, van de Linde IB, Molenaar R, van der Pol CW, Wijtten PJA, Kemp B; et al. Effects of moment of hatch and feed access on chicken development. Poultry Science. 2014, 93, 2604–2614. [CrossRef]
- Proszkowiec-Weglarz, M,Schreier L.L. , Kahl S., Miska K.B. , Russell B. , Elsasser T.H. Effect of delayed feeding post-hatch on expression of tight junction- and gut barrier-related genes in the small intestine of broiler chickens during neonatal development Poult. Sci., 2020, 99, 4714–4729.
- Alexandre L DE, B.M.F., Olivera, C.J.B., Freitas Neto, O.,C., Candice M C G DE,Leon, Saravia, M.M.S., Andrade, M.F.S., White, B. and Givisiez, P.E.N., Intra-Amnionic Threonine Administered to Chicken Embryos Reduces Enteritidis Cecal Counts and Improves Posthatch Intestinal Development. J of Immun Res. 2018, 9.
- Oort J, Scheper RJ. Histopathology of acute and chronic inflammation. Agents Actions Suppl. 1977, 25-30.
- Yonus, M., Nisa, Q., Munir, M., Jamil, T., Kaboudi, K., Rehman, Z., Shah, M. Viral hepatitis in chicken and turkeys. World's Poult Sci J, 2017, 73(2), 379-394. [CrossRef]
