
Submitted:
15 April 2025
Posted:
16 April 2025
You are already at the latest version
Abstract
Climate change-induced drought threatens forest restoration by limiting seedling establishment. To address this, we designed synthetic microbial communities (SynComs) to enhance drought tolerance in Quercus pubescens and Sorbus domestica. Bacteria were isolated from distinct soil compartments and depths, based on drought legacy effects. Functional characterization revealed compartment-specific traits, including exopolysaccharide production, auxin and abscisic acid biosynthesis, siderophore production, and osmotic tolerance. We assembled four-strain SynComs guided by ecological origin and functional complementarity. Biofilm assays showed that weak individual producers could significantly enhance community performance, indicating emergent cooperativity. SynComs and single strains were tested on Arabidopsis thaliana, Q. pubescens, and S. domestica, selected for contrasting root strategies within the root economics space. SynComs reduced drought symptoms by 47% in Q. pubescens (SynCom B) and 71% in S. domestica (SynCom F), outperforming individual strains. Exopolysaccharide-producing SynCom B aligned with Q. pubescens conservative roots, while hormone-producing SynCom F matched S. domestica acquisitive strategy. Predictive modeling identified bacterial identity and symptom timing as key predictors of drought tolerance, with stem diameter increment as a physiological indicator. Overall, SynCom effectiveness reflected compatibility between microbial traits, host strategies, and drought context—supporting their use in climate-adaptive forest restoration.
Keywords:
climate change
; drought
; forest restoration
; microbiome-assisted forest restoration
; plant-bacteria interactions
; synthetic communities
; tree seedlings
; extracellular polymeric substances
; biofilm
1. Introduction
Climate change is profoundly disrupting ecosystems, particularly forests, through rising temperatures and altered precipitation patterns linked to greenhouse gas emissions [1]. These shifts are intensifying extreme weather events, including droughts and floods, which increasingly threaten forest stability and health [2,3,4,5].
Forest restoration remains a key strategy for climate change mitigation and ecological recovery, especially when grounded in the use of native species and community engagement [6]. Yet, its effectiveness is challenged by climate extremes that impair seedling survival during establishment. This growing vulnerability calls for expanded nature-based solutions to improve resilience in forest restoration programs [7,8].
Plants employ complex strategies to recruit beneficial microbiomes, involving chemical signals, immune regulation, and microbial selection [9,10,11]. Under drought, they release specific root metabolites that signal stress and selectively attract plant-associated microorganisms [12,13], thereby enhancing resilience [14,15]. These interactions highlight secondary metabolites as central agents in coordinating plant-microbe-environment communication [10].
Plant-associated microbes (PAMs), such as growth-promoting rhizobacteria (PGPB) and mycorrhizal fungi, are widely recognized for enhancing drought tolerance in crops by improving nutrient uptake, hormone balance, osmotic regulation, and stress protection [16]. However, their application in forest trees remains underexplored. Promising results have been reported: Tiepo et al. [17,18] demonstrated the efficacy of commercial PGPB strains in boosting drought tolerance in Neotropical tree seedlings, while Khosravi et al. [19] identified native drought-tolerant microbial strains that significantly improved oak seedling performance under drought.
Despite this, forest research has largely focused on broad soil processes, often overlooking specific microbe–tree interactions [20,21]. While mycorrhizal symbioses are well-established in both crops and trees [22], the role of bacteria differs. In forests, emphasis is placed on mycorrhization helper bacteria—especially phosphate-solubilizing strains—to support symbiotic networks [23,24], whereas agriculture prioritizes PGPB to counteract soil degradation. This contrast underscores the need for more targeted studies on bacterial support for tree seedlings facing drought.
Synthetic microbial communities (SynComs) generally outperform single-strain inoculations in helping crops face drought stress [25,26,27]. Recent studies highlight diverse strategies, based on bottom-up or top-down strategies for constructing effective SynComs, emphasizing ecological relevance, functional redundancy, and stability [28,29,30]. However, challenges like microbial interactions, and environmental variability add additional layers of complexity, making it clear that SynCom development is highly context-dependent and lacks a one-size-fits-all solution.
Belowground carbon allocation plays a key role in soil processes, with roots and mycorrhizal fungi driving carbon and nutrient cycling. Root strategies are traditionally classified as “fast” (thin, short-lived, high nutrient uptake) or “slow” (thicker, longer-lived, adapted to low-resource conditions) [31]. Recent insights introduce a second axis centered on fungal collaboration: plants either invest in high specific root length to forage independently or “outsource” resource acquisition through symbiotic networks, forming thicker roots in the process [32]. This expanded “root economics space” integrates both root construction and fungal and bacterial symbiotic reliance, offering a more nuanced view of belowground strategy (Prada-Salcedo et al. 2021; Wu et al. 2023) [33,34]. Drought can further shift these strategies by altering root thickness [35] and reducing nutrient mobility, thereby increasing the carbon investment required for root and mycorrhizal structures [36]. Understanding how these strategies interact with SynComs offers promising insights for enhancing drought tolerance.
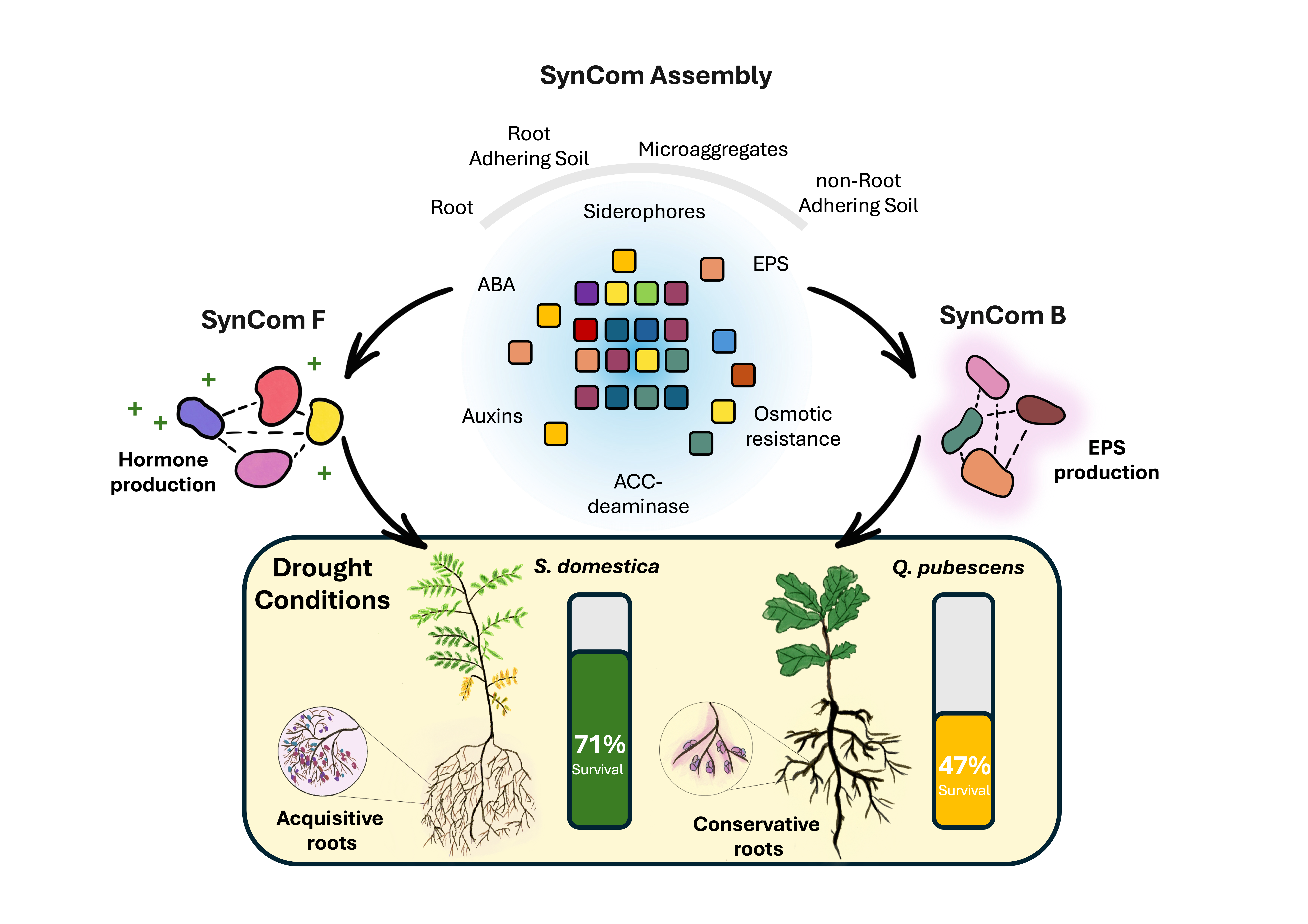
In this study, we developed nature-based solutions by designing tailored bacterial consortia (SynComs) inspired by soil niches and gradients, aiming to enhance drought tolerance in tree seedlings during their vulnerable establishment phase (Figure 1). Strains were isolated from a Mediterranean forest soil subjected to ten years of partial rain exclusion, sampled across depths and root-associated compartments—roots, root-adhering soil, non-root-adhering soil, and root-induced macroaggregates—to capture diverse functional traits. Selection prioritized exopolysaccharide (EPS) producers due to their role in improving soil moisture retention, followed by key PGPB traits like nutrient solubilization, hormone production, and stress tolerance.
We then assembled four-strain SynComs combining complementary functions reflective of forest soil heterogeneity. These were evaluated for cooperative biofilm formation, a proxy for interspecific ecological interactions [37], and for their plant-growth-promoting effects on Arabidopsis thaliana. Multivariate analyses helped identify high-performing and ineffective SynComs to validate our screening strategy.
Finally, we tested selected SynComs on Quercus pubescens and Sorbus domestica, two Mediterranean trees with contrasting root traits. We hypothesized that functional alignment between microbial traits and plant strategies—root architecture, physiology, and symbiosis—is critical for SynCom success.
2. Results
Bacterial Strain Isolation
Soil was collected from the O3HP experimental site (Oak Observatory at the Observatoire de Haute Provence–OHP), a downy oak (Quercus pubescens) forest in southern France. Since 2012, this site has been equipped with a partial rain exclusion system that simulates a 35% reduction in spring and summer rainfall, creating intensified drought conditions for studying ecosystem responses to climate change [38].
To capture the soil functional and spatial diversity, we collected samples from both 0-5 cm (litter removed) and 5-10 cm depths of the A horizon. These layers differ in physicochemical properties such as organic matter, pH, and calcium carbonate content [39], influencing microbial communities. We further subdivided the soil into four compartments based on proximity and interaction with plant roots: (1) the Root itself, which harbors specialized mutualists and endophytes [40,41], (2) Root-Adhering Soil (RAS), enriched in metabolically diverse genera such as Pseudomonas, Bacillus, and Streptomyces [40,42], (3) Root-Induced Macroaggregates (RIM), supporting oligotrophic, fungal-associated taxa [41,43], and (4) non-Root-Adhering Soil, acting as a reservoir of microbial diversity [41,44].
A total of 1,000 microbial strains were isolated under different culture medium nutrient availability (Tryptic Soy Agar (TSA) diluted 10-, 20-, and 50-fold), from soil samples across two depth layers (0-5 cm, 5-10 cm), and four root-related compartments (Roots, RAS, non-RAS, RIM), to capture a wide diversity of growth strategies (Figure 2). From these, 504 isolates were obtained from the upper and 496 from the lower soil layer. In both layers, non-RAS and RIM accounted for the majority of isolates, with smaller contributions from RAS and Roots (Figure 2b and c).
Among the 1,000 strains, 58 (5.8%) showed a strong mucoid phenotype (Figure 2d) and were screened for PGPB core traits (Supplementary File S1), including phytohormone production, nutrient solubilization, and stress tolerance [45]. Auxin (indole-3-acetic acid, IAA) production was strictly tryptophan-dependent, with 18 strains producing ranging from 0.84 to 58.92 µg mL⁻¹. Additionally, 41 isolates (83%) exhibited ACC-deaminase activity, and all mucoid strains grew at -0.51 MPa, indicating high osmotic stress tolerance. Fitness assays identified 54 strains (93%) of interest, including 51 tolerant, 3 resistant (Figure S1), and 28 (48%) producing hydroxamate-type siderophores. Of these, 12 were pyoverdine-producing, typically linked to Pseudomonas under iron-limiting conditions.
Based on their overall traits, twelve top-performing isolates were selected to represent the spatial and nutritional diversity of the root–soil ecosystem (Figure 3). These included strains from four genera—Peribacillus, Pantoea, Pseudomonas, and Caballeronia—identified through full-length 16S rRNA gene sequencing (Supplementary File S1). Two isolates were identified as Peribacillus simplex (s1 and s57), one as Pantoea pleuroti (s14), and one as Caballeronia glathei (s48). Notably, the genus Pseudomonas was dominant, comprising multiple species, including P. umsongensis (s28), P. migulae (s31), P. silesiensis (s39 and s42), P. mandelii (s40 and s54), P. lini (s59), and Pseudomonas sp. (s41).
The isolates were further evaluated for plant growth-promoting properties, including salt tolerance, exopolysaccharide (EPS) production, and abscisic acid (ABA) synthesis. Most strains tolerated 0.5 M NaCl, with P. pleuroti s14 showing halotolerance up to 1.5 M NaCl. ABA production was detected only in P. umsongensis s28 (0.39 µgL-1 at 0.5 M NaCl), while EPS was produced by four strains including P. simplex s12 and s57, C. glathei s48, and P. lini s59 (Supplementary File S1).
Growth dynamics were also assessed using two key parameters: t_mid (time to reach half of carrying capacity) and t_gen (doubling time), summarized in Figure S2. Nine strains—including s12, s14, s28, s31, and s39—exhibited fast growth (t_gen < 1 h and t_mid < 9 h), while three others (s41, s48, s49) displayed slower growth with doubling times over 1.5 hours and t_mid above 10 hours.
Multivariate Analysis of Strain Clustering Based on Functional Traits, Spatial and Nutrient Gradients
We conducted a multiple correspondence analysis (MCA) to explore associations between bacterial strains and their plant growth-promoting traits across spatial and nutrient gradients (Figure 4A). Functional numerical variables (EPS production, IAA and ABA production, ACC-deaminase activity, and osmotic/halotolerance) were converted into ordinal categories ranging from A to D prior to MCA, to enable correspondence-based analysis. Category A corresponded to low or no expression, B to moderate expression, C to high expression, and D to very high expression.
The first two dimensions explained 45.9% of the total variance. Separation along Dimension 1 (Dim 1) was primarily driven by ecological factors—namely compartment, isolation medium, and sampling depth—which were strongly associated together and contributed substantially to this axis structure. Dimension 2 (21.6%) captured variation in functional traits, particularly high EPS production, IAA production, and strong osmotic and halotolerance (NaCl 2 M). These two groups of variables—ecological and functional—varied independently across strains. Interestingly, lower osmotic and halotolerance (NaCl 0.5–1.5 M) and genus-level taxonomic affiliation were highly correlated and contributed almost equally to both dimensions. In contrast, PEG-osmotolerance, ACC deaminase activity, and ABA production showed limited discriminative power in this analysis and were not strongly associated with either axis.
