
Submitted:
03 April 2025
Posted:
04 April 2025
You are already at the latest version
Abstract
Reversine, a synthetic purine analog, has been recognized for its dual role in cellular dedifferentiation and inhibition of mitotic kinases, particularly aurora A and B. Chronic myeloid leukemia (CML) cells exhibit genomic instability associated with dysregulated telomere dynamics. This study investigates the effects of reversine on three-dimensional (3D) telomere architecture of CML cell lines (K-562 and MEG-01), using TeloView® analysis. Our findings demonstrate that reversine treatment significantly alters already dynamically changed 3D telomere organization in CML, reducing the total number of telomeres, telomere aggregates, and nuclear volume while decreasing the expression of AURKA and AURKB. These changes suggest a potential mechanism by which reversine impacts chromatin integrity and leukemic cell viability, namely through the pronounced modulation of 3D telomere architecture. The telomere-dependent destablization of the 3D genome may contribute to reversine’s anti-leukemic effects leading to impaired mitotic progression. Our results support further exploration of reversine as a potential therapeutic agent in CML, particularly in combination with current treatment strategies targeting telomere and chromatin dynamics.
Keywords:
Chronic Myeloid Leukemia
; Reversine
; Telomeres
; TeloView®
; Genomic Instability
; Aurora Kinases
1. Introduction
Reversine is a small synthetic purine analog that has gained attention for its dual role in cellular dedifferentiation and inhibition of mitotic kinases [1]. Initially identified as an agent capable of inducing dedifferentiation of committed cells into multipotent progenitor-like cells [1], reversine has since been recognized for its inhibitory effects on aurora kinases, particularly aurora A and B, which are crucial regulators of mitosis [2]. By targeting these kinases, reversine disrupts mitotic progression, leading to cytokinesis failure, polyploidization, and suppression of tumor cell proliferation [3].
Aurora kinases, including AURKA and AURKB, are serine/threonine kinases that regulate chromosome segregation and cytokinesis during cell division [4]. Overexpression of these kinases has been implicated in some cancers, including hematological malignancies [5,6,7]. In chronic myeloid leukemia (CML), the BCR-ABL oncoprotein upregulates AURKA and AURKB via the Akt signaling pathway, promoting leukemogenesis [8]. Elevated expression of these kinases has been correlated with poor prognosis and resistance to tyrosine kinase inhibitors (TKIs), which are the standard treatment for CML [9].
Several studies have demonstrated that reversine inhibits leukemia cell proliferation by targeting aurora kinases. Research on acute myeloid leukemia (AML) cells found that reversine significantly reduced colony formation and induced apoptosis, while displaying lower toxicity toward healthy donor cells compared to other aurora kinase inhibitors such as VX-680 [2,9]. Additionally, reversine has been found to cause mitotic defects and polyploidization, resulting in the suppression of glioblastoma cells and other tumors [10,11]. Studies have also reported that reversine inhibits the proliferation of chronic myeloid leukemia (CML) cells, supporting its potential as an anti-leukemic agent [2,12]. This selective cytotoxicity suggests that reversine may be a promising candidate for leukemia therapy, including in CML, where dysregulation of aurora kinases plays a role in disease progression [13,14].
Three-dimensional (3D) telomere analysis, performed using TeloView® software, enables quantitative assessment of the 3D telomere architecture by evaluating parameters such as telomere number, length, signal intensity, spatial distribution, and the presence of telomere aggregates [15,16]. In CML, abnormalities in telomere dynamics have been correlated with disease progression, with increased telomere aggregates and nuclear volume alterations being observed in advanced phases of the disease [17]. Studies have shown that as CML progresses from the chronic phase to the blast crisis, there is a shift in telomere organization, which may be linked to genomic instability and resistance to therapy [18].
Although direct studies on reversine’s impact on 3D telomere architecture in CML cells remain unknown, its role as an aurora kinase inhibitor suggests a potential influence on telomere maintenance. Aurora kinases regulate mitotic progression and chromosome segregation, both of which are crucial for telomere stability [19,20]. The inhibition of aurora kinases by reversine has been associated with mitotic defects, aneuploidy, and altered nuclear architecture, which could indirectly affect telomere integrity [20].
In this newly conducted study, we proposed that reversine influences mitotic progression not only by inducing telomere dysfunction but also via its dedifferentiation action, which may promote a more plastic, progenitor-like state in cells. To explore this, we evaluated—for the first time—the effects of reversine on MEG-01 and K-562 chronic myeloid leukemia cells by analyzing changes in their 3D telomere architecture using TeloView® software before and after treatment. Furthermore, we examined the modulation of aurora kinase gene expression to assess reversine’s potential in restoring genomic stability.
2. Material and Methods
2.1. Cell Culture
For this study, MEG-01 and K-562 cells, obtained from ATCC, were cultured in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS), 1% L-glutamine, and 1% penicillin-streptomycin. The cells were maintained at 37°C in a humidified atmosphere with 5% CO₂. To investigate the effects of reversine (Cayman Chemical: Based in Ann Arbor, Michigan, USA), a selective aurora kinase inhibitor, the compound was dissolved in DMSO to prepare a 10mM stock solution. Working concentrations of 5µM, 10µM, 20µM and 30µM were freshly prepared, and cells were treated for 24, 48 and 72 hours, with control groups receiving an equivalent amount of DMSO (<0.1% v/v).