- Ito, NMK & Miyaji, CI & Lima, EA & Okabayashi, S & Claure, RA & Graça, EO. Entero-hepatic pathobiology: Histopathology and semi-quantitative bacteriology of the duodenum. BRAZ J POULT SCI. 2004, 6. 10.1590/S1516-635X2004000100005.
- van der Wagt, I., de Jong, I. C., Mitchell, M. A., Molenaar, R., van den Brand, H. A review on yolk sac utilization in poultry. Poult Sci, 2020, Volume 99, Issue 4, Pages 2162-2175, ISSN 0032-5791. [CrossRef]
- Harmon, B. G. Avian Heterophils in Inflammation and Disease Resistance. Poult. Sci. 1998, 77, 972–977.
Figure 1.
Transversal section of duodenum in a three-day-old chicken, H-E., 100X. Red lines represent the morphometric measurement of villus height and crypt depth
Figure 1.
Transversal section of duodenum in a three-day-old chicken, H-E., 100X. Red lines represent the morphometric measurement of villus height and crypt depth
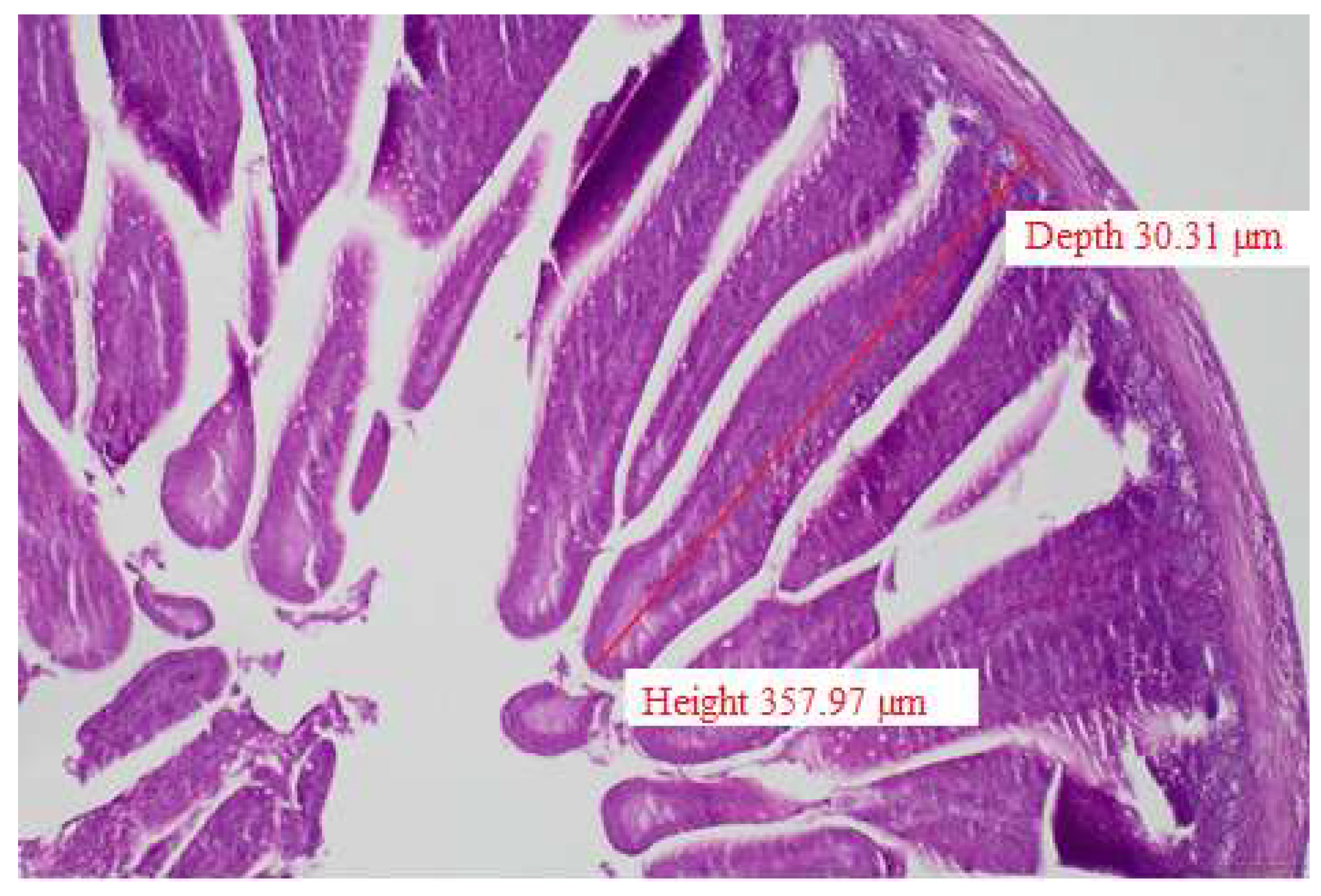
Figure 2.
Hatching weight and live weight at 48 hours post-hatch of broilers in different experimental treatment groups.
Figure 2.
Hatching weight and live weight at 48 hours post-hatch of broilers in different experimental treatment groups.
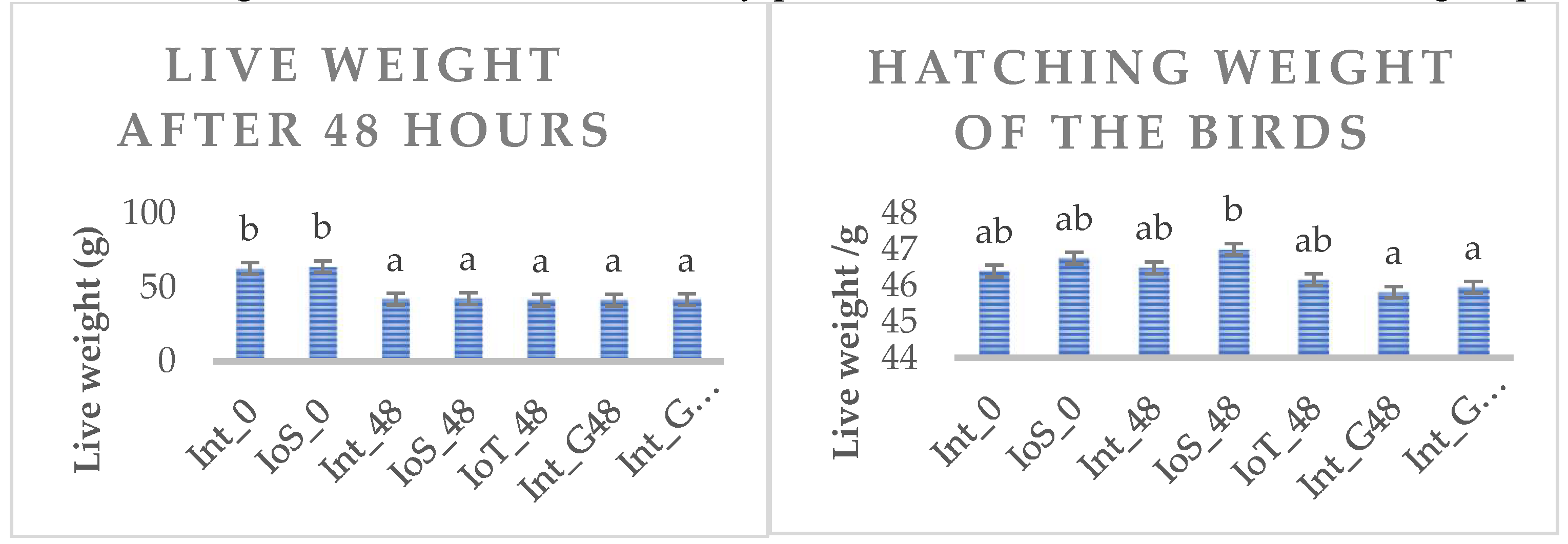

Table 1.