The MCA biplot displays the distribution of bacterial strains in relation to their categorical traits (Figure 4B). The analysis revealed four distinct strain clusters along the two main dimensions (Figure 4B). One cluster, represented by strain s14, appeared isolated on the far right of Dimension 1 and lower end of Dimension 2. The remaining strains formed a gradient from the upper right to the lower left quadrant, reflecting mixed ecological and functional traits. Based on this distribution, four strain classes were defined: i) s14—high IAA production and strong osmotic tolerance, isolated from 0–5 cm Roots; ii) s12 and s57—high EPS producers with strong osmotic tolerance, from 0–5 cm Roots and 5–10 cm RIM, respectively; iii) s31, s48, s59—strong EPS and moderate IAA production with intermediate osmotic tolerance, from 5–10 cm RAS, non-RAS, and RIM; iv) s28, s39, s40, s41, s42, s54—low osmotic and halotolerance, with intermediate IAA, mostly from 5–10 cm RAS or non-RAS compartments.
Designing Synthetic Communities (SynComs)
Recent studies have shown that microbial consortia often exhibit enhanced functional capabilities compared to single-strain inoculants, particularly under complex environmental conditions [46,47,48]. We applied a consortium-based approach using four-strain combinations selected according to the spatial origin and nutrient context of their isolation sites. From an initial pool of twelve strains, 495 theoretical four-strain combinations were possible.
Based on the MCA biplot and the resulting trait-based clustering into four classes, we focused on strains isolated from 0–5 cm Roots and associated with strong functional potential—specifically those including either P. pleuroti s14 (Class 1), P. simplex s12 (Class 2), or both. We first generated 36 combinations by starting with s14 (Class 1), then adding one strain from Class 2, one from Class 3, and one from Class 4. This process was repeated twice: once excluding s12 (18 combinations), and once starting with s12 and excluding s14 (18 combinations), resulting in a total of 72 selected SynComs (Supplementary File S2).
This design aimed to generate consortia spanning a functional and ecological gradient, representative of diverse rhizosphere niches and soil compartments.
Biofilm Assay
We evaluated synergistic/antagonistic biofilm production [49,50] among 12 bacterial strains arranged into 72 four-strain consortia (Table S1). Relative to their best-performing single strains in the consortium, the screening revealed synergistic interactions enhancing biofilm production in 8 consortia and negative interactions reducing biofilm formation in 10 consortia (Figure S3). The remaining consortia showed no significant change in biofilm production (data not shown).
In Vitro Inoculation of Arabidopsis with Single Bacterial Strains or Consortia
We assessed the plant-growth-promoting potential of 12 individual bacterial strains and 8 consortia exhibiting contrasting biofilm formation, categorized as highest (A: 12-48-59-41; B: 12-48-59-54; C: 14-57-59-54; D: 14-57-48-28), lowest (E: 14-12-31-42; F: 12-31-48-28), and intermediate (G: 12-48-59-39; H: 14-48-59-41) biofilm producers. To improve readability, the strain prefix “s” (e.g., s1–s12) was omitted when listing strains within SynCom compositions.
Seedlings were grouped into three developmental categories based on observed growth patterns: (i) germination, (ii) growth (continued development after germination), and (iii) advanced development, characterized by large rosettes (diameter ≥ 0.7 cm), at least six leaves, and more than ten lateral roots. Germination rates remained consistently high (>90%) across all treatments, including controls, with no significant enhancement observed from any consortium or individual strain. (Figure S4-S6). Inoculation, whether with consortia or single strains, did not significantly influence seedling development (Figure S4-S6), regardless of biofilm interactions. Two exceptions were strain 41 (Pseudomonas sp.), which markedly improved seedling establishment, and strain 54 (P. mandelii), which completely inhibited the emergence of well-developed seedlings (Figure S5). Although both strains exhibited similar in vitro traits—intermediate IAA production, osmotic tolerance, and biofilm formation—they produced contrasting effects on Arabidopsis development. Consequently, in this assay, no clear link emerged between biofilm production and plant growth promotion.
Data-Driven Selection of Top-Performing SynCom Candidates
To streamline the selection of the most effective single-strain inoculants and SynComs for young tree experiments, we performed a Principal Component Analysis (PCA) using four quantitative variables related to plant development: germination, growth, development, and biofilm formation. The goal was to reduce dimensionality and identify key associative features. The first two principal components explained 82.9% of the total variance, reflecting strong dimensional reduction and a reliable summary of the data. PC1 accounted for the majority of the variance (61.6%), with PC2 contributing an additional 21.3% (Figure 5a). PCA identified two distinct axes of variation, distinguishing biofilm formation and early-stage traits such as germination from later seedling responses including growth and development. Growth and development in A. thaliana were positively correlated and loaded primarily on PC1, while biofilm production and germination were positively correlated and clustered along PC2. The orthogonality of these vectors indicates that early and late plant responses varied independently across treatments. Contribution values were relatively balanced (as shown by the color scale), indicating that all four variables contributed meaningfully to the PCA dimensions.
The PCA biplot (Figure 5b) revealed two distinct clusters, highlighted by green and violet ellipses. The first cluster grouped SynComs that were positively associated with biofilm formation and plant growth traits—either germination (A, C, and D), growth and development (F and H), or both (B). The second cluster consisted mainly of single strains that showed weak or negative associations with the measured traits, except for s28, s31, and s39, which were strongly linked to growth and development. These results indicate that seed inoculation with synthetic microbial consortia generally enhanced Arabidopsis thaliana performance more effectively than most single-strain inoculations.
To select the best promising treatments, we considered both the projection of each observation onto the trait vectors and the position of this projection relative to the barycentre of the observations in the multivariate space. Based on this analysis, SynComs B, F, and H were identified as the best performers for promoting plant growth and development, whereas SynComs A and D were among the least effective. Among the top-performing individual strains, s31 and s39 were selected over s28 due to their superior tolerance to osmotic stress (Supplementary File S1).
SynComs and Single Inoculants to Mitigate Drought Stress in Tree Seedlings
Five weeks after inoculation for Q. pubescens and four for S. domestica, watering was withheld until soil moisture reached 40% field capacity (FC), which was then maintained. Over the following five weeks, plants were monitored for drought symptoms such as wilting leaves, discoloration, or premature leaf drop (Figure S7). The percentage of plants that remained symptom-free served as a proxy for drought tolerance, reflecting their capacity to maintain normal physiological processes despite reduced water availability (https://foresteurope.org/drought-adaptation-of-forests-in-europe-practical-strategies/). In ecological studies, this metric can serve as an indirect indicator of carbon assimilation efficiency, intrinsic water-use efficiency, and overall forest resilience under drought conditions (Jia et al., 2022).
Kaplan-Meier survival curves (KM curves) and log-rank test (LRT) were used to assess differences between the groups. The hazard function plays a very important role in survival analysis. The Cox proportional hazards model (Hazard Ratio, HR ; Wald’s test) investigates the relationship of predictors and the time-to-event through the hazard function.
Survival analysis of Q. pubescens seedlings subjected to drought conditions (Figure S7a and b) revealed significant differences among bacterial treatments (KM curves, LRT, p-value = 0.0052). Consortium B provided the strongest protective effect with a HR of 0.53 (95% CI [0.30-0.97], p-value = 0.039), indicating that the risk of drought symptoms was reduced by approximately 47% compared to the water control. Single-strain inoculations did not significantly improve drought tolerance in QP seedlings relative to the controls (log-rank test, p-value = 0.12).
In S. domestica seedlings (Figure S7c and d), survival analysis also showed significant differences across bacterial treatments (log-rank test, p-value = 0.017). Consortium F emerged as the most effective treatment with a HR of 0.29 (95% CI [0.11-0.77], p = 0.013), indicating a 71% reduction in risk compared to water. Pairwise comparisons confirmed that Consortium F significantly outperformed both the water control (pairwise log-rank test, p-value = 0.030) and its constituent strains S31 and S39 (pairwise log-rank test, p-value = 0.004 and 0.006, respectively). Strains S31, S39, and S48 alone did not significantly differ from the water control, with HR p-values all above 0.05. This suggests that the enhanced drought protection observed in S. domestica seedlings inoculated with Consortium F likely resulted from synergistic interactions among its component strains. These results contrast with those for Quercus Pubescens, highlighting species-specific responses to bacterial inoculation. No other consortia tested showed improved survival compared to water or TSB/100 controls (LRT, p-value = 0.48).
Prediction of Inoculant Efficacy for Drought Protection of Quercus and Sorbus Seedlings
Multinomial logistic regression model (MLM) is used to predict the relationship between one nominal dependent variable (with more than two categories) and one or more independent variables, which can be either categorical or continuous (https://resources.nu.edu/statsresources/Multinomiallogistic). We used MLM to model the relationship between the outcome of drought symptoms on trees, using the kinetics of symptoms, treatment type, and increments in stem height and diameter.
Quercus Pubescens
In Q. pubescens experiments with consortia A, B, D and H (Figure S8 A), the models showed that symptom kinetics and stem diameter increment were the most crucial factors in predicting the outcome of the inoculation of A, B and D (A, χ² = 486, df = 16, p-value < 0.001; B, χ² = 418.97, p-value < 0.001; D, χ² = 42.53, p-value < 0.001). Height increment showed no significant impact (χ² = 2.192, p-value = 0.335). Treatment with Consortium D significantly reduced the occurrence of partially affected phenotypes from 24.7% (± 3.5%) to 14.2% (± 3.1%) (p-value = 0.033), but showed no significant effect on severe symptoms.
When modelling the inoculation with single strains and Consortium F (Figure S8 B), neither diameter increment (χ² = 2.182, p-value = 0.336) nor height (χ² = 0.0524, p-value = 0.974) were significant according to the likelihood ratio test. Only time remained highly significant (χ² = 143.277, p < 0.001), with treatment effects showing modest impact (χ² = 20.741, p-value = 0.023). This suggests that growth parameters are not predictive variables in the case of moderately effective bacterial treatments, while temporal progression of symptoms remains the dominant factor.
Sorbus Domestica
In S. domestica treated with consortia A, B, D and H (Figure S8 C), the MLM showed that diameter increment, height increment, treatment, and time significantly predicted symptom development (χ² = 146, df = 21, p < 0.001), though with a moderate 11.6% explanatory power (R²N= 0.116). When treated with consortia F and single strains S31, S39 and s48 (Figure S8 D), the predictor variables stayed significant, although with a lower fit (χ² = 154, df = 21, p < 0.001, R²N= 0.121).
Model predicted that consortia A (p-value = 0.001) and D (p-value = 0.04) impacted the development of the partially affected symptoms and consortia F (p-value = 0.008) impacted the development of very affected symptoms. The predicted probability of developing partial symptoms under drought conditions was 24.2 ± 3.5 % in the water control group, which decreased to 12 ± 3% with consortium D inoculation and further declined to 3 ± 1.6 % with consortium A inoculation. The inoculation with the consortium F decreased the probability of the very affected symptoms development, from 14.1 ± 2.8 % to 4 ± 1.8 %.
3. Discussion
This study investigated nature-based solutions using tailored bacterial consortia (SynComs) derived from Mediterranean forest soils to enhance drought tolerance in tree seedlings, particularly during the critical establishment phase. We used a soil that has been subjected to recurrent amplified drought conditions since 2012, through the exclusion of spring and summer rainfalls reducing annual rainfall by approximately 35%—mimicking the climate projections for 2050 and 2100 for RCP 8.5 and 4.5, respectively [51]. The concept of using bacterial inoculants that have already experienced drought is rooted in the 'drought legacy effect' [52,53]. Through repeated exposure to water deficit, these bacterial communities potentially develop or refine mechanisms for drought tolerance and effective host interaction in harsh environments) [54].
From Soil Niches to Synthetic Communities (SynCom)
Bacterial communities associated with plant roots display distinct compositional and functional traits across soil compartments, shaped by gradients in plant influence, microbial interactions, and environmental conditions. By sampling multiple compartments surrounding the root system—Roots, Root-Adhering Soil (RAS), non-Root-Adhering Soil (non-RAS), and Root-Induced Macroaggregates (RIM)—at two depths (topsoil: 0–5 cm, without litter, and subsoil: 5–10 cm), and culturing on a carbon-gradient medium (serial dilutions of Tryptic soil agar, TSA), we isolated bacterial strains spanning a range of metabolic strategies and nutrient requirements. Although both soil layers yielded an equal number of isolates, the subsoil (5–10 cm)—which retains significantly more moisture under drought (water content at 10 cm depth was 100 times higher than at the surface)—provided the majority (84%) of high-performing strains. This depth is known to promote deeper rooting and greater exudation [55], suggesting that it represents a particularly productive niche for isolating drought-alleviating plant growth-promoting bacteria (PGPBs).
The compartment Roots yielded Peribacillus simplex and Pantoea pleuroti, both recovered from the richest culture medium used in this study (TSA/10), highlighting their adaptation to carbon-rich microenvironments at the root interface. P. simplex is a versatile bacterium primarily isolated from soil and crop rhizosphere—where it exhibits plant-growth-promoting and biocontrol properties. P. simplex is also found in harsh environments such as arid soils and stratospheric air samples (reviewed in Manetsberger et al. 2023) [56]. P. pleuroti, associated with Pleurotus mushroom fruiting bodies, is mainly saprotrophic and occasionally pathogenic to its fungal host [57]. These isolates reflect differing ecological niches and carbon usage profiles, with P. simplex demonstrating broader metabolic flexibility ([58], while P. pleuroti requirements are inferred from related species [59].
In RAS, Pseudomonas spp. dominated the culturable bacterial community (Dragone et al., 2024) [60]. Several close relatives from our Pseudomonas collection have been previously isolated from carbon-poor or recalcitrant niches. P. migulae was recovered from mineral water [61] and from the endophytic root tissues of lodgepole pine (Pinus contorta) growing in unreclaimed mining sites in British Columbia [62]. Similarly, P. mandelii and P. lini were isolated from the stem and needles, respectively, of the same pine species [62]. These findings highlight the niche-specific adaptations of Pseudomonas strains, while shared traits—such as exopolymer production (this study) and the ability to degrade complex substrates—underscore the remarkable metabolic diversity of this bacterial genus.
From RIM, we isolated P. lini and P. simplex, both confirmed strong exopolysaccharide (EPS) producers. EPS stabilizes macroaggregates and supports biofilm formation [63,64]. These EPS-producing strains contribute to rhizosheath formation and drought tolerance.
In non-RAS, Caballeronia glathei and P. mandelii were isolated. C. glathei (formerly Burkholderia [65]) is commonly isolated from bulk soil, rhizosphere soil, and contaminated environments such as wastewater sludge. C. glathei has been isolated from acid lateritic relicts in Germany [66] is metabolically versatile, nitrogen-fixing, and tolerant of contaminants [67]. P. mandelii exhibits traits such as polyhydroxybutyrate accumulation under nutrient stress [68], aiding survival and pollutant degradation. This trait is considered a survival strategy, enabling the bacterium to store excess carbon during nutrient scarcity and contributing to the degradation of environmental pollutants [68].