2.2. Cytotoxicity Assessment and Cell Viability Analysis
To assess cytotoxicity, the viability of MEG-01 and K-562 cells was determined using the trypan blue exclusion assay and the MTT assay (Promega Corporation, Madison, USA). For the trypan blue assay, treated and control cells were stained with 0.4% trypan blue solution and counted using a hemocytometer. The MTT assay was performed by incubating the cells with MTT reagent (0.5 mg/mL) for four hours at 37°C, followed by the addition of DMSO to solubilize formazan crystals. Absorbance was measured at 570 nm using a microplate reader. The half-maximal inhibitory concentration (IC₅₀) was determined using non-linear regression analysis in GraphPad Prism 8 software.
2.3. Quantitative Fluorescent In Situ Hybridization (Q-FISH)
For evaluation of telomere architecture before and after reversine treatment, the cells were harvested and fixed with 4% paraformaldehyde. Fixed cells were then placed onto poly-L-lysine-coated coverslips, and nuclei were counterstained with DAPI (0.1 µg/mL) before being mounted with an antifade reagent. Telomere staining was performed using Cy3-labeled telomeric peptide nucleic acid (PNA) probes (Thermo Fisher Scientific, Waltham - Massachusetts, EUA), following a standard quantitative Fluorescence in situ Hybridization (Q-FISH) protocol. The hybridization process involved denaturation at 82°C for three minutes, followed by incubation at 30°C for two hours. Post-hybridization washes were carried out using SSC buffer and PBS to remove non-specific binding.
D Image Acquisition and Processing
Interphase nuclei from each sample were analyzed using an AxioImager M1 microscope (Carl Zeiss, Jena, Germany) equipped with an AxioCam HRm charge-coupled device and a 63x oil objective lens (Carl Zeiss, Jena, Germany). Acquisition times were 500 milliseconds (ms) for Cy3 (telomeres) and 5 ms for DAPI (nuclei). Sixty z-stacks were captured at a sampling distance of x,y: 102 nm and z: 200 nm for each stack slice. Images were deconvolved and converted to TIFF files for 3D analysis using TeloView® (Telo Genomics Corp., Toronto ON, Canada) software [15,16].
2.4. TeloView® Analysis and Statistics
To compare telomere architecture among treated samples, the TeloView® software, proprietary to Telo Genomics, Toronto, Canada, was used [15,16]. TeloView® measures six distinct parameters: (1) telomere length based on signal intensity, (2) the number of telomere signals per nucleus, (3) the number of telomeric aggregates (clusters of telomeres that cannot be resolved further at an optical resolution limit of 200 nm), (4) nuclear volume, (5) a/c ratio (a spatial feature assessing cell cycle progression and proliferation), and (6) the spatial distribution of telomeres within the nuclear space, which reflects gene expression. Telomere dynamics were analyzed across cell cycle stages (G0/G1, S, and G2). Graphical representations were generated for subgroups of treated cells, illustrating telomere signal intensity, aggregate frequency, and signal distribution per cell. Telomeric parameters were compared among subgroups using analysis of variance, and their distributions were analyzed using chi-square tests. Average cell parameters were compared using nested factorial analysis of variance, accounting for both patient and cellular variations. A significant level of 0.05 was used for all tests.
2.5. Aurora Kinase mRNA Analysis
To investigate the modulation of AURKA and AURKB expression levels, after and before reversine treatment, quantitative real-time PCR (qPCR) was performed. Total RNA was extracted from treated and control cells using TRIzol® reagent, and RNA purity was confirmed using a NanoDrop spectrophotometer (260/280 ratio > 1.8). cDNA synthesis was carried out using the High-Capacity cDNA Reverse Transcription Kit. qPCR analysis was conducted using SYBR® Green Master Mix on an ABI 7500 Real-Time PCR System. Specific primers for AURKA (Hs00269212_m1) and AURKB (Hs00177782_m1) were used, with GAPDH as an endogenous control. The reaction conditions included an initial denaturation at 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute. The relative expression levels of AURKA and AURKB were determined using the ΔΔCt method, with control samples serving as the reference group.
Statistical analysis of all experiments was performed using GraphPad Prism 8. Differences between control and treated groups were analyzed using one-way ANOVA with Bonferroni post-hoc tests for multiple comparisons. A p-value of less than 0.05 was considered statistically significant.
3. Results
Our data demonstrate the effect of reversine on cell viability and gene expression in two chronic myeloid leukemia (CML) cell lines, K-562 and MEG-1, as expected, and in accordance with various studies published [9,10,11]. The MTT assay results (Figure 1A,B) reveal a dose- and time-dependent decrease in cell viability upon treatment with reversine. Across all tested concentrations (0, 5, 10, 20, and 30 μM), both cell lines exhibited a progressive reduction in viability, with the most pronounced effect observed at the highest concentration (30 μM). The inhibitory effect became more evident with increased exposure time, suggesting that reversine’s impact on cell viability intensifies over prolonged incubation periods. Notably, statistical analysis using ANOVA followed by Bonferroni post-test confirmed a highly significant effect (p<0.0001), indicating strong reproducibility across independent experiments.
Additionally, qPCR analysis (Figure 1C,D) assessed the relative expression levels of AURKA and AURKB mRNA after 24 hours of reversine treatment. The results indicate a concentration-dependent downregulation of both genes compared to the DMSO-treated control, which served as the calibrator. The decline in AURKA and AURKB expression suggests that reversine interferes with the transcriptional regulation of these mitotic kinases, which are known to be crucial regulators of cell cycle progression and survival in leukemia cells. This finding aligns with previous studies highlighting the role of AURKA and AURKB in maintaining proliferative capacity in cancer cells, supporting the hypothesis that reversine exerts its cytotoxic effects, at least in part, by disrupting their expression.
Together, this data provides strong evidence that reversine effectively impairs CML cell viability while modulating key mitotic regulators. The dose- and time-dependent trends observed in both assays underscore the potential of reversine as a targeted therapeutic agent against leukemia cells. The consistency between cell viability reduction and gene expression downregulation suggests a mechanistic link between reversine’s action and the inhibition of AURKA and AURKB, further reinforcing its potential as an anticancer agent.