Temperature and CO2 level during incubation.
| Hatching day | °C | CO2 concentration % |
|
|---|---|---|---|
| 1 | Incubation | 37.9 | 0.60 |
| 2 | 37.9 | 0.60 | |
| 3 | 37.9 | 0.60 | |
| 4 | 37.9 | 0.60 | |
| 5 | 37.9 | 0.60 | |
| 6 | 37.9 | 0.60 | |
| 7 | 37.8 | 0.60 | |
| 8 | 37.8 | 0.60 | |
| 9 | 37.6 | 0.60 | |
| 10 | Candling | 37.6 | 0.60 |
| 11 | 37.5 | 0.35 | |
| 12 | 37.5 | 0.35 | |
| 13 | 37.4 | 0.35 | |
| 14 | 37.3 | 0.35 | |
| 15 | 37.3 | 0.35 | |
| 16 | 37.2 | 0.35 | |
| 17 | Candling, in-ovo intervention, placing into the incubator | 37.1 | 0.35 |
| 18 | 37.0/36.7 | 0.35/0.60 | |
| 19 | 36.7 | 0.60 | |
| 20 | 36.5 | 0.60 | |
| 21 | 36.2 | 0.60 | |
| 22 | 36.2/35.8 | 0.35 |
Table 2.
Definition of treatment groups in the study.
| Treatment code | Feed access | Early nutrition method | numbr of eggs |
|---|---|---|---|
| Int_0 | Immediate | - | 160 |
| IoS_0 | in ovo, saline | 160 | |
| Int_48 | 48h delayed solid feed access | - | 160 |
| IoS_48 | in ovo, saline | 160 | |
| IoT_48 | in ovo Thr | 160 | |
| Int_G48 | Hydrogel | 160 | |
| Int_GT48 | Hydrogel+Thr | 160 |
Table 3.
Analyzed feed composition in the three feeding phases.