Although EPS supports plant growth by enhancing moisture retention, ion immobilization, and microbial colonization, only 5.8% of our isolates formed mucoid colonies—a common proxy for EPS production. EPS synthesis is typically triggered by osmotic stress and a high carbon-to-nitrogen (C/N) ratio [69]. EPS-producing bacteria contribute to the creation of microenvironments that retain water, immobilize harmful ions, and promote a healthier rhizosphere [70,71,72,73]. They also facilitate soil aggregation by interacting with clay particles, thus helping to maintain the soil’s mechanical and physical properties [74,75]. Santaella et al. [76] demonstrated that Rhizobium sp. YAS34, an EPS-producing strain, promoted the formation of stable microaggregates composed of EPS, mineral particles, and organic matter. These EPS-rich biofilms were mainly localized at the root base, lateral root emergence zones, and root hairs. In contrast, no EPS accumulation was observed at the root apex, where mucilage and root cap-derived cells typically form the first interface with soil [77]. This tri-partite interaction between EPS, microorganisms, and the host plant enhances drought tolerance by improving soil structure, protecting roots, and mitigating environmental stress—ultimately supporting more efficient water uptake under dry conditions [73].
We also assessed classic PGPB traits including auxin and ABA biosynthesis, siderophore production, ACC deaminase activity, and tryptophan dependency. IAA synthesis was strictly tryptophan-dependent, implying reliance on root exudates. Siderophores enhance iron acquisition and can trigger plant immune responses [78]. The interest in ABA-producing bacteria relies on their ability of drought signalling to the plant host [79,80]. We identified P. usmongensis s28 as an ABA-producing bacteria (0.39 µgL-1 under 0.5M NaCl), in the range of Bacillus amyloliquefaciens RWL-(0.32 ng mL-1) [79] , Bacillus pumilus (45 pmol mL-1 [80] and, Bradirhizobium japonicum USDA 110 (0.019 µgmL-1[81]).
Altogether, over 55% of our isolates displayed at least four key PGPB traits, and 8 of the final 12 strains retained 75% of nine tested functions.
The MCA revealed distinct ecological and functional structuring within the bacterial isolate pool, highlighting key factors that drive strain differentiation. Spatial variables—soil compartment, isolation medium, and sampling depth—were the major contributors to the first principal axis, emphasizing the role of niche-specific adaptation in shaping microbial communities. In contrast, functional traits like EPS production, IAA biosynthesis, and salt tolerance—known to support drought tolerance—were primarily associated with the second axis. The independence of these axes suggests that ecological origin and functional capacity represent distinct, yet complementary, dimensions of microbial variation.
This insight guided our SynCom design strategy. Rather than relying solely on individual trait performance, we prioritized strains that not only exhibited beneficial traits but also thrived in different soil compartments. By assembling four-strain SynComs reflecting the spatial structure of the rhizosphere, we aimed to reduce niche overlap and microbial competition, fostering functional complementarity and cooperation. Recognizing the limits of trait-based prediction (Shayanthan et al., 2022), we used a multivariate selection approach to reduce the number of candidate SynComs while maintaining both ecological and functional diversity. While this pragmatic strategy proved effective, it may have excluded other relevant microbial traits—such as phenol oxidase activity [82], broad substrate metabolism [83], or additional plant-interactive capabilities [84].
In summary, combining spatially structured sampling, functional trait screening, and ecologically informed SynCom design [29] enabled us to identify bacterial consortia with strong potential to alleviate drought stress. However, context-dependent validation remains essential, as microbial performance cannot be fully predicted from traits or origin alone [85].
Stronger Together? How Weak Biofilm Formers Drive Collective Performance
Biofilm formation, a key adaptive trait under drought conditions, was used to associate cooperative strains. Although not the sole mechanism of plant growth promotion, biofilm formation enhances root colonization, microbial survival, and stress signalling, ultimately amplifying the effects of traditional PGPB traits such as nutrient solubilization and phytohormone production [86,87,88]. Biofilms serve as structured environments where cooperative metabolic exchanges can be intensified, although resource competition may favour dominant strains, potentially compromising community cohesion [49,89].
Among the 12 bacterial strains tested, seven—mostly from root-associated compartments—formed biofilms individually. P. migulae s31 and P. mandelii s40 were the most prolific producers as single strains. Paradoxically, some of the highest-performing consortia for collective biofilm production included strains that were weak individual producers (e.g., s12, s57, s59, s48). In contrast, strong individual producers such as s31 and s40 often suppressed biofilm formation in consortia, likely due to competitive dominance. This pattern illustrates a trade-off: strains optimized for individual performance may inhibit collective function, while weak biofilm formers may thrive in cooperative networks by benefiting from partner-derived EPS or metabolic scaffolding [90,91,92].
We selected twelve individual strains and eight representative SynComs for further testing on plants, grouped by their biofilm production levels: high (A, B, C, D), low (E and F), and intermediate (G and H). These were screened for their drought-alleviating and plant-growth-promoting capacities, allowing us to evaluate how biofilm-forming ability translates to functional performance in planta.
Teaming Up Underground: Optimizing SynCom by Matching Microbial Skills to Root Strategies
Root strategies have traditionally been described along a fast–slow continuum: fast (acquisitive) roots are thin, short-lived, and specialized for rapid nutrient uptake, whereas slow (conservative) roots are thicker, longer-lived, and adapted to low-resource or stressful environments [31]. More recent frameworks propose a refined two-dimensional model—the “root economics space” (RES)—which integrates structural carbon investment and links root-associated microbial functions with root strategies [32,34,44,93]. Within this model, plants adopt two main strategies for acquiring soil resources: either through the development of long, fine roots with high specific root length—Do It Yourself (DIY) strategy—maximizing direct uptake, or by investing in thicker roots that support microbial symbioses and interactions, thereby outsourcing nutrient acquisition [32,94].
To explore these root strategies and their microbial partnerships, we selected Arabidopsis thaliana (Brassicaceae), Quercus pubescens, and Sorbus domestica. Arabidopsis thaliana is a non-mycorrhizal model plant [95] with fast, acquisitive root traits and genetic tractability [32,96]. The lacks of fungal symbiosis, places it at the “do-it-yourself” end of the RES gradient. Quercus pubescens, an ectomycorrhizal Mediterranean oak, exhibits a conservative root strategy featuring deep, woody roots and drought-adaptive traits like enhanced root tissue density and hydraulic conductance [97,98]. Under drought, it employs a multi-dimensional strategy that includes high root hydraulic conductance to maintain leaf water potential, coupled with moderately regulated stomatal conductance [98,99]. Sorbus domestica develops a shallow, highly ramified root system (https://images.wur.nl/digital/collection/coll13/id/1318/; EUFORGEN; [100]) and adopts a drought-avoidance strategy by accelerating root growth and secondary root development, improving water capture across a larger soil area [101,102,103]. Both Q. pubescens and S. domestica are well adapted to Mediterranean climates and considered future species for forest restoration in Europe under climate change.
Among tested strains, P. silesiensis s39 promoted growth in A. thaliana, whereas P. mandelii s54 inhibited primary root development—possibly due to cyanide production, which can suppress auxin signaling [104]. Nevertheless, under regular conditions, none of the SynComs significantly improved A. thaliana, suggesting its “DIY” strategy may limit benefits from microbial partnerships in non-stressful environments.
For Q. pubescens and S. domestica seedlings, a practical pre-plant soil inoculation method was used differing from the one used for Arabidopsis thaliana and from Tiepo et al. [17,18] and Khosravi et al. 2025 [19]. Pre-plant soil inoculation before outplanting offers a practical advantage, as it can be conducted in the nursery and is more adaptable than direct seed or acorn inoculation. Under drought, two SynComs (F and B) significantly reduced stress symptoms in these tree species. SynCom F, associated with A. thaliana development, lowered the hazard ratio (HR) of drought symptoms by 71% in S. domestica, while SynCom B, linked to enhanced biofilm formation, seed germination, and A. thaliana vigor, reduced the HR by 47% in Q. pubescens. Critically, none of the single-strain treatments matched the efficacy of the full communities, emphasizing the importance of synergistic interactions [37].
Notably, 75% of strains in SynComs B and F were isolated from a medium with moderate carbon content (TSA/10), and 80% from the 5-10 cm depth, suggesting functional potential is not limited to more oligotrophic reservoir or top soil. Also, half of SynCom B and a quarter of SynCom F strains came from non-root-adhering soil, including macroaggregates— a zone of intense microbial, fungal, and root interaction—highlighting overlooked microbial niches with cooperative potential.
Network analyses (Figure S8) revealed SynCom B to be a local optimum for Q. pubescens, where minor compositional changes diminished its performance. A key player was strain P. mandelii s54, detrimental in A. thaliana yet beneficial in Q. pubescens, highlighting the host dependency of microbial function. Likewise, SynCom F proved uniquely effective in S. domestica through the combination of s12, s31, s48, and s28. The ineffectiveness of s31 alone underscores the non-additive nature of SynCom performance.
Overall, SynCom efficacy appears to hinge on how well microbial functions align with a plant’s root strategy (Figure 6). In thin, fast-acquisitive roots like those of S. domestica, SynComs boosting nutrient uptake (e.g., iron solubilization or phytohormone production) are especially advantageous. For conservative, coarser roots, promoting “hyphal-like” biofilm networks or soil-structuring EPS, may be more beneficial. These host-specific interactions suggest that tailoring SynCom design to each plant’s ecological and morphological traits is crucial for improving drought tolerance and resource use efficiency—especially in stress-prone conditions.
Beyond Survival: Predictive Insights into SynCom Function and Drought Tolerance in Trees
Our predictive modeling extended the survival analysis, offering deeper insights into SynCom impacts on drought symptom progression. SynCom D reduced the proportion of Q. pubescens seedlings with partial drought symptoms by 10.5% compared to the water control, while SynCom F significantly alleviated symptoms in S. domestica. Although models also predicted mitigating effects for SynComs A and D in oak, survival analysis showed only marginal changes in HR—29% for A (p = 0.051) and 6% for D (p = 0.725).
Bacterial single strain or SynCom treatment identity consistently emerged as a strong predictor, confirming its influence on symptom development. Time was also significant, reflecting drought symptom dynamics. In contrast, changes in plant height and diameter increments were not consistently predictive, suggesting that unmeasured anatomical or physiological traits may better explain tolerance patterns.
In Q. pubescens, greater stem diameter increment was linked to lower likelihood of severe symptoms, indicating better turgor or growth under drought (Figure 7A). This aligns with literature showing that anatomical traits, such as xylem size and stomatal density, influence drought [105].
Interestingly, S. domestica showed the opposite pattern—greater stem diameter was associated with more partial symptoms (Figure 7B). As an anisohydric species with strong drought recovery [106], it may maintain growth during stress, increasing its risk of hydraulic failure. We observed that S. domestica, often shed basal leaves thereby reducing total leaf area, while sustaining photosynthesis in upper canopy leaves, suggesting a resource allocation towards a higher use efficiency at the whole plant level.
Our findings reinforce that SynComs are generally more effective than single strains in improving drought tolerance. Stem diameter increment, particularly in inoculated seedlings, emerged as a promising trait linked to enhanced drought protection. These results suggest it could serve as a phenotypic marker for SynCom efficacy.
As George Box noted, “all models are wrong, but some are useful.” While informative, our predictive models showed modest R² values, highlighting a need to refine them by incorporating additional variables. Future work should leverage high-throughput phenotyping and multi-omics datasets—metagenomics, proteomics, metabolomics—combined with machine learning [29,107] to improve predictive power and guide SynCom design more effectively.
Overall, the effective SynComs can potentially be used to prevent Q. pubescens and S. domestica from experiencing drought stress, after further investigation and field experiments.
4. Materials and Methods
Soil Sampling
Soil was sampled at O3HP (Oak Observatory at the OHP) experimental facility which is part of the European and French research infrastructure AnaEE-ERIC and AnaEE-France, and is located at Saint-Michel-l’Observatoire in Alpes de Haute-Provence, France (43.9352° N, 5.710667° E). This facility runs a rain exclusion system since 2012 above 300 m² of canopy to simulate an extended summer and an additional spring drought, cumulating to about 35% reduction in annual precipitation, as in line with predictions by climate models [1], in order to create an enhanced drought as compared to natural drought [38]. Exclusion of full rain events raises the number of dry days by about 30 per year [108]. The dominant tree species is Quercus pubescens (downy oak) mixed with co-dominant Acer monspessulanum L. (Montpellier maple), representing about 75% and 25% respectively of the litter production and standing biomass. The soil is a calcosol (leptosol with a mollic horizon), consisting of two distinct horizons [39]: a mollic horizon (rich in organic matter, approximately 0-5 cm deep) and a leptosol horizon (bedrock with little organic matter, rich in clay, approximately 5-10 cm deep).
Groups of trees were used as focal points for subsequent soil sampling. Soil was collected using a stainless steel core (5 cm diameter) from the rain exclusion zone on 22 October 2021. At the time of sampling, the soil had not received any direct rainfall since mid-June and volumetric soil moisture mean had been 0.136 L L-1 for 15 weeks since July 2021, and 13.34 L L-1 at 10 cm depth. The litter layer was discarded before coring and samples were taken from 0-5 cm (upper layer) and 5-10 cm (lower layer) depths. Samples were transported to the laboratory in refrigerated boxes and immediately stored at 4°C. Soils were sieved at 1.2 mm mesh to separate non-root adhering soil (non-RAS) from roots (RT), root-adhering soil (RAS), and root-induced macroaggregates (RIM) (Figure 1b). Roots were vortexed in sterile water, to separate the RAS.
Strain Isolation and Growth
One gram of soil or 100 mg of grinded fresh roots were used to prepare the suspensions in 10 mL or 1ml of sterile distilled water, respectively. The suspensions were homogenised by vortexing and 100 μL were used for dilution plating on a non-selective Tryptic Soy Both (TSB, Soybean-Casein Digest Broth) diluted 10x, 20x and 50x times and solidified with 3 gL-1 of Agar (TSA). Plates were incubated at 30°C for 48 hours. Morphologically different isolated single colonies were picked up and re-streaked to fresh ten-fold diluted TSA plates and incubated in a similar manner. The purified cultures were stored on ten-fold diluted TSA medium at 4°C in a dark chamber.
Strain growth parameters in liquid 10-fold diluted TSB were measured using 96-well plates incubated at 30 °C and 3 mm of orbital amplitude (around 300 rpm) for 24 hours. Optical density (OD) at 600 nm was recorded every 15 minutes using an Infinite® M1000 plate reader (Tecan). Two key growth parameters were extracted: the time to reach half of the carrying capacity (t_mid), which corresponds to the inflection point of the growth curve (Figure S2), and the minimum doubling time (t_gen), representing the fastest generation rate. Both parameters were calculated as indicators of growth performance (Allen & Waclaw, 2018).
Strain Screening for Specific Traits
Exopolysaccharide (EPS) production capacity was estimated by transferring the single colonies to 10-fold diluted TSA supplemented with 20 gL-1 of glucose, followed by incubation at 30°C for 48 hours in the dark. A quantitative estimate of exopolysaccharide production was made from colonies with a strong mucoid phenotype [76].The dry mass was used to estimate the EPS production capacity.
Strains were screened for Plant-Promoting Growth Bacteria (PGPB) traits as siderophore production using casamino acid (CAS) and overlaid CAS (O-CAS) assays [108,109,110], plant hormone production, such as auxins [111,112,113]. Isolates with osmotic tolerance were screened at osmotic potential of −0.51, −1.23, and −2.56 MPa in liquid phase and -0.64, -1.12, -2.28 MPa in solid phase using Polyethylene glycol (PEG 6000) [114,115]. Salt tolerant strains were selected in liquid culture assays with NaCl 0.5, 1, 1.5, 2M according to [115,116].