For 3D telomere archtecture of CML cell lines (K-562 and MEG-01) we determined the IC₅₀ to ensure the preservation of viable cells, and phenotypic changes induced by reversine treatment. The IC₅₀ concentration was established at 28.8 µM for K-562 cells and 28.7 µM for MEG-01 cells. The assessment of telomere organization and structural parameters was conducted using TeloView® software [15,16], which enables a comprehensive evaluation of nuclear telomere distributions, aggregates, signal intensity, and nuclear volume in three-dimensional nuclear space. By employing TeloView®, it was possible to quantify key telomere features such as the total number of telomeres, telomere clustering (aggregates), intensity variations, a/c ratio, and nuclear volume, providing a detailed depiction of how reversine alters nuclear architecture (Table 1).
Statistical analysis revealed significant alterations in telomere organization following reversine exposure in CML cells. The total number of telomeres was markedly reduced, suggesting telomere reorganization, potentially reflecting decreased chromosomal instability. The number of telomere aggregates was also significantly lower in treated cells, indicating a disruption of telomere clustering, which may be linked to impaired chromatin architecture and genome stability (Table 1) (Figure 2). Furthermore, telomere signal intensity decreased significantly after reversine treatment, which could be indicative of telomere attrition, decompaction, or a reduction in telomeric DNA content (Table 1). Moreover, the a/c ratio, a critical parameter reflecting the spatial organization of telomeres and nuclear shape, exhibited significant modifications, suggesting a profound impact of reversine on nuclear integrity. Additionally, nuclear volume was markedly reduced in both K-562 and MEG-01 cells, reinforcing the hypothesis that reversine induces nuclear condensation and structural reorganization (Table 1).
The data presented in Figure 3A,B illustrated the distribution of telomere intensity, a parameter correlating with telomere length, in CML cells under distinct experimental conditions. The analysis included two CML cell lines, K-562 and MEG-01, each assessed in both untreated and reversine-treated states.
The graphical representation demonstrated a three-dimensional telomere profiling approach, capturing variations in telomere length distribution across the different conditions (Figure 3A,B). In untreated cells, the telomere intensity distribution exhibited a characteristic pattern, indicative of a heterogeneous population with varying telomere lengths (Figure 3A,B). Upon reversine treatment, shifts in this distribution were observed, suggesting an influence on telomere integrity or regulatory mechanisms governing telomere length maintenance.
Given the critical role of telomeres in genomic stability and cellular replicative capacity, the observed differences between treated and untreated cells provided insights into the molecular effects of reversine on telomere dynamics in CML. The distinct telomere distribution profiles between the K-562 and MEG-01 cell lines further implied intrinsic differences in telomere maintenance pathways, potentially linked to variations in cellular responses to therapeutic agents.
Utilizing three-dimensional telomere analysis via the TeloView® platform allowed for a more precise evaluation of telomere organization within the nuclear architecture. This detailed analysis not only linked reversine treatment to the inhibition of mitotic progression but also highlighted its role in inducing dedifferentiation. This advanced methodological approach allowed for the assessment of spatial distribution alongside telomere length heterogeneity, facilitating a comprehensive understanding of the impact of reversine on telomere biology in CML cells. These findings may have implications for therapeutic strategies targeting telomere-associated mechanisms in leukemia.
4. Discussion
The data presented in this study provide compelling evidence that reversine, a known aurora kinase inhibitor, significantly impacts the three-dimensional (3D) telomere architecture in CML cell lines, specifically K-562 and MEG-01. The observed alterations in telomere dynamics upon reversine treatment suggest a potential mechanism by which reversine exerts its antileukemic effects [21,22,23].
Telomeres, the protective caps at the ends of chromosomes, play a crucial role in maintaining genomic stability. In CML, telomere dysfunction has been associated with disease progression and poor prognosis [18]. Studies have demonstrated that as CML advances from the chronic phase to the blast crisis, there is a notable shortening of telomere length, which correlates with increased genomic instability [24]. The current findings align with these observations, as reversine treatment resulted in significant changes in telomere length and organization, potentially reflecting a shift toward a less aggressive cellular phenotype [15].
These findings align with reversine’s established dual role as both a cell dedifferentiation agent and a selective anticancer compound [2,24]. The literature suggests that reversine facilitates dedifferentiation by reprogramming lineage-committed cells into progenitor-like states, which involves chromatin remodeling and changes in nuclear organization [22,24]. The observed alterations in telomere structure in CML cells are consistent with this mechanism, as dedifferentiation is frequently associated with telomere instability and nuclear remodeling [25,26]. However, in cancer cells, this dedifferentiation process can also induce mitotic defects, chromosomal missegregation, and cell cycle perturbations, leading to apoptosis or cell cycle arrest [16].
Interestingly, while dedifferentiation is often associated with increased cellular plasticity, reversine may contribute to telomere and chromatin stability in CML cells. This suggests the possibility of a reduced cytotoxic impact on surviving cells rather than necessarily driving a shift toward a less differentiated state. Additionally, the notable reduction in nuclear volume observed in this study could imply that reversine influences nuclear compaction, a phenomenon that is frequently linked to apoptosis or chromatin condensation resulting from mitotic arrest [26]. Aurora kinases, particularly AURKA and AURKB, are pivotal regulators of mitosis and have been implicated in the pathogenesis of various malignancies, including CML [12]. Overexpression of these kinases has been linked to enhanced tumor cell proliferation and survival [13]. Reversine's inhibitory effect on Aurora kinases may disrupt mitotic progression, leading to mitotic defects and subsequent apoptosis in leukemic cells [2,7]. This mechanism is supported by studies showing that Aurora kinase inhibition can induce mitotic catastrophe and cell death in cancer cells [8].