| Ingredients | Starter (1–10) |
Grower (11–21) |
Finisher (22–35) |
|---|---|---|---|
| Corn (grain) | 551 | 577 | 601 |
| Corn gluten (60%) | 32 | 32 | 32 |
| Sunflower meal | 53.5 | 53.5 | 75 |
| Soybean meal (CP 44.2%) | 262 | 230 | 175 |
| Fat, vegetable | 44.7 | 55 | 67.00 |
| MCP | 18.7 | 17.5 | 15 |
| Limestone | 15 | 13.5 | 12.2 |
| NaCl | 2.7 | 2.7 | 2.7 |
| L-Lysin HCl | 5.2 | 4.6 | 4.3 |
| DL-Methionin | 4.5 | 3.9 | 3.2 |
| L-Treonin | 2.6 | 2.3 | 1.8 |
| Premix1 | 5.00 | 5.00 | 5.00 |
| Total | 1000.00 | 1000.00 | 1000.00 |
| Nutrient content (g/kg) | |||
| AMEn (MJ/kg) | 12.5 | 12.9 | 13.4 |
| DM % | 90 | 91.3 | 91.1 |
| Crude protein | 204.2 | 190.7 | 174.9 |
| Crude fat | 71.87 | 82.3 | 94.4 |
| Crude fiber | 41.5 | 41.1 | 44.8 |
| Lysine* | 13.5 | 12,1 | 10,8 |
| M + C* | 10.8 | 9.9 | 9.0 |
| Threonin* | 9.7 | 8,8 | 7,8 |
| Triptophan* | 2.4 | 2.3 | 1.7 |
| Ca | 9.6 | 8.7 | 7.8 |
| P avaliable | 4.7 | 4.5 | 3.9 |
| Na | 1.7 | 1.7 | 1.7 |
1Premix feed contents per kilogram: Zn: 22,032 mg, Cu: 3200 mg, Fe: 16,020 mg, Mn: 21,948 mg, I: 300 mg, Se: 70 mg, Co: 20 mg, Vit. A: 324,0000 IU, Vit. D3: 810,000 IU, Vit. E: 20,800 mg, Vit K3: 810 mg, Vit. B1: 810 mg, Vit. B2: 1890 mg, Vit. B3: 10,800 mg, Vit. B5: 3240 mg, Vit. B6: 1350 mg, Vit B12: 6.8 mg, Folic acid: 270 mg, Biotin: 32 mg.* calculated values
Table 4.
Effect of dietary treatments on body weight at the end of different phases.
| Int_0 | IoS_0 | Int_48 | IoS_48 | IoT_48 | Int_G48 | Int_GT48 | RMSE | P-value | |
|---|---|---|---|---|---|---|---|---|---|
| d10 | 233a | 245b | 191c | 193c | 196c | 194c | 192c | 25.8 | <0.0001 |
| d21 | 855ab | 882a | 766c | 782cd | 809bd | 777cd | 785cd | 95.3 | <0.0001 |
| d35 | 2218a | 2238a | 2072b | 2113ab | 2086b | 2096ab | 2100ab | 257.4 | <0.0001 |
a,b means in the same row with different letters statistically differ (P<0.05).
Table 5.
Effect of dietary treatments on feed intake (FI) feed conversion ratio (FCR) and average daily gain (ADG) in different phases and the whole experiment.
Table 5.
Effect of dietary treatments on feed intake (FI) feed conversion ratio (FCR) and average daily gain (ADG) in different phases and the whole experiment.