Ability to Form Biofilms
To test for synergistic biofilm production among bacterial strains, we conducted in vitro assays using both single strains and 4-strain consortia derived from various soil compartments. Biofilm formation was assessed using the 96-well peg lid system (MBEC Assay® Biofilm Inoculator, Innovotec®), incubated for 24h at 30°C without shaking, based on protocols from [117,118]. After incubation, biofilms were stained with crystal violet, washed, and the dye solubilized in ethanol for OD590 nm measurement using a microplate reader (Infinite® M1000, Tecan). Synergy was evaluated by comparing the OD590 of each consortium to the highest OD590 of its constituent strains. Ratios ≥ 1.9 indicated synergy; ratios < 0.7 suggested antagonism; intermediate values reflected minor differences. Thresholds were set empirically (Figure S9) to select a manageable number of consortia for further testing.
Strain Identification
DNA was obtained by heat shock of a single clone diluted in 20 µL steril milliQ water. The full-length 16S rRNA gene was amplified using bacterial-specific primers 27F (5′-AGAGTTTGATCCTGGCTCG-3') and 1492R (5'-GTTACCTTGTTACGACTT-3'). Amplification was performed using the GoTaq® Flexi DNA Polymerase (Promega, USA, under the following conditions: initial denaturation at 95 °C for 2 min followed by 34 cycles, each set at 95 °C for 30 s, 53 °C for 30 s, and 72 °C for 1:30 min, with a final elongation step at 72 °C for 5 min. Amplified PCR products were transferred to GENEWIZ (France) for two ways Sanger sequencing. The National Centre for Biotechnology Information (NCBI)'s Basic Local Alignment Search Tool (BLAST) software (http://blast.ncbi.nlm.nih.gov) algorithm was used to search for homology using an aligned contiguous consensus sequence of the 16S rRNA gene.
Test of Single Strains and Consortia on Arabidopsis thaliana
Arabidopsis thaliana seeds (Col-0 ecotype) were sown in square Petri dishes (120 mm x 120 mm, Greiner) containing solid medium (two-fold diluted Hoagland with Phytagel®) and inoculated with 10 µL of either bacterial suspensions (10⁶ cells mL-1) or sterile Milli-Q water, following Santaella et al. [76]. Plates were incubated in growth chambers (25 °C/20 °C day/night, 16 h photoperiod, PAR 100 µmol m⁻² s⁻¹, 40% relative humidity) for 3 weeks.
Germination was assessed on day 4, and root development was monitored every second weekday. Seedlings were categorized as germinated, developed, or well-developed (rosette >0.7 cm, >7 leaves, >10 lateral roots). Additionally, the number and total length of first-order lateral roots per cm of primary root were recorded.
Greenhouse Experiments on Trees
Acorns of Q. pubescens Willd. (QPU751), collected in southeastern France (Pignans, Var), and seeds of Sorbus domestica (SDO900 FR), collected throughout mainland France, were provided by the ONF Sècherie de la Joux (France).
The experiment ran from January to November 2023 at the ONF-PNRGF (Pôle National des Ressources Génétiques Forestières) in Cadarache, France. The bacterial strains and consortia were tested in batches of six treatments on Q. pubescens and S. domestica, in two separate trials, each batch including control treatments. A randomized complete block design was then employed to account for potential environmental heterogeneity within the greenhouse. Each block contained all treatments, including controls, enabling within-block comparisons and minimizing the effects of spatial variation (Figure S11).
Seven-month-old Q. pubescens seedlings and five-month-old S. domestica seedlings were grown in 1.4 L pots filled with potting soil NFU 44_551 (Premier Tech, France). This substrate consisted of equal parts white peat and a granular organic fraction (PM bark), amended with 2 kg m-³ of Osmocote Exact 8/9 (ICL Growing Solutions, 15-9-11) at 80% field capacity (FC). In August 2023, the seedlings were inoculated with a 100 mL suspension of either a single strain or a four-strain consortium. Bacterial cells were grown in 10-fold diluted TSB medium and diluted tenfold to achieve a final concentration of 10⁷ cells mL-1. MilliQ water and one hundred-fold diluted TSB (TSB/100) were used as controls (Figure S11).
Five weeks after inoculation for Q. pubescens and four weeks for S. domestica, half of the plants were subjected to drought, while the rest remained well-watered. Drought was induced by gradually reducing soil water content from 80% to 40 ± 5% field capacity (FC), determined gravimetrically. Plant height and trunk diameter were monitored throughout. Drought symptoms—including dry leaves and chlorosis—were assessed under both control (80% FC) and drought (40% FC) conditions. Severity was visually categorized as unaffected (green), partially affected (half symptomatic), or severely affected (fully dry). Controls helped distinguish stress symptoms from seasonal senescence (Figure S6). Each drought-treatment group included 40 replicates for water control, 32 for TSB/100, and 30 for inoculated treatments.
Statistical Analyses
All statistical analyses were done with a significance level of alpha=0.05. Hazard ratios were reported with their corresponding 95% confidence interval and p-value.
Multivariate Analyses-Driven Selection of the Best Candidate Strains for SynComs Selection
Multiple correspondence analysis (MCA), an extension of correspondence analysis (Abdi and Valentin, 2007), was used to assess relationships among several ordinal variables: PGPB traits, EPS production, tolerance to osmotic (PEG 10, PEG 30) and saline (NaCl 0.5–1.5 M) stress, soil depth (0–5 cm, 5–10 cm), isolation compartment (Root, RAS, non-RAS, RIM), and growth medium (TSA/10, TSA/20, TSA/50). Numerical variables were converted to ordinal scales based on performance ranges.
Principal Component Analysis (PCA) was also conducted to explore associations between SynComs/single strains and plant traits, including germination rate, post-germination development, root architecture, and biofilm formation. Data were normalized prior to PCA.
Statistical Analyses on Plant Symptoms and Growth Parameters
Bacterial strain growth parameters were analyzed using the R package Growthcurver [119].
The proportion of germination and developed seedlings of the experiment with single strains and with consortia (synthetic communities) were analyzed with the Chi-Squared test followed by the analysis of the standardized adjusted residuals in RStudio (4.4.2, R Core Team 2024) using the native function chisq.test and the packages dplyr (v1.1.4) [120] and rstatix (v0.7.2) [121].
Principal component analysis (PCA) and Multiple correspondence analysis (MCA) were performed using FactoMineR (v2.7) R package [122].
Survival Models
To evaluate the impact of bacterial inoculants on plant survival under drought, we combined several survival analysis methods. Kaplan–Meier survival curves illustrated the progression of survival probabilities across treatments, and the log-rank test assessed statistical differences between them.
To account for time-dependent variables, we used the Cox proportional hazards model, which estimated the relative mortality risk (hazard ratio, HR) for each treatment, adjusting for covariates like plant height and stem diameter. Hazard Ratio captures changes in risk over time. Multivariable survival analysis helped refine treatment-specific HR estimates by incorporating continuous predictors. The cumulative hazard function described total mortality risk up to a given point, highlighting how risk accumulates over time.
Multinomial Logistic Regression
Multinomial logistic regression was used to assess the relationship between continuous variables (height and diameter increment, time of symptom observation) and the categorical variable Inoculant (SynCom and/or single strain) under drought conditions. The goal was to predict symptom development based on observed growth parameters after inoculation. Model fit was evaluated using the Akaike Information Criterion (AIC), Nagelkerke’s R² (R2N), and p-values. Relationships were expressed via odds ratios, log odds estimates, and marginal means. Analyses were performed with the R package survminer (v0.5.0) [123] and Jamovi (v2.3) [124,125] for additional calculations and visualizations.
Use of GenAI
GenAI was used in the paper, to draft the network in Figure S8, for literature mining and to improve the text readability.
5. Conclusion
Our findings underscore the potential of designing microbial consortia that are ecologically informed and functionally aligned with plant strategies to enhance drought tolerance. Rather than relying on broad-spectrum inoculants, tailoring SynComs to the specific traits of host species and their rhizosphere environment—particularly soil depth and compartment—emerges as a promising approach. This work reinforces the idea that microbial effectiveness is context-dependent, and that trait-based microbial selection holds value for sustainable forest restoration under climate stress. Future applications will benefit from integrating microbial and plant functional traits to guide the formulation of next-generation bioinoculants for forest restoration.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: K-means clustering of bacterial strain tolerance to osmotic stress; Figure. S2: Growth Dynamics of Selected Bacterial Strains. Figure S3: Biofilm formation by individual strains and four-strain consortia. Figure S4: Germination and seedling development of Arabidopsis thaliana under inoculation with single strains (assay N°1; Figure S5: Germination and seedling development of Arabidopsis thaliana under inoculation with single strains (assay N°2); Figure S6: Germination and seedling development of Arabidopsis thaliana under inoculation with synthetic communities (SynCom); Figure S7: Development of drought-related symptoms in plants; Figure S8: Estimated probabilities of symptom development under the most efficient synthetic communities for each tree species; Figure S9: Similarity network of synthetic communities based on shared strain composition; Figure S10: . Evaluation of biofilm synergy in synthetic consortia; Figure S11: Greenhouse experimental setup for tree seedling trials. Table S1: Bacterial strain consortia used in the biofilm assay; File S1: Strain traits. File S2: Syncom assembly.
Author Contributions
Conceptualization, C.S.; Methodology, C.S., J.R., M.d.C., B.L., I.M.R. and I.A.; Software, I.A.; Validation, C.S.; Formal Analysis, I.A. and C.S.; Investigation, I.A., M.F.H., C.S., B.L. and I.M.R.; Resources, C.S., I.M.R.; J.R., M.d.C., B.L.; Data Curation, I.A., and M.F.H.; Writing – Original Draft Preparation, I.A, M.F.H., and C.S.; Writing – Review & Editing, C.S., H.C.O., and I.M.R.; Visualization, I.A., M.F.H. and C.S.; Supervision, C.S.; Project Administration, C.S.; Funding Acquisition, H.C.O., C.S., and I.M.R.”
Funding
This research was funded through the 2019-2020 BiodivERsA+ joint call for research proposals, under the BiodivClim ERA-Net COFUND programme, and with the funding organisations Fundação Araucária/Secretaria de Estado da Ciência, Tecnologia e Ensino Superior do Paraná (NAPI Biodiversidade), FAPESP (Brazil), French National Research Agency (ANR) under grant number ANR-20-EBI5-0008-07, and Federal Ministry of Education and Research (Germany). I.A. received additional funding from the HORIZON EUROPE INFRA-2022-TECH project ‘PHENET’, grant number 101094587. This study was partly funded by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES), grant number 88887.712065/2022-00, for the postdoctoral fellowship of MFH.
Data Availability Statement
Data will be made available on request.
Acknowledgments
We would like to express our sincere gratitude to Sylvain Fochesato and Gilles de Luca for their assistance with strain plating and tree inoculations. Their support was greatly appreciated. Special thanks to Patrice Brahic and to the ONF (Office National des Forêts) for their expertise and for providing the facilities to grow our tree seedlings. Hudge thanks to Agate Gueler, Bastian Charillat and Ugo Furet for their care and attention to the tree seedlings during their development. We acknowledge the use of the COOPERATE database, maintained by I.M.R. and Armand Rotereau.
Conflicts of Interest
The authors declare no conflict of interest.
References
- IPCC Climate Change 2022 – Impacts, Adaptation and Vulnerability: Working Group II Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; 1st ed.; Cambridge University Press, 2023; ISBN 978-1-00-932584-4.
- Senf, C.; Buras, A.; Zang, C.S.; Rammig, A.; Seidl, R. Excess Forest Mortality Is Consistently Linked to Drought across Europe. Nat Commun 2020, 11, 6200. [CrossRef]
- Beloiu, M.; Stahlmann, R.; Beierkuhnlein, C. Drought Impacts in Forest Canopy and Deciduous Tree Saplings in Central European Forests. Forest Ecology and Management 2022, 509, 120075. [CrossRef]
- Gazol, A.; Camarero, J.J. Compound Climate Events Increase Tree Drought Mortality across European Forests. Science of The Total Environment 2022, 816, 151604. [CrossRef]
- Bevacqua, E.; Rakovec, O.; Schumacher, D.; Kumar, R.; Thober, S.; Samaniego, L.; Seneviratne, S.; Zscheischler, J. Direct and Lagged Climate Change Effects Strongly Intensified the Widespread 2022 European Drought 2024.
- Dumroese, R.K.; Balachowski, J.A.; Flores, D.; Horning, M.E. Reforestation to Mitigate Changes to Climate: More than Just Planting Seedlings. In In: XV World Forestry Congress: Building a Green, Healthy, and Resilient Future with Forests.; 2-6 May 2022; Coex, Seoul, Republic of Korea. Food and Agriculture Organization of the United Nations; Korean Forest Service 2022, 7.
- Knutzen, F.; Averbeck, P.; Barrasso, C.; Bouwer, L.M.; Gardiner, B.; Grünzweig, J.M.; Hänel, S.; Haustein, K.; Johannessen, M.R.; Kollet, S.; et al. Impacts on and Damage to European Forests from the 2018–2022 Heat and Drought Events. Natural Hazards and Earth System Sciences 2025, 25, 77–117. [CrossRef]
- Puértolas, J.; Villar-Salvador, P.; Andivia, E.; Ahuja, I.; Cocozza, C.; Cvjetković, B.; Devetaković, J.; Diez, J.J.; Fløistad, I.S.; Ganatsas, P.; et al. Die-Hard Seedlings. A Global Meta-Analysis on the Factors Determining the Effectiveness of Drought Hardening on Growth and Survival of Forest Plantations. Forest Ecology and Management 2024, 572, 122300. [CrossRef]
- Ait-El-Mokhtar, M.; Meddich, A.; Baslam, M. Plant-Microbiome Interactions under Drought—Insights from the Molecular Machinist’s Toolbox. Frontiers in Sustainable Food Systems. 2023, 7, 1253735.