The significant decrease in telomere aggregates observed in this study suggests that reversine prevents telomere clustering, a feature typically associated with chromosomal integrity [27]. Given the role of telomeres in genomic stability, the disruption of telomere clustering may contribute to mitotic defects and cellular stress, leading to a shift toward apoptosis rather than proliferation [28].
To comprehensively analyze the effects of reversine on telomere dynamics, we utilized TeloView® software [16,17], an advanced imaging and analytical tool that enables high-throughput 3D telomere analysis. TeloView® quantitatively assesses telomere organization within interphase nuclei, measuring parameters such as total telomere number, telomere signal intensity, aggregate formation (clustering of telomeres), a/c ratio (a measure of chromatin and cell cycle distribution), and nuclear volume. In cancer research, TeloView® has been instrumental in identifying telomeric signatures associated with genomic instability, disease progression, and response to therapy [15,16,17,18]. The present study demonstrated significant changes in telomere architecture after reversine treatment, as observed through a marked reduction in telomere aggregates, a decrease in overall telomere signal intensity, and alterations in nuclear volume, suggesting a potential role of reversine in modulating chromatin structure and telomere stability.
The clinical implications of these findings are significant. Targeting telomere dynamics and aurora kinase activity presents a promising therapeutic strategy in CML. Telomerase inhibitors have already shown potential in the treatment of myeloid malignancies [29,30,31], and the addition of aurora kinase inhibitors like reversine could enhance therapeutic efficacy by simultaneously disrupting multiple pathways critical for leukemic cell survival [32]. Moreover, the ability of reversine to induce changes in 3D telomere architecture may serve as a biomarker for treatment response, aiding in the stratification of patients and the optimization of therapeutic regimens.
In conclusion, this study underscores the multifaceted role of reversine in modulating telomere dynamics and Aurora kinase activity in CML cells. The integration of TeloView® for 3D telomere analysis provided valuable insights into the structural and functional changes induced by reversine, highlighting its potential as a therapeutic agent in the management of CML.
Acknowledgments
We thank Telo Genomics Corp. for the use of TeloView® software platform. The authors also thank the Genomic Centre for Cancer Research and Diagnosis (GCCRD) for imaging. The GCCRD is funded by the Canada Foundation for Innovation and supported by CancerCare Manitoba Foundation, the University of Manitoba and the Canada Research Chair Tier 1 (S.M.). The GCCRD is a member of the Canadian National Scientific Platforms (CNSP) and of Canada BioImaging.Research group in Molecular Epidemiology (EPIMOL), CNPq, Brazil. Association for Health Education & Research, Brazil. Genomic Medicine Study Group (GMEG).
Ethics Statement
Not applicable.
Data Availability Statement
Research data are not shared.
Conflicts of Interest
SM is founder, director, and chair of the clinical and scientific advisory board of Telo Genomics. No potential conflict of interest was reported by the other authors.
References
- Chen, S.; Zhang, Q.; Wu, X.; Schultz, P.G.; Ding, S. Dedifferentiation of Lineage-Committed Cells by a Small Molecule. J. Am. Chem. Soc. 2003, 126, 410–411. [Google Scholar] [CrossRef] [PubMed]
- D'Alise, A.M.; Amabile, G.; Iovino, M.; Di Giorgio, F.P.; Bartiromo, M.; Sessa, F.; Villa, F.; Musacchio, A.; Cortese, R. Reversine, a novel Aurora kinases inhibitor, inhibits colony formation of human acute myeloid leukemia cells. Mol. Cancer Ther. 2008, 7, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Satinover, D.L.; Leach, C.A.; Stukenberg, P.T.; Brautigan, D.L. Activation of Aurora-A kinase by protein phosphatase inhibitor-2, a bifunctional signaling protein. Proc. Natl. Acad. Sci. 2004, 101, 8625–8630. [Google Scholar] [CrossRef] [PubMed]
- Marumoto, T.; Zhang, D.; Saya, H. Aurora-A — A guardian of poles. Nat. Rev. Cancer 2005, 5, 42–50. [Google Scholar] [CrossRef]
- de Oliveira, F.M.; de Souza, V.G.; Carvalho, A.d.L.; Lizarte, F.S.; Miranda, C.S.S. Amplifications of AURKA and AURKB in a Burkitt lymphoma immunodeficiency-associated type: a case report. Einstein-Sao Paulo 2023, 21, eRC0378. [Google Scholar] [CrossRef]