| Int_0 | IoS_0 | Int_48 | IoS_48 | IoT_48 | Int_G48 | Int_GT48 | RMSE | P-value | |
|---|---|---|---|---|---|---|---|---|---|
| Feed intake (kg/day/pen) | |||||||||
| d1-10 | 23.2a | 24a | 17.9b | 17.7b | 17.8b | 17.1b | 17.9b | 1.72 | <0.0001 |
| d11-21 | 72.8ab | 75.6a | 66.8c | 66.7c | 69.1bc | 67.3c | 66.8c | 1.36 | <0.0001 |
| d22-35 | 145.6a | 144.4a | 136.9b | 138.6ab | 137.6b | 138.6ab | 140.1ab | 6.22 | 0.04 |
| d1-35 | 85.35a | 86.3a | 78.3b | 78.2b | 79.3b | 78.9b | 79.1b | 3.52 | <0.0001 |
| Feed conversion ratio (kg feed/ kg gain) | |||||||||
| d1-10 | 1.24a | 1.21ab | 1.24a | 1.21ab | 1.18b | 1.15b | 1.23ab | 0.13 | 0.04 |
| d11-21 | 1.28ab | 1.3a | 1.28ab | 1.24ab | 1.24b | 1.26ab | 1.23b | 0.77 | 0.001 |
| d22-35 | 1.49b | 1.48b | 1.45ab | 1.45a | 1.5b | 1.46a | 1.48b | 0.13 | 0.03 |
| d1-35 | 1.37ab | 1.37ab | 1.35b | 1.32bc | 1.36ac | 1.34c | 1.34c | 0.13 | 0.043 |
| Average daily gain (g/d) | |||||||||
| d1-10 | 18.6b | 19.8a | 14.4c | 14.6c | 15.0b | 14.8b | 14.5b | 2.57 | <0.0001 |
| d11-21 | 56.5ab | 57.9a | 52.1c | 53.5bc | 55.6ab | 53.0bc | 53.9bc | 7.1 | <0.0001 |
| d22-35 | 97.5 | 97.1 | 93.8 | 95.0 | 91.2 | 94.5 | 94.2 | 14.28 | 0.13 |
| d1-35 | 62.0a | 62.6a | 57.8b | 59ab | 58.3b | 58.5ab | 58.7ab | 7.19 | <0.0001 |
a,b,c means in the same row with different letters statistically differ (P<0.05).
Table 6.
Effect of dietary treatments on villus height, crypt depth, and villus height/crypt depth ratio in the duodenum, ileum, and colon (D, I, and colon, respectively) at hatch.
Table 6.
Effect of dietary treatments on villus height, crypt depth, and villus height/crypt depth ratio in the duodenum, ileum, and colon (D, I, and colon, respectively) at hatch.
| Int_0 | IoS_0 | Int_48 | IoS_48 | IoT_48 | Int_G48 | Int_GT48 | P-value | |
|---|---|---|---|---|---|---|---|---|
| Villus height (µm) | ||||||||
| D | 580.8ab | 506.5a | 673.2b | 639.8ab | 621.8ab | 579.2ab | 586.1ab | 0.032 |
| I | 302.6a | 295.4a | 363.6ab | 375.3b | 358.7ab | 400b | 364.5ab | 0.0011 |
| C | 318.3ab | 377.8a | 347.2ab | 308.1ab | 357.3a | 310.5ab | 258.2b | 0.0055 |
| Crypt depth (µm) | ||||||||
| D | 111.3a | 110.3a | 109.5a | 80.5b | 115.4a | 100.1ab | 103.9ab | 0.001 |
| I | 88.1 | 94.6 | 93.3 | 94.9 | 83.4 | 106.0 | 85.6 | 0.16 |
| C | 92.5ab | 87.4a | 104.5b | 81.4a | 110.5b | 94.4ab | 82a | 0.0004 |
| Villus height/Crypt depth ratio | ||||||||
| D | 4.3a | 4.6a | 4.8a | 8.2b | 5.5a | 5.1a | 3.9a | <0.001 |
| I | 3.4ab | 3a | 3.8ab | 4ab | 3.8ab | 3.8ab | 4.4b | 0.0082 |
| C | 4.0 | 4.2 | 3.5 | 4.0 | 5.5 | 3.4 | 3.3 | 0.28 |
a,b means in the same row with different letters statistically differ (P<0.05).
Table 7.
Effect of dietary treatments on villus height, crypt depth, and villus height/crypt depth ratio in the duodenum, ileum, and colon (D, I, and colon, respectively) 48 hours post-hatch.
Table 7.