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant physiology 2020, 184, 39–52. [CrossRef]
- Zahar Haichar, F.; Santaella, C.; Heulin, T.; Achouak, W. Root Exudates Mediated Interactions Belowground. Soil Biology and Biochemistry 2014, 77, 69–80. [CrossRef]
- Hong, Y.; Zhou, Q.; Hao, Y.; Huang, A.C. Crafting the Plant Root Metabolome for Improved Microbe-assisted Stress Resilience. New Phytologist 2022, 234, 1945–1950. [CrossRef]
- Rolfe, S.A.; Griffiths, J.; Ton, J. Crying out for Help with Root Exudates: Adaptive Mechanisms by Which Stressed Plants Assemble Health-Promoting Soil Microbiomes. Current opinion in microbiology 2019, 49, 73–82. [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of Plant Growth Promoting Rhizobacteria (PGPRs) with Multiple Plant Growth Promoting Traits in Stress Agriculture: Action Mechanisms and Future Prospects. Ecotox. Environ. Safe. 2018, 156, 225–246. [CrossRef]
- Iqbal, S.; Wang, X.; Mubeen, I.; Kamran, M.; Kanwal, I.; Diaz, G.A.; Abbas, A.; Parveen, A.; Atiq, M.N.; Alshaya, H.; et al. Phytohormones Trigger Drought Tolerance in Crop Plants: Outlook and Future Perspectives. Front. Plant Sci. 2022, 12, 799318. [CrossRef]
- Campos, E.; Pereira, A.; Aleksieienko, I.; Carmo, G.; Gohari, G.; Santaella, C.; Fraceto, L.; Oliveira, H. Encapsulated Plant Growth Regulators and Associative Microorganisms: Nature-Based Solutions to Mitigate the Effects of Climate Change on Plants. Plant Science 2023, 331, 111688. [CrossRef]
- Tiepo, A.N.; Constantino, L.V.; Madeira, T.B.; Gonçalves, L.S.A.; Pimenta, J.A.; Bianchini, E.; de Oliveira, A.L.M.; Oliveira, H.C.; Stolf-Moreira, R. Plant Growth-Promoting Bacteria Improve Leaf Antioxidant Metabolism of Drought-Stressed Neotropical Trees. Planta 2020, 251, 83. [CrossRef]
- Tiepo, A.N.; Hertel, M.F.; Rocha, S.S.; Calzavara, A.K.; De Oliveira, A.L.M.; Pimenta, J.A.; Oliveira, H.C.; Bianchini, E.; Stolf-Moreira, R. Enhanced Drought Tolerance in Seedlings of Neotropical Tree Species Inoculated with Plant Growth-Promoting Bacteria. Plant Physiology and Biochemistry 2018, 130, 277–288. [CrossRef]
- Khosravi, M.; Heydari, M.; Alikhani, H.A.; Arani, A.M.; Guidi, L.; Bernard, P. Mitigating Negative Impacts of Drought on Oak Seedlings Performances through Plant Growth-Promoting Rhizobacteria. Journal of Environmental Management 2025, 375, 124163. [CrossRef]
- Gillespie, L.M.; Fromin, N.; Milcu, A.; Buatois, B.; Pontoizeau, C.; Hättenschwiler, S. Higher Tree Diversity Increases Soil Microbial Resistance to Drought. Communications biology 2020, 3, 377. [CrossRef]
- Poudel, M.; Mendes, R.; Costa, L.A.; Bueno, C.G.; Meng, Y.; Folimonova, S.Y.; Martins, S.J. The Role of Plant-Associated Bacteria, Fungi, and Viruses in Drought Stress Mitigation. Frontiers in microbiology 2021, 12, 743512. [CrossRef]
- Carballo-Sánchez, M.P.; Alarcón, A.; Pérez-Moreno, J.; Ferrera-Cerrato, R. Agricultural and Forestry Importance of Microorganism-Plant Symbioses: A Microbial Source for Biotechnological Innovations. Reviews in Agricultural Science 2022, 10, 344–355. [CrossRef]
- Santos, R.F.D.; Cruz, S.P.D.; Botelho, G.R.; Flores, A.V. Inoculation of Pinus Taeda Seedlings with Plant Growth-Promoting Rhizobacteria. Floresta e Ambiente 2018, 25, 20160056.
- Rawat, P.; Das, S.; Shankhdhar, D.; Shankhdhar, S.C. Phosphate-Solubilizing Microorganisms: Mechanism and Their Role in Phosphate Solubilization and Uptake. J Soil Sci Plant Nutr 2021, 21, 49–68. [CrossRef]
- Khan, A.L. The Phytomicrobiome: Solving Plant Stress Tolerance under Climate Change. Frontiers in Plant Science 2023, 14, 1219366. [CrossRef]
- Liu, X.; Mei, S.; Salles, J.F. Inoculated Microbial Consortia Perform Better than Single Strains in Living Soil: A Meta-Analysis. Applied Soil Ecology 2023, 190, 105011. [CrossRef]
- Nicotra, D.; Mosca, A.; Dimaria, G.; Massimino, M.E.; Stabile, M.; Bella, E.; Catara, V. Mitigating Water Stress in Plants with Beneficial Bacteria: Effects on Growth and Rhizosphere Bacterial Communities. International Journal of Molecular Sciences 2025, 26, 1467. [CrossRef]
- Jing, J.; Garbeva, P.; Raaijmakers, J.M.; Medema, M.H. Strategies for Tailoring Functional Microbial Synthetic Communities. The ISME journal 2024, wrae049. [CrossRef]
- Mehlferber, E.C.; Arnault, G.; Joshi, B.; Partida-Martinez, L.P.; Patras, K.A.; Simonin, M.; Koskella, B. A Cross-Systems Primer for Synthetic Microbial Communities. Nature microbiology 2024, 9, 2765–2773. [CrossRef]
- Xu, X.; Dinesen, C.; Pioppi, A.; Kovács, Á.T.; Lozano-Andrade, C.N. Composing a Microbial Symphony: Synthetic Communities for Promoting Plant Growth; Trends in Microbiology, 2025;
- Stock, S.C.; Koester, M.; Boy, J.; Godoy, R.; Nájera, F.; Matus, F.; Dippold, M.A. Plant Carbon Investment in Fine Roots and Arbuscular Mycorrhizal Fungi: A Cross-Biome Study on Nutrient Acquisition Strategies. Science of the Total Environment 2021. [CrossRef]
- Bergmann, J.; Weigelt, A.; Plas, F.; Laughlin, D.C.; Kuyper, T.W.; Guerrero-Ramirez, N.; Valverde-Barrantes, O.J.; Bruelheide, H.; Freschet, G.T.; Iversen, C.M.; et al. The Fungal Collaboration Gradient Dominates the Root Economics Space in Plants. Science Advances 2020, 6, 3756. [CrossRef]
- Prada-Salcedo, L.D.; Goldmann, K.; Heintz-Buschart, A.; Reitz, T.; Wambsganss, J.; Bauhus, J.; Buscot, F. Fungal Guilds and Soil Functionality Respond to Tree Community Traits Rather than to Tree Diversity in European Forests. Molecular Ecology 2021, 30, 572–591. [CrossRef]
- Wu, R.; Zeng, X.; McCormack, M.L.; Fernandez, C.W.; Yang, Y.; Guo, H.; Chen, W. Linking Root-Associated Fungal and Bacterial Functions to Root Economics. bioRxiv 2023.
- Kou, X.; Han, W.; Kang, J. Responses of Root System Architecture to Water Stress at Multiple Levels: A Meta-Analysis of Trials under Controlled Conditions. Frontiers in Plant Science 2022, 13, 1085409. [CrossRef]
- Bauke, S.L.; Amelung, W.; Bol, R.; Brandt, L.; Brüggemann, N.; Kandeler, E.; Vereecken, H. Soil Water Status Shapes Nutrient Cycling in Agroecosystems from Micrometer to Landscape Scales. Journal of Plant Nutrition and Soil Science 2022, 185, 773–792. [CrossRef]
- Platt, T.G. Community Outcomes Depend on Cooperative Biofilm Structure. Proceedings of the National Academy of Sciences 2023, 120, 2221624120. [CrossRef]
- Laoué, J.; Havaux, M.; Ksas, B.; Orts, J.P.; Reiter, I.M.; Fernandez, C.; Ormeno, E. A Decade of Rain Exclusion in a Mediterranean Forest Reveals Trade-Offs of Leaf Chemical Defenses and Drought Legacy Effects. Scientific Reports 2024, 14, 24119. [CrossRef]
- Belviso, S.; Reiter, I.M.; Loubet, B.; Gros, V.; Lathière, J.; Montagne, D.; Delmotte, M.; Ramonet, M.; Kalogridis, C.; Lebegue, B.; et al. A Top-down Approach of Surface Carbonyl Sulfide Exchange by a Mediterranean Oak Forest Ecosystem in Southern France. Atmospheric Chemistry and Physics 2016, 16, 14909–14923. [CrossRef]
- Knights, H.E.; Jorrin, B.; Haskett, T.L.; Poole, P.S. Deciphering Bacterial Mechanisms of Root Colonization. Environmental Microbiology Reports 2021, 13, 428–444. [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, Variation, and Assembly of the Root-Associated Microbiomes of Rice. Proceedings of the National Academy of Sciences 2015, 112, 911–920. [CrossRef]
- Li, M.; Song, Z.; Li, Z.; Qiao, R.; Zhang, P.; Ding, C.; Xie, J.; Chen, Y.; Guo, H. Populus Root Exudates Are Associated with Rhizosphere Microbial Communities and Symbiotic Patterns. Frontiers in Microbiology 2022, 13, 1042944. [CrossRef]
- Baumert, V.L.; Vasilyeva, N.A.; Vladimirov, A.A.; Meier, I.C.; Kögel-Knabner, I.; Mueller, C.W. Root Exudates Induce Soil Macroaggregation Facilitated by Fungi in Subsoil. Frontiers in Environmental Science 2018, 6, 140. [CrossRef]
- Deng, Y.; Kong, W.; Zhang, X.; Zhu, Y.; Xie, T.; Chen, M.; Zhu, L.; Sun, J.; Zhang, Z.; Chen, C.; et al. Rhizosphere Microbial Community Enrichment Processes in Healthy and Diseased Plants: Implications of Soil Properties on Biomarkers. Frontiers in Microbiology 2024, 15, 1333076. [CrossRef]
- Leontidou, K.; Genitsaris, S.; Papadopoulou, A.; Kamou, N.; Bosmali, I.; Matsi, T.; Madesis, P.; Vokou, D.; Karamanoli, K.; Mellidou, I. Plant Growth Promoting Rhizobacteria Isolated from Halophytes and Drought-Tolerant Plants: Genomic Characterisation and Exploration of Phyto-Beneficial Traits. Sci Rep 2020, 10, 14857. [CrossRef]
- Petrillo, C.; Vitale, E.; Ambrosino, P.; Arena, C.; Isticato, R. Plant Growth-Promoting Bacterial Consortia as a Strategy to Alleviate Drought Stress in Spinacia Oleracea. Microorganisms 2022, 10, 1798. [CrossRef]
- Lin, Y.; Zhang, H.; Li, P.; Jin, J.; Li, Z. The Bacterial Consortia Promote Plant Growth and Secondary Metabolite Accumulation in Astragalus Mongholicus under Drought Stress. BMC Plant Biol 2022, 22, 475. [CrossRef]
- Benmrid, B.; Ghoulam, C.; Zeroual, Y.; Kouisni, L.; Bargaz, A. Bioinoculants as a Means of Increasing Crop Tolerance to Drought and Phosphorus Deficiency in Legume-Cereal Intercropping Systems. Commun Biol 2023, 6, 1–15. [CrossRef]
- Ren, D.; Madsen, J.S.; Sørensen, S.J.; Burmølle, M. High Prevalence of Biofilm Synergy among Bacterial Soil Isolates in Cocultures Indicates Bacterial Interspecific Cooperation. The ISME Journal 2015, 9, 81–89. [CrossRef]
- Ren, D.; Madsen, J.S.; Cruz-Perera, C.I.; Bergmark, L.; Sørensen, S.J.; Burmølle, M. High-Throughput Screening of Multispecies Biofilm Formation and Quantitative Pcr-Based Assessment of Individual Species Proportions, Useful for Exploring Interspecific Bacterial Interactions. Microb Ecol 2014, 68, 146–154. [CrossRef]
- Lange, M.A. Climate Change in the Mediterranean: Environmental Impacts and Extreme Events. IEMed Mediterranean Yearbook 2020, 224–229.
- Meisner, A.; Jacquiod, S.; Snoek, B.L.; ten Hooven, F.C.; van der Putten, W.H. Drought Legacy Effects on the Composition of Soil Fungal and Prokaryote Communities. Front. Microbiol. 2018, 9. [CrossRef]
- Vilonen, L.L.; Hoosein, S.; Smith, M.D.; Trivedi, P. Legacy Effects of Intensified Drought on the Soil Microbiome in a Mesic Grassland. Ecosphere 2023, 14, e4545. [CrossRef]
- Tang, Y.; Winterfeldt, S.; Brangarí, A.C.; Hicks, L.C.; Rousk, J. Higher Resistance and Resilience of Bacterial Growth to Drought in Grasslands with Historically Lower Precipitation. Soil Biology and Biochemistry 2023, 177, 108889. [CrossRef]
- Alotaibi, F.; St-Arnaud, M.; Hijri, M. In-Depth Characterization of Plant Growth Promotion Potentials of Selected Alkanes-Degrading Plant Growth-Promoting Bacterial Isolates. Frontiers in Microbiology 2022, 13, 863702. [CrossRef]
- Manetsberger, J.; Caballero Gómez, N.; Soria-Rodríguez, C.; Benomar, N.; Abriouel, H. Simply Versatile: The Use of Peribacillus Simplex in Sustainable Agriculture. Microorganisms 2023, 11, 2540. [CrossRef]
- Ma, Y.; Yin, Y.; Rong, C.; Chen, S.; Liu, Y.; Wang, S.; Xu, F. Pantoea Pleuroti Sp. Nov., Isolated from the Fruiting Bodies of Pleurotus Eryngii. Current Microbiology 2016, 72, 207–212. [CrossRef]
- Sikorski, J.; Pukall, R.; Stackebrandt, E. Carbon Source Utilization Patterns of Bacillus Simplex Ecotypes Do Not Reflect Their Adaptation to Ecologically Divergent Slopes in ‘Evolution Canyon.’ Israel. FEMS microbiology ecology 2008, 66, 38–44.
- Palmer, M.; Steenkamp, E.T.; Coetzee, M.P.; Avontuur, J.R.; Chan, W.Y.; Zyl, E.; Venter, S.N. Mixta Gen. Nov., a New Genus in the Erwiniaceae. International journal of systematic and evolutionary microbiology 2018, 68, 1396–1407.
- Dragone, N.B.; Hoffert, M.; Strickland, M.S.; Fierer, N. Taxonomic and Genomic Attributes of Oligotrophic Soil Bacteria. ISME communications 2024, 4, 081. [CrossRef]
- Verhille, S.; Baida, N.; Dabboussi, F.; Hamze, M.; Izard, D.; Leclerc, H. Pseudomonas Gessardii Sp. Nov. and Pseudomonas Migulae Sp. Nov., Two New Species Isolated from Natural Mineral Waters. International Journal of Systematic and Evolutionary Microbiology 1999, 49, 1559–1572.
- Puri, A.; Padda, K.P.; Chanway, C.P. Evidence of Endophytic Diazotrophic Bacteria in Lodgepole Pine and Hybrid White Spruce Trees Growing in Soils with Different Nutrient Statuses in the West Chilcotin Region of British Columbia, Canada. Forest Ecology and Management 2018, 430, 558–565. [CrossRef]
- Costa, O.Y.A.; Raaijmakers, J.M.; Kuramae, E.E. Microbial Extracellular Polymeric Substances: Ecological Function and Impact on Soil Aggregation. Front. Microbiol. 2018, 9. [CrossRef]
- Waqas, M.; Zelenková, S.; Vogel, C.; Rühmann, B.; Varsadiya, M.; Liebmann, P.; Bárta, J. Stick Together: Isolation and Characterization of Exopolysaccharides Producing Bacteria from Degraded Permafrost Soils. bioRxiv 2024.