- de Oliveira, F.M.; Rodrigues-Alves, A.P.N.; Lucena-Araújo, A.R.; Silva, F.d.P.; da Silva, F.B.; Falcão, R.P. Mantle cell lymphoma harboring Burkitt’s-like translocations presents differential expression of aurora kinase genes compared with others 8q abnormalities. Med Oncol. 2014, 31, 1–7. [Google Scholar] [CrossRef]
- Lucena-Araujo, A.R.; de Oliveira, F.M.; Leite-Cueva, S.D.; dos Santos, G.A.; Falcao, R.P.; Rego, E.M. High expression of AURKA and AURKB is associated with unfavorable cytogenetic abnormalities and high white blood cell count in patients with acute myeloid leukemia. Leuk. Res. 2011, 35, 260–264. [Google Scholar] [CrossRef]
- Yang, J.; Ikezoe, T.; Nishioka, C.; Udaka, K.; Yokoyama, A. Bcr-Abl activates AURKA and AURKB in chronic myeloid leukemia cells via AKT signaling. Int. J. Cancer 2013, 134, 1183–1194. [Google Scholar] [CrossRef]
- O'Hare, T.; Eide, C.A.; Deininger, M.W.N. Bcr-Abl kinase domain mutations, drug resistance, and the road to a cure for chronic myeloid leukemia. Blood 2007, 110, 2242–2249. [Google Scholar] [CrossRef]
- Weng, H.; Yuan, S.; Huang, Q.; Zeng, X.; Wang, X. STAT1 is a key gene in a gene regulatory network related to immune phenotypes in bladder cancer: An integrative analysis of multi-omics data. J. Cell. Mol. Med. 2021, 25, 3258–3271. [Google Scholar] [CrossRef]
- Li, Y.; Wang, S.; Wei, X.; Zhang, S.; Song, Z.; Chen, X.; Zhang, J. Role of inhibitor of yes-associated protein 1 in triple-negative breast cancer with taxol-based chemoresistance. Cancer Sci. 2018, 110, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Alves R, Carneiro-Lobo TC, Palacios A, de Souza W, Morgado-Díaz JA, El-Cheikh MC. Reversine triggers mitotic catastrophe in K562 chronic myeloid leukemia cells. Mol Pharmacol. 2016;90(3):323–35. [CrossRef]
- Williams, A.B.; Nguyen, B.; Li, L.; Brown, P.; Levis, M.; Leahy, D.; Small, D. Mutations of FLT3/ITD confer resistance to multiple tyrosine kinase inhibitors. Leukemia 2012, 27, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Marumoto, T.; Zhang, D.; Saya, H. Aurora-A — A guardian of poles. Nat. Rev. Cancer 2005, 5, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Vermolen, B.J.; Garini, Y.; Mai, S.; Mougey, V.; Fest, T.; Chuang, T.C.; Chuang, A.Y.; Wark, L.; Young, I.T. Characterizing the three-dimensional organization of telomeres. Cytom. Part A 2005, 67A, 144–150. [Google Scholar] [CrossRef]
- Chuang, T.C.Y.; Moshir, S.; Garini, Y.; Chuang, A.Y.-C.; Young, I.T.; Vermolen, B.; Doel, R.v.D.; Mougey, V.; Perrin, M.; Braun, M.; et al. The three-dimensional organization of telomeres in the nucleus of mammalian cells. BMC Biol. 2004, 2, 12–12. [Google Scholar] [CrossRef]
- Nagoshi, N.; Shibata, S.; Nakamura, M.; Matsuzaki, Y.; Toyama, Y.; Okano, H. Neural crest-derived stem cells display a wide variety of characteristics. J. Cell. Biochem. 2009, 107, 1046–1052. [Google Scholar] [CrossRef]
- de Oliveira, F.M.; Jamur, V.R.; Merfort, L.W.; Pozzo, A.R.; Mai, S. Three-dimensional nuclear telomere architecture and differential expression of aurora kinase genes in chronic myeloid leukemia to measure cell transformation. BMC Cancer 2022, 22, 1–11. [Google Scholar] [CrossRef]
- Mukaiyama, H.; Oku, M.; Baba, M.; Samizo, T.; Hammond, A.T.; Glick, B.S.; Kato, N.; Sakai, Y. Paz2 and 13 other PAZ gene products regulate vacuolar engulfment of peroxisomes during micropexophagy. Genes Cells 2002, 7, 75–90. [Google Scholar] [CrossRef]
- Hayashi, M.T.; Cesare, A.J.; Fitzpatrick, J.A.J.; Lazzerini-Denchi, E.; Karlseder, J. A telomere-dependent DNA damage checkpoint induced by prolonged mitotic arrest. Nat. Struct. Mol. Biol. 2012, 19, 387–394. [Google Scholar] [CrossRef]
- Kwon J, Kim Y, Jeong J, Lee H, Kim S. Reversine-induced dedifferentiation disrupts mitotic progression and causes chromosomal instability in cancer cells. Oncotarget. 2017;8(23):37864-78.
- Sun Y, Yang T, Wang N, Liang L, Feng Y. Reversine promotes the dedifferentiation of chondrocytes through chromatin remodeling and nuclear reorganization. Cell Physiol Biochem. 2014;34(1):189-200.
- Kim YJ, Matsumoto K, Kitajima S, Yoshizawa K, Fukui T, Fujii T, et al. Reversine-induced dedifferentiation is associated with telomere shortening and nuclear reorganization in mesenchymal cells. Exp Cell Res. 2015;336(2):277-85.
- Choi YJ, Park YJ, Park YM, Kim HS, Jung E. Reversine induces cell cycle arrest and apoptosis by interfering with mitotic spindle checkpoint in leukemia cells. PLoS One. 2017;12(7):e0182042.