Effect of dietary treatments on villus height, crypt depth, and villus height/crypt depth ratio in the duodenum, ileum, and colon (D, I, and colon, respectively) 48 hours post-hatch.
| Int_0 | IoS_0 | Int_48 | IoS_48 | IoT_48 | Int_G48 | Int_GT48 | P-value | |
|---|---|---|---|---|---|---|---|---|
| Villus height (µm) | ||||||||
| D | 340.6c | 214.9a | 288.5b | 240.8b | 283.4b | 321.6bc | 250.0ab | <0.001 |
| I | 158.6 | 140.2 | 167.4 | 140.8 | 154.7 | 157.1 | 142.9 | 0.057 |
| C | 164.2b | 126.4a | 159.8ab | 146.4ab | 156.5ab | 132.1ab | 130.0ab | 0.0085 |
| Crypt depth (µm) | ||||||||
| D | 36.3 | 29.2 | 35.6 | 34.0 | 36.5 | 31.8 | 35.6 | 0.43 |
| I | 32.5 | 30.3 | 33.7 | 29.6 | 32.6 | 33.6 | 32.8 | 0.14 |
| C | 29.9 | 25.3 | 30.3 | 29.1 | 28.3 | 26.5 | 26.7 | 0.059 |
| Villus height/Crypt depth ratio | ||||||||
| D | 7.7 | 8.1 | 8.1 | 7.6 | 7.8 | 9.2 | 7.2 | 0.28 |
| I | 4.8 | 4.5 | 4.8 | 4.8 | 4.8 | 4.7 | 4.6 | 0.96 |
| C | 5.7b | 5.1b | 3.7a | 4.5ab | 4.5ab | 4.9b | 4.9b | 0.006 |
a,b,c means in the same row with different letters statistically differ (P<0.05)
Table 8.
Effect of dietary treatments on villus height, crypt depth, and villus height/crypt depth ratio in the duodenum, ileum, and colon (D, I, and colon, respectively) at 21 days of age.
Table 8.
Effect of dietary treatments on villus height, crypt depth, and villus height/crypt depth ratio in the duodenum, ileum, and colon (D, I, and colon, respectively) at 21 days of age.
| Int_0 | IoS_0 | Int_48 | IoS_48 | IoT_48 | Int_G48 | Int_GT48 | P-value | |
|---|---|---|---|---|---|---|---|---|
| Villus height (µm) | ||||||||
| D | 768.9ab | 748.8ab | 830.9a | 790.8ab | 755.6ab | 690.1b | 765.8ab | 0.04 |
| I | 210.6a | 221.3ab | 216.1a | 199.3a | 282.8b | 218.7a | 180.4a | 0.0004 |
| C | 214.9 | 235.3 | 220.8 | 237.9 | 205.2 | 211.1 | 217.2 | 0.45 |
| Crypt depth (µm) | ||||||||
| D | 86.1 | 103.5 | 106.8 | 89.5 | 97.4 | 104.5 | 113.2 | 0.09 |
| I | 61.8 | 59.2 | 55.3 | 58.4 | 72 | 66.8 | 60.7 | 0.08 |
| C | 61.9 | 64.3 | 54.3 | 56.1 | 58.8 | 56.1 | 55.2 | 0.26 |
| Villus height/Crypt depth ratio | ||||||||
| D | 9.6a | 7.8ab | 8.1ab | 9.1ab | 8.2ab | 6.6b | 7.4ab | 0.01 |
| I | 3.5ab | 3.8ab | 4ab | 3.5ab | 4.2a | 3.3ab | 3b | 0.01 |
| C | 3.6 | 3.7 | 4.2 | 4.6 | 3.8 | 3.9 | 4.1 | 0.29 |
a,b means in the same row with different letters statistically differ (P<0.05)
Table 9.
Effect of dietary treatment on heterophil granulocyte infiltration, vacuolization, mononuclear infiltration in the liver tissue, and lipid accumulation in liver cells at different time points of the experiment.
Table 9.