- Dobritsa, A.P.; Linardopoulou, E.V.; Samadpour, M. Transfer of 13 Species of the Genus Burkholderia to the Genus Caballeronia and Reclassification of Burkholderia Jirisanensis as Paraburkholderia Jirisanensis Comb. Nov. International Journal of Systematic and Evolutionary Microbiology 2017, 67, 3846–3853. [CrossRef]
- Zolg, W.; Ottow, J.C. Pseudomonas Glathei Sp. Nov., a New Nitrogen-Scavening Rod Isolated from Acid Lateritic Relicts in Germany. J Comp Neurol 1975, 164, 287–299.
- Peeters, C.; Meier-Kolthoff, J.P.; Verheyde, B.; Brandt, E.; Cooper, V.S.; Vandamme, P.; Y., O.; Raaijmakers, J.M.; Kuramae, E.E. Phylogenomic Study of Burkholderia Glathei-like Organisms, Proposal of 13 Novel Burkholderia Species and Emended Descriptions of Burkholderia Sordidicola, Burkholderia Zhejiangensis, and Burkholderia Grimmiae. Frontiers in microbiology 2016, 7, 877 ,.
- Li, R.; Jiang, Y.; Wang, X.; Yang, J.; Gao, Y.; Zi, X.; Hu, N. Psychrotrophic Pseudomonas Mandelii CBS-1 Produces High Levels of Poly-β-Hydroxybutyrate. Springerplus 2013, 2, 1–7. [CrossRef]
- Kimmel, S.A.; Roberts, R.F.; Ziegler, G.R. Optimization of Exopolysaccharide Production by Lactobacillus Delbrueckii Subsp. Bulgaricus RR Grown in a Semidefined Medium. Applied and Environmental Microbiology 1998, 64, 659–664.
- Naseem, H.; Ahsan, M.; Shahid, M.A.; Khan, N. Exopolysaccharides Producing Rhizobacteria and Their Role in Plant Growth and Drought Tolerance. Journal of basic microbiology 2018, 58, 1009–1022. [CrossRef]
- Guo, Y.S.; Furrer, J.M.; Kadilak, A.L.; Hinestroza, H.F.; Gage, D.J.; Cho, Y.K.; Shor, L.M. Bacterial Extracellular Polymeric Substances Amplify Water Content Variability at the Pore Scale. Frontiers in Environmental Science 2018, 6, 93. [CrossRef]
- Bourles, A.; Amir, H.; Gensous, S.; Cussonneau, F.; Medevielle, V. Investigating Some Mechanisms Underlying Stress Metal Adaptations of Two Burkholderia Sensu Lato Species Isolated from New Caledonian Ultramafic Soils. European Journal of Soil Biology 2020, 97, 103166. [CrossRef]
- Paul, S.; Parvez, S.S.; Goswami, A.; Banik, A. Exopolysaccharides from Agriculturally Important Microorganisms: Conferring Soil Nutrient Status and Plant Health. International Journal of Biological Macromolecules 2024, 262, 129954. [CrossRef]
- Vanhaverbeke, C.; Heyraud, A.; Mazeau, K. Conformational Analysis of the Exopolysaccharide from Burkholderia Caribensis Strain MWAP71: Impact on the Interaction with Soils. Biopolymers 2003, 69, 480–497. [CrossRef]
- Lünsdorf, E.; Abraham; Timmis Clay Hutches’: A Novel Interaction between Bacteria and Clay Minerals. Environmental Microbiology 2000, 2, 161–168.
- Santaella, C.; Schue, M.; Berge, O.; Heulin, T.; Achouak, W. The Exopolysaccharide of Rhizobium Sp. YAS34 Is Not Necessary for Biofilm Formation on Arabidopsis Thaliana and Brassica Napus Roots but Contributes to Root Colonization. Environmental Microbiology 2008, 10, 2150–2163. [CrossRef]
- Driouich, A.; Gaudry, A.; Pawlak, B.; Moore, J.P. Root Cap–Derived Cells and Mucilage: A Protective Network at the Root Tip. Protoplasma 2021, 258, 1179–1185. [CrossRef]
- Aznar, A.; Dellagi, A. New Insights into the Role of Siderophores as Triggers of Plant Immunity: What Can We Learn from Animals? Journal of Experimental Botany 2015, 66, 3001–3010. [CrossRef]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Waqas, M.; Kang, S.-M.; Lee, I.-J. Inoculation of Abscisic Acid-Producing Endophytic Bacteria Enhances Salinity Stress Tolerance in Oryza Sativa. Environmental and Experimental Botany 2017, 136, 68–77. [CrossRef]
- Forchetti, G.; Masciarelli, O.; Alemano, S.; Alvarez, D.; Abdala, G. Endophytic Bacteria in Sunflower (Helianthus Annuus L.): Isolation, Characterization, and Production of Jasmonates and Abscisic Acid in Culture Medium. Appl Microbiol Biotechnol 2007, 76, 1145–1152. [CrossRef]
- Boiero, L.; Perrig, D.; Masciarelli, O.; Penna, C.; Cassán, F.; Luna, V. Phytohormone Production by Three Strains of Bradyrhizobium Japonicum and Possible Physiological and Technological Implications. Appl Microbiol Biotechnol 2007, 74, 874–880. [CrossRef]
- Joni, F.R.; Hamid, H.; Yanti, Y. Effect of Plant Growth Promoting Rhizobacteria (PGPR) on Increasing the Activity of Defense Enzymes in Tomato Plants. IJEAB 2020, 5, 1474–1479. [CrossRef]
- Amaya-Gómez, C.V.; Porcel, M.; Mesa-Garriga, L.; Gómez-Álvarez, M.I. A Framework for the Selection of Plant Growth-Promoting Rhizobacteria Based on Bacterial Competence Mechanisms. Appl Environ Microbiol 2020, 86, e00760-20. [CrossRef]
- Fadiji, A.E.; Orozco-Mosqueda, M.D.C.; Santos-Villalobos, S.D.L.; Santoyo, G.; Babalola, O.O. Recent Developments in the Application of Plant Growth-Promoting Drought Adaptive Rhizobacteria for Drought Mitigation. Plants 2022, 11, 3090. [CrossRef]
- Shayanthan, A.; Ordoñez, P.A.C.; Oresnik, I.J. The Role of Synthetic Microbial Communities (SynCom) in Sustainable Agriculture. Frontiers in Agronomy 2022, 4, 896307. [CrossRef]
- Haque, M.M.; Mosharaf, M.K.; Khatun, M.; Haque, M.A.; Biswas, M.S.; Islam, M.S.; Siddiquee, M.A. Biofilm Producing Rhizobacteria with Multiple Plant Growth-Promoting Traits Promote Growth of Tomato under Water-Deficit Stress. Frontiers in Microbiology 2020, 11, 542053. [CrossRef]
- Kumar, A.; Maurya, B.R.; Raghuwanshi, R. Rhizobacteria Genome Sequences: Platform for Defining Functional Traits for Sustainable Agriculture. Journal of Genetic Engineering and Biotechnology 2020, 18, 92.
- Abdelaziz, M.E.; Abdelsattar, M.; Abdeldaym, E.A.; Atia, M.A.M.; Mahmoud, A.W.M.; Saad, M.M.; Hirt, H. Piriformospora Indica Promotes Growth of Maize Plants Under High Salinity Stress: A Study on the Regulatory Mechanisms of Salt Stress Response. Frontiers in Microbiology 2019, 10, 1088.
- Wang, L.; Wang, X.; Wu, H.; Wang, H.; Lu, Z. Interspecies Synergistic Interactions Mediated by Cofactor Exchange Enhance Stress Tolerance by Inducing Biofilm Formation. mSystems 2024, 9, 00884–24.
- Cooperation and Conflict in Microbial Biofilms. Proceedings of the National Academy of Sciences 2007, 104, 876–881. [CrossRef]
- Valiei, A.; Dickson, A.; Aminian-Dehkordi, J.; Mofrad, M.R. Metabolic Interactions Shape Emergent Biofilm Structures in a Conceptual Model of Gut Mucosal Bacterial Communities. npj Biofilms and Microbiomes 2024, 10, 99. [CrossRef]
- Sadiq, F.A.; Reu, K.; Burmølle, M.; Maes, S.; Heyndrickx, M. Synergistic Interactions in Multispecies Biofilm Combinations of Bacterial Isolates Recovered from Diverse Food Processing Industries. Frontiers in microbiology 2023, 14, 1159434. [CrossRef]
- Matthus, E.; Zwetsloot, M.; Delory, B.M.; Hennecke, J.; Andraczek, K.; Henning, T.; Bergmann, J. Revisiting the Root Economics Space—Its Applications, Extensions and Nuances Advance Our Understanding of Fine-Root Functioning; Plant and Soil, 2025;
- Kong, D.; Wang, J.; Wu, H.; Valverde-Barrantes, O.J.; Wang, R.; Zeng, H. Nonlinearity of Root Trait Relationships and the Root Economics Spectrum. Nat. Commun 2019, 10, 1–9. [CrossRef]
- Veiga, R.S.L.; Faccio, A.; Genre, A.; Pieterse, C.M.J.; Bonfante, P.; van der Heijden, M.G.A. Arbuscular Mycorrhizal Fungi Reduce Growth and Infect Roots of the Non-Host Plant Arabidopsis Thaliana. Plant Cell Environ 2013, 36, 1926–1937. [CrossRef]
- Ristova, D.; Rosas, U.; Krouk, G.; Ruffel, S.; Birnbaum, K.D.; Coruzzi, G.M. RootScape: A Landmark-Based System for Rapid Screening of Root Architecture in Arabidopsis. Plant physiology 2013, 161, 1086–1096. [CrossRef]
- Iorio, A.; Lasserre, B.; Scippa, G.S.; Chiatante, D. Root System Architecture of Quercus Pubescens Trees Growing on Different Sloping Conditions. Annals of Botany. 2005, 95, 351–361. [CrossRef]
- Jia, H.; Guan, C.; Zhang, J.; He, C.; Yin, C.; Meng, P. Drought Effects on Tree Growth, Water Use Efficiency, Vulnerability and Canopy Health of Quercus Variabilis-Robinia Pseudoacacia Mixed Plantation 2022.
- Tognetti, R.; Longobucco, A.; Miglietta, F.; Raschi, A. Water Relations, Stomatal Response and Transpiration of Quercus Pubescens Trees during Summer in a Mediterranean Carbon Dioxide Spring. Tree Physiology 1999, 19, 261–270. [CrossRef]
- Pividori, M.; Giannetti, F.; Barbati, A.; Chirici, G. European Forest Types: Tree Species Matrix. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., Ed.; Publications Office of the European Union: Luxembourg, 2016; p. 01 162.
- Šajbidorová, V.; Lichtnerová, H.; Paganová, V. The Impact of Different Water Regime on Chlorophyll Fluorescence of Pyrus Pyraster L. and Sorbus Domestica L. Acta Univ. Agric. Silvic. Mendelianae Brun. 2015, 63, 1575–1579. [CrossRef]
- Paganová, V.; Jureková, Z.; Lichtnerová, H. The Nature and Way of Root Adaptation of Juvenile Woody Plants Sorbus and Pyrus to Drought. Environ Monit Assess 2019, 191, 714. [CrossRef]
- Schmucker, J.; Skovsgaard, J.P.; Uhl, E.; Pretzsch, H. Crown Structure, Growth, and Drought Tolerance of True Service Tree (Sorbus Domestica L.) in Forests and Urban Environments. Urban Forestry & Urban Greening 2024, 91, 128161. [CrossRef]
- Rudrappa, T.; Splaine, R.E.; Biedrzycki, M.L.; Bais, H.P. Cyanogenic pseudomonads influence multitrophic interactions in the rhizosphere. PLoS One 2008, 3, 2073.
- Cheng, J.; Schloerke, B.; Karambelkar, B.; Xie, Y. Leaflet: Create Interactive Web Maps with the JavaScript “Leaflet” Library; 2024;
- Kunz, J.; Räder, A.; Bauhus, J. Effects of Drought and Rewetting on Growth and Gas Exchange of Minor European Broadleaved Tree Species. Forests 2016, 7, 239. [CrossRef]
- Emmenegger, B.; Massoni, J.; Pestalozzi, C.M.; Bortfeld-Miller, M.; Maier, B.A.; Vorholt, J.A. Identifying Microbiota Community Patterns Important for Plant Protection Using Synthetic Communities and Machine Learning. Nature Communications 2023, 14, 7983.
- Polade, S.D.; Pierce, D.W.; Cayan, D.R.; Gershunov, A.; Dettinger, M.D. The Key Role of Dry Days in Changing Regional Climate and Precipitation Regimes. Sci Rep 2014, 4, 4364. [CrossRef]
- Pérez-Miranda, S.; Cabirol, N.; George-Téllez, R.; Zamudio-Rivera, L.S.; Fernández, F.J. O-CAS, a Fast and Universal Method for Siderophore Detection. Journal of Microbiological Methods 2007, 70, 127–131. [CrossRef]
- Ustiatik, R.; Nuraini, Y.; Suharjono, S.; Handayanto, E. Siderophore Production of the Hg-Resistant Endophytic Bacteria Isolated from Local Grass in the Hg-Contaminated Soil. J. Ecol. Eng. 2021, 22, 129–138. [CrossRef]
- Gomes, A.F.R.; Sousa, E.; Resende, D.I.S.P. A Practical Toolkit for the Detection, Isolation, Quantification, and Characterization of Siderophores and Metallophores in Microorganisms. ACS Omega 2024, 9, 26863–26877. [CrossRef]
- Gordon, S.A.; Weber, R.P. COLORIMETRIC ESTIMATION OF INDOLEACETIC ACID. Plant Physiol 1951, 26, 192–195. [CrossRef]
- Gang, S.; Sharma, S.; Saraf, M.; Buck, M.; Schumacher, J. Analysis of Indole-3-Acetic Acid (IAA) Production in Klebsiella by LC-MS/MS and the Salkowski Method. BIO-PROTOCOL 2019, 9. [CrossRef]
- Fricke, W.; Akhiyarova, G.; Veselov, D.; Kudoyarova, G. Rapid and Tissue-specific Changes in ABA and in Growth Rate in Response to Salinity in Barley Leaves. Journal of Experimental Botany 2004, 55, 1115–1123. [CrossRef]
- Verslues, P.E. Quantification of Water Stress-Induced Osmotic Adjustment and Proline Accumulation for Arabidopsis Thaliana Molecular Genetic Studies. Methods Mol Biol 2010, 639, 301–315. [CrossRef]
- Siddique, S.; Naveed, M.; Yaseen, M.; Shahbaz, M. Exploring Potential of Seed Endophytic Bacteria for Enhancing Drought Stress Resilience in Maize (Zea Mays L.). Sustainability 2022, 14, 673. [CrossRef]
- Orhan, F.; Efe, D.; Gormez, A. Chapter 7 - Advantages of Using Halotolerant/Halophilic Bacteria in Agriculture. In Unravelling Plant-Microbe Synergy; Chandra, D., Bhatt, P., Eds.; Developments in Applied Microbiology and Biotechnology; Academic Press, 2023; pp. 133–149 ISBN 978-0-323-99896-3.
- Ren, D.; Madsen, J.S.; Sørensen, S.J.; Burmølle, M. High Prevalence of Biofilm Synergy among Bacterial Soil Isolates in Cocultures Indicates Bacterial Interspecific Cooperation. ISME J 2015, 9, 81–89. [CrossRef]
- Yang, N.; Nesme, J.; Røder, H.L.; Li, X.; Zuo, Z.; Petersen, M.; Sørensen, S.J. Emergent Bacterial Community Properties Induce Enhanced Drought Tolerance in Arabidopsis. npj Biofilms and Microbiomes 2021, 7, 82.