- Brümmendorf, T.H.; Holyoake, T.L.; Rufer, N.; Barnett, M.J.; Schulzer, M.; Eaves, C.J.; Eaves, A.C.; Lansdorp, P.M. Prognostic implications of differences in telomere length between normal and malignant cells from patients with chronic myeloid leukemia measured by flow cytometry. Blood 2000, 95, 1883–1890. [Google Scholar] [CrossRef]
- Ramírez, M.J.; Surralles, J. Laser confocal microscopy analysis of human interphase nuclei by three-dimensional FISH reveals dynamic perinucleolar clustering of telomeres. Cytogenet. Genome Res. 2008, 122, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Rajkumar, S.V.; Jevremovic, D.; Kyle, R.A.; Shifrin, Y.; Nguyen, M.; Husain, Z.; Alikhah, A.; Jafari, A.; Mai, S.; et al. Three-dimensional telomere profiling predicts risk of progression in smoldering multiple myeloma. Am. J. Hematol. 2024, 99, 1532–1539. [Google Scholar] [CrossRef] [PubMed]
- Mai, S.; Garini, Y. The significance of telomeric aggregates in the interphase nuclei of tumor cells. J. Cell. Biochem. 2006, 97, 904–915. [Google Scholar] [CrossRef]
- Keller, G.; Brassat, U.; Braig, M.; Heim, D.; Wege, H.; Brümmendorf, T.H. Telomeres and telomerase in chronic myeloid leukaemia: impact for pathogenesis, disease progression and targeted therapy. Hematol. Oncol. 2009, 27, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Pandita, T.K.; Hunt, C.R.; Sharma, G.G.; Yang, Q. Telomeres and meiosis in health and disease. Cell. Mol. Life Sci. 2007, 64, 131–138. [Google Scholar] [CrossRef]
- Solovei, I.; Kreysing, M.; Lanctôt, C.; Kösem, S.; Peichl, L.; Cremer, T.; Guck, J.; Joffe, B. Nuclear Architecture of Rod Photoreceptor Cells Adapts to Vision in Mammalian Evolution. Cell 2009, 137, 356–368. [Google Scholar] [CrossRef]
- Taylor, S.; Peters, J.-M. Polo and Aurora kinases—lessons derived from chemical biology. Curr. Opin. Cell Biol. 2008, 20, 77–84. [Google Scholar] [CrossRef]
Figure 1.
Reversine decreases cell viability on both CML cell lines (K-562 and MEG-1). (A/B) Cell viability was determined by MTT assay for both cell lines treated with reversine (0, 5, 10, 20 and 30 μM) for 24, 48 and 72 hours. Values were normalized with DMSO. Results are shown as mean±SD of three independent experiments (p<0.0001), ANOVA test and Bonferroni post-test. (C/D) qPCR analysis of AURKA and AURKB mRNA expression in both cell lines treated with reversine (0, 5, 10, 20 and 30 μM) for 24 hours relative to DMSO, which was used as calibrator sample. The bar graph represents the mean±SD of three independent experiments.
Figure 1.
Reversine decreases cell viability on both CML cell lines (K-562 and MEG-1). (A/B) Cell viability was determined by MTT assay for both cell lines treated with reversine (0, 5, 10, 20 and 30 μM) for 24, 48 and 72 hours. Values were normalized with DMSO. Results are shown as mean±SD of three independent experiments (p<0.0001), ANOVA test and Bonferroni post-test. (C/D) qPCR analysis of AURKA and AURKB mRNA expression in both cell lines treated with reversine (0, 5, 10, 20 and 30 μM) for 24 hours relative to DMSO, which was used as calibrator sample. The bar graph represents the mean±SD of three independent experiments.
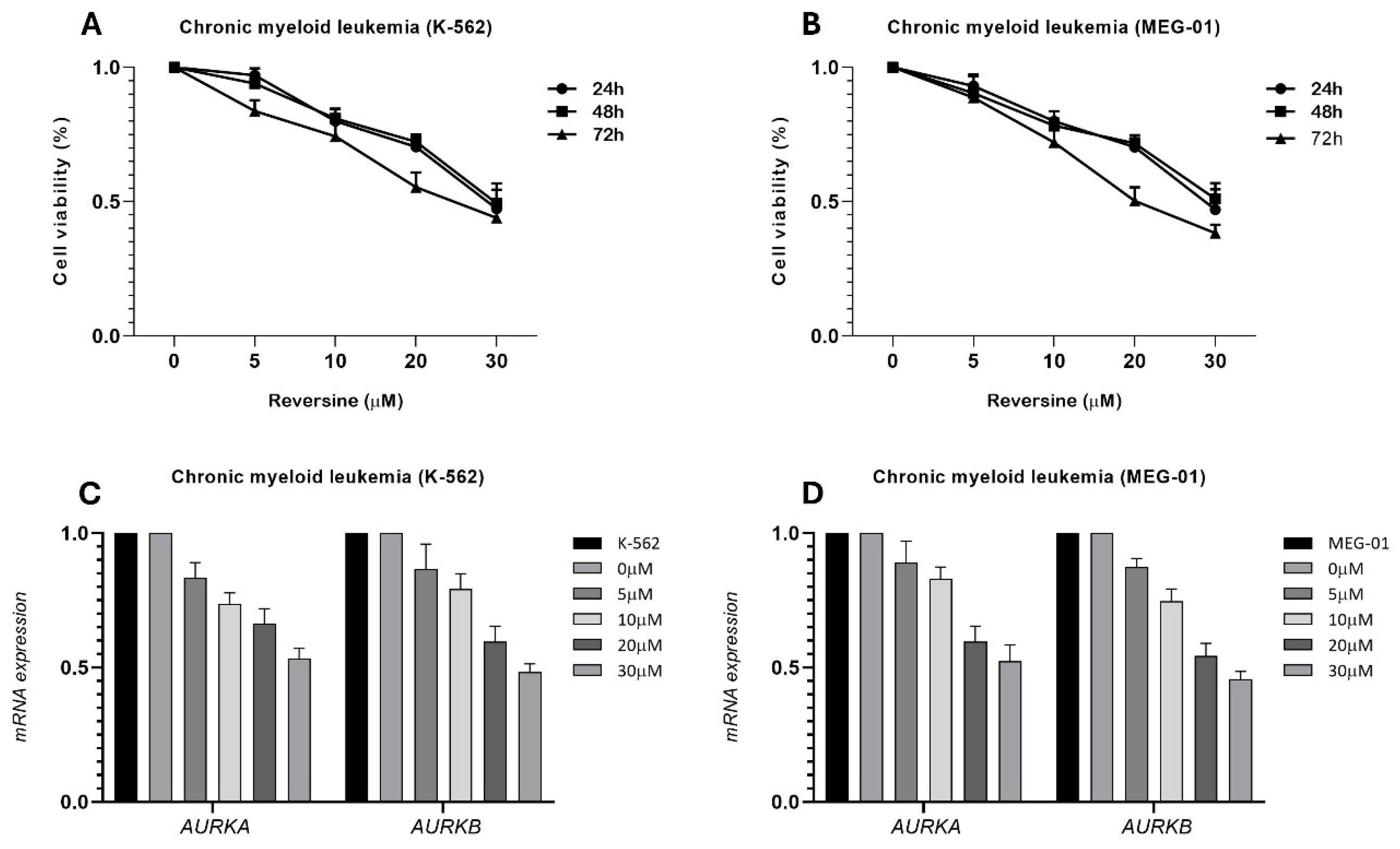
Figure 2.