Effect of dietary treatment on heterophil granulocyte infiltration, vacuolization, mononuclear infiltration in the liver tissue, and lipid accumulation in liver cells at different time points of the experiment.
| Int_0 | IoS_0 | Int_48 | IoS_48 | IoT_48 | Int_G48 | Int_GT48 | P-value* | |
|---|---|---|---|---|---|---|---|---|
| Heterophil granulocyte infiltration | ||||||||
| day 1 | 0.66ab | 0.75ab | 1.0b | 0.58a | 1.0b | 1.0b | 1.0b | 0.01 |
| day 3 | 0.41ab | 1.0b | 0.25a | 0.57ab | 1.0b | 1.0b | 0.75ab | 0.0007 |
| day 21 | 1.1ab | 0.3a | 0.58a | 1.41b | 1.66b | 1.19ab | 1.6b | <0.001 |
| Vacuolization | ||||||||
| day 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - |
| day 3 | 0.16ab | 0a | 0a | 0.42b | 0a | 0a | 0a | 0.017 |
| day 21 | 0a | 0a | 0a | 0.28b | 0a | 0a | 0a | 0.007 |
| Mononuclear infiltration | ||||||||
| day 1 | 0a | 0a | 0a | 0.25 | 0a | 0a | 0a | 0.01 |
| day 3 | 1.41b | 0.33a | 0a | 0.71ab | 0.5a | 1ab | 0a | 0.008 |
| day 21 | 1.77ab | 2b | 1.6ab | 1.3ab | 2.33b | 1.33a | 1.33a | <0.001 |
| Lipid accumulation in liver cells | ||||||||
| day 1 | 1.0a | 1.2b | 1.0a | 1.0a | 1.0a | 1.0a | 1.0a | 0.014 |
| day 3 | 0.25a | 1.66c | 0.75ab | 1.0b | 1.0b | 1.0b | 1.0b | <0.001 |
| day 21 | 0a | 0a | 0a | 0a | 0a | 0.8b | 0.4ab | <0.001 |
* represent Chi2 P-value; a,b means in the same row with different letters statistically differ (P<0.05).
Table 10.
Effect of dietary treatment on the leukocytes* in blood smear at hatch, 48 hours post-hatch, and 21 days of age.
Table 10.
Effect of dietary treatment on the leukocytes* in blood smear at hatch, 48 hours post-hatch, and 21 days of age.
| Int_0 | IoS_0 | Int_48 | IoS_48 | IoT_48 | Int_G48 | Int_GT48 | P-value | |
|---|---|---|---|---|---|---|---|---|
| At hatch | ||||||||
| HE | -- | 0.715 | 0.6 | 0.728 | 0.586 | 0.676 | 0.715 | 0.09 |
| LYM | - | 0.216 | 0.316 | 0.198 | 0.305 | 0.241 | 0.216 | 0.13 |
| MON | - | 0.019 | 0.036 | 0.026 | 0.025 | 0.016 | 0.019 | 0.10 |
| EOS | - | 0.048 | 0.048 | 0.047 | 0.068 | 0.067 | 0.048 | 0.86 |
| 48 hours post-hatch | ||||||||
| HE | 0.629 | 0.529 | 0.619 | 0.597 | 0.610 | 0.525 | 0.629 | 0.052 |
| LYM | 0.291 | 0.386 | 0.305 | 0.339 | 0.314 | 0.397 | 0.291 | 0.79 |
| MON | 0.035 | 0.023 | 0.028 | 0.025 | 0.034 | 0.028 | 0.035 | 0.90 |
| EOS | 0.045 | 0.038 | 0.048 | 0.039 | 0.042 | 0.050 | 0.045 | 0.05 |
| 21 days of age | ||||||||
| HE | 0.314 | 0.323 | 0.296 | 0.346 | 0.317 | 0.357 | 0.314 | 0.33 |
| LYM | 0.578 | 0.579 | 0.613 | 0.570 | 0.571 | 0.563 | 0.578 | 0.82 |
| MON | 0.054 | 0.043 | 0.040 | 0.035 | 0.044 | 0.040 | 0.054 | 0.47 |
| EOS | 0.053 | 0.055 | 0.051 | 0.049 | 0.065 | 0.040 | 0.053 | 0.69 |
Leukocytes: HE= heterophils, LYM= lymphocytes, MON= monocytes, EOS= eosinophyls.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