- Sprouffske, K.; Wagner, A. Growthcurver: An R Package for Obtaining Interpretable Metrics from Microbial Growth Curves. BMC bioinformatics 2016, 17, 1–4.
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. Journal of Open Source Software 2019, 4, 1686. [CrossRef]
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests 2023.
- Lê, S.; Josse, J.; Husson, F. FactoMineR : An R Package for Multivariate Analysis. J. Stat. Soft. 2008, 25. [CrossRef]
- Kassambara, A.; Kosinski, M.; Biecek, P.; Fabian, S. Survminer: Drawing Survival Curves Using “Ggplot2” 2024.
- Balcı, S. ClinicoPathJamoviModule: Justice for All 2020.
- The Jamovi Project (2025). Jamovi (Version 2.6).
Figure 1.
Workflow for designing and evaluating drought-resilient synthetic microbial communities (SynComs). Soil with a drought legacy (partial rain exclusion) were sampled at two depths (0–5 cm and 5–10 cm) and subdivided into four root compartments: Roots, Root-Adhering Soil (RAS), non-Root-Adhering Soil (non-RAS), and Root-Induced Macroaggregates (RIM). Bacterial isolates were screened for key plant growth–promoting traits (Exopolysaccharide, auxins and abscisic acid production, ACC deaminase, siderophores, osmotic tolerance) and grouped via multiple correspondence analysis (MCA).Four-strain SynComs were assembled to maximize soil spatial and functional complementarity and tested for biofilm-based synergy. Top-, lowest- and mid-performing SynComs, identified from a Principal Component Analysis (PCA) on biofilm synergy and Arabidopsis thaliana assays, were further evaluated on Quercus pubescens and Sorbus domestica seedlings under drought and well-watered (40% and 80% field capacity) conditions. Survival modeling (Cox and multinomial logistic regression) assessed SynCom performance and key predictors of resilience. (Created with BioRender).
Figure 1.
Workflow for designing and evaluating drought-resilient synthetic microbial communities (SynComs). Soil with a drought legacy (partial rain exclusion) were sampled at two depths (0–5 cm and 5–10 cm) and subdivided into four root compartments: Roots, Root-Adhering Soil (RAS), non-Root-Adhering Soil (non-RAS), and Root-Induced Macroaggregates (RIM). Bacterial isolates were screened for key plant growth–promoting traits (Exopolysaccharide, auxins and abscisic acid production, ACC deaminase, siderophores, osmotic tolerance) and grouped via multiple correspondence analysis (MCA).Four-strain SynComs were assembled to maximize soil spatial and functional complementarity and tested for biofilm-based synergy. Top-, lowest- and mid-performing SynComs, identified from a Principal Component Analysis (PCA) on biofilm synergy and Arabidopsis thaliana assays, were further evaluated on Quercus pubescens and Sorbus domestica seedlings under drought and well-watered (40% and 80% field capacity) conditions. Survival modeling (Cox and multinomial logistic regression) assessed SynCom performance and key predictors of resilience. (Created with BioRender).
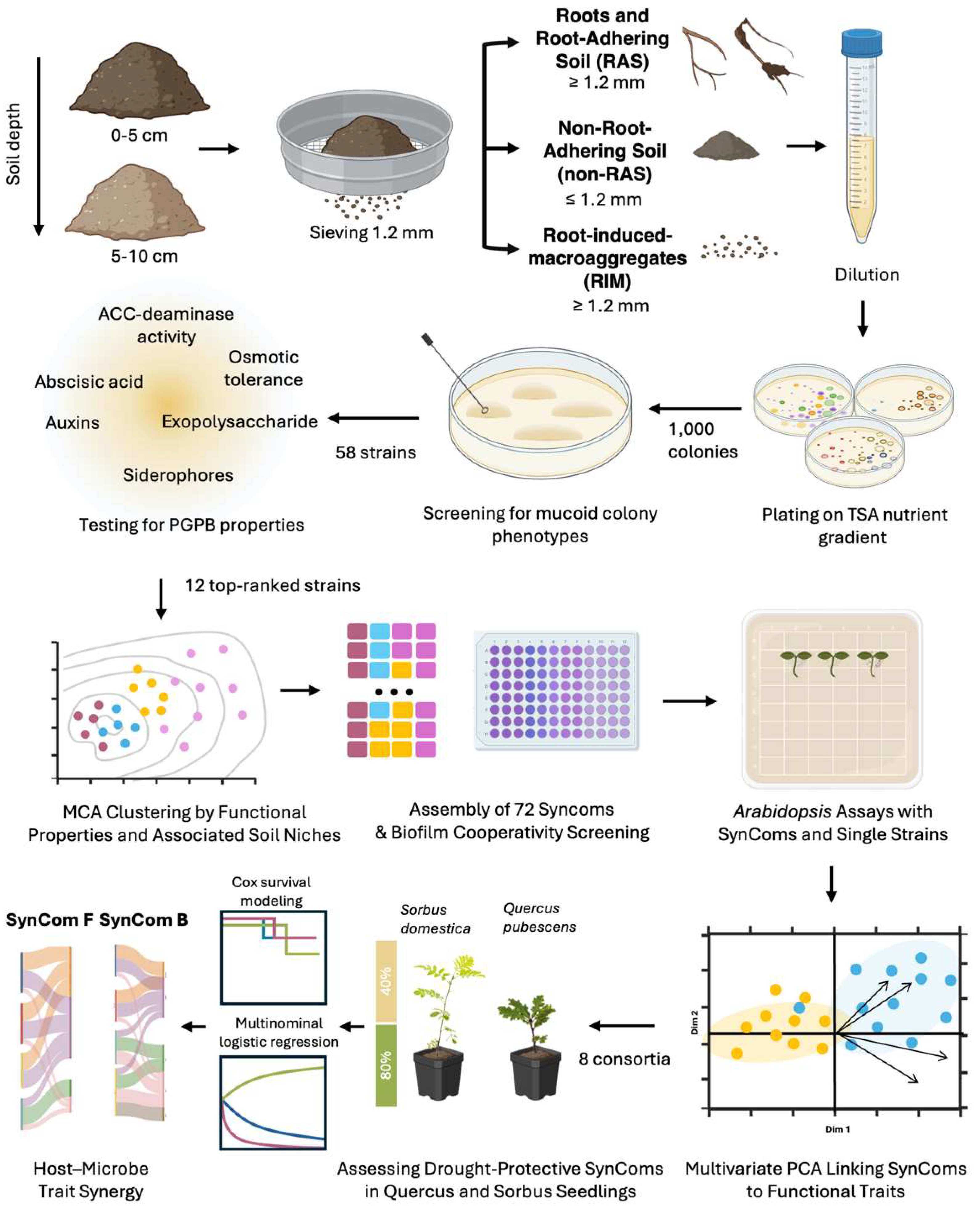
Figure 2.
A) Schematic diagram of sampling from soil submitted to an amplified drought. B) Number of bacterial strains isolated at two soil depths, 0–5 cm and 5–10 cm. We partitioned the soil niche into distinct compartments surrounding the root: the root itself, the root-adhering soil (RAS), the root-induced macroaggregates (RIM), and the non-adhering root soil. C) Distribution of bacterial isolates across soil depths and compartments; (C) Number of mucoid strains isolated from each compartment. Isolation of strains on 10-, 20- and 50-fold diluted tryptic soy broth agar (TSA).
Figure 2.
A) Schematic diagram of sampling from soil submitted to an amplified drought. B) Number of bacterial strains isolated at two soil depths, 0–5 cm and 5–10 cm. We partitioned the soil niche into distinct compartments surrounding the root: the root itself, the root-adhering soil (RAS), the root-induced macroaggregates (RIM), and the non-adhering root soil. C) Distribution of bacterial isolates across soil depths and compartments; (C) Number of mucoid strains isolated from each compartment. Isolation of strains on 10-, 20- and 50-fold diluted tryptic soy broth agar (TSA).
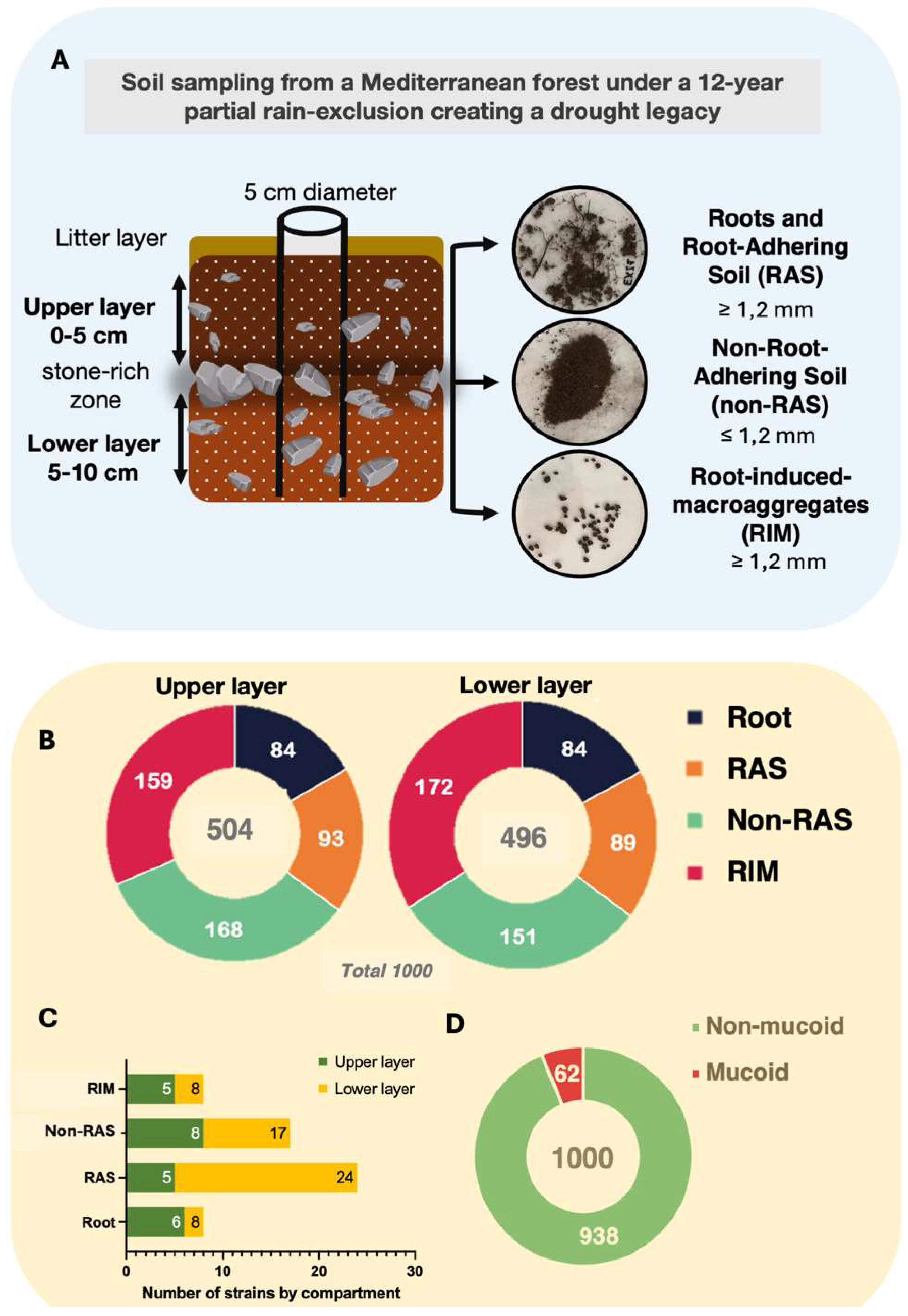
Figure 3.
Twelve top-performing isolates. Distribution and numbers across soil depths, root-associated compartments and culture medium. Soil was sampled at two depths (0-5 cm and 5-10 cm) and divided into four compartments: roots, root-associated soil (RAS), non-root-associated soil (non-RAS) and root-induced macroaggregates (RIM). Strains were grown on tryptic soy agar (TSA) diluted 10×, 20× or 50× to capture different metabolic strategies. Bars (or sections) show the number of isolates obtained from each depth-compartment-medium combination.
Figure 3.
Twelve top-performing isolates. Distribution and numbers across soil depths, root-associated compartments and culture medium. Soil was sampled at two depths (0-5 cm and 5-10 cm) and divided into four compartments: roots, root-associated soil (RAS), non-root-associated soil (non-RAS) and root-induced macroaggregates (RIM). Strains were grown on tryptic soy agar (TSA) diluted 10×, 20× or 50× to capture different metabolic strategies. Bars (or sections) show the number of isolates obtained from each depth-compartment-medium combination.
Figure 4.
Multiple correspondence analysis (MCA). A) MCA Variable plot showing the contribution of ecological and functional variables (centroids) to the first two dimensions Dim 1 and Dim 2. Ecological variables are: soil compartment (Roots; Root-adhering Soil, RAS; non-Root-Adhering Soil, non-RAS); and Root-Induced Macroaggregates, RIM) and depth (0-5 cm and 5-10 cm), culture medium (Tryptic Soil Agar, TSA diluted 10- 20- and 50-fold). Functional variables are exopolysaccharide production, (EPS), osmotic tolerance to Polyethylene Glycol (solid or liquid culture assays, PEG 10 and 30%), and to sodium chloride (NaCl 0.5, 0.1 and 1.5 M), auxin (IAA) and abscisic acid (ABA) production, ACC-deaminase activity (ACC), and strain genus. B) MCA biplot showing the relationships between individual strains (brown dots) and the ecological and functional variables. A, B, C, D Letters indicate expression levels: A = low or none, B = moderate, C = high, and D = very high. Lines represent the iso-response values for the variable indicated.
Figure 4.
Multiple correspondence analysis (MCA). A) MCA Variable plot showing the contribution of ecological and functional variables (centroids) to the first two dimensions Dim 1 and Dim 2. Ecological variables are: soil compartment (Roots; Root-adhering Soil, RAS; non-Root-Adhering Soil, non-RAS); and Root-Induced Macroaggregates, RIM) and depth (0-5 cm and 5-10 cm), culture medium (Tryptic Soil Agar, TSA diluted 10- 20- and 50-fold). Functional variables are exopolysaccharide production, (EPS), osmotic tolerance to Polyethylene Glycol (solid or liquid culture assays, PEG 10 and 30%), and to sodium chloride (NaCl 0.5, 0.1 and 1.5 M), auxin (IAA) and abscisic acid (ABA) production, ACC-deaminase activity (ACC), and strain genus. B) MCA biplot showing the relationships between individual strains (brown dots) and the ecological and functional variables. A, B, C, D Letters indicate expression levels: A = low or none, B = moderate, C = high, and D = very high. Lines represent the iso-response values for the variable indicated.