3D Telomere Architecture in CML Cells. (A) Representative 3D nuclear telomere distribution (red) within the counterstained nucleus (blue) in untreated MEG-01 cells. Orange arrows highlight telomere aggregates. (B) 3D telomere distribution in MEG-01 cells treated with reversine, showing a reduced frequency of telomere aggregates.
Figure 2.
3D Telomere Architecture in CML Cells. (A) Representative 3D nuclear telomere distribution (red) within the counterstained nucleus (blue) in untreated MEG-01 cells. Orange arrows highlight telomere aggregates. (B) 3D telomere distribution in MEG-01 cells treated with reversine, showing a reduced frequency of telomere aggregates.
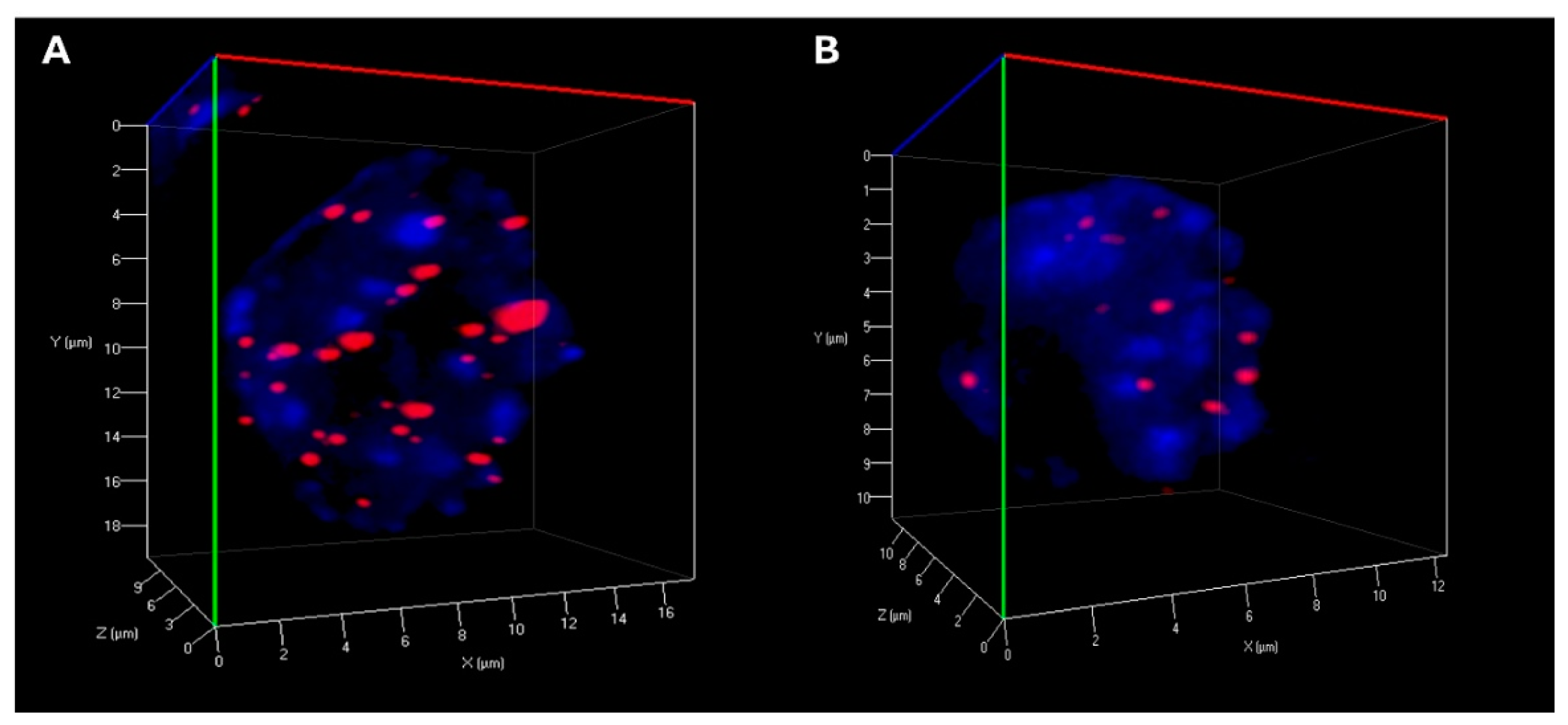
Figure 3.
Graph distribution of number of telomeres according to their intensity (length of telomeres) for CML cell treated and not treated with reversine [(A) K-562(reversine+), K-562, (B) MEG-01, and MEG-01(reversine+)]. The image represents the 3D telomere distribution of the 3D telomeric profile.
Figure 3.
Graph distribution of number of telomeres according to their intensity (length of telomeres) for CML cell treated and not treated with reversine [(A) K-562(reversine+), K-562, (B) MEG-01, and MEG-01(reversine+)]. The image represents the 3D telomere distribution of the 3D telomeric profile.
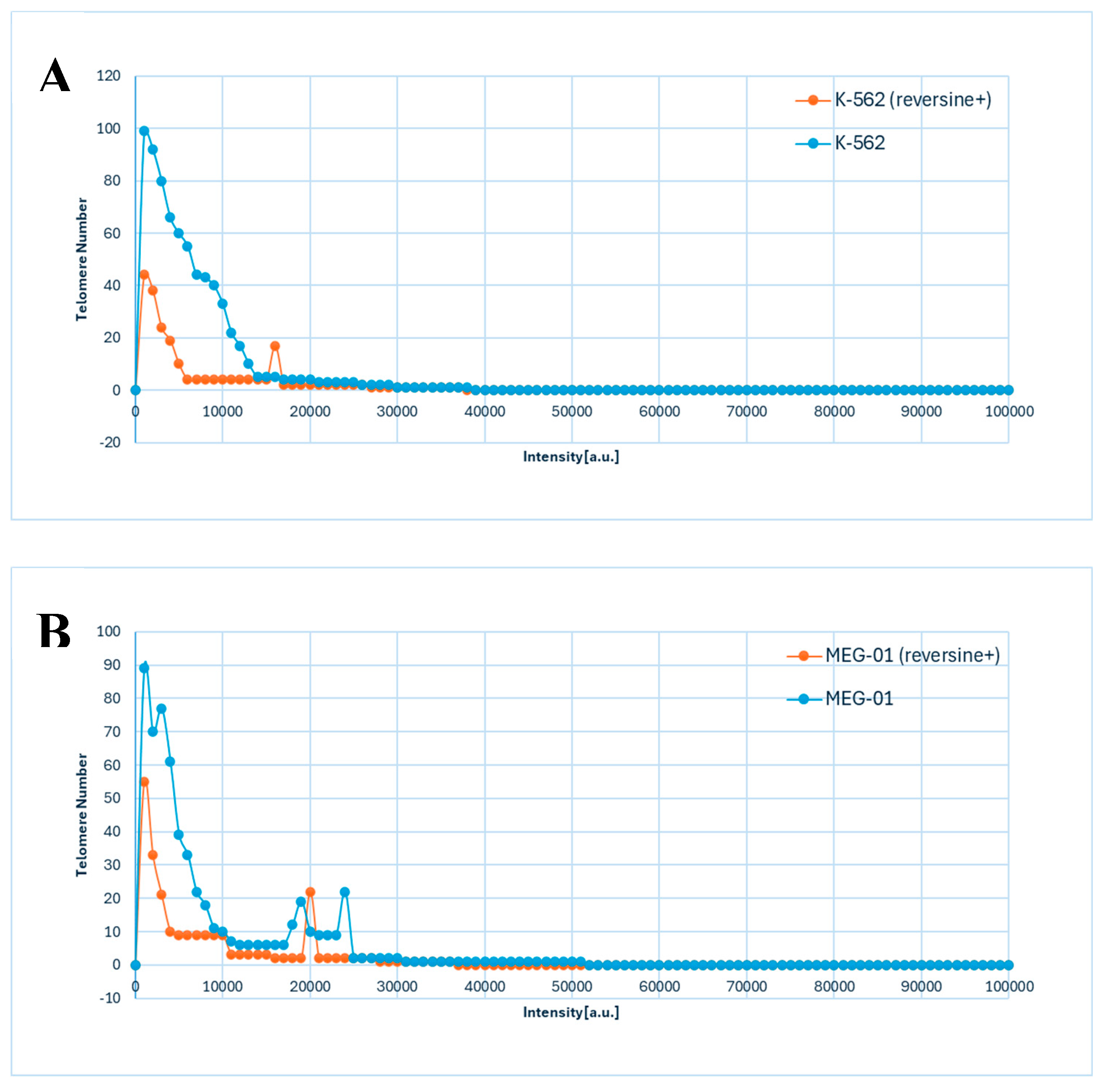

Table 1.
Statistical analysis of telomere parameters for CML cells (K-562 and MEG-01), after and before reversine treatment, based on interphase nuclei information.
Table 1.
Statistical analysis of telomere parameters for CML cells (K-562 and MEG-01), after and before reversine treatment, based on interphase nuclei information.
| CML cells | Total number of telomeres (mean ± SD) | Total number of telomere aggregates (mean ± SD) | Total intensity (mean ± SD) | Intensity of all signals (mean ± SD) | a/c ratio (mean ± SD) | Nuclear volume (mean ± SD) |
| K-562 | 56.27 ± 17.88 | 10.21 ± 3.89 | 643.123 ± 231.337 | 39.543 ± 2562 | 17.88 ± 2.97 | 547621 ± 176234 |
| K-562 (reversine+) | 22.39 ± 11.82 | 4.67 ± 2.65 | 433.332 ± 233.312 | 28.543 ± 2984 | 11.44 ± 3.89 | 266431 ± 134654 |
| p value | p<0.0001 | p<0.0001 | p<0.0001 | p<0.0001 | p<0.0001 | p<0.0001 |
| MEG-01 | 45.22 ± 12.21 | 9.44 ± 3.88 | 543.341 ± 189.132 | 26.876 ± 3241 | 16.43 ± 2.33 | 453765 ± 176432 |
| MEG-01 (reversine+) | 29.33 ± 9.67 | 5.55 ± 2.86 | 312.133 ± 109.102 | 19.432 ± 2987 | 9.66 ± 2.32 | 278543 ± 131762 |
| p value | p<0.0001 | p<0.0001 | p<0.0001 | p<0.0001 | p<0.0001 | p<0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