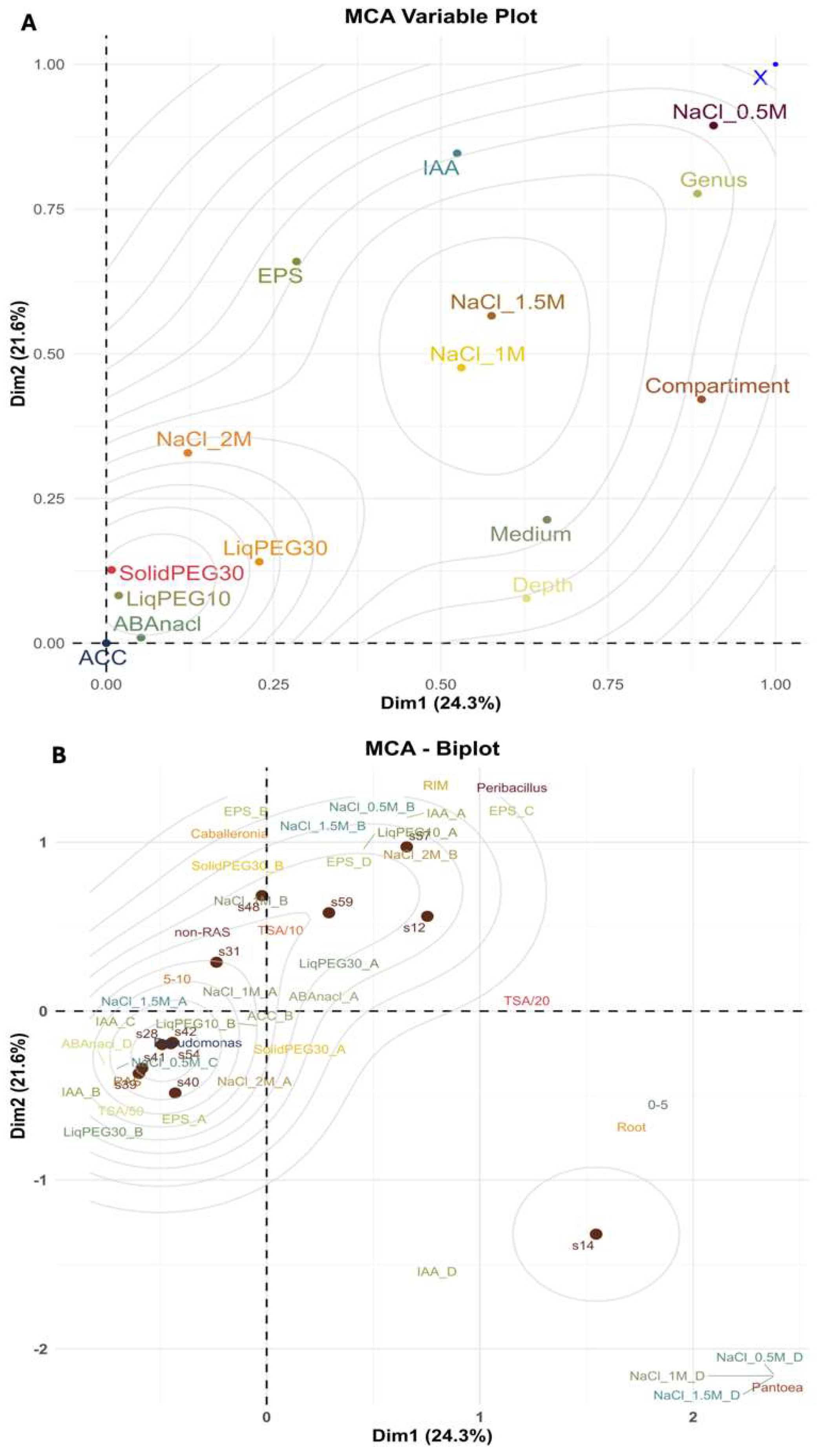
Figure 5.
Principal Components Analysis of the biofilm assay and in vitro inoculation of Arabidopsis thaliana with single strains s1 to s12 or 4-strain synthetic communities (SynCom) A to F. Variables are: Germination (percentage of germinated seeds), Growth (percentage of seedlings that continued growing after germination), Development (percentage of well-developed seedlings – seedlings with > 10 lateral roots and with rosettes ≥ 0.7 cm and at least 6 leaves). Ellipses 95% confidence interval.
Figure 5.
Principal Components Analysis of the biofilm assay and in vitro inoculation of Arabidopsis thaliana with single strains s1 to s12 or 4-strain synthetic communities (SynCom) A to F. Variables are: Germination (percentage of germinated seeds), Growth (percentage of seedlings that continued growing after germination), Development (percentage of well-developed seedlings – seedlings with > 10 lateral roots and with rosettes ≥ 0.7 cm and at least 6 leaves). Ellipses 95% confidence interval.
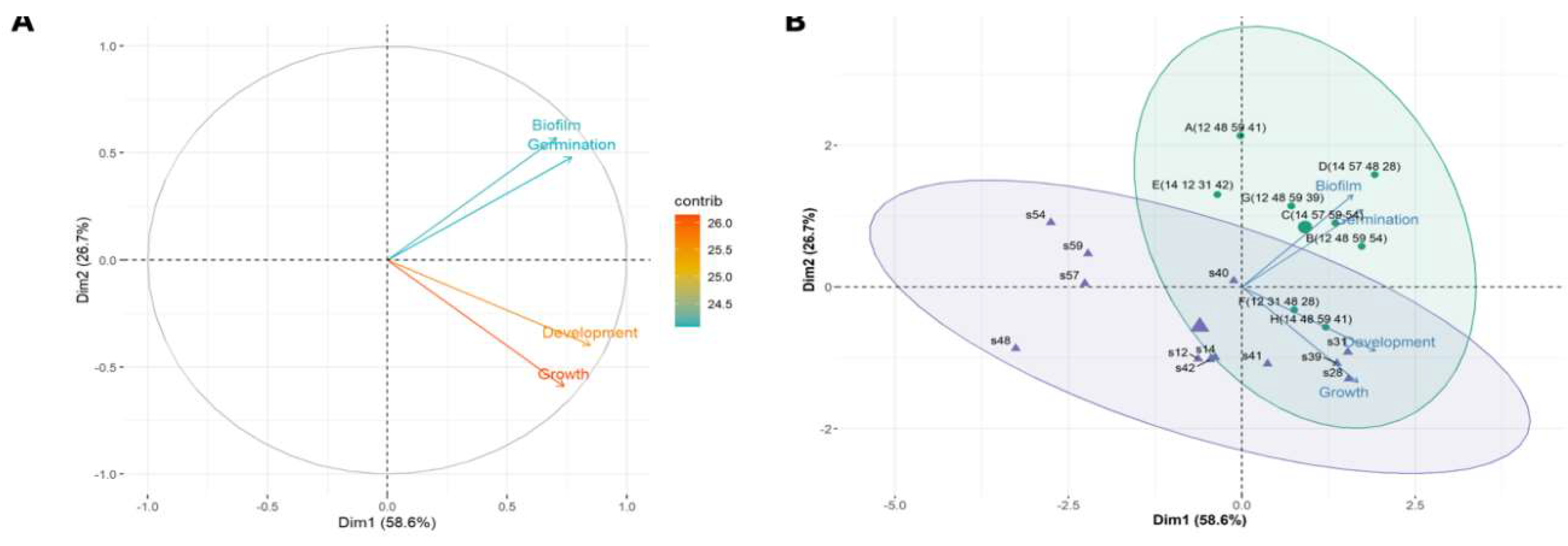
Figure 6.
Alluvial diagram illustrating the compositions of the top-performing SynComs B (left) for Quercus pubescens and F (right) for Sorbus domestica. Flow widths represent the relative contribution of each strain key traits (e.g., exopolysaccharide production, osmotic tolerance, hormone biosynthesis) within each optimized consortium.
Figure 6.
Alluvial diagram illustrating the compositions of the top-performing SynComs B (left) for Quercus pubescens and F (right) for Sorbus domestica. Flow widths represent the relative contribution of each strain key traits (e.g., exopolysaccharide production, osmotic tolerance, hormone biosynthesis) within each optimized consortium.
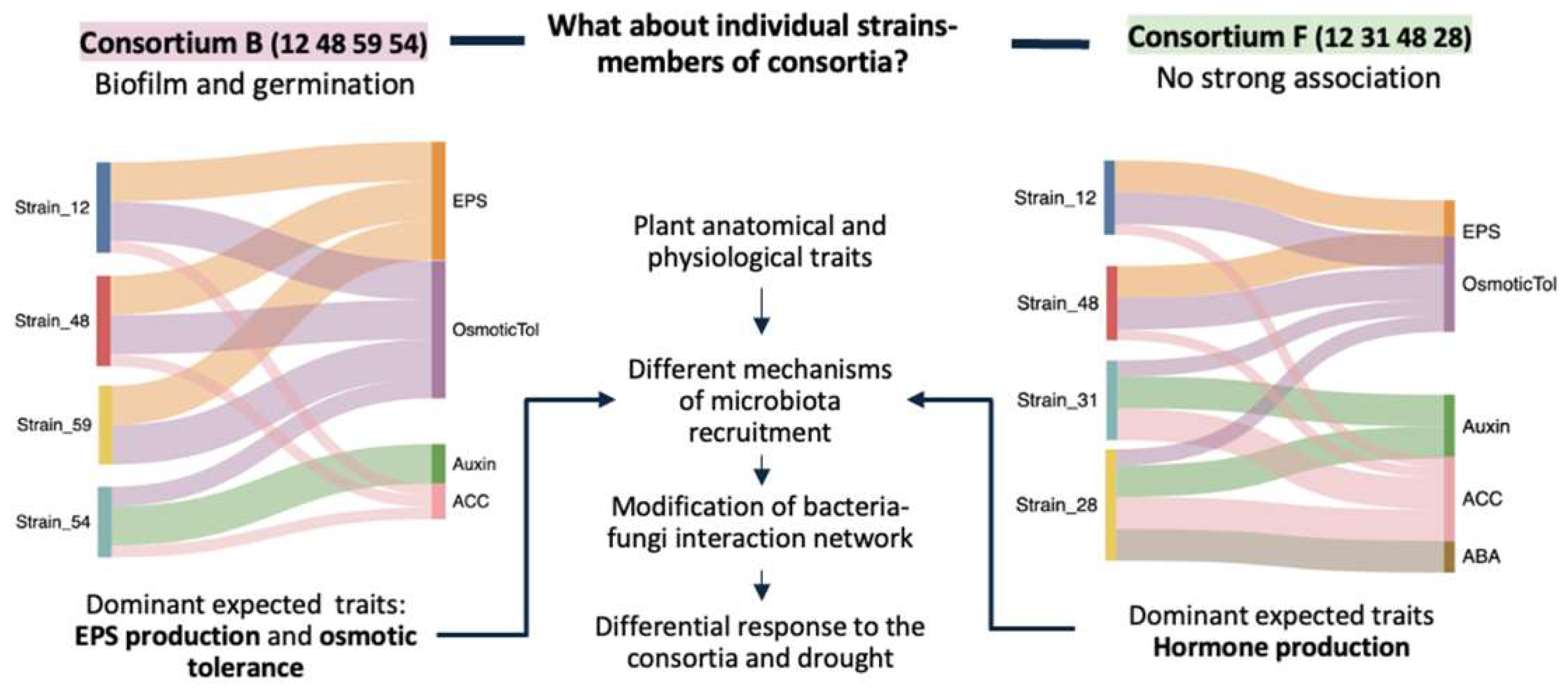
Figure 7.
Probability of symptom development under the most efficient synthetic microbial communities (SynComs) for each tree species, calculated via estimated marginal means from a multinomial logistic regression model. (A) Sorbus domestica inoculated with SynCom F; (B) Quercus pubescens inoculated with SynCom B.
Figure 7.
Probability of symptom development under the most efficient synthetic microbial communities (SynComs) for each tree species, calculated via estimated marginal means from a multinomial logistic regression model. (A) Sorbus domestica inoculated with SynCom F; (B) Quercus pubescens inoculated with SynCom B.
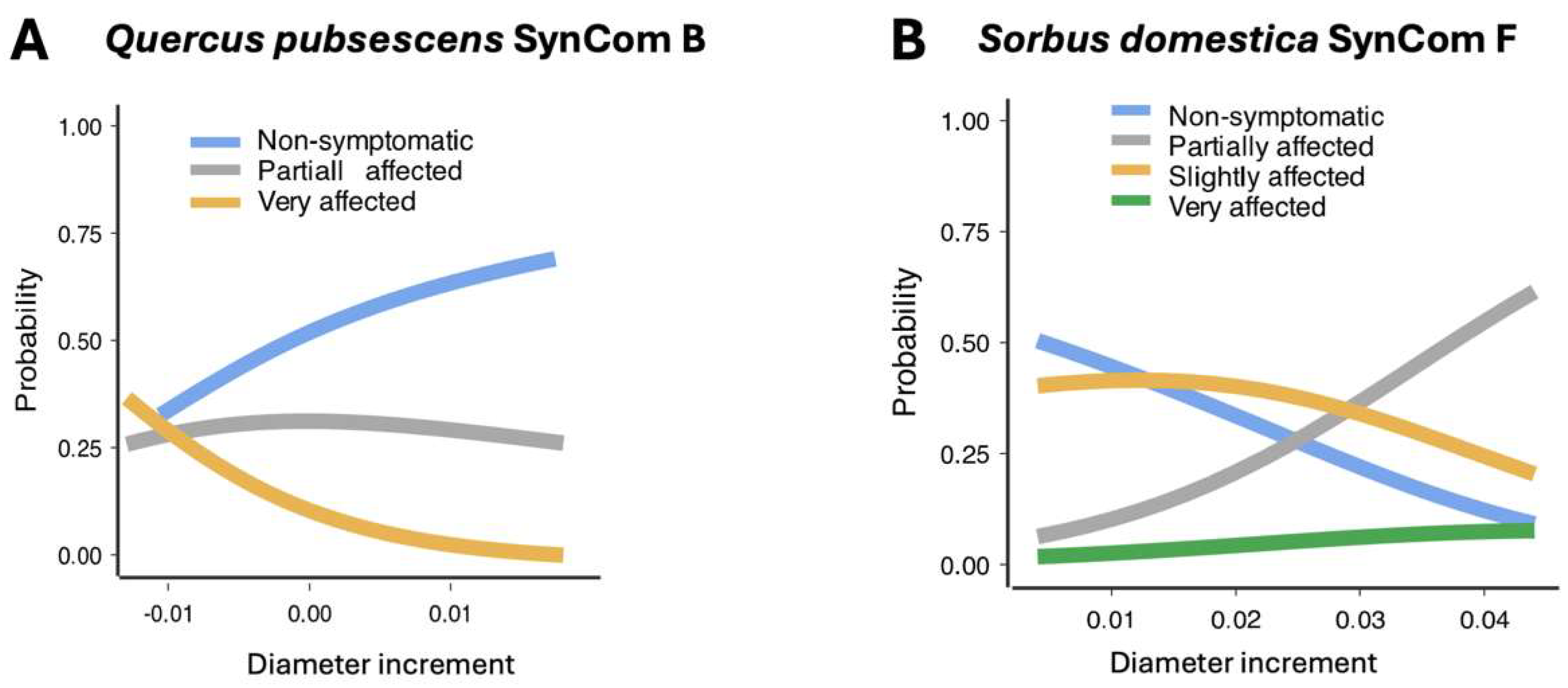
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



